
CARACTERIZACIÓN DEL MUTANTE scs5 DE...
60
“CARACTERIZACIÓN DEL MUTANTE scs5 DE Arabidopsis thaliana Y SU IMPLICACIÓN EN EL MECANISMO DE RESISTENCIA A Pseudomonas syringae DC3000”. Trabajo de Fin de Máster Realizado por: Sofía Gabriela Moya Jiménez Directores: Pablo Vera y Beatriz González Valencia, Enero del 2015
Transcript of CARACTERIZACIÓN DEL MUTANTE scs5 DE...

“CARACTERIZACIÓN DEL
MUTANTE scs5 DE Arabidopsis
thaliana Y SU IMPLICACIÓN EN
EL MECANISMO DE
RESISTENCIA A Pseudomonas
syringae DC3000”.
Trabajo de Fin de Máster
Realizado por:
Sofía Gabriela Moya Jiménez
Directores:
Pablo Vera y Beatriz González
Valencia, Enero del 2015

Pablo Vera, Profesor de Invetigaciòn del Consejo Superior de Investigaciones
Científicas
CERTIFICA
Que el presente trabajo titulado “Caracterización del mutante scs5 de Arabidopsis
thaliana y su implicación en el mecanismo de resistencia a Pseudomonas
syringae DC3000” ha sido realizado por Sofía Gabriela Moya Jiménez, bajo mi
dirección en el Instituto de Biología Molecular y Celular de Plantas (UPV-CSIC) y
autorizo la presentación del Trabajo de Fin de Máster el cual se adecua a los
requisitos formales, metodológicos y de contenido, de acuerdo con la normativa
publicada por la Comisión Académica del Máster de Biotecnología Molecular y Celular
de Plantas de la Universidad Politécnica de Valencia.
Y para que conste a los efectos oportunos lo firmo en Valencia a 23 de Enero del
2015.
Director del TFM

A mis padres, por su amor y apoyo incondicional, por ser mis guías y mi
soporte en cada momento y por darme el mejor modelo de justicia y verdad que
he podido tener. A mis hermanos, Marqui, Eli y Enano por estar siempre a mi
lado y llenar de alegría mi vida.
Sofía Moya

AGRADECIMIENTO
Principalmente a Dios quien ha llenado mi vida de muchas bendiciones y
lecciones importantes que me han permitido crecer y ser mejor.
A mis padres por darme su amor y apoyo incondicional en cada paso de mi
vida, por darme el mejor ejemplo, enseñándome que las cosas grandes se
construyen con pasos pequeños.
A la Secretaría Nacional de Educación Superior, Ciencia y Tecnología del Ecuador
(SENESCYT), por brindarme ésta experiencia maravillosa la cual, me ha permitido
crecer tanto personalmente como progresar en mi formación profesional.
A Pablo Vera y su equipo de trabajo. Gracias por hacerme sentir parte
importante de esa gran familiar que es. Gracias por dejarme aprender de
ustedes, por compartir su conocimiento, por llenar el laboratorio de risas y
felicidad. Y sobre todo por ser unos verdaderos amigos. Pero, especialmente a
Vicente y Bea por su ayuda incondicional, su paciencia y consejos. Vicente fue
la persona que con carisma me ayudó a integrarme en el laboratorio y
encabezó esta investigación. Bea por su parte fue el motor de este trabajo y
sin duda alguna no hubiera conseguido llegar a su fin sin su interés, sus
ánimos, sus consejos, y sus enseñanzas. Nunca podré agradecerles por el
tiempo y el trabajo que invirtieron sin esperanzas de una retribución.

CONTENIDO
1 INTRODUCCIÓN ................................................................................................................... 1
1.1 Sistema de defensa en las plantas .......................................................................... 1
1.1.1 Interacción planta-patógeno ................................................................................... 2
1.1.2 Respuestas de defensa en la planta .................................................................... 8
1.2 Antecedentes del trabajo ............................................................................................. 12
1.2.1 P69C, un gen marcador de la respuesta de defensa en las plantas ............ 12
1.2.2 csb3, un mutante alterado en la resistencia frente a patógenos biotrofos .... 13
1.2.3 scs5, un mutante supresor de la mutación csb3 ............................................... 16
3 MATERIAL Y MÉTODOS ................................................................................................... 19
3.1 Material vegetal y condiciones de crecimiento. ....................................................... 19
3.2 Detección de la actividad de la β-D-glucuronidasa ................................................. 21
3.3 Análisis de susceptibilidad/resistencia. ..................................................................... 21
3.3.1 Inoculación por spray de las plantas con la bacteria fitopatógena
Pseudomonas syringae pv. tomato DC3000 (Pst. DC3000). .................................... 22
3.3.2 Curva de crecimiento bacteriano de Pseudomonas syringae pv. tomato
DC3000 (Pst. DC3000) en plantas mutantes csb3scs5 ............................................ 22
3.4 Mapeo genético de la mutación scs5 ......................................................................... 23
3.4.1 Generación de una población F2 recombinante ............................................... 23
3.4.2 Extracción de ADN genómico .............................................................................. 24
3.4.3 Selección de marcadores moleculares para el mapeo ..................................... 24
3.4.4 PCR .......................................................................................................................... 28
3.4.5 Electroforesis en gel de agarosa .......................................................................... 28
3.4.6 Determinación de la frecuencia de recombinación ........................................... 30
4 RESULTADOS Y DISCUSIÓN .......................................................................................... 31
4.1 Caracterización fenotípica de mutantes scs .............................................................. 32
4.2 Caracterización fenotípica del mutante csb3scs5 .................................................... 35
4.2.1. Características morfológicas ............................................................................... 35
4.2.2 Actividad del transgen P69C::GUS..................................................................... 35
4.3 Mapeo genético de la mutación scs5 ....................................................................... 38
4.3.1 Generación de una población recombinante .................................................... 39
4.3.2 Extracción de ADN genómico ............................................................................... 40
4.3.3 Análisis de las frecuencias de recombinación ................................................... 40
5 CONCLUSIONES ................................................................................................................ 47

6 BIBLIOGRAFIA ..................................................................................................................... 48

RESUMEN
En el presente trabajo se utilizó un abordaje genético con el fin de entender en
profundidad la ruta de señalización patogénica controlada por el gen CSB3 y
descrita anteriormente en plantas de Arabidopsis thaliana. En base a un
escrutinio genético sobre el fondo mutante csb3, se buscaron revertientes del
fenotipo asociado a ésta mutación. Dicha estrategia permitió la identificación de
un selecto grupo de supresores que fueron denominados como “supressors of
csb3” (scs) De estos mutantes se seleccionó a scs5. El mutante scs5, además
de suprimir el fenotipo de csb3 y la resistencia incrementada frente a la bacteria
patógena Pseudomonas syringae DC3000, lleva asociadas alteración
morfológicas consistente principalmente en una clorosis en la zona
meristemática de la roseta, y particularmente intensa en los haces vasculares.
Al analizar la susceptibilidad por parte de las plantas csb3scs5 se determinó
que estas plantas son hipersusceptibles a la infección por Pseudomonas
Syringae DC3000 en comparación con plantas silvestres y a plantas csb3. Se
deduce, por lo tanto, que al ser CSB3 un regulador negativo de la resistencia,
SCS5 tiene una función positiva de la misma. A través de la identificación de la
mutación scs5 se aporta un nuevo componente para el mejor entendimiento de
los elementos que se requieren para la correcta activación de mecanismo de
inmunidad en plantas. La localización del locus SCS5 se acotó mediante el
mapeo posicional a través de la utilización de marcadores moleculares SSLPs
y se determinó que este locus está próximo al marcador CTR1.2,
concretamente, en proximidad con la posición 979764bp del cromosoma 5 de
Arabidopsis cercana a la región telomérica del brazo pequeño de dicho
cromosoma. Ésta localización servirá de punto de partida para el cartografiado
de alta resolución y la eventual identificación y clonación del gen SCS5 en
futuras aproximaciones.

ABSTRACT
A genetic approach was used to understand the pathogenic signaling pathway
controlled by CSB3 and which was described previously in Arabidopsis thaliana.
Based on a second-side genetic screening on a csb3 mutant background, we
identified a select group of suppressors that were named as "supressors of
csb3" (scs). Of these mutants we selected scs5. scs5 mutant suppress csb3
phenotype and its increased resistance to the bacterial pathogen Pseudomonas
syringae DC3000. Inn additional scs5 has associated morphological alterations
consisting mainly of meristematic chlorosis rosette area, and particularly strong
in the vascular bundles. By analyzing the susceptibility of csb3scs5 plants we
observed determined that these plants are hypersusceptible to infection by
Pseudomonas syringae DC3000 compared with wild type and csb3 plants. We
can conclude that CSB3 is a negative regulator of resistance, while SCS5 has a
positive role. Through the identification of scs5, a new component for the better
understanding of the elements required for the successful activation of immunity
mechanism in plants is provided. SCS5 locus localization was narrowed by
positional mapping with SSLPs molecular markers in the proximity to marker
CTR1.2 at position 979764bp in Arabidopsis chromosome 5 close to the
telomeric region of the small arm of the chromosome. This location will serve as
a starting point for a high-resolution mapping and the eventual identification and
subsequent cloning of the SCS5 gene in future approaches.

1
1 INTRODUCCIÓN
1.1 Sistema de defensa en las plantas
Las plantas son organismos sésiles y por ello incapaces de moverse de un
lugar a otro cuando se ven expuestas a condiciones medioambientales poco
favorables para su desarrollo. Destacan dentro de estas condiciones
estresantes las que hacen referencia a la sequía, la salinidad, las inundaciones,
las limitaciones de nutrientes o las derivadas como consecuencia del ataque de
microorganismos patógenos e insectos (De Vos, 2006).
Concretamente, se pueden clasificar dos tipos de estreses que pueden alterar
el correcto desarrollo de las plantas. Por un lado el estrés abiótico, el cual,
dependiendo de la naturaleza del agente causal, puede ser de tipo físico,
como sería el estrés hídrico, temperatura o irradiación; o de tipo químico, tal y
como puede ser la derivada de un crecimiento en lugares con una salinidad
elevada, el desbalance nutricional, la presencia de metales pesados, la
aplicación de pesticidas, o incluso variaciones de pH en el suelo. Por otra lado,
se encuentra el estrés biótico, el cual es causado por la presencia de agentes
fitopatógenos tales como virus, bacterias, hongos, nematodos, o incluso por el
ataque de insectos (Dodds y Rathjen, 2010).
Por esta razón, las plantas han desarrollado mecanismos de defensa
sofisticados que le permiten responder y sobrevivir a estas situaciones de
estrés. Ello, a través del desarrollo de estrategias de adaptación que conllevan
a la superación de dicho estrés, y que están fundamentadas en la activación de
programas moleculares controlados a nivel genético (Hückelhoven, 2007). En lo
que respecta a la interacción planta-patógeno, y en particular a aquello que
llamamos como procesos defensivos o antipatogénicos, cabe destacar el
desarrollo de estrategias preinvasivas, que se corresponden generalmente con
el desarrollo de estructuras que actúan a modo de barreras físicas y que están
establecidas antes de la llegada de los agentes patogénicos o del inicio del
proceso infectivo. Además de este tipo de mecanismos de defensa estructural,
y toda vez que un patógeno consigue sortear las barreras preexistentes e
iniciar el proceso infectivo propiamente dicho, se generan diferentes señales y
respuestas moleculares por parte de la planta a la que llamaremos defensa
postinvasiva. Tanto las defensas preinvasivas como las postinvasivas tienen la
función de impedir la entrada y desarrollo del patógeno y por ello son
consideradas fundamentales para el entendimiento de los mecanismos de
resistencia de las plantas a plagas y enfermedades (De Vos, 2006).

2
1.1.1 Interacción planta-patógeno
Los microorganismos patógenos han desarrollado diferentes estrategias de
infección desplegando distintos estilos de vida que son fundamentales para
infectar y colonizar su planta huésped (León-Reyes, 2009). De acuerdo a los
diferentes estilos de vida y estrategias de infección se pueden clasificar a los
microorganismos patógenos de plantas, de forma grosera, en dos grandes
grupos: patógenos necrotrofos y patógenos biotrofos/hemibiotrofos. (Pieterse,
et al., 2009). Los agentes necrotrofos destruyen la célula huésped y se
alimentan de los detritos celulares, de ahí su nombre (Fig. 1). Dentro de este
grupo, y por su importancia en la agricultura, se puede encontrar a los hongos
causantes de podredumbres tales como Botrytis cinerea y Alternaria
brassicicola, entre otros muchos (De Vos, 2005). Por el contrario los patógenos
biotrofos obtienen los nutrientes exclusivamente de tejidos vivos del huésped a
través del establecimiento de diferentes estructuras especializadas que
permiten el contacto directo con el interior de la célula vegetal y con ello el
aprovisionamiento de nutrientes (Fig. 1). Ejemplos de este grupo de
microorganismos patogénicos son los hongos biotrofos Peronospora parasítica
y Erysiphe oronti, o la bacteria hemibiotrofa Pseudomonas syringae (Anderson
et al., 2004).
Figura 1. Hojas de Arabidopsis thaliana infectadas con diferente clase de patógenos.
Representación en hoja adulta del efecto producido en la planta tras la infección por dos
patógenos con diferente estilo de vida: necrotrofo (Botrytis cinerea) y hemibiotrofo
(Pseudomonas syringae). Fuente: Pieterse et al., 2009.

