
Caracterización ecológica y aprovechamiento de la Paloma...
234
Caracterización ecológica y aprovechamiento de la Paloma Rabiche (Zenaida macroura) en un agrosistema tropical (Cienfuegos, Cuba) Juan Manuel García Bacallao
Transcript of Caracterización ecológica y aprovechamiento de la Paloma...

Caracterización ecológica y aprovechamiento de la Paloma Rabiche (Zenaida macroura) en un agrosistema tropical (Cienfuegos, Cuba)
Juan Manuel García Bacallao

DEPARTAMENTO DE ECOLOGÍA
FACULTAD DE CIENCIAS
Caracterización ecológica y aprovechamiento de la Paloma Rabiche (Zenaida macroura) en un
agrosistema tropical (Cienfuegos, Cuba)
Autor:Juan Manuel García Bacallao
Tesis presentada para aspirar al grado de DOCTOR POR LA UNIVERSIDAD DE ALICANTE
PROGRAMA: CONSERVACIÓN Y RESTAURACIÓN DE ECOSISTEMAS.
Dirigida por:Dr. Victoriano Peiró Clavell


Pensamiento
El fallo en los procedimientos de aprovechamientos prematuros o mal concebidos
constituye buen ejemplo de la necesidad de un conveniente enfoque ecológico como
base para la adecuada conservación o explotación de los recursos biológicos.
George L. Clarke
.


Agradecimientos
Siempre me ha parecido una tarea muy difícil el resumir en unas pocas palabras la
gratitud que siento hacia todas las personas que de muy diferentes maneras me han
prestado su ayuda incondicional para la culminación de este trabajo y que no me es
posible nombrarlos a todos aquí.
Otra de las cuestiones que me genera sentimientos encontrados es el orden, en que
a pesar de todo, es necesario mencionar las personas que resultaron claves en la
realización de este trabajo, por lo que considero indispensable aclarar que el orden
de mención no determina preferencia o importancia.
A lo largo de todos los años que duró la realización del trabajo, una persona y luego
tres más, han tenido que realizar muchos sacrificios que acompañan este tipo de
labor, como dejar de hacer lo planificado, sacrificar su tiempo y horas libre, aguantar
mis historias para que recibiese todo el apoyo necesario: mi esposa Yamila y mis
tres hijos Daniela, Gabriela y Yoan Manuel; sin su apoyo nada de esto hubiese sido
igual.
Deseo hacer una mención especial a mi amigo y profesor Dr. Rubén de Jesús
Chamizo Pérez, quien me inició en esta investigación y confió en mi cuando apenas
nos conocíamos.
También dedico un reconocimiento especial a mi director de tesis Dr. Victoriano
Peiró Clavel por haber asumido una tarea tan difícil desde miles de kilómetros de
distancia y siempre estar dispuesto a dedicar horas a leer mis manuscritos y
ofrecerme comentarios y críticas basadas en sus saberes y propias experiencias, no
hubiese sido posible haber completado el trabajo sin la desinteresada ayuda de él.
No podría dejar de mencionar a dos personas que igualmente han sido claves para
mí:
Nina, muchas gracias por el apoyo y la amistad brindada.

Al Dr. Antonio Escarré Estebes que desde el comienzo de este trabajo se preocupó y
ocupó de encontrar para mí el asesor adecuado y siempre estar al tanto del
desarrollo de este.
A todos expreso mi más profunda gratitud por el tiempo y las ideas que
compartieron.
Muchas gracias.
.

Dedicatoria
La Paloma Rabiche es un símbolo de valor para las comunidades en las que
conviven o están cerca de ellas, son elementos primordiales en el paisaje del campo
cubano, así como en sus ciudades. Su valor es más evidente cuando es la comida
del campesino o cazador, pero además tiene una inmensa riqueza por sus restantes
funciones, tanto en los servicios ambientales como en el imaginario humano. Su
grado de conservación es un buen indicador del estilo de la sociedad, una sociedad
que usa pero protege su fauna es más desarrollada y consciente que aquella que
solo ve en esta el recurso que garantiza su beneficio sin preocuparse por su cuidado
y desarrollo.
Este trabajo está dedicado especialmente para los que desean contribuir al cuidado
y protección no solo de las especies cinegéticas, sino de todas las especies de
nuestra flora y por tanto buscan el conocimiento y las herramientas necesarias para
contribuir en este empeño. Pudiese ser también una opción para los amantes del
medioambiente y cazadores noveles, que no son especialistas en fauna, pero
sencillamente les importan la naturaleza, la aman y cuidan.
Juan Manuel García Bacallao.


Resumen
El conocimiento ecológico de la Paloma Rabiche (Zenaida macroura L.) en Cuba
resulta ser insuficiente, fragmentado y disperso, unido a las diferentes formas de
aprovechamiento a que es actualmente sometida la especie y la falta de control de
su gestión, traen aparejadas importantes dudas sobre los efectos que puedan tener
en sus poblaciones. El objetivo de la investigación fue caracterizar los aspectos
esenciales de la ecología y el aprovechamiento de la Paloma Rabiche (Zenaida
macroura L.) que pueden ser útiles para un manejo sostenible en un agrosistema
tropical. El estudio se realizó en el área de caza “Damují” del municipio de Rodas,
provincia de Cienfuegos, Cuba, cuyo centro se localiza en las coordenadas
geográficas 22° – 21.804' N y 080
° - 33.430' W, abarcando un total de 645,8 ha.
Como resultados fundamentales se obtuvo que el biotopo Agrícola con cobertura
presentó una mejor calidad para la especie estudiada, dado por el predominio de
especies herbáceas, las que constituyen la principal fuente de su alimentación y la
presencia de linderos y árboles aislados que ofrecen condiciones de sombra, refugio
y sustratos para la nidificación. La Paloma Rabiche presentó una distribución
agregada en el biotopo Agrícola con cobertura, no siendo igual en el biotopo Forestal
típico, determinado por la composición y estructura de la vegetación y por tanto la
disponibilidad de alimentos. El periodo reproductivo se extendió desde marzo hasta
agosto, pero una parte de la población se mantuvo en reproducción en el resto de
los meses del año, se detectaron un total de 31 nidos distribuidos en cinco hábitats
de nidificación y sobre 10 especies diferentes de plantas. La especie tuvo un rango
diversificado de sustratos vegetales para la nidificación, aunque se evidenció una
mayor preferencia por los árboles y arbustos espinosos. El éxito reproductor total fue
del 55 %. El análisis de la razón de edad mostró un desequilibrio a favor de los
machos, lo que asegura a través de la selección de estos una mejor calidad en la
población. La razón de edad demostró que la población no es reemplazada
totalmente lo cual debe tenerse en cuenta en su gestión. En el espectro alimentario
se identificaron 33 artículos, de los que el 88 % son de origen vegetal, el 9 %
alimentos de origen mineral y el 3 % de origen animal. Las semillas de Euphorbia

heterphylla y de Panicum maximun fueron los elementos de mayor importancia
alimentaria para la Paloma Rabiche. La base alimentaria estuvo compuesta en un 79
% por semillas de plantas herbáceas, teniendo preferencia por las semillas
pequeñas, de las que solo el 10 % fueron cultivos agrícolas. La Paloma Rabiche
mostró preferencia en su alimentación por áreas donde predominaron las especies
herbáceas con un porciento de suelo desnudo, los cultivos agrícolas ofrecieron la
mejor calidad de sitios, por encima de áreas ganaderas o bosques naturales y
plantaciones forestales. La caracterización morfométrica de la especie difiere
estadísticamente de las descritas en los estudios realizados en el país y para los
E.U.A. Se encontró en la población dos tipos de ectoparásitos, la Mosca de Gavilán
(Lynchia americana) que se hospeda en el plumaje de la especie y mostró una alta
temporalidad, siendo la época lluviosa la de mayor incidencia en la población y un
ácaro desconocido que parasita las patas de la paloma, siendo este último la
especie de más frecuente parasitación, incidiendo en la mayoría de los meses del
año. La Paloma Rabiche es la especie cinegética más importante en el país, por
encima de las anátidas y demás especies del orden Columbiformes, la forma de
aprovechamiento que predominó sobre la Paloma Rabiche fue la caza ilegal (69 %),
siendo el grupo etario más representado en esta los niños y adolescentes (73 %). De
todas las modalidades de caza ilegal encontradas la de mayor incidencia fue la caza
con fusil neumático, seguida por la caza con tirapiedras. Se concluye que el
conocimiento aportado por la presente investigación ofrece los elementos necesarios
para la protección y manejo de la especie en el área de estudio y, en general, en el
país desde una perspectiva que pondere el manejo sostenible de esta paloma.
.

Índice
CAPITULO I - INTRODUCCION GENERAL ......................................................................... 1
1.1.- Evolución e importancia de la actividad cinegética ..................................... 1
1.2.- Descripción de la Paloma Rabiche (Zenaida macroura L.).......................... 3
1.3.- Importancia de la Paloma Rabiche (Zenaida macroura L.) como especie
cinegética en Cuba .................................................................................................. 8
1.4.- Planteamiento del problema y Objetivos de la tesis .................................. 12
1.5.- Contenido de los capítulos ........................................................................... 14
1.6.- Bibliografía ..................................................................................................... 18
CAPÍTULO II – CARACTERISTICAS DEL AREA DE ESTUDIO.................................... 21
2.1.- Ubicación geográfica del área de estudio ................................................... 21
2.2.- Clima............................................................................................................... 22
2.3.- Hidrología ....................................................................................................... 23
2.4.- Relieve y suelos ............................................................................................ 24
2.5.- Usos del suelo ............................................................................................... 26
2.6.- Bibliografía ..................................................................................................... 28
CAPÍTULO III – CARACTERIZACION DE LA VEGETACION COMO BIOTIPO DE
LA PALOMA RABICHE .......................................................................................................... 29
3.1.- Introducción ................................................................................................... 29
3.2.- Materiales y métodos .................................................................................... 30
3.3.- Resultados y discusión ................................................................................ 33
3.4.- Conclusiones y recomendaciones .............................................................. 52
3.5.- Bibliografía ..................................................................................................... 53
CAPÍTULO IV. LA PALOMA RABICHE EN LA COMUNIDAD ORNITÍCA ................... 56
4.1.- Introducción .................................................................................................. 56
4.2.- Materiales y métodos .................................................................................... 57
4.3.- Resultados y discusión ................................................................................ 58
4.4.- Conclusiones y recomendaciones ............................................................. 64
4.4.- Bibliografía ..................................................................................................... 65
CAPÍTULO V. REPRODUCCIÓN Y CONDUCTA REPRODUCTIVA DE LA
PALOMA RABICHE (Zenaida macroura L.) ..................................................................... 67
5.1.- Introducción ................................................................................................... 67

5.2.- Materiales y métodos ................................................................................... 69
5.3.- Resultados y discusión ............................................................................... 71
5.3.1-. Estación reproductora ..................................................................................... 71
5.3.2.- Hábitats de nidificación .................................................................................. 77
5.3.3.- Éxito reproductor ............................................................................................. 80
5.3.4.- Percepción rural de la estación reproductora ......................................... 84
5.3.5.- Aspectos conductuales relevantes de la reproducción ........................ 85
5.4.- Conclusiones y recomendaciones .............................................................. 88
5.4.- Bibliografía ..................................................................................................... 89
Smith D. M., D. M. Finch y D. L. Hawksworth (2012). Nesting characteristics of mourning doves in central New Mexico: Response to riparian forest change. The Journal of Wildlife Management, 76 (2), 382 – 390. .......................................................... 90
CAPÍTULO VI. ESTRUCTURA DE LA POBLACIÓN DE PALOMA RABICHE
(Zenaida macroura L.) ........................................................................................................... 92
6.1.- Introducción .................................................................................................. 92
6.2.- Materiales y métodos ................................................................................... 94
6.3.- Resultados y discusión ............................................................................... 97
6.4.- Conclusiones y recomendaciones ............................................................. 99
6.5.- Bibliografía .................................................................................................. 100
CAPÍTULO VII. ECOLOGÍA TRÓFICA DE LA PALOMA RABICHE (Zenaida
macroura L.) .......................................................................................................................... 102
7.1.- Introducción ................................................................................................ 102
7.2.- Materiales y métodos ................................................................................. 103
7.3.- Resultados y discusión ............................................................................. 105
7.4.- Conclusiones y recomendaciones .......................................................... 116
7.5.- Bibliografía ................................................................................................... 117
CAPÍTULO VIII. CALIDAD DEL HÁBITAT DE LA PALOMA RABICHE (Zenaida
macroura L.) ........................................................................................................................... 121
8.1.- Introducción ................................................................................................. 121
8.2.- Materiales y métodos ................................................................................. 123
8.3.- Resultados y discusión ............................................................................. 124
8.4.- Conclusiones y recomendaciones ........................................................... 129
8.5.- Bibliografía .................................................................................................. 130
CAPÍTULO IX. MORFOMETRÍA DE LA POBLACIÓN DE PALOMA RABICHE
(Zenaida macroura L.) ......................................................................................................... 132

9.1.- Introducción ................................................................................................ 132
9.2.- Materiales y métodos ................................................................................. 134
9.3.- Resultados y discusión ............................................................................. 137
9.3.1.- Caracterización morfométrica y relación con otras áreas ................... 137
9.3.2.- Parámetros morfométricos como indicadores del estado de salud
de la población ............................................................................................................. 143
9.4.- Conclusiones y recomendaciones ........................................................... 147
9.5.- Bibliografía ................................................................................................... 147
CAPÍTULO X. ECTOPARÁSITOS EN LA POBLACIÓN DE PALOMA RABICHE
(Zenaida macroura L.) ......................................................................................................... 151
10.1.- Introducción .............................................................................................. 151
10.2.- Materiales y métodos ................................................................................ 153
10.3.- Resultados y discusión ............................................................................ 154
10.4.- Conclusiones y recomendaciones ......................................................... 170
10.5.- Bibliografía ................................................................................................. 171
CAPÍTULO X I. APROVECHAMIENTO CINEGÉTICO DE LA PALOMA RABICHE
(Zenaida macroura L.) ......................................................................................................... 173
11.1.- Introducción .............................................................................................. 173
11.2.- Materiales y métodos ............................................................................... 174
11.3.- Resultados y discusión ........................................................................... 176
11.3.1.- Formas tradicionales de aprovechamiento de la especie ................. 176
11.3.2.- Importancia de la Paloma Rabiche en la comunidad de especies
cinegéticas cubanas .................................................................................................... 181
11.3.3.- Las especies depredadoras de la Paloma Rabiche .............................. 183
11.3.4.- Propuesta de criterios e indicadores de gestión sostenible ............. 185
11.4.- Conclusiones y recomendaciones ......................................................... 190
11.5.- Bibliografía ................................................................................................ 190
Anexos


1
CAPITULO I - INTRODUCCION GENERAL
1.1.- Evolución e importancia de la actividad cinegética
La fauna silvestre está constituida por el conjunto de animales que viven en una
zona determinada, sus poblaciones son el producto del proceso evolutivo milenario,
diferente en cada región del planeta, lo cual les confiere características únicas y por
tanto valor único y derecho a existir. Por esta razón las especies propias de cada
región constituyen la riqueza y diversidad genética de esos ecosistemas, por lo que
se consideran parte del patrimonio natural de cada país. En la actualidad muchas
personas y organizaciones mundiales reconocen el alto valor intrínseco de la fauna
silvestre dado por sus usos, tanto consuntivos como los no consuntivos. Estos
valores históricamente la sociedad los ha manejado en base al aporte o utilidad que
brinda al hombre, es decir su valor tangible o económico. Uno de los principales usos
que la humanidad ha dado a la fauna silvestre es la caza, actividad que desde
mucho tiempo atrás es considerada en muchas sociedades y estados como
importante renglón económico.
La prosperidad de toda nación depende, en gran medida, del estado y
aprovechamiento de recursos naturales como los bosques, las aguas, los minerales
y la fauna. Los recursos naturales como las aguas, los suelos, la flora y la fauna
constituyen recursos renovables porque si se manejan de manera racional, y son
debidamente protegidos se puede disfrutar permanentemente de sus beneficios. La
forma más elemental en que se ha aprovechado históricamente la fauna silvestre
desde los primeros tiempos de la humanidad ha sido su uso directo: la caza
(Martínez, (1981).
Aramburu y Pons, (1956), destacan la importancia histórica de la caza a través de la
evolución de la humanidad. Esta actividad constituyó una de las primeras realizadas
por el hombre sobre la tierra. La necesidad de sustento en el hombre primitivo como
vía para satisfacer su instinto de conservación y perpetuación de la especie, obligó a
ese ser primitivo a lanzarse sobre las huellas de animales salvajes; y con la primera
captura tuvo lugar el comienzo de la milenaria historia de la caza.

2
La caza ha pasado por diferentes etapas a lo largo de los periodos prehistórico e
histórico. A continuación se presentan de forma resumida los aspectos más
sobresalientes que marcan la evolución del concepto de actividad cinegética: en la
Era Romana, la caza de animales tiene lugar como entretenimiento y entrenamiento
para la guerra. El derecho romano establece el concepto o condición de “Res
Nullíus“ o “Cosa de Nadie” para las especies de caza, pero los romanos también
fueron los primeros en establecer reservas de caza. Durante la Edad Media se
establece el carácter feudal y nobiliario de la caza y se afianza el derecho de
declarar reservas de caza. En el siglo XIV aparecen las armas de fuego y se inicia su
empleo en las actividades venatorias. En el Renacimiento, la caza es todavía una
actividad aristocrática reservada a las clases dominantes. Después de la Revolución
Francesa, a partir de finales del siglo XVIII, el derecho a la caza comenzó a
extenderse paulatinamente a todos los ciudadanos. En cualquier caso, durante estos
siglos, la actividad cinegética ha seguido siendo, legal o ilegalmente una fuente de
aporte alimentario en las economías rurales más desfavorecidas (Peiró, 2003).
En Cuba, la caza forma parte importante de la actividad del hombre en el espacio
rural, y tiene como finalidad principal la obtención de carne para el consumo, lo que
deja en segundo plano los aspectos lúdicos y espirituales (Chamizo, 2004), cosa por
la cual se ve amenazada la sostenibilidad de los aprovechamientos cinegéticos
actuales.
La actividad cinegética está regida por fuertes principios éticos, requiere una estricta
disciplina, y puede convertirse en un verdadero desastre si no obedece a una
planificación y una organización adecuada. Aunque económicamente sea viable, no
es posible una explotación sostenible de los recursos cinegéticos sin la evaluación y
el manejo consecuente de sus impactos ambientales y, particularmente, sin el
control de los efectos que ocasiona sobre la población de la especie aprovechada
(Chamizo, 2000).
En las condiciones actuales en que la actividad humana ha provocado grandes
cambios negativos en la naturaleza, es importante lograr que la caza no sólo excluya
efectos perniciosos, sino que se conciba de manera que contribuya al mejoramiento

3
de los territorios y al beneficio de las poblaciones de animales cinegéticos (Chamizo,
2000). En consecuencia, se hace necesario pasar a una nueva etapa en el
desarrollo cinegético en la que se tomen acciones sobre las deficiencias actuales y
se aprovechen las experiencias y resultados de largos años de trabajo.
1.2.- Descripción de la Paloma Rabiche (Zenaida macroura L.)
La clasificación taxonómica es la siguiente:
Clase: AVIS
Orden: Columbiformes
Familia: Columbidae.
Género: Zenaida.
Especie: macroura
Figura 1. Ejemplar adulto de Paloma Rabiche (Zenaida macroura L.).
Acosta y Ponce de León (2004) en su descripción de la Paloma Rabiche destacan
detalladamente su coloración como aspecto fundamental para la identificación de
esta especie. Refieren que presenta ligeramente colores grises en la parte superior
de la frente y la nuca, por detrás del cuello predomina el carmelita grisáceo con
plumas metálicas en violeta y bronce a ambos lados. Presenta una pequeña estría
detrás de los ojos. El lomo, las coberteras y las alas son de color carmelita grisáceo.
La rabadilla es grisácea, un poco más oscura en los flancos. La cola es larga,

4
escalonada y puntiaguda (las plumas más largas hacia el centro, las más cortas en
los lados y todas ampliamente bordeadas en blanco), de ahí deriva el nombre
específico (macroura) dado por Linneo. El ojo presenta un iris pardo oscuro y
rodeado por una piel orbital entre verdeazulosa y azul. El pico es negro, delgado y
con apariencia delicada. Patas de color rojo opaco. La hembra es más pálida, con
menos iridiscencia en el cuello. Los juveniles son parecidos a las hembras, pero
tienen un reborde carmelita claro en todas las plumas.
La paloma Rabiche (Zenaida macroura L.) es una de las tres especie del género
Zenaida presentes en Cuba (Garrido y Kirkconnell, 2011). Es muy abundante desde
el sur de Canadá y los Estados Unidos hasta el sur de México y las Antillas (Acosta y
Ponce de León, 2004).
Aldrich (1993), Mirarchi and Baskett, (1994), citados por Dolton (2003) plantean que
la especie se reproduce desde la parte sur de Canadá a través de los Estados
Unidos hasta México, las Bermudas, las Bahamas y Antillas Mayores además de
ubicaciones dispersas en América Central. Aluden que aunque algunas palomas
pasan el invierno en la mayor parte del área de reproducción, excepto para el centro
de Canadá y el norte-centro de EE.UU., la mayoría migra hacia el sur de Estados
Unidos, la mayor parte de México, América Central hasta el oeste de Panamá.
(Figura 2).

5
Figura 2. Rango de distribución de la Paloma Rabiche (mapa realizado por el
Laboratorio de Ornitología de la Universidad de Cornell).
Acosta y Ponce de León, (2004) refieren que en Cuba esta especie es frecuente en
potreros y terrenos llanos asociados a parches de bosques, campos cultivados,
zonas residenciales y parques urbanos. Su número se ha incrementado
grandemente en las últimas décadas, debido al creciente desarrollo de áreas para el
cultivo de granos, especialmente arroceras. Los mismos autores plantean que
estudios recientes estiman en unos 500 millones de individuos la población de esta
especie en los Estados Unidos, existiendo 5 subespecies o razas dentro de su rango
de distribución, de las cuales una (Z. m. macroura) es residente permanente en

6
Cuba y otra (Z. m. carolinensis) es visitante ocasional durante la época de migración,
mezclándose con las poblaciones cubanas.
La Paloma Rabiche no está amenazada globalmente y es quizás una de las mejor
manejadas, siendo común en todo su rango de distribución, donde sufre una enorme
presión de caza, a tal punto que solamente en los EUA son cazadas 45 millones de
piezas al año, pero la especie aparentemente resiste esa presión (Acosta y Ponce de
León, 2004). Según la Unión Internacional para la Conservación de la Naturaleza
(UICN), su estado de conservación actual es catalogado como Preocupación Menor
(LC).
La primera descripción que se conoce de la especie en Cuba la realizó el naturalista
alemán Dr. J. Gundlash en su obra “Contribución a la ornitología cubana” impresa en
la Habana en 1876, realizando la siguiente descripción:
Nombre vulgar: Raliche en la parte occidental de la Isla, Guamica en la oriental.
Según Pichardo Mabuda en Puerto Príncipe.
Descripción:
“Esta especie vive en los Estados Unidos, en algunas Antillas menos Puerto Rico
(según parece), y en el Continente meridional de la América. En Cuba es sedentaria,
pero no de paso como se lee en la obra de la Sagra. Prefiere las orillas de los
bosques, los árboles aislados en potreros y sabanas y las arboledas, etc., pero no
los bosques cerrados, y busca su comida en el suelo y en los campos recién arados,
Esta consiste en semillas, con preferencia las del Cardo-santo y del Frailecillo.
En tiempo de la cría anda en parejas ó sola, pero después se reúne en bandadas
chicas ó en familias. Por la tarde se dirige a las lagunas, ojos de agua u orillas de los
arroyos para beber, y después se traslada a sus dormitorios. Es útil al hombre por su
carne sabrosa, aunque a veces tiene un gusto algo amargo, probablemente a causa
de la clase de semillas que come. El daño que causa recogiendo la semilla recién
sembrada, no es de gran importancia, porque no la saca de la tierra, sino que come
solamente la que queda sobre ella, Tampoco quiebra las espigas del arroz o del
millo, ni daña las mazorcas de maíz como lo hacen el Totí, Chambergo, Mayito, etc.

7
Es muy arisca, sin duda por la constante persecución que sufre del hombre; pero se
cría fácilmente en jaulas, y si estas son bastante grandes, anida en ellas. Su canto
es suave, agradable y se parece a las sílabas «'tuíu-tu-tutu." Cuando está asustada
ó en peligro, produce una especie de murmullo. Su vuelo es en extremo rápido,
horizontal y con pocos aleteos, los cuales por la violencia y prontitud del vuelo,
producen un ligero silbido particular. A veces se la ve volar con las alas tendidas, sin
movimiento y la cola muy abierta, hacer evoluciones y descender a lugares donde
hay comida para su alimento. Desde el mes de Marzo hasta Agosto he observado
sus nidos hechos de algunas ramitas o yerbas secas, encima de una ramificación
horizontal, sobre el tronco de un árbol ya retoñado o encima de Curujeyes, y en ellos
2 huevos blancos de 0,026 + 0,919 milím.
No se puede equivocar esta especie con otra, porque la cola es larga, en forma de
cuña y acaba en punta, lo que indica ya los nombres triviales Rabiche y Rabua… El
joven difiere del adulto por los colores menos puros, por los ribetes de las plumas de
sus partes superiores, claros, porque es menos negra y menos pronunciada la
mancha que tiene debajo de la oreja, etc.
Las medidas del adulto son
♂ long. total 0,280; brazas 0,432; cola 0,120 milím.
♀long. total 0,265; brazas 0,420; cola 0,112 milím.”
También fue encontrada una descripción de la especie realizada por Chapman,
(1892) en su libro “Notes on birds and mammals observed near Trinidad, Cuba”,
haciendo referencia a la especie de la siguiente manera:
“Zenaidura macroura (Linn.). Paloma de luto. En todas partes es una especie común.
En San Pablo eran más abundantes, y había pocos intervalos durante el día en que
su llamada no se podía escuchar. Por la tarde al caminar sobre las malezas cerca
del río Agabama, en casi todos los pasos surgían en pequeñas bandadas casi desde
debajo de mis pies. Evidentemente que estaban aquí para proporcionarse grava fina.
En el momento de mi visita estaban criando las aves, la colocación de sus nidos en
los arbustos de guayaba, incluso en las palmeras, donde las hojas se ramifican

8
desde el tronco. Ahora observo con frecuencia aquí una peculiar evolución aérea
probablemente limita a la temporada de reproducción. Anteriormente observado
solamente en Corpus Christi, Texas, donde se agrupan las aves reproductoras. Esta
evolución se compone de un corto vuelo natural, seguido de una vela, a veces
durante una carrera circular. Este cortejo se repite dos o tres veces, y el pájaro luego
planea a la tierra o una percha cerca del nido. Todo el proceso no cubre más de
doscientos pies, y es de suponer que se limita a las proximidades del nido. Por lo
menos he observado en varias ocasiones que ocurre por arriba de un nido. Durante
este vuelo y planeo el ave tanto se parece exactamente al Accipiter que nunca vi uno
que realizara sin tomar involuntariamente mi escopeta para disparar a lo que había
confundido con un halcón. De hecho, es tan cerca la semejanza, que sólo después
de un escrutinio cuidadoso se reconoce la paloma.
Los especímenes cubanos son más pequeños que los ejemplos de Z. macroura del
este de los Estados Unidos, y en su color ligeramente más oscuro y
desproporcionadamente más corta la cola, mostrando una aproximación a Zenaida
cardosantera. Las medidas promedio son los siguientes de cinco ejemplares
cubanos: ala, 5,52; cola, 4.8 pulg. Cinco ejemplares de las proximidades de la ciudad
de Nueva York: ala, 5,92; cola, 5,40 pulg”.
1.3.- Importancia de la Paloma Rabiche (Zenaida macroura L.) como especie
cinegética en Cuba
En Cuba, el origen de la actividad cinegética parte desde la caza de subsistencia
practicada por los aborígenes, única población humana existente en la isla antes de
la colonización española. Según Sejourrné, (1974), citado por Chamizo et al. (2011),
la caza, como la pesca, era “rica y abundante”, aunque en estos tiempos se cazaran
también aves, estas eran menos usadas, siendo más consumidos las jutías [J. conga
(Capromys pilorides), J. carabalí (Capromis prehensilis prehensilis), y el Manatí
(Trichechus manatus L.)] Tabío y Rey, (1979) citado por Chamizo et al. (2011).
La cita más antigua que se conoce sobre el uso de la paloma Rabiche como especie
cinegética es recogida en la obra de Gundlash “Contribución a la ornitología cubana”

9
impresa en la Habana en 1876, donde expresa “Es útil al hombre por su carne
sabrosa”. Más tarde en la obra “Notes on birds and mammals observed near
Trinidad, Cuba” escrita por Chapman, (1892), en el capítulo titulado “General
Impressions of Cuban Bird-life” refiere: “Las restricciones impuestas por el gobierno
sobre el uso de armas de fuego, el alto precio que se cobra para la munición, y la
falta de caza, a excepción de las codornices y palomas, son excelentes razones por
las que deberían ser poco cazadas en Cuba, y durante mi estancia me encontré tres
cazadores los cuales estaban cazando palomas (Zenaidura, Zenaida y Columba
corensis”. Por lo antes descrito por este autor se puede apreciar la abundancia, así
como la estima de esta especie como ave cinegética en nuestro país desde mucho
tiempo atrás.
Actualmente la Paloma Rabiche (Zenaida macroura) es una de las más importantes
especies de caza de Cuba, lo que está relacionado con sus principales
características (Acosta y Ponce de León (2004); García y Chamizo (2004), a saber:
Excelentes aptitudes cinegéticas, que se evidencian a través de un rápido y
zigzagueante vuelo, sobre todo cuando descubre la presencia del cazador.
Carne de excelente calidad.
Su presencia se relaciona con la disponibilidad temporal de alimento, lo que se
traduce en grandes concentraciones (bandos) en determinados lugares y épocas.
Amplia distribución en todo el territorio nacional
Alta abundancia en todo el territorio nacional
Mantiene actividad reproductiva durante todo el año.
Está asociada fundamentalmente a biotopos agrícolas, lo cual facilita la
organización de la caza.
Poblaciones aparentemente estables ante las presiones tanto cinegéticas como
las de otros usos consuntivos y ante los cambios de su hábitat.

10
El interés por la especie como ave cinegética no es solo en Cuba, pues en muchos
países donde habita es considerada como una especie digna de los mejores y más
refinados tiradores. Al respecto Bryant (1926) en Cleveland (1932) expresa que
aparte de su atractivo como un manjar en la mesa, la paloma es un ave de caza
popular, siendo un objetivo que se mueve rápidamente y que exige la máxima
habilidad por parte del cazador. Cuenta que solo estando bien situado cerca de un
abrevadero, abatir tres o cuatro palomas es un excelente promedio para el cazador
más experimentado, su pequeño tamaño y gran velocidad la convierten en un
objetivo difícil. La variedad de tiros posibles es casi interminable, los tiros laterales
son los más difíciles por la velocidad del ave en vuelo, los tiros en torre y los
descendentes, dependen de si el ave va o viene. Los disparos rectos son los menos
frecuentes en esta paloma que en la caza de la Codorniz.
Por tanto, se aprecia que la caza de esta paloma es la mejor para los amantes del
tiro al vuelo. No existe prueba mejor de habilidad en la caza a menos que sea en el
tiro a la Becasina.
La Paloma Rabiche (Zenaida macroura L.) ha sido ampliamente estudiada en los
Estados unidos de América en toda su área de distribución, realizándose
importantes investigaciones, las cuales abarcan aspectos relacionados con la
ecología, el hábitat, la reproducción, las enfermedades y parásitos, la dinámica
poblacional y la respuesta de la especie ante la presión de la caza (Gustswiller et al.,
2001; Ostrand et al., 1998; Haysiette y Mirarchi, 2002; Ostrand et al., 1996; Miller et
al., 2001 y Smith et al., 2012).
Respecto al hábitat de la especie y su relación con otras aves, Gustswiller et al.,
(2001) evaluaron la relación entre las aves y el paisaje a lo largo de la frontera de
México-Texas, encontrando que a menudo las distribuciones de las aves se
relacionaban significativamente con las características del paisaje, por lo que
muchos modelos eran útiles para predecir estas relaciones. También encontraron
que las variaciones aves-paisajes pueden ser influidas por factores como la sucesión
de plantas exóticas, la tolerancia a la variación ambiental de las aves y la diferencia
en la densidad de comida o el clima. Concluyeron que la baja regularidad de los

11
modelos para la mayoría de las especies demuestra la alta diferencias en las
relaciones aves-paisajes con las que se pueden encontrar los decisores, la
incertidumbre en estos modelos estructurales se pueden reducir logrando que estos
cubran los supuestos estadísticos más importantes, probando estos modelos a
través de muestras precisas en el campo y con tamaños muestrales que suministren
datos precisos.
Ostrand et al., (1998), identificaron los hábitat de preferencia de la Paloma Rabiche
para alimentarse, determinaron si el hábitat había sido alterado y evaluaron si los
cambios podían ser los responsables del descenso de los índices de población local.
Encontraron una reducción en los índices poblacionales en un 72 % en el periodo de
1952-1992 y en un 82 % en el periodo de 1952-1993; el cambio más dramático en el
hábitat fue la reducción del 82 % en los terrenos utilizados en la producción de trigo
invernal y una reducción de los comederos para el ganado. Según su hipótesis la
reducción en la disponibilidad de trigo y la consolidación de la industria ganadera ha
contribuido al descenso de la población de palomas en la parte central de Utah,
Estados Unidos de América.
Son de gran interés los trabajos relacionados con la alimentación de la especie,
señalándose el realizado por Haysiette y Mirarchi (2002), donde se determinó la
preferencia alimentaria de la especie en vida silvestre y cautiverio, así como los
elementos contenidos en las semilla, ellos exponen que consumieron la comida de
forma selectiva, tanto las palomas silvestres como las cautivas, estas preferencias
no variaron con las estaciones pero estaban relacionadas de forma positiva con la
presencia del Nitrógeno libre (NFE) en las semillas y de forma negativa con los
niveles de celulosa-lignina, aunque esto no explicó la selección de los alimentos
totalmente, las características físicas de las semillas y la eficiencia metabólica
podrían haber influido en la selección de los alimentos en este estudio
Ostrand et al. (1996), estudiaron el hábitat de forrajeo de Zenaida macroura con un
enfoque estructural, empleando parcelas donde se manipuló la altura de las hierbas,
densidad, cobertura de éstas y cobertura de hojarasca, utilizando parcelas sin
manipular la cobertura como testigos y así conocer la estructura del hábitat preferido,

12
encontrando una mayor preferencia por los lugares manipulados que en las áreas
controles en donde la estructura del hábitat se mantuvo inalterada. Concluyeron que
las palomas basaban su elección de forrajeo en la altura de las hierbas y en su
densidad, estos resultados sugieren que la conversión de campos de Trigo a
herbazales resulta un cambio estructural de menor preferencia para la especie.
Especial importancia tiene el estudio del tema reproductivo, donde se destaca el
realizado por Miller et al. (2001), analizaron la productividad de esta especie durante
la etapa de 1992 hasta 1995 en el Valle Central del Norte de California, obteniendo
que la temporada de anidación osciló entre 105 y 158 días, con un éxito reproductivo
mayor en las temporadas relativamente más cálidas y secas que en las
relativamente más frías y húmedas.
Smith et al. (2012), estudiaron el comportamiento de la nidificación de la especie en
bosques ribereños con diferentes estructuras, encontrando el mayor número de
nidos de la especie en áreas post incendio forestales donde abunda un sotobosque
denso y las intervenciones antrópicas son pocas, las menos utilizadas resultaron ser
aquellas áreas donde se realizaron intervenciones para reducir el material
combustible y evitar los incendio forestales.
En Cuba se ha estudiado la ecología trófica del género Zenaida por parte de Cuesta
(2009), Acosta y Torres (1984), Acosta y Berovides (1982) y Garrido (1986) incluye a
Z. macroura en su descripción de las palomas cubanas. Se ha encontrado una ficha
técnica de esta especie (Hernández, inédito), elaborada con fines de
aprovechamiento cinegético. Pero a pesar de tratarse de una especie sometida a un
elevado uso consuntivo, faltan las publicaciones relacionadas con su
aprovechamiento cinegético.
1.4.- Planteamiento del problema y Objetivos de la tesis
El conocimiento ecológico de la especie en Cuba resulta ser insuficiente,
fragmentado y disperso, unido a las diferentes formas de aprovechamiento a que es
actualmente sometida la especie y la falta de control sobre las mismas traen
aparejadas importantes dudas sobre los efectos que pueden tener en sus

13
poblaciones. En este sentido se hace evidente que existen dificultades o lagunas en
este tema que necesitan responderse mediante la realización de este estudio.
Basado en lo antes expuesto se asume la siguiente interrogante científica: ¿Qué
aspectos esenciales caracterizan la ecología y el aprovechamiento de la Paloma
Rabiche que puedan ser útiles para un manejo sostenible de la especie en un
agrosistema tropical en Cuba?
En este estudio se partió de la siguiente hipótesis: las condiciones ecológicas donde
se desarrollan las poblaciones de Paloma Rabiche determinan su presencia,
reproducción y abundancia relativa, por tanto su conocimiento y gestión favorecen el
aprovechamiento sostenible de la especie.
El presente estudio tiene como objetivo general: Caracterizar los aspectos
esenciales de la ecología y el aprovechamiento de la Paloma Rabiche (Zenaida
macroura L.) que pueden ser útiles para un manejo sostenible en un agrosistema
tropical en Cuba.
Sus objetivos específicos están dirigidos a:
Caracterizar los biotopos presentes en el área de estudio según la densidad y
uso de estos por la Paloma Rabiche.
Identificar aspectos ecológicos de la especie en el área de estudio atendiendo a:
época de reproducción, preferencias para la nidificación, éxito reproductivo,
espectro alimentario, preferencia alimentaria de la especie y presencia de
ectoparásitos.
Relacionar las diferentes asociaciones vegetales y la cobertura del suelo
presentes en el área de estudio con aspectos conductuales como alimentación,
descanso y nidificación.
Describir la morfometría de la especie en el área de estudio.
Caracterizar las distintas formas de aprovechamiento de la especie en el área de
estudio.

14
La novedad científica de la investigación está en que relaciona la distribución y
disponibilidad de alimentos con la abundancia relativa de la especie, ofreciendo
alternativas para predecir su dinámica poblacional y planificar la caza de acuerdo
con los principios de la sostenibilidad, en atención a la capacidad reproductiva del
recurso y a la duración del período crítico de cría. Además, brinda fundamentos
teóricos para determinar los períodos de caza y veda de la especie y aporta
conocimientos sobre el espectro alimentario de esta ave que tienen aplicación en la
gestión de su hábitat.
1.5.- Contenido de los capítulos
El Capítulo I, corresponde a la Introducción general de la tesis. En él se refiere la
importancia del estudio de la actividad cinegética, se describen las principales
características de la Paloma Rabiche, se declara el problema científico abordado en
la investigación, se enuncian la hipótesis general del estudio, se enuncia el objetivo
general, se dan a conocer los objetivos específicos de la tesis y se ofrece un perfil
del contenido de los capítulos desarrollados.
El Capítulo II. Características del área de estudio, describe el área de estudio, su
ubicación geográfica, así como las principales características climáticas,
hidrológicas, forma del relieve y actividades socioeconómicas presentes en ella.
En el Capítulo III, Caracterización de la vegetación como biotopo de la Paloma
Rabiche se presenta un estudio de las principales formaciones vegetales presentes
en el área de estudio, después de describir los materiales y métodos empleados
para ello, y se clasifican y describen los biotopos presentes en esta, así como la
riqueza y diversidad vegetal existente. La caracterización de la vegetación es
fundamental cuando se estudia la ecología de especies cinegéticas, ya que permite
definir la disponibilidad de las especies vegetales más importantes para esta, y por
tanto determinan la calidad del hábitat, contribuyendo al conocimiento de los
requerimientos vitales de esta especie, lo cual deberá considerarse dentro de los
planes de manejo.

15
En el Capítulo IV, La Paloma Rabiche en la comunidad ornitíca, se describen los
materiales y métodos usados en la evaluación del comportamiento de la avifauna del
área de estudio, se determinan parámetros como la abundancia relativa, la
frecuencia, se establecen los grados de fidelidad que señalan la intensidad o
frecuencia de la Paloma Rabiche en los biotopos presentes en el área de estudio.
En el Capítulo V, Reproducción y conducta reproductiva de la Paloma Rabiche
(Zenaida macroura L.). Se evalúa la conducta reproductiva de la especie a través del
año, se presenta el estado de conocimiento sobre la especie de los sectores que
tienen relación con ella (campesinos y cazadores) así como la descripción de los
sustratos usados para la nidificación.
El Capítulo VI. Estructura de la población de Paloma Rabiche (Zenaida macroura
L.). Se determina la estructura de edades y la proporción de sexos en la población
del área de estudio, elementos que permitirá diagnosticar el estado de esta, predecir
su tendencia y fijar racionalmente una gestión sostenible del recurso.
El Capítulo VII, Ecología trófica de la Paloma Rabiche (Zenaida macroura L.), se
determina el espectro alimentario y se calcula el Índice de llenura como indicador
para calcular el tamaño de muestra necesario para el análisis de la dieta de la
especies estudiada, también se ha calculado el Índice de Importancia Alimentaria
para conocer los principales componentes en la dieta de la Paloma Rabiche.
El Capítulo VIII, Calidad del hábitat de la Paloma Rabiche (Zenaida macroura L.),
tuvo como objetivo determinar la relación entre las diferentes características del
hábitat del área de estudio a través de los diferentes grados de selección de hábitat
la especie estudiada y tres parámetros conductuales (descanso, nidificación y
alimentación) lo cual facilitará la predicción de su distribución y abundancia, lo que
permitirá una gestión en función de las características del hábitat.
El Capítulo IX, Morfometría de la población de Paloma Rabiche (Zenaida macroura
L.), se caracterizó morfométricamente la población de la Paloma Rabiche a través de
variables como el peso total del ave, longitud total, longitud del ala plegada, de la
cola y del tarso, largo y ancho del pico. Además se realiza su comparación con datos

16
obtenidos en otras investigaciones sobre la especie en el país y con las medidas que
se describen en la bibliografía de la misma. Esto permitirá conocer la variabilidad de
esta especie en relación con las diferentes latitudes y las condiciones ambientales
que pudiesen provocar dichos cambios, siendo conocido que algunas especies
migratorias presentan patrones de variaciones morfológicas a nivel geográfico.
El Capítulo X, consiste en una Caracterización de la presencia de ectoparásitos en la
población de Paloma Rabiche (Zenaida macroura L.). En él se identifican los
ectoparásitos hallados, así como las principales épocas en que estos aparecen. Fue
calculado el índice de prevalencia, así como su comportamiento por clase de edad y
sexos.
En el Capítulo XI, Aprovechamiento cinegético de la Paloma Rabiche (Zenaida
macroura L.), se describen las principales formas tradicionales de aprovechamiento
cinegéticos de la especie, su estatus legal y una serie de criterio e indicadores como
herramientas para el manejo sostenible de la especie.
Cada capítulo cuenta con una introducción referida a cada tema en cuestión, un
acápite de materiales y métodos, resultados y discusión.
El tema tributa al Programa Nacional de Ciencia y Técnica: Protección al Medio
Ambiente y Desarrollo Sostenible Cubano, Cambios Globales y Evolución del Medio
Ambiente Cubano y al Programa Ramal Científico Técnico de Diversidad Biológica
(CITMA), Sub Programa: Conservación de la Diversidad Biológica.
De la presente investigación han sido presentados resultados en diferentes eventos,
tanto nacionales como internacionales, así como varias publicaciones las que se
relacionan a continuación.
Publicaciones:
“Ecología reproductiva de la Paloma Rabiche (Zenaida macroura L.) en un
agrosistema tropical (Cuba)”. En preparación.

