
4.4.2 Multiplicación de CMV en arabidopsis y en...
169
Resultados 150 Figura A4.1 Origen de los individuos listados en la Tabla A4.1 del Anejo 4. 4.4.2 Multiplicación de CMV en arabidopsis y en melón Todos los genotipos de arabidopsis analizados fueron susceptibles a la infección por los 8 aislados de CMV y en ninguna de las interacciones se observó inmunidad o resistencia cualitativa. La multiplicación viral se cuantificó como acumulación de ARN viral (μg ARN viral/g p.f.) a partir de extracto de ARN total procedente de hojas infectadas sistémicamente recolectadas a los 15 d.p.i. Los valores de acumulación viral en arabidopsis se analizaron, en primer lugar, sin tener en cuenta el huésped de origen de los aislados de CMV. Un análisis GLM en el que se consideraron aleatorios los factores aislado de CMV e individuo de arabidopsis mostró diferencias significativas de acumulación viral entre aislados de CMV (F 7,824 =7.435, P<10 -4 ) e individuos de arabidopsis (F 20,824 =7.697, P<10 -4 ). Además, mostró interacción significativa entre los factores aislado de CMV x individuo de arabidopsis (F 140,824 = 10.394, P<10 -4 ) indicando, como en el Apartado 4.2 y el Apartado 4.3, que la acumulación de CMV en arabidopsis dependió de una interacción genotipo del huésped x genotipo del patógeno. Dicha interacción explicó un 40% de la varianza del caracter (Var(aislado de CMV x individuo de arabidopsis)=39.903, Var(individuo de arabidopsis)=33.051 y Var(aislado de CMV)=12.217). (Anejo 4, Tabla A4.2).
Transcript of 4.4.2 Multiplicación de CMV en arabidopsis y en...

Resultados
150
Figura A4.1 Origen de los individuos listados en la Tabla A4.1 del Anejo 4.
4.4.2 Multiplicación de CMV en arabidopsis y en melón
Todos los genotipos de arabidopsis analizados fueron susceptibles a la infección por
los 8 aislados de CMV y en ninguna de las interacciones se observó inmunidad o resistencia
cualitativa. La multiplicación viral se cuantificó como acumulación de ARN viral (µg ARN
viral/g p.f.) a partir de extracto de ARN total procedente de hojas infectadas sistémicamente
recolectadas a los 15 d.p.i.
Los valores de acumulación viral en arabidopsis se analizaron, en primer lugar, sin
tener en cuenta el huésped de origen de los aislados de CMV. Un análisis GLM en el que se
consideraron aleatorios los factores aislado de CMV e individuo de arabidopsis mostró
diferencias significativas de acumulación viral entre aislados de CMV (F7,824=7.435, P<10-4) e
individuos de arabidopsis (F20,824=7.697, P<10-4). Además, mostró interacción significativa
entre los factores aislado de CMV x individuo de arabidopsis (F140,824= 10.394, P<10-4)
indicando, como en el Apartado 4.2 y el Apartado 4.3, que la acumulación de CMV en
arabidopsis dependió de una interacción genotipo del huésped x genotipo del patógeno. Dicha
interacción explicó un 40% de la varianza del caracter (Var(aislado de CMV x individuo de
arabidopsis)=39.903, Var(individuo de arabidopsis)=33.051 y Var(aislado de CMV)=12.217).
(Anejo 4, Tabla A4.2).

Resultados
151
Figura 4.4.2 Variación de los valores medios de acumulación de los aislados At n y Cu n en los individuos de arabidopsis.
Para estudiar si las diferencias de acumulación entre aislados de CMV se asocian a su
huésped de origen, se realizó un GLM en el que se consideró fijo el factor huésped de origen
de los aislados de CMV y aleatorio el factor individuo de arabidopsis. El resultado mostró
diferencias significativas de acumulación viral entre huéspedes de origen de los aislados de
CMV (F1,824=8.351, P=0.009) (At: 7.917±0.370 y Cu: 6.409±0.331), siendo mayor la
acumulación de los aislados At que la de los aislados Cu; y entre individuos de arabidopsis
(F20,824=13.682, P<10-4) (Figura 4.4.2). El número de individuos de arabidopsis en los que los
aislados At se acumularon más que los aislados Cu fue mayor que el número de individuos en
los que los aislados Cu se acumularon más que los aislados At (6/1, χ2=4.571, P=0.032), (X ~
B(6,7,0.5, P=0.054) (Figura 4.4.2 y 4.4.3). También fue significativa la interacción entre los
factores huésped de origen de los aislados de CMV x individuo de arabidopsis (F20,824=1.828,
P=0.017). El factor que explicó la mayor parte de la varianza fue el factor individuo de
arabidopsis (Var(individuo)=35.000, Var(individuo x huésped de origen)=2.496 y
Var(huésped de origen de los aislados de CMV)=1.983) (Anejo 4, Tabla A4.3).
A diferencia de los individuos de arabidopsis ensayados que provienen de diferentes
zonas de la Península Ibérica (Figura 4.4.1); los aislados de CMV, At y Cu, provienen de una
misma área geográfica, la zona centro de la Península Ibérica (Material y Métodos,
Apartado 3.1.2). Debido a esto, pudiera ser que la mayor multiplicación de lo aislados At que

Resultados
152
de los aislados Cu se debiera a una adaptación de los aislados de CMV a los individuos de
arabidopsis provenientes a la misma zona geográfica, ya que de los 21 individuos analizados,
5 pertenecen a esta zona del centro de la Península Ibérica. Para dilucidar esta cuestión, se
realizó un análisis de correlación entre los valores medios de acumulación de los aislados At y
Cu por individuo de arabidopsis y los valores de distancia de la localidad de procedencia de
cada individuo al centro de la Península (40.416º N, -3.703º O). Los resultados no mostraron
correlación significativa ni para los valores de acumulación de los aislados At (rp=-0.221,
rs=0.062, P>0.335) ni para los aislados Cu (rp=-0.054, rs=0.131, P>0.573), por lo que la
mayor multiplicación en arabidopsis de los aislados At respecto a los aislados Cu no se debe a
la procedencia geográfica de los aislados.
Además, se analizó si existía relación entre los valores de acumulación de los aislados
At y Cu en cada individuo de arabidopsis. Un análisis de correlación mostró correlación
positiva entre ambas variables (Figura 4.4.3), lo que indicó que los individuos de arabidopsis
más resistentes a los aislados At también los son, en conjunto, a los aislados Cu.
Figura 4.4.2 Correlación entre los valores medios de acumulación de los aislados At y Cu por individuo. La recta de color rojo indica la bisectriz.
rS=0.808, P<10-4
rP=0.895, P<10-4

Resultados
153
A continuación, se analizó la multiplicación de los aislados de CMV en melón. Un
GLM en el que se consideró aleatorio el factor aislado de CMV mostró diferencias
significativas de acumulación entre aislados de CMV (F7,72=10.679, P<10-4) (Var(aislado de
CMV=50.344)). Para analizar si éstas diferencias se asocian a su huésped de origen, se realizó
un GLM en el que se consideró fijo el factor huésped de origen de los aislados de CMV. Este
análisis mostró diferencias significativas de acumulación viral según el huésped de origen de
los aislados de CMV (F1,72=25.985, P<10-4) (Var(huésped de origen de los aislados de
CMV)=41.045)). Los mismos resultados se obtuvieron mediante el test no paramétrico
Kruskal-Wallis (P<10-4). La acumulación de los aislados Cu (32.669±2.327) fue mayor que la
de los aislados At (19.250±4.909), (Figura 4.4.4). Por tanto, estos resultados indican que
existe adaptación a huésped en los aislados de CMV.
Figura 4.4.4. Media±error estándar de la acumulación de los aislados At y los aislado Cu en arabidopsis y melón.
Es posible que un cambio de huésped implique un coste biológico para el aislado de
CMV, existiendo compromisos de adaptación. Para analizar esta hipótesis se realizó un GLM
en el que se consideraron como fijos los factores huésped de origen de los aislados y huésped
de ensayo. La acumulación viral difirió entre huésped de origen de los aislados
(F1,896=45.959, P<10-4) y entre huésped de ensayo (F1,896=460.048, P<10-4), y la interacción
entre los dos factores fue significativa (F1,890=64.249, P<10-4). Dada la interacción
significativa, se analizó por separado la acumulación de los aislados At y los aislados Cu. La
acumulación de los aislados At (en arabidopsis: 6.459±0.388 y en melón: 19.132±1.764); y los

Resultados
154
Cu (en arabidopsis: 5.293±0.319 y en melón: 33.090±2.119) fue mayor en melón que en
arabidopsis (At: F1,452=83.245, P<10-4, Cu: F1,444=482.980, P<10-4) (Figura 4.4.4), mostrando
que melón es mejor huésped de CMV que arabidopsis, independientemente del origen de los
aislados.
En conclusión, los resultados indican que tanto los aislados At como los aislados Cu
están adaptados a su huésped de origen. Además, tal adaptación está asociada a un
compromiso de adaptación a huésped en la multiplicación viral, ya que el cambio de huésped
de los aislados respecto a su huésped de origen implica una reducción de su multiplicación.
4.4.3 Virulencia de CMV en arabidopsis y melón
La virulencia en arabidopsis de los diferentes aislados de CMV se evaluó mediante dos
parámetros: i) el efecto de la infección de CMV sobre la producción de semillas (SWi/SWm), y
ii) el efecto de la infección de CMV sobre la biomasa (BMi/BMm), donde los subíndices i y m
corresponden a las plantas infectadas y a la media de los controles negativos, respectivamente.
Los datos de SWi/SWm y BMi/BMm se analizaron, en primer lugar, sin tener en cuenta el
huésped de origen de los aisaldos de CMV. Un análisis GLM en el que se consideraron
aleatorios los factores aislado de CMV e individuo de arabidopsis, mostró diferencias
significativas de SWi/SWm entre aislados de CMV (F7,824=21.130, P<10-4) e individuos de
arabidopsis (F20,824=5.154, P<10-4). La variable BMi/BMm no dirió significativamente según
aislado de CMV (F7,824=0.816, P=0.575) pero sí según individuo de arabidopsis
(F20,824=13.749, P<10-4). Además, el análisis mostró interacción significativa entre los factores
aislado de CMV x individuo de arabidopsis tanto para SWi/SWm como para BMi/BMm
(F140,824=4.785, P<10-4; F140,824=1.282, P=0.025, respectivamente) indicando, como en el
Apartado 4.2 y el Apartado 4.3, que el efecto de la infección por CMV sobre la producción
de semillas de arabidopsis dependió de una interacción genotipo del huésped x genotipo del
patógeno. El factor aislado de CMV explicó un 29% de la varianza de SWi/SWm
Var(individuo)=15.555, Var(aislado de CMV)=28.888 y Var(individuo x aislado de
CMV)=24.444) (Anejo 4, Tabla A4.4). El factor individuo de arabidopsis explicó un 14.7% y
la interacción de los dos factores analizados un 15.9% de la varianza de BMi/BMm
(Var(individuo)=14.772, Var(aislado de CMV)=19.318 y Var(individuo x aislado de
CMV=15.909)) (Anejo 4, Tabla A4.5).

Resultados
155
Para estudiar si las diferencias de SWi/SWm y BMi/BMm entre aislados de CMV se
asocian a su huésped de origen, se realizó un GLM para cada variable en el que se consideró
fijo el factor huésped de origen de los aislados y aleatorio el factor individuo de arabidopsis.
El resultado mostró diferencias significativas de SWi/SWm según el huésped de origen de los
aislados (F1,824= 50.020, P<10-4); siendo más virulentos los aislados At (0.482±0.013) que los
aislados Cu (0.640±0.014) y según el individuo de arabidopsis (F20,824=7.116, P<10-4) (Figura
4.4.5 y Anejo 4, Tabla A4.6). De acuerdo con estos resultados, el número de individuos en los
que los aislados At fueron más virulentos que los aislados Cu fue mayor que el número de
individuos en los que los aislados Cu fueron más virulentos que los aislados At (12/0,
χ2=20.167, P<10-4), (X ~ B(12,12,0.5, P<10-3) (Figura 4.4.5 y Figura 4.4.6). La relación
BMi/BMm no difirió significativamente según el huésped de origen de los aislados
(F1,824=1.603, P=0.315) pero sí según individuo de arabidopsis (F20,824=15.971, P<10-4). La
interacción entre los factores individuo de arabidopsis x huésped de origen fue marginalmente
significativa para SWi/SWm (F140,824=1.517, P=0.068) y no fue significativa para BMi/BMm
F140,824=1.052, P=0.397) (Anejo 4, Tabla A4.7). El factor huésped de origen de los aislados
explicó el 12.9% de la varianza observada de SWi/SWm (SWi/SWm Var(individuo)=16.129,
Var(huésped de origen)=12.903 y Var(individuo x huésped de origen)=2.150). El factor
individuo de arabidopsis explicó un 27.1% de la varianza de BMi/BMm
(Var(individuo)=27.142, Var(huésped de origen)=8.571 y Var(individuo x aislado de
CMV)=1.428) (Anejo 4, Tabla A4.7).
Al igual que con la multiplicación viral se analizó si las diferencias en el efecto de los
aislados At sobre la producción de semillas de los individuos de arabidopsis se debe a una
adaptación de los aislados a los individuos pertenecientes a la misma zona geográfica. El
análisis no mostró correlación significativa para los valores del efecto de los aislados At
(rp=0.001, rs=-0.091, P>0.696) y tampoco para el de los aislados Cu (rp=-0.111, rs=-0.140,
P>0.544). Por tanto, la mayor virulencia, medida como el efecto de la infección sobre la
producción de semillas, de los aislados At respecto a los aislados Cu no se debe a la
procedencia geográfica de los aislados.

Resultados
156
Figura 4.4.5. Variación de los valores medios de SWi/SWm (A) y BMi/BMm (B) de los aislados At n y Cu n en arabidopsis.
Además, se analizó si existía una relación entre los valores de virulencia
(SWi/SWm y BMi/BMm) de los aislados At y de los aislados Cu en cada individuo de
arabidopsis. Los análisis mostraron correlación positiva tanto para SWi/SWm como para
BMi/BMm. (Figura 4.4.6).
En conclusión, estos análisis revelaron que los aislados de CMV de arabidopsis fueron
más virulentos en arabidopsis que los aislados de cucurbitáceas, y que los individuos de
arabidopsis en los que los aislados At fueron más virulentos, en conjunto son los individuos de
arabidopsis en los que los aislados Cu fueron también más virulentos.
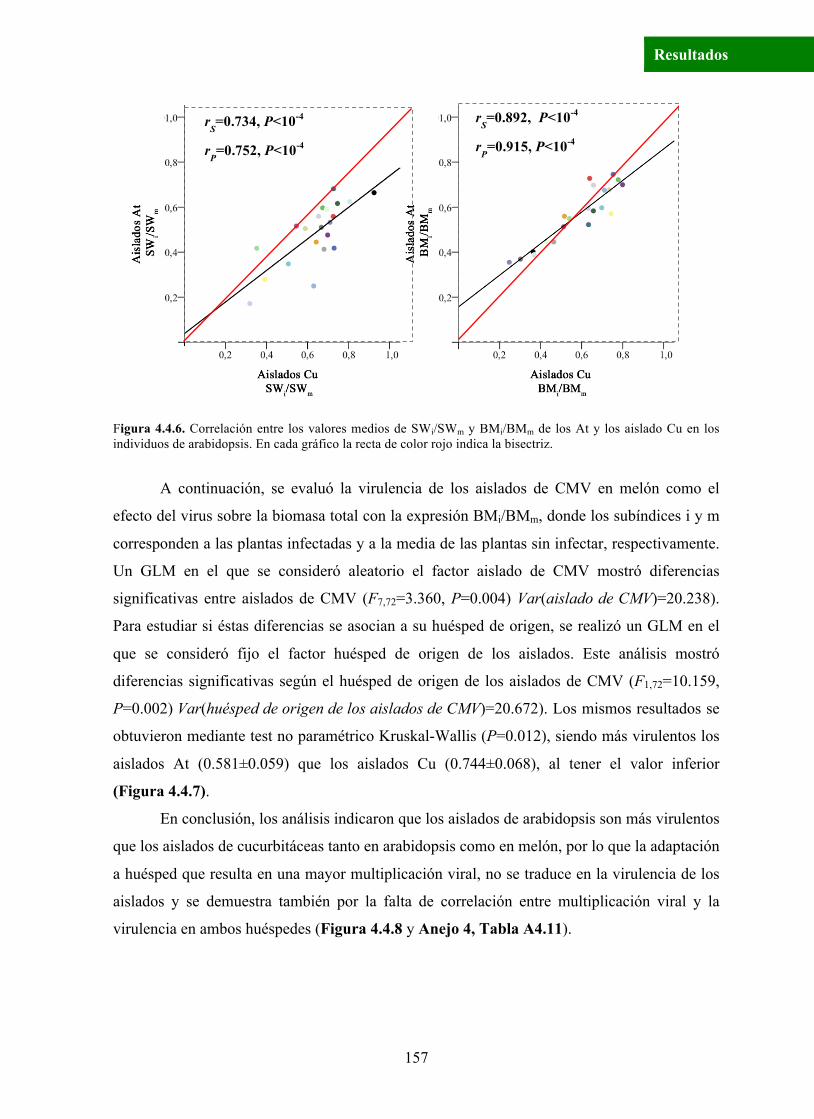
Resultados
157
Figura 4.4.6. Correlación entre los valores medios de SWi/SWm y BMi/BMm de los At y los aislado Cu en los individuos de arabidopsis. En cada gráfico la recta de color rojo indica la bisectriz.
A continuación, se evaluó la virulencia de los aislados de CMV en melón como el
efecto del virus sobre la biomasa total con la expresión BMi/BMm, donde los subíndices i y m
corresponden a las plantas infectadas y a la media de las plantas sin infectar, respectivamente.
Un GLM en el que se consideró aleatorio el factor aislado de CMV mostró diferencias
significativas entre aislados de CMV (F7,72=3.360, P=0.004) Var(aislado de CMV)=20.238).
Para estudiar si éstas diferencias se asocian a su huésped de origen, se realizó un GLM en el
que se consideró fijo el factor huésped de origen de los aislados. Este análisis mostró
diferencias significativas según el huésped de origen de los aislados de CMV (F1,72=10.159,
P=0.002) Var(huésped de origen de los aislados de CMV)=20.672). Los mismos resultados se
obtuvieron mediante test no paramétrico Kruskal-Wallis (P=0.012), siendo más virulentos los
aislados At (0.581±0.059) que los aislados Cu (0.744±0.068), al tener el valor inferior
(Figura 4.4.7).
En conclusión, los análisis indicaron que los aislados de arabidopsis son más virulentos
que los aislados de cucurbitáceas tanto en arabidopsis como en melón, por lo que la adaptación
a huésped que resulta en una mayor multiplicación viral, no se traduce en la virulencia de los
aislados y se demuestra también por la falta de correlación entre multiplicación viral y la
virulencia en ambos huéspedes (Figura 4.4.8 y Anejo 4, Tabla A4.11).
rS=0.734, P<10-4
rP=0.752, P<10-4
rS=0.892, P<10-4
rP=0.915, P<10-4

Resultados
158
Figura 4.4.7. Medias±error estándar de la virulencia (BMi/BMm) de los aislados At y los aislado Cu en arabidopsis y melón.
Figura 4.4.8. Relación entre la acumulación viral y SWi/SWm y la acumulación viral y BMi/BMm en los individuos de arabidopsis.
4.4.4 Relación entre los aislados de CMV de arabidopsis y cucurbitáceas
La adaptación a huésped de los aislados de CMV, puede generar que la diversidad
genética de los aislados esté estructurada según su huésped de origen. Para estimar la
diversidad genética y analizar las relaciones entre los aislados de CMV según su huésped de
BMi/B
Mm

Resultados
159
origen, se obtuvieron secuencias parciales de los tres ARNs (ARN 1: fragmento de 2397
nucleótidos, ARN 2: fragmento de 2847 nucleótidos y ARN 3: fragmento de 1918
nucleótidos) de cada uno de los 8 aislados CMV (Material y Métodos, Apartado 3.7). A
partir de los alineamientos de las secuencias nucleotídicas concatenadas de los fragmentos de
los 3 ARN se infirió un árbol filogenético por ML (Material y Métodos, Apartado 3.9.5).
Dicho análisis mostró que los 8 aislado de CMV analizados pertenecen al Subgrupo IA al estar
en el mismo clado que el aislado Fny-CMV (NC_002034, NC_002035, NC_001440), aislado
tipo del Subgrupo IA (Figura 4.4.9). Este resultado además indica que los aislados At y Cu
son del mismo grupo que los descritos de otros huéspedes en esta misma región (Fraile et al.,
1997). El mismo resultado se obtuvo al analizar cada fragmento de ARN (Anejo 4, Figura
A4.1). En la inspección visual del árbol se aprecia que los aislados están agrupados
genéticamente en función del huésped de origen de los mismos (Figura 4.4.9 y Anejo 4,
Figura A4.1). Para confirmar esta observación, se realizó un análisis cuantitativo de
asociación entre la filogenia de los aislados de CMV y su huésped de origen.
Figura 4.4.9. Árbol filogenético a partir de la secuencia de los fragmentos de los 3 ARN concatenados de los 8 aislados de CMV inferido por ML y enraizado en Ls. El aislado Ls es un aislado tipo del Grupo II y el aislado Fny del subgrupo IA. Los asteriscos indican nodos con una significación mayor al 75% basada en 1000 réplicas.
Resultados
160
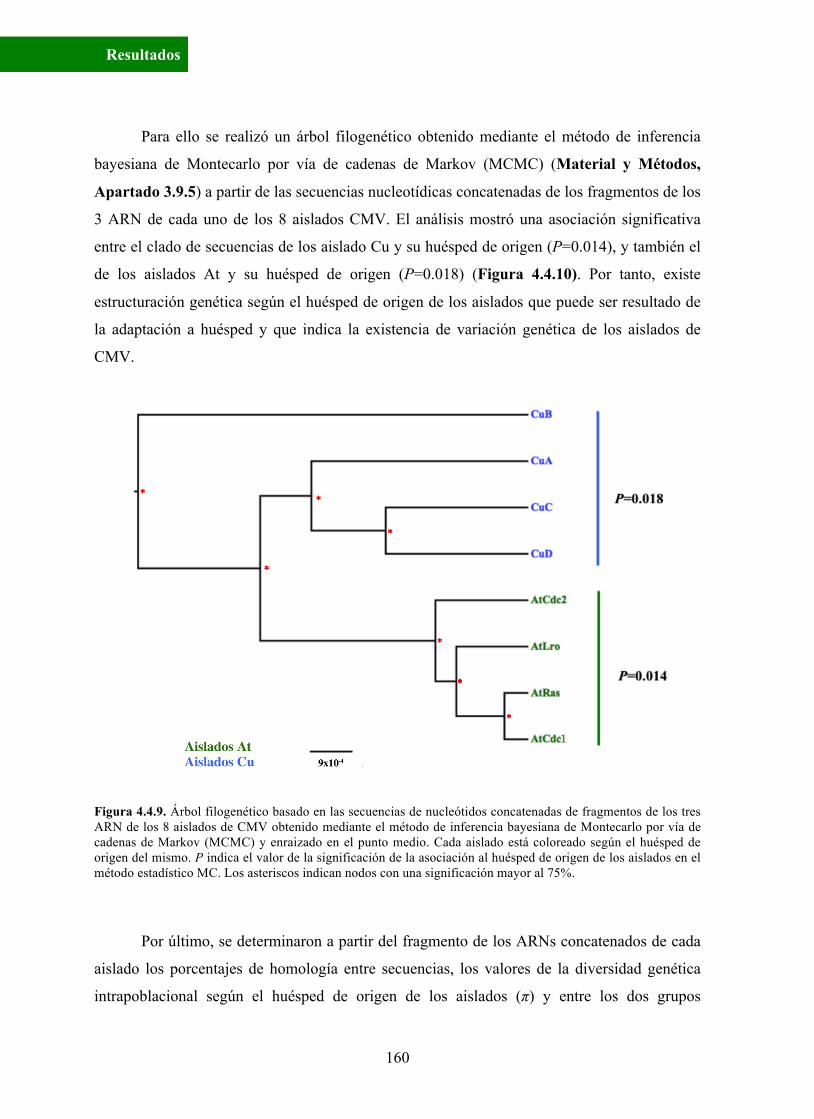
Para ello se realizó un árbol filogenético obtenido mediante el método de inferencia
bayesiana de Montecarlo por vía de cadenas de Markov (MCMC) (Material y Métodos,
Apartado 3.9.5) a partir de las secuencias nucleotídicas concatenadas de los fragmentos de los
3 ARN de cada uno de los 8 aislados CMV. El análisis mostró una asociación significativa
entre el clado de secuencias de los aislado Cu y su huésped de origen (P=0.014), y también el
de los aislados At y su huésped de origen (P=0.018) (Figura 4.4.10). Por tanto, existe
estructuración genética según el huésped de origen de los aislados que puede ser resultado de
la adaptación a huésped y que indica la existencia de variación genética de los aislados de
CMV.
Figura 4.4.9. Árbol filogenético basado en las secuencias de nucleótidos concatenadas de fragmentos de los tres ARN de los 8 aislados de CMV obtenido mediante el método de inferencia bayesiana de Montecarlo por vía de cadenas de Markov (MCMC) y enraizado en el punto medio. Cada aislado está coloreado según el huésped de origen del mismo. P indica el valor de la significación de la asociación al huésped de origen de los aislados en el método estadístico MC. Los asteriscos indican nodos con una significación mayor al 75%.
Por último, se determinaron a partir del fragmento de los ARNs concatenados de cada
aislado los porcentajes de homología entre secuencias, los valores de la diversidad genética
intrapoblacional según el huésped de origen de los aislados (π) y entre los dos grupos
Resultados
161
(diversidad genética interpoblacional) y el valor del coeficiente de diferenciación nucleotídica
(Nst). Los porcentajes de homología entre parejas de secuencias fueron de 98% y 99% (Tabla
4.1), la diversidad genética media en el conjunto de los aislados At (0.003±0.001) fue
significativamente menor que en el conjunto de los aislados Cu (0.011±0.001), al menos 0.5
veces, el valor de diversidad genética entre los dos grupos fue 0.013±0.001 y el valor Nst,
0.319±0.039. Estos resultados indicaron que la relación genética entre los aislados de CMV
está determinada por el huésped de origen de los mismos y que entre un 13% y un 30% de la
diversidad existente entre los aislados es explicada por su huésped de origen de los aislados
ensayados.
Tabla 4.1. Porcentaje de homología entre parejas de las secuencias del fragmento de los ARNs concatenados de cada asilado de CMV analizado.
AtCdc1 AtCdc2 AtLro AtRas CuA CuB CuC
AtCdc1
AtCdc2 99%
AtLro 99% 99%
AtRas 99% 99% 99%
CuA 99% 99% 99% 99%
CuB 98% 98% 99% 98% 99%
CuC 99% 98% 99% 99% 99% 99%
CuD 99% 98% 99% 99% 99% 98% 99%

Resultados
162

5. DISCUSIÓN

Discusión
164

Discusión
165
Los estudios de coevolución planta-virus requieren un ecosistema silvestre en el cual
no haya existido, ni exista, manipulación humana sobre el huésped o el patógeno. Aunque la
evidencia derivada de estos ecosistemas es escasa, la coevolución planta-virus es aceptada
comúnmente (Fraile & García-Arenal, 2010, Pagán et al., 2010). La selección de un
patosistema silvestre requiere del conocimiento de los virus que infectan las poblaciones de
plantas, su incidencia y los posibles efectos de las infecciones virales sobre las plantas y las
poblaciones de estas. Dichas informaciones no son numerosas debido a la escasez de este tipo
de estudios (Alexander et al., 2014; Cooper & Jones, 2006; Roossinck & García-Arenal,
2015), aunque los existentes muestran que las infecciones virales son frecuentes en
ecosistemas silvestres (Haydon et al., 2002; Roossinck & García-Arenal, 2015). De hecho,
con el desarrollo de nuevas técnicas como la secuenciación masiva ha aumentado el
conocimiento sobre las especies virales que infectan las plantas silvestres (Roossinck &
García-Arenal, 2015). La evidencia sobre el efecto de las infecciones virales sobre las plantas
silvestres y sus poblaciones también es escasa (Power et al., 2011; Prendeville et al., 2014;
Rodelo-Urrego et al., 2013; Rua et al., 2011), pero algunos trabajos muestran que tal efecto
puede ser diverso dependiendo del ambiente (Malmstrom et al., 2011, Prendeville et al.,
2014), negativo (Kelley, 1994; Malmstrom et al., 2006; Maskell et al., 1999; Remold, 2002),
positivo (Melcher et al., 2008; Muthukumar et al., 2009; Roossinck, 2005) o no existir
(Malmstrom et al., 2005). Dichos efectos virales pueden influir en la germinación, el
crecimiento, la supervivencia, la redistribución de recursos o a la fecundidad de las plantas; y
determinar por tanto la dinámica de sus poblaciones (Funayama et al., 2001; Maskell et al.,
1999; Malmstrom et al., 2005a, b, y 2006; Power & Mitchell, 2004; Seabloom et al., 2009;
Yahara & Oyama 1993).
En esta tesis se ha seleccionado arabidopsis como planta silvestre para llevar a cabo un
estudio de coevolución planta-virus. En el trabajo realizado por Pagán y col (2010), se
detectaron infecciones por cinco especies virales en poblaciones silvestres de arabidopsis
localizadas en la Meseta Central de la Península Ibérica. Por ello, se han seleccionado cinco de
estas poblaciones para llevar a cabo un estudio sobre la incidencia de cinco virus y su efecto
en la demografía del huésped, considerando distintos factores del medio abiótico.

Discusión
166
5.1. Dinámica de las infecciones virales en poblaciones silvestres de
arabidopsis
La primera parte de este trabajo de tesis se centra en el estudio la demografía de
arabidopsis. El ciclo natural de arabidopsis está condicionado por el ambiente: en las
poblaciones naturales de esta planta sometidas a inviernos muy fríos, como las de alta
montaña, los individuos nacidos en otoño apenas sobreviven al invierno por lo que la
población fundamentalmente la componen las plantas nacidas en primavera (Picó, 2012). En
cambio, en las poblaciones localizadas en zonas con inviernos suaves, las plantas nacidas en
otoño sobreviven durante el invierno. En estas condiciones, el frío tiene un papel central en el
ciclo de vida de arabidopsis ya que activa la ruta de vernalización, gracias a la cual las plantas
que permanecen en estado vegetativo durante el invierno y florecen en primavera. Como
consecuencia, en primavera solapan dos cohortes: las plantas nacidas en otoño y las nacidas a
principios de primavera (Méndez-Vigo et al., 2010). Los datos del Apartado 4.1 muestran
que, en general, en todas las poblaciones se han observado dos cohortes de germinación, en
otoño y en primavera, y que la mayor parte de las plantas de la primera cohorte sobreviven al
invierno y florecen en primavera. Estos resultados coinciden con lo descrito en otras
poblaciones con inviernos suaves o moderados de la Península Ibérica y del Mundo (Callahan
& Pigliucci, 2002; Griffith et al., 2004; Méndez-Vigo et al., 2012; Montesinos et al., 2009;
Picó, 2012; Shimizu & Purugganan, 2005).
El clima es un factor fundamental en la distribución geográfica de muchas especies de
plantas (Archibold, 1995; Woodward, 1987). De hecho, las variaciones climáticas pueden
producir alteraciones en procesos fisiológicos (Berry & Björkman 1980; Körner, 2007;
Williams & Black, 1993), caracteres morfológicos, y parámetros demográficos como la
geminación o el periodo de dormancia de las semillas (Cavieres & Arroyo, 2001; Cummins &
Miller, 2002; Ma et al., 2010; Phillips et al., 2011, Picó, 2012). En el caso de arabidopsis, se
ha propuesto que la temperatura y la precipitación pueden ser los factores más importantes
(Hoffman, 2005). Por ejemplo, las temperaturas invernales son un factor abiótico que influye
en le ciclo de vida de arabidopsis (Picó, 2012). Se ha demostrado experimentalmente que los
individuos de arabidopsis procedentes de altitudes elevadas poseen tasas de crecimiento
mayores en las primeras etapas de su desarrollo para aumentar el tamaño de la planta antes de
la llegada del invierno. Sin embargo, su ritmo de crecimiento es más lento en las etapas

Discusión
167
posteriores para alargar el período pre-reproductor antes de la floración (Montesinos et al.,
2009; Montesinos-Navarro et al., 2011). En el presente trabajo, se analizó si las
precipitaciones y las temperaturas pudieran ser los factores causantes de la variación de los
valores de densidad de plantas en las poblaciones de arabidopsis seleccionadas. Los resultados
muestran que las precipitaciones no influyen en los valores de densidad. Esta escasa
importancia de las precipitaciones podría deberse a que tienen un efecto cualitativo, y no tanto
cuantitativo. Es decir, que la germinación de las semillas y el mantenimiento de las plantas
requeriría niveles bajos de precipitación; y una vez alcanzado ese umbral mínimo, cualquier
aumento de las precipitaciones tendría poca importancia para germinación de las semillas y la
supervivencia de las plantas. En cambio, los resultados indican que la temperatura tiene un
papel fundamental en la demografía de arabidopsis. Los valores de densidad de plantas en
otoño son mayores cuando las temperaturas de Septiembre, Octubre y Noviembre son
moderadas. Por tanto, se podría pensar que las temperaturas suaves de finales de verano y
otoño favorecen la germinación de las semillas y la supervivencia de las plantas. Además, los
valores de densidad de la especie en primavera están relacionados con las temperaturas
invernales, ya que los valores de densidad de Marzo-Mayo son mayores cuando las
temperaturas de Diciembre-Febrero son moderadas. Estas observaciones concuerdan con lo
sugerido por Montesino y col (2009), que proponen que los inviernos extremos contribuirían a
un fuerte periodo secundario de dormancia, reduciendo la germinación de la segunda cohorte y
reducirían la supervivencia de las plantas durante este periodo. Los resultados indican también
que las temperaturas suaves durante Marzo-Mayo favorecen una elevada densidad de plantas
en primavera. Por tanto, los factor meteorológicos explican parcialmente los valores de
densidad de plantas. Es obvio que otros factores además de la temperatura podrían tener un
papel importante en la demografía de arabidopsis. Un claro ejemplo pueden ser las
interacciones bióticas. Así, sería necesario considerar en futuros estudios el efecto de las
especies florales acompañantes y sus densidades, ya que arabidopsis es mala competidora y
necesita colonizar lugares abiertos con poca vegetación (Bergelson et al., 1998; Mauricio,
1998; Montesinos et al., 2009, Picó, 2012). Otro factor a considerar serían las infecciones por
patógenos, ya que no solo afectan a la supervivencia y la eficacia biológica de arabidopsis
(Pagán et al., 2007 y 2008), sino también a su habilidad competitiva (Pagán et al., 2009). Este
último factor es en el que se centra esta tesis, y en concreto en las infecciones virales.
En las cinco poblaciones de arabidopsis consideradas se analizó la incidencia de cinco
virus, CaMV, CMV, TCV, TuMV y TYMV. Estos virus se seleccionaron porque se han

Discusión
168
demostrado sus infecciones en especies silvestres de la familia Brassicaceae en Europa, en la
Meseta Central de la Península Ibérica (Malpica et al., 2006; Maskell et al., 1999; Pallett et al.,
2002; Sacristán et al., 2004; Thurston et al., 2001) y en poblaciones de arabidopsis del mismo
área (Pagán et al., 2010). Los cinco virus se detectaron en las cinco poblaciones analizadas y
en la mayoría de los años en los que se realizó el estudio (Pagán et al., 2010). En ninguna de
las plantas infectadas se observaron síntomas, lo que ha sido descrito en numerosos de trabajos
en los que se han analizado las infecciones virales en plantas silvestres (Pagán et al., 2010;
Prendeville et al., 2012; Roossinck, 2012; Roossinck et al., 2008; Stobbe & Roossinck, 2014).
Los valores de incidencias máximas de virus variaron según los años y las poblaciones,
llegando en ocasiones hasta un 80% de plantas infectadas. Dichos valores ocurren en
primavera cuando en las poblaciones de arabidopsis se encuentran plantas en periodo pre-
reproductor y en el inicio del periodo reproductor. Es decir, los valores más elevados de
incidencia se dan en las primeras fases del ciclo biológico de arabidopsis, cuando las
infecciones virales tienen un efecto negativo mayor sobre las plantas (Larsen et al., 2002;
Matthews, 2014), inclusive arabidopsis (Pagán et al., 2007). Esto sugiere que las infecciones
virales pueden ser un factor importante en la dinámica poblacional de arabidopsis, como se ha
demostrado en otros huéspedes (Kelley, 1994; Maskell et al., 1999; Remold, 2002;
Malmstrom et al., 2005a). El efecto de las infecciones virales en la dinámica poblacional de
arabidopsis se analizó utilizando como parámetro poblacional la densidad del huésped.
Además, este parámetro se seleccionó porque se ha propuesto que un incremento de la
densidad del huésped puede aumentar la incidencia del patógeno debido al aumento del
contacto entre huéspedes infectados y huéspedes no infectados (Burdon & Chilvers, 1982).
Los resultados no muestran relación entre los valores de densidad y los valores de la
incidencia de virus, lo que no apoyaría la hipótesis anterior.
Los efectos de diferentes factores que no han sido considerados en esta tesis podrían
explicar la ausencia de asociación entre densidad de arabidopsis e incidencia de virus. Por
ejemplo, factores bióticos como la herbivoría, de la cual se han encontrados signos en todas
las poblaciones, o el efecto de otros patógenos como las infecciones por los oomicetos Albugo
sp. y Hyaloperonospora arabidopsis Gäum., que han sido detectadas en algunas de las
poblaciones analizadas (Cdc, Lro, Mar) y descritas en otras poblaciones (Holub et al., 1994;
Thines et al., 2014, van Damme et al., 2005). También cabe la posibilidad de que los valores
de incidencia de virus estuvieran relacionados con los valores de densidad total de plantas
huéspedes y no con los valores de densidad de arabidopsis, sobre todo en virus generalistas

Discusión
169
como CMV. Otro factor a considerar sería el efecto de las condiciones climatológicas. En
estudios realizados en cultivos de la misma región, se ha demostrado que las condiciones
climatológicas, principalmente las temperaturas mínimas invernales, mínimas y medias
primaverales, y la pluviometría primaveral, afectan a la dinámica de la incidencia de los virus
transmitidos por vectores (Alonso-Prados et al., 2003; Luis-Arteaga et al., 1998; Moreno et al.,
2004). Sin embargo, en diferentes periodos del ciclo vital de arabidopsis no se encuentra
relación significativa entre los valores de incidencia máxima de virus en las poblaciones y los
de diferentes factores meteorológicos anuales. Por tanto, los factores meteorológicos no
explican la variación de la incidencia máxima de los diferentes virus entre años ni a lo largo
del año. Por último, otra posible causa de la variación espacio-temporal de la incidencia de
virus, y de no detectar el efecto de las infecciones por virus sobre la densidad del huésped
puede ser existencia de variación genética de las defensas de las poblaciones del huésped a
cada virus. La diversidad genética de las poblaciones silvestres de plantas es mayor que la de
los cultivos; además, los caracteres de defensa, como la resistencia son de los más variables en
el genoma de las plantas (Raybould et al., 2001). Por tanto, en las poblaciones silvestres
existirán individuos diferenciados por distintos niveles de susceptibilidad a las infecciones,
mientras que en los cultivos no. Arabidopsis es una especie con una amplia variación genética
para diversos caracteres entre los cuales también se encuentran la resistencia y la tolerancia a
patógenos (Karasov et al., 2014; Salvaudon, 2008; Pagán et al., 2007 y 2010). Por ello, es
posible que el efecto de las infecciones de virus sobre la dinámica de las poblaciones no sea
evidente debido a las diferentes respuestas de defensa de los individuos que las componen.
En resumen, la intensidad del efecto de las infecciones virales sobre las poblaciones de
arabidopsis podría depender del efecto, individual o en combinación, de distintos factores
como la estructura genética de la población del huésped y la del patógeno, el tiempo en el cual
se produce la infección, y los costes directos e indirectos de la infección en cada una de las
interacciones entre genotipos del huésped y del patógeno. En consecuencia, sería necesario
abordar un análisis de los efectos de las infecciones virales en las poblaciones de arabidopsis
desde otro enfoque. Esta tesis aborda el análisis de la coevolución planta-virus en condiciones
experimentales, ya que el efecto de algunos de los factores anteriormente mencionados se
relaciona directamente con los procesos de coevolución.

Discusión
170
5.2 Evidencia de coevolución arabidopsis-CMV
De los cinco virus detectados en las poblaciones de arabidopsis analizadas, CMV es el
de mayor incidencia en la mayoría de los años y las poblaciones. Además, se ha demostrado
experimentalmente el efecto negativo de las infecciones de diferentes aislados de CMV sobre
la eficacia biológica de distintos individuos (Pagán et al., 2007 y 2008). Por tanto, de los cinco
virus seleccionados, es probable que CMV sea uno de los que tenga un efecto mayor sobre la
evolución de las poblaciones de arabidopsis de la Meseta Central de la Península Ibérica. En
consecuencia, se ha seleccionado el patosistema arabidopsis-CMV para llevar a cabo un
estudio de coevolución planta-virus. Las poblaciones de arabidopsis utilizadas en esta tesis se
seleccionaron por ser representativas de la variación genética de la especie y del ambiente en
la Península Ibérica (Méndez-Vigo et al., 2011; Picó et al., 2008). La variación genética de
CMV se representó mediante el uso de dos aislados, Cdc1-CMV aislado del Subgrupo IA
procedente de arabidopsis y Fny-CMV aislado tipo del mismo subgrupo procedente de melón;
grupo al cual pertenecen la mayoría de los aislados de CMV de la Meseta Central de la
Península Ibérica (Fraile et al., 1997). Para analizar la evidencia de coevolución entre
arabidopsis y CMV se han planteado distintos enfoques experimentales que en conjunto dan
respuestas a los objetivos planteados.
5.2.1 Variación genética de las defensas de arabidopsis a CMV e interacción
entre los genotipos del huésped y del patógeno
El objetivo 2 de esta tesis está ligado a dos de las condiciones necesarias para que se de
coevolución huésped-patógeno que son: 1) la existencia de variación genética de los caracteres
relevantes para la interacción entre el huésped y el patógeno, y 2) el resultado de la interacción
huésped-patógeno ha de depender de la naturaleza de los genotipos del huésped y del patógeno
(Woolhouse et al., 2002). Como la resistencia y la tolerancia son los mecanismos principales
de defensa de las plantas frente a virus (Mauricio et al., 1997; Simms & Triplett, 1994), son
caracteres fundamentales en la interacción planta-virus. Por tanto, esta tesis se ha centrado en
los dos principales mecanismos de defensa. Dado que los caracteres cuantitativos, tales como
la resistencia y la tolerancia, tienen una fuerte componente ambiental (Falconer & Mackay,

Discusión
171
1996), se analizó la variabilidad de ambos caracteres en dos condiciones experimentales
diferentes, Ambiente 1 y Ambiente 2.
En primer lugar, se comprobó que los mecanismos de tolerancia a CMV descritos
previamente en arabidopsis se daban en los individuos utilizados en las dos condiciones
experimentales. En general, los mecanismos de tolerancia de los huéspedes a los patógenos
pueden estar asociados a cambios en la historia vital del huésped (Mauricio et al., 1997;
Simms & Triplett., 1994). Existen modelos basados en la teoría de la historia vital que
proponen la existencia de compromisos en la distribución de recursos entre los diferentes
componentes de la eficacia biológica de un organismo como la reproducción, el crecimiento o
la supervivencia (Stearns, 1976). El patrón de distribución de recursos dependerá de las
condiciones ambientales, entre ellas las infecciones por patógenos (van Noordwijk & de Jong,
1986; Michalakis & Hochberg, 1994; Koella & Agnew, 1999; Agnew et al., 2000).
Previamente, Pagán y col (2008) han descrito que los niveles de tolerancia a CMV de los
individuos de arabidopsis están relacionados con la arquitectura de la planta, de tal modo que
existen dos grupos alométricos que difieren en su arquitectura y en su tolerancia a CMV:
Grupo 1 y Grupo 2. Los individuos del Grupo 1 se definen por tener un mayor periodo pre-
reproductor que reproductor, mientras que los individuos del Grupo 2 ocurre a la inversa. Los
individuos del Grupo 1 son más tolerantes a CMV que los del Grupo 2, debido en parte, a que
la infección por CMV provoca que se destine mayor cantidad de recursos a la producción de
semillas en vez de al desarrollo de estructuras vegetativas, en comparación con las plantas no
infectadas (Pagán et al., 2008). Los resultados del Apartado 4.2 indican que los individuos del
Grupo 1 fueron más tolerantes a CMV, y que tal tolerancia se debió, en parte, a una
redistribución de recursos de crecimiento vegetativo a la reproducción.
La infección por patógenos en las plantas también puede alterar la duración de su ciclo
biológico (Hily et al., 2015; Pagán et al., 2008; Salvaudon & Shykoff, 2013). Según los
modelos sobre la teoría de la historia vital, los huéspedes menos tolerantes sufrirían una
reducción de su periodo pre-reproductor a causa de la infección por el patógeno para producir
la progenie antes de su muerte; en cambio, en los huéspedes más tolerantes, la infección
aumentaría el periodo pre-reproductor para permitir la compensación del daño producido por
el patógeno y por tanto se retrasaría la reproducción del huésped (Hochberget al., 1992;
Gandon et al., 2002). La infección por CMV en arabidopsis, también tiene un efecto sobre la
duración del ciclo vital de las plantas que difiere según el grupo alométrico al que pertenezca
el individuo. En los individuos más tolerantes a CMV, pertenecientes al Grupo 1, la infección

Discusión
172
prolonga el periodo pre-reproductor; no así en los individuos menos tolerantes, los del Grupo
2 (Pagán et al., 2008). En los individuos de arabidopsis analizados en el Apartado 4.2, se
pudieron diferenciar, en los dos ambientes diferentes, los dos grupos alométricos descritos en
arabidopsis. Los resultados del Apartado 4.2 concuerdan con resultados previos en el mismo
patosistema (Pagán et al., 2008; Hily et al., 2015) que evidencian experimentalmente que los
individuos menos tolerantes a la infección reducen su periodo pre-reproductor, mientras que
los del Grupo 1 lo aumentan, frente a la infección por CMV.
Se ha demostrado que el ambiente influye en las respuestas de defensa de arabidopsis a
CMV (Hily et al., 2015). Los resultados del Apartado 4.2 muestran que el ambiente produce
variaciones en las relaciones arquitectónicas de las plantas ya que 24 individuos de 56 se
clasificaron en distinto grupo alométrico según el ambiente. Pero a pesar de estas variaciones,
los grupos alométricos identificados se comportan de igual modo frente a la infección por
CMV en el Ambiente 1 y en el Ambiente 2, como se han descrito en otros trabajos (Hily et
al., 2015; Pagán et al., 2008). Por tanto, los resultados de los dos ambientes muestran que la
tolerancia a CMV de los individuos de distintas poblaciones de la Península Ibérica es
producida, en parte, por un mecanismo de redistribución de recursos y son compatibles con los
modelos de la teoría de la historia vital así como con los descritos previamente en el
patosistema arabidopsis-CMV.
Demostrado, por tanto, que en los individuos de arabidopsis analizados se expresa la
tolerancia a CMV, se analizó la variación genética de los dos caracteres de defensa a CMV, la
resistencia y la tolerancia. La variación genética de ambos caracteres se analizó entre y dentro
de poblaciones de la Península Ibérica, y entre haplotipos, para evitar la posible sobreestima
de la variación genética en los dos caracteres entre individuos, causada por factores
ambientales no controlados (Apartado 4.2). También se analizó la variación genética de la
resistencia y la tolerancia a CMV entre individuos de diferentes poblaciones, obteniéndose
resultados similares (Apartado 4.3 y 4.4). En ninguno de los individuos de arabidopsis
utilizados en los diferentes experimentos de esta tesis se detectó resistencia cualitativa a
ninguno de los aislados de CMV, por ello se evaluó en todos los casos la resistencia
cuantitativa. Además, los resultados del Apartado 4.2 se obtuvieron de dos condiciones
experimentales diferentes, Ambiente 1 y Ambiente 2, de este modo se pudo analizar el efecto
del ambiente en los caracteres de defensa a CMV (Apartado 4.2). En el Ambiente 1, los
resultados muestran variación genética de la resistencia a Cdc1-CMV y a Fny-CMV entre los
individuos, los haplotipos y las poblaciones de arabidopsis. Esta variación tiene una fuerte

Discusión
173
componente genética ya que el 50% de la variación se debe a diferencias entre individuos y el
30% entre haplotipos. Además, los valores de heredabilidad de las poblaciones para ambas
resistencias fueron de moderados a elevados (0.272-0.899). Al igual que la resistencia, el nivel
de tolerancia a Cdc1-CMV y a Fny-CMV en el Ambiente 1 fue variable entre individuos,
haplotipos y poblaciones con una fuerte componente genética, ya que el 49% de la variación
se debió a diferencias entre individuos y el 27% entre haplotipos. Los valores de heredabilidad
de las poblaciones para la tolerancia a CMV fueron menores que los de la resistencia, pero
también de moderados a altos (0.103-0.803). La menor heredabilidad de la tolerancia puede
deberse a que es un carácter con una influencia mayor del ambiente que la resistencia (Hily et
al., 2015). En el Ambiente 2 los resultados indican conclusiones similares a las del Ambiente 1
(Apartado 4.2). Los experimentos del Apartado 4.3 y el Apartado 4.4 también mostraron
variación genética de los dos caracteres entre individuos de arabidopsis de diferentes
poblaciones. Los trabajos que analizan la variación de la resistencia a patógenos de plantas
dentro (e.j. Burdon & Jarosz, 1991; Jarosz & Burdon, 1990; Parker, 1989; Sorci et al., 1997) o
entre poblaciones (e.j. Kaltz & Shykoff, 1998; van Zandt & Mopper, 1998) son numerosos,
pero los trabajos que estudian ambas variaciones simultáneamente son más escasos
(Antonovics et al., 1994; Burdon, 1987; Ebert et al., 1998; Imhoof & Schmid-Hempel, 1998;
Laine, 2004; Thrall et al., 2001). Aún más escasos son los análisis que incluyen también el
efecto de la variación genética del patógeno (Bevan et al., 1993a, b; de Nooij & van Damme,
1988a, b; Jarosz & Burdon, 1991; Pagán et al., 2010), y sólo uno, que conozca, esta basado en
la interacción planta-virus, en concreto CMV-arabidopsis (Pagán et al., 2010). Los
mecanismos de tolerancia a patógenos de plantas y la variación genética de la tolerancia entre
y dentro de las poblaciones se ha estudiado mucho menos que la de la resistencia (Fraile &
García-Arenal, 2010; Little et al., 2010). Los únicos estudios publicados se centran en la
variación genética de la tolerancia a patógenos entre individuos de diferentes poblaciones
(Kover & Schaal, 2002, Pagán et al., 2007; Salvaudon et al., 2005) pero no entre poblaciones
diferentes. Por tanto, los resultados de esta tesis no sólo son coherentes con trabajos previos
que evidencian la existencia de polimorfismos para la resistencia en las poblaciones del
huésped, sino que también demuestran por primera vez la existencia de polimorfismos para la
tolerancia en las interacciones planta-virus, una de las condiciones necesarias para que exista
coevolución. Además, la existencia de esta diversidad genética de las defensas frente a CMV
en las poblaciones de arabidopsis podría explicar la ausencia de relación entre la incidencia de
CMV y la densidad de arabidopsis en poblaciones silvestres observada en el Aparado 4.1. De

Discusión
174
hecho, los resultados del Apartado 4.2 muestran que dos de las poblaciones analizadas en el
Apartado 4.1 (Cdc y Mar) son variables genéticamente para los dos caracteres de defensa a
CMV analizados.
Otra de las condiciones para que se de coevolución planta-virus es que el resultado de
la interacción huésped-patógeno dependa de la interacción entre los genotipos del huésped y
del patógeno. Los resultados de todos los experimentos realizados con diferentes individuos de
arabidopsis y diferentes aislados de CMV en el presente trabajo (Apartados 4.2, 4.3 y 4.4)
muestran que la resistencia y la tolerancia a CMV dependen de la interacción genotipo de
arabidopsis x genotipo de CMV. Estos resultados son coherentes con trabajos previos en
interacciones planta-patógeno (Pagán et al., 2010; Salvaudon et al., 2008). En consecuencia,
nuevamente los resultados argumentan que hay coevolución entre arabidopsis-CMV.
La expresión simultánea de la resistencia y la tolerancia a patógenos supondrían para el
huésped una inversión de recursos en dos funciones con un mismo fin, y por tanto
redundantes. Por ello, se han propuesto varios modelos que predicen un compromiso entre
resistencia y tolerancia, y por tanto una correlación negativa entre ambos caracteres (Fineblum
& Rausher, 1995; Mauricio, 2000; Simms & Triplett, 1994; Stowe, 1998; Strauss & Agrawal,
1999; Tiffin, 2000; van der Meijden, et al, 1988), que podría estar modulada por las
condiciones ambientales (Fornoni et al., 2003). Los resultados del Apartado 4.2, Apartado
4.3 y del Apartado 4.4 no muestran correlación negativa entre la resistencia y la tolerancia a
cualquiera de los aislados de CMV en ninguno de los ambientes analizados. Por tanto, los
resultados no indican un compromiso entre la resistencia y la tolerancia a CMV en
arabidopsis, lo que es coherente con resultados obtenidos en otros trabajos donde se ha
empleado el mismo patosistema (Pagán et al., 2008) o el mismo virus (Carr et al., 2005). Esta
ausencia de correlación se ha explicado por la inexistencia de costes de la resistencia y la
tolerancia, o porque la tolerancia esté ligada a otros caracteres del huésped (Mauricio et al.,
1997 y 2000). En los diferentes experimentos realizados en esta tesis, no se hayan costes de la
resistencia ni de la tolerancia a ninguno de los aislados de CMV, ya que los individuos sin
infectar más resistentes o más tolerantes no muestran una menor producción de semillas
(rP<0.075, rS<0.053, P>0.461). Por tanto, esta ausencia de costes podría explicar al menos en
parte la ausencia de correlación entre tolerancia y resistencia. No se puede descartar que estos
dos caracteres de defensa de arabidopsis a CMV estén ligados a otros caracteres con funciones
importantes para la biología de arabidopsis. Sin embargo, dicha posibilidad no se ha analizado
en este trabajo.