3
1.1.1.1 Defensa preinvasiva
En el curso de la evolución, y como consecuencia de su permanente
interacción con los microorganismos patógenos, las plantas han conseguido
desarrollar una serie de barreras estructurales, así como la síntesis de un
conjunto de compuestos antimicrobianos que eventualmente permiten prevenir
o atenuar la invasión por un gran número de patógenos. Todo ello constituye la
defensa preinvasiva o también llamada defensa constitutiva, siendo ésta la
primera barrera de las plantas para evitar el paso de los microorganismos. Esta
defensa consta tanto de estrategias físicas, tales como la aparición de barreras
estructurales (e.g., cutícula), como químicas, tales como la síntesis de
metabolitos secundarios de naturaleza antimicrobiana (e.g., camalexina)
(Hückelhoven, 2007). La defensa constitutiva se considera propia de cada
especie y se caracteriza por no involucrar una respuesta activa del huésped
ante la presencia del patógeno. La defensa preinvasiva tiene lugar previamente
al reconocimiento del patógeno y sirve como obstáculo para evitar la entrada
de parásitos de tipo inespecífico (Van der Ent et al., 2009). De esta forma, para
que el patógeno pueda llegar a invadir la planta debe penetrar primeramente a
través de las aperturas naturales (estomas) o heridas existentes en la cutícula
protectora que se encuentra sobre las células de la epidermis de las hojas o
raíces. Posteriormente, deberá conseguir disolver o dañar la pared celular, la
misma que a su vez también contiene gran cantidad de componentes o
subestructuras moleculares que a su vez generan resistencia (e.g., lignina,
pectina, callosa) (Agrios, 2005). Los patógenos por su lado también han
desarrollado herramientas que les facilita el acceso a la célula vegetal y a sus
nutrientes. Dentro de estas herramientas cabe destacar la secreción de
enzimas hidrolíticas tipo cutinasas y pectinasas que tiene como el objetivo de
degradar las estructuras protectoras de la planta, y también enzimas
detoxificadoras de determinados metabolitos producidos por la planta como
defensa a estos agentes patogénicos (Dodds y Rathjen, 2010). Una vez
superadas estas barreras, se activa un complejo mecanismo de comunicación
y de interacción célula-célula y que incluye la síntesis de sustancias tóxicas o
efectores que actúan de manera hormonal para así manipular el metabolismo
celular y poder alimentarse de las células de la planta (Van Hulten, 2009).
1.1.1.2 Defensa postinvasiva
Una vez que el patógeno ha superado estas primeras barreras, entra en
funcionamiento el verdadero reconocimiento de éste por parte de la célula
vegetal y se activan diferentes mecanismos de señalización que conllevan a la
activación de la respuesta defensiva inducible o también llamada defensa
postinvasiva. En tal caso, la interacción planta-patógeno puede ser del tipo
compatible o incompatible. Cuando es del tipo compatible, la planta permite el
paso del patógeno (Nurnberger y Brunner, 2002). El microorganismo se replica

4
y avanza hasta el sistema vascular para invadir toda la planta. Por otro lado,
cuando la planta posee los mecanismos necesarios para reconocer y
desencadenar las señales necesarias para evitar el avance del patógeno a los
tejidos vasculares y confinarlo en el sitio de infección se da una interacción
incompatible. El resultado final de esta interacción es alcanzar la resistencia
(Hückelhoven, 2007).
Dentro de este tipo de defensa, las plantas disponen de dos estrategias
sofisticadas para percibir y traducir la señal en una respuesta inmune efectiva.
La primera estrategia de defensa que tiene lugar es la denominada Inmunidad
activada por PAMPs, o PTI, del inglés PAMP- Triggered Immunity. Una vez el
patógeno ha superado esta barrera, se desarrolla la segunda estrategia
denominada Inmunidad activada por Efectores, o ETI, del inglés Effector
Triggered Immunity (Dodds y Rathjen, 2010).
- Inmunidad activada por PAMPs (PTI)
Los PAMPs son patrones moleculares conservados evolutivamente y
esenciales de los patógenos microbianos. Las flagelina, quitina, glicoproteínas,
y lipopolisacáridos son algunos ejemplos. Estas características comunes
permiten identificar patógenos de un mismo grupo, ya que comparte una
respuesta similar por parte de la planta. Los PAMPs son un subgrupo dentro de
los patrones moleculares asociados a microorganismos llamados MAMPs
(Microbe-Associated Molecular Patterns), donde se encuentran incluso
microrganismos no patogénicos (Hückelhoven, 2007). Los PAMPs son
reconocidos por las proteínas receptoras transmembrana de reconocimiento de
patrones (PRR, Pattern Recognition Receptors) caracterizadas por tener una
región rica en leucinas (LRR) localizada en la parte externa de la membrana
plasmática (Fig. 2). Este es el lugar de reconocimiento y unión entre los PAMPs
y los PRR. Estos receptores, dependiendo de la región o estructura existente
en el lado citoplasmático pueden ser de dos tipos. Un grupo son los LRR-RLK
que tienen un dominio serin/treonin quinasa intracelular, mientras que el otro
grupo, denominado LRR-RLP, carece de este dominio (Jones y Dangl, 2006). A
partir del momento en que la planta reconoce los PAMPs se desencadena una
serie eventos de señalización mediados por MAPK (Mitogen Activation Protein
Kinase) hasta permitir la activación de la defensa PTI (Verhage et al., 2010).
Este evento ocasiona la producción de especies reactivas de oxígeno y
compuestos antimicrobianos, reforzamiento de la pared celular mediante la
síntesis de callosa y lignina en los sitios de infección y la activación
transcripcional de genes de defensa con la finalidad de atenuar o impedir el
avance de la infección (Nurnberger y Brunner, 2002).

5
Con el fin de superar estos mecanismos de defensa los patógenos han
desarrollado singulares estrategias que pretenden evitar la activación de la
inmunidad activada por PAMPs. Un ejemplo de esto lo constituyen las
proteínas denominadas “efectoras” que son secretadas o inyectadas en el
citoplasma de la célula huésped mediante dos formas diferentes. Una ocurre a
través de los sistemas de secreción de tipo III (TTSS, TYPE III Secretion
System) en las bacterias, mientras que la otra manera se genera por la
producción de factores de virulencia activados por moléculas de RNA para
suprimir la PTI y promover la virulencia del patógeno. En el caso que el
patógeno logre superar la primera barrera, la planta activa el mecanismo ETI
definido anteriormente (y que desarrollaremos a continuación) logrando con ello
su supervivencia y permitiéndole superar la infección (Dodds & Rathjem, 2011).
- Inmunidad activada por efectores (ETI)
Durante los procesos co-evolutivos entre patógenos y la planta huésped, los
microorganismos han podido desarrollar moléculas efectoras que son patrones
propios de cada microorganismo. Éstas son transportadas a la célula huésped
para que ejerzan su función de suprimir la primera barrera (la PTI) y promover
la virulencia del patógeno. Esto puede dar como resultado la activación de una
segunda barrera que es la inmunidad activada por efectores (ETI). Esta barrera
es específica para la detección del agente infeccioso y tiene como finalidad
proveer a la planta de mecanismos de resistencia frente a patógenos
determinada por sus efectores. Además, estos patrones pueden modificar
proteínas o genes relacionados a la defensa de la célula huésped para su
beneficio (Chisholm et al., 2006). A partir del reconocimiento de los efectores
por parte de proteínas de resistencia, proteínas R (Fig. 2), puede ocurrir que la
planta no sea capaz de reconocer los efectores y activar esta barrera
defensiva, por lo tanto, se producirá la infección y la susceptibilidad activada
por efectores (ETS, Effector-Triggered Susceptibility) (Jones y Dangl, 2006).
El resultado final en esta batalla depende del equilibrio entre la capacidad del
patógeno para suprimir el sistema inmune de la planta y la capacidad de ésta
para reconocer al patógeno y activar de formar eficaz los diferentes
mecanismos de defensa.

6
Figura 2. Representación esquemática del sistema inmune de las plantas. Fuente:
Pieterse et al., 2009. A) A partir del ataque del patógeno, los PAMPs son reconocidos por los
receptores PRRs. En la planta se genera la cascada de señalización para activar la PTI. B) El
patógeno virulento posee efectores para suprimir la PTI, dando como resultado la activación de
la ETS. C) La panta posee proteínas de resistencia adquirida (R) que reconocen a los efectores
de los patógenos, dando como resultado ETI.
Profundizando en lo anterior, el mecanismo de reconocimiento que tiene lugar
en la ETI, también conocido como el modelo de “Interacción gen a gen”, es el
que mejor explica estos eventos. En esta interacción, las proteínas R
codificadas por los genes R por parte de la célula huésped reconocen las
proteínas de avirulencia (Avr) de los patógenos, para desencadenar un
cascada de señalización y alcanzar una respuesta inmunitaria de la planta,
llamada también Respuesta de Hipersensibilidad (HR) (Dangl, et al., 1996 y
Van der Ent, 2009). La Respuesta de Hipersensibilidad (HR) conlleva,
generalmente, activación de muerte celular mediada por la producción de
especies reactivas de oxígeno (ROS), e impide el avance de la infección al
resto de la planta (Pieterse et al., 1999). Por lo tanto, cuando el patógeno
supera la primera barrera de defensa PTI, la planta impide el avance de la
infección a través de la HR y genera resistencia, dándose lugar una interacción
planta-patógeno incompatible. Además, se generan moléculas de carácter
defensivo con la finalidad de proteger a la planta de futuros ataques del mismo
patógeno. Por el contrario, una interacción compatible tendrá lugar cuando no
haya reconocimiento entre los genes R y Avr. Por lo tanto, no se desarrolla la
ETI y la infección se expande por toda la planta produciéndose la enfermedad.
Es decir, la planta será susceptible y el patógeno habrá ganado la batalla
dentro de la interacción planta-patógeno (Verhage et al., 2010).

7
Una vez establecido este modelo y basándose en el estudio de la interacción
entre P. syringae y Arabidopsis, Bienzen y Jones en 1998 propusieron la
“hipótesis del gen guardián”. Este modelo explica como la planta puede
responder a una gran variedad de efectores, es decir, genes Avr, con un
número determinado de genes R, sin necesidad de una interacción física entre
ellos. Las proteínas R están asociadas a otras proteínas de la planta, de las
cuales el patógeno se beneficia para generar la infección. El patógeno utiliza
los efectores para generar modificaciones en estas proteínas asociadas a las
proteínas R, pero estas últimas se encuentran monitoreando constantemente
las modificaciones que se puedan generar. Una vez que se da el
reconocimiento de estas modificaciones por parte de las proteínas R se
desencadena una serie de señales de defensa que tienen como finalidad
detener al patógeno. Explicando así como las plantas pueden reconocer a
miles de patógenos diferentes usando solo una cantidad menor de genes R
(Dangl y Jones, 2001).
Este sistema de inmunidad inducida que se ha descrito anteriormente se
resume en el modelo zig-zag (Fig.3) establecido por Jones y Dangl en el 2006.
Este modelo consiste en cuatro fases. La primera fase ocurre cuando los
PAMPs son reconocidos por los PRR, lo que resulta en un respuesta defensiva
del tipo PTI que impide el avance del patógeno por toda la planta. En la
segunda fase, si el patógeno supera la primera fase, produce efectores
específicos para promover la infección y por lo tanto, hace a la planta
susceptible, es decir se produce una respuesta de tipo ETS. En la tercera fase,
los efectores son reconocidos por las proteínas R dándose una interacción
específica según la hipótesis de la interacción gen a gen o indirecta según la
teoría del gen guardián, lo que desencadena la resistencia a través de la
respuesta de hipersensibilidad (HR). En la última fase, los mecanismos de
adaptación y procesos evolutivos intervienen, haciendo que algunos patógenos
pierdan el efector reconocido y que puedan ganar nuevos efectores,
permitiendo así superar la ETI. Pero, la selección natural también favorece a
las plantas mediante la producción de nuevas proteínas R que puedan
reconocer a los efectores, lo que resulta en la activación nuevamente de la ETI
(Koornneef y Pieterse, 2008).

8
Figura 3. Esquema de la teoría del zig-zag. Fuente: Jones y Dangl, 2006.
1.1.2 Respuestas de defensa en la planta
Una vez que la respuesta defensiva está activa en el sitio de infección, una
respuesta defensiva sistémica se dispara también en las partes distales para
proteger el tejido que no ha sido dañado y conferir resistencia a la planta contra
la invasión del patógeno. Además estos mecanismos tienen también como
objetivo preparar a la planta, manteniendo activas determinadas moléculas
señalizadoras, para futuras infecciones que le permitan reaccionar con mayor
rapidez y/o efectividad y evitar así la enfermedad en la planta. Este fenómeno
constituye la esencia de la resistencia sistémica adquirida, denominada SAR, y
en sí constituye la base de la resistencia inducida o IR. Por lo tanto, en las
plantas hay dos tipos de respuesta: una del tipo local relacionada con la
respuesta de hipersensibilidad (HR). Y otra respuesta, que se produce de
forma sistémica, y que tiene lugar en zonas distales que no han sido infectadas
directamente (Jones y Dangl, 2006).
1.1.2.1 Respuesta local
La respuesta local por parte de la planta al ataque del patógeno ocurre
inicialmente en el sitio de infección después del reconocimiento por parte del
gen de resistencia (R) de la planta al gen de avirulencia (Avr) del patógeno. En
este evento primero se altera el flujo de iones y se activan las vías de
señalización producidas principalmente por proteínas quinasas como las
MAPKs (Mitogen-Activated Protein Kinase). También, se dan cambios en la
regulación transcripcional de genes del hospedador, se refuerza la pared

9
celular, se generan especies reactivas de oxígeno (ROS) y se produce óxido
nítrico (NO). Todo ello como mecanismos de respuestas defensivas. Estos
fenómenos provocan la respuesta de hipersensibilidad (HR) y por ende la
muerte celular del tejido infectado con el objeto de limitar e impedir el avance
del patógeno (Van der Ent et al., 2009).
Las MAPKs (tras fosforilaciones progresivas) permiten la activación
postraduccional de factores de transcripción específicos para promover la
consecuente activación de la expresión de genes relacionados con la defensa.
Por su parte, los procesos oxidativos producidos en la célula infectada se
caracterizan por la producción y acumulación de especies reactivas de oxígeno
como el superóxido (O2) y el peróxido de hidrógeno (H2O2) (De Vos et al.,
2005). La forma de acción de los ROS dependen de la concentración de éstos,
actuando como moléculas señal a bajas dosis o, alternativamente, si sus
niveles son elevados, conducen a la activación de un mecanismo de muerte
celular programada (HR) de tipo apoptótico en las células infectadas. Existe
una relación directa entre el aumento de ROS y NO debido a que este último
tiene la capacidad de inhibir la función de las catalasas y peroxidasas
encargadas de la detoxificación de los ROS (De Wit, 1997).
1.1.2.2 Señalización hormonal
Gracias a estudios bioquímicos, genéticos y moleculares utilizando como
modelo a especies de Arabidopsis thaliana y Nicotiana tabacum se demostró
que las hormonas juegan un papel importante en la regulación del desarrollo y
vías de señalización como respuesta a un amplio rango de situaciones de
estrés (Krouk et al., 2011). La regulación de los mecanismos de defensa
encargados de traducir los eventos de señalización en la planta después del
ataque de un patógeno dependen de la acción de las hormonas. Por lo tanto, la
infección del patógeno estimula a la planta a sintetizar una o varias señales
hormonales dependiendo del tipo de atacante. De los estudios antes
mencionados, se evidenció que los patógenos biotrofos como Pseudomona
syringae y Hyaloperonospera parasítica son generalmente sensibles a la
respuesta defensiva que está regulada por la síntesis y acumulación de Ácido
Salicílico (SA). Sin embargo, los patógenos necrotrofos como Botrytis cinerea y
Alternaria brassicicola están comúnmente controlados por el Ácido Jásmónico
(JA) y el etileno (ET). Además, la respuesta a una herida producida por
insectos herbívoros también está regulada por la ruta de señalización del JA
(Glazebrook, 2005 y De Vos et al., 2005).
El SA es una señal primordial en la defensa sistémica inducida (SAR) como
respuesta al ataque de patógenos. Sin embargo, existen otras hormonas como
las auxinas, o el ácido ascórbico que trabajan de forma conjunta en la compleja
red de interacción que permitirá a la planta tener una adecuada respuesta

10
frente a la presencia de diferentes agentes patogénicos (Chen et al., 2009, y
Pieterse et al., 2009).
Concretamente, el ácido salicílico tiene un papel importante en la fotosíntesis y
la transpiración entre otros procesos fisiológicos necesarios para el óptimo
desarrollo de las plantas. Pero además, el SA tiene un rol esencial en la
respuesta defensiva. La concentración de SA aumenta en el tejido local así
como en el distal después del ataque del patógeno (Koornneef y Pieterse,
2008). Por lo tanto, el SA interviene directamente en la activación de la
resistencia sistémica adquirida (SAR). La ruta dependiente del SA participa en
la expresión de proteínas relacionada a la patogénesis (PR, Pathogenesis
Related) como los genes PR-1 y PR-2. Varios estudios realizados en el
mutante npr1 (nonexpressor of PR genes) en Arabidopsis indican que NPR1 es
un regulador clave en la resistencia sistémica adquirida (SAR) dependiente de
SA. Como respuesta a una infección, se incrementan los niveles de SA en el
citosol y por lo tanto se incrementa la inducción de NPR1, el cual gracias a un
cambio en el potencial redox se reduce de su forma de olígomérica a su forma
monomérica, permitiendo así su paso al núcleo donde finalmente, NPR1
monomérico actúa como factor de transcripción permitiendo la activación del
gen PR-1 (Wu et al., 2012).
El ácido jasmónico está relacionado con la expresión de genes de respuesta a
JA como es PDF1.2 de las siglas en inglés PLANT DEFENSIN 1.2 o LOX2 por
LIPOXIGENASE 2. Al trabajar con el mutante insensible a JA, coi1, se
estableció que el JA-Ile se une al receptor COI1 (Coronatine Insensitive1), que
codifica para una proteína F-box con actividad E3 ubiquitina ligasa de tipo
SKP1-CUL1-F-box (SCF). Este complejo interacciona con proteínas con
dominio-ZIM (JAZ), las cuales son ubiquitinadas permitiendo su degradación
vía proteosoma 26S. Puesto que los JAZ actúan como represores de la
respuesta a jasmónico, al degradarse se liberan los factores de transcripción
tipo MYC y se activa la respuesta a daños mecánicos. Además se activan los
factores de transcripción ERF-1 y ORA59 que regulan la respuesta a
patógenos necrotrofos (Howe, 2004 y De Vos, 2006).
Otra hormona implicada en los mecanismos de defensa es el etileno el cual
actúa de forma sinérgica con el JA, ya que puede inducir genes de respuesta a
JA. La interacción de estas dos hormonas permite la activación de la respuesta
sistémica inducida (ISR) (Van Hulten, 2009).
Las tres hormonas actúan de forma sinérgica o antagónica, permitiendo a la
planta tener una eficiente capacidad reguladora. Por lo tanto, la señalización de
la defensa es específica para patógenos biotrofos o necrotrofos lo que lleva a
la activación de la expresión de los diferentes tipos de genes de defensa.

