
3 proteínas iii
36
Proteínas III: Estructura y función Dra. Ana I. Sotelo 21 de agosto de 2013
Transcript of 3 proteínas iii

Proteínas III: Estructura y función
Dra. Ana I. Sotelo 21 de agosto de 2013

Proteínas conjugadas
el grupo prostético de una proteína se vincula con su función

interacción ligando-proteína interacción reversible
Ligando = molécula que se une a reversiblemente a otra
Sitio de fijación = sitio de la proteína donde se une el
ligando
es complementario al ligando en tamaño, forma y carácter
la interacción es específica
Las proteínas son flexibles =
pueden cambiar de
conformación encaje
inducido = cuando se
adaptan al ligando

interacción ligando-proteína interacción reversible
Expresión de equilibrio P + L PL
Ka = [PL] = ka
[P] [L] kd
Constante de asociación
[PL] = Ka [L] [P] θ = sitios de fijación ocupados = [PL]
sitios de fijación totales [PL] + [P]
θ = Ka [L] [P] = [L]
Ka [L] [P] + [P] [L] + 1/ Ka 1/ Ka

interacción ligando-proteína constantes de disociación (Kd)

cuando el ligando es un gas
cuando el oxígeno es el ligando se mide la presión parcial de
oxígeno (pO2) porque la concentración de un gas en solución es
proporcional a su presión parcial local en la fase gaseosa
θ = pO2
pO2 + P50
θ = [L]
[L] + Kd

proteínas de unión a oxígeno
el oxígeno puede estar unido a un grupo prostético hemo

grupo hemo
grupo hemo: anillo protoporfirínico que
coordina un ion Fe2+
El Fe2+ une oxígeno, el Fe3+ no
el CO y el NO se unen al Fe2+ del hemo
con mayor afinidad que el O2

mioglobina
el grupo hemo es coordinado por la His en posición 93
(en la hélice H)

la estructura proteica afecta la unión del ligando
el CO presenta afinidad 20.000 veces
mayor que el O2 por el hemo libre
pero solo 200x por el hemo en la
mioglobina
la His 67 (His E7 o histidina distal)
coordina al O2 del hemo

hemoglobina
los GR tienen 34% de hemoglobina
la Hb de sangre arterial está saturada con O2 en un 96 %; la venosa en un 64 %

hemoglobina la hemoglobina es un
tetrámero formado por dos
subunidades α (de 141
residuos) y dos
subunidades β (de 146
residuos)
la estructura de cada
subunidad es parecida a la
de la mioglobina, aunque
solo tienen 27 aa en la
misma posición
la interfase α1-β1 involucra 30
residuos, la α1-β2 19

conformación de la hemoglobina
la hemoglobina en estado R presenta mayor afinidad por el O2; el O2 estabiliza el estado R
en ausencia de O2 predomina el estado T o desoxihemoglobina
el estado T (tenso) está estabilizado por mayor número de enlaces iónicos en la interfase α1-β2 que el estado R (relajado)

unión cooperativa de oxígeno
la Hb debe unir O2 en los
pulmones y liberarlo en los
tejidos
la Hb pasa de un estado de
baja afinidad (T) a uno de
alta afinidad (R) cuando une
más moléculas de O2: la
unión de O2 cambia la
afinidad de las subunidades
adyacentes

unión cooperativa de ligando
proteína alostérica: la unión de un ligando afecta las
propiedades de otros sitios de unión

unión cooperativa de ligando ecuación de Hill
Equilibrio P + nL PLn
Ka = [PLn]
[P] [L] n
Cte asociación
θ = [L]n
[L]n + Kd
θ = [L]n
1- θ Kd
Log θ = n log [L] – log Kd
1- θ

carboxihemoglobina
la unión de CO a una o
dos subunidades de Hb
aumenta la afinidad por
el O2 en las demás
subunidades

modelos de mecanismo de unión en la unión cooperativa
modelo concertado modelo secuencial

la hemoglobina también une CO2 y H+
Efecto Bohr
el CO2 es convertido en
HCO3- + H+ por la
anhidrasa carbónica; la
hidratación del CO2 hace
que descienda el pH de la
sangre
a bajo pH disminuye la
afinidad de la Hb por el O2

la hemoglobina también une CO2 y H+
Efecto Bohr el H+ se une a la His HC3
de la cadena b
este residuo, protonado a bajo pH, forma par iónico con el Asp FG1, estabilizando el estado T
también se protona el NH2 terminal
el CO2 se une al NH2 terminal para dar carbaminohemoglobina
la unión de CO2 disminuye la afinidad de la Hb por el O2

