
“Actividad antimicrobiana de los compuestos …00:20Z-115… · de la División de Estudios para...
64
REPÚBLICA BOLIVARIANA DE VENEZUELA LA UNIVERSIDAD DEL ZULIA FACULTAD DE INGENIERÍA DIVISIÓN DE ESTUDIOS PARA GRADUADOS MAESTRÍA EN CIENCIA Y TECNOLOGÍA DE LOS ALIMENTOS “Actividad antimicrobiana de los compuestos producidos por Lactobacillus acidophilus ATCC 4356 sobre microorganismos contaminantes de alimentos” Trabajo presentado por la Ing. Mawill Rodríguez Marval ante La Universidad del Zulia para optar al título de Magister Scientarium en Ciencia y Tecnología de Alimentos Maracaibo; marzo de 2003
Transcript of “Actividad antimicrobiana de los compuestos …00:20Z-115… · de la División de Estudios para...

REPÚBLICA BOLIVARIANA DE VENEZUELA LA UNIVERSIDAD DEL ZULIA FACULTAD DE INGENIERÍA
DIVISIÓN DE ESTUDIOS PARA GRADUADOS MAESTRÍA EN CIENCIA Y TECNOLOGÍA DE LOS ALIMENTOS
“Actividad antimicrobiana de los compuestos producidos por Lactobacillus
acidophilus ATCC 4356 sobre microorganismos contaminantes de
alimentos”
Trabajo presentado por la Ing. Mawill Rodríguez Marval ante La Universidad del Zulia para optar al título de
Magister Scientarium en Ciencia y Tecnología de Alimentos
Maracaibo; marzo de 2003

REPÚBLICA BOLIVARIANA DE VENEZUELA LA UNIVERSIDAD DEL ZULIA FACULTAD DE INGENIERÍA
DIVISIÓN DE ESTUDIOS PARA GRADUADOS MAESTRÍA EN CIENCIA Y TECNOLOGÍA DE LOS ALIMENTOS
“Actividad antimicrobiana de los compuestos producidos por Lactobacillus
acidophilus ATCC 4356 sobre microorganismos contaminantes de
alimentos”
Realizado por Ing. Mawill Rodríguez Marval
C.I. 12620142
Tutora: Prof. Gisela Páez
Maracaibo; marzo de 2003

Este jurado aprueba el Trabajo de Grado titulado “Actividad
antimicrobiana de los compuestos producidos por Lactobacillus
acidophilus ATCC 4356 sobre microorganismos contaminantes de los
alimentos”, que presenta la Ing. Mawill Rodríguez Marval al Consejo Técnico
de la División de Estudios para graduados de La Universidad del Zulia, en
cumplimiento con los requisitos señalados en el artículo 51, aparte 51.6,
sección segunda, capítulo III del Reglamento de Estudios para Graduados,
para optar por el título de Magister Scientiarium en Ciencia y Tecnología de
Alimentos.
Maracaibo, marzo de 2003
Jurado:
__________________________________________
Prof. Gisela Páez
__________________________________________
Profa. Zulay Mármol
__________________________________________
Prof. José R. Ferrer
__________________________________________
Prof. Carlos Rincón
Director de Postgrado
I

AGRADECIMIENTO
Quiero expresar mi más profundo agradecimiento a todas las
personas e instituciones que de alguna manera colaboraron en la exitosa
culminación de este Trabajo de Grado.
A La Universidad del Zulia.
A FUNDADESARROLLO, por el financiamiento de mi Becaría.
A la Profesora Gisela Páez quien siempre ha tenido confianza en mi y
me animó cuando pensé que las cosas no salían bien. Gracias por todo,
especialmente por hacerme sentir como “estrella”.
A todas las personas del Laboratorio de Tecnología de Alimentos y
Fermentaciones Industriales. Todos los tesistas: Jasmín, Kena, Mireya.
Niohelis, David, Ronald, Ninoska, Karen y Eleana. Gracias a todos no sólo por
su colaboración con mi trabajo, sino también por construir momentos tan
agradables a pesar de las limitaciones y las circunstancias. A los profesores
del Laboratorio: José Ferrer, Cynthia Chandler, Zulay Mármol, Karelen Araujo
y a la Sra Bárbara.
A Eduardo y Marisela que siempre están allí no sólo con la respuesta,
sino también con el consejo.
Al Personal del laboratorio de Tecnología de Lácteos de la Facultad de
Veterinaria.
Al Sr. Edinson y Venelácteos por suministrar el suero de leche y
hacerlo con las ganas de ayudar de siempre.
A las Prof. Sofia de Valero y Luzmila Meza de la Facultad de Medicina
por su asesoría en la parte experimental.
A todos muchas gracias!!!
Mawill
DEDICATORIA
II

“Es justamente la posibilidad de realizar los sueños lo que hace la vida
interesante; … seas quien seas o hagas lo que hagas, cuando deseas con
firmeza alguna cosa, es porque este deseo nació en el alma del Universo. Es
tu misión en la tierra …. Y cuando quieres algo, todo el Universo conspira
para que realices tu deseo … La gente siempre está en condiciones de realizar
lo que sueña.”
Paulo Coelho en “El Alquimista”
Este trabajo está dedicado a todas esas personas que han creído en mi y que
me han ayudado a cumplir mis sueños; a todos aquellos con su amor y apoyo
han estado allí para mí, y se alegran conmigo cuando yo soy feliz y me
ayudan cada día a no desistir de mi “Leyenda Personal”.
A Dios quien ha sido bueno y generoso conmigo… siempre.
A Crispina, quien es la más feliz de todos por cada uno de mis éxitos.
A María y William, por su apoyo, su amor incondicional y su enorme
paciencia
A Melissa y Marcel, a quienes quiero ver cumplir todos sus sueños y a
quienes pretendo inspirar para seguir adelante y darse cuenta que vale la
pena el esfuerzo.
A Harvey, por su amor, paciencia, comprensión y apoyo en todos mis
proyectos.
A mis amigos de siempre… Gracias por tantos momentos de alegría.
RESUMEN
III

Rodríguez, Mawill. “Actividad antimicrobiana de los compuestos producidos por Lactobacillus acidophilus ATCC 4356 sobre microorganismos contaminantes de los alimentos” Trabajo de Grado. La Universidad del Zulia. Facultad de Ingeniería. Laboratorios de Tecnología de Alimentos y Fermentaciones Industriales. Maracaibo, 2003
Se estudió la actividad antimicrobiana de los compuestos producidos por Lactobacillus acidophilus ATCC 4356 sobre microorganismos contaminantes de los alimentos. Como medio de crecimiento para Lacobacillus acidophilus se emplearon caldo MRS y lactosuero suplementado con 0,3% p/v de extracto de levadura y 0,15% de peptona trípsica. Las fermentaciones se realizaron en erlenmeyers de 500 ml de capacidad a 37 ºC y pH 6,0 no controlado, 20% de inóculo y agitación de 50 rpm y sin agitación. Se evaluó el efecto de los dos medios de crecimiento, caldo MRS y lactosuero suplementado, y de la agitación sobre la constante específica de crecimiento () y la concentración de biomasa final (X
f). La detección de la actividad
antimicrobiana se llevó a cabo mediante la técnica de la difusión en el agar y la técnica de la difusión en el agar modificada, utilizando como microorganismos indicadores cepas de Candida, Kluyveromyces y Lactobacillus creciendo en caldo YM en el caso de las levaduras y caldo MRS en el caso de los lactobacilos. La máxima concentración de biomasa final se obtuvo en lactosuero suplementado sin agitación (2,9694 ± 0,2892 Kg/m3). La máxima velocidad de crecimiento se produjo en caldo MRS con 50 rpm de agitación (0,2131 ± 0,0192 h-1). No se detectó actividad antimicrobiana a ningún nivel sobre las cepas indicadoras utilizadas mediante las técnicas de análisis empleadas, posiblemente debido a la sensibilidad de la técnica.
Palabras clave: actividad antimicrobiana, Lactobacillus acidophilus,
microorganismos deterioradores
ABSTRACT
Rodríguez, Mawill. “Antimicrobial activity of compounds produced by
IV

University of Zulia. Faculty of Engineering. Food Technology and Industrial Fermentations Laboratories. Maracaibo 2003
Antimicrobian activity of Lactobacillus acidophilus ATCC 4356 against foodborne microorganism was studied. Lactobacillus acidophilus was grown in MRS broth and supplemented whey with 0,3% of yeast extract and 0,15% of peptone. Fermentation were carried out in 500 ml flasks at 37ºC and uncontrolled pH 6,O, 20% of inoculum, 50 rpm agitation and without agitation. The effect of the medium and agitation over specific growth rate () and biomass final concentration (X
f). Antimicrobial activity detection was
carried out using the agar diffusion assay and the agar diffusion assay modified, using strains of Candida, Kluyveromyces and Lactobacillus as indicator microorganisms. Indicator microorganisms were grown in MRS broth (lactobacillus) and YM broth (yeasts). Maximum biomas final concentration was obtained in supplemented whey without agitation (0, 9694 ± 0,2892 Kg/m3) and maximum specific growth rate was obtained in MRS broth at 50 rpm (0,2131± 0,092 h-1). Antimicrobial activity was not detected with the assays using in this study over any of the indicator microorganism, maybe because of the sensibility of the technique.
Key words: antimicrobial activity, Lactobacillus acidophilus, food
borne microorganism
ÍNDICE GENERAL
Pág.
V

Página de aprobación I
Agradecimiento II
Dedicatoria III
Resumen IV
Abstract V
índice de figuras VIII
índice de tablas IX
Introducción 1
Capítulo I. Revisión bibliográfica
1.1. Antecedentes históricos del uso de las bacterias ácido
lácticas (BAL) en la preservación de alimentos 5
1.2. Sustancias con actividad antimicrobiana de las
bacterias ácido lácticas y su uso como preservativos en
alimentos 6
1.2.1. Peróxido de hidrógeno 6
1.2.2. Diacetil 7
1.2.3. Reuterina 9
1.2.4. Dióxido de carbono 9
1.2.5. Productos finales de procesos de fermentación 10
1.2.6. Bacteriocinas 11
1.2.6.1. Definición 11
1.2.6.2. Clasificación 11
1.2.6.3. Diferencias entre las bacteriocinas y
los antibióticos 12
1.2.6.4. Mecanismo de acción 13
1.2.6.5. Usos 14
1.2.6.6. Consideraciones sobre la salud 16
1.3 Bacterias ácido lácticas y levaduras como
microorganismos contaminantes de los alimentos. 17
1.4. El Lactobacillus acidophilus 21
VI

1.4.1. Potencial terapéutico y beneficios a la salud
del Lactobacillus acidophilus 20
1.4.2. Bacteriocinas producidas por Lactobacillus
acidophilus 23
1.5. Suero de leche 24
1.5.1. Generalidades y composición 24
1.5.2. Usos del suero de leche 25
Capítulo II. Procedimiento Experimental
2.1. Materiales 30
2.1.1. Microorganismos 30
2.1.2. Medios de mantenimiento 30
2.2. Ultrafiltración y suplementación del lactosuero 30
2.3. Métodos de análisis 31
2.3.1. Nitrógeno 31
2.3.2. Proteína 31
2.3.3. Lactosa 31
2.3.4. Humedad 31
2.3.5. Biomasa 31
2.3.6. Actividad antimicrobiana 32
2.4. Fermentaciones 33
Capítulo III. Resultados y discusión 34
Capítulo IV. Conclusiones 46
Referencias bibliográficas 48
ÍNDICE DE FIGURAS
Figura Pág.
VII

Figura 1. Mecanismos para la generación de peróxido de hidrógeno 7
Figura 2. Curvas de crecimiento del Lactobacillus acidophilus ATCC
4356 a pH 6,0 y 37 ºC 36
Figura 3. Determinación de la velocidad específica de crecimiento del
Lactobacillus acidophilus ATCC 4356 a pH 6,0 y 37 ºC 37
ÍNDICE DE TABLAS
Tabla Pág.
VIII

IX
Tabla 1. Diferencias entre los antibióticos y las bacteriocinas 12
Tabla 2. Levaduras y bacterias ácido lácticas existentes en los alimentos 18
Tabla 3. Utilización del lactosuero 28
Tabla 4. Composición porcentual promedio del lactosuero fresco y desproteinizado 35
Tabla 5. Velocidad específica de crecimiento () del Lactobacillus acidophilus ATCC 4356 a 37 ºC y pH 6,0 39
Tabla 6. Concentraciones de biomas finales (Xf) obtenidas durante el
crecimiento de Lactobacillus acidophilus ATCC 4356 a 37 ºC y pH 6,0 39
Tabla 7. Detección de la actividad inhibitoria del Lactobacillus acidophilus ATCC 4356 mediante la técnica de la difusión en el agar 41
Tabla 8. Detección de la actividad inhibitoria del Lactobacillus acidophilus ATCC 4356 mediante la técnica de la difusión en el agar modificada 44

