Centro: NEIKER Participantes: Entidades …...Informe técnico final CLONCITO 2 Clonación de...
12
Informe técnico final CLONCITO 1 Informe final de proyecto Clonación de individuos adultos del género Pinus spp. Identificación de genes implicados en los procesos organogénicos Centro: NEIKER Participantes: Paloma Moncalean ([email protected]) Itziar Montalbán, Nuria de Diego, Sonia Castañón, Pablo G.Goikoetxea Entidades participantes: Universidad de Oviedo Año 2006
Transcript of Centro: NEIKER Participantes: Entidades …...Informe técnico final CLONCITO 2 Clonación de...

Informe técnico final CLONCITO 1
Informe final de proyecto
Clonación de individuos adultos del género Pinus spp.
Identificación de genes implicados en los procesos organogénicos
Centro: NEIKER
Participantes:
Paloma Moncalean ([email protected])
Itziar Montalbán, Nuria de Diego,
Sonia Castañón, Pablo G.Goikoetxea
Entidades participantes:
Universidad de Oviedo
Año 2006

Informe técnico final CLONCITO 2
Clonación de individuos adultos del género Pinus spp. Identificación de genes implicados en los procesos organogénicos
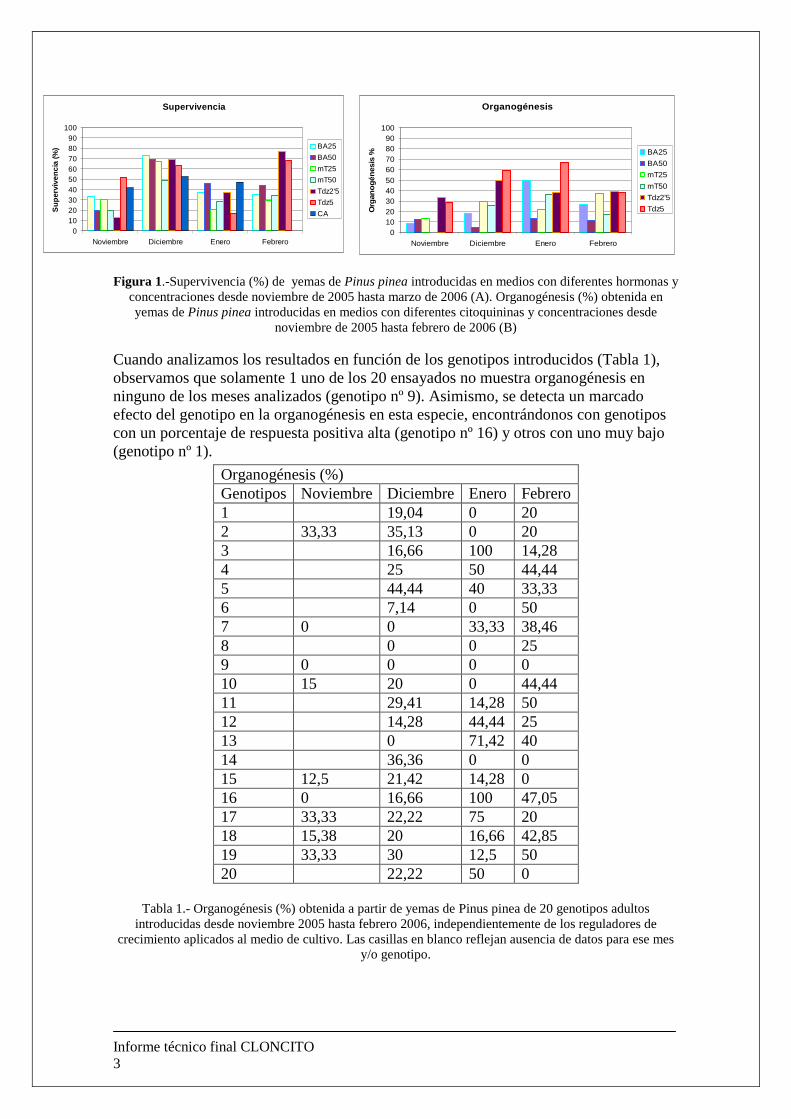
1.- Introducción El objetivo principal de este proyecto era el de la clonación de individuos adultos de especies del género Pinus spp así como la identificación de genes implicados en procesos de organogénesis. 2.-Clonación de individuos adultos del género Pinus spp Establecimiento de las condiciones de cultivo, la época de recogida de las muestras, el medio de cultivo y los reguladores de crecimiento necesario para lograr la organogénesis de yemas de individuos adultos de P. pinea, P. pinaster y P. sylvestris. Pinus pinea Se seleccionaron 20 individuos de más de 15 años instalados en una plantación de recreo de la Comunidad Autónoma Vasca. Debido al desconocimiento del momento óptimo en el que deben encontrarse las yemas para la inducción de los procesos organogénicos en esta especie, pero basándonos en la experiencia adquirida con P. radiata, realizamos una recogida de las yemas caulinares al menos una vez al mes desde noviembre de 2005 hasta marzo de 2006. Una vez escindidas del árbol y en un plazo nunca superior a 72 horas, las yemas se sometieron a un proceso de asepsia. Dicho proceso consistió en la inmersión de las yemas durante 20 minutos en hipoclorito sódico comercial al 50% (v/v) y 