11
1.1.2.3 Respuesta Sistémica
Una vez que se ha desarrollado la respuesta de defensa local de la planta
después del ataque del patógeno, se produce una serie de respuestas
defensivas sistémicas activadas por señalización hormonal localizadas en
partes distales de la planta para proteger el tejido no dañado de la invasión
subsecuente del patógeno, así como de futuras infecciones (Sticher et al.,
1997). Esta resistencia puede ser de larga duración y de amplio espectro y es
conocida como Resistencia Sistémica Adquirida (SAR). Mientras que, la
resistencia inducida por microorganismos del suelo que establecen una
relación de mutualismo con la planta como las rizobacterias es llamada
Resistencia Sistémica Inducida (ISR) (Van Loon, 1997).
- SAR
SAR es la respuesta sistémica activada en las células no infectadas, por ello
distales al sitio donde ocurrió el ataque del patógeno, y confiere resistencia y
protección de larga duración. La activación de SAR puede ser provocada
durante la interacción planta-patógeno, tanto en la PTI como en la ETI. Gracias
al estudio de mutantes y plantas transgénicas que tienen alterada la
señalización de SA, incapaces de desarrollar SAR y que no muestran activos
los genes PR se determinó que la activación de SAR está generalmente
asociado con el incremento de los niveles del SA, de forma local en el sitio de
la infección y también en el tejido distal (Durrant y Dong, 2004). Y que además,
SAR se caracteriza por la activación coordinada por genes de defensa
pertenecientes de forma mayoritaria a los que codifican proteínas PR. Los
genes PR (Pathogenesis-Related) codifican para una gran cantidad de
proteínas efectivas en la inhibición del crecimiento, multiplicación y propagación
de infecciones y se expresan tanto de forma local como distal. (Van Loon,
1997).
- ISR
Estudios realizados con la rizobacteria no patógena Pseudomons fluorescens
WCS417 y la planta modelo Arabidopsis thaliana permitieron identificar una
nueva forma de resistencia denominada respuesta sistémica inducida o ISR. La
misma se basa en que la colonización vía radicular por esta bacteria induce
resistencia frente a posibles infecciones posteriores. En contraste con la
dependencia de SAR por el ácido salicílico (SA) y el gen regulador NPR1, el
ISR es activada por microoganismos no patogénicos que viven en estado de
mutualismo con la planta, y está regulada por las vías de señalización
dependientes al JA y el ET (Kunkel y Brooks, 2002). Mientras que el SAR es
efectivo frente al ataque de patógenos biotrofos que son sensibles a la defensa
dependientes de SA, el ISR es efectivo frente a patógenos e insectos sensibles
a la defensa dependiente de JA y ET (Van Oosten et al., 2008).

12
1.2 Antecedentes del trabajo
Como se ha dicho anteriormente, la respuesta defensiva de las plantas al
ataque de patógenos implica cambios en la transcripción de numerosos genes.
Y entre ellos se encuentra P69C.
1.2.1 P69C, un gen marcador de la respuesta de defensa en las plantas
Como se describe en Zhao et al., 2000 las proteasas tiene una función
importante en varios procesos celulares incluyendo la diferenciación celular, la
adaptación a diferentes factores ambientales y la misma activación de
mecanismos de defensa. Un ejemplo de proteasas involucradas en la defensa
lo constituyen las subtilasas, en particular las correspondientes al grupo de
proteínas denominadas P69 (Meichtry et al., 1999). Dichas subtilasas son
glicoproteínas que se secretan a la matriz extracelular donde se acumulan y
realizan las funciones, supuestamente, de reconocimiento y procesamiento de
sustratos pericelulares (Matarasso et al., 2005). De los seis miembros de P69
caracterizados en detalle hasta el momento, y que están fuertemente regulados
por factores ambientales y de desarrollo, cuatro (i.e., P69A, P69D, P69E y
P69F) no parecen participar en la respuesta inmune. A pesar de esto, no se
descarta que participen en la primera línea de defensa. Los otros dos
miembros, i.e., P69B y P69C, han sido relacionados con mecanismos de
defensa (Tornero et al., 1996, Jordá et al., 2009, Ramírez et al., 2013). Con el
propósito de profundizar en el entendimiento del control transcripcional que
modula la expresión de genes de defensa en el laboratorio se seleccionó al gen
P69C como un buen candidato para realizar desarrollos de genética reversa y
directa. Para tal fin, en trabajos anteriores en nuestro laboratorio se generaron
plantas transgénicas de Arabidopsis que tenían el gen marcador GUS bajo el
control de la región promotora del gen P69C (Jordá y Vera, 2000). Al realizar
un estudio comparativo de la expresión de la actividad GUS en las plantas
P69C::GUS mediante tinción histoquímica con el sustrato X-Gluc se observó
una activación transcripcional cuando las plantas se inoculan tanto con la
bacteria virulenta Pseudomonas syringae DC3000 (Pst. DC3000) como con la
estirpe avirulenta Pst. DC3000 (avrRpm1). La expresión de GUS dirigida por el
promotor P69C se induce después de la infección y se observa este patrón de
expresión en forma de punteaduras en toda la superficie foliar y no alrededor
de la zona de infección. La expresión acontecía tanto en hojas inoculadas
como en hojas distales al punto de inoculación (Gil, 2005) (Fig. 4).

13
Figura 4: Patrón de expresión del gen marcador GUS bajo el control del promotor de
P69C después de un estímulo patogénico por Pst. DC3000, a nivel local y distal. Fuente:
Gil, 2005.
1.2.2 csb3, un mutante alterado en la resistencia frente a patógenos
biotrofos
Para un mejor entendimiento de la regulación y modo de acción de P69C se
desarrolló una estrategia genética con el fin de identificar genes implicados en
las rutas de señalización de la respuesta defensiva basada en la identificación
de mutantes en base al patrón de expresión de P69C::GUS. Uno de los
criterios de selección aplicado al escrutinio de mutantes estaba basado en la
identificación de plantas en las que, de forma constitutiva y sin la existencia de
ningún estímulo externo, se encontrara activada la expresión del gen GUS
dirigida por el promotor de P69C. De un primer rastreo de 200.000 plantas M2
se seleccionaron 13 individuos que tenían expresión constitutiva del gen
marcador GUS. Más tarde, se comprobó la expresión del gen GUS en estos
individuos en la generación M3 y M4 y, finalmente, se seleccionaron los
mutantes csb1, csb2 y csb3, los cuales llevan este nombre por sus siglas en
inglés constitutive subtilisin (Gil, 2005). En los ensayos histoquímicos de la
expresión del gen marcador GUS, los tres mutantes tienen expresión
constitutiva de GUS. Sin embargo, csb2 y csb3 tienen un patrón de expresión
del gen marcador similar al de la línea parental P69C::GUS, la cual ha sido
infectada previamente con Pst. DC3000 (Fig. 5). Estos datos sugieren que las
plantas csb2 y csb3 tienen alterado alguno de los componentes que participan
en la regulación patogénica del gen P69C. Los análisis genéticos de los
mutantes csb revelan que los tres exhiben una mutación nuclear monogénica
de carácter recesivo con expresión constitutiva de la actividad GUS (Ramírez et
al., 2013). Finalmente, el análisis de la expresión de genes de la ruta de
señalización dependiente al SA, tales como PR-1 y PR-2 y GST6, se observó

14
en el mutante csb3 una mayor expresión de los mismos en ausencia de
estímulo patogénico. Además, este mutante mostró resistencia incrementada a
Pst. DC3000, lo que concuerda con el hecho de que se observe una activación
de estos marcadores de la respuesta defensiva (Fig. 5). Estas características
permitieron seleccionar a csb3 para realizar futuros estudios por su relación
con el sistema de defensa de las plantas (Gil, 2005).
Figura 5 Caracterización del mutante csb3, Fuente (Gil, 2004).
A) Aspecto morfológico de las plantas silvestres y plantas csb3 de cuatro semanas) Tinción
histoquímica de la actividad GUS dirigida por el promotor P69C en plantas silvestres y en
plantas csb3 tras 10 días de crecimiento in vitro. C) Detección de la actividad GUS en hojas de
plantas silvestres y plantas csb3 de cuatro semanas. D) Actividad de la los genes PR-1, PR-2,
GST y PDF1.2 en plantas silvestres y csb3 E) Estudio comparativo del crecimiento bacteriano
en plantas inoculadas a los 3 y 5 días después de la infección con Pst. DC3000.
Con la finalidad de ampliar y corroborar la relación de la resistencia mostrada
por el mutante csb3 con la activación de genes de defensa, se realizaron
análisis transcriptómicos que demostraran este fenómeno. De los 21000 genes
analizados en un chip de Affymetrix AT1 se identificaron 3843 genes que se
sobreexpresan y 2626 genes se reprimen en plantas csb3 en comparación con
el control. Los genes más representativos están ligados al estrés biótico (Gil,
2005). Un ejemplo de genes ligados al estrés biótico encontrados en plantas

15
csb3 mediante el análisis transcriptómico son los genes R involucrados en el
reconocimiento de los genes de avirulencia del patógeno. También, está PAD4
y EDS1 que participan en las primeras etapas de la ruta de transducción de la
señalización mediada por SA. Otro grupo representativo son los pertenecientes
a la síntesis de SA involucrados en la biosíntesis de los fenilpropanoides (Spoel
et al., 2003) así como de la ruta de biosíntesis de SA a partir de isocorismato
en el cloroplasto (ICS1 y EDS5) (Chen et al., 2009 y Dempsey et al., 2011).
Una vez caracterizado fenotípica y molecularmente, tras su clonación por
posición en mapa y paseo cromosómico se estableció que la mutación csb3 es
una pérdida parcial de la función del gen CSB3. Una mutación de pérdida total
en la función del gen CSB3 resulta en letalidad debido a que, según la
descripción de Xiao et al. (2012), CSB3 interviene en la síntesis de
isoprenoides, moléculas estas de cardinal importancia para el desarrollo y
crecimiento de las plantas. CSB3 es un gen de copia única y codifica la enzima
1-hidroxi-2-metil-2-butenil 4-difosfato [HMBPP] sintasa (HDS) de localización
cloroplástica (Fig. 6). Esta enzima cataliza la formación de HMBPP a partir de
2-C-metil-Deritritol 2,4-ciclodifosfato (ME-CPP) y además controla una de las
etapas finales de la síntesis de isopentenil difosfato (IPP) y su isómero
dimetilalil difosfato (DMAPP) a través de la ruta de síntesis de isoprenoides
independiente de mevalonato o ruta llamada MEP (nombre derivado del 2-C-
metil-D-eritritol-4-fosfato) y que se encuentra en el cloroplasto. Tanto la ruta
MEP como la ruta citosólica dependiente de mevalonato (MVA) sintetizan los
precursores de los isoprenoides (Xiao et al., 2012).

16
Figura 6: Ruta de MEP para la biosíntesis de isoprenoides. Fuente: Xiao et al., 2012.
1.2.3 scs5, un mutante supresor de la mutación csb3
La identificación de genes que participan en la ruta de señalización y
resistencia a patógenos biotrofos en la que participa el gen CSB3 es una
estrategia utilizada para comprender el por qué este gen es un regulador
negativo de la resistencia de las plantas. Para llevar a cabo esto, se necesita
identificar mutantes supresores del fenotipo determinado por la mutación csb3.
Para este paso, se mutagenizaron semillas csb3 con EMS siguiendo el mismo
procedimiento realizado para la obtención de los mutantes csb3. Para la
identificación de los mutantes se tuvieron en cuenta dos criterios principales:
reversión del fenotipo característico del mutante csb3 (planta enana con hojas
aserradas) y la supresión de la expresión constitutiva del transgen P69C::GUS.
Tras aplicar el primer criterio de selección se identificaron 35 individuos, a los
cuales posteriormente se les aplicó la prueba histoquímica GUS para
determinar la supresión de la expresión del gen marcador GUS. Gracias a este
ensayo, se descartaron 23 individuos que finalmente resultaron en 11 mutantes
con comportamiento estable tras analizar en M3 y M4. A estos mutantes se les
llamó scs (suppressor of csb3).

17
Dependiendo de la respuesta fenotípica de los supresores scs a Pst. DC3000
se establecieron tres categorías de mutantes scs: mutantes scs que se
comportan como la línea parental csb3 manteniendo la misma respuesta frente
a Pst. DC3000; mutantes scs que revierten la resistencia de csb3 a niveles
comparables a los detectados para plantas silvestres (wt); y mutantes scs que
muestran hipersusceptibilidad a la bacteria (Gil & Vera, 2009). Uno de los
mutantes scs, en los que se basará nuestro trabajo es csb3scs5.
La finalidad de este trabajo consiste en caracterizar al mutante scs5 y
determinar la función reguladora del gen SCS5 en base a la respuesta del
mutante de este gen frente a Pst. DC3000 y la realización de un mapeo
genético grosero para sentar las bases de su futura clonación.

18
2 OBJETIVOS
Las plantas han desarrollado sistemas de defensa complejos con el objetivo de
interactuar y superar el ataque de diversos patógenos. Estos sistemas de
defensa son altamente sofisticados y van dirigidos a reconocer y activar
respuestas frente a las diferentes estrategias de infección. Con las ventajas
que proporciona trabajar con Arabidopsis thaliana, se puede elucidar y
entender de una manera más rápida y sencilla cómo funciona la interacción
planta-patógeno y la compleja regulación que ocurre dentro de las plantas con
el fin de sobrevivir a diferentes tipos de estrés.
Un ejemplo de este tipo de investigaciones relacionada con la interacción
planta-patógeno, lo constituye el estudio de la función de CSB3, el cual posee
un papel de regulación negativa importante en el mecanismo de defensa frente
a patógeno biotrofos mediante la vía de señalización dependiente del SA. Por
lo tanto, en trabajos anteriores en nuestro laboratorio se ha desarrollado una
colección de mutantes con alteraciones en la susceptibilidad y resistencia al
patógeno Pseudomonas syringae DC3000. De estos mutantes, se estudiará
uno en particular. Por ello, para dicha investigación se han planteado los
siguientes objetivos:
- Caracterización del mutante scs5, mutante supresor de la mutación
csb3, en la defensa frente a Pst. DC3000.
- Establecer un rango de la localización de la mutación scs5 mediante
mapeo posicional.

