Celula musculo - virus - bacterias
56
BIOLOGIA CELULAR Y MOLECULAR FACULTAD DE SALUD II SEMESTRE
-
Upload
liliana-gonzalez -
Category
Documents
-
view
25 -
download
2
Transcript of Celula musculo - virus - bacterias

BIOLOGIA CELULAR Y MOLECULAR
FACULTAD DE SALUDII SEMESTRE

LA CELULA
LIC. LILIANA GONZALEZ B.
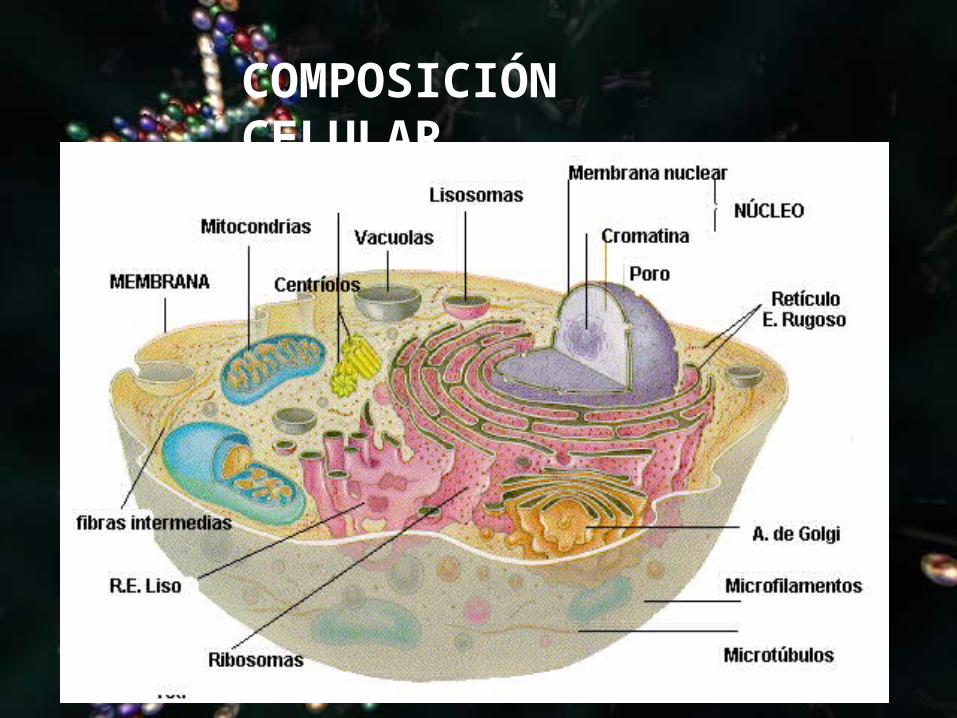
COMPOSICIÓN CELULAR

ORGANELAS CELULARES
• El citoplasma en una célula eucariota, se encuentra entre el núcleo celular y la membrana plasmática.
• Consiste en una emulsión coloidal muy fina de aspecto
granuloso, el citosol o hialoplasma, y en una diversidad de orgánulos celulares que desempeñan diferentes funciones.
• Su función es albergar los orgánulos celulares y contribuir al movimiento de los mismos. Este es la sede de muchos de los procesos metabólicos que se dan en las células.
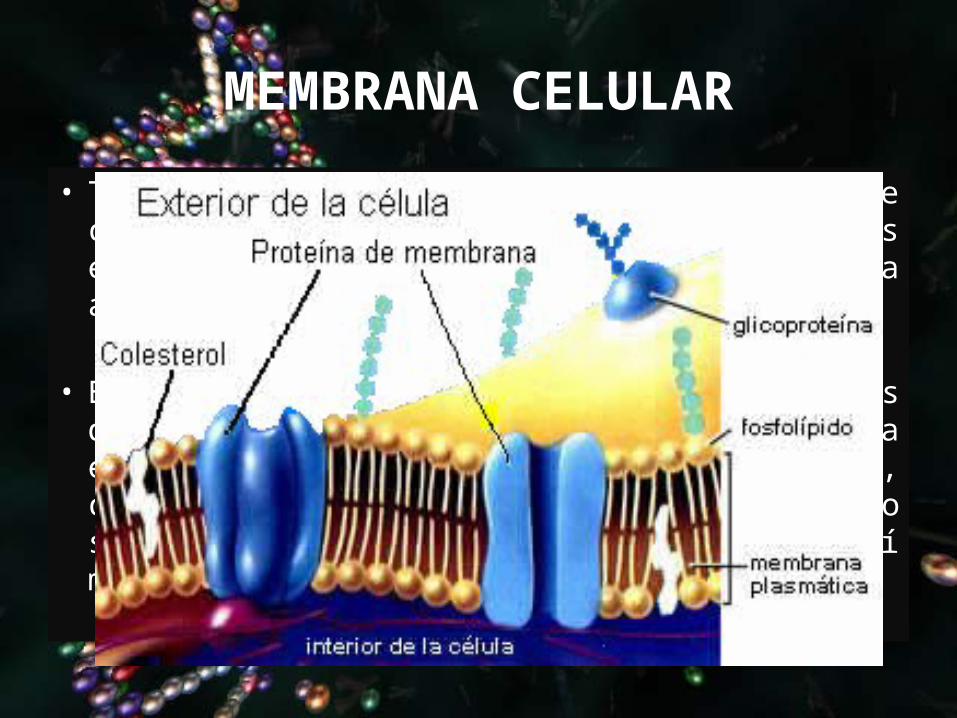
MEMBRANA CELULAR
• Tienen una estructura formada por una doble capa de lípidos que contiene proteínas especializadas, asociadas a su vez a azúcares de superficie.
• Esta estructura trilaminar (dos líneas densas delgadas -capa interna y capa externa- y una zona más clara entre ellas), conocida como modelo del mosaico fluido no se ve con el microscopio óptico pero sí mediante el microscopio electrónico.

Las principales funciones de la membrana plasmática de la célula son:
• Confiere a la célula su individualidad, al separarla de su entorno
• Constituye una barrera con permeabilidad muy selectiva, controlando el intercambio de sustancias
• Controla el flujo de información entre las células y su entorno
• Proporciona el medio apropiado para el funcionamiento de las proteínas de membrana
Lípidos
Los principales lípidos son:
Cada tipo de lípido de membrana posee una cabeza polar superficial (hidrofílica) y dos cadenas de ácidos grasos orientadas hacia el interior de la membrana (hidrofóbicas), por lo que se dice que esa molécula es anfipática.
FOSFOLÍPIDOS: representan en torno al 50% del componente lipídico. Las débiles fuerzas que unen entre sí a la bicapa permiten a las moléculas de fosfolípidos moverse con cierta libertad en el seno de cada capa, lo que confiere una gran movilidad a la membrana.
COLESTEROL: hace que la membrana sea menos fluida, pero mecánicamente más estable, La mayoría de las células bacterianas no contiene colesterol. Tampoco las células vegetales.
GLUCOLÍPIDOS: sólo se encuentran en la cara externa de la membrana celular, con los azúcares expuestos hacia el espacio extracelular.