3 lavados en agua destilada estéril de 5 minutos cada uno. Una vez realizada la asepsia, las yemas se introdujeron en medio LP modificado (Aitken J., comunicación personal). Con el fin de determinar la importancia del tipo de citoquinina empleada para inducir organogénesis se ensayaron: Benciladenina (BA), meta-topolina (mT) y Tidiazuron (Tdz) así como diversas concentraciones para elegir la óptima (25 y 50 µM). Además, se ensayó un medio de similares características al anterior pero sin ninguna citoquinina presente en el medio y con carbon activo (3 g/L). Se introdujeron entre 300 y 500 segmentos de yemas caulinares en cada uno de los meses ensayados. Como podemos observar en la Figura 1 A, la supervivencia de las yemas introducidas in vitro fue mayor en el mes de Diciembre generalmente, con lo que podemos decir que independientemente de cual sea el contenido del medio de cultivo utilizado para la organogénesis de yemas, diciembre fue el mes óptimo para lograr el mayor porcentaje de éxito en la introducción. Si analizamos la organogénesis obtenida (Figura 1 B), observamos que el Tidiazuron es la hormona que mejores resultados produce en todos los meses ensayados, llegándose a observar un 60-70% de organogénesis durante los meses de Enero y Febrero (Figura 2 A).
Informe técnico final CLONCITO 3
Supervivencia
0102030405060708090
100
Noviembre Diciembre Enero Febrero
Sup
ervi
venc
ia (%
)
BA25
BA50
mT25
mT50
Tdz2'5
Tdz5
CA
Organogénesis
0102030405060708090
100
Noviembre Diciembre Enero Febrero
Org
anog
énes
is %
BA25
BA50
mT25
mT50
Tdz2'5
Tdz5
Figura 1.-Supervivencia (%) de yemas de Pinus pinea introducidas en medios con diferentes hormonas y concentraciones desde noviembre de 2005 hasta marzo de 2006 (A). Organogénesis (%) obtenida en yemas de Pinus pinea introducidas en medios con diferentes citoquininas y concentraciones desde
noviembre de 2005 hasta febrero de 2006 (B)
Cuando analizamos los resultados en función de los genotipos introducidos (Tabla 1), observamos que solamente 1 uno de los 20 ensayados no muestra organogénesis en ninguno de los meses analizados (genotipo nº 9). Asimismo, se detecta un marcado efecto del genotipo en la organogénesis en esta especie, encontrándonos con genotipos con un porcentaje de respuesta positiva alta (genotipo nº 16) y otros con uno muy bajo (genotipo nº 1).
