1 Alcance y objetivos. - Muralmural.uv.es/rata3/arginine/Vision evolutiva de la Biosintesis de...
45
Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética 1 Alcance y objetivos. El principal objetivo de este resumen es revisar los aspectos principales de la biosíntesis de arginina con especial atención a su evolución filogenética. Igualmente, cuando sea posible, se prestará especial atención a los diferencies requerimientos de arginina en mamíferos centrándose en las causas que originan las diferencias. Hay que tener en cuenta que desde el punto de vista del análisis filogenético de genomas los límites entre especies pueden no coincidir con la visión bioquímica tradicional. No es objeto de este resumen, la revisión de los aspectos funcionales de la Arginina como tal, ni revisar las implicaciones del ciclo de la urea (que se expondrá en otro tema), ni repetir lo que ya está explicado en los diferentes artículos que tratan el tema. De esta forma para cada concepto expuesto se facilitaran las fuentes bibliográficas originales en donde se puede ampliar el tema, y únicamente de detallaran las figuras y conclusiones más importantes. Debido a las abundantes interrelaciones de la biosíntesis de la arginina con otros procesos metabólicos, el resumen se centrara únicamente en pocos conceptos importantes que sigan un hilo conductor y que puedan ser expuestos de forma amena en el espacio de media hora. Hoy día, el uso de las tecnologías de la información aplicadas a la biología es un campo innovador. Por tanto, en los ejemplos introduciremos el uso de algunas de las herramientas disponibles para la investigación de las reacciones metabólicas. 2 Importancia de la biosíntesis de Arginina. Aminoácidos esenciales. Aminoácidos funcionales. Aminoácidos proteicos. El pool de aminoácidos. 2.1 Los aminoácidos tienen dos facetas bioquímicas. Por una parte, los aminoácidos son los componentes constructivos de las proteínas ( los llamados “aminoácidos comunes”). pero no todos los aminoácidos existentes en los organismos forman parte de las proteínas. Cada especie tiene una composición diferente de aminoácidos. En el caso de la arginina se pueden observar diferencias sustanciales entre organismos superiores (Ball et al., 2007). El principal deposito de proteína corporal en todos los organismos es el músculo. Particularmente, el músculo tiene una estructura y composición bioquímica similar en todas las especies. Cuando se compara la composición de arginina en los músculos de los animales comprobamos que varia desde los 69 mg/gr del cerdo a 77 mg/gr en humanos, mientras que en peces el contenido es de 62mg/gr. Como se describe en el articulo de Ball (Ball et al., 2007), la composición de aminoácidos conviene representarla como porcentaje respecto al contenido en Lisina. Debido a que: (i) es el aminoácido limitante en la mayoría de los alimentos de la dieta de animales (sobre todo en piensos), (i) define la ingesta de proteínas necesarias para satisfacer las necesidades globales de aminoácidos en los animales. ( Pag 1 de 45). 12/01/2010 10:28:00
Transcript of 1 Alcance y objetivos. - Muralmural.uv.es/rata3/arginine/Vision evolutiva de la Biosintesis de...

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
1 Alcance y objetivos.
El principal objetivo de este resumen es revisar los aspectos principales de la biosíntesis de arginina con
especial atención a su evolución filogenética. Igualmente, cuando sea posible, se prestará especial
atención a los diferencies requerimientos de arginina en mamíferos centrándose en las causas que
originan las diferencias. Hay que tener en cuenta que desde el punto de vista del análisis filogenético de
genomas los límites entre especies pueden no coincidir con la visión bioquímica tradicional. No es objeto
de este resumen, la revisión de los aspectos funcionales de la Arginina como tal, ni revisar las
implicaciones del ciclo de la urea (que se expondrá en otro tema), ni repetir lo que ya está explicado en
los diferentes artículos que tratan el tema. De esta forma para cada concepto expuesto se facilitaran las
fuentes bibliográficas originales en donde se puede ampliar el tema, y únicamente de detallaran las figuras
y conclusiones más importantes. Debido a las abundantes interrelaciones de la biosíntesis de la arginina
con otros procesos metabólicos, el resumen se centrara únicamente en pocos conceptos importantes que
sigan un hilo conductor y que puedan ser expuestos de forma amena en el espacio de media hora.
Hoy día, el uso de las tecnologías de la información aplicadas a la biología es un campo innovador. Por
tanto, en los ejemplos introduciremos el uso de algunas de las herramientas disponibles para la
investigación de las reacciones metabólicas.
2 Importancia de la biosíntesis de Arginina. Aminoácidos esenciales. Aminoácidos
funcionales. Aminoácidos proteicos. El pool de aminoácidos.
2.1 Los aminoácidos tienen dos facetas bioquímicas.
Por una parte, los aminoácidos son los componentes constructivos de las proteínas ( los llamados
“aminoácidos comunes”). pero no todos los aminoácidos existentes en los organismos forman parte de
las proteínas. Cada especie tiene una composición diferente de aminoácidos. En el caso de la arginina se
pueden observar diferencias sustanciales entre organismos superiores (Ball et al., 2007).
El principal deposito de proteína corporal en todos los organismos es el músculo. Particularmente, el
músculo tiene una estructura y composición bioquímica similar en todas las especies. Cuando se compara
la composición de arginina en los músculos de los animales comprobamos que varia desde los 69 mg/gr
del cerdo a 77 mg/gr en humanos, mientras que en peces el contenido es de 62mg/gr.
Como se describe en el articulo de Ball (Ball et al., 2007), la composición de aminoácidos conviene
representarla como porcentaje respecto al contenido en Lisina. Debido a que: (i) es el aminoácido
limitante en la mayoría de los alimentos de la dieta de animales (sobre todo en piensos), (i) define la
ingesta de proteínas necesarias para satisfacer las necesidades globales de aminoácidos en los animales.
( Pag 1 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Figura 1.- Comparativa de la composición de aminoácidos en diferentes organismos. Fuente.- Ball, R. O., Urschel, K. L., and Pencharz, P. B. Nutritional consequences of interspecies
differences in arginine and lysine metabolism. J.Nutr. 137(6 Suppl 2), 1626S-1641S. 2007.
En la Figura 1 los valores relativos a la composición de lisina aparecen entre paréntesis. Es curioso
comprobar que de los cuatro mamíferos comparados, tres tienen una composición muy similar de
Arginina (105-109%), mientras que la rata (en rosa) tiene una composición similar al observado en aves
(94%). En el caso de los peces las diferencias se hacen mucho más grandes (únicamente un 65%).
Como primera conclusión podemos decir que los diferentes requerimientos de arginina ínterespecies van
a estar ligados a la demanda de proteína muscular.
En un segundo termino los aminoácidos tienen funciones distintas del componente estructural. En
concreto los aminoácidos pueden actuar como: (i) moléculas de señalización celular, (ii) reguladores de la
expresión génica (iii) precursores para la síntesis de hormonas y de de sustancias nitrogenadas de bajo
peso molecular . Los metabolitos de sus reacciones catabólicas y anabólicas (por ejemplo, el óxido
nítrico, poliaminas, glutatión, taurina, hormonas tiroideas, y la serotonina) son igualmente necesarios en
algunas funciones vitales del organismo.
De esta forma, algunos aminoácidos regulan las vías metabólicas claves necesarias para el crecimiento,
reproducción, y la inmunidad. Estos aminoácidos se denominan funcionales, e incluyen la arginina,
cisteína, glutamina, leucina, prolina y el triptófano. Suplementos dietéticos con uno o una mezcla de estos
aminoácidos puede ser beneficioso para (i) aminorar los problemas de salud en las distintas etapas del
ciclo de vida (por ejemplo, el crecimiento fetal, la mortalidad neonatal, la disfunción intestinal asociada al
destete, obesidad, diabetes, la infertilidad); (2) la optimización del crecimiento muscular, la producción de
( Pag 2 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
leche, huevos, mejorar la calidad de la carne y el rendimiento deportivo, evitando la acumulación de grasa
en exceso.
Figura 2.- Funciones asociadas a la arginina y los metabolitos resultantes de su biosíntesis. Fuente.-
En el articulo de (Wu et al., 2009) “Arginine metabolism and nutrition in growth, health and disease ”
podemos encontrar una extensa lista de las diferentes funciones asociadas a la arginina y a otros
aminoácidos.
2.2 Aminoácidos esenciales y no esenciales.
Los aminoácidos esenciales son aquellos que el organismo no puede generar mediante el metabolismo. La
fuente de estos aminoácidos es la ingesta directa a través de la dieta. Por ejemplo, el metabolismo de los
animales no es capaz de llevar a cabo la síntesis endógena de los aminoácidos aromáticos, los de cadena
ramificada, así como la metionina y lisina, que también pertenecen a los aminoácidos esenciales.
El valor nutritivo de las proteínas de la dieta es en muchos casos decisivo y depende del contenido en
aminoácidos esenciales. Por ejemplo, las proteínas vegetales tiene un bajo contenido en lisina y
( Pag 3 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
metionina, mientras que las proteínas de origen animal contienen todos los aminoácidos en proporciones
equilibradas.
Los aminoácidos proteinogénicos (que forman parte de la proteínas) no esenciales son los que el
organismo es capaz de sintetizar para mantener los requerimientos metabólicos. Se pueden dividir en
cinco familias en relación con su biosíntesis. Los miembros de cada familia son derivados de precursores
comunes, que son producidos en el ciclo de los ácidos tricarboxílicos o en el metabolismo de los hidratos
de carbono ( Figura 4 ).