INTRODUCCIÓN
El Lactobacillus acidophilus, es una bacteria acidoláctica que produce
diversos metabolitos con actividad antimicrobiana, tales como peróxido de
hidrógeno y bacteriocinas, capaces de inhibir el crecimiento de cepas de los
géneros Kluyveromyces, Candida, Cladosporium (Plockova, Chumuchalova, y
Tomanova, 1997) y otros Lactobacillus (Kawai, Saito, Uemura, y Itoh, 1997;
Lewus, y Montville, 1991). Este microorganismo tiene la capacidad de crecer en
medios a base de suero de leche (Dave y Shah 1998), por lo cual este desecho
puede utilizarse como medio de cultivo para la producción de esos
metabolitos; la utilización del suero de leche como medio de crecimiento,
resulta en una disminución de su potencial contaminante, en el
aprovechamiento de sus componentes de alto valor biológico (como la lactosa
y las proteínas) y en la generación de productos tales como bebidas
fermentadas, ácidos orgánicos (Oberto y Carrasqueño, 1986), proteína
unicelular y sustancias con actividad antimicrobiana que pueden ser utilizadas
como preservativos contra el desarrollo de microorganismos contaminantes de
los alimentos.
En Venezuela y especialmente en la región zuliana, se cuenta con
cantidades importantes de suero de leche, un subproducto de la fabricación
de queso; la eliminación de este se ha convertido en uno de los problemas de
mayor importancia desde el punto de vista industrial y de la salud pública, ya
que el suero posee una Demanda Bioquímica de Oxígeno (DBO) de 4000 –
4800 ppm (Scott, 1991), convirtiéndolo en un poluente de elevado potencial
contaminante cuando se descarga en corrientes y reservorios de agua.
Para el año 1999, se produjeron en el Estado Zulia 91346542 L de
suero de leche (Ministerio de la Producción y el Comercio, 1999). Sin embargo,
sólo una pequeña fracción de este gran total es aprovechado principalmente
como alimento animal y suplemento alimenticio. El resto es dispuesto en los
1

suelos y cuerpos de agua, lo que representa un grave problema de
contaminación; por estas razones resulta importante y necesario diseñar
sistemas de disposición y utilización de este subproducto, que en nuestra
región es vertido principalmente en las aguas del Lago de Maracaibo,
aumentado los niveles de contaminación de este estuario.
Debido a la elevada incidencia de levaduras y mohos en los productos
lácteos y tomando en cuenta el peligro que representa la contaminación de
alimentos con micotoxinas, resulta importante la búsqueda de cepas de uso
seguro que produzcan compuestos inhibidores del crecimiento de esos
microorganismos. Una cepa reconocida como de uso seguro es el Lactobacillus
acidophilus ATCC 4356, la cual es utilizada ampliamente en la elaboración de
alimentos fermentados tales como el yogurt y actualmente es reconocido como
“probiótico”. (Lindgren y Dobrogosz, 1990; Mital y Satyendra, 1995)
La bioconversión del suero de leche para producir compuestos con
actividad antimicrobiana partir del Lactobacillus acidophilus ATCC 4356,
permite producir biopreservativos usando una materia prima muy económica,
al mismo tiempo que disminuye el potencial contaminante del suero. Las
sustancias con actividad antimicrobiana obtenidas a partir de este proceso
pueden utilizarse como preservativos alimenticios en sustitución de los
sulfitos, nitritos, parabenos y diacetatos, lo cual se corresponde con los
intereses de los consumidores que actualmente exigen alimentos procesados
más seguros. Por ejemplo, se conoce que algunas levaduras del género
Candida son capaces de resistir y adicionalmente atacar conservadores tales
como el ácido benzóico y ácido sórbico, los cuales son utilizados en la
industria de las bebidas alcohólicas (Mossell y Moreno, 1982).
Tradicionalmente los antibióticos como la nistatina y la anfotericina se utilizan
para combatir las infecciones por Candida en humanos (Leveau y Bouix, 2000).
Sin embargo estos compuestos no pueden ser utilizados como preservativos
alimentarios.
2

3
El objetivo de este trabajo es determinar si el Lactobacillus acidophilus
ATCC 4356 es capaz de producir, durante su crecimiento en suero de leche y
caldo MRS, compuestos con actividad inhibitoria del desarrollo de
microorganismos contaminantes de los alimentos de los géneros
Kluyveromyces, Candida y Lactobacillus.


REVISIÓN BIBLIOGRÁFICA
1.1. Antecedentes históricos del uso de las bacterias acidolácticas
(BAL) en la preservación de alimentos
Las bacterias ácido lácticas (BAL) han sido utilizadas en la producción
de alimentos fermentados durante varios siglos; esto ha sido posible ya que
estos microorganismos poseen un efectivo sistema enzimático que les
permite la degradación de los componentes de la leche y otros alimentos
(lactosa, proteínas, lípidos, citrato, etc.), produciendo metabolitos que alteran
el olor y la textura de los alimentos, así como varios metabolitos secundarios.
Ambos, los metabolitos primarios y secundarios, pueden poseer carácter
antimicrobiano (Lindgren y Dobrogosz, 1990).
Históricamente, la preservación de alimentos mediante la
fermentación ácido láctica ha sido un proceso empírico, en el cual se
observaron los cambios en los alimentos que generaban alimentos diferentes
con cualidades de conservación mejoradas. Existe una amplia variedad de
alimentos crudos que son preservados a través de la fermentación ácido
láctica, los cuales incluyen la leche, carnes, frutas, vegetales y piensos
(Lindgren y Dobrogosz).
A pesar de que los alimentos fermentados son conocidos desde la
antigüedad, la observación de que ciertos microorganismos son responsables
del efecto preservativo no se conoció de manera general sino hasta
comienzos del siglo XIX. Las primeras investigaciones confirmaron que la
preservación de los alimentos fermentados se debía al efecto acidificante que
ciertas bacterias le impartían a los alimentos por la conversión de los
azúcares en ácidos orgánicos. Las acciones de preservación que estos
microorganismos imparten a los alimentos son principalmente la disminución
del pH y la remoción de grandes cantidades de carbohidratos por
fermentación. Sin embargo, es también reconocido que las BAL son capaces
de producir sustancias inhibitorias además de ácidos orgánicos, que son
5

antagónicas hacia otros microorganismos (Lindgren y Dobrogosz; Mital y
Satyendra, 1995; Kawai et al., 1997; Plockova et al. 1997). Estas sustancias
son producidas en cantidades mucho menores e incluyen el peróxido de
hidrógeno, diacetilo, bacteriocinas y productos de reacciones secundarias
(por ejemplo, el hipotiocianato generado por la acción de la lactoperoxidasa
sobre el peróxido de hidrógeno y el tiocianato). Otros metabolitos que
pueden producirse, especialmente durante procesos heterofermentativos y
que pueden presentar actividad antagónica son el ácido fórmico, acetoín y
2,3-butanediol (Lindgren y Dobrogosz).
Se cree que las bacteriocinas son importantes en la habilidad de las
BAL de competir en ecosistemas no fermentativos tales como el intestino
humano (Lindgren y Dobrogosz). Durante las décadas pasadas se ha
incrementado el interés por el uso de la actividad antagónica de las BAL para
extender la vida útil de alimentos ricos en proteínas tales como pescados y
carnes.
1.2. Sustancias con actividad antimicrobiana de las bacterias ácido
lácticas y su uso como preservativos en alimentos
1.2.1. Peróxido de hidrógeno
El peróxido de hidrógeno es uno de los principales metabolitos
producidos por los lactobacilos durante su crecimiento, el cual puede ser
generado a través de diferentes mecanismos (Figura 1). Usualmente es
producido por la reducción directa del oxígeno, a través de una reacción
catalizada por la -glicerolfostato oxidasa o por la lactato oxidasa. (Mital y
Satyendra, 1995).
6

Piruvato + O2 + PO
4
3- Acetilfosfato + CO2 + H
2O
2piruvato oxidasa
Lactato + O2
L-lactato oxidasaPiruvato + H
2O
2
Lactato + O2 Piruvato + H
2O
2
NAD-independienteD-lactato deshidrogenasa
NADH oxidasaNAD + H
2O
2NADH + H+ + O
2
Figura 1: Mecanismos de generación de peróxido de hidrógeno
por bacterias ácido lácticas
La acumulación de peróxido de hidrógeno en el medio de crecimiento
se debe a que los lactobacilos no poseen la enzima catalasa (Daeschel,
1998). Esta acumulación puede alcanzar niveles capaces de inhibir el
crecimiento de otros microorganismos, ya que el H2O
2 es un compuesto
citotóxico debido a su capacidad para generar redicales hidroxil activos, los
cuales tienen un efecto fuertemente oxidante. (Mital y Satyendra). Otro efecto
producido por el peróxido de hidrógeno es la destrucción de las estructuras
básicas de las proteínas de las células. (Lindgren y Dobrogosz).
El peróxido de hidrógeno puede reaccionar con otros componentes
para formar sustancias inhibitorias. En la leche cruda, el peróxido de
hidrógeno generado por las bacterias ácido lácticas puede oxidar el
tiocianato endógeno, reacción que es catalizada por la lactoperoxidasa para
formar productos intermediarios de oxidación, los cuales tienen capacidad
inhibitoria hacia algunos microorganismos patógenos y deterioradores de los
alimentos. Este proceso se conoce como “sistema antibacterial de la
7

lactoperoxidasa” (LPS, por sus siglas en inglés), o “sistema lactoperoxidasa
tiocianato” (Mital y Satyendra), y ciertos estudios han indicado que este
sistema puede ser útil para extender la vida útil de la leche cruda no
refrigerada (Daeschel, 1998; Rodríguez, Tomillo, Núñez y Medina, 1997). El
efecto del LPS es muy variable; las bacterias Gram-positivas, incluyendo las
bacterias ácido lácticas, son mínimamente afectadas por el LPS, mientras que
bacterias Gram-negativas tales como Escherichia coli, Salmonella y
Pseudomonas son muy sensibles. El mecanismo antibacterial del LPS es
complejo y el mayor efecto ha sido atribuido a la oxidación de los grupos
SH- en enzimas vitales para el metabolismo, como la hexoquinasa, aldolasa y
la gliceraldehido-3 fosfato deshidrogenasa. (Lindgren y Dobrogosz).
La producción de peróxido de hidrógeno por las BAL depende no
solamente de la disponibilidad de oxígeno, sino también de la cepa en
particular (Lindaren y Dobrogosz). Su actividad antimicrobiana ha sido bien
reconocida y documentada. Su formación, acumulación y subsiguiente acción
antagonista ha sido demostrada contra especies tales como Staphylococcus
aureus, Pseudomonas sp. (Daeschel, 1998). Candida, Penicillium, y
Alternaria (Paraje, Albesa y Eraso, 2000).
1.2.2. Diacetil
El diacetil (2,3-butadieno) el un producto final del metabolismo de
las bacterias ácido lácticas, el cual es sintetizado a partir del piruvato. El
diacetil es bien conocido por su aroma a mantequilla, característico de los
productos lácteos (Lindgren y Dobrogosz); también es bien conocido por su
actividad antimicrobiana ya que tiene actividad inhibitoria a niveles de
200g/ml hacia levaduras y bacterias Gram-negativas, y a niveles de
300g/ml hacia bacterias no lácticas Gram-positivas (Daeschel)
A pesar de que el diacetil está ampliamente reconocido como seguro,
su utilidad como preservativo está limitada debido a las grandes cantidades
que se requieren para proveer efecto preservador, además de que su intenso
8

aroma no permite su uso en algunos alimentos. Su uso es recomendado para
la limpieza de superficies de trabajo debido a su alta volatilidad (Daeschel)
1.2.3. Reuterina
Se cree que la reuterina es un producto de bajo peso molecular, no
protéico, altamente soluble y de pH neutro producido por las especies
heterofermentativas Lactobacillus reuterii. Este compuesto ha sido descrito
como un agente antimicrobial de amplio espectro, el cual es activo contra
ciertas bacterias Gram-positivas y Gram-megativas, levaduras, hongos y
protozoarios. Algunos microorganismos de significación para la salud pública
que son inhibidos por la reuterina son Salmonella, Shiguella, Clostridium,
Staphilococcus, Listeria, Candida y Tripanosoma. Los lactobacilos
productores de reuterina pueden tener aplicación en la conservación de
alimentos humanos y animales, al reducir la carga de microorganismos
deterioradores de los mismos (Daeschel).
1.2.4. Dióxido de carbono
El dióxido de carbono es el resultado de la respiración endógena de
las células. La influencia del CO2 en la preservación de productos alimenticios
se debe a dos razones. Primero, juega un papel en la creación de un ambiente
anaeróbico remplazando el oxígeno molecular existente en el producto. En
segundo lugar, el CO2 en si mismo posee actividad antimicrobiana. El rol
protector del CO2 es especialmente importante en la fermentación de piensos
y vegetales para prevenir el crecimiento de mohos (Lindgren y Dobrogosz).
El CO2 en bajas concentraciones puede estimular el crecimiento de
algunos microorganismos, mientras que en altas concentraciones previene el
crecimiento de otros. La sensibilidad puede variar considerablemente, e
incluso existen microorganismos que no son sensibles al CO2 a ningún nivel
de concentración (Lindgren y Dobrogosz ).
9