Tabla 1.- Organogénesis (%) obtenida a partir de yemas de Pinus pinea de 20 genotipos adultos introducidas desde noviembre 2005 hasta febrero 2006, independientemente de los reguladores de
crecimiento aplicados al medio de cultivo. Las casillas en blanco reflejan ausencia de datos para ese mes y/o genotipo.
Organogénesis (%) Genotipos Noviembre Diciembre Enero Febrero 1 19,04 0 20 2 33,33 35,13 0 20 3 16,66 100 14,28 4 25 50 44,44 5 44,44 40 33,33 6 7,14 0 50 7 0 0 33,33 38,46 8 0 0 25 9 0 0 0 0 10 15 20 0 44,44 11 29,41 14,28 50 12 14,28 44,44 25 13 0 71,42 40 14 36,36 0 0 15 12,5 21,42 14,28 0 16 0 16,66 100 47,05 17 33,33 22,22 75 20 18 15,38 20 16,66 42,85 19 33,33 30 12,5 50 20 22,22 50 0

Informe técnico final CLONCITO 4
Como mencionamos anteriormente, los mejores resultados se han obtenido cuando se ha utilizado como citoquinina el Tidiazuron a una concentración de 5 µM, observándose que durante el mes de enero el 70% de las yemas introducidas se muestran organogénicas (Figura 1 A y B). Posteriormente y para lograr la elongación de los primordios inducidos, introdujimos las yemas en un medio LP modificado (Aitken J., comunicación personal) de las mismas características al utilizado en la fase de inducción pero sin reguladores de crecimiento y con carbono activo. Tras 3 subcultivos en dicho medio, los explantos muestran un aspecto juvenil como el que podemos observar en la figura 2 B y C.
Figura 2.- Yemas de Pinus pinea en fase de inducción organogénica (A). Microtallos de Pinus pinea procedentes de yemas de un individuo adulto tras la elongación de las yemas cultivadas en un medio con ausencia de reguladores de crecimiento y en presencia de carbon activo (B). Microtallos obtenidos tras 3 subcultivos en medio de elongación con carbon activo antes de pasar a la etapa de inducción rizogénica
(C)
Pinus pinaster Se seleccionaron 20 individuos (denominados A-1 hasta A-20) de más de 15 años instalados en una plantación productiva de la Comunidad Autónoma Vasca. Debido al desconocimiento del momento óptimo en el que deben encontrarse las yemas para la inducción de los procesos organogénicos en esta especie pero basándonos en la experiencia adquirida con P. radiata, realizamos una recogida de las yemas caulinares al menos una vez al mes desde noviembre de 2005 hasta marzo de 2006. Una vez escindidas del árbol y en un plazo nunca superior a 72 horas, las yemas se sometieron a un proceso de asepsia. Dicho proceso consistió en la inmersión de las yemas durante 20 minutos en hipoclorito sódico comercial al 50% (v/v) y 3 lavados en agua destilada estéril de 5 minutos cada uno. Una vez realizada la asepsia, las yemas se introdujeron en medio diferentes medios de cultivo: DCR (Gupta y Durzan, 1987), LP (Quorin y Lepoivre, 1977) y WP (Lloyd y McCown, 1980).