17
“Feeding use of a tropical agroecosystem (Cuba) by Mourning dove (Zenaida
macroura)”. The Journal of Animal and Plant Sciences (The JAPS). Pakistan.
2016.
Libro impreso “Caza y conservación en el ámbito rural de Cuba”. Publicaciones
Universidad de Alicante. España. ISBN 978-84-9717-181-6. 2011.
“Caracterización del comportamiento reproductivo y alimentario de la Paloma
Rabiche (Zenaida macroura L.) en el territorio de Rodas. V Simposio Internaional
Sobre manejo Sostenible de los Recursos Forestales 2008. Universidad de Pinar
del Río. Cuba. ISBN978-959-16-0655-6.
“Caracterización del comportamiento reproductivo y alimentario de la Paloma
Rabiche (Zenaida macroura L.) en el territorio de Rodas. ISBN 978-959-257-195-
2. Universidad “Carlos Rafael Rodríguez”. Cienfuegos. Anuario 2008.
Eventos y talleres científicos:
“Incidencia de dos especies de interés cinegético en la agricultura cubana
(Zenaida macroura y Colinus virginianus)”. Taller local “Caza y conservación en el
ámbito rural de Cuba”. Universidad de Alicante, España y Jardín Botánico de
Cienfuegos. Cienfuegos. 2012.
“Caracterización del comportamiento reproductivo y alimentario de la Paloma
Rabiche (Zenaida macroura L.) en el territorio de Rodas. V Simposio Internaional
Sobre manejo Sostenible de los Recursos Forestales 2008. Universidad de pinar
del Río. Cuba.
“Principales características ecológicas de la Paloma Rabiche (Zenaida macroura)
en un área de caza dl municipio Rodas, Cienfuegos. XV pleno Nacional de la
Federación Cubana de Caza Deportiva. Ciudad de la Habana. Cuba. 2005.
“Caracterización del comportamiento reproductivo y alimentario de la Paloma
Rabiche (Zenaida macroura L.) en el territorio de Rodas. Tercer Congreso
Forestal de Cuba. 2004.

18
-“Caracterización del comportamiento reproductivo y alimentario de la Paloma
Rabiche (Zenaida macroura L.) en el territorio de Rodas. Tercer Pre Congreso
Forestal de Cuba. Cienfuegos. Cuba.2004.
El presente estudio se basó en el principio de evitar a toda costa la pérdida
innecesaria de ejemplares y huevos, así como minimizar en lo posible el estrés a las
aves. Por tal motivo algunos aspectos no fueron abordados a profundidad o los
métodos utilizados fueron adaptados a este principio. Solo fueron colectados los
ejemplares necesarios para este estudio.
1.6.- Bibliografía
Acosta, M. y Berovides. V. (1982). Ecología trófica de las palomas del género
Zenaida en el sur de Pinar del Río. Ciencias Biológicas, 7, 113-123.
Acosta, M. y O. Torres. (1984). Ecología trófica de las palomas del género Zenaida
en el Jardín Botánico de Cienfuegos, Cuba. Ciencias Biológicas, 11, 107-116.
Acosta, M; P. de león, J.L. (2004). Historia Natural de la Ploma Rabiche (Zenaida
macroura). (Inedito). Facultad de Biología. Universidad de la Habana. Cuba.
Aramburo, A. y S. Pons. (1956). Adiestramiento del perro de muestra. Editora Latino
Americana. México. 269 p.
Chamizo, R. (2000). Curso de capacitación sobre gestión de caza menor. MINAGRI.
Chamizo, R. (2004). Caza y sostenibilidad: una polémica de actualidad también en
Cuba. Revista Forestal Baracoa. La Habana. 85–92.
Chamizo, R. et al. (2011). Caza y conservación en el ámbito rural en Cuba.
Publicaciones Universidad de Alicante. España. p. 13.
Chapman, F. M. (1892). Notes on birds and mammals observed near Trinidad, Cuba
with remarks on the origen of West Indian bird–life. Bulletin American Museum of
Natural History. N. I, v. IV. 291– 292.

19
Cuesta, L. A. (2009). Perspectivas hacia una Gestión Sostenible de la Actividad
Cinegética en la Provincia de Pinar del Río, Cuba. Universidad de Alicante,
España, Universidad de Pinar del Río, Cuba.
Dolton, D. D., and R. D. Rau. (2003). Mourning dove population status, 2003. U.S.
Fish and Wildlife Service, Laurel, Maryland. 27 p.
Garrido, O. (1986). Las palomas. Editorial Científico–Técnica. Ciudad de la Habana.
52 p.
Garrido, O. H. y Kirkcornnell, A. (2011). Aves de Cuba. Comstock Publishing
Associates a división of Cornell University Press. Ithaca. Estados Unidos de
América. 287 p.
Gundlash, J. (1876). Contribución a la ornitología cubana. Editora La antillana. La
Habana. p 139 – 140.
Gutzwiller, K. et al. (2001). Bird-landscape relations in the Chihuahuan Desert:
Coping with uncertainties about predictive models. Journal Ecological
Applications, 11 (5), 1517-1532.
Hayslette, S. E.; Mirarchi, R. E. (2002). Foraging-patch use and within-patch diet
selectivity in Mourning Doves, Zenaida macroura. Ecology, 83 (9), 2637-2641.
Martinez, J. (1981). Protección y desarrollo de la fauna. Editorial Libros para la
Educación. Ciudad de la Habana. 124 p.
Miller et al. (2001). Mourning dove productivity in California during 1992-95: Was it
sufficient to balance mortality?, Journal of Wildlife Management, 65 (2), 300 –
311.
Ostrand, W. D., Bissonette, J. A. & Conover, M. R. (1996). Selection of foraging
habitat by Mourning Doves: a structural approach. Journal of Field Ornithology, 6
(4), 691-697.
Ostrand, W. D. et al. (1998). Changes in Land Use as a Possible Factor in Mourning
Dove population Decline in Central Uthah. Journal of Field Ornithology, 69 (2),
192-200.

20
Peiró, V. (2003). Gestión ecológica de los recursos cinegéticos. Publicaciones de la
Universidad de Alicante. España. 136 p.
Smith, D. M., Finch, D. M. & Hawksworth , D. L. (2012). Nesting characteristics of
mourning doves in central New Mexico: Response to riparian forest change. The
Journal of Wildlife management, 76 (2), 382-390.
.

21
CAPÍTULO II – CARACTERISTICAS DEL AREA DE ESTUDIO
2.1.- Ubicación geográfica del área de estudio
La gran diversidad y características propias de cada ecosistema están dadas por las
diferencias geográficas, físicas y biológicas propias de cada región. Esta gran
diversidad está amenazadas tanto por los fenómenos tanto naturales como los
generados por el hombre, por tanto cada uno de los componentes que integran un
ecosistema debe ser gestionado de acuerdo con sus propias características para
evitar su deterioro o pérdida total.
Cada vez está más difundida y aceptada entre profesionales y parte de la sociedad,
la necesidad de una gestión integradora de los bosques que permita el
mantenimiento no solo del recurso explotable (madera, leñas, setas…), sino del
conjunto de seres vivos que componen el ecosistema. Tendencia que surge de la
aceptación de la multifuncionalidad de los bosques y de los sistemas agroforestales
en general, donde la conservación de la función biológica es crucial para el
mantenimiento de las demás funciones: productividad, ambiental y recreativa.
Además, es posible que en un futuro próximo, la potenciación de la biodiversidad
constituya un beneficio económico, directo o indirecto, para las rentas de las
poblaciones rurales (Camprodon y Subirachs, 2001).
El conocimiento de los ecosistemas y su conservación, constituyen un reto para la
investigación científica. No es posible el aprovechamiento sostenido de un recurso
natural sin el conocimiento del ecosistema en que se efectuará éste.
La investigación se desarrolló el área de caza “Damují” del municipio de Rodas,
provincia de Cienfuegos, Cuba (Figura 3), su centro se localiza en las coordenadas
geográficas 22° – 21.804' N y 080
° - 33.430' W, abarcando un total de 645,8 ha. A
efectos de la presente investigación se seleccionó esta zona porque reúne las
características representativas de los biotopos más usados por la especie objeto de
estudio en Cuba.

22
Figura 3. Ubicación geográfica del área de estudio.
2.2.- Clima
El clima de la zona, corresponde al Tropical poco húmedo de llanura. Este tipo de
climático está ampliamente difundido hacia la zona central y occidental de la
provincia, el que se caracteriza por ser caliente, húmedo y lluvioso entre los meses
de mayo a octubre, y seco y poco húmedo, con temperaturas menos calientes de
noviembre a abril. La temperatura media anual fluctúa entre los 24 a 26 0C, en el
periodo seco oscila entre los 22 a 240C y en el periodo húmedo mayor de 26
0C. Las
precipitaciones tienen una media anual entre los 1000 y 1100 mm, siendo menor de
200 mm en el periodo seco y entre los 1000 y 1200 mm en el húmedo. La intensidad
media de lluvias fluctúa entre los 15 y 25 mm. La mayor parte de las precipitaciones
están asociadas con frentes convectivos de verano. La presión atmosférica se
mantiene de relativamente baja a muy baja, la nubosidad es media entre 4/8 y 5/8 a

23
cielo cubierto. La zona es afectada por la trayectoria de los huracanes,
fundamentalmente en el mes de octubre. Durante la época seca es frecuente que se
originen remolinos de polvo (Sulroca, 1976).
2.3.- Hidrología
La red hídrica del área de estudio está compuesta por el río “Damují”, con aguas de
excelente calidad disponibles durante todo el año, tanto para el consumo de la fauna
silvestre, como para el humano; también existen algunas cañadas intermitentes que
en época de lluvias constituyen fuente de abasto para la fauna silvestre y el ganado.
Los cultivos disponen de agua de buena calidad permanentemente gracias a los
sistemas riego existente en la zona.
Figura 4. Mapa hidrológico de la zona.

24
2.4.- Relieve y suelos
Predomina un relieve llano (pendientes de 0 – 20 %) en casi toda el área, solo en la
parte este, próxima al río “Damují” presenta algunas ondulaciones.
Figura 5. Mapa del relieve de la zona.

25
Los suelos cambian de un lugar a otro. Su estructura física y composición química
está determinada por el tipo de material geológico del que se origina, la cubierta
vegetal, la topografía y los cambios resultantes de las actividades humanas.
Los requerimientos de suelo de las distintas plantas varía mucho, plantas como el
Arroz (Oriza sativa) o la Cortadera (Cladium jamaicense) requieren suelos húmedos
que estarían insuficiente mente drenados para Caimitillo (Chysophyllum oliviforme) o
la Baria (Cordia gerascanthus). Es entonces evidente que el tipo de suelo determina
en gran medida la vegetación del área y esta a su vez en la fauna silvestre.
En toda el área de estudio existe un total de cuatro tipos de suelo: ferralítico rojo
típico (II-A), fersialítico pardo rojizo típico (VIII-A), húmico calsimórfico (XII-A), y el
húmico calsimórfico (XIII-A), con predominio del ferralítico rojo típico y el fersialítico
pardo rojizo.
Los suelo ferralítico rojo típico (II-A) son suelos rojos que se forman por el proceso
de ferralitización, el que se caracteriza por una alteración intensa de los minerales,
con lavado de la mayor parte de las bases alcalinas y alcalinotérreas y una parte de
la sílice. Su PH se clasifica como ligeramente acido (6-6,5) y el contenido de materia
orgánica oscila entre un 2-5 %.
Los fersialítico pardo rojizo típico (VIII-A) son suelos rojos que se forman bajo el
proceso de fersialitización caracterizado por la presencia de minerales arcillosos y un
contenido de Fe2O3 con colores rojos o amarillentos en el perfil o en algunos de los
horizontes). Su PH se clasifica como ligeramente acido (6-6,5) y el contenido de
materia orgánica oscila entre un 2-5 %.
Los suelos húmico calsimórfico (XII-A) y el húmico calsimórfico (XIII-A) en ellos
predomina el proceso de humificación favorecido por la presencia de grandes
cantidades de calcio activo y de arcillas. Estos suelos se desarrollan sobre roca
caliza; casi todos presentan CaCO3 en todo el perfil. Su PH es clasificado como
neutro (7) y el contenido de materia orgánica oscila alrededor del 6 %.

26
Figura 6. Mapa de suelo de la zona de estudio.
2.5.- Usos del suelo
El uso mayoritario del suelo es el cultivo agrícola y la ganadería con una amplia red
de linderos o setos que dividen estas áreas. Una gran parte del área está dedicada a
los cultivos agrícolas de forma intensiva (Manihot esculenta, Ipomea batatas y Zea
mays entre otros), donde predominan las rotaciones de cultivos menores con riego
artificial durante todo el año, pero también presentan pastos y plantas herbáceas
bordeando los canales de riego que se mantienen con agua la mayor parte del año.
Existen áreas dedicadas a la ganadería en las cuales predominan los pastos, tanto
naturales como artificiales (Andropogon annulatus, Panicum maximun, Eleusine
indica), aunque en algunos casos aparecen áreas de potreros invadidas por el
Marabú (Dichostachys cynerea), y otras malas hierbas. La parte restante de la zona

27
(63 ha) está ocupada por un bosque natural semicaducifolio (Samanea saman,
Bursera simaruba, Guazuma tomentosa) sobre suelo calizo que en algunos sitios
presenta un aceptable grado de conservación y en otros aparecen áreas abiertas
(claros) de cultivos abandonados.
Figura 7. Mapa de los usos de suelos del área de estudio.

28
2.6.- Bibliografía
Bisse, J. (1988). Árboles de Cuba. Editora Científico-Técnica. Ciudad de la Habana.
Cuba. 384 pp.
Camprodon, J; Subirachs, E. P. (2001). Conservación de la biodiversidad y gestión
forestal. Su aplicación en la fauna vertebrada. Ediciones de la Universidad de
Barcelona. España. 469 pp.
Catasús, L. J. (2002). Las gramíneas (Poaceae) de Cuba, II. Editorial Cavanillesia
Altera 3. Madrid. España. 165 pp.
León, H. y Alain, H. (1957). Flora de Cuba. Vol.IV. Contr. Mus. Hist. Nat. Colegio de
La Salle, No. 16, Imp. P. Fernández y Cía, La Habana. 556 pp.
León, H. y Alain, H. (1953). Flora de Cuba. Vol. III.Contr. Mus. Hist. Nat. Colegio de
La Salle, No.13, Imp. P. Fernández y Cía, La Habana. 502 pp.
León, H. y Alain, H. (1951). Flora de Cuba. Vol. II. Contr. Mus. Hist. Nat. Colegio de
La Salle, No. 10, Imp. P. Fernández y Cía, La Habana. 456 pp.
Rodríguez, S., Rodríguez, J. y Pérez, L. (1988). Plantas indeseables en el cultivo de
la caña de azúcar. Editora Científico-Técnica. Ciudad de la Habana. Cuba.196
pp.
Roig, J. T. (1965). Diccionario Botánico de Nombres Vulgares Cubanos. Tomo I y II.
Editorial del Consejo Nacional de Universidades. La Habana. Cuba. 1142p.
Sulroca O. (1976). Zonificación climática de Cienfuegos, DPPF, Cienfuegos. 34 pp.

29
CAPÍTULO III – CARACTERIZACION DE LA VEGETACION COMO BIOTIPO DE
LA PALOMA RABICHE
3.1.- Introducción
La vegetación es un recurso natural clave para el equilibrio del ecosistema por lo que
es necesario disponer de información cuantitativa sobre sus características y
distribución. Las comunidades de aves, su abundancia y diversidad están
estrechamente influenciadas por la composición de plantas, así como por la
altura de la vegetación . Un gran número de plantas nativas mantienen una
estrecha relación con las aves debido a que su dispersión y polinización
depende de las aves. Algunos frutos maduran en la misma estación que las
aves migratorias almacenan reservas de energía para la migración. Esta temática
ha sido abordada por diferentes autores (Bautista, 2013; Marques et al., 2005;
Ramírez y Ramírez, 2002 y Zaragoza et al., 2004).
Algunos estudios muestran una relación positiva entre las aves y la vegetación
nativa, que mantiene una alta diversidad y riqueza de especies de aves;
igualmente las aves no nativas responden positivamente a la vegetación exótica
(Emlen, 1974; Mills et al., 1989, Chace et al., 2004; Day, 1995; en French et al.
2005, citados por (Bautista, 2013).
Por tal motivo la caracterización de la vegetación es fundamental cuando se estudia
la ecología de especies cinegéticas, ya que proporciona información relevante sobre
las condiciones naturales, principalmente de alimentación y refugio que ésta le
brinda a la especie estudiada.
La presente investigación se circunscribe al estudio de dos hábitats representativos
de Zenaida macroura en esta región: las áreas agrícolas tradicionales y el bosque
semicaducifolio sobre suelo calizo. La primera es considerada de importancia para
muchas especies de aves, ya que sus linderos actúan como corredor de dispersión
y albergue de fauna silvestre; adicionalmente, proveen el hábitat a invertebrados y
especies vegetales típicas de estas que son fuente importante para la fauna
asociada a ellos. El segundo hábitat figura como otro tipo de ecosistema con

30
factores y recursos distintos. En él se destaca el predominio de la flora arbórea, una
gran cobertura del suelo y un régimen menor de radiación solar directa sobre el
suelo. Ambos cuentan con composición florística, estructura y diversidad distinta,
pudiendo llegar a hospedar organismos que, en algunos casos se clasifican en algún
tipo de categoría de amenaza.
El objetivo de este capítulo fue realizar una caracterización de los biotopos presentes
en el área de estudio que permitirá definir la disponibilidad de las especies vegetales
más importantes para la Paloma Rabiche, las cuales influyen en la dinámica
poblacional, y por tanto determinan la calidad del hábitat. Se contribuirá así al
conocimiento de los requerimientos vitales de esta especie, la que ha sido poco
estudiada en Cuba. La información que se obtenga de esta investigación deberá
considerarse dentro de los planes de manejo de la especie, en beneficio de su
conservación y de las condiciones ecológicas de la región.
3.2.- Materiales y métodos
Los inventarios describen la estructura y función de la vegetación para su aplicación
en el uso y manejo de la misma (Álvarez et al., 2006, en Campo y Soledad, 2014).
La caracterización de sus propiedades fisonómicas permite el reconocimiento de la
complejidad estructural presente mientras que su representación mediante fórmulas
resume la información en un solo valor. También es posible realizar comparaciones
entre la diversidad de distintos hábitats o la diversidad de un mismo hábitat a través
del tiempo. Los índices cuantitativos muestran la relevancia de su conservación en
áreas protegidas (Suárez y Vischi, 1997 en Campo y Soledad, 2014).
El inventario de la flora en los biotopos presentes en la zona se desarrolló por el
método propuesto por Mostacedo y Fredericksen (2000). En las áreas de cultivos
agrícolas y potreros se utilizó el método de la selección de parcelas en cuadrantes,
de 1,30 x 1,30 m; se realizaron un total de 56 parcelas, distribuidas al azar,
garantizando así la representación de los diferentes hábitats donde se identificaron
cada una de las especies presentes. En el área boscosa se realizaron 9 transectos

31
lineales de 500 m de longitud y 4 m de ancho distribuidos al azar, garantizando así la
representación de todas las especies (Anexo 1).
La determinación del número de muestras adecuado para el estudio, se efectuó a
través del método de la curva especie-área, la cual permite visualizar la
representatividad de un muestreo. Cuando la curva tiende a mantenerse horizontal,
indica que el número de especies se mantendrá constante aunque aumente el
tamaño de muestreo (Mostacedo y Fredericksen, 2000).
En la identificación de la especies vegetales se emplearon las obras de León y Alain,
1951; 1953 y 1957; Bisse, 1998; Rodríguez, Rodríguez y Pérez, 1988 y Sánchez y
Uranga, 1993.
En el análisis de la estructura horizontal de la vegetación se tuvieron en cuenta los
criterios de Mostacedo y Fredericksen (2000) donde: se valora la participación de
cada especie en relación a las demás presentes, que da idea de su distribución
espacial, tanto en cantidad como en frecuencia. Este aspecto puede ser determinado
a través de parámetros como la abundancia relativa y la frecuencia con que se
encuentra representadas las especies
La abundancia relativa indica la participación que tiene cada una de las especies de
un área determinada y nos permite identificar aquellas especies que presentan
escasa representatividad (Magurran, 1988).
Ar = ni / u
donde:
ni representa el promedio de individuos de la especie i
u representa la unidad de medida utilizada (tiempo, longitud área etc.)
La frecuencia se define como la proporción de veces en que aparece una especie en
determinadas unidades en relación con todas las unidades muestreadas.
F = ni / N
donde:

32
ni representa el número de unidades muestreadas donde aparece la especie i
N representa el número total de unidades muestreadas
Según Moreno (2001), la diversidad en un área específica se mide a través de dos
métodos: aquellos basados en la cuantificación del número de especies presentes
(riqueza específica) y los métodos basados en la estructura de la comunidad, es
decir los que toman en cuenta la distribución proporcional de la especies
(dominancia y equidad).
Los índices de diversidad son aquellos que describen lo diverso que puede ser un
determinado lugar, considerando el número de especies (riqueza) y el número de
individuos de cada especie (Mostacedo y Fredericksen, 2000).
En este estudio fueron utilizados para caracterizar la vegetación los siguientes
índices ecológicos:
Riqueza específica, es la forma más sencilla de evaluar la diversidad puntual y
provee información suficiente sobre la expresión de procesos ecológicos e históricos:
El índice de Margalef (DMg), es una forma sencilla de medir la biodiversidad ya que
proporciona datos de riqueza de especies de la vegetación. Mide el número de
especies por número de individuos especificados o la cantidad de especies por área
en una muestra (Moreno, 2001). Su fórmula es:
DMg = S – 1 / Ln N
donde:
S= número de especies
N=número total de individuos
Índice de diversidad de Shannon – Wiener (con base logaritmo natural), es uno de
los índices más utilizados para determinar la diversidad de especies de plantas de un
determinado hábitat. Este índice se calcula mediante la siguiente fórmula:
H' = Σpi x ln pi
donde:

33
H' representa el índice de Shannon-Wiener
pi representa la abundancia relativa de la especie vegetal i
pi = n / M
n representa el número de individuos de la especie vegetal i
N representa el número total de individuos en toda la muestra o comunidad vegetal
El índice de equitatividad de Pielou (J´), se expresa como el grado de uniformidad en
la distribución de individuos entre especies. Se puede medir comparando la
diversidad observada en una comunidad contra la diversidad máxima posible de una
comunidad hipotética con el mismo número de especies, posee valores que pueden
variar de 0 a 1 siendo cercanos a 1 los que corresponden a situaciones donde todas
las especies son igualmente abundantes (Moreno, 2001).
E = J' = H' / H máx.
H' representa el Índice de Shannon – Wiener
Hmáx. = Ln S
S representa el número total de especies en la muestra.
3.3.- Resultados y discusión
En la caracterización de la vegetación se ofrece en un listado de las especies
vegetales presentes en el área de estudio (Anexo 2). Atendiendo al predomino y la
distribución de las especies vegetales presentes se llegó a la conclusión de que en el
área de estudio, según clasificación de Chamizo (2001), existen dos biotopos usados
por la fauna silvestre:
Agrícola con cobertura: formado por cultivos menores y/o pastizales con linderos,
árboles dispersos, bosquetes o matorrales, cubriendo de un 65 % del área y
suelos bien drenados.
Forestal típico: bosques semicaducifolios o matorrales ralos, continuos con
algunas áreas abiertas, cubriendo un 10 % del área. Buen desarrollo del
sotobosque y suelos bien drenados.

34
Basado en la hipótesis de Hilden (1965), en Carrascal y Telleria (1998): en general,
la abundancia de las especies se relacionan con la disponibilidad de sus
hábitats preferidos, de manera que la estructura de la vegetación es un factor
determinante de los niveles poblacionales de las aves. Teniendo en cuenta lo
antes expuesto se procedió al análisis de la vegetación en el área de estudio.
Figura 8. Momento en que se realizaban los muestreos en las áreas agrícolas.
Una vez definido la forma en que serían realizados los muestreos, surgió la pregunta
de cuántas parcelas eran necesarias para que nuestra información fuese
representativa y confiable sobre el biotopo o área en cuestión a evaluar.
En las figuras 9 y 10 se puede apreciar el comportamiento de las curvas especie -
área en cada biotopo. En el agrícola con cobertura con un total de 20 parcelas se
obtuvo un tamaño de muestra representativo de los diferentes hábitats que
predominaron, en el caso del biotopo forestal típico al ser un hábitat más homogéneo
con 6 trancectos la curva tendió a mantenerse horizontal, indicativo de la no
incorporación de nuevas especies aunque se aumentase el tamaño de muestra.

35
Figura 9. Comportamiento de la relación especie-unidades de muestreo en el biotopo
Agrícola con cobertura.
Figura 10. Comportamiento de la relación especie-unidades de muestreo en el
biotopo Forestal típico.
En el área de estudio se encontraron un total de 101 especies vegetales, agrupadas
en cinco categorías según su forma de vida, distribuidas en 38 familias y 74 géneros.
(Figura 11).

36
Figura 11. Participación de las distintas categorías de especies vegetales presentes
en el área de estudio.
Al realizar el análisis de la abundancia relativa de cada una de las familias presentes
en el área de estudio, así como la frecuencia en que estas están presentes (Figura
11 y Tabla 1) se apreciaron 5 familias que sobresalen del resto por su alta frecuencia
y abundancia relativa. La primera de las familias más abundantes y con una mayor
frecuencia es la Graminaceae lo que está determinado por su forma de vida (gran
número de ejemplares por metros cuadrados y gran capacidad de dispersión).
Dentro de esta familia están presentes en el área un total de 11 especies vegetales,
siendo la más abundante el Andropongon pertussus (Pasto Camagueyano). Esta
especie es ampliamente utilizada como pasto para el ganado en todo el país, por lo
que en las áreas dedicadas a la ganadería existe un gran número de ejemplares. El
resto de las especies que componen esta familia en el biotopo aunque presentan un
elevado número no pueden igualarse con la población de este pasto. Es importante
destacar que dentro de esta familia se encuentran cuatro especies que forman parte
del espectro alimentario de la Paloma Rabiche: la Hierba de Guinea (Panicum
maximun), el arroz (Oriza sativa), Canutillo (Rottboellia conchinchinensis) y el Maíz
(Zea mays).

37
Le siguen en orden de importancia la familia Malvaceae, está representada con ocho
especies de las que solo dos no son usadas por la Paloma Rabiche: el Algodón
cimarrón (Gossypium barbadensis), especie que se encontró al borde de un camino
rural y el Quimbombó (Hibiscus esculentus) especie de gran importancia para la
alimentación de la población. El resto de las seis especies lo constituyen las
llamadas Malvas (géneros Sida, Malvastrum, Walteria y Malacara) consideradas de
gran importancia, tanto para la Paloma Rabiche como para el hombre, ya que
forman parte del espectro alimentario de esta paloma, ocupando uno de los primeros
lugares en el orden de importancia. Todas estas malvas son consideradas especies
de importancia económica por el hombre debido a su forma de colonizar las áreas de
cultivos agrícolas, los gastos y esfuerzos que se requieren para su control.
La familia Poaceae, está representada por nueve especies vegetales, de las que dos
del género Paspalum forman parte del espectro alimentario de la Paloma Rabiche:
Paspalum conjugatum, (Cambute) y Paspalum distichum (Saca sebo), ambas
especies además de ser comunes presentan una alta frecuencia debido a su forma
de distribución uniforme o regular.
La familia Anacardiaceae la componen en el área de estudio solo tres especies:
Comocladia dentata, Spondias mombin Y Spondias sp., ninguna es usada como
alimento por la Paloma Rabiche. Comocladia dentata (Guao prieto), arbusto
frecuente en las áreas de bosques donde bajo determinadas condiciones ecológicas
puede llegar a ser muy abundante y es considerada una especie indeseable por sus
propiedades cáusticas al contacto con la piel de las personas. No forma parte de
espectro alimentario de la especie, pero sus frutos si son comidos por otra tipo de
paloma, la Torcaza Cabeciblanca (Patagioenas leucocephala) en los lugares donde
está habita. El Jobo (Spondias mombin), se considera espontánea en los bosques
naturales y aunque su madera no es de gran de valor económico sus frutos sirven de
alimento a diferentes especies de la fauna silvestre. También es usada para formar
parte de los linderos y por tanto sirve a la Paloma Rabiche como refugio, descanso y
sustrato para la nidificación. Por último Spondias sp (Ciruelón) es una especie de
crecimiento rápido y muy fácil de propagar por estacas por lo que es usada por los

38
campesinos en la confección de linderos, aunque en mucho menor cuantía que las
especies tradicionales (Bursera simaruba, Gliricidia cepium, Erythrina sp. etc.).
La familia Erythroxylaceae representada por dos especies (Erythroxylon havanensis
y Erythroxylon sp.). De ellas la Jíba (Erythroxylon havanensis) es un arbusto que
forma parte de la dieta de la Paloma Rabiche y es el único fruto que fue hallado en
su espectro alimentario. En zonas agrícolas de cultivos intensivos se considera un
arbusto espontáneo no abundante, solo presente en la vegetación secundaria de los
linderos. Algo muy diferente a lo que ocurre en las áreas boscosas donde es muy
abundante y frecuente. Erythroxylon sp. (Arabo) es un arbusto espontaneo en
bosques semicaducifolios sobre suelo calizo que hasta el momento no se ha
reportado ningún usos para le PalomaRabiche.
Figura 12. Comportamiento de la abundancia relativa y la frecuencia de las cinco
familias más representadas en el área de estudio.

39
Tabla 1. Comportamiento de la abundancia relativa y la frecuencia por familias de las
especies vegetales presentes en el área de estudio.
Familia botánica Ar F
Amaranthaceae 0,12 0,41
Anacardiaceae 0,25 0,23
Apocinaceae 0,07 0,2
Asteraceae 0,11 0,09
Boraginaceae 0,04 0,23
Bromeliaceae 0,28 0,12
Burseraceae 0,19 0,21
Cesalpinaceae 0,27 0,14
Compositae 0,25 0,06
Convolvulaceae 0,04 0,2
Cruciferae 0,01 0,01
Cucurbitaceae 0,06 0,14
Cyperaceae 0,14 0,2
Ericaceae 0,02 0,03
Erythroxylaceae 0,25 0,2
Euphorbiaceae 0,05 0,21
Fabaceae 0,04 0,21
Flacourtiaceae 0,2 0,17
Graminaceae 3,14 0,85
Lamiaceae 0,02 0,03
Leguminoceae 0,06 0,51
Malpighiaceae 0,05 0,05
Malvaceae 0,3 0,52
Meliaceae 0,13 0,21
Mimosaceae 0,26 0,32
Myrtaceae 0,04 0,08
Nyctaginaceae 0,01 0,03
Papilonaceae 0,02 0,05
Poaceae 0,25 0,8
Rutaceae 0,21 0,14
Sapindaceae 0,12 0,17
Solanaceae 0,07 0,26
Sterculiaceae 0,14 0,21
Teofrastaceae 0,09 0,12
Tliaceae 0,02 0,05
Unbelliferae 0,07 0,08

40
Verbenaceae 0,12 0,18
Vitaceae 0,05 0,12
Es importante resaltar el comportamiento de ciertas Familias presentes en el área de
estudio: Euphorbiaceae, Papaveraceae, Fabaceae y Graminaceae dentro de las
cuales se hallan especies de gran importancia en el espectro alimentario de la
Paloma Rbiche como la Tua-tuta (Croton lobatus), Lechoza (Euphorbia heterophylla),
el Cardo santo (Argemone mexicana) y el Frijol silvestre (Phaceolus sp.) aunque
estas aparecen con valores no significativos de abundancia y frecuencia a nivel de
área total. Estos resultados muestran comportamientos diferentes al realizar el
análisis individual por biotopo.
Un caso diferente muestra el Arroz (Oriza sativa), importante alimento que puede
llegar a concentrar grandes bandos de palomas en torno a sus áreas de cultivo en
determinadas épocas del año. Se encuentra dentro de la familia más abundante del
área, presenta alta abundancia relativa pero baja frecuencia, dado su carácter de
cultivo muy localizado por sus exigencias ecológicas y por tanto no generalizado en
el área de estudio.
De todas las especies que componen el espectro alimentario de la Paloma Rabiche
la más representada en el área de estudio es Sida rhombifolia (Malva de Puerco),
planta herbácea de la familia Malvaceae. Se encuentra presente en el biotopo
Agrícola con cobertura. Según Sánchez y Uranga (1993) es una planta perenne, que
hace su mayor aparición en los meses de primavera y verano, aunque también se la
observa en las demás épocas del año en sus diferentes estadios. Su propagación se
realiza sólo por semillas, las cuales son diseminadas por animales, equipos
agrícolas, el agua etc. Es común en toda Cuba, presenta gran resistencia y fortaleza
en la competencia por lo que se considera de importancia económica al necesitar
medios y tiempo en su control.
Al realizar el mismo análisis en cada uno de los biotopos representados, aparecen
reflejadas las características de los mismos, observándose notables diferencias.

41
El biotopo agrícola con cobertura cuenta con un total de 84 especies vegetales,
agrupadas en 34 familias botánicas y un total de 60 géneros. En este biotopo al igual
que a nivel de área de estudio existe un total predominio de las especies herbáceas
(70 %), las que constituyen la principal fuente de alimentación de la Paloma Rabiche
(Zenaida macroura L.) en la zona de estudio.
Al observar el comportamiento de la abundancia relativa y la frecuencia con que
aparecen las especies en las diferentes unidades de muestreos podemos apreciar el
dominio de las Garaminaceae, estando presente en este biótopo las mismas once
especies descritas en el área total y predominando igualmente el Andropongon
pertussus. Como es evidente el peso total de la especie vegetal más abundante del
área de estudio lo aporta este biotopo, pues esta especie de gramínea aunque se
propaga de manera natural es un pasto cultivado por el hombre para alimento del
ganado y no está presente en los linderos o en zonas de bosques naturales. En el
caso de la especie Panicum maximun de gran importancia en la alimentación de la
Paloma Rabiche, su número resulta menor que a nivel de área total, debido a que
este pasto aunque se desarrolla a pleno sol, está presente no solo en zonas
agrícolas sino también en lugares de sombra como linderos y áreas de borde de
bosques, claros o parches de bosques ralos.

42
Figura 13. Comportamiento de la abundancia relativa y la frecuencia de las cinco
familias más representadas en el biotopo Agrícola con cobertura.
Le siguen en orden la familia Malvaceae representadas de la misma manera que en
el área total debida que solo se encontró en este biotopo. No existió ejemplar alguno
en el biotopo Forestal típico. Esta familia según Sánchez y Uranga (1993) sólo se
reproduce por semillas la cuales son diseminadas por animales, equipos agrícolas y
el agua por lo que se debe considerar la Paloma Rabiche como una especie
controladora de estas plantas consideradas indeseables por los agricultores.
La familia Poaceae está compuesta en este biotopo por nueve especies todas de
una alta frecuencia debido a su distribución uniforme por todo las áreas cultivadas,
así como las de barbecho. Dentro de esta familia dos especies del género Paspalum
forman parte del espectro alimentario de la Paloma Rabiche, ellas son por su orden
de importancia Paspalum distichum (Saca sebo) y Paspalum conjugatum (Cambute).
Ambas especies son consideradas de importancia económica en la agricultura por el
esfuerzo y gastos necesarios para su control.
La familia Cyperaceae está representada en este biotopo por dos especies
vegetales: Cyperus rotundus (Cebolleta), especie considerada indeseable en los

43
cultivos agrícolas donde es común encontrarla formando en determinados lugares
grupos muy densos, pero su distribución es agregada y por tanto; presenta una baja
frecuencia en las unidades de muestreos. Esta especie forma parte del espectro
alimentario de la Paloma Rabiche y constituye el único artículo alimentario que es
consumido en forma de bulbo. La hierba Cortadera (Cladium jamaicense) es una
especie que requiere zonas con una alta humedad por lo que está muy asociada en
el país al cultivo del Arroz. Por estas características presenta al igual que la
Cebolleta una distribución agregada y por tanto baja frecuencia. De manera similar
que el Arroz está presente en el espectro alimentario de la Paloma Rabiche. Es
conocido en el “mundo” de los cazadores que en áreas donde existen arrozales y
junto a estos la Cortadera, constituyen excelentes lugares donde lograr “buenas
perchas” de Paloma Rabiche, por la atracción que provocan estas especies en está
paloma.
La familia Mimosaceae, representada en este biotopo por cuatro especies vegetales
de las cuales solo una no se reporta como de uso por la Paloma Rabiche, la Mimosa
asperata (Aroma Weiler). Esta especie está asociada a terrenos con un alto
contenido de humedad o con inundaciones temporales. El resto lo constituyen
especies de importancia ecológica para la paloma estudiada. El Marabú y la Aroma
blanca están consideradas especies invasoras en la agricultura cubana y ocupan
diferentes hábitats invadidos como potreros, realengos y bordes de caminos rurales,
lugares donde es común encontrar la especie anidando. La otra especie presente
en esta familia es Samanea saman (Algarrobo del país) de amplia distribución, pero
con pocos representantes. Aparece en las áreas agrícolas como árboles dispersos o
formando parte de los linderos donde se considera una especie espontánea ya que
no es utilizada por los campesinos para formar parte de estos linderos; constituye
una importante especie usada tanto para el descanso como para refugio y
nidificación.

44
Tabla 2. Comportamiento de la abundancia relativa y la frecuencia por familias de las
especies vegetales presentes en el biotopo Agrícola con cobertura.
Familia botánica Ar F
Amaranthaceae 0,05 0,36
Anacardiaceae 0,03 0,11
Apocinaceae 0,02 0,12
Asteraceae 0,11 0,12
Bromeliaceae 0,05 0,05
Burceraceae 0,06 0,1
Compositae 0,02 0,12
Convolvulaceae 0,03 0,11
Crusiferaceae 0,02 0,03
Cucurvitaceae 0,02 0,07
Cyperaceae 0,16 0,23
Erythrxylaceae 0,03 0,07
Euphorbiaceae 0,06 0,25
Fabaceae 0,04 0,16
Flacourtiaceae 0,02 0,05
Graminaceae 3,94 0,89
Lamiaceae 0,03 0,04
Leguminoceae 0,46 0,07
Malvaceae 0,91 0,61
Meliaceae 0,06 0,11
Mimosaceae 0,23 0,21
Myrtaceae 0,02 0,02
Papilonaceae 0,03 0,05
Poaceae 1,12 0,57
Portulacaceae 0,03 0,12
Rubiaceae 0,04 0,11
Sapindaceae 0,02 0,03
Sapotaceae 0,03 0,07
Solanacea 0,03 0,12
Sterculiaceae 0,03 0,11
Teofrastaceae 0,02 0,02
Tiliaceae 0,02 0,05
Vervenaceae 0,04 0,14
vitaceae 0,02 0,03

45
A pesar de existir un claro predominio de las plantas herbáceas, también se
encuentra un considerable número de especies arbóreas y arbustivas (Anexo 2), las
que se ubican en linderos que bordean los caminos rurales y las cercas perimetrales
presentes en el área y en forma de árboles aislados en las áreas de potreros que
sirven a esta especie como percha para el descanso, refugio y sustrato para la
nidificación.
Todas estas características se traducen en una excelente calidad de hábitat para la
población de la especie estudiada. Este hábitat ocupa el mayor por ciento (65 %) del
área de estudio. Estos resultados son consistentes con los obtenidos por Cuesta
(2009) donde al clasificar las áreas de estudio de Zenaida macroura en la provincia
de Pinar del Río le asigna a las zonas de cultivos agrícolas con ganadería como
actividad económica secundaria una calidad de sitio alta.
Ostrand, et al. (1996) al estudiar la selección del hábitat de alimentación de Zenaida
macroura con un enfoque en las diferentes estructuras de los sitios de alimentación,
concluyeron que esta paloma basa su selección de hábitat de forrajeo en la altura de
las hierbas y la densidad de estas.
Un caso diferente se refleja en el resultado obtenido para el biotopo Forestal típico.
Aquí existe una marcada desproporción en los valores porcentuales en comparación
con el biotopo agrícola con cobertura que favorece a las especies arbóreas.
El biotopo Forestal típico cuenta con un total de 29 familias botánicas, 39 géneros y
44 especies vegetales. Al realizar el análisis del comportamiento de las diferentes
familias presentes en el biotopo con respecto a los valores de abundancia relativa y
la frecuencia en que se encuentran distribuidas en el , se observó un
comportamiento diferente al agrícola con cobertura: de las cinco familias más
abundantes, en las cuatro primeras no se encontraron ejemplares de especies
vegetales herbáceas, y en la de menor representación solo existió una especie
herbácea de las dos de esta familia, la Cassia occidentalis, que solo ocupó el 30 %
del total de ejemplares hallados. Otro aspecto que reafirma el peso de estas familias
arbóreas en este biotopo es el hecho de que en cuatro de las cinco familias aquí

46
descritas la frecuencia obtuvo el máximo valor (1), porque en cada trancecto siempre
estuvo presente algún miembro de estas.
Figura 14. Comportamiento de la abundancia relativa y la frecuencia de las cinco
familias más representadas en el biotopo Forestal típico.
La familia con mayor representación en este biotopo resultó ser la Mimosaceae, y
dentro de esta la especie de mayor número, fue Samanea saman (Algarrobo del
país). Especie muy común en los bosques semicaducifolios de todo el país, presenta
una amplia distribución por todo el biotopo, pero también aunque en menor cuantía
puede hallarse en el biotopo Agrícola con cobertura. Logra alcanzar una gran altura y
es muy usada por la avifauna de los lugares donde habita. La Paloma Rabiche la
emplea como sustrato para nidificar, aunque no es la más usada por la especie para
este fin. No se encuentra dentro del espectro alimentario de la especie.
Le sigue en orden de importancia de acuerdo a su número la familia Anacardiaceae,
representada en este biotopo solo por dos especies vegetales: Spondia mombin
(Jobo) y Comocladia dentata (Guao prieto). Ninguna de las dos especies se

47
encuentra dentro del espectro alimentario de la Paloma Rabiche. Ambas se
consideran especies comunes en este tipo de vegetación y en el caso del Jobo
puede ser usada como sustrato para la nidificación.
La familia Erythroxylaceae se encuentra representada en este biotopo por dos
especies: Erythroxylum havanensis (Jibá), especie arbustiva común en los bosques
semicaducifolios del país. Forma parte del espectro alimentario de la Paloma
Rabiche y también es común en los linderos que forman parte de las zonas
agrícolas. La otra especie vegetal que representa esta familia en este biotopo es
Erythroxylum sp. (Arabo); especie espontánea y muy común en los bosques
semicaducifolios de todo el país, no forma parte del espectro alimentario de la
Paloma Rabiche ni ha sido reportado algún uso para la especie.
La familia Burseraceae está representada en el biotopo por la especie Bursera
simaruba (Almacigo). Árbol común en todo el biotopo, no forma parte del espectro
alimentario de la especie pero le sirve como refugio y descanso; cuando este árbol
se encuentra en el borde de las áreas boscosas es frecuente observar las Palomas
posadas en él. También puede ser usado como sustrato para la nidificación de la
especie. Constituye una especie muy empleada por los campesinos en la
construcción de los linderos.
Por último la familia Sterculiaceae se encuentra representada por una especie
vegetal arbórea, Guazuma tomentosa (Guásima), árbol muy común en todo el país,
tanto en bosques como en áreas agrícolas. Según Roig (1962) se distribuye en toda
la isla, y tal vez la más común y la más ampliamente distribuida especie cubana,
crece en toda clase de terreno. Su fruto constituye un alimento para cerdos y ganado
vacuno y equino. Hasta la fecha no se ha reportado su uso como alimento para la
Paloma Rabiche. Al estar presente de manera espontánea en linderos y en forma de
árbol aislado en zonas agrícolas sirve de refugio, descanso y sustrato para la
nidificación a está paloma.