Discusión
175
5.2.2 Selección de la resistencia y la tolerancia de arabidopsis a CMV
Otra cuestión relevante, por su implicación en la coevolución planta-patógeno, es la
determinación de la contribución de la selección en la evolución de la tolerancia y la
resistencia. La determinación de la contribución de la selección natural a la evolución de un
carácter cuantitativo se puede llevar a acabo mediante la comparación de los patrones de
diferenciación genética entre poblaciones para marcadores supuestamente neutrales y para el
carácter estudiado, y así determinar si la evolución de ambos caracteres ha seguido el mismo
patrón (Rogers, 1986; Spitze, 1993). Para abordar esta cuestión, uno de los métodos más
adecuados es la comparación entre la cantidad de diferenciación genética de caracteres
cuantitativos entre poblaciones (QST) y la cantidad de diferenciación genética de marcadores
neutrales entre poblaciones (FST). Si se realiza la comparación entre ambos puede darse alguna
de las situaciones siguientes:
- QST < FST: La diferenciación genética observada para el carácter cuantitativo es menor
que la diferenciación genética neutra. La selección actuaría favoreciendo a los fenotipos
más cercanos a la media poblacional de manera que existe un efecto uniformador.
- QST = FST: En este caso la cantidad de diferenciación genética debida a procesos
demográficos es igual a la del carácter cuantitativo, por lo tanto, no se puede concluir
que el caracter esté bajo selección.
- QST > FST: la diferenciación genética del carácter cuantitativo entre poblaciones es
mayor que la diferenciación genética neutra. Esto indica que la selección es direccional,
favoreciendo a distintos fenotipos en distintas poblaciones.
Cuando se estiman valores de QST, es necesario prestar especial atención a cómo se
estima la diferenciación genética en los caracteres cuantitativos analizados. Todo lo que no sea
varianza genética aditiva, es decir, la varianza fenotípica debida a los distintos ambientes y la
varianza debida a efectos maternos, puede ocultar la diferenciación poblacional. También es
necesario analizar un número elevado de poblaciones ya que se mejora la precisión y las
propiedades estadísticas del índice QST (Edelaar et al., 2011; Holsinger & Weir, 2009;
Leinonen et al., 2008 y 2013; Miller et al., 2008; O’Hara & Merilä, 2005). En el Apartado 4.2
se han analizado 10 y 12 poblaciones, según el ambiente, lo que podría parecer un numero
bajo. Sin embargo, en este tipo de estudios no sólo es importante el número de poblaciones

Discusión
176
utilizadas, sino también la representatividad de la variación genética de la especie. Las
poblaciones utilizadas representan la variación genética de la especie en la Península Ibérica y
por tanto son válidas para realizar la comparación QST y FST como se ha demostrado en el
estudio realizado por Méndez-Vigo y col (2012).
En el presente trabajo, de los análisis llevados a cabo con los valores de FST y de QST se
obtienen tres resultados relevantes. En primer lugar, los valores de FST estimados indican una
elevada diferenciación genética de los marcadores neutrales entre las poblaciones de
arabidopsis de la Península Ibérica. Los valores estimados de QST de la resistencia y de la
tolerancia a CMV son inferiores a los valores de FST, lo que indica que la diferenciación
genética de la resistencia y la tolerancia a CMV entre poblaciones es menor de lo esperado
debido al azar. En segundo lugar, la diferenciación genética poblacional para los marcadores
neutrales sigue un patrón de aislamiento por distancia, en cambio, este patrón no se detecta
para la resistencia y la tolerancia a CMV. Esto sugiere que otros factores evolutivos, distintos
de los que afectan a los marcadores neutrales, afectan a la resistencia y la tolerancia de las
poblaciones de arabidopsis a CMV. Y por último, las historias evolutivas de la resistencia y la
tolerancia a CMV son diferentes debido a la ausencia de correlación entre las matrices de los
valores de QST de ambos caracteres (Cdc1-CMV: r=-0.233, P=0.239; Fny-CMV: r=0.059,
P=0.751). Por tanto, la resistencia y la tolerancia a CMV están sometidas a una selección
uniforme y la evolución favorece una mayor diversidad genética de ambos caracteres dentro
que entre las poblaciones.
La explicación más obvia para la selección uniforme observada sobre la tolerancia y la
resistencia a CMV en arabidopsis sería que se debe al efecto directo de la infección viral.
Alternativamente, podría ocurrir que la expresión de la resistencia y la tolerancia estuvieran
condicionadas por los factores ambientales del lugar de procedencia de los genotipos, de modo
que la magnitud de estos dos caracteres difiriera entre genotipos según los factores
ambientales de sus lugares de procedencia, lo que conllevaría la existencia de una selección
heterogénea (Byers, 2005; Gillespie & Turelli, 1989). A este respecto, se ha demostrado que
los factores ambientales pueden modular la variación genética de arabidopsis en los caracteres
relacionados con el crecimiento vegetativo, la distribución de la biomasa o la fenología de la
Península Ibérica (Manzano-Piedras et al., 2014; Méndez-Vigo et al., 2011). Por tanto, es
posible que también la variación genética de la resistencia y la tolerancia a CMV, y en
consecuencia su selección, esté influida por los factores ambientales o ligada a otros caracteres
cuya variación esté relacionada con los factores ambientales. Cualquiera de las dos situaciones

Discusión
177
tendrían la misma consecuencia, la selección de la resistencia y la tolerancia a CMV
demostrada en el Apartado 4.2 no estaría producida por el efecto de la infección.
Para estudiar esta cuestión se han utilizado 77 individuos de arabidopsis procedentes de
distintos hábitats que representan la distribución de la especie en la Península Ibérica. Con esta
aproximación experimental queda representada la variación geográfica continua de los
factores del medio abiótico que podrían ejercer presiones de selección sobre la resistencia y la
tolerancia. Los resultados del Apartado 4.3, que además de considerar la variación ambiental
y fenotípica de los caracteres de defensa también integran la variación genética de arabidopsis,
indican que la variación genética de los caracteres relacionados con la distribución de recursos
de las plantas no infectadas, el peso de la roseta, la inflorescencia y las semillas, está
relacionada con la variación de las variables térmicas, como ya ha sido demostrado
previamente para otros caracteres de arabidopsis (Manzano-Piedras et al., 2014; Méndez-Vigo
et al., 2011). En cambio, los resultados no indican relación entre la variación de los factores
del medio abiótico y la variación genética de la resistencia y la tolerancia a diferentes
genotipos de CMV aislados de arabidopsis. Es necesario destacar que la tolerancia de
arabidopsis a CMV depende en parte, de una redistribución de recursos de la planta desde las
estructuras vegetativas a las reproductoras. Los resultados del Apartado 4.3 indican que los
caracteres de la planta asociados a tal redistribución están relacionados con la estructura
genética de la especie en la Península Ibérica. Por el contrario, la resistencia y la tolerancia a
CMV no lo están. Por tanto, de los resultados del Apartado 4.3 se derivan dos conclusiones:
1) los resultados apoyan la posibilidad de que la presión de selección a la que está sometida la
resistencia y la tolerancia de arabidopsis a CMV sea producida por la infección de CMV. 2) Se
confirman las conclusiones de los experimentos descritos en el Apartado 4.2, y se evidencia
que la tolerancia pueda producirse por otros mecanismos además de la redistribución de
recursos.
Estos resultados son coherentes con los obtenidos en otros estudios realizados en
sistemas silvestres planta-hongo, en los que la selección de la resistencia se debe a la infección
del patógeno (Salvaudon et al., 2008). Uno de estos estudios se realizó en una de las
interacciones planta-patógeno mejor caracterizadas, formada por la especie silvestre
australiana de lino, Linum marginale, y el hongo del lino, Melampsora lini, en poblaciones del
sur de Australia. Esta especie de lino se encuentra en diferentes hábitats, desde vegetación
costera hasta pantanos e incluso vegetación subalpina. M. lini disminuye significativamente el
ciclo de vital del huésped. En dichas poblaciones, se ha demostrado variación genética de la

Discusión
178
resistencia de lino a M. lini y de la virulencia de M. lini tanto espacial (dentro y entre
poblaciones) como temporal (Thrall et al., 2001). Además, en la misma área geográfica, las
poblaciones de M. lini se diferencian en función de su virulencia: algunas poblaciones del
huésped son susceptibles a la infección por la mayoría de los genotipos del patógeno, otras
poblaciones son resistentes sólo a algunos genotipos del patógeno, y por último, hay
poblaciones resistentes a los genotipos del hongo que no son del mismo área del huésped, lo
que es evidencia de adaptación local. Por tanto, el patógeno debe estar ejerciendo una presión
de selección fuerte que provocaría una mejora de la resistencia del huésped (Thrall et al.,
2002) ya que el patógeno está adaptado a su huésped localmente. La dinámica de la resistencia
de L. marginale a M. lini dentro de las poblaciones parece estar influida por otros factores
además de la presión de selección ejercida por el patógeno. En teoría, una epidemia de un
patógeno debería seleccionar aquellos huéspedes más resistentes a los patógenos locales
responsables de la epidemia. Sin embargo, una de las epidemias de M. lini que causó la
reducción del 80% de la población de lino, produjo una reducción de la frecuencia de
huéspedes resistentes aumentando la frecuencia de huéspedes susceptibles (Burdon &
Thompson, 1995). Por tanto, la resistencia de L. marginale a M. lini puede estar vinculada a
genes sometidos a una presión de selección mayor que la ejercida por el patógeno o a efectos
demográficos tales como la estructura de edad.
Otro de los sistemas bien caracterizados en el que se ha demostrado evidencia de
coevolución planta-hongo es el compuesto por Plantago lanceolata-Podospera planaginis
sphaera. En las poblaciones P. lanceolata localizadas en el Archipiélago Aland en Finlandia,
se ha demostrado variación genética de la resistencia a Podospera planaginis sphaera dentro y
entre poblaciones del huésped. Dicha variación genética no mostró un patrón de aislamiento
por distancia ya que la similitud de los fenotipos de resistencia no aumentó con la proximidad
de las poblaciones del huésped (Laine, 2004 y 2006). Además, se ha demostrado adaptación
local del patógeno, ya que las poblaciones de éstos muestran una infectividad mayor en las
plantas de su población local que los de otras poblaciones cercanas (Laine, 2005). Por tanto, al
igual que en la interacción L. marginale–M. lini, el patógeno está ejerciendo una presión de
selección sobre la resistencia del huésped. En conjunto, los resultados obtenidos en el análisis
de la resistencia y de la tolerancia de las poblaciones de arabidopsis a CMV son coherentes
con los obtenidos en otros patosistemas planta-hongo y son evidencia de coevolución planta-
virus.

Discusión
179
5.2.3. Respuesta evolutiva de CMV a las defesas de arabidopsis
Hasta este punto, todos los resultados discutidos demuestran la evolución de las
defensas de arabidopsis en respuesta a la infección por CMV. Pero un proceso coevolutivo
requiere también de una respuesta evolutiva del patógeno para contrarrestar las defensas del
huésped. Una consecuencia de dicho proceso puede ser la adaptación de un patógeno a su
huésped. Tal adaptación puede ser producida por la variación de los genes de patogenicidad y
de virulencia en respuesta a la selección ejercida por las defensas del huésped (Woolhouse et
al., 2002). La evidencia de adaptación a huésped puede derivar de diferencias en la eficacia
biológica del patógeno en diferentes huéspedes, cuando la eficacia biológica del patógeno es
mayor en su huésped de origen respecto a otros huéspedes. CMV es un virus generalista que
por definición, es capaz de infectar a distintos huéspedes. Por tanto, se esperaría que los
mecanismos de patogenicidad y de virulencia evolucionarán en respuesta a los mecanismos de
defensa de sus múltiples huéspedes, de modo que la selección actuará manteniendo un valor
medio de eficacia biológica de CMV en todos ellos. Sin embargo, existe evidencia de
adaptación a huésped de distintos aislados de CMV. Por ejemplo, en el estudio realizado por
Malpica y col (2006) sobre la incidencia de cinco virus generalistas (incluido CMV) en 21
especies silvestres de plantas, los análisis indicaron la preferencia de cada virus por algunos de
los huéspedes analizados. También, existen trabajos en los que algunos aislados de CMV,
como los de azucena y soja, tiene un número inferior de especies de huéspedes en
comparación a otros aislados (Hong et al., 2007; Lee et al., 2007; Masuta et al., 2002; Ryu et
al., 2002). Luego existen argumentos que son evidencia de que algunos aislados de CMV
están adaptados a sus huéspedes, por lo que es posible que los aislados de CMV procedentes
de arabidopsis estuvieran adaptados a arabidopsis como consecuencia de un proceso
coevolutivo. Si esto fuera cierto, un cambio de huésped de estos aislados reduciría sus
eficacias biológicas. Los resultados del Apartado 4.4 abordan esta cuestión, y para ello se
utilizaron 8 aislados de CMV (4 procedentes de arabidopsis y 4 de especies de la familia de las
cucurbitáceas) los cuales se inocularon en 21 individuos de diferentes poblaciones de
arabidopsis de la Península Ibérica y en la cucurbitácea melón. Todos los aislados infectaron a
todos los huéspedes, por lo que no existen diferencias en sus infectividades. En cambio,
existen diferencias en la multiplicación viral. Los resultados muestran que melón es mejor
huésped para la multiplicación de CMV. Además, los aislados de arabidopsis se multiplican
más que los de cucurbitáceas en arabidopsis, y a la inversa en melón. Estos resultados no

Discusión
180
discrepan con los obtenidos en otros virus generalistas (Cronin et al., 2010; Moreno-Pérez et
al., 2014) y también en trabajos realizados con diferentes aislados de CMV (Betancourt et al.,
2011; Escriu et al., 2003; Sacristán et al., 2005). Por ejemplo, Agudelo y col (2008) analizaron
experimentalmente la adaptación del aislado 7DA de TEV procedente de N.tabacum var
Xanthi a C. Annuum var Marconi. El aislado 7DA-TEV evolucionado tras 15 pases seriados
en C. Annuum var Marconi, fue más virulento y su multiplicación fue mayor en C. Annuum
var Marconi que en su huésped original, N.tabacum var Xanthi, en el cual fue además menos
virulento y su multiplicación viral fue menor respecto al aislado no evolucionado 7DA-TEV.
Los resultados del Apartado 4.4 indican que los aislados de arabidopsis son más virulentos
que los aislados de cucurbitáceas tanto en arabidopsis como en melón, por lo que la adaptación
a huésped que resulta en una mayor multiplicación viral, no se asocia con cambios en la
virulencia de los aislados de CMV. La adaptación a huésped puede tener asociados
compromisos de adaptación aunque no siempre (Betancourt et al., 2011; Escriu et al., 2003;
García-Arenal & Fraile et al., 2013). De hecho, los compromisos de adaptación pueden existir
entre algunos huéspedes y sólo para algunos de los componentes de la eficacia biológica viral.
Por ejemplo, en el estudio realizado por Sacristán y col (2005) se compararon diferentes
aislados de CMV procedentes de diferentes huéspedes en tres familias botánicas y se
encontraron evidencias de compromisos de adaptación en la infectividad entre algunos
huéspedes pero no en la multiplicación. Por ello, es posible que la adaptación observada para
la multiplicación viral y no para la virulencia se deba a la existencia de compromisos con otras
especies de huéspedes no consideradas, pero no rechazan la hipótesis de adaptación a huésped
de los aislados de CMV.
Otra evidencia de adaptación a huésped puede ser la estructuración genética del
patógeno según el huésped (García-Arenal et al., 2001; Jeger et al 2006; Moury et al., 2006;
Oshima et al., 2002), o según las poblaciones del huésped (Kawecki & Ebert, 2004). En este
caso, diferentes aislados de CMV con diferentes huéspedes se estructuran genéticamente
según el huésped de origen (Berniak et al., 2010; Bonnet et al., 2005; Chen et al., 2001; Hong
et al., 2007), como ocurre con los aislados de CMV de azucena, los cuales forman un grupo
monofilético (Masuta et al., 2002). Los resultados de Apartado 4.4 muestran que todos los
aislados de CMV, independientemente del huésped de origen, son del Subgrupo IA como la
mayoría de los aislados descritos en la zona (Fraile et al., 1997). A pesar del elevado
porcentaje de homología entre las secuencias de nucleótidos de los genomas de los aislados,
los resultados indican que los aislados se estructuran genéticamente según su huésped de

Discusión
181
origen. La diversidad genética media del conjunto de aislados procedentes de arabidopsis es
menor que la de los procedentes de cucurbitáceas y entre un 13% y un 30% de la diversidad
genética se explica por el huésped de origen de los aislados. Además, aislados de CMV de un
mismo huésped están agrupados en un mismo clado filogenético que a su vez se asocia
significativamente con el factor huésped de origen. Por tanto, existe estructuración genética de
los aislados de CMV según su huésped de origen, lo que es evidencia de adaptación a huésped.
En definitiva, los aislados de CMV de arabidopsis están adaptados a su huésped de
origen ya que presentan una multiplicación mayor que otros aislados de CMV procedentes de
cucurbitáceas, además de una estructuración genética según el huésped de procedencia.
También, los resultados del Apartado 4.4 indican variación de la multiplicación y del efecto
de diferentes aislados de CMV de arabidopsis sobre el peso de las semillas y que ambos
caracteres dependen de la interacción entre los diferentes genotipos de arabidopsis y CMV. En
conjunto estos resultados evidencian la variación genética del patógeno y la interacción entre
los genotipos del huésped de patógenos, condiciones necesarias e indicadoras de la existencia
de coevolución arabidopsis-CMV. Por tanto, la adaptación a huésped demostrada de los
aislados de CMV pueden ser evidencia de que la selección a la que está sometida la resistencia
y la tolerancia de arabidopsis a CMV sea producida por la infección de CMV.

Discusión
182

Conclusiones
183
6. CONCLUSIONES

Conclusiones
184

Conclusiones
185
En esta tesis se han analizado diferentes aspectos de la interacción entre Arabidopsis
thaliana y el virus del mosaico del pepino (CMV) cuyo conocimiento es necesario para
demostrar que se de coevolución planta-virus. Los resultados obtenidos arrojan las siguientes
conclusiones:
1. El virus con mayor incidencia en las poblaciones silvestres de arabidopsis de la Meseta
Central de la Península Ibérica es CMV, y su máxima incidencia ocurre cuando las
plantas están en las primeras fases de su ciclo biológico. Estos hechos son argumentos
para suponer un efecto de la infección por CMV sobre la dinámica poblacional de
arabidopsis, que no resulta evidente del análisis de su demografía.
2. La tolerancia a CMV detectada en individuos de diferentes poblaciones de arabidopsis
de la Península Ibérica se de debe en parte, a cambios en los caracteres de la historia
vital tras la infección, que dan lugar a la redistribución de recursos de crecimiento
vegetativo a reproducción y al retraso de la reproducción.
3. La resistencia y la tolerancia a CMV son caracteres cuantitativos con variación
genética dentro y entre poblaciones. La magnitud de la resistencia y la tolerancia de
arabidopsis a CMV depende de la interacción entre los genotipos de arabidopsis y
CMV.
4. No se ha hallado evidencia de que la resistencia y la tolerancia de arabidopsis a CMV
esté asociada a costes en ausencia de infección. Tampoco se ha hallado evidencia de
compromisos entre ambos mecanismos de defensa.
5. La historias evolutivas de la resistencia y la tolerancia a CMV de arabidopsis son
diferentes entre si y también de la historia evolutiva de marcadores neutrales. Estos
resultados sugieren que los factores que determinan la evolución de la resistencia y la
tolerancia de arabidopsis a CMV son diferentes de los que determinan la evolución de
los marcadores neutrales, y que la resistencia y la tolerancia están bajo selección. La
variación genética de la resistencia y la tolerancia a CMV de arabidopsis es mayor
dentro que entre poblaciones, sugiriendo que ambos caracteres están sometidos a
selección uniforme.

Conclusiones
186
6. La variación genética de la resistencia y la tolerancia de arabidopsis a CMV no está
relacionada con la variación de un amplio conjunto de factores del medio abiótico, por
lo que es posible que la infección por CMV sea el factor que de la selección de ambos
caracteres.
7. La comparación de aislados de CMV procedentes de arabidopsis y de cucurbitáceas
muestra adaptación en este virus generalista. La adaptación a CMV a arabidopsis es
compatible de nuevo, con una hipótesis de coevolución planta-virus.
8. Todos los resultados de esta tesis apoyan la hipótesis de que la infección por CMV sea
un factor en la dinámica de las poblaciones de arabidopsis de la Península Ibérica, y
aportan evidencia a favor de que se den dinámicas coevolutivas entre las plantas y los
virus.

7. BIBLIOGRAFÍA

Bibliografía
188

Bibliografía
189
4Peaks Griekspoor, A. & Groothuis, T. http://nucleobytes.com/4peaks/index.htmlpor A., mekentosj.com.
Abbott, R. J. & Gomes, M. F. (1989) Population genetic structure and outcrossing rate of Arabidopsis thaliana (L.) Heynh. Heredity. 62: 411–418.
Agnew, P., Koella, J. C. & Michalakis, Y. (2000) Host life history responses to parasitism. Microbes and Infection, 2: 891–896.
Agrawal, A. (2000) Specificity of induced resistance in wild radish: causes and consequences for two specialist and two generalist caterpillars. Oikos. 89: 493-500.
Agrawal, A. & Lively C. M. (2002) Infection genetics: gene-for-gene versus matching-alleles models and all points in between. Evolutionary Ecology Research. 4: 79-90.
Agrios, G. N. (2005) Plant pathology. 5ª Ed. Elsevier Academic Press. Amsterdam. Agudelo-Romero, P., de la Iglesia, F. & Elena, S. E. (2008) The pleiotropic cost of host-
specialization in Tobacco etch potyvirus. Infection Genetics and Evolution. 8: 806–814. Al-Shehbaza I. A. & O’Kane, S. L. Jr. (2002) Taxonomy and phylogeny of Arabidopsis
(Brassicaceae) The Arabidopsis Book. American Society of Plant Biologists. Rockville. Alexander, H., Mauck, K., Whitfield, A., Garrett, K. & Malmstrom, C. (2014) Plant-virus
interactions and the agro-ecological interface. European journal of plant pathology, 138: 529-547.
Alexander, H.M. (1989) An experimental field study of anther- smut disease of Silene alba caused by Ustilago violacea: geno- typic variation and disease incidence. Evolution 43: 835–847.
Ali, A. & Kobayashi, M. (2010) Seed transmission of Cucumber mosaic virus in pepper. Journal of Virology Methods. 163: 234–237.
Ali, A., Li, H., Schneider, W. L., Sherman, D. J., Gray, S., Smith, D. & Roossinck, M. J. (2006) Analysis of genetic bottlenecks during horizontal transmission of Cucumber mosaic virus. Journal of Virology. 80: 8345–8350.
Alonso-Blanco C, Bentsink L, Hanhart CJ, Blankestijn-de Vries H & Koornneef M. (2003) Analysis of natural allelic variation at seed dormancy loci of Arabidopsis thaliana. Genetics. 164: 711-29.
Alonso-Blanco, C. & Koornneef, M. (2000) Naturally occurring variation in Arabidopsis: an underexploited resource for plant genetics. Trends in Plant Science. 5: 22-29
Alonso-Prados, J. L., Luis-Arteaga, M. & Alvarez, J. M. (2003) Epidemics of aphid-transmitted viruses in melon crops in Spain. European Journal of Plant Pathology. 109: 129-138.
Anderson, R. M. & May, R. M. (1978) Regulation and stablity of host- parasite population interactions; regulatory processes. Journal of Animal Ecology. 47: 219-247.
Anderson, R. M. & May, R. M. (1982) Coevolution of hosts and parasites. Parasitology 85: 411-426. Anderson, P. K., Cunningham, A. A., Patel, N. G., Morales, F. J., Epstein, P. R. & Daszak, P.
(2004) Emerging infectious diseases of plants: pathogen pollution, climate change and agrotechnology drivers. Trends in Ecology & Evolution. 19: 535-544.
Antonovics, J., Thrall, P., Jarosz, A. & Stratton, D. (1994) Ecological genetics of metapopulations: the Silene–Ustilago plant–pathogen system. Ecological genetics. Princeton: Princeton University Press.
Archibold, O. W. (1995) Ecology of world vegetation. London: Chapman and Hall. Arnott, S. A., Barber, I. & Huntingford, F. A. (2000) Parasite-associated growth enhancement in a
fish-cestode system. Proceeding of the Royal Society of London Serie B. 267: 657-663.

Bibliografía
190
Ayme, V., Souche, S., Caranta, C., Jacquemond, M., Chadoeuf, J., Palloix A & Benoît, M. (2006) Different mutations in the genome-linked protein VPg of potato virus y confer virulence on the pvr23 resistance in pepper. Molecular Plant Microbe Interactions. 19: 557–563.
Baltrus, D. A., Nishimura, M. T., Dougherty, K. M., Biswas, S., Mukhtar, M. S., Vicente, J., Holub, E. B. & Dangl, J. L. (2012) The molecular basis of host specialization in bean pathovars of Pseudomonas syringae. Molecular Plant Microbe Interactions., 25: 877-888.
Baltrus, D. A., Nishimura, M. T., Romanchuk, A., Chang, J. H., Mukhtar, M. S., Cherkis, K., Roach, J., Grant, S. R., Jones, C. D. & Dangl, J. L. (2011) Dynamic evolution of pathogenicity revealed by sequencing and comparative genomics of 19 Pseudomonas syringae isolates. PLoS Pathogen, 7:e1002132.
Banik, M. T. & Zitter, T. A. (1990) Determination of Cucumber mosaic virus titer in muskmelon by enzyme-linked immunosorbent assay and correlation with aphid transmission. Plant Disease. 74: 857-859.
Bao,Y., Flasinski, S. & Roossinck, M. (1999) The 1a protein of Cucumber mosaic virus contains an adenosylmethionine-binding domain. En: “Abstract of the XIth International Congress of Virology”, Sydney.
Barrett, L. G., Thrall, P. H., Burdon, J. J. & Linde, C. C. (2008) Life history determines genetic structure and evolutionary potential of host–parasite interactions. Trends in Ecology & Evolution. 23: 678–685.
Baskin, J. M. & Baskin, C. C. (1983) Seasonal changes in the germination responses of buried seeds of Arabidopsis thaliana and ecological interpretation. Botanical Gazette. 144: 540–543.
Baucom, R. S. & de Roode J. C. (2011) Ecological immunology and tolerance in plants and animals. Functional Ecology. 25: 18– 28.
Beale, C. M., Lennon, J. J., Yearsley, J. M., Brewer, M. J. & Elston, D. A. (2010) Regression analysis of spatial data. Ecology Letters, 13(2), 246–264.
Beck, J.B., Schmuths, H. & Schaal, B. A. (2008) Native range genetic variation in Arabidopsis thaliana is strongly geographically structured and reflects Pleistocene glacial dynamics. Molecular Ecology. 17: 902–915.
Becklin, K. M. & Kirkpatrick, E. (2006) Compensation through rosette formation: the response of scarlet gilia (Ipomopsis aggregata: Polemoniaceae) to mammalian herbivory. Canadian Journal of Botany. 84: 1298-1303.
Ben-Khalifa, M., Simon, V., Fakhfakh, H. & Moury, B. (2012) Tunisian Potato virus Y isolates with unnecessary pathogenicity towards pepper: support for the matching allele model in eIF4E resistance–potyvirus interactions. Plant Pathology. 61: 441-447.
Bentsink, L., Jowett, J., Hanhart, C. J. & Koornneef M. (2006) Cloning of DOG1, a quantitative trait locus controlling seed dormancy in Arabidopsis. Proceedings of the National Academy of Sciences of USA. 103: 17042-7.
Bergelson, J. & Purrington, C. B. (1996) Surveying patterns in the cost of resistance in plants. The American Naturalist. 148: 536 –558.
Bergelson, J., Dwyer, G. & Emerson, J. J. (2001) Models and data on plant-enemy coevolution. Annual Review of Genetics. 35: 469–499.
Bergelson, J., Stahl, E., Dudek, S. & Kreitman, M. (1998) Genetic Variation Within and Among Populations of Arabidopsis thaliana. Genetics. 148: 1311-1323.
Berniak, H., Kamin ska, M. & Malinowski, T. (2010) Cucumber mosaic virus groups IA and II are represented among isolates from naturally infected lilies. European Journal of Plant Pathology. 127: 305–309.

Bibliografía
191
Berry, J. & Björkman, O. (1980) Photosynthetic response and adaptation to temperature in higher plants. Annual Review of Plant Physiology. 31: 491–543.
Berzal-Herranz, A., de la Cruz, A., Tenllado, F., Díaz-Ruíz, J. R., López, L., Sanz, A. I., Vaquero, C., Serra, M. T. & García-Luque, I. (1995) The Capsicum L3 gene-mediated resistance against the tobamoviruses is elicited by the coat protein. Virology. 209: 498-505.
Betancourt, M., Fereres, A., Fraile, A. & García-Arenal, F. (2008) Estimation of the effective number of founders that initiate an infection after aphid transmission of a multipartite plant virus. Journal of Virology. 82: 12416–12421.
Betancourt, M., Escriu, F., Fraile, A. & García-Arenal, F. (2013) Virulence evolution of a generalist plant virus in a heterogeneous host system. Evolutionary Applications. 6: 875-890.
Betancourt, M., Fraile, A. & García-Arenal, F. (2011) Cucumber mosaic virus satellite RNAs that induce similar symptoms in melon plants show large differences in fitness. Journal of General Virology. 92: 1930-1938.
Bevan, J. R., Clarke, D. D. & Crute, I. R. (1993b) Resistance to Erysiphe fischeri in two populations of Senecio vulgaris. Plant Pathology. 42: 636–46.
Bevan, J. R., Crute, I. R. & Clarke, D. D. (1993a) Variation for virulence in Erysiphe fischeri from Senecio vulgaris. Plant Pathology. 42: 622–35.
Biere, A. & Antonovics, J. (1996) Sex specific costs of resistance to the fungal pathogen Ustilago violacea (Microbotryum violaceum) in Silene alba. Evolution. 50: 1098–1110.
Blanchard, C. L., Boyce, P. M. & Anderson, B. J. (1996) Cucumber mosaic virus RNA 5 is a mixed population derived from the conserved 3 -́terminal regions of genomic RNAs 2 and 3. Virology. 217: 598-601.
Blanchard, C. L., Higgins, T. J. & Anderson, T. J. (1997) RNAs 4A and 5 are present in tomato aspermy virus and both subgroups of Cucumber mosaic virus. Archives of Virology. 142: 1273-1283.
Boccard, F. & Baulcombe, D. (1993) Mutational analysis of cis-acting sequences and gene function in RNA 3 of Cucumber mosaic virus. Virology. 193: 563-578.
Bonnet, J., Fraile, A., Sacristán, S., Malpica, J. M. & García-Arenal, F. (2005) Role of recombination in the evolution of natural populations of Cucumber mosaic virus, a tripartite RNA plant virus. Virology. 332: 359–368.
Bonnin, I., J. M. Prosperi & I. Olivieri. (1996) Genetic markers and quantitative genetic variation in Medicago truncatula (Leguminosae): A comparative analysis of population structure. Genetics. 143: 1795-1805.
Borevitz, J. O., Hazen, S. P., Michael, T. P., Morris, G. P., Baxter, I. R., Hu, T. T., Chen, H., Werner, J. D., Nordborg, M., Salt, D. E., Kay, S. A., Chory, J., Weigel, D., Jones, J. D. G. & Ecker, J. R. (2007) Genome-wide patterns of single-feature polymorphism in Arabidopsis thaliana. Proceedings of the National Academy of Sciences of USA. 104: 12057-12062.7.
Brennan, A. C., Méndez-Vigo, B., Haddioui, A., Martínez-Zapater, J. M., Picó, F. X. & Alonso-Blanco, C. (2014) The genetic structure of Arabidopsis thaliana in the south-western Mediterranean range reveals a shared history between North Africa and southern Europe. BMC Plant Biology. 14:17.
Brown, J. K. M. (2003) A cost of disease resistance: paradigm or peculiarity? Trends in Genetics. 19: 667–671.
Brown, J. K. M. & Rant, J. C. (2013) Fitness costs and trade-offs of disease resistance and their consequences for breeding arable crops. Plant Pathology. 62: 83–95.

Bibliografía
192
Brown, J. K. M. & Tellier, A. (2011) Plant-Parasite Coevolution: Bridging the Gap between Genetics and Ecology. Annual Review of Phytopathology. 49: 345–367.
Bruenn, J.A. (1991) Relationships among the positive strand and double-strand RNA viruses as viewed through their RNA-dependent RNA polymerases. Nucleic Acids Research. 19: 217-226.
Boyes, D. C., Zayed, A. M., Ascenzi, R., McCaskill, A. J., Hoffman, N.E., Davis, K. R. & Görlach, J. (2001) Growth stage-based phenotypic analysis of Arabidopsis: A model for high throughput functional genomics in plants. Plant Cell. 13: 1499-1510.
Buell, C. R. & Somerville, S. C. (1997) Use of Arabidopsis recombinant inbred lines reveals amonogenic and a novel digenic resistance mechanism to Xanthomonas campestris pv campestris. The Plant Journal. 12: 21-29.
Burdon, J. J. & Thrall, P. H. (2003) The Fitness Costs to Plants of Resistance to Pathogens. Genome Biology. 4: 227.
Burdon, J. J. & Chilvers, G. A. (1982) Host density as a factor in plant disease ecology. Annual Review of Phytopathology. 20: 143–166.
Burdon, J. J. & Jarosz, A. M. (1991) Host–pathogen interactions in natural populations of Linum marginale and Melampsora lini. I. Patterns of resistance and racial variation in a large host population. Evolution 45: 205–217.
Burdon, J. J. & Thrall, P. H. (2009) Coevolution of Plants and Their Pathogens in Natural Habitats. Science, 324: 755–756.
Burdon, J. J. (1987) Phenotypic and genetic patterns of resistance to the pathogen Phakopsora pachyrhizi in populations of Glycine canescens. Oecologia 73: 257–267.
Byers, D. L. (2005) Evolution in heterogeneous environments and the potential of maintenance of genetic variation in traits of adaptive significance. Genetica, 123, 107–124.
Callahan, H.S. & Pigliucci, M. (2002) Shade-induced plasticity and its ecologi- cal significance in wild populations of Arabidopsis thaliana. Ecology. 83: 1965–1980.
Cao, J., Schneeberger, K., Ossowski, S., Günther, T., Bender, S., Fitz, J., Koenig, D., Lanz, C., Stegle, O., Lippert, C., Wang, X., Ott, F., Müller, J., Alonso-Blanco, C., Borgwardt, K., Schmid, K.J. & Weigel, D. (2011) Whole-genome sequencing of multiple Arabidopsis thaliana populations. Nature Genetics. 43: 956–963.
Carr, D.E. & Eubanks, M.D. (2002) Inbreeding alters resistance to insect herbivory and host plant quality in Mimulus guttatus (Scrophulariaceae) Evolution 17: 85–103.
Carr, D. E., Murphy, J. F. & Eubanks, M. D. (2006) Genetic variation and covariation for resistance and tolerance to Cucumber mosaic virus in Mimulus guttatus (Phrymaceae): a test for costs and constraints. Heredity. 96: 29-38.
Cavieres, L. A. & Arroyo, M. T. K. (2001) Persistent soil seed banks in Phacelia secunda (Hydrophyllaceae): experimental detection of variation along an altitudinal gradient in the Andes of Central Chile (33°S) Journal of Ecology. 89: 31–39.
Charlesworth, D. (2006) Balancing selection and its effects on sequences in nearby genome regions. PLoS Genetics. 2:e64.
Chen, B. & Francki, R. I. B. (1990) Cucumovirus transmission by the aphid Myzus persicae is determined solely by the viral coat protein. Journal of General Virology. 71: 939-944.
Chen, Y. K., Derks, A. F. L. M., Langeveld, S., Goldbach, R. & Prins, M. (2001) High sequence conservation among Cucumber mosaic virus isolates from lily. Archives of Virology. 146: 1631–1636.

Bibliografía
193
Choi, S. K., Palukaitis, P., Min, B. E., Lee, M. Y., Choi, J. K. & Ryu, K. H. (2005) Cucumber mosaic virus 2a polymerase and 3a movement proteins independently affect both virus movement and the timing of symptom development in zucchini squash. Journal of General Virology. 86: 1213–1222.
Christie, P., Richner, H. & Oppliger, A. (1996) Begging, food provisioning, and nestling competition in great tit broods infested with ectoparasites. Behavioral Ecology. 7: 127-131
Clair, D.A.St. (2010) Quantitative disease resistance and quantitative resistance loci in breeding. Annual Review of Phytopathology. 48: 247–268.
Clarke, D. D. (1986) Tolerance of parasites and disease in plants and its significance in host-parasite interactions. Advance in Plant Pathology. 5: 161-198.
Clay, K. & Kove, P. X. (1996) The Red Queen hypothesis and plant/pathogen intereactions. Annual Reviews of Phytopathology. 34: 29-50.
Clay, K. & Putten, W. H. V. (1999) in Life History Evolution in Plants. Vuorisalo, T. & Mutikainen, P. Kluwer, Dordrecht, The Netherlands.
Cliff, A.D. & J.K. Ord. (1981) Spatial processes: models and applications. Taylor & Francis. Collins, N., Drake, J., Ayliffe, M., Sun, Q., Ellis, J., Hulbert, S. & Pryor, T. (1999) Molecular
characterization of the maize Rp1-D rust resistance haplotype and its mutants. Plant Cell. 11: 1365–1376.
Cookson, S.J. & Granier, C. (2006) A dynamic analysis of the shade-induced plasticity in Arabidopsis thaliana rosette leaf development reveals new components shade-adaptive response. Annals of Botany. 97: 443–452.
Cooper, I. & Jones, R. A. C. (2006) Wild Plants and Viruses: Under‐Investigated Ecosystems. In Advances in Virus Research (Vol. 67) Elsevier.
Cosson, P., Schurdi-Levraud, V., Le, Q. H., Sicard, O., Caballero, M., Roux, F., Le Gall, O., Candresse, T. & Revers, F. (2012) The RTM Resistance to Potyviruses in Arabidopsis thaliana: Natural Variation of the RTM Genes and Evidence for the Implication of Additional Genes. PLoS ONE, 7(6), e39169.
Cressie, N. (1993) Statistics for spatial data. Wiley Interscience. Cronin, J. P., Welsh, M. E., Dekkers, M. G., Abercrombie, S. T. & Mitchell, C. E. (2010) Host
physiological phenotype explains pathogen reservoir potential. Ecology Letters. 13: 1221–32. Cronin, J. P., Rúa, M. A. & Mitchell, C. E. (2014) Why Is Living Fast Dangerous? Disentangling the
Roles of Resistance and Tolerance of Disease. The American Naturalist. 184: 172–187. Cummins, R. P. & Miller, G. (2002) Altitudinal gradients in seed dynamics of Calluna vulgaris in
eastern Scotland. Journal of Vegetation Science. 13: 859–866. D’Arcy, C. J., Eastburn, D. M. & Schumann, G. L. (2001) Illustrated glossary of plant pathology.
Plant Health Instructor. Darriba, D., Taboada, G. L, Doallo, R. & Posada, D. (2012) jModelTest 2: more models, new
heuristics and parallel computing. Nature Methods. 9: 772. Davies, N. B. & Brooke, M. D. L. (1989) An experimental study of co-evolution between the cuckoo,
Cuculus canorus, and its hosts. II. Host egg markings, chick discrimination and general discussion. The Journal of Animal Ecology. 58: 225–236.
Dawkins, R. & Krebs, J. R. (1979) Arms races between and within species. Proceedings of the Royal Society London Biological Society. 205: 489-511.
de Meaux, J. & Mitchell-Olds, T. (2003) Evolution of Plant Resistance at the Molecular Level: Ecological Context of Species Interactions. Heredity: 345-52.

Bibliografía
194
de Nooij, M.P. & van Damme, J.M.M. (1988a) Variation in host susceptibility among and within populations of Plantago lanceolata L. infected by the fungus Phomopsis subordinaria (Desm.) Trav. Oecologia 75: 535–538.
de Nooij, M.P. & van Damme, J.M.M. (1988b) Variation in pathogenicity among and within populations of the fungus Phomopsis subordinaria infecting Plantago lanceolata. Evolution. 42: 1166–1171.
de Ronde, D., Butterbach, P. & Kormelink, R. (2014) Dominant resistance against plant viruses. Frontiers in Plant Science. 5: 307.
de Roode, J. C., Yates, A. J. &Altizer, S. (2008) Virulence-transmission trade-offs and population divergence in virulence in a naturally occurring butterfly parasite. Proceedings of the National Academy of Sciences of the United States of America. 105: 7489-7494.
de Wispelaere, M. & Rao, A. L. N. (2009) Production of Cucumber mosaic virus RNA5 and its role in RNA recombination. Virology. 384: 179–191.
Decroocq, V., Salvador, B., Sicard, O., Glasa, M., Cosson, P., Svanella-Dumas, L., Revers, F., Garcia, J.A. & Candresse, T. (2009) The determinant of potyvirus ability to overcome the RTM resistance of Arabidopsis thaliana maps to the N-terminal region of the coat protein. Molecular Plant–Microbe Interaction. 22: 1302–1311.
Decroocq, V., Sicard, O., Alamillo, J. M., Lansac, M., Eyquard, J. P., García, J., Candresse, T., Le Gall, O. & Revers, F. (2006) Multiple resistance traits control Plum pox virus infection in Arabidopsis thaliana. Molecular Plant–Microbe Interaction. 19: 541–549.
Delseny, M. & Hull, R. Isolation and characterization of faithful and altered clones of the genomes of cauliflower mosaic virus isolates Cabb B-JI, CM4-184, and Bari I. Plasmid. 1983: 31–41.
Desbiez, C., Gal-On, A., Girard, M., Wipf-Scheibel, C. & Lecoq, H. (2003) Increase in Zucchini yellow mosaic virus symptom severity in tolerant zucchini cultivars is related to a point mutation in P3 protein and is associated with a loss of relative fitness on susceptible plants. Phytopathology 93: 1478–1484.
Desbiez, C., Wipf-Scheibel, C. & Lecoq, H. (2002) Biological and serological variability, evolution and molecular epidemiology of Zucchini yellow mosaic virus (ZYMV, Potyvirus) with special reference to Caribbean islands. Virus Research. 85: 5–16.
Díaz-Pendón, J. A., Li, F., Li, W.-X. & Ding, S.-W. (2007) Suppression of antiviral silencing by Cucumber mosaic virus 2b protein in Arabidopsis is associated with drastically reduced accumulation of three classes of small interfering RNAs. Plant Cell. 19: 2053–2063.
Ding, S. –W., Anderson, B. J., Haase, H. R. & Symons, R. H. (1994) New overlapping gene encoded by the Cucumber mosaic virus genome. Virology. 198: 593-601.
Ding, S. –W., Li, W. –X. & Symons, R. H. (1995) A novel naturally occurring hybrid gene encoded by a plant RNA virus facilitates long distance virus movement. The EMBO Journal. 14: 5762-5772.
Dinoor, A. (1970) Sources of oat crown rust resistance in hexaploid and tetraploid wild oats in Israel. Canadian Journal of Botany. 48: 153–61.
Donohue, K. (2002) Germination timing influences natural selection on life-history characters in Arabidopsis thaliana. Ecology. 83: 1006-1016.
Dobson, A., Lafferty, K.D., Kuris, A.M., Hechinger, R.F. & Jetz, W. (2008) Homage to Linnaeus: How many parasites? How many hosts? Proceedings of the National Academy of Sciences of the United States of America. 105: 11482-11489.
Doolittle, S. P. (1916) A new infectious mosaic disease of cucumber. Phytopathology. 6: 145-147.

Bibliografía
195
Douine, L., Quiot, J. B., Marchoux G. & Archange, P. (1979) Index of plants susceptible to Cucumber mosaic virus (CMV) Annales de phytopathologie. 11: 439-475.
Doumayrou, J., Leblaye, S., Froissart, R. & Michalakis, Y. (2013) Reduction of leaf area and symptom severity as proxies of disease-induced plant mortality: The example of the Cauliflower mosaic virus infecting two Brassicaceae hosts. Virus Research. 176: 91–100.
Drummond, A. J. & Rambaut, A. (2007) BEAST: Bayesian evolutionary analysis by sampling trees. BMC evolutionary biology. 7:214.
Dutilleul, P., Clifford, P., Richardson, S. & Hemon, D. (1993) Modifying the t test for assessing the correlation between two spatial processes. Biometrics. 49: 305–314.
Ebert D. (1998) Experimental evolution of parasites. Science. 282: 1432–5. Ebert, D., Zschokke-Rohringer, C.D. & Carius, H.J. (1998) Within- and between-population
variation for resistance of Daphnia magna to the bacterial endoparasite Pasteuria ramosa. Philosophical Transactions of the Royal Society of London. 265: 2127–2134.
Edelaar, P., Burraco, P. & Gomez-Mestre, I. (2011) Comparisons between QST and FST—how wrong have we been? Molecular Ecology. 20: 4830–4839.
Edwardson J. R. & Christie, R. G. (1991) CRC Handbook of viruses infecting legumes, Boca Raton, CRC Press.
Eiras, M., Boari, A. J., Colariccio, A., Chaves, A. L. R., Briones, M. R. S., Figueira, A. R. & Harakava, R. (2004) Characterization of isolates of the cucumovirus Cucumber mosaic virus present in Brazil. The Journal of Plant Pathology. 86: 61–69.
Elena, S. F. & Sanjuán, R. (2003) Climb every mountain? Science. 302: 2074-2075. Elena, S. F., Carrasco, P., Daròs, J. A. & Sanjuán, R. (2006) Mechanisms of genetic robust- ness in
RNA viruses. EMBO Reports. 7: 168–173. Elena, S. F., Fraile, A. & García-Arenal, F. (2014) Evolution and emergence of plant viruses.
Advances in Virus Research. 88: 161-191. Elton, C.S. (1927) Animal Ecology. Sidgewick and Jackson, London. Ennos, R.A. & Swales, K. W. (1991) Genetic variability and population structure in the canker
pathogen Crumenulopsis sororia. Mycological Research. 95: 521–525. Escriu, F., Fraile, A. & García-Arenal, F. (2000a) Evolution of virulence in natural populations of
the satellite RNA of Cucumber mosaic virus. Phytopathology. 90: 480-485. Escriu, F., Fraile, A. & Garcia-Arenal, F. (2003) The evolution of virulence in a plant virus.
Evolution, International journal of organic evolution. 57: 755–65. Escriu, F., Perry, K. L. & García-Arenal, F. (2000b) Transmissibility of Cucumber mosaic virus by
Aphis gossypii correlates with viral accumulation and is affected by the presence of its satellite RNA. Phytopathology. 90: 1068–1072.
Excoffier, L. & Lischer, H. E. L. (2010) Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Molecular Ecology Resources. 10: 564-567.
Excoffier, L., Smouse, P. E., Quattro, J.M. (1992) Analysis of molecular variance inferred from metric distances among DNA haplotypes: application to human mitochondrial DNA restriction data. Genetics. 131: 479–491.
Falconer, D. S. & Mackay, T. F. C. (1996) Introduction to Quantitative Genetics, 4ª Ed Longman. Londres.
Fargette, D., Pinel, A., Traoré, O., Ghesquière, A. & Konaté, G. (2002) Emergence of resistance-breaking isolates of Rice yellow mottle virus during serial inoculations. European Journal of Plant Pathology. 108: 585-591.

Bibliografía
196
Farnham, G. & Baulcombe, D.C. (2006) Artificial evolution extends the spectrum of viruses that are targeted by a disease-resistance gene from portato. Proceeding of the National Academy os Sciences USA. 103: 18828–18833.
Finch, J. T., Klug, A. & van Regenmortel, M. H. V. (1967) The structure of Cucumber mosaic virus. Journal of Molecular Biology. 24: 269-273.
Fineblum, W.L. & Rausher, M.D. (1995) Trade-off between resistance and tolerance to herbivore damage in a morning glory. Nature. 377: 517–520.
Fitch, W. M. (1971) Toward defining the course of evolution: Minimal change for a specific tree topology. Systematic Zoology. 20: 406-416.
Flor, H. H. (1955) Host-parasite interaction in flax rust -its geneticsand other implications. Phytopathology. 45: 680-685.
Forbes, M. R. L. (1993) Parasitism and host reproductive effort. Oikos. 67: 444-450. Fornoni, J., Valverde, P. L. & Núñez-Farfán, J. (2003) Quantitative genetics of plant tolerance and
resistance against natural enemies of two natural populations of Datura stramonium. Evolutionary Ecology Research. 5: 1049–1065.
Fornoni, J., Valverde, P. L. & Núñez-Farfán, J. (2004) Population variation in the cost and benefit of tolerance and resistance against herbivory in datura stramonium. Evolution, 58: 1696-1704.
Fraile, A. & García-Arenal, F. (2010) The coevolution of plants and viruses: Resistance and pathogenicity. Advances in Virus Research.76: 1-32.
Fraile, A., Alonso-Prados, J. L., Aranda, M. A., Bernal, J. J., Malpica, J. M. & García-Arenal, F. (1997) Genetic exchange by recombination or reassortment is infrequent in natural populations of a tripartite RNA plant virus. Journal of Virology.71: 934-940.
Fraile, A., Hily, J.M., Pagán, I., Pacios, L.F. & García-Arenal, F. (2014) Host resistance selects for traits unrelated to resistance-breaking that affect fitness in a plant virus. Molecular Biology and Evolution. 31: 928-939.
Fraile, A., Pagán, I., Anastasio, G., Saez, E. & García-Arenal, F. (2011) Rapid genetic diversification and high fitness penalties associated with pathogenicity evolution in a plant virus. Molecular Biology and Evolution. 28: 1425-1437.
François, O., Blum, M. G., Jakobsson, M., Rosenberg, N. A. (2008) Demographic history of European populations of Arabidopsis thaliana. PLoS Genetics. 4:e1000075.
Frank, S. A. (1992) Models of plant pathogen co-evolution. Trends in Genetics. 8: 213–219. Frank, S.A. (1994) Recognition of polymorphisms in host–parasite genetics. Philosophical
Transactions Royal Society of Lond B. 346: 283–293. Frank, S. A. (1996) Models of parasite virulence. The Quarterly Review of Biology. 71: 37– 78. Fraser, R. S. S. (1990) The genetics of resistance to plant viruses. Annual Review of Phytopathology.
28:179–200. Frank, S. A. (2000) Specific and non-specific defense against parasitic attack. Journal of Theoretical
Biology. 202: 283–304. Friess, N. & Maillet, J. (1996) Influence of cucumber mosaic virus infection on the intraspecific
competitive ability and fitness of purslane (Portulaca oleracea) New Phytology. 132: 103-111.
Fritz, R. S. & Simms, E. L. (1992) Plant Resistance to Herbivores and Pathogens: Ecology, evolution and genetics. Chicago University Press. Chicago.
Fritzsche, R., Lehmann, K. E. & Proesler, G. (1972) Tierische vectoren pflanzen pathogener viren. Fischer. Gena.

Bibliografía
197
Fukuzawa, N., Itchoda, N., Ishihara, T., Goto, K., Masuta, C. & Matsumura, T. (2010) HC-Pro, a potyvirus RNA silencing suppressor, cancels cycling of Cucumber mosaic virus in Nicotiana benthamiana plants. Virus Genes. 40: 440–446.
Funayama, S., Terashima, I. & Yahara, T. (2001) Effects of virus infection and light environment on population dynamics of Eupatorium makinoi (Asteraceae) American Journal of Botany. 84: 823–829.
Futuyma, D. J. (1998) Evolutionary Biology. 3rd ed. Sinauer Associates, Sunderland, Massachusetts. Gallitelli, D. (2000) The ecology of Cucumber mosaic virus and sustainable agriculture. Virus
Research. 71: 9–21 Gan, X., Stegle, O., Behr, J., Steffen, J. G., Drewe, P., Hildebrand, K. L., Lyngsoe, R.,
Schultheiss, S. J., Osborne, E. J., Sreedharan, V. T., Kahles, A., Bohnert, R., Jean, G., Derwent, P., Kersey, P., Belfield, E., Harberd, N. P., Kemen, E.,Toomajian, C., Kover, P. X., Clark, R. M., Rätsch, M., & Mott, R. (2011) Multiple reference genomes and transcriptomes for Arabidopsis thaliana. Nature. 477: 419-423.
Gandon, S., Jansen, V. A. A. & van Baalen, M. (2001) Host life history and the evolution of parasite virulence. Evolution. 55: 1056-1062.
Gandon, S., Agnew, P., Michalakis, Y. (2002) Coevolution between parasite virulence and host life-history traits. The Americal Naturalist. 160: 374–388.
Gao, L., Roux, F. & Bergelson, J. (2009) Quantitative fitness effects of infection in a gene-for-gene system. New Phytologist. 184: 485–494.
García-Arenal, F. (1997) Genetic exchange by recombination or reassortment is infrequent in natural populations of a tripartite RNA plant virus. Journal of Virology. 71: 934- 940.
García-Arenal, F. & Palukaitis, P. (1999) Structure and functional relationships of satellite RNAs of Cucumber mosaic virus. Current Topics in Microbiology and Inmunology. 239: 37-63.
García-Arenal, F. & Fraile, A. (2013) Trade-offs in host range evolution of plant viruses. Plant Pathology, 62: 2–9.
García-Arenal, F., Fraile, A. & Malpica, J. M. (2001) Variability and genetic structure of plant virus populations. Annual Review of Phytopathology. 39: 157-186.
García-Arenal, F. & McDonald, B. A. (2003) An analysis of the durability of resistance to plant viruses. Phytopathology, 93: 941–952.
Garcı́a-Arenal, F., Escriu, F. & Aranda, M. A. (2000) Molecular epidemiology of Cucumber mosaic virus and its satellite RNA. Virus Research. 71: 1-8.
Gera, A. Loebenstein, G. & Raccah, B. (1979) Protein coats of two strains of Cucumber mosaic virus affect transmission by Aphis gossypii. Phytopathology. 69: 396-399.
Gibbs, A. (1980) Plant virus that partially protects its wild legume host against herbivores. Intervirology. 13: 42-47.
Gillespie, J. H., Turelli, M. (1989) Genotype–environment interactions and the maintenance of polygenic variation. Genetics, 121: 129–138.
Goldbach, R. & Peters, D. (1994) Possible causes of the emergence of tospovirus diseases. Seminar in Virology. 5: 113–120.
Gonsalves, D., Providenti, R. & Edwards, M. C. (1982) Tomato white leaf: the relation of an apparent satellite RNA and Cucumber mosaic virus. Phytopathology. 72: 1533-1538.
Goto, K., Kobori, T., Kosaka, Y., Natsuaki, T. & Masuta, C. (2007) Characterization of silencing suppressor 2b of Cucumber mosaic virus based on examination of its small RNA-binding abilities. Plant Cell Physiology. 48:1050–1060.
Goss, E. M. & Bergelson, J. (2006) Variation in resistance and virulence in the interaction between

Bibliografía
198
Arabidopsis thaliana and a bacterial pathogen. Evolution. 60: 1562-1572. Griffith, C., Kim, E., Donohue, K. (2004) Life-history variation and adaptation in the historically
mobile plant Arabidopsis thaliana (Brassicaceae) in North America. American Journal of Botany. 91: 837–849.
Grosberg, R. K. & Hart, M. W. (2000) Mate selection and the evolution of highly polymorphic self/nonself recognition genes. Science. 289: 2111–2114
Guindon S. & Gascuel O. (2003) A simple, fast and accurate algorithm to estimate large phylogenies by maximum likehood. Systematic Biology. 52: 696-704.
Guo, Y. L., Fitz, J., Schneeberger, K., Ossowski, S., Cao, J., Weigel, D. (2011) Genome-wide comparison of nucleotide-binding site-leucinerich repeat-encoding genes in Arabidopsis. Plant Physiology. 157: 757-769.
Habili, N. & Francki, R. I. B. (1974) Comparative studies on tomato aspermy and cucumber mosaic virases. III. Further studies on relationship and costruction of a virus from parts of the two viral genomes. Virology. 61: 443-449.
Habili, N. & Symons, R. H. (1989) Evolutionary relationship between luteoviruses and other RNA plant viruses based on sequence motifs in their putative RNA polymerases and nucleic acid helicases. Nucleic Acids Research. 17: 9543-9555.
Haining, R.P. (2003) Spatial Data Analysis: Theory and Practice. Cambridge University Press. Cambridge.
Haldane, J. B. S. (1949) Disease and evolution. Ricerca Scientifica. 19: 168-174. Halterman, D. A., Wei, F. & Wise, R.P. (2003) Powdery mildew-induced Mla mRNAs are
alternatively spliced and contain multiple upstream open reading frames. Plant Physiology. 131: 558–567.
Hamada, H., Tomita, R., Iwadate, Y., Kobayashi, K., Minemura, I., Takeuchi, S., Hikichi, Y. & Suzuki, K. (2007) Cooperative effect of two amino acid mutations in the coat protein of Pepper mild mottle virus overcomes L3-mediated resistance in Capsicum plants. Virus Genes. 34: 205-214.
Hamada, K. & Harrison, B. D. (1977) Effects of virus genotype and temperature on seed transmission of nepoviruses. Annals of Applied Biology. 85, 79-92.
Hampton, R. O. & Francki, R. I. B. (1992) RNA-1 dependent seed transmissibility of Cucumber mosaic virus in Phaseolus vulgaris. Phytopathology. 82: 127-130.
Haydon, D. T., Cleaveland, S., Taylor, L. H. & Laurenson, M. K. (2002) Identifying reservoirs of infection: a conceptual and practical challenge. Emerging Infectios Deseases Journal.8: 1468–1473.
Hayes, R. J. & Buck, K. W. (1990) Complete replication of a eukaryotic virus RNA in vitro by a purified RNA-dependent RNA polymerase. Cell. 63: 363-368.
Herms, D. A. & Mattson, W. J. (1992) The dilemma of plants - to grow or defend. Quarterly Review of Biology. 67: 283–335.
Hily, J. M., García, A., Moreno, A., Plaza, M., Wilkinson, M. D., Fereres, A., Fraile, A. & García-Arenal, F. (2014) The Relationship between Host Lifespan and Pathogen Reservoir Potential: An Analysis in the System Arabidopsis thaliana-Cucumber mosaic virus. PLoS Pathogens. 10(11), e1004492.
Hily, J. M., Poulicard, N., Mora, M.-Á., Pagán, I. & García-Arenal, F. (2015) Environment and host genotype determine the outcome of a plant-virus interaction: from antagonism to mutualism. New Phytologist. n/a–n/a.