Existen dos explicaciones acerca del mecanismo de acción del CO2;
uno implica la inhibición de las descarboxilaciones enzimáticas y el otro
apunta hacia la acumulación del CO2 en la membrana lipídica, lo cual resulta
en una disfunción en la permeabilidad de la misma (Lindgren y Dobrogosz).
1.2.5. Productos finales de procesos de fermentación
La acumulación de productos finales ácidos incrementa la actividad
antimicrobiana en los productos fermentados. La producción de ácido y la
correspondiente disminución del pH prolongan la fase de adaptación de los
microorganismos sensibles. Se han definido tres factores importantes para la
acción preservativa por sustancias ácidas: (1) el efecto del pH, (2) el alcance
de la disociación del ácido y (3) un efecto específico de la molécula de ácido
en si misma. La actividad antimicrobiana de los ácidos orgánicos de más de
cuatro carbonos generalmente se incrementa a un pH constante a medida
que aumenta la longitud de la cadena. Sin embargo, debido a su baja
solubilidad en agua, los ácidos de cadenas largas mayores de C10
o C11 no son
particularmente eficientes con respecto a la actividad antimicrobiana, y
aquellos cuyas cadenas son mayores de C8 son usualmente inefectivos contra
bacterias Gram-negativas. Los ácidos lipofílicos tales como el acético y el
láctico en sus formas no disociadas pueden penetrar la célula microbiana e
interferir con las funciones metabólicas esenciales tales como translocaciones
del sustrato y fosforilación oxidativa, y reducir el pH intracelular (Lindgren y
Dobrogosz).
El ácido acético ha sido reportado de producir mayor inhibición que el
ácido láctico, especialmente contra levaduras y mohos. Esto se debe al
alcance de la disociación, ya que el ácido acético posee entre dos y cuatro
veces más ácido en el estado no disociado comparado con el ácido láctico, en
un intervalo de pH de 4,0 a 4,6.
1.2. 6. Bacteriocinas
10

1.2.6.1. Definición
Las bacteriocinas son varias clases de pequeños péptidos que son
producidos naturalmente por varias clases de microorganismos, con efecto
antimicrobiano, efectivos generalmente contra especies cercanamente
relacionadas (Brink, Minekus, Van der Vossen, Leer y in’t Veld, 1994). Por
definición todas las bacteriocinas poseen una proteína o componente
protéico que es esencial para su acción antimicrobiana (Jack, Tagg y Ray,
1995), pero ciertos análisis químicos demuestran que algunas bacteriocinas
son moléculas complejas formadas por combinaciones de varias proteínas o
por la adición de lípidos y/o carbohidratos al componente protéico (Burianek
y Yousef, 2000).
1.2.6.2. Clasificación
Las bacteriocinas se agrupan en cuatro clases:
Clase I o lantabióticos
Son moléculas pequeñas (< 5 kDa), encapsuladas y que
presentan un amplio espectro de acción, dentro de este grupo la
nisina es la más conocida. Pueden ser producidas por bacterias no
ácido lácticas. Son generalmente producidas al final de la fase
logarítmica de crecimiento. Su nombre se deriva del hecho de que este
tipo de bacteriocinas contienen lantionina, -metil lantionina y
residuos deshidratados (Burianek y Yousef 2000).
Clase II
Incluye péptidos pequeños (< 10 kDa), estables al calor (80 –
120ºC), no contienen lantionina y contienen un péptido activo en la
membrana (Burianek y Yousef).
Clase III
11

Son moléculas largas (> 30 kDa), estables al calor ((Burianek y
Yousef).
Clase IV
Son moléculas grandes de proteínas o de compuestos de
proteínas con otros factores como carbohidratos o lípidos, que son
necesarios para la funcionalidad de la molécula
1.2.6.3. Diferencias entre las bacteriocinas y los antibióticos
Tabla 1: Diferencias entre las bacteriocinas y los antibióticos.
Bacteriocinas Antibióticos
Síntesis ribosómica
Existe un gen precursor que ha podido ser clonado y secuenciado
No son utilizadas para tratar o inhibir enfermedades infecciosas
Son utilizadas como preservativos en alimentos
No producen sensibilización, son atóxicas e inocuas
Son producidas por muchas bacterias
Síntesis enzimática.
Su estructura proviene de la síntesis enzimática por diferentes vías metabólicas
Se utilizan para inhibir o tratar enfermedades
Su uso como preservativo en alimentos no está permitido
Producen sensibilización, pueden ser tóxicos
Son producidos por pocos microorganismos
1.2.6.4. Mecanismo de acción de las bacteriocinas
12

La acción antimicrobiana de las bacteriocinas está asociada a la
despolimerización de los componentes de la membrana celular, lo cual
conduce a la salida de los componentes celulares y a la pérdida del potencial
a través de la membrana.
Sin embargo, las bacteriocinas presentan diferencias en cuanto a su
mecanismo de acción de acuerdo al microorganismo por el cual son
producidas. Así se tiene que las moléculas de bacteriocinas producidas por
los lactococos, son transportadas en el interior de la célula como péptidos
inactivos, los cuales contienen un extremo N-terminal en la secuencia líder
del péptido y C-terminal en el propéptido, siendo removido el péptido líder
por una peptidasa específica, dando origen al propéptido (Adams y Moss
1997). Se requiere de otras modificaciones antes de que los propéptidos sean
convertidos en moléculas activas, tal como la formación de la lantionina en el
caso de la nisina. Las moléculas activas de bacteriocinas son segregadas
desde el interior al medio a valores de pH 5,0 o superior. Las moléculas
tienen carga positiva y tienen la tendencia de formar agregados.
En general, la acción bacteriocida de las bacteriocinas que no
contienen lantionina contra las células sensibles es producida principalmente
por la desestabilización de las funciones de la membrana, más que por la
pérdida de la integridad de la estructura de la misma. Este efecto es el
resultado de la disipación de la fuerza impulsora del protón y pérdida de la
permeabilidad de la barrera independientemente de la energía, en contraste
con la acción dependiente de la energía de las bacteriocinas que contienen
lantionina (Burianek y Yousef).
Ambos tipos de bacteriocinas parecen afectar la permeabilidad de la
membrana por la formación de canales llenos de agua o poros,
probablemente por un mecanismo de ataque a la membrana. Adicionalmente,
previo a la formación de los poros, todas las bacteriocinas que no contienen
lantionina parecen interactuar con los receptores de proteínas asociados a la
13

membrana, otra vez en contraste directo con lo que ocurre con las
bacteriocinas que si contienen lantionina, las cuales no tienen ese
requerimiento (Jack et al., 1995).
Las cepas productoras de bacteriocinas poseen un gen de inmunidad
que parece producir proteínas de inmunidad específica que previenen la
formación de poros en la célula productora a través de un mecanismo que
hasta ahora no ha sido aclarado. Sin embargo este mecanismo debe estar
acompañado por un escudo de los receptores de proteínas, o por una
interacción competitiva con las moléculas de bacteriocinas o por un bloqueo
de los poros (Jack et al.; Entian y de Vos, 1996).
Las colonias sensibles difieren en el grado de sensibilidad hacia una
bacteriocina específica. Las bacterias Gram-negativas son resistentes a la
nisina y a la pediocina AcH, pero si se exponen a condiciones de estrés tales
como congelamiento, secado, tratamiento con EDTA o ácido, se pueden hacer
susceptibles a estas bacteriocinas. Las células subletalmente estresadas, con
una membrana externa lesionada, probablemente permiten la entrada de la
molécula de bacteriocina a su interior estableciendo contacto con la
membrana citoplasmática y desestabilizando sus funciones.
Las bacteriocinas producidas por los miembros del género
Lactobacillus generalmente tienen espectro de acción reducido o limitado y
usualmente actúan contra los miembros de la familia Lactobacillaceae. Su
acción es generalmente bactericida.
1.2.6.5. Usos de las bacteriocinas
Las investigaciones sobre bacteriocinas producidas por BAL han
aumentado en las últimas décadas, en la medida que han surgido varias
aplicaciones potenciales muy interesantes:
1. Las bacteriocinas de los microorganismos productores pueden
utilizarse como preservativos
14

2. Las determinantes genéticas para la producción de las bacteriocinas
y para la inmunidad del microorganismo productor, pueden utilizarse en la
producción de vectores para ser usados a nivel de alimentos, mejorando así
las cepas mediante el uso de la tecnología del ADN y
3. La producción de bacteriocinas puede utilizarse en estudios
taxonómicos.
En vista de la creciente demanda de los consumidores por
preservativos naturales, la mayor parte de la investigación está enfocada en la
primera posibilidad.
Existen varias estrategias posibles para la aplicación de las
bacteriocinas en la biopreservación de alimentos. Por una parte es posible la
inoculación de los alimentos con bacterias ácido lácticas (cultivos iniciadores)
que produzcan la bacteriocina en el producto (producción in situ). También
es posible agregar la bacteriocina purificada o semipurificada como
preservativo alimentario (Milo, 2000), o el uso de un producto previamente
fermentado con una cepa productora de bacteriocina como un ingrediente en
el procesamiento de alimentos (Schillinger, Geisen y Holzapfel, 1996).
Existen varios factores que afectan la producción de bacteriocinas por
las bacterias ácido lácticas (Schillinger et al., 1996), entre los cuales podemos
citar:
1. Inadecuadas condiciones ambientales (pH, temperatura, nutrientes,
etc.) para la producción de bacteriocinas
2. Pérdida espontánea de la capacidad de producción
3. Infección por fagos
4. Antagonismo con otros microorganismos presentes en el alimento
Adicionalmente existen factores que pueden afectar que interfieren
con la actividad de las bacteriocinas ya producidas:
15

1. Bacterias patógenas o deteriorativas emergentes resistentes a las
bacteriocinas
2. Condiciones que desestabilizan la actividad biológica de proteínas
como proteasas y procesos de oxidación.
3. Enlazamiento a componentes del alimento tales como partículas de
grasa y superficies protéicas
4. Inactivación por otros aditivos
5. Baja solubilidad de la bacteriocina que ocasiona una desigual
distribución en el alimento
6. Efecto del pH sobre la estabilidad y actividad de la bacteriocina
1.2.6.6. Consideraciones sobre la seguridad
El uso intencional o incidental de BAL en la producción de alimentos
fermentados se ha considerado como un proceso seguro ya que se carece de
evidencia que demuestre lo contrario. Por lo tanto es razonable asumir que
las sustancias antimicrobianas producidas por las BAL a los niveles
encontrados en los alimentos fermentados son seguras cuando se ingieren
como parte del alimento en el cual son producidas.
La nisina (cuyo nombre deriva de “group N inhibitory susbtance”), es la
única bacteriocina aprobada para su uso como preservativo en alimentos.
Esto se logró en U.S.A. en 1988, sólo después de 25 años de uso seguro en
varios países europeos. Se utiliza como agente inhibidor del crecimiento de
esporas de C. botulinum y la formación de toxinas en ciertos quesos
pasteurizados (Daeschel). El uso de la nisina en la preservación de alimentos
se considera seguro en más de 50 países alrededor del mundo e incluso hay
investigaciones que señalan su uso para controlar úlceras en humanos y
combatir infecciones como la mastitis en ganado (Delves-Broughton,
Blackburn, Evans, y Hugenholtz, 1996).
16

1.3. Bacterias ácido lácticas y levaduras como microorganismos
contaminantes de los alimentos.
Los microorganismos existentes en un alimento proceden tanto de la
microflora propia de la materia prima como de los microorganismos
introducidos durante las operaciones de recolección, sacrificio, tratamiento,
almacenamiento y distribución. En la mayoría de los casos, esta microflora no
ejerce un efecto aparente por lo que el alimento es consumido sin reparos y
sin consecuencias adversas. No obstante, algunas veces los microorganismos
manifiestan su presencia en una de las siguientes formas (Adams y Moss,
1997):
1. Causan alteración;
2. Causan una enfermedad transmitida por el alimento;
3. Transforman las propiedades de un alimento de una forma
beneficiosa, como es el caso de los alimentos fermentados.
En la actualidad se conoce que las condiciones del tratamiento y del
almacenamiento crean un medio en el que pueden prosperar determinados
tipos de organismos. La inmensa mayoría de los alimentos fermentados es
producida por la actividad de bacterias acidolácticas y de hongos,
principalmente levaduras pero también, aunque en menor medida, por
mohos. Ambos grupos de organismos comparten un nicho ecológico común,
siendo capaces de crecer bajo condiciones de pH bajo y actividad de agua
reducida, aunque en anaerobiosis sólo prosperarán las bacterias ácido
lácticas y las levaduras facultativas. En consecuencia, con frecuencia estos
organismos se hallan juntos en los alimentos fermentados; en algunos casos
representantes de ambos grupos actúan conjuntamente para producir un
determinado producto mientras que en otros, uno de los grupos desempeña
la función de organismo causante de alteración. En la Tabla 2 se presentan
algunos ejemplos de estos últimos (Adams y Moss).
17