Proteínas
Las proteínas pueden formar parte de esa bicapa en forma de proteínas integrales, que atraviesan todo su espesor, o en forma de proteínas periféricas, unidas a la superficie citoplasmática de la bicapa. Algunas de las intrínsecas atraviesan todo el espesor de la membrana (proteínas transmembranosas) y quedan expuestas en las dos superficies; otras proteínas no están fijas y "flotan" en el espesor de la membrana, como icebergs en un mar de lípidos. Aunque los diferentes tipos de proteínas que pueden encontrarse dependen del tipo celular de que se trate.
Funciones
•Fijan los filamentos del citoesqueleto a la membrana celular
•Fijan las células a la matriz extracelular
•Forman canales iónicos que facilitan el paso de iones y moléculas específicas a través de la membrana
•Actúan como receptores en los procesos de comunicación entre células
•Poseen actividades enzimáticas específicas
•Reconocen, por medio de receptores, a antígenos y células extrañas

AZUCARES
Se encuentran en su mayor parte limitados a la superficie de la membrana celular, formando el glucocáliz. Se puede poner en evidencia mediante microscopio electrónico, en forma de una capa blanca por fuera de la membrana celular, formada por azúcares unidos a las proteínas de esa membrana, a los fosfolípidos de la cara externa, o a ambos.
FUNCIONES•Proteger la superficie celular contra la interacción de otras proteínas extrañas o lesiones físicas o químicas•Papel en el reconocimiento celular, y en los procesos de rechazos de injertos y transplantes•Participa en los procesos de coagulación de la sangre y en las reacciones inflamatorias, entre otras.•Fecundación: los espermatozoides distinguen los óvulos de la propia especie de los de especies diferentes
TRANSPORTE PASIVO
TRANSPORTE ACTIVO
TRANSPORTE DE MACROMOLECULAS Y PARTICULAS
TRANSPORTE CELULA
R
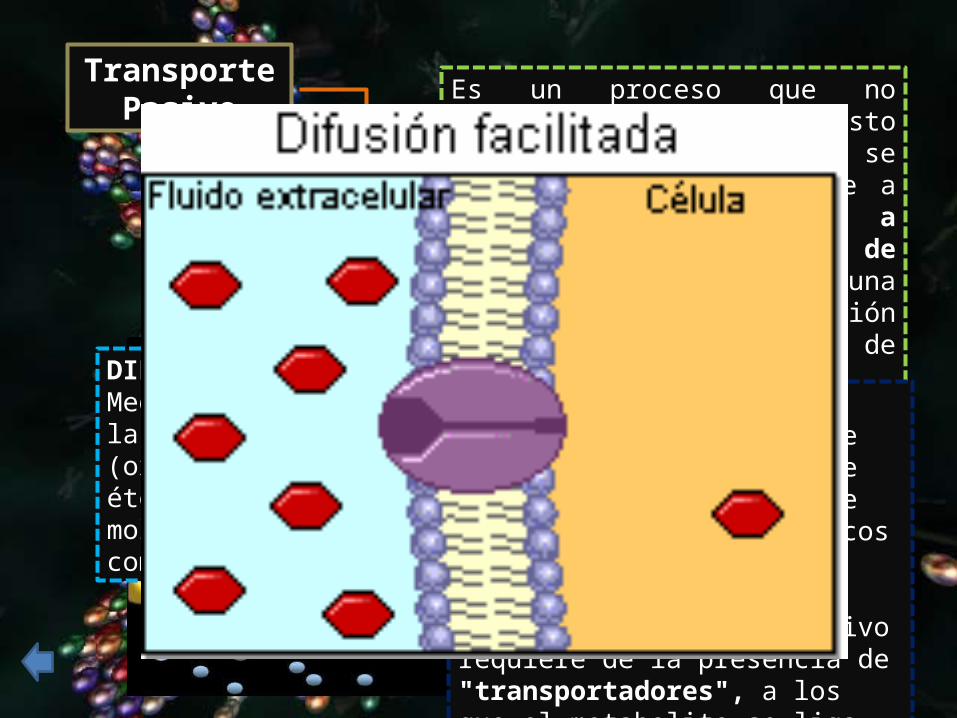
Transporte Pasivo
Es un proceso que no requiere energía, puesto que las moléculas se desplazan espontáneamente a través de la membrana a favor de gradiente de concentración (desde una zona de concentración elevada a otra de concentración más baja)
DIFUSIÓN SIMPLE O PASIVA:Mediante este sistema pasan las moléculas no polares (oxígeno, nitrógeno, éter...), y algunas moléculas polares pequeñas, como urea, agua y CO2.
DIFUSIÓN FACILITADA: también es dependiente de la concentración, y tiene que ver con el transporte de metabolitos hidrofílicos de mayor tamaño, como glucosa y aminoácidos. Aunque es un proceso pasivo requiere de la presencia de "transportadores", a los que el metabolito se liga, como son las proteínas transportadoras específicas.
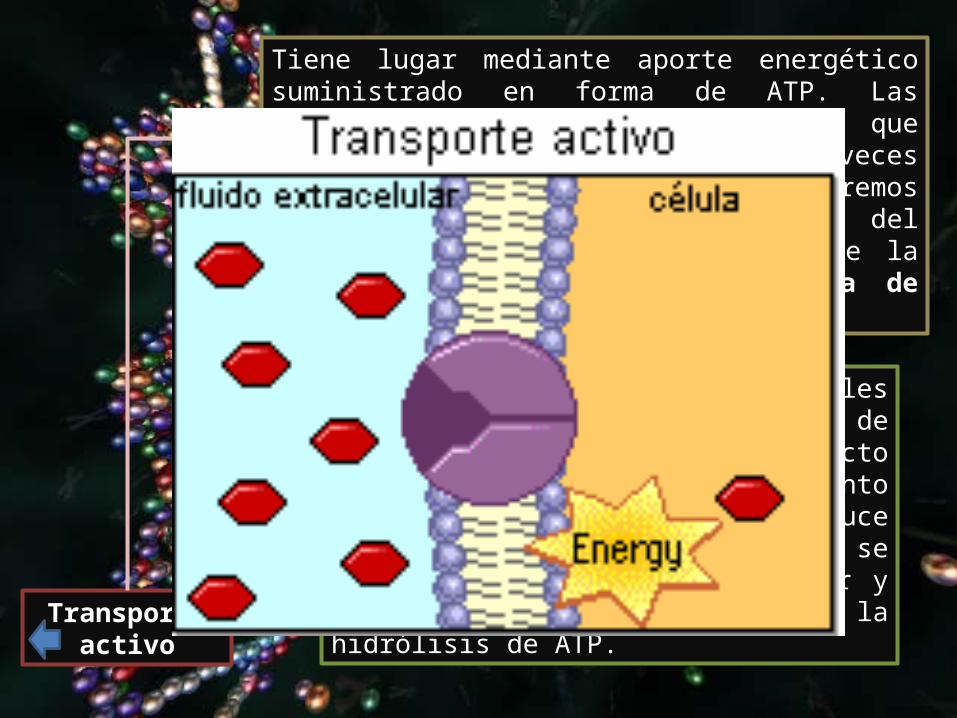
Transporte activo
Tiene lugar mediante aporte energético suministrado en forma de ATP. Las proteínas transportadoras que intervienen se llaman "bombas". A veces ocurre en contra de gradientes extremos de concentración, como es el caso del transporte de sodio hacia fuera de la célula mediante la llamada "bomba de Na+-K+“.
La mayoría de las células animales tienen altas concentraciones de potasio y bajas de sodio respecto a su medio externo; el movimiento de ambos iones se produce simultáneamente, de manera que se bombean 3 Na+ hacia el exterior y 2 K+ hacia el interior, con la hidrólisis de ATP.
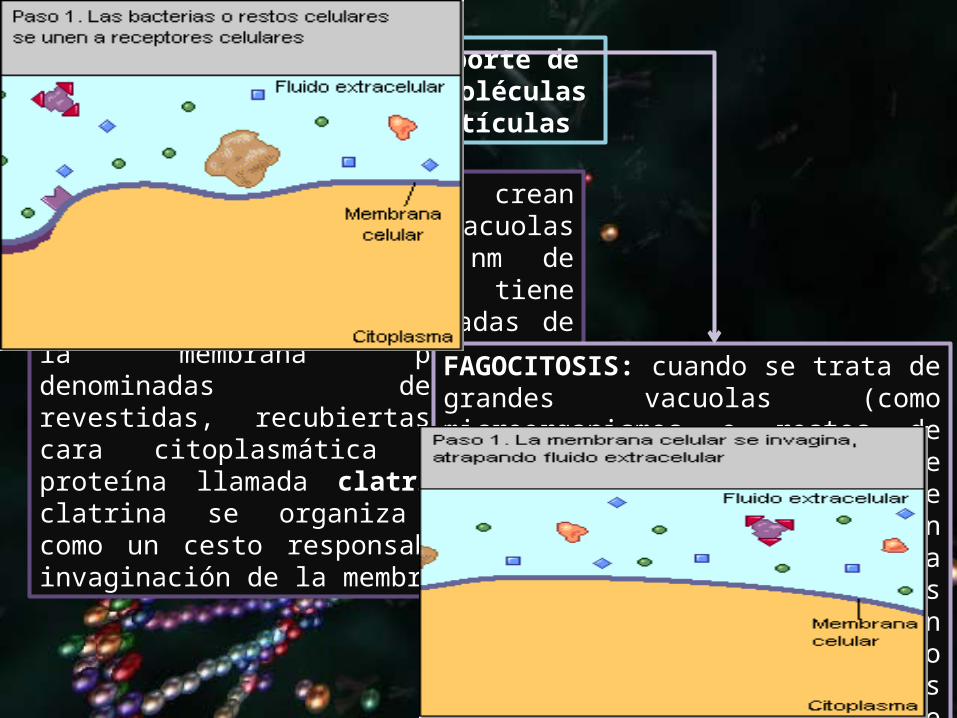
Transporte de macromoléculas y
partículas
Las grandes moléculas o pequeñas partículas que quedan atrapadas por la membrana plasmática, forman vacuolas o endosomas (vesículas) limitadas por membranas. El proceso por el que se ingieren moléculas se este tipo se llama endocitosis, y el proceso por el que se segregan se llama exocitosis.
Endocitosis
PINOCITOSIS: cuando se crean pequeñas vacuolas (vacuolas líquidas de unos 150 nm de diámetro). La pinocitosis tiene lugar en zonas especializadas de la membrana plasmática denominadas depresiones revestidas, recubiertas en su cara citoplasmática por una proteína llamada clatrina. Esta clatrina se organiza formando como un cesto responsable de la invaginación de la membrana.
FAGOCITOSIS: cuando se trata de grandes vacuolas (como microorganismos o restos de células, endosomas de más de 250 nm de diámetro). Para que ocurra la fagocitosis deben existir en la superficie de la célula receptores específicos para las sustancias que se van a englobar. De ese modo, cuando una partícula se une a los receptores de la superficie de la célula, ésta emite pseudópodos que engloban a esa partícula formando un fagosoma.