2,3-DPG (difosfoglicerato)
se forma a partir del 1,3-DPG (vía glucolítica) y se encuentra en grandes cantidades en el eritrocito funciona como efector alostérico de la Hb (modulador heterotrópico) disminuye la afinidad de la Hb por el O2

unión de 2,3-DPG a la hemoglobina
en las alturas aumenta la
[2,3-DPG]
también aumenta en hipoxia
se ubica entre subunidades β
en el estado T

hemoglobina fetal La Hb fetal posee mayor afinidad
por el O2 que la Hb materna ya que
debe obtenerlo de la sangre de la
madre
posee subunidades γ en vez de β

anemia falciforme
La Hb S es resultado de una única sustitución E6V, que hacen que sea insoluble cuando se desoxigena

interacciones complementarias entre proteínas y ligandos: el sistema inmune
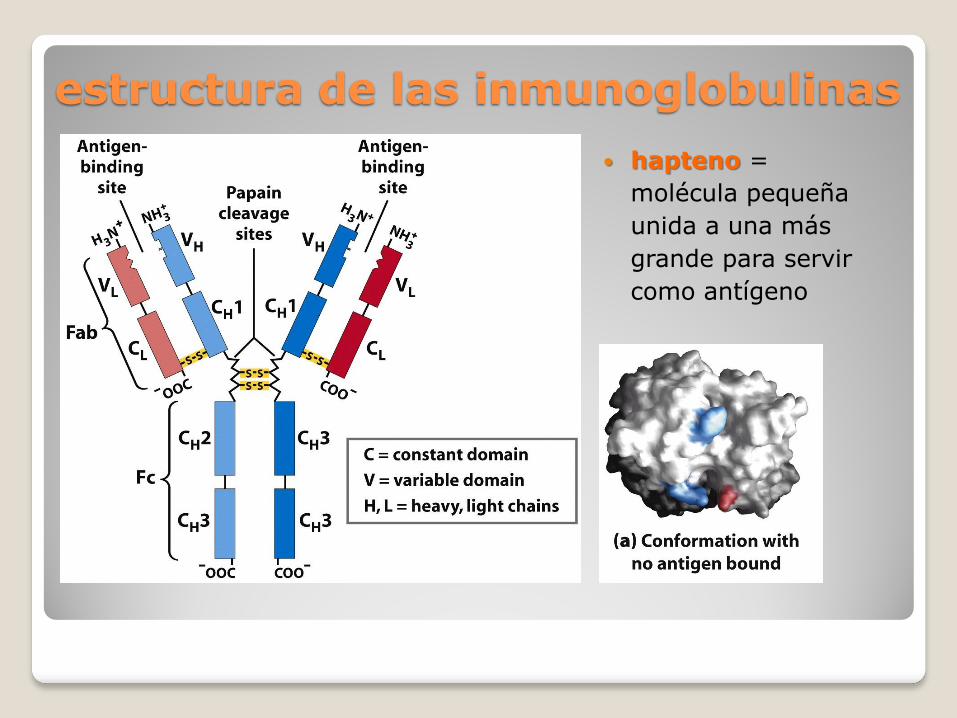
estructura de las inmunoglobulinas
hapteno =
molécula pequeña
unida a una más
grande para servir
como antígeno

interacción antígeno-anticuerpo
epitope es la porción del antígeno que reconoce el anticuerpo
ajuste inducido: para adaptarse al antígeno, el anticuerpo sufre
ligeros cambios conformacionales
especificidad: dada por la complementariedad entre el antígeno y
el sitio de fijación (en el anticuerpo)

fagocitosis
cuando la inmunoglobulina reconoce a un agente extraño (virus, bacterias, etc), lo une y lo dirige a degradación por células, los macrófagos, que los incorporan por fagocitosis