Tabla 2: Levaduras y bacterias acidolácticas existentes en
diferentes alimentos
Levaduras1 Bacterias acidolácticas2 Levaduras y bacterias ácido lácticas
Cervezas europeas actuales
Pan
Vino
Sidra
Yogur
Chucruta
Salami
Queso
Pan de masa ácida
Kefir
Salsa de soja
Cervezas africanas
Cerveza Lambic 1La presencia de bacterias ácido lácticas con frecuencia va acompañada de alteración 2La presencia de levaduras con frecuencia va acompañada de alteración
En general, las levaduras se encuentran en diversos alimentos, como
un ingrediente o como contaminante. En este último caso, se produce una
fermentación no deseada de los azúcares del alimento, lo cual no sólo
deteriora el producto, dándole un sabor “alcohólico”, sino que también puede
ser peligroso, ya que la fermentación genera dióxido de carbono, capaz de
presurizar las botellas y otros envases hasta hacerlos explotar (Berger y Welt,
2001; The Society of Food Higiene Technology, 2003)
Las levaduras del género Kluyveromyces se encuentran en diversos
alimentos tales como frutas, conservas, maíz, leche y productos lácteos en
general (Banwart, 1981). Las especies capaces de fermentar la lactosa tales
como K. lactis y K. fragilis (Quintero et al., 2001; Montiel et al., 2000) se
encuentran en los productos lácteos y producen CO2, etanol y acetaldehído
(Müller, 1981). También se encuentran presentes, en gran proporción, en la
flora de los granos de kefir (Simova et al., 2002). Algunas especies son
osmofílicas y otras son capaces de sobrevivir en pulpa de frutas congeladas
(Trindade, Resende, Silva, y Rosa, 2002).
Las levaduras del género Candida se encuentran ampliamente
dispersas en la naturaleza. La C.albicans se encuentra en animales y humanos
y se presume que su presencia en otros ambientes es provocada por la
18

contaminación a partir de estos (Baker, 1997). Esta levadura ha sido aislada
de hojas de plantas, flores, agua y suelos. Se ha reportado como causa
común de infecciones en la superficie de la piel, boca y vagina. Las células de
este microorganismo generalmente no son transportadas por el aire y se
consideran componentes normales de las mucosas de la boca y otras
membranas del cuerpo. La mayoría de las infecciones por esta levadura
ocurren después de tratamientos prolongados con antibióticos o esteroides.
La C. krusei es uno de los microorganismos responsables de la
fermentación del cacao, y ha sido aislada de cerveza, productos lácteos y
salmueras (Micology OnLine, 2003). También ha sido encontrada en el aire,
hombre y otros mamíferos.
La C. parasilopsis se encuentra en humanos y otros mamíferos, en el
té, jugos de frutas y agua.
Las infecciones por Candida en humanos representan un grave
problema, especialmente para aquellas personas con deficiencias del sistema
inmunológico, tales como pacientes con HIV, diabetes mellitus, transplantes
de órganos, etc. La sobrepoblación en humanos de diferentes especies de
Candida de se conoce como candidiasis, que puede presentarse en diferentes
órganos del cuerpo humano. La infección por cándida es la más frecuente de
las infecciones micóticas del esófago, en particular las ocasionadas por C.
albicans. Otras especies como la C. tropicalis, C. glabrata, C. krusei o C.
parasilopsis pueden ser patógenas ocasionalmente, ya que pueden ser
contaminantes de los alimentos y también poseen la capacidad de adherirse a
las mucosas (Gastroenterología, 2003).
Otra infección producida por diferentes especies de Candida en la
candidemia, una complicación infecciosa grave en pacientes con neoplasias
hematológicas, con una mortalidad relacionada cercana al 40% (Jarque et al.,
2002)). Estas infecciones pueden ser causadas no sólo por Candida albicans,
que es bien conocida como patógena, sino también por las especies C. krusei
19

y C. parasilopsis (Frasquet et al., 2002). Las infecciones por C. albicans y C.
krusei también son comunes en pacientes con quemaduras severas (Khardori,
2002).
Se han reportado a las carnes ahumadas, frutas y quesos como
fuentes importantes de la contaminación por Candida en pacientes
inmunocomprometidos (Tomsikova, 2002).
En general, antimicóticos como el clotrimazol, itraconazol y la
nistatina, y antibióticos como el fluconazol, la anfotericina y la flucitosina se
utilizan para combatir las infecciones por Candida en humanos (Leveau y
Bouix, 2000; Gastroenterología, 2003). La introducción de la profilaxis con
fluconazol a principio de la década de 1990 se ha asociado con un
incremento en las infecciones por las especies de C. krusei que muestran
resistencia intrínseca al fluconzol (Jarque et al., 2002; Baker, 1997). Sin
embargo estos compuestos no pueden utilizarse como preservativos
alimentarios.
La natamicina es una bacteriocina producida por Streptomyces
natelensis, la cual muestra un efecto antimicótico hacia levaduras y mohos.
Se ha probado que previene el crecimiento de este tipo de microorganismos
en la superficie de quesos (Milo, 2000). También se ha reportado la inhibición
de diferentes especies de Candida por varias especies de Lactobacillus
(Paraje, Albesa y Eraso, 2000), que incluyen el Lactobacillus acidophilus (Mital
y Satyendra).
1.4. El Lactobacillus acidophilus
Los lactobacilos forman parte de la familia de las bacterias
ácidolácticas (BAL), la cual incluye además los géneros Leuconostoc,
Pediococcus y Lactococcus (grupo N estreptococos). Es bien conocido que las
BAL están involucradas en la preservación de ciertos alimentos y que son
responsables de algunas características sensoriales únicas de los mismos
(tales como textura y aroma), las cuales no es posible obtener por métodos
20

de preservación distintos de la fermentación por BAL. Adicionalmente las
pérdidas del valor nutricional se consideran mínimas durante los procesos de
fermentación mientras se extiende la vida útil de los alimentos al ser estos
menos propensos al ataque de microorganismos patógenos.
El grupo de Lactobacillus acidophilus ha sido recientemente dividido
en 6 subgrupos de acuerdo a la homología en el ADN y a los análisis de
composición de la pared celular (Kawai et al.). Estos subgrupos son:
Grupo A1: L. acidophilus
Grupo A2: L. crispatus
Grupo Ag3: L. amylovorus
Grupo A4: L. gallinarium
Grupo B1: L. gasseri
Grupo B2: L. johnsonii
1.4.1. Potencial terapéutico y beneficios a la salud del
Lactobacillus acidophilus.
Se ha propuesto que la longevidad de los búlgaros se debe en parte al
consumo de grandes cantidades de leches fermentadas que contienen
lactobacilos. Esta observación inició un rápido crecimiento en la búsqueda del
rol de los cultivos ácido lácticos y productos lácteos fermentados en el alivio
de desórdenes gastrointestinales en animales y humanos.
Los recientes avances en el conocimiento de las actividades
biosintéticas de las bacterias ácido lácticas y su acción antagónica contra
organismos patógenos, tóxicos y putrefactores, han añadido una nueva
dimensión a la importancia de las leches fermentadas en la nutrición y salud
humanas.
El surgimiento de bacterias resistentes a los antibióticos y de vías
naturales para la supresión del crecimiento de patógenos ha contribuido al
21

concepto de “probióticos”. Las bacterias probióticas no sólo compiten y
suprimen las “fermentaciones no saludables” en el intestino humano, sino
que también producen efectos beneficiosos en la salud por ellas mismas
(Kailaspathy y Chin, 2000).
Los microorganismos que han sido usados comúnmente como
probióticos en humanos son Lactobacillus acidophilus, L. casei,
Bifidobacterium bifidum, B. longum y la levadura Saccharomyces boulardii
(Jack et al.). Las bacterias probióticas se pueden aplicar en los desórdenes del
balance microbiano intestinal y otras disfunciones del tracto gastrointestinal.
El Lactobacillus acidophilus es ampliamente utilizado en productos
lácteos tales como yogur y leches ácidas, como probiótico, para proveer de
funciones fisiológicas eficientes en el intestino humano (Kawai et al.).
Adicionalmente, propiedades tales como actividad antitumoral y
anticolesterolémica, reacciones químicas asociadas con la reducción de
nitritos, mejoramiento del estado inmunológico y disminución de los
desórdenes gatrointestinales, han sido atribuídos al consumo de productos
lácteos fermentados (Lindgren y Dobrogosz). Los principales beneficios
terapéuticos y a la salud del Lactobacillus acidophilus son (Kailasapathy y
Chin, 2000):
1. Mejoramiento de la inmunidad contra infecciones intestinales
2. Mejoramiento inmune
3. Prevención de enfermedades diarreicas
4. Prevención de cáncer de colon
5. Prevención de hipercolesterolemia
6. Mejoramiento de la utilización de la lactosa
7. Prevención de enfermedades del tracto gastrointestinal
8. Estabilización de la barrera de la mucosa intestinal
22

La contribución de las bacterias presentes en el yogur en el
mejoramiento de la microflora intestinal ha sido ampliamente reconocida. La
incorporación de L. acidophilus y B. bifidum en los cultivos inciadores del
yogur resulta en un producto lácteo de excelente valor terapéutico. El
consumo regular de yogur (400 – 500 g/semana) provee de 1.0x106 UFC/g de
L. acidophilus y B. bifidum, que son capaces de sobrevivir en las regiones
más altas del tracto gastrointestinal humano, lo cual es esencial para lograr el
efecto terapéutico (Kailasapathy y Chin).
1.4.2. Bacteriocinas producidas por L. acidophilus.
La mayoría de las bacteriocinas producidas por las distintas especies
de lactobacilos, pertenecen a la clase II, las cuales son péptidos pequeños,
hidrofóbicos, estables al calor y sintetizados como precursores (Bogovic-
Matijasic, Rogelj, Nes y Holo, 1998).
La producción de bacteriocinas está ampliamente distribuída dentro
de los Lactobacillus acidophilus y sus especies relacionadas, tales como L.
gasseri y L. johnsonii (Bogovic-Matijasic et al., 1998).
La lactacina F fue la primera bacteriocina del Lactobacillus acidophilus
caracterizada genética y bioquímicamente; es en realidad una bacteriocina de
dos componentes activa contra otras especies de lactobacilos y Enterococcus
faecalis (Bogovic-Matijasic et al.).
Posteriormente un estudio sobre 52 cepas de Lactobacillus acidophilus
reveló que la mayoría de ellos producía un compuesto que inhibía el
crecimiento de otros lactobacilos y se identificó como lactacina B (Lindgren y
Dobrogosz).
Otra bacteriocina producida por L. acidophilus es la acidocina B, que
tiene un espectro de inhibición que incluye algunas bacterias no
acidolácticas, tales como Clostridium sporogenes, Listeria monocitogenes y
Brochotix thermosphacta. La acidocina A es otra bactericiocina producida por
23

Lactobacillus acidophilus con un interesante espectro de actividad, que
incluye algunos patógenos de los alimentos tales como Listeria
monocitogenes. (Bogovic-Matijasic et al.).
La bacteriocina CH5 también es producida por Lactobacilus
acidophilus y su espectro de inhibición incluye varias especies de levaduras y
hongos, lo cual resulta de gran interés, ya que estos microorganismos son
responsables de gran parte del deterioro de los alimentos (Plockova et al.,
1997).
Más recientemente se han descrito otras bacteriocinas, tales como la
acidocina J1132, acidocina LF221 A y acidocina LF221 B (Bogovic-Matijasic, B.
et al.) y la lacidina es producida por Lactobacillus acidophilus OSU1333 y
tiene actividad contra Staphylococcus aureus y Enterococcus faecalis
(Burianek y Yousef).
1.5. Suero de leche
1.5.1. Generalidades y composición
El suero es un subproducto líquido de la industria de elaboración del
queso; es el fluido que se obtiene cuando se separa el coágulo de la leche
entera, precipitando las grasas y las proteínas (González, 1996)
Los sueros procedentes de la elaboración de quesos blandos o de
quesos de veteado azul son ácidos (0.18 – 1.25% de acidez), pero los
procedentes de quesos muy prensados son más ácidos (0.22 – 0.4% de
acidez) (Scott, 1991).
En general el suero de leche contiene lactosa (3,3 – 6%), proteínas
(0,32 – 0,7%), grasas (0,15 – 1%) y trazas de sales. Si está seco, está
constituido principalmente por 70% de lactosa, 9 a 14% de proteínas y 9% de
cenizas. La proteína consiste de -lactobulina (50%), -lactobulina (25%) y
otras proteínas (25%), incluyendo inmunoglobulinas (25%) y es el componente
económicamente más valiosos del suero. A partir de estas proteínas es
24

posible preparar concentrados de proteínas con un amplio rango de
propiedades funcionales (González, 1996).
La composición de los sueros de quesería varía de acuerdo con el tipo
de queso elaborado y por tanto también su contenido de proteínas, sales
ácidos grasos, lactosa y ácido láctico. La concentración de lactosa en el suero
suele ser bastante constante pero depende de la proporción de lactosa
original que ha sido degradada a ácido láctico. El contenido de proteínas
depende en su mayor parte del tipo de coágulo y de su tratamiento, y puede
incrementarse debido a la presencia en el mismo de partículas de cuajada así
como una manipulación inadecuada de ésta. Aunque el contenido de sales es
usualmente constante, depende de la adición a la leche de algunos
compuestos como nitratos, etc., pero muy especialmente de cloruro o
hidróxidos cálcicos. El contenido de grasa, así como también el de proteínas
se ve modificado por procesos como el tamizado o la centrifugación del
suero (Scott).
1.5.2. Usos del suero
El uso que más frecuentemente se le da al suero de leche es la
alimentación de cerdos. Sin embargo, dado su bajo contenido de nitrógeno,
su administración en grandes cantidades puede ocasionar un desequilibrio
nutritivo; por lo tanto es importante complementar cuidadosamente la ración
de cereales, alimentos concentrados, etc., para cubrir todos los
requerimientos nutricionales de los animales. Adicionalmente el consumo
excesivo de suero puede provocar diarrea a los cerdos; esto se evita
añadiendo al suero amonio cuaternario (Veisseyre, 1972).
En algunos casos el queso es de importancia secundaria siendo de
mayor interés la obtención de un suero con poca acidez. Tal es el caso
cuando se trata de la suplementación de dietas o de la elaboración de
alimentos para niños, para lo que se requiere un suero de bajo contenido en
ácido láctico (0.12 – 0.13% de acidez). Este tipo de sueros resulta muy
25