Figura 3.- Clasificación nutricional de los aminoácidos.
Figura 4.- Visión general de la síntesis de aminoácidos.
( Pag 4 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Las plantas y los microorganismos son capaces de sintetizar todos los aminoácidos, pero durante el curso
de la evolución, los mamíferos han perdido la capacidad de sintetizar aproximadamente la mitad de los 20
amino ácidos proteinogénicos (Caetano-Anolles et al., 2009; Payne and Loomis, 2006; Peregrin-Alvarez
et al., 2009). Por ejemplo, en el caso de la arginina muchos de los enzimas de la ruta biosintética se han
perdido durante la evolución, Figura 5.
Figura 5.- Perdida de las rutas metabólicas en el transcurso de la evolución. (Payne and Loomis, 2006)
En conclusión: (i) En los organismos existen dos fuentes diferenciadas de suministro de Arginina (y en
general de aminoácidos esenciales), las proteínas de la dieta y la síntesis endógena. (ii) El consumo de
arginina del organismo esta dirigido principalmente a la síntesis de proteínas musculares y en un segundo
termino a regular funciones metabólicas.
( Pag 5 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
2.3 El “pool” de aminoácidos.
Los aminoácidos libres en el organismo se encuentran en solución en los diferentes líquidos corporales:
intracelular, intersticial, plasma, linfa y otros. Si bien las concentraciones de estos compuestos son
diferentes en cada uno de los compartimientos líquidos considerados existe un continuo intercambio entre
éstos a través de las distintas membranas celulares, capilares y otras.
El concepto de pool es perfectamente aplicable para el conjunto de todos los aminoácidos libres presentes
en el organismo incluida la arginina. La cantidad y concentración de cada uno de los aminoácidos del
pool es biológicamente constante en el sentido de que sus variaciones se producen dentro de unos
determinados márgenes. La constancia del pool de aminoácidos refleja un estado de equilibrio dinámico
entre los procesos que le aportan aminoácidos al pool y los que consumen.
Figura 6.- El pool de aminoácidos. Este modelo muestra cómo el equilibrio entre los procesos catabólicos y anabólicos ejerce un control homeostático sobre el pool de aminoácidos, Fuente.- Nature. 1998 Feb 26;391(6670):843, 845.
Protein metabolism. Slow and fast dietary proteins.
Frühbeck G. PMID: 9495333 [PubMed - indexed for MEDLINE]
El pool de aminoácidos en las células se genera por (i)el aporte de aminoácidos en la dieta, (ii)como
resultado de la degradación de las proteínas existentes dentro de la célula y (ii) mediante síntesis
endógena. Todas las proteínas dentro de las células tienen una vida media (t1 / 2) determinada, algunas de
muy corta duración (de 5 a 20 minutos) y otras están presentes durante períodos prolongados, con una
vida media de muchas horas o incluso días. Así, las proteínas son continuamente sintetizadas y
degradadas en el organismo.
( Pag 6 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
( Pag 7 de 45). 12/01/2010 10:28:00
La proteína muscular se degrada durante los períodos de ayuno, y los aminoácidos son utilizados para la
gluconeogénesis. Después de la ingestión de proteínas en la dieta, se resintetiza proteína muscular. Sólo
aproximadamente el 6% (aproximadamente 10 g) de la proteína que entra en el tracto digestivo se excreta
en las heces cada día. El resto se recicla. Los lisosomas participar en el proceso de degradación de las
proteínas celulares. Dentro de los lisosomas, la familia de la catepsina proteásas degrada las proteínas
ingeridas por autofagia en aminoácidos individuales. Los amino ácidos reciclado pueden salir del
lisosoma y reincorporarse al pool intracelular de aminoácidos. Aunque los detalles de cómo se induce la
autofagia aún se desconocen,
2.4 Como modelizar las diferencias entre especies y los requisitos dietarios de arginina.
Las diferentes especies tienen diferentes capacidades de síntesis endógena de arginina, y por lo tanto, el
requisito dietario de la arginina varía ampliamente desde el 1,4% de la proteína cruda en la especie
porcina (en ejemplares de 100 kg),a más del 5% en pollos de engorde y en el salmón del Pacífico. Este
efecto se agudiza durante las etapas de crecimiento debido a la necesidad extra de requerimiento proteico.
Estas observaciones resultan en la imposibilidad de comparar los requerimientos de arginina de una forma
objetiva de una especie a otra. Es decir, el metabolismo funcional del cerdo no tiene porque coincidir con
el de los humanos, y por tanto los requerimientos mínimos serán distintos, y más aún si comparamos entre
especimenes en diferentes estadios de crecimiento.
Para medir los requerimientos mínimos la dieta (Ball et al., 2007) proponen un modelo que aglutina los
requerimientos metabólicos, la síntesis endógena y el aporte dietario. El gráfico de la Figura 7 representa
los requisitos metabólicos de arginina y su relación con la contribución de la dieta y la síntesis endógena
según el modelo propuesto. La diferencia entre el requerimiento metabólico y la tasa máxima de síntesis
endógena corresponde al requerimiento mínimo de arginina en la dieta. El gráfico de la Figura 8
representa las posibles respuestas metabólicas en función de la ingesta de arginina.
Figura 7.- Particionamiento de los requerimientos metabólicos y dietario de de arginina en los animales
Figura 8.- Las respuestas metabólicas a la ingesta dietética de arginina en relación con los cambios en la síntesis de proteínas.
Fuente.- Nutritional consequences of interspecies differences in arginine and lysine metabolism. Ball RO, Urschel KL, Pencharz PB. J Nutr. 2007 Jun;137(6 Suppl 2):1626S-1641S. Review.PMID: 17513439 Free article

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
La cuantificación de todos estos niveles en las diferentes especies es de gran ayuda para completar la
comprensión del metabolismo de la arginina, y permite comparaciones de datos que este momento no son
posibles.
Como conclusión , de momento los datos de “dieta mínima” únicamente sirven como punto de partida, y
en ningún caso son directamente extrapolables entre especies.
2.5 Los diferencias de requerimientos de arginina entre especies.
Las diferencias en los requerimientos de arginina entre las diversas especies como los omnívoros (los
seres humanos, cerdos, ratas, perros), carnívoros (gatos), herbívoros (conejos, caballos), rumiantes
(ganado vacuno), aves de corral, y el pescado, se revisan en detalle en el artículo de Ball. (Ball et al.,
2007).
2.5.1 El requerimiento de arginina en felinos.
En el articulo de Baker en 2005 (Baker, 2005) se formulan algunas cuestiones clave acerca de los
requerimientos de aminoácidos en la dieta. Una de ellas es ¿Por qué algunas especies de felinos mueren a
las 24 horas de recibir una dieta libre de Arginina?.
Figura 9.- Actividad de las enzimas P5CS y OAT en gatos y ratas
Los felinos han evolucionado como carnívoros, y como tales, tienen muchas particularidades
nutricionales (Morris, 2002; Morris, 1985). A diferencia de los mamíferos omnívoros como por ejemplo
los perros y cerdos, los gatos tienen bajos niveles de enzimas clave para la síntesis ornitina a partir del
ácido glutámico. La ingesta de una dieta libre de arginina causa graves efectos letales, como la anorexia,
hiperamonemia, vómitos, ataxia, e incluso la muerte. Los gatos tienen una baja capacidad para la
biosíntesis de ornitina en la mucosa intestinal a partir de ácido glutámico, debido a la baja actividad del
enzima pirrolina-5-carboxilato sintetasa (P5CS) y ornitina aminotransferasa (OAT). Así la ornitina se
vuelve crítica para el hígado para tomar el amoníaco como carbamilfosfato y eliminarlo mediante el ciclo
de la urea. La consecuencia inmediata es la rápida aparición de hiperamonémia cuando reciben una dieta
libre de arginina. Por lo tanto, en los gatos la principal limitación en la síntesis de novo arginina es el bajo
( Pag 8 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
nivel de las enzimas intestinales necesarios para la síntesis de la ornitina, que limita la formación de
citrulina y las concentraciones de citrulina en circulación y, posteriormente, renal la síntesis de arginina.
Figura 10.- Síntesis de arginina en animales.
2.5.2 Los requisitos de arginina en perros.
Los perros tienen necesidades dietéticas de arginina (0,36 g / kg-1× d-1 en cachorros y 0,06 g / kg-1× d-1
en perros adultos, suponen un 3,5% de CP en) intermedias a las del gato y el ratón. A diferencia de los
( Pag 9 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
gatos, los perros son estrictamente carnívoros, y esto puede explicar en parte los menores requerimientos
de arginina.
Figura 11.- Requerimientos de arginina en carnívoros.
2.5.3 Los requisitos de arginina en los cerdos.
La arginina es considerada como un aminoácido condicionalmente indispensables en los cerdos. Hay un
requisito de la dieta en el recién nacido, pero en el adulto sano, la síntesis endógena es suficiente para
satisfacer todos los requerimientos metabólicos. El metabolismo de la arginina en cerdos es muy similar
al de humanos.
2.5.4 Los requisitos de arginina en ratas y ratones.
Gran parte de la investigación pionera sobre el metabolismo de la arginina y del ciclo urea se ha llevado
a cabo en roedores. Sin embargo, hay elementos característicos en el metabolismo de la arginina en las
rata que tienen que ser tenidos en cuenta al comparar los requisitos de las ratas con otras especies. En
concreto, se debe considerar que las ratas practican coprofagía. La coprofagía proporciona una cantidad
desconocida de aminoácidos a la dieta, a través la flora microbiana. Por tanto en los estudios de
requerimientos de la dieta en las ratas puede haber una subestimación de la obligación alimentaria real.