Bibliografía
199
Hochberg, M.E., Michalakis, Y. & de Meeus, T. (1992) Parasitism as a constraint on the rate of life-history evolution. Journal of Evolutionary Biology. 5: 491–504.
Hochwender, C., Marquis, R. & Stowe, K. (2000) The potential forand constraints on the evolution of compensatory ability in Asclepias syriaca. Oecologia. 122: 361-370.
Hoffmann, M. H. (2002) Biogeography of Arabidopsis thaliana (L.) Heynh. (Brassicaceae) Journal of Biogeography. 29: 125–134.
Hoffmann, M. H. (2005) Evolution of the realized climatic niche in the genus Arabidopsis (Brassicaceae) Evolution. 59: 1425–1436.
Hoffmann, M. H., Glaß, A. S., Tomiuk, J., Schmuths, H., Fritsch, R. M. & Bachmann, K. (2003) Analysis of molecular data of Arabidopsis thaliana (L.) Heynh. (Brassicaceae) with Geographical Information Systems (GIS) Molecular Ecology. 12: 1007-1019.
Holsinger, K. E. & Weir, B. S. (2009) Genetics in geographically structured populations: defining, estimating and interpreting FST. Nature Reviews Genetics. 10: 639–650.
Holub, E. B., Beynon, L. J., CruteI, R. (1994) Phenotypic and genotypic characterization of interactions between isolates of Peronospora parasitica and accessions of Arabidopsis thaliana. Molecular Plant-Microbe Interacttions. 7: 223–39.
Hong, J. S., Ohnishi, S., Masuta, C., Choi, J. K. & Ryu, K. H. (2007) Infection of soybean by Cucumber mosaic virus as determined by viral movement protein. Archives of Virology. 152: 321–328.
Hou, W.-N., Duan, C.-G., Fang, R.-X., Zhou, X.-Y. & Guo, H.-S. (2011) Satellite RNA reduces expression of the 2b suppressor protein resulting in the attenuation of symptoms caused by Cucumber mosaic virus infection. Molecular Plant Pathology. 12: 595-605.
Huh, S. U., Kim, M. J., Ham, B.-K. & Paek, K.-E. (2011) A zinc finger protein Tsip1 controls Cucumber mosaic virus infection by interacting with the replication complex on vacuolar membranes of the tobacco plant. New phytologist. 191: 746–762.
Hwang, M. S., Kim, K. N., Lee, J. H. & Park, Y. I. (2007) Identification of amino acid sequences determining interaction between the cucumber mosaic virus-encoded 2a polymerase and 3a movement proteins. Journal of General Virology. 88: 3445–3451.
Imhoff, M. W., Leonard, K. J. & Main, C. E. (1982) Patterns of bean rust lesion size increase and spore production. Phytopathology. 72: 441-446.
Imhoof, B. & Schmid-Hempel, P. (1998) Patterns of local adaptation of a protozoan parasite to its bumblebee host. Oikos 82: 59–66.
Ishibashi, K., Kezuka, Y., Kobayashi, C., Kato, M., Inoue, T., Nonaka, T., Ishikawa, M., Matsumura, H. & Katoh, E. (2014) Structural basis for the recognition-evasion arms race between Tomato mosaic virus and the resistance gene Tm-1. Proceedings of the National Academy os Science USA. 111: 3486-3495.
Ishibashi, K., Mawatari, N., Miyashita, S., Kishino, H., Meshi, T. & Ishikawa, M. (2012) Coevolution and hierarchical interactions of Tomato mosaic virus and the resistance gene Tm-1. PLoS Pathogens. 8, e1002975.
Kim, M. J., Kim, H. R. & Paek, K.-H. (2006) Arabidopsis tonoplast proteins TIP1 and TIP2 interact with the Cucumber mosaic virus 1a replication protein. Journal of General Virology. 87: 3425–3431.
Jacquemond, M. (2012) Cucumber mosaic virus. Advances in Virus Research. 84: 439–504. Jagger, I. C. (1916) Experiments with the cucumber mosaic disease. Phytopathology. 6: 148-151.

Bibliografía
200
Janzac, B., Fabre, M. F., Palloix, A. & Moury, B. (2009) Phenotype and spectrum of action of the Pvr4 resistance in pepper against potyviruses, and selection for virulent variants. Plant Pathology. 58: 443–449.
Janzac, B., Montarry, J., Palloix, A., Navaud, O. & Moury, B. (2010) A point mutation in the polymerase of Potato virus Y confers virulence toward the Pvr4 resistance of pepper and a high competitiveness cost in susceptible cultivar. Molecular Plant–Microbe Interactions. 23: 823–30.
Janzen, D. H. (1980) When is it coevolution? Evolution. 34: 611–612. Jarosz, A. M. & Burdon, J. J. (1990) Race specific resistance to Phakopsora pachyrhizi and Glycine
argyrea: Phenotypic and genetic patterns. Heredity. 64: 347–353. Jarosz, A. M. & Burdon, J. J. (1991) Host-pathogen interactions in natural populations of Linum
marginale and Melampsora lini. II. Local and regional variation in patterns of resistance and racial structure. Evolution. 45: 1618–27.
Jarozs, A. M. & Davelos, A. I. (1995) Effects of disease in wild plant populations and the evolution of pathogen aggressiveness. New Phytolist. 129:371–387.
Jeger, M. J., Seal, S. E. & van den Bosch, F. (2006) Evolutionary epidemiology of plant virus disease. Advances in Virus Research. 67: 163–203.
Ji, L.-H. & Ding, S.-W. (2001) The suppressor of transgene RNA silencing encoded by Cucumber mosaic virus interferes with salicylic acid-mediated virus resistance. Molecular Plant-Microbe Interactions. 14: 715–724.
Jiménez-Martínez E. S & Bosque-Pérez N. A. (2004) Variation in Barley Yellow Dwarf Virus Transmission Efficiency by Rhopalosiphum padi (Homoptera: Aphididae) after Acquisition from Transgenic and Nontransformed Wheat Genotypes. Journal of Economic Entomology. 97: 1790-1796.
Johanson, U., West, J., Lister, C., Michaels, S. D., Amasino, R. M. & Dean, C. (2000) Molecular analysis of FRIGIDA, a major determinant of natural varition in Arabidopsis flowering time. Science. 290: 344-47.
Jones, J. D. G. & Dangl, J. L. (2006) The plant immune system. Nature. 444: 323-329. Jorgensen, S. & Mauricio, R. (2004) Neutral genetic variation among wild North American
populations of the weedy plant Arabidopsis thaliana is not geographically structured. Molecular Ecology. 13: 3403–3413.
Juenger, T. & Bergelson, J. (2000) The evolution of compensation to herbivory in scarlet gilia, Ipomopsis aggregata: herbivore-imposed natural selectionand the quantitative genetics of tolerance. Evolution. 54: 764-777.
Kaltz, O. & Shykoff, J. A. (1998) Local adaptation in host–parasite systems. Heredity. 81: 361–370. Kang, B. C., Yeam, I., Frantz, J. D., Murphy, J. F. & Jahn, M. M. (2005) The pvr1 locus in pepper
encodes a translation initiation factor eIF4E that interacts with Tobacco etch virus VPg. The Plant Journal. 41: 392-405.
Kaplan, I. B., Lee, K.-C., Canto, T., Wong, S.-K. & Palukaitis, P. (2004) Host-specific encapsidation of a defective RNA3 of Cucumber mosaic virus. Journal of General Virology. 85: 3757–3763.
Karasev, A. V. & Gray, S. M. (2013) Continuous and Emerging Challenges of Potato virus Yin Potato. Annual Review of Phytopathology. 51: 571–586.
Karasov, T. L., Horton, M. W. & Bergelson, J. (2014) Genomic variability as a driver of plant–pathogen coevolution? Current Opinion in Plant Biology. 18: 24-30.
Kawecki, T. J. & Ebert, D. (2004) Conceptual issues of local adaptation. Ecology Letters. 7: 1225–

Bibliografía
201
1241. Kearsey, M. J. & Farquar, A. G. L. (1998) QTL analysis in plants; where are we now? Heredity. 80:
137-142. Kelly, S. E. (1994) Viral pathogens and the advantage of sex in the perennial grass Anthoxantum
odoratum: A review. Philosophical transaction of the Royal Society B. 346: 295-302. Khetarpal R. K., Maisonneuve B., Maury Y., Chalhoub B., Di- nant S., Lecoq H. & Varma A.
(1998) Breeding for resistance to plant viruses. In: Hadidi A., Khetarpal R.K., Koganezawa H. (eds) Plant Virus Disease Control. APS Press, St. Paul, MN, USA.
Kirchner, J. W. & Roy B. A. (2002) Evolutionary implications of host–pathogen specificity: fitness consequences of pathogen virulence traits. Evolutionary Ecology Research. 4: 27-48.
Koella, J. C. & Agnew, P. (1999) A correlated response of a parasite's virulence and life cycle to selection on its host's life history. Journal of Evolutionary Biology. 12: 70-79.
Koörner, Ch. (2007) The use of ‘altitude’ in ecological research. Trends in Ecology and Evolution. 22: 569–574.
Korves, T. M. & Bergelson J. (2003) A Developmental Response to Pathogen Infection in Arabidopsis. Plant Physiology. 133: 339-347.
Kover, P. X. & Schaal, B. A. (2002) Genetic variation for disease resistance and tolerance among Arabidopsis thaliana accessions. Proceedings of the National Academy of Sciences of the United States of America. 99: 11270–11274.
Krasileva, K. V., Zheng, C., Leonelli, L., Goritschnig, S., Dahlbeck, D. & Staskawicz, B. J. (2011) Global Analysis of Arabidopsis/Downy Mildew Interactions Reveals Prevalence of Incomplete Resistance and Rapid Evolution of Pathogen Recognition. PLoS ONE, 6(12), e28765.
Koornneef, M., Alonso-Blanco, C. & Vreugdenhil, D. (2004) Naturally occurring genetic variation in Arabidopsis thaliana. Annual Review of Plant Biology. 55: 141-172.
Laibach, F. (1907) Bot Centbl Beihefte (I) 22:191–210. Laibach, F. (1943) Arabidopsis thaliana (L.) HEYNH. als Objekt fur genetische und
entwicklungsphysiologische Untersuchungen. Bot. Archiv. 44: 439–455. Laine, A. L. (2004) Resistance variation within and among host populations in a plant–pathogen
metapopulation— implications for regional pathogen dynamics. Journal of Ecology. 92: 990–1000.
Laine, A. L. (2005) Spatial scale of local adaptation in a plant-pathogen. Journal of Evolutionary Biology. 18: 1665-1673.
Laine, A. L. (2006) Evolutionary of host resistance: looking for coevolutionry hotspot at small spatial scales. Philosophical Transactions of the Royal Society of London Serie: B: Biological Sciences. 273: 267-273.
Laine, A.L. (2007) Pathogen fitness components and genotypes differin their sensitivity to nutrient and temperature variation in a wild plant-pathogen association. Journal of Evolutionary Biology. 20: 2371–2378.
Lakshman, D. K. & Gonsalves, D. (1985) Genetic analyses of two large-lesion isolates of Cucumber mosaic virus. Phytopathology. 75: 758-762.
Lambrechts, l., Fellous, S. & Koella, J. C. (2006) Coevolutionary interactions between host and parasite genotypes. Trends Parasitoly. 22: 12-16.
Lanfermeijer, F.C., Warmink, J. & Hille, J. (2005) The products of the broken Tm-2 and the durable Tm-22 resistance genes from tomato differ in four amino acids. Journal of Experimental Botany. 56: 2925-2933.

Bibliografía
202
Lasky, J. R., Marais, Des, D. L., Lowry, D. B., Povolotskaya, I., McKay, J. K., Richards, J. H., Keitt, T. H. & Juenger T. E. (2014) Natural Variation in Abiotic Stress Responsive Gene Expression and Local Adaptation to Climate in Arabidopsis thaliana. Molecular Biology and Evolution. 31: 2283–2296.
Lawrence, M. J. (1976) Variations in natural populations of Arabidopsis thaliana (L.) Heynh. The Biology and Chemistry of the Cruciferae. Academic Press. New York. EEUU.
Legendre, P. (1993) Spatial autocorrelation: Trouble or new paradigm? Ecology 74: 1659-1673. Legendre, P. & Legendre, L. (1978) Numerical Ecology. Elsevier. Legendre, P. & M.-J. Fortin. (1989) Spatial pattern and ecological analysis. Vegetatio, 80: 107-138. Legendre, P., M. R. T. Dale, M.-J. Fortin, J. Gurevitch, M. Hohn & D. Myers. (2002) The
consequences of spatial structure for the design and analysis of ecological field surveys. Ecography. 25:601–615.
Le Corre, V. (2005) Variation at two flowering time genes within and among populations of Arabidopsis thaliana: comparison with markers and traits. Molecular Ecology. 14: 4181–4192.
Le Gall, O., Aranda, M. & Caranta, C. (2011) Plant resistance to viruses mediated by translation initiation factors. Recent Advances in Plant Virology. Caister Academic Press. Norfolk, UK.
Lee, J.-A., Choi, S. K., Yoon, J. Y., Hong, J. S., Ryu, K. H., Lee, S. Y. & Choi, J. K. (2007) Variation in the pathogenicity of lily isolates of Cucumber mosaic virus. Plant Pathology Journal. 23: 251–259.
Legendre, P., Dale, M. R. T., Fortin, M. J., Gurevitch, J., Hohn, M. & Myers, D. (2002) The consequences of spatial structure for the design and analysis of ecological field surveys. Ecography 25: 601–615.
Leinonen, T., O’Hara, R. B., Cano, J. M. & Merilä, J. (2008) Comparative studies of quantitative trait and neutral marker divergence: a meta-analysis. Journal of Evolutionary Biology. 21: 1–17.
Leinonen, T., McCairns, R. J. S., O'Hara, R. B. & Merilä, J. (2013) Q(ST)-F(ST) comparisons: evolutionary and ecological insights from genomic heterogeneity. Nature Reviews Genetics, 14: 179–190.
Lenski, R. E. & May, R. M. (1994) The evolution of virulence in parasites and pathogens: reconciliation between two competing hypotheses. Journal of theoretical Biology. 169: 253-265.
Leonard, K. J. & Czochor, R. J. (1989) Theory of genetic interactions among populations of plants and their pathogens. Annual Review of Phytopathology. 18: 237–258.
Leutwiler, L. S., Houghevans, B. R. & Meyerowitz, E. M. (1984) The DNA of Arabidopsis thaliana. Molecular Genetics and Genomic. 194: 15–23.
Lewandowska-Sabat, A. M., Fjellheim, S. & Rognli, O. A. (2010) Extremely low genetic variability and highly structured local populations of Arabidopsis thaliana at higher latitudes. Molecular Ecology. 19: 4753–4764.
Lewsey, M. G., Murphy, A. M., MacLean, D., Dalchau, N., Westwood, J. H., Macaulay, K., Bennett, M. H., Moulin, M., Hanke, D. E., Powell, G., Smith, A. G. & Carr, J. P. (2010) Disruption of two defensive signaling pathways by a viral RNA silencing suppressor. Molecular Plant-Microbe Interacttions. 23: 835–845.
Lewsey, M., Surette, M., Robertson, F. C., Ziebell, H., Choi, S. H., Ryu, K. H., Canto, T., Palukaitis, P., Payne, T., Walsh, J. A. & Carr, J. P. (2009) The role of the Cucumber

Bibliografía
203
mosaic virus 2b protein in viral movement and symptom induction. Molecular Plant-Microbe Interacttions. 6: 642–654.
Li, Q., Ryu, K. H. & Palukaitis, P. (2001) Cucumber mosaic virus-plant interactions: Identification of 3a protein sequences affecting infectivity, cell-to-cell movement, and long-distance movement. Molecular Plant-Microbe Interacttions. 14: 378-385.
Lin, H. X., Rubio, L., Smythe, A., Jiminez, M. & Falk, B. W. (2003) Genetic diversity and biological variation among California isolates of Cucumber mosaic virus. Journal of General Virology. 84: 249-258.
Lipsitch, M., Siller, S. & Nowak, M. A. (1996) The evolution of virulence in pathogens with vertical and horizontal transmission. Evolution. 50: 1729-1741.
Little, T. J., Shuker, D. M., Colegrave, N., Day, T. & Graham, A. L. (2010) The Coevolution of Virulence: Tolerance in Perspective. PLoS Pathogens. 6(9) e1001006.
Lively, C. M. & Dybdahl, M. F. (2000) Parasite adaptation to locally common host genotypes. Nature. 405: 679–681.
Loebenstein, G. (2009) Plant virus diseases: Economic aspects. En: Mahy B.W.J., Van Regenmortel M.H.V. (eds.) Desk Encyclopedia of Plant and Fungal Virology. Elsevier Academic Press, San Diego, EEUU.
López, C., Aramburu, J., Galipienso, L. & Nuez, F. (2007) Characterisation of several heterogeneous species of defective RNAs derived from RNA3 of Cucumber mosaic virus. Archives of Virology. 152: 621–627.
Lot, H. & Kaper, J. M. (1976) Further studies on the RNA component distribution among the nucleoproteins of Cucumber mosaic virus. Virology. 74: 223-226.
Lot, H., Marrou, J., Quiot, J. B. & Esvan, C. (1972) Contribution à l ́étude du virus de la mosaïque du concombre (CMV) Méthode de purification rapide du virus. Annales de phytopathologie. 4: 25-38.
Luis-Arteaga, M., Alvarez, J. M., Alonso-Prados, J. L., Bernal, J. J., García-Arenal, F., Lavin a, A., Batlle, A. & Moriones, E. (1998) Occurrence, distribution, and relative incidence of mosaic viruses infecting field-grown melon in Spain. Plant Disease. 82: 979-982.
Lynch, M. & Walsh, B. (1998) Genetics and Analysis of Quantitative Traits. Sinauer Associates, Sunderland, MA
Ma, M., Zhou, X., Wang, G., Ma, Z. & Du, G. (2010) Seasonal dynamics in alpine meadow seed banks along an altitudinal gradient on the Tibetan Plateau. Plant and Soil. 336: 291–302.
Mackinnon, M.J. & Read, A.F. (2004) Virulence in malaria: an evolutionary viewpoint. Philosophical Transactions of the Royal Society of London Serie: B Biological Sciences, 359, 965–986.
Mahy, B.W. J. & van Regenmortel H. V. (2009) Desk Encyclopedia of Plant and Fungal Virology. 1ª Ed. Academic Press. Amsterdam.
Makkouk, K. M. & Attar, N. (2003) Seed transmission of Cucumber mosaic virus and Alfalfa mosaic virus in lentil seeds. Arab Society Plant Protection. 21:49–52.
Malmstrom, C. M., Hughes, C. C., Newton, L. A. & Stoner, C. J. (2005) Virus infection in remnant native bunchgrasses from invaded California grasslands. New Phytologist. 168: 217–230.
Malmstrom, C. M., McCullough, A. J., Newton, L. A., Johnson, H. A. & Borer, E. T. (2005b) Invasive annual grasses indirectly increase virus incidence in California native perennial bunch grasses. Oecologia. 145: 153–164.

Bibliografía
204
Malmstrom, C. M., Melcher, U. & Bosque-Pérez, N. A. (2011) The expanding field of plant virus ecology: Historical foundations, knowledge gaps, and research directions. Virus Research, 159: 84–94.
Malmstrom, C. M., Stoner, C. J., Brandenburg, S. & Newton, L. A. (2006) Virus infection and grazing exert counteracting influences on survivorship of native bunchgrass seedlings competing with invasive exotics. Journal of Ecology. 94: 264–275.
Malpica, J.M., Sacristán, S., Fraile, A. & García-Arenal, F. (2006) Association and host selectivity in multi-host pathogens. PLoS ONE. 1, e41.
Mantel, N. (1967) The detection of disease clustering and a generalized regression approach. Cancer Research. 27: 209–220.
Mantel, N. & Valand, R. S. (1970) A technique of nonparametric multivariate analysis. Biometrics. 26: 47–558.
Manzano-Piedras, E., Marcer, A., Alonso-Blanco, C. & Picó, F. X. (2014) Deciphering the Adjustment between Environment and Life History in Annuals: Lessons from a Geographically-Explicit Approach in Arabidopsis thaliana. PLoS ONE, 9(2) e87836.
Margaria, P., Ciuffo, M., Pacifico, D. & Turina, M. (2007) Evidence that the nonstructural protein of Tomato spotted wilt virus is the avirulence determinant in the interaction with resistant pepper carrying the Tsw gene. Molecular Plant-Microbe Interactions. 20: 547-558.
Martin, D. P., van der Walt, E., Posada, D. & Rybicki, E. P. (2005) The evolutionary value of recombination is constrained by genome modularity. PLoS Genet. 1: e51.
Mascia, T., Cillo, F., Fanelli, V., Finetti-Sialer, M. M., De Stradis, A., Palukaitis, P. & Gallitelli, D. (2010) Characterization of the interactions between Cucumber mosaic virus and Potato virus Y in mixed infections in tomato. Molecular Plant-Microbe Interactions. 23:1514–1524.
Maskell, L. C., Raybould, A. F., Cooper, J. I., Edwards, M. L. & Gray, A. J. (1999) Effects of turnip mosaic virus and turnip yellow mosaic virus on the survival, growth and reproduction of wild cabbage (Brassica oleracea) Annals Applied Biology. 135: 401-407.
Maskell, L. C., Raybould, A. F., Cooper, J. I., Edwards, M. L. & Gray, A. J. (1999) Effects of turnip mosaic virus and turnip yellow mosaic virus on the survival, growth and reproduction of wild cabbage (Brassica oleracea) Annals of Applied Biology 135, 401-407.
Masuta, C., Seshimo, Y., Mukohara, M., Jung, H. J., Ueda, S., Ryu, K. H., and Choi, J. K. (2002) Evolutionary characterization of two lily isolates of Cucumber mosaic virus isolated in Japan and Korea. Journal of General Plant Pathology. 68: 163–168.
Matthews, R. E. F. (2014) Plant virology, 5ª ed. CA: Academic Press. San Diego. Mauck, K. E., De Moraes, C. M. & Mescher, M. C. (2010) Deceptive chemical signals induced by a
plant virus attract insect vectors to inferior hosts. Proceeding of National Academy of Sciences. 107: 3600–3605.
Maule, A., Caranta, C. & Boulton, M. (2007) Sources of natural resistance to plant viruses: Status and prospects. Molecular Plant Pathology. 8: 223-231.
Mauricio, R. (2000) Natural selection and the joint evolution of tolerance and resistance as plant defenses. Evolutionary Ecology. 14:491–507.
Mauricio, R. (1998) Costs of resistance to natural enemies in field populations of the annual plant Arabidopsis thaliana. The American Naturalist. 151: 20–28
Mauricio, R., Rausher, M. D. & Burdick, D. S. (1997) Variation in the defense strategies of plants: are resistance and tolerance mutually exclusive? Ecology. 78: 1301-1311.
May, R. & Anderson, R.M. (1983) Epidemiolgy and genetics in the coevolution of

Bibliografía
205
parasites and hosts. Proceeding of the Royal Society of London Serie: B. Biological Science. 219: 281-313.
Medzhitov, R., D. S. Schneider & Soares M. P. (2012) Disease tolerance as a defense strategy. Science. 335: 936–941.
Melcher, U., Muthukumar, V., Wiley, G. B., Min, B.E., Palmer, M. W., Verchot-Lubicz, J., Ali, A., Nelson, R. S., Roe, B. A., Thapa, V. & Pierce, M. L. (2008) Evidence for novel viruses by analysis of nucleic acids in virus-like particle fractions from Ambrosia psilostachya. Journal of Virology Methods. 152: 49 –55.
Méndez-Vigo, B., de Andres, M. T., Ramiro, M., Martinez-Zapater, J. M. & Alonso-Blanco, C. (2010) Temporal analysis of natural variation for the rate of leaf production and its relationship with flowering initiation in Arabidopsis thaliana. Journal of Experimental Botany. 61: 1611–1623.
Méndez-Vigo, B., Gomaa, N. H., Alonso-Blanco, C. & Xavier Picó, F. (2012) Among- and within-population variation in flowering time of Iberian Arabidopsis thalianaestimated in field and glasshouse conditions. New Phytologist. 197: 1332–1343.
Méndez-Vigo, B., Pico, F. X., Ramiro, M., Martinez-Zapater, J. M. & Alonso-Blanco, C. (2011) Altitudinal and Climatic Adaptation Is Mediated by Flowering Traits and FRI, FLC, and PHYC Genes in Arabidopsis. Plant Physiology. 157: 1942–1955.
Mestre, P., Brigneti, G., Durrant, M. C. & Baulcombe, D. C. (2003) Potato virus Y Nia protease activity is not sufficient for elicitation of Ry-mediated disease resistance in potato. The Plant Journal. 36: 755 –761.
Michaels, S. D., He, Y., Scortecci, K. C. & Amasino, R. M. (2003) Attenuation of FLOWERING LOCUS C activity as a mechanism for the evolution of summer-annual flowering behavior in Arabidopsis. Proceedings of the National Academy of Sciences of USA. 100: 10102-7.
Michalakis, Y. & Hochberg, M. E. (1994) Parasitic effects on host life-history traits: a review of recent studies. Parasite. 1: 291-294.
Miller, J. R., Wood, B. P. & Hamilton, M. B. (2008) FST and QST Under Neutrality. Genetics, 180: 1023–1037.
Miller, M.R. ,White, A. & Boots, M. (2006) The evolution of parasites in response to tolerance in their hosts: The good, the bad and apparent commensalism. Evolution. 60:945–956.
Miller, M.R., White, A. & Boots, M. (2005) The evolution of host resistance: tolerance and control as distinct strategies. Journal Theoretical Biology. 236: 198– 207.
Minchella, D. J. (1985) Host life-history variation in response to parasitism. Parasitology. 90: 205-216.
Mishra, Y., Jänkäpäa, H. J., Kiss, A. Z., Funk, C., Schröder, W. P. & Jansson, S. (2012) Arabidopsis plants grown in the field and in climate chambers significantly differ in leaf morphology and photosystems components. BMC Plant Biology 12: 6.
Mitchell-Olds, T. & Schmitt, J. (2006) Genetic mechanisms and evolutionary significance of natural variation in Arabidopsis. Nature. 441: 947-52.
MochizukI, T. & Ohki, S. T. (2011) Cucumber mosaic virus: viral genes as virulence determinants. Molecular Plant Pathology. 13: 217–225.
Montarry, J., Cartier, E., Jacquemond, M., Palloix, A. & Moury, B. (2012) Virus adaptation to quantitative plant resistance: erosion or breakdown? Journal of Evolutionary Biology, 25: 2242–2252.

Bibliografía
206
Montarry, J., Corbiere, R., Lesueur, S., Glais, I. & Andrivon, D. (2006) Does selection by resistant hosts trigger local adaptation in plant-pathogen systems? Journal of Evolutionary Biology. 19: 522-531.
Montesinos-Navarro, A., Picó, F. X. & Tonsor, S. J. (2012) Clinal variation in seed traits influencing life cycle timing in arabidopsis thaliana. Evolution. 66: 3417–3431.
Montesinos, A., Tonsor, S. J., Alonso-Blanco, C. & Picó, F. X. (2009) Demographic and Genetic Patterns of Variation among Populations of Arabidopsis thaliana from Contrasting Native Environments. PLoS ONE, 4(9) e7213.
Moran P. A. P. (1950) Notes on continuous stochastic phenomena. Biometrika 37: 17–23 Moreno-Pérez, M.G., Pagán, I., Aragón-Caballero, L., Cáceres, F., Fraile, A. & García-Arenal,
F. (2014) Ecological and genetic determinants of Pepino mosaic virus emergence. Journal of Virology. 88: 3359-3368.
Moreno, A., de Blas, C., Biurrun, R., Nebreda, M., Palacios, I., Duque, M. & Fereres, A. (2004) The incidence and dis- tribution of viruses infecting lettuce, cultivated Brassica and associated natural vegetation. Spain. Annals of Applied Biology 144: 339 – 346.
Moreno, P., Ambrós, S., Albiach-Martí, M. R., Guerri, J. & Peña, L. (2008) Citrus tristeza virus: a pathogen that changed the course of the citrus industry. Molecular Plant Pathology. 9: 251–268.
Mossop, D. W. & Francki, R. I. (1978) Survival of a satellite RNA in vivo and its dependence on Cucumber mosaic virus for replication. Virology. 86: 562-566.
Moury, B., Desbiez, C., Jacquemond, M. & Lecoq, H. (2006) Genetic diversity of plant virus populations: towards hypothesis testing in molecular epidemiology. Advances in Virus Research. 67: 49-87.
Moury, B., Janzac, B., Ruellan, Y., Simon, V., Ben Khalifa, M., Fakhfakh, H., Fabre, F. & Palloix, A. (2014) Interaction patterns between Potato virus Y and eIF4E-mediated recessive resistance in the Solanaceae. Journal of General Virology. 88: 9799-9807.
Murren, C. J., Denning, W. & Pigliucci, M. (2005) Relationships between vegetative and life history traits and fitness in a novel field environment: impacts of herbivores. Evolutionary Ecology. 19: 583-601.
Murant, A.F., Taylor, C.E.& Chambers, J. (1968) Properties, relationships and transmission of a strain of raspberry ringspot virus infecting raspberry cultivars immune to the common Scottish strain. Annals of Applied Biology. 61:175 –186.
Muthukumar, V., Melcher, U., Pierce, M., Wiley, G. B., Roe, B. A., Palmer, M. W., Thapa, V., Ali, A. & Ding, T. (2009) Non-cultivated plants of the Tallgrass Prairie Preserve of northeastern Oklahoma frequently contain virus-like sequences in particulate fractions. Virus Research 141: 169–173.
Nei, M. & Kumar, S. (2000) Molecular evolution and phylogenetics. Oxford University Press. Nueva York.
Nemri, A., Atwell, S. & Tarone, A. M. (2010) Genome-wide survey of Arabidopsis natural variation in downy mildew resistance using combined association and linkage mapping. Proceedings of the National Academy of Science. 107: 10302-10307.
Netsu, O., Hiratsuka, K., Kuwata, S., Hibi, T., Ugaki, M. & Suzuki, M. (2008) Peanut stunt virus 2b cistron plays a role in viral local and systemic accumulation and virulence in Nicotiana benthamiana. Archives Virology. 153: 1731-1735.
Ng, J. C. K., Josefsson, C., Clark, A. J., Franz, A. W. E. & Perry, K. L. (2005) Virion stability and aphid vector transmissibility of Cucumber mosaic virus mutants. Virology. 332:397–405.

Bibliografía
207
Nieto, C., Morales, M., Orjeda, G., Clepet, C., Monfort, A., Sturbois, B., Puigdomènech, P., Pitrat, M., Caboche, M., Dogimont, C., Garcia-Mas, J., Aranda, M. A., Bendahmane, A. (2006) An eIF4Eallele confers resistance to an uncapped and non-polyadenylated RNA virus in melon. The Plant Journal. 48: 452–462.
Nieto, C., Piron, F., Dalmais, M., Marco, C. F., Moriones, E., Gómez-Guillamón, M. L., Truniger, V., Gómez, P., García-Mas, J., Aranda, M.A. & Bendahmane, A. (2007) EcoTILLING for the identification of allelic variants of melon eIF4E, a factor that controls virus susceptibility. BMC Plant Biology. 7:34.
Ninyerola, M., Pons, X. & Roure, J. M. (2000) A methodological approach of climatological modelling of air temperature and precipitation through GIS techniques. International Journal of Climatology. 20: 1823-1841.
Ninyerola, M., Pons, X. & Roure, J. M. (2005) Atlas Climático Digital de la Península Ibérica. Metodología y aplicaciones en bioclimatología y geobotánica. Universidad Autónoma de Barcelona, Bellaterra.
Núñez-Farfán, J., Fornoni, J. & Valverde, P. L. (2007) The Evolution of Resistance and Tolerance to Herbivores. Annual Review of Ecology, Evolution, and Systematics. 38: 541–566.
O ́Reilly, E. K., Tang, N., Ahlquist, P. & Kao, C. C. (1995) Biochemical and genetic analices of the interaction between the helicase-like and polymerase-like proteins of the Brome mosaic virus. Virology. 214: 59-71.
O'Hara, R. B. & Merila, J. (2005) Bias and Precision in QST Estimates: Problems and Some Solutions. Genetics, 171: 1331–1339.
O’Keefe, D. C., Berryman, D. I., Coutts, B. A. & Jones, R. A. C. (2007) Lack of seed coat contamination with Cucumber mosaic virus in lupin permits reliable, large-scale detection of seed transmission in seed samples. Plant Disease. 91:504–508.
Oerke E.C. (2006) Crop losses to pests. Journal of Agricultural Science. 144: 31 - 43. Oh, J. W., Kong, Q., Song, C., Carpenter, C. D., Simon, A. E. (1995) Open reading frames of turnip
crinkle virus involved in satellite expression and incompatibility with Arabidposis thaliana ecotype Dijon. Molecular Plant-Microbe Interactions. 8: 979–987.
Ohshima, K., Yamaguchi, Y., Hirota, R., Hamamoto, T., Tomimura, K., Tan, Z., Sano, T., Azuhata, F., Walsh, J.A., Fletcher, J., Chen, J., Gera, A. & Gibbs, A. (2002) Molecular evolution of Turnip mosaic virus: evidence of host adaptation, genetic recombination and geographical spread. Journal of General Virology. 83: 1511-1521.
Ouibrahim, L. & Caranta, C. (2013) Exploitation of natural genetic diversity to study plant-virus interactions: what can we learn from Arabidopsis thaliana? Molecular Plant Pathology, 14: 844–854.
Pagán, I., Alonso-Blanco, C. & García-Arenal, F. (2007) The Relationship of Within-Host Multiplication and Virulence in a Plant-Virus System. PLoS ONE, 2(8) e786.
Pagán, I., Alonso-Blanco, C. & García-Arenal, F. (2008) Host Responses in Life-History Traits and Tolerance to Virus Infection in Arabidopsis thaliana. PLoS Pathogens, 4(8) e1000124.
Pagán, I., Alonso-Blanco, C. & García-Arenal, F. (2009) Differential tolerance to direct and indirect density-dependent costs of viral infection in Arabidopsis thaliana. PLoS One 5: e1000531.
Pagán, I., Fraile, A., Fernandez-Fueyo, E., Montes, N., Alonso-Blanco, C. & García-Arenal, F. (2010) Arabidopsis thaliana as a model for the study of plant-virus co-evolution. Philosophical Transactions of the Royal Society of London Serie: B: Biological Sciences. 365: 1983–1995.

Bibliografía
208
Pagán, I., Fraile, A. & García-Arenal, F. (2016) Virus Evolution Current Research and Future Directions. Ed Caister Academic Press, UK.
Pagán, I., Montes, N., Milgroom, M. G. & García-Arenal, F. (2014) Vertical Transmission Selects for Reduced Virulence in a Plant Virus and for Increased Resistance in the Host. PLoS Pathogens, 10(7) e1004293.
Pagny, G., Paulstephenraj, P. S., Poque, S., Sicard, O., Cosson, P., Eyquard, J. P., Caballero, M., Chague, A., Gourdon, G., Negrel, L., Candresse, T., Mariette, S. & Decroocq, V. (2012) Family-based linkage and association mapping reveals novel genes affecting plum pox virus infection in Arabidopsis thaliana. New Phytolist. 196: 873–86.
Pallett, D. W., Thurston, M. I., Cortina-Borja, M., Edwards, M. L., Alexander, M., Mitchell, E., Raybould, A. F. & Cooper, J. I. (2002) The incidence of viruses in wild Brassica rapa ssp. sylvestris in southern England. Annals of Applied Biology. 141:163–170.
Palukaitis, P. & García-Arenal, F. (2003a) Cucumber mosaic virus. No. 400. En: Description of plant viruses.
Palukaitis, P. & García-Arenal, F. (2003b) Cucumoviruses. Advances in Virus Research. 62: 241-323.
Palukaitis, P. & Carr, J. P. (2008) Plant resistance responses to viruses. Journal of Plant Pathology, 90: 153–171.
Palukaitis, P., Roossinck, M. J., Dietzgen, R.G. & Francki, R. I. B. (1992) Cucumber Mosaic Virus. Advances in Virus Research. 41: 281-348.
Parker, I.M. & Gilbert, G.S. (2004) The evolutionary ecology of novel plant–pathogen interactions. Annual Review of Ecology, Evolution, and Systematics. 35: 675– 700.
Parker, J., Rambaut, A. R. & Pybus, O. G. (2008) Correlating viral phenotypes with phylogeny: accounting for phylogenetic uncertainty. MEEGID. 8:239-246.
Parker, M. A. (1985) Local population differentiation for compa- tibility in an annual legume and its host-specific fungal pathogen. Evolution. 39: 713–723.
Parker, M. A. (1988) Polymorphism to disease resistance in the annual legume Amphicarpaea bracteata. Heredity. 60: 27–31.
Parker, M. A. (1989) Disease impact and local genetic diversity in the clonal plant Podophyllum peltatum. Evolution 43: 540–547.
Parker, M. A. (1994) Pathogens and sex in plants. Evolutionary Ecology. 8: 560 –584. Parker, M. A. (1996) The nature of plant–parasite specificity. Evolutionary Ecology. 10: 319-322. Peever, T. L., Liu, Y. C., Cortesi, P. & Milgroom, M.G. (2000) Variation in toler- ance and
virulence in the chestnut blight fungus-hypovirus interaction. Applied Environmental Microbiology. 66: 4863-4869.
Perrin, N. & Christe, P. (1996) On host life-history response to parasitism. Oikos. 75: 317-320. Perry, K. L., Zhang, L. & Palukaitis, P. (1998) Amino acid changes in the coat protein of Cucumber
mosaic virus differentially affect transmission by the aphids Myzus persicae and Aphis gossypii. Virology. 242: 204-210.
Perry, K. L., Zhang, L. Shintaku, M. H. & Palukaitis, P. (1994) Mapping determinants in Cucumber mosaic virus for transmission by Aphis gossypii. Virology. 206: 591- 595.
Pfenning, K. S. (2001) Evolution of pathogen virulence: The role of variation in host phenotype. Proceeding of the Royal Society of London Serie: B Biological Science. 268:755-760.
Phan, M. S. V., Seo, J.-K., Choi, H.-S., Lee, S.-H. & Kim, K.-H. (2014) Molecular and Biological Characterization of an Isolate of Cucumber mosaic virus from Glycine soja by Generating its Infectious Full-genome cDNA Clones. The Plant Pathology Journal, 30: 159–167.

Bibliografía
209
Phillips, N.C., Drost, D. T., Varga, W. A. & Shultz, L. M. (2011) Demography, reproduction, and dormancy along altitudinal gradients in three intermoun-tain Allium species with contrasting abundance and distribution. Flora. 206: 164–171.
Picó, F. X. (2012) Demographic fate of Arabidopsis thaliana cohorts of autumn‐and spring‐germinated plants along an altitudinal gradient. Journal of Ecology. 100:1009-1018.
Picó, F. X., Mendez-Vigo, B., Martinez-Zapater, J. M. & Alonso-Blanco, C. (2008) Natural Genetic Variation of Arabidopsis thaliana Is Geographically Structured in the Iberian Peninsula. Genetics, 180: 1009–1021.
Pierrugues, O., Guilbaud, L., Fernandez-Delmond, I., Fabre, F., Tepfer, M. & Jacquemond, M. (2007) Biological properties and relative fitness of inter-subgroup cucumber mosaic virus RNA3 recombinants produced in vitro. Journal of General Virology. 88: 2852–2861.
Pietravalle, S., Lemarié, S. & van den Bosch, F. (2006) Durability of resistance and cost of virulence. European Journal of Plant Pathology. 114, 107–116.
Pigliucci, M. & Kolodynska, A. (2002) Phenotypic plasticity to light intensity in Arabidopsis thaliana: invariance of reaction norms and phenotypic integration. Evolutionary Ecology. 16: 27–47.
Pigliucci, M. & Kolodynska, A. (2006) Phenotypic integration and response to stress in Arabidopsis thaliana: a path analytical approach. Evolutionary Ecology Research. 8: 415–433.
Poland, J. A., Balint-Kurti, P. J., Wisser, R. J., Pratt, R. C. & Nelson, R. J. (2009) Shades of gray: the world of quantitative disease resistance. Trends in Plant Science, 14: 21–29.
Pilson D. (2000) The evolution of plant response to herbivory: simultaneously considering resistance and tolerance in Brassica rapa. Evol Ecol 14:457-489.
Pimentel, D., Feinberg, E. H., Wood, P. W. & Hayes, J. T. (1965) Selection, spatial distribution and the coexistence of competing fly species. The American Naturalist. 99: 97-109.
Pineau, C., Loubet, S., Lefoulon, C., Chalies, C., Fizames, C., Lacombe, B., Ferrand, M., Loudet, O., Berthomieu, P. & Richard O. (2012) Natural Variation at the FRD3 MATE Transporter Locus Reveals Cross-Talk between Fe Homeostasis and Zn Tolerance in Arabidopsis thaliana. PLoS Genetics, 8(12)
Platt A, Horton, M., Huang, Y. S., Li Y., Anastasio, A. E., Mulyati, N. W., Agren, J., Bossdorf, O., Byers, D., Donohue, K., Dunning, M., Holub, E. B., Hudson, A., Le Corre, V., Loudet, O., Roux, R., Warthmann, N., Weigel, D., Rivero, L., Scholl, R., Nordborg, M., Bergelson, J. & Borevitz, J. O. (2010) The scale of population structure in Arabidopsis thaliana. PLoS Genetics. 6:e1000843.
Poch, O., Sauvaget, I., Delarue, M. & Tordo, N. (1989) Identification of four conserved motifs among the RNA-dependent polymerase encoding elements. The EMBO Journal. 8: 3867-3874.
Poulicard, N., Pinel-Galzi, A., Hebrard, E. & Fargette, D. (2010) Why Rice yellow mottle virus, a rapidly evolving RNA plant virus, is not efficient at breaking rymv1-2 resistance. Molecular Plant Pathology. 11: 145-154.
Poulicard, N., Pinel-Galzi, A., Traoré, O., Vignols, F., Ghesquière, A, Konaté, G., Hébrard, E. & Fargette, D. (2012) Historical contingencies modulate the adaptability of Rice yellow mottle virus. PLoS Pathog. 8, e1002482.
Poulin, R. (2014) Parasite biodiversity revisited: frontiers and constraints. International Journal for Parasitology. 44: 581-589.
Poulin, R. & Morand, S. (2000) The diversity of parasites. The Quarterly Review of Biology. 75: 277-293.

Bibliografía
210
Poulin, R. & Morand, S. (2004) Parasite Biodiversity. Smithsonian Institution Press,Washington, USA.
Poulin, R., Fredensborg B. L., Hansen, E. & Leung, L.F. (2005) The true cost of host manipulation by parasites. Behavioural Processes. 68: 241-244
Power, A.G. & Mitchell, C.E. (2004) Pathogen spillover in disease epidemics. The American Naturalist. 164: S79-S89.
Power, A.G., Borer, E.T., Hosseini, P., Mitchell, C.E. & Seabloom, E. W. (2011) The community ecology of barley/cereal yellow dwarf viruses in Western US grasslands. Virus Research. 159: 95-100.
Prendeville, H. R., Tenhumberg, B. & Pilson, D. (2014) Effects of virus on plant fecundity and population dynamics. New Phytologist, 202(4) 1346–1356.
Prendeville, H. R., Ye, X., Jack Morris, T. & Pilson, D. (2012) Virus infections in wild plant populations are both frequent and often unapparent. American Journal of Botany. 99: 1033–1042.
Preston, D. & Johnson, P. (2010) Ecological Consequences of Parasitism. Nature Education Knowledge. 3:47.
Price, J.S., Bever, J.D. & Clay, K. (2004) Genotype, environment, and genotype by environment interactions determine quantitative resistance to leaf rust (Coleosporium asterum) in Euthamia graminifolia (Asteraceae) New Phytholist. 162: 729-743.
Price, P. W., Westoby, M., Rice, B., Arsatt, P. R.., Fritz, R. S., Thompson, J. N. & Mobley, K. (1986) Parasite mediation in ecological interactions. Annual review of ecology and systematic. 17: 487-505
Price, P.W. (1980) Evolutionary Biology of Parasites. Princeton University Press. Princeton, NJ Quenouille, J., Montarry, J., Palloix, A. & Moury, B. (2013) Farther, slower, stronger: how the
plant genetic background protects a major resistance gene from breakdown. Molecular Plant Pathology. 14: 109–18.
Quenouille, J., Paulhiac, E., Moury, B. & Palloix, A. (2014) Quantitative trait loci from the host genetic background modulate the durability of resistance gene: a rational basis for sustainable resistance breedindin in plant. Heredity, 112: 579-587.
Quinn, G. P. & Keough, M. J. (2002) Experimental Design and Data Analysis for Biologists. Cambridge University Press.
Quiot, J. B., Marchoux, G., Douine, L. & Vigoroux, A. (1979) Ecologie et Epidémiologie du Virus de la Mosaique du Cocombre dans le Sud-East de la France. V — Role des espéces spontanées dans da conservation du virus. Annales de phytopathologie. P11: 325–348.
Rangel, T.F., Diniz-Filho, J.A.F. & Bini, L.M. (2010) SAM: A comprehensive aplication for Spatail Analysis in Macroecology. Ecography. 33: 1-5.
Raybould, A. F., Alexander, M. J., Mitchell, E., Thurston, M.I., Pallett, D.W., Hunter, P., Walsh, J.A., Edwards, M.-L., Jones, A. M. E., Moyes, C. L. Gray, A. J. &Cooper, J. I. (2001) The ecology of turnip mosaic virus in wild populations of Brassica species. Hails RS, Beringer JE, Charles H, Godfray J, eds. Genes in the environment: 15th Special Symposium of the British Ecological Society. Cambridge University Press. Oxford, UK
Read, A. F. (1994) The evolution of virulence. Trends in Microbiology. 2: 73–76. Remold, S. K. (2002) Unapparent virus infection and host fitness in three weedy grass species. Journal
of Ecology. 90: 967–977. Restif, O. & Koella, J. C. (2003) Shared control of epidemiological traits in a coevolutionary model
of host-parasite interactions. American Naturalist, 161: 827–836.

Bibliografía
211
Revers, F., Guiraud, T., Houvenaghel, M.C., Mauduit, T., Le Gall, O. & Candresse, T. (2003) Multiple resistance phenotypes to Lettuce mosaic virus among Arabidopsis thaliana accessions. Mol. Plant–Microbe Interactions. 16: 608–616.
Ridley, M. (1993) Evolution. Blackwell Scientific Publications, London. Rietveld, K., Pleij, C. W. & Bosch, L. (1983) Three-dimensional models of the tRNA-like 3'termini
of some plant viral RNAs. The EMBO Journal 2: 1079. Rizzo, T.M. & Palukaitis, P. (1990) Construction of full-length cDNA of Cucumber mosaic virus RNAs 1, 2 and 3: generation of infectious RNA transcripts. Molecular Genetics and
Genomics. 222: 249-256. Robert, C., Bancal, M. O. & Lannou, C. (2002) Sheath leaf rust uredospore production and carbon
and nitrogen export in relation to lesion size and density. Phytopathology. 92: 762-768. Rodelo-Urrego, M., Pagán, I., González-Jara, P., Betancourt, M., Moreno-Letelier, A., Ayllón,
M.A., Fraile, A., Piñero, D. & García-Arenal, F. (2013) Landscape heterogeneity shapes host-parasite interactions and results in apparent plantvirus codivergence. Molecular Ecology. 22: 2325-2340.
Rogers, A. R. & Harpending, H. C. (1983) Population structure and quantitative characters. Genetics. 105: 985-1002.
Roossinck, M. J., Saha, P., Wiley, G. B, Quan, J, White, J. D, Lai, H, Chavarría, F, Shen, G. & Roe, B. A. (2010) Ecogenomics: using massively parallel pyrosequencing to understand virus ecology.Molecular Ecology. 19:81-88.
Roossinck, M. J. (2002) Evolutionary history of Cucumber mosaic virus deduced by phylogenetic analyses. Journal of Virology.76: 3382-3387.
Roossinck, M. J. (2005) Symbiosis versus competition in plant virus evolution. Nature Reviews Microbiology. 3: 917–924.
Roossinck M. J. (2012) Persistent plant viruses: molecular hitchhikers or epigenetic elements? In Viruses: Essential Agents of Life. Edited by Witzany G. Springer.
Roossinck, M. J. & García-Arenal, F. (2015) Ecosystem simplification, biodiversity loss and plant virus emergence. Current Opinion in Virology. 10: 56–62.
Roossinck, M. J., Zhang, L. & Hellwald, K. H. (1999) Rearrangements in the 5 ́ nontranslated region and phylogenetic analyses of Cucumber mosaic virus RNA 3 indicete radial evolution of three subgroups. Journal of Virology.73: 6752-6758.
Rosenberg, M. S. & Anderson, C. D. (2011) PASSaGE: pattern analysis, spatial statistics and geographic exegesis. Version 2. Methods in Ecology and Evolution. 2: 229–232.
Roy, B.A. & Kirchner, J. W. (2000) Evolutionary dynamics of pathogen resistance and tolerance. Evolution. 54: 51–63.
Rua, M.A., Pollina, E. C., Power, A. G. & Mitchell, C. E. (2011) The role of viruses in biological invasions: friend or foe? Current Opinion in Virology. 1: 68-72.
Ryang, B.-S., Kobori, T., Matsumoto, T., Kosaka, Y. & Ohki, S. T. (2004) Cucumber mosaic virus 2b protein compensates for restricted spread of Potato virus Y in doubly infected tobacco. Journal of General Virology. 85:3405–3414.
Ryu, K. H., Park, H. W. & Choi, J. K. (2002) Characterization and sequence analysis of a lily isolate of Cucumber mosaic virus from Lilium tsingtauense. Plant Pathology Journal 18: 85–92.
Sambrook, J. & Russell, D. W. (2001) Molecular cloning: A laboratory manual. 3a Ed. Cold Spring Harbor Laboratory Press. Cold Spring Harbor. New York.

Bibliografía
212
Sacristán, S., Fraile, A. & García-Arenal, F. (2004) Population dynamics of Cucumber mosaic virus in melon crops and in weeds in Central Spain. Phytopathology. 94: 992–998.
Sacristán, S. & García-Arenal, F. (2008) The evolution of virulence and pathogenicity in plant pathogen populations. Molecular Plant Pathology. 9: 369-384.
Sacristán, S., Fraile, A., Malpica, J. M. & García-Arenal, F. (2005) An Analysis of Host Adaptation and Its Relationship with Virulence in Cucumber mosaic virus. Phytopathology. 95: 827–833.
Salánki, K., Gellert, A., Huppert, E., Náray-Szabó, G. & Balázs, E. (2004) Compatibility of the movement protein and the coat protein of cucumoviruses is required for cell-to-cell movement. Journal of General Virology. 85: 1039–1048.
Salánki, K., Gellert, A., Náray-Szabó, G. & Balázs, E. (2007) Modeling-based characterization of the elicitor function of amino acid 461 of Cucumber mosaic virus 1a protein in the hypersensitive response. Virology. 358: 109-118.
Salvaudon L. & Shykoff J.A. (2013) Variation in Arabidopsis developmental responses to oomycete infection: resilience vs changes in life history traits. New Phytologist. 197: 919–926.
Salvaudon, L., Héraudet, V. & Shykoff, J. A. (2005) Parasite-host fitness trade-offs change with parasite identity: genotype-specific interactions in a plant-pathogen system. Evolution. 59: 2518-2524.
Salvaudon, L., Heraudet, V. & Shykoff, J. A. (2008b) Arabidopsis thaliana and the Robin Hood parasite: a chivalrous oomycete that steals fitness from fecund hosts and benefits the poorest one? Biology Letters. 4: 526-529.
Salvaudon, L., Giraud, T. & Shykoff, J. A. (2008a) Genetic diversity in natural populations: a fundamental component of plant–microbe interactions. Current Opinion in Plant Biology, 11: 135–143.
Sánchez-Bermejo, E., Castrillo, G., del Llano, B. A. R., Navarro, C., Zarco-Fernández, S., Martinez-Herrera, D. J., Leo-del Puerto, Y., Muñoz, R., Cámara, C., Paz-Ares, J., Alonso-Blanco, C. & Leyva, A. (2015) Natural variation in arsenate tolerance identifies an arsenate reductase in Arabidopsis thaliana. Nature Communications. 5: 1–9.
Sandland, G.J. & Minchella, D.J. (2003) Costs of immune defense: an enigma wrapped in an environmental cloak? Trends in Parasitology. 19: 571–574.
Sasaki, N., Park, J.-W., Maule, A. J. & Nelson, R. S. (2006) The cysteine-histidine-rich region of the movement protein of Cucumber mosaic virus contributes to plasmodesmal targeting, zinc binding and pathogenesis. Virology. 349: 396-408.
Schmalhausen, I.I. (1947) The Factors of Evolution. Blakiston. Schmid, K.J., Torjek, O., Meyer, R., Schmuths, H., Hoffmann, M.H. & Altmann, T. (2006)
Evidence for a large-scale population structure of Arabidopsis thaliana from genome-wide single nucleotide polymorphism markers. Theoretical Applied Genetics. 112:1104–1114.
Schürch, S. & Roy, B. A. (2004) Comparing single- vs. mixed-genotype infections of Mycosphaerella graminicola on wheat: effects of pathogen virulence and host tolerance. Evolutionary Ecology. 18: 1-14.
Schwinghamer, M. W. & Symons, R. H. (1975) Fractionation of Cucumber mosaic virus RNA and its translation in a wheat embrio cell-free system. Virology. 63: 252-262.
Sclavounos, A. P., Voloudakis, A. E., Arabatzis, C. H. & Kyriakopoulou, P. E. (2006) A severe Hellenic CMV tomato isolate: Symptom variability in tobacco, characterization and discrimination of variants. European Journal of Plant Pathology. 115: 163–172.

Bibliografía
213
Seabloom, E. W., Borer, E. T., Mitchell, C. E. & Power, A. G. (2010) Viral diversity and prevalence gradients in North American Pacific Coast grasslands. Ecology. 91: 721-732.
Seabloom, E. W., Borer, E. T., Jolles, A. & Mitchell, C. E. (2009) Direct and indirect effects of viral pathogens and the environment on invasive grass fecundity in Pacific Coast grasslands. Journal of Ecology 97: 1264–1273.
Senthil-Kumar, M. & Mysore, K. S. (2013) Nonhost Resistance Against Bacterial Pathogens: Retrospectives and Prospects. Annual Review of Phytopathology, 51: 407–427.
Sharbel, T.F., Haubold, B. & Mitchell-Olds, T. (2000) Genetic isolation by distance in Arabidopsis thaliana: biogeography and postglacial colonization of Europe. Molecular Ecology. 9: 2109–2118.
Shi, B. –J., Ding, S. –W. & Symons, R. H. (1997) In vivo expression of an overlapping gene encoded by the cucumoviruses. Journal of General Virology. 78: 237-241.
Shi, B. –J., Miller, J., Symons, R. H. & Palukaitis, P. (2003) The 2b protein of cucumoviruses has a role in promoting cell-to-cell movement of pseudorecombinant viruses. Mol. Plant-Microbe Interactions. 16: 261-267.
Shimizu, K.K. & Purugganan, M. D. (2005) Evolutionary and ecological genomics of Arabidopsis. Plant Physiology. 138: 578–584.
Shindo, C., Bernasconi, G. & Hardtke, C. S. (2007) Natural Genetic Variation in Arabidopsis: Tools, Traits and Prospects for Evolutionary Ecology. Annals of Botany. 99: 1043–1054.
Shykoff, J. A. & Kaltz, O. (1997) Effects of the anther smut fungus Microbotryum violaceum on host life-history patterns in Silene latifolia (Caryophyllaceae) International Journal of Plant Science. 158: 164-171.
Shykoff, J. A. & Kaltz, O. (1998) Phenotypic changes in host plants diseased by Microbotryum violaceum: parasite manipulation, side effects, and trade-offs. International Journal of Plant Science. 159: 236-243
Siddiqui, S. A., Valkonen, J. P. T., Rajamáki, M.-L. & Lehto, K. (2011) The 2b silencing suppressor of a mild strain of Cucumber mosaic virus alone is sufficient for synergistic interaction with Tobacco mosaic virus and induction of severe leaf malformation in 2b-transgenic tobacco plants. Molecular Plant-Microbe Interactions. 24: 685–693.
Siemens, D.H., Lischke, H., Maggiulli, N., Schurch, S. & Roy, B.A. (2003) Cost of resistance and tolerance under competition: the defence-stress benefit hypothesis. Evolutionary Ecology. 17: 247–263.
Simms, E. & Triplett, J. (1994) Costs and benefits of plant responses to disease: resistance and tolerance. Evolution. 48: 1973-1985.
Simons, J. N. (1957) Three strains of Cucumber mosaic virus affecting bell pepper in the everglades area of south Florida. Phytopathology. 47: 145-150.
Simons, J. N. (1958) Titers of three nonpersistent aphid-borne viruses affecting pepper in south Florida. Phytopathology. 48: 265-268.
Smith, T. J., Chase, E., Schmidt, T. & Perry, K. L. (2000) The structure of Cucumber mosaic virus and comparison to cowpea chlorotic mottle virus. Journal of Virology.74: 7578- 7586.
Sokal, R.R. & Oden, N. L. (1978) Spatial autocorrelation in biology 1. Methodology, Biological Journal of the Linnean Society, pp. 10-199.
Sokal, R. R. & Rohlf, F. J. (2012) Biometry: the principles and practice of statistics in biological research. 4ª Ed. W. H. Freeman and Co. New York.
Somerville, C. R. & Meyerowitz, E. M. (2002) The Arabidopsis Book. American Society of Plant Biologists. Rockville. EEUU.

