
Cambios en el espermatozoide desde la eyaculación hasta la...
12
38 Luño Lázaro, Victoria y Matas Parra, Carmen Cambios en el espermatozoide desde la eyaculación hasta la fecundación Victoria Luño Lázaro 1 y Carmen Matas Parra 2 1 Dpto. Patología Animal (Área de Reproducción y Obstetricia). Facultad de Veterinaria. Universidad de Zaragoza 2 Dpto. Fisiología (Fisiología Veterinaria).Facultad de Veterinaria. Universidad de Murcia. [email protected]. http://www.um.es/grupo-fisiovet Foto Axón Comunicación ❱ ❱ Introducción Los espermatozoides tras ser eyaculados ascienden por el tracto genital femenino, donde solo una pequeña proporción es capaz de unirse a las células epiteliales del oviducto en el denominado reservorio espermático. Aquí, permanecen hasta el momento de la ovulación en el que son liberados secuencialmente gracias a modi- ficaciones endocrinas, que conllevan una serie de cambios a nivel de membrana plas- mática del epitelio oviductal o en el fluido intraluminal (Fazeli et al., 1999) (Figura 1). El oviducto juega un papel modulador importante en el proceso reproductivo, controlando y sincronizando a los gametos hasta su unión. La interacción entre los espermatozoides y las células oviductales durante dicho ascenso permite que los espermatozoides adquieran la capacidad fecundante a través de un proceso denomi- nado capacitación, el cual fue descrito por primera vez simultáneamente por Austin y Chang (1951), y le confiere a los esper- matozoides la capacidad de penetrar al ovocito (Figura 2). Las modificaciones producidas por este proceso tienen varias consecuencias en el gameto. Entre ellas podemos encontrarnos la modificación en el patrón de movimien- to, la generación de radicales libres, el incremento del nivel de calcio, o la modi- ficación de lípidos de membrana (Matas et al., 2010). No obstante, parece ser que una de las modificaciones más importantes que se producen es el aumento los niveles de fosforilación en tirosina de las proteínas (Figura 3). En varias especies de mamíferos la fosforilación en residuos en tirosina ha sido correlacionada con el estado de capa- citación del espermatozoide. (Visconti et al., 1995; 1999; Tadiff et al., 2001; Pommer et al., 2003; Grasa et al., 2006). Éste se plantea como uno de los principales criterios para la unión entre las células oviductales y los espermatozoides, representando un meca- nismo de selección de la población esper- mática más competente para la fecunda- ción (Fazeli et al., 1999).
Transcript of Cambios en el espermatozoide desde la eyaculación hasta la...
38
Luño Lázaro, Victoria y Matas Parra, Carmen
Cambios en el espermatozoide desde la eyaculación hasta la fecundación
Victoria Luño Lázaro1 y Carmen Matas Parra2
1Dpto. Patología Animal (Área de Reproducción y Obstetricia). Facultad de Veterinaria. Universidad de Zaragoza
2Dpto. Fisiología (Fisiología Veterinaria).Facultad de Veterinaria. Universidad de Murcia. [email protected]. http://www.um.es/grupo-fisiovet
Foto Axón Comunicación
❱❱ Introducción
Los espermatozoides tras ser eyaculados ascienden por el tracto genital femenino, donde solo una pequeña proporción es capaz de unirse a las células epiteliales del oviducto en el denominado reservorio espermático. Aquí, permanecen hasta el momento de la ovulación en el que son liberados secuencialmente gracias a modi-ficaciones endocrinas, que conllevan una serie de cambios a nivel de membrana plas-mática del epitelio oviductal o en el fluido intraluminal (Fazeli et al., 1999) (Figura 1).
El oviducto juega un papel modulador importante en el proceso reproductivo, controlando y sincronizando a los gametos hasta su unión. La interacción entre los espermatozoides y las células oviductales durante dicho ascenso permite que los espermatozoides adquieran la capacidad fecundante a través de un proceso denomi-nado capacitación, el cual fue descrito por primera vez simultáneamente por Austin y Chang (1951), y le confiere a los esper-matozoides la capacidad de penetrar al ovocito (Figura 2).
Las modificaciones producidas por este proceso tienen varias consecuencias en el gameto. Entre ellas podemos encontrarnos la modificación en el patrón de movimien-to, la generación de radicales libres, el incremento del nivel de calcio, o la modi-ficación de lípidos de membrana (Matas et al., 2010). No obstante, parece ser que una de las modificaciones más importantes que se producen es el aumento los niveles de fosforilación en tirosina de las proteínas (Figura 3).
En varias especies de mamíferos la fosforilación en residuos en tirosina ha sido correlacionada con el estado de capa-citación del espermatozoide. (Visconti et al., 1995; 1999; Tadiff et al., 2001; Pommer et al., 2003; Grasa et al., 2006). Éste se plantea como uno de los principales criterios para la unión entre las células oviductales y los espermatozoides, representando un meca-nismo de selección de la población esper-mática más competente para la fecunda-ción (Fazeli et al., 1999).
39
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
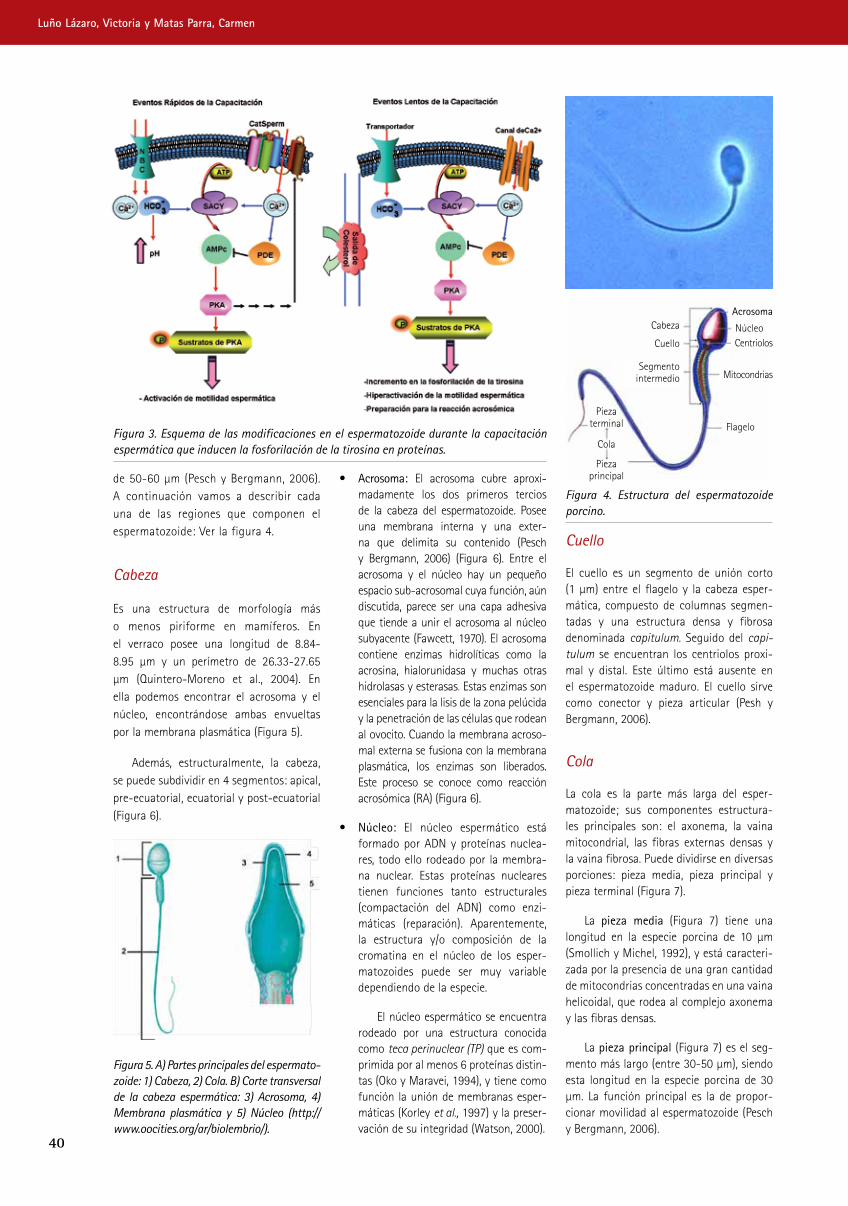
NBC: Cotransportador de Na+/HCO3, SACY: Adenilato Ciclasa Soluble; PKA: Protein kinasa A (Adaptado de Visconti, 2009).
In vitro se han desarrollado diversas técnicas para capacitar a los espermato-zoides intentando imitar las condiciones fisiológicas. Entre ellas se encuentran, los lavados con albúmina, lavados en gradien-tes de diferente densidad o simplemente centrifugación e incubación en un medio rico en calcio y bicarbonato (Matás et al., 2003). De la misma forma, median-te cultivos in vitro de células oviductales porcinas (POEC) se ha intentado simular las condiciones que los espermatozoides encuentran en el tracto genital femenino. Se ha determinado un mayor porcentaje de
espermatozoides unidos con un nivel bajo de fosforilación en tirosina y de concen-tración de calcio intracelular en diferen-tes especies, representando la población más viable destinada a la fecundación (Petrunkina et al., 2001; 2003a; Zumoffen et al., 2010). Otra posibilidad es el uso de explantes procedentes de oviductos de hembras recién ovuladas en el momento del sacrificio en mataderos, o la utilización de oviductos de hembras que han sido inseminadas bajo condiciones de campo. También se ha visualizado los esperma-tozoides in situ tras inseminaciones en cerda y vaca, demostrándose interacciones entre el epitelio y la membrana plasmática espermática en la zona caudal del istmo (Hunter, 1991).
❱❱ El espermatozoide. Generalidades
El espermatozoide (del griego esperma-semilla, y zoon-animal) es una célula haploide que constituye el gameto mascu-lino y cuya función es la formación de un cigoto totipotente, resultado de la fusión de su núcleo con el del gameto femenino, fenómeno que dará lugar, posteriormente, al embrión y al feto.