Para ratas la arginina es un aminoácido indispensable durante el crecimiento, pero no para las ratas
adultas. Sin embargo, en ratas adultas anteriormente sometido a la malnutrición proteica, el aumento de
incrementos de arginina la dieta resultó en un lineal positiva de respuesta con respecto al aumento de peso
( Pag 10 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
y la retención de nitrógeno y de una respuesta lineal negativa para la excreción renal. Por lo tanto, en
algunos casos la arginina puede ser indispensable para las ratas maduras. Aunque este efecto no es
extrapolable directamente en los seres humanos u otras especies.
El requisito de la arginina dietética para ratas en crecimiento se basa en una ingesta de proteína cruda del
15% . Como en otras especies, los requisito de arginina para la generación de músculo y de excreción de
los metabolitos de nitrógeno, en ratas aumentan junto con el contenido de proteínas de la dieta. Esto es
probablemente debido al papel de la arginina en el ciclo de la urea, y el hecho de que el aumento del
nivel de proteínas hace que aumente la actividad de la enzimas del ciclo de la urea. Por lo tanto, el
contenido de proteína de de la dieta, especialmente el exceso de aminoácidos, debe tenerse en cuenta al
determinar si la ingesta de la arginina es adecuada.
2.5.5 Arginina metabolismo y las necesidades de los rumiantes.
Se calcula que el requisito de la arginina en la dieta (vaca 0,17 g / kg-1× d-1) es muy bajo en comparación
con las necesidades alimenticias de otros más mamíferos (0.30-0.50 g / kg-1× d-1, Como se discutió
anteriormente). Sin embargo, este dato se basa únicamente en la composición de aminoácidos de la
proteína del cuerpo, y puede estar subestimado. El hígado de los rumiantes tiene una actividad detectable
de todos los enzimas del ciclo de la urea . Sin embargo, a diferencia de otras especies de mamíferos, se ha
comprobado que un aumento en el consumo de arginina no afecta a la actividad hepática de la arginasa.
Los rumiantes reutilizan el nitrógeno excretando la urea en el tracto gastrointestinal, donde es utilizado
por la microflora para sintetizar aminoácidos.
2.5.6 Observaciones generales sobre el metabolismo de la arginina a través de las especies.
La investigación sobre el metabolismo de arginina en las especies de mamíferos plantea muchos puntos
interesantes para el debate:
En todas las especies estudiadas, la citrulina es un precursor de la arginina más eficaz que cualquiera de
ornitina y prolina, que implica que es la formación la citrulina y no la conversión de citrulina en arginina
es el paso limitante para la síntesis endógena de arginina.
1. La causa de la limitación de la síntesis de citrulina varía entre las especies en función de las
enzimas presentes. Por ejemplo, la limitación en los pollos y el pescado se debe a que la urea no
se excretan como sus residuos nitrogenados y la falta de actividad mitocondrial de CPS I,
Mientras que en los carnívoros que la razón es la baja actividad de P5CS y OAT. En los
mamíferos, la razón por la que la formación de citrulina es limitante para la síntesis de arginina
no se ha determinado de manera concluyente, pero las investigaciones recientes sugieren que el es
la síntesis de N-acetilglutamato la que a su vez limita la síntesis de la carbamilfosfato.
2. Los síntomas de deficiencia de arginina son más graves en los carnívoros (caso de los felinos) ,
seguido de los omnívoros, y son menos graves en los herbívoros. Los omnívoros pueden mostrar
( Pag 11 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
una amplia gama de síntomas relacionados con la carencia. Los síntomas de la deficiencia son
mas graves en los jóvenes que en los adultos.
3. Las especies con prácticamente ninguna capacidad para la síntesis endógena de arginina, de
pescado y pollos, son las más altas a las necesidades alimenticias de arginina a pesar de que no
requieren de arginina para la función de ciclo de la urea. Por tanto, de la capacidad para la síntesis
endógena de arginina es el principal determinante de la obligación de arginina dietética. Esto
implica que todas las futuras investigaciones sobre los requisitos de la arginina y el metabolismo,
independientemente de la especie, debería incluir una medición de la in vivo tasa de síntesis
endógena.
4. Debido a los requisitos de arginina son más altos en las especies sin la función del ciclo de la
urea, se demuestra que las principales metabólicos el uso de arginina esta destinado a la síntesis
de proteínas. Se estima que El 70% del uso diario es de arginina para las proteínas de síntesis. Por
lo tanto, aunque las otras funciones metabólicas de la arginina son fundamentales para el
crecimiento y la salud, la síntesis de proteínas sigue siendo el componente principal de la
obligación de arginina metabólico.
5. La contribución potencial de la síntesis microbiana de arginina en el intestino de los animales ,
parece que no se han investigado a fondo. Los conejos y los roedores pueden adquirir el consumo
significativo de los aminoácidos de coprofagía, y los rumiantes como hemos visto por reciclado
en el intestino.
2.6 Implicaciones de las comparaciones entre especies para la nutrición humana.
Actualmente no hay recomendaciones dietéticas para los seres humanos. Sin embargo, en las principales
especies experimentales (cerdos y ratas) existen requisitos de dietarios de arginina durante el
crecimiento, ¿Son los seres humanos la excepción?. Dada la dificultad de la experimentación en seres
humanos, las conclusiones (o supuestos requerimientos) tienen que ser extrapoladas a partir de
investigaciones en animales y de datos clínicos.
Un breve resumen de los datos clínicos en humanos es la siguiente.
1. Los bebés prematuros pueden ser particularmente susceptibles a la deficiencia de arginina. Por
ejemplo, recién nacidos con algunas enfermedades como la enterocolitis necrotizante y la
hipertensión pulmonar persistente se han relacionado con una deficiencia de arginina.
2. El tratamiento con suplementos de arginina ha resultado en una mejoría de los cuadros clínicos.
Igualemenete, los bebes con hiperamonémia tratados con suplementos de arginina aliviaron los
síntomas.
( Pag 12 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
3. Los adultos sanos no parecen verse afectados negativamente por una dieta libre de arginina sin
embargo no hay datos concluyentes sobre los posibles requerimientos metabólicos durante
enfermedades o traumatismos prolongados.
En conjunto, estas observaciones sugieren que incluso la combinación de la síntesis endógena de arginina
con arginina de la dieta, puede no ser suficiente para la salud neonatal. En particular cuando la salud está
comprometida ya sea por un parto prematuro o por una complicación intestinal. La pregunta sigue siendo:
¿cuáles son las metabólicas y dietéticas los requisitos de arginina para los recién nacidos y los niños en
crecimiento?
Recientemente, se ha publicado un análisis de los límites de seguridad en suplementos de diversos
aminoácidos (Shao and Hathcock, 2008), resultando ser de 20 gramos día.
Figura 12.- Límites de seguridad de suplementos de aminoácidos.
2.7 El ciclo de la urea y su implicación en la biosíntesis de arginina.
Las consecuencias evolutivas del ciclo de la urea han sido tratadas en diversos artículos históricos
(Mommsen and Walsh, 1989; Takiguchi et al., 1989; Wright, 1995).
La mayoría de especies acuáticas, como por ejemplo los peces óseos, excretan el nitrógeno amínico en
forma de amoníaco por lo que se les llama animales amonotélicos; la mayoría de animales terrestres son
ureotélicos, excretan el nitrógeno amínico en forma de urea; las aves y los reptiles son uricotélicos,
excretan el nitrógeno amínico en forma de ácido úrico. En los organismos ureotélicos, el amoníaco
depositado en las mitocondrias de los hepatocitos se convierte en urea mediante el ciclo de la urea. La
producción de urea tiene lugar casi exclusivamente en el hígado y representa el destino de la mayor parte
del amoníaco allí canalizado. La urea pasa al torrente sanguíneo y de ahí a los riñones y se excreta en la
orina.
Las diferencias entre especies en el mecanismo de excreción de desechos nitrogenados tienen un impacto
significativo sobre los requisitos de la arginina metabólicas y de dieta. Por ejemplo, sólo los animales
( Pag 13 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
ureotélicos requieren arginina el funcionamiento del ciclo de la urea, la hiperamonémia es la
consecuencia metabólica de un aporte insuficiente de arginina en la dieta de estas especies. Sin embargo,
especies como aves y peces usan medios alternativos para la excreción de los de residuos de nitrógeno, y
la deficiencia en la dieta de arginina reduce la tasa de crecimiento, pero no causa hiperamonemia.
La arginina desempeña un papel clave en la regulación de la urea génesis en mamíferos. La principal
prueba de ello es el hecho de que de todos los intermediarios de ciclo de la urea, la arginina es el más
consumido por el hígado dado que la absorción de la ornitina es menor y la de la citrulina es
insignificante, incluso en los altos niveles fisiológicos.
La capacidad metabólica para tratar el exceso de consumo de arginina en la dieta no parece estar
relacionada con el método de excreción de nitrógeno, sino más bien con la capacidad para metabolizar el
exceso de proteínas. Las especies carnívoras, por ejemplo, tienen altos requerimientos dietéticos de
arginina, pero fácilmente puede degradar y excretar grandes cantidades de residuos de nitrógeno sin
efectos perjudiciales, mientras que los animales omnívoros suelen tener un menor requerimiento debido a
una mayor síntesis endógena.