48
Tabla 3. Comportamiento de la abundancia relativa y la frecuencia por familias de las
especies vegetales presentes en el biotopo Forestal típico.
Familia botánica Ar F
Amaranthaceae 1,2 0,78
Anacardiaceae 2,84 1
Apocinaceae 0,91 0,67
Boraginaceae 0,58 0,89
Bromeliaceae 2,25 0,78
Burseraceae 2 1
Cesalpinaceae 1,1 0,78
Compositae 0,63 0,33
Convolvulaceae 0,43 0,78
Cucurbitaceae 0,64 0,55
Ericaceae 0,17 0,22
Erythroxylaceae 2,5 1
Fabaceae 0,36 0,44
Flacourtiaceae 0,84 1
Graminaceae 0,64 0,55
Leguminoceae 0,38 0,55
Malpighiaceae 0,37 0,33
Meliaceae 0,8 0,89
Mimosaceae 4,2 1
Myrtaceae 0,33 0,44 Nyctaginaceae 0,11 0,22
Rutaceae 1,01 1
Sapindaceae 1,11 1
Sapotaceae 1,11 0,67
Sterculiaceae 1,6 0,9
Teofrastaceae 0,77 0,77
Unbelliferae 0,51 0,5
Verbenaceae 1 0,44
Vitaceae 0,42 0,67
Toda esta situación se traduce en un biotopo de baja calidad para la especie
estudiada, dado su pobre potencial de alimentación, así como la propia estructura de
la vegetación. Estos resultados son consistentes con los obtenidos por Ostrand et al.
(1998), al estudiar los cambios en el uso de la tierra como un posible factor en la
disminución de las poblaciones de la Paloma Rabiche en la parte central de Utha,
USA. Ellos plantean como principales factores: la disminución de los campos

49
cultivados de trigo durante la primavera, áreas donde se alimenta principalmente la
especie, además de usar también los comederos de ganados y parches con
yerbajos. Esta afirmación se demuestra en los resultados de los análisis del
comportamiento de las densidades de especies vegetales que sirven de alimento a
la Paloma Rabiche en el área de estudio (tabla 1); por tal razón debe tenerse en
cuenta en el momento de una planificación en función de su aprovechamiento.
Tabla 4. Porcentaje de especies vegetales disponibles por biotopos y su
representación en la alimentación de la Paloma Rabiche.
Biotopos
%
Herbácea
s
%
Arbustivas
%
Arbóreas
%
Trepadoras
%
Epifitas
Total
No
Sp.
% Sp
Aliment
o
paloma
Agrícola
con cobertura 70 13 11 6 0 84 25
Forestal
típico 16 32 36 11 5 44 5
Total Área
estudio 57 17 18 6 2 101 21
*los porcentajes de cada biotopo no necesariamente tienen que coincidir con el del área total porque existen
especies repetidas y otras que no aparecen en un biotopo, pero está presente en el área total.
En general, de lo antes dicho y del análisis de la Tabla 1 se evidencia que en el área
de estudio la flora en general ofrece excelentes condiciones para el desarrollo de la
Paloma Rabiche. La anterior afirmación se fundamenta porque predominaron en
toda el área de estudio las especies herbáceas, que constituyen el mayor por ciento
de la dieta de la especie estudiada, y por la dominancia del biotopo Agrícola con
cobertura (65 %) respecto al forestal típico (10 %), ya que el resto corresponde a
viales rurales y áreas urbanas.
Como se aprecia en la figura 15 en el biotopo Agrícola con cobertura el 54 % de las
especies vegetales del área de estudio están clasificadas como muy abundantes en
este biotopo, representadas aquí por especies herbáceas en su gran mayoría, las
que constituyen el mayor por ciento de la dieta de la especie estudiada, y solo el 18

50
% son escasas y un 15 % ausentes, referidas en ambos casos a especies arbóreas
que su presencia está asociada a linderos o en forma de árboles aislados en
potreros.
Figura 15. Comportamiento de la abundancia relativa de la flora del área de estudio
en el biotopo Agrícola con cobertura.
La figura 16 muestra el comportamiento de la abundancia relativa de la flora en el
biotopo Forestal típico, donde el mayor porcentaje correspondió a las especies
ausentes debido al gran número de herbáceas inventariadas en el área total, sin
embargo están muy poco representadas en este biotopo; resultados que pudiesen
estar condicionados por las exigencias ecológicas de dichas especie.
Es importante destacar que los términos ausentes, escasas, abundantes y muy
abundantes antes mencionados, hacen referencia a su condición o número de
ejemplares descritos solo dentro de los diferentes biotopos del área de estudio, por
tanto no se refieren a que estén reportadas en la actualidad con este status en el
país.

51
Figura 16. Comportamiento de la abundancia relativa de la flora del área de estudio
en el biotopo Forestal típico.
Los índices de Diversidad de Shannon (H`), Riqueza de Especies de Margalef (M) y
Equitatividad (J), se encuentran dentro de los principales indicadores a tener en
cuenta en los estudios de comunidades tanto vegetales como animales ya que
expresan de conjunto los valores de la biodiversidad presentes, y por tanto, permiten
la caracterización de los biotopos presentes.
Los valores de H´ y M se comportan de forma bastante similar en ambos biotopos,
pero con una ligera superioridad en el Forestal Típico lo que está dado por el mayor
número de especies forestales que lo forman.
La Equitatividad (J) refleja la distribución de los individuos por especies (Tabla 1), al
comparar ambos biotopos se aprecia el mayor valor en el Forestal Típico (0,885) que
en el Agrícola con Cobertura (0,797). Es importante destacar que esta pequeña
diferencia permite deducir que las especies vegetales están distribuidas de forma
bastante similar en ambos biotopos, ya que cuando los valores obtenidos son
superiores a 0,5 esto indica la no existencia de una alta dominancia por parte de
ninguna de las especies presentes.

52
Tabla 5. Comportamiento de los principales Índices de diversidad biológica usados
en el área de estudio.
Biotopos Diversidad
de Shannon (H')
Riqueza de
especies
(M)
Equitatividad
(J')
Agrícola con Cobertura 1,463 23,948 0,797
Forestal típico 1,466 32,064 0,885
3.4.- Conclusiones y recomendaciones
1. De los dos biotopos presentes en el área de estudio (Agrícola con cobertura y
Forestal típico), el de mayor calidad para la especie estudiada es el agrícola con
cobertura que ocupa además, el 65 % del territorio estudiado.
2. En el agrícola con cobertura a pesar de existir un elevado número de especies
arbóreas y arbustivas, predominan las especies herbáceas (70 %), las que
constituyen la principal fuente de alimentación de la Paloma Rabiche (Zenaida
macroura L.). Todas estas características se traducen en una excelente calidad
de hábitat para la población de la especie estudiada.
3. La presencia de linderos en el biotopo clasificado como agrícola con cobertura es
un factor importante y favorece el uso de los mismos por la especie, pues ofrece
condiciones de sombra, refugio y perchas para el descanso de las aves.
4. Se recomienda determinar, a través de una serie de parcelas (permanentes y no
permanentes), la dinámica de la vegetación herbácea en los biotopos presentes
en el área por ser la principal fuente de alimentos para la especie.

53
3.5.- Bibliografía
Bautista, L. (2013). Manual de plantas útiles para las aves en la ciudad de
Querétaro. Universidad Autónoma de Querétaro. Primea Edición. México. 64 pp.
Bisse, J. (1998). Árboles de Cuba. Editora Científico–Técnica. Ciudad de la Habana.
384 pp.
Campo, A. M. y Soledad, V. D. (2014). Diversidad y valor de importancia para la
conservación de la vegetación natural. Parque Nacional Lihué Calel. Argentina.
Revista Anales de Geografía, 34 (2), 25-42.
Carrascal, L. M. y Telleria, J. L. (1998). Relación entre avifauna y estructura de la
vegetación en los medios agrícolas del norte de la Península Ibérica (País Vasco
atlántico). MUNIBE (Ciencias Naturales). 40. 9-17.
Cuesta, L. A. (2009): Perspectivas hacia una Gestión Sostenible de la Actividad
Cinegética en la Provincia de Pinar del Río, Cuba. Universidad de Alicante,
España, Universidad de Pinar del Río, Cuba.
Chamizo, R. (2001). Análisis de Sostenibilidad de la Actividad Cinegética en Cuba.
Tesis Doctoral. Universidad de Alicante, España, Universidad de Pinar del Río,
Cuba.
Gunn C. R. & C. A. Ritchie. 1988. Identification of disseminules listed in the Federal
Noxious Weed Act. U. S. Department of Agriculture, Technical Bulletin No. 1719.
313 pp.
León, H. y Alain, H. (1951). Flora de Cuba. Vol. II. Contr. Mus. Hist. Nat. Colegio de
La Salle, No. 10, Imp. P. Fernández y Cía, La Habana. 456 pp.
León, H. y Alain, H. (1953). Flora de Cuba. Vol. III.Contr. Mus. Hist. Nat. Colegio de
La Salle, No.13, Imp. P. Fernández y Cía, La Habana. 502 pp.
León, H. y Alain, H. (1957). Flora de Cuba. Vol.IV. Contr. Mus. Hist. Nat. Colegio de
La Salle, No. 16, Imp. P. Fernández y Cía, La Habana. 556 pp.

54
Magurran, A. (1988). Ecological diversity and its meansurements. Princeton
University Press. New Jersey. U.S.A. 179 pp.
Marquez, M., García, E., González, C. y Tarango, L. (2005). Composición de la dieta
del guajalote silvestre (Meleagris gallopavo mexicana, Gould, 1856) reintroducido
en “Sierra Fría”, Aguascalientes, México. Veterinaria México, 36 (4). 395-409.
Moreno, C. E. (2001). Métodos para medir la biodiversidad. M&T–Manuales y Tesis
SEA, vol. 1. Zaragoza, España. 84 pp.
Mostacedo, B; Fredericksen, T. S. (2000). Manual de métodos básicos de muestreo
y análisis en ecología Vegetal. Santa Cruz, Bolivia. 10-12.
Ostrand, W. D., Bissonette, J. A. y Conover, M. R. (1996). Selection of Foraging
Habitat by Mourning Doves: a structural approach. Journal of Field Ornithology,
67 (4), 691-697.
Ostrand, W. D., Meyers, P. M., Bissonette, J. A. y Conover, M. R. (1998). Changes in
land use as posible factor in Mourning Dove population decline in Central Utah.
Journal of Field Ornithology, 69 (2), 192- 200.
Ramírez, J. Ramírez, M. (2002). Avifauna de la región de la sierra de Huautla,
Morelos, México.
Anales del instituto de Biología, Universidad nacional autónoma de México, Serie
Zoología, 73 (1). 91-111.
Rodríguez, S., Rodríguez, J. y Pérez, L. (1988). Plantas indeseables en el cultivo de
la caña de azúcar. Editora Científico–Técnica. Ciudad de la Habana. 196 pp.
ROIG, J. T. (1962). Diccionario botánico de nombres vulgares cubanos. Editorial
Nacional de Cuba, Editora del Consejo Nacional de Universidades. La
Habana. 1t. 1142 pp.
Sánchez, P. y Uranga, H. (1993). Plantas indeseables de importancia económica en
los cultivos tropicales. Editora Científico-Técnica. Ciudad de la Habana. 166 pp.

55
Zaragoza, C. et al. (2004). Composición de la dieta de la Codorníz Moctezuma
(Cyrtonyx montezumae) en el noreste del estado de México, México, Veterinaria
México.35 (003). 215-236..


56
CAPÍTULO IV. LA PALOMA RABICHE EN LA COMUNIDAD ORNITÍCA
4.1.- Introducción
La fauna silvestre es un recurso natural que pertenece a todos y que nos legaron
nuestros antepasados con la idea que este era inagotable y por tanto su uso podía
ser sin control e ilimitado. Actualmente está demostrado que aunque esta sea
renovable sin un uso ordenado la perderemos para siempre con todas las
consecuencias colaterales que esta situación traería aparejada. Es por tanto
obligación nuestra legarle a las nuevas generaciones no solo una amplia gama de
especies para su gestión y aprovechamiento, sino la convicción que solo un uso
sostenible de este recurso siga siendo fuente inagotable de riqueza.
Según Gallina y López (2011) por muchas décadas se consideró a la fauna como un
recurso natural inagotable, el cual era explotado de una forma irracional al grado que
llegaron a extinguirse especies, y muchas se encuentran actualmente en peligro de
extinción. En esta situación incidieron factores como la intensa persecución que el
hombre les dio con fines diversos: para comida, como fuente de comercio y
básicamente por la destrucción de sus hábitats. Sin embargo, el criterio de recurso
natural inagotable ha cambiado y afortunadamente en la actualidad sabemos que la
fauna debe ser explotada en forma racional y se le está dando la importancia
debida, al menos reconocemos en nuestros días que los animales silvestres tienen
valor desde muchos puntos de vista como el estético, económico, comercial y
cinegético entre otros.
La conservación de la biodiversidad no solo implica la protección estricta del recurso
en las áreas naturales, sino también promover su conocimiento y cuidado por parte
de todos. Para llevar a cabo una conservación efectiva, debemos conocer a
profundidad que tenemos en nuestras áreas en términos de especies, número de
individuos, dinámica de sus poblaciones, relaciones entre ellas, además de otros
factores que puedan afectar su sostenibilidad a largo plazo (Acosta, Mugica y
Aguilar, 2013).

57
Los estudios sobre la fauna silvestre constituyen la base principal para la
conservación de este recurso natural. Uno de los pasos esenciales para el
conocimiento de la fauna en el territorio es realizar su inventario.
El objetivo de este capítulo es caracterizar la avifauna del área de estudio a través
de la dinámica poblacional, grado de utilización del hábitat y la forma de
distribución de ésta en el área de estudio. Este objetivo permitirá conocer la
importancia ecológica que ocupa la Paloma Rabiche dentro de la comunidad ornítica
de Cuba.
4.2.- Materiales y métodos
Los conteos de la avifauna local, se realizaron mediante un total de 11 trancectos,
siguiendo el método de trancectos lineales fijos propuesto por Jarvinen y Väisänen
en López (1989), con un ancho de 50 m y una longitud de 500 m, distribuidos al azar
en la zona de estudio. Los conteos fueron efectuados en los periodos de mayo-
agosto y diciembre-febrero, a fin de incluir la etapa principal de reproducción y
obtener mayor información sobre la dinámica de reproducción de la especie
estudiada y la temporada de caza, coincidente con la época de migración de las
aves.
La determinación del número de muestras adecuado para el estudio, se efectuó a
través del método de la curva especie-área, la cual permite visualizar la
representatividad de un muestreo. Cuando la curva tiende a mantenerse horizontal,
indica que el número de especies se mantendrá constante aunque aumente el
tamaño de muestreo (Mostacedo y Fredericksen, 2000).
La identificación de las diferentes especies de la avifauna fue realizada con la ayuda
del manual de Garrido y Kirkconnell (2010), y el catálogo de Garrido y García
(1975).
Se determinó la Abundancia relativa de las aves (AR), donde:
AR = ni / u
ni representa el número promedio de individuos de la especie i

58
u representa la unidad de medida utilizada (hectáreas)
Se determinó la frecuencia (F) con que una especie de ave es detectada a través de
la siguiente expresión:
F = ni / N
ni representa el número de unidades de muestreos en que ha sido detectada la
especie i
N representa en número total de unidades (trancectos)
Se establecieron los grados de fidelidad que señalan la intensidad o frecuencia con
que la especie se encuentra incluida en las unidades ambientales de los biotopos
presentes en el área de estudio y se expresaron en función de los valores de
ocurrencia según la siguiente expresión (Beltzer 1984; 1987, citado por Belzer,
1990):
Fo = Xe x 100 / Xt
donde:
Xe representa el número de observaciones con presencia de la especie en estudio
Xt representa el número total de observaciones
4.3.- Resultados y discusión
La composición de especies en un ecosistema depende de factores que actúan a
nivel espacial y temporal. En las aves, la riqueza y diversidad ha sido correlacionada
con el clima y la heterogeneidad del paisaje, sugiriéndose que tales factores
influyen con diferente magnitud en relación a su escala de análisis (Bohning-
Gaese, 1997; Cueto y de Casenave, 1999; Gillespie y Walter, 2001 en Solis y
Ena, 2008). En este sentido, el clima tendría un efecto muy marcado a una
escala macrogeográfica mientras que la heterogeneidad del paisaje actuaría con
una mayor incidencia a nivel local (Bohning-Gaese, 1997; Cueto y Casenave, 1999
en Solis y Ena, 2008).

59
En la figura 17 se puede apreciar el comportamiento de la curva especie – área,
evidenciándose que con un total de 8 trancectos la curva tendió a mantenerse
horizontal, indicativo de la no incorporación de nuevas especies aunque se
aumentase el tamaño de muestra y por tanto representativo de las especies de
avifauna presentes en el área de estudio, así como su distribución en los diferentes
biotopos.
Figura 17. Comportamiento de la relación aves -unidades de muestreo en el área de
estudio.
En el Anexo 3 se presenta el listado de especies de la avifauna en el territorio y su
clasificación.
Un descriptor que permite conocer el grado de utilización que realiza una especie de
un determinado hábitat es la abundancia relativa (AR), este parámetro por su gran
sencillez ha sido ampliamente utilizado en muchos estudios ecológicos. Su
determinación se basa en la relación existente entre el número de individuos
promedio de cada especie observado u oída y la unidad de medida utilizada (tiempo,
longitud, área etc.).
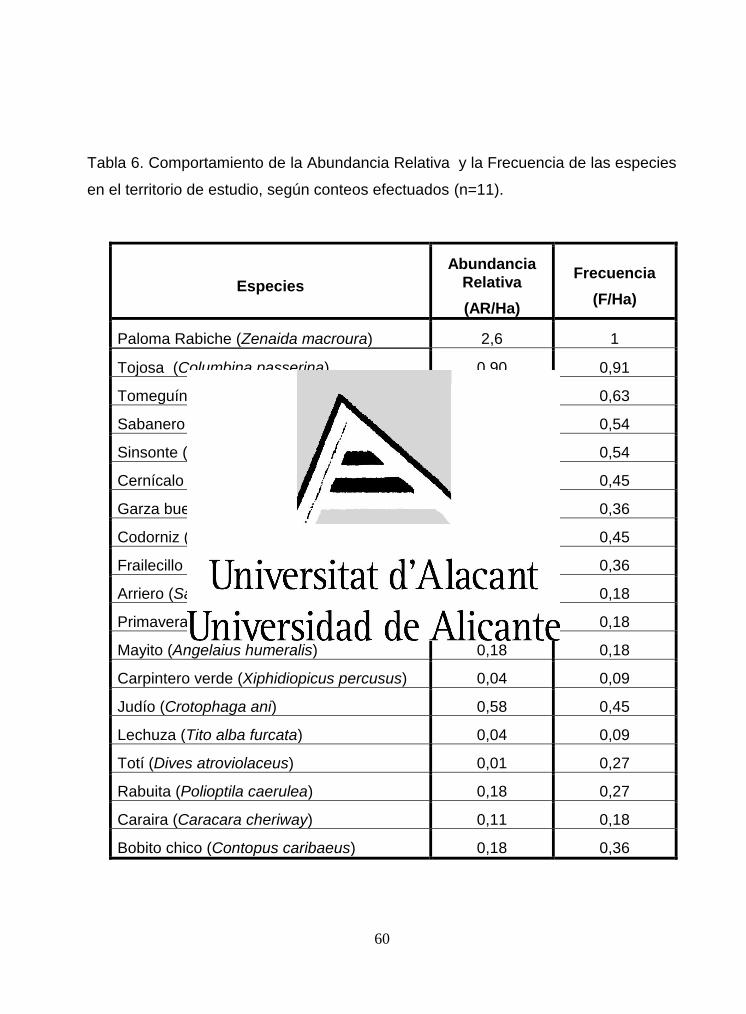
60
Tabla 6. Comportamiento de la Abundancia Relativa y la Frecuencia de las especies
en el territorio de estudio, según conteos efectuados (n=11).
Especies
Abundancia
Relativa
(AR/Ha)
Frecuencia
(F/Ha)
Paloma Rabiche (Zenaida macroura) 2,6 1
Tojosa (Columbina passerina) 0,90 0,91
Tomeguín de la tierra (Tiaris olivácea) 0,83 0,63
Sabanero (Sturnella magna) 0,36 0,54
Sinsonte (Mimus polyglottos) 0,44 0,54
Cernícalo (Falco sparverius) 0,22 0,45
Garza bueyera (Bubulcus ibis) 0,76 0,36
Codorniz (Colinus virginianus) 0,94 0,45
Frailecillo (Charadrius vociferus) 0,29 0,36
Arriero (Sauropthera merlini) 0,14 0,18
Primavera (Coccyzus americanus) 0,07 0,18
Mayito (Angelaius humeralis) 0,18 0,18
Carpintero verde (Xiphidiopicus percusus) 0,04 0,09
Judío (Crotophaga ani) 0,58 0,45
Lechuza (Tito alba furcata) 0,04 0,09
Totí (Dives atroviolaceus) 0,01 0,27
Rabuita (Polioptila caerulea) 0,18 0,27
Caraira (Caracara cheriway) 0,11 0,18
Bobito chico (Contopus caribaeus) 0,18 0,36

61
La tabla 6, presenta los resultados de los conteos efectuados en el área de estudio
(n = 11). Los valores de la AR evidencian el elevado grado de utilización por la
Paloma Rabiche del biotopo predominante en el área de estudio (Agrícola con
cobertura). La abundancia relativa muestra un valor bastante alto (2,6), por lo que
se estima una buena abundancia real (casi tres palomas por hectárea), en
comparación con las otras especies presentes en la zona.
El cálculo de la frecuencia de las especies encontradas al realizar los conteos de
fauna a pesar de ser de aplicación simple, puede cumplir con los requerimientos de
muchos estudios ecológicos (Acosta et al., 2013), esta frecuencia al estar
estrechamente relacionada con la abundancia relativa, de esta relación o
comportamiento entre ambas es posible conocer el tipo de distribución que
presentan las especies inventariadas.
En la figura 18, se muestra el comportamiento de la abundancia relativa y la
frecuencia de las especies del área de estudio, es evidente, además el alto grado de
utilización que hace la Paloma Rabiche del área de estudio. Se refleja además, que
la misma mantiene una distribución uniforme ya que presenta un elevado valor de
frecuencia (dado por su presencia en la mayoría de los conteos realizados), lo cual
está relacionado con sus hábitos. Según refieren Acosta y Ponce de León (2004),
en Cuba esta especie es frecuente en potreros y terrenos llanos asociados a
parches de bosques, campos cultivados. Con esa misma distribución también se
hallan la Tojosa (Columbina passerina) y el Tomeguín de la tierra (Tiaris olivácea).
Un caso diferente se puede observar en los valores de la Garza bueyera (Bubulcus
ibis) con una alta abundancia relativa, pero una baja frecuencia, revelando una forma
de distribución agrupada, dada por sus hábitos gregarios. Otro comportamiento
reflejado son los casos de la Lechuza (Tito alba furcata), Carpintero verde
(Xiphidiopicus percusus) y la Primavera (Coccyzus americanus) ejemplares poco
comunes y de hábitos solitarios, por lo que aparecen con valores muy bajos de
abundancia relativa y frecuencia.

62
Figura 18. Comportamiento de los valores de Abundancia Relativa y Frecuencia de
las especies de aves en el área de estudio.
Conocer los niveles de asociación de las especies con su hábitat es un aspecto
importante de su ecología, ya que se refiere a su nicho estructural y brinda
información básica sobre el comportamiento de la especie y sus requerimientos
naturales. Determinar estos niveles de asociación permitiría establecer manejos
correctos para un aprovechamiento sostenible, tanto de la Paloma Rabiche como de
su hábitat.
La figura 19, ofrece los resultados obtenidos en función de los valores de ocurrencia,
donde se aprecia el uso que hace la especie estudiada de las áreas de cultivos
agrícolas (40 %) y las áreas ganaderas (35 %), la cual está dada por la gran
disponibilidad de alimentos y las características de estas áreas (zonas abiertas, con
árboles dispersos y en ciertos lugares parches de suelo desnudos). El 22 % de las
observaciones corresponden a los linderos o setos, estos presentan una
combinación de vegetación arbórea y arbustiva, factor que favorece el uso de sitios
de alimentación por la especie que encuentra perchas y sombra para el descanso,

63
mientras que en último lugar de preferencia con solo un 3 % están las áreas de
bosques, donde escasean las especies herbáceas que constituyen el 79 % de la
dieta de la especie. Estos resultados apoyan la descripción de los hábitos de esta
especie descritos por Gundlash (1876): “Prefiere los árboles aislados en potreros y
sabanas y las arboledas, etc., pero no los bosques cerrados, y busca su comida en
el suelo y en los campos recién arados”.
Los resultados obtenidos coinciden con lo expuesto por Solis y Ena (2008), la
disposición espacial de aves dentro de un hábitat (nivel local) está
determinado por las características fisonómicas del hábitat (abierto o cerrado,
plano u ondulatorio, continuo o dividido en zonas). Sin embargo, esta
distribución está estrechamente ligada con la conducta de las especies, ya que
existen aves que presentan una conducta de selección de hábitat fija, es decir
son especialistas de hábitat, encontrándolas por lo tanto, sólo en hábitats
específicos y sucede lo contrario con las especies generalistas de hábitats que
no presentan una dependencia estricta por un tipo de hábitat, utilizando más
de uno, siempre y cuando estos les proporcionen los recursos necesarios para
su supervivencia (alimento y sitios de nidificación).

64
Figura 19. Grados de fidelidad, dado por los valores de ocurrencia para Zenaida
macroura en las unidades del paisaje agrario del área de estudio. 1= cultivos
agrícolas, 2= linderos o setos, 3= potreros, 4= bosque. Los valores señalados por las
flechas indican los porcentajes obtenidos.
4.4.- Conclusiones y recomendaciones
1. La Paloma Rabiche resultó ser la especie de mayor frecuencia de observación y
mayor densidad poblacional en la zona de estudio.
2. La Paloma Rabiche presentó una distribución agregada en el biotopo Agrícola
con cobertura, no ocurriendo igual en el biotopo Forestal típico, donde presentó
una menor frecuencia determinada por el tipo de densidad y vegetación existente
en cada área.
3. Dentro del biotopo agrícola con cobertura, en función de la composición y
distribución de la vegetación se manifiestan diferencias en cuanto a la densidad
de la especie estudiada, debido esencialmente, a la disponibilidad de alimentos.

65
4. Los cultivos agrícolas obtuvieron los mayores valores de ocurrencia, seguidos por
las áreas de potreros y linderos, siendo menos frecuente en las áreas boscosas.
5. Se recomienda continuar con los estudios de la dinámica de la especie en los
diferentes hábitats teniendo en cuenta que la estructura de los hábitat agrícolas
está en constante modificaciones por el hombre y este factor pudiese variar a su
vez la estructura de las comunidades de aves; de esta manera se podrá predecir
con mayor confiabilidad la presencia de la especie y lograr un manejo más eficaz
4.4.- Bibliografía
Acosta, M., Mugica, L. y Aguilar, S. (2013). Protocolo para el monitoreo de aves
acuáticas y marinas. Centro Nacional de Áreas Protegidas. La Habana. Cuba.
142 pp.
Acosta, M. y P. de león, J.L. (2004). Historia Natural de la Ploma Rabiche (Zenaida
macroura). (Inedito). Facultad de Biología. Universidad de la Habana. Cuba.
Beltzer, A. H. (1990). Biología alimentaria del Gavilán común Buteo magnirostris
(aves: Accipitridae) en el valle aluvial del río Paraná medio, Argentina.
Ornitología Neotropical, 1 (1-2), 3-8.
Gallina, S. y López, C. (2011). Manual de técnicas para el estudio de la fauna.
Volumen I. universidad Autónoma de Querétaro–Instituto de ecología. México.
377 pp.
Garrido, O. y García, H. (1975). Catálogo de las aves de Cuba. La Habana.
Academia de Ciencias. 149 pp.
Garrido, O. y Kirkconnell, A. (2010). Aves de Cuba. Edición Comstock Publishing
Associates, Cornell University Press. USA. 287 pp.
Gundlash, J. (1876). Contribución a la ornitología cubana. Editora La antillana. La
Habana. 139 – 140.
Mostacedo, B; Fredericksen, T. S. (2000). Manual de métodos básicos de muestreo
y análisis en ecología Vegetal. Santa Cruz, Bolivia. 10-12.

66
Solis, P. y Ena, E. (2008). Relación entre la estructura del hábitat y la comunidad de
aves en el fundo agroecológico Huaquina, Chincha- Perú. (Abril-Septiembre
2007). Tesis para optar el título de: Biólogo. Universidad Nacional “San Luís
Gonzaga” de ICA. Facultad de Ciencias, Escuela Académico Profesional de
Biología. Perú.


67
CAPÍTULO V. REPRODUCCIÓN Y CONDUCTA REPRODUCTIVA DE LA
PALOMA RABICHE (Zenaida macroura L.)
5.1.- Introducción
La conducta de los animales está marcada por diferentes variables tanto de carácter
climático como de su propia biología. En cada especie una particularidad muy
singular resultan ser los patrones conductuales que estas desarrollan. En particular
los patrones reproductivos en las aves, pudiesen estar relacionados con factores
como la abundancia de alimentos, las condiciones climáticas presentes, la
selección de áreas de nidificación en lugares que brinden una mayor protección ante
depredadores, etc.
De acuerdo con Berovides (1988), al comportamiento de los animales en
determinadas actividades que realizan con secuencia en el tiempo se denomina
conducta. Dicha conducta es parte de su fenotipo, es decir del conjunto de todas sus
características morfológicas, fisiológicas, visibles o no, que son controladas por el
genotipo y el ambiente. La conducta se hace evidente a través de diferentes
manifestaciones, a las que se les llama Patrones de Conducta animal.
Estos patrones han sido agrupados en tres grandes grupos (Opus cit.):
Patrones de Conducta Individual
Patrones de Conducta Social
Patrones de Conducta reproductiva
Los Patrones de conducta Reproductiva, a diferencia de todos los demás están
relacionados no con la supervivencia del individuo o del grupo, sino con la
supervivencia de la especie (Opus cit.). “La reproducción es probablemente la más
notable y la más característica actividad de los sistemas vivientes. Fácilmente se
comprende la eficacia de la reproducción como mecanismo de autoperpetuación”
(Weisz, 1968).

68
Según Yarrow (2009) la época de reproducción en la Paloma Rabiche, comienza
con el cortejo, en la llegada de la primavera, cuando el macho se eleva en el aire
con aleteos lentos y laboriosos que le pueden llevar a alcanzar 100 pies o más. Al
llegar a la cúspide de su vuelo, extiende sus alas y se desliza hacia la tierra en un
planeo en círculos. Sobre el terreno, se pavonea con movimientos como asintiendo
con la cabeza, pronunciando una serie de llamadas a su aspirante a compañera. En
Carolina del Sur, este comportamiento de cortejo es común desde finales de febrero
a octubre, alcanzando un pico a finales de primavera y principios del verano, aunque
también puede ser observado durante cualquier mes. Una vez que el macho ha
atraído a una hembra, forman una pareja y permanecen juntos durante toda la
temporada de reproducción. Las tareas de construcción del nido son compartidos.
Cuando un sitio adecuado ha sido seleccionado, por lo general en los pinos, cedros,
cerezos silvestres, robles o arbustos de bajo crecimiento.
Un paso esencial para llevar a cabo un programa de conservación o manejo
adecuado en especies de interés cinegético, es identificar la época de reproducción
de las mismas, así como las áreas de reproducción más importantes, la cual varía de
una especie a otra y dedicar a ellas un sustancial esfuerzo de vigilancia y monitoreo.
Existen varios indicadores que permiten conocer el éxito reproductivo de una
población; la natalidad y el éxito reproductor son buenos indicadores de la “salud” de
una población y de la intensidad de la mortalidad que pueden llegar a sufrir sin llegar
a declinar (Peiro, 2003).
El objetivo de este capítulo es determinar la duración de la estación reproductora y
el periodo de máxima reproducción de la especie en el área de estudio, utilizando
descriptores fisiológicos de los progenitores y de la presencia de jóvenes palomas.
Además, se pretende analizar el hábitat de nidificación según elementos del paisaje
agroforestal y la diversidad de sustratos vegetales de emplazamiento del nido, así
como el éxito reproductor.

69
5.2.- Materiales y métodos
Los trabajos de campo consistieron en la toma de muestras de los ejemplares
cazados de esta paloma. La caza fue realizada al azar con escopeta de cartuchos en
recorridos a pie que abarcaron toda el área y cada ejemplar abatido fue etiquetado
siguiendo el orden de captura y clasificado según la clase de edad (adulto o pichón).
Se estudiaron un total de 76 ejemplares. A cada ejemplar capturado se le efectuó la
disección para conocer el sexo y el estado de desarrollo de los testículos o de los
huevos en el interior del abdomen, la presencia y desarrollo de las glándulas del
buche o “de la leche”.
Se realizaron una serie de recorridos por el área de estudio para analizar el
comportamiento según la época del año y el hábitat de nidificación atendiendo al el
biotopo en que se encuentra, elementos del paisaje agrícola y especies vegetales
como sustratos de nidificación, a razón de dos recorridos por mes durante todo el
año.
En cada nido encontrado se clasificó la planta sobre la que se encontraba. Con un
clinómetro Sunto se calculó la altura total de la planta y la altura a la cual se
encontraba el nido respecto al suelo; además de agrupar las plantas en dos
categorías (espinosas y no espinosas) para conocer las influencias de estas
categorías en la selección de nidificación. Se utilizó como hipótesis de partida que
las plantas espinosas darían mayor protección al nido frente a depredadores y
aumentarían el éxito reproductor de la especie.
El éxito reproductivo de la Paloma Rabiche se determinó mediante el seguimiento
de los nidos que según su ubicación se podían chequear sin que fuese necesario
realizar grandes esfuerzos o colocar artefactos que pudiesen provocar el abandono
de las aves del nido producto de las perturbaciones asociadas al muestreo.
A pesar de que constituye una limitación del estudio el no seguir protocolos más
precisos para la obtención del éxito reproductivo de la especie, y decidir emplear el
método anteriormente presentado; consideramos que sus resultados aún con los

70
posibles sesgos que pudiesen ofrecer resultan de gran beneficio para investigadores
que deseen aproximarse al fenómeno y no cuenten con la tecnología requerida.
La duración del proceso de incubación se determinó mediante el seguimiento de los
nidos donde al menos un pichón llegó a volar, determinándose el máximo, mínimo y
media de días de incubación. Se desecharon aquellos nidos que fueron destruidos
por causas naturales o antrópicas durante este período.
Se realizaron un total de cuatro muestreos, a razón de dos por cada mes,
seleccionando las áreas de mayor densidad de nidos. Se observó en cada estación
con unos binoculares por espacio de 3 horas todas las palomas que se movían, se
clasificaron y anotaron todos los vuelos vistos.
En la zona de estudio se realizó una encuesta para obtener información sobre la
percepción que tienen los sectores sociales rurales (cazadores deportivos,
campesinos) relacionados con la especie sobre la reproducción, sitios de nidificación
e identificación de esta (Anexo 4). La información obtenida en este proceso es
esencial para fortalecer los resultados que se obtengan de la presente investigación.
A los datos obtenidos se aplicaron primeramente técnicas de estadística descriptiva,
con la finalidad de resumirlos, describirlos numéricamente y facilitar su interpretación
(medidas de tendencia central y de variabilidad). Con el objetivo de comparar la
proporción de sucesos entre las diferentes muestras se utilizó el test de “Chi-
cuadrado”. A los 31 datos obtenidos sobre la nidificación se les aplicó el test de
correlación de Pearson (r) para analizar la relación entre la altura total de las
plantas utilizadas como sustrato para la nidificación y la altura del nido al suelo.
Teniendo como hipótesis de partida que esta relación podría estar asociada con una
conducta de la especie en busca de protección al nido frente a depredadores aéreos,
terrestres y fenómenos meteorológicos adversos. En el procesamiento de la
información se emplearon los programas Excel y SPSS versión 15.0 para Windows.

71
5.3.- Resultados y discusión
5.3.1-. Estación reproductora
El ciclo gonadal de las aves está dado por las variaciones que ocurren en el tamaño
de las gónadas (ovarios o testículos). Al realizar un seguimiento periódico de estos
cambios se podría contar con la aptitud reproductiva en determinada época, por lo
que este análisis gonadal podría ser válido para conocer el ciclo reproductivo de la
Paloma Rabiche. Otro aspecto que se tuvo en cuenta para conocer el ciclo
reproductivo de esta especie fue el desarrollo de las glándulas del buche; según
Marshall, 1969 en Acosta y Torres (1984), su desarrollo solo se presenta en estas
aves con la etapa de incubación hasta que termina la ceba de los pichones.
La evolución mensual del estado reproductivo de esta paloma mediante el análisis
gonadal, glándulas del buche, desarrollo testicular y presencia de juveniles en una
muestra de 78 ejemplares cazados se presentan en la Figura 20 (A y B).
Figura 20. Variación mensual del desarrollo de los testículos y presencia de huevos
ováricos maduros (A) y de las glándulas del buche desarrolladas y la presencia de
pichones (B) de la Paloma Rabiche (n=76) en Cuba.

72
La Figura 20 A muestra un periodo reproductivo que se extiende desde marzo
hasta agosto, pero una parte de la población se mantiene en reproducción en el
resto de los meses del año, lo que queda evidenciado por la presencia de huevos
ováricos maduros en los meses de enero y diciembre. La Figura 20 B muestra
concordancia entre el periodo más elevado de desarrollo glandular del buche de los
progenitores y los valores más altos de frecuencias de pichones en la población
estudiada, marcando así una etapa claramente definida de mayor actividad
reproductiva correspondiendo respectivamente con la cría de pollos y su posterior
abandono del nido.
Al analizar si existen diferencias estadísticas en cuanto al comportamiento
reproductivo de esta columbida en el tiempo, se agruparon los datos obtenidos de
los cuatro indicadores reproductivos en dos periodos: primavera-verano (marzo-
agosto), época donde debido a la alta humedad y temperatura predomina la rotación
y maduración de cultivos, así como la presencia y fructificación de plantas silvestres
herbáceas presentes en el espectro alimentario. Otoño-invierno (septiembre-febrero),
época caracterizada por ser poco húmeda, seca y se registran las menores
temperaturas del país. Al aplicar el test estadístico de Chi-cuadrado se evidenció una

73
estacionalidad definida en la reproducción, dada por la época de primavera-verano
(X2 = 36,750; p=0,000; gl=1; p<0,05) como se muestra en la Figura 20 A y B.
Estos resultados apoyan lo planteado por Berovides (1998), donde expone que la
época de reproducción es desencadenada en la mayoría de las especies que
presentan una marcada conducta sexual en primavera. Este autor considera que
este fenómeno se produce por el alargamiento de las horas de luz y por el comienzo
de las lluvias que condicionan la disponibilidad del alimento durante la estación
reproductiva o previa a la misma. En nuestro caso considero que el alargamiento de
las horas de luz no es el factor que desencadena la época de reproducción, por ser
Cuba una región tropical, donde la magnitud de las horas de luz varía poco durante
el año (Quintero y Alonzo, 2007).
En este estudio los resultados son coincidentes con los obtenidos por Rivera
(2001), en Puerto Rico, Vieques y Cayo Culebra donde relacionó la densidad de
nidos con la lluvia, abundancia de comida y abundancia de rapaces. Siendo la
abundancia de comida el factor más importante en los cambios anuales en la
densidad de nidos. Es consistente también con lo planteado por Camprodon y
Subirachs (2001): “a lo largo de un año y sincronizadamente con la sucesión de las
estaciones climáticas, las especies de caza menor organizan su vida. Los ciclos
anuales de la fauna se pueden dividir en dos periodos distintos y consecutivos: el
periodo reproductor (en el que sucede la cría) y el periodo no reproductor (sin
actividad sexual). Las aves y los mamíferos utilizan distintas estrategias en la
asignación de tiempos a lo largo del año a las tareas reproductoras y no
reproductoras”.
Estos resultados han reforzado nuestro punto de vista de que al ser el comienzo de
las lluvias un aspecto bien marcado en el área de estudio, condicionan un aumento
de la disponibilidad de alimentos para la Paloma Rabiche. Es en esta época
primavera – verano, donde se aprecia de manera evidente el comienzo y el período
de máxima reproducción de la especie.

74
El análisis del desarrollo gonadal, tanto en machos como hembras evidencia un pico
reproductivo en el mes de junio, aunque la época de máxima reproducción comienza
en marzo y se extiende hasta agosto. El desarrollo de los testículos en los machos,
se observa desde el mes de febrero. Acosta y Torres (1984) obtuvieron resultados
que demuestran las variaciones que sufrieron los testículos de la Paloma Rabiche
entre los meses de enero y septiembre. Al examinar la presencia de huevos ováricos
en las hembras, además de obtenerse el mayor valor de la muestra en abril, se
observó también en los meses de diciembre y enero. Es importante señalar que en
los recorridos de campo realizados en el área de estudio se observaron ejemplares
de la especie en nidos activos fuera de la época de máxima reproducción. Según
Camprodon y Subirach (2001) la amplitud del periodo reproductor condiciona las
oportunidades de tener éxito en los intentos procreadores de los animales. Esto hace
que las estrategias reproductoras que utilizan las distintas especies de caza menor
tengan diferencias fundamentales. Por otro lado, no sólo la amplitud del periodo hábil
para la reproducción condiciona su éxito, también es relevante el sincronismo que se
consigue con el mejor momento anual para tener éxito en la cría.
Similares resultados obtuvieron Acosta y Torres (1984) en el Jardín Botánico de
Cienfuegos, Cuba, donde determinaron que la época de máxima reproducción de la
Paloma Rabiche se encuentra entre los meses de marzo a octubre. Peters (1961)
citado por Westmoreland et al. (1986), plantearon que Zenaida macroura tiene una
de las más largas épocas de reproducción de todas las aves de América del Norte.
Estos autores refieren en su estudio una serie de características evolutivas que
garantizan una estrategia en su reproducción como: alimentación de pichones con
“leche de paloma”, que les permite a estas palomas iniciar nuevas reproducciones
de forma rápida, incubación constante, rápido crecimiento de los pichones,
construcción de pequeños nidos y a veces reutilización de viejos y un menor papel
de la hembra en la producción de la “leche de paloma” que permite a éstas iniciar
nuevas nidificaciones de forma rápida.
En las columbidas el desarrollo de las glándulas del buche ocurre en ambos sexos,
este comienza antes de la etapa de ceba de los pichones, alcanzando su máximo

75
desarrollo en el momento de la eclosión de los huevos, siendo un buen indicador de
la actividad reproductiva de la especie. Igual criterio refiere Marshall (1960) citado
por Acosta y Torres (1984), al exponer “… la presencia de las “glándulas de la
leche” es otro factor que puede contribuir al esclarecimiento del ciclo reproductivo de
las palomas, ya que se presentan únicamente cuando el ave se encuentra criando y
se origina una proliferación de células especiales del epitelio del buche que se van
descamando, y que están constituidas por grasas (25-30 %), lectina (5 %) y proteína
(10-15 %)”.
Al comparar los resultados obtenidos del desarrollo de las glándulas del buche y la
presencia de pichones en la muestra obtenida existe una concordancia entre el
periodo de valores más elevados de desarrollo glandular del buche de los
progenitores y los valores más altos de frecuencia de pichones en la población
estudiada, marcando así una etapa claramente definida de mayor actividad
reproductiva correspondiente con la cría de los pollos y su posterior abandono del
nido.
Esta etapa puede definirse desde marzo hasta agosto, con un pico reproductivo
(dado por la mayor cantidad de pichones en la muestra) en junio. Estos resultados
presentan concordancia con lo expuesto por Tomlinson (1994) en los EUA, donde la
época de reproducción comienza en marzo y el pico reproductivo tiene lugar en
junio, finalizando para el mes de agosto. Miller et al. (2001) en un estudio realizado
en el Norte del valle central de California, obtuvieron que la especie inició la
reproducción a mediados de marzo, manteniendo esa actividad hasta finales de
agosto, coincidiendo estos meses con la época de primavera-verano, la cual
presenta condiciones favorables para la reproducción de la especie. En relación a
estos análisis Camprodon y Subirachs (2001) afirman que el conocimiento de la
fenología de las especies de la zona en que vamos a trabajar es de suma
importancia ya que nos permitirá valorar no sólo el grado de incidencia de
determinadas actuaciones, sino también escoger el momento idóneo para realizar
una determinada actuación.