19
3 MATERIAL Y MÉTODOS
3.1 Material vegetal y condiciones de crecimiento.
Arabidopsis thaliana ha sido considerada mundialmente como un organismo
modelo (Pieterse et al., 2009), debido a su corto ciclo de reproducción, tamaño
pequeño, alta producción de semillas viables y por su genoma que además de
ser pequeño se encuentra secuenciado y disponible en bases de datos de
acceso libre (De Vos, 2006). Arabidopsis, como el resto de la plantas, se
encuentra afectada por varios tipos de estreses tanto bióticos como abióticos.
Por lo tanto, la utilización de esta planta en nuestra investigación, será una
base para entender los mecanismos de desarrollo y los procesos moleculares
que están implicados en la defensa. De esta forma, la información obtenida
partir de esta investigación se podrá aplicar para futuros planes de
fitomejoramiento de otras especies vegetales de importancia agronómica y
comercial, garantizando producciones de alta calidad a bajos costos.
Para la caracterización y mapeo de las plantas csb3scs5, mutante objeto de
estudio en este trabajo, se emplearon como controles las plantas silvestres
(WT) de Arabidopsis thaliana (Arabidopsis), del ecotipo Columbia-0 (Col-0) y
Landsberg erecta (Ler). También se utilizó la línea mutante csb3, la cual
proviene de una mutación de semillas de la línea transgénica P69C::GUS.
Esta línea contiene una construcción génica formada por el promotor P69C,
cuyo gen codifica una subtilisina inducible por Pseudomonas syringae
perteneciente a la familia de las P69 de tomate, y el gen reportero GUS. La
utilización de dicha línea para este estudio se basaba en resultados previos
obtenidos en nuestro laboratorio que revelaban una singularidad en el patrón
de expresión de dicho gen tras la infección patogénica en plantas de
Arabidopsis thaliana (Jordá y Vera, 2000), como se ha mencionado en la
introducción. Gracias al trabajo de María José Gil (2005) y de otros
colaboradores del laboratorio se caracterizó tanto física como molecularmente
a csb3. Las plantas csb3 de forma constitutiva y sin la existencia de ningún
estímulo externo, tienen activada la expresión del gen GUS dirigida por el
promotor de P69C. Además, estas plantas se caracterizan por exhibir una
elevada resistencia frente a patógenos biotrofos como P. syringae y H.
parasitica, mientras que, la susceptibilidad frente a patógenos necrotrofos
permanece inalterada.
La línea transgénica de A. thaliana que se pretende caracterizar en este trabajo
es el doble mutante csb3scs5, el cual proviene de un conjunto de 11 mutantes
denominados scs (suppressor of csb3) a partir de semillas csb3. Esta
mutagénesis se realizó con la finalidad de encontrar un supresor del fenotipo
conferido por csb3 que permita identificar genes que participen en la resistencia

20
a patógenos biotrofos mediado por el gen CSB3. Por lo tanto, la relación que
puede tener uno de estos mutantes, debido a su procedencia, con los
mecanismos de resistencia o susceptibilidad de las plantas es lo
suficientemente evidente como para poner de manifiesto la importancia del
objeto de este estudio.
Las semillas de Col-0, Ler, csb3, csb3scs9 y csb3scs5 se obtuvieron de líneas
ya establecidas en el laboratorio. Estas semillas fueron sembradas en macetas
de 12 cm de diámetro con Jiffy-7 (Clause-Tesier Ibérica) de 44 cm de
diámetro. Las semillas fueron sembradas, después de seguir un procedimiento
de estratificación de 3 a 5 días a 4 °C en oscuridad y en humedad con la
finalidad de romper la dormancia de las semillas. Se sembró en macetas
individuales para facilitar el análisis y recogida de material de los ensayos de
susceptibilidad, prueba histoquímica GUS, selección de recombinantes para el
mapeo grosero y recogida de semilla.
Cada bandeja de macetas sembradas se mantuvo cubierta con film
transparente durante 4-5 días para mantener la humedad necesaria para la
germinación y aparición de los cotiledones. Éste se fue retirando gradualmente
durante 4-5 días más para evitar una condensación excesiva de agua y
favorecer la aclimatación. El aporte nutricional se realizó con el riego de las
plantas con una solución nutritiva dos veces por semana. Las plantas
permanecieron de 4 a 5 semanas en cámaras de cultivo, para los ensayos
realizados en este trabajo. Estos fitotrones proporcionan el ambiente adecuado
para este óptimo desarrollo de las plantas en condiciones de 23-25°C con un
fotoperíodo de 8 horas de luz y 16 horas de oscuridad (día corto), intensidad
lumínica de 200 µE m2 seg -1 y humedad relativa del 60%.
Otra parte de la población de plantas obtenidas fue destinada para disponer de
un stock de semillas adecuado. Por tal motivo, las plantas se pasaron a
cámaras de cultivo con día largo para favorecer la floración y maduración de
las plantas. Una vez las silicuas maduraron, las plantas fueron embolsadas y
una vez secas, se limpiaron con coladores de malla fina hasta obtener semillas
libres de impurezas y eliminar los desechos del material vegetal. Las semillas
fueron almacenadas en tubos de microcentrífuga con gel de sílice naranja (T3Q
química) para mantener condiciones adecuadas de ambiente seco que no
perjudique la germinación de las mismas.

21
3.2 Detección de la actividad de la β-D-glucuronidasa
Dentro de las características de reversión del fenotipo de csb3 por parte de los
supresores es que reviertan la expresión constitutiva del gen GUS. Por lo tanto
se comprobó este hecho con la detección de la actividad del gen GUS en las
plantas de csb3scs5. La prueba histoquímica β-D-glucuronidasa (GUS) es un
sistema que permite evaluar los diferentes niveles de expresión de un
determinado gen tras analizar la actividad del gen GUS bajo el control del
promotor del gen de estudio en plantas transgénicas con dicha construcción. La
proteína GUS es una enzima cuya función es catalizar el β-glucorónido, un
producto disponible comercialmente en forma de sustrato. El más utilizado para
esta tinción histoquímica es el X-Gluc y es el que se utilizó en estos ensayos
(5-bromo-4-cloro-3-indolil-ß-D-glucurónido sodio sal; Duchefa Biochemie B.V.;
disuelto previamente en DMSO). Tras la catálisis de este sustrato por parte de
la enzima, se produce un precipitado azul en el sitio en el que se presenta la
actividad enzimática, lo que permite detectar in situ el punto donde se expresa
la enzima GUS y por tanto el gen de estudio (Jefferson et al., 1987). Para
realizar este ensayo, se seleccionaron 3-4 hojas de cada planta Col-0, csb3,
csb3csc5 y csb3scs9 y se infiltraron en tubos de cultivo con la solución de
tinción (Tabla 1), la cual cubría completamente todas las hojas. Para conseguir
una entrada adecuada del compuesto X-Gluc en todos los tejidos de las hojas,
la infiltración se realizó con vacío durante 20 min, el cual se fue rompiendo con
intervalos de 1 minuto aproximadamente. Posteriormente, las hojas infiltradas
con la solución se incubaron a 37°C en oscuridad durante 24 horas para
terminar de mejorar la entrada del sustrato. Finalmente, el revelado de la
tinción se realizó con varios lavados con etanol al 70% (v/v) y observado a las
24 horas.
Tabla 1. Solución de tinción para medir la actividad GUS.
COMPONENTE CONCENTRACIÓN
Tampón fosfato p H 7.2 100 mM
K3Fe(Cn)6 0.5 mM
K4Fe(Cn)6 0.5 mM
EDTA 10 mM
Tritón X-100 0.1% (v/v)
Metanol 0.1% (v/v)
x-Gluc 1 mM
3.3 Análisis de susceptibilidad/resistencia.
Dentro de la caracterización del mutante csb3scs5 y por su relación con el
sistema de defensa de las plantas (mencionado en la introducción) es
importante estudiar la respuesta de este mutante a determinados fitopatógenos
biotrofos. Por ellos, se analizó la respuesta de csb3scs5, supresor de la

22
mutación csb3, frente a la bacteria Pseudomonas syringae pv. tomato DC3000
(Pst. DC3000) que nos permitirá tener una idea del comportamiento del gen
SCS5 en el mecanismo defensivo contra agentes biotrofos.
3.3.1 Inoculación por spray de las plantas con la bacteria fitopatógena
Pseudomonas syringae pv. tomato DC3000 (Pst. DC3000).
El inóculo utilizado proviene de un glicerinado de Pseudomonas syringae pv.
tomato DC3000 (Pst. DC3000) almacenado a -80°C. Este inóculo fue
sembrado en placas de 14 cm de diámetro de King’s B (KB: extracto de carne,
glicerol, 10mL de una solución al 10% de K2HPO4 y 10mL de 10% de MgSO4),
en presencia del antibiótico rifampicina (0.5µl/mL), con ayuda de un asa de
siembra para cubrir todo el área de la placa. Después, ésta fue incubada a
temperatura ambiente durante 48 horas. El manto de colonias crecido en estas
placas fue diluido en una solución de 35mL de sulfato de magnesio (MgSO4) a
10 mM. Posteriormente se midió la Densidad Óptica (DO) de la solución en un
espectofotómetro Eppendorf Biophotometer y se hizo la dilución necesaria para
alcanzar una DO final de 0.1 en 500mL de MgSO4 con 0.02% (v/v) de Silwet.
Una vez preparado el inóculo las plantas fueron inoculadas por spray siguiendo
el método de Kim et al. (2005) en plantas de 4-5 semanas. Las plantas ya
inoculadas permanecieron tapadas con film transparente durante 4-5 horas
para crear el ambiente de humedad y temperatura necesario para garantizar la
entrada del patógeno a la planta.
3.3.2 Curva de crecimiento bacteriano de Pseudomonas syringae pv.
tomato DC3000 (Pst. DC3000) en plantas mutantes csb3scs5
La curva de crecimiento bacteriano fue realizada a los 3 y 5 días post
inoculación. Para ello se usó el extracto crudo por planta de 4 discos de hojas
de 7-9 mm de diámetro, obtenidos con la ayuda de un sacabocados, inmersos
en 600µL de MgSO4 (10 mM). Para cada día de curva se analizaron un total de
12 plantas por genotipo. Una vez los discos fueron triturados en el tampón para
extraer la bacteria, se prosiguió a hacer 6 diluciones seriadas 1:5 en placas de
96 pocillos a partir del extracto crudo inicial. Con ayuda de un replicador, se
cultivaron 5µL de estas diluciones, así como del extracto inicial en placas KB
suplementadas con rifampicina durante 24 horas a temperatura ambiente en
oscuridad. Para el análisis comparativo de la sintomatología se observó cómo
el proceso de infección se desarrolla a los 3 y 5 días después de ser inoculadas
las plantas con la bacteria. Para el estudio del crecimiento bacteriano en la
planta, se analizó el número de unidades formadoras de colonias (ufc)
presentes en el extracto inicial, tomando en cuenta para ello la dilución de
conteo de las ufc, el número de discos utilizados y el número de colonias
contadas. Finalmente, las ufc obtenidas a partir de este cálculo se relacionan a

23
la superficie de hoja que se utilizó, reflejando la media y la desviación estándar
del log de u.f.c/cm2 (unidades formadoras de colonias/cm2). Para la parte
estadística, primero se analizó la existencia de valores aberrantes que se
alejan de una distribución normal. Y finalmente, se realizó una ANOVA estudiar
las diferencias -significativas entre las muestras de los diferentes genotipos
Col-0, csb3, csb3scs5 y csb3scs9.
3.4 Mapeo genético de la mutación scs5
Arabidopsis es una especie modelo para el desarrollo de mapas genéticos
primero porque tiene un tiempo generacional muy corto y produce gran
cantidad de semillas viables, lo que permite tener grandes poblaciones F1, F2 o
F3 y segundo porque existe una cantidad enorme de líneas genéticamente
definidas, además de miles de mutaciones y un gran número de
reordenaciones cromosómicas, información disponibles en grandes bases de
datos. Por otra parte, scs5 es un importante regulador de la defensa estudiado
en Arabidopsis por lo tanto aprovechando las ventajas de trabajar con esta
planta se procedió a hacer un mapeo grosero de la mutación.
3.4.1 Generación de una población F2 recombinante
Una vez que se conoce la respuesta fenotípica morfológica que tienen las
plantas mutantes csb3scs5, se prosiguió a realizar el mapeo de la mutación
mediante un mapa de ligamiento (linkage maps) (Kucuktas et al., 2009). Este
mapa es conocido también como mapa meiótico, ya que durante la meiosis, los
loci de diferentes cromosomas se separan al azar en los gametos. Mientras
que, los loci que están en un mismo cromosoma tienden a cosegregar, al
menos que se produzcan eventos de recombinación que rompan esta
asociación procedente del parental y hayan entrecruzamientos (crossing-over)
entre diferentes partes de los cromosomas (Azofeifa-Delgado, 2006). De esta
forma, la ligación entre dos loci se mide por su frecuencia de recombinación. Por
tanto, se dice que están ligados cuando ésta es menor del 50%. Si esta frecuencia
es de 0, el ligamiento es completo. Teniendo en cuenta la frecuencia de
recombinación entre genes se puede determinar la distancia aproximada que
separa a esos genes en los cromosomas y generar mapas de ligamiento.
Aplicando esta misma premisa para la distancia entre un gen y un marcador
polimórfico de posición conocida en el genoma, se podría determinar con este tipo
de mapeo un posible rango de localización de la mutación scs5. Por lo tanto, para
desarrollar este mapeo hace falta en primer lugar realizar el cruzamiento entre
plantas csb3scs5 con un ecotipo diferente al parental (Col-0). En este caso fue
utilizado el ecotipo Landsberg erecta (Ler) para asegurar plantas con regiones
con alelos de diferente fondo con respecto al parental y determinar las regiones
en las que se habían producido eventos de recombinación. Resultante de la

24
autofecundación de la F1 de este cruce, se obtuvo una población F2. De ésta,
se sembraron 200 plantas, de las cuales fueron seleccionados un conjunto de
recombinantes, en base a seguir mostrando el fenotipo característico de la
mutación scs5.
3.4.2 Extracción de ADN genómico
El ADN genómico de los parentales utilizados (Col-0 y Ler) y de las plantas
recombinantes seleccionadas fue extraído por el método de Rogers y Bendich
(1988), basado en una rápida lisis celular con un tampón de extracción (Tabla
2) (compuesto principalmente por sales y detergente primordial SDS), una
posterior precipitación del ADN mediante isopropanol y la disolución final en
agua.
Tabla 2. Componentes del tampón de extracción de ADN genómico
COMPONENTE CONCENTRACIÓN
Tris HCl, pH 7,8 1M
EDTA 0,5M
SDS 20%
NaCl 5M
Por último, se cuantificó la concentración de ADN extraído mediante un equipo
Nanodrop ND-100.
3.4.3 Selección de marcadores moleculares para el mapeo
Para proceder a mapear genéticamente y obtener una aproximación inicial de
localización cromosómica en el que se encuentra la mutación csb3scs5 primero
se seleccionar los marcadores moleculares a usar. De acuerdo a la naturaleza
del marcador se pueden clasificar en marcadores de proteínas o de ADN. Los
marcadores proteicos determinan variaciones en las cargas eléctricas de
algunas proteínas que comporten el mismo sustrato. Mientras que los
marcadores de ADN son secuencias no codificantes y por lo tanto muy
variables, usados para la construcción de mapas de ligamiento. Existen varias
técnicas para identificar marcadores de ADN, como es por PCR. Dentro de la
categoría que emplea PCR están los microsatélites de tipo SSLPs (“Single
Sequence Length Polymorphism”) que es el tipo de marcador que usamos en
este trabajo. Este tipo de marcadores son polimorfismos de ADN se basan en
tener diferente número de repeticiones en tándem de dos o más nucleótidos
que en general son (CA)n y (GA)n. Los marcadores moleculares utilizados
(Tabla 3) para analizar en los recombinantes escogidos fueron seleccionados a
partir de una base de datos disponibles en el INRA (Institut National de la