( Pag 14 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
3 La biosíntesis de arginina
3.1 Visión general de la biosíntesis de arginina
Como hemos visto en secciones anteriores, L-Arginina se clasifica normalmente como un aminoácido
condicionalmente esencial o semi-esencial porque la capacidad del cuerpo para sintetizar cantidades
suficientes para satisfacer sus necesidades varía según la edad, el estadio de desarrollo y las posibles
enfermedades o lesiones. Es uno de los aminoácidos metabólicamente más versátiles, dando lugar a óxido
nítrico (NO), la urea, ornitina, citrulina, creatina, agmatina, glutamato, prolina y poliaminas.
Figura 13.- Fuentes y destinos de arginina
Por lo tanto, no sorprende que su metabolismo sea complejo y altamente regulado. Esta complejidad es
consecuencia no sólo de la diversidad de las enzimas implicadas en el metabolismo de la arginina y sus
metabolitos, sino también de los patrones específicos de expresión de las enzimas.
Básicamente, la L-arginina se sintetiza a partir de la citrulina por la acción secuencial de las enzimas
citosólicas argininosuccinato sintetasa (ASS - EC 6.3.4.5) y la argininosuccinato liasa ( ASL - EC
4.3.2.1). Particularmente, la Citrulina en los mamíferos puede sintetizarse por diversas rutas:
• a partir de L-arginina a través de NOS, (cíclica)
• ornitina vía catabolismo de la prolina y glutamina / glutamato,
• a través de ADMA DDAH (degradación de proteinas).
( Pag 15 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Figura 14.- Visión general de las rutas de biosíntesis de arginina en mamiferos ADC, arginina decarboxilasa; AGAT, arginina: glicina amidinotransferasa; ARG, arginasa, ASL, argininosuccinato liasa, ASS,
argininosuccinato sintetasa; DDAH, dimetilarginina dimethylaminohydrolase; Me2, dimetil, NO, óxido nítrico; OAT, ornitina
aminotransferasa; ODC, ornitina descarboxilasa; OTC, ornitinatranscarbamilasa; P5C, L-1-pirrolina-5-carboxilato; PRMT, proteína-
metiltransferasa arginina.
Nota: P5C está en equilibrio químico con L-glutamato semialdehído a través de una reacción enzimática espontánea (no mostrado). Las
lineas a trazos señalan la inhibición de la óxido nítrico sintasa (NOS) y arginasa por dimetilarginina (ADMA) y NG-hidroxi-L-arginina
(NOHA)
Es importante señalar que las vías que unen a la L-arginina, el glutamato y la prolina son bidireccionales.
De esta forma, la utilización neta o producción de estos aminoácidos es altamente dependiente del tipo
celular y del grado de desarrollo (Figura 15).
En los mamíferos, la vía más importante de aporte de arginina es la ruta a través del eje intestino-renal. Es
decir:
1. las células epiteliales del intestino delgado producen citrulina principalmente a partir de la
glutamina y el glutamato. La citrulina es vertida al riego sanguíneo y transportado al riñón,
2. las células del túbulo proximal del riñón, extraen citrulina de la circulación y la convierte a la L-
arginina, que se devuelve a la circulación.
( Pag 16 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
La Síntesis de L-arginina desde la citrulina también ocurre otros tejidos, pero la aportación que supone al
pool de arginina es muy baja. Se ha observado también que el deterioro del intestino delgado o de la
función renal puede reducir la síntesis endógena de arginina, lo que aumenta la exigencia de esta en la
dieta.
La capacidad para la síntesis de arginina puede verse afectada marcadamente por la actividad de las NOS,
ya que la citrulina puede ser reciclada a L-arginina en una vía conocida como el ciclo citrulina-NO o
arginina- citrulina. Esto se demuestra por el hecho de que en muchos tipos de células la citrulina puede
sustituir a la L-arginina en cierta medida en el apoyo a la síntesis de NO. Sin embargo, el reciclado no es
cuantitativo, porque la citrulina se
acumula junto con nitratos y
nitritos, los productos finales
estables de del metabolismo del
NO.
Figura 15.- Comparación de las vías biosintéticas de arginina en neonatos y adultos.
Figura 1.- Implicación de P5CS en la biosíntesis de Arginina.
( Pag 17 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
4 Evolución filogenética de la síntesis de Arginina.
En la bibliografía la evolución filogenética de la biosíntesis de Arginina se puede ha abordado desde dos
puntos de vista: (i) la evolución de las rutas biosintéticas globales de los aminoácidos, y (ii) la evolución
independiente de cada uno de los enzimas implicados.
4.1 Evolución filogenética de la ruta de síntesis de Arginina.
Como hemos visto en puntos anteriores (2.2), la evolución filogenética de la síntesis de Arginina se ha
tratado en diversos artículos recientes (Bono et al., 1998; Caetano-Anolles et al., 2009; Cunchillos and
Lecointre, 2003; Cunchillos and Lecointre, 2005; Cunchillos and Lecointre, 2007; Fondi et al., 2007;
Hernandez-Montes et al., 2008; Peregrin-Alvarez et al., 2009; Perlstein et al., 2007; Xu et al., 2000). No
obstante aun sigue siendo un tema “candente” en biología evolutiva de mamíferos debido a que la
cantidad de “genomas” publicados sigue siendo limitada. Por otra parte, algunos de los mecanismos de
reacción de los enzimas no están bien determinados, lo que complica el análisis de los árboles
filogenéticos.
4.2 La evolución desde el LUCA.
En 1997 Brown (Brown and Doolittle, 1997) ya habían tratado la evolución filogenética de la síntesis de
arginina desde el punto de vista de la transición de los organismos eucariotas a procariotas. El articulo de
Xu en 2007 (Xu et al., 2007) abre de nuevo el debate de evolución filogenética de la biosíntesis de
arginina desde el punto de vista de los microorganismos.
Como ya comentamos en un epígrafe anterior en el transcurso de la evolución algunas de las vías
metabólicas de los aminoácidos se han perdido (Payne and Loomis, 2006), y otras han evolucionado
desde el LUCA (last universal common ancestor). El caso de la arginina es un buen ejemplo que ilustra
todas las formas de evolución en un solo sustrato. Los estudios evolutivos suponen que el LUCA es el
hipotético último organismo del cual descendemos todos los existentes. Este organismo disponía de un
reducido numero de genes que codificaban un reducido numero de proteínas y por tanto enzimas. Desde
este punto de vista los enzimas estos deberían ser multifuncionales, es decir, un enzima era capaz de
catalizar diversas reacciones metabólicas. Los genes originales, pueden duplicarse dando lugar a genes o
proteínas “homologas” y estas evolucionar independientemente, e incluso alguna de ellas perderse en el
trascurso de la evolución. De esta forma, la evolución independiente de los genes duplicados puede dar
lugar a la especialización de algunos de los enzimas, y a la divergencia de las rutas metabólicas.
La Figura 16 muestra la relación entre la síntesis de arginina en los microorganismos y en los mamíferos.
Téngase en cuenta que la vía de síntesis hepática en mamíferos forma parte del mecanismo de eliminación
de nitrógeno mediante urea, y no aporta Arginina al “pool”, por lo que no tendremos en cuenta las rutas
catabólicas de arginina (Arginasas y NOS).
( Pag 18 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Figura 16.- Biosíntesis de arginina en los microorganismos, y comparación con la ruta fisiológica de biosíntesis en mamíferos.
La primera conclusión que se observa es que tanto en microorganismos como en mamíferos la ruta
biosintética parte de Glutamato y converge en Ornitina. La segunda es que en microorganismos existen
varias alternativas para sintetizar Arginina a partir de glutamato.
4.3 Perspectiva Genómica de los orígenes y evolución de las redes de síntesis de aminoácidos.
Mediante un analisis de las rutas metabolicas y la información disponible en las bases de datos
metabolicas, Montes et. al. (Hernandez-Montes et al., 2008) reconstruyen las redes biosinteticas de los 20
aminoacidos estandar y la densidad de los enzimas homologos implicados en las rutas.