Exocitosis• Se trata del proceso inverso y ocurre en
dirección opuesta, de manera que una vesícula revestida de membrana se fusiona con la membrana plasmática para descargar su contenido al espacio extracelular. Las moléculas segregadas se adhieren a la superficie celular y pasan a formar parte del glucocáliz o bien se incorporan directamente a la matriz extracelular.

NUCLEO
El nucleo es la porción del citoplasma rodeada de la membrana nuclear, constituido por gránulos de cromatina, partículas de ribonucleoproteínas y matriz nuclear. Desde el punto de vista bioquímico, esta última contiene en torno al 10% de proteínas totales, 30% del RNA, 1-3% del DNA y 2-5% de todo el fosfato nuclear total.
La membrana nuclear está formada por dos cubiertas: la membrana nuclear interna y externa, separadas entre sí por un espacio (cisterna perinuclear). La cubierta externa está orientada hacia el citoplasma y se continúa con el RER, y su superficie está recubierta por ribosomas que sintetizan proteínas que irán a formar parte de esas membranas nucleares.
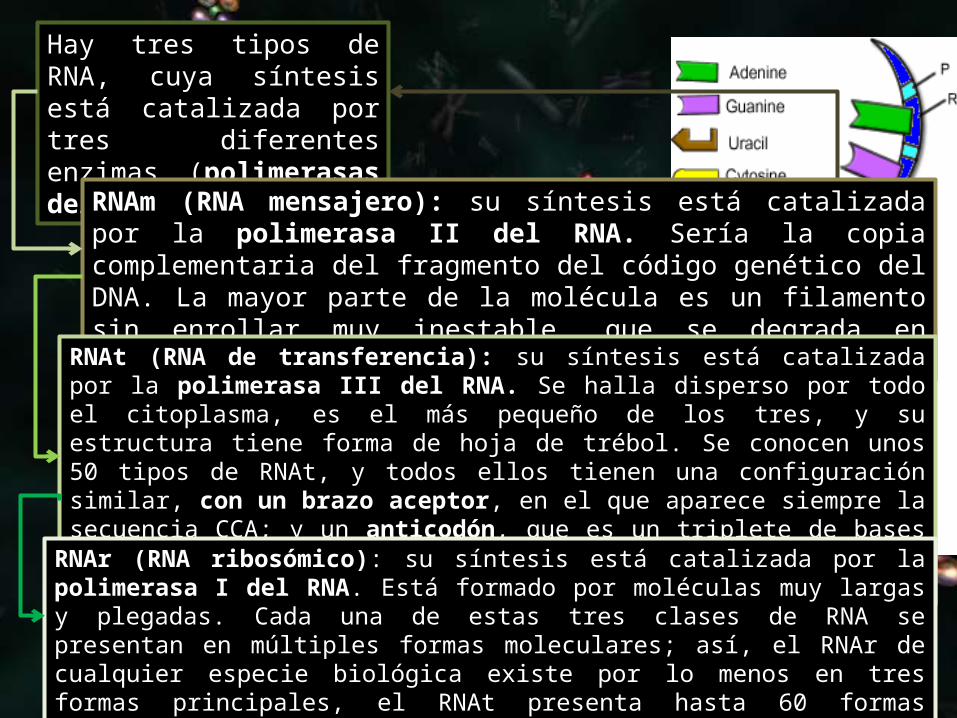
RNA
El ácido ribonucleico (RNA) es similar al DNA porque también está compuesto por una secuencia lineal de nucleótidos. Sin embargo, tiene una sola cadena y el azúcar es la ribosa en lugar de la desoxirribosa. Además, la timina está sustituida por uracilo, que también es complementario de la adenina. La mayor parte de las células contienen de 2 a 8 veces más de RNA que de DNA.
Hay tres tipos de RNA, cuya síntesis está catalizada por tres diferentes enzimas (polimerasas del RNA):
RNAm (RNA mensajero): su síntesis está catalizada por la polimerasa II del RNA. Sería la copia complementaria del fragmento del código genético del DNA. La mayor parte de la molécula es un filamento sin enrollar muy inestable, que se degrada en cuestión de horas.
RNAt (RNA de transferencia): su síntesis está catalizada por la polimerasa III del RNA. Se halla disperso por todo el citoplasma, es el más pequeño de los tres, y su estructura tiene forma de hoja de trébol. Se conocen unos 50 tipos de RNAt, y todos ellos tienen una configuración similar, con un brazo aceptor, en el que aparece siempre la secuencia CCA; y un anticodón, que es un triplete de bases nitrogenadas que determina el aminoácido que se va a unir a esa molécula de RNAt.
RNAr (RNA ribosómico): su síntesis está catalizada por la polimerasa I del RNA. Está formado por moléculas muy largas y plegadas. Cada una de estas tres clases de RNA se presentan en múltiples formas moleculares; así, el RNAr de cualquier especie biológica existe por lo menos en tres formas principales, el RNAt presenta hasta 60 formas diferentes, y el RNAm aparece en centenares e incluso millares de formas diferentes.
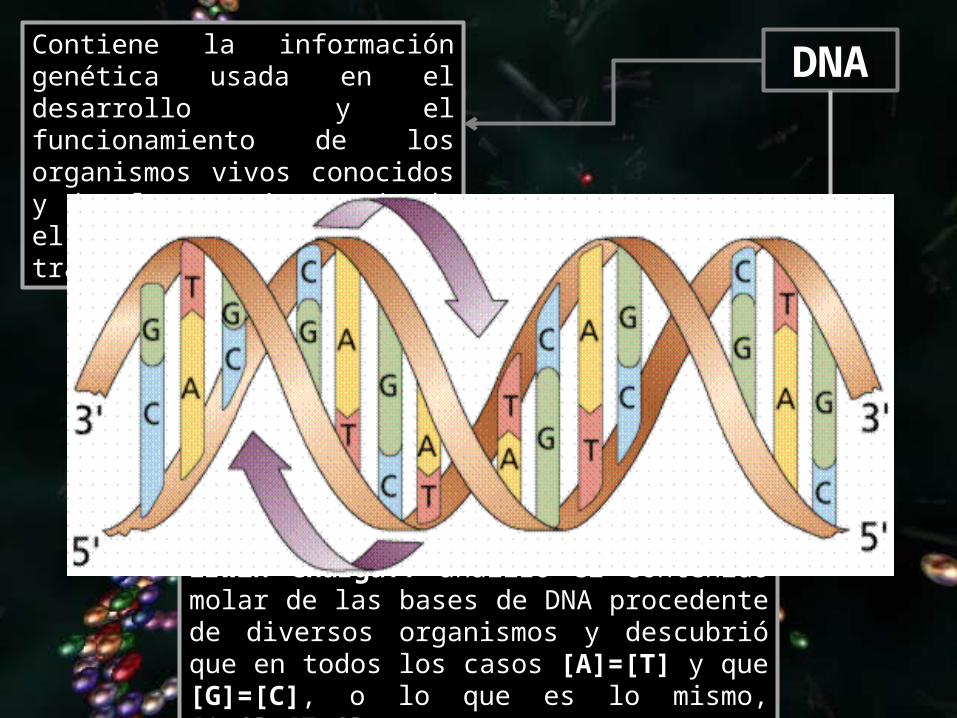
DNAContiene la información genética usada en el desarrollo y el funcionamiento de los organismos vivos conocidos y de algunos virus, siendo el responsable de su transmisión hereditaria.
LA ESTRUCTURA DEL DNAEn los años 20, el bioquímico Phoebus Levene determinó que el DNA estaba formado por 4 tipos distintos de nucleótidos. Cada nucleótido estaba formado por desoxirribosa, fosfato y una base nitrogenada (A, C, T o G). En 1949, el bioquímico Erwin Chargaff analizó el contenido molar de las bases de DNA procedente de diversos organismos y descubrió que en todos los casos [A]=[T] y que [G]=[C], o lo que es lo mismo, [A+G]=[T+C] ([purinas]=[pirimidinas]). Esta es la llamada ley de Chargaff.