Informe técnico final CLONCITO 5
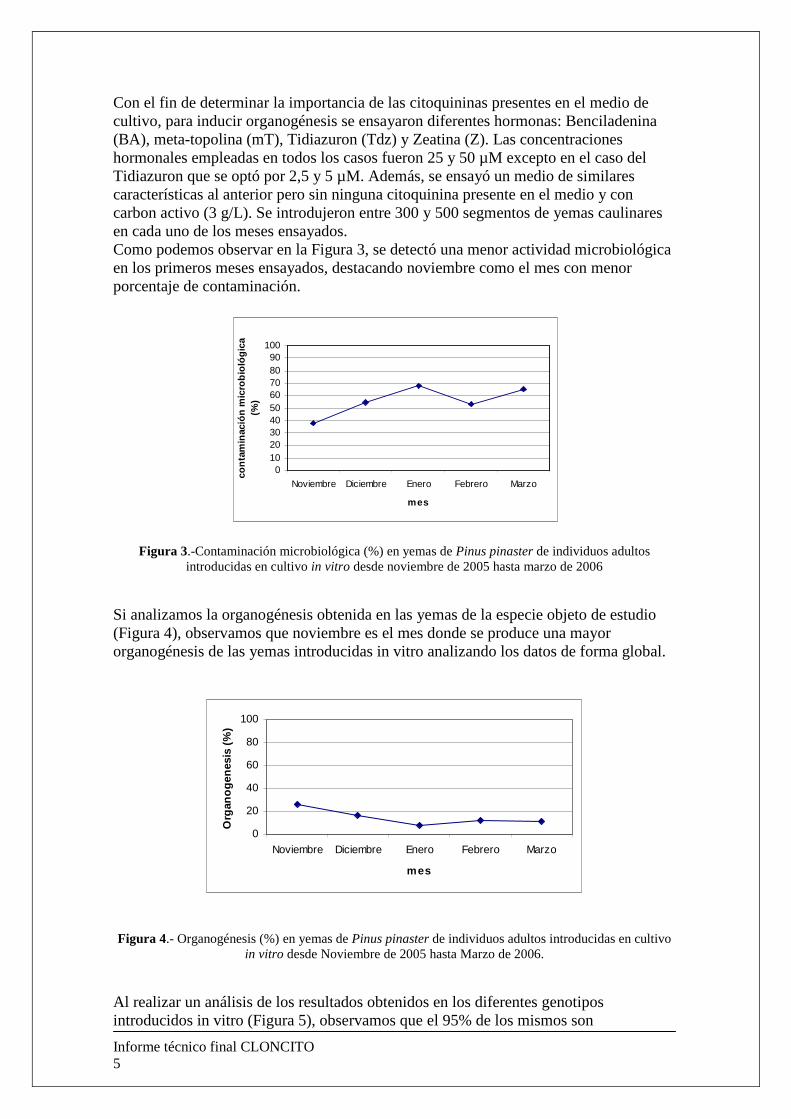
Con el fin de determinar la importancia de las citoquininas presentes en el medio de cultivo, para inducir organogénesis se ensayaron diferentes hormonas: Benciladenina (BA), meta-topolina (mT), Tidiazuron (Tdz) y Zeatina (Z). Las concentraciones hormonales empleadas en todos los casos fueron 25 y 50 µM excepto en el caso del Tidiazuron que se optó por 2,5 y 5 µM. Además, se ensayó un medio de similares características al anterior pero sin ninguna citoquinina presente en el medio y con carbon activo (3 g/L). Se introdujeron entre 300 y 500 segmentos de yemas caulinares en cada uno de los meses ensayados. Como podemos observar en la Figura 3, se detectó una menor actividad microbiológica en los primeros meses ensayados, destacando noviembre como el mes con menor porcentaje de contaminación.
0102030405060708090
100
Noviembre Diciembre Enero Febrero Marzo
mes
con
tam
inac
ión
mic
robi
ológ
ica
(%)
Figura 3.-Contaminación microbiológica (%) en yemas de Pinus pinaster de individuos adultos introducidas en cultivo in vitro desde noviembre de 2005 hasta marzo de 2006
Si analizamos la organogénesis obtenida en las yemas de la especie objeto de estudio (Figura 4), observamos que noviembre es el mes donde se produce una mayor organogénesis de las yemas introducidas in vitro analizando los datos de forma global.
0
20
40
60
80
100
Noviembre Diciembre Enero Febrero Marzo
mes
Org
anog
enes
is (
%)
Figura 4.- Organogénesis (%) en yemas de Pinus pinaster de individuos adultos introducidas en cultivo in vitro desde Noviembre de 2005 hasta Marzo de 2006.