adecuado cuando lo que se persigue es una mayor concentración de lactosa
no degradada como componente de los mismos. (Scott).
Como el suero es un líquido que fermenta rápidamente, debe ser
tratado adecuadamente para garantizar su conservación. La concentración al
vacío y la deshidratación son métodos excelentes desde el punto de vista
técnico, mediante los cuales es posible obtener jarabe o almíbar de suero,
con un contenido de agua siete veces menor que el original. También es
posible concentrar el suero hasta obtener una pasta con más de 60% en peso
de extracto seco y se puede obtener suero concentrado y azucarado, para
confitería, pastelería o para la fabricación de quesos fundidos. Mediante la
deshidratación profunda del suero por el método de los cilindros o por
atomización permite obtener suero en polvo que se utiliza en la preparación
de alimentos para el ganado (Veisseyre, 1972).
El suero de leche constituye un excelente medio de cultivo para una
gran variedad de microorganismos. El lactosuero suplementado ha sido
utilizado previamente en el crecimiento de cepas de lactobacilus (Jakymec et
al., 2001; Vielma y Urribarrí, 1999); Ramones, 2001) y en la producción de
sustancias tales como ácido láctico, al cual se le atribuye actividad
antimicrobiana, por la disminución del pH que ocasiona en el medio de
crecimiento (Daeschel).
Es posible también someter el suero a la fermentación butírica
provocada por Clostridium acetobutylicum. A través de este proceso de
obtienen butanol y acetona (González).
Otros microorganismos que pueden producirse usando como sustrato
el suero de leche, son las levaduras capaces de metabolizar la lactosa del
suero y que se utilizan como fuente de proteínas y vitaminas para animales.
Entre estas se encuentran algunas cepas de Torula y de Kluyveromyces. El
suero se utiliza como medio de mantenimiento y propagación de
microorganismos tales como Penicillium glaucum y Penicillium candidum
26

27
para la producción que quesos Camembert, Brie, Carré, etc. (Quintero et al.,
2001).
Recientemente se ha desarrollado la utilización de las proteínas del
suero de leche en la confección de películas que actúan como barreras
protectoras de los alimentos de baja o intermedia humedad contra factores
como oxígeno y aromas. Las películas de proteína de suero proveen una
mayor vida útil del alimento cuando se utilizan como recubrimiento de los
mismos. Los enlaces disulfuro y enlaces iónicos ayudan a la formación de
geles cristalinos a base de suero de leche. Bajo las apropiadas condiciones de
calentamiento, las proteínas del suero contribuyen a las características de
viscosidad y estabilidad al calor del gel. Las potenciales aplicaciones de esta
tecnología incluyen recubrimientos para separar ingredientes ricos en lípidos
de otros componentes en alimentos heterogéneos, y barreras para el oxígeno
y aromas en alimentos de baja humedad cuando se utiliza en combinación
con películas de recubrimiento simples. Las películas de proteína de suero
podría potencialmente eliminar el uso de barreras sintéticas para el oxígeno y
aromas en empaques con múltiples capas, además de reducir las pérdidas
por deterioro y extender la vida útil de los alimentos (Milo). En la Tabla 2 se
resumen los usos más importantes del lactosuero
Tabla 2. Utilización del suero (Scott).
Piensos para cerdos y aves
Suplementación del valor nutritivo del pan
Inclusión en alimentos para niños y alimentos dietéticos
Bebidas carbónicas y bebidas fermentadas
Precipitados de albúminas y globulinas como aditivos alimentarios

Preparados de albúminas utilizados como suplemento del valor nutritivo de algunos alimentos
Preparados cosméticos y farmacéuticos
Fabricación de alcohol y lactosa
Fabricación de jarabes de galactosa/glucosa
Queso de suero, Ziger, Urda, Ricotta, etc.
Aislamiento de riboflavina
Fabricación de ácido láctico para la industria en general, farmacéutica o alimentaria
Como medio de fermentación para la fabricación de antibióticos, combustibles (metano), biomasa para la producción de alimentos, o fabricación de jarabes de galactosa/glucosa para pastelería o fabricación de cerveza
28


PROCEDIMIENTO EXPERIMENTAL
2.1. Materiales 2.1.1. Microorganismos
Se utilizó como microorganismo productor de compuestos inhibitorios
el Lactobacillus acidophilus ATCC 4356.
Como microorganismos indicadores se utilizaron cepas de
Lactobacillus delbrueckii helveticus, Lactobacillus delbruekii bulgaricus,
Lactobacillus helveticus, Candida parasilopsis, Candida krusei, Candida
albicans y Kluyveromyces fragilis.
2.1.2. Medios de mantenimiento
Las levaduras se mantuvieron en cuñas con YMA (Yeast Medium Agar)
en nevera a 4ºC antes de su utilización y se replicaron cada cuatro días.
Los lactobacilos se mantuvieron en tubos con caldo MRS (De Man,
Rogosa y Sharpe), en nevera a 4ºC antes de su uso y se replicaron una vez
por semana.
2.2. Ultrafiltración y suplementación del lactosuero
Se utilizó suero de leche resultado de la producción de queso blanco
prensado, suministrado por la empresa Venelácteos, ubicada en Santa Rosa
de Agua, Maracaibo, Edo. Zulia. Este suero fue sometido a un proceso de
centrifugación, en una desnatadora para eliminar la mayor parte de la grasa
(Laboratorio de Lácteos, Fac. de Ciencias Veterinarias, LUZ). Luego fue
ultrafiltrado para separar las proteínas. La ultrafiltración se realizó en una
planta piloto de filtración tangencial (Laboratorio de Fermentaciones
Industriales y Tecnología de Alimentos, Fac. de Ingeniería, LUZ). Después de
pasar a través del equipo de ultrafiltración, el suero fue sometido a un
tratamiento termoácido (pH 4,5, 90 ºC, 10 minutos) para precipitar las
proteínas que aún permanecieran en solución. Luego de este tratamiento se
30

enfrió hasta 35 ºC y se paso nuevamente a través de las membranas de
ultrafiltración con el objetivo de remover las proteínas precipitadas con el
tratamiento termoácido. Posteriormente se procedió a la suplementación con
0,3% p/v de extracto de levadura y 0,15% p/v de peptona trípsica, y a su
esterilización en autoclave a 118 ºC durante 15 minutos.
2.3. Métodos de análisis
2.3.1. Nitrógeno
El contenido de nitrógeno del suero de leche se determinó por el
método Kjeldah (Association of Oficial Analitical Chemist, 1990).
2.3.2. Proteínas
El contenido de proteínas del suero de leche se determinó como el
porcentaje de nitrógeno multiplicado por el factor 6,38 (A.O.A.C., 1990).
2.3.3. Lactosa
El contenido de lactosa del suero de leche se determinó utilizando el
método espectrocolorimétrico de Dubois y colaboradores, previa
construcción de una curva de calibración (Dubois, Pilles, Hamilton, Rebers y
Smith, 1956).
2.3.4. Humedad
Se determinó el contenido de humedad del suero utilizando el método
de la A.O.A.C. (A.O.A.C.).
2.3.5. Biomasa
La concentración de biomasa en el medio se determinó
turbidimétricamente, midiendo la absorbancia de la muestra en un
espectrofotómetro Cary 50 (Australia) a una longitud de onda de 660 nm
(Parente y Hill, 1992; Shimizu et al., 1999), previa construcción de una curva
de calibración.
2.3.6. Actividad antimicrobiana
31

La detección de la actividad antimicrobiana del sobrenadante se llevó
a cabo utilizando la técnica de la difusión en el agar (Parente, Brienza, Moles
y Ricciardi, 1995).
Para la técnica de la difusión en el agar se prepararon 20 ml de agar
(YMB para el caso de las levaduras y MRS para los lactobacilos) que se
colocaron en una placa de Petri de 90 mm de diámetro y se dejaron secar
durante 30 minutos. Luego se prepararon las diluciones del sobrenadante,
comenzando con el sobrenadante puro, y utilizando el mismo medio utilizado
para su producción (caldo MRS o suero de leche) por duplicado. Se colocaron
10l de la dilución sobre el agar ya seco y se dejó difundir durante 30
minutos. Luego se cubrió el agar con 5 ml de agar semisólido (YMB para el
caso de las levaduras y MRS para los lactobacilos) previamente inoculado con
la cepa indicadora, con un inóculo estandarizado (D.O. 0,9 1,0 a 660 nm). Las
placas se incubaron en condiciones de acuerdo con el microorganismo
indicador utilizado: para los lactobacilos la incubación se realizó durante 16h
y a 37 ºC; para las levaduras la incubación se realizó a 30 ºC durante 2 días.
Luego de completar el periodo de incubación correspondiente se
revisaron las placas para verificar la existencia de zonas de inhibición.
Para la técnica de la difusión en el agar modificada, se preparó el agar
(YMB para las levaduras y MRS para los lactobacilos) y se distribuyó en tubos
de vidrio de 25x150 mm. Luego se agregó a los tubos el sobrenadante en la
dilución deseada y se colocó en placas de Petri de 90 mm y se dejó secar. Una
vez solidificado el agar se procedió a colocar sobre este una dilución de un
cultivo de los microorganismos indicadores correspondientes. Las placas se
incubaron y se compararon con los controles para determinar si existe
inhibición. El grado de inhibición se describió como: sin inhibición, inhibición
leve, inhibición moderada, inhibición completa.
2.4. Fermentaciones
32

33
Para las fermentaciones se utilizaron dos medios de crecimiento: caldo
MRS y en suero de leche.
Las fermentaciones se llevaron cabo en fiolas de 500 ml de capacidad
utilizando 300 ml de medio de crecimiento y un inóculo del 20%. Se permitió
el crecimiento del microorganismo productor durante 12 -14 horas, con un
pH inicial de 6,0, el cual no fue controlado durante la fermentación.
Transcurrido este tiempo, el sobrenadante se separó del medio de
crecimiento por centrifugación a 10000 g por 15 minutos. Luego se ajustó el
pH a 7,0. El sobrenadante así obtenido se esterilizó por filtración con filtros
Millipore de 0.2 de diámetro del poro y se llevó a congelación a -18 ºC,
cuando no se utilizó inmediatamente.

RESULTADOS Y DISCUSIÓN
Composición del suero de leche
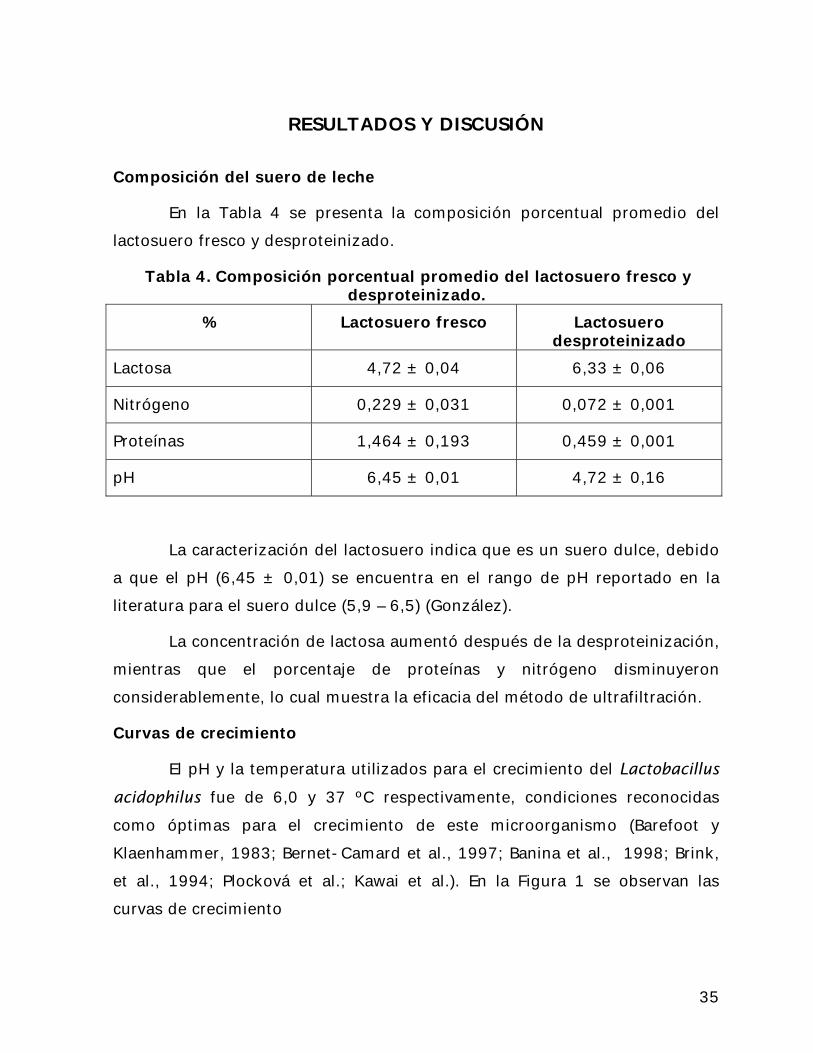
En la Tabla 4 se presenta la composición porcentual promedio del
lactosuero fresco y desproteinizado.
Tabla 4. Composición porcentual promedio del lactosuero fresco y desproteinizado.
% Lactosuero fresco Lactosuero desproteinizado
Lactosa 4,72 ± 0,04 6,33 ± 0,06
Nitrógeno 0,229 ± 0,031 0,072 ± 0,001
Proteínas 1,464 ± 0,193 0,459 ± 0,001
pH 6,45 ± 0,01 4,72 ± 0,16
La caracterización del lactosuero indica que es un suero dulce, debido
a que el pH (6,45 ± 0,01) se encuentra en el rango de pH reportado en la
literatura para el suero dulce (5,9 – 6,5) (González).
La concentración de lactosa aumentó después de la desproteinización,
mientras que el porcentaje de proteínas y nitrógeno disminuyeron
considerablemente, lo cual muestra la eficacia del método de ultrafiltración.
Curvas de crecimiento
El pH y la temperatura utilizados para el crecimiento del Lactobacillus
acidophilus fue de 6,0 y 37 ºC respectivamente, condiciones reconocidas
como óptimas para el crecimiento de este microorganismo (Barefoot y
Klaenhammer, 1983; Bernet-Camard et al., 1997; Banina et al., 1998; Brink,
et al., 1994; Plocková et al.; Kawai et al.). En la Figura 1 se observan las
curvas de crecimiento
35