MITOCONDRIA
Energía Las mitocondrias constituyen la fuente energética de las células, ya que mediante el proceso de fosforilación oxidativa producen trifosfato de adenosina (ATP), que es la forma estable de almacenamiento de energía que puede utilizar la célula para llevar a cabo las actividades que la requieren. Se trata de organelas flexibles cuya morfología varía de unas células a otras: en las que tienen un elevado nivel de metabolismo oxidativo suelen ser grandes y serpenteantes, en forma de bastoncillo; en otras tienen un aspecto más redondeado. Una célula eucariótica típica puede contener del orden de unas 2000 mitocondrias, ocupando en torno a un 20% de todo el volúmen celular.

Reticulo endoplasmatico• El Retículo Endoplásmico consiste en una red de
túbulos, vesículas y sacos interconectados, que se extienden por todo el citoplasma y que llevan a cabo diversas funciones celulares, incluida la síntesis proteica, la producción de esteroides, almacenamiento de glucógeno, etc.

Retículo Endoplásmico Rugoso (RER) Toma su nombre de la presencia de ribosomas sobre sus membranas. Junto al aparato de Golgi conforman dos regiones diferenciadas de un mismo compartimento membranoso intercomunicado, que participa en la biosíntesis y transporte de proteínas y lípidos celulares. Su membrana se continúa con la membrana externa de la envoltura nuclear.

Son pequeñas partículas compuestas por proteínas ribosomales (sintetizadas en el citosol) y RNA ribosomal (RNAr, sintetizado en el nucléolo), que funcionan como superficie para la síntesis de proteínas. Cada ribosoma consta de una subunidad grande y otra pequeña, que se elaboran en el nucléolo y se vierten como entidades separadas hacia el citosol, no formando un ribosoma como tal hasta que no se inicie la síntesis de proteínas. La subunidad pequeña tiene un valor de sedimentación de 40S; la subunidad grande tiene un valor de sedimentación de 60S
RIBOSOMAS

Aparato de golgi El aparato de Golgi es una organela que interviene tanto en la síntesis de azúcares (especialmente polisacáridos) como en la ordenación de las proteínas elaboradas en el retículo endoplásmico rugoso Su desarrollo varía según el tipo de célula y su estado fisiológico, al igual que el RER, está muy desarrollado en aquellas células especializadas en procesos de secreción.
Está formado por una o más series de cisternas ligeramente curvas y aplanadas limitadas por membranas, y a este conjunto se conoce como apilamiento de Golgi o dictiosoma. Los extremos de cada cisterna están dilatados y rodeados de vesículas que o se fusionan con este comportamiento, o se separan del mismo mediante gemación. Cada pila tiene una región más cercana al RER que es la cara cis (convexa), orientada hacia el núcleo, y la superficie opuesta, la cara trans (cóncava). Entre ambas caras se encuentran varios compartimentos mediales.
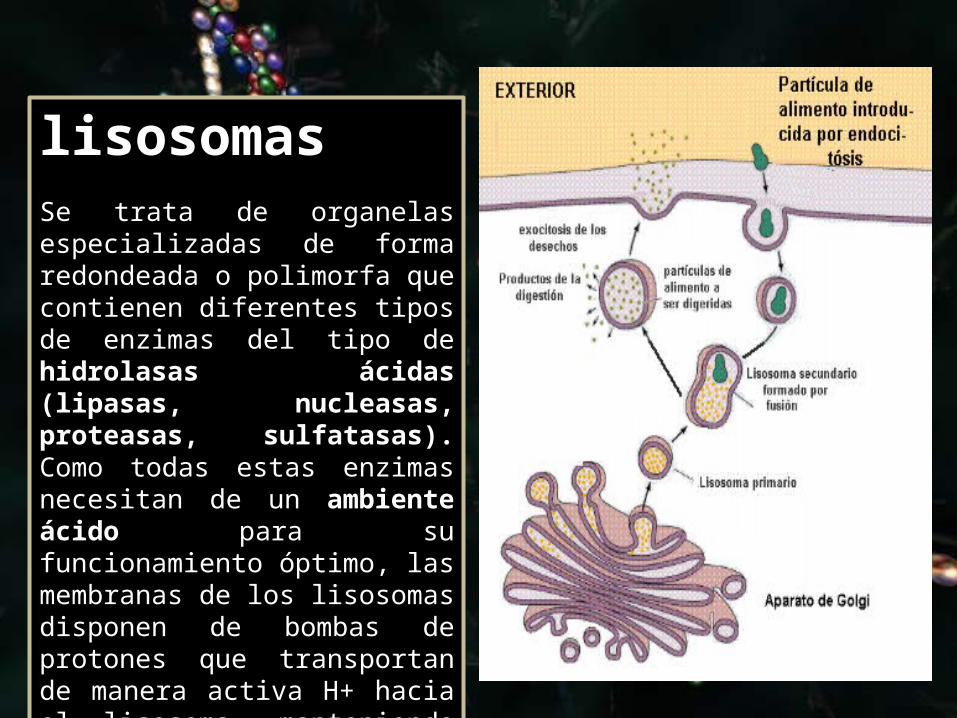
lisosomas Se trata de organelas especializadas de forma redondeada o polimorfa que contienen diferentes tipos de enzimas del tipo de hidrolasas ácidas (lipasas, nucleasas, proteasas, sulfatasas). Como todas estas enzimas necesitan de un ambiente ácido para su funcionamiento óptimo, las membranas de los lisosomas disponen de bombas de protones que transportan de manera activa H+ hacia el lisosoma, manteniendo así un pH de 5.