76
En el área de estudio, aunque la frecuencia de ocurrencia de pichones en las
muestras cazadas es nula en los meses de noviembre a enero, se sabe, por
observaciones directas de nidos activos realizadas por el propio autor, que en estos
meses la especie mantiene cierto nivel de actividad reproductiva (Tabla 7).
Tabla 7. Nidos activos encontrados durante los meses del año en el área de estudio.
Meses Nidos activos
enero -
febrero 1
marzo 3
abril 3
Mayo 5
junio 7
julio 5
agosto 2
septiembre 2
octubre 1
noviembre -
diciembre 2
Acosta y Berovides (1982), también encontraron palomas Rabiche con glándulas del
buche desarrolladas en el mes de diciembre en el Sur de Pinar del Río, Cuba,
evidencia de cierto nivel de reproducción de la especie en esa época. Estos
resultados confirman los expuestos por Skutch (1950); Tompson (1950) citados
por Westmoreland et al. (1986) al exponer que la mayoría de las aves sincronizan
la reproducción con un breve período de abundancia de alimentos de alta calidad
para los pichones. Sin embargo las Columbidas al producir alimentos como la “leche
de palomas” para los primeros días de nacidos los pichones y luego alimentarlos con
una variada dieta de semillas cuando son algo mayores (Browning (1959) en
Westmoreland et al. (1986), presentan por tanto prolongadas temporadas de
reproducción.
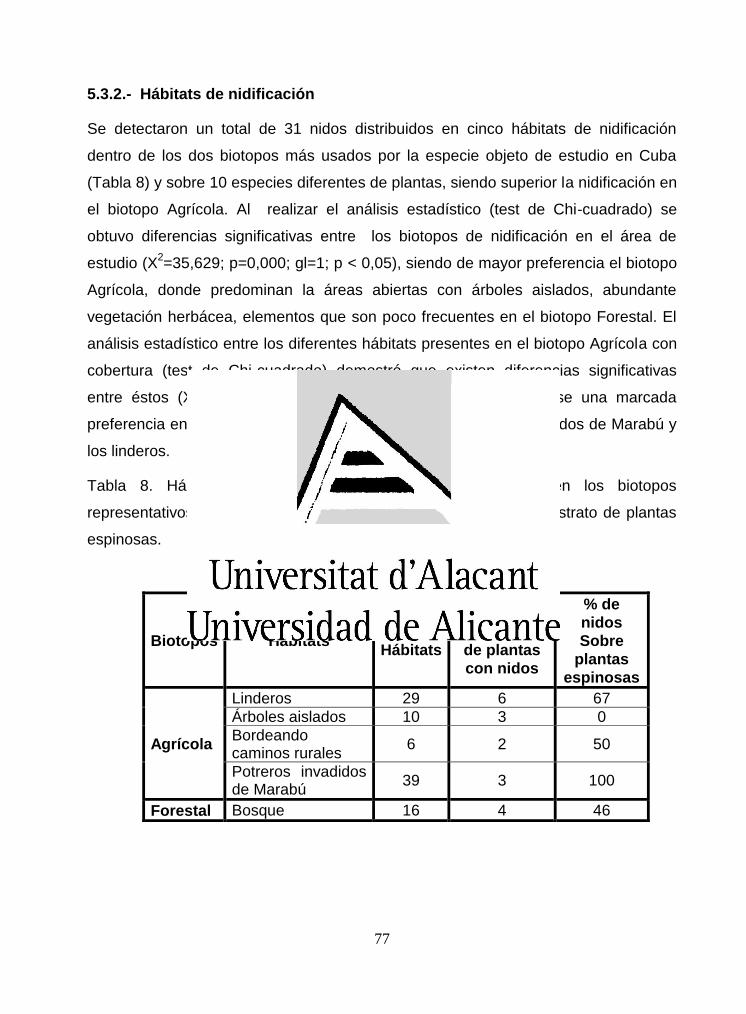
77
5.3.2.- Hábitats de nidificación
Se detectaron un total de 31 nidos distribuidos en cinco hábitats de nidificación
dentro de los dos biotopos más usados por la especie objeto de estudio en Cuba
(Tabla 8) y sobre 10 especies diferentes de plantas, siendo superior la nidificación en
el biotopo Agrícola. Al realizar el análisis estadístico (test de Chi-cuadrado) se
obtuvo diferencias significativas entre los biotopos de nidificación en el área de
estudio (X2=35,629; p=0,000; gl=1; p < 0,05), siendo de mayor preferencia el biotopo
Agrícola, donde predominan la áreas abiertas con árboles aislados, abundante
vegetación herbácea, elementos que son poco frecuentes en el biotopo Forestal. El
análisis estadístico entre los diferentes hábitats presentes en el biotopo Agrícola con
cobertura (test de Chi-cuadrado) demostró que existen diferencias significativas
entre éstos (X2=21,244; p=0,000,gl=1, p < 0,05), evidenciándose una marcada
preferencia en la nidificación de la especie hacia los potreros invadidos de Marabú y
los linderos.
Tabla 8. Hábitats de nidificación de la Paloma Rabiche en los biotopos
representativos de la especie en Cuba y comportamiento sobre sustrato de plantas
espinosas.
Biotopos Hábitats
% nidos
por
Hábitats
Número
especies
de plantas
con nidos
% de
nidos
Sobre
plantas
espinosas
Agrícola
Linderos 29 6 67
Árboles aislados 10 3 0
Bordeando caminos rurales
6 2 50
Potreros invadidos de Marabú
39 3 100
Forestal Bosque 16 4 46

78
El análisis estadístico (test de Chi-cuadrado) de las categorías de plantas sobre las
que se encontraron ubicados los nidos muestra una marcada preferencia (71% de
nidos) de las plantas espinosas (X2=19,758; p=0,000; gl= 1 p < 0,05), donde las
plantas de mayor uso como sustrato de emplazamiento del nido resultaron ser dos
especies espinosas, el Marabú (Dichostachys cynerea) y la Aroma blanca (Leucaena
glauca).
Aunque existen diferencias entre las diferentes especies de aves a la hora de
seleccionar los sitios de nidificación, estos emplazamientos generalmente tienen
características en común, como son adecuada estabilidad en las estructuras del nido
frente a fenómenos meteorológicos, abundante material de construcción del nido,
protección contra depredadores y daños causados por el hombre y la cercanía a las
áreas de alimentación.
El mayor número de nidos se hallaban en el hábitat de potreros invadidos de Marabú
(Dichostachys cynerea) del agrosistema tropical. Esto pone de manifiesto la
tendencia de la especie a usar con mayor frecuencia áreas con una cubierta vegetal
muy difícil de penetrar y por tanto que ofrece una mayor protección al nido frente a
factores meteorológicos adversos, a depredadores y molestias antrópicas. Estos
resultados difieren de los obtenidos por Drobney et al. (1998) en la parte central de
Missouri, EUA, al estudiar la reproducción de la Paloma Rabiche, el 81,9 % se
encontraron en hábitats de bordes, aunque los hábitats continuos fueron menos
seleccionados, no se encontraron diferencias significativas en la proporción de nidos
existentes entre los dos tipos de hábitats.
En Cuba la Paloma Rabiche tiene un rango amplio de selección de plantas como
soporte para la ubicación del nido, aunque utiliza fundamentalmente especies de
arbustos y árboles. Basándonos en estos resultados se analizó las categorías de
plantas sobre las cuales estaban ubicados los nidos en que al menos un pollo llegó a
abandonar el nido. En este análisis se obtuvo que más del 70 % de estos nidos se
encontraban sobre plantas espinosas (Anexos 5 y 6). Estos resultados proporcionan
un sólido soporte al argumento de la protección brindada por esta categoría de
plantas en la reproducción de la especie. Concuerda este resultado con los

79
presentados por Cisneros et al. (2006) en el evento Talleres sobre conservación y
uso sustentable de aves y mamíferos silvestres, en relación con las Unidades de
Conservación y Manejo de Vida Silvestre (UMA) en México, donde exponen que al
realizar una breve descripción de los rasgos más relevantes de la historia natural de
la especie los nidos suelen ubicarse, al menos en el noreste de México en las
especies vegetales del matorral Tamaulipeco (Mezquites (Prosopis glandulosa,
Prosopis velutina), Huizaches (Acacia pennatula) y plantas espinosas en general),
son las más utilizadas para construir los nidos.
En la revisión de la literatura no se ha podido encontrar estudios que analicen el
éxito reproductor de la Paloma Rabiche en relación a la selección de esta categoría
de plantas en Cuba, siendo nuestros resultados los primeros obtenidos. No obstante,
surge la necesidad de nuevas investigaciones sobre el conocimiento de otros
factores que influyen en el éxito reproductivo de la especie que proporcionarían una
mejor base para la gestión de su hábitat de reproducción. Estos resultados difieren
de los descritos para los EUA por Tomlinon (1994), donde las características y
preferencias de la especie para la nidificación varían sustancialmente, prefiriendo las
coníferas a principios de año (cuando aún es invierno), antes que los árboles
deciduos hayan desarrollado las hojas, lo que les brinda una mayor cobertura y
protección ante inclemencias del tiempo y el ataque de los depredadores.
Al comparar la altura media de los nidos con la altura total de la vegetación, el
comportamiento de la especie muestra preferencia por situar los nidos en la parte
central de la altura de la vegetación utilizada como sustrato para la nidificación
(altura media de vegetación = 4,1 m, altura media del nido = 2,5 m).
Según la clasificación dada por Martínez et al. (2009), existe una correlación positiva
entre estas dos variables de altura, clasificándose como fuerte y directa (r= 0,777;
p=0,000; p<0,01). Este mismo procedimiento fue usado para analizar el
comportamiento de la especie ante los árboles y arbustos usados como sustrato
para la nidificación, obteniéndose en ambas categorías de plantas los siguientes
resultados: una correlación positiva, moderada y directa (rho= 0,550, p=0,125;

80
p>0,05) en los árboles y una correlación positiva, fuerte y directa (rho= 0,832;
p=0,000; p<0,01) en los arbustos.
Durante todo el periodo que duró la realización de este estudio, ni en los años de
experiencia del autor ha sido detectado un nido de esta Columbida en el suelo, lo
cual difiere de lo descrito por Cleveland (1932) al describir que “… este pájaro
anida con frecuencia en el suelo, incluso en un grupo de hierbas’’. Igualmente
Drobney et al. (1998), expresaron “Nuestros datos demuestran que estas palomas
utilizan todo el espectro de tipos de hábitats para la nidificación, que van desde nidos
en el suelo en campos abiertos al dosel del interior de los bosques’’.
5.3.3.- Éxito reproductor
El análisis del seguimiento reproductor de los nidos (Tabla 9) muestra que el éxito de
eclosión (% de nidos que alcanzaron la fase de eclosión de los huevos) fue del 67 %
y las causas naturales de pérdida superaron a las producidas por daños antrópicos
de los nidos perdidos en fase de incubación. Ningún nido tuvo un tamaño de puesta
de un solo huevo o más de dos huevos. Ningún huevo quedó en el nido sin
eclosionar por motivo de infertilidad. El éxito de cría de pollos (% de nidos con
eclosiones en que algún pollo llega a abandonar el nido) fue del 81 %, siendo mayor
la pérdida de pollos por daños antrópicos que por depredación natural. El éxito
reproductor total fue del 55 %, pero uno de los nidos sufrió una depredación parcial
de un pollo. Los hábitats de nidificación con mayor éxito reproductor son los potreros
y el bosque, mientras que los linderos presentan el menor éxito de reproducción.

81
Tabla 9. Éxito reproductor de la Raloma Rabiche en hábitats de agrosistema de
Cuba.
Hábitat de
nidificación
% Nidos con pérdida
de huevos %
nidos
con
eclosiones
% Nidos con pérdida
de pichones % Nidos
con
pichones Natural Antrópico Natural
(Depre.) Antrópico
Borde del camino
0 50 50 0 0 50
Potreros invadidos de Marabú
25 75 12,5* 0 62,5*
Linderos 33 0 67 0 0 33
Bosque 33 67 0 0 33
TOTAL 25 8 67 4* 8 55*
*Nido con depredación parcial de un pollo contado como pérdida de 0,5
Con el objetivo de determinar la influencia de las categorías de plantas (espinosas y
no espinosas) seleccionadas por la especie para nidificar sobre el éxito reproductor
total se aplicó el test estadístico de Chi-cuadrado, obteniéndose diferencias
significativas entre ambas categorías (X2=4,571; p=0,033; gl=1, p<0,05), siendo
mayor el éxito reproductor total en las plantas espinosas.
Las principales causas del fracaso de las nidadas de esta Columbida fueron los
factores naturales (64 %), estos resultados concuerdan con los expuestos por Acosta
y Ponce de León (2004), cuando describen que las condiciones climáticas extremas
como vientos fuertes y lluvias intensas provocan entre 4 y 5 veces más pérdidas de
las nidadas que las causadas por las cacerías organizadas.
Las causas antrópicas que propiciaron la pérdida de los nidos corresponden a su
ubicación en los bordes de caminos rurales o en linderos de campos de cultivo.
Estos lugares presentan un tránsito frecuente de campesinos y en los linderos se
realizan tareas de mantenimiento que causan destrucción de estos nidos (Anexos 7

82
y 8). Entre estas tareas agrícolas, destacan el impacto de la poda de los árboles de
los linderos cuando éstos son muy frondosos y proporcionan mucha sombra a
cultivos o pastizales aledaños. Estos resultados coinciden con los obtenidos por
Smith et al. (2012) al estudiar el comportamiento de la nidificación de la especie en
bosques ribereños con diferentes estructuras, el área con menor porcentaje de
nidificación fue aquella en que se realizaron intervenciones como medidas para la
reducción del material combustible y evitar los incendios forestales.
Miller et al. (2001), obtuvieron que el éxito reproductivo de la Paloma Rabiche fue de
un 22 a un 45 % en California, siendo inferior al éxito en un agrosistema de Cuba
obtenido en este estudio (55 %). Los autores citados refieren en su estudio que el
éxito de incubación obtenido fue de un 35 %. Este mayor éxito reproductivo en Cuba,
puede ser debido a las diferencias en cuanto al tipo de plantas utilizadas en la
nidificación, ya que en el país se conoce que un importante número de nidos son
construidos sobre plantas espinosas, obteniendo una mayor protección en los
mismos. Otro factor que debe tenerse presente al analizar el éxito reproductivo de la
especie es la altura media a la que el nido es construido, al utilizar la parte central
de las plantas evita así a depredadores aéreos que sobrevuelan las zonas de
anidación, factores meteorológicos adversos y daños antrópicos. Como es posible
apreciar el éxito reproductivo de la especie tiende a comportarse de diversas
maneras según sean las condiciones en las diferentes zonas donde habita. Rivera y
Frank (2001) al evaluar la relación entre la densidad de nidos y factores como las
precipitaciones, la abundancia, diversidad de alimentos y depredadores, obtuvo que
la abundancia de alimentos fue el predictor más abundante en los cambios anuales
en la densidad de nidos.
En todos los nidos que llegaron a la fase de eclosión de los huevos, la misma fue
exitosa en un 100 %.
La Tabla 10 muestra el tiempo total empleado por la Paloma Rabiche desde que
comenzó la incubación hasta el abandono del nido por los pichones. Solo fueron
tomados en cuenta para esta información los 7 nidos donde llegaron los pichones a
volar, no se tuvo en cuenta el tiempo de construcción del nido, ni el periodo de

83
puesta, porque algunos nidos al ser descubiertos ya tenían un huevo o en otros
estaban a punto de iniciar la incubación.
Una vez que se detectó la eclosión de los huevos, se redujo el número de
observaciones y las realizadas se hicieron a mayor distancia con el objetivo de no
provocar una perturbación en el animal que provocase el abandono de las crías.
Tabla 10. Periodo de incubación y estancia en el nido de los pichones de paloma
Rabiche en el área de estudio (n=7).
I D*
Días
incubación
Estancia
en nido
Tiempo
Total
B 12 11 23
C 16 13 29
E 14 12 26
F 13 15 28
I 14 12 26
J 15 13 28
L 14 14 24
TOTAL
Media 14 13 26
Máximo 16 15 29
Mínimo 12 11 23
ID* código de identificación de cada nido que permite conocer el biotopo, especie de planta y altura
donde se encontró.
El periodo de estancia en el nido fue calculado a partir de la eclosión de ambos
huevos, siendo el mínimo de 11 días, un máximo de 15 días y como promedio para
el área de estudio de 13 días.
En resumen el tiempo total transcurrido desde el inicio de la incubación hasta el
abandono del nido por los pichones fue de 26 días como promedio. Miller et al.
(2001) en estudio realizado en el norte del valle central de California, EUA obtuvo

84
que el éxito de la incubación fue de un 35 a un 59 %. En México según Cisneros et
al. (2006) la incubación requiere de 14 a 15 días; el tiempo de vuelo inicial
normalmente es de 14 a 15 días después de la eclosión del huevo, pero en
algunas ocasiones puede ser de 10 días, El ciclo completo de anidación requiere
cerca de 32 días.
5.3.4.- Percepción rural de la estación reproductora
El conocimiento tradicional de cazadores y campesinos de la zona que tienen cierto
nivel de relación con la especie, ayudan a conocer los hábitos y características de la
especie. Con esta información se pueden inferir comportamientos, tendencias y
mayores usos en los diferentes hábitat, además permite evaluar la relación social
con la especie, elemento útil en la conservación, pues revelan el tipo de interacción
con la especie y su hábitat, señalando áreas o sectores donde trabajar hacia una
mejor perspectiva de estas personas hacia los recursos.
La encuesta realizada en la investigación con el fin de obtener información sobre la
percepción que tienen estos sectores sociales sobre la reproducción, sitios de
nidificación e identificación de la especie fue esencial en este proceso ya que
permitió triangular los datos registrados con el trabajo de campo, proporcionando de
esta manera mayor robustez a los resultados presentados.
Tabla 11. Resultado de las encuestas aplicadas a sectores rurales vinculados con la
especie (n=27).
Sectores
No
. e
nc
ue
sta
do
s
ID Sp (%) EC
(%)
UN
(%)
¿A cambiado la
época de cría?
Si
No
Pri
ma
v.
Se
ca
Am
ba
s
Lin
de
ro
Bo
sq
ue
Po
tre
ros
Inv
ad
id.
Árb
ole
s
ais
lad
os
Si
No
No
sé
Cazadores 11 100 91 9 60 30 30 9 73 18
Campesinos 16 100 87 13 31 6 19 44 12 87
Total 27 100 89 7 4 44 4 15 37 4 37 59
ID SP: Identificación de la especie. EC: Época de cría. UN: Ubicación del nido.

85
La Tabla 11 muestra los resultados obtenidos en las encuestas realizadas por el
autor en el área de estudio, aquí aparece reflejado como la especie objeto de estudio
es identificada por todos los encuestados, lo cual demuestra la importancia que ha
tenido la especie en los campos cubanos, que ha sido históricamente usada tanto
por cazadores como por campesinos, principalmente como fuente de alimento
alternativa. Este resultado es válido ya que demuestra que la especie no ha sido
confundida con otra paloma, y así lo expresado es el resultado de su conocimiento
sobre la Paloma Rabiche.
Otro importante resultado que avala los datos obtenidos en este trabajo lo refleja la
información obtenida en la encuesta respecto a la época de cría de las Paloma
Rabiche, donde a pesar que la mayoría de las respuestas coinciden que es en
primavera, existen opiniones donde se afirma que en seca (en Cuba los
campesinos y muchas personas le llaman así al invierno por ser una época de muy
pocas precipitaciones) aún se pueden encontrar nidos activos de Paloma Rabiche
(Tabla 7).
5.3.5.- Aspectos conductuales relevantes de la reproducción
Con el objetivo de corroborar de manera práctica ciertas conductas de vuelo
observadas en la especie en una primera etapa de ejecución de este estudio, se
decidió realizar una serie de muestreos en los meses de mayo y junio, ya que en
estos dos meses la especie se encuentra en el periodo de máxima reproducción y es
posible observar más fácilmente los tipos de vuelos relacionados con esta etapa, que
en otros meses. Por lo antes señalado sería más difícil encontrar parejas activas en
el resto del año.
A continuación se describen los tipos de vuelos clasificados, así como su
descripción:
De traslación: vuelo rápido, con golpes de alas casi constantes, fuertes y rápidos,
nunca planean y dan la impresión de que si dejan de batir las alas se caerán.
Pueden ser bajos o los más altos en esta especie de paloma, sobre todo estos
últimos cuando lo realizan en pequeños bandos. Estos vuelos se pueden

86
observar a cualquier hora del día cuando las palomas se dirigen a las áreas de
alimentación o descanso, generalmente es recto, se dirigen directo a un punto o
lugar. Durante todo el vuelo la paloma nunca parece deambular. Se puede
observar solas, en parejas o en época de sequías (invierno) formando pequeños
bandos en movimiento.
De cortejo: se subdividió en dos variantes:
De seguimiento: vuelo muy rápido, golpes de alas rápidos, continuos y fuertes,
además de cortos. A veces es recto, a veces en forma de zig-zag, caracterizado
por los cambios rápidos y bruscos de dirección, generalmente son vuelos por
encima de las copas de los árboles, nuca se observó a gran altura. Siempre
ocurren cuando una paloma persigue a otra y en algunas ocasiones se han visto
hasta 3 palomas persiguiéndose. Este vuelo está relacionado con la época de
reproducción.
De planeo: vuelo con golpes de ala lentos y metódicos, luego son seguidos por
un planeo que nunca llega a ser tan largo. Es casi siempre en círculos y puede
observarse realizándolo a una paloma o una pareja, son ejecutados por encima
de las copas de los árboles, pero no a gran altura. Este es vuelo de cortejo por lo
que está estrechamente relacionado con la época de reproducción.
Tabla 12. Clasificación de los tipos de vuelos usados por la Paloma Rabiche en el
área de estudio y su frecuencia quincenal (Q).
Tipos
vuelos
mayo junio Población
1ra
Q
2da
Q Total Media
1ra
Q
2da
Q Total
Media Total Media
Traslación 5 9 14 7 8 11 19 9.5 33 8.2
Seguimiento 1 3 4 2 5 3 8 4 12 3
Planeo 2 3 5 2.5 1 1 2 1 7 1.7

87
En la Tabla 12, se puede apreciar que los vuelos de traslación son más evidentes,
reflejando una gran actividad durante todo el día; aspecto relacionado con sus
hábitos alimentarios, evidenciado en un constante buscar comida. Al respecto
Acosta y Berovides (1982) refieren “… la Rabiche puede observarse todo el día en
una actividad constante que, por consiguiente, trae aparejado un consumo máximo
de alimentos para suplir este gasto energético”.
Otras categorías de vuelos que pueden considerarse indicativo de la especie en
reproducción y fácil de identificar son los de cortejo; esta categoría a su vez se
subdivide en dos variantes: vuelos de seguimiento y de planeo. El primero resultó
mucho más frecuente que el segundo, lo que puede estar relacionado con la
competencia en la búsqueda de pareja, donde la presencia de varios machos los
condiciona. El vuelo de planeo resultó ser menos frecuente y está quizás
relacionado con la aceptación de los machos por las hembras. Los vuelos de
cortejo de la Paloma Rabiche deberían tenerse más presentes en la actividad
reproductiva de la especie.
Otro aspecto que se tuvo en cuenta para poder dar una más amplia caracterización
de la conducta de la Paloma Rabiche fue su comportamiento ante la perturbación
humana cuando está en el nido. Con este objetivo se determinó el nivel de tolerancia
de la especie ante la presencia del observador. Del total de los 12 nidos que fueron
seguidos periódicamente, fue tenido en cuenta la distancia mínima a la que se pudo
llegar sin que la paloma abandonase el nido. Cuando la Paloma Rabiche está en el
nido es bastante tolerante a la perturbación, pues si se le acerca con cuidado y sin
mucho ruidos, admite la presencia humana hasta los 2 o 3 metros, siempre en los
primeros acercamientos al nido. Como media de distancia de acercamiento se
obtuvo 3,7 m. Es importante aclarar que la distancia de tolerancia de la especie
estuvo marcada por la altura del nido, mientras más alto estuviesen los nidos, la
paloma toleraba más la presencia humana debajo del árbol, en los nidos
construidos sobre arbustos a poca altura del suelo, luego de las dos primeras visitas,
ya al notar la presencia humana abandonaba el nido rápidamente.

88
Tabla 13. Distancia media de huida de la Paloma Rabiche en el área de estudio
(n=12).
Tipo de plantas Distancia media de huida (m)
Arbustos 4,5
Árboles 2,9
TOTAL 3,7
Cuando en periodo de incubación o con pichones la Paloma Rabiche fue molestada
de tal manera que abandonó el nido, siempre empleó el “truco del ala rota” en las
primeras perturbaciones, luego de estas primeras veces ya su conducta fue de huida
directamente. Este “truco”, consiste en una conducta típica de la especie de dejarse
caer a cierta distancia del nido y aletear como si tuviese un ala rota, algún golpe. Si
se le persigue y se trata de agarrar, vuela unos metros más adelante y repite lo
mismo con la intención de desviar la atención del lugar del nido.
Nunca se observó una Paloma Rabiche defendiendo el nido, ni cuando tenía huevos,
ni cuando tenía pichones al ser molestada por el hombre.
Este comportamiento en la huida descrito anteriormente no ha sido encontrado en
ningún trabajo realizado en la especie, por lo que debería tenerse en cuenta al
analizar la conducta reproductiva en próximos estudios.
5.4.- Conclusiones y recomendaciones
1. La Paloma Rabiche puede considerarse como una especie de reproducción
continua en agrosistemas tropicales, pero la etapa de mayor actividad
reproductiva de la especie se extiende de marzo a agosto.
2. El desarrollo de los testículos en los machos y el desarrollo de los huevos
ováricos en las hembras, así como el desarrollo de las glándulas del buche en
ambos sexos, constituyen buenos indicadores para evaluar la actividad
reproductiva de la especie en áreas tropicales.

89
3. La mayor ocurrencia de palomas juveniles en la muestra cazada se ubica desde
marzo hasta agosto, siendo junio el mes de mayor frecuencia de ejemplares
jóvenes que han abandonado el nido.
4. La especie tiene un rango diversificado de sustratos vegetales para la
nidificación, aunque se evidencia una mayor preferencia por los árboles y
arbustos espinosos.
5. El éxito reproductivo de la especie estudiada fue de un 55 %, siendo las
principales causas de las pérdidas de los nidos las naturales como fuertes vientos
y lluvias intensas.
6. Se recomienda utilizar por personal adiestrado as diferentes conductas
adoptadas en el vuelo como indicadores de las diferentes etapas del ciclo de
vida en la Paloma Rabiche.
5.4.- Bibliografía
Acosta, M. y O. Torres. (1984). Ecología trófica de las palomas del género Zenaida
en el Jardín Botánico de Cienfuegos, Cuba. Ciencias Biológicas, 11. 107-116.
Acosta, M. y V. Berovides. (1982). Ecología trófica de las palomas del genero
Zenaida en el sur de Pinar del Río. Ciencias Biológicas, 7. 113-123.
Acosta, M; P. de león, J.L. (2004): Historia Natural de la Ploma Rabiche (Zenaida
macroura). (Inedito). Facultad de Biología. Universidad de la Habana. Cuba.
Berovides, V. (1988). ¿Por qué los animales se comportan así?. Editorial Científico-
Técnica. La Habana. 226 pp.
Camprodon, J; Subirachs, E. P. (2001). Conservación de la biodiversidad y gestión
forestal. Su aplicación en la fauna vertebrada. Ediciones de la Unibersitat de
Barcelona. España. 469 pp.
Cisnero, E., Zamora, H., Mellink, E., Peterson, E., Ochoa, J., Franco, J. y Sánchez, J.
(2006). DGVS, 2006. Talleres sobre conservación y uso sustentable de aves y
mamíferos silvestres, en relación con las Unidades de Conservación y Manejo
de Vida Silvestre (UMA) en México. INE-SEMARNAT-UPC. 73 – 106.

90
Cleveland, A. (1932). Historias de vida familiar de las aves de Norteamérica.
Capítulo “La Paloma de Luto del Este (Zenaida macroura)”. Smithsonian
Institution Estados Unidos. Bolettin Museo nacional 162. Imprenta del Gobierno
de los Estados Unidos. 402-416.
Droney, R. D., Schulz, J. H., Sheriff, S. L. & Fuemmeler, W. (1998). Mourning Dove
nesting habitat and nest success in central Missouri. Journal of Field Ornithology,
69 (2), 299-305.
Martínez, M., Seguí, M. y López, C. (2009): Bioestadística amigable. Ediciones Díaz
de Santos. España. 293-342 pp.
Miller, M. R., Stemler, C. L. & Blankenship, D. S. (2001). Mournig Dove productivity in
California during 1992-95: was it sufficient to balance mortality?. Journal of
Wildlife Management, 65 (2), 300-311.
Peiró, V. (2003). Gestión ecológica de los recursos cinegéticos. Publicaciones de la
Universidad de Alicante. España. 136 pp.
Quintero, E. y Alonso, A. (2007). Ecología Agrícola. Editorial Félix Varela. La
Habana. Cuba. 183 pp.
Rivera, M. & Frank, F. (2001). Trancect surveys of Columbid nest on Vieques, and
Culebra islands, Puerto Rico. The Condor, 103 (2), 332-342.
Smith D. M., D. M. Finch y D. L. Hawksworth (2012). Nesting characteristics of
mourning doves in central New Mexico: Response to riparian forest change. The
Journal of Wildlife Management, 76 (2), 382 – 390.
Tomlinson, R. et al. (1994). Mournig Dove. Migratory shore an upland game bird
management in North. America. International Association of Fish and Wildlife
agencies. Allen Press, Lawrence, Kansas. E.U.A. 5-26.
Weiz, P. (1968). Biología. Edición Revolución. La Habana. 227-268 pp.
Westmoreland, D., Best, L. B & Blockstein, D. E. (1986). Multiple brooding as a
reproductive strategy: time-conserving adaptations in Mourning Doves. The Auk,
103 (1), 196- 203.

91
Yarrow, G. (2009). Biología y manejo de la Paloma de Luto. Extensión de Recursos
Naturales y Forestales de la Universidad de Clemson. Carolina del Sur. USA.
5pp.


92
CAPÍTULO VI. ESTRUCTURA DE LA POBLACIÓN DE PALOMA RABICHE
(Zenaida macroura L.)
6.1.- Introducción
Según Gonzales et al. (2013), entre los múltiples aspectos para la planificación y
manejo de un área protegida, se encuentra el conocimiento de la abundancia de las
poblaciones de las especies que se desean manejar y proteger o que puedan ser
indicadoras de cambios espaciales y/o temporales, en los hábitats o ecosistemas
donde las mismas habitan. Un paso inicial necesario es responder a la pregunta
¿Cuántos son?, o sea, cuántos individuos (adultos y juveniles, machos y hembras,
etc.) componen la población que se desea conocer, si estas cantidades se
encuentran entre los rangos normales registrados para la especie y si se mantienen
adecuadamente estables, se incrementan o disminuyen. Sin este dato básico es
imposible tomar decisiones de manejo o catalogar las especies como abundante o
escasas, entre otros aspectos.
La estructura de una población se define como su composición en grupos de edades
(age-ratio, que es el número de individuos juveniles partido por el número de
individuos adultos) y sexo (sex-ratio, que es el número de machos partido por el
número de hembras). El examen de las aves abatidas durante la caza o las muestras
biológicas obtenidas de ellas se pueden usar para determinar el sexo y la edad de
los animales, con estos datos se pueden calcular los dos cocientes: razón de edad y
razón de sexos. Conocer la estructura de la población es necesario para diagnosticar
el estado de ésta, predecir su tendencia y fijar racionalmente el cupo de capturas
(Nadal, 2004 citado por Cereza, 2005).
Se han publicado en la literatura científica varios estudios relacionados con las
razones de edad y su utilidad en el estudio poblacional, siendo ampliamente
aceptado el tema como importante indicador a tener en cuenta, tanto en la
conservación de especies, como en los planes de ordenación y aprovechamientos
cinegéticos:

93
Según Peiró (2003) la relación de edades o razón de edad (J/A) se expresa por la
proporción entre el número de jóvenes y el número de adultos, expresado bien en
tanto por uno (1 joven / adulto) o en porcentajes (50 % jovenes:50 % adultos). Este
parámetro es considerado como un indicador o medida de la natalidad y el éxito
reproductor, esta relación debe examinarse periódicamente para asegurar que se
mantenga dentro del rango de valores apropiados para la reproducción o para los
objetivos del método de caza.
Para Cereza (2005) la razón de edades en las cacerías es un buen indicador del
nivel de productividad reproductora anual. Teniendo en cuenta que la gestión de
capturas en caza menor descansa básicamente en el incremento anual por
reproducción, se comprende fácilmente el interés de este parámetro para la
ordenación cinegética.
Nadal (1994) citado por Chalaux (2005) expresó: cuando el índice de razón de edad
es mayor que 1, la población se encuentra en expansión, cuando el índice es igual a
1, la población es estable y cuando es menor que 1, no se renueva el total de la
población adulta.
Según Fuentes et al. (1991) citado por Cereza (2005) la razón de sexos (M/H) es un
indicativo de la estructura de la población y debe ser valorada en función del sistema
de apareamiento de cada especie estudiada. El análisis de este índice es de gran
interés, pues informa de un parámetro poblacional difícil de conocer por otros
medios.
Cuando la razón de sexos en menor que 1, existe un mayor número de hembras y
en consecuencia se trata de una población con una alta productividad potencial. En
el caso contrario, cuando la razón de sexos es mayor que 1, el número de hembras
es menor que el de machos, por lo que la productividad potencial es baja. Según
Nadal (1995) en Cereza (2005) en aquellas poblaciones estables y en expansión el
índice de sexo es igual o menor que 1.
El vacío de información científica en estos aspectos de las poblaciones de Paloma
Rabiche en Cuba impide un efectivo manejo para su aprovechamiento y

94
conservación, por esta razón el objetivo de este capítulo es determinar la estructura
de edades y la proporción de sexos en la población estudiada.
6.2.- Materiales y métodos
Como fue descrito anteriormente en el Capítulo V. Conducta y Reproducción, los
trabajos de campo consistieron en la toma de muestras de los ejemplares cazados
de esta paloma. La caza fue realizada al azar con escopeta de cartuchos en
recorridos a pie que abarcaron toda el área y cada ejemplar abatido fue etiquetado
siguiendo el orden de captura y clasificado según la clase de edad (adulto o pichón).
A cada ejemplar capturado se le efectuó la disección para conocer el sexo, se
estudiaron un total de 78 ejemplares. Con esos datos se calcularon la razón de edad
y de sexos.
La edad de los ejemplares capturados se determinó según las características
descritas en la literatura propias de cada clase de edad (joven o pichón y adulto) y la
experiencia del autor:
Joven o pichón: tienen como características distintivas en los extremos terminales de
las plumas una pequeña banda de color claro, las patas son bastante diferentes de
los adultos pues no presentan la típica coloración rojo, siendo de un color más opaco
y carmelitoso, el pico es de un tono más carmelita y apariencia débil. No presentan la
mancha iridiscente en el cuello.

95
Figura 21. Ejemplar joven de Paloma Rabiche.
Adultos: como se describió en el capítulo 1 los adultos de esta especie presentan la
parte superior de la frente y la nuca ligeramente grises. Por detrás el cuello es
carmelita grisáceo, con plumas metálicas en violeta y bronce a ambos lados, El
lomo, las coberteras y las alas son de color carmelita grisáceo. La rabadilla es
grisácea, un poco más oscura en los flancos. El ojo presenta un iris pardo oscuro y
rodeado por una piel orbital entre verdeazulosa y azul. El pico es negro, delgado y
con apariencia delicada. Patas de color rojo. La hembra es más pálida, con menos
iridiscencia en el cuello.

96
Figura 22. Ejemplar adulto de Paloma Rabiche.
La razón de edad (Re), indica la relación existente entre individuos jóvenes e
individuos adultos, nos da una idea de la capacidad de reproducción de los
individuos adultos, refleja la tendencia poblacional de la especie en función de la
tasa de renovación de los individuos adultos e indica su productividad (Nadal, 2004
citado por Cereza, 2005).
Su fórmula es:
Re = J / A
donde:
J = número de juveniles en la muestra cazada.
A = número de adultos en la muestra cazada.
La razón de sexo (Rs), es una forma sencilla de determinar la relación existente
entre sexos en la población estudiada. Esta proporción de sexos es un indicativo de
la estructura poblacional y debe ser valorada en función del sistema de

97
apareamiento de cada especie (Fuentes et al. 1991 en Cereza 2005). Su fórmula
es:
Rs = M / H
donde:
M = número de machos en la muestra cazada.
H = número de hembras en la muestra cazada.
6.3.- Resultados y discusión
Al realizar un análisis de estos dos importantes índices en la población de Paloma
Rabiche en el área de estudio se obtuvo que la estructura total del número de
muestra estudiado, contó con un 63 % de machos, un 37 % de hembras, un 18 %
de juveniles y un 82 % de adultos.
Tabla 14. Estructura de la población de Paloma Rabiche en el área de estudio
(n=78).
Razón de edad (J/A) Razón de sexos (M/H)
Ejemplares Razón
Ejemplares Razón
Joven Adulto Macho Hembra
14 62 0,23 48 28 1,71
Al realizar un análisis del resultado del índice de razón de sexos el primer elemento a
tener en cuenta será el tipo de relación que presenta la especie estudiada; la Paloma
Rabiche es una especie monógama, por lo que la relación de sexos esperada o
idónea sería de 1:1, un macho por cada hembra. En la muestra del área de estudio
el valor obtenido es superior a 1 (1,71), lo que según Fuentes et al. (1991) citado por
Cereza (2005) la población muestra un desequilibrio a favor de los machos, por lo
que tiene una productividad potencial reducida, dado por el hecho de quedar un
determinado número de machos sin formar parejas y por tanto sin producir
descendencia. Esta situación también a nuestro juicio tiene ciertas ventajas ya que

98
serán escogidos los mejores dotados, asegurando una mejor calidad en la población.
Cuesta (2009) al estudiar la especie en Pinar del Río, obtuvo un valor similar en
adultos, aunque algo superior (1,76). Esto evidencia la confiabilidad de los datos
obtenidos en la investigación, siendo de esperar en similares condiciones climáticas,
con una política agraria uniforme y con idénticos manejo y forma tradicional de
aprovechamiento de la especie, una similitud de resultados.
La razón de edad (J/A), es un indicador de la tasa de renovación de una población,
según Nadal (1994) citado por Calaux (2005), esta relación asegura la supervivencia
en el tiempo de la población. La relación esperada o adecuada para el mínimo
reemplazamiento de cualquier población es de 1:1, ósea, 1 joven por cada adulto. Al
calcular este índice en la población estudiada se obtuvo un valor inferior a 1 (0,23), lo
que evidencia la no renovación del total de individuos adultos (0,23 juveniles por
cada 1 adulto). Similares resultados obtuvo Cuesta (2009) en Pinar del Río, donde
encontró 0,17 juveniles por cada adulto. Acosta y Ponce de León (2004) exponen
que para los juveniles el porcentaje de mortalidad es alrededor del 75 %. Miller et al.
(2001) en estudio realizado en el norte del valle central de California, EUA obtuvo
que el éxito de reproducción para la especie fue de un 22 a un 44 %. Tomlinson
(1994) describe que la supervivencia de los juveniles de Paloma Rabiche está entre
un 24 y un 44 %. La relación de edades media de esta paloma en USA, obtenida de
muestras al azar de cazadores, es de 1,9 jóvenes/adulto (Miller and Otis, 2010) y con
estimaciones de reclutamiento de hasta 4,1 pichones/hembra adulta (Schuz et al.,
2013).
Esta relación tan baja encontrada en este agrosistema cubano no se debe tomar de
forma categórica y estos resultados deben ser interpretados con cautela, ya que
según Nadal (1994) citado por Chalaux (2005), teóricamente, las razones de edad
tendrían que estar relacionadas con la calidad del hábitat; los hábitats de buena
calidad permiten que el ave desarrolle una eficiencia biológica alta; es decir, que
tenga un éxito reproductor y produzca excedentes poblacionales. Por lo que sería de
esperar que debido a la calidad del área donde se desarrolló el estudio tuviera un
mayor éxito reproductor. En estos resultados pudiese estar influyendo la movilidad

99
de los juveniles, los que según nuestro juicio durante los primeros tiempos luego de
la época de independencia de los padres, presentan una menor movilidad que los
adultos. Otro aspecto que pudiese tener influencia en estos resultados podría estar
relacionado con diferentes factores como el tipo de aprovechamiento a que es
sometida la especie, los daños antrópicos presentes en el área de estudio y factores
meteorológicos adversos. Se sugiere que en futuros estudios sobre la especie se
tomen en cuenta estos elementos.
6.4.- Conclusiones y recomendaciones
1. A pesar de la presión de la caza y la mortalidad natural, la especie resulta ser la
más abundante en el área de estudio.
2. La razón de sexos (M/H) calculados en la población de la especie estudiada
(1,17), a pesar de mostrar desequilibrio a favor de los machos, pudiese presentar
ventajas como la elección de los mejores dotados, asegurando una mejor calidad
en la población ya que es una tendencia evolutiva de todas las especies elegir a
la pareja mejor dotada.
3. La razón de edad (J/A) calculados en la población de la especie estudiada (0,23),
demuestra el no reemplazo total de la población, lo que debe tenerse en cuenta
en la planificación de las capacidades de aprovechamiento.
4. Los valores obtenidos en las razones de sexo y edad en el área de estudio son
similares a los obtenidos en Pinar del Río al estudiar la especie, lo que evidencia
confiabilidad en estos resultados, siendo de esperar en similares condiciones
climáticas, con una política agraria uniforme y con idénticos manejo y forma
tradicional de aprovechamiento de la especie esta similitud.
5. A pesar de la aparente no existencia de dimorfismo sexual en la especie, se
recomienda su empleo por personal adiestrado en el manejo de la especie, el
uso de las diferencias en el plumaje en las clasificaciones por sexos en adultos y
jóvenes o adultos en la clase de edad.

100
6.5.- Bibliografía
Acosta, M; P. de león, J.L. (2004). Historia Natural de la Ploma Rabiche (Zenaida
macroura). (Inedito). Facultad de Biología. Universidad de la Habana. Cuba.
Cereza, J. (2005). Estudio de la dinámica poblacional de la codorniz (Coturnix c.
coturnix) en cotos de Burgos, León, Palencia, Soria, Teruel, Badajoz, Sevilla,
Cadiz, Ceuta, Cuenca y Valencia durante la media veda. 2003. TPT. Ingeniería
Técnica Forestal. Universidad de Lleida. España.
Chalaux, J. (2005). Dinámica poblacional de la perdiz roja (Alectoris rufa) en la
provincia de Burgos mediante el análisis de muestras biológicas de animales
capturados durante la temporada 2002-2003. TPT. Ingeniería Técnica Forestal.
Universidad de Lérida. España.
Cuesta, L. A. (2009). Perspectivas hacia una Gestión Sostenible de la Actividad
Cinegética en la Provincia de Pinar del Río, Cuba. Universidad de Alicante,
España, Universidad de Pinar del Río, Cuba.
Gonzáles, A., Berovides, V., Alonso, M. y López, M. (2013). Protocolo para el
monitoreo de Cyclura nubila nubila. Centro Nacional de Áreas Protegidas. La
Habana. Cuba. 53pp.
Miller M. R, C. L. Stemiery D. S. Blankenship (2001). Mourning dove productivity in
California during 1992-95: Was it sufficient to balance mortality?, Journal of
Wildlife Management, 65 (2), 300 – 311.
Miller, D.A.; Otis, D.L. (2010). Calibrating Recruitment Estimates for Mourning Doves
From Harvest Age Ratios. The Journal of Wildlife Management, 74 (5), 1070-
1078.
Peiró, V. (2003). Gestión ecológica de los recursos cinegéticos. Publicaciones de la
Universidad de Alicante. España. 136 pp.
Schulz, J. H., T. W. Bonnot, T. W. Mong and J. J. Millspaugh. (2013). Statewide and
local recruitment of mourning doves in Missouri. The Journal of Wildlife
Management, 77 (2), 336-345.

101
Tomlinson, R. et al. (1994). Mournig Dove. Migratory shore an upland game bird
management in North. America. International Association of Fish and Wildlife
agencies. Allen Pres Lawrence, Kansas. E.U.A. 5-26 pp.
.