La célula espermática tiene 3 regio-nes altamente diferenciadas y especia-lizadas: cabeza, cuello y cola o flagelo (Figura 4). En el caso del verraco la longitud total del espermatozoide es
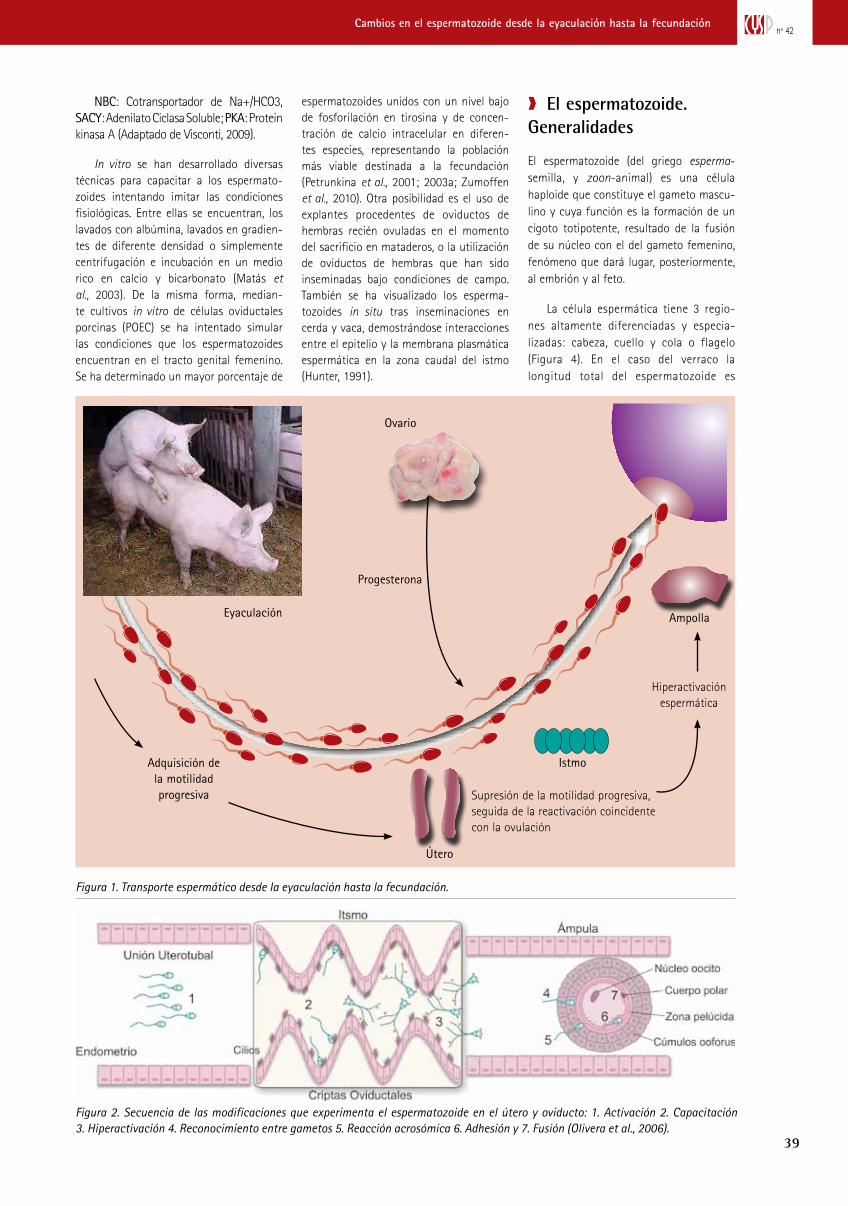
Figura 1. Transporte espermático desde la eyaculación hasta la fecundación.
Eyaculación
Progesterona
Útero
Adquisición de la motilidad progresiva Supresión de la motilidad progresiva,
seguida de la reactivación coincidente con la ovulación
Hiperactivación espermática
Ovario
Istmo
Ampolla
Figura 2. Secuencia de las modificaciones que experimenta el espermatozoide en el útero y oviducto: 1. Activación 2. Capacitación 3. Hiperactivación 4. Reconocimiento entre gametos 5. Reacción acrosómica 6. Adhesión y 7. Fusión (Olivera et al., 2006).
40
Luño Lázaro, Victoria y Matas Parra, Carmen
de 50-60 µm (Pesch y Bergmann, 2006). A continuación vamos a describir cada una de las regiones que componen el espermatozoide: Ver la figura 4.
Cabeza
Es una estructura de morfología más o menos piriforme en mamíferos. En el verraco posee una longitud de 8.84-8.95 µm y un perímetro de 26.33-27.65 µm (Quintero-Moreno et al., 2004). En ella podemos encontrar el acrosoma y el núcleo, encontrándose ambas envueltas por la membrana plasmática (Figura 5).
Además, estructuralmente, la cabeza, se puede subdividir en 4 segmentos: apical, pre-ecuatorial, ecuatorial y post-ecuatorial (Figura 6).
• Acrosoma: El acrosoma cubre aproxi-madamente los dos primeros tercios de la cabeza del espermatozoide. Posee una membrana interna y una exter-na que delimita su contenido (Pesch y Bergmann, 2006) (Figura 6). Entre el acrosoma y el núcleo hay un pequeño espacio sub-acrosomal cuya función, aún discutida, parece ser una capa adhesiva que tiende a unir el acrosoma al núcleo subyacente (Fawcett, 1970). El acrosoma contiene enzimas hidrolíticas como la acrosina, hialorunidasa y muchas otras hidrolasas y esterasas. Estas enzimas son esenciales para la lisis de la zona pelúcida y la penetración de las células que rodean al ovocito. Cuando la membrana acroso-mal externa se fusiona con la membrana plasmática, los enzimas son liberados. Este proceso se conoce como reacción acrosómica (RA) (Figura 6).
• Núcleo: El núcleo espermático está formado por ADN y proteínas nuclea-res, todo ello rodeado por la membra-na nuclear. Estas proteínas nucleares tienen funciones tanto estructurales (compactación del ADN) como enzi-máticas (reparación). Aparentemente, la estructura y/o composición de la cromatina en el núcleo de los esper-matozoides puede ser muy variable dependiendo de la especie.
El núcleo espermático se encuentra rodeado por una estructura conocida como teca perinuclear (TP) que es com-primida por al menos 6 proteínas distin-tas (Oko y Maravei, 1994), y tiene como función la unión de membranas esper-máticas (Korley et al., 1997) y la preser-vación de su integridad (Watson, 2000).
Cuello
El cuello es un segmento de unión corto (1 µm) entre el flagelo y la cabeza esper-mática, compuesto de columnas segmen-tadas y una estructura densa y fibrosa denominada capitulum. Seguido del capi-tulum se encuentran los centriolos proxi-mal y distal. Este último está ausente en el espermatozoide maduro. El cuello sirve como conector y pieza articular (Pesh y Bergmann, 2006).
Cola
La cola es la parte más larga del esper-matozoide; sus componentes estructura-les principales son: el axonema, la vaina mitocondrial, las fibras externas densas y la vaina fibrosa. Puede dividirse en diversas porciones: pieza media, pieza principal y pieza terminal (Figura 7).
La pieza media (Figura 7) tiene una longitud en la especie porcina de 10 µm (Smollich y Michel, 1992), y está caracteri-zada por la presencia de una gran cantidad de mitocondrias concentradas en una vaina helicoidal, que rodea al complejo axonema y las fibras densas.
La pieza principal (Figura 7) es el seg-mento más largo (entre 30-50 µm), siendo esta longitud en la especie porcina de 30 µm. La función principal es la de propor-cionar movilidad al espermatozoide (Pesch y Bergmann, 2006).
Figura 3. Esquema de las modificaciones en el espermatozoide durante la capacitación espermática que inducen la fosforilación de la tirosina en proteínas.
Figura 5. A) Partes principales del espermato-zoide: 1) Cabeza, 2) Cola. B) Corte transversal de la cabeza espermática: 3) Acrosoma, 4) Membrana plasmática y 5) Núcleo (http://www.oocities.org/ar/biolembrio/).
Figura 4. Estructura del espermatozoide porcino.
Cabeza
Cuello
Flagelo
Pieza terminal
Cola
Pieza principal
Mitocondrias
CentriolosNúcleo
Acrosoma
Segmento intermedio

41
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
La pieza terminal (Figura 7) es la parte distal del flagelo, es una porción corta y tiene una longitud de 3 µm.
❱❱ Eyaculación
La eyaculación es el reflejo de expulsión del semen fuera del tracto reproductivo a través de la uretra mediante contracciones musculares coordinadas. El cerdo eyacula un volumen que varía entre 200 y 250 ml y se encuentra compuesto por una sus-pensión de espermatozoides en un fluido denominado plasma seminal.
Se diferencian tres fracciones en el eyaculado en función de la procedencia de las mismas (Garner y Hafez, 1996):
1. Fracción pre-espermática: formada por las secreciones procedentes de la prós-tata, las vesículas seminales y de las glándulas bulbouretrales.
2. Fracción espermática: presenta una concentración elevada de espermato-zoides junto con secreciones de la próstata y de las vesículas seminales.
3. Fracción post-espermática: tiene una baja concentración espermática y está constituida por las secreciones de la próstata y de las glándulas bul-bouretrales. Éstas últimas producen abundantes corpúsculos gelatinosos denominados comúnmente “tapio-ca”, que van a formar una especie de tapón para sellar el cérvix de la hembra, evitando en la monta natural el reflujo del eyaculado del verraco (Geisert, 1999).