Vacuolas Los peroxisomas o microcuerpos son organelas pequeñas y esféricas, limitadas por membranas, muy parecidos a los lisosomas, aunque se distinguen de éstos porque disponen de contenidos enzimáticos muy diferentes: en concreto oxidasas (productoras de peróxido de hidrógeno) y catalasas (que lo eliminan).
Las principales funciones son: - llevan a cabo reacciones oxidativas de degradación de ácidos grasos y aminoácidos - intervienen en reacciones de detoxificación (por ejemplo, gran parte del etanol que bebemos es detoxificado por peroxisomas de células hepáticas)

Citoesqueleto Consiste en una malla tridimensional de filamentos proteicos cuyas principales funciones son: •Proporcionar el soporte estructural para la membrana plasmática y los orgánulos celulares•Proporcionar el medio para el movimiento intracelular de organelas y otros componentes del citosol•Proporcionar el soporte para las estructuras celulares móviles especializadas, como cilios y flagelos, responsables de la propiedad contráctil de las células en tejidos especializados como el músculo
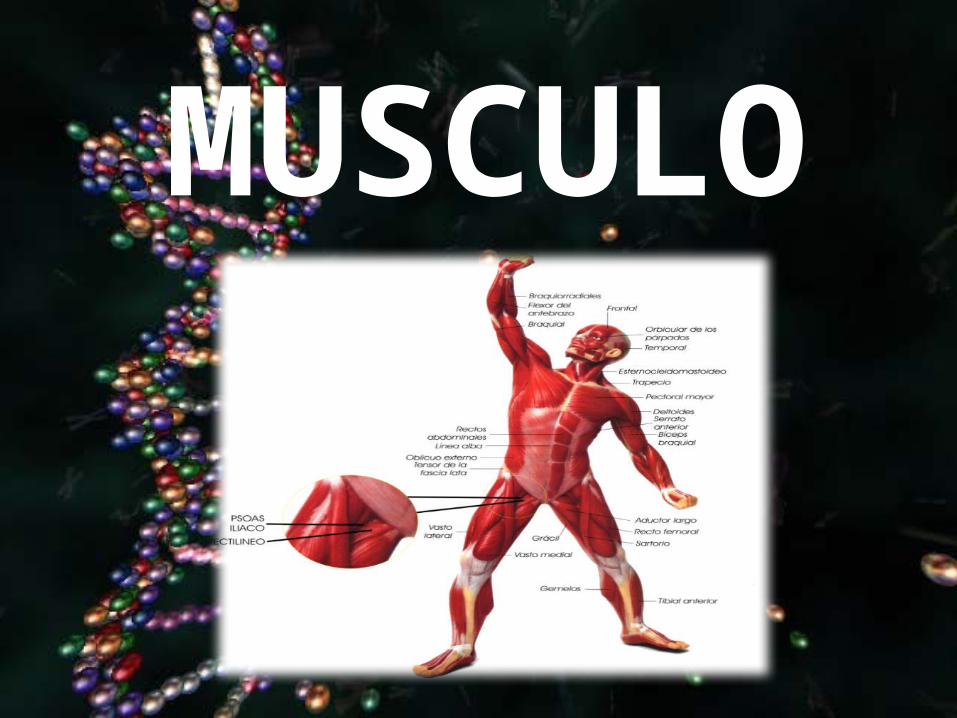
MUSCULO
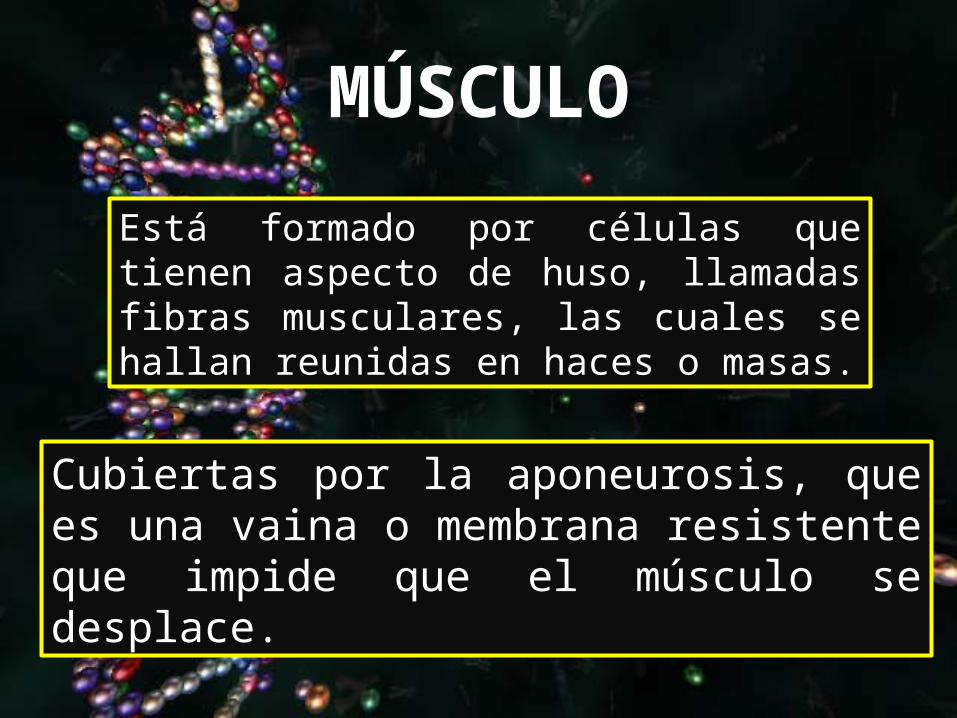
MÚSCULO
Cubiertas por la aponeurosis, que es una vaina o membrana resistente que impide que el músculo se desplace.
Está formado por células que tienen aspecto de huso, llamadas fibras musculares, las cuales se hallan reunidas en haces o masas.