102
CAPÍTULO VII. ECOLOGÍA TRÓFICA DE LA PALOMA RABICHE (Zenaida
macroura L.)
7.1.- Introducción
Uno de los aspectos determinantes en el desarrollo de las poblaciones es la
posibilidad de uso de los recursos tróficos, por lo que el estudio de la dieta se ha
convertido en uno de los aspectos principales de la autoecología de las especies
animales (Dennis (2000) en Acosta y García 2005).
Además esta información resulta indispensable para conocer la función que tiene
cada especie en los ecosistemas, así como para tomar medidas concretas para su
manejo (González et al. (2001) en Acosta y García 2005).
Dentro de los aspectos de mayor importancia respecto a el hábitat de una especie,
está el estudio del nicho trófico por su importancia para el manejo y utilización de las
poblaciones naturales, ya que según Peiró (2003) “La variación en la calidad del
hábitat tiene efecto sobre los parámetros de la dinámica poblacional, alterando la
supervivencia y la producción de jóvenes, y limitando el tamaño de la población”.
Al identificar la composición y estructura de la dieta de la Paloma Rabiche nos
permitió conocer el impacto agrícola sobre los agrosistemas donde esta se
desarrolla, además de poder realizar una buena gestión en su aprovechamiento
cinegético.
A pesar de tratarse de una especie sometida a un elevado uso consuntivo, faltan
publicaciones relacionadas con su ecología, y en particular sobre sus hábitos
alimentarios, lo que motivó la realización del presente estudio. Por estas razones el
objetivo de este trabajo es analizar la composición y estructura de la dieta de la
Paloma Rabiche, para a partir de esto conocer el uso alimentario e impacto agrícola
de la especie sobre el agrosistema cubano, lo que permitirá en un futuro realizar
propuestas de mejora de los paisajes agrarios tropicales. Este capítulo ha sido objeto
de un artículo científico publicado en la revista Journal of Animal and Plant Sciences

103
en 2016, indexada el Cuartil Q3 por el Institute for Scienctific Information/Journal
Citation Reports. (ISI/JCR) de Thomson-Reuters (Anexo 9).
7.2.- Materiales y métodos
Los trabajos de campo consistieron en la toma de muestras de los ejemplares
cazados de esta paloma, los cuales son descritas en el capítulo V. Conducta y
Reproducción, a cada ejemplar se le extrajo el buche, colocando su contenido en un
recipiente etiquetado con alcohol al 70 % durante unas 72 horas antes de proceder a
su procesamiento. El pesaje de los contenidos alimentarios se hizo en condiciones
de laboratorio y se empleó una balanza analítica de 0,01 g. de precisión. El
contenido de cada buche se colocó sobre papel de filtro, después de seco se separó
y pesó cada tipo de alimento, identificándolo hasta la categoría de familia y especie.
Para determinar el tamaño de muestra adecuado se usó el índice de llenura (Ill)
según Godínez (1985), permitiendo determinar si la muestra fue suficiente para
analizar la composición del espectro alimentario y la frecuencia de especies que
consume esta paloma
Ill = Pt / Pe
donde:
Pt = representa el peso seco total de todos los elementos encontrados en un ave
Pe = representa el peso del ejemplar
Este índice ofrece una muestra de la cantidad de biomasa de alimentos que necesita
una especie consumidora para mantener su peso corporal, además del uso que
hace esta paloma de un determinado hábitat y la calidad de este, al tener en cuenta
su relación con el peso de las aves.
El inventario de la flora en los biotopos presentes en la zona se desarrolló por el
método propuesto por Mostacedo y Fredericksen (2000). Así, en las áreas de
cultivos agrícolas y potreros se realizó por el método de la selección de parcelas en
cuadrantes, de 1,30 x 1,30 m; donde se identificaron cada una de las especies
presentes. En el área boscosa se realizaron transectos lineales de 500m de longitud

104
y 4m de ancho distribuidos al azar, garantizando así la representación de todas las
especies.
Las muestras de los diferentes artículos alimentarios encontrados en la dieta de la
especie estudiada fueron identificadas por la DraC. Ramona Oviedo, en el Herbario
HAC del Instituto de Ecología y Sistemática (IES), del CITMA. Cuba.
La caracterización de la vegetación es fundamental cuando se estudia la ecología de
especies cinegéticas, porque proporciona información relevante sobre las
condiciones naturales, principalmente de alimentación y refugio que ésta le brinda a
la especie estudiada. Realizar esta indagación permitirá definir la disponibilidad de
las especies vegetales más importantes en la zona en estudio para la Paloma
Rabiche, las cuales influyen en la dinámica poblacional, y por tanto determinan la
calidad del hábitat.
El análisis de la componente mayoritaria del espectro alimentario de esta especie
de paloma se realizó mediante los índices ecológicos de riqueza y diversidad de los
artículos vegetales consumidos.
Riqueza: S = Número de especies vegetales
Índice de diversidad de Shannon – Wiener (con base logaritmo natural):
H' = Σ pi x ln pi
donde:
pi = Ni / N
pi = representa la abundancia de la especie vegetal i
Ni = representa el número de individuos de la especie i
N representa el número total de individuos en toda la muestra o comunidad vegetal
La preferencia trófica de la especie se determinó por el índice de importancia
alimentaria (Ia) según Acosta y Berovides (1982):
Ia = ai / N + pi / PN + ni / Nt

105
donde:
ai representa la abundancia del elemento i en la muestra
N representa la cantidad de elementos encontrados en la muestra
pi representa el peso del elemento i en la muestra
PN representa el peso total de todos los elementos de la muestra
ni representa el número de casos donde aparece el elemento i
Nt representa el número de casos totales
Este índice constituye un importante objeto de estudio por la posibilidad que ofrece
de identificar los principales componentes de la dieta de una especie y así proyectar
el manejo de su hábitat, de manera que se garantice una disponibilidad adecuada y
permanente de los alimentos más importantes.
A los datos obtenidos se aplicaron primeramente técnicas de estadística descriptiva,
con la finalidad de resumirlos, describirlos numéricamente y facilitar su interpretación
(medidas de tendencia central y de variabilidad). Luego se les realizó un análisis
exploratorio de los datos a través de los test de normalidad Shapiro-Wilk W. y
Kolmogorov-Smirno, y con el fin de conocer el comportamiento de la distribución y
así determinar el tipo de análisis estadístico a utilizar (paramétrico o no paramétrico).
Para analizar el efecto que puede tener en una variable aleatoria, una variable
cualitativa que actúe como factor se aplicó finalmente el test de U Man-Whitney. En
el procesamiento de la información se emplearon los programas Excel y SPSS
versión 15.0 para Windows.
7.3.- Resultados y discusión
Del total de los 78 ejemplares obtenidos, todas las muestras fueron válidas para el
estudio de la dieta de la Paloma Rabiche. El índice de llenura de Godínez (1985)
presenta una estabilidad en el comportamiento de sus valores medios con un
tamaño de muestra alrededor de los 20 ejemplares, lo cual es indicativo de que se
está en posesión de una muestra suficiente para analizar la composición del
espectro alimentario y la frecuencia de los artículos que la componen.

106
Figura 23. Comportamiento de los valores medios del Índice de llenura (Ill) según el
tamaño de muestra.
Del comportamiento del índice de llenura en los diferentes meses del año (Tabla 15)
se destaca los valores altos consecutivos en los meses de abril, mayo y junio. Este
periodo se asocia con la época de máxima reproducción de la especie, con
presencia de pollos (pichones), lo que requiere un mayor consumo energético.
Además coincide con la primavera, época del año donde a pesar de no existir
temperaturas tan altas y lluvias tan abundantes como en verano, estos factores en
dicha estación determinan la rotación y maduración de los cultivos, así como la
presencia y fructificación de plantas silvestres presentes en el espectro alimentario
de la Paloma Rabiche, condicionando la disponibilidad de alimentos para la especie.
Similares resultados obtuvieron Acosta y Torres (1984) en el Jardín Botánico de
Cienfuegos, Cuba, donde la Paloma Rabiche consumió casi el doble (26) de tipos
diferentes de alimentos en época de lluvias. Ante esta evidente sincronización
estacional puede inferirse en la especie un carácter oportunista y un gran éxito
adaptativo, lo que es concordante con los estudios de Abu Baker y Brown (2009).

107
Tabla 15. Cambios mensuales del índice de llenura (Ill) de la Paloma Rabiche.
Meses Media Desviación
Estándar
Intervalo de
confianza (95%)
Inferior Superior
enero 0.0775 0.0183 0.0558 0.1143
febrero 0.0462 0.0409 0.0099 0.0824
marzo 0.0381 0.0244 0.0176 0.0585
abril 0.0687 0.0490 0.0277 0.1096
mayo 0.0595 0.0405 0.0170 0.1022
junio 0.0648 0.0321 0.0378 0.0916
julio 0.0200 0.0188 0.0017 0.0332
agosto 0.0458 0.0279 0.0163 0.0754
septiembre 0.0601 0.0326 0.0078 0.1118
octubre 0.0442 0.0337 0.0022 0.0861
noviembre 0.0325 0.0263 0.0034 0.0522
diciembre 0.0264 0.0283 -0.0088 0.0616
El tratamiento estadístico del Índice de llenura según el sexo de las palomas
mediante el test no paramétrico de U Man-Whitney no muestra diferencias
significativas (p= 0,580, p > 0,05) lo que concuerda con una estrategia de
alimentación similar en ambos sexos constatada en otras especies de género
Zenaida en ambientes tropicales (Cezilly and Keddar, 2012). El mismo análisis
estadístico aplicado a este índice comparando entre las clases de edad
(jóvenes/adultos) tampoco fue significativo (p= 0,486, p > 0,05). Es importante
destacar que índice de llenura medio (0,0617) en palomas jóvenes (pichones) es
algo mayor que el consumo promedio (0,0466) en aves adultas, coincidiendo con los
resultados obtenidos por Best y Smartt (1986) en Nuevo México. La relación de
edades media de esta paloma en USA, obtenida de muestras al azar de cazadores,

108
es 1,9 jóvenes/adulto (Miller y Otis, 2010) y con estimaciones de reclutamiento de
hasta 4,1 pichones/hembra adulta (Schulz et al., 2013). Estas diferencias en las dos
clases de edad están relacionadas con los requerimientos metabólicos del proceso
de crecimiento y desarrollo que experimentan los pichones (Miller, 2010; 2011).
Tabla 16. Comparación de las medias de Índice de llenura para sexo y clase de
edad.
Sexo Población
Clase de edad
Adultos Pichón
N Media N Media Máx Mín N Media Máx Mín
Macho 47 0.0468 37 0.0418 0.0991 0.0010 10 0.0655 0.1052 0.0203
Hembra 29 0.0534 25 0.0536 0.1299 0.0054 4 0.0522 0.1685 0.0064
TOTAL 76 0.0494 62 0.0466 0.1299 0.0010 14 0.0617 0.1686 0.0064
En la Tabla 17 aparecen reflejados los porcentajes de cada categoría de especies
vegetales obtenidas en los inventarios de vegetación realizados en el área de
estudio, así como el total de especies presentes en la zona, y de éstas cuantas
utiliza la Paloma Rabiche como alimento.
El 25 % de las especies vegetales inventariadas sirven de alimento a la Paloma
Rabiche en el área agrícola con cobertura donde existe un predominio de las
especies herbáceas (70%), que constituyen la principal fuente de alimentos de esta
paloma en la zona de estudio. También se encuentra una disponibilidad
representativa de especies arbóreas y arbustivas en este biotopo, pero sólo una
arbustiva (Erytroxilum havanensis) tiene uso alimentario. Las plantas arbustivas y
arbóreas se ubican fundamentalmente en linderos o setos que bordean los caminos
rurales y las cercas aisladas en las áreas de potreros, que sirven a esta paloma
como perchas para el descanso, refugio y sustrato para la nidificación. Todas estas
características del paisaje agrario las podemos traducir en una excelente calidad de
hábitat para la población de la especie estudiada.

109
Tabla 17. Porcentaje de especies vegetales disponibles por biotopos y su
representación en la alimentación de la Paloma Rabiche.
Biotopos
%
Herbáceas
%
Arbustivas
Especies
Arbóreas
Sp
Trepadoras
Epifitas
Total
No
Sp
% Sp
Alimento
paloma
Agrícola con cobertura
70 13 11 6 0 84 25
Forestal Típico
16 32 36 11 5 44 5
Total Área estudio
57 17 18 6 2 101 21
* El total de cada biotopo y de la zona de estudio procede del sumatorio de especies en cada fila
En el biotopo Forestal típico sólo un 5 % de especies vegetales sirven de alimento a
la Paloma Rabiche y con un predominio en el medio de las especies arbóreas y
arbustivas (68%), lo que muestra un biotopo de baja calidad para la especie
estudiada, dado su pobre potencial alimentario, excepto en los claros presentes
donde existe abundancia de especies herbáceas.
Estos resultados son apoyados en los consejos expuestos por Yarrow (2009) a los
propietarios de EUA que deseen atraer a la especie:
Fácilmente en la mayoría de los casos se pueden combinar la atracción de las
palomas con el resultado de las prácticas agrícolas y recolección de cosechas
normales.
Las siguientes son algunas maneras adicionales de mejorar la tierra para la Paloma
de Rabiche:
1. Favorecer el crecimiento de arbustos, matas de pinos y cercas tupidas para
anidar.
2. Mantener áreas aradas y campos de barbecho para fomentar el crecimiento de
pastos nativos y las malas hierbas como la alimentación paloma.
3. Deja esquinas de campos cultivados con vegetación sucesional temprana para
favorecer a las plantas productoras de alimentos preferidos por las palomas.

110
4. Como se puede apreciar todos estas recomendaciones se sustentan en la base
de una adecuada gestión de un sistema agrícola donde predomine cierto nivel de
árboles dispersos, cercas vivas plantas herbáceas silvestres (el típico biotopo
Agrícola con cobertura).
La Tabla 18 presenta el contenido alimentario con un total de 33 artículos, de los que
el 88 % son de origen vegetal (semillas, frutos, brotes tiernos y bulbo), el 9%
corresponden a alimentos de origen mineral (fragmentos de cuarzo, perdigones y
pequeños terrones de arcilla) y el 3 % de origen animal (fragmentos de conchas de
caracoles). Al comparar los resultados obtenidos en los alimentos de origen animal
con los expuestos por Acosta y Ponce de León (2004) sólo existe coincidencia en el
consumo de caracoles terrestres; ellos plantean que además ingieren huevos de
saltamontes, hormigas, escarabajos e isópodos (Cochinillas).
Cuesta (2009) al analizar la ecología trófica de la especie en Pinar del Río, encontró
un total de 37 artículos, de los que el 87 % son de origen vegetal (semillas y granos)
y un 13 % de origen mineral (gastrolitos de 5 tipos diferentes). Al comparar ambas
localidades a pesar del menor número de artículos hallados en el área de estudio,
esta posee una mayor diversidad que en Pinar del Río. Es importante destacar que
la base fundamental de la dieta de la especie en las dos áreas estudiadas consistió
en alimentos de origen vegetal.
Tabla 18. Clasificación de los artículos, ordenados según el índice de importancia
alimentaria (Ia), que conforman el espectro de alimentación de la Paloma Rabiche.
I.A. Artículo
alimentario
Nombre
común en
Cuba
Familia
botánica Especie
1.7367 Semillas
Lechosa Euphorbiaceae Euphorbia heterophylla
1.2952 Semillas
Hierba guinea Graminaceae Panicum maximun
0.7130 Semillas
Tua - tua Euphorbiaceae Croton lobatus
0.6839 Semillas
Arroz Graminaceae Oryza sativa
0.6832 Semillas
Canutillo Graminaceae Rottboellia conchinchinensis
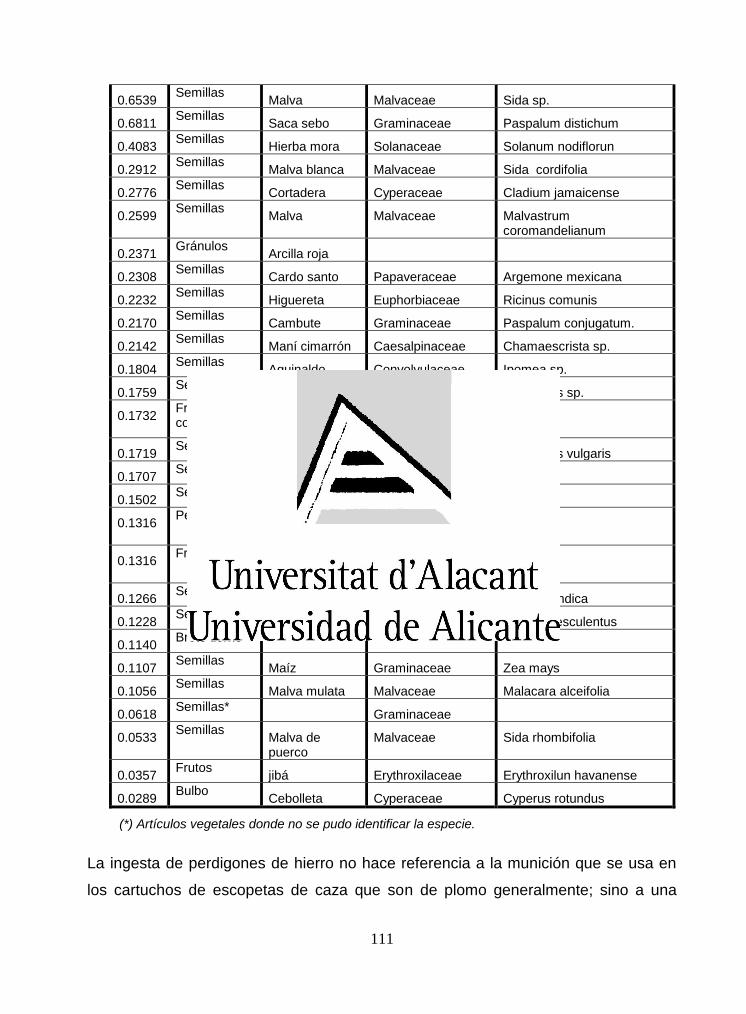
111
0.6539 Semillas
Malva Malvaceae Sida sp.
0.6811 Semillas
Saca sebo Graminaceae Paspalum distichum
0.4083 Semillas
Hierba mora Solanaceae Solanum nodiflorun
0.2912 Semillas
Malva blanca Malvaceae Sida cordifolia
0.2776 Semillas
Cortadera Cyperaceae Cladium jamaicense
0.2599 Semillas
Malva Malvaceae Malvastrum coromandelianum
0.2371 Gránulos
Arcilla roja
0.2308 Semillas
Cardo santo Papaveraceae Argemone mexicana
0.2232 Semillas
Higuereta Euphorbiaceae Ricinus comunis
0.2170 Semillas
Cambute Graminaceae Paspalum conjugatum.
0.2142 Semillas
Maní cimarrón Caesalpinaceae Chamaescrista sp.
0.1804 Semillas
Aguinaldo Convolvulaceae Ipomea sp.
0.1759 Semillas
Frjol silvestre Phabaceae Phaceolus sp.
0.1732 Frag, conchas
Caracoles terrestres
0.1719 Semillas
Frijol negro Phabaceae Phaceolus vulgaris
0.1707 Semillas
*
Passifloraceae
0.1502 Semillas
Malva Malvaceae Sida sp
0.1316 Perdigones
Perdigones de hierro
0.1316 Fragmentos
Piedras de Cuarzo
0.1266 Semillas
Malva blanca Sterculiaceae Walteria indica
0.1228 Semillas
Chufa Cyperaceae Cuperus esculentus
0.1140 Brote tierno
0.1107 Semillas
Maíz Graminaceae Zea mays
0.1056 Semillas
Malva mulata Malvaceae Malacara alceifolia
0.0618 Semillas*
Graminaceae
0.0533 Semillas
Malva de puerco
Malvaceae Sida rhombifolia
0.0357 Frutos
jibá Erythroxilaceae Erythroxilun havanense
0.0289 Bulbo
Cebolleta Cyperaceae Cyperus rotundus
(*) Artículos vegetales donde no se pudo identificar la especie.
La ingesta de perdigones de hierro no hace referencia a la munición que se usa en
los cartuchos de escopetas de caza que son de plomo generalmente; sino a una

112
pequeña concreción de una roca ferrosa que abunda mucho en los suelos de Cuba.
Los perdigones de caza no han aparecido en nuestro análisis alimentario de esta
paloma en Cuba. No obstante estudios de campo con individuos de Zenaida
macroura cazados han mostrado que el porcentaje de aves con perdigones de plomo
ingeridos es de 0,2 a 6,5% (Plautz et al., 2011).
La base alimentaria de la especie en el área de estudio se caracteriza por ser
fundamentalmente de origen vegetal, compuesta en un 79 % por semillas de plantas
herbáceas y mostrando preferencia por las semillas pequeñas sobre otros alimentos
de origen vegetal.
Similares resultados exponen Acosta y Ponce de León (2004), quienes afirman que
el 99 % de los alimentos consumidos por la especie son de origen vegetal (semillas y
otras partes de plantas).
Al realizar una comparación entre los diversos elementos de origen vegetal que
forman parte de la dieta de Zenaida macroura en el área de estudio y los resultados
obtenidos por Cuesta (2009) en Pinar del Río (Tabla 19) se obtiene un espectro
alimentario de esta paloma compuesto por un total de 15 familias botánicas, no
obstante aparece una diferencia en cuanto a la representación de 7 familias
botánicas entre ambos estudios, contando el área de estudio con 1 familia más que
Pinar del Río. Un caso diferente se muestra si se analiza el espectro alimentario
respecto a la variedad de especies vegetales, siendo superior este caso la dieta en
Pinar del Río (4 especies más). Los resultados obtenidos evidencian como la
disponibilidad y acceso a los alimentos según las características de cada territorio
pueden determinar ciertas variaciones en la dieta de Zenaida macroura, aunque se
mantenga constante su base alimentaria.

113
Tabla 19. Representación de familias botánicas presentes en el espectro alimnetario
de Zenaida macroura en el área de estudio y Pinar del Río.
Familias
botánicas
Cienfuegos Pinar del Río
Presencia No.
especies Presencia
No.
especies
Amaranthaceae X 1
Caesalpinaceae X 1
Convolvulaceae X 1 X 1
Cyperaceae X 3 X 7
Erythroxilaceae X 1
Euphorbiaceae X 3 X 5
Graminaceae X 7 X 6
Malvaceae X 6
Papaveraceae X 1
Passifloraceae X 1 X 1
Phabaceae X 2 X 7
Scrophulariaceae X 1
Solanaceae X 1 X 1
Sterculiaceae X 1 X 1
No clasificada X 1
TOTAL 15 12 28 11 32
Se constata (Tabla 19) que hay dos elementos tróficos cuyo índice de importancia
alimentaria es superior (> 1) al resto: las semillas de Lechosa (Euphorbia
heterphylla) y de Hierba Guinea (Panicum maximun). La Lechosa, especie herbácea
que aparece espontáneamente en campos de cultivos, tierras en barbecho, terrenos
arados y en bordes de caminos, es considerada una mala hierba por los agricultores.
La Hierba de Guinea produce abundantes semillas de pequeño tamaño y es una
planta perenne que crece espontáneamente en terrenos fértiles y húmedos, además

114
se considera el mejor pasto de Cuba. Este Panicum se puede encontrar en las áreas
de cultivos agrícolas en zonas de barbecho, cultivos abandonados y bordeando
caminos, en ocasiones también aparece como especie asociada en los linderos o
setos y en los claros de los bosques. También es de destacar el Arroz (Oryza
sativa), un cultivo ampliamente diseminado en el territorio que, aunque tiene ciertas
épocas de cosechas preferenciales, está más o menos presente todo el año en el
área de estudio y constituye un alimento de importancia para la Paloma Rabiche.
Acosta y Berovides (1982) refieren en el estudio realizado al sur de la provincia de
Pinar del Río, Cuba, que el arroz ocupó el primer lugar en cuanto a volumen en la
dieta de esta especie de paloma.
Del análisis de los resultados obtenidos por Cuesta (2009) y su comparación con los
de este trabajo, resulta evidente la importancia de la familia botánica Graminaceae
en la dieta de esta paloma, representada en ambas áreas de estudio por más de 5
especies de plantas. Otro resultado que confirma lo antes expuesto es que en
ambos casos esta familia ocupa los primeros lugares en orden de importancia de los
índices de preferencia alimentaria.
En USA, Hayslette (2006) y Hayslette y Mirarchi (2002) estudiaron la selección
trófica según el tamaño de las semillas y encontraron también una preferencia de la
Paloma Rabiche por semillas pequeñas como el Trigo (Triticum aestivum) y el Mijo
(Panicum miliaceum) sobre otras de mayor tamaño como el Maíz (Zea mays). Según
Ostrand et al. (1998), esta paloma se alimenta principalmente en los campos de
cultivo de trigo y en los parches de malezas, y que una disminución en la
disponibilidad de trigo durante la primavera ha contribuido a una disminución de la
población de este especie. Nuestro estudio también muestra algunos artículos
vegetales de mayor tamaño como el Maíz y el fruto de Jibá (Erythroxilun havanense)
en el espectro alimentario de esta paloma, pero están dentro de las de menor
importancia alimentaria.
La riqueza vegetal del espectro alimentario de la Paloma Rabiche en el área de
estudio fue de 29 especies 28 de semillas de diferentes plantas y 1 brote tierno) de
las cuales solo 3 (10%) son cultivos agrícolas. Este resultado pone de manifiesto la

115
importancia de la Paloma Rabiche por su bajo impacto para la agricultura, también,
porque ésta toma las semillas directamente del suelo, no escarba ni quiebra espigas
de cultivos y sólo consume restos de cosechas. Además, esta Columbidae consume
gran cantidad de semillas de especies herbáceas que crecen en las zonas de
cultivos agrícolas y que son consideradas maleza por los agricultores, y al poseer un
estómago muscular bien desarrollado, tienen la capacidad de desintegrar
prácticamente todas las semillas ingeridas, evitando así la propagación de estas
“malas hierbas”. Blockstein et al. (1987) determinaron que muy pocas semillas de
todas las consumidas atravesaron el sistema digestivo de las palomas, y que sólo
una pequeña cantidad de semillas se recuperaron en la materia fecal encontrada en
los nidos, por lo que ellos consideran que esta especie de paloma raramente actúa
como agente dispersante de semillas, y en particular de la dispersión de plantas
invasoras de medios agrarios a excepción de algunas plantas exóticas introducidas
en América como Commelina benghalensis (Goddard et al., 2009). La dieta de la
Paloma Rabiche en el área de estudio es moderadamente diversa (H’ = 1,738
nats/individuo), lo que puede ser una respuesta a la cantidad y distribución en que se
encuentra disponible el alimento vegetal en su hábitat, lo que garantiza la
supervivencia de la especie al permitirle una utilización más amplia de los recursos
presentes en el hábitat.
Según nuestro estudio, la Paloma Rabiche es una especie de caza menor muy
asociada a agroecosistemas en Cuba. Esto contrasta con los estudios sobre otras
especies de la familia Columbidae en los que se ha estudiado la interacción con
diferentes unidades estructurales del paisaje según Jiménez et al. (2006) y Belda et
al. (2011). Así, en paisajes donde domina la matriz de vegetación natural, la
densidad de Paloma Torcaz (Columba palumbus) está influenciada positivamente
por extensas zonas forestales, pero también se acomoda a ambientes deforestados
por la agricultura (Larruy; Burgas, 2004). No obstante, la biodiversidad animal en los
paisajes agrícolas se puede aumentar con la conversión de algunos terrenos de baja
producción en tierras más naturales o gestionadas extensivamente; pero no se sabe
en qué medida esta biodiversidad y la abundancia de palomas se puede mejorar

116
mediante la alteración patrón del paisaje sin reducir la producción agrícola (Belda et
al,. 2011; Fahrig et al., 2011). La Tórtola Europea (Streptopelia turtur) prefiere una
matriz de matorral arbolado con intercalación de frutales de regadío, pero evita el
matorral denso debido a la falta de recursos tróficos adecuados. Esta especie de
tórtola coincide en su preferencia de hábitat con la subespecie occidental de la
Paloma Rabiche en USA, Zenaida macroura marginella (Meyers et al., 2005). La
Tórtola Turca (Streptopelia decaocto) habita zonas urbanas con jardines con árboles
o con parches de bosque, cerca de las zonas de asentamiento urbano (que son
preferidos como áreas de nidificación). Un paisaje de mosaico preferido por esta
Columbidae incluye tierras agrícolas cerca de bosques y parches urbanos. Esta
intercalación de hábitats proporciona áreas de alimento, nidificación y refugio. El
aumento de la urbanización dentro masas de repoblación forestal densa está
favoreciendo la densidad de esta tórtola (Belda et al., 2011). Sin embargo, los
estudios experimentales en cautividad muestran un alto grado de solapamiento de la
dieta y preferencias alimentarias similares de la Tórtola Turca con la Paloma Rabiche
(Poling and Hayslette, 2006).
7.4.- Conclusiones y recomendaciones
1. De los biotopos presentes en el área de estudio, el de mayor calidad trófica para
la especie de paloma estudiada es el Agrícola con cobertura.
2. A partir de los 20 ejemplares se cuenta con un tamaño de muestra que puede
proporcionar información cuantitativa confiable sobre el contenido del buche en la
población de la Paloma Rabiche.
3. El uso trófico de la Paloma Rabiche se caracteriza por ser fundamentalmente de
origen vegetal y compuesto en su mayoría por semillas de plantas herbáceas.
Este espectro alimentario es moderadamente amplio según el índice de
diversidad. Las semillas de Euphorbia heterphylla y de Panicum maximun son los
elementos de mayor importancia alimentaria para la Paloma Rabiche.

117
4. El índice de llenura varía con la estación (mes) del año. Ambos sexos se
comportan de manera similar respecto a la cantidad de alimentos ingeridos.
Corresponde a los jóvenes (pichones) el mayor índice de consumo de alimentos.
5. La Paloma Rabiche causa un impacto trófico agrícola (daño en los cultivos) bajo,
además de consumir semillas de plantas consideradas maleza por los
agricultores, por lo que se puede concluir que es una especie beneficiosa en los
agrosistemas tropicales.
6. La utilización que hace la Paloma Rabiche de los artículos de mayor importancia
alimentaria presenta diferencias significativas respecto a los meses del año. Se
recomienda continuar con el estudio de la actividad trófica de la especie,
haciendo énfasis en las variaciones estacionales de las especies que forman su
espectro alimentario en función de un manejo sostenible de la especie.
7.5.- Bibliografía
Abu Baker, M. A., Brown, J. S. (2009). Patch area, substrate depth, and richness
affect giving-up densities: a test with mourning doves and cottontail rabbits.
Oikos, 118 (11), 1721-1731.
Acosta, M. y García, L. (2005). Morfometría y alimentación de la codorniz (Colinus
virginianus) en dos áreas del occidente de Cuba. Journal of Caribbean
Ornitholog, 18. 54-68.
Acosta, M; Berovides, V. (1982). Ecología trófica de las palomas del género Zenaida
en el sur de Pinar del Río, Cuba. Ciencias Biológicas, 7, 113-123.
Acosta, M; Ponce de León, J.L. (2004). Historia Natural de la Paloma Rabiche
(Zenaida macroura). (Inédito). Facultad de Biología. Universidad de La Habana.
Cuba.
Acosta, M; Torres, O. (1984). Ecología trófica de las palomas del género Zenaida en
el Jardín Botánico de Cienfuegos, Cuba. Ciencias Biológicas, 11, 107-116.

118
Belda, A., J. E. Martinez-Perez, V. Peiro, E. Seva, and J. Arques. (2011). Main
landscape metrics affecting abundance and diversity of game species in a semi-
arid agroecosystem in the Mediterranean region. Spanish Journal of Agricultural
Research, 9 (4), 1197-1212.
Best, T; Smartt, R. A. (1986). Feending Ecology of Mourning Doves (Zenida
macroura) in Southeastem New Mexico. The Southwestern Naturalist, 31 (1), 33-
38.
Blockstein, D. E; Fay, P. K; Maxwell, B. D. (1987). Dispersal of leafy Spurge Seeds
(Euphorbia esula) by Mourning Doves (Zenaida macroura). Weed Science, 35
(2), 160-162.
Cuesta, L. A. (2009): Perspectivas hacia una Gestión Sostenible de la Actividad
Cinegética en la Provincia de Pinar del Río, Cuba. Universidad de Alicante,
España, Universidad de Pinar del Río, Cuba.
Cezilly, F.; Keddar, I. (2012). Vigilance and food intake rate in paired and solitary
Zenaida Doves Zenaida aurita. Ibi, 154 (1), 161-166.
Fahrig, L., J. Baudry, L. Brotons, F. G. Burel, T. O. Crist, R. J. Fuller, C. Sirami, G. M.
Siriwardena and J-L. Martin. (2011). Functional landscape heterogeneity and
animal biodiversity in agricultural landscapes. Ecology Letters,14, 101-112.
Goddard, R. H., T. M. Webster, R. Carter, and T. L. Grey. (2009). Resistance of
Benghal Dayflower (Commelina benghalensis) Seeds to Harsh Environments and
the Implications for Dispersal by Mourning Doves (Zenaida macroura) in Georgia,
U.S.A. Weed Science, 57 (6) 603-612.
Godínez, E. (1985). Tamaño de muestra de la Torcaza Cabesiblanca (Columba
leucocephala) para estudios biométricos y alimentarios en dos poblaciones de
Cuba. Revista Forestal Baracoa, 15 (2), 55-64.
Hayslette, S. E.; Mirarchi, R. E. (2002). Foraging-patch use and within-patch diet
selectivity in Mourning Doves, Zenaida macroura. Ecology. 83 (9), 2637-2641.

119
Haystte, S. E. (2006). Seed-size selection in Mourning Doves and Euroasian
Collored-doves. The Wilson Journal of Ornithology, 118 (1), 64-69.
Jiménez-García, D.; Martínez-Pérez, J.E.; Peiró; V. (2006). Relationship between
game species and landscape structure in the SE of Spain. Wildlife Biology in
Practice, 2 (2), 48-62.
Larruy, X. and Burgas, D., J. Estarda. V. Petrocchi., L. Brotons and S. Herrando
(2004). Tudó Columba palumbus. In: Atlas dels ocells nidificants de Catalunya.
1999-2002. Institut Català d’Ornitologia. Lynx Edicions, Barcelona (España).
264-265.
Meyers, P. M., Conover, M. R., Bissonette, J. A. (2005). Effect of perch sites on
mourning dove nest distribution. Western North American Naturalist, 65 (1), 64-
69.
Miller, D. A. (2010). Morphological plasticity reduces the effect of poor developmental
conditions on fledging age in mourning doves. Proceedings Royal Society
London B., 277, 1659–1665.
Miller, D. A. (2011). Immediate and delayed effects of poor developmental conditions
on growth and flight ability of juvenile mourning doves Zenaida macroura. Journal
of Avian Biology, 42 (2), 151-158.
Miller, D.A.; Otis, D.L. (2010). Calibrating Recruitment Estimates for Mourning Doves
From Harvest Age Ratios. The Journal of Wildlife Management, 74 (5), 1070-
1078.
Mostacedo, B; Fredericksen, T. S. (2000). Manual de métodos básicos de muestreo
y análisis en ecología Vegetal. Santa Cruz, Bolivia. 10-12.
Ostrand, W. D., P. M. Meyers, J. A. Bissonette and M. R. Conover. (1998). Changes
in Land Use as a Possible Factor in Mourning Dove population Decline in Central
Uthah. Journal of Field Ornithology, 69 (2), 192-200.
Peiró V (2003) Gestión ecológica de los recursos cinegéticos. Publicaciones de la
Universidad de Alicante. España. 136 pp.

120
Plautz, S. C.; Halbrook, R. S.; Sparling, D. W. (2011). Lead shot ingestion by
mourning doves on a disked field. The Journal of Wildlife Management, 75 (4),
779-785.
Poling, T. D.; Hayslette, S. E. (2006). Dietary Overlap and Foraging Competition
Between Mourning Doves and Eurasian Collared-Doves. The Journal of Wildlife
Management, 70 (4), 998-1004.
Schulz, J. H., T. W. Bonnot, T. W. Mong and J. J. Millspaugh. (2013). Statewide and
local recruitment of mourning doves in Missouri. The Journal of Wildlife
Management, 77 (2), 336-345.
Yarrow, G. (2009): Biología y manejo de la Paloma de Luto. Extensión de Recursos
Naturales y Forestales de la Universidad de Clemson. Carolina del Sur. USA.
.


121
CAPÍTULO VIII. CALIDAD DEL HÁBITAT DE LA PALOMA RABICHE (Zenaida
macroura L.)
8.1.- Introducción
El hábitat se define como el espacio ocupado por una determinada especie
(Morrison et al., 1992 citado por Camprodon y Subirachs, 2001). Debido a las
características propias de cada región en no toda su extensión será igual el
comportamiento poblacional, existiendo diferencias según la estructura y
composición de cada sector. Al respecto Camprodon y Subirachs (2001) exponen
“…no ha de extrañarnos, por lo tanto, que la modificación de la estructura de un
hábitat produzca cambios en la comunidad de vertebrados tanto más drásticos en
cuanto más se altere la composición florística y fisionómica de la vegetación”.
Al respecto Block et al. (1987) en Peiró et al. (1993) expresan que uno de los
aspectos importantes en el estudio del manejo de la fauna silvestre es el análisis del
uso o selección del hábitat. Dicho estudio asume que los organismos seleccionan las
partes de su medio que satisfacen mejor sus requisitos vitales, y que estos hábitats
comparten algunas características comunes.
Según Peiró 2003 en el estudio o análisis del hábitat se deben considerar los
factores que puedan ser importantes para la población cinegética (suelo, vegetación
y uso del territorio etc.). Muchos de los procedimientos para analizar el hábitat de las
poblaciones cinegéticas se basan en realizar censos mediante observaciones,
considerando solo los tipos de vegetación-cultivos y obtener índices de selección del
hábitat.
El conocimiento de los principios del funcionamiento natural de los ecosistemas,
comunidades y poblaciones biológicas es esencial para lograr una conservación
efectiva a todos los niveles (composición de especies, integridad estructural e
integridad ecológica), por ello hoy resulta urgente tanto para los técnicos de campo,
el personal que supervisa los programas de conservación y los cazadores dispongan
de un conocimiento común y claro; ya que difícilmente los esfuerzos por lograr un
manejo sostenible puedan atenuar o eliminar las amenazas presentes sin estas

122
herramientas. Es por tanto indispensable generar información de calidad sobre los
requerimientos de hábitat y de área de cada especie bajo régimen de manejo, pues
estas especies deberán contar con el hábitat natural necesario para sostener
poblaciones reproductivas estables.
Chamizo et al. (2011) concluyen que la evaluación de la correspondencia entre la
calidad de los sitios que utilizan las especies cinegéticas y su densidad poblacional
es de gran interés para la planificación de la caza y el uso sostenible de los recursos
cinegéticos.
En los estudios de vegetación en áreas de manejo de especies es necesario ir un
poco más allá de los clásicos inventarios que solo ofrecen datos sobre la existencia
de plantas en las diferentes formaciones vegetales, lo más razonable sería tener en
cuenta parámetros estructurales que permitirían medir los diferentes grados de uso
de las especies bajo manejo de cada biotopo, lo que constituiría una herramienta
para la planificación y el manejo.
La importancia de la calidad del hábitat sobre la Paloma Rabiche ha sido estudiada
en los Estados Unidos de América por diversos autores (Ostrand et al., (1998);
Haysiette y Mirarchi, (2002) y Ostrand et al., (1996). Al revisar el tema en Cuba son
pocos los estudios relacionados con el tema. Estos solo abordan aspectos
relacionados con los diferentes alimentos usados, la importancia de estos en la
dieta de la especie sin tener en cuenta la influencia de la estructura y características
específicas de la vegetación, así como, la cobertura del suelo, por lo que estos
antecedentes en nuestro país resultan insuficientes (Cuesta ,2009; Chamizo et. al.,
2011).
Esta aproximación al estudio de la distribución y uso de las aves con respecto a
diferentes elementos del paisaje agrario y sus diferentes estructuras permitirá la
creación de modelos que, con carácter predictivo, sean aplicables al conocimiento
del impacto que la gestión humana del medio tiene sobre la Paloma Rabiche.
El objetivo de este capítulo es determinar la relación entre las diferentes
características del hábitats del área de estudio a través de los grados de selección

123
de hábitat de la especie estudiada y tres parámetros conductuales (descanso,
nidificación y alimentación), lo cual facilitará la predicción de su distribución y
abundancia y la toma de decisiones prudentes respecto a la viabilidad o no de una
gestión en función de las características del hábitat.
8.2.- Materiales y métodos
Según Peiró (2003), muchos de los procedimientos para analizar el hábitat de las
poblaciones cinegéticas se basan en realizar censos mediante observación, donde
para cada contacto de los animales, se realiza un análisis considerando solo los
tipos de vegetación, cultivos etc. Y con estos datos obtener índices de selección de
hábitat para cada tipo mediante el cociente entre el porcentaje de contactos
observados de la especie cinegética y el porcentaje de superficie de cada tipo de
vegetación, cultivo etc. De este modo, se puede obtener información sobre los tipos
de hábitat seleccionados para alimentación, refugio, reproducción; si se considera el
tipo de actividad realizada por los contactos.
Se realizaron recorridos a pie por toda el área de estudio con el objetivo de evaluar
aspectos conductuales de la paloma Rabiche, luego fueron clasificados los hábitats
donde se realizaron los contactos y según el objetivo de cada conducta a evaluar:
Uso de los biotopos del área de estudio: alimentación, descanso y nidificación
Conducta alimentaria según la cobertura del suelo: suelo completamente desnudo,
cobertura parcial de vegetación y cubierta total vegetación.
La relación de la Paloma Rabiche con los diferentes biotopos presentes en el área
de estudio, así como con la cobertura del suelo se realizó a través del cálculo de
Índice de selección o uso (Isu) y el Índice de selección de cobertura (Isc) según Peiró
(2003) modificado por el autor, cuya fórmula es:
Isx = ni / Nt x 100
donde:
Isx = índice de selección de uso o sitio

124
ni = número de palomas vistas en la asociación vegetal i o determinado grado de
cobertura del suelo
Nt = total de observaciones de palomas en toda el área
Estos índices facilitaron la determinación de la calidad de los biotopos presentes en
el área de estudio para la Paloma Rabiche, a través de los diferentes grados de
selección de hábitat de la especie y en tres parámetros conductuales (alimentación,
descanso y nidificación).
8.3.- Resultados y discusión
En la Tabla 20 se refleja que el 88 % de los contactos detectados durante el estudio
del uso de los biotopos fue en el Agrícola con cobertura y solo un 12 % ocurrieron en
el Forestal típico. Este resultado confirma la importancia del biotopo Agrícola con
cobertura para la especie estudiada y reafirma lo descrito por Camprodon y
Subirachs (2001) cuando describen que con el examen de los elementos que
conforman los hábitats y su estructura, se comprende por qué ciertos lugares son
óptimos para las especies y otros no lo son, el hábitat óptimo, en primer lugar
incluye todos aquellos elementos que son necesarios para la vida de la especie.
Tabla 20. Índice de selección de uso (Isu) de la Paloma Rabiche en el área de
estudio (n=164 contactos).
Como se observa en la Tabla 21, resulta evidente el hecho de que cada uno de las
diferentes asociaciones vegetales presentes en el área de estudio cuente con
diferentes opciones y acceso a los recursos usados por la especie estudiada en
diferentes composición y número, condiciona en mayor o menor medida la
accesibilidad y utilización de estos por la especie estudiada. Esta diferenciación
Biotopos No.
contactos Isu
Agrícola con cobertura 145 88
Forestal típico 19 12
TOTAL 164 100

125
permite un uso más marcado de cada uno de los hábitats presentes, así como de los
biotopos en el área de estudio.
Tabla 21. Comportamiento del Índice de selección de uso (Isu) para tres conductas
de la Paloma Rabiche y en diferentes usos del territorio en el área de estudio
(n=164).
Elementos del
paisaje vegetal
Isu Total
Contactos Alimentación Descans
o
Nidificació
n
Cultivos agrícolas 100 - - 41 (25 %)
Linderos - 72 28 36 (22 %)
Potreros 72 28 - 25 (15 %)
Potreros inv. Marabú 23 42 35 43 (26 %)
Bosque 16 58 26 19 (12 5)
TOTAL 44 38 18 164 (100)
Al analizar de forma más detallada el Índice de selección por cada elemento del
paisaje vegetal del área de estudio (Tabla 21) y conocer el comportamiento de las
conductas antes descritas (alimentación, descanso y nidificación) se observa que la
alimentación fue la actividad con mayor frecuencia de observación (41), siendo la
única conducta observada en los cultivos agrícolas. Coincidiendo con lo descrito por
Acosta y Berovides (1982), esta paloma puede observarse todo el día en una
actividad constante que, por consiguiente, trae aparejado un consumo máximo de
alimento para suplir este gasto energético.
En cuanto a las selección de áreas dedicadas al descanso, las más importantes
resultaron ser los linderos o setos con un 72 % de índice de selección de sitios,
seguidos por las áreas de bosques, lo que está determinado por la presencia de
árboles de porte mediano y grande que sirven de atalayas a la especie (esta
conducta es más acentuada en los árboles del borde del bosque, ya dentro del
mismo la presencia de la especie es muy poca). Es importante destacar la ausencia
total de palomas en descanso en las áreas de cultivos agrícolas, donde la mayoría

126
de los cultivos son plantas herbáceas o tubérculos. En las áreas de potreros con
ausencia de invasión de marabú la especie presentó un valor bajo del índice de
selección, los contactos en esta conducta se realizaron en algún árbol aislado o en
los alambres de las cercas utilizados por los ganaderos para dividir las áreas de
potreros en cuartones.
En el área de estudio el comportamiento de la nidificación tuvo un resultado sin
grandes diferencias entre los linderos y los bosques, es importante aclarar que en las
áreas de bosque, donde mayor número de palomas con nidos se detectaron fueron
en los bordes del mismo. El mayor índice de selección (35 %) se detectó en los
potreros invadidos de marabú, donde la especie encuentra protección contra
depredadores naturales y daños antrópicos, así como abundante material para la
construcción del nido. El comportamiento de este índice fue nulo en las zonas de
cultivos agrícolas y las áreas de potreros donde no existen las condiciones
necesarias (ausencia de árboles, y cuando estos se encuentran fácil acceso de las
personas y depredadores).
Un aspecto que se considera juega un papel muy importante en la utilización de los
diferentes asociaciones vegetales por parte de la Paloma Rabiche al momento de
forrajear y alimentarse lo constituye el porcentaje de cobertura del suelo. Por esta
razón se decide realizar un primer intento de buscar un patrón entre los contactos
tenidos con la especie en los recorridos donde se evaluó la relación de la especie
con su hábitat a través del empleo del índice de selección de cobertura del suelo.