36

del Lactobacillus acidophilus ATCC 4356, creciendo bajo estas condiciones en
caldo MRS y en lactosuero suplementado.
37

Se realizaron las curvas de crecimiento con y sin agitación, ya que este
es un factor primordial en la producción de H2O
2, debido a que favorece la
incorporación de O2 en el medio de crecimiento. Se utilizaron 50 rpm de
agitación para garantizar la suspensión homogénea de las células en el medio
de crecimiento y la incorporación de cierta cantidad de aire que permitiera la
producción de peróxido de hidrógeno pero no inhibiera el crecimiento del
microorganismo por exceso de oxígeno disuelto, ya que el Lactobacillus
acidophilus se conoce como microaerofílico y se ha reportado su crecimiento
en presencia de mezclas de gases tales como H2, CO
2 y N
2 para promover
condiciones anaerobias (Barefoot y Klaenhammer, 1983; Bernet-Camard et
al., 1997; Brink et al.).
En la Tabla 5 se presentan los valores de la velocidad específica de
crecimiento () de este microorganismo a las condiciones antes citadas, y el
efecto de la agitación y del medio de crecimiento sobre este parámetro. Se
realizó un análisis de la varianza y no se encontraron diferencias
significativas entre los valores encontrados. Se puede observar que el mayor
valor se obtiene cuando este microorganismo se desarrolla en caldo MRS a 50
rpm de agitación. Sin embargo en la Tabla 6, donde se presentan los valores
de concentración final de biomasa del Lactobacillus acidophilus ATCC 4356 a
37 ºC y pH 6,0, puede observarse que los valores de máximos de
concentración de biomasa final se obtuvieron en el lactosuero sin agitación, a
pesar de que el caldo MRS es el medio específico utilizado para el crecimiento
de lactobacilos. Esto indica que el lactosuero es un medio de crecimiento
adecuado para el desarrollo del Lactobacillus acidophilus ATCC 4356, lo cual
representa una alternativa a los altos costos de disposición de este desecho y
de los medios específicos para el crecimiento de este microorganismo. En la
Tabla 6 también se observa el efecto de la agitación sobre la velocidad de
crecimiento, la cual aumento cuando se utilizaron 50 rpm de agitación. Sin
embargo el efecto sobre la concentración de biomasa final fue inverso, ya que
esta disminuyó por efecto de la agitación.
38
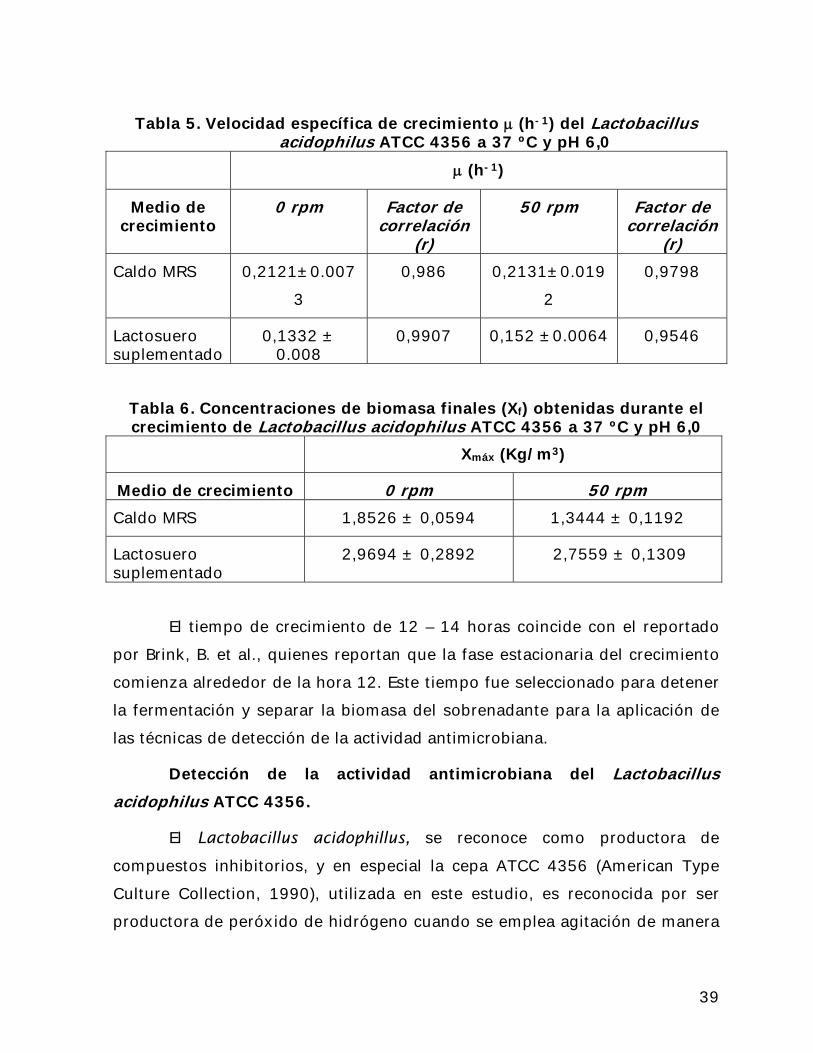
Tabla 5. Velocidad específica de crecimiento (h-1) del Lactobacillus acidophilus ATCC 4356 a 37 ºC y pH 6,0
(h-1)
Medio de crecimiento
0 rpm Factor de correlación
(r)
50 rpm Factor de correlación
(r)
Caldo MRS 0,2121±0.007
3
0,986 0,2131±0.019
2
0,9798
Lactosuero suplementado
0,1332 ± 0.008
0,9907 0,152 ±0.0064 0,9546
Tabla 6. Concentraciones de biomasa finales (Xf) obtenidas durante el crecimiento de Lactobacillus acidophilus ATCC 4356 a 37 ºC y pH 6,0
Xmáx (Kg/m3)
Medio de crecimiento 0 rpm 50 rpm
Caldo MRS 1,8526 ± 0,0594 1,3444 ± 0,1192
Lactosuero suplementado
2,9694 ± 0,2892 2,7559 ± 0,1309
El tiempo de crecimiento de 12 – 14 horas coincide con el reportado
por Brink, B. et al., quienes reportan que la fase estacionaria del crecimiento
comienza alrededor de la hora 12. Este tiempo fue seleccionado para detener
la fermentación y separar la biomasa del sobrenadante para la aplicación de
las técnicas de detección de la actividad antimicrobiana.
Detección de la actividad antimicrobiana del Lactobacillus
acidophilus ATCC 4356.
El Lactobacillus acidophillus, se reconoce como productora de
compuestos inhibitorios, y en especial la cepa ATCC 4356 (American Type
Culture Collection, 1990), utilizada en este estudio, es reconocida por ser
productora de peróxido de hidrógeno cuando se emplea agitación de manera
39

que se incorpore oxígeno al medio de crecimiento, necesario para la
producción de este compuesto (Mital y Satyendra).
La selección de las cepas de Candida, Kluyveromyces y Lactobacillus
como microorganismos indicadores se basó en diversos factores,
principalmente: incidencia en los alimentos como microorganismos
deterioradores, utilización previa como microorganismos indicadores,
disponibilidad y seguridad para su manipulación (Plocová et al.; Nagendra, y
Rajiv, 2001).
En la Tabla 5 se presentan los resultados obtenidos al utilizar el
método de la difusión en el agar. Se observa que en ninguno de los casos el
sobrenadante inhibió, en ningún grado, el crecimiento normal de la cepa
indicadora utilizada.
Plocková et al., reportan la inhibición de cepas de Kluyveromyces,
Candida, Lactobacillus, Cladosporium, Alternaria y Penicillium, todas cepas
con aplicaciones en la industria de los productos lácteos. En ese estudio se
reporta que la inhibición era claramente visible por el método de la difusión
en el agar, al apreciarse sólo un débil crecimiento en el caso de las levaduras
y cambios en la tasa de crecimiento y en la apariencia típica del desarrollo de
los micelios de los hongos. Estos resultados son atribuidos a sustancias
antimicrobianas diferentes de los ácidos orgánicos y el peróxido de
hidrógeno, ya que estos autores trataron térmicamente el sobrenadante
obtenido para evitar la acción del peróxido de hidrógeno y cualquier otra
sustancia termoestable. Además garantizaron la completa remoción de las
células viables del sobrenadante para evitar la producción de ácido láctico
durante el periodo de incubación requerido por la técnica de análisis, que
pudiera ser causante de inhibición de la cepa indicadora.
En el caso de este estudio todas las células fueron removidas del
sobrenadante por filtración. Sin embargo el sobrenadante no fue tratado
térmicamente, de manera que cualquier efecto inhibitorio sobre las cepas
40
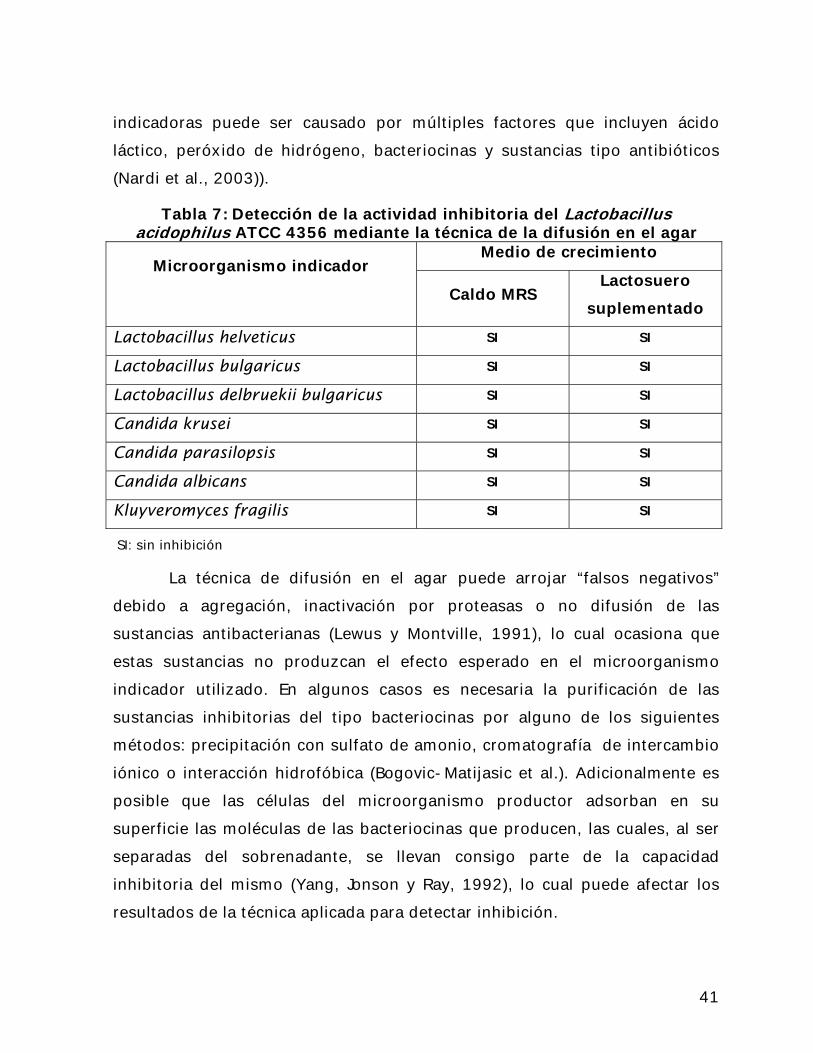
indicadoras puede ser causado por múltiples factores que incluyen ácido
láctico, peróxido de hidrógeno, bacteriocinas y sustancias tipo antibióticos
(Nardi et al., 2003)).
Tabla 7: Detección de la actividad inhibitoria del Lactobacillus acidophilus ATCC 4356 mediante la técnica de la difusión en el agar
Medio de crecimiento Microorganismo indicador
Caldo MRS Lactosuero
suplementado
Lactobacillus helveticus SI SI
Lactobacillus bulgaricus SI SI
Lactobacillus delbruekii bulgaricus SI SI
Candida krusei SI SI
Candida parasilopsis SI SI
Candida albicans SI SI
Kluyveromyces fragilis SI SI
SI: sin inhibición
La técnica de difusión en el agar puede arrojar “falsos negativos”
debido a agregación, inactivación por proteasas o no difusión de las
sustancias antibacterianas (Lewus y Montville, 1991), lo cual ocasiona que
estas sustancias no produzcan el efecto esperado en el microorganismo
indicador utilizado. En algunos casos es necesaria la purificación de las
sustancias inhibitorias del tipo bacteriocinas por alguno de los siguientes
métodos: precipitación con sulfato de amonio, cromatografía de intercambio
iónico o interacción hidrofóbica (Bogovic-Matijasic et al.). Adicionalmente es
posible que las células del microorganismo productor adsorban en su
superficie las moléculas de las bacteriocinas que producen, las cuales, al ser
separadas del sobrenadante, se llevan consigo parte de la capacidad
inhibitoria del mismo (Yang, Jonson y Ray, 1992), lo cual puede afectar los
resultados de la técnica aplicada para detectar inhibición.
41