25
Recherche Agronomique, París, Francia) según la posición que tienen el
genoma (para que sean equidistantes dentro de cada cromosoma) y en base
también a trabajos previos realizados en el laboratorio. Finalmente, se
seleccionaron 25 marcadores en total (5 marcadores por cromosoma) con la
finalidad de abarcar toda la longitud del mismo y se analizaron para todos los
recombinantes seleccionados. Para acotar más la posible región donde se
encontraba la mutación, se analizaron sobre los recombinantes 3 marcadores
más, cercanos a esa zona determinada.
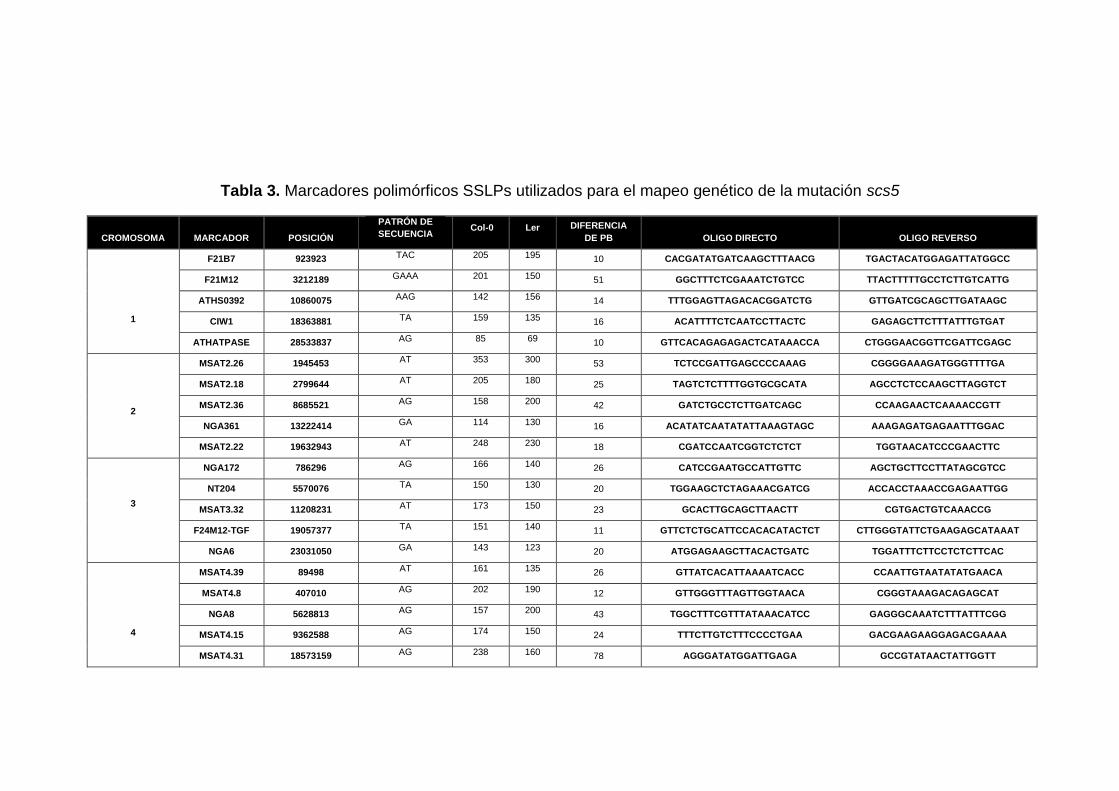
Tabla 3. Marcadores polimórficos SSLPs utilizados para el mapeo genético de la mutación scs5
CROMOSOMA
MARCADOR
POSICIÓN
PATRÓN DE
SECUENCIA Col-0 Ler DIFERENCIA
DE PB
OLIGO DIRECTO
OLIGO REVERSO
1
F21B7 923923 TAC 205 195 10 CACGATATGATCAAGCTTTAACG TGACTACATGGAGATTATGGCC
F21M12 3212189 GAAA 201 150 51 GGCTTTCTCGAAATCTGTCC TTACTTTTTGCCTCTTGTCATTG
ATHS0392 10860075 AAG 142 156 14 TTTGGAGTTAGACACGGATCTG GTTGATCGCAGCTTGATAAGC
CIW1 18363881 TA 159 135 16 ACATTTTCTCAATCCTTACTC GAGAGCTTCTTTATTTGTGAT
ATHATPASE 28533837 AG 85 69 10 GTTCACAGAGAGACTCATAAACCA CTGGGAACGGTTCGATTCGAGC
2
MSAT2.26 1945453 AT 353 300 53 TCTCCGATTGAGCCCCAAAG CGGGGAAAGATGGGTTTTGA
MSAT2.18 2799644 AT 205 180 25 TAGTCTCTTTTGGTGCGCATA AGCCTCTCCAAGCTTAGGTCT
MSAT2.36 8685521 AG 158 200 42 GATCTGCCTCTTGATCAGC CCAAGAACTCAAAACCGTT
NGA361 13222414 GA 114 130 16 ACATATCAATATATTAAAGTAGC AAAGAGATGAGAATTTGGAC
MSAT2.22 19632943 AT 248 230 18 CGATCCAATCGGTCTCTCT TGGTAACATCCCGAACTTC
3
NGA172 786296 AG 166 140 26 CATCCGAATGCCATTGTTC AGCTGCTTCCTTATAGCGTCC
NT204 5570076 TA 150 130 20 TGGAAGCTCTAGAAACGATCG ACCACCTAAACCGAGAATTGG
MSAT3.32 11208231 AT 173 150 23 GCACTTGCAGCTTAACTT CGTGACTGTCAAACCG
F24M12-TGF 19057377 TA 151 140 11 GTTCTCTGCATTCCACACATACTCT CTTGGGTATTCTGAAGAGCATAAAT
NGA6 23031050 GA 143 123 20 ATGGAGAAGCTTACACTGATC TGGATTTCTTCCTCTCTTCAC
4
MSAT4.39 89498 AT 161 135 26 GTTATCACATTAAAATCACC CCAATTGTAATATATGAACA
MSAT4.8 407010 AG 202 190 12 GTTGGGTTTAGTTGGTAACA CGGGTAAAGACAGAGCAT
NGA8 5628813 AG 157 200 43 TGGCTTTCGTTTATAAACATCC GAGGGCAAATCTTTATTTCGG
MSAT4.15 9362588 AG 174 150 24 TTTCTTGTCTTTCCCCTGAA GACGAAGAAGGAGACGAAAA
MSAT4.31 18573159 AG 238 160 78 AGGGATATGGATTGAGA GCCGTATAACTATTGGTT

CROMOSOMA
MARCADOR
POSICIÓN
PATRÓN DE
SECUENCIA Col-0 Ler DIFERENCIA
DE PB
OLIGO DIRECTO
OLIGO REVERSO
5
CTR1.2 979764 CT 159 143 16 CCACTTGTTTCTCTCTCTAG TATCAACAGAAACGCACCGAG
MED24D 1002257 T 302 287 11 GGGGGACCTTTTTCTTGATTACC GCAGAGTCTCACTCTCATCTCC
NGA249 2270216 AG 125 115
10 TACCGTCAATTTCATCGCC GGATCCCTAACTGTAAAATCCC
NGA106 5397351 GA 157 123
34 GTTATGGAGTTTCTAGGGCACG TGCCCCATTTTGTTCTTCTC
MSAT5.14 7498509 AT 221 210 11 AACAACCCTATCTTCTTCTG TGTGACCCCTTACTCAATA
MSAT5.25 13722916 At 162 180 18 GCTTAATTTGGGTTAAAT GCACGCAAGTGACT
CIW9 17044001 TA 165 145 20 CAGACGTATCAAATGACAAATG GACTACTGCTCAAACTATTCGG
MSAT5.19 25924795 AT 208 190 18 AACGCATTTGCTGTTTCCCA ATGGTTATCTCATCTGGTCT
28
3.4.4 PCR
Continuando con el proceso de mapeo, se realizaron PCRs para determinar la
longitud de cada marcador SSLPs y evaluar la región perteneciente a los
recombinantes homocigotos de Col-0 y/o de Ler. Las reacciones de PCR
fueron realizadas en un volumen final de 20µL donde se añadían los
componentes necesarios como son: el par de oligos (directo y reverso) con una
concentración cada uno de 0,4µM para flanquear la región que será
amplificada, tampón 1x para mantener el pH y permitir el normal
funcionamiento de la Taq polimerasa, 100ng/µL del ADN de la muestra
aproximadamente, 0,2mM de un mix de nucleótidos libres y 1unidad/µL de Taq
polimerasa. El programa utilizado para cada PCR (Tabla 4) fue optimizado para
cada marcador, teniendo en cuenta la temperatura media de fusión del par de
oligos utilizado (Tabla 5). adecuada que permite la unión de los oligos a las
cadenas de ADN. Además se optimizó el número de ciclos de la PCR, con la
finalidad de saturar más el ADN amplificado y tener bandas definidas en la
electroforesis. El programa de PCR a excepción de estos dos parámetros
siguió con los siguientes ciclos:
Tabla 4. Programa de PCR
FASE TEMPERATURA TIEMPO
Inicio 95°C 5 min
Desnaturalización 95°C 30 seg
Alineamiento 45-60°C 20 seg
Extensión 72°C 30 seg
Enlongación 72°C 10 min
Con el objetivo de resuspender el producto de PCR y poderlo visualizar se
realizó una electroforesis en gel de agarosa
3.4.5 Electroforesis en gel de agarosa
Después de la realización de la electroforesis para separar los productos de la
PCR en base a la diferencia de tamaño de los polimorfismos determinados por
la longitud de los marcadores determinado para las regiones con alelos
diferentes al parental (Ler) o regiones de recombinación (Col-0). La
concentración de agarosa en el gel se estableció entre 2 y 4% para tener
mayor o menor resolución dependiendo de la diferencia de pares de bases en
la longitud de los microsatélites analizados entre los dos diferentes fondos
genéticos (Col-0 y Ler) (Tabla 5). Las plantas csb3scs5 son de fondo Col-0, el

29
cual, corresponde a uno de los controles utilizados. Las muestras fueron
cargadas en el gel, con un volumen de 15 µL aproximadamente, así como se
utilizaron 6 µL del marcador de 100 pares de bases para poder diferenciar el
tamaño del producto de la PCR en base a los datos teóricos dados por los
marcadores SSLPs para cada ecotipo. Para la electroforesis se aplicó una
corriente de 80 voltios durante una hora aproximadamente para permitir la
correcta diferenciación de las bandas y, comparando con las de los controles,
determinar si los recombinantes eran homocigotos (ya sea Col-0 o Ler) o
heterocigotos para cada marcador. Para la detección de bandas en el gel de
agarosa, se utilizó bromuro de etidio al 6% y el revelado se realizó en un
transiluminador tras exposición con luz UV.
Tabla 5. Programa de PCR y electroforesis para el mapeo de la mutación scs5
CROMOSOMA MARCADOR CAMBIOS EN LAS CONDICIONES DE
PCR % GEL DE AGAROSA
F21B7 Tm 55'' 40ciclos 4
F21M12 Tm 60'' 40ciclos 2
1 ATHS0392 Tm 55'' 40ciclos 4
CIW1 Tm 50'' 40ciclos 4
ATHATPASE Tm 55'' 40ciclos 4
MSAT2.26 Tm 55'' 40ciclos 2
MSAT2.18 Tm 60'' 40ciclos 2
2 MSAT2.36 Tm 55'' 40ciclos 2
NGA361 Tm 60'' 40ciclos 4
MSAT2.22 Tm 55'' 40ciclos 3
NGA172 Tm 55'' 40ciclos 2
NT204 Tm 55'' 40ciclos 3
3 MSAT3.32 Tm 60'' 40ciclos 2
F24M12-TGF Tm 60'' 40ciclos 4
NGA6 Tm 55'' 40ciclos 2
4
MSAT4.39 Tm 55'' 50ciclos 2
MSAT4.8 Tm 55'' 50ciclos 3
NGA8 Tm 55'' 50ciclos 2
MSAT4.15 Tm 55'' 50ciclos 2
MSAT4.31 Tm 55'' 50ciclos 2
CTR1.2 Tm 55'' 55ciclos 4
MED24D Tm 55'' 55ciclos 4
NGA249 Tm 55'' 55ciclos 4
NGA106 Tm 55'' 55ciclos 2
5 MSAT5.14 Tm 60'' 40ciclos 4
MSAT5.25 Tm 50'' 60ciclos 4
CIW9 Tm 55'' 60ciclos 3
MSAT5.19 Tm 45'' 60ciclos 4

30
3.4.6 Determinación de la frecuencia de recombinación
Finalmente, para cada marcador analizado, se cuantificó el número de plantas
recombinantes que eran homocigotos Col-0 (C), homocigotos Ler (L) o
heterocigotos (H). Con el número total de eventos de recombinación
producidos (determinado por la presencia de L y H y el total de recombinantes
analizados (T) se obtiene el porcentaje de recombinación dado por la siguiente
fórmula:
% 𝑑𝑒 𝑅𝑒𝑐𝑜𝑚𝑏𝑖𝑛𝑎𝑐𝑖ó𝑛 = (𝐿 + 𝐻
𝑇) ∗ 100
Con el cálculo de este porcentaje de recombinación, se podrá determinar la
distancia entre el marcador analizado y la mutación. De forma, que cuanto
menor sea este porcentaje, menor es la distancia entre ambos ya que ya que la
probabilidad de recombinación es menor. Así, como se ha explicado en el
apartado anterior (3.4.1) por el mapeo por ligamiento, se podrá establecer una
región posible donde se encuentra la mutación.