127
Tabla 22. Resultados del Índice de selección de cobertura del suelo (Isc) en la
alimentación en los diferentes elementos del paisaje vegetales en el área de estudio
por la Paloma Rabiche (n=72 contactos).
Elementos del
paisaje vegetal
Isc
Total
contactos
Suelo
completamente
desnudo
Cobertur
a
parcial
Cubierta
total
vegetación
Cultivos agrícolas 34 44 22 41 (57 %)
Linderos - - - -
Potreros 22 56 22 18 (25 %)
Potreros invadidos Marabú
10 90 - 10 (14 %)
Bosque 67 33 - 3 (4 %)
TOTAL CONTACTOS 21 (29 %) 38 (53 %) 13 (18 %) 72 (100 %)
Al realizar el análisis de los resultados del índice de selección de cobertura durante
el proceso de alimentación de la especie (Tabla 22), se aprecia una clara
preferencia por las áreas con cierto nivel de cobertura del suelo y una menor
preferencia por aquellos sitios con una cubierta vegetal total del suelo. Esta
preferencia alimentaria está condicionada por su modo de alimentación.
Como se explicó en el capítulo 7, la base alimentaria de la especie se caracteriza
por ser fundamentalmente de origen vegetal, compuesta mayoritariamente por
semillas pequeñas de plantas herbáceas. Al tomar las semillas directamente del
suelo, sin escarbar ni quebrar las espigas de los cultivos o restos de cosechas se
explica fácilmente su preferencia por aquellos lugares donde existen las especies
vegetales que le sirven de alimento y están disponibles las semillas en un suelo no
totalmente cubierto por donde le es posible desplazarse y localizar fácilmente las
semillas. Situación muy diferente ocurre cuando está paloma se encuentra en áreas

128
donde abundan las especies vegetales que sirven de alimento y encontrándose este
no tiene acceso al mismo o lo logran con dificultad, ya que las pequeñas semillas
permanecen bajo una cubierta vegetal. Similares resultados obtuvo Cuesta (2009)
donde al comparar los datos obtenidos, del peso de los alimentos consumido y del
índice de llenura con diferentes calidades de sitios para la Paloma Rabiche señala
como un factor clave la densidad del estrato herbáceo. El autor citado concluye que
esta densidad puede dificultar el acceso a los recursos, como es el caso de las
áreas ganaderas que a menudo imponen muchas dificultades a la especie para
acceder a los granos bajo una densa cubierta de pastos y vegetación espontánea.
Yarrow (2009) expone en sus recomendaciones a los propietarios de tierras que
deseen atraer a la especie la eliminación del exceso de vegetación mediante
incineración o aradura. Esta práctica expone el grano disponible en el suelo y hace
que sea fácilmente accesible para las aves, debido que estas prefieren bajar,
caminar y alimentarse en un suelo relativamente libre de vegetación densa.
Ostrand et al. (1996) con el objetivo de conocer la estructura del hábitat preferido de
la paloma Zenaida macroura estudiaron su hábitat de forrajeo con un enfoque
estructural en Utah, EUA; empleando por una parte parcelas donde se manipuló la
altura de las hierbas, densidad, cobertura de estas y cobertura de hojarasca y por
otra parcelas como testigos en las que no se manipuló la cobertura. Los resultados
de este estudio registraron una mayor preferencia de esta especie por los lugares
manipulados, que en las áreas controles donde la estructura del hábitat se mantuvo
inalterado. Ellos concluyeron que las palomas basaban su elección de forrajeo según
la altura de las hierbas y su densidad.
Como se aprecia en la Tabla 22 la mayoría de los contactos realizados con la
especie al estudiar la relación entre la alimentación y la cobertura del suelo, fueron
en las áreas dedicadas a los cultivos agrícolas (57 %), seguidos de las áreas de
potreros (25 %). Como indican los diferentes índices calculados los valores más altos
se obtuvieron donde existía una combinación de cierto grado de cobertura vegetal y
exposición de suelo desnudo. Esto asegura una buena localización de las semillas
de especies herbáceas que constituyen el 79 % de la dieta de la especie, siendo un

129
rasgo característico de estas semillas su pequeño tamaño, por lo que sería muy
difícil de acceder a ellas con todo el suelo cubierto de vegetación para una especie.
Según Acosta y Ponce de León (2004) la especie toma las semillas directamente
del suelo, no escarba en la tierra, ni quiebra las espigas.
Es importante señalar que estas características vinculadas a condiciones
alimentarias que determinan la calidad de los diferentes biotopos para la especie
están sujetas a diferentes cambios en función de la época del año, condiciones
climáticas y el manejo del hombre en las zonas de cultivos agrícolas y ganaderos, al
respecto. Cuesta (2009) al referirse a las condiciones de las diferentes áreas de
alimentación expresa "… pero estas condiciones varían de acuerdo con la
intensidad del uso de la tierra, la época del año y otros factores. Algunas áreas
ganaderas se convierten en proveedoras de abundante alimento en ciertas épocas,
o bajo determinadas condiciones de uso que favorecen la presencia de plantas
alimenticias y el acceso a los granos".
8.4.- Conclusiones y recomendaciones
1. Las asociaciones vegetales presentes en los biotopos estudiados según la
disponibilidad y accesibilidad a estos por la especie, condicionan su presencia y
abundancia destacándose en el área de estudio un marcado uso del biotopo
Agrícola con cobertura.
2. La conducta con mayor incidencia resultó ser la alimentación. El uso del espacio
y el acceso a los recursos de esta paloma está condicionado por la cobertura del
suelo donde se alimenta, siendo de mayor frecuencia en las áreas de cultivos
agrícolas y en menor número en las áreas de bosques.
3. La presencia de linderos y árboles dispersos constituyen un factor importante en
la calidad de los biotopos agrícolas para la Paloma Rabiche.
4. La Paloma Rabiche prefiere para su alimentación áreas donde predominen las
especies herbáceas, pero en estas ha de existir un porciento de suelo desnudo
para poder acceder a las semillas que le sirven de alimento.

130
5. Las tierras dedicadas a los cultivos agrícolas han beneficiado el desarrollo de las
poblaciones de Paloma Rabiche más que sectores como la ganadería, áreas
forestales y frutales.
6. La utilización que hace la Paloma Rabiche de las diferentes asociaciones
vegetales está condicionada con la época del año, ya que esta determina la
rotación y cosechas tanto de plantas silvestres como cultivos agrícolas, y por
tanto la calidad para la especie.
7. Se recomienda un estudio detallado para poder identificar la dinámica que
inciden en la cobertura del suelo, herramienta indispensable para predecir y
manejar las poblaciones de la Paloma Rabiche en función de su calidad.
8.5.- Bibliografía
Acosta, M. y V. Berovides. (1982). Ecología trófica de las palomas del genero
Zenaida en el sur de Pinar del Río. Ciencias Biológicas, 7, 113-123.
Acosta, M; P. de león, J.L. (2004). Historia Natural de la Ploma Rabiche (Zenaida
macroura). (Inedito). Facultad de Biología. Universidad de la Habana. Cuba.
Camprodon, J; Subirachs, E. P. (2001). Conservación de la biodiversidad y gestión
forestal. Su aplicación en la fauna vertebrada. Ediciones de la Unibersitat de
Barcelona. España. 469 pp.
Chamizo, R., Escarré, A., Herrera, P., García, J., Cuesta, L., Martínez, L., Pino, …
Berovides, V. (2011). Caza y conservación en el ámbito rural en Cuba.
Publicaciones Universidad de Alicante. España. 13-20.
Cuesta, L. A. (2009). Perspectivas hacia una Gestión Sostenible de la Actividad
Cinegética en la Provincia de Pinar del Río, Cuba. Universidad de Alicante,
España, Universidad de Pinar del Río, Cuba.
Hayslette, S. E.; Mirarchi, R. E. (2002). Foraging-patch use and within-patch diet
selectivity in Mourning Doves, Zenaida macroura. Ecology, 83 (9), 2637-2641.

131
Ostrand, W. D. et al. (1998). Changes in Land Use as a Possible Factor in Mourning
Dove population Decline in Central Uthah. Journal of Field Ornithology, 69 (2),
192-200.
Ostrand, W. D., Bissonette, J. A. & Conover, M. R. (1996). Selection of foraging
habitat by Mourning Doves: a structural approach. Journal of Field Ornithology, 6
(4), 691-697.
Peiró, V. (2003). Gestión ecológica de los recursos cinegéticos. Publicaciones de la
Universidad de Alicante. España. 136 pp.
Peiró, V; E. Seva y M. Almiñana (1993). Selección de hábitat de una población de
Perdiz Roja (Alectoris rufa) en una zona de sierra del sur de la provincia de
Alicante. Mediterránea Serv. Biol., 14, 5-22.
Yarrow, G. (2009). Biología y manejo de la Paloma de Luto. Extensión de Recursos
Naturales y Forestales de la Universidad de Clemson. Carolina del Sur. USA.


132
CAPÍTULO IX. MORFOMETRÍA DE LA POBLACIÓN DE PALOMA RABICHE
(Zenaida macroura L.)
9.1.- Introducción
Desde época tan temprana como 1859 Darwin señaló que las diferencias
morfológicas entre las especies son indicadores de diferencias entre hábitat.
Selander (1971) citado por Rodríguez (2008) expone que los estudios morfométricos
en sus inicios fueron enfocados en la descripción y comparación de la morfología de
las aves, para entender principalmente la clasificación y las relaciones evolutivas que
se establecen entre las especies. Más tarde comienza a relacionarse la morfometría
como un indicador de los requerimientos ecológicos de las especies.
Dado que las condiciones ambientales y de hábitat de cada especie no son estables.
Estas condiciones varían no solo con las estaciones del año, sino también con
cambios provocados por el hombre y otros efectos estocásticos. Estos cambios
pueden generar transformaciones en los patrones de alimentación y con el tiempo
reflejarse en las características morfométricas de las especies.
En las aves el seguimiento en los cambios periódicos de los valores morfométricos,
ayudan a entender los estudios sobre los procesos de crecimiento, migración,
alimentación, influencia de las fluctuaciones ambientales, que provocan variabilidad
en la disponibilidad de los recursos, el desgaste de las plumas y las actividades
reproductivas (McMillen y Carpenter, 1976; Arendt y Faaborg, 1989; Rodríguez, 2001
en Rodríguez, 2008). El estudio de las diferencias morfométricas entre poblaciones
aporta elementos sobre los patrones de distribución espacial, calidad del hábitat y
límites de distribución de las especies, uno de los objetivos fundamentales de la
ecología (Carbonell, 1998).
De los estudios realizados por estos autores podemos inferir que la selección de
diferentes hábitats por las especies está condicionada por la morfología de estas,
por tanto el uso de los valores descritos en la literatura para la especie proveniente
de otras latitudes puede no reflejar las verdaderas características de la población
estudiada.

133
Otros aspectos importantes en el uso de la caracterización morfométrica en
poblaciones de aves es la diferenciación sexual de individuos de especies donde no
existe dimorfismo sexual o este es muy sutil. El estado o condición de salud de los
individuos, es otra condición en la que una caracterización morfométrica puede ser
muy útil, y así ahorrar tiempo y recursos en análisis mucho más complejos para la
determinación del sexo.
Es indiscutible por tanto, que el estudio ecológico de cualquier especie precisa de
una básica descripción morfológica, así como de la variabilidad que aparece en
estos caracteres y las proporciones e índices que relacionan las principales medidas;
estos aspectos son las herramientas principales para establecer comparaciones y
diagnosis descriptivas entre especies, poblaciones e individuos (Antelo, 2008 en
Ramos, 2014).
Los estudios biométricos en Cuba han sido más abundantes en las comunidades de
aves acuáticas, haciendo énfasis en las diferencias morfológicas entre las especies
que conviven en un mismo biotopo (Acosta et al., 2002; Mujica et al., 2003; Estévez,
2004). Las aves cinegéticas debido a la importancia que representan en el país no
han estado exentas de este tipo de estudio donde podemos citar los realizados en la
Paloma Torcaza Cabeciblanca (Columba leucocephala) (Chamizo et al., 1983;
Godinez, 1993), las palomas del género Zenaida (Acosta y Torres, 1984; Acosta y
Berovides, 1982 y Cuesta, 2009) y a la Gallina de Guinea (Numida meleagris)(
Torres et al., 1986).
Según Acosta et al. (2013) a cada ejemplar, es posible tomar un elevado número de
variables morfométricas, pero esto no siempre resulta útil, ya que por lo general un
grupo de ellas estarán altamente correlacionadas y, por tanto, con solo tener en
cuenta alguna de ellas podremos hallar los mismos resultados.
Por las razones antes expuestas es esencial caracterizar, sobre la base de los
caracteres morfométricos la población de Paloma Rabiche del área de estudio. Se
establece como objetivo de este capítulo la caracterización morfométrica de la
población a través de variables como el peso total del ave, longitud total, longitud del

134
ala plegada, de la cola y del tarso, largo y ancho del pico. Se realizará además la
comparación con datos obtenidos en otras investigaciones sobre la especie en el
país y con las medidas que se describen en la bibliografía de la misma. La
información recabada permitirá conocer la variabilidad de esta especie en relación
con las diferentes latitudes y las condiciones ambientales que pudiesen provocar
dichos cambios, siendo conocido qué algunas especies migratorias presentan
patrones de variaciones morfológicas a nivel geográfico.
9.2.- Materiales y métodos
Como fue descrito anteriormente en el Capítulo V “Conducta y Reproducción”, los
trabajos de campo consistieron en la toma de muestras de los ejemplares cazados
de esta paloma. La caza fue realizada al azar con escopeta de cartuchos en
recorridos a pie que abarcaron toda el área y cada ejemplar abatido fue etiquetado
siguiendo el orden de captura y clasificado según la clase de edad (adulto o pichón),
a cada ejemplar capturado se le tomó las medidas morfométricas. Se estudiaron un
total de 78 ejemplares.
Para garantizar el éxito del trabajo, se tuvieron en cuenta las recomendaciones
dadas por Acosta et al. (2013):
Antes de iniciar una investigación en la cual tengamos que capturar aves y tomar
sobre ellas un grupo de medidas, resulta importante delimitar cuales serán estas
medidas, en función del tipo de resultados que pretendamos obtener, y cómo las
vamos a tomar, de manera que, en sucesivos muestreos, se tomen de la misma
manera, y evitar con ello una fuente de error. Otro elemento a tener en cuenta es
que las medidas siempre deben ser tomadas por la misma persona, ya que el error
asociado al proceso de medición se incrementa, cuando las medidas son tomadas
por diferentes personas. Se ha probado, además, que períodos largos de trabajo
reducen la exactitud de las mediciones.
Para el caso de estructuras pareadas como las alas o los tarsos es importante
siempre elegir el mismo lado, ya que resulta frecuente para algunas especies cierta
variabilidad entre la derecha y la izquierda (Niethammer, 1962). En los casos en que

135
las alas tengan cierta diferencia en longitud, generalmente, la derecha será algo
mayor que la izquierda (Vaurie, 1959). Usualmente, han sido detectadas también
diferencias entre las medidas obtenidas por personas zurdas y derechas.
Resulta importante también tener en cuenta la precisión de los instrumentos con que
estamos tomando cada una de las medidas y no ir más allá del valor real que
podemos obtener en su lectura. Por ejemplo, si al medir la longitud del ala con una
regla graduada en milímetros, el extremo de la pluma cae entre los valores 128 y
129, debemos dar la medida como 128 mm y no como 128,5 mm, ya que esta última
se encuentra fuera de la precisión del instrumento utilizado… En el caso del peso,
resulta importante utilizar la balanza más cercana al peso del ave que estemos
procesando, ya que de eso dependerá la precisión del resultado.
Las medidas que se han tomado se pueden agrupar en dos categorías: descriptores
generales del tamaño (longitud total, longitud del pico y ancho del pico) y
descriptores de la morfología alar y caudal (longitud del ala plegada, longitud del
tarso y longitud de la cola). La masa se ha tomado con una precisión de ± 0,1 g, las
restantes medidas con una precisión de ± 0,1 mm.
A continuación se exponen las medidas morfométricas empleadas en nuestro trabajo
y su importancia según (Acosta et al., 2013).
-Peso total del ave (g): es una de las más simples que se pueden obtener, resulta de
gran utilidad para conocer las necesidades energéticas de la especie en cuestión. Se
utilizó una balanza con precisión de 0,1 g.
El peso total del ave en esta investigación fue realizado al momento de realizar la
captura por lo que incluye el contenido del buche.
Longitud total (cm): de importancia general, nos da una medida de la posición de
la especie en el ecosistema, de las cantidades de alimentos que necesita para su
mantenimiento y desarrollo, además, resulta muy útil, cuando queremos calcular
algunas medidas relativas destinadas a obtener información sobre la forma
general del ave. Se midió con una regla de tope graduada en mm.

136
Medidas del pico: revelan información clave sobre los tipos y tamaños de los
recursos que pueden ser a por cada especie, tomadas todas con pie de rey con
0,01 mm de precisión.
Longitud del pico (mm): se empleó la variante “culmen expuesto”, o sea la
distancia desde la punta del pico hasta en nacimiento de las primeras plumas en
la frente,
Ancho del pico (mm): tomado en la base del pico, al mismo nivel en que se midió
el “culmen expuesto”.
Longitud del ala plegada (cm): se usó la variante del ala plegada. Medida con una
regla de tope graduada en mm.
Longitud del tarso (mm): medida siempre muy ligada al movimiento que tenga el
ave en el medio en que desarrolla sus actividades vitales, tomadas con pie de
rey con 0,01 mm de precisión.
Longitud de la cola (cm): asociada con los movimientos de la especie durante el
vuelo, así como en el equilibrio a la hora de posarse. En algunas especies en la
elección de pareja, medidas con una con una regla de tope graduada en mm.
Dada la conveniencia de contar con algún indicador del estado de salud de la
población de la especie objeto de estudio, fueron obtenidos unos índices que tienen
en cuenta el tamaño del ave (longitud del cuerpo sin la cola) y su peso total.
Se trata del índice de masa corporal (IMC), apropiado para relacionar sobre el
estado de salud de la especie y calculado mediante la expresión:
IMC = P/Ta
Ta = Lt – Lc
donde:
P representa el peso total del ave.
Ta representa el tamaño total del ave .

137
Lt representa la longitud total del ave.
Lc representa el largo de la cola.
A los datos obtenidos se aplicaron primeramente técnicas de estadística descriptiva,
con la finalidad de resumirlos, describirlos numéricamente y facilitar su interpretación
(medidas de tendencia central y de variabilidad). Luego se les realizó un análisis
exploratorio de los datos a través de los test de normalidad Shapiro-Wilk W. y
Kolmogorov-Smirno, con el fin de conocer el comportamiento de la distribución y así
determinar el tipo de análisis estadístico a utilizar (paramétrico o no paramétrico).
Con el objetivo de comparar la media de la muestra obtenida con un valor de
referencia de la especie en la literatura se utilizó el test de “T de Student”. Al
realizar el análisis entre el peso y las diferentes categorías (sexo, clase edad y las
estaciones del año) para analizar el efecto que puede tener en una variable
aleatoria, una variable cuantitativa que actúe como factor se aplicó finalmente los
tests de la T de Student y el de U Man-Whitney según el comportamiento de
normalidad en los datos obtenidos (Martínez et al., 2009). En el procesamiento de la
información se emplearon los programas Excel y SPSS versión 15.0 para Windows.
9.3.- Resultados y discusión
9.3.1.- Caracterización morfométrica y relación con otras áreas
Del total de las 78 muestras obtenidas, solo en la variable longitud total no fue
empleada el total de la muestra cazada, ya que en algunos pocos casos y en
determinada época del año se capturaron ejemplares sin cola producto de la muda
del plumaje.

138
Tabla 23. Características morfométricas de la Paloma Rabiche en el área de estudio
(n= 78).
Parámetros
Promed
de la
especie
Clase de edad y sexo
Adulto Pichón
media macho hembra media macho hembra
Peso total del
ave (g) 112.20 114.8 116.3 112.2 100.6 104.11 91.6
máximo 137.1 137.1 137.1 120.3 118.4 118.4 94.5
mínimo 89.0 93.5 93.5 102.5 89.0 91.2 89.0
Longitud total
(cm) 27.6 27.9 28.2 27.4 26.2 26.6 25.2
máximo 30.0 30.02 30.0 28.5 29.0 29.0 25.5
mínimo 24.3 26.2 26.5 26.2 24.3 25.5 24.3
Largo del pico
(mm) 4.78 4.9 4.9 4.7 4.4 4.5 4.2
máximo 6.0 6.0 6.0 5.2 5.3 5.3 4.2
mínimo 4.0 4.0 4.2 4.0 4.0 4.0 4.0
Ancho del pico
(mm) 12.89 13.0 13.4 12.4 12.4 12.4 12.4
máximo 15.1 15.1 15.0 15.0 15.0 15.0 13.1
mínimo 7.3 10.4 11.4 7.3 7.3 7.3 11.7
Longitud ala
pleg. cm) 13.66 13.8 13.9 13.5 13.2 13.4 12.8
máximo 14.6 14.6 14.6 14.2 13.9 13.9 12.7
mínimo 12.1 12.1 13.0 12.1 12.5 13.0 12.5
Longitud tarso
(mm) 21.7 21.8 22.1 21.3 21.2 21.5 20.5
máximo 23.0 22.6 22.6 22.0 23.0 23.0 21.0
mínimo 20.0 20.0 20.7 20.0 20.0 20.1 20.0
Longitud cola
(cm) 11.74 12.1 12.3 11.6 10.2 10.4 9.7

139
máximo 9.0 13.3 13.3 12.8 13.0 13.0 10.5
mínimo 13.3 10.6 10.6 10.9 9.4 9.4 9.0
Una vez determinadas las características morfométricas de la población del área de
estudio, se comprobó la normalidad del peso total de las aves estudiadas y a través
del tests de la T de Studen utilizado para comparar una media con un valor de
referencia (Martínez et al., 2009) se analizó de manera independiente los datos del
área de estudio vs los establecidos para Cuba y los EUA.
Tabla 24. Comparación de la media de la población del área de estudio con dos
valores de referencia (α= 0,5).
Categorías
Media Significación (p)
Área estudio
(A)
Cuba
(B) EUA (C) A vs B A vs C
Peso total
especie 112,20 107,80 126,28
p=0,001 p=0,000
Machos 113,83 109,10 130,00 p=0,010 p=0,000
Hembras 109,28 105,80 123,00 p=0,045 p=0,000
Como se aprecia en la Tabla 24 al comparar el peso total de la población del área
de estudio, así como por sexos con los valores de referencia de los EUA, expuestos
por Tomlinson (1994) y los obtenidos en Cuba por Acosta y Torres (1984) en su
trabajo realizado en el Jardín Botánico de Cienfuegos, en todas las categorías (peso
total de la especie y por ambos sexos) presentan diferencias significativas entre el
área de estudio y Cuba (p= 0,001, p<0,05) y entre el área de estudio y EUA (p0,000,
p<0,05); aunque un análisis más detallado evidencia una menor diferencia entre el
área de estudio y los valores de Cuba que entre el área de estudio y los valores
descritos para los EUA en todas las categorías. Esto denota una diferencia menos
marcada entre las poblaciones del mismo país, no ocurriendo lo mismo al
compararla con las continentales.

140
Hasta el momento no hay suficientes publicaciones sobre la morfometría de la
Paloma Rabiche en diferentes regiones del país que permitan una amplia
comparación entre las diferentes poblaciones en Cuba. Sería necesario realizar
estudios ecomorfométricos de las poblaciones cubanas de esta especie para
dilucidar estas diferencias y sus posibles causas ecológicas.
Al realizar una evaluación del peso corporal, hay que tener en cuenta que esta
variable depende del estado nutricional del ave, que a su vez está en función de la
disponibilidad del alimento y la cantidad de alimento diario ingerido en el momento
de la captura; por lo tanto posee una variabilidad biológica intrínseca alta,
conociéndose que en otros columbiformes estas diferencias llegan ser superiores al
resto de las variables (Acosta y Torres, 1984; Godinez, 1993 en Rodríguez, 2008).
Una situación semejante es descrita para otra especie del género Columbiforme por
Rodríguez (2008) en su estudio sobre la Paloma Perdiz (Starnoenas cyanocephala)
al citar que Garrido, (2005) considera que las poblaciones de Paloma Perdiz de
Guantánamo, Santiago de Cuba y Pinar del Río; son similares en sus patrones de
medidas, mientras que las de Camagüey, Matanzas y provincia La Habana, son más
pequeños (refiriéndose a los datos morfométricos y al peso).
En cuanto a las diferencias obtenidas en relación al área de estudio y los EUA, es
conocido que las poblaciones de una misma especie que habitan en diferentes
regiones difieren entre sí por las condiciones ecológicas, tanto las históricas como
las actuales. La selección a las que han estado sometidas y las que operan en el
presente se puede traducir en una divergencia que puede verse reflejada en
diferencias morfológicas externas.
Hoyo et al. (1994) en Acosta y García (2005), sugieren en la Codorniz (Colinus
virginianus) una disminución del peso corporal para latitudes más bajas que los EUA,
al mirar los resultados antes expuestos pudiésemos estar ante una situación similar
en la Paloma Rabiche.
Se realizó un análisis estadístico en la población del área de estudio entre el peso
total del ave y el sexo, y la clase de edad. Del análisis entre el peso del ave y el

141
sexo se pudo obtener, al aplicar el test de U de Man-Whitney por no existir
normalidad en uno de los grupos (hembras), que la relación existente no presentó
diferencias significativas (p=0,245; P>0,05), pero como se aprecia en la Tabla 24
existe una ligera superioridad en todos los valores a favor de los machos, aunque
esta no sea significativa. Debido a la conducta reproductora de la especie, donde
existe una competencia por el acceso a una pareja entre los machos, es de
esperarse que esta selección sexual a través del tiempo conlleve a un aumento en
el tamaño en el sexo más competitivo.
Similares resultados encontraron Acosta y Torres (1984), donde los valores más
elevados del peso corporal correspondieron a los machos, aunque no se apreciaron
diferencias estadísticas significativas. Igualmente al estudiar esta especie en el sur
de Pinar del Río Acosta y Berovides (1982) encontraron un peso superior en los
machos. Rodríguez (2008) al analizar en su estudio las características
morfométricas de la Paloma Perdiz expresa que aunque no existe un dimorfismo
sexual evidente para el hombre en esta especie, dentro de las variables
morfométricas estudiadas hay una tendencia de los machos a tener mayor peso que
las hembras, quizás relacionado con el papel que desempeñan cada sexo dentro de
la reproducción, ya que pueden ser un indicador para las hembras, de las
habilidades competitivas de los machos
Un comportamiento similar es descrito por Di Mare (2003) para otra paloma del
género Zenaida en áreas de Tempisque, Costa Rica al encontrar valores
ligeramente inferiores en las hembras de la Paloma de Alas blancas (Zenaida
asiática), en tres variables morfométricas (peso total del ave, longitud del ala y el
largo total del cuerpo).
Estos resultados son consistentes con lo expuesto por Székely et al. (2004), ellos
encontraron que en un sistema de apareamiento con mayor competitividad entre
machos existe una alta correlación con el aumento en su tamaño (cuerda alar y
masa corporal) y viceversa con las hembras. Este comportamiento de aumento de
tamaño en respuesta a la competitividad intra-sexual, ha sido descrito en otras

142
especies de animales como ácaros, serpientes, tortugas, lagartos, colibríes y aves
canoras (Abouhief y Fairbairn 1997, Cox et al. 2003, Székely et al. 2000, 2004).
Generalmente las diferencias o semejanzas en cuanto al tamaño, peso y colorido
entre machos y hembras se relacionan con la estrategia reproductiva de la especie
(Maier, 2001). Independientemente de la existencia o no de dimorfismo sexual, las
hembras tienden a seleccionar a los machos por su mayor vistosidad, dado por el
mayor peso, tamaño y condiciones del plumaje; que sin dudas redundará en
beneficio del éxito reproductivo (Badii, 2004 en Rodríguez, 2008).
Por otro lado, Székely et al. (2000), consideran que la intensidad de la competencia
entre los machos adultos, puede provocar a través de mecanismos de selección
sexual un aumento de los valores en las medidas dentro de este sexo. En el caso de
la Paloma Perdiz, está descrito que al comienzo de la época reproductiva se
producen conflictos territoriales entre los machos (Sánchez et al, 1992). Es probable
que la tendencia de los machos a ser mayores que las hembras en la Paloma Perdiz,
pueda estar relacionado con esta conducta.
Al comparar el peso del ave con las clases de edad, utilizando el test de Student
(ambos grupos siguen una distribución normal), se obtuvo que existen diferencias
significativas (p=0,000; p<0,05), correspondiendo a los adultos los mayores valores.
Como se explicó en capítulos anteriores en Cuba están bien definidas solo dos
épocas (lluvias y sequía) las que además de otros factores condicionan la época de
máxima reproducción de la especie. Al realizar el análisis estadístico utilizando el test
de Student (ambos grupos siguen una distribución normal), se obtuvo que no
existen diferencias significativas (p=0,318; p>0,05) entre el peso de las aves y
ambas estaciones. Sin embargo al analizar los valores del comportamiento del peso
corporal en ambas épocas se aprecia una ligera superioridad en la época de sequía.
Este resultado pudiera estar relacionado con el periodo de máximo pico
reproductivo de la especie, que se desarrolla en la época de lluvias y las palomas
deben dedicar un tiempo considerable a la misma, con la consiguiente disminución
del tiempo para el forrajeo y la alimentación. Otro factor importante y que debería

143
tenerse en cuenta en próximos estudios de la especie es la influencia de la cobertura
del suelo en la alimentación de la especie en ambas épocas. En época de lluvias
existe una gran cantidad de alimentos para la especie, pero en época de sequía, el
alimento disponible en el suelo desnudo o parcialmente desnudo es más asequible a
la paloma que no lo toma de las plantas directamente.
Estos supuestos pueden ser apoyados por los resultados obtenidos en el capítulo 8
de esta investigación donde se analiza la influencia de la cobertura del suelo en la
especie, obteniéndose un mayor índice de selección en aquellos hábitats donde su
cobertura es parcial y con la investigación de Ostrand et al. (1996) anteriormente
referenciados en el capítulo 8.
9.3.2.- Parámetros morfométricos como indicadores del estado de salud de la
población
Para todo personal encargado del manejo de poblaciones silvestres, tanto en su
aprovechamiento como en su protección es de interés conocer el estado de salud de
dicha población. Al mismo tiempo, diagnosticar en cada etapa esta condición puede
convertirse en una herramienta fundamental para alcanzar el éxito en el manejo. Sin
embargo Núnez et al. (2011) alegan que la medición de esta variable presenta
dificultades debidas al gran número de técnicas utilizadas y a la controversia sobre la
interpretación de los resultados; La condición física es sin duda una característica
determinante a la hora de modular la conducta de un individuo y es por lo tanto una
variable a tener en cuenta en los estudios etológicos. Medir la condición física
puede también ser importante para evaluar los beneficios que aportan determinadas
conductas.
Ya acentuada la necesidad de conocer la salud tanto individual como poblacional en
el manejo de las especies, es importante tener en cuenta las dificultades para su
realización. En estos estudios lo más conveniente sería la aplicación de métodos que
no implicasen el sacrificio de los individuos examinados y que por tanto no incidirán
directamente en la densidad poblacional.

144
Sin embargo, es preciso mencionar que los análisis post mortem proporcionan la
información más completa sobre la composición de un organismo (e.g.
composición y cantidad de reservas nutricionales, estado de los tejidos, etc.) y
pueden ser importantes, en algunos casos, para validar otros índices de condición
físicas no invasivos (Brown, 1996).
Según Núnez et al. (2011) en general, una mayor masa corporal se interpreta como
índice de mejor condición física porque indica, presumiblemente, la
disponibilidad de reservas de nutrientes que permiten hacer frente a situaciones de
elevada demanda. Pero no siempre una menor masa corporal es indicadora de una
deficiente condición de salud, ya que pueden existir otros factores que incidan
directamente sobre el peso corporal.
Witter y Cuthill (1993) consideran que una reducción de la masa puede ser
adaptativa en condiciones de elevado riesgo de depredación porque mejora la
respuesta de huida de un individuo. Esta hipótesis es debatida, ya que la pérdida de
peso en condiciones de alta depredación puede ser una mera consecuencia de un
elevado nivel de estrés o de una reducción de la tasa de ingesta de alimento debido
a una mayor inversión en vigilancia.
Tabla 25. Índices de masa corporal de la población total de Paloma Rabiche, así
como la parasitada por las diferentes categorías (n= 78).
Parámetros
Categorías poblacionales
Población
total
Población
parasitada
Machos
parasitados
Hembras
parasitadas
Adultos
parasitados
Pichón
parasitados
Media 7,04 6,92 6,95 6,80 7,18 6,36
Al realizar el análisis entre el índice de masa corporal de la población y el peso de la
población de Paloma Rabiche (correlación de Pearson r= 0,761; p= 0,000) se obtuvo
que existe una relación buena y directa, por lo que al aumentar o disminuir la masa
corporal en las aves, este cambio se reflejará en el índice de masa corporal en ese

145
mismo sentido. Este resultado podría ser indicativo de situaciones que repercutan
en la salud de las aves y por tanto en su condición física.
Una vez comprobado la relación positiva y directa entre el índice de masa corporal y
el peso de las aves, se procedió a determinar si existen diferencias significativas
entre las diferentes medias de los índices de masa corporal de la población
parasitada por categorías y, el índice de masa corporal de la población según el
comportamiento de la normalidad de los datos en cada variable; ya que estas
diferencias pudiesen ser indicadoras de la salud poblacional.
Tabla 26. Comparación entre el IMC de la población y los diferentes índices de masa
corporal por categorías.
Pt vs Pp Pt vs Pap Pt vs Ppp Pt vs Pmp Pt vs Php
Test aplicado T Student U- ManW. T Student U-Man W. T Student
Valor de p 0,489 0,658 0,001 0,281 0,344
Significación
estadística
p > 0,05 No dif.
p > 0,05 No dif.
p < 0,05
exist. dif.
p > 0,05 No dif.
p > 0,05 No dif.
Pt: población total, Pp: población parasitada, Pap: población adulta parasitada, Ppp: población de pichones parasitados, Pmp: población de machos parasitados, Php: población de hembras parasitadas.
En la Tabla 26 se aprecia que solo una categoría presenta diferencias significativas
entre los índices comparados (pichón), el resto no mostró diferencias
significativas.
Estas diferencias encontradas en la clase de edad pudiesen estar dadas por las
propias variaciones de peso debidas a las diferencias de tamaño de entre ambas
clase, pero también pudiese estar influyendo la respuesta de los jóvenes ante las
patologías asociadas al parasitismo. En la población estudiada encontramos una
tendencia en las aves parasitadas de la intensidad de infección se incrementa
con la disminución de la edad y/o el tamaño del hospedador. Esta es una
característica que quizás pueda explicarse debido a que una vez que los
parásitos ingresan al hospedador existen filtros (ecológicos y/o evolutivos)

146
que determinan cuál o cuáles podrán instalarse, desarrollarse y prosperar
en el hospedador, y por tanto los efectos no serán iguales en ambas clases.
En el resto de las demás categorías no se observaron diferencias estadísticamente
significativas entre los índices de masa muscular de la población y los índices de
masa muscular de las aves parasitadas (T de Student p > 0,05; U de Man-Whitney p
> 0,05). A pesar de que no se observaron diferencias estadísticamente significativas
entre las categorías, sí que se puede establecer una tendencia negativa en el peso
de las aves parasitadas. Un análisis de varios años y con una muestra más grande
podría revelar un comportamiento más claro de las variables evaluadas, ya que, se
cree que hubiera sido posible demostrar estadísticamente el efecto negativo del
parasitismo sobre la condición corporal, si hubiera sido posible realizar un estudio
más detallado y de duración mayor, apoyado en un análisis más exhaustivo de
laboratorio.
Conocer tales relaciones resulta de utilidad para predecir potenciales cambios en
poblaciones bajo manejo, amenazadas o que habitan sistemas perturbados. En este
sentido, las relaciones peso-salud son empleadas con frecuencia para modelar las
tendencias de la biomasa basados en la talla y son ampliamente usados en el
manejo de poblaciones de importancia económica.
Otros estudios, sin embargo, prueban que la pérdida de peso puede ser una
respuesta adaptativa eficaz en determinadas condiciones. Por estas razones hay
que tener siempre presente los objetivos del trabajo que se está realizando y las
especies con las que se trabaja ya que estos pueden variar entre diferentes especies
(Núnez et al., 2011).
Frente a todas estas dificultades, es recomendable mantener cierta cautela en la
interpretación de los resultados y, siempre que sea posible, combinar diferentes
técnicas y averiguar la repetibilidad de los índices utilizados para obtener un mejor
contraste de las hipótesis que estamos investigando.

147
9.4.- Conclusiones y recomendaciones
1. Existen diferencias estadísticamente significativas entre el peso total de las aves
de la población estudiada y las descritas en la bibliografía para Cuba y los EUA,
estas diferencias son menos marcadas respecto a los valores descritos para
Cuba que para las continentales.
2. No existen diferencia estadísticamente significativas entre el peso corporal entre
los sexos, no obstante existe una pequeña diferencia a favor de los machos.
3. A pesar de no existir diferencias estadísticamente significativas entre el peso de
las aves y las estaciones del año, se aprecia una ligera superioridad a favor de la
época de sequía.
4. Se apreció un menor peso en las aves parasitadas, si bien no hubo relación
estadísticamente significativa.
5. Se recomienda realizar un estudio donde sea posible determinar la validación del
estado general de salud de la Paloma Rabiche a través del índice de masa
corporal, pues este constituiría un importante indicador de la condición de salud
de las poblaciones de la especie y es relacionable con la calidad de los territorios
de asentamiento. Resulta necesario combinar diferentes técnicas para obtener un
resultado de un mejor contraste de la hipótesis que estamos investigando.
9.5.- Bibliografía
Abouhief, E. y D.J. Fairbairn (1997). A comparative analysis of allometry for sexual
size dimorphism: Assessing Rensch's rule. The American Naturalist, 149 (3),
540-562.
Acosta, M., Mugica, L. y Aguilar, S. (2013). Protocolo para el monitoreo de aves
acuáticas y marinas. Centro Nacional de Áreas Protegidas. La Habana. Cuba.
142 pp.
Acosta, M.; A. C. López y L. Mugica (2002). Influencia del peso corporal y la longitud
del tarso en las relaciones ecológicas de las comunidades de aves acuáticas del
agroecosistema arrocero. Biología, 16 (2), 109-122.

148
Acosta, M. (1998). Segregación del nicho en la comunidad de aves acuáticas del
agroecosistema arrocero en Cuba. Tesis presentada en opción al grado de
Doctor en Ciencias Biológicas. Universidad de la Habana, Cuba.
Acosta, M; Berovides, V. (1982). Ecología trófica de las palomas del género Zenaida
en el sur de Pinar del Río, Cuba. Ciencias Biológicas, 7, 113-123.
Acosta, M. y García, L. (2005). Morfometría y alimentación de la codorniz (Colinus
virginianus) en dos áreas del occidente de Cuba. Journal of Caribbean
Ornithology, 18, 54-68.
Acosta, M. y O. Torres. (1984). Ecología trófica de las palomas del género Zenaida
en el Jardín Botánico de Cienfuegos, Cuba. Ciencias Biológicas, 11, 107-116.
Brown, M. E. 1996. Assessing body condition in birds. Current Ornithology, 13, 67–
135.
Carbonell, R. (1998). Variaciones Morfométricas de cinco poblaciones de Currucas
Capirotadas (Silvia atricapilla) Ibéricas. Journal of Avian Biology, 29.
Chamizo, R., R. Rodríguez, C. Maros, y E. Godinez. (1983). Aspectos de la
reproducción de la Torcaza Cabesiblanca (Columba leucocephala) en cayos “Los
Indios”, Isla de la juventud. Revista Forestal Baracoa, 65-81.
Cox, R.M., S.L., Skelly, y H.B. John-Alder. (2003). A comparative test of adaptive
hypotheses For sexual size dimorphism in lizards. Evolution, 57 (7), 1653-1669.
Cuesta, L. A. (2009). Perspectivas hacia una Gestión Sostenible de la Actividad
Cinegética en la Provincia de Pinar del Río, Cuba. Universidad de Alicante,
España, Universidad de Pinar del Río, Cuba.
Darwin, C. (1859). The origin of species by means of natural selection. London: In:
Murray (1897): Voyage of a Naturalist. London: Murray.)
Di Mare, M. I. (2003). Densidad poblacional y uso sostenible de la Paloma de alas
blanca (Zenaida asiática) en áreas costeras de Bagaces a Abangares en el área
de conservación Tempisque, Costa Rica. Informe al área de conservación de
Tempisque. http://www.inbio.ac.cr/es/estudios/pdf/informe-PalomaAlaBlanca.pdf

149
Estévez, Y.(2004). Estudio preliminar de patrones osteológicos en nueve especies
de garzas (Aves: Ardeidae). Tesis de Diploma. Facultad de Biología, Universidad
de la Habana, Cuba.
Godinez, E. (1993). Situación de las poblaciones de Columba leucocephala (Aves:
Columbidae) en Cuba entre 1979 y 1987. Editorial Academia. 78 pp.
Maier, R. (2001). Comportamiento animal. Un enfoque evolutivo y ecológico. Mc
Graw Hill-Interamericana de España, S.A.U., Madrid, España. 581pp.
Martínez, M., Seguí, M. y López, C. (2009). Bioestadística amigable. Ediciones Díaz
de Santos. España. 293-342.
Mujica, L.; M. Acosta y D.D. Avila (2003). Variaciones espacio temporales y uso del
hábitat por la comunidad de aves en la arrocera sur del Jíbaro, Sancti Spiritus,
Cuba. Biología, 17 (2): 105-119.
Nuñes, C., Canestrari, D., Baglione, V. (2011). Evaluación de la condición física en
las aves y su aplicación a los estudios etológicos. Etologuia, 23, 13-42.
Ostrand, W. D., Bissonette, J. A., Conover, M. R. (1996). Selection of foraging hábitat
by mourning doves: a structural approach. Journal Field Ornithology, 67 (4), 691-
697.
Ramos, R. (2014). Protocolo para el monitoreo del cocodrilo cubano (Crocodylus
rhombifer). Centro nacional de Áreas Protegidas. La Habana. Cuba. 25 pp.
Rodríguez, Carrillo, Yamilet (2008). Morfometría y conducta en cautiverio de la
Paloma Perdiz (Columbiforme: Starnoenas cyanocephala, Linnnaeus, 1758).
Tesis en opción al grado de Master en Zoología y Ecología Animal, mención:
Vertebrados. Zoológico Nacional. Ciudad de la Habana. Cuba.
Sánchez, B.; D. Rodríguez y M. Acosta (1992). Algunos aspectos de la reproducción
de la Paloma Perdiz (Starnoenas cyanocephala) en cautiverio. Serv. Zool, 15, 1-
8.