La técnica de la difusión en el agar y sus diferentes modificaciones
han sido ampliamente utilizadas en la detección de actividad antimicrobiana
(Plocková et al.; Bogovic-Matijasic et al.; Brink et al., 1994; Yang, et al., 1992;
Burianek y Yousef; Banina et al., 1998; Parente y Hill, 1922; Rodríguez et al.,
1997). Sin embargo, en un estudio realizado por Lewus y Montville (1991) se
compararon diversos métodos para la detección de bacteriocinas producidas
por BAL. Los resultados de este estudio muestran que el método de la
difusión en el agar produce una mayor cantidad de falsos negativos
comparados con otros métodos de detección de antimicrobianos tales como
la técnica del los puntos sobre el agar y de la capa de agar invertida.
Adicionalmente Parente y Hill reportan que los ensayos fotométricos proveen
resultados más reproducibles y precisos que aquellos obtenidos con los
métodos de difusión en el agar.
Lewus y Montville sugieren que la sensibilidad de la técnica podría
incrementarse aumentando el tiempo de difusión de la solución del
componente inhibitorio en el agar antes de la incubación; también sugiere
aumentar la cantidad de muestra del componente antimicrobiano aplicado en
cada ensayo.
En base a estas observaciones se realizaron las modificaciones a la
técnica de la difusión en al agar utilizada previamente. Estas modificaciones
consistieron en aumentar hasta 20% la cantidad de sobrenadante utilizado en
cada ensayo y adicionalmente se combinó el sobrenadante con el agar
licuado a 37 ºC, antes de que este último solidificara. Por otra parte es
extendió de 30 minutos hasta 2 horas el lapso de tiempo entre la aplicación
del sobrenadante al agar y la inoculación de las placas con el microorganismo
indicador correspondiente. Con estas modificaciones se esperó mejorar la
difusión del sobrenadante en el agar, ya que al encontrarse este en forma
líquida, la distribución podía hacerse de manera más uniforme.
42

Debido a que parte del efecto inhibitorio puede ser ocasionado por la
disminución del pH producida por la presencia de ácido láctico en el medio,
se realizaron una serie de experimentos omitiendo la neutralización del pH
final del sobrenadante (el cual osciló entre 5,70 y 4,32), previa a la aplicación
de la técnica de detección. Adicionalmente y debido a que la congelación
podría también afectar la actividad de los compuestos antimicrobianos,
también se realizaron pruebas utilizando el sobrenadante inmediatamente
después de finalizada la fermentación (SC) y después de 2 días de
congelación a -18 ºC (C).
La Tabla 6 presenta los resultados obtenidos al aplicar la técnica de
difusión en el agar modificada. Puede apreciarse que las modificaciones
realizadas no arrojaron resultados diferentes de los obtenidos en las pruebas
realizadas previamente. Adicionalmente la neutralización del pH parece no
tener efecto sobre la actividad del sobrenadante. Sin embargo es posible que
esto afecte la concentración de ácido láctico en el medio, el cual es uno de
los principales agentes inhibitorios del crecimiento de otros microorganismos
(Mataragas, Drosinos y Metaxopoulos, 2003), especialmente los lactobacilos,
ya que en el caso de las levaduras se conoce que estas pueden desarrollarse
en un amplio rango de pH (Adams y Moss)
El hecho de que las cepas utilizadas en este estudio no presenten
sensibilidad a los compuestos antimicrobianos producidos por L. acidophilus
ATCC 4356, no significa que otras cepas contaminantes de alimentos no sean
sensibles y puedan inhibirse. Plockova et al. (1997) reportan que la cepa de
Candida famata, no fue afectada por la bacteriocina CH5; en cambio las
cepas de Kluyveromyces marxianus y Candida kefir fueron inhibidas
levemente. En el caso de los mohos, algunas cepas de Penicillium no fueron
inhibidas por la bacteriocina y las cepas de Cladosporium y Alternaria
resultaron ser las más sensibles, al mostrar una fuerte supresión de su
crecimiento en presencia de la bacteriocina CH5.
43
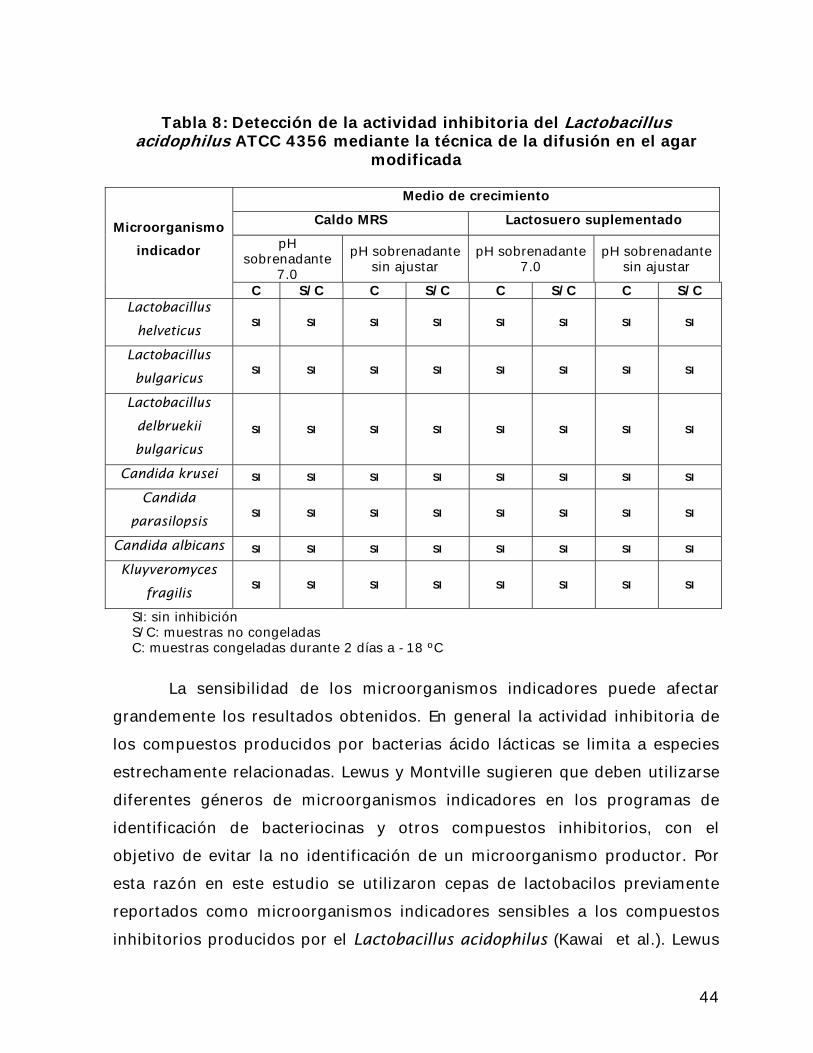
Tabla 8: Detección de la actividad inhibitoria del Lactobacillus acidophilus ATCC 4356 mediante la técnica de la difusión en el agar
modificada
Medio de crecimiento
Caldo MRS Lactosuero suplementado
pH sobrenadante
7.0
pH sobrenadante sin ajustar
pH sobrenadante 7.0
pH sobrenadante sin ajustar
Microorganismo
indicador
C S/C C S/C C S/C C S/C Lactobacillus
helveticus SI SI SI SI SI SI SI SI
Lactobacillus
bulgaricus SI SI SI SI SI SI SI SI
Lactobacillus
delbruekii
bulgaricus
SI SI SI SI SI SI SI SI
Candida krusei SI SI SI SI SI SI SI SI
Candida
parasilopsis SI SI SI SI SI SI SI SI
Candida albicans SI SI SI SI SI SI SI SI
Kluyveromyces
fragilis SI SI SI SI SI SI SI SI
SI: sin inhibición S/C: muestras no congeladas C: muestras congeladas durante 2 días a -18 ºC
La sensibilidad de los microorganismos indicadores puede afectar
grandemente los resultados obtenidos. En general la actividad inhibitoria de
los compuestos producidos por bacterias ácido lácticas se limita a especies
estrechamente relacionadas. Lewus y Montville sugieren que deben utilizarse
diferentes géneros de microorganismos indicadores en los programas de
identificación de bacteriocinas y otros compuestos inhibitorios, con el
objetivo de evitar la no identificación de un microorganismo productor. Por
esta razón en este estudio se utilizaron cepas de lactobacilos previamente
reportados como microorganismos indicadores sensibles a los compuestos
inhibitorios producidos por el Lactobacillus acidophilus (Kawai et al.). Lewus
44

45
y Montville reportan el Lactobacillus sake Lb 706 como el microorganismo
más sensible a los compuestos antimicrobianos producidos por otras cepas
de Lactobacillus, ya que en su estudio esa cepa fue inhibida por la mayoría de
los microorganismos productores utilizados.
Brink et al., reportan que Lactobacillus acidophilus M46 no inhibió el
crecimiento de Candida albicans ni de los microorganismos indicadores
Gram-negativos utilizados en su estudio. También reporta que los productos
formados por L. acidophilus M46 inhibieron sólo una cepa de Lactobacillus,
mientras que la actividad inhibitoria fue mayor contra Clostridium
sporogenes.
La eficacia de los compuestos inhibitorios también puede verse
afectada por la aparición espontánea de microorganismos mutantes
resistentes a los compuestos antimicrobianos, dentro de cepas sensibles a
estos (Schillinger et al., 1996).
La efectividad de los compuestos antimicrobianos puede
incrementarse mediante la simbiosis de estas sustancias con otros factores.
Por ejemplo Banina et al. reportan un aumentos del crecimiento del
Lactobacillus acidophilus cuando crece en simbiosis con otros
microorganismos. Por su parte Dykes, Amarowicz y Pegg (2003) reportan un
aumento en la actividad antimicrobiana de la nisina cuando esta se usa
mezclada con extracto de hojas de bayas.