❱❱ Plasma seminal
El plasma seminal es el fluido en el que se encuentran inmersos los espermato-zoides de los mamíferos domésticos. Está compuesto por una mezcla compleja de secreciones procedentes de los testícu-los, el epidídimo y las glándulas sexuales accesorias (vesículas seminales, próstata y glándulas bulbouretrales). El conteni-do de estas glándulas es específico de cada especie y muy variable incluso entre individuos de la misma especie o entre eyaculados del mismo animal. Entre las causas más importantes de esta variación, aparecen diversos procesos patológicos, la estación del año, el estado fisiológico del animal o las variaciones de temperatura (Pérez-Pé et al., 2001; Cardozo et al., 2006; Colenbrander y Kemp, 1990). Entre los compuestos secretados encontramos dis-tintos iones como por ejemplo, el Zn2+ que tiene propiedades antibacterianas (Strzezek y Cieresko, 1987) y que en el caso del plasma seminal de verraco alcanza concen-traciones muy altas (0.3-0.7 nM) (Arver y Eliasoson, 1980); y el Ca2+, que participa en los procesos de capacitación espermática y reacción acrosómica (Visconti et al., 2002). También vamos a encontrar distintos sus-tratos energéticos para el metabolismo de la célula, como fructosa, inositol, ácido cítrico y ácido ascórbico (Frei et al., 1990), aminoácidos (ácido glutámico, carnitina, taurina, hipotaurina) que servirán como fuente de energía y de protección contra sustancias oxígeno reactivas y distintas enzimas (proteasas, acrosina, nucleasas, fosfatasa ácida y alcalina y superóxido dismutasa) que intervendrán en la licue-facción seminal, penetración del ovocito
por parte del espermatozoide, digestión de espermatozoides muertos y dañados, y protección contra sustancias oxígeno reactivas (Strzezek, 2002). Por tanto, el papel del plasma seminal no es el de ser un líquido meramente nutritivo y de trans-porte, sino que también posee funciones de protección, regulación de la motilidad y capacitación de los espermatozoides, reconocimiento y unión entre gametos (Johnson et al., 2000), (Figura 9).
De todos los componentes del plasma seminal, parece ser que son las proteínas las que contribuyen de forma más impor-tante en el proceso reproductivo (Töpfer-Petersen, 1999). Además, el plasma seminal va a contener tanto factores que van a prevenir reacciones acrosómicas espontá-neas o prematuras (factores decapacitan-tes) como otros factores que van a propi-ciar la fertilización de los espermatozoides (Thérien et al., 1995). Estudios recientes sugieren que la acción concertada de los factores seminales reguladores del plas-ma, modulan el estado de capacitación de los espermatozoides de las especies de mamíferos. Los efectos biológicos de estos factores del plasma en la funcionalidad espermática, no están hoy en día del todo
Figura 7. A) Dibujo esquemático de un espermatozoide. B) Sección longitudinal de un espermatozoide; C) y H) secciones transversales de la cola del espermato-zoide: (C) pieza media proximal, (D) pieza media distal, E) y F) pieza principal, G) y H) pieza terminal. (1) Membrana plasmá-tica, (2) acrosoma, (3) núcleo, (4) vacuola nuclear, (5) centriolo proximal, (6) mito-condria, (7) fibras externas densas (ODF), (8) túbulo exterior doble, (9) vaina o envol-tura fibrosa; (←) unión de la pieza media con la hélice mitocondrial y la pieza prin-cipal con la vaina fibrosa (Imagen tomada de Pesch y Bergmann, 2006).
Figura 6. A) Esquema de la estructura del espermatozoide. B) Corte sagital de la célula espermática: 1: Membrana espermática, 2: Membrana acrosomal externa, 3: Contenido acrosomal, 4: Membrana acrosomal interna, 5: Núcleo celular, 6: Contenido de ADN altamente condensado, 7: Anillo posterior o placa basal, 8: Pieza media, 9: Mitocondria, 10: Anillo anular, 11: Flagelo. C) 12: Vesícula (fusión de la membrana plasmática con la membrana acrosomal externa, 13: Secreción acrosomal. D) 14: Estructura en horquilla (Imagen de Flesch y Gadella 2000).
1234
5
6
7
8
9
1
1
C.
D.
E.
F.
G.
H.1
1
1
1
2
6
6
3
3
1
4
4
4
4
2
8
7
1
8
9
4
A. B.
42
Luño Lázaro, Victoria y Matas Parra, Carmen
claros debido en parte a la alta variabilidad en la composición del plasma seminal entre especies, entre machos dentro de la misma especie y entre eyaculados de un mismo macho (Killian et al., 1993; Zhu et al., 2000).
❱❱ Ascenso por el tracto genital femenino
Tras la monta o la inseminación artificial, alrededor de 3000 millones de espermato-zoides son depositados en el cérvix uterino, de los cuales solo una pequeña proporción es capaz de alcanzar el oviducto (Figura 9).
Durante el ascenso la mayoría de los espermatozoides (>70%) son eliminados en la luz uterina, parte de ellos mediante el reflujo retrógrado (35%), por lo que solo unos pocos son capaces de atravesar la unión útero-tubárica (Rodriguez-.Martinez et al., 2005). También se produce una fago-citosis, mediada principalmente por leu-cocitos que migran al lumen procedentes del endometrio en los primeros 30 minu-tos tras la monta (Matthijs et al., 2003). Hay varios factores implicados que inician esta repuesta, como la propia distensión uterina, los espermatozoides o el plasma seminal (Bischof et al., 1994). Proteínas como el heterodímero glicosilado PSP-I/
PSP-II posee actividad inmuno-estimula-dora, y es capaz de inducir la migración de leucocitos (Leshin et al., 1998). Ese periodo de latencia hasta que se produce la migración al lumen uterino, permite que cierta población espermática atraviese el útero y no sea fagocitada (Rodríguez-.Martínez et al., 2005). Durante dicho ascenso se produce también la progresiva y coordinada modificación de ciertos fac-tores procedentes del plasma seminal que están unidos a la superficie plasmática del espermatozoide (Rodríguez-.Martínez et al., 2004). Estos, previenen capacitaciones prematuras, intervienen en la tolerancia inmunitaria, a la vez que están involucra-dos en la unión con las células oviductales (Rodríguez-Martínez, 2009). Finalmente solo una pequeña proporción atravesará la unión útero-tubárica.
Selección de la población espermática
Existen diferentes mecanismos a lo largo del tracto genital femenino que permiten una selección progresiva de la población más apta para fecundar, estableciéndose diferentes subpoblaciones espermáticas. Estas, son parcial o totalmente deficientes en uno o más aspectos fenotípicos o fun-cionales necesarios para participar en los diferentes aspectos de la fecundación. Si inseminamos con la misma concentración de espermatozoides, el número capaz de acceder al oviducto varía de un individuo a otro (Satake et al., 2006), ya que cada macho produce su particular proporción de subpoblaciones (Holt, 2009).
Existen variaciones fenotípicas que determinan diferentes tipos de esperma-tozoides, éstas van desde la forma de la cabeza, la motilidad o la amplitud fla-gelar (Thurston et al., 2001), a otras que involucran la eficiencia de las señales de transducción (Holt y Harrison 2002) y la integridad de la información genética y epigenética que transmite al ovocito (Miller, 2007).
El paso por la (UTJ) es uno de los prin-cipales mecanismos de selección (Figura 9), ya que alrededor del 96% de los esper-matozoides localizados en el oviducto se encuentran allí (Mburu et al., 1996). Es atravesada por gametos que presentan una motilidad progresiva y una determinada composición bioquímica a nivel de mem-brana. La integridad molecular de la super-ficie del espermatozoide es determinante para su entrada, por lo que sucesos como la capacitación que producen modificacio-nes estructurales, disminuyen la capacidad del espermatozoide de llegar al oviducto y
Figura 8. Representación esquemática de la membrana plasmática bajo distintas circuns-tancias: en el epidídimo, en contacto con el plasma seminal (eyaculado) y en el genital de la hembra (capacitado).
Figura 9. Transporte espermático en el genital femenino y eventos que acontecen en las distintas secciones del tracto reproductivo de la hembra (Modificado de Seger 1999).
4. OviductoCapacitación completada
Hiperactividad
3. ÚteroInicio de capacitación
Fagocitos
2. CérvixBarrera espermática
Reservorio espermático
1. Transporte inmediatoPérdida retrógrada
FagocitosEntrada en cérvix y útero
5. FecundaciónReacción acrosómica
Penetración espermáticaFormación de pronúcleos
UT
Lumen

43
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
unirse a las células oviductales (Fazeli et al., 1999). Se ha determinado que aproxi-madamente el 70 % de los espermatozoi-des en el oviducto están no capacitados (Tienthai et al., 2004). El daño del material genético presente en el espermatozoide ha sido correlacionado con el incremento de la ubiquitinación de la superficie espermática (Sutovsky et al., 2002), lo cual produciría alteraciones en la membrana similares a las ocurridas en procesos como la capacita-ción, lo que disminuiría su paso por la UTJ.
Podemos concluir que la selección de la población espermática que llega a oviducto (Figura 9) está determinada por diferentes factores clasificados como:
• Tipo 1 (factores fijos): específicos del testículo, están determinados gené-ticamente o afectados por las con-diciones como elevada temperatura testicular.
• Tipo 2 (factores variables): influencia-dos por tratamientos como el lavado, la congelación…La motilidad se incluye en ambos porque está determinada por aspectos genéticos y puede ser modifi-cada su expresión.
• Tipo 3 (factores variables): determinada por señales endocrinas dentro del trac-to genital femenino.
Oviducto
El oviducto tiene un papel significativo en el proceso reproductivo de los mamíferos (Figura 10), ya que proporciona un ambiente apropiado para el transporte y maduración de gametos, la fecundación y el desarrollo embrionario temprano (Hunter, 2005). Está dividido en cuatro segmentos funcionales: el infundíbulo, la ampolla, el istmo y la unión útero-tubárica, cada una de ellas con una función determinada (Hunter ,1994). El infundíbulo es el encargado de captar al ovocito en el momento de la ovulación gracias a la estructura que presenta en forma de dedos. De este modo, el gameto es capaz de llegar hasta la ampolla donde tendrá lugar la fecundación. En el istmo se produce la formación del reservorio espermático (Suarez, 2002), que permite el mantenimiento de la población viable apara la fecundación. Por último la unión útero-tubárica representa la conexión con el cuerno uterino y permite regu-lar y controlar el transporte espermático (Hunter, 1991).
Histológicamente el oviducto consta de tres capas, una serosa externa, una muscular media y una mucosa interna.
El epitelio está formado por células cilia-das y secretoras, distribuidas en diferentes proporciones según la región del oviducto (Gandolfi, 1995). Así, en el infundíbulo y la ampolla se encuentran en mayor pro-porción las células ciliadas y en menor las secretoras, mientras que en la región del istmo ocurre lo contrario (Abe, 1996). Se ha demostrado la presencia de receptores para estrógenos y progesterona a lo largo de todo el oviducto (Amso et al., 1994), que modulan la cantidad celular e intensidad de movimiento ciliar según la fase del ciclo estral. Las células secretoras del istmo acu-mulan gránulos secretores durante la fase folicular, los cuales son eliminados al fluido folicular tras la ovulación (Abe, 1996). Estas células vierten al lumen diferentes macro-moléculas (Abe, 1996), que proporcionan el ambiente idóneo para las diferentes fases del proceso de fecundación.