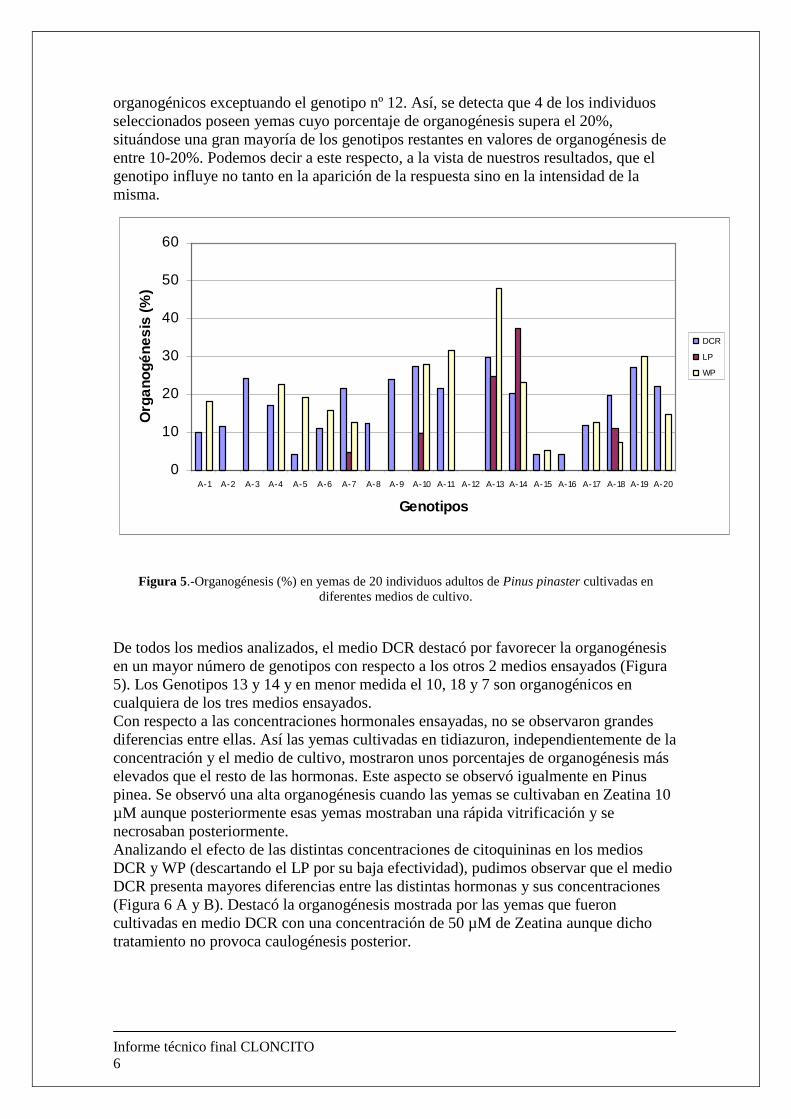
Al realizar un análisis de los resultados obtenidos en los diferentes genotipos introducidos in vitro (Figura 5), observamos que el 95% de los mismos son
Informe técnico final CLONCITO 6
organogénicos exceptuando el genotipo nº 12. Así, se detecta que 4 de los individuos seleccionados poseen yemas cuyo porcentaje de organogénesis supera el 20%, situándose una gran mayoría de los genotipos restantes en valores de organogénesis de entre 10-20%. Podemos decir a este respecto, a la vista de nuestros resultados, que el genotipo influye no tanto en la aparición de la respuesta sino en la intensidad de la misma.
0
10
20
30
40
50
60
A-1 A-2 A-3 A-4 A-5 A-6 A-7 A-8 A-9 A-10 A-11 A-12 A-13 A-14 A-15 A-16 A-17 A-18 A-19 A-20
Genotipos
Org
anog
énes
is (%
)
DCR
LP
WP
Figura 5.-Organogénesis (%) en yemas de 20 individuos adultos de Pinus pinaster cultivadas en diferentes medios de cultivo.