CONCLUSIONES Y RECOMENDACIONES
No se detectó actividad antimicrobiana en el sobrenadante de los
cultivos de Lactobacillus acidophilus ATCC 4356 en caldo MRS o lactosuero
suplementado sobre los microorganismos indicadores utilizados,
posiblemente debido a la sensibilidad de la técnica. Ninguna de las
modificaciones efectuadas a la técnica original tuvieron efecto sobre los
resultados obtenidos previamente.
Se recomienda el uso de técnicas más sensibles en la detección de
actividad antimicrobiana, así como el uso de una mayor variedad de
microorganismos indicadores que incluyan los reportados como más
sensibles a los compuestos antimicrobianos. Adicionalmente se recomienda
la purificación de los compuestos antimicrobianos por técnicas tales como
HPLC, interacción hidrofóbica y precipitación con sulfato de amonio.
A pesar de que la velocidad de crecimiento fue mayor en el caldo MRS,
los mayores valores de concentración de biomasa se obtuvieron en lactosuero
suplementado. La velocidad de crecimiento del microorganismo fue mayor
con agitación de 50 rpm que sin agitación. Las concentraciones de biomasa
finales fueron menores cuando se utilizaron 50 rpm de agitación.
El lactosuero suplementado es un medio adecuado para el crecimiento
de Lactobacillus acidophilus ATCC 4356 ya que con su uso se obtienen
valores de velocidad de crecimiento y biomasa comparables a los obtenidos
con el medio específico para el crecimiento de lactobacilus (caldo MRS). Esto
constituye una alternativa de utilización del lactosuero, disminuyendo su
potencial contaminante y los altos costos de su disposición como desecho.
Adicionalmente es una alternativa como medio de crecimiento ante los costos
de los medios específicos para este microorganismo (caldo MRS).
47

REFERENCIAS BIBLIOGRÁFICAS
1. ADAMS, M.R. y MOSS, M. O. (1997). Microbiología de los alimentos. Zaragoza, España: Editorial Acribia.
2. American Type Culture Collection (ATCC) (1990). Catalogue of bacteria and bacteriophages. 18th Edition, 1992
3. Association of Official Analytical Chemists (A.O.A.C.) (1990). Official methods of analysis. Fifteenth Edition, 1990
4. BAKER; B. (1997). The Johns Hopkins Microbiology Newsletter, 16, (5). Disponible en: www.pathology5.pathology.jhmi.edu/micro/
5. BANINA, A., VUKASINOVIC, S., BRANKOVIC, S., FIRA, D., KOJIC, M. y TOPISIROVIC, L. (1998). Characterization of natural isolate Lactobacillus acidophilus BGRA43 useful for acidophilus milk production. Journal of Applied Microbiology, 84, 593-599.
6. BANWART, G. J. (1981). Basic Food Microbiology. New York, USA: Abridged Edition
7. BAREFOOT, S. y KLAENHAMMER, T. (1983). Detection and activity of lactacin B, a bacteriocin produced by Lactobacillus acidophilus. Applied and Environmental Microbiology, 45 (6), 1808-1815.
8. BERGER, K. y WELT, B. (2001). Food product quality: it isn’t just food safety that counyt. Case study: Food contamination issues and the influence of packaging. University of Florida. Cooperative Extension Service. Disponible en: www.edis.ifas.ufl.edu/BODY_AE203
9. BERNET-CAMARD, M., LIEVIN, V., BRASSART, D., NEESER, J., SERVIN, A. y HUDAULT, S. (1997). The human Lactobacillus acidophilus strain LA1 secretes a nonbacteriocin antibacterial substance(s) active in vitro and in vivo. Applied and environmental microbiology, 67 (7), 2747-2753.
10. BOGOVIC-MATIJASIC, B., ROGELJ, I., NES, I. y HOLO, H. (1998).Isolation and characterization of two bacteriocins of Lactobacillus acidophilus LF221. Applied Microbiology and Biotechnology, 49, 606-612.
11. BRINK, B., MINEKUS, M., VAN DER VOSSEN, J., LEER, R. y IN’T VELD, J. (1994). Antimicrobial Activity of lactobacilli: preliminary characterization and optimization of production of acidocin B, a novel bacteriocin produced by Lactobacllus acidophilus M46. Journal of
48

Applied Bacteriology, 77, 140-148.
12. BURIANEK, L. y YOUSEF, A. (2000). Solvent extraction of bacteriocins from liquid cultures. Letters in Applied Microbiology, 31, 193-197.
13. DAESCHEL, M. (1998, Enero)). Antimicrobial substances from lactic acid bacteria for use as food preservative. Food Technology, 164-167.
14. DAVE, R. y SHAH, N. Characteristics of bacteriocin produced by Lactobacillus acidophilus LA1.(1998). International Dairy Journal, 7 (11), 707-715.
15. DELVES-BOUGHTON, J., BLACKBURN, P., EVANS, R. J. y HUGENHOLTZ, J. (1996) Applications of the bacteriocin, nisin. Antonie van Leeuwenhoek, 69, 193-202.
16. DUBOIS, M., GILLES, K., HAMILTON, J., REBERS, P., SMITH, F. (1956) Colorimetric method for determination of sugars and related substances. Analitical Chemestry, 28.
17. DYKES, G. A., AMAROWICZ, R. y PEGG, R. B. (2003). Enhancement of nisin antibactyerial activity by a bearberry (Arctostaphylos uva-ursi) leaf axtract. Food Microbiology, 20 (2), 211-216.
18. ENTIAN, K. y DE VOS, W. (1996). Genetics of subtilin and nisin biosynteses. Antonie van Leeuwenhoek, 69, 109-117.
19. FRASQUET, J., VALENTÍN, A., APARISI, N., PÉREZ, C., PERNÁN, J. CANTÓN, E., y GOBERNADO, M. (2002). Cuatro años de candidemias en el hospital La Fe, 1998-2001. IV Congreso Nacional de Micrología. Disponible en: www.reviberoammicol.com/CNM2002
20. GASTROENTEROLOGÍA (2003). Esofagitis. Disponible en: www.iqb.es/ICD10/K20a.htm
21. GONZÁLEZ SISO, M.I. (1996). The biotechnological utilization of cheese whey: a review. Bioresource Technology, 57, 1-11.
22. JACK, R., TAGG, J. y RAY, B. (1995). Bacteriocins from Gram-Positive bacteria. Microbiological Reviews, 59 (2), 171-200.
23. JAKYMEC, M., MORÁN, H., PÁEZ, G., FERRER, J. R., MÁRMOL, Z. y RAMONES, E. (2001). Cinética de la producción de ácido láctico por fermentación sumergida con lactosuero como sustrato. Revista Científica, FCV-LUZ, XI (1), 53-59.
24. JARQUE, I., ANDREU, R., SALAVERT, M., SANZ. G., MARTÍN, G., PÉREZ,
49

B., PERNÁN, J., GOBERNADO, M. y SANZ, M. (2002). Epidemiología de la candidemia en pacientes con neoplasias hematológicas. Experiencia en un solo centro en un periodo de 16 años. VI Congreso Nacional de Micología. Disponible en: www.reviberoammicol.com/CNM2002
25. KAILASAPATHY, K. y CHIN, J. (2000). Survival and therapeutic potential of probiotic organisms with reference to Lactobacillus acidophilus and Bifidobacterium spp. Inmunilogy and Cell Biology 78, 80-88
26. KAWAI, Y., SAITO, T., UEMURA, J. y ITOH, T. (1997). Rapid detection for bacteriocin and distribution of bacteriocin-producin strains in Lactobacillus acidophilus group lactic acid bacteria isolated from human feces. Bioscience, Biotechecnology and Biochemestry, 61 (1), 179-182.
27. KHARDORI, N. (2002). Management of infections in patients with severe burns: impacto f multiresistant pathogens. Journal of Burns, 1 (1), 17. Disponible en: www.journalofburns.com/read
28. LEVEAU, J. Y. y BOUIX, M. (2000). Microbiología Industrial, Zaragoza, España: Editorial Acribia.
29. LEWUS, C., y MONTVILLE, T. (1991). Detection of bacteriocins produced by lactic acid bacteria. Journal of Microbiological Methods, 13, 145 -150.
30. LINDGREN, S. y DOBROGOSZ W. (1990). Antagonistic activities of lactic acid bacteria in food and feed fermentations. FEMS Microbiology Reviews, 87, 149-164
31. MARTÍNEZ, R. Curso de Microbiología de los Alimentos. Notas de clase
32. MATARAGAS, M., DROSINOS, E. H. y METAXOPOULOS, J. (2003). Antagonistic activity of lactic acid bacteria against Listeria monocytogenes in sliced cooked cured pork shoulder stored vacuum or modified atmosphere at 4±2 ºC. Food Microbiology, 20 (2), 259-265.
33. MICOLOGY ONLINE. (2003). Candida krusei. Disponible en www.mycology.adelaide.edu
34. MILO, L. (2000). A fresh look at food safety. Disponible en: www.preparedfood.com
35. MINISTERIO DE LA PRODUCCIÓN Y EL COMERCIO. (1999). Anuario estadístico agropecuario. División de Planificación Agrícola. UEMPC-
50

Zulia.
36. MITAL, B.K. y SATYENDRA, K. G. (1995). Anticarcinogenic, hipocholesterolemic and antagonistic activities of Lactobacillus acidophilus, 21 (3), 175-214.
37. MONTIEL, X., CARRUYO, I., FERRER, J., MARCANO, L., MÁRMOL, Z. y PAÉZ, G. (2000). Producción de -galactosidasa en cultivo por carga de Kluyveromyces fragilis empleando como sustrato suero de leche. Revista Técnica de la Facultad de Ingeniería, 23 (2), 134-140.
38. MOSSEL, D. A. y MORENO GARCÍA, B. (1982). Microbiología de los Alimentos. Zaragoza, España: Editorial Acribia.
39. MÜLLER, G. (1981). Microbiología de los Alimentos Vegetales. Zaragoza, España: Editorial Acribia.
40. NAGENDRA, P. S. y RAJIV, I. D. (2001). Characteristics of bacteriocin like inhibitory substances produced by Lactobacillus acidophilus (BDLA-1, 2409 and MOLA-2), Lactobacillus fermentum (5174) and Lactobacillus plantarum (2903). (2001). Disponible en: www.bifidus.bcasj.or.jp
41. NARDI, R. D., SANTOS, A. R., CARVALHO, M. A., FARIAS, L. M., BECHETRIT, L. C. y NICOLI, J. R. (2003). Antagonism against anaerobic and facultative bacteria through a diffusible inhibitory compound produced by a Lactobacillus sp. Isolated from the rat fecal microbiota. Disponible en: www.anaerobe.org
42. OBERTO, A. y CARRASQUERO Z. (1986). Producción de etanol por fermentación del suero de leche. Trabajo especial de grado. Facultad de Ingeniería. LUZ. Maracaibo.
43. PARAJE, M., ALBESA, I. y ERASO, A. (2000). Conservation in probiotic preparation of Lactobacillus with inhibitory capacity on other species. New Microbiology, 23 (4), 423-431.
44. PARENTE, E. y HILL, C. (1992). A comparison of factors affecting the production of two bacteriocins from lactic acid bacteria. Journal of Applied Bacteriology, 73, 290-298.
45. PARENTE, E., BRIENZA, C., MOLES, M. y RICCIARDI A. (1995). A comparison of methods for the measurement of bacteriocin activity. Journal of Microbiological Methods, 22, 95-108.
46. PLOCKOVA, M., CHUMUCHALOVA, J. y TOMANOVA, J. (1997).
51

Antifungal activity of Lactobacillus acidophilus CH5 metabolites. Czech Journal of Food Science, 15 (1,) 39-48.
47. QUINTERO, H., RODRÍGUEZ, M., PÁEZ, G., FERRER, J., MÁRMOL, Z. y RINCÓN, M. (2001) Producción continua de proteína microbiana (K. fragilis) a partir de suero de leche. Revista Científica, FCV-LUZ, XI (2), 87-94.
48. RAMONES, E. (2000). Producción de ácido láctico por un cultivo mixto de L. helveticus y L. bulgaricus en suero de leche. Trabajo de grado. La Universidad del Zulia. Facultad de Ingeniería. División de Estudios para Graduados.
49. RODRÍGUEZ, E., TOMILLO, J., NÚÑEZ, M. y MEDINA, M. (1997). Combined effect of bacteriocin-producing lactic acid bacteria and lactoperoxidase system activation on Listeria monocitogenes in refrigerated raw milk. Journal of Applied Microbiology, 83, 389-395.
50. SCHILLINGER, U., GEISEN, R. y HOLZAPFEL, W. H. (1996, Mayo). Potencial of antagonistic microorganism and bacteriocins for the biological preservation of foods. Trends in Food Science & Technology, (7), 158-164.
51. SCOTT, R. (1991). Fabricación de queso. Zaragoza, España: Editorial Acribia.
52. SHIMIZU, H., MIZUGUCHI, T., TANAKA, E. y SHIOYA, S. (1999). Nisin production by a mixed-culture system consisting of Lactococcus lactis and Kluyveromyces marxianus. Applied and Environmental Microbiology, 65 (7), 3134-3141.
53. SIMOVA, E., BESHKOVA, D., ANGELOV, A., HRISTOZOVA, T., FRENGOVA, G. y SPASOV, Z. (2002). Lactic acid bacteria and yeast in kefir grains and kefir made from them. Journal of Industrial Microbiology and Biotechnology, 28 (1), 1-6.
54. THE SOCIETY OF FOOD HYGIENE TECHNOLOGY (2003). Yeast. Disponible en: www.sofht.co.uk/technical/yeast.htm
55. TOMSIKOVA, A. (2002). Risk of fungal infection from foods, particulary in immunocompromised patients. Epidemiology, Microbiology and Imunology, 51 (2), 78-81.
56. TRINDADES, R., RESENDE, M., SILVA, C. y ROSA, C. (2002). Yeast associated with fresh and frozen pulps of Brazilian tropical fruits. Syst Applied Microbiology, 25 (2), 294-300
52

53
57. VEISSEYRE, R. (1972). Lactología Técnica. Zaragoza España: Editorial Acribia.
58. VIELMA, A. y URRIBARRÍ, L. (1999). Producción de ácido láctico en cultivo continuo utilizando Lactobacillus helveticos a partir de suero de leche. Trabajo de Grado. La Universidad del Zulia. Facultad de Ingeniería. Escuela de Ingeniería Química.
59. YANG, R., JONSON, M. y RAY, B. (1992). Novel method to extract large amounts of bacteriocins from lactic acid bacteria. Applied and Environmental Microbiology, 58 (10), 3355-3359.


