Bibliografía
214
Smouse, P., Long, J. & Sokal, R. (1986) Multiple regression and correlation extensions of the Mantel test of matrix correspondence. Systematic Zoology. 35: 627–632.
Soosaar, J. L. M., Burch-Smith, T. M. & Dinesh-Kumar, S. P. (2005) Mechanisms of plant resistance to viruses. Nature Reviews Microbiology. 3: 789–798.
Sorci, G., Møller, A.P. & Boulinier, T. 1997. Genetics of host– parasite interactions. Trends in Ecology & Evolution. 12: 196–200.
Sorensen, R.E. & Minchella, D. J. (1998) Parasite influences on host life history: Echinostoma revolutum parasitism of Lymnaea elodes snails. Oecologia. 115: 188-195.
Spitze, K. (1993) Population structure in Daphnia obtusa: quantitative genetic and allozymic variation. Genetics. 135: 367–374.
Stearns, S. C. (1976) Life-history tactics: A review of the ideas. The Quarterly Review of Biology. 51: 3-47.
Stenøien, H. K., Fenster, C. B., Tonteri, A. & Savolainen, O. (2005) Genetic variability in natural populations of Arabidopsis thaliana in northern Europe. Molecular Ecology. 14:137–148.
Stevens, R.B. (1960) In Plant Pathology, an Advanced Treatise (Vol. 3) Horsfall, J.G. and Dimond, A.E. Academic Press
Stobbe, A.H. & Roossinck, M.J. (2014) Plant virus metagenomics: what we know and why we need to know more. Frontiers in Plant Science. 5: 150.
Stowe, K. A., Marquis, R. J., Hochwener, C. G., Simms, E. L. (2000) The evolutionary ecology of tolerance to consumer damage. Annual Review of Ecology and Systematics. 31: 565–595.
Stowe, K.A. (1998) Experimental evolution of resistance in Brassica rapa: correlated response of tolerance in lines selected for glucosinolate content. Evolution. 52: 703–712.
Strauss, S. Y. & Agrawal, A. A. (1999) The ecology and evolution of plant tolerance to herbivory. Trends in Ecology & Evolution. 14: 179–185.
Su, S., Liu, Z., Chen, C., Zhang, Y., Wang, X., Zhu, L., Miao, L., Wang, X.-C. & Yuan, M. (2010) Cucumber mosaic virus movement protein severes actin filaments to increase the plasmodesmal size exclusion limit in tobacco. Plant Cell. 22: 1373–1387.
Svensson, E. I. & Raberg, L. (2010) Resistance and toler- ance in animal enemy-victim coevolution. Trends in Ecology & Evolution. 25: 267–274.
Swarup, K., Alonso-Blanco, C., Lynn, J. R., Michaels, S. D., Amasino, R. M., Koornneef, M. & Millar, A. J. (1999) Natural allelic variation identifies new genes in the Arabidopsis circadian system. Plant Journal. 20: 67-77.
Symons, R. H. (1979) Extensive sequence homology at the 3 ́-termini of the four RNAs. Takahashi, H., Goto, N. & Ehara, Y. (1994) Hypersensitive response in cucumber mosaic virus-
inoculated Arabidopsis thaliana. The Plant Journal. 6: 369-377. Takahashi, H., Miller, J., Nonaki, Y., Takeda, M., Shah, J., Hase, S., Ikegami, M., Ehara, Y. &
Dinesh-Kumar, S. P. (2002) RCY1, an Arabidopsis thaliana RPP8/HRT family resistance gene, conferring resistance to cucumber mosaic virus requires salicylic acid, ethylene and a novel signal transduction mechanism. The Plant Journal. 32:655–667.
Takeshita, M., Matsuo, Y., Yoshikawa, T., Suzuki, M., Furuya, N., Tsuchiya, K. & Takanami, Y. (2008) Characterisation of a defective RNA derived from RNA3 of the Y strain of Cucumber mosaic virus. Archives of Virology. 153: 579–583.
Tamura, K. & Nei, M. (1993) Estimation of the number of nucleotide substitutions in the control region of mitochondrial DNA in humans and chimpanzees. Molecular Biology and Evolution. 10: 512–526.

Bibliografía
215
Tamura, K., Stecher, G., Peterson, D., Filipski, A. & Kumar, S. (2013) MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Molecular biology and Evolution 30: 2725-2729.
Tellier, A. & Brown, J. K. M. (2007) Stability of genetic polymorphism in host–parasite interactions. Proceeding of the Royal Society of London Serie B: Biological Science. 274: 809 –817.
The Arabidopsis Genome Initiative (2000) Analysis of the genome sequence of the flowering plant Arabidopsis thaliana. Nature. 408: 796–815.
Thines, M. (2014) Phylogeny and evolution of plant pathogenic oomycetes—a global overview. European Journal of Plant Pathology. 138: 431–447.
Thompson, J. R., Buratti, E., de Wispelaere, M. & Tepfer, M. (2008) Structural and functional characterization of the 50 region of subgenomic RNA5 of Cucumber mosaic virus. Journal of General Virology. 89:1729–1738.
Thompson, J. N. 1994. The coevolutionary process. The University of Chicago Press, Chicago. Thompson, R. (1982) Methods of estimation of genetics parametres. 2ª Ed. World Congress of
Genetics Applied to Livestock Production. Editorial Garsi, Madrid. Thrall, P. H., Hochberg, M. E., Burdon, J. J. & Bever, J.D. (2007) Coevolution of symbiotic
mutualists and parasites in a community context. Trends in Ecology & Evolution. 22: 120–126.
Thrall, P. H. & Burdon, J. J. (2002) Evolution of gene-for- gene systems in metapopulations: the effect of spatial scale of host and pathogen dispersal. Plant Pathology. 51: 169–184.
Thrall, P. H., Burdon, J. J. & Young, A. (2001) Variation in resistance and virulence among demes of a plant host– pathogen metapopulation. Journal of Ecology. 89: 736–748.
Thurston, M. I., Pallet, D. W., Cortina-Borja, M., Edwards, M. L., Raybould, A. F. & Cooper, J. I. (2001) The inci- dence of viruses in wild Brassica nigra in Dorset (UK) Annals of Applied Biology 139: 277–284.
Tian, D.M., Traw, B., Chen, J.Q., Kreitman, M. & Bergelson, J. (2003) Fitness costs of R-gene-mediated resistance in Arabidopsis thaliana. Nature. 423:74 –77.
Tiffin, P. (2000) Are tolerance, avoidance and antibiosis evolutionary and ecologically equivalent responses of plants to herbivores? The American Naturalist. 155: 128–138.
Tiffin, P. & Rausher, M.D. (1999) Genetic constraints and selection acting on tolerance to herbivory in the common morning glory Ipomoea purpurea. The American Naturalist. 154: 700–716
Todesco, M., Balasubramanian, S., Hu, T. T., Traw, M. B., Horton, M., Epple, P., Kuhns, C., Sureshkumar, S., Schwartz, C., Lanz, C., Roosa, A. Laitinen, E., Huang, Y., Chory, J., Lipka, V., Borevitz, J., Dangl, J. L. Bergelson, B., Nordborg, M & Weigel, W. (2010) Natural allelic variation underlying a major fitness trade-off in Arabidopsis thaliana. Nature, 465: 632-636.
Todokoro, S., Terauchi, R. & Kawano, S. (1995) Microsatellite polymorphisms in natural populations of Arabidopsis thaliana in Japan. The Japanese Journal of Genetics. 70: 543–554.
Tomita, R., Murai, J., Miura, Y., Ishikara, H., Liu, S., Kubotera, Y., Honda, A., Hatta, R., Kuroda, T., Hamada, H., Sakamoto, M., Munemura, I., Nunomura, O., Ishikawa, K., Genda, Y., Kawasaki, S., Suzuki, K., Meksem, K., and Kobayashi, K. (2008) Fine mapping and DNA fiber FISH analysis locates the tobamovirus resistance gene L3 of Capsicum chinense in a 400-kb region of R-like genes cluster embedded in highly repetitive sequences. Theorical and Applied Genetics. 117: 1107-1118.

Bibliografía
216
Tsuda, S., Kirita, M. & Watanabe, Y. (1998) Characterization of a pepper mild mottle tobamovirus strain capable of overcoming the L3 gene-mediated resistance, distinct from the resistance-breaking Italian isolate. Molecular Plant-Microbe Interactions. 11: 327-331.
Ueda, H., Yamaguchi, Y. & Sano, H. (2006) Direct interaction between the Tobacco mosaic virus helicase domain and the ATP-bound resistance protein, N factor during the hypersensitive response in tobacco plants. Plant Molecular Biology 61: 31–45.
Upton, G. & Fingleton, B. (1985) Spatial data analysis by example. Vol. I: point pattern and quantitative data, Wiley. Chichester, UK.
van Berloo R. (2008) GGT 2.0: versatile software for visualization and analysis of genetic data. The Journal of Heredity 99: 232-6.
van Damme, M., Andel, A., Huibers, R. P., Panstruga, R., Weisbeek, P. J. & van den Ackerveken. (2005) Identification of arabidopsis loci required for susceptibility to the downy mildew pathogen Hyaloperonospora parasitica. Molecular Plant-Microbe Interactions. 18: 583–592.
van der Meijden, E., Wijn, H. & Verkaar, J. (1988) Defence and regrowth: alternative plant strategies in the struggle against herbivores. Oikos. 51: 355–363.
van der Plank, J.E. (1968) Disease Resistance in Plants. Academic Press. van Noordwijk, A. J. & de Jong, G. (1986) Acquisition and allocation of resources: their influence
on variation in life history tactics. The American Naturalist. 128: 137-142. van Zandt, P.A. & Mopper, S. (1998) A meta-analysis of the adaptive deme formation analysis in
phytophagous insect populations. The American Naturalist. 152: 595–604. Vila-aiub, M. M., Neve, P., Roux, F. (2011) A unified approach to the esti- mation and interpretation
of resistance costs in plants. Heredity. 107: 386–394. Wang, W., Barnaby, J.Y., Tada, Y., Li, H., Toer, M., Caldelari, D., Lee, D.u., Fu, X.D. & Dong,
X. (2011) Timing of plant immune responses by a central circadian regulator. Nature. 470: 110-114.
Wang, T.H., Donaldson, Y.K., Brettle, R.P., Bell, J.E. & Simmonds, P. (2001) Identification of shared populations of Human immunodeficiency virus Type 1 infecting microglia and tissue macrophages outside the central nervous system. Journal Virology. 75: 11686-11699.
Wang, Y., Lee, K. C., Gaba, V., Wong, S. M., Palukaitis, P. & Gal-On, A. (2004a) Breakage of resistance to Cucumber mosaic virus by co-infection with Zucchini yellow mosaic virus: Enhancement of CMV accumulation independent of symptom expression. Archives of Virology. 149: 379–396.
Wang, Y., Tzfira, T., Gaba, V., Citovsky, V., Palukaitis, P. & Gal-On, A. (2004b) Functional analysis of the Cucumber mosaic virus 2b protein: Pathogenicity and nuclear localization. Journal of General Virology. 85: 3135–3147.
Warthmann, N., Fitz, J. & Weigel, D. (2007) MSQT for choosing SNP assays from multiple DNA alignments. Bioinformatics 23: 2784–2787.
Weinig, C., Stinchcombe, J. R. & Schmitt, J. (2003) QTL architecture of resistance and tolerance traits in Arabidopsis thaliana in natural environments. Molecular Ecology. 12: 1153-1163.
Weir, B.S. & Cockerham, C.C. (1984) Estimating F-statistics for the analysis of population structure. Evolution. 38: 1358-1370.
Wetter C. & Conti M. (1988) Pepper mild mottle virus. No. 330. Description of plant viruses. Whitham, S., Dinesh-Kumar, S. P., Choi, D., Hehl, R., Corr, C. & Baker, B. (1994) The product of
the tobacco mosaic virus resistance gene N: Similarity to Toll and the interleukin-1 receptor. Cell. 78:1101–1115.

Bibliografía
217
Whitlock, M. C. (2008) Evolutionary inference from QST. Molecular Ecology. 17: 1885–1896. Wilcox, R. R. (2012) Introduction to Robust Estimation and Hypothesis Testing (3rd Edition)
Academic Press. San Diego. Williams, G. C. (1966) Adaptation and natural selection. Princeton University Press. Princeton. Williams, G. C. (1957) Pleiotropy, natural selection, and the evolution of senescence. Evolution. 11:
398–411. Williams, D.G. & Black, R.A. (1993) Phenotypic variation in contrasting tem- perature environments:
growth and photosynthesis in Pennisetum Setaceum from different altitudes on Hawaii. Functional Ecology. 7: 623–633.
Wilson, I. W., Schiff, C. L., Hughes, D. E. & Somerville, S. C. (2001) Quantitative trait loci analysis of powdery mildew disease resistance in the Arabidopsis thaliana accession Kashmir-1. Genetics. 158: 1301-1309.
Windsor, D.A. (1998) Most of the species on Earth are parasites. International Journal for Parasitology. 28:1939-1941.
Woodward, F.I. (1987) Climate and Plant Distribution. Cambridge University Press, Cambridge. Woolhouse M.E.J., Webster J.P., Domingo E., Charlesworth B. & Levin B.R. (2002) Biological
and biomedical implications of the co-evolution of pathogens and their hosts. Nature Genetics. 32: 569–577.
Woolhouse, M.E.J., Taylor, L.H. & Haydon, D.T. (2001) Population biology of multi-host pathogens. Science. 292: 1109-1112.
Wren, J. D., Roossinck, M. J., Nelson, R. S., Scheets, K., Palmer, M. W. & Melcher, U. (2006) Plant virus biodiversity and ecology. PLoS Biol. 4, 314–315.
Wright, S. (1943) Isolation by Distance. Genetics, 38:114-138. Xu, P., Chen, F., Mannas, J.P., Feldman, T., Sumner, L.W. & Roossinck, M.J. (2008) Virus
infection improves drought tolerance. New Phytologist. 180: 911-921. Yahara, T. & Oyama, K. (1993) Effects of virus infection on demographic traits of an
agamospermous population of Eupatorium chinense (Asteraceae) Oecologia. 96: 310–315. Yang, Y., Kim, K. S. & Anderson, E. J. (1997) Seed transmission of Cucumber mosaic virus in
spinach. Phytopathology. 87: 924–931. Young, N.D. (1996) QTL mapping and quantitative disease resistance in plants. Annual Review of
Phytopathology. 34: 479–501. Zhang, L., Hanada, K. & Palukaitis, P. (1994) Mapping local and systemic symptom determinants
of cucumber mosaic cucumovirus in tobacco. Journal of General Virology. 75: 3185- 3191. Zhang, N., Lariviere, A., Tonsor, S. J. & Traw, M. B. (2014) Constitutive camalexin production and
environmental stress response variation in Arabidopsis populations from the Iberian Peninsula. Plant Science, 225: 77–85.
Zhang, Y., Lubberstedt, T. & Xu, M. (2013) The Genetic and Molecular Basis of Plant Resistance to Pathogens. Journal of Genetics and Genomics, 40: 23-35.
Zhu, Y., Chen, H., Fan, J., Wang, Y., Li, Y., Chen, J., Yang, S., Hu, L., Leung, H., Mew, T. W., Teng, S. P., Wang, Z. & Mundt, C. C. (2000) Genetic diversity and disease control in rice. Nature 406: 718-722.10.
Zhu, W., Ausin, I., Seleznev, A., Méndez-Vigo, B., Picó, F. X., Sureshkumar, S., Sureshkumar, S., Sundaramoorthi, V., Bulach, D., Powell, D., Seemann, T., Alonso-Blanco, C. & Balasubramanian, S. (2015) Natural Variation Identifies ICARUS1, a Universal Gene Required for Cell Proliferation and Growth at High Temperatures in Arabidopsis thaliana. PLoS Genetics, 11(5).


ANEJO 1

Anejo 1
200

Anejo 1
221
Tabla A1.1. Incidencia de virus en las poblaciones silvestres de arabidopsis listadas en Material y Métodos, Apartado 3.2 y factores demográficos del huésped.
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Ciruelos de Coca 2005 Abr - - - - 0.029 (1/35) 0.200 (7/35) 0.000 (0/35) 0.000 (0/35) 0.000 (0/35) 0.229 (8/35) Ciruelos de Coca 2005 May - - - - - - - - - - Ciruelos de Coca 2006 Abr - - - - 0.032 (1/31) 0.194 (6/31) 0.000 (0/31) 0.000 (0/31) 0.065 (2/31) 0.323 (10/31) Ciruelos de Coca 2006 May - - - - 0.136 (3/22) 0.455 (10/22) 0.045 (1/22) 0.000 (0/22) 0.000 (0/22) 0.591 (13/22) Ciruelos de Coca 2007 Abr - - - - 0.100 (3/30) 0.167 (5/30) 0.200 (6/30) 0.000 (0/30) 0.067 (2/30) 0.500 (15/30) Ciruelos de Coca 2007 May - - - - - - - - - - Ciruelos de Coca 2008 Feb 10 100 0 0 0.300 (3/10) 0.300 (3/10) 0.100 (1/10) 0.100 (1/10) 0.000 (0/10) 0.500 (5/10) Ciruelos de Coca 2008 Mar 20 68 32 0 0.600 (15/25) 0.120 (3/25) 0.600 (15/25) 0.560 (14/25) 0.040 (1/25) 0.760 (19/25) Ciruelos de Coca 2008 Abr 26 0 100 0 0.150 (5/33) 0.120 (4/33) 0.360 (12/33) 0.120 (4/33) 0.000 (0/33) 0.550 (18/33) Ciruelos de Coca 2008 May 20 0 96 4 0.000 (0/25) 0.000 (0/25) 0.400 (10/25) 0.360 (9/25) 0.000 (0/25) 0.400 (10/25) Ciruelos de Coca 2008 Jun - - - - - - - - - - Ciruelos de Coca 2008 Nov - - - - 0.000 (0/1) 1.000 (1/1) 1.000 (1/1) 0.000 (0/1) 0.000 (0/1) 1.000 (1/1) Ciruelos de Coca 2010 Mar 22.667 66.66 33.33 0 0.063 (1/16) 0.438 (7/16) 0.000 (0/16) 0.000 (0/16) - 0.500 (8/16) Ciruelos de Coca 2010 Abr 16 0 100 0 0.171 (7/41) 0.073 (3/41) 0.244 (10/41) 0.098 (4/41) - 0.585 (24/41) Ciruelos de Coca 2010 May 32.8 0 0 100 1.000 (5/5) 1.000 (5/5) 0.000 (0/5) 0.000 (0/5) - 1.000 (5/5) Ciruelos de Coca 2010 Nov 4 100 0 0 0.000 (0/2) 0.000 (0/2) 0.000 (0/2) 0.000 (0/2) - 0.000 (0/2) Ciruelos de Coca 2011 Feb 1.333 100 0 0 1.000 (1/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 1.000 (1/1) Ciruelos de Coca 2011 Mar 6.667 50 50 0 0.000 (0/18) 0.333 (6/18) 0.389 (7/18) 0.278 (5/18) 0.556 (10/18) 0.500 (9/18) Ciruelos de Coca 2011 Abr 9.6 0 0 100 0.000 (0/11) 0.000 (0/11) 0.182 (2/11) 0.182 (2/11) 0.000 (0/11) 0.455 (5/11) Ciruelos de Coca 2011 May 3.2 0 0 100 - - - - - -
2011 Nov 104 100 0 0 0.000 (0/55) 0.145 (8/55) 0.000 (0/55) 0.000 (0/55) 0.000 (0/55) 0.000 (0/55) 2012 Feb 125.778 100 0 0 0.000 (0/75) 0.000 (0/75) 0.000 (0/75) 0.000 (0/75) 0.000 (0/75) 0.000 (0/75)

Anejo 1
222
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Ciruelos de Coca 2012 Mar 246.667 100 0 0 0.036 (5/139) 0.129 (18/139) 0.000 (0/139) 0.022 (3/139) 0.000 (0/139) 0.173 (24/139)
2012 Abr a 225.778 89.82 10.18 0 0.037 (4/107) 0.187 (20/107) 0.065 (7/107) 0.056 (6/107) 0.009 (1/107) 0.318 (34/107) Ciruelos de Coca 2012 Abr b 247.429 3.46 96.3 0.23 0.017 (2/121) 0.884 (3/107) 0.025 (5/107) 0.041 (0/107) 0.000 (0/107) 0.264 (32/107) Ciruelos de Coca 2012 May 170.857 0 0 100 - - - - - - Ciruelos de Coca 2012 Nov 216 100 0 0 0.000 (0/17) 0.059 (1/17) 0.000 (0/17) 0.000 (0/17) 0.000 (0/17) 0.059 (1/17) Ciruelos de Coca 2013 Ene 175.333 100 0 0 0.125 (1/8) 0.625 (5/8) 0.000 (0/8) 0.000 (0/8) 0.000 (0/8) 0.750 (6/8) Ciruelos de Coca 2013 Feb 130 100 0 0 0.053 (2/38) 0.342 (13/38) 0.000 (0/38) 0.000 (0/38) 0.000 (0/38) 0.395 (15/38) Ciruelos de Coca 2013 Mar 98.666 16.89 83.11 0 0.040 (1/25) 0.080 (2/25) 0.040 (2/25) 0.000 (0/25) 0.000 (0/25) 0.160 (4/25) Ciruelos de Coca 2013 Abr 132 0 0 100 0.069 (2/29) 0.172 (5/25) 0.069 (2/25) 0.034 (1/25) 0.000 (0/25) 0.345 (10/25) Ciruelos de Coca 2013 May 0 0 0 0 - - - - - - Ciruelos de Coca 2013 Nov 92 100 0 0 0.000 (0/25) 0.000 (0/25) 0.000 (0/25) 0.000 (0/25) 0.000 (0/25) 0.000 (0/25) Ciruelos de Coca 2014 Feb 190.667 100 0 0 0.103 (7/68) 0.309 (21/68) 0.088 (6/68) 0.044 (3/68) 0.000 (0/68) 0.544 (37/68) Ciruelos de Coca 2014 Mar 100.571 0 100 0 0.083 (1/12) 0.167 (2/12) 0.083 (1/12) 0.000 (0/12) 0.000 (0/12) 0.333 (4/12) Ciruelos de Coca 2014 Abr 58.857 0 0 100 0.111 (2/18) 0.000 (0/12) 0.000 (0/12) 0.056 (1/12) 0.000 (0/12) 0.167 (3/12)
Las Rozas 2005 Abr - - - - 0.000 (0/21) 0.667 (14/21) 0.000 (0/21) 0.238 (5/21) 0.000 (0/21) 0.905 (19/21) Las Rozas 2005 May - - - - - - - - - - Las Rozas 2006 Abr - - - - - - - - - - Las Rozas 2006 May - - - - 0.167 (3/18) 0.222 (4/18) 0.111 (2/18) 0.111 (2/18) 0.000 (0/18) 0.500 (9/18) Las Rozas 2007 Abr - - - - 0.154 (4/26) 0.115 (3/26) 0.038 (1/26) 0.038 (1/26) 0.077 (2/26) 0.385 (10/26) Las Rozas 2007 May - - - - 0.074 (2/27) 0.222 (6/27) 0.000 (0/27) 0.000 (0/27) 0.074 (2/27) 0.333 (9/27) Las Rozas 2008 Feb 150 100 0 0 0.070 (2/30) 0.070 (2/30) 0.070 (2/30) 0.070 (2/30) 0.070 (2/30) 0.230 (7/30) Las Rozas 2008 Mar 42 37.92 62.08 0 0.310 (9/29) 0.070 (2/29) 0.280 (8/29) 0.450 (19/29) 0.380 (11/29) 0.720 (21/29)
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.
Anejo 1
223
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Las Rozas 2008 Abr 248 7.14 92.83 0 0.020 (1/42) 0.020 (1/42) 0.260 (11/42) 0.380 (16/42) 0.070 (3/42) 0.500 (21/42) Las Rozas 2008 May 205 0 44.12 55.88 0.150 (4/24) 0.350 (8/24) 0.150 (4/24) 0.060 (1/24) 0.030 (1/24) 0.380 (9/24) Las Rozas 2008 Jun - - - - - - - - - - Las Rozas 2008 Nov - - - - 0.000 (0/11) 0.000 (0/11) 0.000 (0/11) 0.000 (0/11) 0.000 (0/11) 0.000 (0/11) Las Rozas 2010 Mar 186.4 4.05 95.95 0 0.192 (0/52) 0.327 (17/52) 0.000 (0/52) 0.000 (0/52) - 0.519 (17/52)
2010 Abr 24.8 91.66 8.34 0 0.071 (1/14) 0.286 (4/14) 0.429 (6/14) 0.071 (1/14) - 0.857 (12/14) Las Rozas 2010 May 10.4 0 61.53 38.47 0.000 (0/8) 0.000 (0/8) 0.750 (6/8) 0.000 (6/8) - 0.750 (6/8) Las Rozas 2010 Nov 35.2 100 0 0 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) - 0.000 (0/5) Las Rozas 2011 Feb 19.2 91.66 8.34 0 0.000 (0/2) 0.000 (0/2) 1.000 (2/2) 0.000 (0/2) 0.000 (0/2) 0.500 (1/2)
Las Rozas 2011 Mar 133.6 0 100 0 0.277 (44/159)
0.503 (80/159) 0.635 (101/159) 0.132 (21/159) 0.101 (16/159) 0.868
(138/159) Las Rozas 2011 Abr 40 0 76 24 0.059 (1/17) 0.059 (1/17) 0.000 (0/17) 0.000 (0/17) 0.118 (2/17) 0.176 (3/17) Las Rozas 2011 May - - - - - - - - - - Las Rozas 2011 Nov 155.2 100 0 0 0.000 (0/17) 0.000 (0/17) 0.000 (0/17) 0.000 (0/17) 0.000 (0/17) 0.000 (0/17) Las Rozas 2012 Feb 73.6 100 0 0 0.000 (0/57) 0.000 (0/57) 0.000 (0/57) 0.000 (0/57) 0.000 (0/57) 0.000 (0/57) Las Rozas 2012 Mar 158 67.08 32.92 0 0.013 (1/76) 0.026 (2/76) 0.000 (0/76) 0.000 (0/76) 0.000 (0/76) 0.039 (3/76) Las Rozas 2012 Abr a 144.667 36.07 69.93 0 0.097 (6/62) 0.081 (5/62) 0.081 (5/62) 0.032 (2/62) 0.016 (1/62) 0.290 (18/62) Las Rozas 2012 Abr b 194 0 100 0 0.042 (3/71) 0.099 (7/71) 0.042 (3/71) 0.014 (1/71) 0.014 (1/71) 0.197 (14/71) Las Rozas 2012 May 48.666 0 0 100 - - - - - - Las Rozas 2012 Nov 75.2 100 0 0 0.000 (0/36) 0.000 (0/36) 0.000 (0/36) 0.000 (0/36) 0.000 (0/36) 0.000 (0/36) Las Rozas 2013 Ene 100 100 0 0 0.000 (0/7) 0.143 (1/7) 0.000 (0/7) 0.000 (0/7) 0.000 (0/7) 0.143 (1/7) Las Rozas 2013 Feb 80.666 27.5 72.5 0 0.059 (2/34) 0.147 (5/34) 0.059 (2/34) 0.029 (1/34) 0.029 (1/34) 0.324 (11/34) Las Rozas 2013 Mar 104.666 0 100 0 0.044 (2/45) 0.022 (1/45) 0.067 (3/45) 0.044 (2/45) 0.022 (1/45) 0.200 (9/45)
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.
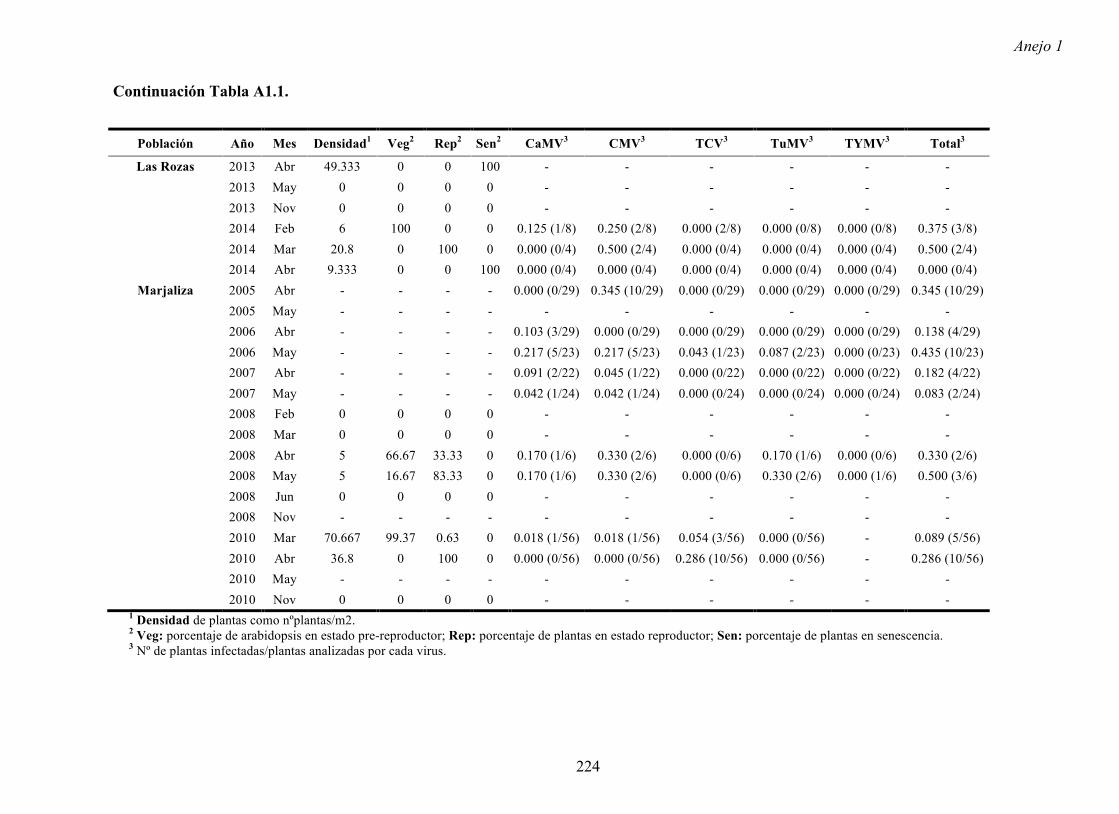
Anejo 1
224
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Las Rozas 2013 Abr 49.333 0 0 100 - - - - - - Las Rozas 2013 May 0 0 0 0 - - - - - - Las Rozas 2013 Nov 0 0 0 0 - - - - - - Las Rozas 2014 Feb 6 100 0 0 0.125 (1/8) 0.250 (2/8) 0.000 (2/8) 0.000 (0/8) 0.000 (0/8) 0.375 (3/8) Las Rozas 2014 Mar 20.8 0 100 0 0.000 (0/4) 0.500 (2/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.500 (2/4) Las Rozas 2014 Abr 9.333 0 0 100 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) Marjaliza 2005 Abr - - - - 0.000 (0/29) 0.345 (10/29) 0.000 (0/29) 0.000 (0/29) 0.000 (0/29) 0.345 (10/29)
2005 May - - - - - - - - - - 2006 Abr - - - - 0.103 (3/29) 0.000 (0/29) 0.000 (0/29) 0.000 (0/29) 0.000 (0/29) 0.138 (4/29)
2006 May - - - - 0.217 (5/23) 0.217 (5/23) 0.043 (1/23) 0.087 (2/23) 0.000 (0/23) 0.435 (10/23) Marjaliza 2007 Abr - - - - 0.091 (2/22) 0.045 (1/22) 0.000 (0/22) 0.000 (0/22) 0.000 (0/22) 0.182 (4/22) Marjaliza 2007 May - - - - 0.042 (1/24) 0.042 (1/24) 0.000 (0/24) 0.000 (0/24) 0.000 (0/24) 0.083 (2/24) Marjaliza 2008 Feb 0 0 0 0 - - - - - - Marjaliza 2008 Mar 0 0 0 0 - - - - - - Marjaliza 2008 Abr 5 66.67 33.33 0 0.170 (1/6) 0.330 (2/6) 0.000 (0/6) 0.170 (1/6) 0.000 (0/6) 0.330 (2/6) Marjaliza 2008 May 5 16.67 83.33 0 0.170 (1/6) 0.330 (2/6) 0.000 (0/6) 0.330 (2/6) 0.000 (1/6) 0.500 (3/6) Marjaliza 2008 Jun 0 0 0 0 - - - - - - Marjaliza 2008 Nov - - - - - - - - - - Marjaliza 2010 Mar 70.667 99.37 0.63 0 0.018 (1/56) 0.018 (1/56) 0.054 (3/56) 0.000 (0/56) - 0.089 (5/56) Marjaliza 2010 Abr 36.8 0 100 0 0.000 (0/56) 0.000 (0/56) 0.286 (10/56) 0.000 (0/56) - 0.286 (10/56) Marjaliza 2010 May - - - - - - - - - - Marjaliza 2010 Nov 0 0 0 0 - - - - - -
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.

Anejo 1
225
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Marjaliza 2011 Feb 11.2 100 0 0 0.333 (1/3) 0.333 (1/3) 0.333 (1/3) 0.333 (1/3) 0.000 (0/3) 1.000 (3/3) Marjaliza 2011 Mar 4.8 100 0 0 0.000 (0/8) 0.250 (2/8) 0.625 (5/8) 0.250 (2/8) 0.000 (0/8) 1.000 (8/8) Marjaliza 2011 Abr 8 0 0 100 0.000 (0/7) 0.143 (1/7) 0.143 (1/7) 0.143 (1/7) 0.000 (0/7) 0.428 (3/7) Marjaliza 2011 May 0 0 0 0 - - - - - - Marjaliza 2011 Nov 25.6 100 0 0 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) Marjaliza 2012 Feb 2.285 100 0 0 0.000 (0/2) 0.000 (0/2) 0.000 (0/2) 0.000 (0/2) 0.000 (0/2) 0.000 (0/2) Marjaliza 2012 Mar 16.444 91.89 8.11 0 0.000 (0/12) 0.000 (0/12) 0.000 (0/12) 0.000 (0/12) 0.000 (0/12) 0.000 (0/12) Marjaliza 2012 Abr a 19.111 16.27 83.73 0 0.000 (0/14) 0.000 (0/14) 0.071 (1/14) 0.000 (0/14) 0.000 (0/14) 0.071 (1/14) Marjaliza 2012 Abr b 12.444 0 100 0 0.000 (0/11) 0.182 (2/14) 0.000 (0/14) 0.000 (0/14) 0.000 (0/14) 0.182 (2/14) Marjaliza 2012 May 7.555 0 5.55 94.45 - - - - - -
2012 Nov 25.333 100 0 0 0.000 (0/12) 0.167 (0/12 ) 0.000 (0/14) 0.000 (0/14) 0.000 (0/14) 0.167 (2/14) 2013 Ene 133.333 100 0 0 0.143 (2/14) 0.429 (6/14) 0.071 (1/14) 0.071 (1/14) 0.000 (0/14) 0.714 (10/14) 2013 Feb 143.666 35.26 64.74 0 0.043 (2/47) 0.000 (0/47) 0.021 (1/47) 0.000 (0/47) 0.000 (0/47) 0.064 (3/47)
2013 Mar 158 1.26 98.74 0 0.030 (2/66) 0.045 (3/66) 0.015 (1/66) 0.000 (0/66) 0.015 (1/66) 0.106 (7/66) Marjaliza 2013 Abr 127.428 0 41.42 58.58 0.050 (2/40) 0.175 (7/40) 0.050 (2/40) 0.025 (1/40) 0.000 (0/40) 0.300 (12/40) Marjaliza 2013 May 77.89 0 0 100 - - - - - - Marjaliza 2013 Nov 239.555 100 0 0 0.000 (0/31) 0.000 (0/31) 0.000 (0/31) 0.000 (0/31) 0.000 (0/31) 0.000 (0/31) Marjaliza 2014 Feb 207.556 100 0 0 0.095 (6/63) 0.175 (11/63) 0.048 (3/63) 0.032 (2/63) 0.016 (1/63) 0.365 (23/63) Marjaliza 2014 Mar 160 0.55 98.05 1.39 0.040 (2/50) 0.080 (4/50) 0.040 (2/50) 0.000 (0/50) 0.020 (1/50) 0.180 (9/50) Marjaliza 2014 Abr 147.556 0 0 100 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4)
Menasalbas 2005 Abr - - - - 0.000 (0/33) 0.364 (12/33) 0.000 (0/33) 0.000 (0/33) 0.000 (0/33) 0.364 (0/33) Menasalbas 2005 May - - - - - - - - - -
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.

Anejo 1
226
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Menasalbas 2006 Abr - - - - 0.273 (9/33) 0.091 (3/33) 0.000 (0/33) 0.000 (0/33) 0.061 (2/33) 0.424 (14/33) Menasalbas 2006 May - - - - 0.042 (1/24) 0.042 (1/24) 0.000 (0/24) 0.042 (1/24) 0.000 (0/24) 0.125 (3/24) Menasalbas 2007 Abr - - - - 0.250 (5/20) 0.000 (0/20) 0.000 (0/20) 0.000 (0/20) 0.000 (0/20) 0.350 (7/20) Menasalbas 2007 May - - - - 0.000 (0/16) 0.000 (0/16) 0.000 (0/16) 0.000 (0/16) 0.063 (1/16) 0.063 (1/16) Menasalbas 2008 Feb 0 0 0 0 - - - - - - Menasalbas 2008 Mar 0 0 0 0 - - - - - - Menasalbas 2008 Abr 12 6.67 93.33 0 0.200 (3/16) 0.200 (3/16) 0.330 (5/16) 0.530 (8/16) 0.000 (0/16) 0.500 (8/16) Menasalbas 2008 May 19 0 91.17 8.3 0.040 (1/24) 0.380 (9/24) 0.080 (2/24) 0.080 (2/24) 0.000 (0/24) 0.500 (12/24) Menasalbas 2008 Jun 5 0 33.33 66.67 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) Menasalbas 2008 Nov - - - - 0.000 (0/3) 0.200 (2/3) 0.200 (2/3) 0.000 (0/3) 0.000 (0/3) 0.300 (2/3) Menasalbas 2010 Mar 50 97.6 2.4 0 0.097 (3/31) 0.000 (0/31) 0.065 (2/31) 0.000 (0/31) - 0.161 (5/31) Menasalbas 2010 Abr 44.8 0 100 0 0.040 (1/25) 0.000 (0/25) 0.080 (2/25) 0.000 (0/25) - 0.120 (3/25) Menasalbas 2010 May 30.4 0 0 100 - - - - - - Menasalbas 2010 Nov 58.4 100 0 0 0.000 (0/8) 0.000 (0/8) 0.000 (0/8) 0.000 (0/8) - 0.000 (0/8)
2011 Feb 15.2 78.94 21.06 0 0.214 (3/14) 0.500 (7/14) 0.143 (2/14) 0.000 (0/14) 0.143 (2/14) 0.929 (13/14)
2011 Mar 130.4 86.5 13.5 0 0.273 (35/128) 0.383 (49/128) 0.633 (81/128) 0.305 (39/128) 0.125 (16/128) 0.891 (114/128)
2011 Abr 89.6 5.35 91.07 3.58 0.132 (9/68) 0.250 (17/68) 0.132 (9/68) 0.162 (11/68) 0.191 (13/68) 0.941 (64/68)
2011 May 14 0 0 100 - - - - - - Menasalbas 2011 Nov 29 100 0 0 0.000 (0/18) 0.056 (1/18) 0.000 (0/18) 0.000 (0/18) 0.000 (0/18) 0.000 (0/18) Menasalbas 2012 Feb 29.333 100 0 0 0.000 (0/18) 0.000 (0/18) 0.111 (2/18) 0.000 (0/18) 0.000 (0/18) 0.111 (2/18) Menasalbas 2012 Mar 20.666 96.77 3.23 0 0.000 (0/15) 0.067 (1/15) 0.000 (0/15) 0.000 (0/15) 0.000 (0/15) 0.067 (1/15) Menasalbas 2012 Abr a 28.5 14.03 85.97 0 0.038 (1/26) 0.346 (9/26) 0.115 (3/26) 0.077 (2/26) 0.077 (2/26) 0.615 (16/26)
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.

Anejo 1
227
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Menasalbas 2006 Abr - - - - 0.273 (9/33) 0.091 (3/33) 0.000 (0/33) 0.000 (0/33) 0.061 (2/33) 0.424 (14/33) Menasalbas 2006 May - - - - 0.042 (1/24) 0.042 (1/24) 0.000 (0/24) 0.042 (1/24) 0.000 (0/24) 0.125 (3/24) Menasalbas 2007 Abr - - - - 0.250 (5/20) 0.000 (0/20) 0.000 (0/20) 0.000 (0/20) 0.000 (0/20) 0.350 (7/20) Menasalbas 2007 May - - - - 0.000 (0/16) 0.000 (0/16) 0.000 (0/16) 0.000 (0/16) 0.063 (1/16) 0.063 (1/16) Menasalbas 2008 Feb 0 0 0 0 - - - - - - Menasalbas 2008 Mar 0 0 0 0 - - - - - - Menasalbas 2008 Abr 12 6.67 93.33 0 0.200 (3/16) 0.200 (3/16) 0.330 (5/16) 0.530 (8/16) 0.000 (0/16) 0.500 (8/16) Menasalbas 2008 May 19 0 91.17 8.3 0.040 (1/24) 0.380 (9/24) 0.080 (2/24) 0.080 (2/24) 0.000 (0/24) 0.500 (12/24) Menasalbas 2008 Jun 5 0 33.33 66.67 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) Menasalbas 2008 Nov - - - - 0.000 (0/3) 0.200 (2/3) 0.200 (2/3) 0.000 (0/3) 0.000 (0/3) 0.300 (2/3) Menasalbas 2010 Mar 50 97.6 2.4 0 0.097 (3/31) 0.000 (0/31) 0.065 (2/31) 0.000 (0/31) - 0.161 (5/31) Menasalbas 2010 Abr 44.8 0 100 0 0.040 (1/25) 0.000 (0/25) 0.080 (2/25) 0.000 (0/25) - 0.120 (3/25) Menasalbas 2010 May 30.4 0 0 100 - - - - - - Menasalbas 2010 Nov 58.4 100 0 0 0.000 (0/8) 0.000 (0/8) 0.000 (0/8) 0.000 (0/8) - 0.000 (0/8)
2011 Feb 15.2 78.94 21.06 0 0.214 (3/14) 0.500 (7/14) 0.143 (2/14) 0.000 (0/14) 0.143 (2/14) 0.929 (13/14)
2011 Mar 130.4 86.5 13.5 0 0.273 (35/128) 0.383 (49/128) 0.633 (81/128) 0.305 (39/128) 0.125 (16/128) 0.891 (114/128)
2011 Abr 89.6 5.35 91.07 3.58 0.132 (9/68) 0.250 (17/68) 0.132 (9/68) 0.162 (11/68) 0.191 (13/68) 0.941 (64/68)
2011 May 14 0 0 100 - - - - - - Menasalbas 2011 Nov 29 100 0 0 0.000 (0/18) 0.056 (1/18) 0.000 (0/18) 0.000 (0/18) 0.000 (0/18) 0.000 (0/18) Menasalbas 2012 Feb 29.333 100 0 0 0.000 (0/18) 0.000 (0/18) 0.111 (2/18) 0.000 (0/18) 0.000 (0/18) 0.111 (2/18) Menasalbas 2012 Mar 20.666 96.77 3.23 0 0.000 (0/15) 0.067 (1/15) 0.000 (0/15) 0.000 (0/15) 0.000 (0/15) 0.067 (1/15) Menasalbas 2012 Abr a 28.5 14.03 85.97 0 0.038 (1/26) 0.346 (9/26) 0.115 (3/26) 0.077 (2/26) 0.077 (2/26) 0.615 (16/26)
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.
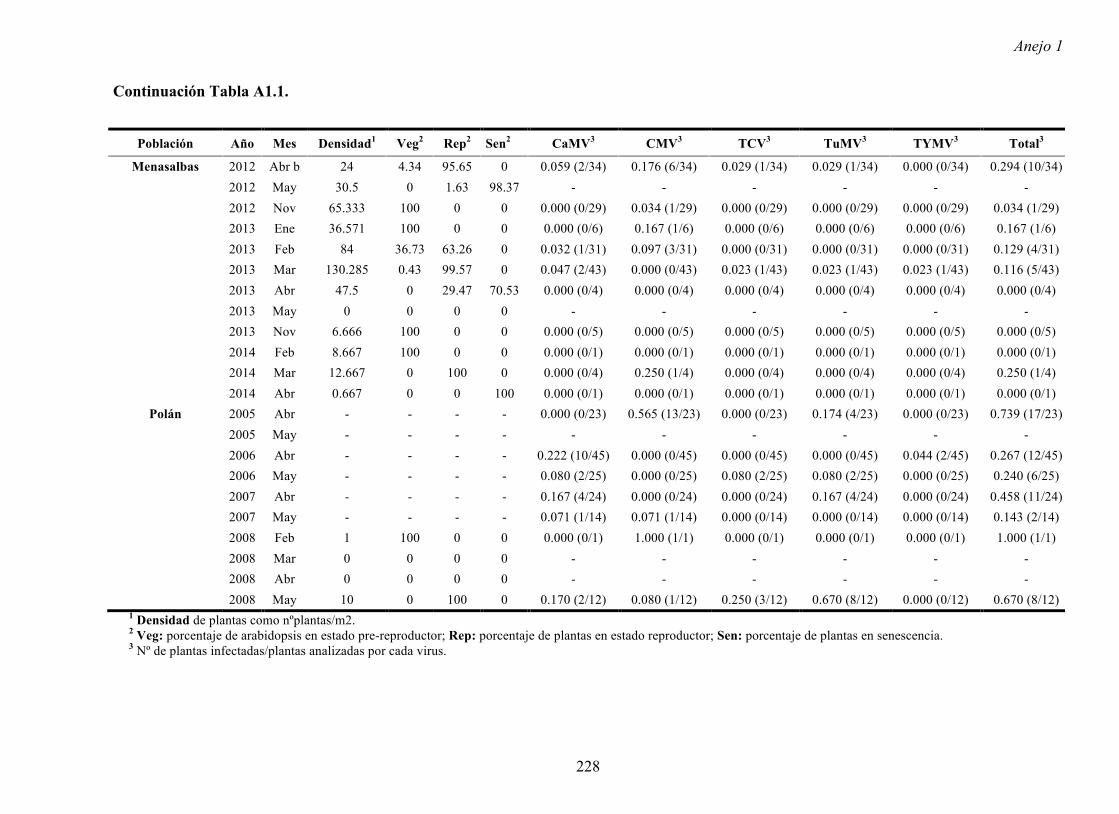
Anejo 1
228
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Menasalbas 2012 Abr b 24 4.34 95.65 0 0.059 (2/34) 0.176 (6/34) 0.029 (1/34) 0.029 (1/34) 0.000 (0/34) 0.294 (10/34) Menasalbas 2012 May 30.5 0 1.63 98.37 - - - - - - Menasalbas 2012 Nov 65.333 100 0 0 0.000 (0/29) 0.034 (1/29) 0.000 (0/29) 0.000 (0/29) 0.000 (0/29) 0.034 (1/29) Menasalbas 2013 Ene 36.571 100 0 0 0.000 (0/6) 0.167 (1/6) 0.000 (0/6) 0.000 (0/6) 0.000 (0/6) 0.167 (1/6) Menasalbas 2013 Feb 84 36.73 63.26 0 0.032 (1/31) 0.097 (3/31) 0.000 (0/31) 0.000 (0/31) 0.000 (0/31) 0.129 (4/31) Menasalbas 2013 Mar 130.285 0.43 99.57 0 0.047 (2/43) 0.000 (0/43) 0.023 (1/43) 0.023 (1/43) 0.023 (1/43) 0.116 (5/43) Menasalbas 2013 Abr 47.5 0 29.47 70.53 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) Menasalbas 2013 May 0 0 0 0 - - - - - - Menasalbas 2013 Nov 6.666 100 0 0 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) 0.000 (0/5) Menasalbas 2014 Feb 8.667 100 0 0 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) Menasalbas 2014 Mar 12.667 0 100 0 0.000 (0/4) 0.250 (1/4) 0.000 (0/4) 0.000 (0/4) 0.000 (0/4) 0.250 (1/4) Menasalbas 2014 Abr 0.667 0 0 100 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1)
Polán 2005 Abr - - - - 0.000 (0/23) 0.565 (13/23) 0.000 (0/23) 0.174 (4/23) 0.000 (0/23) 0.739 (17/23) Polán 2005 May - - - - - - - - - - Polán 2006 Abr - - - - 0.222 (10/45) 0.000 (0/45) 0.000 (0/45) 0.000 (0/45) 0.044 (2/45) 0.267 (12/45) Polán 2006 May - - - - 0.080 (2/25) 0.000 (0/25) 0.080 (2/25) 0.080 (2/25) 0.000 (0/25) 0.240 (6/25) Polán 2007 Abr - - - - 0.167 (4/24) 0.000 (0/24) 0.000 (0/24) 0.167 (4/24) 0.000 (0/24) 0.458 (11/24) Polán 2007 May - - - - 0.071 (1/14) 0.071 (1/14) 0.000 (0/14) 0.000 (0/14) 0.000 (0/14) 0.143 (2/14)
2008 Feb 1 100 0 0 0.000 (0/1) 1.000 (1/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 1.000 (1/1)
2008 Mar 0 0 0 0 - - - - - -
2008 Abr 0 0 0 0 - - - - - -
2008 May 10 0 100 0 0.170 (2/12) 0.080 (1/12) 0.250 (3/12) 0.670 (8/12) 0.000 (0/12) 0.670 (8/12) 1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.
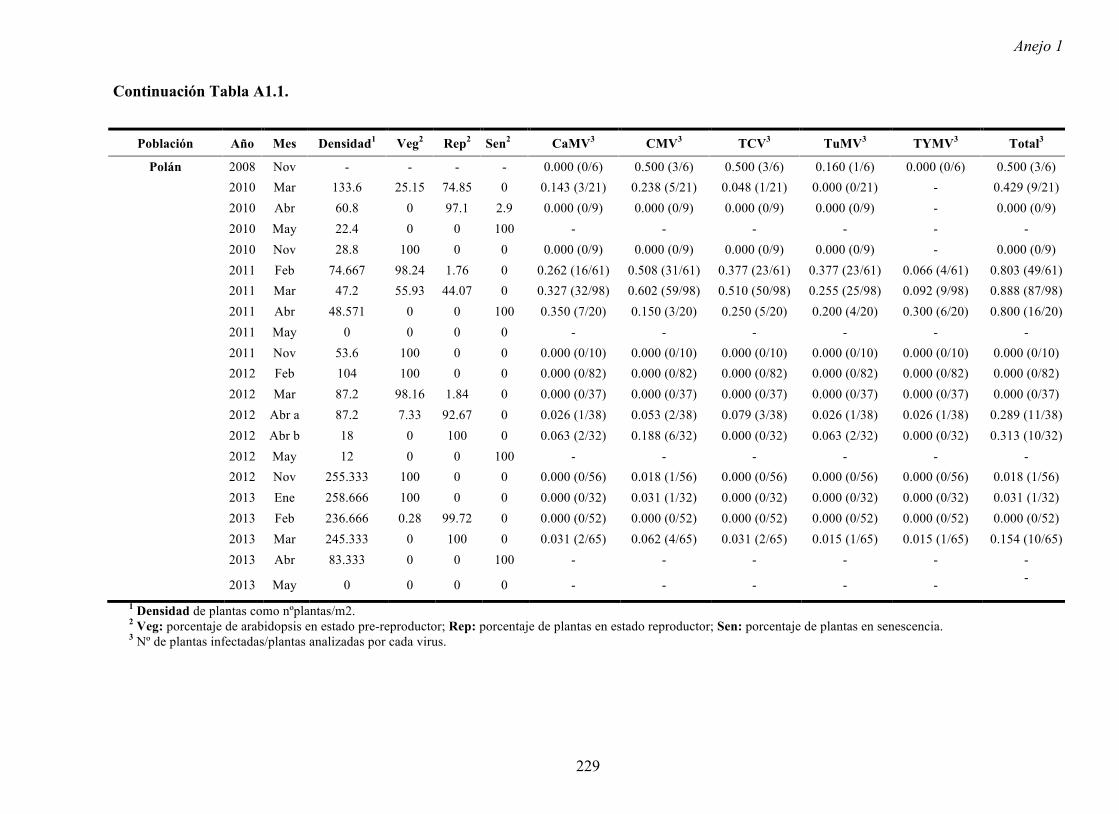
Anejo 1
229
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Polán 2008 Nov - - - - 0.000 (0/6) 0.500 (3/6) 0.500 (3/6) 0.160 (1/6) 0.000 (0/6) 0.500 (3/6) Polán 2010 Mar 133.6 25.15 74.85 0 0.143 (3/21) 0.238 (5/21) 0.048 (1/21) 0.000 (0/21) - 0.429 (9/21) Polán 2010 Abr 60.8 0 97.1 2.9 0.000 (0/9) 0.000 (0/9) 0.000 (0/9) 0.000 (0/9) - 0.000 (0/9) Polán 2010 May 22.4 0 0 100 - - - - - - Polán 2010 Nov 28.8 100 0 0 0.000 (0/9) 0.000 (0/9) 0.000 (0/9) 0.000 (0/9) - 0.000 (0/9) Polán 2011 Feb 74.667 98.24 1.76 0 0.262 (16/61) 0.508 (31/61) 0.377 (23/61) 0.377 (23/61) 0.066 (4/61) 0.803 (49/61) Polán 2011 Mar 47.2 55.93 44.07 0 0.327 (32/98) 0.602 (59/98) 0.510 (50/98) 0.255 (25/98) 0.092 (9/98) 0.888 (87/98) Polán 2011 Abr 48.571 0 0 100 0.350 (7/20) 0.150 (3/20) 0.250 (5/20) 0.200 (4/20) 0.300 (6/20) 0.800 (16/20) Polán 2011 May 0 0 0 0 - - - - - - Polán 2011 Nov 53.6 100 0 0 0.000 (0/10) 0.000 (0/10) 0.000 (0/10) 0.000 (0/10) 0.000 (0/10) 0.000 (0/10) Polán 2012 Feb 104 100 0 0 0.000 (0/82) 0.000 (0/82) 0.000 (0/82) 0.000 (0/82) 0.000 (0/82) 0.000 (0/82) Polán 2012 Mar 87.2 98.16 1.84 0 0.000 (0/37) 0.000 (0/37) 0.000 (0/37) 0.000 (0/37) 0.000 (0/37) 0.000 (0/37) Polán 2012 Abr a 87.2 7.33 92.67 0 0.026 (1/38) 0.053 (2/38) 0.079 (3/38) 0.026 (1/38) 0.026 (1/38) 0.289 (11/38) Polán 2012 Abr b 18 0 100 0 0.063 (2/32) 0.188 (6/32) 0.000 (0/32) 0.063 (2/32) 0.000 (0/32) 0.313 (10/32) Polán 2012 May 12 0 0 100 - - - - - - Polán 2012 Nov 255.333 100 0 0 0.000 (0/56) 0.018 (1/56) 0.000 (0/56) 0.000 (0/56) 0.000 (0/56) 0.018 (1/56) Polán 2013 Ene 258.666 100 0 0 0.000 (0/32) 0.031 (1/32) 0.000 (0/32) 0.000 (0/32) 0.000 (0/32) 0.031 (1/32) Polán 2013 Feb 236.666 0.28 99.72 0 0.000 (0/52) 0.000 (0/52) 0.000 (0/52) 0.000 (0/52) 0.000 (0/52) 0.000 (0/52) Polán 2013 Mar 245.333 0 100 0 0.031 (2/65) 0.062 (4/65) 0.031 (2/65) 0.015 (1/65) 0.015 (1/65) 0.154 (10/65) Polán 2013 Abr 83.333 0 0 100 - - - - - -
Polán 2013 May 0 0 0 0 - - - - - -
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.