Fluido oviductal
El fluido oviductal representa el medio acuo-so en el que gametos y embriones están sus-pendidos durante su recorrido por el oviduc-to. Está formado por una secreción específica de origen epitelial y un exudado procedente del plasma sanguíneo (Leese et al., 2001). Sin embargo, posee una concentración iónica, pH, osmolaridad y contenido macromolecular diferente a la que se encuentra en el plasma sanguíneo (Leese, 1988). El pH, determinado principalmente por la concentración de bicar-bonato, varían según el momento del ciclo estral y la región oviductal (Zhou et al., 2005). En la cerda, hacia mitad del ciclo, el pH fluc-túa entre 0,3 y 0,7 unidades entre la ampolla y el istmo respectivamente, aumentando en el momento de la ovulación hasta 0,4 uni-dades (Nichol et al., 1997). Estas variaciones pueden tener un gran significado, dado que
ese pH alcalino podría influenciar en la moti-lidad y capacitación espermática cuando está en la proximidad el gameto femenino.
El fluido oviductal también varía en volumen y composición según el momento del ciclo, bajo influencia hormonal (Leese et al., 2001). Está compuesto por una varia-da cantidad de proteínas, entre las que destacan un grupo de glicoproteínas espe-cificas del oviducto (OGP) u oviductinas (Killian, 2004), que podrían estar involucra-das en modificaciones estructurales de la ZP del ovocito (Hunter, 1994). También posee un alto contenido en glicosaminoglicanos (GAG), sulfatados (acido hialurónico) y no sulfatados (Tienthai et al., 2000). Las con-centraciones varían según el momento del ciclo estral y entre las regiones oviductales. Estas se incrementan durante los momentos preovulatorios, principalmente a nivel del istmo en el cerdo (Tienthai et al., 2000). Parece que están involucrados en la super-vivencia espermática la capacitación y la unión-liberación del reservorio espermático (Rodríguez-Martínez et al., 2001; 2005).
Reservorio espermático
En la porción caudal del istmo oviductal se produce la formación del denomina-do reservorio oviductal (Figura10) durante el periodo pre y peri-ovulatorio (Suarez et al., 1991; Töpfer-Petersen et al., 2002). La permanencia de los espermatozoides depende del momento del ciclo estral, variando entre 36 y 48 horas (Hunter, 1984), situándose tanto en el lumen como unidos al epitelio oviductal, principalmente a las células ciliadas (Mburu et al., 1996).
Tiene carácter multifuncional, ya que modula la capacitación espermática, man-
Figura 10. Esquema del estado de los espermatozoides en las distintas secciones del oviducto.

44
Luño Lázaro, Victoria y Matas Parra, Carmen
tiene la viabilidad del gameto masculino, sincroniza el tiempo de maduración esper-mática con el momento de la ovulación y selecciona los espermatozoides con mejor morfología (Töpfer-Petersen, 1999). La unión con las células oviductales permite que un número adecuado de gametos viables y fértiles estén disponibles para la fecundación en el momento que tiene lugar la ovulación, y por otro reducir el riesgo de polispermia al permitir que un número reducido de espermatozoides asciendan a la vez hasta el lugar de fecundación.
Presenta una capacidad de unión selec-tiva, escogiendo la población espermática más competente para la fecundación, la cual presenta determinadas características relacionadas con la morfología, motili-dad, integridad de membrana, nivel de calcio citosólico o estado de capacitación y fosforilación en tirosina de las pro-teínas (Ellington et al., 1998; Fazeli et al., 1999; Petrunkina et al., 2001; Gualtieri y Talevi, 2003).
Varios factores pueden estar involucra-dos en la formación del reservorio esper-mático. La unión se produce por contacto directo mediante una interacción ligando-receptor entre las moléculas presentes en la cara rostral del espermatozoide y car-bohidratos de la membrana plasmática de las células oviductales de manera especie-especifico (Lefebvre y Suarez, 1996; Suarez et al., 1998). Dicha unión es un proce-so reversible que involucra en todas las especies estudiadas a diferentes azúcares (Dobrinski et al., 1996a, Suarez et al., 1998). La relativa contribución de otros factores como las secreciones mucosas, las propie-dades químicas del fluido oviductal o los gradientes de temperatura, podría contri-buir en diferente grado a la formación del reservorio.
En el porcino las espermadesinas, en especial AQN1, se encuentran íntimamen-te unidas a través de fosfolípidos a la
Cambios en los patrones de movimiento: hiperactivación
El espermatozoide adquiere la capacidad de mover el flagelo en su tránsito por el epidí-dimo, pero el movimiento empieza después de la eyaculación. Este proceso es conocido como “activación del espermatozoide”. El movimiento del flagelo es característico y consiste en un batido simétrico de la cola que hace que el espermatozoide se despla-ce de forma progresiva. El semen pasa rápi-damente a través del cuello y el útero de la hembra; a los 10 minutos de su deposición encontramos espermatozoides en la unión útero-tubárica. De aquí, los espermatozoi-des pasan al reservorio del oviducto (istmo) caracterizado por cilios luminares epitelia-les y plegamientos o criptas de la mucosa oviductal (Figura 11).
Los espermatozoides son retenidos en las criptas oviductales y allí pierden los fac-tores decapacitantes que habían aportado las glándulas accesorias, experimentando la capacitación.
In vivo, los espermatozoides de verra-co se acumulan en el istmo del oviducto uniéndose a la membrana de las célu-las del epitelio (Suarez, 2002; Tienthai et al., 2004). Después, estos espermatozoides pueden ser liberados de este lugar cuan-do son capacitados (Fazeli et al., 1999; Rodríguez-Martínez et al., 2001). El proceso de capacitación espermática está asociado al desarrollo de un patrón de movimiento diferente conocido con el nombre de hipe-ractivación y que se observa en el momen-to cercano a la fecundación. Se considera que la hiperactivación es crítica para la fecundación (Stauss et al., 1995) porque permite al espermatozoide desprenderse de la pared del oviducto, nadar en el lumen del oviducto o penetrar las células del cumulus oophorus para alcanzar el ovocito. Además, se va a considerar como uno de los pará-metros más importantes en la valoración de la capacidad fecundante de un eyacula-do (Flowers, 1997). Este movimiento apare-ce en diferentes condiciones físicas y varía según especies, pero básicamente consiste en un movimiento no lineal de la cabeza espermática, con aumento de la amplitud de batido de la cola y movimientos asimé-tricos (Yanagimachi, 1994), una marcada trayectoria en giro y baja progresión del movimiento (Figura 12).
La hiperactividad de los espermato-zoides ha sido observada in vitro en varias especies de mamíferos, sobre todo si se induce la capacitación de los espermatozoi-des con Ca2+ y HCO3
- (Yanagimachi, 1994). Para ello se han utilizado sistemas de aná-lisis de imagen computerizado (CASA) que
membrana espermática del espermatozoi-de (Calvete et al., 1997), reconociendo diferentes carbohidratos como manosa y galactosa (Ekhlasi-Hundriesen et al., 2005). La adición de AQN1 inhibe in vitro la unión del espermatozoide a las células epitelia-les de explantes oviductales de manera dosis dependiente (Ekhlasi-Hundriesen et al., 2005). Además, los sitios de unión están localizados en la región rostral de la cabeza del espermatozoide, perdiéndose duran-te la capacitación espermática (Ekhlasi-Hundriesen et al., 2005).
Mediante el análisis de las células de la superficie oviductal se ha deter-minado la interacción de la membrana periacrosomal del espermatozoide con una proteína presente en las células epi-teliales del istmo (Marini y Cabada 2003), la proteína de unión espermática (SBG). Está localizada en la cara apical, cerca del lumen oviductal (Pérez et al., 2006), exponiendo el disacárido o-unido Galβ1-3GalNac, el cual tiene capacidad de unión con la espermadesina AQN1 (Marini y Cabada 2003). SBG podría estar involu-crada en una selección espermática nega-tiva, ya que esta proteína solo se une a espermatozoides capacitados, alterando la integridad acrosomal y suprimiendo la motilidad (Teijeiro et al., 2008). Además produce fosforilación en tirosina de una proteína con peso molecular de 97 KDa, que podría estar involucrada en el meca-nismo de acción de la proteína (Marini y Cabada 2003). Las anexinas aisladas de las células del oviducto porcino también presentan afinidad por proteínas de la membrana periacrosomal. En particular, la anexina A2, localizada en la cara apical del epitelio oviductal puede estar involucrada en la formación del reservorio espermá-tico a través de la interacción con AQN1 (Teijeiro et al., 2009). Otras proteínas pre-sentes en la membrana apical del epitelio oviductal, como HSPA8 tienen capacidad de unión a los glicolípidos sulfatados a la región rostral del espermatozoide, además de aumentar la capacidad de superviven-cia del espermatozoide (Elliot et al., 2009).
Figura 11. Esquema de los diferentes tipos de movimiento del espermatozoide según su localización: a) epidídimo, b) eyaculado y c) en el oviducto (hiperactivado).
a b c Figura 12. Representación esquemática del movimiento de los espermatozoides capacitados y no capacitados.
Parámetros de movilidad regulares
Alta frecuenciaBaja amplitud
Baja frecuenciaGran amplitud
Parámetros de movilidad post-capacitación

45
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
permiten evaluar tanto el tipo de movi-miento como la velocidad de las células (Figura 13).
Presumiblemente, el inicio de la hipe-ractivación se debe a una señal que se pro-duce en el oviducto en un momento exacto, pero todavía no ha sido identificada.