la reacción antígeno-anticuerpo es la base de numerosas técnicas que permiten identificar o cuantificar proteínas específicas en muestras complejas
interacción antígeno-anticuerpo: importante herramienta bioquímica
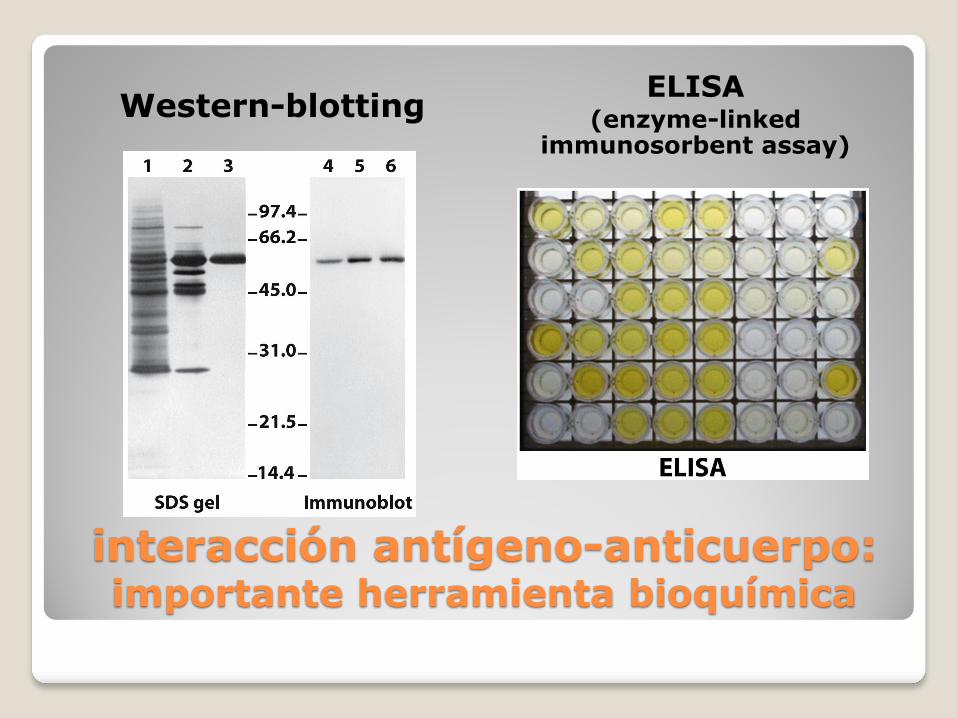
interacción antígeno-anticuerpo: importante herramienta bioquímica
Western-blotting ELISA
(enzyme-linked immunosorbent assay)
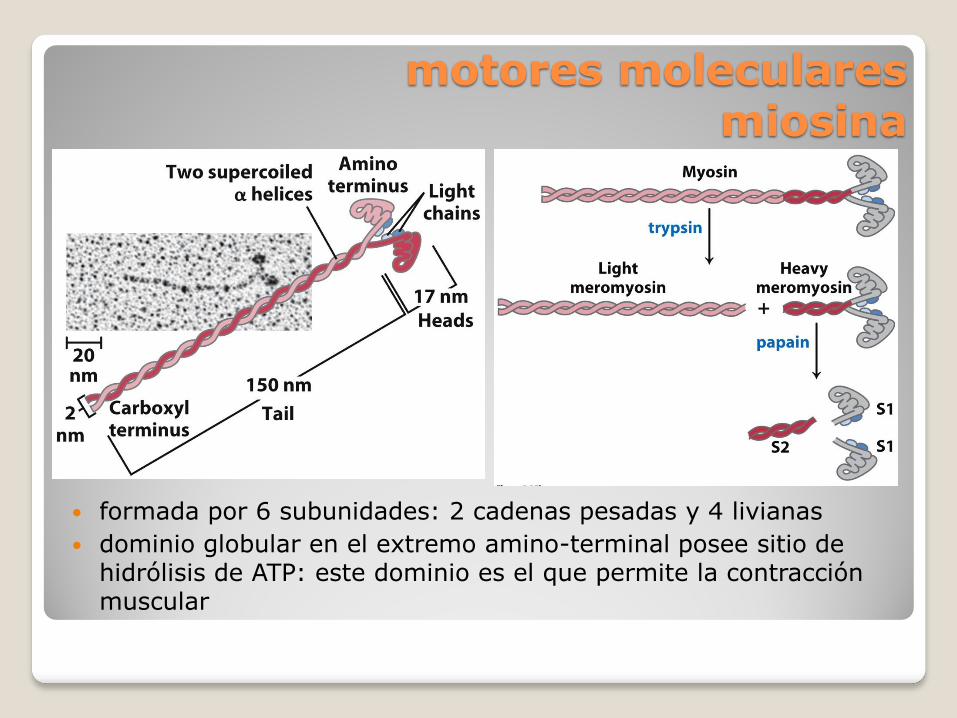
motores moleculares miosina
formada por 6 subunidades: 2 cadenas pesadas y 4 livianas
dominio globular en el extremo amino-terminal posee sitio de hidrólisis de ATP: este dominio es el que permite la contracción muscular