Anejo 1
230
Continuación Tabla A1.1.
Población Año Mes Densidad1 Veg2 Rep2 Sen2 CaMV3 CMV3 TCV3 TuMV3 TYMV3 Total3
Polán 2013 Nov 8.666 100 0 0 0.000 (0/6) 0.000 (0/6) 0.000 (0/6) 0.000 (0/6) 0.000 (0/6) 0.000 (0/6) Polán 2014 Feb 46.667 100 0 0 0.026 (1/38) 0.079 (3/38) 0.000 (0/38) 0.000 (0/38) 0.000 (0/38) 0.105 (4/38) Polán 2014 Mar 25.33 0 100 0 0.143 (1/7) 0.286 (2/7) 0.000 (0/7) 0.000 (0/7) 0.143 (1/7) 0.571 (4/7) Polán 2014 Abr 12.667 0 0 100 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1) 0.000 (0/1)
1 Densidad de plantas como nºplantas/m2. 2 Veg: porcentaje de arabidopsis en estado pre-reproductor; Rep: porcentaje de plantas en estado reproductor; Sen: porcentaje de plantas en senescencia. 3 Nº de plantas infectadas/plantas analizadas por cada virus.
Anejo 1
231
Tabla A1.2. Densidad máxima de plantas y VM medio de cada población de arabidopsis por año.
Población Año Densidad máxima1 VM medio2
Ciruelos de Coca 2010 32.800 200.175
2011 9.600 200.932
2012 247.429 12.473.728
2013 175.333 3.091.484
2014 190.667 1.169.073
Las Rozas 2010 186.400 1.294.207
2011 155.200 591.888
2012 194 545.975
2013 104.666 189.455
2014 20.800 282.008
Marjaliza 2010 70.667 461.110
2011 25.600 214.910
2012 25.333 472.185
2013 239.555 1.879.935
2014 207.556 7.200.360
Menasalbas 2010 58.400 926.313
2011 130.400 953.394
2012 65.333 722.377
2013 84 763.598
2014 12.667 134.280
Polán 2010 133.600 274.105
2011 74.667 583.003
2012 255.333 590.452
2013 258.666 437.202
2014 46.667 376.013 1 Densidad máxima de arabidopsis como nºplantas/m2. 2 VM: Coeficiente de distribución espacial estimado como VM=s2/x.
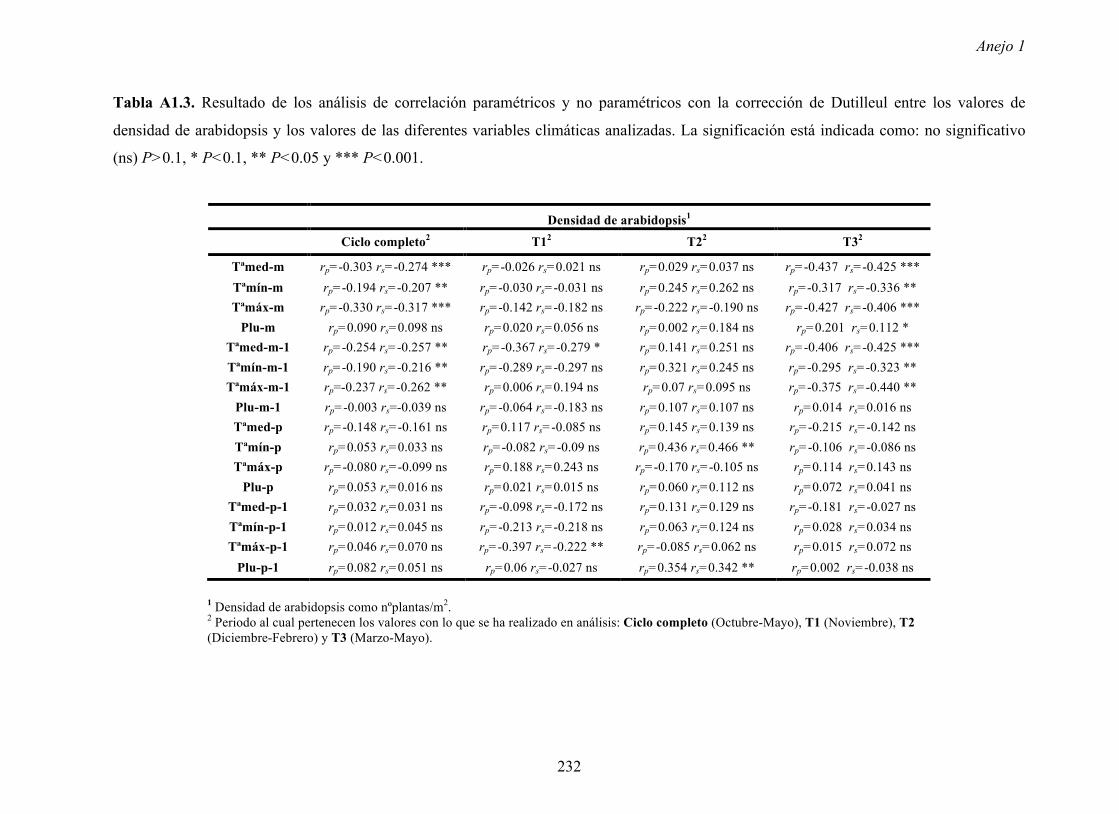
Anejo 1
232
Tabla A1.3. Resultado de los análisis de correlación paramétricos y no paramétricos con la corrección de Dutilleul entre los valores de
densidad de arabidopsis y los valores de las diferentes variables climáticas analizadas. La significación está indicada como: no significativo
(ns) P>0.1, * P<0.1, ** P<0.05 y *** P<0.001.
Densidad de arabidopsis1
Ciclo completo2 T12 T22 T32
Tªmed-m rp=-0.303 rs=-0.274 *** rp=-0.026 rs=0.021 ns rp=0.029 rs=0.037 ns rp=-0.437 rs=-0.425 *** Tªmín-m rp=-0.194 rs=-0.207 ** rp=-0.030 rs=-0.031 ns rp=0.245 rs=0.262 ns rp=-0.317 rs=-0.336 ** Tªmáx-m rp=-0.330 rs=-0.317 *** rp=-0.142 rs=-0.182 ns rp=-0.222 rs=-0.190 ns rp=-0.427 rs=-0.406 ***
Plu-m rp=0.090 rs=0.098 ns rp=0.020 rs=0.056 ns rp=0.002 rs=0.184 ns rp=0.201 rs=0.112 * Tªmed-m-1 rp=-0.254 rs=-0.257 ** rp=-0.367 rs=-0.279 * rp=0.141 rs=0.251 ns rp=-0.406 rs=-0.425 *** Tªmín-m-1 rp=-0.190 rs=-0.216 ** rp=-0.289 rs=-0.297 ns rp=0.321 rs=0.245 ns rp=-0.295 rs=-0.323 ** Tªmáx-m-1 rp=-0.237 rs=-0.262 ** rp=0.006 rs=0.194 ns rp=0.07 rs=0.095 ns rp=-0.375 rs=-0.440 **
Plu-m-1 rp=-0.003 rs=-0.039 ns rp=-0.064 rs=-0.183 ns rp=0.107 rs=0.107 ns rp=0.014 rs=0.016 ns Tªmed-p rp=-0.148 rs=-0.161 ns rp=0.117 rs=-0.085 ns rp=0.145 rs=0.139 ns rp=-0.215 rs=-0.142 ns Tªmín-p rp=0.053 rs=0.033 ns rp=-0.082 rs=-0.09 ns rp=0.436 rs=0.466 ** rp=-0.106 rs=-0.086 ns Tªmáx-p rp=-0.080 rs=-0.099 ns rp=0.188 rs=0.243 ns rp=-0.170 rs=-0.105 ns rp=0.114 rs=0.143 ns
Plu-p rp=0.053 rs=0.016 ns rp=0.021 rs=0.015 ns rp=0.060 rs=0.112 ns rp=0.072 rs=0.041 ns Tªmed-p-1 rp=0.032 rs=0.031 ns rp=-0.098 rs=-0.172 ns rp=0.131 rs=0.129 ns rp=-0.181 rs=-0.027 ns Tªmín-p-1 rp=0.012 rs=0.045 ns rp=-0.213 rs=-0.218 ns rp=0.063 rs=0.124 ns rp=0.028 rs=0.034 ns Tªmáx-p-1 rp=0.046 rs=0.070 ns rp=-0.397 rs=-0.222 ** rp=-0.085 rs=0.062 ns rp=0.015 rs=0.072 ns
Plu-p-1 rp=0.082 rs=0.051 ns rp=0.06 rs=-0.027 ns rp=0.354 rs=0.342 ** rp=0.002 rs=-0.038 ns
1 Densidad de arabidopsis como nºplantas/m2. 2 Periodo al cual pertenecen los valores con lo que se ha realizado en análisis: Ciclo completo (Octubre-Mayo), T1 (Noviembre), T2 (Diciembre-Febrero) y T3 (Marzo-Mayo).

Anejo 1
233
Figura A1.1. Correlaciones considerando el conjunto de todas las poblaciones entre los
valores de densidad de arabidopsis y los de las diferentes variables climáticas pertenecientes a
periodo T1 (Noviembre) de los años 2010, 2011, 2012, 2013 y 2014 de todas las poblaciones
analizadas.

Anejo 1
234
Figura A1.2. Correlaciones considerando el conjunto de todas las poblaciones entre los
valores de densidad de arabidopsis y los de la temperatura máxima correspondiente al periodo
anterior a T1 (Septiembre y Octubre) de los años 2010, 2011, 2012, 2013 y 2014.
Figura A1.4. Correlaciones considerando el conjunto de todas las poblaciones entre los
valores de densidad de arabidopsis y los de las diferentes variables climáticas del periodo T2
(Diciembre-Febrero) de los años 2010, 2011, 2012, 2013 y 2014.

Anejo 1
235
Figura A1.5. Correlaciones considerando el conjunto de todas las poblaciones entre los
valores de densidad de arabidopsis y los de las diferentes variables climáticas del periodo T2
(Diciembre-Febrero) de los años 2010, 2011, 2012, 2013 y 2014.

Anejo 1
236
Tabla A1.4. Modelos obtenidos a partir del análisis SAR considerando todas las poblaciones
entre los valores de densidad de plantas y los de las diferentes variables climáticas en
diferentes periodos del ciclo de vida de las plantas. La significación está indicada como: no
significativo (ns) P>0.1, * P<0.1, ** P<0.05 y *** P>0.001.
Ciclo completo1 T11 T21 T31
Tªmed-m -0.226 ns 1.167 ns 0.170 ns Tªmín-m 0.235 ns -2.195 ns 0.094 ns Tªmáx-m -0.290 ns 0.510 ** -0.440 * Plu-m 0.021 ns 2.434 ns 0.131 ns Tªmed-m-1 -0.562 ns -2.180 ns 0.118 ns Tªmín-m-1 0.211 ns -0.484 ns -0.074 ns Tªmáx-m-1 0.086 ns 2.457 ns -0.404 ns Plu-m-1 -0.026 ns 2.198 ns 0.015 ns Tªmed-p -0.373 ns 4.249 ns -0.325 * Tªmín-p -0.061 ns 1.544 ns -0.328 * Tªmáx-p 0.714 ns -3.531 ns 0.472 *** Plu-p 0.067 ns -2.197 ns 0.073 ns Tªmed-p-1 0.439 ns -1.613 ** -0.505 ** Tªmín-p-1 -0.055 ns 0.526 ns 0.687 *** Tªmáx-p-1 -0.140 ns -1.085 ** 0.535 ** Plu-p-1 0.049 ns -1.472 ns -0.157 ns
R2 0.217 0.879 0.542 0.517 F 1.833 ** 3.551 ** 0.570 ns 3.751 ***
1 Periodo al cual pertenecen los valores con lo que se ha realizado en análisis: Ciclo completo (Octubre-Mayo), T1 (Noviembre), T2 (Diciembre-Febrero) y T3 (Marzo-Mayo).
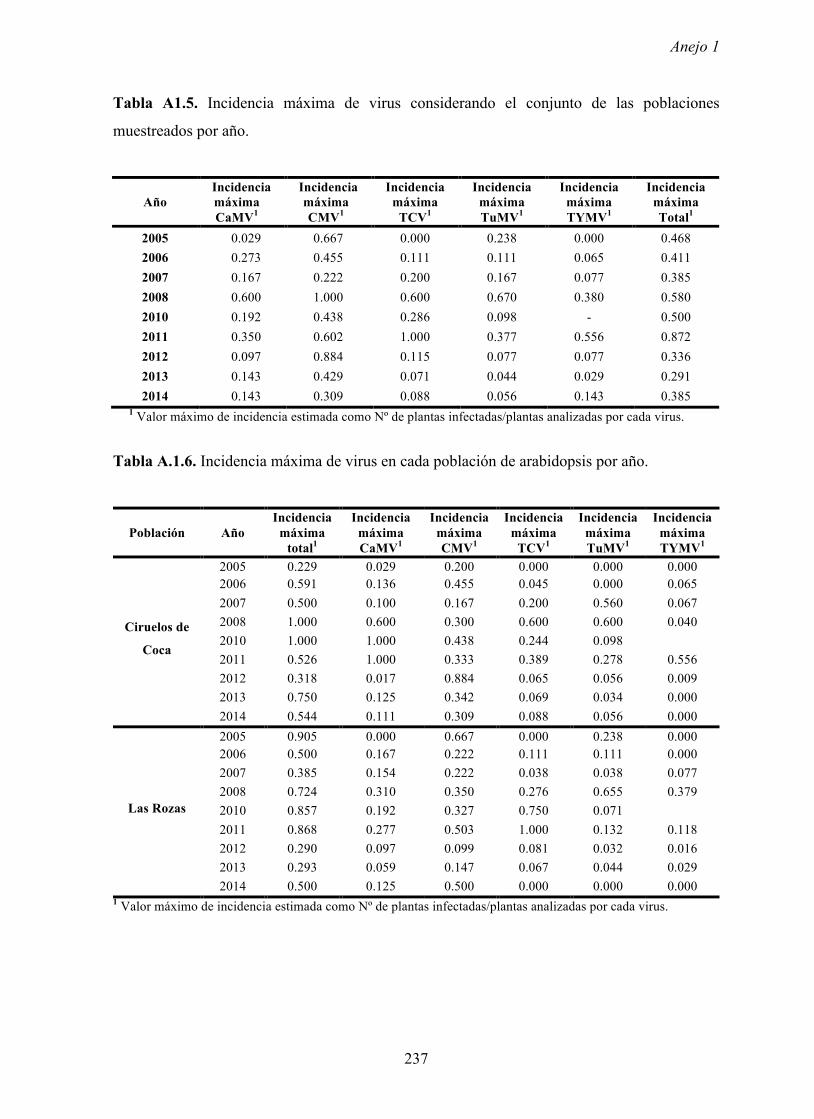
Anejo 1
237
Tabla A1.5. Incidencia máxima de virus considerando el conjunto de las poblaciones
muestreados por año.
Año Incidencia máxima CaMV1
Incidencia máxima CMV1
Incidencia máxima
TCV1
Incidencia máxima TuMV1
Incidencia máxima TYMV1
Incidencia máxima Total1
2005 0.029 0.667 0.000 0.238 0.000 0.468 2006 0.273 0.455 0.111 0.111 0.065 0.411 2007 0.167 0.222 0.200 0.167 0.077 0.385 2008 0.600 1.000 0.600 0.670 0.380 0.580 2010 0.192 0.438 0.286 0.098 - 0.500 2011 0.350 0.602 1.000 0.377 0.556 0.872 2012 0.097 0.884 0.115 0.077 0.077 0.336 2013 0.143 0.429 0.071 0.044 0.029 0.291 2014 0.143 0.309 0.088 0.056 0.143 0.385
1 Valor máximo de incidencia estimada como Nº de plantas infectadas/plantas analizadas por cada virus.
Tabla A.1.6. Incidencia máxima de virus en cada población de arabidopsis por año.
Población Año Incidencia
máxima total1
Incidencia máxima CaMV1
Incidencia máxima CMV1
Incidencia máxima
TCV1
Incidencia máxima TuMV1
Incidencia máxima TYMV1
Ciruelos de
Coca
2005 0.229 0.029 0.200 0.000 0.000 0.000 2006 0.591 0.136 0.455 0.045 0.000 0.065 2007 0.500 0.100 0.167 0.200 0.560 0.067 2008 1.000 0.600 0.300 0.600 0.600 0.040 2010 1.000 1.000 0.438 0.244 0.098 2011 0.526 1.000 0.333 0.389 0.278 0.556 2012 0.318 0.017 0.884 0.065 0.056 0.009 2013 0.750 0.125 0.342 0.069 0.034 0.000 2014 0.544 0.111 0.309 0.088 0.056 0.000
Las Rozas
2005 0.905 0.000 0.667 0.000 0.238 0.000 2006 0.500 0.167 0.222 0.111 0.111 0.000 2007 0.385 0.154 0.222 0.038 0.038 0.077 2008 0.724 0.310 0.350 0.276 0.655 0.379 2010 0.857 0.192 0.327 0.750 0.071 2011 0.868 0.277 0.503 1.000 0.132 0.118 2012 0.290 0.097 0.099 0.081 0.032 0.016 2013 0.293 0.059 0.147 0.067 0.044 0.029 2014 0.500 0.125 0.500 0.000 0.000 0.000
1 Valor máximo de incidencia estimada como Nº de plantas infectadas/plantas analizadas por cada virus.
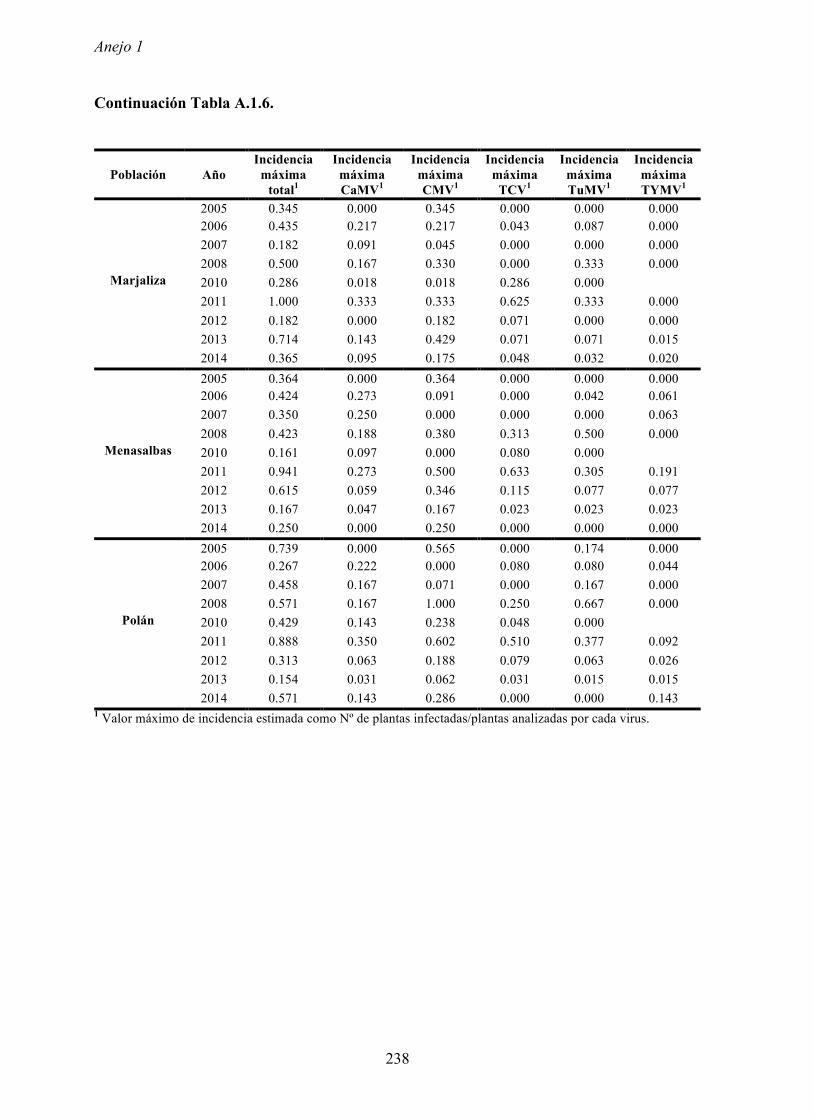
Anejo 1
238
Continuación Tabla A.1.6.
Población Año Incidencia
máxima total1
Incidencia máxima CaMV1
Incidencia máxima CMV1
Incidencia máxima
TCV1
Incidencia máxima TuMV1
Incidencia máxima TYMV1
Marjaliza
2005 0.345 0.000 0.345 0.000 0.000 0.000 2006 0.435 0.217 0.217 0.043 0.087 0.000 2007 0.182 0.091 0.045 0.000 0.000 0.000 2008 0.500 0.167 0.330 0.000 0.333 0.000 2010 0.286 0.018 0.018 0.286 0.000 2011 1.000 0.333 0.333 0.625 0.333 0.000 2012 0.182 0.000 0.182 0.071 0.000 0.000 2013 0.714 0.143 0.429 0.071 0.071 0.015 2014 0.365 0.095 0.175 0.048 0.032 0.020
Menasalbas
2005 0.364 0.000 0.364 0.000 0.000 0.000 2006 0.424 0.273 0.091 0.000 0.042 0.061 2007 0.350 0.250 0.000 0.000 0.000 0.063 2008 0.423 0.188 0.380 0.313 0.500 0.000 2010 0.161 0.097 0.000 0.080 0.000 2011 0.941 0.273 0.500 0.633 0.305 0.191 2012 0.615 0.059 0.346 0.115 0.077 0.077 2013 0.167 0.047 0.167 0.023 0.023 0.023 2014 0.250 0.000 0.250 0.000 0.000 0.000
Polán
2005 0.739 0.000 0.565 0.000 0.174 0.000 2006 0.267 0.222 0.000 0.080 0.080 0.044 2007 0.458 0.167 0.071 0.000 0.167 0.000 2008 0.571 0.167 1.000 0.250 0.667 0.000 2010 0.429 0.143 0.238 0.048 0.000 2011 0.888 0.350 0.602 0.510 0.377 0.092 2012 0.313 0.063 0.188 0.079 0.063 0.026 2013 0.154 0.031 0.062 0.031 0.015 0.015 2014 0.571 0.143 0.286 0.000 0.000 0.143
1 Valor máximo de incidencia estimada como Nº de plantas infectadas/plantas analizadas por cada virus.
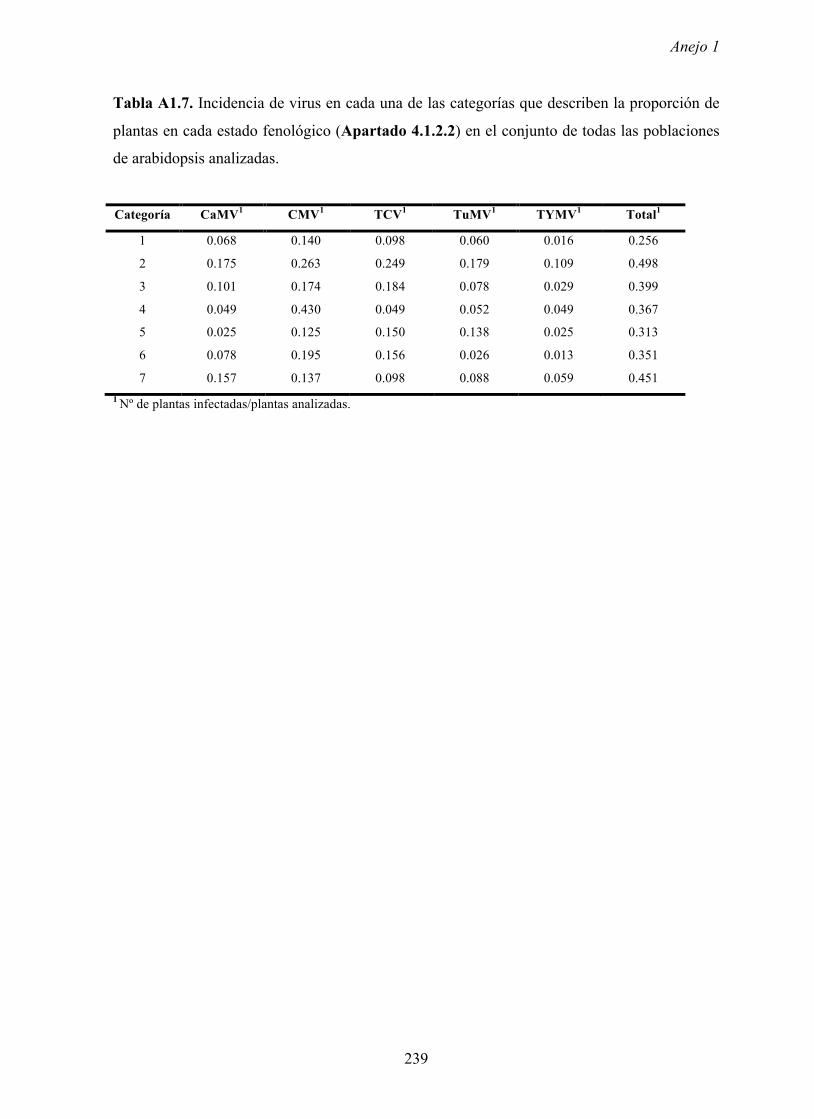
Anejo 1
239
Tabla A1.7. Incidencia de virus en cada una de las categorías que describen la proporción de
plantas en cada estado fenológico (Apartado 4.1.2.2) en el conjunto de todas las poblaciones
de arabidopsis analizadas.
Categoría CaMV1 CMV1 TCV1 TuMV1 TYMV1 Total1
1 0.068 0.140 0.098 0.060 0.016 0.256
2 0.175 0.263 0.249 0.179 0.109 0.498
3 0.101 0.174 0.184 0.078 0.029 0.399
4 0.049 0.430 0.049 0.052 0.049 0.367
5 0.025 0.125 0.150 0.138 0.025 0.313
6 0.078 0.195 0.156 0.026 0.013 0.351
7 0.157 0.137 0.098 0.088 0.059 0.451 1 Nº de plantas infectadas/plantas analizadas.

Anejo 1
240
Figura A1.6. Correlaciones considerando el conjunto de todas las poblaciones entre los
valores de densidad de plantas y los de incidencia de cada virus de los años 2010, 2011, 2012,
2013 y 2014.

Anejo 1
241
Tabla A1.8. Resultado de los análisis de correlación paramétrica y no paramétrica con la corrección de Detilleul considerando el conjunto de
las poblaciones entre los valores incidencia máxima de CaMV, CMV, TCV, TuMV, TYMV y total, y los de las variables climáticas
(Apartado 4.1.1 Tabla 4.1.1) de los años 2005-2008 y 2010-2014 en el periodo T2 (Diciembre-Febrero). La significación está indicada
como: no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y *** P>0.001.
Incidencia máxima
Total Incidencia máxima
CaMV Incidencia máxima
CMV Incidencia máxima
TCV Incidencia máxima
TuMV
Tª med-m rp=0.387 rs=0.352** ns rp=0.404 rs=0.458 ** ns rp=0.496 rs=0.337 ** Tª mín-m rp=0.354 rs=0.371* ns rp=0.382 rs=0.489 ** ns ns Tª máx-m ns ns ns rp=0.384 rs=0.128 * rp=0.436 rs=0.216 ** Plu-m ns ns ns ns ns Tª med-m-1 rp=-0.360 rs=-0.450* ns ns ns ns Tª mín-m-1 rp=-0.641 rs=-0.581*** rp=-0.596 rs=-0.432 ** rp=-0.311 rs=-0.327 * ns ns Tªmáx-m-1 ns ns ns ns ns Plu-m-1 ns ns ns ns ns Tª med-p ns rp=-0.460 rs=-0.528 ** ns ns ns Tª mín-p ns ns ns ns ns Tª máx-p ns ns ns ns ns Plu-p ns ns ns ns ns Tª med-p-1 rp=-0.349 rs=-0.449** rp=-0.409 rs=-0.456 * ns ns ns Tª mín-p-1 ns ns ns ns ns Tª máx-p-1 ns ns ns ns ns Plu-p-1 ns ns ns ns ns

Anejo 1
242
Tabla A1.9. Modelos obtenidos a partir del análisis SAR considerando el conjunto de todas
las poblaciones entre los valores de incidencia máxima de CaMV, CMV, TCV, TuMV,
TYMV y los de las diferentes las diferentes variables climáticas (Apartado 4.1.1 Tabla 4.1.1)
del periodo T2 (Diciembre-Febrero) de los años 2005-2008 y 2010-2014. La significación está
indicada como: no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y *** P>0.001.
T2 Incidencia máxima total
Incidencia máxima CaMV
Incidencia máxima CMV
Incidencia máxima TCV
Incidencia máxima TuMV
Tªmed-m 0.801 ns Tªmín-m 0.484 * Tªmáx-m -1.515 ** Plu-m -0.056 ns Tªmed-m-1 0.940 ns Tªmín-m-1 -1.027 ** Tªmáx-m-1 -0.788 ns Plu-m-1 2.831 ** Tªmed-p 0.114 ns Tªmín-p -1.464 ** Tªmáx-p 0.720 ns Plu-p -2.158 ** Tªmed-p-1 -0.801 * Tªmín-p-1 0.849 ** Tªmáx-p-1 0.574 ** Plu-p-1 -0.171 ns R2 0.884 0.908 0.519 0.754 0.710 F 2.476 ns 3.693 * 0.397 ns 1.068 ns 0.971 ns

Anejo 1
243
Figura A1.7. Correlaciones considerando el conjunto de todas las poblaciones entre los
valores de incidencia máxima total (�) y los de diferentes variables climáticas (Apartado 4.1.1
Tabla 4.1.1) del periodo T2 (Diciembre-Febrero) de los años 2005-2008 y 2010-2014.
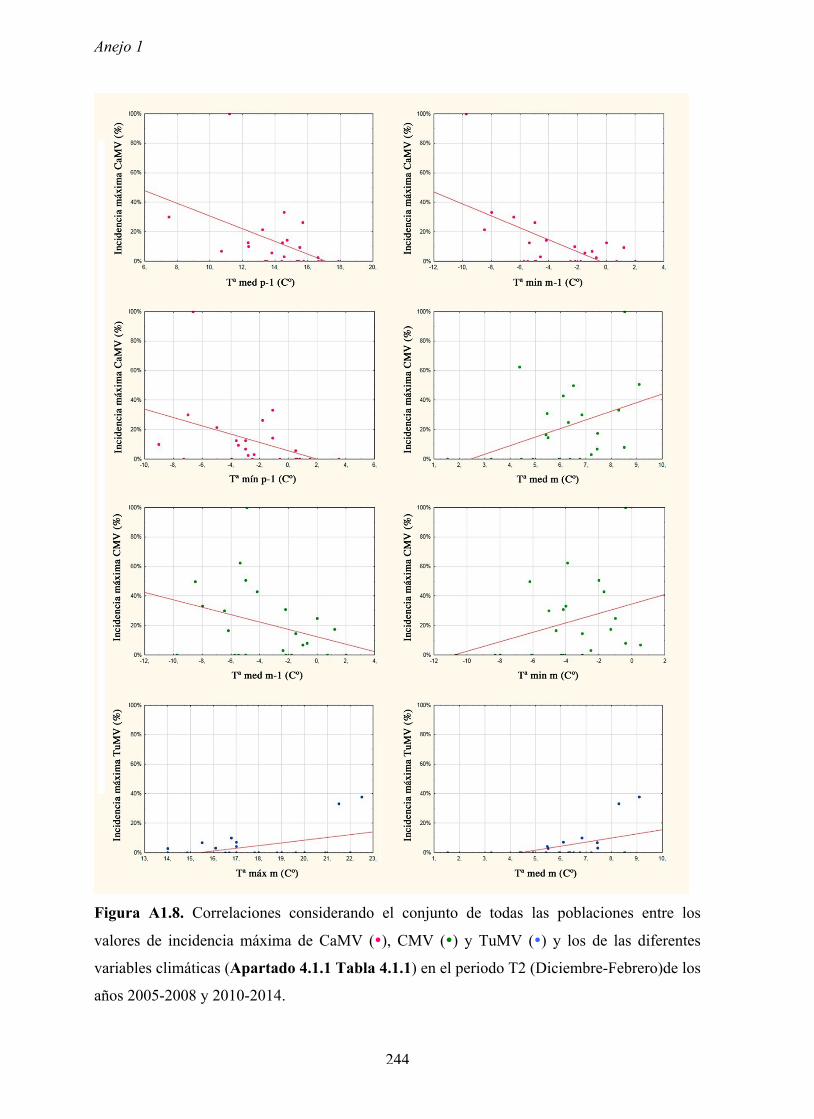
Anejo 1
244
Figura A1.8. Correlaciones considerando el conjunto de todas las poblaciones entre los
valores de incidencia máxima de CaMV (�), CMV (�) y TuMV (�) y los de las diferentes
variables climáticas (Apartado 4.1.1 Tabla 4.1.1) en el periodo T2 (Diciembre-Febrero)de los
años 2005-2008 y 2010-2014.

Anejo 1
245
Tabla A1.10. Resultado de los análisis de correlación paramétrico y no paramétrico con la corrección de Detilleul considerando el conjunto de
las poblaciones entre los valores de incidencia máxima de CaMV, CMV, TCV, TuMV y TYMV y total y los de las variables climáticas
(Apartado 4.1.1 Tabla 4.1.1) del periodo T3 (Marzo-Mayo) de los años 2005-2008 y 2010-2014. La significación está indicada como: no
significativa (ns) P>0.1, * P<0.1, ** P<0.05 y *** P>0.001.
Incidencia máxima Total
Incidencia máxima CaMV
Incidencia máxima CMV
Incidencia máxima TCV
Incidencia máxima TuMV
Tªmed-m ns ns ns rp=-0.249, rs=-0.254 * ns
Tªmín-m ns ns ns rp=-0.248, rs=-0.130 * ns
Tªmáx-m ns ns ns ns ns
Plu-m ns ns ns ns ns
Tªmed-m-1 ns rp=0.257, rs=0.295 * ns ns ns
Tªmín-m-1 ns ns ns ns ns
Tªmáx-m-1 ns rp=0.314, rs=-0.289 ** ns ns ns
Plu-m-1 ns ns ns ns ns
Tªmed-p ns ns ns ns ns
Tªmín-p ns ns rp=-0.245, rs=-0.206 * ns ns
Tªmáx-p rp=0.258, rs=-0.020 * ns ns ns ns
Plu-p ns ns ns ns ns
Tªmed-p-1 ns ns rp=-0.364 , rs=-0.246 * ns ns
Tªmín-p-1 ns ns ns ns ns
Tªmáx-p-1 ns ns ns ns ns
Plu-p-1 ns ns ns ns ns

Anejo 2
246
Tabla A1.11. Modelos obtenidos a partir del análisis SAR considerando el conjunto de todas
las poblaciones entre los valores de incidencia máxima de CaMV, CMV, TCV, TuMV,
TYMV y total y las diferentes variables climáticas (Apartado 4.1.1 Tabla 4.1.1) del periodo
T3 (Marzo-Mayo) de los años 2005-2008 y 2010-2014. La significación está indicada como:
no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y *** P>0.001.
Incidencia
máxima total
Incidencia máxima CaMV
Incidencia máxima
CMV
Incidencia máxima
TCV
Incidencia máxima TuMV
Tªmed-m -1.06 ** -1.683 ** -1.183 ** Tªmín-m -0.085 ns -0.354 ns -0.174 ns Tªmáx-m 0.798 ** 1.137 ** 0.756 ** Plu-m -0.481 ** 0.321 ns -0.397 * Tªmed-m-1 0.566 ns 0.829 ns -0.416 ns Tªmín-m-1 0.106 ns 0.488 * 0.615 ** Tªmáx-m-1 -0.011 ns -0.143 ns 0.746 ** Plu-m-1 -0.179 ns -0.089 ns -0.216 * Tªmed-p 1.207 ** 0.699 ** 0.294 ns Tªmín-p -0.169 ns -0.270 ns -0.059 ns Tªmáx-p -1.373 ** -0.373 ns -0.541 * Plu-p -0.058 ns -0.243 ns 0.031 ns Tªmed-p-1 -0.812 ** -0.599 ** -0.007 ns Tªmín-p-1 0.593 ** 0.349 ns 0.125 ns Tªmáx-p-1 0.966 ** 0.091 ns 0.229 ns Plu-p-1 0.363 ** 0.112 ns 0.459 ** R2 0.562 0.358 0.514 0.626 0.371 F 2.159 ** 0.944 ns 1.785 * 2.727 ** 0.936
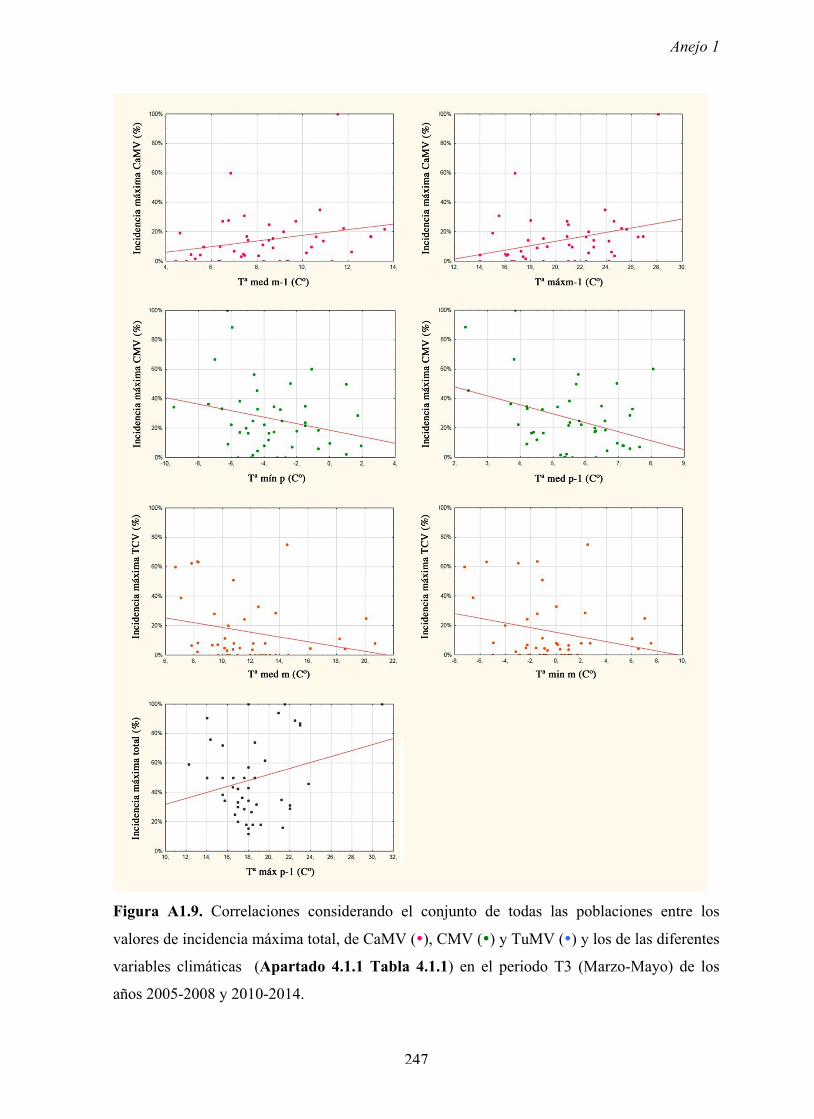
Anejo 1
247
Figura A1.9. Correlaciones considerando el conjunto de todas las poblaciones entre los
valores de incidencia máxima total, de CaMV (�), CMV (�) y TuMV (�) y los de las diferentes
variables climáticas (Apartado 4.1.1 Tabla 4.1.1) en el periodo T3 (Marzo-Mayo) de los
años 2005-2008 y 2010-2014.


ANEJO 2

Anejo 2
250

Anejo 2
251
Tabla A2.1. Localización geográfica, altitud y ecosistema de las poblaciones de arabidopsis analizadas en el Apartado 4.2
Población Longitud Latitud Altitud Ecosistema
Aguarón (Agu) 1.340 41.321 1045 Encinar mediterráneo
Bisaurri (Bis) 0.537 42.490 1450 Pradera de montaña
Ciruelos de Coca (Cdc) 4.544 41.215 800 Pinar mediterráneo
Grazalema (Gra) 5.394 36.769 1284 Monte bajo mediterráneo
San Leo nardo de Yagüe (Leo) 3.113 41.797 1010 Llanura aluvial
Marjaliza (Mar) 3.930 39.583 1030 Encinar mediterráneo
Mérida (Mer) 6.336 38.917 238 Área arenosa cercana a muro de piedra
Molacillos (Moc 5.640 41.566 665 Linde de cultivos
Poblet (Pob) 1.027 41.355 677 Bosque mixto de pino y encima
Prádena del Rincón (Pra) 3.538 41.052 1106 Zona de rivera
Quintela (Qui) 6.929 42.693 700 Borde de camino
Santa Elena (San) 3.508 38.333 700 Encinar mediterráneo

Anejo 2
252
Tabla A2.2. Poblaciones, individuos y haplotipos de arabidopsis
Población Agu Bis Cdc Gra Leo Mar Mer Moc Pob Pra Qui San
Individuo
Agu 0 1-6 Bis 0 1-6 Cdc 1 2, Gra 0 1-6 Leo 1 1-4 Mar 2 2 Mer 0 1-6 Moc 0 1-6 Pob 1 2, 5 Pra 0 1-4 Qui 0 1-6 San 0 1-6 Agu 1 1-4 Bis 1 1-4 Cdc 3 2, 5 Gra 1 1-6 Leo 2 1-4 Mar 6 2, 5 Mer 1 1, 4 Moc 1 1-6 Pob 2 1-6 Pra 1 1-4 Qui 1 1-6 San 2 1, 4 Agu 2 1-6 Bis 2 1-6 Cdc 7 2, Gra 2 1-6 Leo 3 1-4 Mar 8 2, 5 Mer 3 1-6 Moc 2 1-4 Pob 3 1-6 Pra 2 1-4 Qui 2 1-6 San 4 1-6 Agu 4 1, 3, 4 Bis 3 1-6 Cdc 10 2, Gra 3 1-6 Leo 4 1-4 Mar 102, 5 Mer 5 1-6 Moc 5 1-4 Pob 4 1-4 Pra 3 1-4 Qui 3 1-6 San 5 1-6 Agu 6 1-4 Bis 4 1-6 Cdc 12 2, 5 Gra 4 1-6 Leo 6 1-4 Mar 12 2, 5 Mer 6 1, 4 Moc 7 1-6 Pob 5 1-4 Pra 4 1-4 Qui 4 1-6 San 6 1-6 Agu 7 1-6 Bis 5 1-6 Cdc 13 2 Gra 5 1-6 Leo 7 1-4 Mar 13 2, 5 Mer 7 1, 4 Moc 8 1-6 Pob 6 2, 5 Pra 5 1-4 Qui 5 1-6 San 8 1-6 Agu 8 1-4 Bis 6 1-6 Cdc 1-6 2 Gra 6 1-6 Leo 8 1-4 Mar 15 2, 5 Mer 9 1-4 Moc 9 1-4 Pob 7 1-6 Pra 6 1-4 Qui 6 1-6 San 9 1-6 Agu 11 1-6 Bis 7 1-6 Cdc 22 2, 5 Gra 7 1-6 Leo 9 1-4 Mar 23 2, 5 Mer 10 1-6 Moc 14 1-6 Pob 8 1-4 Pra 7 1-4 Qui 7 1-6 San 10 1-6 Agu 12 1-4 Bis 8 1-4 Cdc 27 2, 5 Gra 8 1-6 Leo 10 1-4 Mar 25 2, 5 Mer 12 1-6 Moc 15 1-4 Pob 9 1-6 Pra 8 1-4 Qui 8 1-6 San 12 1-6 Agu 18 1-4 Bis 9 1-4 Gra 9 1-6 Leo 11 1-4 Mar 29 2 Mer 1-4 1-6 Moc 19 1-4 Pob 10 1-4 Pra 10 1-4 Qui 9 1-6 San 13 1-6
Haplotipo
AGU H11-6 BIS H11-6 CDC H1 2, 5 GRA H11-6 LEO H11-4 MAR H12, 5 MER H11-6 MOC H11-6 POB H11-6 PRA H11-4 QUI H11-6 SAN H11-6 AGU H21-6 BIS H21-4 CDC H2 2 GRA H21-6 LEO H21-4 MAR H2 2, 5 MER H21, 4 MOC H21-6 POB H21-6 PRA H21-4 QUI H21-6 SAN H21, 4 AGUH31, 4 BIS H31-4 CDC H4 2 GRA H31-6 LEO H31-4 MAR H3 2, 5 MOC H31-4 POB H31-6 PRA H31-4 QUI H31-6 SAN H31-6 AGU H41-6 CDC H5 2 GRA H41-6 LEO H41-4 MAR H4 2, 5 MOC H41-6 POB H41-4 PRA H41-4 QUI H41-6 SAN H41-6 AGU H51-6 CDC H6 2, 5 LEO H51-4 MAR H5 2, 5 MOC H51-6 POB H51-4 PRA H51-4 QUI H51-6 SAN H51-6 CDC H7 2, MAR H6 2, 5 MOC H61-4 POB H61-6 PRA H61-4 SAN H61-6 CDC H8 2, 5 MAR H7 2 MOC H71-6 PRA H71-4 SAN H71-6 CDC H9 2, 5 MOC H81-4 PRA H81-4 SAN H81-6 MOC H91-4 PRA H91-4 SAN H91-6
1) Individuo/haplotipo ensayado en el análisis de la resistencia a CMV en el Ambiente 1 2) Individuo/haplotipo ensayado en el análisis de la resistencia a CMV en el Ambiente 2 3) Individuo/haplotipo ensayado en el análisis del efecto del ambiente en la resistencia a CMV 4) Individuo/haplotipo ensayado en el análisis de la tolerancia a CMV en el Ambiente 1 5) Individuo/haplotipo ensayado en el análisis de la tolerancia a CMV en el Ambiente 2 6) Individuo/haplotipo ensayado en el análisis del efecto del ambiente en la tolerancia a CMV

Anejo 2
253
Tabla A2.2a. Individuos de arabidopsis que conforman cada uno de los haplotipos ensayados
en este trabajo.
Población Haplotipo: Individuo
Agu
AGU H1:Agu 0, Agu 1, Agu 6, Agu 8, Agu 12 AGU H2: Agu 2, Agu 18 AGU H3:Agu 4 AGU H4:Agu 7 AGU H5:Agu 11
Bis BIS H1: Bis0, Bis1, Bis2, Bis3, Bis4, Bis5, Bis6, Bis7 BIS H2: Bis8 BIS H3: Bis9
Cdc
CDC H1: Cdc 3, Cdc 13 CDC H2: Cdc 1 CDC H3: Cdc 5 CDC H4: Cdc 7 CDC H5: Cdc 10 CDC H6: Cdc 12 CDC H7: Cdc 16 CDC H8: Cdc 22 CDC H9: Cdc 27
Gra
GRA H1: Gra 0, Gra 1, Gra 3, Gra 4, Gra 8, Gra 9 GRA H2: Gra 2, Gra 7 GRA H3: Gra 5 GRA H4: Gra 6
Leo
LEO H1: Leo 1, Leo 4, Leo 6 LEO H2: Leo 9, Leo 10, Leo 11 LEO H3: Leo 2, Leo 7 LEO H4: Leo 3 LEO H5: Leo 8
Mar
MAR H1: Mar 2, Mar 8, Mar 23 MAR H2: Mar 10, Mar 15 MAR H3: Mar 6 MAR H4: Mar 12 MAR H5: Mar 13 MAR H6: Mar 25 MAR H7: Mar 29

Anejo 2
254
Continuación Tabla A2.2a.
Población Haplotipo: Individuo
Mer MER H1: Mer 0, Mer 3, Mer 5, Mer 7, Mer 9, Mer 10, Mer 12, Mer 14 MER H2: Mer 1, Mer 6
Moc
MOC H1: Moc 0, Moc 2 MOC H2: Moc 1 MOC H3: Moc 5 MOC H4: Moc 7 MOC H5: Moc 8 MOC H6: Moc 9 MOC H7: Moc 14 MOC H8: Moc 15 MOC H9: Moc 19
Pob
POB H1: Pob 1, Pob 3 POB H2: Pob 9, Pob 10 POB H3: Pob 6, Pob 7 POB H4: Pob 4 POB H5: Pob 8 POB H6: Pob 2 POB10: Pob 5
Pra
PRA H1: Pra 5, Pra 6 PRA H2: Pra 0 PRA H3: Pra 1 PRA H4: Pra 2 PRA H5: Pra 3 PRA H6: Pra 4 PRA H7: Pra 7 PRA H8: Pra 8 PRA H9: Pra 10
Qui
QUI H1: Qui 0, Qui 3, Qui 6, Qui 9 QUI H2: Qui 1, Qui 4, Qui 5 QUI H3: Qui 2 QUI H4: Qui 7 QUI H5: Qui 8
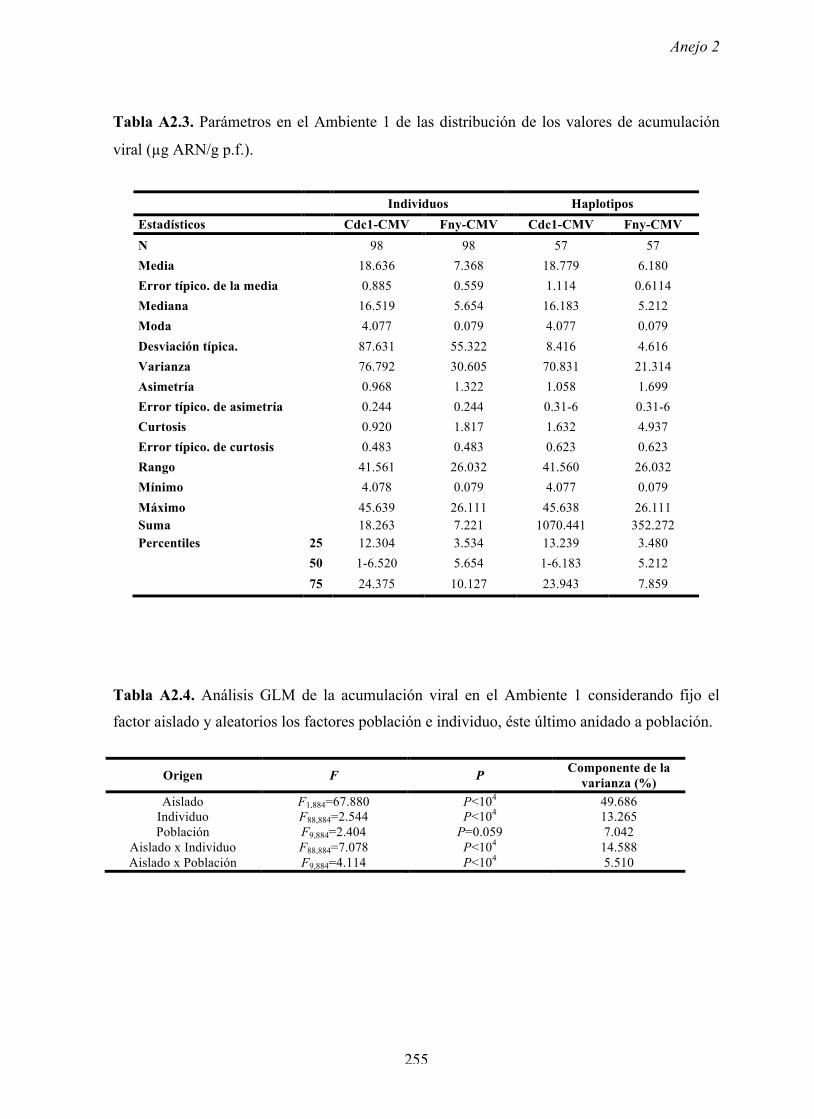
Anejo 2
255
Tabla A2.3. Parámetros en el Ambiente 1 de las distribución de los valores de acumulación
viral (µg ARN/g p.f.).
Individuos Haplotipos Estadísticos Cdc1-CMV Fny-CMV Cdc1-CMV Fny-CMV N 98 98 57 57 Media 18.636 7.368 18.779 6.180 Error típico. de la media 0.885 0.559 1.114 0.6114 Mediana 16.519 5.654 16.183 5.212 Moda 4.077 0.079 4.077 0.079 Desviación típica. 87.631 55.322 8.416 4.616 Varianza 76.792 30.605 70.831 21.314 Asimetría 0.968 1.322 1.058 1.699 Error típico. de asimetría 0.244 0.244 0.31-6 0.31-6 Curtosis 0.920 1.817 1.632 4.937 Error típico. de curtosis 0.483 0.483 0.623 0.623 Rango 41.561 26.032 41.560 26.032 Mínimo 4.078 0.079 4.077 0.079 Máximo 45.639 26.111 45.638 26.111 Suma 18.263 7.221 1070.441 352.272 Percentiles 25 12.304 3.534 13.239 3.480 50 1-6.520 5.654 1-6.183 5.212 75 24.375 10.127 23.943 7.859
Tabla A2.4. Análisis GLM de la acumulación viral en el Ambiente 1 considerando fijo el
factor aislado y aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,884=67.880 P<104 49.686 Individuo F88,884=2.544 P<104 13.265 Población F9,884=2.404 P=0.059 7.042
Aislado x Individuo F88,884=7.078 P<104 14.588 Aislado x Población F9,884=4.114 P<104 5.510