150
Székely, T, R.P. Freckleton y J.D. Reynolds. (2004). Sexual selection explains
Rensch's rule of size dimorphism in shorebirds. Proceedings of the National
Academy of Sciences of the United States of America, 101 (33), 12224-12227.
Székely, T, J.D. Reynolds y J. Figueroa. (2000). Sexual size dimorphism in
shorebirds, gulls and alcids: the influence of sexual and natural selection.
Evolution. 54(4), 1404-1413.
Tomlinson, R. et al. (1994). Mournig Dove. Migratory shore an upland game bird
management in North. America. International Association of Fish and Wildlife
agencies. Allen Press, Lawrence, Kansas. E.U.A. 5-26.


151
CAPÍTULO X. ECTOPARÁSITOS EN LA POBLACIÓN DE PALOMA RABICHE
(Zenaida macroura L.)
10.1.- Introducción
Uno de los principales aspectos que deben considerarse en la gestión de los
recursos cinegéticos es el conocimiento del estado actual de sanidad y vitalidad de
los mismos, así como los factores que inciden en la dinámica poblacional de las
especies. Según Peiró, (2003) el análisis biosanitario de las poblaciones cinegéticas
ha de ser considerado como una parte de su aprovechamiento sostenible.
Núñez et al. (2011) expone que la evaluación de la carga parasitaria individual
debería proporcionar información relevante acerca del estado de salud en las aves.
Sin embargo, el estudio del parasitismo en aves silvestres todavía tiene algunas
limitaciones importantes. En primer lugar, el conocimiento taxonómico de los
parásitos aviares es todavía muy incompleto Clayton & Moore (1997)
consideran que la mayoría de ellos todavía está por descubrir.
Debido a que Zenaida macroura es una especie no solo con valor ecológico, sino
también cinegético y por tanto económico y recreativo, es importante conocer su
estado sanitario, así como los parásitos que intervienen en este.
A las enfermedades de las especies de vida silvestre no se ha dado suficiente
importancia ecológica, sino hasta hace poco (Simonette (1995) en Di Mare (2003) y
sobre todo con especies en vías de extinción (Cooper, 1989 en Di Mare 2003).
De acuerdo con Schmidt y Roberts (1984) citados por Pazmiño (2007) la avifauna
silvestre generalmente se encuentra infectada por varias especies de parásitos. En
raras ocasiones sufren muertes masivas o epizootias, debido a la dispersión y
territorialismo que la mayor parte de las especies presentan. Se considera que la
mayoría de las aves silvestres toleran su carga de parásitos adecuadamente, sin
embargo, estos animales mueren cuando la infección es alta, a causa de los
diversos estragos causados por estos.

152
Existen tres grupos de parásitos que pueden afectar las palomas: los artrópodos
(ácaros, garrapatas y piojos), protozoarios (parásitos unicelulares que viven dentro
de los hospederos) y helmintos (gusanos redondos). Dentro de estos grupos, se
encuentran representantes con diversos requerimientos microambientales que les
permiten invadir áreas externas del cuerpo como plumas, pecho, ano; así como
áreas internas a lo largo del tracto digestivo y el sistema sanguíneo (Di Mare, 2003).
El parasitismo, es uno de los factores que regulan las poblaciones en muchas
especies. Cada parásito suele habitar sobre un órgano determinado del huésped.
Los ectoparásitos son aquellos que viven en la piel, pelo o pluma de sus huéspedes
(Storer y Usinger, 1972).
Los ectoparásitos son importantes en la adaptabilidad de las aves, aunque la
presencia de algunos pocos de ellos pueden tener poco efecto en las aves, una gran
infestación o carga parasitaria puede deteriorar la salud de un ave, así como la
reproducción de ésta. Las aves son anfitrionas de muchas clases de parásitos
“chupadores de sangre”, especialmente las moscas del plumaje (familia
Hippoboscidae), las pulgas, piojos, ácaros y garrapatas (Podulka et al., 2004).
En el presente estudio solo se tuvo en cuenta la presencia de ectoparásitos debido a
que su localización y clasificación no supondría del empleo de medios
especializados o el envío de las muestras a un laboratorio con ese fin.
Los estudios existentes sobre las parasitosis en la Paloma Rabiche son exiguos y
fragmentados en diferentes regiones, destacándose los realizados en los EUA. A
partir de las fuentes consultadas se encontró que existe escasa información acerca
de los ectoparásitos que afectan las poblaciones y la condición corporal de esta
especie en nuestro país. El objetivo de este capítulo es determinar los ectoparásitos
presentes en la población de Paloma Rabiche estudiada, así como la incidencia
según las clases de edad, sexo de la Paloma Rabiche y época del año.

153
10.2.- Materiales y métodos
Como fue descrito anteriormente en el Capítulo V. “Conducta y Reproducción”, los
trabajos de campo consistieron en la toma de muestras de los ejemplares cazados
de esta paloma. La caza fue realizada al azar con escopeta de cartuchos en
recorridos a pie que abarcaron toda el área y cada ejemplar abatido fue etiquetado
siguiendo el orden de captura y clasificado según la clase de edad (adulto o pichón)
y de sexo, a cada ejemplar capturado inmediatamente fue etiquetada y colocada en
una bolsa plástica individual sellada, (con el cuidado de cerrar inmediatamente la
bolsa para evitar escape de ectoparásitos), luego se procedió a revisar
minuciosamente las plumas, alas, pecho, dorso, ano, patas, pico y cabeza, para
detectar parásitos externos. Con una pinza fina se colectaron los parásitos y
fueron colocados en un frasco con alcohol al 70 % hasta logrado su identificación.
Se estudiaron un total de 76 ejemplares. Según lo expuesto por Brown (1996) los
análisis post mortem proporcionan la información más completa sobre la
composición de un organismo (e.g. composición y cantidad de reservas
nutricionales, estado de los tejidos, etc.) y pueden ser importantes, en algunos
casos, para validar otros índices de condición físicas no invasivos.
Con estos resultados se procedió a valorar la prevalencia de estos en la población
estudiada mediante el índice de prevalencia.
El índice de prevalencia (Ip) permite conocer cuántas aves de un grupo definido
están enfermas en un determinado momento, por lo tanto nos indica el peso o la
abundancia del evento que soporta la población. Margolis et al. (1982) y revisadas
por Bush et al. (1997). Fue calculado por la siguiente fórmula:
Ip = ni / N x 100
donde:
ni = representa el número de palomas infestadas por la especie i de parásito
N = representa el número total de palomas analizadas

154
A los datos obtenidos se aplicaron primeramente técnicas de estadística descriptiva,
con la finalidad de resumirlos y facilitar su interpretación (medidas de tendencia
central, cálculo de porcentajes por clases de edad y sexos). Luego se les realizó un
análisis exploratorio de los datos a través de los test de normalidad Shapiro-Wilk W.
y Kolmogorov-Smirno, con el fin de conocer el comportamiento de la distribución y
así determinar el tipo de análisis estadístico a utilizar (paramétrico o no paramétrico).
Al realizar el análisis entre el peso total de los ejemplares parasitados y los no
parasitados en las diferentes categorías, se aplicó el test de la T de Student o el de
U Man-Whitney según el comportamiento de normalidad en los datos obtenidos. Con
el objetivo de comparar la media de la muestra obtenida con un valor de referencia
de la especie en la literatura se utilizó el test de “T de Student” (Martínez et al.,
2009). En el procesamiento de la información se emplearon los programas Excel y
SPSS versión 15.0 para Windows.
10.3.- Resultados y discusión
En el total de muestras colectadas, solo se encontraron dos ectoparásitos, la “mosca
de gavilán” (Lynchia americana) y un ácaro (aún sin clasificar) que se encontró en los
tarsos de la mayoría de los ejemplares capturados. De las 78 aves estudiadas, 26
(34 %) presentó alguna clase de ectoparásito, el grupo predominante fue el de los
ácaros podales, los cuales constituyeron el 22 % de las aves parasitadas, seguidos
por la “mosca de gavilán” con el 8 % de la población parasitada.
Dentro de las llamadas “moscas planas” existen varias tipos de moscas que también
son conocidas como “moscas del plumaje”. Las moscas del plumaje son un grupo de
ectoparásitos de la familia Hippoboscidae que se alimentan de la sangre de las
aves, son parecidas a las moscas domésticas, tienen el cuerpo sumamente
aplastado, lo que le permite moverse con holgura entre el contorno de las plumas,
son conocidas también junto a otros ectoparásitos como los mosquitos, piojos y
pulgas por ser trasmisores de enfermedades como la viruela.

155
Figura 24. Comportamiento del Índice de prevalencia de Lynchia americana en la
población de Paloma Rabiche estudiada a través del año (n= 78).
Como se aprecia en la Figura 24 la “mosca de gavilán”, solo se encontró en los
meses de marzo, mayo, junio y agosto, correspondiendo al mes de junio el mayor
Índice de prevalencia (Ip) (37,5 %), coincidiendo con la época de máxima
reproducción de la especie y el mayor número de pichones en la población. El Índice
de prevalencia en adultos fue de un 4,7 % y en los pichones de un 21,4 %. Estos
meses corresponden a la época de lluvias y al analizar el comportamiento del Índice
de prevalencia en ambas épocas se aprecia una notable diferencia (Figura 25).

156
Figura 25. Comportamiento del Índice de prevalencia de Lynchia americana en dos
épocas en el área de estudio (n= 78).
Similares resultados expone Di Mare (2003) en su estudio sobre Zenaida asiática
(paloma de alas blanca) en Tempisque, Costa Rica, donde reporta dentro de los
ectoparásitos encontrados una mosca hematófaga en el plumaje de las palomas
analizadas (Pseudolynchia canariensis); aunque su incidencia en la muestra
estudiada fue muy baja (1 % de índice de prevalencia).En cuanto al análisis de la
prevalencia mensual de los ectoparásitos en Tempisque, Costa Rica, los meses de
mayor afectación en la población correspondieron a los de la época de lluvias
(marzo, julio, agosto y octubre). Leherman (1993) en Di Mare (2003), indica que
una gran incidencia de ectoparásitos en la época de reproducción puede provocar el
abandono de nidos activos en respuesta a una alta densidad de ectoparásitos.
Finalmente en los resultados de este estudio pueden encontrarse similitudes con
los ya obtenidos por Herman (1937), quien encontró en los EUA, que las moscas
de la familia Hippoboscidae son más abundantes durante finales de julio y el mes de
agosto, sugiriendo que las Hippoboscidaes crecen prolíferamente mejor en una
atmósfera seca. A pesar de existir coincidencia en los meses del año con la
tendencia observada en este estudio, en nuestro país estos meses no resultan ser

157
los más secos del año, todo lo contrario, es donde se registran con más frecuencia
lluvias, pero un factor común en ambos países resultan ser las altas temperaturas de
esta época del año que pueden determinar un aumento en la proliferación del
ectoparásito estudiado.
En la literatura revisada hasta el momento en Cuba no se han desarrollado estudios
de forma sistemática sobre el índice de prevalencia de este ectoparásito en la
poblaciones de Paloma Rabiche, ni de ninguna otro especie de paloma del país, por
lo que no se cuenta con parámetros que permitan identificar qué resultado de índice
de prevalencia en un población puede acarrear problemas de salud en está.
Al analizar la relación de parasitismo por esta mosca entre ambos sexos en la
población estudiada se comprobó la no existencia de grandes diferencias en los
resultados, siendo más bien bajos estos índices de prevalencia en ambos sexos,
aunque existió una ligera superioridad de los valores obtenidos en los machos con
respecto a las hembras (Figura 26). Estos resultados registrados coinciden con los
obtenidos por González et al. (2004a) en el estudio de Parásitos gastrointestinales y
externos de la paloma domestica (Columba livia) en la ciudad de Chillan, Chile,
donde comprobó que no hubo diferencias significativas entre la infestación por
ectoparásitos entre los sexos, lo que según autores podría estar relacionado con el
escaso dimorfismo sexual entre palomas, además de presentar hábitos similares en
individuos de ambos sexos. No ocurrió igual en estudio realizado en Ñuble, Chile por
González et al. (2004b) al estudiar la fauna parasitaria de la tórtola común (Zenaida
auriculata, de Murs 1847); expone que en la relación hembra-macho se pudo
observar una predominancia de las hembras con respecto a los machos, siendo esta
más notoria en los ectoparásitos menos frecuentes (Hohors-tiella sp. y Bonomiella
sp).

158
Figura 26. Comportamiento del Índice de prevalencia de Lynchia americana entre
sexos en la población de paloma Rabiche estudiada (n= 78).
En la Figura 27 se puede observar el comportamiento del Índice de prevalencia en
ambas clases de edad en la población de Paloma Rabiche estudiada, destacándose
la gran incidencia de este ectoparásito en las palomas jóvenes. Leherman (1993) en
Di Mare (2003) plantea que los ectoparásitos chupadores como las moscas del
plumaje, al chupar sangre producen heridas muy dolorosas, especialmente en los
pichones jóvenes. Es por tanto esta incidencia, un factor clave para el éxito
reproductivo de la especie. Al respecto Núñez et al. (2011) expuso que en las aves,
la condición física influye notablemente sobre el comportamiento petitorio de los
pollos en el nido.

159
Figura 27. Comportamiento de Índice de prevalencia de Lynchia americana en las
clase de edad en la población de paloma Rabiche estudiada (n= 78).
En la presente investigación es importante destacar que en las aves parasitadas por
Lynchia americana no se detectaron cambios patológicos notorios al momento de
las necropsias en las muestras estudiadas, ni al realizar la prueba de T de Student
para comparar una media con un valor de referencia, escogido en este caso los
valores medios del peso corporal de las aves parasitadas vs peso promedio de la
población del área de estudio (Tabla 21). Como se explicó en el capítulo dedicado al
estudio morfométrico de la especie el peso total de un ave a pesar de ser una de las
medidas más simples, es de gran importancia ya que además de ser de gran utilidad
para conocer las necesidades energéticas de la especie, esta medida es un buen
indicador de las posibilidades de supervivencia de la especie por ser un indicador de
las reservas necesaria para procesos vitales como la migración, reproducción y
muda Acosta et al. (2013). Por tanto valores por debajo de los medios o normales
para una especie deben ser tomados como anomalía en algún aspecto de la calidad
de hábitat o la salud de los individuos.

160
Tabla 27. Resultados obtenidos al realizar la prueba de T de Student para comparar
una media con un valor de referencia entre el peso promedio de la población de
Paloma Rabiche en el área de estudio vs el peso promedio de las aves parasitadas
por Lynchia americana.
Estadísticos Peso total medio
de la población
Peso total medio
de las aves parasitadas
Media 112.20 106.07
Valor de p 0.297
Significación
estadística P > 0.05, No existen diferencias significativas
A pesar de no existir diferencias estadísticas significativas se puede observar
superioridad en el peso promedio de las aves no infestadas por Lynchia americana,
lo que está dado por la posible depauperación progresiva que pueden sufrir las aves
infestadas debido a las enfermedades que pueden contraer producto de las picadas
de estas moscas o a la pérdida contante de sangre por la alimentación de los
ectoparásitos.
Entre los artrópodos parásitos más comunes en las aves que pueden afectar la
condición física individual están los malófagos, que se alimentan normalmente de las
plumas, la piel o los productos de la piel. Los ectoparásitos más frecuentes, sin
embargo, son los ácaros (Núñez et al., 2011).
La Figura 28. nos permite apreciar el comportamiento del índice de prevalencia en
los ejemplares de paloma afectados con ácaros podales en relación con los meses
del año, Así, este ectoparásito se encuentra prácticamente en ambas épocas del
año; siendo abril y agosto los meses de mayor prevalencia, los que corresponden
con la época lluviosa. Los ácaros podales hallados no mostraron un patrón
estacional definido. Si bien se observaron fluctuaciones en los índices de

161
prevalencia a lo largo del año, podría denominarse como una especie común
en la comunidad.
Figura 28. Comportamiento del Índice de prevalencia de ácaros podales en la
población de Paloma Rabiche estudiada a través del año (n= 78).
Los resultados expuestos anteriormente son confirmados con la relación obtenida al
calcular el índice de prevalencia en las dos épocas bien marcadas de nuestro país
(sequía y lluvias). Resulta evidente la poca diferencia entre los valores obtenidos,
aunque se considera oportuno mencionar que dentro de la población parasitada las
aves con mayor infestación fueron en los meses de marzo, septiembre y octubre;
coincidiendo estos meses de alto índice con la época de lluvia, caracterizada por
altas temperaturas y abundantes precipitaciones.

162
Figura 29. Comportamiento del Índice de prevalencia de ácaros podales en dos
épocas en el área de estudio (n= 78).
Al analizar el Índice de prevalencia de los ácaros podales entre sexos en la
población parasitada de Paloma Rabiche estudiada a través del año, se aprecia que
ninguno superó el 50 % o más, lo que constituiría una clara preferencia sobre algún
sexo. No obstante, aunque no se demostró diferencias estadísticamente
significativas entre sexos, se registró una ligera diferencia a favor de los machos
(Figura 30). Vale destacar que los más altos grados de infestación por este ácaro en
las aves parasitadas correspondieron a ejemplares machos.

163
Figura 30. Comportamiento del Índice de prevalencia de los ácaros podales entre
sexos en la población parasitada de paloma Rabiche estudiada (n= 78).
En la figura 31 se puede observar los resultados obtenidos al calcular el índice de
prevalencia del ataque de este ácaro en relación con las clases de edad,
evidenciándose una mayor incidencia en la población joven. Estos resultados son
similares a los obtenidos en el análisis realizado de los ejemplares afectados por
Lynchia americana, donde también los índices de infestación por este ectoparásito
fueron superiores en la población joven. Constituye pues, una evidencia de la
influencia de esta clase de edad (jóvenes) sobre la estructura de las comunidades
parásitas en esta Columbidae.

164
Figura 31. Comportamiento del índice de prevalencia (Ip) de ácaros podales según la
clase de edad en la población de Paloma Rabiche (n = 78).
La diferencia observada entre ambas clases de edad, y coincidente con el
comportamiento de Lynchia americana en la población estudiada, podría explicarse
por el poco tiempo de vida que tienen los pichones, que pudiese conducir a una
actitud diferente ante el ataque de los parásitos, con una respuesta más débil ante
agentes patógenos.
Según Núñez et al., (2011) el sistema inmune de las aves consta de un componente
innato, y uno específico (o adquirido). El sistema innato responde de forma
inmediata pero no específica a muchos agentes patógenos, representa una
primera línea de defensa que no confiere inmunidad a largo plazo. La inmunidad
adquirida, sin embargo, es capaz de producir una “memoria inmunológica” que
proporciona una defensa mejorada en sucesivos encuentros con ese mismo
patógeno. El sistema inmunitario innato está menos sujeto a variaciones
ambientales, nutricionales o a los niveles de estrés que el sistema adquirido, y por
lo tanto es un buen indicador de inmunocompetencia. Además, refleja la acción de
los genes ligados a la producción de anticuerpos naturales, que están sujetos a
selección natural.

165
Como ya se explicó anteriormente en todas las aves examinadas se tuvo en cuenta
la presencia de cambios patológicos notorios al momento de su examen, tanto en el
físico como en las necropsias. En ninguno de los casos de ejemplares parasitados
por ácaros podales fueron detectadas algún tipo de anomalías. Para conocer la
relación entre el peso de las aves parasitadas por los ácaros podales y la media
general de la población (Tabla 28) se realizó el test de T de Student (comparación
de la media de una variable con distribución normal con un valor de referencia)
Tabla 28. Resultados obtenidos al realizar la prueba de T de Student para comparar
una media con un valor de referencia entre el peso promedio de la población de
Paloma Rabiche en el área de estudio vs el peso promedio de las aves parasitadas
por ácaro podal.
Estadísticos Peso total medio
de la población
Peso total medio
de las aves parasitadas
Media 112.20 111.40
Valor de p 0.694
Significación
estadística P > 0.05, No existen diferencias significativas
El resultado de esta prueba es similar al obtenido en el análisis de la “mosca de
gavilán”. La media de la población es superior a la de la población infestada y
aunque no existan significación estadística se aprecia una ligera superioridad en el
peso de la población sana, aspecto este a tener en cuenta. Según Núñez et al.
(2011) el estado físico de un animal influye sobre su conducta y por lo tanto ha de
considerarse en los estudios etológicos. Medir la condición física en una población
puede también ser importante para evaluar los beneficios que aportan determinadas
variables morfométricas y ayudar con ello a comprender su función adaptativa.
Una vez realizados los análisis de cada ectoparásito encontrado en la población de
Paloma Rabiche es indispensable contar con un análisis general de la incidencia de

166
ambos en la población, de conjunto ambos determinan el estado general de la salud
poblacional.
Es importante señalar que actualmente se desconoce hasta qué punto los parásitos
estudiados afectan o limitan la población de la especie, así como los trastornos
fisiológicos que provocan, por lo que se considera un tema a continuar investigando
en el futuro.
Al comparar la incidencia de los dos ectoparásitos hallados en la población de
Zenaida macroura se aprecia que la especie dominante resulto ser los ácaros
podales, con valores del índice de prevalencia más constantes en las
diferentes épocas del año. A pesar de las variaciones observadas en este ácaro
la especie mantuvo la condición de común en todos los periodos
analizados. Pocos cambios en la composición estacional pudiesen indicar una
gran estabilidad tanto en el tiempo como en diversas características del
hospedador.
Ningún de las dos especies de ectoparásitos hallados superó el 55 % o más de
índice de prevalencia, lo que supondría una gran afectación en la población de
Paloma Rabiche. No obstante los ácaros podales fue la especie con los
mayores valores de índice prevalencia (Ip= 50 %) en la población, mientras que
Lynchia americana mostró como máximo valor el 37,5 5 de índice de prevalencia.
De las dos especies de ectoparásitos hallados Lynchia americana mostró una
tendencia a encontrarse en pocos hospedadores con intensidades de 1 ó 2
parásitos por hospedador, excepto en un solo ejemplar. Este resultado indicaría
que estos ectoparásitos pudiesen clasificarse como estacionales. Así
pudiésemos estar ante un grupo de ectoparásitos que confieren estabilidad al
sistema.
Estos resultados son coincidentes con los obtenidos por Dimare (2003) en su
estudio sobre Zenaida asiática en Tampisque, Costa Rica, donde el índice de
prevalencia del ácaro Falculifer spp fue del 18 % y solo el 1 % de la mosca
hematófaga Pseudolynchia canariensis.

167
En las fluctuaciones observadas en los índices de prevalencias de los
ectoparásitos entre estaciones del año se observó una tendencia de Lynchia
americana a variar de manera inversa al aumento de la época seca, esta
diferencia coincide con las mayores abundancias de este ectoparásito observadas
en un ejemplar. Contrario a la conducta seguida por los ácaros podales que se
mantuvieron presentes en la mayoría de los meses del año.
Estas conductas pueden ser el reflejo de los patrones de colonización
estacionales de los parásitos, de los cambios en la dieta de los
hospedadores entre las dos épocas y/o de cambios en las condiciones
climáticas. Las condiciones climáticas cambiantes y la distinta disposición de
recursos en las diferentes épocas pueden producir efectos muy importantes sobre
las poblaciones parásitas. La alta incidencia de un parásito en una estación
determinada le confiere una alta predictibilidad en esta época y por tanto una labor
preventiva consecuente con la estación anual.
Dado que no se observaron grandes diferencias en los índices de prevalencia
entre las especies parásitas respecto a los sexos, se sugiere que no existen
grandes diferencias tróficas o de comportamiento entre machos y hembras, y
por tanto grandes diferencias ecológicas entre sexos en la población de esta
Columbida que influyan en las cargas parasitarias. Estos resultados sugieren que
el sexo del huésped no es un factor importante en la estructura de las comunidades
parasitadas.
El análisis de los resultados hallados en las relaciones entre ambos ectoparásitos y
la clase de edad refleja una mayor incidencia de ambos ectoparásitos hacia la
población joven, lo que constituye una evidencia de la influencia de la clase edad de
los hospedadores sobre la estructura de las comunidades parásitas en estas
palomas.
Quizás estas diferencias observadas puedan explicarse por las características
propias de cada clase de edad y sus respuestas ante los parásitos, además existen
otros factores como la habilidad de colonización, capacidad infectiva de las especies

168
parasitas, factores ambientales y ecológicos que condicionan el acceso al
hospedador. Una vez que el parásito llega al hospedero existen “filtros”
inmunológicos típicos de cada clase de edad que determinan cuáles podrán
instalarse y prosperar y cuáles no.
Para Núñez et al. (2011) en general, una mayor masa corporal se interpreta como
índice de mejor condición física porque indica, presumiblemente, la disponibilidad de
reservas de nutrientes que permiten hacer frente a situaciones de elevada demanda.
Es entonces indispensable determinar si existen diferencias en la población
estudiada entre el peso corporal de los ejemplares parasitados y los no parasitados.
Una vez realizada la prueba de normalidad al peso de los ejemplares capturados
separados por las categorías sanos y parasitados (incluye todos los ejemplares con
cualquiera de los dos ectoparásitos hallados en este estudio), se procedió a realizar
la comparación de las medias obtenidas en ambos grupos aplicando el test de T de
Student para muestras independientes.
A pesar de que no se observaron diferencias estadísticamente significativas
entre los grupos de aves (parasitadas y no parasitadas; T de Student (p=
0,189; p >0.05), se puede apreciar una tendencia negativa en el peso de las
aves parasitadas.

169
Figura 32. Comparación de la media (g) de las aves parasitadas y no parasitadas en
relación a la media poblacional de Zenaida macroura (n = 78).
A pesar de no ser fácil establecer las incidencias que pueden generar la parasitosis
en un ave, los cambios en la disminución de la masa corporal encontrados en este
estudio pueden sugerir deficiencias en el estado de salud general de las aves
parasitadas. Por tales razones, es aconsejable para determinar el estado de salud
de la población de Paloma Rabiche calcular índices relacionados con la masa
corporal que puedan ser indicadores confiables del estado de salud poblacional.
Como se ha descrito antes el estudio de la condición de salud de las poblaciones
bajo algún tipo de manejo es necesario, pero este presenta limitaciones, por su
complejidad: técnicas que se aplican, difícil aplicación en el campo, costo económico
de algunos análisis y la existencia de lagunas en el estado de conocimiento de los
patógenos que pudiesen encontrarse. Siempre que sea posible la aplicación de
diferentes técnicas y su combinación con índices darán resultados más confiables.
Por lo tanto aplicar la repetitividad en el tiempo de un índice de condición es clave en
la interpretación de sus resultados. Además, la aplicabilidad de los índices puede
variar entre diferentes especies, por lo que sería importante validar el método en

170
cada caso, algo que en la realidad ocurre con muy poca frecuencia (Núñez et al.,
2011).
10.4.- Conclusiones y recomendaciones
1. Las palomas estudiadas tuvieron un porcentaje moderado de ectoparásitos (34
%). Se encontró en la población dos tipos de ectoparásitos, la Mosca de Gavilán
(Lynchia americana), que se hospeda en el plumaje de la especie y un ácaro
desconocido que parasita las patas de la paloma.
2. El índice de prevalencia de la mosca de gavilán (Lynchia americana) mostró una
alta temporalidad, siendo la época lluviosa la de mayor incidencia en la
población. Correspondió al mes de junio el mayor número de ejemplares
afectados, lo que coincide con la época de máxima reproducción de la especie y
de mayor número de pichones en la población.
3. El ácaro podal es la especie de más frecuente parasitación, aunque existieron
ejemplares con afectación de ambos parásitos. Si bien hubo afectación del
ácaro podal en la mayoría de los meses del año, sus mayores incidencias
coincidieron junto a (Lynchia americana) en la época de lluvias.
4. La ausencia de marcadas diferencias en las cargas parasitarias de ambos
ectoparásitos entre los sexos de los hospedadores, sugiere la no existencia
de diferencias tróficas o de comportamiento entre machos y hembras que
influyan en los índices de prevalencia.
5. La clase de edad más afectada por ambos ectoparásitos resultó ser la población
joven, esta incidencia fue más acentuada en las aves parasitadas por (Lynchia
americana) que por los ácaros podales. Constituye una evidencia de la influencia
de la clase edad de los hospedadores sobre la estructura de las comunidades
parásitas en estas palomas.
6. Debida a la importancia cinegética de estas aves exige continuar realizando
estudios acerca de su estado sanitario para conocer cuáles son las amenazas
que pueden sufrir sus poblaciones y evaluar su evolución en el tiempo.

171
10.5.- Bibliografía
Acosta, M., Mujica, L., Aguilar, S. (2013). Protocolo para el monitoreo de aves
acuáticas y marinas. Centro Nacional de Áreas Protegidas. La Habana. Cuba.
Brown, M. E. (1996). Assessing body condition in birds. Current Ornithology, 13, 67–
135.
Bush, A., K. Lafferty, J. Lotz, A. Shostack. (1997). Parasitology meets ecology on its
own terms: Margolis et al. revisited. J Parasitol, 83, 575-583.
Clayton, D. H. & Moore, J. (1997). Host–parasite evolution: general principles and
avian models. Oxford University Press, Oxford.
Di Mare, M. I. (2003). Densidad poblacional y uso sostenible de la Paloma de alas
blanca (Zenaida asiática) en áreas costeras de Bagaces a Abangares en el área
de conservación Tempisque, Costa Rica. Informe al área de conservación de
Tempisque. http://www.inbio.ac.cr/es/estudios/pdf/informe-PalomaAlaBlanca.pdf
González, D., Castillo, G., López, J., Moreno, L., Donoso, S., Skewes, O., Martínez,
R. y Cabello, J. (2004a). Parásitos gastrointestinales y externos de la paloma
domestica (Columbia livia) en la ciudad de Chillan, Chile. Agro-Ciencia, 20 (2),
107-112.
González, D., Dugschies, A., Rubilar, L., Pohlmeyer, K., Skewes, O. y Eberhard, M.
(2004b). Fauna parasitaria de la tórtola común (Zenaida auriculata, de Murs
1847) (Columbiformes: Columbidae) en Ñuble, Chile. Parasitol Latinoam,
59, 37–41.
Herman, C. M. (1937). Notes Hippoboscid Flies. Departament of Protozoology,
Jhons Hopkins University School of Hygiene and Public Healh. Bird-Banding, 8,
161-166.
Martínez, M., Seguí, M. y López, C. (2009). Bioestadística amigable. Ediciones Díaz
de Santos. España. 293-342.
Nuñes, C., Canestrari, D., Baglione, V. (2011). Evaluación de la condición física en
las aves y su aplicación a los estudios etológicos. Etologuia, 23. 13-42.

172
Pazmiño, A. (2007). Estudio de la fauna parasitaria intestinal en palomas (Columba
livia Gmelin, 1789) de la ciudad de Valdivia. Tesis para optar por el título de
Médico Veterinario. Universidad Austral de Chile, facultad de Ciencias
Veterinaria. Valdivia, Chile.
Peiró, V. (2003). Gestión ecológica de los recursos cinegéticos. Publicaciones de la
Universidad de alicante. España. 136 pp.
Podulka, S., Rohrbaugh, R. and Bonney, R. (2004). Handbook of bird biology.
Segund Editión, CORNELL LAB. OF ORNITOLOGY. USA.
Storer, T. y R. Usinger. (1972). Zoología General. Editora Revolucionaria. La
Habana, Cuba. 273–313.

173
CAPÍTULO X I. APROVECHAMIENTO CINEGÉTICO DE LA PALOMA RABICHE
(Zenaida macroura L.)
11.1.- Introducción
Es bien conocido que el aprovechamiento de las aves en Cuba tiene una gran
tradición y se remonta a tiempos antes de la conquista. Los usos tanto de
ejemplares así como de diferentes partes de animales o sus derivados están
enraizados en nuestra cultura (usados desde los tocados de plumas por nuestros
aborígenes para ceremonias religiosas hasta el uso de diferentes ejemplares para la
alimentación). Actualmente en nuestro país es muy común la tenencia de aves
canoras y de ornato, además de existir gran variedad de especies de aves de interés
cinegético, estando dentro de este grupo la Paloma Rabiche.
Las poblaciones de Paloma Rabiche a pesar de no encontrarse en condiciones de
riesgo, poseen características particulares que requieren un ordenamiento de su
aprovechamiento tradicional, ya que éste puede generar un fuerte impacto sobre sus
poblaciones. A pesar de no contarse con datos exactos sobre el aprovechamiento
de subsistencia realizado sobre esta especie, especialmente el realizado por la
población rural, es importante señalar que éste significa un aporte proteico de gran
relevancia en estas comunidades.
Mientras el hombre no era sino recolector de frutos y plantas silvestres y cazador
ocasional, sus actividades no afectaban al medio ambiente más que las de
cualquiera otra especie de mamífero. Pero comenzó a modificarlo desde el momento
en que, hace miles de años descubrió el fuego e inventó las herramientas y la
agricultura (e. g., El CORREO de la UNESCO, 1980).
Para De Camino y Sabine (1995) la meta buscada no es la no utilización de los
recursos naturales, sino una utilización de forma racional, la cual propicie el
desarrollo y asegure la perpetuidad de los ecosistemas y lo recursos.
Según Peiró (2003), el principal objetivo de la explotación cinegética es optimizar la
utilización del recurso por el hombre, centrados en la obtención de un máximo

174
rendimiento a largo plazo. Para poder llevar a cabo un rendimiento sostenido, es
necesario conocer la forma o modalidades en que son aprovechados estos recursos,
así como cuanto afecta la población objeto de aprovechamiento cada modalidad de
caza.
En consecuencia con lo planteado en párrafos anteriores, los criterios e indicadores
se consideran como un instrumento para evaluar si se ha logrado un progreso hacia
el manejo sostenible o no. Tienen el potencial de ayudar a la orientación de las
políticas forestales, cinegéticas, agrícolas, ambientales y de la investigación, para
guiar las prácticas hacia el manejo sostenible de acuerdo con las expectativas de la
sociedad. Estos no son un fin en sí, sino deberían ser considerados como
herramientas para alcanzar un manejo sostenible, para observar las tendencias
sobre el estado o condiciones del territorio y el manejo del mismo. Una vez
establecidos los indicadores deberán ser monitoreados periódicamente y utilizarse
para mejorar la planificación, ejecución y control del manejo de los recursos del
territorio y del medio ambiente en general.
El objetivo de este capítulo es identificar las principales formas de aprovechamiento
de la Paloma Rabiche y su incidencia en la población, analizar la importancia de esta
paloma en el conjunto de las especies cinegéticas de Cuba, relacionar la comunidad
de depredadores de esta colúmbida, así como proponer criterios e indicadores que
sirvan de herramienta para el manejo sostenible de la especie.
11.2.- Materiales y métodos
Se realizaron varios recorridos en diferentes épocas del año por el área de estudio
para obtener información mediante entrevistas a miembros de la población rural
residentes o campesinos propietarios de tierra que laboran diariamente en ellas y
cazadores ilegales o deportivos sobre las formas de aprovechamiento de la especie
(Anexo 9); se registraron, clasificaron y resumieron las características de estas:
Caza legal, no es más que la realizada al amparo de la ley, sobre especies
cinegéticas silvestre, sin valerse de medios ilícitos, engaños ni trampas de ninguna
clase. Solo existe una forma legal y muy tradicional de aprovechamiento de la

175
especie, la caza con escopetas de cartuchos de diferentes calibres, desde la salida
del sol hasta su puesta, solo los días fines de semanas, feriados y festivos. Hace
unos años (década de los 70 y 80 del pasado siglo) tuvo gran auge, pero hoy está
muy disminuida. Es importante destacar que esta modalidad de caza si no se realiza
de forma racional y organizada puede acarrear grandes desastres, pues se pueden
llegar a diezmar las poblaciones en muy poco tiempo.
Caza ilegal, se realiza de forma ilícita por cazadores furtivos, sin tener en cuenta
vedas, especies autorizadas o restringidas, por lo que va contra todo
aprovechamiento sostenible de la especie capturada. Como forma tradicional de
caza ilegal podemos citar:
Caza con casillas o caravana: modalidad muy antigua y poco usada, llevada a cabo
en zonas rurales, principalmente por campesinos o menores de edad, es una
modalidad que causa poco daño a las poblaciones de la especie y generalmente se
hace para consumo familiar. En la mayoría de los campos de Cuba es usada para la
captura de Codornices (Colinus virginianus), aunque también es usada para la
captura de palomas, pero en menor grado. Es una de las más apropiadas para
estudios donde se requiera capturar el ejemplar sin daños o cuando se requiere
devolverlo a la vida libre (captura y recaptura).
Caza con fusil neumático: modalidad bastante generalizada, que puede ser
practicada de día en “burladeros” (sitios camuflados desde donde se les dispara a
las palomas sin ser visto) donde existen los llamados “comederos” (lugares que por
la concentración de determinados alimentos atrae grandes concentraciones de aves)
de la especie y puede llegar a tener una alta y muy negativa repercusión en las
poblaciones. También se emplea el fusil neumático para la caza nocturna en los
dormitorios con la ayuda de una luz artificial, esta modalidad es igualmente muy
perjudicial, pues generalmente los que la emplean tienen como fin la
comercialización de las piezas cazadas y el lucro sin consideración de los daños
ocasionados.

176
Caza con tira-piedras: modalidad de caza muy popular en todo el ámbito rural del
país, practicada principalmente por niños y adolescentes. Tiene muy bajo impacto
sobre poblaciones cuando se realiza fuera de los picos reproductivos de la especie,
aunque si puede ocasionar daños considerables en épocas de reproducción, pues
puede ocasionar daños considerables en época de reproducción, pues se matan las
palomas en los nidos, lo que las convierte en blanco fácil.
Se clasificaron a los cazadores atendiendo a la edad (menores hasta 19 años, de 20
a 35 años y mayores de 35 años), ocupación y legalidad de la caza.
La importancia cinegética de la Paloma Rabiche dentro de la comunidad de especies
cinegéticas de Cuba se ha obtenido a partir de estadísticas oficiales de caza
disponibles.
Por último, se propone una clasificación de los depredadores de la Paloma Rabiche
en la zona de estudio, adaptada de Peiró (2003), considerando la información sobre
la presencia de estos depredadores por la experiencia acumulada del autor como
cazador y durante el desarrollo de esta tesis.
11.3.- Resultados y discusión
11.3.1.- Formas tradicionales de aprovechamiento de la especie
La Figura 31 refleja el comportamiento de las formas de caza de la Paloma Rabiche
en el área de estudio, donde se puede apreciar un claro dominio de la actividad
ilegal 69 % que se lleva a cabo sobre la especie estudiada. Es importante destacar
que en el área de estudio, al clasificar los grupos de personas que se han observado
practicando la caza ilegal, la incidencia mayor (73 %) es de los menores
(principalmente por niños y adolescentes). Similares resultados encontró Chamizo
(2001) en su trabajo “Análisis de la sostenibilidad de la actividad cinegética en
Cuba”.