• Se logra mediante la contracción de células musculares, que hacen trabajo mecánico al contraerse, en cuyo acto se acortan y ensanchan. Están formadas por las proteínas miosina y actina.
MOVIMIENTO MUSCULAR
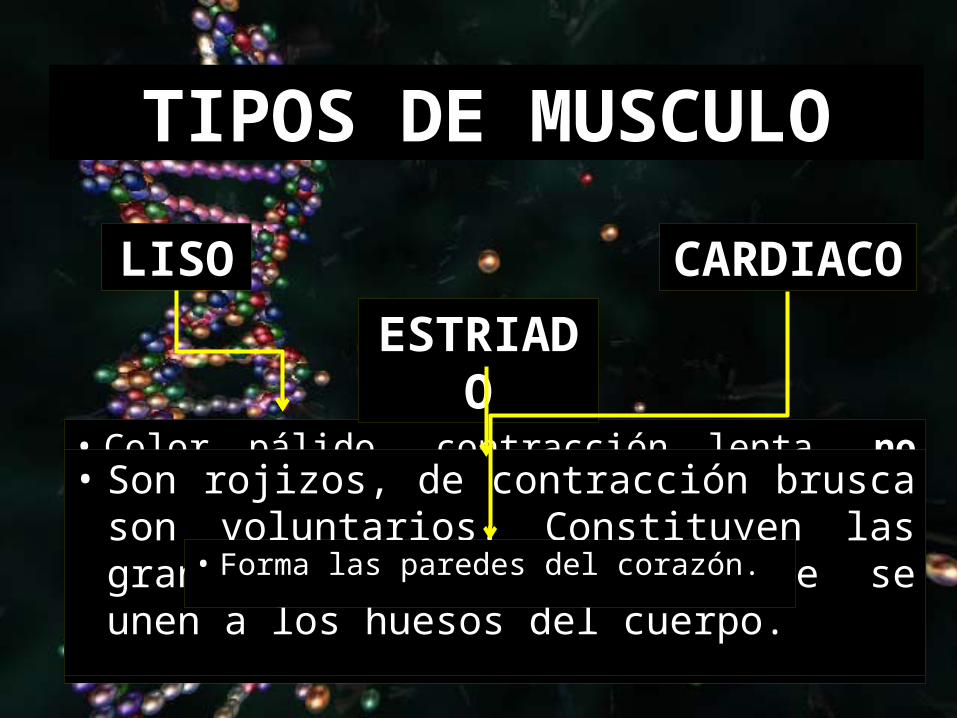
• Color pálido, contracción lenta, no son voluntarios. Se encuentran en las paredes del tubo digestivo, en las capas medias de las paredes de los vasos arteriales y demás órganos internos.
TIPOS DE MUSCULO
LISO
ESTRIADO
CARDIACO
• Son rojizos, de contracción brusca son voluntarios. Constituyen las grandes masa musculares que se unen a los huesos del cuerpo.
• Forma las paredes del corazón.
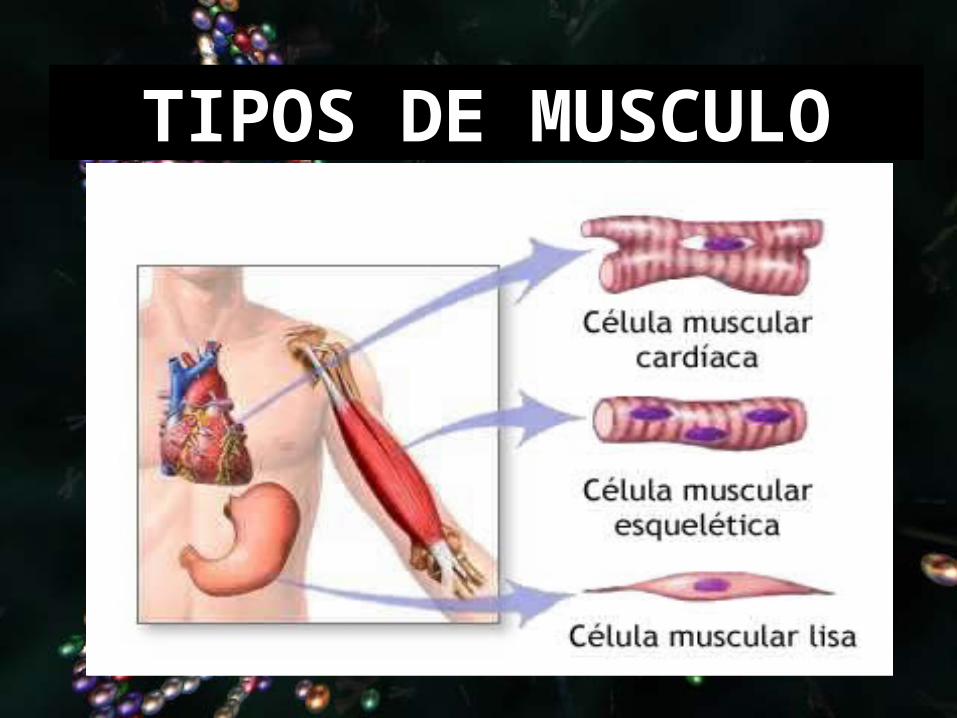
TIPOS DE MUSCULO

FUNCIÓN
Contractilidad, mediante la que se contrae al acortar sus fibras
Elasticidad, que permite que un músculo contraído recupere su forma
Tonicidad, gracias a la cual el músculo queda siempre semi contraido, ejerciendo de modo permanente una acción sobre los huesos a los que está adherido.

• Los músculos se contraen en grupos; están dispuestos en pares antagonistas, uno tira de un hueso en una dirección y el otro a la inversa, estos pares antagónicos se encuentran en la muñeca, rodilla, tobillo y otras articulaciones.
CONTRACCIÓN MUSCULAR
Siempre que un flexor se contrae, deberá relajarse el extensor en oposición mediante la coordinación de los impulsos nerviosos dirigidos a los músculos antagonistas.

CONTRACCIÓN MUSCULAR

• El músculo está compuesto de agua en un 80% de la masa con un resto principalmente
proteínico y pequeñas cantidades de grasas y glucógeno, así como dos sustancias
fosforadas, la fosfocreatina y el trifosfato de adenosina.
BIOQUIMICA DE LA CONTRACCIÓN

• En el músculo hay dos proteínas, miosina y actina, que actúan en forma conjunta.
BIOQUIMICA DE LA CONTRACCIÓN
• Durante la contracción muscular hay sustancias que disminuyen su cantidad: glucógeno, oxígeno, fosfocreatina y trifosfato de adenosina
• Otros elementos que aumentan: anhídrido carbónico, ácido láctico, difosfato de adenosina y fósforo inorgánico
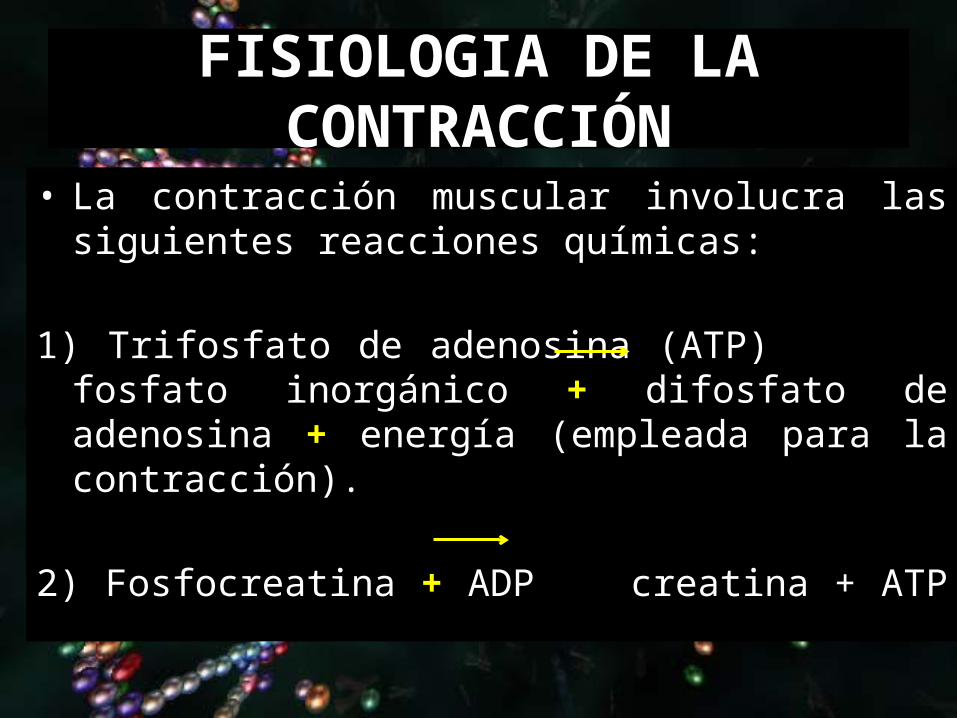
FISIOLOGIA DE LA CONTRACCIÓN
• La contracción muscular involucra las siguientes reacciones químicas:
1) Trifosfato de adenosina (ATP) fosfato inorgánico +
difosfato de adenosina + energía (empleada para la contracción).
2) Fosfocreatina + ADP creatina + ATP

3) Glucógeno + intermediarios ácido láctico + energía (empleada para la resíntesis de los fosfatos orgánicos).
4) Parte del ácido láctico + O2 > CO2 + H2 +
energía (empleada para resintetizar el resto del ácido, glucógeno y en la re síntesis de ATP y fosfocreatina).
FISIOLOGIA DE LA CONTRACCIÓN

DEUDA DE OXÍGENO
• Durante los momentos de violenta actividad, los músculos utilizan la energía que no necesita oxígeno, al cesar el movimiento, el sistema muscular y otros tejidos pagan la "deuda“, con el fin de restaurar los compuestos fosfóricos energéticos y el glucógeno a su estado original.