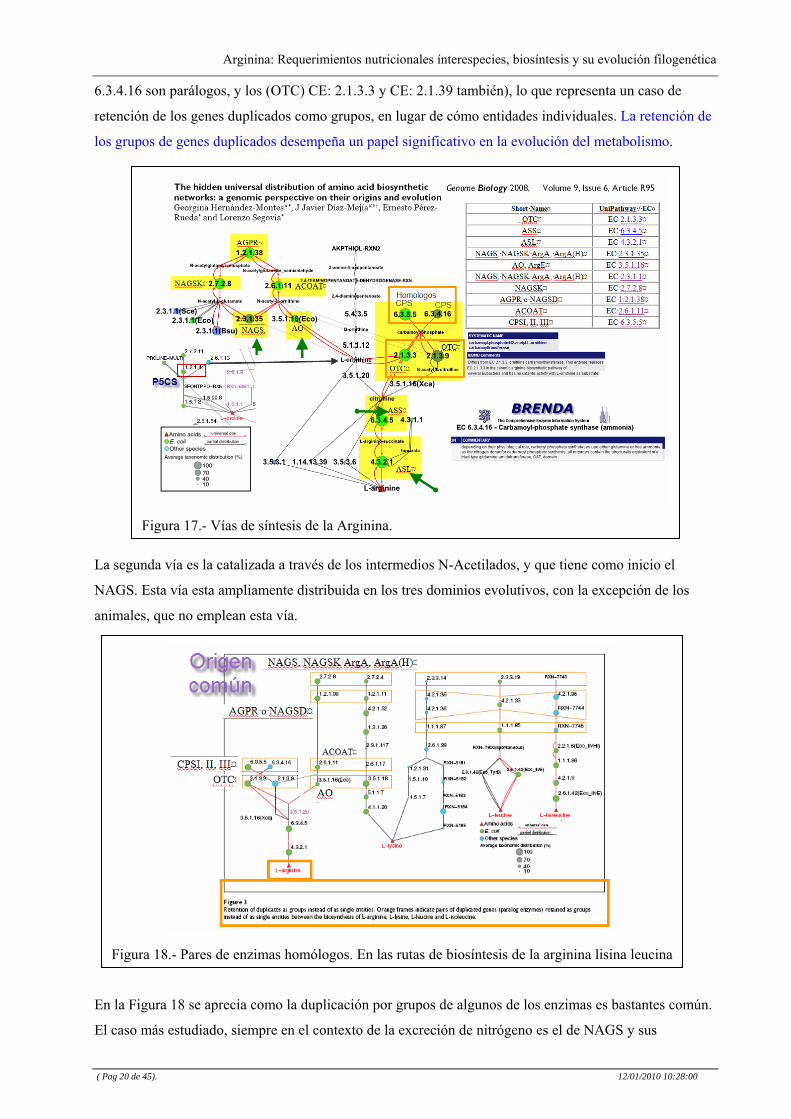
A nivel de los microorganismos, hay al menos cuatro vías de la síntesis de L-arginina, todos ellos
relacionados con la interconversión de L-ornitina y citrulina, aunque pueden ser agrupadas en dos grandes
- vías (Figura 17). La primera puede proceder a través de dos ramas alternó logas.
La primera rama, es la vía clásica de E. coli, catalizada por dos enzimas ampliamente distribuidas, la
carbamilfosfato sintetasa (CPS EC: 6.3.5.5) y ornitin carbamoil transferasa (OCT EC: 2.1.3.3).
La segunda utiliza tres enzimas (EC: 6.3.4.16, EC: 2.1.3.9 y la EC: 3.5.1.16), de los cuales dos son
también ampliamente distribuidos (Figura 2). Curiosamente, los enzimas EC: 6.3.5.5 (CPS) y EC:
( Pag 19 de 45). 12/01/2010 10:28:00
Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
6.3.4.16 son parálogos, y los (OTC) CE: 2.1.3.3 y CE: 2.1.39 también), lo que representa un caso de
retención de los genes duplicados como grupos, en lugar de cómo entidades individuales. La retención de
los grupos de genes duplicados desempeña un papel significativo en la evolución del metabolismo.
La segunda vía es la catalizada a través de los intermedios N-Acetilados, y que tiene como inicio el
NAGS. Esta vía esta ampliamente distribuida en los tres dominios evolutivos, con la excepción de los
animales, que no emplean esta vía.
Figura 17.- Vías de síntesis de la Arginina.
Figura 18.- Pares de enzimas homólogos. En las rutas de biosíntesis de la arginina lisina leucina
En la Figura 18 se aprecia como la duplicación por grupos de algunos de los enzimas es bastantes común.
El caso más estudiado, siempre en el contexto de la excreción de nitrógeno es el de NAGS y sus
( Pag 20 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
( Pag 21 de 45). 12/01/2010 10:28:00
variantes, implicado en vías cruciales desde el punto de vista de evolutivo(Caldovic and Tuchman, 2003;
Haskins et al., 2008; Pekkala et al., 2009; Qu et al., 2007).
4.3.1 La interconexión de las vías de síntesis de lisina, arginina y leucina.
Fondi et al. (Fondi et al., 2007) han realizado un estudio más detallado de la interconexión de la vías de
síntesis de lisina, arginina y leucina. En este estudio, comparan los genes implicados en organismos
inferiores (Bacteria y Archaea) en busca de la ruta común de biosíntesis en el LUCA. Como se aprecia en
la figura, quedan fuera del estudio NAGS (gen ArgA) y P5CS. Este último interconectaría las rutas de la
arginina y la prolina.
Figura 19.- Interconexión entre las vías biosintéticas de lisina, arginina y leucina.
Actualmente, se dispone de gran cantidad de información acerca de los enzimas implicados en la
biosíntesis de Arginina (y de prácticamente cualquier proceso metabólico). La información es fácilmente
accesible a través de los servidores de bases de datos metabólicas como por ejemplo MetaCyt o
UniPathway. Cada reacción metabólica y su enzima ( o posibles enzimas) correspondiente esta indexada
mediante su identificador EC, de esta forma la secuencia de las reacciones enzimáticas del proceso de
biosíntesis de arginina en mamíferos queda resumido en la Tabla 1. En esta sección trataremos algunos de
los aspectos evolutivos más importantes de cada uno de los enzimas.
4.4 Evolución de los enzimas implicados en la biosíntesis de Arginina en mamíferos.

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
( Pag 22 de 45). 12/01/2010 10:28:00
Reacciones de Biosíntesis de Arginina en mamíferos
Nombre Reacción Gen EC
NAGS: N-acetylglutamate
synthase Acetyl-CoA + L-Glutamate <=> CoA + N-Acetyl-L-glutamate (NAG) NAGS 2.3.1.1
CPS I: carbamoyl-phosphate
synthetase 1 2 ATP + NH(3) + CO(2) + H(2)O <=> 2 ADP + phosphate + carbamoyl phosphate CPS1 6.3.4.16
P5CS: pyrroline-5-carboxylate
synthetase
ATP + L-glutamate <=> ADP + L-glutamate 5-phosphate (Ver nota)
L-glutamyl 5-phosphate + NADPH <=> L-glutamate 5-semialdehyde + phosphate + NADP(+) ALDH18A1
1.2.1.41
2.7.2.11
OAT: ornithine
aminotransferase L-glutamate + L-glutamate γ-semialdehyde <=> L-ornithine + 2-oxoglutarate OAT 2.6.1.13
OTC: ornithine
carbamoyltransferase L-ornithine + carbamoyl-phosphate <=> L-citrulline + phosphate + H+ OTC 2.1.3.3
ASS: Argininosuccinate lyase L-aspartate + L-citrulline + ATP <=> L-arginino-succinate + diphosphate + AMP + 2 H+ ASS1 6.3.4.5
ASL: argininosuccinate lyase L-arginino-succinate <=> L-arginine + fumarate ASL 4.3.2.1
Tabla 1.- Reacciones de Biosíntesis de Arginina. Nota.- Fuente (http://biocyc.org/META/NEW-IMAGE?type=ENZYME-IN-RXN-DISPLAY&object=MONOMER-11424 )
Formation of (S)-1-pyrroline-5-carboxylate (or L-glutamate γ-semialdehyde , which is in nonenzymatic equilibrium with it) from glutamate in humans is catalyzed by a single bifunctional enzyme with both γ-glutamyl kinase (EC 2.7.2.11) and γ-glutamyl phosphate reductase (EC 1.2.1.41) activities [ Aral96 , Hu99 ], as is the case in plants [ Hu92 ]. This is in contrast to prokaryotes and lower eukaryotes such as Saccharomyces cerevisiae S288c , in which these two reactions are catalyzed by two different enzymes [ Smith80 ]. This bifunctional enzyme exists in two forms, with the longer form (HsP5CS.long) containing two more amino acids than the short form (HsP5CS.short), a result of alternative splicing [ Hu99 ]. The combined enzymatic activity of this enzyme has been named (S)-1-pyrroline-5-carboxylate synthase, or P5CS for short. The two forms are found in many tissues, in a ratio of about 1:4 (short to long). The enzyme shows the highest expression in pancreas, ovary, testis, and kidney, followed by colon, small intestine, placenta, heart and skeletal muscle. Both forms were able to complement mutations in yeast in both functions. The short form was sensitive to inhibition by ornithine, while the long form was not [ Hu99 ].
http://www.ncbi.nlm.nih.gov/entrez/utils/fref.fcgi?http://www.expasy.org/cgi-bin/nicezyme.pl?2.3.1.1
http://www.ncbi.nlm.nih.gov/entrez/utils/fref.fcgi?http://www.expasy.org/cgi-bin/nicezyme.pl?2.1.3.3
http://www.ncbi.nlm.nih.gov/entrez/utils/fref.fcgi?http://www.expasy.org/cgi-bin/nicezyme.pl?4.3.2.1
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&list_uids=8761662&dopt=Abstract
http://www.ncbi.nlm.nih.gov/entrez/query.fcgi?cmd=Retrieve&db=PubMed&list_uids=1384052&dopt=Abstract

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
4.4.1 Evolución de NAGS: N-acetylglutamate synthase y OAT: ornithine aminotransferase
Diversos artículos han tratado la evolución del N-acetilglutamato sintasa (NAGS) y ornithine
aminotransferase (OAT) (Haskins et al., 2008; Pekkala et al., 2009; Qu et al., 2007; Sancho-Vaello et al.,
2008; Shi et al., 2006). Como hemos visto, la biosíntesis de arginina en a través del N-acetilglutamato
(NAG) es una característica común de muchos procariotas, hongos y plantas, pero no de los animales.
Tanto NAGS como OAT tienen capacidad para sintetizar los intermedios N-acetilados por ello los
trataremos conjuntamente.