Fosforilación en tirosina de las proteínas espermáticas
La fosforilación de las proteínas es un evento implicado en la regulación de una gran variedad de funciones celulares como el crecimiento, el control del ciclo celular, cambios en el citoesqueleto, exocitosis, etc. (Pawson, 2004). La adición y/o elimina-ción de los grupos fosfato a aminoácidos de distintas proteínas es un mecanismo que tiene como resultado la producción de cambios conformacionales de dichas proteínas, conduciendo a su activación o inactivación. La fosforilación puede ocurrir en los residuos de serina, treonina, y tiro-sina de las proteínas. Sin embargo, la más ampliamente descrita y estudiada ha sido la fosforilación en tirosina, ya que puede ser el primer indicador de las vías de seña-lización de transducción en las células (Naz y Rajesh, 2004).
En espermatozoides maduros se pro-ducen fenómenos de fosforilación/desfos-forilación de proteínas en la regulación de fenómenos importantes, como la capa-citación espermática, la hipermotilidad y la reacción acrosómica que van a ser indispensables para que el espermatozoi-de pueda alcanzar y penetrar al ovocito (Galantino- Homer et al., 1997).
En varias especies de mamíferos la fosforilación en residuos en tirosina ha sido correlacionada con el estado de capa-citación del espermatozoide. (Visconti et al., 1995; 1999; Tadiff et al., 2001; Pommer et al., 2003; Grasa et al., 2006). Dichos autores observaron un incremento tiem-po-dependiente en la fosforilación de la tirosina de varias proteínas específicas en
rangos moleculares de 40-120 kDa, con el estado de capacitación de los espermato-zoides. Evidenciaron que la fosforilación de la tirosina es un evento importante en la modulación de los cambios bioquímicos asociados a la capacitación.
Se han descrito numerosas proteínas en las que se produce la fosforilación en tirosina en las diferentes especies animales. En la cerda se ha descrito la fosforilación de la tirosina de 2 sets de proteínas de la membrana plasmática, las mayores (27 kDa, 37 kDa y 40 kDa) y las menores (34 kDa, 47 kDa y 55 kDa) (Flesch et al, 1999) (Figura 14).
La fosforilación en tirosina de las pro-teínas espermáticas ha sido localizada principalmente a nivel del flagelo, estando relacionada la fosforilación de la pieza principal con la adquisición de la hiper-motilidad (Nassar et al., 1999; Petrunkina et al., 2003). Además, la localización de determinadas proteínas fosforiladas en el área acrosomal pueden estar relacionadas con la interacción con la zona pelúcida y la penetración (Dube et al., 2005). También se ha descrito la aparición de un área triangular dentro del segmento ecuatorial en la unión con la zona postacrosomal, el subsegmento ecuatorial (Figura 15). Éste, presenta proteínas fosforiladas en residuos en tirosina, las cuales podrían jugar un
papel clave como centro organizador de ensamblaje de complejos multi-molecula-res mediando en la fusión de gametos y la reacción acrosómica (Jones et al., 2008).
Modulación de la capacitación espermática en el oviducto
Tras la ascensión por el tracto genital feme-nino, los gametos van a quedar retenidos en el reservorio espermático donde completarán el periodo de capacitación (Figura 2). Como es un proceso de desestabilización gradual controlado, que reduce la vida fértil del espermatozoide (Harrison, 1996), el manteni-miento de la viabilidad durante cierto tiempo y el control de la capacitación son pro-cesos que están íntimamente relacionados (Töper-Petersen 1999; Hunter y Rodríguez-Marínez, 2004). Se produce una sincroni-zación entre el momento de ovulación y la adquisición de la capacidad fecundante por parte del espermatozoide, gracias a la unión con las células oviductales en el reser-vorio espermático (Dobrinski et al., 1996b; Petrunkina et al., 2007) (Figura 16).
El estado de capacitación se plan-tea como uno de los principales criterios para la unión entre las células oviducta-les y los espermatozoides, representando un mecanismo de selección de la pobla-ción espermática más competente para la fecundación (Fazeli et al., 1999). También se determinan otros aspectos relacionados con el estado o morfología del espermato-zoide. Existe una correlación negativa entre el porcentaje de espermatozoides unidos y la presencia de alteraciones morfológi-cas o gotas citoplasmáticas (Petrunkina et al., 2001). Es necesario cierto estado de maduración o integridad morfológica para que se produzca la interacción. Además los espermatozoides unidos presentan bajas concentraciones de calcio (Petrunkina et al., 2001), lo que se correlaciona con estadíos de baja capacitación. Se produce una supresión de la capacitación hasta el momento de la ovulación (Rodríguez-Martínez et al., 2001).
Figura 13. Sistema de análisis computerizado de la motilidad espermática. a) microscopio de contraste de fases con placa térmica adaptado a un ordenador, b) imagen del software empleado para la valoración del movimiento espermático mediante análisis de imagen (CASA).
a b
Figura 14. Imagen 1: espermatozoides lavados con proteínas fosforiladas en tirosina evaluados mediante inmunofluorescencia indirecta (1a microscopia de contraste de fases, 1b microscopia de fluorescencia). En la imagen 2 se observan diferencias en las proteínas fosforiladas en tirosina de los espermatozoides lavados (L) y eyaculados (E) sin tratar evaluados mediante western blot.
1a 1b 2
46
Luño Lázaro, Victoria y Matas Parra, Carmen
Existe una regulación hormonal gona-dal que produce los diferentes cambios en el tracto genital (Hunter, 2008), permitien-do a los espermatozoides viables:
• Unirse al epitelio oviductal en el reser-vorio espermático durante la fase preovulatoria.
• Mantener la viabilidad y un estado de baja capacitación.
• Activarse y liberarse durante la fase periovulatoria y postovulatoria.
Este control endocrino permite que un número pequeño de espermatozoides sean liberados del reservorio espermático funcional sincrónicamente con el momento de la ovulación, permitiendo un bajo ratio espermatozoide:ovocito. (Hunter, 1994).
Bajo condiciones fisiológicas, se pro-duce una transición de la dominancia hormonal del estradiol, asociada con la maduración folicular, a la secreción de progesterona procedente de los folículos preovulatorios, la cual será determinante
en la dinámica oviductal. La secreción de progesterona, junto con prostaglandinas y diferentes péptidos llega al oviducto en grandes concentraciones y de manera rápida mediante el flujo contracorriente (Hunter, 1984). Estos pueden tener efecto en la modulación de la composición del fluido oviductal, así como en la degrada-ción de los sitios de unión a las células oviductales. Además, producen una remo-delación estructural de la membrana plas-mática espermática debido a la producción de un gradiente de calcio en los momentos periovulatorios (Flesch y Gadella, 2000), que estaría relacionada junto con la ini-ciación de la capacitación a la liberación al lumen oviductal de los espermatozoides.
También existe cierta regulación autó-noma por parte del espermatozoide. El con-tacto entre el gameto y las células oviducta-les establece un diálogo mutuo, que permite controlar los flujos de calcio (Dobrinski et al., 1996b), manteniendo viable la población y en un estadío bajo de capacitación, a la vez que se modifica el espectro de proteínas secretadas por las células epiteliales del ovi-ducto (Thomas et al., 1995).
Cultivo de células oviductales
Mediante cultivos in vitro de células ovi-ductales (OEC) (Figura 17) se ha intentado simular las condiciones que los esperma-tozoides encuentran en el tracto genital femenino, y así poder estudiar los cambios que allí acontecen. Estos cultivos se han realizado tanto para el estudio de las inte-racciones espermatozoides-oviducto como para recrear las condiciones apropiadas para una correcta fecundación, aproxi-mando el proceso de fecundación a las condiciones in vivo.
Cocultivos de células oviductales y espermatozoides
La interacción entre espermatozoide-célula oviductal permite que se manten-ga la viabilidad, motilidad y fertilidad de los espermatozoides (Suarez et al., 1991, Yeste et al., 2009). Cuando las pobla-ciones de espermatozoides unidos y no unidos son analizadas tras 24 horas de cocultivo, solo un 10% de los que per-manecieron en contacto con las células oviductales presentan alteraciones en la motilidad o viabilidad, mientras que esta cifra se torna entre el 50 y 80 % en los que permanecieron no unidos (Yeste et al., 2009). Además, dicha unión estabi-liza la membrana acrosomal, aumenta la supervivencia y permite mantener un nivel bajo de capacitación (Petrunkina et al., 2001).
El estado de capacitación se plantea como uno de los principales criterios para la unión entre las células oviducta-les y los espermatozoides, representando un mecanismo de selección de la pobla-ción espermática más competente para la fecundación (Fazeli et al., 1999). Se ha determinado que un mayor porcentaje de espermatozoides unidos presentan un nivel bajo de fosforilación en tirosina y de concentración de calcio intracelular en diferentes especies (Petrunkina et al., 2001; 2003b; Zumoffen et al., 2010). La unión permite mantener un esta-do de baja fosforilación en tirosina durante periodos de incubación largos
Figura 15. Distribución de proteínas fosforiladas en espermatozoides porcinos. (A) Sin fosforilar o fosforilación leve, (B) Fosforilación en acrosoma y/o cola, (C) Fosforilación en segmento ecuatorial, (D) Fosforilación en segmento ecuatorial, acrosoma y cola. Evaluación mediante inmunofluorescencia indirecta con microscopia de fluorescencia.
Figura 17. Células oviductales porcinas a los 7 días de cultivo, procesadas mediante inmunofluorescencia indirecta con anticuerpo antifosfotirosina. Las muestras fueron evaluadas con microscopio de contraste de fases bajo luz de fluorescencia (A) y campo claro (B) (100x).
Figura 16. Imagen de espermatozoides adheridos a las células del oviducto.

47
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
· Aumenta los beneficios en cerdos de engorde*.
· Mejora los parámetros reproductivos en cerdas**.
· La vacunación frente a PRRS de cerdas y lechones multiplica el beneficio total de la granja.
R e p o s i c i ó n L e c h o n e sR e p r o d u c t o r a s
* M
arte
lli 1
7º IP
VS
Con
gres
s 20
02 *
* A
lexo
pou
lus
Vet
Mic
200
5
Consulte con nuestro Departamento Técnico para un programa personalizado
A4 PUBLICIDAD 2 PORCILIS PRRS.indd 1 13/03/12 12:19

48
Luño Lázaro, Victoria y Matas Parra, Carmen
(Figura 18), permitiendo prolongar la viabilidad espermática y disminuir el estado de capacitación para sincronizar la liberación con el momento exacto de la ovulación.