31
4 RESULTADOS Y DISCUSIÓN
En los últimos años, y más en el ámbito de la investigación básica, se ha
generado un notable interés en la comprensión de los mecanismos moleculares
implicados en las rutas de señalización que controlan la activación de los
sistemas de respuestas defensivas de las plantas frente al ataque de los
patógenos. Todo ello, con el fin de conocer en profundidad los procesos de la
inmunidad vegetal y de esta forma mejorar la capacidad de las plantas para
defenderse de este tipo de estrés biótico (Glazebrook, 2001). Una de las
formas de abordar este problema es utilizar Arabidopis thaliana ya que, como
sistema modelo, facilitará la realización de estos estudios por las ventajas que
conlleva el uso de la misma. De esta forma, podremos identificar nuevos genes
implicados en las rutas de señalización que regulan el sistema de defensa de
las plantas.
Dentro de este contexto, se trabajó en nuestro laboratorio con el mutante csb3,
alterado en la expresión del gen P69C. Las plantas csb3 muestran expresión
constitutiva del transgen P69C::GUS, y tienen la capacidad de conferir
resistencia frente a la bacteria fitopatógena Pseudomonas syringae DC3000.
Además, estas plantas tienen un marcador morfológico (son plantas enanas
con hojas aserradas) (Fig. 6), que facilita su reconocimiento y utilización en
posteriores estudios (Jordá y Vera, 2000). Por lo tanto, una forma de
profundizar e identificar los genes que participan en la ruta de señalización
mediada por el gen CSB3 y el mecanismo de acción frente a agentes
biotróficos, es a través de la identificación de mutantes supresores del fenotipo
dado por la mutación csb3.
Tras llevar a cabo esta estrategia genética de identificación de supresores de
un fenotipo mutante, se identificaron los mutantes scs (suppressor of csb3).
Estos mutantes revierten la morfología y la resistencia asociada a la mutación
csb3 a niveles de plantas silvestres. Dentro de este conjunto, se escogió para
este trabajo el mutante csb3scs5, con el fin de caracterizarlo y establecer una
localización aproximada del locus responsable de dicha mutación. Estos
estudios permitirán entonces conocer más acerca de los mecanismos
implicados en la defensa de las plantas frente a patógenos biotrofos.
Existen varios trabajos que han seguido esta misma estrategia de búsqueda y
estudio de supresores de un fenotipo. Un ejemplo es la identificación de genes
que intervienen en la señalización de NPR1. Este gen es un co-regulador
transcripcional importante para la activación de los genes dependientes de SA
y por lo tanto relacionado con la defensa frente a agentes biotrofos (Wu, et al.,
2012). Tras estos estudios se identificó el mutante ssi2, supresor de npr1,
afectado en la señalización de la ruta de defensa dependiente a NPR1 (Shah et

32
al., 2001). Con la finalidad entender mejor este mecanismo, se buscaron
supresores de este supresor. Así se identificó el mutante sfd que suprime el
fenotipo de ssi2 (enanismo), la expresión de NPR1 y la resistencia a
Pseudomonas syringae. Gracias a este abordaje genético que permitió
identificar al mutante sfd2, (el gen SFD codifica enzimas involucradas en la
biosíntesis de lípidos), se demostró que los lípidos son mediadores importantes
en la activación de SAR, además de tener importancia en procesos de
desarrollo y crecimiento en Arabiodpsis (Nandi et al., 2003). Estos trabajos
demuestran las ventajas que conlleva desarrollar este tipo de abordajes y
además, generan gran cantidad de información relevante acerca del
funcionamiento de los mecanismos de defensa.
4.1 Caracterización fenotípica de mutantes scs
De la identificación de los mutantes scs, que se llevó a cabo en trabajos previos
de este laboratorio, en base a los dos criterios mencionados: reversión del
fenotipo morfológico y la supresión de la expresión constitutiva del transgen
P69C::GUS características de csb3. En este trabajo se utilizaron 7 de ellos
para realizar una previa caracterización fenotípica, siendo éstos los mutantes
csb3scs2, csb3scs5, csb3scs6, csb3scs7, csb3scs9, csb3scs11 y csb3scs12.
Estos mutantes fueron los de mayor relevancia debido a sus características
morfológicas y diferentes grados de susceptibilidad frente a Pst. DC3000. En
esa caracterización morfológica de los mutantes (Fig. 7) se observó que los
mutantes scs revierten el fenotipo de csb3 en relación a tamaño y forma de las
hojas en diferentes grados. Por ejemplo los mutantes csb3scs2, csb3sc7,
csb3scs11 y csb3scs12 se asemejan más a plantas Col-0. Estos mutantes
antes mencionados tienen las hojas de mayor tamaño y además no son
aserradas como csb3. Los mutantes csb3scs5 y csb3scs9 además de revertir
el fenotipo característico de csb3 tienen otros aspectos adicionales asociados,
tal y como puede ser el presentar una clorosis tenue en el centro de la roseta
de la planta.

33
Figura 7. Caracterización de 7 mutantes scs, supresores del mutante csb3.
Continuando con la caracterización de estos mutantes, se analizó la respuesta
de las plantas frente a la infección con la bacteria compatible Pst. DC3000.
Para ello, se inocularon por spray los mutantes scs, plantas silvestres (Col-0) y
el mutante csb3 (ambos como controles de infección y resistencia,
respectivamente). Finalmente, en estos ensayos, se observó la sintomatología
de las plantas a los 3, 5 y 7 días post inoculación (Fig. 8). Como es sabido, los
síntomas característicos de la infección con Pst. DC3000 en las hojas de
Arabidopsis corresponden a manchas cloróticas de pequeño tamaño que
pueden observarse a los dos o tres días después de la infección. Las lesiones
se expanden y a menudo éstas áreas se necrosan y adquieren un color
castaño claro a blanco en una semana (Cao et al., 1997 y Katagiria et al.,
2002).
Según la sintomatología observada, los mutantes scs se pudieron dividir en dos
categorías. La primera categoría englobaba a los mutantes csb3scs2,
csb3scs6, csb3scs7, csb3scs11 y csb3scs12 que corresponde a los mutantes
que adquierieron un nivel similar a plantas silvestres, sin muchos síntomas de
infección. Y en la segunda categoría encontramos los mutantes csb3scs5 y
csb3scs9, que corresponde a aquellos que mostraron una elevada
sintomatología (manchas cloróticas) en las hojas tras la infección (3 y 5 días
después de la infección), la cual aumenta con el tiempo a lo largo de las hojas.
Además, los síntomas de csb3 scs5 y csb3 scs9 son comparables a las plantas
mutantes npr1, eds, pad4. Éstos mutantes se ven afectados en la activación de

34
las respuestas defensivas durante la interacción compatible por lo que
muestran elevada susceptibilidad a la infección P. syringae (Glazebrook, 2001).
En trabajos que se estaban llevando a cabo en este grupo, se había estudiado
ya en el mutante csb3scs9 el nivel de crecimiento de la bacteria tras este tipo
de inoculación y se observó que las plantas eran hipersusceptibles con
respecto a Col-0. Por lo tanto, dada la similitud en la sintomatología con
respecto al mutante csb3scs5, era interesante estudiar el fenotipo que se
obtendría en este mutante, ya que podría también estar relacionado con la
susceptibilidad a Pst DC3000. Además, csb3scs9 podría ser utilizado como un
buen control positivo de susceptibilidad para los ensayos de estudio de
crecimiento de la bacteria.
Figura 8. Sintomatología de los mutantes csb3scs2, csb3scs5, csb3scs6, csb3scs7,
csb3scs9, csb3scs11, csb3scs12, csb3 y en plantas silvestres (Col-0) a los 3, 5 y 7 días
post inoculación (dpi) con Pst. DC3000.
Por lo tanto, como resultado de esta caracterización previa de los mutantes
aquí mostrados, se seleccionó el mutante csb3scs5 para llevar a cabo en este
trabajo su caracterización y mapeo grosero. Debido a las características
mostradas de clorosis y una mayor sintomatología tras la infección con la
bacteria. Todo ello, daba a entender que posiblemente se trataba de una
mutación relacionada con el establecimiento de los sistemas de defensa de la

35
planta. Con lo que se justificaría el interés del estudio presentado en este
trabajo.
4.2 Caracterización fenotípica del mutante csb3scs5
4.2.1. Características morfológicas
Este mutante revirtió el fenotipo morfológico de la mutación csb3 y además
mostró una característica morfológica particular como es la clorosis en la zona
meristemática de la roseta, a diferencia de la mutación scs9 que la desarrolla
por toda la zona foliar (Fig. 9). Esta característica de la mutación scs5 es
bastante singular y única, lo que podrá ser utilizado como un marcador
morfológico y permitirá la identificación de este mutante de una manera sencilla
y rápida en los futuros ensayos. El fenotipo de las plantas dado por la mutación
scs5 se caracterizó, además, en que las hojas eran globulares y con contorno
liso, y no aserrado como ocurre en las plantas csb3. Las plantas csb3scs5
también difirieron de csb3 en el tamaño de la planta, ya que revertía el fenotipo
enano de éste y se asemejaba más a las plantas silvestres (Col-0).
4.2.2 Actividad del transgen P69C::GUS
Además de analizar la supresión del fenotipo morfológico de csb3, se
caracterizó también la capacidad del mutante scs5 para suprimir la expresión
constitutiva del transgen que contiene (debida a la acción de csb3 sobre el
transgen P69C::GUS). Para ello se realizó una tinción histoquímica para
determinar la actividad GUS en hojas de roseta procedentes de plantas
P69C::GUS, csb3, csb3scs9 y csb3scs5 (Fig. 9). Ésta es una forma fácil y
rápida de evaluar los diferentes niveles de expresión de un gen determinado
(Jefferson, et al., 1987). Tras este ensayo se pudo observar que la mutación
scs5 consiguió reprimir la expresión constitutiva del transgen P69C::GUS que
muestra el mutante csb3, con respecto a línea parental. Sin embargo, esta
supresión no fue totalmente nula como sí ocurrió para la mutación scs9. Por lo
tanto, las plantas csb3scs5 fueron capaces de revertir estas dos características
fenotípicas ligadas a la mutación csb3.

36
Figura 9. Características fenotípicas de plantas de 4 semanas de Col-0, csb3, csb3scs9 y
csb3scs5. Panel superior: aspectos morfológicos. Panel inferior: tinción histoquímica de una
hoja representativa de cada fenotipo.
4.2.3 Respuesta a Pst. DC3000
Es importante recordar que además de la reversión de estas características
fenotípicas ligadas a csb3, las plantas csb3scs5 pertenecen al grupo de los scs
que presentaron mayores signos de infección a Pst. DC3000, indicando que
SCS5 es clave para el establecimiento de la resistencia a esta bacteria. Por lo
que se vio necesario comprobar y cuantificar la susceptibilidad de las plantas
csb3scs5 a Pst. DC3000, ya que una de las características más relevantes de
las plantas csb3 es la resistencia a patógenos biotrofos, debido a la mutación
del locus CSB3 (Gil. 2005). En un estudio previo por parte del laboratorio se vio
que plantas csb3 muestran un crecimiento de Pst DC3000 de 30 a 60 veces
menor en comparación con las plantas silvestres (Gil, et al., 2005). Para
corroborar lo anteriormente mencionado se analizó la curva de crecimiento
bacteriano en plantas de csb3 de 5 semanas inoculadas por spray con Pst.
DC3000 con una OD de 0.1. Como resultado se obtuvo que el crecimiento
bacteriano es aproximadamente 10 veces menor que las plantas silvestres
como se observa en la Fig. 10, confirmando así lo esperado y demostrando una
vez más la resistencia incrementada de las plantas csb3 a Pst. DC3000.
El resultado de la curva de crecimiento bacteriano realizada a los 3 y 5 días
después de la infección (dpi) en plantas de la mutación estudiada en este
trabajo, csb3scs5, el crecimiento de la bacteria es 20 veces más en
comparación de las plantas csb3 y 10 veces mayor que en plantas silvestres
(Fig. 10). Está es una manera de cuantificar la sintomatología observada en
estas plantas después de la infección. Ensayos anteriores se habían realizado
en el laboratorio con otro miembro del conjunto de scs (csb3scs9). Al realizar
la curva bacteriana en condiciones similares se observó el carácter de
hipersusceptible de las plantas csb3scs9 a Pst. DC3000. Al comparar los
resultados de susceptibilidad después de analizar la curva bacteriana en
plantas csb3scs5 con las plantas csb3scs9 se observa que estos dos mutantes
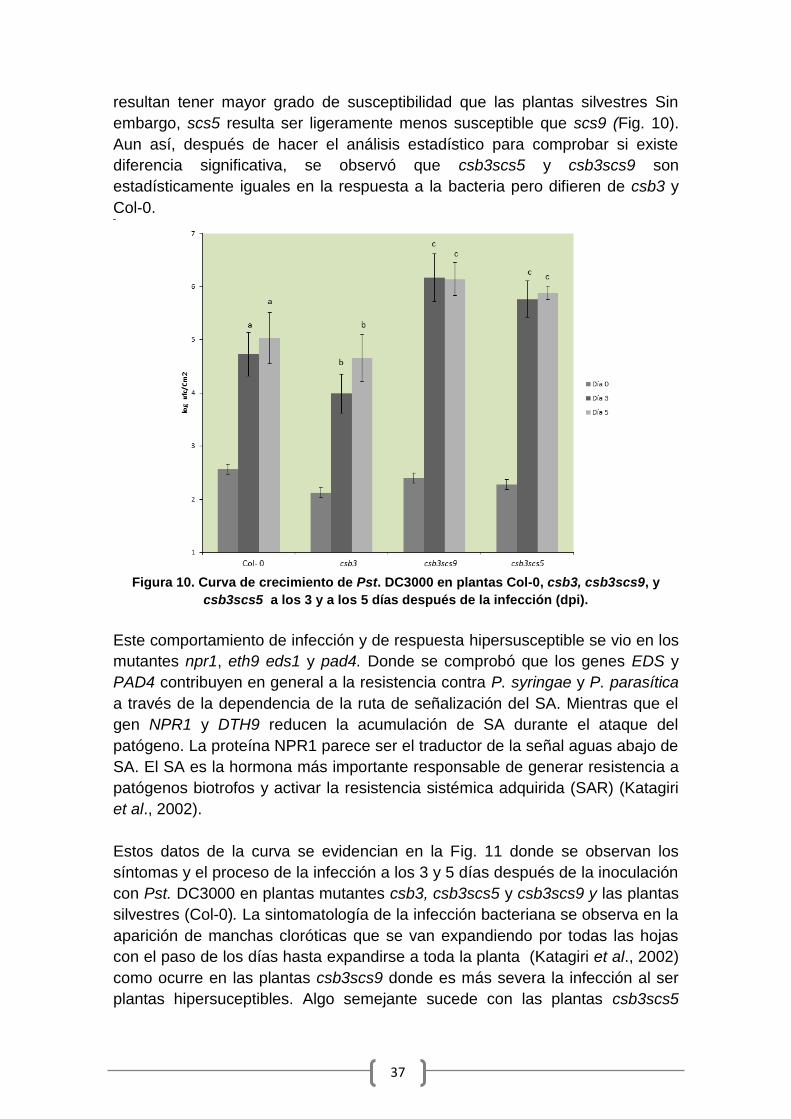
37
resultan tener mayor grado de susceptibilidad que las plantas silvestres Sin
embargo, scs5 resulta ser ligeramente menos susceptible que scs9 (Fig. 10).
Aun así, después de hacer el análisis estadístico para comprobar si existe
diferencia significativa, se observó que csb3scs5 y csb3scs9 son
estadísticamente iguales en la respuesta a la bacteria pero difieren de csb3 y
Col-0. Este co s
Figura 10. Curva de crecimiento de Pst. DC3000 en plantas Col-0, csb3, csb3scs9, y
csb3scs5 a los 3 y a los 5 días después de la infección (dpi).
Este comportamiento de infección y de respuesta hipersusceptible se vio en los
mutantes npr1, eth9 eds1 y pad4. Donde se comprobó que los genes EDS y
PAD4 contribuyen en general a la resistencia contra P. syringae y P. parasítica
a través de la dependencia de la ruta de señalización del SA. Mientras que el
gen NPR1 y DTH9 reducen la acumulación de SA durante el ataque del
patógeno. La proteína NPR1 parece ser el traductor de la señal aguas abajo de
SA. El SA es la hormona más importante responsable de generar resistencia a
patógenos biotrofos y activar la resistencia sistémica adquirida (SAR) (Katagiri
et al., 2002).
Estos datos de la curva se evidencian en la Fig. 11 donde se observan los
síntomas y el proceso de la infección a los 3 y 5 días después de la inoculación
con Pst. DC3000 en plantas mutantes csb3, csb3scs5 y csb3scs9 y las plantas
silvestres (Col-0). La sintomatología de la infección bacteriana se observa en la
aparición de manchas cloróticas que se van expandiendo por todas las hojas
con el paso de los días hasta expandirse a toda la planta (Katagiri et al., 2002)
como ocurre en las plantas csb3scs9 donde es más severa la infección al ser
plantas hipersuceptibles. Algo semejante sucede con las plantas csb3scs5
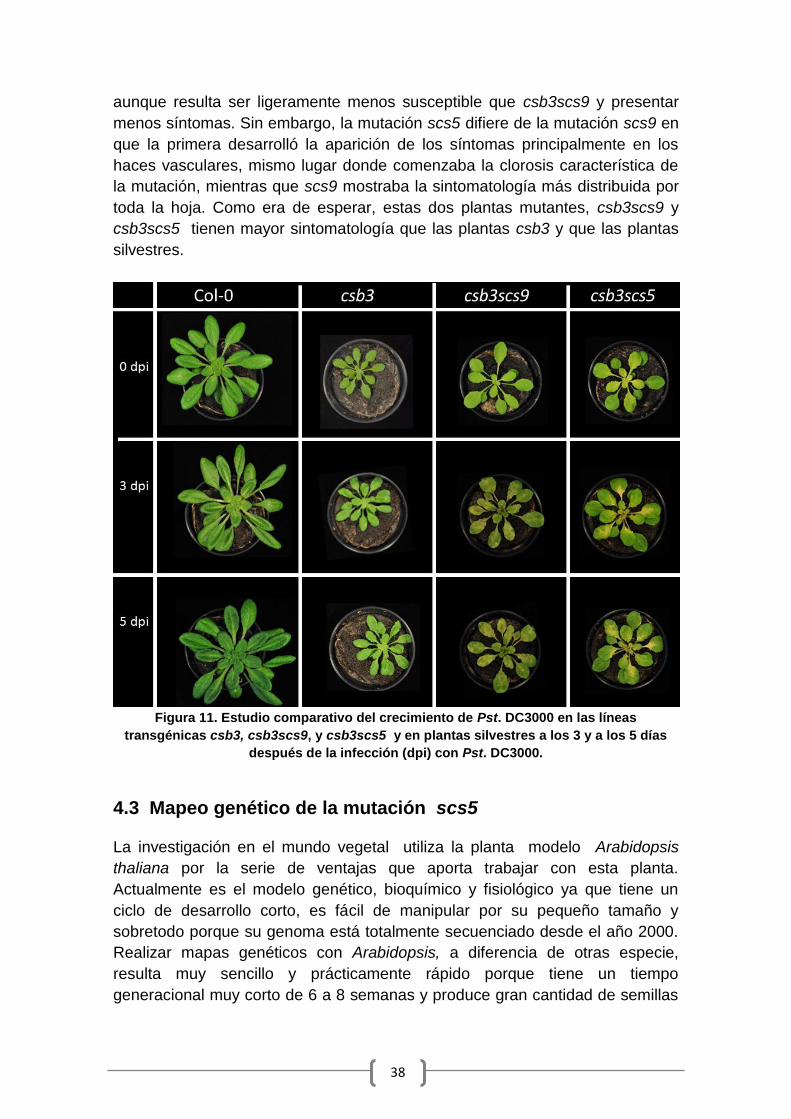
38
aunque resulta ser ligeramente menos susceptible que csb3scs9 y presentar
menos síntomas. Sin embargo, la mutación scs5 difiere de la mutación scs9 en
que la primera desarrolló la aparición de los síntomas principalmente en los
haces vasculares, mismo lugar donde comenzaba la clorosis característica de
la mutación, mientras que scs9 mostraba la sintomatología más distribuida por
toda la hoja. Como era de esperar, estas dos plantas mutantes, csb3scs9 y
csb3scs5 tienen mayor sintomatología que las plantas csb3 y que las plantas
silvestres.
Figura 11. Estudio comparativo del crecimiento de Pst. DC3000 en las líneas
transgénicas csb3, csb3scs9, y csb3scs5 y en plantas silvestres a los 3 y a los 5 días
después de la infección (dpi) con Pst. DC3000.
4.3 Mapeo genético de la mutación scs5
La investigación en el mundo vegetal utiliza la planta modelo Arabidopsis
thaliana por la serie de ventajas que aporta trabajar con esta planta.
Actualmente es el modelo genético, bioquímico y fisiológico ya que tiene un
ciclo de desarrollo corto, es fácil de manipular por su pequeño tamaño y
sobretodo porque su genoma está totalmente secuenciado desde el año 2000.
Realizar mapas genéticos con Arabidopsis, a diferencia de otras especie,
resulta muy sencillo y prácticamente rápido porque tiene un tiempo
generacional muy corto de 6 a 8 semanas y produce gran cantidad de semillas