4.4.1.1 Evolución de NAGS en los microorganismos.
En los microorganismos (Bacteria – E. Coli) NAGS está codificada por un único gen (ArgA), y la
proteína presenta dos dominios catalíticos: (i) el carbamate kinase (CK) que también está presente en la
enzima NAGK (ArgB) y que cataliza la siguiente reacción de la biosíntesis, y el (ii) dominio de unión con
Acetil-CoA común de la familia de las N-acetiltransferasas (NAT). Esta estructura génica sugiere que el
dominio NAT es el sito catalítico de unión para el glutamato, como ocurre en el resto de los enzimas de la
familia. Algunos bacterias y archea disponen de un enzima alternativo, S-NAGS, que es una versión
recortada de NAGS, presentando únicamente el dominio NAT.
La opción alternativa para sintetizar N-acetil-glutamato es mediante ornithine N-acetyltrasnferase, (OAT)
codificada en microorganismos por el gen ArgJ. Sin embargo no existe ninguna similitud ni entre el
mecanismo , ni similitud de secuencias entre NAGS y OAT. Algunos OAT son bifuncionales, es decir
pueden sintetizar ademas acetil-glutamate.
En algunos organismos (Alternomonas-Vibrio) se ha detectado fusiones del gen ArgA y del ArgJ,
resultado en un gen multifuncional denominado Arg(A). (Figura 21)
Figura 20.- Opciones para sintetizar NAG en microorganismos.
( Pag 23 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
4.4.1.2 NAGS un marcador molecular de la evolución de los tetrápodos.
El ciclo de la urea en los mamíferos, anfibios, tortugas, caracoles, gusanos y muchos animales acuáticos,
requiere N-acetilglutamato (NAG), un activador alostérico de carbamylfosfato sintetasa I (CPSI) en los
mamíferos y anfibios, y carbamylfosfato sintetasa III (CPSIII) en peces e invertebrados. De esta forma los
NAG-CPSI y CPSIII catalizan la formación de carbamylfosfato el primer paso limitante de la
ureagénesis.
El NAG se produce en una reacción catalizada por
(NAGS). La arginina es un inhibidor alostérico de la
NAGS microbiana y vegetal, y activador alostérico de
NAGS mamíferos (Figura 23). La Figura 22 muestra la
diferente actividad de la arginina en varios organismos.
Se observa que la actividad es 500 veces menos en
anfibios que en mamíferos, en los peces es la mitad y en
bacterias existe mucha diversidad.
Haskins et al. ((Haskins et al., 2008)) combinando
información estructural recuperada de la NAGK de
Thermotoga maritima enlazada con arginina, y de
estudios de mutagénesis, han identificado los residuos del
sitio de unión de la arginina en NAGS. Los residuos fueron mutados en el gen NAGS de ratón y en el
NAGS-K de Xantomonas c. y el resultado fue la pérdida del la capacidad activadora de la arginina sobre
Figura 21.- Genes y enzimas de la síntesis de N-acetil-glutamato.
Figura 22.- Actividad de la arginina como activador de NAGS.
( Pag 24 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
NAGS y NAGS-K. También comprobaron que la respuesta de clones de NAGS son inhibidos por la
arginina en el caso de peces y parcialmente inhibidos en ranas.
Figura 23.- Inversión del efecto alostérico de NAGS en tetrápodos.
El efecto alostérico está regulado por los cambios conformacionales resultantes de la unión de la arginina
en el sitio activo. Como demuestran, el cambio alostérico fue es gradual, desde la completa inhibición en
bacterias, pasando por la parcial inhibición en peces a la activación en ranas. El cambio funcional en la
proteína, coincide con el evolutivo de peces a tetrápodos.
4.4.2 Evolución de CPS I: carbamoyl-phosphate synthetase 1
A diferencia de NAGS, la evolución de CPS I no ha sido muy estudiada en los últimos años. Los estudios
existentes (Hong et al., 1994; Lawson et al., 1996; Lindley et al., 2007; Rubio and Cervera, 1995; Simmer
et al., 1990) muestran un patrón evolutivo complejo, principalmente debido a la fusión y duplicación del
gen. Sin embargo el centro activo del enzima muestra una alta conservación del (Figura 24). Las CPS se
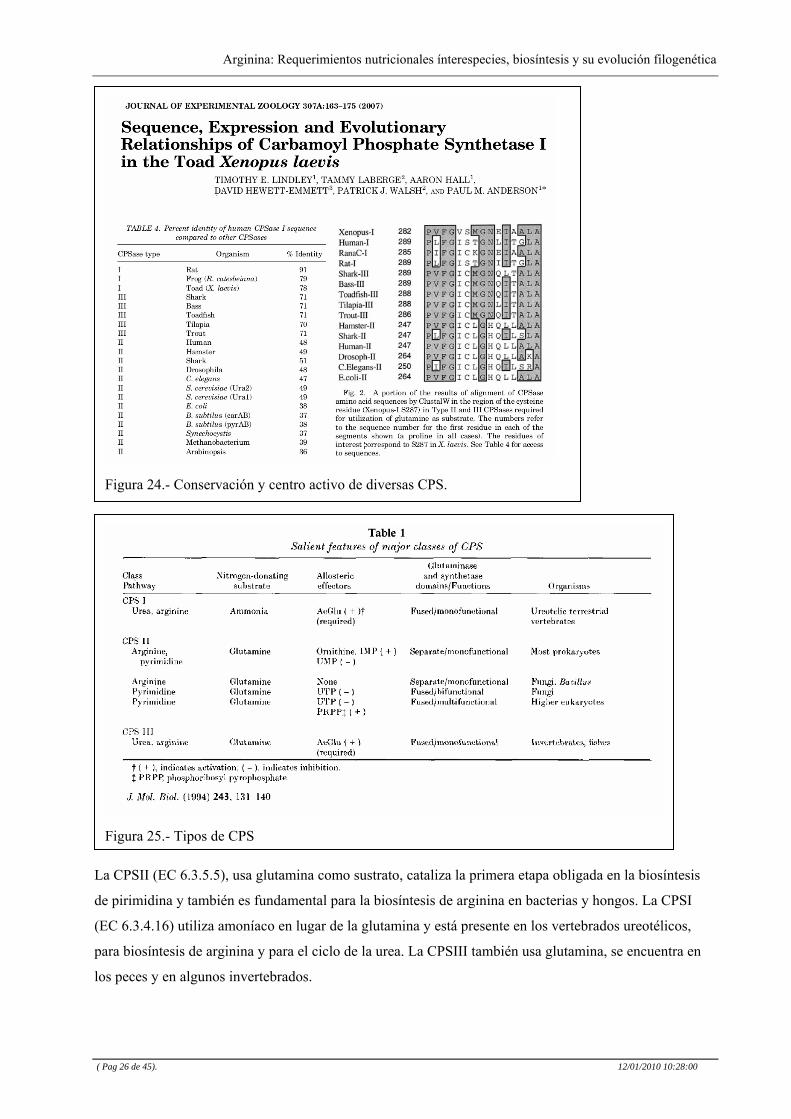
clasifican en tres grupos I, II y III, (Ver ) dependiendo del sustrato donador de nitrógeno. Figura 25
( Pag 25 de 45). 12/01/2010 10:28:00
Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Figura 25.- Tipos de CPS
Figura 24.- Conservación y centro activo de diversas CPS.
La CPSII (EC 6.3.5.5), usa glutamina como sustrato, cataliza la primera etapa obligada en la biosíntesis
de pirimidina y también es fundamental para la biosíntesis de arginina en bacterias y hongos. La CPSI
(EC 6.3.4.16) utiliza amoníaco en lugar de la glutamina y está presente en los vertebrados ureotélicos,
para biosíntesis de arginina y para el ciclo de la urea. La CPSIII también usa glutamina, se encuentra en
los peces y en algunos invertebrados.
( Pag 26 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
( Pag 27 de 45). 12/01/2010 10:28:00
4.4.3 Evolución de P5CS: pyrroline-5-carboxylate synthetase
El P5CS es un enzima clave en la evolución de biosíntesis de arginina en los mamíferos (¡Error! No se
encuentra el origen de la referencia.), ya que permite el aporte extra de ornitina para la biosíntesis de
Arginina. El gen responsable de la codificación del enzima es el ALDH18A1.
El gen Forma parte de la super-familia de aldehide dehidrogenase (ALDH) , ampliamente distribuida en
todos los organismos .
Los enzimas producidos a partir de los genes de la familia ALDH están involucrados en la catabolismos
de diferentes moléculas esenciales en las células, principalmente ciertas grasas (colesterol y otros ácidos
grasos) y aminoácidos como la prolina y arginina. Una función adicional de estos enzimas es desintoxicar
el organismo de sustancias externas, como el alcohol y los xenobióticos y sustancias endogenes, como las
toxinas que se forman en las células. El papel específico de las enzimas ALDH es modificar los grupos
aldehído de estas moléculas. Hasta el momento se han identificado 20 genes ALDH en humanos.
Figura 26 - Familias 18 y 19 de ALDH
Figura 27. Superfamila ALDH

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
El nombre de cada gen de la aldehído deshidrogenasa comienza con ALDH, lo que indica que es parte de
la familia de genes aldehído deshidrogenasa. El gen también se da un número asociado a un grupo
específico dentro de la familia del gen, una letra que representa el gen de la subfamilia, y un número
asignado a la gen específico dentro de la subfamilia. Por ejemplo, el primer gen de la aldehído
deshidrogenasa en el grupo 7, subfamilia A, se escribe como ALDH7A1.