❱❱ Bibliografía1. ABE H. THE MAMMALIAN OVIDUCTAL EPITHELIUM:
REGIONAL VARIATIONS IN CYTOLOGICAL AND FUNC-TIONAL ASPECTS OF THE OVIDUCTAL SECRETORY CELLS. HISTOL HISTOPATHOL 1996; 11: 743-768.
2. AMSO NN, CROW J, SHAW RW. COMPARATIVE IMMUNOHISTOCHEMICAL STUDY OF OESTROGEN AND PROGESTERONE RECEPTORS IN THE FALLOPIAN TUBE AND UTERUS AT DIFFERENT STAGES OF THE MENSTRUAL CYCLE AND THE MENOPAUSE. HUM REPROD 1994; 9: 1027-1037.
3. ARVER S, ELIASSON R. ZINC AND MAGNESIUM IN BULL AND BOAR SPERMATOZOA. J REPROD FERTIL 1980; 60: 481-484.
4. AUSTIN CR. OBSERVATIONS ON THE PENETRATION OF THE SPERM IN THE MAMMALIAN EGG. AUST J SCI RES B 1951; 4: 581-596.
5. BISCHOF RJ, LEE CS, BRANDON MR, MEEUSEN E. INFLAMMATORY RESPONSE IN THE PIG UTERUS INDUCED BY SEMINAL PLASMA. J REPROD IMMUNOL 1994; 26: 131-146.
6. CALVETE JJ, ENSSLIN M, MBURU J, IBORRA A, MARTíNEZ P, ADERMANN K, WABERSKI D, SANZ L, TöPFER-PETERSEN E, WEITZE KF, EINARSSON S, RODRíGUEZ-MARTíNEZ H. MONOCLONAL ANTI-BODIES AGAINST BOAR SPERM ZONA PELLUCIDA-BINDING PROTEIN AWN-1.CHARACTERIZATION OF A CONTINUOUS ANTIGENIC DETERMINANT AND IMMUNOLOCALIZATION OF AWN EPITOPES IN INSEMINATED SOWS. BIOL REPROD 1997; 57: 735-742.
7. CARDOZO JA, FERNÁNDEZ-JUAN M, FORCADA F, ABECIA A, MUIñO-BLANCO T, CEBRIÁN-PÉREZ JA. MONTHLY VARIATIONS IN OVINE SEMINAL PLASMA PROTEINS ANALYZED BY TWO-DIMEN-SIONAL POLYACRYLAMIDE GEL ELECTROPHORESIS. THERIOGENOLOGY 2006; 66: 841-850.
8. CHANG MC. FERTILIZING CAPACITY OF SPERMATO-ZOA DEPOSITED INTO THE FALLOPIAN TUBES. NATURE 1951; 168: 697-698.
9. COLENBRANDER B, KEMP B. FACTORS INFLUENCING SEMEN QUALITY IN PIGS. J REPROD FERTIL SUPPL 1990; 40: 105-115.
10. DOBRINSKI I, IGNOTZ GG, THOMAS PG, BALL BA. ROLE OF CARBOHYDRATES IN THE ATTACHMENT OF EQUINE SPERMATOZOA TO UTERINE TUBAL (OVIDUC-TAL) EPITHELIAL CELLS IN VITRO. AM J VET RES 1996A; 57: 1635-1639.
11. DOBRINSKI I, SUAREZ SS, BALL BA. INTRACELLULAR CALCIUM CONCENTRATION IN EQUINE SPERMATO-ZOA ATTACHED TO OVIDUCTAL EPITHELIAL CELLS IN VITRO. BIOL REPROD 1996B; 54: 783-788.
12. DUBÉ C, LECLERC P, BABA T, REYES-MORENO C, BAILEY JL. THE PROACROSIN BINDING PROTEIN, SP32, IS TYROSINE PHOSPHORYLATED DURING CAPACITATION OF PIG SPERM. J ANDROL 2005; 26: 519-528.
13. EKHLASI-HUNDRIESER M, GOHR K, WAGNER A, TSOLOVA M, PETRUNKINA A, TöPFER-PETERSEN E. SPERMADHESIN AQN1 IS A CANDIDATE RECEPTOR MOLECULE INVOLVED IN THE FORMATION OF THE OVIDUCTAL SPERM RESERVOIR IN THE PIG. BIOL REPROD 2005; 73: 536-545.
14. ELLINGTON JE, EVENSON DP, FLEMING JE, BRISBOIS RS, HISS GA, BRODER SJ, WRIGHT RW. COCULTURE OF HUMAN SPERM WITH BOVINE OVIDUCT EPITHE-LIAL CELLS DECREASES SPERM CHROMATIN STRUC-TURAL CHANGES SEEN DURING CULTURE IN MEDIA ALONE. FERTIL STERIL 1998; 69: 643-649.
15. ELLIOTT RM, LLOYD RE, FAZELI A, SOSTARIC E, GEORGIOU AS, SATAKE N, WATSON PF, HOLT WV. EFFECTS OF HSPA8, AN EVOLUTIONARILY CONSERVED OVIDUCTAL PROTEIN, ON BOAR AND BULL SPERMA-TOZOA. REPRODUCTION 2009; 137: 191-203.
16. FAWCETT DW. A COMPARATIVE VIEW OF SPERM ULTRASTRUCTURE. BIOL REPROD 1970; 2: SUPPL 2:90-127.
17. FAZELI A, DUNCAN AE, WATSON PF, HOLT WV. SPERM-OVIDUCT INTERACTION: INDUCTION OF CAPACITATION AND PREFERENTIAL BINDING OF UNCAPACITATED SPERMATOZOA TO OVIDUCTAL EPI-THELIAL CELLS IN PORCINE SPECIES. BIOL REPROD 1999; 60: 879-886.
18. FLESCH FM, COLENBRANDER B, VAN GOLDE LM, GADELLA BM. CAPACITATION INDUCES TYROSINE PHOSPHORYLATION OF PROTEINS IN THE BOAR SPERM PLASMA MEMBRANE. BIOCHEM BIOPHYS RES COMMUN. 1999; 262(3):787-92.
19. FLESCH FM, GADELLA BM. DYNAMICS OF THE MAM-MALIAN SPERM PLASMA MEMBRANE IN THE PRO-CESS OF FERTILIZATION. BIOCHIM BIOPHYS ACTA 2000; 1469: 197-235.
20. FLOWERS WL. MANAGEMENT OF BOARS FOR EFFI-CIENT SEMEN PRODUCTION. J REPROD FERTIL SUPPL 1997; 52: 67-78.
21. FREI B, STOCKER R, ENGLAND L, AMES BN. ASCORBATE: THE MOST EFFECTIVE ANTIOxIDANT IN
HUMAN BLOOD PLASMA. ADV ExP MED BIOL 1990; 264: 155-163.
22. GALANTINO-HOMER HL, VISCONTI PE, KOPF GS. REGULATION OF PROTEIN TYROSINE PHOSPHORY-LATION DURING BOVINE SPERM CAPACITATION BY A CYCLIC ADENOSINE 3’5’-MONOPHOSPHATE-DEPENDENT PATHWAY. BIOL REPROD 1997; 56: 707-719.
23. GANDOLFI F. FUNCTIONS OF PROTEINS SECRETED BY OVIDUCT EPITHELIAL CELLS. MICROSC RES TECH 1995; 32: 1-12.
24. GARNER DL, HAFEZ ESE (1996) ESPERMATOZOIDES Y PLASMA SEMINAL. EN: REPRODUCCIóN E INSEMI-NACIóN ARTIFICIAL EN ANIMALES. HAFEZ ESE (ED.) INTERAMERICANA MCGRAW-HILL (MÉxICO). PP 158-179.
25. GEISERT RD (1999) PIGS. EN: ENCYCLOPEDIA OF REPRODUCTION. KNOBIL E, NEILL JD (EDS.) ACADEMIC PRESS (SAN DIEGO, USA). PP 792-799.
26. GRASA P, CEBRIÁN-PÉREZ JA, MUIñO-BLANCO T. SIGNAL TRANSDUCTION MECHANISMS INVOLVED IN IN VITRO RAM SPERM CAPACITATION. REPRODUCTION 2006; 132: 721-732.
27. GUALTIERI R, TALEVI R. SELECTION OF HIGHLY FERTILIZATION-COMPETENT BOVINE SPERMATOZOA THROUGH ADHESION TO THE FALLOPIAN TUBE EPITHELIUM IN VITRO. REPRODUCTION 2003; 125: 251-258.
28. HARRISON RA. CAPACITATION MECHANISMS, AND THE ROLE OF CAPACITATION AS SEEN IN EUTHERIAN MAMMALS. REPROD FERTIL DEV 1996; 8: 581-594.
29. HOLT WV, HARRISON RA. BICARBONATE STIMULA-TION OF BOAR SPERM MOTILITY VIA A PROTEIN KINASE A-DEPENDENT PATHWAY: BETWEEN-CELL AND BETWEEN-EJACULATE DIFFERENCES ARE NOT DUE TO DEFICIENCIES IN PROTEIN KINASE A ACTI-VATION. J ANDROL 2002; 23: 557-565.
30. HOLT WV. IS SEMEN ANALYSIS USEFUL TO PREDICT THE ODDS THAT THE SPERM WILL MEET THE EGG? REPROD DOMEST ANIM 2009; 44 SUPPL 3: 31-38.
31. HUNTER RH. PRE-OVULATORY ARREST AND PERI-OVULATORY REDISTRIBUTION OF COMPETENT SPER-MATOZOA IN THE ISTHMUS OF THE PIG OVIDUCT. J REPROD FERTIL 1984; 72: 203-211.
32. HUNTER RH, FLÉCHON B, FLÉCHON JE. DISTRIBUTION, MORPHOLOGY AND EPITHELIAL INTERACTIONS OF BOVINE SPERMATOZOA IN THE OVIDUCT BEFORE AND AFTER OVULATION: A SCANNING ELECTRON MICROSCOPE STUDY. TISSUE CELL 1991; 23: 641-656.
33. HUNTER RH. OVARIAN REGULATION OF SPERM PROGRESSION IN THE FALLOPIAN TUBES. ZYGOTE 1994; 2: 363-366.
34. HUNTER RH, RODRIGUEZ-MARTINEZ H. CAPACITATION OF MAMMALIAN SPERMATOZOA IN VIVO, WITH A SPECIFIC FOCUS ON EVENTS IN THE FALLOPIAN TUBES. MOL REPROD DEV 2004; 67: 243-250.