177
En la investigación sólo el 31 % correspondió a la caza legal, donde todas las
personas encontradas correspondieron a la categoría “mayores de 35 años”.
Figura 32. Comportamiento del carácter legal o ilegal de la caza de la Paloma
Rabiche en el área de estudio (n= 16).
El aprovechamiento de una especie cinegética en época de veda es una de las
cuestiones que repercuten de manera considerable en la estabilidad o desarrollo de
estas, ya que limita en gran medida la compensación de las pérdidas por mortalidad
natural, más las creadas por el propio aprovechamiento.
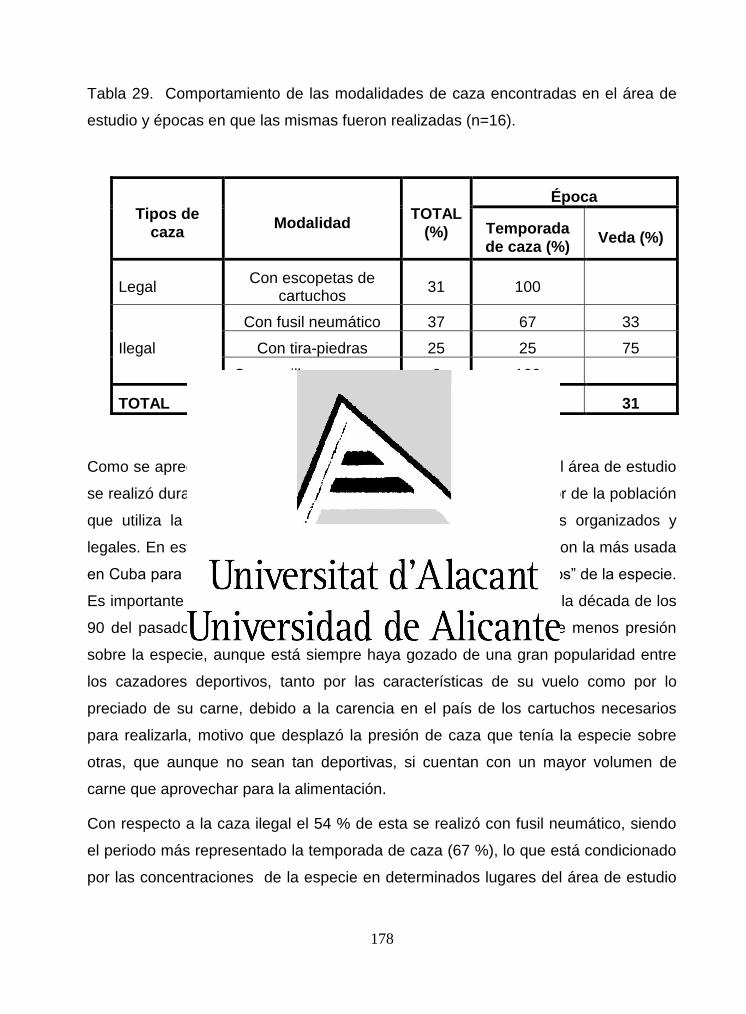
178
Tabla 29. Comportamiento de las modalidades de caza encontradas en el área de
estudio y épocas en que las mismas fueron realizadas (n=16).
Como se aprecia en la Tabla 29, toda la caza legal encontrada en el área de estudio
se realizó durante la temporada de caza, la que representa un sector de la población
que utiliza la Paloma Rabiche que corresponde a los cazadores organizados y
legales. En este caso la modalidad de caza empleada, concuerda con la más usada
en Cuba para la especie estudiada. La caza al acecho en “comederos” de la especie.
Es importante destacar que en la actualidad y desde comienzos de la década de los
90 del pasado siglo, esta modalidad de caza legal cada día ejerce menos presión
sobre la especie, aunque está siempre haya gozado de una gran popularidad entre
los cazadores deportivos, tanto por las características de su vuelo como por lo
preciado de su carne, debido a la carencia en el país de los cartuchos necesarios
para realizarla, motivo que desplazó la presión de caza que tenía la especie sobre
otras, que aunque no sean tan deportivas, si cuentan con un mayor volumen de
carne que aprovechar para la alimentación.
Con respecto a la caza ilegal el 54 % de esta se realizó con fusil neumático, siendo
el periodo más representado la temporada de caza (67 %), lo que está condicionado
por las concentraciones de la especie en determinados lugares del área de estudio
Tipos de
caza Modalidad
TOTAL
(%)
Época
Temporada
de caza (%) Veda (%)
Legal Con escopetas de
cartuchos 31 100
Ilegal
Con fusil neumático 37 67 33
Con tira-piedras 25 25 75
Con casilla o caravana 6 100
TOTAL 100 69 31

179
donde se formaron los “comederos” como campos de Arroz (Oryza sativa), Ajonjolí
(Sesamun indicum), Frijoles (Phaseolus vulgaris) entre otros. De las formas en que
esta se realizaba se hallan el tirar a las palomas posadas y tirar desde “burladeros”
ubicados cerca de las áreas donde se concentran las palomas para comer cuando
están en el suelo o cerca de un árbol con ramas secas o desnudas en las cercanías
de las áreas de alimentación que las palomas usan como perchas y se agrupan en él
un gran número de ellas.
Un resultado nada alentador es el hecho que el 37 % de la caza ilegal realizada con
tira-piedras ocurrió en la temporada de veda para la especie (en Cuba del primero de
abril al tres de septiembre), este tipo de caza es realizada como se describió
anteriormente por personas jóvenes (niños y adolescentes) que como opción más
fácil tiran a las palomas cuando incuban en los nidos y son muy tolerantes a la
presencia humana. Otra forma de realizarla también, es tirando a las palomas
posadas o en el suelo, cuando están distraídas comiendo.
De todas las formas de caza ilegal encontradas en el área de estudio, la de menor
frecuencia (9 %) fue la utilización de casillas o caravana que en nuestra área solo se
encontró un caso. Esta modalidad al ser muy antigua y poco productiva es poco
usada en la actualidad, en el caso encontrado la realizaba un campesino de edad
avanzada. Esta modalidad es generalmente empleada en temporada de caza,
cuando por ser época de sequía existen un considerable porciento de suelo desnudo
y surgen los llamados “bañaderos” (lugares de suelos desnudo y suelto, donde las
aves se revuelcan y asean) que los ojos “bien entrenados” no tienen dificultad para
hallar e incluso hasta conocer las aves que lo usan, por las plumas que en ellos se
encuentran.
Como se aprecia de la situación antes descrita, es evidente que este recurso
renovable no está siendo bien manejado en el área de estudio, dentro de los
principales factores que impiden afectan el uso sostenible de la especie podemos
citar:

180
Escaso personal facultado para hacer cumplir las regulaciones vigentes sobre la
fauna silvestre.
La confiabilidad de los estimados poblacionales, siendo las posibles causas de su
origen:
La confiabilidad de los observadores. Existe inestabilidad con relación a las
personas que realizan los conteos, en algunos casos son cazadores con poco
o ningún conocimiento sobre la materia, en otros casos son personal del
Servicio Estatal Forestal (S.E.F.) o miembros del Cuerpo de Guarda Bosques
(C.G.B.)
Poco seguimiento de las áreas efectivas de la especie. Estas pueden variar en el
transcurso de un periodo de tiempo relativamente corto.
Realización de los conteos en áreas de caza poco representativas de la
distribución territorial de la especie, lo cual tiende a falsear la información.
Existen errores y dudas en cuanto a la clasificación de las áreas dentro de los
territorios cinegéticos (áreas de caza, zona de descanso y nidificación, parcelas
de caza, etc.).
Se desconoce el área efectiva real de la especie (dada por el espacio vital de una
población).
Ausencia de una estadística de caza confiable, lo cual en la mayoría de los casos
evita un monitoreo real de la relación Plan vs Real.
Existencia de la posibilidad de una disminución de los efectivos poblacionales al
controlar los reales cazados por la norma de caza o número de piezas por
cazador (lo que solo incluye las piezas cobradas), no analizando el total de
piezas abatidas, cuestión esta que es la definitoria (Chamizo, 2001).
La influencia del poco o ningún uso de los perros de cobro, lo que hace que se
pierdan muchas piezas, pues aunque las palomas se cazan en sitios de poca
complejidad por sus características de vuelo rápido e inestable, al abatirlas

181
muchas veces caen distantes del cazador y luego el color del plumaje hace el
resto para que la pieza quede camuflada y sea prácticamente imposible de
cobrar (Chamizo, 2001).
11.3.2.- Importancia de la Paloma Rabiche en la comunidad de especies
cinegéticas cubanas
Las palomas forman el grupo de especies cinegéticas más abundantes en el país,
dado esencialmente por las del género Zenaida; además están dentro de las
especies más cazadas, como se observa en la Figura 33 (adaptada de Chamizo,
2001) y confeccionada con datos de siete temporadas de caza entre los años 1999 y
el 2005, tomados de la estadística de caza del Servicio Estatal Forestal del
municipio de Rodas, provincia de Cienfuegos, Cuba.
Figura 33. Participación de especies cinegéticas del municipio de Rodas, provincia
de Cienfuegos de siete temporadas de caza (desde 1999 hasta el 2005).
El género Zenaida se considera de amplia distribución en América e importancia
cinegética, y dentro de este Z. macroura es el ave cinegética más abundante y más
ampliamente distribuida en Norteamérica (Cisneros et al., (2006); Se encuentra
representado en nuestro país por 3 especies, la Paloma Rabiche ( Z. macroura), la
Paloma Aliblanca (Z. asiatica) y la Paloma Sanjuanera o Guanaro (Z. aurita), siendo

182
la primera la de mayor abundancia de todas y uno de los recursos cinegéticos de
mayor disponibilidad en el país. En el área de estudio solo están presentes las dos
primeras especies. La Figura 34, construida con datos oficiales de la estadística del
Servicio Estatal Forestal del municipio de Rodas, provincia de Cienfuegos, Cuba,
refleja la importancia de la Paloma Rabiche dentro del género Zenaida (adaptado de
Chamizo, 2001).
Figura 34. Representación de la Paloma Rabiche en las estadísticas oficiales de
caza del municipio de Rodas (resumen de siete temporadas de caza, desde 1999
hasta el año 2005).
Es importante destacar que la Paloma Rabiche es una especie que tradicionalmente
está presente todos los años en el calendario de caza nacional, y con las cifras más
altas de ejemplares a capturar de todas las especies presentes. Además es la
especie más empleada en la realización de competencias de caza a nivel nacional,
debido a sus características.
Desde fechas tan lejanas como el siglo XIX es descrita como una especie de utilidad
para el hombre por su sabrosa carne (Gundlash, 1876) y luego más tarde Cleveland
(1932) afirma que en la mesa esta paloma resulta ser un manjar muy atractivo.

183
Considerando estos resultados es fácil inferir que la Paloma Rabiche constituye uno
de los recursos cinegéticos más importantes del país.
11.3.3.- Las especies depredadoras de la Paloma Rabiche
En muchos países, los depredadores de las especies cinegéticas han sido, y todavía
lo son en muchos casos, considerados como “alimañas” debido a falsas creencias
tradicionales. Hasta tiempos recientes (década de los 70), se les consideraba como
el principal factor que reducía en número a las especies de caza. Sin embargo, ya
algunos estudios (Errington, 1943, 1946) habían demostrado que los depredadores
no constituyen un factor importante en el control de las poblaciones de las especies
presa, pero sí la disponibilidad de ciertos recursos del hábitat. Hoy se sabe que la
importancia de los depredadores varía según los ecosistemas y que estos pueden,
en ciertas condiciones, ser el principal regulador de la dinámica poblacional de las
especies presa (Sih et al., 1986).
Las especies depredadoras deben considerarse como un recurso natural renovable
tan valioso como el de las especies cinegéticas ya que, aparte de su importancia
ecológica y de algunas estar protegidas, pueden ayudar a mantener a las especies
presa en una buena calidad poblacional (eliminación de enfermos, débiles, etc.). Sin
embargo, esto no implica que deba excluirse un control de aquellas poblaciones de
depredadores abundantes en tal exceso que disminuyan peligrosamente la población
de una especie presa (Peiró, 2003).
Según Peiró (2003), en la gestión ecológica de los depredadores el control de estos
debe realizarse en aquellas áreas en que está claramente justificado por el alto
impacto de la depredación. Pero, en primer lugar, se debe establecer una
clasificación objetiva de los mismos, la cual tendrá en consideración su status y su
potencial impacto sobre las especies cinegéticas. Proponiendo el siguiente modelo
de clasificación:
Domésticos: se trata de animales criados por el hombre, que luego son
abandonados y se asilvestran. Son principalmente los perros y los gatos.

184
Antropófilos: especies silvestres muy adaptadas a explotar los medios antrópicos,
aprovechando los residuos orgánicos de basureros y vertederos incontrolados,
cadáveres de ganados y los encontrados en carreteras. Son principalmente en
Cuba, las ratas (Rattus sp.), el Aura Tiñosa (Cathartes aura) y la Caraira
(Caracara planchus), los que pueden depredar ocasionalmente sobre huevos y
pollos.
Silvestres no protegidos: habitan en medios naturales o poco antropizados y no
gozan de un status de estricta protección. En Cuba, los principales son la
Mangosta (Herpestes auropunctatus), el Jubo Gris (Alsophis cantherigerus), el
Chichinguaco (Quiscalus niger) y el Arriero (Saurothera merlini).
Protegidos: principalmente rapaces diurnas y nocturnas, y ciertas aves y
mamíferos. En Cuba, se encuentran entre ellos el Gavilán Colilargo (Accipiter
gundlachi), el Halconcito (Accipiter striatus), Carpintero Jabao (Melanerpes
superciliaris) y el Zorzal Real (Turdus plumbeus).
En caso de ser necesario el control de los depredadores en un área de caza debe
realizarse sobre las dos primeras categorías, excluyendo siempre a las especies
protegidas. En Cuba, incluyendo la zona de estudio, a no ser en casos muy
puntuales, solo se han considerado históricamente como depredadores altamente
dañinos la Mangosta (Herpestes auropunctatus), el Gato Cimarrón (Felis catus) y el
Perro Jíbaro (Canis familiaris), los que según los calendarios de caza nacionales se
exceptuaban de la veda, estando su caza permitida en todas las épocas del año y en
cualquier lugar.
En el área de estudio no existe ningún trabajo que avale o justifique la caza de
depredadores, aunque, como se explicó anteriormente, la Paloma Rabiche tiene aquí
varias especies depredadoras, incluidos algunos de los más importantes en las dos
categorías que sugiere Peiró (2003) se deben cazar (domésticos y silvestres no
protegidos).

185
11.3.4.- Propuesta de criterios e indicadores de gestión sostenible
La fauna cinegética es un recurso renovable si se logra mantener la calidad de su
hábitat, el éxito reproductor está garantizado; la calidad depende de la disponibilidad
de recursos, alimentos, refugios, agua y perturbaciones humanas entre otros
factores. Como recurso renovable que son, tienen que ser gestionadas de forma que
su aprovechamiento no comprometa su regeneración biológica (Nadal, 1994).
Para un aprovechamiento cinegético sostenible puede resultar muy viable y útil la
delimitación de directrices generales a través del empleo de criterios e indicadores.
Aun cuando esta problemática es muy compleja, es fundamental el planteamiento
correcto y meditado de los indicadores para que permitan establecer correctamente
el protocolo a seguir, siendo éste la guía clave para la aplicación práctica del método
a las necesidades y circunstancias de cada caso. Según el autor de este trabajo,
partiendo de un enfoque holístico se deben considerar los criterios ecológico,
económico, social e institucional, como pilares fundamentales de la sostenibilidad.
La utilización de estos criterios y sus correspondientes indicadores posibilitarían
encuadrar en puntos específicos y también de manera global las bases medibles de
la sostenibilidad para la planificación y el trazado de estrategias de manejo y
aprovechamiento sostenible de los recursos cinegéticos (Tabla 30).
Tabla 30. Criterios e indicadores para el manejo sostenible de la Paloma Rabiche
(Zenaida macroura) en el territorio estudiado propuestos por el autor.
CRITERIOS INDICADORES COMENTARIOS
Ecológico
Calidad del sitio Los diferentes biotopos presentes y su grado de conservación determinan la distribución y la abundancia de la especie.
Tasa de crecimiento poblacional
Determina la planificación del aprovechamiento y la estrategia de manejo de la especie.
Densidad
poblacional Requiere mantenerse en ciertos límites para obtener el máximo rendimiento sostenible.
Estado sanitario de la población
Su monitoreo sistemático posibilita conocer los límites de densidad poblacional más favorables.

186
Nivel de
perturbación Las perturbaciones afectan la conducta y la permanencia de la especie en el territorio.
Económico
Rendimiento Los datos seriados de rendimiento anual permiten evaluar la gestión de caza y hacer reajustes para mejorar la productividad.
Demanda de
mercado
La demanda de caza y consumo de la carne define las posibilidades de comercialización y obtención de ingresos.
Rentabilidad El análisis coste-beneficio permite los reajustes necesarios para conseguir la sostenibilidad económica en la gestión de la especie
Externalidades y
sinergias
El análisis de las interacciones entre la caza y otras actividades económicas del territorio evita conflictos de uso-gestión y potencia las sinergias que elevan la productividad del suelo.
Social
Oportunidad de
empleo
La gestión económica del recurso genera necesidades de plazas de cuidadores, guías de caza, comercializadores y técnicos.
Incorporación de
la mujer
Algunas de las plazas pueden ser ocupadas por mujeres, pues son las que mayor índice de desempleo tienen en el espacio rural.
Beneficios sociales derivados del
manejo
Entre los beneficios más comunes de la gestión cinegética están la disponibilidad de una producción cárnica de alta calidad alimenticia y el aumento de la renta familiar.
Comprensión social de la importancia del recurso
La participación comunitaria en la gestión cinegética conlleva a la interiorización de los conceptos de sostenibilidad y conservación, al sentido de pertenencia y a la percepción de propiedad sobre el recurso natural por parte de la población local.
Institucional
Colaboración interinstitucional
Sin la disposición al entendimiento y la colaboración entre las instituciones que convergen en el espacio rural, desde la familia hasta los organismos del Estado no es posible la sostenibilidad.
Delimitación de responsabilidades, derechos, deberes y obligaciones
El respeto a los derechos individuales e institucionales, así como la delimitación de responsabilidades y el cumplimiento de los deberes y obligaciones son la garantía de un funcionamiento institucional eficiente.
Completamiento y cumplimiento de las regulaciones
necesarias
La actividad cinegética y el aprovechamiento de la fauna silvestre en general, están carentes de regulaciones técnicas, metodológicas, administrativas y legales.
Adecuación a las actuales
Sobre todo para las instituciones responsabilizadas con la gestión del recurso, es muy importante tener

187
condiciones del entorno institucional.
en cuenta las condiciones que favorecen el desarrollo de la actividad y las que tienden a frenarlo o lo niegan, desde el plano material hasta el conceptual.
El criterio ecológico se debe sustentar en los indicadores: calidad del sitio, tasa de
crecimiento poblacional, densidad poblacional, estado sanitario de la población y
nivel de perturbación. El monitoreo de estos indicadores posibilitaría una mejora
integral del hábitat, el establecimiento de unas buenas prácticas agrícolas y
ganaderas, el control de depredadores así como la regulación de la presión de caza,
a partir de la aplicación de unas adecuadas medidas de manejo.
El criterio económico contemplaría como indicadores: rendimiento, demanda de
mercado, rentabilidad, externalidades y sinergias. Con las conclusiones que se
extraen de los datos económicos se posibilitaría detectar la problemática existente
desde el punto de vista de la economía y la actividad humana. El fin último de esta
medición seria corregir los errores y las carencias más significativas para conseguir
la mayor compatibilidad posible entre todos los usos y aprovechamientos.
El criterio social contemplaría dentro de sus indicadores aquellos que reflejarían la
mayor repercusión social de la actividad cinegética: oportunidad de empleo,
incorporación de la mujer, beneficios sociales derivados del manejo y comprensión
social de la importancia del recurso. Esto posibilitaría, a la postre, una mejor
comprensión de la actividad, influyendo positivamente en la conservación de la
naturaleza, el incremento de la renta agraria, la creación de empleos y la fijación de
la población humana en el medio.
Por último, dentro del criterio institucional deben usarse como indicadores:
colaboración interinstitucional; delimitación de responsabilidades, derechos, deberes
y obligaciones; completamiento y cumplimiento de las regulaciones necesarias y
adecuación a las actuales condiciones del entorno institucional. La necesidad de
contar con este criterio se fundamenta sobre todo en la conciencia de que es
necesario un cambio sustancial en las forma de entender la función social de la

188
actividad, su manera de operar y de organizarse. El ejercicio de análisis de este
criterio a la larga ayudaría también a generar un conocimiento más preciso sobre la
misión de las instituciones respecto a la sostenibilidad del aprovechamiento
cinegético, así como a priorizar mejor los objetivos y acciones en este aspecto, a una
mejor planificación del trabajo a corto, mediano y largo plazo y al fortalecimiento del
trabajo en equipos.
Esta propuesta es coherente con lo expuesto por Rojo et. al. (2007) al establecer las
metas e indicadores de éxitos para la UMA en México: “ Las metas a corto, mediano
y largo plazo e indicadores de éxito de las UMA, deberán preverse en función de
las condiciones del hábitat, poblaciones y ejemplares, así como del contexto social y
económico por lo cual se deberán plantear metas e indicadores particulares de
acuerdo con lo establecido en e l artículo 38 del Reglamento”.
A pesar de la reconocida importancia de la Paloma Rabiche como un recurso
cinegético, aún en el país la actividad cinegética no recibe el apoyo y aseguramiento
necesario que garantice obtener el máximo rendimiento sin que el recurso vaya en
detrimento.
Lo anterior se evidencia, según las propias experiencias del autor y lo referido por
Chamizo (2001), a través de los siguientes hechos:
Gran escasez de cartuchos a los cazadores nacionales.
Muy poca o ninguna preocupación por los organismos o instancias que pueden
desarrollar la actividad cinegética.
Desconocimiento de la importancia de la caza.
Un mal enfoque de la política ecológica hacia la actividad de la caza, lo cual crea
una imagen negativa de la misma.
Poco respeto hacia la actividad.
Falta de una ordenación adecuada en la actividad.
Falta de infraestructura capaz de permitir un correcto desarrollo de la actividad.

189
Falta de personal capacitado para desarrollar la actividad.
Existencia de una política que solo ve ganancias en los productos madereros de
los bosques, marginando todos los demás.
Debido a la gran importancia de la especie como recurso cinegético renovable y de
gran interés por parte de los cazadores, tanto nacionales como internacionales se
debería continuar el estudio de la misma con un enfoque holístico, estudio que haga
posible considerar la cinegética como un elemento de interés en el futuro desarrollo
de la agricultura y el desarrollo rural, considerando así la caza como una actividad
más en la zona, junto con la ganadería y los cultivos menores que podrían llevar al
nivel de desarrollo deseado en el medio rural. Basada esta creencia en las siguientes
razones de peso:
La actividad cinegética constituye un sector económico de notable importancia,
generadora de rentas y riquezas, siendo una actividad tan amplia, pero además,
la práctica de la caza, al ser necesario el desplazamiento fuera del núcleo de
residencia, da lugar también a un importante volumen de renta en el sector de
los servicios, hospedajes y gastronomía.
Las profundas raíces sociales que tiene la actividad cinegética, demostrada en la
heterogeneidad (tanto en edades como en profesiones) de sus practicantes, así
como su distribución en todo el territorio nacional (Chamizo, 2001).
La caza genera un número notable de empleo que se ubican en zonas con pocas
opciones y que pueden contribuir a un aumento del nivel económico de muchos
núcleos familiares del medio rural.
La actividad cinegética puede contribuir a un incremento del turismo, sector muy
importante en la economía de nuestro país, favoreciendo su diversificación.
La caza y quienes la practican tienen un compromiso firme y sólido con el
mantenimiento y conservación del medio ambiente, ya que los principales
interesados en la conservación de los espacios naturales en condiciones óptimas

190
son los cazadores, algo imprescindible para la conservación de las poblaciones
de fauna y para el desarrollo rural, contribuyendo a fijar la población humana.
11.4.- Conclusiones y recomendaciones
1. Dentro de las formas tradicionales de aprovechamiento de la Paloma Rabiche
predomina la caza ilegal (69 %), siendo el grupo etario más representado en esta
los niños y adolescentes (73 %).
2. La época de mayor incidencia de caza en el área de estudio resultó ser la
temporada de caza (de septiembre hasta marzo) pero con un predominio de la
caza ilegal en esta época, ya que las modalidades en que se realiza esta
actividad no son las legisladas en el país.
3. De todas las modalidades de caza ilegal encontradas la de mayor incidencia es la
caza con fusil neumático, seguida por la caza con tirapiedras.
4. La caza deportiva organizada y legal solo se encontró en la temporada de caza,
lo que pudiese ser un factor para incentivar un mayor cuidado del recurso pero el
Ministerio de la Agricultura y la Federación Cubana de Caza Deportiva no cuenta
con medios para un uso sostenible de la especie.
5. La Paloma Rabiche es la especie cinegética más importante en el país, por
encima de las anátidas y demás especies del orden Columbiformes.
6. Se recomienda la ejecución de un trabajo que profundice en las relaciones entre
los depredadores y la población de la Paloma Rabiche en el área de estudio para
determinar el grado de incidencia de estos, tanto los domésticos como los
protegidos y no protegidos.
11.5.- Bibliografía
Chamizo, R. (2001). Análisis de Sostenibilidad de la Actividad Cinegética en Cuba.
Tesis Doctoral. Universidad de Alicante, España, Universidad de Pinar del Río,
Cuba. 276 pp.
Cisnero, E., Zamora, H., Mellink, E., Peterson, E., Ochoa, J., Franco, J. y Sánchez, J.
(2006). DGVS, 2006. Talleres sobre conservación y uso sustentable de aves y

191
mamíferos silvestres, en relación con las Unidades de Conservación y Manejo
de Vida Silvestre (UMA) en México. INE-SEMARNAT-UPC. 73 – 106.
Cleveland, A. (1932). Historias de vida familiar de las aves de Norteamérica.
Capítulo “La Paloma de Luto del Este (Zenaida macroura)”. Smithsonian
Institution Estados Unidos. Bolettin Museo nacional 162. Imprenta del Gobierno
de los Estados Unidos. 402-416.
De camino, R. y S. Muller. (1995). Sostenibilidad de la Agricultura y los Recursos
Naturales. 38 Serie Documentos de Programas. Instituto Interamericano de
Cooperación para la Agricultura (IICA). Costa Rica.
Erringon, P. L., (1943). An analysis of mink predation upon muskrats in North Central
U.S.A. Agricultural Experimental Station, Iowa State College of Agriculture,
Research Bull., 320, 798-924.
Erringon, P. L., (1946). Predation and vertebrate populations. Quaterly Review of
Biology, 21, 144-177.
El Correo de la UNESCO. (1980). El hombre y la tierra, una estrategia para
sobrevivir. Año XXXIII.
Gundlash, J. (1876). Contribución a la ornitología cubana. Editora La antillana. La
Habana. 139 – 140.
Nadal, J. (1994). Demografía de la Perdiz roja (Alectoris rufa) en el Bajo Cinca y los
alrededores de Huesca aplicada a la gestión de sus poblaciones durante 1983-
1988. Tesis Doctoral. Facultad de Biología. Universidad de Barcelona.
Peiró, V. (2003). Gestión ecológica de los recursos cinegéticos. Publicaciones de la
Universidad de Alicante. España. 136 pp.
Rojo, A., Cruz, R., Hernández, R. y Moya, E. (2007). Plan de manejo tipo de
Guajalote silvestre (Meleagris gallopavo Lin naeus, 1758). Secretaría de Medio
Ambiente, Recursos Naturales y Pesca. Subsecretaría de Gestión para la
Protección ambiental. Dirección general de vida silvestre. D. F., México.

192
Shi, A., Crowley, P., Mcpeek, M., Petranka, J. y Strohmeier, K. (1986). Predation,
competition and prey communities. A review of field experiments. Annual Review
of Ecology and Systematic, 16, 269-311.

Anexo 1. Ubicación de los muestreos de vegetación en los dos biotopos presentes
en el área de estudio.
.


Anexo 2 - Descripción y clasificación de las especies florísticas presentes en el área
de estudio.
No
Nombre Nombre científico Familia
Cla
s
Abundanci
a Alimento
vulgar relativa Paloma
Agr For
1 Aguinaldo Ipomea sp. Convolvulaceae T 3 2 X
2 Algarrobo Samanea saman Mimosaceae A 2 3
3 Algodón cimarrón
Gossypium barbadensis Malvaceae Arb 1 0
4 Almacigo Bursera simaruba Burseraceae A 2 3
5 Amor seco Aechynomente americana
Leguminoceae H 2 0
6 Arabo Erythroxylon sp. Erythroxylaceae Arb 0 3
7 Aroma blanca Mimosa glauca Mimosaceae Arb 3 3
8 Aroma weiler Mimosa asperata Mimosaceae Arb 2 0
9 Arrocillo Echinochloa crusgalli Poaceae H 3 0
10 Arroz Oriza sativa Graminaceae H 3 0 X
11 Ateje americano
Cordia collococca Boraginaceae A 0 2
12 Ateje de goma
Cordia alba Boraginaceae A 0 1
13 Ayúa Xanthoxylon martniscensis
Rutaceae A 0 3
14 Baria Cordia gerascanthus Boraginaceae A 0 2
15 Bayúa Xanthoxylon elephantiasis
Rutaceae A 0 1
16 Bejuco angarilla
Serjania diversifolia Sapindaceae T 2 3
17 Bejuco uví Cissus cisyoides Vitaceae T 1 2
18 Bien vestido Glyricidia cepium Fabaceae A 1 2
19 Bleo blanco (no esp)
Amaranthus viridis Amaranthaceae H 3 0
20 Bleo espinoso Amaranthus sp. Amaranthaceae H 3 0
21 Boniato Ipomea batatas Convolvulaceae H 3 0
22 Cafetillo Rauwolfia sp. Apocinaceae Arb 0 1
23 Caimitillo Chysophyllum oliviforme Sapotaceae Arb 2 3
24 Cambute Paspalum conjugatum Poaceae H 3 0 X
25 Canutillo Rottboellia conchinchinensis
Graminaceae H 3 0 X

No
Nombre Nombre científico Familia
Cla
s
Abundanci
a Alimento
vulgar relativa Paloma
Agr For
26 Cardo santo Argemone mexicana Papaveraceae H 3 0 X
27 Cebolleta Cyperus rotundus Cyperaceae H 3 0 X
28 Ciguaraya Trichilia havanensis Meliaceae A 2 2
29 Ciruelón Spondias sp. Anacardiaseae A 1 0
30 Cortadera Cladium jamaicense Cyperaceae H 2 0 X
31 Cuero buey Paspalum notatum Graminaceae H 3 0
32 Culantrillo cimarrón
Eryngium foetidum Unbelliferae H 0 2
33 Cundiaol Momordica charantia Cucurbitaceae T 3 2
34 Curujey Tillandsia sp. Bromeliaceae E 1 3
35 Curujey fino Tillandsia sp. Bromeliaceae E 0 1
36 Don carlos Sorghum alepense Graminaceae H 3 0
37 Dormidera Mimosa pudica Leguminoceae H 3 0
38 Escoba amarga
Parthenium hysterophorus
Asteraceae H 3 0
39 Espartillo Sporobolus indicus Poaceae H 3 0
40 Espuela caballero
Jacquina aculeata Teofrastaceae Arb 2 3
41 Frijol Phaceolus vulgaris Fabaceae H 3 0 X
42 Frijol silvestre Phaceolus sp. Fabaceae H 1 0 X
43 Galan Cestrum diurnum Solanaceae Arb 3 0
44 Garro blanco Borreria spinosa Rubiaceae H 3 0
45 Grama caballo
Eleusine indica Poaceae H 3 0
46 Guanina Cassia obtusifolia Leguminoceae H 2 1
47 Guao prieto Comocladia dentata Anacardiaceae A 1 3
48 Guárana Cupania americana Sapindaceae A 0 2
49 Guásima Guazuma tomentosa Sterculiaceae A 2 3
50 Guayaba cotorrera
Psidium guajava Myrtaceae Arb 1 1
51 Guisaso caballo
Triumfetta heterophylla Tliaceae H 3 0
52 Guisaso común
Cenchrus sp. Graminaceae H 3 0
53 Hierba fina Cynodon dactylon Graminaceae H 3 0
54 Hierba guinea Panicum maximun Graminaceae H 3 1 X
55 Hierba hedionda
Cassia occidentalis Leguminoceae H 3 0

No
Nombre Nombre científico Familia
Cla
s
Abundanci
a Alimento
vulgar relativa Paloma
Agr For
56 Higuereta Ricinus comunis Euphorbiaceae H 1 0 X
57 Huevo gallo Tabernamontana citrifolia
Apocinaceae Arb 1 2
58 Jibá Erythroxylon havanensis Erythroxylaceae Arb 1 3 X
59 Jobo Spondias mombin Anacardiaceae A 2 3
60 Lechoza Euphorbia heterophylla Euphorbiaceae H 3 0 X
61 Maíz Zea mays Graminaceae H 3 0 X
62 Malva (1) Sida sp. Malvaceae H 3 0 X
63 Malva (3) Malvastrum coromandelianum
Malvaceae H 3 0 X
64 Malva blanca (2)
Sida cordifolia Malvaceae H 3 0 X
65 Malva blanca (5)
Waltheria indica Malvaceae H 3 0 X
66 Malva mulata (6)
Malacara alceifolia Malvaceae H 3 0 X
67 Malva puerco (7)
Sida rhombifolia Malvaceae H 3 0 X
68 Mamoncillo Mellicocca bijuga Sapindaceae A 1 2
69 Marabú Dichostachys cynerea Mimosaceae Arb 3 3
70 Mastuerzo Lepidium virginicum Cruciferae H 3 0
71 Metebravo Echinochloa colona Poaceae H 3 0
72 Oro azul Phylla strigillosa Verbenaceae H 3 2
73 Palo bronco Malpighia sp. Malpighiaceae Arb 0 2
74 Palo mexicano
Andropongon annulatus Cesalpinaceae A 0 2
75 Pangola Digitaria decumbes Poaceae H 3 0
76 Pasa negro Phylla angulata Verbenaceae H 3 0
77 Pasto camagueyano
Andropongon pertussus Graminaceae H 3 0
78 Pasto mexicano
Andropongon annulatus Graminaceae H 3 0
79 Pata gallina Digitaria saguinalis Poaceae H 3 0
80 Pepino cimarrón
Melothriagua dalupensis Cucurbitaceae T 2 1
81 Piña ratón Bromelia pinguin Bromeliaceae H 1 0
82 Pitilla sabana Andropongon sp. Graminaceae H 3 0
83 Quimbobó Hibiscus esculentus Malvaceae H 3 0
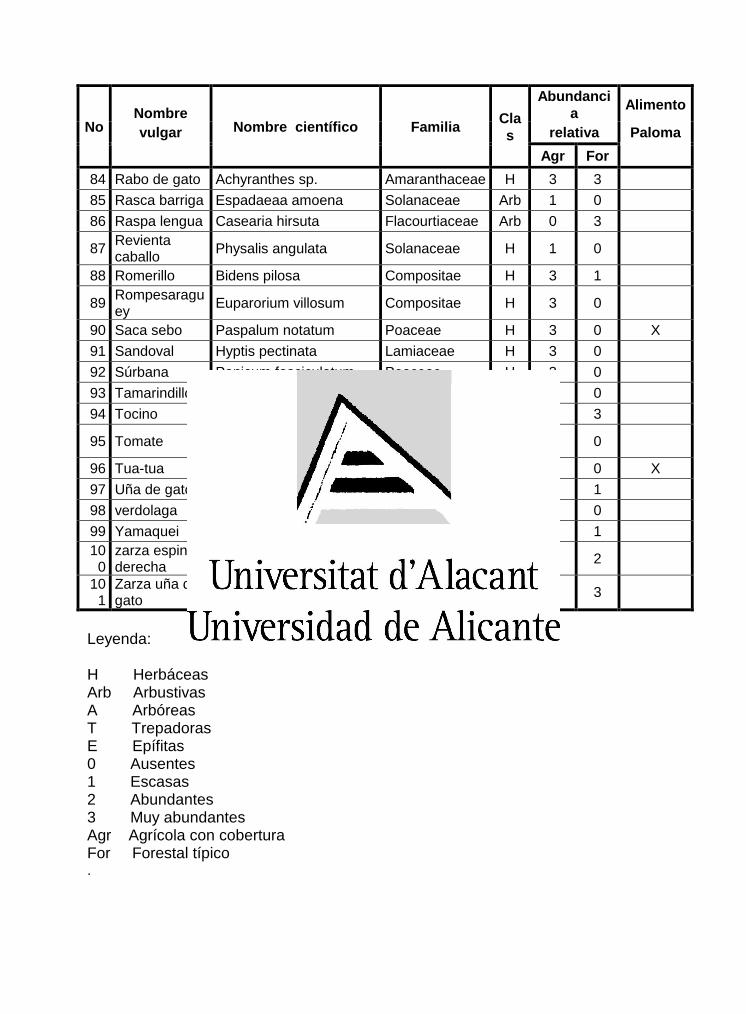
No
Nombre Nombre científico Familia
Cla
s
Abundanci
a Alimento
vulgar relativa Paloma
Agr For
84 Rabo de gato Achyranthes sp. Amaranthaceae H 3 3
85 Rasca barriga Espadaeaa amoena Solanaceae Arb 1 0
86 Raspa lengua Casearia hirsuta Flacourtiaceae Arb 0 3
87 Revienta caballo
Physalis angulata Solanaceae H 1 0
88 Romerillo Bidens pilosa Compositae H 3 1
89 Rompesaraguey
Euparorium villosum Compositae H 3 0
90 Saca sebo Paspalum notatum Poaceae H 3 0 X
91 Sandoval Hyptis pectinata Lamiaceae H 3 0
92 Súrbana Panicum fasciculatum Poaceae H 3 0
93 Tamarindillo Aeschynomene sp. Papilonaceae Arb 1 0
94 Tocino Acacia paniculata Mimosaceae Arb 0 3
95 Tomate Lycopoersicum esculentum
Solanaceae H 3 0
96 Tua-tua Croton lobatus Euphorbiaceae H 3 0 X
97 Uña de gato Pisonia aculeata Nyctaginaceae H 0 1
98 verdolaga Portulaca oleraceae Portulacaceae H 3 0
99 Yamaquei Belairia spinosa Ericaceae A 0 1
100
zarza espina derecha
Casearia sp. Flacourtiaceae Arb 1 2
101
Zarza uña de gato
Zanthoxylum fagara Flacourtiaceae Arb 1 3
Leyenda: H Herbáceas Arb Arbustivas A Arbóreas T Trepadoras E Epífitas 0 Ausentes 1 Escasas 2 Abundantes 3 Muy abundantes Agr Agrícola con cobertura For Forestal típico .

Anexo 3 - Listado de la avifauna y su clasificación.
Nombre vulgar Nombre
científico Orden Familia
Presencia M S
C Pc E
Arriero Sauropthera merlini
Cculiformes Cuculidae X X
Aura Cathartes aura Ciconiiforrmes Cathartidae X X
Azulejo Paserina cyanea Passeriforrmes Fringilidae X X
Bijirita Azul garg. negra
Dendroica caerulescens
Passeriformes Parulidae X X
Bijirita Candelita Sethophaga ruticilla
Passeriformes Parulidae X X
Bijirita Trepadora Mniotilta varia Passeriformes Parulidae X X
Bobito Chico Contopus caribaeus
Passeriformes Tyrannidae X X
Bobito Grande Myarchus stolidus Passeriformes Tyrannidaes X X
Cachiporra Himantopus himantopus
Charadriiformes Recyrvirrostridae
X X
Aguita caiman Butorides virescen
Ciconiiformes Ardeidae X X
Caraira Caracara plancus Falconiformes Falconidae X X
Carpintero de Paso
Sphyrapicus varius
Piciformes Picidae X X
Carpinteo Jabao Melanerpes superficiliaris
Passeriformes Picidae X X
Cartacuba Todus muticolor Coraciformes Todidae X X
Cernícalo Falco sparverius Falconiformes Falconidae X X
Chichinguaco Quiscalus niger Passeriformes Icteridae X X
Codorniz Colinus virginianus
Galliformes Phasianidae X X
Degollado Pheucticus ludovicianus
Passeriformes Frigillidae X X
Frailecillo Sabanero
Charadrius vociferus
Charadriiformes Charadriidae X X
Gallareta Azul Porphyrula martinica
Gruiformes Rallidae X X
Gallareta P. Blanco
Fulica americana Guiformes Rallidae X X
Gallareta P. Rojo Gallinula chloropus
Gruiformes Rallidae X X
Gallito de Río Jacana spinosa spinosa
Charadriiformes Jacanidae X X
Garza Azul Florida caeruela Ciconiiformes Ardeidae X X
Garza Bueyera Bubulcus ibis Ciconiiformes Ardeidae X X
Garza Real Egrretta thula thula
Ciconiiformes Ardeidae X X
Garzón Blanco Ardea herodias repens
Ciconiiformes Ardeidae X X
Gavilán de Monte Buteo jamaicensis Falconiformes Accipitridae X X
Guanabá de Florida
Nycticorax nycticorax
Ciconiiformes Ardeidae X X
Guanabá Real Nycticorax violacea
Ciconiiformes Ardeidae X X

Nombre vulgar Nombre
científico Orden Familia
Presencia M S
C Pc E
Judío Crotophaga ani Cuculiformes Cuculidae X X
Lechuza Tyto alba furcata Strigiforme Tytonidae X X
Malvero Dendroica palmarun
Passeriformes Parulidae X X
Mariposa Passerina ciris ciris
Passeriformes Fringillidae X X
Martín Pescador Ceryle alcyon alcyon
Coraciiformes Alcenidae X X
Mayito Angelaius humeralis
Passeriformes Icteridae X X
Paloma Aliblanca Zenaida asiatica Columbiformes Columbidae X X
Paloma Rabiche Zenaida macroura Columbiformes Columbidae X X
Pato de Florida Anas discor Anseriformes Anatidae X X
Pitirre Abejero Tyrannus dominicensis
Passeriformes Tyrannidae X X
Primavera Coccysus americanus
Cuculiformes Cuculidae X X
Rabuita Polioptila caerulea Passeriformes Polioptilidae X X
Sabanero Sturnella magna Passeriformes Icteridae X X
Sinsonte Mimus polyglottos Passeriformes Mimidae X X
Tojosa Columbina passerina
Columbiforme Columbidae X X
Totí Dives atroviolaceus
Passeriformes Icteridae X X
Tomeguín de la Tierra
Tiaris olivacea Passeriformes Fringilidae X X
Tórtola Turca Streptopelia decaoto
Columbiformes Columbidae X X
Yaguasa Dendrocygna arborea
Anseriformes Anatidae X X
Zorzal Gato Dumetella carolinensis
Passeriformes Turdidae X X
Zorzal Real Tordus plumbeus Passeriformes
Zunzún Chorostilbon ricordii
Apodiformes Apodidae X X
Leyenda:
C Común Pc Poco común E Escaso M Migratoria S Sedentaria

Anexo 4 - Encuesta realizada a campesinos y cazadores.
Usted ha sido seleccionado para participar con su criterio en el desarrollo del
proyecto de investigación sobre la actividad reproductiva de la paloma Rabiche en el
municipio de Rodas que desarrolla la Federación Cubana de Caza Deportiva.
Su criterio resultará muy valioso por lo que esperamos constar con su opinión y
sinceridad en el siguiente cuestionario.
Cordialmente Ing. Forestal Juan Manuel García Bacallao.
Nombre y Apellidos: ____________________________________ Edad: ____
Ocupación: _______________________________Cazador: Si ___ No ____
en caso de serlo años de experiencia: ______
1- ¿Es capaz de identificar la Paloma Rabiche entre otras especies de palomas?
Si ____ No ____
2- ¿En qué época el año es común observar las palomas Rabiches en nidos?
___________________________________________________________________
_________________________________________________________________
3- ¿Dónde ha observado con mayor frecuencia nidos de Palomas Rabiches?
___________________________________________________________________
_________________________________________________________________
4- ¿Ha observado nidos de Palomas Rabiches fuera de la época que usted
considera se reproduce esta especie?

Si ___ No __ en qué época
___________________________________________________________________
_________________________________________________________________
5- ¿Considera usted que ha cambiado la época en que tradicionalmente anida la
especie?
Si _____ No ____ explique:
___________________________________________________________________
_________________________________________________________________
.

Anexo 5 - Localización de nido de Paloma Rabiche sobre una planta espinosa
Zanthoxylum fagara (Zarza uña de gato). Foto del autor
.


Anexo 6 - Localización de nido de Paloma Rabiche sobre una planta espinosa
Dychrostachys cynerea (Marabú). Foto del autor
.


Anexo 7 – Localización de nido de Paloma Rabiche sobre una poste naciente de
Bursera simaruba (Almacigo) en un Lindero. Foto del autor


Anexo 8 – Localización de nido de Paloma Rabiche sobre Cordia alba (Ateje de
goma) al borde de un camino rural. Foto del autor.


Anexo 9 – Primera página del artículo publicado en 2016 sobre la Paloma Rabiche.
The Journal of Animal & Plant Sciences, 26(6): 2016, Page: 1879-1885
ISSN: 1018-7081
FEEDING USE OF A TROPICAL AGROECOSYSTEM (CUBA) BY MOURNING DOVE
(Zenaida macrouraL)
J. M. García 1,2
, V. Peiró3*
1Chapter of Doctorate Thesis of the first author in the doctorate program about “Dvelopment of Tropical
Woodlands, Forest and Tourist Management” from University of Pinar del Rio (Cuba) and University of Alicante
(Spain). 2Forest State Service. Municipal Delegation of Agricultural Ministry (MINAGRY) 39 number 52 street, between
55 and 57, Encanto, Rodas, Cienfuegos, Cuba, [email protected]
3*Director of Thesis from Department of Ecology, Faculty of Sciences, Alicante University Apto. 99, 03080,
Spain *Corresponding Authore-mail:[email protected]
ABSTRACT
The Mourning dove (Zenaida macrouraL.) is a species subjected to high consumptive use in Cuba, but
there area few publications related to its feeding habits in tropical regions. The goal of the
present study was feeding analysis of Mourning dove to know its food spectrum, dietary importance
and impact on a Caribbean agroecosystem. Seventy six samples of dove’s crop were collected in a
hunting area of Cienfuegos province. The farming habitat presented higher feeding quality for this
dove. The diet spectrum showed 88% plant, 9% mineral and 3% from animal origin. The seeds of
Euphorbia heterphylla and Panicum máximum were the most important food items consumed by this
species. Its main diet composed of 79% of seeds from herbaceous plants, with a preference for thes
mall seeds. Crop damage caused by this dove was low, because only 10% of consumed food came
from agricultural species and this dove ate seeds from plants considered weeds by farmers. Thus we
concluded that this dove is a beneficial species in tropical agrosystems.
Keywords: Zenaida macroura, fullness index, food spectrum, index of dietary importance,
agricultural impact.
Introduction
The Rabiche pigeon (Zenaida macroura L.) is an abundant specie from the south of Canada and
United States to the south of Mexico and the Antillas. Zenaida macroura is the only one from five
subspecies that lives in Cuba permanently according to Acosta and Ponce León(2004).This pigeon
frequents pastures, and lands associated to forests, cultivated fields, residential areas and also in urban
parks. As the rest of the pigeons its diet is mainly herbivore, but it has adapted very well to the
agricultural ecosystems, which give it a differential agro ecological characteristic from the rest of the
Columbidae family species. (BELDA et al., 2011; FAHRIG et al., 2011; JIMÉNEZ et al., 2006;
LARRUY; BURGAS, 2004; MEYERS et al., 2005).
Rabiche Pigeon abundance has increased considerably in the last decade that is why it is considered
one of the most important shooting specie of the country. This has to deal with its principal
characteristics (Acosta y Ponce de León, 2004): excellent synergetic aptitude that is evident through
its fast and zigzag fly especially


Anexo 10 – Guía de Entrevista Semi-estructurada para las personas encontradas
realizando la caza de Paloma Rabiche.
Estimado compañero/ a:
La presente entrevista tiene la finalidad de obtener conocimientos sobre el
comportamiento de la población de palomas rabiche en el territorio, debido a su
importancia ecológica y como especie de caza. Deseamos conocer su opinión
acerca de diferentes aspectos relacionados con esta especie y su caza. Las
respuestas ofrecidas en este estudio solo serán utilizadas con fines investigativos.
Le solicitamos su colaboración y aprobación para participar en la investigación y le
agradecemos su contribución. Usted tiene total libertad de participar o no.
CUESTIONARIO.
-Edad
-Sexo
-Ocupación
-¿Dónde reside?
- ¿Con qué medios Ud realiza la caza?
-¿Cuál es la época en que realiza la caza?
-¿Pertenece usted a la Federación Cubana de Caza Deportiva?
-¿Sabe usted cuáles son las formas legales de uso de la especie?


