El gen ALDH18A1 codifica para una enzima bifuncional, con actividad de gamma-glutamil-cinasa y de
fosfato reductasa. La enzima es tradicionalmente conocida como Δ1-pirrolina-5-carboxilato sintetasa
(P5CS), que cataliza la reducción de la L-glutamato a Δ1-pirrolina-5-carboxilato de metilo, un paso
crítico en la biosíntesis endógena de la prolina, ornitina y arginina.
El splicing alternativo de ALDH18A1 genera dos isoformas, una corta (P5CS.Short) y una isoforma larga
(P5CS.long) que difieren en la inserción de dos aminoácidos en el extremo N, justo en sitio activa
quinasa. La isoforma P5CS.Short tiene una alta actividad en el intestino y cataliza el paso esencial en la
biosíntesis de la arginina. Se inhibe en ornitina proporcionando así un mecanismo para la regulación de la
síntesis de arginina. La isoforma larga se expresa en múltiples tejidos y es necesaria para la síntesis de la
prolina de glutamato. Es insensible a la inhibición de la ornitina.
Figura 28.- Splicing alternativo de P5CS
La ruta de biosíntesis de la prolina a través de glutamato implica dos pasos enzimáticos catalizados por la
gamma-glutamil-cinasa (GK, EC 2.7.2.11) y Gamma-glutamil-fosfato reductasa (GPR) GPR (EC
1.2.1.41). Estas dos enzimas se funden en la enzima P5CS en las plantas y animales, mientras que son las
enzimas separadas en las bacterias y levaduras. En los seres humanos, la P5CS . El tomate por ejemplo,
tiene tanto el gen procariota como el P5CS bifuncional (ALDH18B1).
( Pag 28 de 45). 12/01/2010 10:28:00
Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
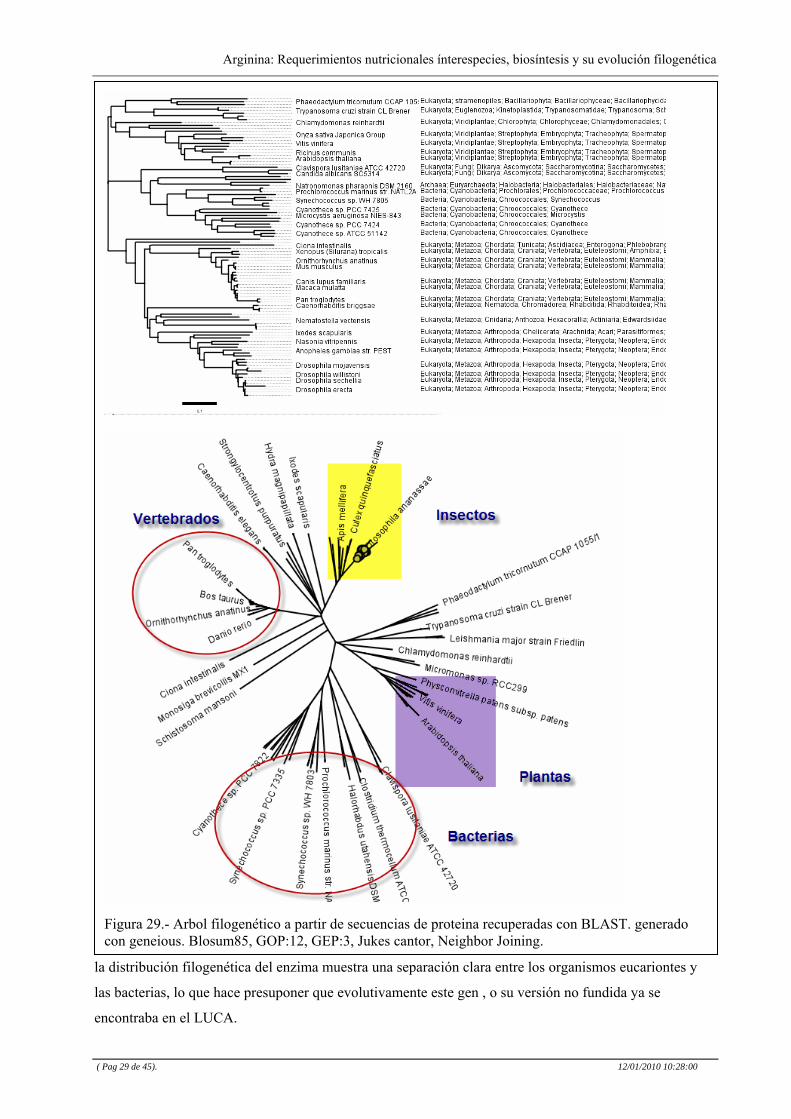
Figura 29.- Arbol filogenético a partir de secuencias de proteina recuperadas con BLAST. generado con geneious. Blosum85, GOP:12, GEP:3, Jukes cantor, Neighbor Joining.
la distribución filogenética del enzima muestra una separación clara entre los organismos eucariontes y
las bacterias, lo que hace presuponer que evolutivamente este gen , o su versión no fundida ya se
encontraba en el LUCA.
( Pag 29 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
En los mamíferos las enzimas ALDH se encuentran principalmente en el hígado y las células del riñón,
pero también se pueden encontar en las células de todo el cuerpo. Variaciones comunes (polimorfismos)
en los genes ALDH puede afectar a la función de las enzimas. Las enfermedades causadas por mutaciones
en los genes ALDH típicamente involucran la acumulación de sustancias en el cuerpo que son
perjudiciales en grandes cantidades o que alteran la función o la producción de otras moléculas
necesarias. Por ejemplo, se ha identificado que una mutación en uno de los aminoácidos del dominio C-
terminal es el causante del “síndrome neurocutaneo”.
Figura 30.- Alineamiento de secuencias de P5CS de diferentes organismos.
( Pag 30 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
4.4.4 Evolución de OTC: ornithine carbamoyltransferase
La OTC es una enzima altamente conservada como muestran los alineamientos de secuencias demuestran
una alta conservación en todos los organismos, especialmente en los residuos implicados en la unión
carbamil fosfato y ornitina (Labedan et al., 1999; Labedan et al., 2004). Sin embargo el gen codificarte
esta sujeto a múltiples polimorfismos y mutaciones, que han abierto la puerta a nuevas investigaciones
relacionadas con su implicación con la enfermedad de alzeimer((Bensemain et al., 2009)).
Las enzimas OTCase y ATCases, que se encuentran en muchas bacterias (gen ArgF) y eucariotas (gen
OTC en mamiferos), así como en arqueas y catalizan reacciones análogas. La ATCasa (el gen arcB)
cataliza la reacción entre la carbamilfosfato y aspartato que inicia la ruta para la biosíntesis de pirimidina,
y la OTCase que cataliza la transferencia de la fracción carbamoil de carbamilfosfato a la 5-amino de la
ornitina para formar citrulina. La similitud estructural real de los dos carbamoyltransferases fue
recientemente confirmada por el análisis tridimensional de la OTCase de Pseudomonas aeruginosa (el gen
arcB) y la comparación de su estructura globular a la de la ATCasa de E. coli sugiere que las
carbamoyltransferasas surgieron por la duplicación de un gen ancestral.
Figura 31.- Relación de OTC con la enfermedad de alzeimer.
( Pag 31 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
4.4.5 Evolución de ASS: Argininosuccinate lyase
El enzima ASS esta codificado por el gen ASS1. Este gen presenta características un tanto especiales,
como observamos en su entrada de Entrez. En el genoma humano existen entre 10 y 14 pseudo genes
ASS. El gen activo esta localizado en el cromosoma 9 y hay identificado dos transcritos que codifican la
misma proteína.
Estas
caracteristicas
especiales le
confieren un
especial valor
evolutivo.
Otra característica especial es que esta altamente conservado en eucariotas y en especial en mamíferos,
por lo que se convierte en el candidato ideal para estudiar las diferentes actividades del enzima de los
mamíferos.
Figura 33.- Conservación del Gen ASS1
Figura 32.- Entrada en Entrez del gen ASS1.
( Pag 32 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Un ejemplo de cómo han evolucionado de diferente forma dos de los genes implicados en la biosíntesis
de la arginina, lo encontramos en el articulo de Brown (Brown and Doolittle, 1997). En el caso del gen
ASS en eucariontes esta más cerca evolutivamente de Arquea que de las bacterias, mientras que en el caso
de P5CR (implicado en la síntesis de prolina) ocurre lo contrario.
Figura 34.- Diferente evolución de los genes ASS y P5CR.
Dado que este estudio es relativamente antiguo, podemos reconstruir el árbol filogenético incluyendo las
todas las secuencias existentes actualmente (Figura 36).
Figura 35.- Comparación del árbol filogenético del artículo de Brown con el generado con datos actuales.
( Pag 33 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Efectivamente, si únicamente dejamos del árbol de la Figura 36 las secuencias de los organismos de
brown obtenemos la misma topología (Figura 35).
Figura 36.- Arbol filogenético de ASS reconstruido con las secuencias actuales
Si comprobamos el resultado de nuestro alineamiento con clustalW con el Logo obtenido de Expansy
podemos identificar fácilmente el centro catalítico del enzima. Las implicaciones biomédicas de este
hecho son inmediatas: Si se puede conocer la estructura y el sitio catalítico, podemos inferir el mecanismo
de reacción, y si juntamos todos estos factores, es posible evaluar la diferente actividad de la enzima en
diferentes especies.