35. HUNTER RH. THE FALLOPIAN TUBES IN DOMESTIC MAMMALS: HOW VITAL IS THEIR PHYSIOLOGICAL ACTIVITY? REPROD NUTR DEV 2005; 45: 281-290.
36. HUNTER RH. SPERM RELEASE FROM OVIDUCT EPI-THELIAL BINDING IS CONTROLLED HORMONALLY BY PERI-OVULATORY GRAAFIAN FOLLICLES. MOL REPROD DEV 2008; 75: 167-174.
37. JOHNSON LA, WEITZE KF, FISER P, MAxWELL WM. STORAGE OF BOAR SEMEN. ANIM REPROD SCI 2000; 62: 143-172.
38. JONES R, JAMES PS, OxLEY D, COADWELL J, SUZUKI-TOYOTA F, HOWES EA. THE EQUATORIAL SUBSEG-MENT IN MAMMALIAN SPERMATOZOA IS ENRICHED IN TYROSINE PHOSPHORYLATED PROTEINS. BIOL REPROD 2008; 79: 421-431.
39. KILLIAN GJ, CHAPMAN DA, ROGOWSKI LA. FERTILITY-ASSOCIATED PROTEINS IN HOLSTEIN BULL SEMINAL PLASMA. BIOL REPROD 1993; 49: 1202-1207.
40. KILLIAN GJ. EVIDENCE FOR THE ROLE OF OVIDUCT SECRETIONS IN SPERM FUNCTION, FERTILIZATION AND EMBRYO DEVELOPMENT. ANIM REPROD SCI 2004; 82-83: 141-153.
41. KORLEY R, POURESMAEILI F, OKO R. ANALYSIS OF THE PROTEIN COMPOSITION OF THE MOUSE SPERM PERINUCLEAR THECA AND CHARACTERIZATION OF ITS MAJOR PROTEIN CONSTITUENT. BIOL REPROD 1997; 57: 1426-1432.
42. LEESE HJ. THE FORMATION AND FUNCTION OF OVI-DUCT FLUID. J REPROD FERTIL 1988; 82: 843-856.
Figura 18. Espermatozoides unidos a las células oviductales in vitro. La imagen A, muestra una fosforilación en tirosina de proteínas en la región la ecuatorial y el flagelo. La ima-gen B presenta espermatozoides fosforilados en la región ecuatorial y sin fosforilar (*). Muestras evaluadas mediante inmunofluorescencia indirecta. Imagenes obtenidas con microscopio de contraste de fases bajo luz de fluorescencia y campo claro (100x).
A
B

49
nº 42Cambios en el espermatozoide desde la eyaculación hasta la fecundación
43. LEESE HJ, TAY JI, REISCHL J, DOWNING SJ. FORMATION OF FALLOPIAN TUBAL FLUID: ROLE OF A NEGLECTED EPITHELIUM. REPRODUCTION 2001; 121: 339-346.
44. LEFEBVRE R, SUAREZ SS. EFFECT OF CAPACITA-TION ON BULL SPERM BINDING TO HOMOLOGOUS OVIDUCTAL EPITHELIUM. BIOL REPROD 1996; 54: 575-582.
45. LESHIN LS, RAJ SM, SMITH CK, KWOK SC, KRAELING RR, LI WI. IMMUNOSTIMULATORY EFFECTS OF PIG SEMINAL PROTEINS ON PIG LYMPHOCYTES. J REPROD FERTIL 1998; 114: 77-84.
46. MARINI PE, CABADA MO. ONE STEP PURIFICA-TION AND BIOCHEMICAL CHARACTERIZATION OF A SPERMATOZOA-BINDING PROTEIN FROM PORCINE OVIDUCTAL EPITHELIAL CELLS. MOL REPROD DEV 2003; 66: 383-390.
47. MATTHIJS A, ENGEL B, WOELDERS H. NEUTROPHIL RECRUITMENT AND PHAGOCYTOSIS OF BOAR SPER-MATOZOA AFTER ARTIFICIAL INSEMINATION OF SOWS, AND THE EFFECTS OF INSEMINATE VOLUME, SPERM DOSE AND SPECIFIC ADDITIVES IN THE ExTENDER. REPRODUCTION 2003; 125: 357-367.
48. MATÁS C, COY P, ROMAR R, MARCO M, GADEA J, RUIZ S. EFFECT OF SPERM PREPARATION METHOD ON IN VITRO FERTILIZATION IN PIGS. REPRODUCTION 2003; 125: 133-141.
49. MATÁS C, SANSEGUNDO M, RUIZ S, GARCíA-VÁZQUEZ FA, GADEA J, ROMAR R, COY P. SPERM TREATMENT AFFECTS CAPACITATION PARAM-ETERS AND PENETRATION ABILITY OF EJACU-LATED AND EPIDIDYMAL BOAR SPERMATOZOA. THERIOGENOLOGY 2010; 74: 1327-1340.
50. MBURU JN, EINARSSON S, LUNDEHEIM N, RODRIGUEZ-MARTINEZ H. DISTRIBUTION, NUMBER AND MEMBRANE INTEGRITY OF SPERMATOZOA IN THE PIG OVIDUCT IN RELATION TO SPONTANEOUS OVULATION. ANIM REPROD SCI 1996; 45: 109-121.
51. MILLER D. SPERMATOZOAL RNA AS RESERVOIR, MARKER AND CARRIER OF EPIGENETIC INFOR-MATION: IMPLICATIONS FOR CLONING. REPROD DOMEST ANIM 2007; 42 SUPPL 2: 2-9.
52. NAZ RK, RAJESH PB. ROLE OF TYROSINE PHOS-PHORYLATION IN SPERM CAPACITATION / ACRO-SOME REACTION. REPROD BIOL ENDOCRINOL 2004; 2: 75.
53. NICHOL R, HUNTER RH, COOKE GM. OVIDUCT FLUID PH IN INTACT AND UNILATERALLY OVARIECTO-MIZED PIGS. CAN J PHYSIOL PHARMACOL 1997; 75: 1069-1074.
54. OLIVERA M, RUIZ T, TARAZONA A, GIRALDO C (2006) EL ESPERMATOZOIDE, DESDE LA EYACU-LACIóN HASTA LA FERTILIZACIóN. REV COL CIENC PEC. 19:426-436.
55. OKO R, MARAVEI D. PROTEIN COMPOSITION OF THE PERINUCLEAR THECA OF BULL SPERMATOZOA. BIOL REPROD 1994; 50: 1000-1014.
56. PAWSON T. SPECIFICITY IN SIGNAL TRANSDUCTION: FROM PHOSPHOTYROSINE-SH2 DOMAIN INTERAC-TIONS TO COMPLEx CELLULAR SYSTEMS. CELL 2004; 116: 191-203.
57. PESCH S, BERGMANN M. STRUCTURE OF MAM-MALIAN SPERMATOZOA IN RESPECT TO VIABILITY, FERTILITY AND CRYOPRESERVATION. MICRON 2006; 37: 597-612.
58. PETRUNKINA A, FRIEDRICH J, DROMMER W, BICKER G, WABERSKI D, TöPFER-PETERSEN E. KINETIC CHARACTERIZATION OF THE CHANGES IN PROTEIN TYROSINE PHOSPHORYLATION OF MEMBRANES, CYTOSOLIC CA2+ CONCENTRATION AND VIABIL-ITY IN BOAR SPERM POPULATIONS SELECTED BY BINDING TO OVIDUCTAL EPITHELIAL CELLS. REPRODUCTION 2001; 122: 469-480.
59. PETRUNKINA A, SIMON K, GüNZEL-APEL A, TöPFER-PETERSEN E. REGULATION OF CAPACITATION OF CANINE SPERMATOZOA DURING CO-CULTURE WITH HETEROLOGOUS OVIDUCTAL EPITHELIAL CELLS. REPROD DOMEST ANIM 2003A; 38: 455-463.
60. PETRUNKINA AM, SIMON K, GüNZEL-APEL AR, TöPFER-PETERSEN E. SPECIFIC ORDER IN THE APPEARANCE OF PROTEIN TYROSINE PHOSPHORY-LATION PATTERNS IS FUNCTIONALLY COORDINATED WITH DOG SPERM HYPERACTIVATION AND CAPACI-TATION. J ANDROL 2003B; 24: 423-437.
61. PETRUNKINA AM, WABERSKI D, GüNZEL-APEL AR, TöPFER-PETERSEN E. DETERMINANTS OF SPERM
QUALITY AND FERTILITY IN DOMESTIC SPECIES. REPRODUCTION 2007; 134: 3-17.
62. POMMER AC, RUTLLANT J, MEYERS SA. PHOSPHORYLATION OF PROTEIN TYROSINE RESI-DUES IN FRESH AND CRYOPRESERVED STALLION SPERMATOZOA UNDER CAPACITATING CONDI-TIONS. BIOL REPROD 2003; 68: 1208-1214.
63. PÉREZ FA, ROMA SM, CABADA MO, MARINI PE. SPERM BINDING GLYCOPROTEIN IS DIFFERENTIALLY PRESENT SURROUNDING THE LUMEN OF ISTH-MUS AND AMPULLA OF THE PIG’S OVIDUCT. ANAT EMBRYOL (BERL) 2006; 211: 619-624.
64. PÉREZ-PÉ R, BARRIOS B, MUIñO-BLANCO T, CEBRIÁN-PÉREZ JA. SEASONAL DIFFERENCES IN RAM SEMINAL PLASMA REVEALED BY PARTITION IN AN AQUEOUS TWO-PHASE SYSTEM. J CHROMATOGR B BIOMED SCI APPL 2001; 760: 113-121.
65. QUINTERO-MORENO A, RIGAU T, RODRíGUEZ-GIL JE. REGRESSION ANALYSES AND MOTILE SPERM SUBPOPULATION STRUCTURE STUDY AS IMPROV-ING TOOLS IN BOAR SEMEN QUALITY ANALYSIS. THERIOGENOLOGY 2004; 61: 673-690.