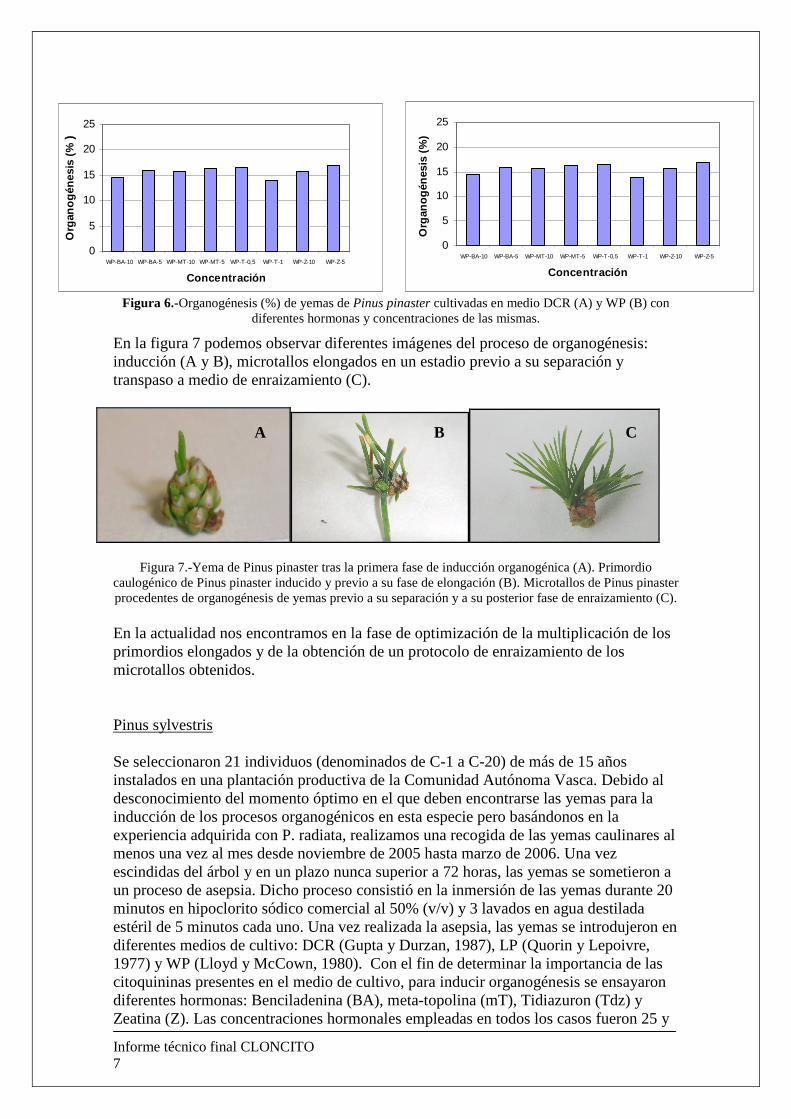
De todos los medios analizados, el medio DCR destacó por favorecer la organogénesis en un mayor número de genotipos con respecto a los otros 2 medios ensayados (Figura 5). Los Genotipos 13 y 14 y en menor medida el 10, 18 y 7 son organogénicos en cualquiera de los tres medios ensayados. Con respecto a las concentraciones hormonales ensayadas, no se observaron grandes diferencias entre ellas. Así las yemas cultivadas en tidiazuron, independientemente de la concentración y el medio de cultivo, mostraron unos porcentajes de organogénesis más elevados que el resto de las hormonas. Este aspecto se observó igualmente en Pinus pinea. Se observó una alta organogénesis cuando las yemas se cultivaban en Zeatina 10 µM aunque posteriormente esas yemas mostraban una rápida vitrificación y se necrosaban posteriormente. Analizando el efecto de las distintas concentraciones de citoquininas en los medios DCR y WP (descartando el LP por su baja efectividad), pudimos observar que el medio DCR presenta mayores diferencias entre las distintas hormonas y sus concentraciones (Figura 6 A y B). Destacó la organogénesis mostrada por las yemas que fueron cultivadas en medio DCR con una concentración de 50 µM de Zeatina aunque dicho tratamiento no provoca caulogénesis posterior.
Informe técnico final CLONCITO 7
Figura 6.-Organogénesis (%) de yemas de Pinus pinaster cultivadas en medio DCR (A) y WP (B) con diferentes hormonas y concentraciones de las mismas.