Anejo 2
256
Tabla A2.5. Análisis GLM de la acumulación viral en el Ambiente 1 considerando fijo el
factor aislado y aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,884=70.859 P<104 49.627 Haplotipo F47,884=2.473 P=0.001 8.761 Población F9,884=2.268 P=0.070 6.986
Aislado x Haplotipo F47,884=3.136 P<104 7.814 Aislado x Población F9,884=3.241 P=0.004 4.763
Tabla A2.6. Análisis GLM en el Ambiente 1 de la acumulación de Cdc1-CMV considerando
aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F88,453=11.583 P<104 52.901 Población F9,453=4.938 P<104 23.984
Tabla A2.7. Análisis GLM en el Ambiente 1 de la acumulación de Fny-CMV considerando
aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F88,431=17.850 P<104 58.813 Población F9,431=4.984 P<104 26.090
Tabla A2.8. Análisis GLM en el Ambiente 1 de la acumulación de Cdc1-CMV considerando
aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F47,453=5.629 P<104 34.242 Población F9,453=4.119 P=0.001 23.992
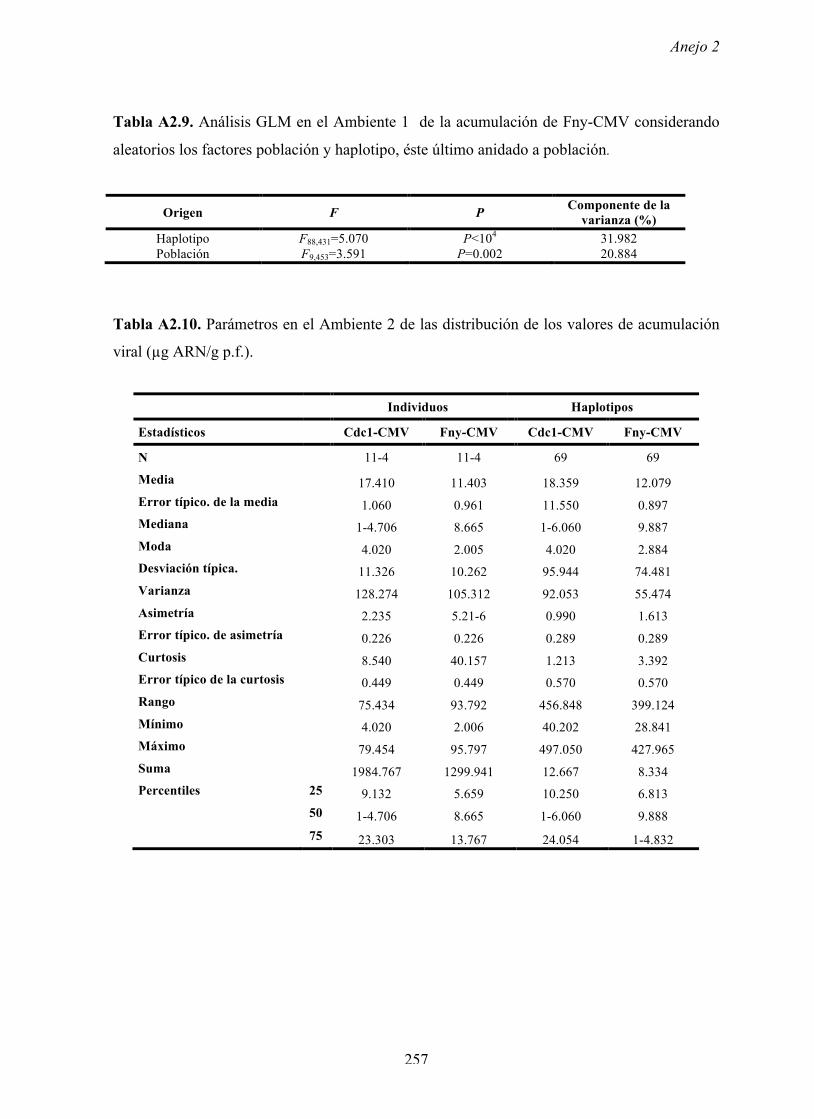
Anejo 2
257
Tabla A2.9. Análisis GLM en el Ambiente 1 de la acumulación de Fny-CMV considerando
aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F88,431=5.070 P<104 31.982 Población F9,453=3.591 P=0.002 20.884
Tabla A2.10. Parámetros en el Ambiente 2 de las distribución de los valores de acumulación
viral (µg ARN/g p.f.).
Individuos Haplotipos
Estadísticos Cdc1-CMV Fny-CMV Cdc1-CMV Fny-CMV
N 11-4 11-4 69 69
Media 17.410 11.403 18.359 12.079 Error típico. de la media 1.060 0.961 11.550 0.897 Mediana 1-4.706 8.665 1-6.060 9.887 Moda 4.020 2.005 4.020 2.884 Desviación típica. 11.326 10.262 95.944 74.481 Varianza 128.274 105.312 92.053 55.474 Asimetría 2.235 5.21-6 0.990 1.613 Error típico. de asimetría 0.226 0.226 0.289 0.289 Curtosis 8.540 40.157 1.213 3.392 Error típico de la curtosis 0.449 0.449 0.570 0.570 Rango 75.434 93.792 456.848 399.124 Mínimo 4.020 2.006 40.202 28.841 Máximo 79.454 95.797 497.050 427.965 Suma 1984.767 1299.941 12.667 8.334 Percentiles 25 9.132 5.659 10.250 6.813 50 1-4.706 8.665 1-6.060 9.888 75 23.303 13.767 24.054 1-4.832

Anejo 2
258
Tabla A2.11. Análisis GLM de la acumulación viral en el Ambiente 2 considerando fijo el
factor aislado y aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,1-459= 42.529 P < 104 7.814 Individuo F102,1-459 = 7.921 P < 104 30.635 Población F11,1-459= 4.015 P < 104 12.996
Aislado x Individuo F102,1-459= 1.479 P = 0.026 3.506 Aislado x Población F11,1-459= 2.102 P = 0.002 1.180
Tabla A2.12. Análisis GLM de la acumulación viral en el Ambiente 2 considerando fijo el
factor aislado y aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,1-459= 37.859 P < 104 7.421 Haplotipo F58,1-459= 8.479 P < 104 15.795 Población F11,1-459= 3.31-6 P = 0.002 11.470
Aislado x Haplotipo F58,1-459 = 0.665 P = 0.975 0 Aislado x Población F11,1-459= 2.887 P = 0.007 1.079
Tabla A2.13. Análisis GLM de la acumulación de Cdc1-CMV en el Ambiente 2 considerando
aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F102,733= 4.197 P < 104 23.319 Población F11,733 = 5.889 P < 104 17.206
Tabla A2.14. Análisis GLM de la acumulación de Fny-CMV en el Ambiente 2 considerando
aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F102,726= 1-4.21-4 P < 104 56.363 Población F11,726 = 2.849 P =0.003 12.201

Anejo 2
259
Tabla A2.15. Análisis GLM de la acumulación de Cdc1-CMV en el Ambiente 2 considerando
aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F57,733 = 2.888 P < 104 15.903 Población F11,733 = 4.51-4 P < 104 13.227
Tabla A2.16. Análisis GLM de la acumulación de Fny-CMV en el Ambiente 2 considerando
aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F57,726= 3.992 P < 104 19.899 Población F11,726 = 32.548 P = 0.011 10.322
Tabla A2.17. Análisis GLM del efecto del ambiente en la acumulación viral considerando
fijos los factores aislado y ambiente y aleatorios los factores población y individuo, éste
último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,1984 = 97.986 P < 104 35.643 Individuo F83,1984 = 1.304 P = 0.133 2.445 Población F9,1984 = 0.318 P = 0.949 0 Ambiente F1,1984 = 0.036 P = 0.854 0
Aislado x Individuo F83,1984 = 1.019 P = 0.466 0 Aislado x Población F9,1984 = 1.051 P = 0.479 0 Ambiente x Aislado F1,1984 = 8.324 P = 0.018 2.604
Ambiente x Individuo F83,1984 = 2.643 P < 104 11.228 Ambiente x Población F9,1984 = 5.958 P = 0.001 10.803 Aislado x Ambiente
Individuo F83,1984 = 2.123 P < 104 6.917
Aislado x Ambiente Población F9,1984 = 3.907 P < 104 4.521

Anejo 2
260
Tabla A2.18. Análisis GLM del efecto del ambiente en la acumulación de Cdc1-CMV
considerando fijos los factores aislado y ambiente y aleatorios los factores población e
individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F83,906 = 1.405 P = 0.062 5.770 Población F9,906 = 0.407 P = 0.903 0 Ambiente F1,906 = 0.407 P = 0.539 0
Ambiente x Individo F83,906 = 3.381 P < 104 21.930 Ambiente x Población F9,906 = 11.851 P < 104 23.258
Tabla A2.19. Análisis GLM del efecto del ambiente en la acumulación de Fny-CMV
considerando fijos los factores aislado y ambiente y aleatorios los factores población e
individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F83,978 = 0.945 P = 0.602 0 Población F9,978 = 0.375 P = 0.920 0 Ambiente F1,978 = 0.1-66 P = 0.920 0
Ambiente x Individo F83,978 = 5.1-69 P < 104 33.460 Ambiente x Población F9,978 = 7.381 P < 104 18.985
Tabla A2.20. Análisis GLM del efecto del ambiente en la acumulación de Cdc1-CMV
considerando fijos los factores aislado y ambiente y aleatorios los factores población y
haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F44,906 = 1.325 P = 0.177 3.377 Población F9,906 = 0.345 P = 0.937 0 Ambiente F1,906 = 0.206 P = 0.661 0
Ambiente x Haplotipo F44,906 = 3.099 P < 104 16.584 Ambiente x Población F9,906 = 8.597 P < 104 24.541
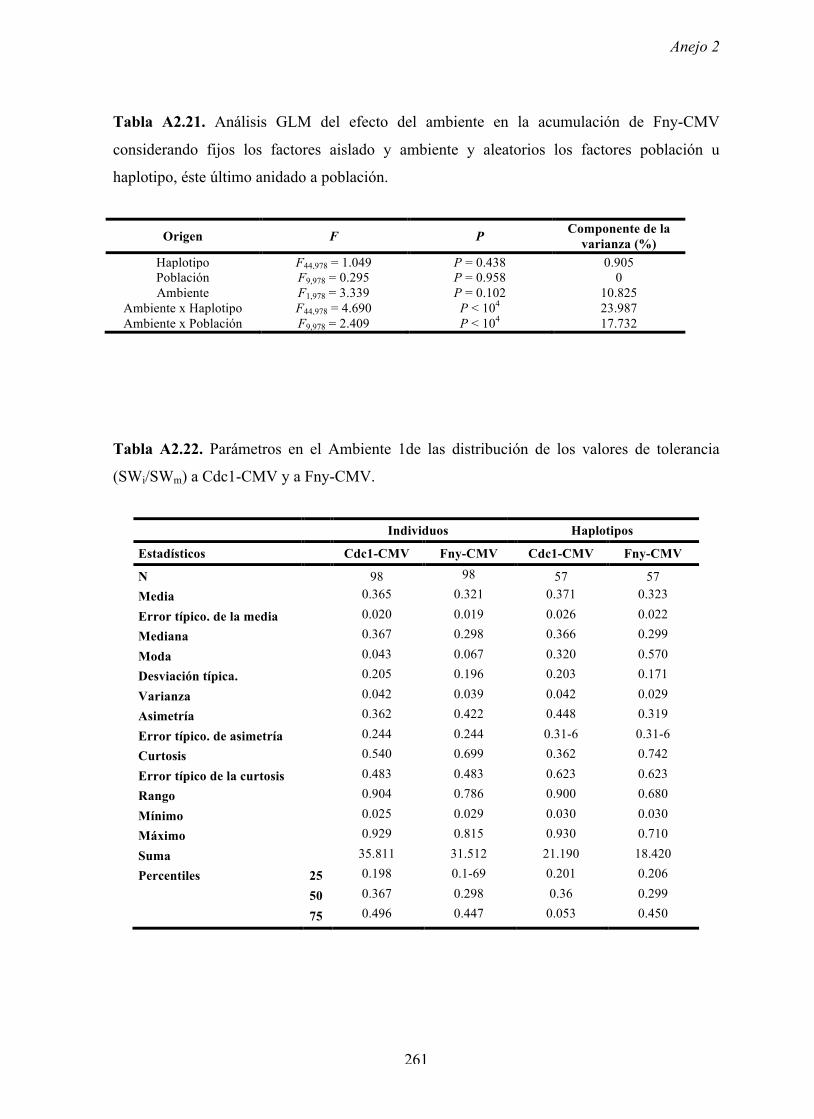
Anejo 2
261
Tabla A2.21. Análisis GLM del efecto del ambiente en la acumulación de Fny-CMV
considerando fijos los factores aislado y ambiente y aleatorios los factores población u
haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F44,978 = 1.049 P = 0.438 0.905 Población F9,978 = 0.295 P = 0.958 0 Ambiente F1,978 = 3.339 P = 0.102 10.825
Ambiente x Haplotipo F44,978 = 4.690 P < 104 23.987 Ambiente x Población F9,978 = 2.409 P < 104 17.732
Tabla A2.22. Parámetros en el Ambiente 1de las distribución de los valores de tolerancia
(SWi/SWm) a Cdc1-CMV y a Fny-CMV.
Individuos Haplotipos
Estadísticos Cdc1-CMV Fny-CMV Cdc1-CMV Fny-CMV
N 98 98 57 57 Media
0.365 0.321 0.371 0.323
Error típico. de la media 0.020 0.019 0.026 0.022
Mediana 0.367 0.298 0.366 0.299
Moda 0.043 0.067 0.320 0.570
Desviación típica. 0.205 0.196 0.203 0.171
Varianza 0.042 0.039 0.042 0.029
Asimetría 0.362 0.422 0.448 0.319
Error típico. de asimetría 0.244 0.244 0.31-6 0.31-6
Curtosis 0.540 0.699 0.362 0.742
Error típico de la curtosis 0.483 0.483 0.623 0.623
Rango 0.904 0.786 0.900 0.680
Mínimo 0.025 0.029 0.030 0.030
Máximo 0.929 0.815 0.930 0.710
Suma 35.811 31.512 21.190 18.420
Percentiles 25 0.198 0.1-69 0.201 0.206
50 0.367 0.298 0.36 0.299
75 0.496 0.447 0.053 0.450

Anejo 2
262
Tabla A2.23. Análisis GLM de la tolerancia a CMV en el Ambiente 1 considerando fijo el
factor aislado y aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,884 = 5.305 P = 0.046 1.611 Individuo F88,884 = 2.445 P = 0.023 32.237 Población F9,884 = 2.404 P = 0.059 6.447
Aislado x Individuo F88,884 = 2.649 P < 104 16.118 Aislado x Población F9,884 = 1.010 P = 0.439 0.641
Tabla A2.24. Análisis GLM en el Ambiente 1 de la tolerancia a CMV considerando fijo el
factor aislado y aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,884 = 7.358 P = 0.023 1.428 Haplotipo F47,884 = 3.205 P < 104 24.285 Población F9,884 = 2.808 P = 0.017 5.714
Aislado x Haplotipo F47,884 2.086 P < 104 8.571 Aislado x Población F9,884 = 0.596 P = 0.794 0
Tabla A2.25. Análisis GLM en el Ambiente 1 de la tolerancia a Cdc1-CMV considerando
aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F88,453 = 6.340 P < 104 49.230 Población F9,453 = 2.288 P = 0.023 7.692
Tabla A2.26. Análisis GLM en el Ambiente 1 de la tolerancia a Fny-CMV considerando
aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F47,431 = 5.702 P < 104 50.000 Población F9,431 = 1.979 P = 0.051 5.172
Anejo 2
263
Tabla A2.27. Análisis GLM en el Ambiente 1 de la tolerancia a Cdc1-CMV considerando
aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F47,431 = 5.497 P < 104 40.540 Población F9,431 = 1.895 P = 0.076 4.540
Tabla A2.28. Análisis GLM de la tolerancia a Fny-CMV en el Ambiente 1 considerando
aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F47,417 = 3.590 P < 104 27.219 Población F9,417 = 2.115 P = 0.047 4.838
Tabla A2.29. Parámetros de las distribución de los valores de tolerancia (SWi/SWm) a Cdc1-
CMV y a Fny-CMV en el Ambiente 2.
Individuos Haplotipos
Estadísticos Cdc1-CMV Fny-CMV Cdc1-CMV Fny-CMV
N 71 71 42 42 Media
0.324 0 .303 0.342 0.318
Error típico. de la media 0.027 0.025 0.028 0.028
Mediana 0.305 0.259 0.332 0.286
Moda 0.000 0.1-45 0.000 0.173
Desviación típica. 0.232 0.21-4 0.186 0.186
Varianza 0.054 0.046 0.035 0.035
Asimetría 1.583 1.1-60 0.113 0.520
Error típico. de asimetría 0.285 0.285 0.365 0.365
Curtosis 6.399 1.793 0.574 0.206
Error típico de la curtosis 0.563 0.563 0.717 0.717
Rango 1.447 1.092 0.690 0.790
Mínimo 0.000 0.001 0.000 0.011
Máximo 1.447 1.093 0.690 0.801
Suma 23.012 21.535 1-4.380 13.383
Percentiles 25 0.180 0.1-49 0.228 0.180
50 0.305 0.259 0.332 0.286
75 0.430 0.430 0.456 0.478

Anejo 2
264
Tabla A2.30. Análisis GLM de la tolerancia a CMV en el Ambiente 2 considerando fijo el
factor aislado y aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,838= 1.1-46 P =0.308 1.612 Individuo F61,838 = 2.409 P < 104 32.258 Población F9,838= 4.731 P = 0.001 6.451
Aislado x Individuo F61,838= 2.445 P < 104 16.129 Aislado x Población F9,838= 0.490 P = 0.875 0
Tabla A2.31. Análisis GLM en el Ambiente 2 de la tolerancia a CMV considerando fijo el
factor aislado y aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,838= 3.112 P = 0.1-47 0 Haplotipo F32,838= 4.353 P < 104 26.744 Población F9,838= 1.305 P = 0.291 0
Aislado x Haplotipo F32,838= 1.782 P = 0.058 3.488 Aislado x Población F9,838= 0.388 P = 0.927 0
Tabla A2.32. Análisis GLM en el Ambiente 2 de la tolerancia a Cdc1-CMV considerando
aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F61,421= 5.543 P < 104 35.560 Población F9,421= 2.297 P = 0.027 9.890
Tabla A2.33. Análisis GLM en el Ambiente 2 de la tolerancia a Fny-CMV considerando
aleatorios los factores población e individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F61,417= 3.442 P < 104 23.469 Población F9,417 = P =0.002 13.265

Anejo 2
265
Tabla A2.34. Análisis GLM en el Ambiente 2 de la tolerancia a Cdc1-CMV considerando
aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F32,421 = 5.017 P < 104 32.530 Población F9,421 = 0.938 P = 0.509 0
Tabla A2.35. Análisis GLM de la tolerancia a Fny-CMV en el Ambiente 2 considerando
aleatorios los factores población y haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F32,417= 4.535 P < 104 30.000 Población F9,417 = 1.120 P = 0.382 0
Tabla A2.36. Análisis GLM del efecto del ambiente en la tolerancia a CMV considerando
fijos los factores aislado y ambiente y aleatorios los factores población y individuo, éste
último anidado a población.
Origen F P Componente de la varianza (%)
Aislado F1,1111 = 0.998 P = 0.348 0 Individuo F48,1111 = 1.301 P = 0.245 5.505 Población F7,1111 = 2.585 P = 0.077 1.683 Ambiente F1,1111 = 0.357 P = 0.567 0
Aislado x Individuo F48,1111 = 0.776 P = 0.808 1.010 Aislado x Población F9,1111 = 22.406 P = 0.862 0 Ambiente x Aislado F1,1111 = 0.030 P = 0.865 0 Ambiente x Individo F48,1111 = 1.301 P = 0.026 5.050
Ambiente x Población F9,1111 = 2.630 P = 0.073 2.356 Aislado x Ambiente Individuo F48,1111 = 3.047 P < 104 0 Aislado x Ambiente Población F7,1111 = 0.030 P = 0.865 0

Anejo 2
266
Tabla A2.37. Análisis GLM del efecto del ambiente en la tolerancia a Cdc1-CMV
considerando fijos los factores aislado y ambiente y aleatorios los factores población e
individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F48,572= 0.988 P = 0.51-6 0 Población F7,572 = 3.656 P = 0.082 10.256 Ambiente F1,572 = 0382 P = 0.554 0
Ambiente x Individo F48,572= 5.712 P < 104 43.589 Ambiente x Población F7,572 = 0.968 P = 0.465 0
Tabla A2.38. Análisis GLM del efecto del ambiente en la tolerancia a Fny-CMV
considerando fijos los factores aislado y ambiente y aleatorios los factores población e
individuo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Individuo F48,549 = 1.003 P = 0.496 2.380 Población F7,549 = 3.881 P = 0.062 10.71-4 Ambiente F1,549 = 0.260 P = 0.624 0
Ambiente x Individo F48,549= 3.150 P < 104 25.000 Ambiente x Población F7,549 = 1.207 P = 0.317 1.190
Tabla A2.39. Análisis GLM del efecto del ambiente en la tolerancia a Cdc1-CMV
considerando fijos los factores aislado y ambiente y aleatorios los factores población y
haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F24,572 = 0.850 P = 0.653 0 Población F7,572 = 0.567 P =0.201 2.597 Ambiente F1,572 = 4.713 P = 0.586 0
Ambiente x Haplotipo F24,572 = 5.434 P < 104 36.365 Ambiente x Población F7,572= 0.342 P = 0.925 0

Anejo 2
267
Tabla A2.40. Análisis GLM del efecto del ambiente en la tolerancia a Fny-CMV
considerando fijos los factores aislado y ambiente y aleatorios los factores población y
haplotipo, éste último anidado a población.
Origen F P Componente de la varianza (%)
Haplotipo F24,549 =1.027 P = 0.474 3.797 Población F7,549 = 3.883 P = 0.204 2.531 Ambiente F1,549 = 1.21-4 P = 0.360 0
Ambiente x Haplotipo F1,549 = 2.806 P < 104 18.987 Ambiente x Población F7,549 = 0.572 P = 0.769 0
Tabla A2.41. Medias±error estándar en el Ambiente 1 de la relación entre el peso de la parte
reproductiva y el de la parte vegetativa, y del peso (g) de las diferentes partes de las plantas sin
infectar según el grupo alométrico.
Grupo alométrico (SW+IW)/RW1 RW2 IW3 SW4 BM5
Grupo 1 7.158±0.229 0.648±0.037 3.070±0.120 0.923±0.053 4.642±0.153 Grupo 2 34.879±1.786 0.174±0.003 2.708±0.026 0.944±0.013 3.827±0.038
1(SW+IW)/RW: Relación entre el peso (g) de la parte reproductiva y la parte vegetativa. 2RW: Peso de la roseta (g). 3IW: Peso de la inflorescencia sin las semillas (g). 4SW: Peso de las semillas (g). 5BM: Peso de la biomasa (g).
Tabla A2.42. Composición en el Ambiente 1 de individuos de cada grupo alométrico.
Individuos del Grupo 1
Agu1, Agu11, Agu12, Agu4, Agu6, Agu8, Bis4, Bis9, Gra1, Gra2, Gra3, Gra4, Gra5, Gra6, Gra7, Gra9, Leo1, Leo10, Leo11, Leo2, Leo3, Leo4, Leo6, Leo7, Leo8, Leo9, Moc15, Moc19, Moc9, Pob10, Pob2, Pob3, Pob4, Pob8, Pob9, Pra0, Pra10, Pra2, Pra3, Pra4, Pra7, Pra8, Qui6, Qui8, Qui9, San2
Individuos del Grupo 2
Agu0, Agu18, Agu2, Agu7, Bis0, Bis1, Bis2, Bis3, Bis5, Bis6, Bis7, Bis8, Gra0, Gra8, Mer0, Mer1, Mer10, Mer12, Mer14, Mer3, Mer5, Mer6, Mer7, Mer9, Moc0, Moc1, Moc14, Moc2, Moc5, Moc7, Moc8, Pob5, Pob7, Pra1, Pra5, Pra6, Qui0, Qui1, Qui2, Qui3, Qui4, Qui5, Qui7, San0, San10, San12, San13, San4, San5, San6, San8, San9, San13, San4
Anejo 2
268
Tabla A2.43. Medias±error estándar en el Ambiente 2 de la relación entre el peso de la parte
reproductiva y el de la parte vegetativa, y del peso (g) de las diferentes partes de las plantas sin
infectar según el grupo alométrico.
Grupo
alométrico (SW+IW)/RW1 RW2 IW3 SW4 BM5
Grupo 1 1.402±0.035 1.491±0.031 1.611±0.032 0.303±0.012 3.406±0.052
Grupo 2 8.169±0.484 0.509±0.030 1.836±0.053 0.529±0.021 2.875±0.087 1(SW+IW)/RW: Relación entre el peso (g) de la parte reproductiva y la parte vegetativa. 2RW: Peso de la roseta (g). 3IW: Peso de la inflorescencia sin las semillas (g). 4SW: Peso de las semillas (g). 5BM: Peso de la biomasa (g).
Tabla A2.44. Composición en el Ambiente 2de individuos de cada grupo alométrico.
Individuos del Grupo 1
Agu11, Agu7, Bis0, Bis2, Bis3, Bis4, Bis5, Bis6, Bis7, Cdc12, Cdc22, Cdc3, Gra0, Gra1, Gra2, Gra3, Gra4, Gra5, Gra6, Gra7, Gra8, Gra9, Mar13, Mar6, Moc0, Moc1, Moc14, Moc7, Pob1, Pob3, Pob6, Pob7, Pob9, Qui5, San10, San12, San4, San9
Individuos del Grupo 2
Agu0, Agu2, Cdc27, Mar10, Mar12, Mar15, Mer0, Mer10, Mer12, Mer14, Mer3, Mer5, Mer9, Moc8, Qui0, Qui1, Qui2, Qui3, Qui4, Qui6, Qui7, Qui8, Qui9, San0, San13, San5, San6, San8
Tabla A2.45. Medias±error estándar en el Ambiente 1 del efecto de la infección por CMV
sobre el peso de la roseta, la inflorescencia sin semillas, las semillas y la biomasa total de la
planta según el grupo alométrico.
Grupo
alométrico Tratamiento RWm/RWi
1 IWi/IWm2 SWi/SWm
3 BMi/BMm4
Grupo 1 CMV 0.155±0.008 0.493±0.012 0.365±0.012 0.416±0.010
Grupo 2 CMV 0.331±0.015 0.503±0.012 0.320±0.019 0.448±0.011 1RWi/RWm: Efecto de la infección por CMV sobre el peso de la roseta. 2IWi/IWm: Efecto de la infección por CMV sobre el peso de la inflorescencia sin las semillas. 3SWi/SWm: Efecto de la infección por CMV sobre el peso de las semillas. 4BWi/BWm: Efecto de la infección por CMV sobre el peso de biomasa total.
Anejo 2
269
Tabla A2.46. Medias±error estándar en el Ambiente 2 del efecto de la infección por CMV
sobre el peso de la roseta, la inflorescencia sin semillas, las semillas y la biomasa total de la
planta según el grupo alométrico.
Grupo
alométrico Tratamiento RWm/RWi
1 IWi/IWm2 SWi/SWm
3 BMi/BMm4
Grupo 1 CMV 0.155±0.007 0.418±0.013 0.332±0.015 0.297±0.009
Grupo 2 CMV 0.202±0.008 0.352±0.0101 0.287±0.011 0.306±0.011 1RWi/RWm: Efecto de la infección por CMV sobre el peso de la roseta. 2IWi/IWm: Efecto de la infección por CMV sobre el peso de la inflorescencia sin las semillas. 3SWi/SWm: Efecto de la infección por CMV sobre el peso de las semillas. 4BWi/BWm: Efecto de la infección por CMV sobre el peso de biomasa total.
Tabla A2.47 Medias±error estándar en el Ambiente 1 de las variables que describen el ciclo
vital (días) de las plantas según grupo alométrico.
1GP: Duración (días) del periodo pre-reproductor. 2FT: Duración (días) del periodo reproductor.
Tabla A2.48. Medias±error estándar en el Ambiente 2 de las variables que describen el ciclo
vital (días) de las plantas según grupo alométrico.
1GP: Duración (días) del periodo pre-reproductor. 2FT: Duración (días) del periodo reproductor. 3TP: Duración (días) del ciclo vital.
Grupo alométrico Tratamiento GP1 FT2
Grupo 1 Control 108.066±0.402 20.08±0.350 Grupo 1 CMV 110.175±0.322 23.024±0.243 Grupo 2 Control 101.037±0.353 20.154±0.353 Grupo 2 CMV 101.508±0.254 21.817±0.174
Grupo alométrico Tratamiento GP1 FT2 TP3
Grupo 1 Control 95.04±0.661 23.84±0.253 166.26±0.752 Grupo 1 CMV 100.169±0.387 22.767±0.309 175.45±0.500 Grupo 2 Control 72.86±0.489 22.46±0.393 158.78±0.682 Grupo 2 CMV 73.415±0.429 24.286±0.471 168.57±0.595

Anejo 2
270
Tabla A2.49. Análisis de correlación en el Ambiente 2 entre los valores medios de cada
individuo de acumulación de CMV y/o tolerancia a CMV y las variables que describen el
ciclo vital de las plantas infectadas.
GP1 RP2 TP3
Acumulación de CMV
rS=0.247, P=0.040
rP=0.237, P=0.055
rS=0.227, P=0.067
rP=0.260, P=0.035
rS=0.191, P=0.124
rP=0.223, P=0.072
Tolerancia a CMV r
S=0.119 P=0.342
rP=0.192 P=0.122
rS=0.205 P=0.099
rP=0.113 P=0.368
rS=0.024 P=0.847 r
P=0.000 P=1
1GP: Duración (días) del periodo pre-reproductor. 2FT: Duración (días) del periodo reproductor. 3TP: Duración (días) del ciclo vital.
Tabla A2.50. Valores en el Ambiente 2 de QST para la resistencia a Cdc1-CMV y para la
resistencia a Fny-CMV.
Cdc1-CMV Fny-CMV
QST 0.395
(IC 95%: 0.584 0.296)
0.178 (IC 95%: 0.3190.123)
FST 0.568
(IC 95%: 0.601 0.532)
Tabla A2.51. Valores en el Ambiente 2 de QST para la tolerancia a Cdc1-CMV y para la
tolerancia a Fny-CMV.
Cdc1-CMV Fny-CMV
QST 0.200
(IC 95%: 0.360 0.1-40)
0.361 (IC 95%: 0.55600.270)
FST 0.651
(IC 95%: 0.690 0.610)

ANEJO 3

Anejo 3
272

Anejo 3
273
Tabla A3.1. Listado de individuos de arabidopsis analizados en el Apartado 4.3, indicando su procedencia y hábitat.
Individuo Localidad Longitud (Oº)
Latitud (Nº)
Altitud (m) Hábitat
Adc-5 Argamasilla de Calatrava -4.072 38.766 701 Olivar Adm-0 Arroba de los Montes -4.543 39.146 763 Olivar Agu-1 Aguarón -1.340 41.321 1045 Encinar Ala-0 Alcántara -6.891 39.722 158 Pared de piedra al lado de un río en el pueblo Ali-1 Alcañizo -5.089 39.895 392 Camino agrícola y pradera xérico All-0 Allariz -7.803 42.189 450 Pared de piedra al lado de un río en el pueblo
Alm-0 Almedíjar -0.364 39.885 678 Camino rocoso en bosque de pinos y encinas Alo-0 Alcongosta -7.475 40.111 763 Paredes rocosas en un arroyo Alr-0
-0.346 39.095 123
ArA3 Aranda de Duero -3.682 41.702 859 Claro en un bosque de Quercus caducifolios y perennes Bea-0 Benarrabá -5.268 36.521 140 Plano aluvial y pradera Ber-0 Bernués -0.561 42.517 1080 Camino flanqueante a un encinar Bis-0 Bisaurri 0.537 42.490 1450 Pradera subalpina Cad-0 Candelario -5.741 40.366 1170 Muro de piedra dentro del pueblo Cal-0 Calamocha -1.369 40.942 1120 Encinar Cap-1 Capileira -3.359 36.969 1460 Muro de piedra cercano a terrenos agrícolas abandonados Car-1 Cardeña -4.324 38.253 756 Muro de piedra junto a un encinar Cdc3 Ciruelos de Coca -4.544 41.215 800 Camino arenoso junto a pinar Cdo0 Castrillejos de la Olma -4.637 42.232 820 Lindes de terrenos agrícolas Cho-0 La Chopera de las Rozas -3.897 40.515 725 Pradera xérica y matorral mediterráneo cerca de la cartera Coa-0 Corval -7.498 38.446 208 Linde del camino Coc0 Cabo de Creus 3.195 42.312 245 Matorral mediterráneo costero Cor0 Corduente -2.000 40.831 1080 Ladera rocosa y borde de la carretera en un pinar
Cum-1 Cumbres Mayores -6.656 38.068 640 Muro de piedra junto a pradera
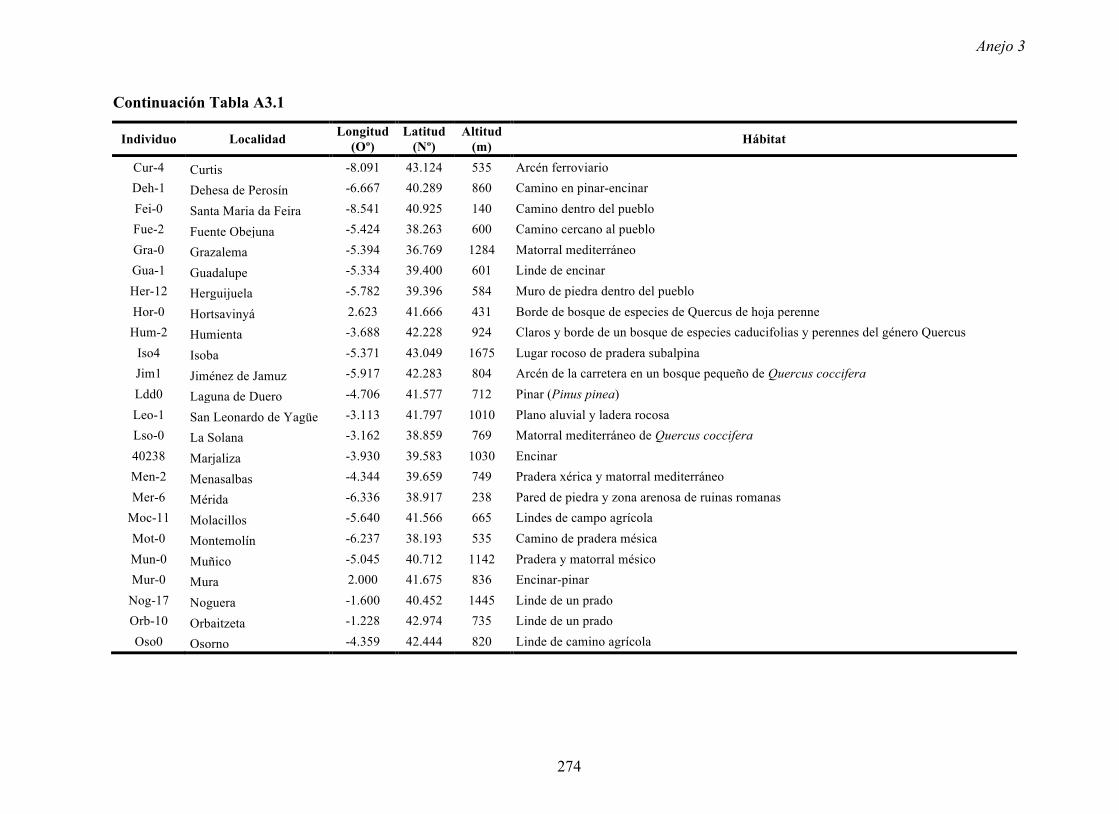
Anejo 3
274
Continuación Tabla A3.1
Individuo Localidad Longitud (Oº)
Latitud (Nº)
Altitud (m) Hábitat
Cur-4 Curtis -8.091 43.124 535 Arcén ferroviario Deh-1 Dehesa de Perosín -6.667 40.289 860 Camino en pinar-encinar Fei-0 Santa Maria da Feira -8.541 40.925 140 Camino dentro del pueblo Fue-2 Fuente Obejuna -5.424 38.263 600 Camino cercano al pueblo Gra-0 Grazalema -5.394 36.769 1284 Matorral mediterráneo Gua-1 Guadalupe -5.334 39.400 601 Linde de encinar Her-12 Herguijuela -5.782 39.396 584 Muro de piedra dentro del pueblo Hor-0 Hortsavinyá 2.623 41.666 431 Borde de bosque de especies de Quercus de hoja perenne Hum-2 Humienta -3.688 42.228 924 Claros y borde de un bosque de especies caducifolias y perennes del género Quercus
Iso4 Isoba -5.371 43.049 1675 Lugar rocoso de pradera subalpina Jim1 Jiménez de Jamuz -5.917 42.283 804 Arcén de la carretera en un bosque pequeño de Quercus coccifera Ldd0 Laguna de Duero -4.706 41.577 712 Pinar (Pinus pinea) Leo-1 San Leonardo de Yagüe -3.113 41.797 1010 Plano aluvial y ladera rocosa Lso-0 La Solana -3.162 38.859 769 Matorral mediterráneo de Quercus coccifera 40238 Marjaliza -3.930 39.583 1030 Encinar Men-2 Menasalbas -4.344 39.659 749 Pradera xérica y matorral mediterráneo Mer-6 Mérida -6.336 38.917 238 Pared de piedra y zona arenosa de ruinas romanas
Moc-11 Molacillos -5.640 41.566 665 Lindes de campo agrícola Mot-0 Montemolín -6.237 38.193 535 Camino de pradera mésica Mun-0 Muñico -5.045 40.712 1142 Pradera y matorral mésico Mur-0 Mura 2.000 41.675 836 Encinar-pinar
Nog-17 Noguera -1.600 40.452 1445 Linde de un prado Orb-10 Orbaitzeta -1.228 42.974 735 Linde de un prado Oso0 Osorno -4.359 42.444 820 Linde de camino agrícola
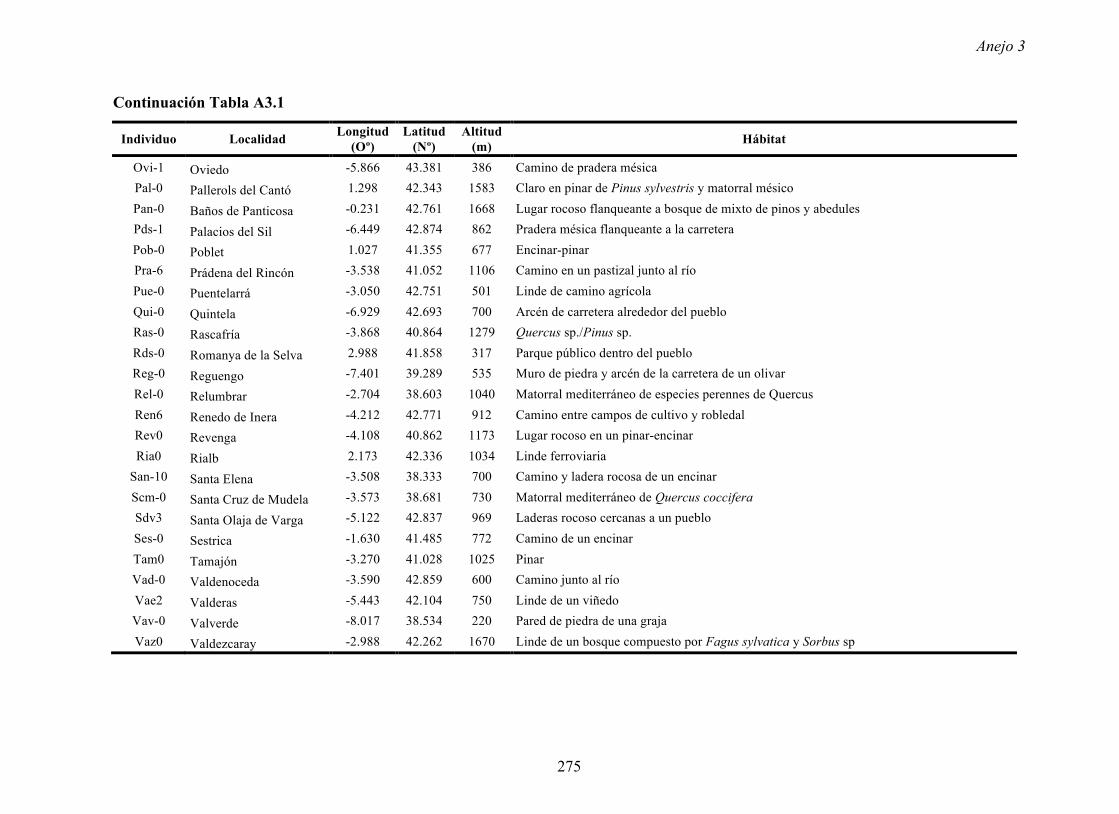
Anejo 3
275
Continuación Tabla A3.1
Individuo Localidad Longitud (Oº)
Latitud (Nº)
Altitud (m) Hábitat
Ovi-1 Oviedo -5.866 43.381 386 Camino de pradera mésica Pal-0 Pallerols del Cantó 1.298 42.343 1583 Claro en pinar de Pinus sylvestris y matorral mésico Pan-0 Baños de Panticosa -0.231 42.761 1668 Lugar rocoso flanqueante a bosque de mixto de pinos y abedules Pds-1 Palacios del Sil -6.449 42.874 862 Pradera mésica flanqueante a la carretera Pob-0 Poblet 1.027 41.355 677 Encinar-pinar Pra-6 Prádena del Rincón -3.538 41.052 1106 Camino en un pastizal junto al río Pue-0 Puentelarrá -3.050 42.751 501 Linde de camino agrícola Qui-0 Quintela -6.929 42.693 700 Arcén de carretera alrededor del pueblo Ras-0 Rascafría -3.868 40.864 1279 Quercus sp./Pinus sp. Rds-0 Romanya de la Selva 2.988 41.858 317 Parque público dentro del pueblo Reg-0 Reguengo -7.401 39.289 535 Muro de piedra y arcén de la carretera de un olivar Rel-0 Relumbrar -2.704 38.603 1040 Matorral mediterráneo de especies perennes de Quercus Ren6 Renedo de Inera -4.212 42.771 912 Camino entre campos de cultivo y robledal Rev0 Revenga -4.108 40.862 1173 Lugar rocoso en un pinar-encinar Ria0 Rialb 2.173 42.336 1034 Linde ferroviaria
San-10 Santa Elena -3.508 38.333 700 Camino y ladera rocosa de un encinar Scm-0 Santa Cruz de Mudela -3.573 38.681 730 Matorral mediterráneo de Quercus coccifera Sdv3 Santa Olaja de Varga -5.122 42.837 969 Laderas rocoso cercanas a un pueblo Ses-0 Sestrica -1.630 41.485 772 Camino de un encinar Tam0 Tamajón -3.270 41.028 1025 Pinar Vad-0 Valdenoceda -3.590 42.859 600 Camino junto al río Vae2 Valderas -5.443 42.104 750 Linde de un viñedo Vav-0 Valverde -8.017 38.534 220 Pared de piedra de una graja Vaz0 Valdezcaray -2.988 42.262 1670 Linde de un bosque compuesto por Fagus sylvatica y Sorbus sp

Anejo 3
276
Continuación Tabla A3.1
Individuo Localidad Longitud (Oº)
Latitud (Nº)
Altitud (m) Hábitat
Vdm0 Vilanova de Meiá 1.007 42.037 1077 Matorral mediterráneo Ver-5 Verín -7.448 41.949 510 Pared de piedra de una granja abandonada Vim-0 Villarino de Manzanas -6.509 41.878 652 Lugar rocoso en matorral mediterráneo Vis-0 Villareal de San Carlos -6.044 39.847 244 Camino junto a un arroyo Voz0 Vozmediano -1.878 41.846 940 Pradera cercana a un arroyo

Anejo 3
277
Tabla A3.2. Coeficientes de correlación de Pearson de las correlaciones con la corrección de
Dutilleul entre los valores de las distintas variables ambientales (Apartado 4.3 Tabla 4.3.1).
La significancia está indicada como: ns (no significativa) P>0.1, * P<0.1, ** P<0.05 y ***
P>0.001.
BIO1 BIO2 BIO3 BIO4 BIO5 BIO6
BIO1 BIO2 0.107ns BIO3 -0.077ns 0.131ns BIO4 0.175ns 0.642** -0.662** BIO5 0.822** 0.557** -0.294ns 0.665** BIO6 0.927*** -0.184ns 0.096ns -0.178ns 0.576** BIO7 0.143ns 0.831*** -0.438** 0.955*** 0.673** -0.217ns BIO8 0.393ns -0.055ns -0.081ns 0.069ns 0.247ns 0.336ns BIO9 0.735** 0.079ns -0.122ns 0.166ns 0.655** 0.707** BIO10 0.955*** 0.285ns -0.273** 0.459ns 0.942*** 0.785** BIO11 0.956*** -0.087ns 0.110ns -0.116ns 0.634** 0.989*** BIO12 -0.349* -0.599** 0.071ns -0.536** -0.581** -0.128ns BIO13 -0.093ns -0.572** 0.019ns -0.466** -0.349ns 0.118ns BIO14 -0.769** -0.468** 0.043ns -0.404* -0.860** -0.636** BIO15 0.686** 0.096ns -0.011ns 0.078ns 0.597* 0.679** BIO16 -0.109ns -0.552** 0.046ns -0.477** -0.358ns 0.103ns BIO17 -0.734** -0.519** 0.038ns -0.437** -0.860** -0.589** BIO18 -0.682** -0.474** 0.067ns -0.416* -0.818** -0.561** BIO19 -0.035ns -0.467** 0.070ns -0.435** -0.251ns 0.171ns CLI1 -0.008ns -0.480** 0.096ns -0.469** -0.246ns 0.208ns CLI2 -0.648*** -0.527** -0.003ns -0.430** -0.750** -0.472** CLI3 0.009ns -0.520** 0.130ns -0.525** -0.272ns 0.238ns CLI4 0.866*** -0.072ns -0.118ns 0.080ns 0.682** 0.892*** CLI5 0.866** -0.073ns 0.034ns -0.07ns 0.509* 0.765*** CLI6 0.908*** -0.059ns -0.165ns 0.130ns 0.698** 0.871*** CLI7 0.889*** -0.153ns -0.009ns -0.069ns 0.594** 0.926*** CLI8 0.734*** -0.073ns 0.034ns -0.070ns 0.509* 0.765*** CLI9 0.889*** 0.343ns -0.072ns 0.360ns 0.861** 0.740** CLI10 0.918*** 0.067ns 0.071ns 0.039ns 0.697** 0.889*** CLI11 0.911*** 0.292ns -0.036ns 0.286ns 0.839** 0.785*** CLI12 0.902*** -0.035ns 0.071ns -0.037ns 0.638** 0.914** CLI13 0.927*** 0.171ns -0.126ns 0.273ns 0.817** 0.822*** CLI14 0.914*** -0.045ns 0.026ns -0.013ns 0.653** 0.917*** HUH 0.396** 0.265ns 0.303* -0.030ns 0.350* 0.385** PHS -0.174ns 0.088ns -0.234ns 0.282ns -0.058ns -0.295ns

Anejo 3
278
Continuación Tabla A3.2.
BIO7 BIO8 BIO9 BIO10 BIO11 BIO12
BIO1 BIO2 BIO3 BIO4 BIO5 BIO6 BIO7 BIO8 -0.009ns BIO9 0.143ns -0.010ns
BIO10 0.415ns 0.368ns 0.718** BIO11 -0.137ns 0.364ns 0.703** 0.829** BIO12 -0.578** -0.392** -0.276ns -0.466** -0.182ns BIO13 -0.523** -0.273* -0.077ns -0.209ns 0.059ns 0.921*** BIO14 -0.452** -0.178ns -0.696** -0.819** -0.661** 0.593*** BIO15 0.098ns -0.040ns 0.568* 0.657* 0.684** 0.097ns BIO16 -0.521** -0.331** -0.080ns -0.228ns 0.045ns 0.938*** BIO17 -0.494** -0.183ns -0.650** -0.798** -0.615** 0.647*** BIO18 -0.470** 0.005ns -0.763** -0.745** -0.572** 0.584** BIO19 -0.454** -0.463** 0.074ns -0.148ns 0.109ns 0.885*** CLI1 -0.482** -0.398** 0.059ns -0.136ns 0.146ns 0.867*** CLI2 -0.469** -0.367* -0.525** -0.709** -0.518** 0.842*** CLI3 -0.540** -0.360** 0.058ns -0.137ns 0.180ns 0.878*** CLI4 0.008ns 0.429* 0.641** 0.850** 0.899*** -0.198ns CLI5 -0.084ns 0.295ns 0.547** 0.646** 0.766*** -0.085ns CLI6 0.045ns 0.480** 0.637** 0.863** 0.883*** -0.245** CLI7 -0.129ns 0.403* 0.653** 0.784** 0.921*** -0.142ns CLI8 -0.084ns 0.295ns 0.547** 0.646** 0.766*** -0.085ns CLI9 0.358ns 0.428* 0.615** 0.908*** 0.788** -0.457**
CLI10 0.028ns 0.434* 0.642** 0.841** 0.915*** -0.257ns CLI11 0.292ns 0.405ns 0.621** 0.907*** 0.833** -0.391* CLI12 -0.065ns 0.406* 0.652** 0.805** 0.922*** -0.197ns CLI13 0.232ns 0.457* 0.650** 0.918*** 0.854*** -0.383* CLI14 -0.050ns 0.424* 0.655** 0.823** 0.928*** -0.205ns HUH 0.070ns 0.024ns 0.339* 0.346* 0.406** -0.278** PHS 0.197ns 0.344ns -0.251ns -0.075ns -0.266ns -0.401**

Anejo 3
279
Continuación Tabla A3.2.
BIO13 BIO14 BIO15 BIO16 BIO17 BIO18
BIO1 BIO2 BIO3 BIO4 BIO5 BIO6 BIO7 BIO8 BIO9
BIO10 BIO11 BIO12 BIO13 BIO14 0.295* BIO15 0.403** -0.692** BIO16 0.990*** 0.305* 0.405** BIO17 0.355** 0.981*** -0.651* 0.365** BIO18 0.320* 0.938*** -0.623* 0.325* 0.945*** BIO19 0.951*** 0.188ns 0.477** 0.964*** 0.246ns 0.151ns CLI1 0.916*** 0.186ns 0.447** 0.935*** 0.245ns 0.166ns CLI2 0.636*** 0.809*** -0.299ns 0.664*** 0.828*** 0.767*** CLI3 0.929*** 0.195ns 0.447** 0.944*** 0.269ns 0.200ns CLI4 0.083ns -0.659** 0.674** 0.042ns -0.618** -0.573** CLI5 0.149ns -0.544** 0.619** 0.125ns -0.503** -0.485* CLI6 0.023ns -0.650** 0.628** -0.019ns -0.607** -0.559** CLI7 0.123ns -0.624** 0.665** 0.087ns -0.573** -0.546** CLI8 0.149ns -0.544** 0.619** 0.125ns -0.503** -0.485* CLI9 -0.19ns -0.793** 0.630** -0.226ns -0.771** -0.708**
CLI10 0.003ns -0.685** 0.666** -0.030ns -0.650** -0.600** CLI11 -0.136ns -0.758** 0.646** -0.166ns -0.741** -0.672** CLI12 0.075ns -0.669** 0.686** 0.041ns -0.627** -0.591** CLI13 -0.109ns -0.756** 0.645** -0.146ns -0.726** -0.667** CLI14 0.062ns -0.662** 0.672** 0.026ns -0.619** -0.580** HUH -0.203ns -0.416** 0.304ns -0.191ns -0.430** -0.441** PHS -0.422** 0.095ns -0.399ns -0.461** 0.073ns 0.144ns
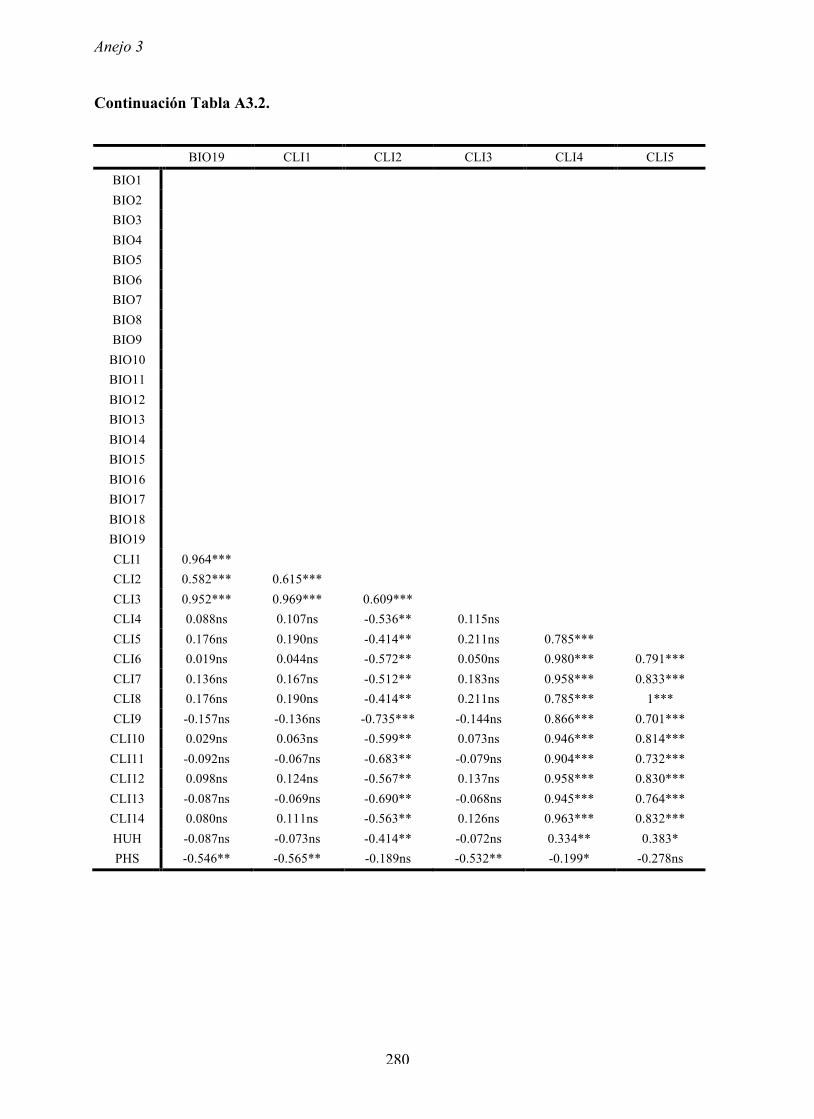
Anejo 3
280
Continuación Tabla A3.2.
BIO19 CLI1 CLI2 CLI3 CLI4 CLI5
BIO1 BIO2 BIO3 BIO4 BIO5 BIO6 BIO7 BIO8 BIO9
BIO10 BIO11 BIO12 BIO13 BIO14 BIO15 BIO16 BIO17 BIO18 BIO19 CLI1 0.964*** CLI2 0.582*** 0.615*** CLI3 0.952*** 0.969*** 0.609*** CLI4 0.088ns 0.107ns -0.536** 0.115ns CLI5 0.176ns 0.190ns -0.414** 0.211ns 0.785*** CLI6 0.019ns 0.044ns -0.572** 0.050ns 0.980*** 0.791*** CLI7 0.136ns 0.167ns -0.512** 0.183ns 0.958*** 0.833*** CLI8 0.176ns 0.190ns -0.414** 0.211ns 0.785*** 1*** CLI9 -0.157ns -0.136ns -0.735*** -0.144ns 0.866*** 0.701***
CLI10 0.029ns 0.063ns -0.599** 0.073ns 0.946*** 0.814*** CLI11 -0.092ns -0.067ns -0.683** -0.079ns 0.904*** 0.732*** CLI12 0.098ns 0.124ns -0.567** 0.137ns 0.958*** 0.830*** CLI13 -0.087ns -0.069ns -0.690** -0.068ns 0.945*** 0.764*** CLI14 0.080ns 0.111ns -0.563** 0.126ns 0.963*** 0.832*** HUH -0.087ns -0.073ns -0.414** -0.072ns 0.334** 0.383* PHS -0.546** -0.565** -0.189ns -0.532** -0.199* -0.278ns

Anejo 3
281
Continuación Tabla A3.2.
CLI6 CLI7 CLI8 CLI9 CLI10 CLI11
BIO1 BIO2 BIO3 BIO4 BIO5 BIO6 BIO7 BIO8 BIO9
BIO10 BIO11 BIO12 BIO13 BIO14 BIO15 BIO16 BIO17 BIO18 BIO19 CLI1 CLI2 CLI3 CLI4 CLI5 CLI6 CLI7 0.965*** CLI8 0.791*** 0.833*** CLI9 0.896*** 0.835*** 0.701**
CLI10 0.954** 0.833*** 0.814*** 0.920*** CLI11 0.909** 0.860*** 0.732** 0.965*** 0.947*** CLI12 0.962*** 0.985*** 0.830*** 0.886*** 0.987*** 0.908*** CLI13 0.970** 0.918*** 0.764** 0.969*** 0.961*** 0.966*** CLI14 0.971*** 0.990*** 0.832*** 0.887*** 0.989*** 0.913*** HUH 0.331** 0.376** 0.383** 0.440** 0.430** 0.415** PHS -0.145ns -0.244ns -0.278ns -0.109ns -0.213ns -0.161ns

Anejo 3
282
Continuación Tabla A3.2.
CLI12 CLI13 CLI14 HUH
BIO1 BIO2 BIO3 BIO4 BIO5 BIO6 BIO7 BIO8 BIO9
BIO10 BIO11 BIO12 BIO13 BIO14 BIO15 BIO16 BIO17 BIO18 BIO19 CLI1 CLI2 CLI3 CLI4 CLI5 CLI6 CLI7 CLI8 CLI9
CLI10 CLI11 CLI12 CLI13 0.945*** CLI14 0.997*** 0.951*** HUH 0.418** 0.390** 0.407** PHS -0.238ns -0.113ns -0.224ns -0.191ns