66. RODRIGUEZ-MARTINEZ H, TIENTHAI P, SUZUKI K, FUNAHASHI H, EKWALL H, JOHANNISSON A. INVOLVEMENT OF OVIDUCT IN SPERM CAPACITA-TION AND OOCYTE DEVELOPMENT IN PIGS. REPROD SUPPL 2001; 58: 129-145.
67. RODRíGUEZ-MARTíNEZ H, SARAVIA F, WALLGREN M, TIENTHAI P, JOHANNISSON A, VÁZQUEZ JM, MARTíNEZ E, ROCA J, SANZ L, CALVETE JJ. BOAR SPERMATOZOA IN THE OVIDUCT. THERIOGENOLOGY 2005; 63: 514-535.64.
68. RODRíGUEZ-MARTíNEZ H, KVIST U, SARAVIA F, WALLGREN M, JOHANNISSON A, SANZ L, PEñA FJ, MARTíNEZ EA, ROCA J, VÁZQUEZ JM, CALVETE JJ. THE PHYSIOLOGICAL ROLES OF THE BOAR EJACU-LATE. SOC REPROD FERTIL SUPPL 2009; 66: 1-21.
69. SATAKE N, ELLIOTT RM, WATSON PF, HOLT WV. SPERM SELECTION AND COMPETITION IN PIGS MAY BE MEDIATED BY THE DIFFERENTIAL MOTIL-ITY ACTIVATION AND SUPPRESSION OF SPERM SUBPOPULATIONS WITHIN THE OVIDUCT. J ExP BIOL 2006; 209: 1560-1572.
70. SENGER PL. PATHWAYS TO PREGNANCY AND PARTURITION.1ST EDITION.CURRENT CONCEPTIONS, INC, 1999, 1997. WASHINGTON STATE UNIVERSITYRESEARCH & TECHNOLOGY PARK. 1615NE EASTGATE BLVD PULLMAN, WA 99163-5607.
71. SMOLLICH A, MICHEL G. MICROSCOPICAL ANAT-OMY OF DOMESTIC ANIMALS. GUSTAV FISCHER, (STUTTGART) 1992. PP 245.
72. STAUSS CR, VOTTA TJ, SUAREZ SS. SPERM MOTIL-ITY HYPERACTIVATION FACILITATES PENETRATION OF THE HAMSTER ZONA PELLUCIDA. BIOL REPROD 1995; 53: 1280-1285.
73. STRZEZEK J, CIERESZKO A. HETEROGENEITY OF ASPARTATE AMINOTRANSFERASE (AAT) IN BULL SEMEN. COMP BIOCHEM PHYSIOL B 1987; 86: 373-375.
74. STRZEZEK J. SECRETORY ACTIVITY OF BOAR SEMINAL VESICLE GLANDS. REPROD BIOL 2002; 2: 243-266.
75. SUAREZ SS, KATZ DF, OWEN DH, ANDREW JB, POWELL RL. EVIDENCE FOR THE FUNCTION OF HYPERACTIVATED MOTILITY IN SPERM. BIOL REPROD 1991; 44: 375-381.
76. SUAREZ SS, REVAH I, LO M, KöLLE S. BULL SPERM BINDING TO OVIDUCTAL EPITHELIUM IS MEDI-ATED BY A CA2+-DEPENDENT LECTIN ON SPERM THAT RECOGNIZES LEWIS-A TRISACCHARIDE. BIOL REPROD 1998; 59: 39-44.
77. SUAREZ SS. FORMATION OF A RESERVOIR OF SPERM IN THE OVIDUCT. REPROD DOMEST ANIM 2002; 37: 140-143.
78. SUTOVSKY P, NEUBER E, SCHATTEN G. UBIQUITIN-DEPENDENT SPERM QUALITY CONTROL MECHA-NISM RECOGNIZES SPERMATOZOA WITH DNA DEFECTS AS REVEALED BY DUAL UBIQUITIN-TUNEL ASSAY. MOL REPROD DEV 2002; 61: 406-413.
79. TARDIF S, DUBÉ C, CHEVALIER S, BAILEY JL. CAPACITATION IS ASSOCIATED WITH TYROSINE PHOSPHORYLATION AND TYROSINE KINASE-LIKE ACTIVITY OF PIG SPERM PROTEINS. BIOL REPROD 2001; 65: 784-792.
80. TEIJEIRO JM, CABADA MO, MARINI PE. SPERM BIND-ING GLYCOPROTEIN (SBG) PRODUCES CALCIUM AND BICARBONATE DEPENDENT ALTERATION OF
ACROSOME MORPHOLOGY AND PROTEIN TYRO-SINE PHOSPHORYLATION ON BOAR SPERM. J CELL BIOCHEM 2008; 103: 1413-1423.
81. TEIJEIRO JM, IGNOTZ GG, MARINI PE. ANNExIN A2 IS INVOLVED IN PIG (SUS SCROFA) SPERM-OVIDUCT INTERACTION. MOL REPROD DEV 2009; 76: 334-341.
82. THOMAS PG, IGNOTZ GG, BALL BA, BRINSKO SP, CURRIE WB. EFFECT OF COCULTURE WITH STALLION SPERMATOZOA ON DE NOVO PROTEIN SYNTHESIS AND SECRETION BY EQUINE OVIDUCT EPITHELIAL CELLS. AM J VET RES 1995; 56: 1657-1662.
83. THURSTON LM, WATSON PF, MILEHAM AJ, HOLT WV. MORPHOLOGICALLY DISTINCT SPERM SUBPOPULA-TIONS DEFINED BY FOURIER SHAPE DESCRIPTORS IN FRESH EJACULATES CORRELATE WITH VARIA-TION IN BOAR SEMEN QUALITY FOLLOWING CRYO-PRESERVATION. J ANDROL 2001; 22: 382-394.
84. THÉRIEN I, BLEAU G, MANJUNATH P. PHOSPHATIDYLCHOLINE-BINDING PROTEINS OF BOVINE SEMINAL PLASMA MODULATE CAPACITA-TION OF SPERMATOZOA BY HEPARIN. BIOL REPROD 1995; 52: 1372-1379.
85. TIENTHAI P, KJELLÉN L, PERTOFT H, SUZUKI K, RODRIGUEZ-MARTINEZ H. LOCALIZATION AND QUANTITATION OF HYALURONAN AND SUL-FATED GLYCOSAMINOGLYCANS IN THE TISSUES AND INTRALUMINAL FLUID OF THE PIG OVIDUCT. REPROD FERTIL DEV 2000; 12: 173-182.
86. TIENTHAI P, JOHANNISSON A, RODRIGUEZ-MARTINEZ H. SPERM CAPACITATION IN THE POR-CINE OVIDUCT. ANIM REPROD SCI 2004; 80: 131-146.
87. TöPFER-PETERSEN E. MOLECULES ON THE SPERM’S ROUTE TO FERTILIZATION. J ExP ZOOL 1999; 285: 259-266.
88. TöPFER-PETERSEN E, WAGNER A, FRIEDRICH J, PETRUNKINA A, EKHLASI-HUNDRIESER M, WABERSKI D, DROMMER W. FUNCTION OF THE MAMMALIAN OVIDUCTAL SPERM RESERVOIR. J ExP ZOOL 2002; 292: 210-215.
89. VISCONTI P, BAILEY J, MOORE G, PAN D, OLDS-CLARKE P, KOPF G. CAPACITATION OF MOUSE SPERMATOZOA. I. CORRELATION BETWEEN THE CAPACITATION STATE AND PROTEIN TYROSINE PHOSPHORYLATION. DEVELOPMENT 1995; 121: 1129-1137.
90. VISCONTI PE, STEWART-SAVAGE J, BLASCO A, BATTAGLIA L, MIRANDA P, KOPF GS, TEZóN JG. ROLES OF BICARBONATE, CAMP, AND PROTEIN TYROSINE PHOSPHORYLATION ON CAPACITATION AND THE SPONTANEOUS ACROSOME REACTION OF HAMSTER SPERM. BIOL REPROD 1999; 61: 76-84.
91. VISCONTI PE, WESTBROOK VA, CHERTIHIN O, DEMARCO I, SLEIGHT S, DIEKMAN AB. NOVEL SIG-NALING PATHWAYS INVOLVED IN SPERM ACQUI-SITION OF FERTILIZING CAPACITY. J REPROD IMMUNOL 2002; 53: 133-150.
92. VISCONTI PE. UNDERSTANDING THE MOLECU-LAR BASIS OF SPERM CAPACITATION THROUGH KINASE DESIGN. PROC NATL ACAD SCI U S A. 2009; 106(3):667-668.
93. WATSON PF. THE CAUSES OF REDUCED FERTILITY WITH CRYOPRESERVED SEMEN. ANIM REPROD SCI 2000; 60-61: 481-492.
94. YANAGIMACHI R. FERTILITY OF MAMMALIAN SPER-MATOZOA: ITS DEVELOPMENT AND RELATIVITY. ZYGOTE 1994; 2: 371-372.
95. YESTE M, LLOYD RE, BADIA E, BRIZ M, BONET S, HOLT WV. DIRECT CONTACT BETWEEN BOAR SPERMA-TOZOA AND PORCINE OVIDUCTAL EPITHELIAL CELL (OEC) CULTURES IS NEEDED FOR OPTIMAL SPERM SURVIVAL IN VITRO. ANIM REPROD SCI 2009; 113: 263-278.
96. ZHOU Cx, WANG xF, CHAN HC. BICARBONATE SECRETION BY THE FEMALE REPRODUCTIVE TRACT AND ITS IMPACT ON SPERM FERTILIZING CAPACITY. SHENG LI xUE BAO 2005; 57: 115-124.
97. ZHU J, xU x, COSGROVE JR, FOxEROFT GR. EFFECTS OF SEMEN PLASMA FROM DIFFERENT FRACTIONS OF INDIVIDUAL EJACULATES ON IVF IN PIGS. THERIOGENOLOGY 2000; 54: 1443-1452.
98. ZUMOFFEN CM, CAILLE AM, MUNUCE MJ, CABADA MO, GHERSEVICH SA. PROTEINS FROM HUMAN OVIDUCTAL TISSUE-CONDITIONED MEDIUM MODU-LATE SPERM CAPACITATION. HUM REPROD 2010; 25: 1504-1512.


