FATIGA
• El músculo que se contrajo repetidamente y por ello agotó sus reservas de glucógeno y fosfatos orgánicos y acumuló ácido láctico, habrá perdido su poder de contracción por lo que se dice que está fatigado.

CONTRACCIÓN DEL MÚSCULO LISO
• El músculo liso está compuesto por fibras mucho más pequeñas que las del esquelético.
• TIPO DE MÚSCULO LISO
• Músculo liso multiunitario: compuesto de fibras musculares lisas discretas. Están revestidas por una sustancia de colágeno fibrillas glucoprotéicas. Cada fibra puede contraerse independientemente de las otras, y su control se ejerce principalmente por señales nerviosas.
Músculo liso unitario: También conocido como visceral o sinicital. Significa que una masa de miles de fibras se contraen juntas como si fueran una unidad. Las fibras están dispuestas en capas y unidas por uniones intracelulares, que permite que una fuerza generada en la fibra se transmita a la siguiente.
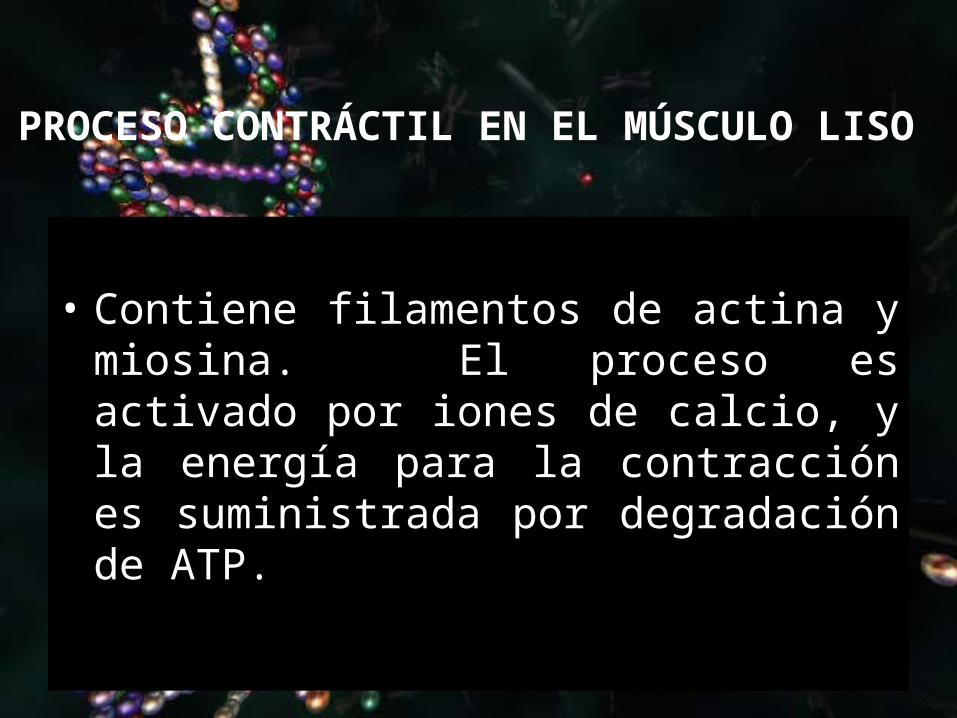
PROCESO CONTRÁCTIL EN EL MÚSCULO LISO
• Contiene filamentos de actina y miosina. El proceso es activado por iones de calcio, y la energía para la contracción es suministrada por degradación de ATP.

COMPARACIÓN ENTRE LA CONTRACCIÓN MUSCULAR LISA Y LA ESQUELÉTICA
1. Ciclo lento de los puentes transversales: significa la unión de la actina, después su liberación de la actina, y la nueva unión para el ciclo siguiente. Es mucho más lenta en el m liso

COMPARACIÓN ENTRE LA CONTRACCIÓN MUSCULAR LISA Y LA ESQUELÉTICA
2. Energía requerida para mantener la contracción muscular lisa: Sólo se requiere 1 ATP por cada ciclo, independientemente de su duración. Economía energética. Ejemplo: intestino que mantiene su contracción tónica indefinidamente.

COMPARACIÓN ENTRE LA CONTRACCIÓN MUSCULAR LISA Y LA ESQUELÉTICA
3. Lentitud de la contracción y relajación del músculo liso: Causados por la lentitud del establecimiento y rotura de los puentes transversales.
4. Fuerza de contracción muscular: Es mucho más grande que la del esquelético. Se debe al prolongado periodo de anclaje de los puentes transversales de miosina a los filamentos de actina.

COMPARACIÓN ENTRE LA CONTRACCIÓN MUSCULAR LISA Y LA ESQUELÉTICA
5. Porcentaje de acortamiento del músculo liso durante la contracción: Se puede acortar mucho, hasta 2/3 partes de su longitud estirada. Le permite hacer funciones específicas en vísceras huecas (vejiga)

COMPARACIÓN ENTRE LA CONTRACCIÓN MUSCULAR LISA Y LA ESQUELÉTICA
6. Mecanismo de cerrojo para contracciones sostenidas en el músculo liso: Una vez que el m. liso ha desarrollado la contracción máxima, el grado de activación puede reducirse a un nivel menor del inicial, Lo importante de este mecanismo es que permite la contracción prolongada en m. liso durante bastante tiempo y con gasto mínimo de energía.