Anejo 3
283
Tabla A3.3. Análisis GLM de las variables RWm, IWm y SWm de las plantas sin infectar
considerando aleatorio el factor individuo de arabidopsis.
Variable F P RWm
1 F77, 410=57.558 P<10-4
IWm2 F77, 410=12.179 P<10-4
SWm3 F77, 410=9.158 P<10-4
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar.
Tabla A3.4. Valores (media ± error estándar) de RWm, IWm y SWm (g) de los individuos de
arabidopsis sin infectar.
Individuos RWm1 IWm
2 SWm3
Adc-5 0.131 ± 0.008 1.489 ± 0.141 0.846 ± 0.074 Adm-0 0.148 ± 0.014 1.138 ± 0.163 0.739 ± 0.083 Agu-1 0.488 ± 0.063 1.537 ± 0.121 0.866 ± 0.052 Ala-0 0.094 ± 0.016 0.839 ± 0.092 0.539 ± 0.063 Ali-1 0.174 ± 0.027 1.681 ± 0.231 0.735 ± 0.120 All-0 0.041 ± 0.010 0.709 ± 0.084 0.439 ± 0.037
Alm-0 0.700 ± 0.055 2.220 ± 0.341 0.915 ± 0.059 Alo-0 0.432 ± 0.045 1.671 ± 0.184 0.941 ± 0.081 Alr-0 0.063 ± 0.009 0.868 ± 0.060 0.569 ± 0.045 ArA3 0.778 ± 0.063 2.030 ± 0.074 0.915 ± 0.073 Bea-0 0.097 ± 0.013 1.099 ± 0.055 0.341 ± 0.014 Ber-0 0.543 ± 0.047 2.311 ± 0.061 1.089 ± 0.098 Bis-0 0.851 ± 0.080 2.507 ± 0.207 0.957 ± 0.057 Cad-0 0.607 ± 0.063 2.178 ± 0.081 0.963 ± 0.045 Cal-0 0.471 ± 0.061 1.826 ± 0.138 0.862 ± 0.034 Cap-1 0.453 ± 0.028 2.226 ± 0.059 1.160 ± 0.039 Car-1 0.300 ± 0.019 1.680 ± 0.084 1.027 ± 0.069 Cdc3 0.413 ± 0.021 2.134 ± 0.083 0.929 ± 0.041 Cdo0 0.591 ± 0.059 1.810 ± 0.093 0.774 ± 0.033 Cho-0 0.301 ± 0.024 1.682 ± 0.105 0.975 ± 0.073 Coa-0 0.122 ± 0.014 1.146 ± 0.069 0.644 ± 0.019 Coc0 0.503 ± 0.059 1.913 ± 0.156 0.845 ± 0.054 Cor0 0.333 ± 0.035 1.932 ± 0.242 0.894 ± 0.029
Cum-1 0.066 ± 0.008 0.854 ± 0.030 0.492 ± 0.047 Cur-4 0.290 ± 0.015 2.049 ± 0.284 0.731 ± 0.120 Deh-1 0.262 ± 0.016 1.801 ± 0.092 0.812 ± 0.062 Fei-0 0.081 ± 0.014 1.288 ± 0.050 0.658 ± 0.048
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar.
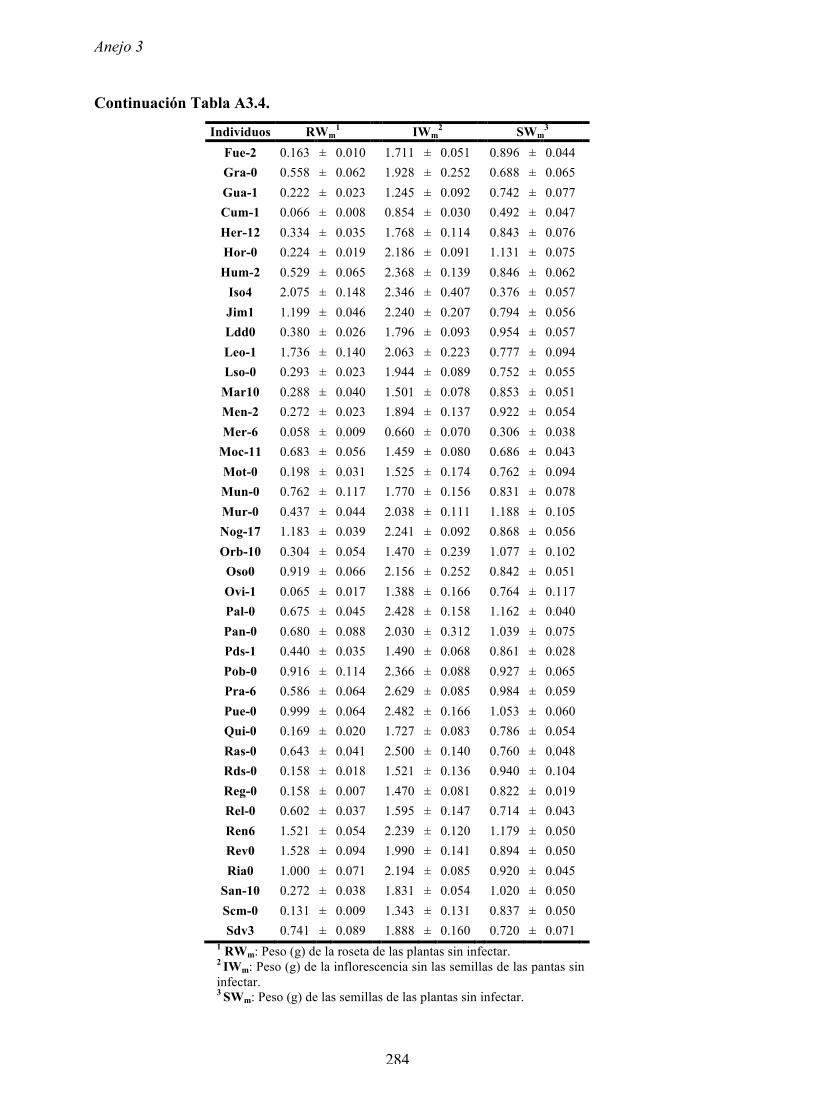
Anejo 3
284
Continuación Tabla A3.4.
Individuos RWm1 IWm
2 SWm3
Fue-2 0.163 ± 0.010 1.711 ± 0.051 0.896 ± 0.044
Gra-0 0.558 ± 0.062 1.928 ± 0.252 0.688 ± 0.065
Gua-1 0.222 ± 0.023 1.245 ± 0.092 0.742 ± 0.077
Cum-1 0.066 ± 0.008 0.854 ± 0.030 0.492 ± 0.047
Her-12 0.334 ± 0.035 1.768 ± 0.114 0.843 ± 0.076
Hor-0 0.224 ± 0.019 2.186 ± 0.091 1.131 ± 0.075
Hum-2 0.529 ± 0.065 2.368 ± 0.139 0.846 ± 0.062
Iso4 2.075 ± 0.148 2.346 ± 0.407 0.376 ± 0.057
Jim1 1.199 ± 0.046 2.240 ± 0.207 0.794 ± 0.056
Ldd0 0.380 ± 0.026 1.796 ± 0.093 0.954 ± 0.057
Leo-1 1.736 ± 0.140 2.063 ± 0.223 0.777 ± 0.094
Lso-0 0.293 ± 0.023 1.944 ± 0.089 0.752 ± 0.055
Mar10 0.288 ± 0.040 1.501 ± 0.078 0.853 ± 0.051
Men-2 0.272 ± 0.023 1.894 ± 0.137 0.922 ± 0.054
Mer-6 0.058 ± 0.009 0.660 ± 0.070 0.306 ± 0.038
Moc-11 0.683 ± 0.056 1.459 ± 0.080 0.686 ± 0.043
Mot-0 0.198 ± 0.031 1.525 ± 0.174 0.762 ± 0.094
Mun-0 0.762 ± 0.117 1.770 ± 0.156 0.831 ± 0.078
Mur-0 0.437 ± 0.044 2.038 ± 0.111 1.188 ± 0.105
Nog-17 1.183 ± 0.039 2.241 ± 0.092 0.868 ± 0.056
Orb-10 0.304 ± 0.054 1.470 ± 0.239 1.077 ± 0.102
Oso0 0.919 ± 0.066 2.156 ± 0.252 0.842 ± 0.051
Ovi-1 0.065 ± 0.017 1.388 ± 0.166 0.764 ± 0.117
Pal-0 0.675 ± 0.045 2.428 ± 0.158 1.162 ± 0.040
Pan-0 0.680 ± 0.088 2.030 ± 0.312 1.039 ± 0.075
Pds-1 0.440 ± 0.035 1.490 ± 0.068 0.861 ± 0.028
Pob-0 0.916 ± 0.114 2.366 ± 0.088 0.927 ± 0.065
Pra-6 0.586 ± 0.064 2.629 ± 0.085 0.984 ± 0.059
Pue-0 0.999 ± 0.064 2.482 ± 0.166 1.053 ± 0.060
Qui-0 0.169 ± 0.020 1.727 ± 0.083 0.786 ± 0.054
Ras-0 0.643 ± 0.041 2.500 ± 0.140 0.760 ± 0.048
Rds-0 0.158 ± 0.018 1.521 ± 0.136 0.940 ± 0.104
Reg-0 0.158 ± 0.007 1.470 ± 0.081 0.822 ± 0.019
Rel-0 0.602 ± 0.037 1.595 ± 0.147 0.714 ± 0.043
Ren6 1.521 ± 0.054 2.239 ± 0.120 1.179 ± 0.050
Rev0 1.528 ± 0.094 1.990 ± 0.141 0.894 ± 0.050
Ria0 1.000 ± 0.071 2.194 ± 0.085 0.920 ± 0.045
San-10 0.272 ± 0.038 1.831 ± 0.054 1.020 ± 0.050
Scm-0 0.131 ± 0.009 1.343 ± 0.131 0.837 ± 0.050
Sdv3 0.741 ± 0.089 1.888 ± 0.160 0.720 ± 0.071 1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar.

Anejo 3
285
Continuación Tabla A3.4.
Individuos RWm1 IWm
2 SWm3
Ses-0 0.458 ± 0.039 2.259 ± 0.129 0.814 ± 0.085
Tam0 0.722 ± 0.065 1.679 ± 0.085 0.869 ± 0.063
Vad-0 0.219 ± 0.029 2.131 ± 0.143 0.958 ± 0.046
Vae2 1.086 ± 0.064 3.133 ± 0.087 0.649 ± 0.033
Vav-0 0.078 ± 0.009 0.908 ± 0.080 0.500 ± 0.036
Vaz0 1.802 ± 0.113 3.481 ± 0.165 1.114 ± 0.126
Vdm0 0.738 ± 0.066 2.441 ± 0.140 1.238 ± 0.064
Ver-5 0.195 ± 0.012 1.619 ± 0.056 0.992 ± 0.070
Vim-0 0.561 ± 0.059 1.427 ± 0.094 0.731 ± 0.047
Vis-0 0.484 ± 0.066 1.521 ± 0.266 1.104 ± 0.069
Voz0 0.634 ± 0.050 2.735 ± 0.056 1.216 ± 0.042 1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar.
Tabla A3.5. Valores (media±error estándar) de la acumulación de Cdc1-CMV y Lro-CMV y
de tolerancia a sus infecciones de cada individuo de arabidopsis.
Individuo Acumulación de Cdc1-CMV1
Acumulación de Lro-CMV1
Tolerancia a Cdc1-CMV2
Tolerancia a Lro-CMV2
Adc-5 15.548 ± 1.260 9.905 ± 0.471 0.622 ± 0.062 0.646 ± 0.078
Adm-0 18.357 ± 0.627 3.236 ± 0.914 0.490 ± 0.087 0.443 ± 0.090
Agu-1 19.720 ± 0.917 11.835 ± 1.133 0.476 ± 0.076 0.336 ± 0.065
Ala-0 23.528 ± 1.085 9.529 ± 0.974 0.553 ± 0.133 0.397 ± 0.053
Ali-1 41.252 ± 1.800 17.315 ± 1.634 0.360 ± 0.037 0.486 ± 0.028
All-0 56.283 ± 2.214 16.633 ± 1.300 0.347 ± 0.069 0.329 ± 0.074
Alm-0 30.306 ± 1.446 9.509 ± 0.564 0.477 ± 0.049 0.334 ± 0.072
Alo-0 34.657 ± 1.363 5.297 ± 0.327 0.487 ± 0.054 0.490 ± 0.063
Alr-0 1.700 ± 0.135 2.394 ± 0.395 0.390 ± 0.023 0.393 ± 0.051
Ara4 22.176 ± 3.728 5.429 ± 0.481 0.338 ± 0.090 0.360 ± 0.071
Bea-0 29.115 ± 3.180 14.333 ± 1.131 0.352 ± 0.084 0.419 ± 0.080
Ber-0 14.569 ± 2.403 9.300 ± 0.855 0.548 ± 0.075 0.501 ± 0.078
Bis-0 35.486 ± 2.796 10.448 ± 1.039 0.619 ± 0.056 0.588 ± 0.082
Cad-0 21.557 ± 1.582 7.515 ± 0.670 0.652 ± 0.074 0.680 ± 0.029
Cal-0 7.767 ± 0.506 4.239 ± 0.381 0.531 ± 0.038 0.538 ± 0.026
Cap-1 20.434 ± 2.363 3.513 ± 0.194 0.511 ± 0.023 0.506 ± 0.044
Car-1 25.052 ± 0.646 2.838 ± 0.194 0.332 ± 0.128 0.290 ± 0.066
Cdc3 19.033 ± 3.223 19.922 ± 1.738 0.351 ± 0.047 0.234 ± 0.037 1 Valores de acumulación medida en µgARN/g p.f. 2 Valores de tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm).
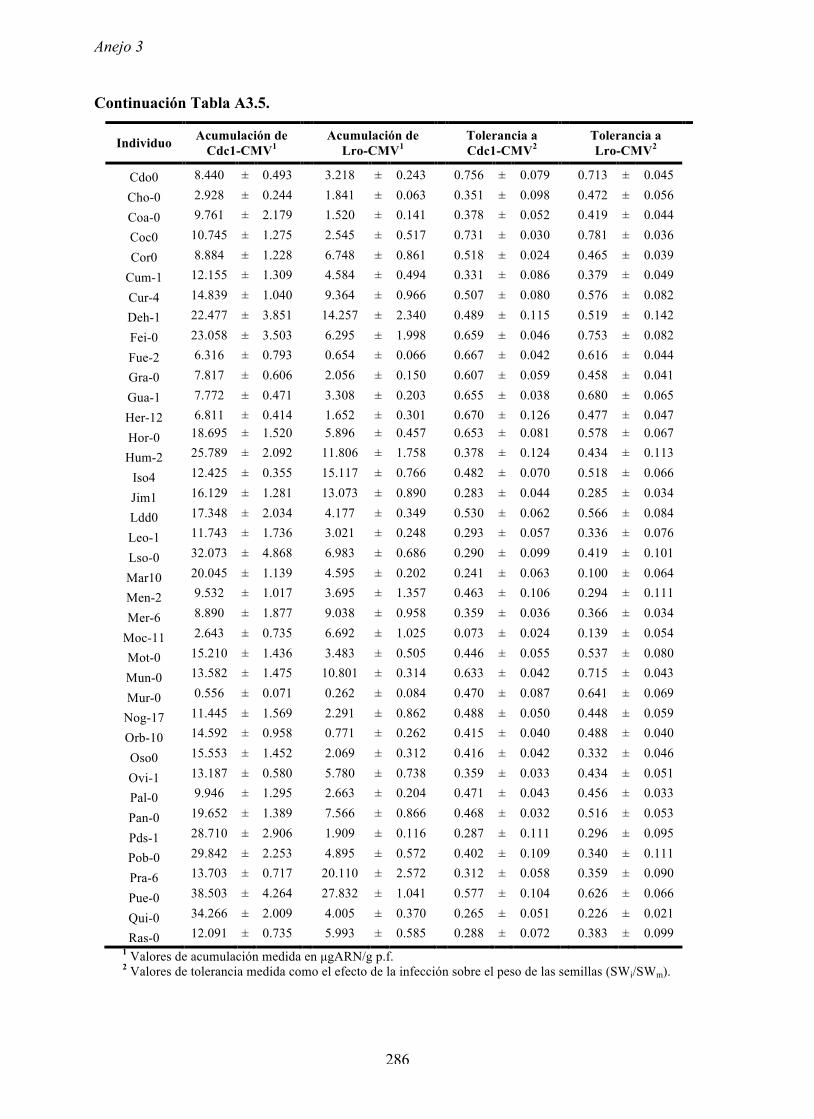
Anejo 3
286
Continuación Tabla A3.5.
Individuo Acumulación de Cdc1-CMV1
Acumulación de Lro-CMV1
Tolerancia a Cdc1-CMV2
Tolerancia a Lro-CMV2
Cdo0 8.440 ± 0.493 3.218 ± 0.243 0.756 ± 0.079 0.713 ± 0.045
Cho-0 2.928 ± 0.244 1.841 ± 0.063 0.351 ± 0.098 0.472 ± 0.056
Coa-0 9.761 ± 2.179 1.520 ± 0.141 0.378 ± 0.052 0.419 ± 0.044
Coc0 10.745 ± 1.275 2.545 ± 0.517 0.731 ± 0.030 0.781 ± 0.036
Cor0 8.884 ± 1.228 6.748 ± 0.861 0.518 ± 0.024 0.465 ± 0.039
Cum-1 12.155 ± 1.309 4.584 ± 0.494 0.331 ± 0.086 0.379 ± 0.049
Cur-4 14.839 ± 1.040 9.364 ± 0.966 0.507 ± 0.080 0.576 ± 0.082
Deh-1 22.477 ± 3.851 14.257 ± 2.340 0.489 ± 0.115 0.519 ± 0.142
Fei-0 23.058 ± 3.503 6.295 ± 1.998 0.659 ± 0.046 0.753 ± 0.082
Fue-2 6.316 ± 0.793 0.654 ± 0.066 0.667 ± 0.042 0.616 ± 0.044
Gra-0 7.817 ± 0.606 2.056 ± 0.150 0.607 ± 0.059 0.458 ± 0.041
Gua-1 7.772 ± 0.471 3.308 ± 0.203 0.655 ± 0.038 0.680 ± 0.065
Her-12 6.811 ± 0.414 1.652 ± 0.301 0.670 ± 0.126 0.477 ± 0.047
Hor-0 18.695 ± 1.520 5.896 ± 0.457 0.653 ± 0.081 0.578 ± 0.067
Hum-2 25.789 ± 2.092 11.806 ± 1.758 0.378 ± 0.124 0.434 ± 0.113
Iso4 12.425 ± 0.355 15.117 ± 0.766 0.482 ± 0.070 0.518 ± 0.066
Jim1 16.129 ± 1.281 13.073 ± 0.890 0.283 ± 0.044 0.285 ± 0.034
Ldd0 17.348 ± 2.034 4.177 ± 0.349 0.530 ± 0.062 0.566 ± 0.084
Leo-1 11.743 ± 1.736 3.021 ± 0.248 0.293 ± 0.057 0.336 ± 0.076
Lso-0 32.073 ± 4.868 6.983 ± 0.686 0.290 ± 0.099 0.419 ± 0.101
Mar10 20.045 ± 1.139 4.595 ± 0.202 0.241 ± 0.063 0.100 ± 0.064
Men-2 9.532 ± 1.017 3.695 ± 1.357 0.463 ± 0.106 0.294 ± 0.111
Mer-6 8.890 ± 1.877 9.038 ± 0.958 0.359 ± 0.036 0.366 ± 0.034
Moc-11 2.643 ± 0.735 6.692 ± 1.025 0.073 ± 0.024 0.139 ± 0.054
Mot-0 15.210 ± 1.436 3.483 ± 0.505 0.446 ± 0.055 0.537 ± 0.080
Mun-0 13.582 ± 1.475 10.801 ± 0.314 0.633 ± 0.042 0.715 ± 0.043
Mur-0 0.556 ± 0.071 0.262 ± 0.084 0.470 ± 0.087 0.641 ± 0.069
Nog-17 11.445 ± 1.569 2.291 ± 0.862 0.488 ± 0.050 0.448 ± 0.059
Orb-10 14.592 ± 0.958 0.771 ± 0.262 0.415 ± 0.040 0.488 ± 0.040
Oso0 15.553 ± 1.452 2.069 ± 0.312 0.416 ± 0.042 0.332 ± 0.046
Ovi-1 13.187 ± 0.580 5.780 ± 0.738 0.359 ± 0.033 0.434 ± 0.051
Pal-0 9.946 ± 1.295 2.663 ± 0.204 0.471 ± 0.043 0.456 ± 0.033
Pan-0 19.652 ± 1.389 7.566 ± 0.866 0.468 ± 0.032 0.516 ± 0.053
Pds-1 28.710 ± 2.906 1.909 ± 0.116 0.287 ± 0.111 0.296 ± 0.095
Pob-0 29.842 ± 2.253 4.895 ± 0.572 0.402 ± 0.109 0.340 ± 0.111
Pra-6 13.703 ± 0.717 20.110 ± 2.572 0.312 ± 0.058 0.359 ± 0.090
Pue-0 38.503 ± 4.264 27.832 ± 1.041 0.577 ± 0.104 0.626 ± 0.066
Qui-0 34.266 ± 2.009 4.005 ± 0.370 0.265 ± 0.051 0.226 ± 0.021
Ras-0 12.091 ± 0.735 5.993 ± 0.585 0.288 ± 0.072 0.383 ± 0.099 1 Valores de acumulación medida en µgARN/g p.f. 2 Valores de tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm).
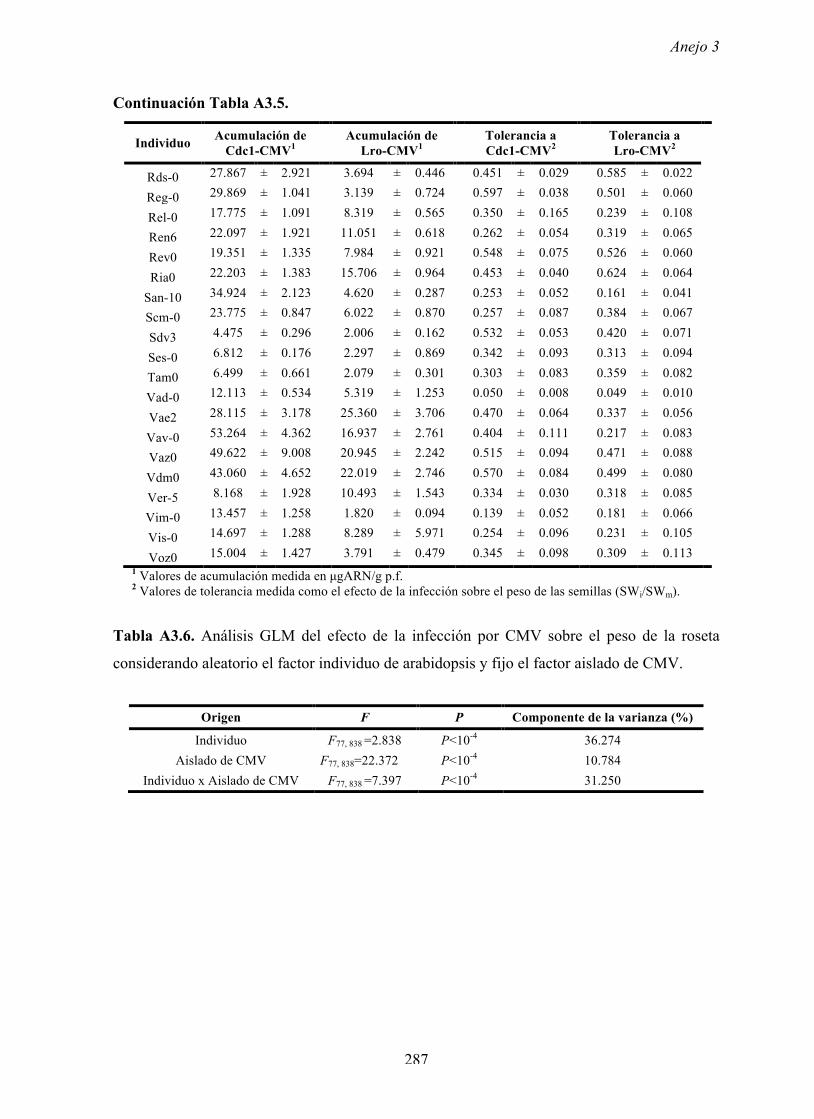
Anejo 3
287
Continuación Tabla A3.5.
Individuo Acumulación de Cdc1-CMV1
Acumulación de Lro-CMV1
Tolerancia a Cdc1-CMV2
Tolerancia a Lro-CMV2
Rds-0 27.867 ± 2.921 3.694 ± 0.446 0.451 ± 0.029 0.585 ± 0.022
Reg-0 29.869 ± 1.041 3.139 ± 0.724 0.597 ± 0.038 0.501 ± 0.060
Rel-0 17.775 ± 1.091 8.319 ± 0.565 0.350 ± 0.165 0.239 ± 0.108
Ren6 22.097 ± 1.921 11.051 ± 0.618 0.262 ± 0.054 0.319 ± 0.065
Rev0 19.351 ± 1.335 7.984 ± 0.921 0.548 ± 0.075 0.526 ± 0.060
Ria0 22.203 ± 1.383 15.706 ± 0.964 0.453 ± 0.040 0.624 ± 0.064
San-10 34.924 ± 2.123 4.620 ± 0.287 0.253 ± 0.052 0.161 ± 0.041
Scm-0 23.775 ± 0.847 6.022 ± 0.870 0.257 ± 0.087 0.384 ± 0.067
Sdv3 4.475 ± 0.296 2.006 ± 0.162 0.532 ± 0.053 0.420 ± 0.071
Ses-0 6.812 ± 0.176 2.297 ± 0.869 0.342 ± 0.093 0.313 ± 0.094
Tam0 6.499 ± 0.661 2.079 ± 0.301 0.303 ± 0.083 0.359 ± 0.082
Vad-0 12.113 ± 0.534 5.319 ± 1.253 0.050 ± 0.008 0.049 ± 0.010
Vae2 28.115 ± 3.178 25.360 ± 3.706 0.470 ± 0.064 0.337 ± 0.056
Vav-0 53.264 ± 4.362 16.937 ± 2.761 0.404 ± 0.111 0.217 ± 0.083
Vaz0 49.622 ± 9.008 20.945 ± 2.242 0.515 ± 0.094 0.471 ± 0.088
Vdm0 43.060 ± 4.652 22.019 ± 2.746 0.570 ± 0.084 0.499 ± 0.080
Ver-5 8.168 ± 1.928 10.493 ± 1.543 0.334 ± 0.030 0.318 ± 0.085
Vim-0 13.457 ± 1.258 1.820 ± 0.094 0.139 ± 0.052 0.181 ± 0.066
Vis-0 14.697 ± 1.288 8.289 ± 5.971 0.254 ± 0.096 0.231 ± 0.105
Voz0 15.004 ± 1.427 3.791 ± 0.479 0.345 ± 0.098 0.309 ± 0.113 1 Valores de acumulación medida en µgARN/g p.f. 2 Valores de tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm).
Tabla A3.6. Análisis GLM del efecto de la infección por CMV sobre el peso de la roseta
considerando aleatorio el factor individuo de arabidopsis y fijo el factor aislado de CMV.
Origen F P Componente de la varianza (%)
Individuo F77, 838 =2.838 P<10-4 36.274 Aislado de CMV F77, 838=22.372 P<10-4 10.784
Individuo x Aislado de CMV F77, 838 =7.397 P<10-4 31.250

Anejo 3
288
Tabla A3.7. Análisis GLM del efecto de la infección por CMV sobre el peso de la
inflorescencia sin semillas considerando aleatorio el factor individuo de arabidopsis y fijo el
factor aislado de CMV.
Origen F P Componente de la varianza (%)
Individuo F77, 838 =1.937 P=0.002 19.791 Aislado de CMV F77, 838 =33.564 P<10-4 18.750
Individuo x Aislado de CMV F77, 838 =11.429 P<10-4 40.625
Tabla A3.8. Medias±error estándar del efecto de la infección de Cdc1-CMV y Lro-CMV
sobre el peso de la roseta y el peso de la inflorescencia sin semillas estimadas a partir de los
valores de los 77 individuos.
Aislado de CMV RWi/RWm1 IWi/IWm
2
Cdc1-CMV
Lro-CMV
0.360±0.015
0.512±0.015
0.447±0.013
0.640±0.013 1 RWi/RWm Efecto de la infección sobre el peso de la roseta 2 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas
Tabla A3.9. Coeficiente de correlación de Pearson y de Spearman con la corrección de
Dutilleul de las correlaciones entre los valores de RWm, IWm y SWm de las plantas sin infectar.
La significancia está indicada como: no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y ***
P>0.001.
RWm
1 IWm2
RWm1
IWm2 0.668/0.750***
SWm3 0.199/0.340ns 0.582/0.524**
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar.

Anejo 3
289
Tabla A3.10. Coeficiente de correlación de Pearson y de Spearman con la corrección de
Dutilleul de las correlaciones entre los valores de las diferentes variables de las plantas
infectadas por Cdc1-CMV. La significancia está indicada como: no significatva (ns) P>0.1, *
P<0.1, ** P<0.05 y *** P>0.001.
Acumulación1 RWi/RWm
2 IWi/IWm3
Acumulación1
RWi/RWm
2 0.086/-0.038ns
IWi/IWm3 -0.149/-0.122ns 0.641/0.734***
Tolerancia4 -0.018/-0.051ns 0.184/0.324ns 0.550/0.564*** 1 Valores de acumulación medida en µgARN/g p.f.. 2 RWi/RWm Efecto de la infección sobre el peso de la roseta. 3 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas. 4 Valores de tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm).
Tabla A3.11. Coeficiente de correlación de Pearson y Spearman con la corrección de
Dutilleul de las correlaciones entre los valores de las diferentes variables de las plantas
infectadas por Lro-CMV. La significancia está indicada como: no significativa (ns) P>0.1, *
P<0.1, ** P<0.05 y *** P>0.001.
Acumulación1 RWi/RWm
2 IWi/IWm3
Acumulación1
RWi/RWm2 -0.043/0.009ns
IWi/IWm3 0.015/0.033ns 0.411/0.419**
Tolerancia4 -0.006/-0.026ns 0.303/0.266** 0.481/0.433 *** 1 Acumulación medida en µgARN/g p.f.. 2 RWi/RWm Efecto de la infección sobre el peso de la roseta. 3 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas. 4 Valores de tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm).
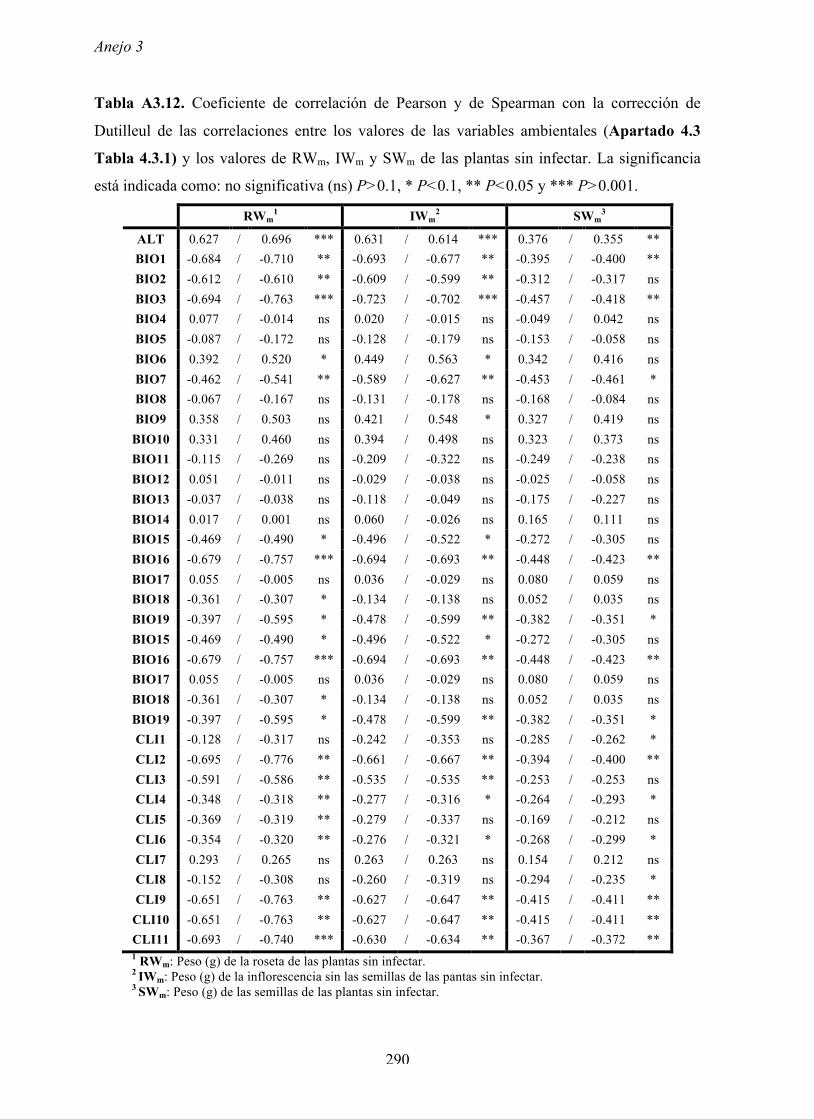
Anejo 3
290
Tabla A3.12. Coeficiente de correlación de Pearson y de Spearman con la corrección de
Dutilleul de las correlaciones entre los valores de las variables ambientales (Apartado 4.3
Tabla 4.3.1) y los valores de RWm, IWm y SWm de las plantas sin infectar. La significancia
está indicada como: no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y *** P>0.001.
RWm1 IWm
2 SWm3
ALT 0.627 / 0.696 *** 0.631 / 0.614 *** 0.376 / 0.355 ** BIO1 -0.684 / -0.710 ** -0.693 / -0.677 ** -0.395 / -0.400 ** BIO2 -0.612 / -0.610 ** -0.609 / -0.599 ** -0.312 / -0.317 ns BIO3 -0.694 / -0.763 *** -0.723 / -0.702 *** -0.457 / -0.418 ** BIO4 0.077 / -0.014 ns 0.020 / -0.015 ns -0.049 / 0.042 ns BIO5 -0.087 / -0.172 ns -0.128 / -0.179 ns -0.153 / -0.058 ns BIO6 0.392 / 0.520 * 0.449 / 0.563 * 0.342 / 0.416 ns BIO7 -0.462 / -0.541 ** -0.589 / -0.627 ** -0.453 / -0.461 * BIO8 -0.067 / -0.167 ns -0.131 / -0.178 ns -0.168 / -0.084 ns BIO9 0.358 / 0.503 ns 0.421 / 0.548 * 0.327 / 0.419 ns
BIO10 0.331 / 0.460 ns 0.394 / 0.498 ns 0.323 / 0.373 ns BIO11 -0.115 / -0.269 ns -0.209 / -0.322 ns -0.249 / -0.238 ns BIO12 0.051 / -0.011 ns -0.029 / -0.038 ns -0.025 / -0.058 ns BIO13 -0.037 / -0.038 ns -0.118 / -0.049 ns -0.175 / -0.227 ns BIO14 0.017 / 0.001 ns 0.060 / -0.026 ns 0.165 / 0.111 ns BIO15 -0.469 / -0.490 * -0.496 / -0.522 * -0.272 / -0.305 ns BIO16 -0.679 / -0.757 *** -0.694 / -0.693 ** -0.448 / -0.423 ** BIO17 0.055 / -0.005 ns 0.036 / -0.029 ns 0.080 / 0.059 ns BIO18 -0.361 / -0.307 * -0.134 / -0.138 ns 0.052 / 0.035 ns BIO19 -0.397 / -0.595 * -0.478 / -0.599 ** -0.382 / -0.351 * BIO15 -0.469 / -0.490 * -0.496 / -0.522 * -0.272 / -0.305 ns BIO16 -0.679 / -0.757 *** -0.694 / -0.693 ** -0.448 / -0.423 ** BIO17 0.055 / -0.005 ns 0.036 / -0.029 ns 0.080 / 0.059 ns BIO18 -0.361 / -0.307 * -0.134 / -0.138 ns 0.052 / 0.035 ns BIO19 -0.397 / -0.595 * -0.478 / -0.599 ** -0.382 / -0.351 * CLI1 -0.128 / -0.317 ns -0.242 / -0.353 ns -0.285 / -0.262 * CLI2 -0.695 / -0.776 ** -0.661 / -0.667 ** -0.394 / -0.400 ** CLI3 -0.591 / -0.586 ** -0.535 / -0.535 ** -0.253 / -0.253 ns CLI4 -0.348 / -0.318 ** -0.277 / -0.316 * -0.264 / -0.293 * CLI5 -0.369 / -0.319 ** -0.279 / -0.337 ns -0.169 / -0.212 ns CLI6 -0.354 / -0.320 ** -0.276 / -0.321 * -0.268 / -0.299 * CLI7 0.293 / 0.265 ns 0.263 / 0.263 ns 0.154 / 0.212 ns CLI8 -0.152 / -0.308 ns -0.260 / -0.319 ns -0.294 / -0.235 * CLI9 -0.651 / -0.763 ** -0.627 / -0.647 ** -0.415 / -0.411 **
CLI10 -0.651 / -0.763 ** -0.627 / -0.647 ** -0.415 / -0.411 ** CLI11 -0.693 / -0.740 *** -0.630 / -0.634 ** -0.367 / -0.372 ** 1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar.
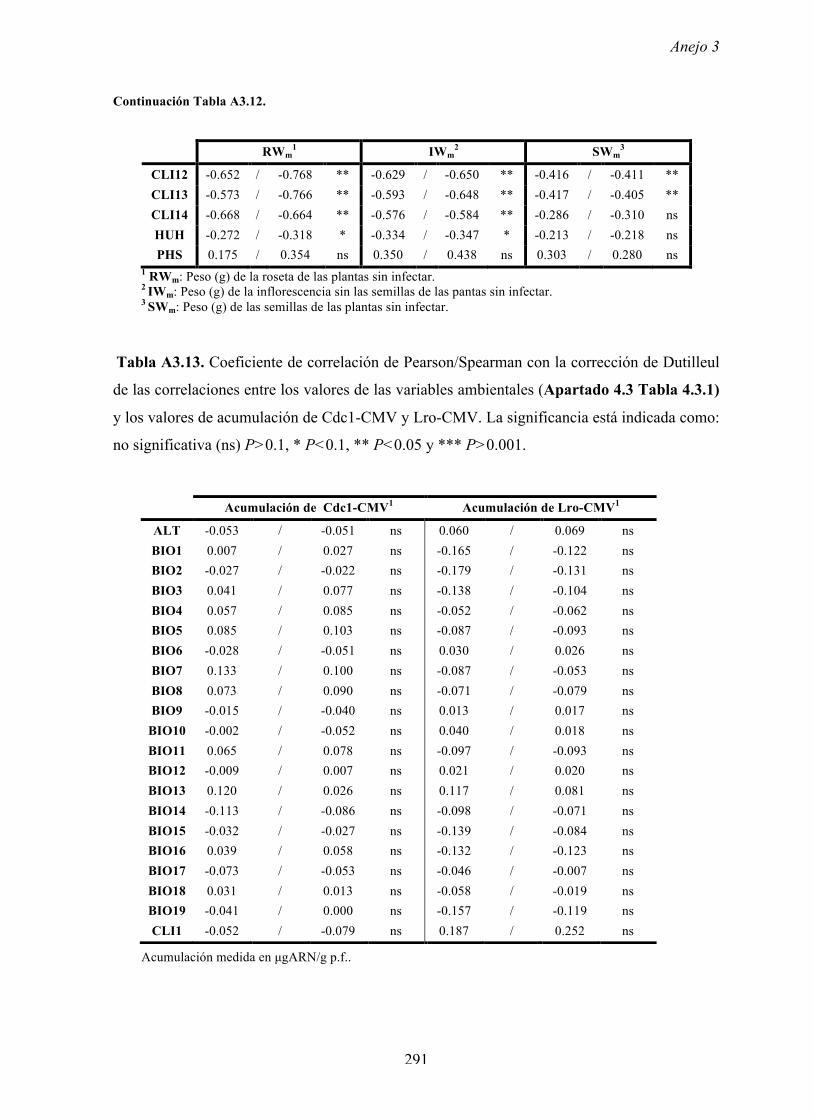
Anejo 3
291
Continuación Tabla A3.12.
RWm1 IWm
2 SWm3
CLI12 -0.652 / -0.768 ** -0.629 / -0.650 ** -0.416 / -0.411 ** CLI13 -0.573 / -0.766 ** -0.593 / -0.648 ** -0.417 / -0.405 ** CLI14 -0.668 / -0.664 ** -0.576 / -0.584 ** -0.286 / -0.310 ns HUH -0.272 / -0.318 * -0.334 / -0.347 * -0.213 / -0.218 ns PHS 0.175 / 0.354 ns 0.350 / 0.438 ns 0.303 / 0.280 ns
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar.
Tabla A3.13. Coeficiente de correlación de Pearson/Spearman con la corrección de Dutilleul
de las correlaciones entre los valores de las variables ambientales (Apartado 4.3 Tabla 4.3.1)
y los valores de acumulación de Cdc1-CMV y Lro-CMV. La significancia está indicada como:
no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y *** P>0.001.
Acumulación medida en µgARN/g p.f..
Acumulación de Cdc1-CMV1 Acumulación de Lro-CMV1
ALT -0.053 / -0.051 ns 0.060 / 0.069 ns BIO1 0.007 / 0.027 ns -0.165 / -0.122 ns BIO2 -0.027 / -0.022 ns -0.179 / -0.131 ns BIO3 0.041 / 0.077 ns -0.138 / -0.104 ns BIO4 0.057 / 0.085 ns -0.052 / -0.062 ns BIO5 0.085 / 0.103 ns -0.087 / -0.093 ns BIO6 -0.028 / -0.051 ns 0.030 / 0.026 ns BIO7 0.133 / 0.100 ns -0.087 / -0.053 ns BIO8 0.073 / 0.090 ns -0.071 / -0.079 ns BIO9 -0.015 / -0.040 ns 0.013 / 0.017 ns
BIO10 -0.002 / -0.052 ns 0.040 / 0.018 ns BIO11 0.065 / 0.078 ns -0.097 / -0.093 ns BIO12 -0.009 / 0.007 ns 0.021 / 0.020 ns BIO13 0.120 / 0.026 ns 0.117 / 0.081 ns BIO14 -0.113 / -0.086 ns -0.098 / -0.071 ns BIO15 -0.032 / -0.027 ns -0.139 / -0.084 ns BIO16 0.039 / 0.058 ns -0.132 / -0.123 ns BIO17 -0.073 / -0.053 ns -0.046 / -0.007 ns BIO18 0.031 / 0.013 ns -0.058 / -0.019 ns BIO19 -0.041 / 0.000 ns -0.157 / -0.119 ns CLI1 -0.052 / -0.079 ns 0.187 / 0.252 ns

Anejo 3
292
Continuación Tabla A3.13.
Acumulación medida en µgARN/g p.f..
Acumulación de Cdc1-CMV1 Acumulación de Lro-CMV1
ALT -0.053 / -0.051 ns 0.060 / 0.069 ns BIO1 0.007 / 0.027 ns -0.165 / -0.122 ns BIO2 -0.027 / -0.022 ns -0.179 / -0.131 ns BIO3 0.041 / 0.077 ns -0.138 / -0.104 ns BIO4 0.057 / 0.085 ns -0.052 / -0.062 ns BIO5 0.085 / 0.103 ns -0.087 / -0.093 ns BIO6 -0.028 / -0.051 ns 0.030 / 0.026 ns BIO7 0.133 / 0.100 ns -0.087 / -0.053 ns BIO8 0.073 / 0.090 ns -0.071 / -0.079 ns BIO9 -0.015 / -0.040 ns 0.013 / 0.017 ns
BIO10 -0.002 / -0.052 ns 0.040 / 0.018 ns BIO11 0.065 / 0.078 ns -0.097 / -0.093 ns BIO12 -0.009 / 0.007 ns 0.021 / 0.020 ns BIO13 0.120 / 0.026 ns 0.117 / 0.081 ns BIO14 -0.113 / -0.086 ns -0.098 / -0.071 ns BIO15 -0.032 / -0.027 ns -0.139 / -0.084 ns BIO16 0.039 / 0.058 ns -0.132 / -0.123 ns BIO17 -0.073 / -0.053 ns -0.046 / -0.007 ns BIO18 0.031 / 0.013 ns -0.058 / -0.019 ns BIO19 -0.041 / 0.000 ns -0.157 / -0.119 ns CLI1 -0.052 / -0.079 ns 0.187 / 0.252 ns CLI2 -0.106 / -0.089 ns 0.045 / 0.054 ns CLI3 -0.087 / -0.048 ns -0.087 / -0.039 ns CLI4 0.027 / 0.046 ns 0.088 / 0.075 ns CLI5 0.004 / -0.013 ns 0.057 / 0.067 ns CLI6 0.019 / 0.028 ns 0.085 / 0.077 ns CLI7 0.043 / -0.036 ns 0.091 / 0.207 ns CLI8 -0.065 / -0.056 ns 0.228 / 0.256 ** CLI9 -0.099 / -0.093 ns 0.080 / 0.073 ns
CLI10 -0.099 / -0.093 ns 0.080 / 0.073 ns CLI11 -0.099 / -0.079 ns 0.023 / 0.032 ns CLI12 -0.108 / -0.091 ns 0.083 / 0.078 ns CLI13 -0.096 / -0.104 ns 0.232 / 0.072 ** CLI14 -0.093 / -0.060 ns -0.051 / -0.043 ns HUH 0.063 / 0.010 ns 0.038 / -0.048 ns PHS -0.117 / -0.107 ns -0.069 / -0.090 ns
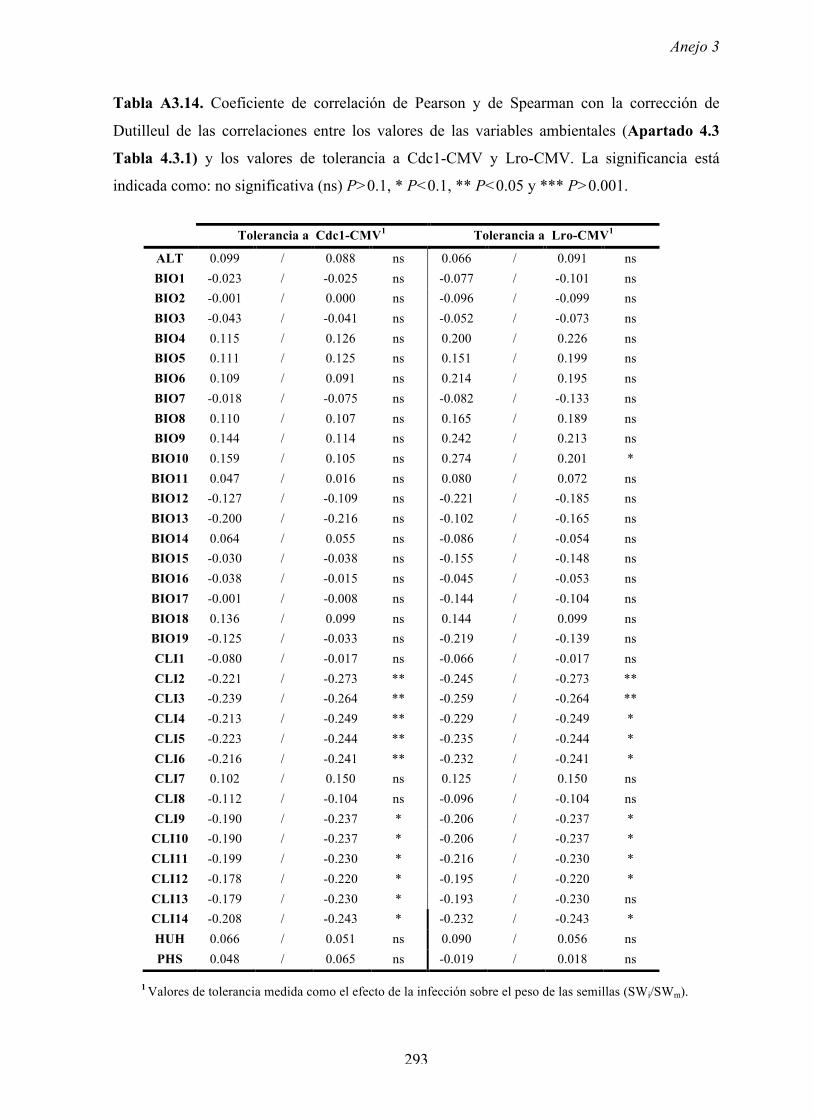
Anejo 3
293
Tabla A3.14. Coeficiente de correlación de Pearson y de Spearman con la corrección de
Dutilleul de las correlaciones entre los valores de las variables ambientales (Apartado 4.3
Tabla 4.3.1) y los valores de tolerancia a Cdc1-CMV y Lro-CMV. La significancia está
indicada como: no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y *** P>0.001.
1 Valores de tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm).
Tolerancia a Cdc1-CMV1 Tolerancia a Lro-CMV1
ALT 0.099 / 0.088 ns 0.066 / 0.091 ns BIO1 -0.023 / -0.025 ns -0.077 / -0.101 ns BIO2 -0.001 / 0.000 ns -0.096 / -0.099 ns BIO3 -0.043 / -0.041 ns -0.052 / -0.073 ns BIO4 0.115 / 0.126 ns 0.200 / 0.226 ns BIO5 0.111 / 0.125 ns 0.151 / 0.199 ns BIO6 0.109 / 0.091 ns 0.214 / 0.195 ns BIO7 -0.018 / -0.075 ns -0.082 / -0.133 ns BIO8 0.110 / 0.107 ns 0.165 / 0.189 ns BIO9 0.144 / 0.114 ns 0.242 / 0.213 ns
BIO10 0.159 / 0.105 ns 0.274 / 0.201 * BIO11 0.047 / 0.016 ns 0.080 / 0.072 ns BIO12 -0.127 / -0.109 ns -0.221 / -0.185 ns BIO13 -0.200 / -0.216 ns -0.102 / -0.165 ns BIO14 0.064 / 0.055 ns -0.086 / -0.054 ns BIO15 -0.030 / -0.038 ns -0.155 / -0.148 ns BIO16 -0.038 / -0.015 ns -0.045 / -0.053 ns BIO17 -0.001 / -0.008 ns -0.144 / -0.104 ns BIO18 0.136 / 0.099 ns 0.144 / 0.099 ns BIO19 -0.125 / -0.033 ns -0.219 / -0.139 ns CLI1 -0.080 / -0.017 ns -0.066 / -0.017 ns CLI2 -0.221 / -0.273 ** -0.245 / -0.273 ** CLI3 -0.239 / -0.264 ** -0.259 / -0.264 ** CLI4 -0.213 / -0.249 ** -0.229 / -0.249 * CLI5 -0.223 / -0.244 ** -0.235 / -0.244 * CLI6 -0.216 / -0.241 ** -0.232 / -0.241 * CLI7 0.102 / 0.150 ns 0.125 / 0.150 ns CLI8 -0.112 / -0.104 ns -0.096 / -0.104 ns CLI9 -0.190 / -0.237 * -0.206 / -0.237 *
CLI10 -0.190 / -0.237 * -0.206 / -0.237 * CLI11 -0.199 / -0.230 * -0.216 / -0.230 * CLI12 -0.178 / -0.220 * -0.195 / -0.220 * CLI13 -0.179 / -0.230 * -0.193 / -0.230 ns CLI14 -0.208 / -0.243 * -0.232 / -0.243 * HUH 0.066 / 0.051 ns 0.090 / 0.056 ns PHS 0.048 / 0.065 ns -0.019 / 0.018 ns