filamentos
la miosina forma los filamentos gruesos
monómeros de actina G polimerizan dando
largos filamentos de actina F
actina F, junto a troponina y tropomiosina
forman los filamentos delgados

fibra muscular
actina y miosina forman el 80 % de la masa muscular

contracción muscular
sarcómero = unidad contráctil: banda A + banda I
en la contracción muscular los discos Z se acercan entre sí
los filamentos de actina se unen al disco Z, en la unión participan α-actinina,
desmina y vimentina
en la línea M se organizan los filamentos gruesos; incluyen paramiosina, proteína C y proteína M; la titina une el filamento grueso a la línea Z

contracción muscular
los filamentos gruesos de miosina se
deslizan sobre los filamentos
delgados de actina
la miosina está unida a la actina
por unión e hidrólisis de ATP la
miosina se libera de la actina, se
mueve y se une a otra molécula de
actina
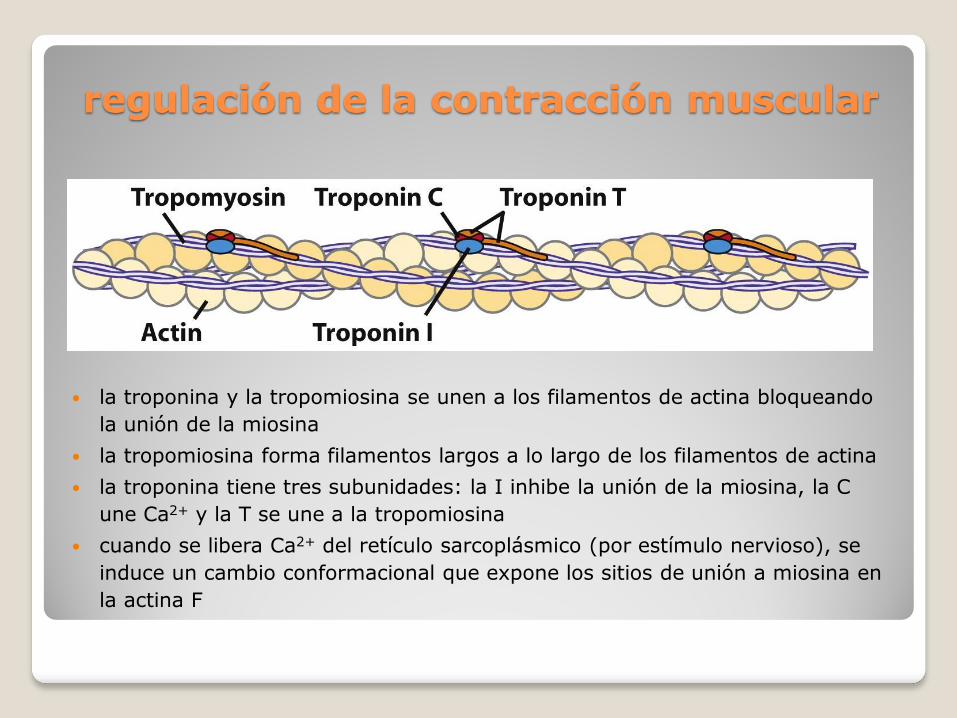
regulación de la contracción muscular
la troponina y la tropomiosina se unen a los filamentos de actina bloqueando
la unión de la miosina
la tropomiosina forma filamentos largos a lo largo de los filamentos de actina
la troponina tiene tres subunidades: la I inhibe la unión de la miosina, la C
une Ca2+ y la T se une a la tropomiosina
cuando se libera Ca2+ del retículo sarcoplásmico (por estímulo nervioso), se
induce un cambio conformacional que expone los sitios de unión a miosina en
la actina F


