39
viables, lo que permite tener grandes poblaciones en poco tiempo. En esta
especie se han desarrollado varias técnicas de mapeo, programas de
informátios y bases de datos completas y disponibles. Además, gracias a la
información que se ha obtenido acerca de esta planta se ha generado una gran
cantidad de líneas genéticamente definidas y un gran número de mutaciones.
Enfocándonos más en nuestro tema de estudio, en Arabidopsis se han descrito
la mayoría de mecanismos de respuesta frente a diferente tipos de estrés como
es el estrés ocasionado por agentes patogénicos (Jones y Takemoto, 2004).
Pero la mayor importancia de trabajar con Arabidopsis es que los resultados
se pueden aplicar en otras especies ya sean de interés comercial o agrícola.
(Glazebrook, et al, 1997).
4.3.1 Generación de una población recombinante
Una vez que se tiene una idea de la respuesta fenotípica por parte de la
mutación scs5 y la relación que tiene ésta en la defensa frente Pst. DC3000 se
prosiguió a realizar el mapeo genético para definir de forma aproximada la
posición cromosómica en la que se encuentra el gen SCS5. Para realizar el
mapeo genético era necesario primeramente obtener la población
recombinante mediante el cruzamiento entre las plantas csb3scs5 x Landsberg
erecta (Ler). Es importante recordar que las plantas csb3scs5 son el producto
de la mutagénesis de plantas csb3 y éstas a su vez son el producto de la
mutagénesis de las plantas transgénicas P69C::GUS generadas a partir de
plantas silvestres Col-0, Por lo tanto, las plantas scs5 tienen fijada la mutación
en el ecotipo Col-0. En este caso se utilizó el ecotipo Landsberg erecta (Ler)
por ser una línea diferente a la línea parental Col-0 y así asegurar que existan
regiones cromosómicas que tengan una combinación de alelos diferente al
parental. De esta manera se obtuvo una población F2 resultante de la
autofecundación de la F1. De la población F2 (200 plantas) fueron
seleccionadas las plantas recombinantes (37 en total) que mostraban el
fenotipo de la mutación scs5. Este número se ajusta a la proporción esperada
de ¼ de plantas con fenotipo mutante (scs5), en una F2 de 200 plantas para
un gen recesivo (Fig. 12). La selección de los recombinantes con fenotipo de la
mutación scs5 se realizó basándonos en las características singulares de estas
plantas. Como se explicó en apartados anteriores, las plantas scs5 tienen una
clorosis en la roseta particular que facilita su identificación.

40
Figura 12. Proceso de selección de plantas recombinantes procedentes de una F2 del
cruce de plantas csb3scs5xLer utilizado para el posterior mapeo de la mutación scs5. A
la izquierda está la representación esquemática del cruzamiento. En la parte derecha está una
imagen de un grupo de plantas F2 donde claramente se diferencia el fenotipo mutante scs5.
4.3.2 Extracción de ADN genómico
El ADN genómico de los ecotipos Col-0 y Ler y de las plantas recombinantes
seleccionadas fue extraído y cuantificado según el protocolo descrito en el
apartado 3.4.2. La concentración de cada muestra se encontró entre 100 a
300ng/µl. Esta concentración fue adecuada para proceder a realizar la PCR.
4.3.3 Análisis de las frecuencias de recombinación
Se analizaron por PCR los 25 marcadores SSLPs seleccionados en un primer
momento (descritos en 3.4.3) para todos los recombinantes identificados en la
población F2. Para asociar la posición de la mutación a alguno de estos
marcadores, se esperaba observar en el gel de agarosa que el mayor número
de recombinantes tuvieran el mismo tamaño de banda correspondiente al del
control Col-0, debido a que las plantas scs5 tienen fijada la mutación en este
ecotipo. Finalmente, tras el análisis de los 25 marcadores (Tabla 6), se observó
que el marcador MED24D del cromosoma 5 cosegrega con el fenotipo scs5,

41
debido al bajo número de eventos de recombinación observados. Este
resultado proviene del análisis de la frecuencia de recombinación donde se
cuantificó el número de recombinantes Col-0 (C), el número de recombinantes
Ler (L) y el número de recombinantes heterocigotos (H). El número total de
recombinantes analizados corresponde al total (T). Para el caso del marcador
MED24D tenemos 34 recombinantes Col-0, 1 recombinante Ler y 2
recombinantes heterocigotos, con un total de 37 individuos analizados (Fig. 13).
Al realizar el cálculo de la frecuencia de recombinación (tal y como se describe
en 3.4.6), se obtuvo que para este marcador era del 8% (Tabla 6). Por lo tanto,
según la posición del marcador MED24D, ésto nos indicaba que la mutación
scs5 estaba alrededor de la posición 1002257pb del cromosoma 5. Dado que el
porcentaje de recombinación se puede corresponder a la distancia entre el
marcador y la mutación. En este caso la mutación está más cerca del marcador
MED24D en el genoma ya que la probabilidad de recombinación es menor.
Figura 13. Gel de agarosa al 4% a partir de un PCR con el marcador molecular MED24D
para el mapeo genético de la mutación scs5
Ler

42
Tabla 6. Análisis de los marcadores SSLPs para el mapeo del mutante
csb3scs5. Número de recombinantes Col-0, Ler y heterocigotos y las
frecuencias de recombinación
CROMOSOMA
MARCADORES MOLECULARES
F21B7 F21M12 ATHS0392 CIW1 ATHATPASE
Col-0 14 8 13 9 7
Ler 8 13 7 16 13
1 HETEROCIGOTOS 15 16 17 12 17
TOTAL 37 37 37 37 37
FRECUENCIA DE
RECOMBINACION 62,16 78,38 64,86 75,68 81,08
MSAT2.26 MSAT2.18 MSAT2.36 NGA361 MSAT2.22
Col-0 10 9 8 11 9
Ler 15 8 9 9 11
2 HETEROCIGOTOS 12 20 20 17 17
TOTAL 37 37 37 37 37
FRECUENCIA DE
RECOMBINACION 72,97 75,68 78,38 70,27 75,67
NGA172 NT204 MSAT3.32 F24M2.TGF NGA6
Col-0 10 3 8 9 4
Ler 10 14 14 9 23
3 HETEROCIGOTOS 17 20 15 19 10
TOTAL 37 37 37 37 37
FRECUENCIA DE
RECOMBINACION 72,97 91,89 78,38 75,68 89,19
MSAT4.39 MSAT4.8 NGA8 MSAT4.15 MSAT4.31
Col-0 8 8 7 7 5
Ler 9 12 10 12 10
4 HETEROCIGOTOS 20 17 20 18 22
TOTAL 37 37 37 37 37
FRECUENCIA DE
RECOMBINACION 78,37 78,38 81,08 81,08 86,48
MED24D MSAT5.14 MSAT5,25 CIW9 MSAT5.19
Col-0 34 24 16 12 13
Ler 1 4 5 14 9
5 HETEROCIGOTOS 2 9 16 11 15
TOTAL 37 37 37 37 37
FRECUENCIA DE
RECOMBINACION 8,11 35,14 56,76 67,57 64,86

43
Con la finalidad de acotar más la localización de la mutación scs5 se
seleccionaron 3 nuevos marcadores SSLPs, que fueron CTR1.2, NGA249 Y
NGA106. Estos marcadores se encuentran en posiciones cercanas a la región
determinada por MED24D.
Tras el análisis realizado (Tabla 7), siguiendo la misma base que para los
anteriores marcadores, se observó que el primer marcador CTR1.2
determinaba un porcentaje de recombinación aproximado del 3%, el más bajo
de los 3 nuevos marcadores.
Tabla 7. Análisis de los nuevos marcadores SSLPs para el mapeo del
mutante csb3scs5. Número de recombinantes Col-0, Ler y heterocigotos y las
frecuencias de recombinación
CROMOSOMA MARCADORES MOLECULARES
5
CTR1.2 NGA249 NGA106
Col-0 29 27 22
Ler 0 0 2
HETEROCIGOTOS 1 3 6
TOTAL 30 30 30
FRECUENCIA DE
RECOMBINACION
3,33 10,00 26,67
Esto nos proporcionaba entonces una región grosera más próxima, que el
marcador MED24D, de la localización de la mutación scs5. Este resultado se
debe a que 29 individuos fueron recombinantes Col-0, ninguno Ler y un
recombinante en heterocigosis (Fig. 14). Por lo tanto, estos datos nos dieron
como resultado el porcentaje más bajo de recombinación y nos indican que el
gen SCS5 probablemente se encuentre más próximo a la posición 979764bp
en el cromosoma 5. Ya que la frecuencia de recombinación es la menor, tanto
el marcador con la mutación están más cerca en el genoma porque la
probabilidad de recombinación es menor cuanto menor distancia hay entre
ellos.
Figura 14. Gel de agarosa al 4% a partir de un PCR con el marcador molecular MED24D
para el mapeo genético de la mutación scs5

44
El mapa físico de los marcadores SSLPs utilizados para el mapeo de la
mutación scs5 queda definido como se indica en la Fig. 15. Este mapa nos
indica la posición relativa del gen SCS5 dado por el marcador CTR1.2 el cual
está a 979764bp en el cromosoma 5 de Arabidopsis.
Para acotar más la posición del locus responsable de la mutación scs5, sería
necesario aumentar la población de recombinantes a analizar en una F2 así
como el número de marcadores analizados en ella.
Figura 15. Mapa físico de los marcadores SSPLs utilizado para el mapeo de la mutación
scs5. A la izquierda de cada cromosoma se indica la posición del marcador en el genoma
(pares de bases) y a la derecha el nombre del marcador correspondiente. El marcador CTR1.2
está más próximo al gen SCS5.
Como discusión general de los resultados obtenidos en este trabajo decir que
la interacción planta patógeno es un tema que cada vez toma más fuerza y ha
generado interés en la comunidad científica, no solo con el objetivo de realizar
investigación y satisfacer la curiosidad científica sino a nivel económico. Al
entender cómo funcionan los mecanismos de defensa de los que disponen las
plantas se podrá mejorar mediantes técnicas biotecnológicas la calidad y la
producción de los cultivos y a su vez a la agricultura. A parte, se disminuirá el
uso de pesticidas y productos químicos en este sector, los cuales no solo
causan daños al medio ambiente sino a la salud de los seres humanos.
Enfocándonos en el ámbito de la investigación, que es el área que nos
corresponde, el entendimiento del complejo conjunto de rutas y mecanismos
que forman parte de la respuesta de defensa resulta un campo todavía incierto.