Anejo 3
294
Tabla A3.15. Coeficiente de correlación de Pearson y de Spearman con la corrección de
Dutilleul de las correlaciones entre los valores de las variables ambientales (Apartado 4.3
Tabla 4.3.1) y los valores del efecto de Cdc1-CMV y Lro-CMV sobre la roseta. La
significancia está indicada como: no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y ***
P>0.001.
1 RWi/RWm Efecto de la infección sobre el peso de la roseta.
RWi/RWm Cdc1-CMV1 RWi/RWm Lro-CMV1
ALT 0.542 / 0.513 ** 0.443 / 0.376 ** BIO1 0.555 / 0.524 ** 0.466 / 0.405 ** BIO2 0.501 / 0.454 ** 0.460 / 0.401 ** BIO3 -0.493 / -0.455 ** -0.369 / -0.289 ** BIO4 -0.035 / -0.098 ns -0.280 / -0.300 ** BIO5 -0.037 / -0.112 ns -0.254 / -0.294 * BIO6 -0.014 / -0.078 ns -0.256 / -0.322 * BIO7 0.408 / 0.391 * 0.419 / 0.405 * BIO8 0.387 / 0.331 ** 0.343 / 0.293 * BIO9 0.289 / 0.351 ns 0.267 / 0.247 ns
BIO10 0.269 / 0.356 ns 0.252 / 0.389 ns BIO11 0.284 / 0.276 ns 0.315 / 0.297 ns BIO12 -0.237 / -0.289 ns -0.244 / -0.273 ns BIO13 -0.216 / -0.267 ns -0.194 / -0.208 ns BIO14 0.389 / 0.354 ** 0.144 / 0.098 ns BIO15 -0.186 / -0.224 ns -0.157 / -0.192 ns BIO16 0.147 / 0.111 ns 0.066 / 0.001 ns BIO17 0.002 / -0.069 ns -0.089 / -0.064 ns BIO18 -0.022 / -0.090 ns -0.091 / -0.044 ns BIO19 0.060 / 0.140 ns -0.083 / -0.015 ns CLI1 0.111 / 0.230 ns 0.111 / 0.148 ns CLI2 -0.191 / -0.181 ns -0.123 / -0.032 ns CLI3 0.125 / 0.201 ns 0.094 / 0.128 ns CLI4 0.106 / 0.213 ns 0.098 / 0.131 ns CLI5 0.103 / 0.001 ns -0.084 / -0.189 ns CLI6 -0.103 / -0.060 ns -0.083 / -0.054 ns CLI7 0.136 / 0.103 ns 0.037 / -0.010 ns CLI8 -0.118 / -0.092 ns -0.040 / 0.007 ns CLI9 0.181 / 0.146 ns 0.022 / -0.012 ns
CLI10 0.166 / 0.124 ns 0.021 / -0.006 ns CLI11 0.166 / 0.124 ns 0.021 / -0.006 ns CLI12 0.159 / 0.114 ns 0.017 / -0.016 ns CLI13 -0.152 / -0.076 ns -0.025 / 0.041 ns CLI14 0.156 / 0.117 ns -0.006 / -0.044 ns
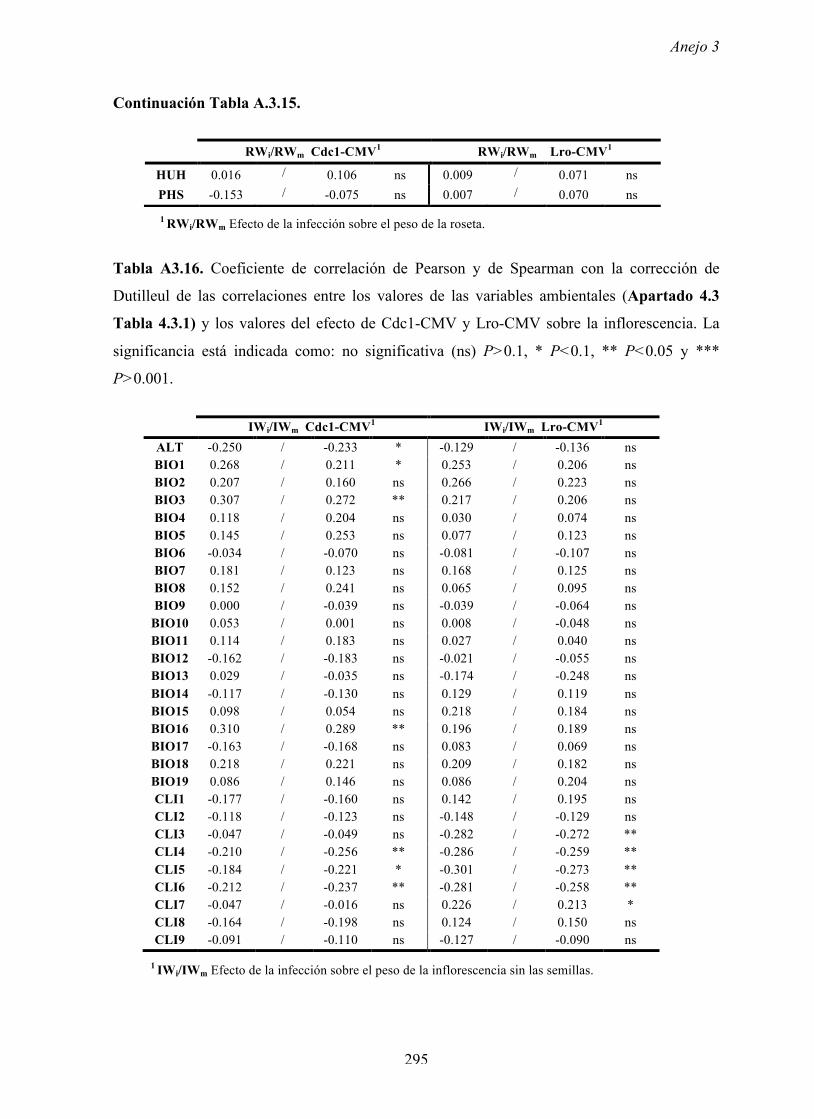
Anejo 3
295
Continuación Tabla A.3.15.
1 RWi/RWm Efecto de la infección sobre el peso de la roseta.
Tabla A3.16. Coeficiente de correlación de Pearson y de Spearman con la corrección de
Dutilleul de las correlaciones entre los valores de las variables ambientales (Apartado 4.3
Tabla 4.3.1) y los valores del efecto de Cdc1-CMV y Lro-CMV sobre la inflorescencia. La
significancia está indicada como: no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y ***
P>0.001.
1 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
RWi/RWm Cdc1-CMV1 RWi/RWm Lro-CMV1
HUH 0.016 / 0.106 ns 0.009 / 0.071 ns PHS -0.153 / -0.075 ns 0.007 / 0.070 ns
IWi/IWm Cdc1-CMV1 IWi/IWm Lro-CMV1
ALT -0.250 / -0.233 * -0.129 / -0.136 ns BIO1 0.268 / 0.211 * 0.253 / 0.206 ns BIO2 0.207 / 0.160 ns 0.266 / 0.223 ns BIO3 0.307 / 0.272 ** 0.217 / 0.206 ns BIO4 0.118 / 0.204 ns 0.030 / 0.074 ns BIO5 0.145 / 0.253 ns 0.077 / 0.123 ns BIO6 -0.034 / -0.070 ns -0.081 / -0.107 ns BIO7 0.181 / 0.123 ns 0.168 / 0.125 ns BIO8 0.152 / 0.241 ns 0.065 / 0.095 ns BIO9 0.000 / -0.039 ns -0.039 / -0.064 ns
BIO10 0.053 / 0.001 ns 0.008 / -0.048 ns BIO11 0.114 / 0.183 ns 0.027 / 0.040 ns BIO12 -0.162 / -0.183 ns -0.021 / -0.055 ns BIO13 0.029 / -0.035 ns -0.174 / -0.248 ns BIO14 -0.117 / -0.130 ns 0.129 / 0.119 ns BIO15 0.098 / 0.054 ns 0.218 / 0.184 ns BIO16 0.310 / 0.289 ** 0.196 / 0.189 ns BIO17 -0.163 / -0.168 ns 0.083 / 0.069 ns BIO18 0.218 / 0.221 ns 0.209 / 0.182 ns BIO19 0.086 / 0.146 ns 0.086 / 0.204 ns CLI1 -0.177 / -0.160 ns 0.142 / 0.195 ns CLI2 -0.118 / -0.123 ns -0.148 / -0.129 ns CLI3 -0.047 / -0.049 ns -0.282 / -0.272 ** CLI4 -0.210 / -0.256 ** -0.286 / -0.259 ** CLI5 -0.184 / -0.221 * -0.301 / -0.273 ** CLI6 -0.212 / -0.237 ** -0.281 / -0.258 ** CLI7 -0.047 / -0.016 ns 0.226 / 0.213 * CLI8 -0.164 / -0.198 ns 0.124 / 0.150 ns CLI9 -0.091 / -0.110 ns -0.127 / -0.090 ns

Anejo 3
296
Continuación Tabla A.3.16.
1 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
Tabla A3.17. Coeficientes de correlación estándar de los análisis univariantes SAR entre los
valores de las variables ambientales (Apartado 4.3 Tabla 4.3.1) y los de RWm, IWm y SWm
de los individuos sin infectar, acumulación de Cdc1-CMV y Lro1-CMV, tolerancia a Cdc1-
CMV y Lro-CMV y el efecto de la infección de Cdc1-CMV y Lro-CMV sobre la roseta y la
inflorescencia. La significancia está indicada como: no significativa (ns) P>0.1, * P<0.1, **
P<0.05 y *** P>0.001.
1RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar. 4 Acumulación medida en µgARN/g p.f.. 5Tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm). 6 RWi/RWm Efecto de la infección sobre el peso de la roseta. 7 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
IWi/IWm Cdc1-CMV1 IWi/IWm Lro-CMV1
CLI10 -0.091 / -0.110 ns -0.127 / -0.090 ns CLI11 -0.071 / -0.074 ns -0.159 / -0.137 * CLI12 -0.088 / -0.110 ns -0.111 / -0.076 ns CLI13 -0.150 / -0.088 ns -0.220 / -0.107 ** CLI14 -0.052 / -0.071 ns -0.186 / -0.158 * HUH 0.295 / 0.261 ** 0.091 / 0.064 ns PHS -0.059 / -0.082 ns 0.040 / 0.005 ns
RWm1 IWm
2 SWm3 Acumulación
Cdc1-CMV4 Acumulación
Lro-CMV4
ALT 0.602*** 0.579*** 0.332** -0.057ns 0.069ns BIO1 -0.684*** -0.674*** -0.365** 0.009ns -0.171ns BIO2 0.090ns 0.018ns 0.005ns -0.013ns 0.015ns BIO3 -0.090ns -0.150ns -0.165ns 0.120ns 0.123ns BIO4 -0.090ns 0.126ns 0.192ns -0.013ns -0.105ns BIO5 -0.450*** -0.475*** -0.239* -0.031ns -0.144ns BIO6 -0.671*** -0.656*** -0.402*** 0.042ns -0.138ns BIO7 0.127ns 0.103ns -0.363** -0.076ns -0.055ns BIO8 -0.378*** -0.158ns 0.000ns 0.029ns -0.073ns BIO9 -0.376*** -0.426*** 0.108ns -0.040ns -0.155ns
BIO10 -0.614*** -0.597*** -0.286** -0.024ns -0.183* BIO11 -0.686*** -0.692*** -0.421*** 0.043ns -0.145ns BIO12 0.087*** 0.031ns -0.0421ns 0.066ns -0.040ns BIO13 -0.044ns -0.073ns -0.109ns 0.092ns -0.082ns
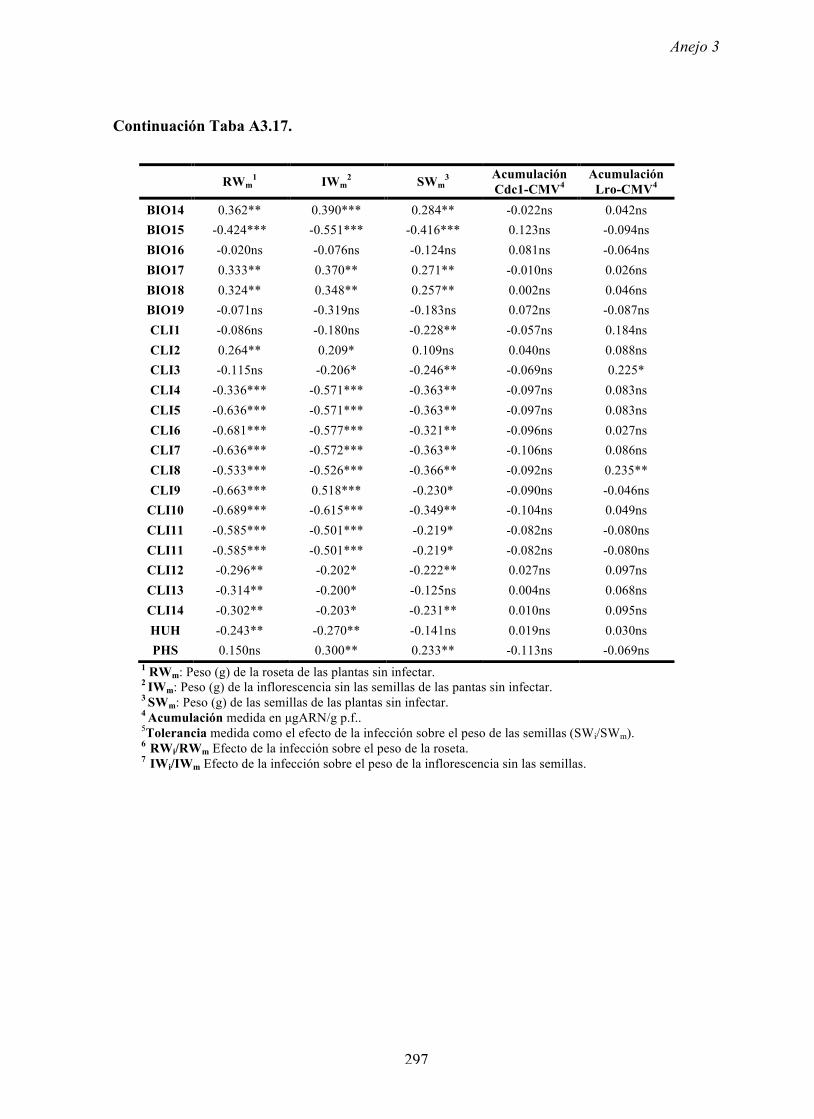
Anejo 3
297
Continuación Taba A3.17.
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar. 4 Acumulación medida en µgARN/g p.f.. 5Tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm). 6 RWi/RWm Efecto de la infección sobre el peso de la roseta. 7 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
RWm1 IWm
2 SWm3 Acumulación
Cdc1-CMV4 Acumulación
Lro-CMV4
BIO14 0.362** 0.390*** 0.284** -0.022ns 0.042ns BIO15 -0.424*** -0.551*** -0.416*** 0.123ns -0.094ns BIO16 -0.020ns -0.076ns -0.124ns 0.081ns -0.064ns BIO17 0.333** 0.370** 0.271** -0.010ns 0.026ns BIO18 0.324** 0.348** 0.257** 0.002ns 0.046ns BIO19 -0.071ns -0.319ns -0.183ns 0.072ns -0.087ns CLI1 -0.086ns -0.180ns -0.228** -0.057ns 0.184ns CLI2 0.264** 0.209* 0.109ns 0.040ns 0.088ns CLI3 -0.115ns -0.206* -0.246** -0.069ns 0.225* CLI4 -0.336*** -0.571*** -0.363** -0.097ns 0.083ns CLI5 -0.636*** -0.571*** -0.363** -0.097ns 0.083ns CLI6 -0.681*** -0.577*** -0.321** -0.096ns 0.027ns CLI7 -0.636*** -0.572*** -0.363** -0.106ns 0.086ns CLI8 -0.533*** -0.526*** -0.366** -0.092ns 0.235** CLI9 -0.663*** 0.518*** -0.230* -0.090ns -0.046ns
CLI10 -0.689*** -0.615*** -0.349** -0.104ns 0.049ns CLI11 -0.585*** -0.501*** -0.219* -0.082ns -0.080ns CLI11 -0.585*** -0.501*** -0.219* -0.082ns -0.080ns CLI12 -0.296** -0.202* -0.222** 0.027ns 0.097ns CLI13 -0.314** -0.200* -0.125ns 0.004ns 0.068ns CLI14 -0.302** -0.203* -0.231** 0.010ns 0.095ns HUH -0.243** -0.270** -0.141ns 0.019ns 0.030ns PHS 0.150ns 0.300** 0.233** -0.113ns -0.069ns

Anejo 3
298
Continuación Tabla A3.17.
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar. 4 Acumulación medida en µgARN/g p.f.. 5 Tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm). 6 RWi/RWm Efecto de la infección sobre el peso de la roseta. 7 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
Tolerancia
Cdc1-CMV5 Tolerancia Lro-CMV5
RWi/RWm Cdc1-CMV6
RWi/RWm Lro-CMV6
IWi/IWm Cdc1-CMV7
IWi/IWm Lro-CMV7
ALT 0.107ns 0.071ns -0.484*** 0.352*** -0.249** -0.119ns BIO1 -0.030ns .0.082ns 0.503*** 0-448*** 0.271** 0.239** BIO2 -0.123ns -0.214* -0.113ns -0.099ns -0.164ns -0.027ns BIO3 -0.195* -0.089ns 0.132ns -0.041ns 0.042ns -0.138ns BIO4 0.063ns -0.094ns -0.189ns -0.042ns -0.130ns 0.097ns BIO5 -0.034ns -0.165ns 0.274** 0.294*** 0.094ns 0.205* BIO6 -0.043ns -0.045ns 0.542*** 0.430*** 0.314** 0.188ns BIO7 0.001ns -0.147ns -0.182ns -0.007ns -0.174ns 0.058ns BIO8 0.124ns 0.128ns 0.296** 0.256*** 0.215* 0.184ns BIO9 -0.124ns -0.218* 0.263** 0.256* 0.088ns 0.184ns
BIO10 -0.008ns -0.105ns 0.406*** 0.405*** 0.207* 0.252** BIO11 -0.049ns -0.054ns 0.557*** 0.454*** 0.311** 0.207* BIO12 0.116ns 0.201ns 0.007ns 0.005ns 0.117ns 0.038ns BIO13 0.111ns 0.153ns 0.088ns 0.087ns 0.143ns 0.076ns BIO14 0.111ns 0.217* -0.226* -0.225* -0.031ns -0.069ns BIO15 -0.019ns -0.079* 0.382*** 0.326** 0.184ns 0.162ns BIO16 0.111ns 0.169ns 0.083ns 0.075ns 0.151ns -0.060ns BIO17 0.147ns 0.246** -0.208* -0.174ns 0.003ns -0.025ns BIO18 0.159ns 0.277** -0.181ns -0.148ns 0.056ns 0.017ns BIO19 0.052ns 0.091ns 0.103ns 0.074ns 0.114ns 0.033ns CLI1 -0.080ns -0.070ns -0.018ns -0.114ns -0.175ns 0.130ns CLI2 0.101ns 0.123ns -0.121ns -0.062ns -0.048ns 0.214* CLI3 -0.111ns -0.099ns 0.005ns -0.103ns -0.163ns 0.119ns CLI4 -0.190* -0.205* 0.168ns 0.025ns -0.088ns -0.124ns CLI5 -0.190* -0.205* 0.168ns 0.025ns -0.088ns -0.124ns CLI6 -0.199* -0.215* 0.183ns 0.029ns -0.069ns 0.154ns CLI7 -0.178ns -0.194* 0.160ns 0.021ns -0.086ns -0.109ns CLI8 -0.176ns -0.190* 0.060ns -0.067ns 0.147ns -0.210* CLI9 -0.209* -0.234** 0.150ns 0.082ns -0.050ns -0.178ns
CLI10 .0.222** -0.244** 0.137ns 0.045ns -0.116ns -0.143ns CLI11 -0.241** -0.260** 0.162ns 0.011ns -0.045ns -0.270** CLI12 -0.212* 0.225** -0.035ns -0.241*** -0.208* -0.267** CLI13 -0.222** 0.230** -0.011ns -0.231*** -0.182ns -0.275** CLI14 -0.214* -0.227** -0.032ns -0.265*** -0.209* -0.259** HUH 0.076ns 0.112ns 0.391*** 0.146ns 0.305** 0.104ns PHS 0.044ns -0.034ns -0.188ns -0.126ns -0.060ns 0.034ns

Anejo 3
299
Tabla A3.18. Coeficientes de correlación de los test de Mantel parciales entre los valores de
las variables ambientales (Apartado 4.3 Tabla 4.3.1) de RWm, IWm y SWm de los individuos
sin infectar, la acumulación de Cdc1-CMV y Lro1-CMV, la tolerancia a Cdc1-CMV y Lro-
CMV y el efecto de la infección de Cdc1-CMV y Lro-CMV sobre la roseta y la inflorescencia.
La significancia está indicada como: no significativa (ns) P>0.1, * P<0.1, ** P<0.05 y ***
P>0.001.
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar. 4 Acumulación medida en µgARN/g p.f.. 5Tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm). 6 RWi/RWm Efecto de la infección sobre el peso de la roseta. 7 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
RWm1 IWm
2 SWm3 Acumulación
Cdc1-CMV4 Acumulación
Lro-CMV4 ALT 0.363** 0.330** 0.249** 0.058ns 0.036ns BIO1 0.430** 0.397** 0.256** 0.091ns 0.063ns BIO2 -0.078ns -0.114** -0.043ns -0.090ns -0.058ns BIO3 -0.018ns -0.003ns 0.059ns 0.094ns 0.004ns BIO4 -0.048ns -0.058ns -0.051ns 0.044ns -0.003ns BIO5 0.211** 0.143** 0.030ns -0.029ns -0.011ns BIO6 0.410** 0.390** 0.304** 0.121ns 0.086ns BIO7 -0.073ns -0.101* -0.082ns -0.030ns -0.034ns BIO8 0.212** -0.095* -0.016ns -0.111ns -0.058ns BIO9 0.104ns 0.089ns 0.095ns 0.108ns 0.109ns
BIO10 0.358** 0.287** 0.147** 0.033ns 0.030ns BIO11 0.431** 0.445** 0.314** 0.143ns 0.087ns BIO12 0.039ns -0.119ns -0.016ns -0.085ns -0.043ns BIO13 -0.026ns -0.102ns -0.013ns -0.062ns -0.033ns BIO14 0.156** 0.080ns 0.067ns -0.007ns 0.000ns BIO15 0.060ns 0.227** 0.165** 0.023ns -0.003ns BIO16 -0.015ns -0.107* -0.001ns -0.071ns -0.035ns BIO17 0.127* 0.037ns 0.051ns -0.029ns -0.022ns BIO18 0.085ns -0.102* -0.032ns -0.044ns 0.005ns BIO19 -0.030ns 0.099ns 0.003ns -0.056ns -0.043ns CLI1 -0.014ns -0.085ns 0.021ns -0.019ns -0.045ns CLI2 0.115* -0.010ns 0.014ns -0.045ns -0.017ns CLI3 0.001ns -0.063ns 0.014ns -0.043ns -0.012ns CLI4 0.340** 0.252** 0.062ns 0.023ns -0.025ns CLI5 0.340** 0.252** 0.155** 0.023ns -0.025ns CLI6 0.421** 0.260** 0.155** 0.015ns 0.019ns CLI7 0.342** 0.255** 0.156** 0.030ns -0.032ns
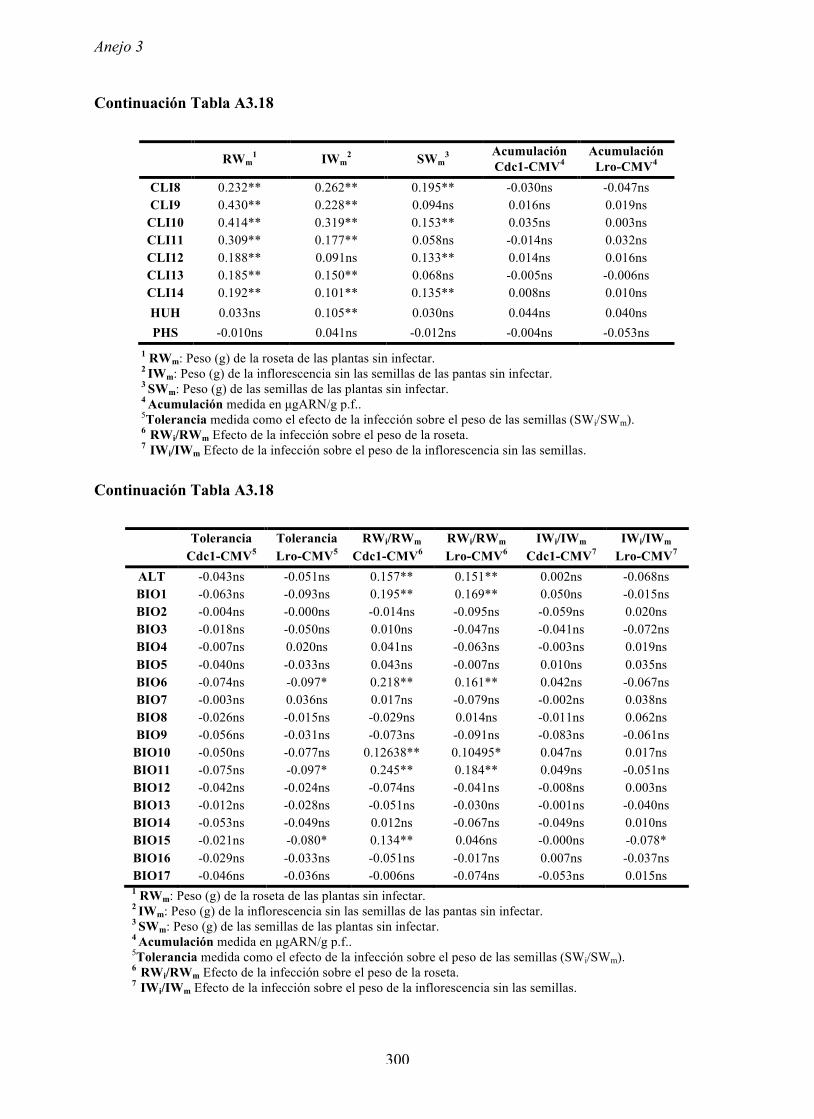
Anejo 3
300
Continuación Tabla A3.18
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar. 4 Acumulación medida en µgARN/g p.f.. 5Tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm). 6 RWi/RWm Efecto de la infección sobre el peso de la roseta. 7 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
Continuación Tabla A3.18
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar. 4 Acumulación medida en µgARN/g p.f.. 5Tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm). 6 RWi/RWm Efecto de la infección sobre el peso de la roseta. 7 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
RWm1 IWm
2 SWm3 Acumulación
Cdc1-CMV4 Acumulación
Lro-CMV4 CLI8 0.232** 0.262** 0.195** -0.030ns -0.047ns CLI9 0.430** 0.228** 0.094ns 0.016ns 0.019ns
CLI10 0.414** 0.319** 0.153** 0.035ns 0.003ns CLI11 0.309** 0.177** 0.058ns -0.014ns 0.032ns CLI12 0.188** 0.091ns 0.133** 0.014ns 0.016ns CLI13 0.185** 0.150** 0.068ns -0.005ns -0.006ns CLI14 0.192** 0.101** 0.135** 0.008ns 0.010ns HUH 0.033ns 0.105** 0.030ns 0.044ns 0.040ns PHS -0.010ns 0.041ns -0.012ns -0.004ns -0.053ns
Tolerancia
Cdc1-CMV5 Tolerancia Lro-CMV5
RWi/RWm Cdc1-CMV6
RWi/RWm Lro-CMV6
IWi/IWm Cdc1-CMV7
IWi/IWm Lro-CMV7
ALT -0.043ns -0.051ns 0.157** 0.151** 0.002ns -0.068ns BIO1 -0.063ns -0.093ns 0.195** 0.169** 0.050ns -0.015ns BIO2 -0.004ns -0.000ns -0.014ns -0.095ns -0.059ns 0.020ns BIO3 -0.018ns -0.050ns 0.010ns -0.047ns -0.041ns -0.072ns BIO4 -0.007ns 0.020ns 0.041ns -0.063ns -0.003ns 0.019ns BIO5 -0.040ns -0.033ns 0.043ns -0.007ns 0.010ns 0.035ns BIO6 -0.074ns -0.097* 0.218** 0.161** 0.042ns -0.067ns BIO7 -0.003ns 0.036ns 0.017ns -0.079ns -0.002ns 0.038ns BIO8 -0.026ns -0.015ns -0.029ns 0.014ns -0.011ns 0.062ns BIO9 -0.056ns -0.031ns -0.073ns -0.091ns -0.083ns -0.061ns
BIO10 -0.050ns -0.077ns 0.12638** 0.10495* 0.047ns 0.017ns BIO11 -0.075ns -0.097* 0.245** 0.184** 0.049ns -0.051ns BIO12 -0.042ns -0.024ns -0.074ns -0.041ns -0.008ns 0.003ns BIO13 -0.012ns -0.028ns -0.051ns -0.030ns -0.001ns -0.040ns BIO14 -0.053ns -0.049ns 0.012ns -0.067ns -0.049ns 0.010ns BIO15 -0.021ns -0.080* 0.134** 0.046ns -0.000ns -0.078* BIO16 -0.029ns -0.033ns -0.051ns -0.017ns 0.007ns -0.037ns BIO17 -0.046ns -0.036ns -0.006ns -0.074ns -0.053ns 0.015ns
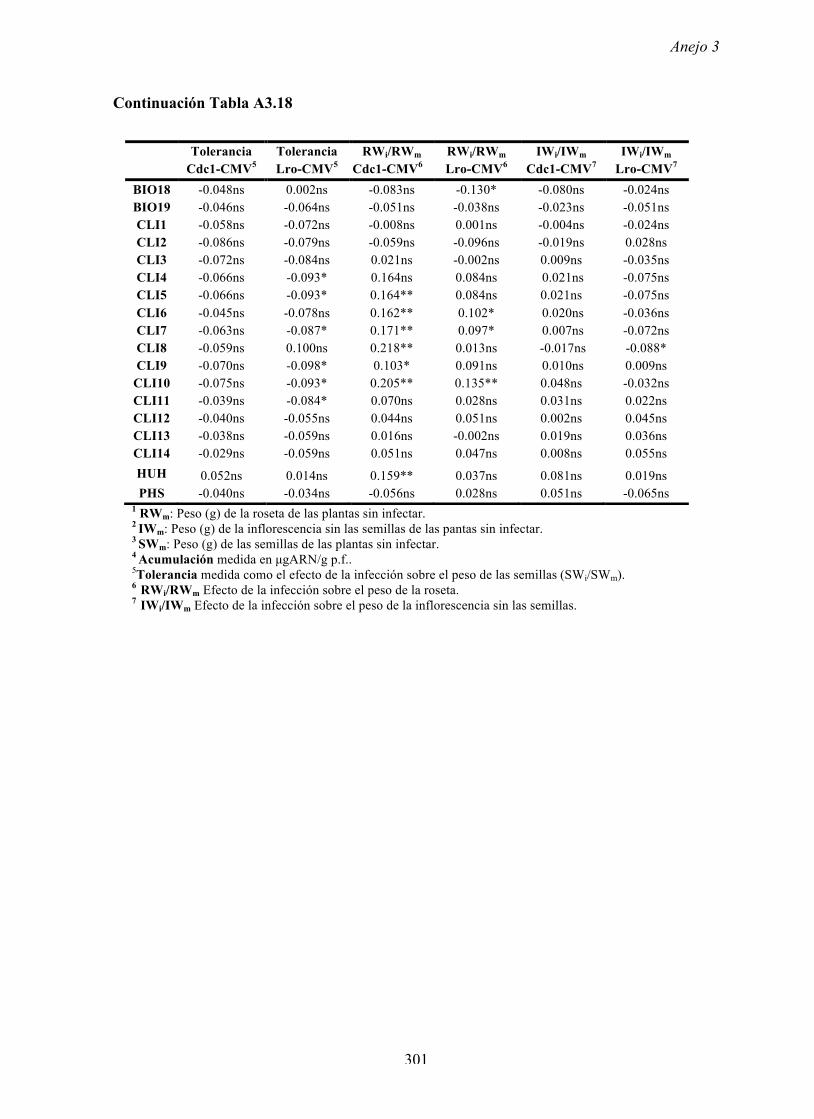
Anejo 3
301
Continuación Tabla A3.18
1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar. 4 Acumulación medida en µgARN/g p.f.. 5Tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm). 6 RWi/RWm Efecto de la infección sobre el peso de la roseta. 7 IWi/IWm Efecto de la infección sobre el peso de la inflorescencia sin las semillas.
Tolerancia
Cdc1-CMV5 Tolerancia Lro-CMV5
RWi/RWm Cdc1-CMV6
RWi/RWm Lro-CMV6
IWi/IWm Cdc1-CMV7
IWi/IWm Lro-CMV7
BIO18 -0.048ns 0.002ns -0.083ns -0.130* -0.080ns -0.024ns BIO19 -0.046ns -0.064ns -0.051ns -0.038ns -0.023ns -0.051ns CLI1 -0.058ns -0.072ns -0.008ns 0.001ns -0.004ns -0.024ns CLI2 -0.086ns -0.079ns -0.059ns -0.096ns -0.019ns 0.028ns CLI3 -0.072ns -0.084ns 0.021ns -0.002ns 0.009ns -0.035ns CLI4 -0.066ns -0.093* 0.164ns 0.084ns 0.021ns -0.075ns CLI5 -0.066ns -0.093* 0.164** 0.084ns 0.021ns -0.075ns CLI6 -0.045ns -0.078ns 0.162** 0.102* 0.020ns -0.036ns CLI7 -0.063ns -0.087* 0.171** 0.097* 0.007ns -0.072ns CLI8 -0.059ns 0.100ns 0.218** 0.013ns -0.017ns -0.088* CLI9 -0.070ns -0.098* 0.103* 0.091ns 0.010ns 0.009ns
CLI10 -0.075ns -0.093* 0.205** 0.135** 0.048ns -0.032ns CLI11 -0.039ns -0.084* 0.070ns 0.028ns 0.031ns 0.022ns CLI12 -0.040ns -0.055ns 0.044ns 0.051ns 0.002ns 0.045ns CLI13 -0.038ns -0.059ns 0.016ns -0.002ns 0.019ns 0.036ns CLI14 -0.029ns -0.059ns 0.051ns 0.047ns 0.008ns 0.055ns HUH 0.052ns 0.014ns 0.159** 0.037ns 0.081ns 0.019ns PHS -0.040ns -0.034ns -0.056ns 0.028ns 0.051ns -0.065ns
Anejo 3
302
Figura A3.1. Correlaciones entre los valores medios de RWm, IWm, SWm de cada individuo y
los valores de las variables ambientales temperatura anual media, máxima, mínima y la
precipitación anual acumulada de los lugares de procedencia de los individuos.
Anejo 3
303
Figura A3.2. Correlaciones entre los valores medios de acumulación de Cdc1-CMV,
tolerancia a Cdc1-CMV (SWi/SWm) y el efecto de Cdc1-CMV sobre la roseta (RWi/RWm) y la
inflorescencia (IWi/IWm) en de cada individuo y los valores de las variables ambientales
temperatura anual media, máxima, mínima y la precipitación anual acumulada de los lugares
de procedencia de los individuos.
Anejo 3
304
Figura A3.3. Correlaciones entre los valores medios de acumulación de Cdc1-CMV (�) y Lro-
CMV (�), tolerancia a Cdc1-CMV y a Lro-CMV (SWi/SWm) y el efecto de Cdc1-CMV y de
Lro-CMV sobre la roseta (RWi/RWm) y la inflorescencia (IWi/IWm) en de cada individuo y los
valores de la precipitación anual acumulada de los lugares de procedencia de los individuos.
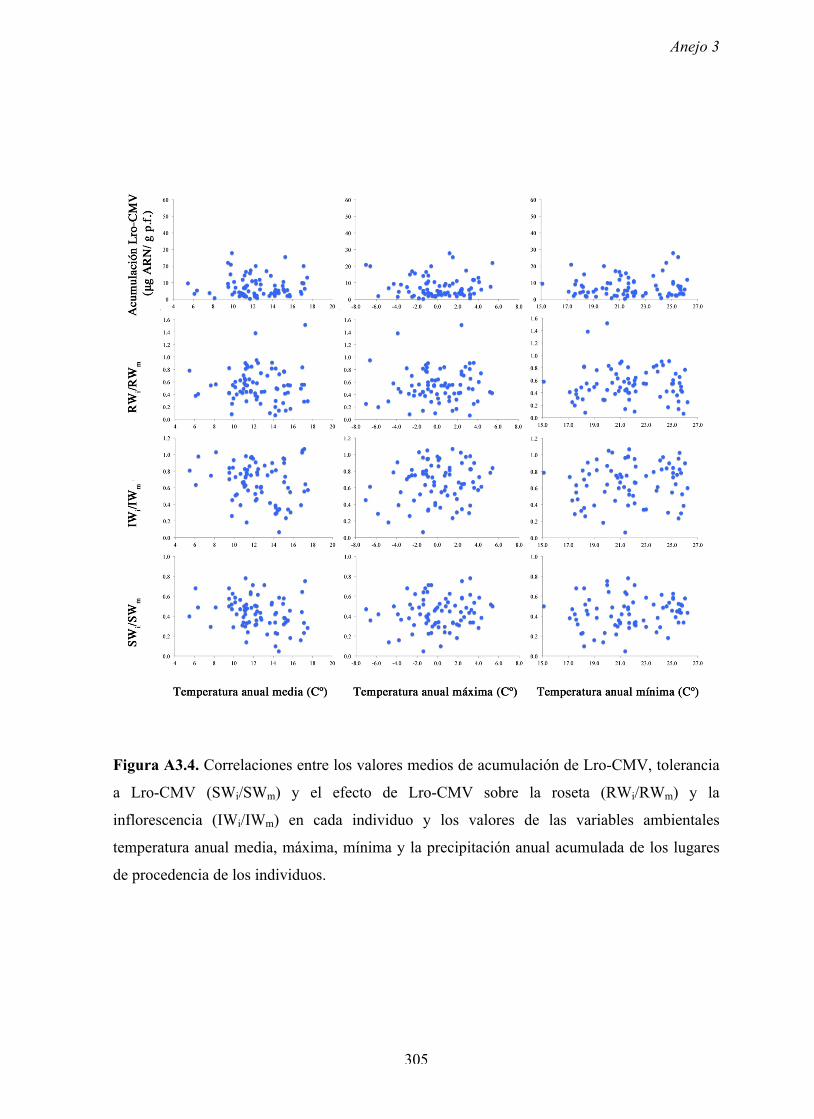
Anejo 3
305
Figura A3.4. Correlaciones entre los valores medios de acumulación de Lro-CMV, tolerancia
a Lro-CMV (SWi/SWm) y el efecto de Lro-CMV sobre la roseta (RWi/RWm) y la
inflorescencia (IWi/IWm) en cada individuo y los valores de las variables ambientales
temperatura anual media, máxima, mínima y la precipitación anual acumulada de los lugares
de procedencia de los individuos.
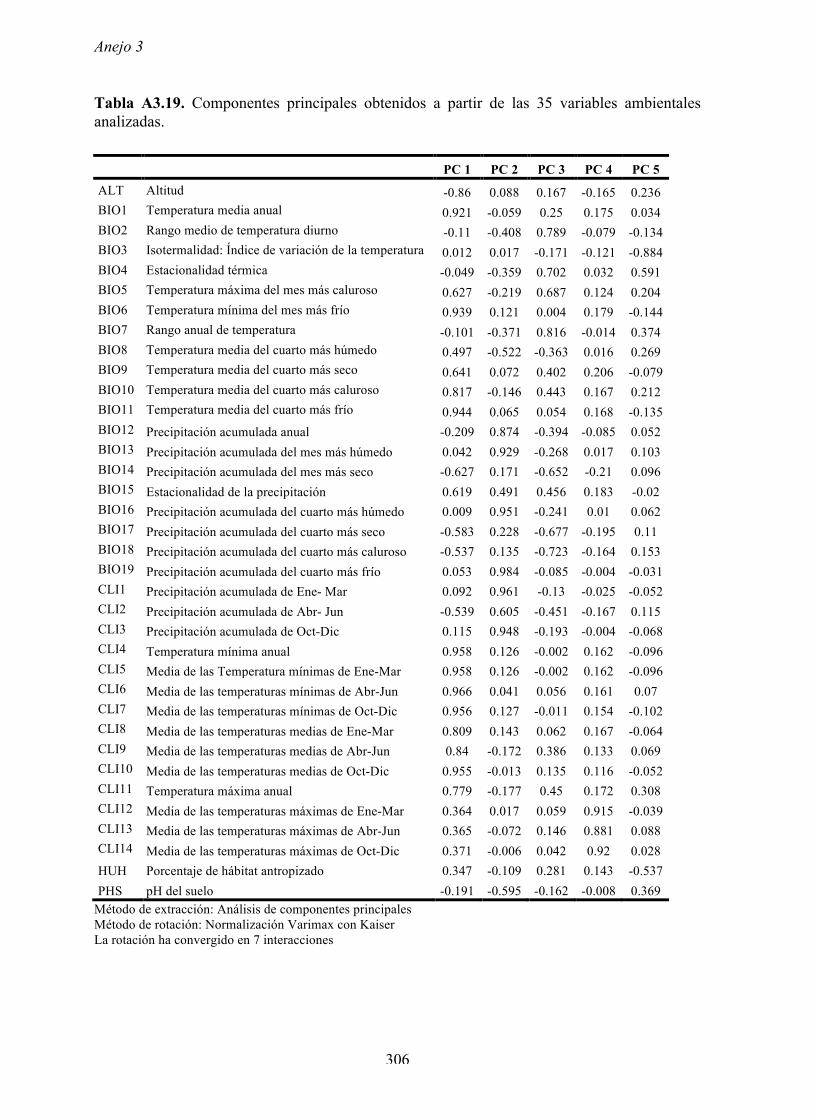
Anejo 3
306
Tabla A3.19. Componentes principales obtenidos a partir de las 35 variables ambientales analizadas.
PC 1 PC 2 PC 3 PC 4 PC 5
ALT Altitud -0.86 0.088 0.167 -0.165 0.236 BIO1 Temperatura media anual 0.921 -0.059 0.25 0.175 0.034 BIO2 Rango medio de temperatura diurno -0.11 -0.408 0.789 -0.079 -0.134 BIO3 Isotermalidad: Índice de variación de la temperatura 0.012 0.017 -0.171 -0.121 -0.884 BIO4 Estacionalidad térmica -0.049 -0.359 0.702 0.032 0.591 BIO5 Temperatura máxima del mes más caluroso 0.627 -0.219 0.687 0.124 0.204 BIO6 Temperatura mínima del mes más frío 0.939 0.121 0.004 0.179 -0.144 BIO7 Rango anual de temperatura -0.101 -0.371 0.816 -0.014 0.374 BIO8 Temperatura media del cuarto más húmedo 0.497 -0.522 -0.363 0.016 0.269 BIO9 Temperatura media del cuarto más seco 0.641 0.072 0.402 0.206 -0.079 BIO10 Temperatura media del cuarto más caluroso 0.817 -0.146 0.443 0.167 0.212 BIO11 Temperatura media del cuarto más frío 0.944 0.065 0.054 0.168 -0.135 BIO12 Precipitación acumulada anual -0.209 0.874 -0.394 -0.085 0.052 BIO13 Precipitación acumulada del mes más húmedo 0.042 0.929 -0.268 0.017 0.103 BIO14 Precipitación acumulada del mes más seco -0.627 0.171 -0.652 -0.21 0.096 BIO15 Estacionalidad de la precipitación 0.619 0.491 0.456 0.183 -0.02 BIO16 Precipitación acumulada del cuarto más húmedo 0.009 0.951 -0.241 0.01 0.062 BIO17 Precipitación acumulada del cuarto más seco -0.583 0.228 -0.677 -0.195 0.11 BIO18 Precipitación acumulada del cuarto más caluroso -0.537 0.135 -0.723 -0.164 0.153 BIO19 Precipitación acumulada del cuarto más frío 0.053 0.984 -0.085 -0.004 -0.031 CLI1 Precipitación acumulada de Ene- Mar 0.092 0.961 -0.13 -0.025 -0.052 CLI2 Precipitación acumulada de Abr- Jun -0.539 0.605 -0.451 -0.167 0.115 CLI3 Precipitación acumulada de Oct-Dic 0.115 0.948 -0.193 -0.004 -0.068 CLI4 Temperatura mínima anual 0.958 0.126 -0.002 0.162 -0.096 CLI5 Media de las Temperatura mínimas de Ene-Mar 0.958 0.126 -0.002 0.162 -0.096 CLI6 Media de las temperaturas mínimas de Abr-Jun 0.966 0.041 0.056 0.161 0.07 CLI7 Media de las temperaturas mínimas de Oct-Dic 0.956 0.127 -0.011 0.154 -0.102 CLI8 Media de las temperaturas medias de Ene-Mar 0.809 0.143 0.062 0.167 -0.064 CLI9 Media de las temperaturas medias de Abr-Jun 0.84 -0.172 0.386 0.133 0.069 CLI10 Media de las temperaturas medias de Oct-Dic 0.955 -0.013 0.135 0.116 -0.052 CLI11 Temperatura máxima anual 0.779 -0.177 0.45 0.172 0.308 CLI12 Media de las temperaturas máximas de Ene-Mar 0.364 0.017 0.059 0.915 -0.039 CLI13 Media de las temperaturas máximas de Abr-Jun 0.365 -0.072 0.146 0.881 0.088 CLI14 Media de las temperaturas máximas de Oct-Dic 0.371 -0.006 0.042 0.92 0.028 HUH Porcentaje de hábitat antropizado 0.347 -0.109 0.281 0.143 -0.537 PHS pH del suelo -0.191 -0.595 -0.162 -0.008 0.369
Método de extracción: Análisis de componentes principales Método de rotación: Normalización Varimax con Kaiser La rotación ha convergido en 7 interacciones

Anejo 3
307
Tabla A3.20. Valores (media±error estándar) de RWm, IWm y SWm según el grupo genético
de los individuos de arabidopsis sin infectar.
Grupo genético RWm1 IWm
2 SWm3
1 0.659±0.046 1.820±0.053 0.818±0.019
2 0.747±0.040 2.292±0.059 1.004±0.021
3 0.440±0.026 1.859±0.069 0.825±0.022
4 0.173±0.015 1.254±0.046 0.670±0.026
5 0.589±0.062 2.031±0.055 0.909±0.027 1 RWm: Peso (g) de la roseta de las plantas sin infectar. 2 IWm: Peso (g) de la inflorescencia sin las semillas de las pantas sin infectar. 3 SWm: Peso (g) de las semillas de las plantas sin infectar.
Tabla A3.21. Valores (media±error estándar) de la acumulación de Cdc1-CMV y Lro-CMV y
de la tolerancia a Cdc-CMV y Lro-CMV según el grupo genético de los individuos de
arabidopsis.
Grupo genético Acumulación
Cdc1-CMV1
Acumulación
Lro-CMV1
Tolerancia
Cdc-CMV2
Tolerancia
Lro-CMV2
1 19.159±1.119 8.821±0.649 0.363±0.018 0.438±0.015
2 21.135±1.658 8.833±0.773 0.409±0.021 0.574±0.014
3 15.696±1.232 5.743±0.717 0.256±0.029 0.521±0.026
4 21.441±1.555 6.950±0.661 0.360±0.024 0.500±0.018
5 17.237±1.265 8.256±0.907 0.402±0.027 0.516±0.026 1Acumulación medida en µgARN/g p.f.. 2Tolerancia medida como el efecto de la infección sobre el peso de las semillas (SWi/SWm).

Anejo 3
308

ANEJO 4

Anejo 3
310

Anejo 4
311
Tabla A4.1 Listado de individuos de arabidopsis analizados en el Apartado 4.3.
Población Individuo Longitud
(Oº) Latitud
(Nº) Altitud
(m) Hábitat
Aguarón Agu-8 -1.340 41.321 1045 Encinar Alcira Alr-0 -0.346 39.095 123 Bisaurri Bis-7 0.537 42.490 1450 Pradera de montaña Capileira Cap-1 -3.359 36.969 1460 Muro de piedra cercano a terrenos agrícolas abandonados Ciruelos de Coca Cdc-5 -4.544 41.215 800 Pinar La Chopera de las Rozas Cho-0 -3.897 40.515 725 Pradera xérica y matorral mediterráneo cerca de la cartera Corduentes Cor-0 -2.000 40.831 1080 Ladera rocosa y borde de la carretera en un pinar Santa María da Feira Fei-0 -8.541 40.925 140 Camino dentro del pueblo Grazalema Gra-9 -5.394 36.769 1284 Monte bajo mediterráneo Hortsavinyá Hor-0 2.623 41.666 431 Borde de bosque de especies de Quercus de hoja perenne San Leonardo de Yagüe Leo-8 -3.113 41.797 1010 Llanura aluvial Mérida Mer-3 -6.336 38.917 238 Área arenosa cercana a muro de piedra Molacillos Moc-8 -5.640 41.566 665 Linde de cultivos Orbaitzeta Orb-10 -1.228 42.974 735 Linde de un prado Oviedo Ovi-1 -5.866 43.381 386 Camino de pradera mésica Poblet Pob-1 1.027 41.355 677 Bosque mixto de pino y encima Prádena del Rincón Pra-3 -3.538 41.052 1106 Zona de rivera Quintela Qui-2 -6.929 42.693 700 Borde de camino Rascafría Ras-0 -3.868 40.864 1279 Quercus sp./Pine sp. Santa Elena San-10 -3.508 38.333 700 Encinar Valverde Vav-0 -8.107 38.534 220 Muro de piedra en una granja

Anejo 4
312
Tabla A4.2 Análisis GLM de la acumulación viral considerando aleatorios los factores
individuo de arabidopsis y aislado de CMV.
Origen F P Componente de la
varianza (%) Aislado de CMV F7,824=7.435 P <10-4 12.217
Individuo de arabidopsis F20,824=7.697 P <10-4 33.051 Aislado de CMV x
Individuo de arabidopsis F140,824 = 10.394 P <10-4 39.903
Tabla A4.3 Análisis GLM de la acumulación viral considerando fijo el factor huésped de
origen de los aislados de CMV y aleatorio el factor individuo de arabidopsis.
Origen F P Componente de la
varianza (%) Huésped de origen F1,824 = 8.351 P = 0.009 1.983
Individuo de arabidopsis F20,824=13.682 P <10-4 35.000 Huésped de origen x
Individuos de arabidopsis F20,824=1.828 P=0.017 2.496
Tabla A4.4. Análisis GLM de la tolerancia a CMV (SWi/SWm ) considerando aleatorios los
factores individuo de arabidopsis y aislado de CMV.
Origen F P Componente de la varianza (%)
Aislado de CMV F7,824=21.130 P <10-4 28.888 Individuo de arabidopsis F20,824=5.154 P <10-4 15.555
Aislado de CMV x Individuos de arabidopsis
F140,824=4.785 P <10-4 24.444
Tabla A4.5. Análisis GLM del efecto de la infección sobre el peso de la biomasa total (BMi/BMm) considerando aleatorios los factores individuo de arabidopsis y aislado de CMV.
Origen F P Componente de la
varianza (%) Aislado de CMV F7,824=0.816, P=0.575 19.318
Individuo de arabidopsis F20,824=13.749 P <10-4 14.772 Aislado de CMV x
Individuos de arabidopsis F140,824=1.282 P=0.025 15.909

Anejo 4
313
Tabla A4.6 Análisis GLM de la tolerancia a CMV (SWi/SWm) considerando fijo el factor
huésped de origen de los aislados de CMV y aleatorio el factor individuo de arabidopsis.
Origen F P Componente de la
varianza (%) Huésped de origen F20,824 = 50.020 P<10-4 12.903
Individuo de arabidopsis F20,824=7.116 P <10-4 16.129 Huésped de origen x
Individuos de arabidopsis F140,824=1.517 P=0.068 2.150
Tabla A4.7 Análisis GLM del efecto de la infección de CMV sobre la biomasa total (BMi/BMm) considerando fijo el factor huésped de origen de los aislados de CMV y aleatorio el factor individuo de arabidopsis.
Origen F P Componente de la
varianza (%) Huésped de origen F1,824=1.603 P=0.315 5.617
Individuo de arabidopsis F20,824=15.971 P <10-4 15.729 Huésped de origen x
Individuos de arabidopsis F140,824=1.052 P=0.397 0.002
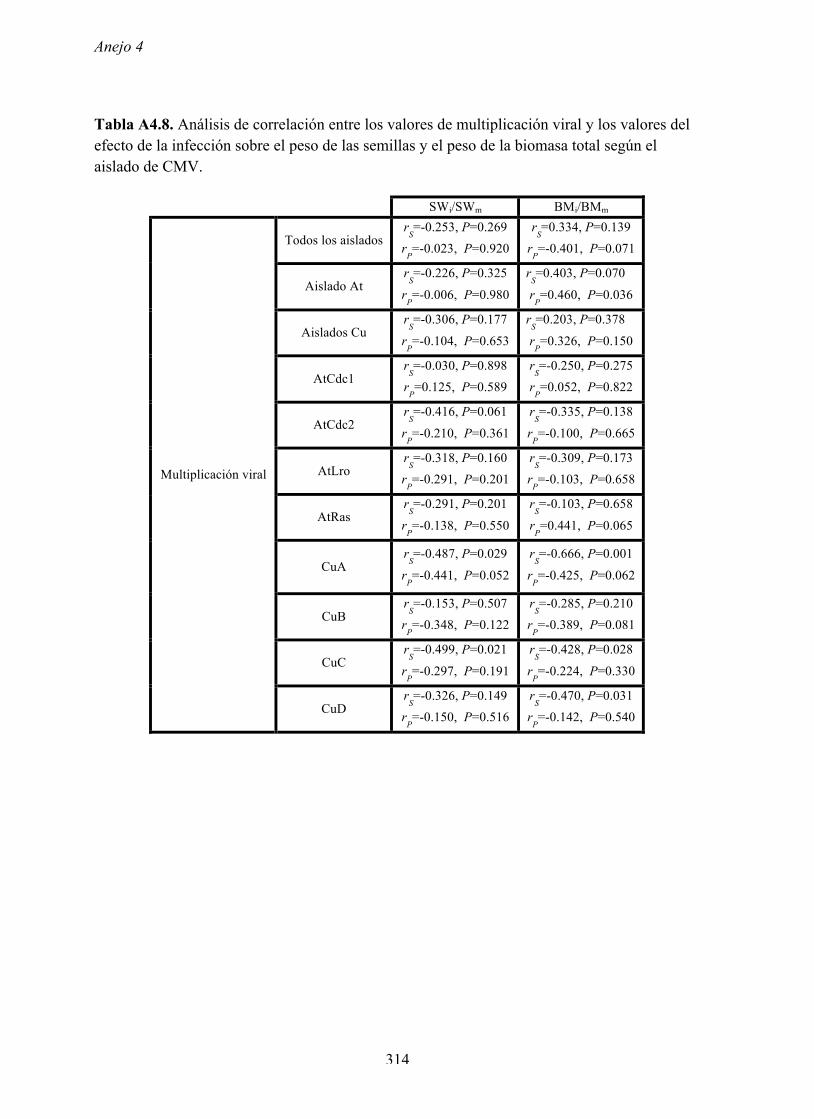
Anejo 4
314
Tabla A4.8. Análisis de correlación entre los valores de multiplicación viral y los valores del efecto de la infección sobre el peso de las semillas y el peso de la biomasa total según el aislado de CMV.
SWi/SWm BMi/BMm
Multiplicación viral
Todos los aislados r
S=-0.253, P=0.269
rP=-0.023, P=0.920
rS=0.334, P=0.139
rP=-0.401, P=0.071
Aislado At r
S=-0.226, P=0.325
rP=-0.006, P=0.980
rS=0.403, P=0.070
rP=0.460, P=0.036
Aislados Cu r
S=-0.306, P=0.177
rP=-0.104, P=0.653
rS=0.203, P=0.378
rP=0.326, P=0.150
AtCdc1 r
S=-0.030, P=0.898
rP=0.125, P=0.589
rS=-0.250, P=0.275
rP=0.052, P=0.822
AtCdc2 r
S=-0.416, P=0.061
rP=-0.210, P=0.361
rS=-0.335, P=0.138
rP=-0.100, P=0.665
AtLro r
S=-0.318, P=0.160
rP=-0.291, P=0.201
rS=-0.309, P=0.173
rP=-0.103, P=0.658
AtRas r
S=-0.291, P=0.201
rP=-0.138, P=0.550
rS=-0.103, P=0.658
rP=0.441, P=0.065
CuA r
S=-0.487, P=0.029
rP=-0.441, P=0.052
rS=-0.666, P=0.001
rP=-0.425, P=0.062
CuB r
S=-0.153, P=0.507
rP=-0.348, P=0.122
rS=-0.285, P=0.210
rP=-0.389, P=0.081
CuC r
S=-0.499, P=0.021
rP=-0.297, P=0.191
rS=-0.428, P=0.028
rP=-0.224, P=0.330
CuD r
S=-0.326, P=0.149
rP=-0.150, P=0.516
rS=-0.470, P=0.031
rP=-0.142, P=0.540
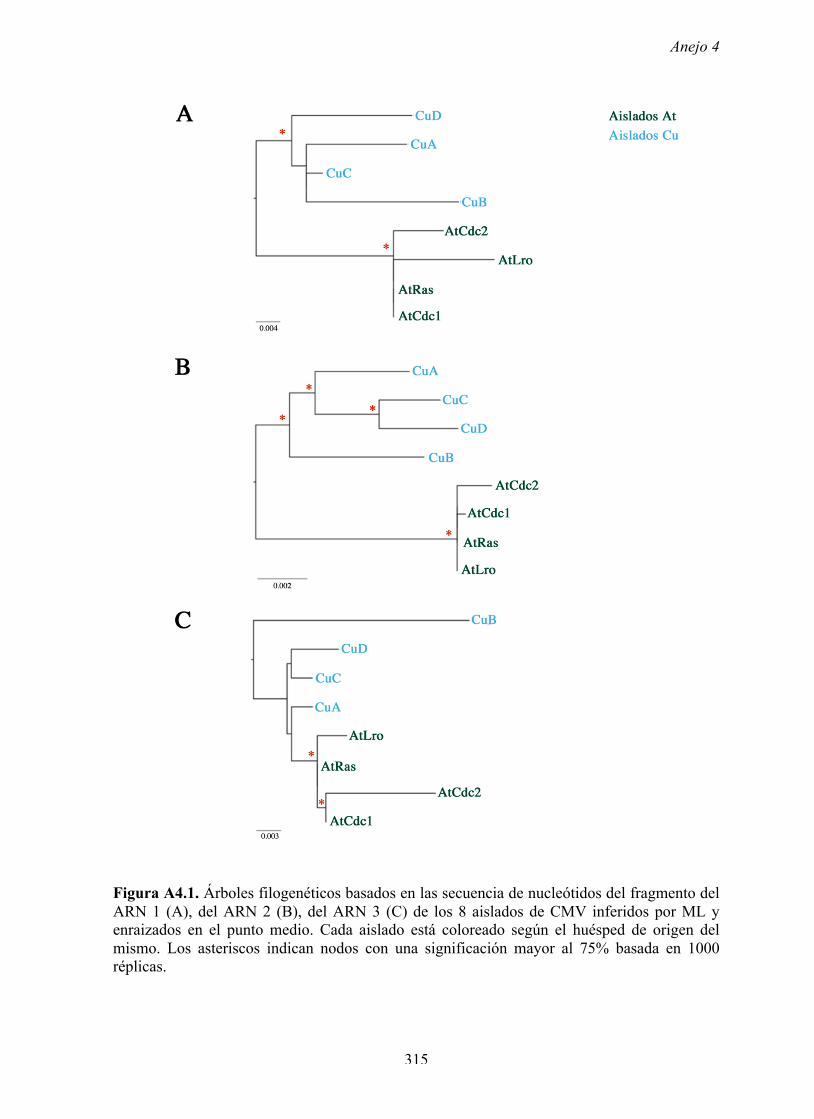
Anejo 4
315
Figura A4.1. Árboles filogenéticos basados en las secuencia de nucleótidos del fragmento del ARN 1 (A), del ARN 2 (B), del ARN 3 (C) de los 8 aislados de CMV inferidos por ML y enraizados en el punto medio. Cada aislado está coloreado según el huésped de origen del mismo. Los asteriscos indican nodos con una significación mayor al 75% basada en 1000 réplicas.

Anejo 4
316

Anejo 4
317



