Efectivamente, si consultamos en BRENDA, rápidamente encontramos la referencia al mecanismo de la
reacción enzimática (Lemke and Howell, 2002; Lemke and Howell, 2001).
( Pag 34 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Figura 37.- Alineamientos de ASS y comprobación del LOGO de ExPASy
( Pag 35 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
( Pag 36 de 45). 12/01/2010 10:28:00
Figura 38.- Mecanismo de reacción de ASS.

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
4.4.6 Evolución de ASL: argininosuccinate lyase
El ASL esta codificado por un gen con el mismo nombre (ASL). La evolución filogenética de ASL es
similar a la de ASS, de echo se tratan conjuntamente en un reciente articulo(Wagemaker et al., 2007).
ASL pertenece a la superfamilia de enzimas homotetramericos que incluye a fumarasas, aspartasas, ASL,
y D-cristalin entre otras, que catalizan reacciones homólogas. A pesar de la similitud de secuencia es baja,
se han identificado tres dominios activos perfectamente definidos.
( Pag 37 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Figura 39.- Identificación de los sitios activos de ASL
( Pag 38 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
( Pag 39 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Reference List
Baker,D.H. (2005) Comparative nutrition and metabolism: explication of open questions with emphasis on protein and amino acids. Proc. Natl. Acad. Sci. U. S. A, 102, 17897-17902.
Ball,R.O., Urschel,K.L. and Pencharz,P.B. (2007) Nutritional consequences of interspecies differences in arginine and lysine metabolism. J. Nutr., 137, 1626S-1641S.
Bensemain,F. et al. (2009) Evidence for induction of the ornithine transcarbamylase expression in Alzheimer's disease. Mol. Psychiatry, 14, 106-116.
Bono,H. et al. (1998) Reconstruction of amino acid biosynthesis pathways from the complete genome sequence. Genome Res., 8, 203-210.
Brown,J.R. and Doolittle,W.F. (1997) Archaea and the prokaryote-to-eukaryote transition. Microbiol. Mol. Biol. Rev., 61, 456-502.
Caetano-Anolles,G. et al. (2009) The origin and evolution of modern metabolism. Int. J. Biochem. Cell Biol., 41, 285-297.
Caldovic,L. and Tuchman,M. (2003) N-acetylglutamate and its changing role through evolution. Biochem. J., 372, 279-290.
Cunchillos,C. and Lecointre,G. (2003) Evolution of amino acid metabolism inferred through cladistic analysis. J. Biol. Chem., 278, 47960-47970.
Cunchillos,C. and Lecointre,G. (2005) Integrating the universal metabolism into a phylogenetic analysis. Mol. Biol. Evol., 22, 1-11.
Cunchillos,C. and Lecointre,G. (2007) Ordering events of biochemical evolution. Biochimie., 89, 555-573.
Fondi,M. et al. (2007) The primordial metabolism: an ancestral interconnection between leucine, arginine, and lysine biosynthesis. BMC. Evol. Biol., 7 Suppl 2:S3., S3.
Haskins,N. et al. (2008) Inversion of allosteric effect of arginine on N-acetylglutamate synthase, a molecular marker for evolution of tetrapods. BMC. Biochem., 9:24., 24.
Hernandez-Montes,G. et al. (2008) The hidden universal distribution of amino acid biosynthetic networks: a genomic perspective on their origins and evolution. Genome Biol., 9, R95.
Hong,J. et al. (1994) Carbamyl phosphate synthetase III, an evolutionary intermediate in the transition between glutamine-dependent and ammonia-dependent carbamyl phosphate synthetases. J. Mol. Biol., 243, 131-140.
( Pag 40 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Labedan,B. et al. (1999) The evolutionary history of carbamoyltransferases: A complex set of paralogous genes was already present in the last universal common ancestor. J. Mol. Evol., 49, 461-473.
Labedan,B. et al. (2004) Using quaternary structures to assess the evolutionary history of proteins: the case of the aspartate carbamoyltransferase. Mol. Biol. Evol., 21, 364-373.
Lawson,F.S., Charlebois,R.L. and Dillon,J.A. (1996) Phylogenetic analysis of carbamoylphosphate synthetase genes: complex evolutionary history includes an internal duplication within a gene which can root the tree of life. Mol. Biol. Evol., 13, 970-977.
Lemke,C.T. and Howell,P.L. (2001) The 1.6 A crystal structure of E. coli argininosuccinate synthetase suggests a conformational change during catalysis. Structure., 9, 1153-1164.
Lemke,C.T. and Howell,P.L. (2002) Substrate induced conformational changes in argininosuccinate synthetase. J. Biol. Chem., 277, 13074-13081.
Lindley,T.E. et al. (2007) Sequence, expression and evolutionary relationships of carbamoyl phosphate synthetase I in the toad Xenopus laevis. J. Exp. Zool. A Ecol. Genet. Physiol., 307, 163-175.
Mommsen,T.P. and Walsh,P.J. (1989) Evolution of urea synthesis in vertebrates: the piscine connection. Science., 243, 72-75.
Morris,J.G. (1985) Nutritional and metabolic responses to arginine deficiency in carnivores. J. Nutr., 115, 524-531.
Morris,J.G. (2002) Idiosyncratic nutrient requirements of cats appear to be diet-induced evolutionary adaptations. Nutr. Res. Rev., 15, 153-168.
Payne,S.H. and Loomis,W.F. (2006) Retention and loss of amino acid biosynthetic pathways based on analysis of whole-genome sequences. Eukaryot. Cell, 5, 272-276.
Pekkala,S. et al. (2009) Structural insight on the control of urea synthesis: identification of the binding site for N-acetyl-L-glutamate, the essential allosteric activator of mitochondrial carbamoyl phosphate synthetase. Biochem. J., 424, 211-220.
Peregrin-Alvarez,J.M., Sanford,C. and Parkinson,J. (2009) The conservation and evolutionary modularity of metabolism. Genome Biol., 10, R63.
Perlstein,E.O. et al. (2007) Evolutionarily conserved optimization of amino acid biosynthesis. J. Mol. Evol., 65, 186-196.
Qu,Q. et al. (2007) A novel bifunctional N-acetylglutamate synthase-kinase from Xanthomonas campestris that is closely related to mammalian N-acetylglutamate synthase. BMC. Biochem., 8:4., 4.
Rubio,V. and Cervera,J. (1995) The carbamoyl-phosphate synthase family and carbamate kinase: structure-function studies. Biochem. Soc. Trans., 23, 879-883.
( Pag 41 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Sancho-Vaello,E., Fernandez-Murga,M.L. and Rubio,V. (2008) Site-directed mutagenesis studies of acetylglutamate synthase delineate the site for the arginine inhibitor. FEBS Lett., 582, 1081-1086.
Shao,A. and Hathcock,J.N. (2008) Risk assessment for the amino acids taurine, L-glutamine and L-arginine. Regul. Toxicol. Pharmacol., 50, 376-399.
Shi,D. et al. (2006) Structure and catalytic mechanism of a novel N-succinyl-L-ornithine transcarbamylase in arginine biosynthesis of Bacteroides fragilis. J. Biol. Chem., 281, 20623-20631.
Simmer,J.P. et al. (1990) Mammalian carbamyl phosphate synthetase (CPS). DNA sequence and evolution of the CPS domain of the Syrian hamster multifunctional protein CAD. J. Biol. Chem., 265, 10395-10402.
Takiguchi,M. et al. (1989) Evolutionary aspects of urea cycle enzyme genes. Bioessays., 10, 163-166.
Wagemaker,M.J. et al. (2007) Argininosuccinate synthetase and argininosuccinate lyase: two ornithine cycle enzymes from Agaricus bisporus. Mycol. Res., 111, 493-502.
Wright,P.A. (1995) Nitrogen excretion: three end products, many physiological roles. J. Exp. Biol., 198, 273-281.
Wu,G. et al. (2009) Arginine metabolism and nutrition in growth, health and disease. Amino. Acids., 37, 153-168.
Xu,Y., Labedan,B. and Glansdorff,N. (2007) Surprising arginine biosynthesis: a reappraisal of the enzymology and evolution of the pathway in microorganisms. Microbiol. Mol. Biol. Rev., 71, 36-47.
Xu,Y. et al. (2000) Evolution of arginine biosynthesis in the bacterial domain: novel gene-enzyme relationships from psychrophilic Moritella strains (Vibrionaceae) and evolutionary significance of N-alpha-acetyl ornithinase. J. Bacteriol., 182, 1609-1615.
( Pag 42 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Figuras adicionales.
( Pag 43 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
Figura 40.- Distribución de la P5CS en mamíferos. http://www.ensembl.org/Homo_sapiens/Gene/Compara_Tree?db=core;g=ENSG00000059573;r=10:97365696-97416463;t=ENST00000371224
( Pag 44 de 45). 12/01/2010 10:28:00

Arginina: Requerimientos nutricionales ínterespecies, biosíntesis y su evolución filogenética
( Pag 45 de 45). 12/01/2010 10:28:00


![AMINO ACIDOS [Modo de compatibilidad]medicinaunivalle14.weebly.com/uploads/2/6/7/2/26724831/aminoacidos... · BIOSINTESIS DE LOS AMINO ACIDOS. ... Histidine Arginine Asparagine Citrate](https://static.fdocumento.com/doc/165x107/5b93fc4309d3f22b0a8c6c34/amino-acidos-modo-de-compatibilidad-biosintesis-de-los-amino-acidos-histidine.jpg)