45
Por lo que resulta de gran interés conocer los genes y las moléculas implicadas
en estos mecanismos que sirvan de pistas para entender el modo de acción de
los mecanismos moleculares implicado en las rutas de señalización que
controla la activación de los sistemas de respuesta de defensa de las plantas.
Arabidopsis, como el resto de plantas se ve afectada por diferentes tipos de
estrés como es el estrés biótico (De Vos et al, 2005). Gracias a las facilidades
que brinda trabajar con esta planta modelo se podrá abordar estrategias
genéticas que nos permitan elucidar de una forma más rápida el
funcionamiento de la interacción planta-patógeno, para posteriormente
extrapolar estos resultados a otras especies.
La aproximación experimental realizada en este trabajo tiene como finalidad
identificar genes implicados en la regulación del gen CSB3 y por consiguiente
en la respuesta de defensa de las plantas. El abordaje genético que se aplicó
para este propósito consistió en el aislamiento de mutantes que reviertan el
fenotipo de plantas csb3 mediante la mutagénesis de semillas de esta línea con
EMS. Las plantas csb3 tienen también un fenotipo peculiar, ya que tienen un
tamaño reducido y sus hojas son aserradas. Este fenotipo refleja un gasto
energético enorme que realiza al mantener activa la ruta de señalización
defensiva mediada por SA frente a patógenos de vida biotrofos como es la
bacteria P. syringae (Gil, 2005).
Los criterios para la selección de los mutante scs se basaron en que debían
revertir el fenotipo morfológico de csb3 y suprimir la expresión constitutiva del
gen reportero GUS. Así es como se identificaron al conjunto de mutantes scs
de los cuales se escogió al mutante csb3scs5 para realizar este trabajo. Tras la
caracterización realizada, las plantas csb3scs5 tenían un marcador morfológico
importante, el mismo que las hace ideales para trabajar en esta clase de
investigaciones, el cual es una clorosis marcada en la zona meristemática de la
roseta, y al revertir el fenotipo de las plantas csb3 se puede observar que las
hojas son de forma oval semejantes a las de las plantas silvestres y que son de
mayor tamaño que las hojas de csb3. Como csb3scs5 es un supresor de csb3
se esperaría que fuera un regulador positivo de la defensa, al contrario de
csb3. Por ello, se cuantificaron los niveles de resistencia o susceptibilidad que
tenía csb3scs5 a la infección con Pst. DC3000. En estos ensayos se demostró
el carácter hipersusceptible de las plantas csb3scs5 y se confirmaron los
síntomas observados en plantas inoculadas a los 3 y 5 días después de ser
inoculadas. En estas plantas se ve como la infección se presentaba en forma
de manchas cloróticas en los haces vasculares que se expanden con el tiempo.
También se observó que la expresión del gen GUS en las plantas csb3scs5
estaba inactivo (Gil, 2005).
Futuros ensayos con este mutante permitirían aclarar de una mejor manera la
forma en la que actúa en los mecanismos de la planta contra agentes biotrofos,

46
ya que por el momento sólo se ha probado la susceptibilidad con la bacteria
Pst. DC3000 por lo que se podría tratar con otras especie como Hyalonospora
parasítica e inclusive se podría probar con patógenos de estilo de vida
necrotrofos como Botrytis cinerea o Alternaria brassicicola y así descartar una
relación con la ruta de señalización del JA. Además sería importante cuantificar
los niveles de SA o del conjugado glucosilado (SAG) ya que tenemos como
premisa que el SA es la hormona señalizadora de la defensa contra patógenos
biotrofos como es Pst. DC3000. Al encontrar alguna relación de la mutación
scs5 con el SA se debería profundizar y evaluar la expresión de genes
relacionados a la señalización de esta hormona como es el gen PR-1.
También, se podría probar ensayos en los que se infecte a las plantas scs5 con
Pst. DC3000 (avrRpm1) o analizar la respuesta a bacteria tras una aplicación
exógena con SA u otros análogos como BTH y determinar la inducción de
genes PR. Finalmente, una vez localizada una región cercana a la mutación
csb3scs5, esto cual servirá como primer paso para realizar un cartografiado y
clonaje posicional del gen SCS5 con la finalidad de definir la posición
cromosómica exacta del locus que define la mutación. Esto nos permitirá
conocer a que gen está relacionado y como está involucrado en la defensa de
las plantas y así entender de una mejor manera los mecanismos sofisticados y
complejos de la interacción planta patógeno.

47
5 CONCLUSIONES
- Se ha caracterizado a la mutación recesiva scs5 como un supresor
del fenotipo de resistencia asociado a la mutación csb3.
- El mutante scs5 parece tener un carácter pleitrópico ya que las
plantas portadoras de la mutación scs5 tienen un marcador
morfológico único el cual se caracteriza por tener clorosis en la zona
meristemática de la roseta, en el comienzo de los haces vasculares,
que será de gran utilidad para la identificación de la mutación en
futuros ensayos.
- Las plantas csb3scs5 se caracterizan por ser hipersusceptibles a la
infección por el patógenos biotrofo Pseudomonas Syringae DC3000
en comparación con plantas silvestres y plantas csb3. Ello demuestra
que esta mutación suprime al fenotipo de resistencia acusada de las
plantas csb3 frente a patógenos (hemi)biotrofos. Por ello, mientras
que CSB3 es un regulador negativo de la resistencia, SCS5
demuestra su papel como regulador positivo de la misma. A través
de la identificación de la mutación scs5 se aporta un nuevo regulador
para el mejor entendimiento de los componentes que se requieren
para la correcta activación de mecanismo de inmunidad en plantas.
- Mediante el mapeo posicional del locus SCS5, a través de la
utilización de marcadores moleculares SSLPs, se ha podido acotar la
posición cromosómica de dicho locus en proximidad con el marcador
CTR1.2 y con ello, concretamente, en proximidad con la posición
979764bp del cromosoma 5 de Arabidopsis cercana a la región
telomérica del brazo pequeño de dicho cromosoma. Dicha
localización servirá de punto de partida para el cartografiado de alta
resolución y la eventual identificación y clonación del gen SCS5 en
futuras aproximaciones.

48
6 BIBLIOGRAFIA
- Anderson, J., Badruzsaufari, E., Schenk, P., Manners J., Desmond, O.,
Ehlert C., Maclean, D., Eber, P. & Kazan, K. (2004). Antagonistic
Interaction between Abscisic Acid and Jasmonate-Ethylene Signaling
Pathways Modulates Defense Gene Expression and Disease Resistance
in Arabidopsis. The Plant Cell, 16: 3460–3479.
- Azofeifa-Delgado, A. (2006). Uso de marcadores moleculares en
plantas: aplicaciones en frutales del trópico. Agronomía Mesoamericana,
17(2): 221-242.
- Cao, H., Bowling, S., Gordon, A & Dong, X. (1997).Characterization of an
Arabidopsis Mutant That Is Nonresponsive to Inducers of Systemic
Acquired Resistance. Cell: 88, 57–63,
- Chen, Z., Zheng, Z., Huang, J., Lai, Z., & Fan, B. (2009). Biosynthesis of
salicylic acid in plants. Plant Signaling & Behavior, 4(6), 493–496.
- Chistiakov, D., Hellemans, B. & Volckaert, F. (2006). Microsatellites and
their genomic distribution, evolution, function and applications: A review
with special reference to fish genetics. Aquaculture, 255:1-4.
- Dangl, J.L., Dietrich, R.A., Richberg, M.H. (1996). Death Don‟t Have No
Mercy: Cell Death Programs in Plant-Microbe Interactions. Plant Cell, 8:
1793– 1807.
- De Vos, M., Van Oosten, v., Van Poecke, R., Van Pelt, J., Pozo, M.,
J. Mueller, M., Buchala, A., Métraux, J., Van Loon, J., Dicke, M.
& Pieterse, C. (2005). Signal Signature and Transcriptome Changes
of Arabidopsis During Pathogen and Insect Attack. Molecular Plant
Microbe Interactions (18) 9:923-937
- Dempsey, D. A., Vlot, A. C., Wildermuth, M. C., & Klessig, D. F. (2011).
Salicylic Acid Biosynthesis and Metabolism. The Arabidopsis Book /
American Society of Plant Biologists, 9, e0156. doi:10.1199/tab.0156
- Dodds, P.N. y Rathjen, J.P. (2010). Plant immunity: towards an
integrated view of plantpathogen interactions. Nature Reviews.
Gennetics, 11: 539-548
- Durrant, W. E., & Dong, X. (2004). Systemic Acquired Resistance.
Annual Review Phytopathology, 42: 185-209.

49
- Glazebrook, J.( 2001). Genes controlling expression of defense
responses in Arabidopsis status. Current Opinion in Plant Biology: 4,
301-308.
- Howe, G. (2004). Jasmonates as signals in the wound response. Journal
of Plant Growth Regulation 23(3):223-237.
- Hückelhoven, R. (2007). Cell Wall–Associated Mechanisms of Disease
Resistance and Susceptibility. Annual Review of Phytopathology, 45:
101–127.
- Jefferson, R., Kavanagh, T. & Bevan, M. (1987). GUS fusions: ß-
glucuronidase as a sensitive and versatile gene fusion marker in higher
plants. EMBO Journal, 6:3901-3907.
- Jordá, L. & Vera, P. (2000). Local and Systemic Induction of Two
Defense-Related Subtilisin-Like Protease Promoters in Transgenic
Arabidopsis Plants. Luciferin Induction of PR Gene Expression. Plant
Physiology,124;124-156.
- Jordá, L., Coego, A., & Vera, P. (1999). A genomic cluster containing
four differentially regulated subtilisin-like processing protease genes is in
tomato plants. Journal Biological Chemistry, 274: 2360–2365.
- Katagiri, F., Thilmony, R., & Yang, S. (2002). The Arabidopsis Thaliana-
Pseudomonas Syringae Interaction. Arabidopsis Book. 2002; 1: e0039.
- Kinkema, M., Fan, W., & Dong, X. (2000). Nuclear localization of NPR1
is required for activation of PR gene expression. Plant Cell, 12: 2339-
2350.
- Koornneef, A. & Pieterse, C. (2008). Cross Talk in Defense Signaling.
Plant Physiology, 146 (3) 839-844.
- Krouk, G., Ruffel, S., Gutiérrez, R., Gojon, A., Crawford, N., Coruzzi, G.,
Lacombe B. (2011). A framework integrating plant growth with hormones
and nutrients. Trends Plant Sci 16: 178–182
- Kucuktas, H., Wang, S.,Li, P., He, C., Xu, P., Sha Y., Liu, H., Jiang, H.,
Baoprasertkul, P., Somridhivej, B.,Wang, Y., Abernathy, J., Guo, X., Liu,
L., Muir, M.& Liu, Z. (2009). Construction of Genetic Linkage Maps and
Comparative Genome Analysis of Catfish Using Gene-Associated
Markers. Genetics, 181: 1649–1660.

50
- Kunkel, B., y Brooks, D. (2002). Cross talk between signaling pathways
in pathogen defense. Plant Biology , 5:325–331.
- León-Reyes, A. (2009). Making sense out of signaling during plant
defense. Ph.D. Thesis Utrecht University, ISBN 978-90-393-509739.
- Matarasso, N., Schuster, S., & Avni. A. (2005). A Novel Plant Cysteine
Protease Has a Dual Function as a Regulator of 1-Aminocyclopropane-1-
Carboxylic Acid Synthase Gene Expression. The Plant Cell, 17; 1205–
1216.
- Meichtry, J., Amrhein, N., Schaller, A. (1999). Characterization of the
subtilase gene family in tomato (Lycopersicon esculentum Mill.). Plant
Molecular Biology, 39: 749–760.
- Nandi, A., Krothapalli, K. Buseman, K., Li,. M., Welti, R., Enyedi, A.,
& Shah, J. (2003). Arabidopsis sfd Mutants Affect Plastidic Lipid
Composition and Suppress Dwarfing, Cell Death, and the Enhanced
Disease Resistance Phenotypes Resulting from the Deficiency of a Fatty
Acid Desaturase. The Plant Cell, 15(10): 2383-2398.
- Nurnberger, T. y Brunner, F. (2002). Innate immunity in plants and
animals: emerging parallels between the recognition of general elicitors
and pathogenassociated molecular patterns. Plant Biology, 5: 318-324.
- Pieterse, C., Leon-Reyes., A., Van der Ent., S. and Van Wees. (2009).
Networking by small-molecule hormones in plant immunity. Nature
Chemistry Biology, 5; 308-316.
- Ramírez V, López A, Mauch-Mani B, Gil MJ, Vera P. 2013. An
Extracellular Subtilase Switch for Immune Priming in Arabidopsis. PLoS
Pathogens, 9(6):e1003445. doi:10.1371/journal.ppat.1003445.
- Shah, J., Kachroo, P., Nandi, A., & Klessig, D. (2001). A recessive
mutation in the Arabidopsis SSI2 gene confers SA- and NPR1-
independent expression of PR genes and resistance against bacterial
and oomycete pathogens. Plant Journal, 25(5):563-74.
- Spoel, S., Koornneef, A., Claessens, S., Korzelius, J., Van Pelt, J.,
Mueller, M., Buchala, A., Métraux, J., Brown, R., Kazan, K., Van Loon,
L., Dong, X., Pieterse, C. (2003). NPR1 Modulates Cross-Talk between
Salicylate- and Jasmonate-Dependent Defense Pathways through a
Novel Function in the Cytosol.The Plant Cell, 15: 760–770.

51
- Sticher, L., Mauch-Mani, B., y Métraux, J.P. (1997). Systemic acquired
resistance. Annual Review Phytopathology, 35: 235-270.
- Stintzi, A., Heitz, T., Prasad, V., Wiedemann-Merdinoglu, S., Kauffmann,
S., Geoffroy, P., Legry, M., & Fritig, B. (1993). Plant 'pathogenesis-
related' proteins and their role in defense against pathogens.
Biochimiestry,75: 687-706.
- Tornero, P., Conejero, V., & Vera, P. (1996). Primary structure and
expression of a pathogen-induced protease (P69) in tomato plants:
Similarity of functional domains to subtilisin-like endoproteases.
Proceedings of the National Academy of Sciences, 93: 6332–6337.
- Van Der Biezen, E.A. y Jones, J.D.G., (1998). Plant disease-resistance
proteins and the gene-for-gene concept. Trends in Biochemical
Sciences, 23: 454–456.
- Van der Ent, S., Koornneef, A. & Pieterse, J. (2009). Induced
Resistance-Orchestrating defence mecanisms through rosstalk and
priming. Annual Plant Reviews 34: 334-370.
- Van Hulten, M. (2009). Costs and benefits of priming for defense in
Arabidopsis. Proceedings of the National Academy of Science of The
United Stated of America, 103: 5602-5607.
- Van Lon, L.C. (1997). Induced resitance in plants and the role of
pathogenesi-related proteins. European Journal. Plant Pathology, 103:
753-765.
- Van Oosten, V., Bodenhausen, N., Reymond, F., Van Pelt, J., Van Loon,
L., Dicke, M., y Pieterse, C. (2008). Differential Effectiveness of
Microbially Induced Resistance Against Herbivorous Insects in
Arabidopsis. The American Phytopathological Society, 21(7): 919–930.
- Verhage, A.; van Wees, S. C. M.; Pieterse, C. M. J. (2010). Plant
Immunity: It’s the Hormones Talking, But What Do They Say? Plant
Physiology 154(2):536-540.
- Wu, Y., Zhang, D., Chu, J., Boyle, P., Wang, Y., Brindle, I., De Luca, V.,
& Després, C. (2012). The Arabidopsis NPR1 protein is a receptor for the
plant defense hormone salicylic acid. Cell Reports 1(6):639-47.
- Xiao, Y., Savchenko,T., Baidoo, E.,Chehab, W., Hayden, D., Tolstikov,
V., Corwin, J. Kliebenstein, D., Keasling, J., & Dehesh, K. (2012).

52
Retrograde Signaling by the Plastidial Metabolite MEcPP Regulates
Expression of Nuclear Stress-Response Genes.Cell,149;1525’1535 doi
10.1016/j.cell.2012.04.038
- Zhao, C., Johnson, B.J., Kositsup, B., y Beers, E.P. (2000). Exploiting
secondary growth in Arabidopsis. Construction of xylem and bark cDNA
libraries and cloning of three xylem endopeptidases. Plant Physiology.
123, 1185-1196


