COMPARACIÓN ENTRE LA CONTRACCIÓN MUSCULAR LISA Y LA ESQUELÉTICA
7. Relajación del estrés del músculo liso: También se caracteriza por la capacidad de retornar a una fuerza de contracción casi idéntica a la original segundos después de ser alargado o acortado. Permite a un órgano hueco mantener aproximadamente la misma presión independientemente de la longitud de las fibras musculares
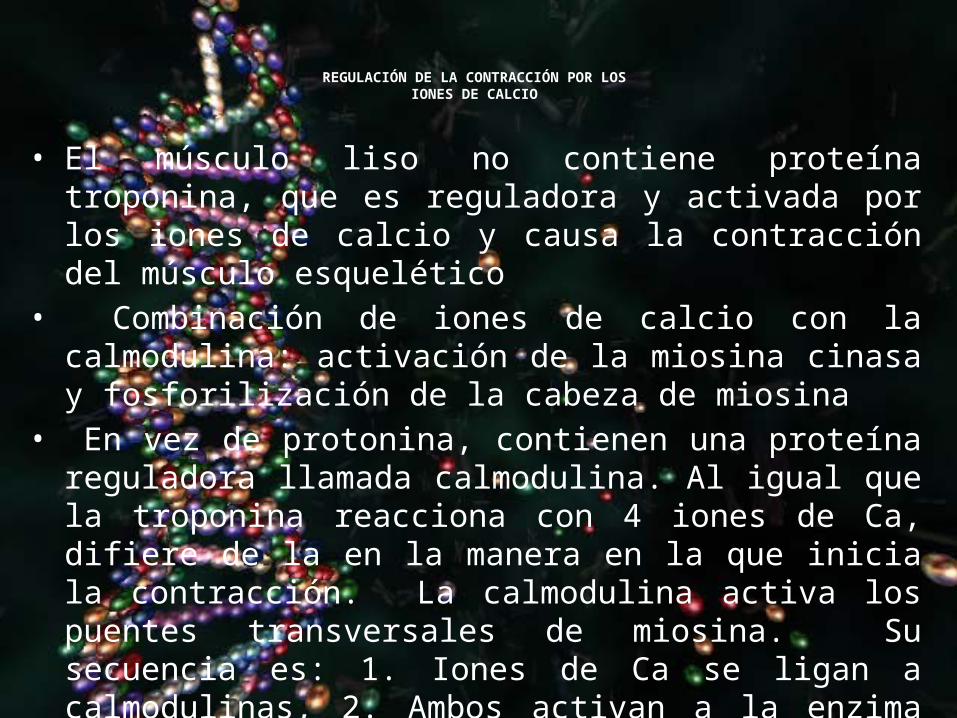
REGULACIÓN DE LA CONTRACCIÓN POR LOS IONES DE CALCIO
• El músculo liso no contiene proteína troponina, que es reguladora y activada por los iones de calcio y causa la contracción del músculo esquelético
• Combinación de iones de calcio con la calmodulina: activación de la miosina cinasa y fosforilización de la cabeza de miosina
• En vez de protonina, contienen una proteína reguladora llamada calmodulina. Al igual que la troponina reacciona con 4 iones de Ca, difiere de la en la manera en la que inicia la contracción. La calmodulina activa los puentes transversales de miosina. Su secuencia es: 1. Iones de Ca se ligan a calmodulinas, 2. Ambos activan a la enzima miosina cinasa, 3. Una de las cabezas de las cabezas de misosina sefosforila por la enzima
•
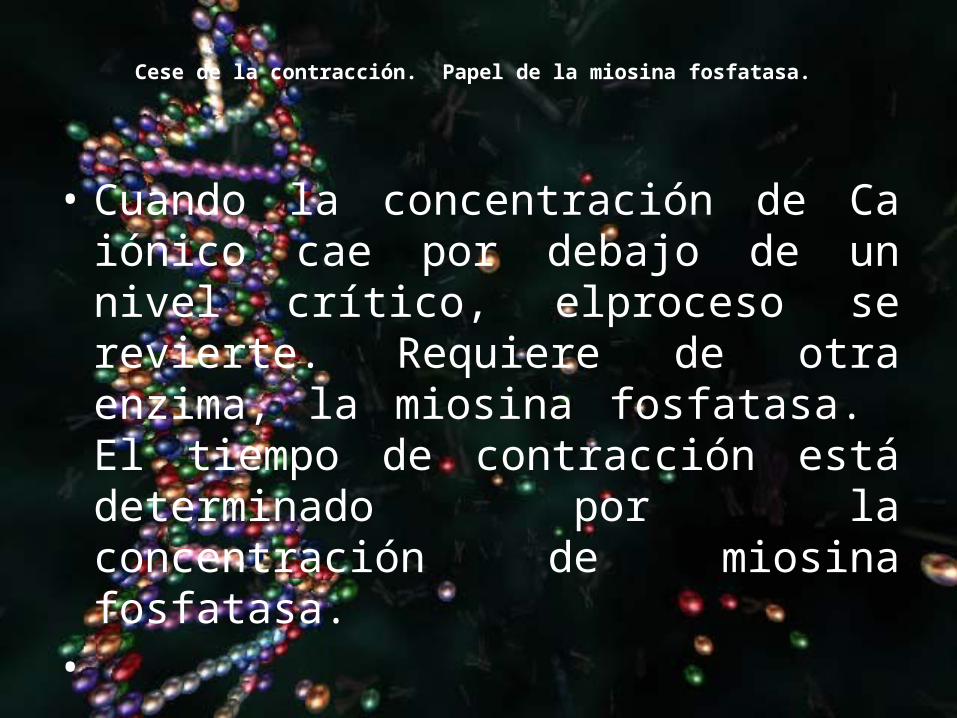
Cese de la contracción. Papel de la miosina fosfatasa.
• Cuando la concentración de Ca iónico cae por debajo de un nivel crítico, elproceso se revierte. Requiere de otra enzima, la miosina fosfatasa. El tiempo de contracción está determinado por la concentración de miosina fosfatasa.
•

CONTROL NEUROLÓGICO DE LA CONTRACCIÓN DEL MÚSCULO LISO
• Se diferencia básicamente en que el m liso tiene una membrana que contiene muchos tipos de proteínas receptoras que pueden comenzar el proceso de contracción, o inhibirlo.
• •

UNIONES NEUROMUSCULARES DEL MÚSCULO LISO
• • En el m liso no existen uniones neuromusculares tan
complejas. Las fibras del sistema nervioso autónomo se distribuyen en forma difusa sobre una capa de fibras musculares. Estas fibras no entran en contacto directo con las fibras musculares lisas, si no que forman las uniones difusas que segregan sustancias transmisoras. Los axones terminales tienen múltiples varicosidades, por las que son secretadas las sustancias transmisoras (acetilcolina, noradrenalina)
•
Sustancias transmisoras excitadoras e inhibidoras en la unión neuromuscular del músculo liso
• • Son la acetilcolina y la noradrenalina. Cuando
la acetilcolina excita una fibra muscular, la noradrenalina la inhibe, y viceversa.
• Esto es debido a que ambas se ligan de primero a una proteína receptora de la membrana de la célula muscular. De las proteínas, hay receptores inhibidores y receptores excitadores.

POTENCIALES DE MEMBRANA Y DE ACCIÓN EN EL M. LISO
• Potencial de membrana: entre 50 y 60 milivoltios.
• Potencial de acción en el músculo liso unitario: • En punta: la mayoría de los músculos unitarios. Se
pueden desencadenar de muchas maneras. • Con mesetas: Inicia igual que el de punta,, pero la
repolarización de la membrana se atrasa unos milisegundos.
• La importancia de la meseta es que puede ser responsable delos periodos largos de contracción del m liso.
Importancia de los canales de calcio en la génesis del potencial de acción del músculo liso
•
• La membrana de la célula lisa contiene muchos más canales de calcio que la de la esquelética, pero pocos canales de sodio. El flujo principal de los iones de Ca es el responsable del potencial de acción. Los canales de calcio se abren muchas veces más lentamente que los canales de sodio, pero permanecen abiertos más tiempo
• Algunos músculos lisos como el intestino, el útero, son autoexcitables. Los potenciales de acción surgen en el m liso sin impulso externo
• Cuando el m liso visceral se distiende suficientemente, se generan
habitualmente potenciales de acción
• Las fibras musculares lisas del músculo liso multi-unitario se contraen en respuesta a estímulos nerviosos

CONTRACCIÓN DEL MÚSCULO LISO SIN POTENCIAL DE ACCIÓN. EFECTO DE FACTORES TISULARES LOCALES Y DE HORMONAS
• • Quizá la mitad de la contracción no es desencadenada por
potenciales de acción, sino por factores estimuladores que actúan directamente. Estos factores son:
• • Tisulares locales: Falta de oxígeno, exceso de dióxido de
carbono, aumento de la concentración local de hidrógeno, causan aumento en la vasodilatación.
• • Diversas hormonas: Causan contracción cuando una
membrana tiene receptores específicos excitadores para ellas. Entre ellas: noradrenalina, acetilcolina, adrenalina, serotonina, histamina, etc.
PROCEDENCIA DE LOS IONES DE CALCIO QUE CAUSAN LA CONTRACCIÓN
• Aunque el m liso y el esquelético requieren de calcio para el proceso contráctil, la fuente de éste es diferente. La diferencia básica es que los iones de calcio usados en el esquelético provienen del retículo sarcoplásmico, y en el m liso provienen del líquido extracelular en el momento del potencial de acción. Son los iones de calcio los que causan la contracción, y NO el potencial de membrana.
• • Para que se relaje el m contraído es necesario eliminarlos
iones de calcio de los líquidos intracelulares que rodean filamentos de actina y miosina. Esto se lograra por la bomba de calcio que bombea de nuevo los iones al líquido extracelular o al retículo sarcoplásmico.
• •