En la figura 7 podemos observar diferentes imágenes del proceso de organogénesis: inducción (A y B), microtallos elongados en un estadio previo a su separación y transpaso a medio de enraizamiento (C).
Figura 7.-Yema de Pinus pinaster tras la primera fase de inducción organogénica (A). Primordio caulogénico de Pinus pinaster inducido y previo a su fase de elongación (B). Microtallos de Pinus pinaster procedentes de organogénesis de yemas previo a su separación y a su posterior fase de enraizamiento (C). En la actualidad nos encontramos en la fase de optimización de la multiplicación de los primordios elongados y de la obtención de un protocolo de enraizamiento de los microtallos obtenidos. Pinus sylvestris Se seleccionaron 21 individuos (denominados de C-1 a C-20) de más de 15 años instalados en una plantación productiva de la Comunidad Autónoma Vasca. Debido al desconocimiento del momento óptimo en el que deben encontrarse las yemas para la inducción de los procesos organogénicos en esta especie pero basándonos en la experiencia adquirida con P. radiata, realizamos una recogida de las yemas caulinares al menos una vez al mes desde noviembre de 2005 hasta marzo de 2006. Una vez escindidas del árbol y en un plazo nunca superior a 72 horas, las yemas se sometieron a un proceso de asepsia. Dicho proceso consistió en la inmersión de las yemas durante 20 minutos en hipoclorito sódico comercial al 50% (v/v) y 3 lavados en agua destilada estéril de 5 minutos cada uno. Una vez realizada la asepsia, las yemas se introdujeron en diferentes medios de cultivo: DCR (Gupta y Durzan, 1987), LP (Quorin y Lepoivre, 1977) y WP (Lloyd y McCown, 1980). Con el fin de determinar la importancia de las citoquininas presentes en el medio de cultivo, para inducir organogénesis se ensayaron diferentes hormonas: Benciladenina (BA), meta-topolina (mT), Tidiazuron (Tdz) y Zeatina (Z). Las concentraciones hormonales empleadas en todos los casos fueron 25 y
0
5
10
15
20
25
WP-BA-10 WP-BA-5 WP-MT-10 WP-MT-5 WP-T-0,5 WP-T-1 WP-Z-10 WP-Z-5
Concentración
Org
anog
énes
is (
%)
0
5
10
15
20
25
WP-BA-10 WP-BA-5 WP-MT-10 WP-MT-5 WP-T-0,5 WP-T-1 WP-Z-10 WP-Z-5
Concentración
Org
anog
énes
is (
%)
A B C

Informe técnico final CLONCITO 8
50 µM excepto en el caso del Tidiazuron que se optó por 2,5 y 5 µM. Además, se ensayó un medio de similares características al anterior pero sin ninguna citoquinina presente en el medio y con carbon activo (3 g/L). Se introdujeron entre 300 y 500 segmentos de yemas caulinares en cada uno de los meses ensayados. Como podemos observar en la figura 8, se observa un menor porcentaje de contaminación microbiológica en las yemas introducidas en el mes de Diciembre.
0
20
40
60
80
100
NOVIEMBRE DICIEMBRE ENERO MARZO
Meses
Con
tam
inac
ión
mic
robi
oló
gica
(%
)
Figura 8.- Contaminación microbiológica (%) en yemas de individuos adultos de Pinus sylvestris introducidas in vitro desde noviembre de 2005 hasta marzo de 2006.
Paralelamente, se observó (Figura 9) que el porcentaje de organogénesis en las yemas de esta especie disminuyó desde noviembre a marzo, siendo el primero de éstos el mes más óptimo para lograr la organogénesis en individuos adultos de esta especie.
0
20
40
60
80
100
NOVIEMBRE DICIEMBRE ENERO MARZO
Meses
Org
anog
énes
is (
%)
Figura 9.-Organogénesis (%) en yemas de Pinus pinaster de individuos adultos introducidas en cultivo in vitro desde Noviembre de 2005 hasta Marzo de 2006.
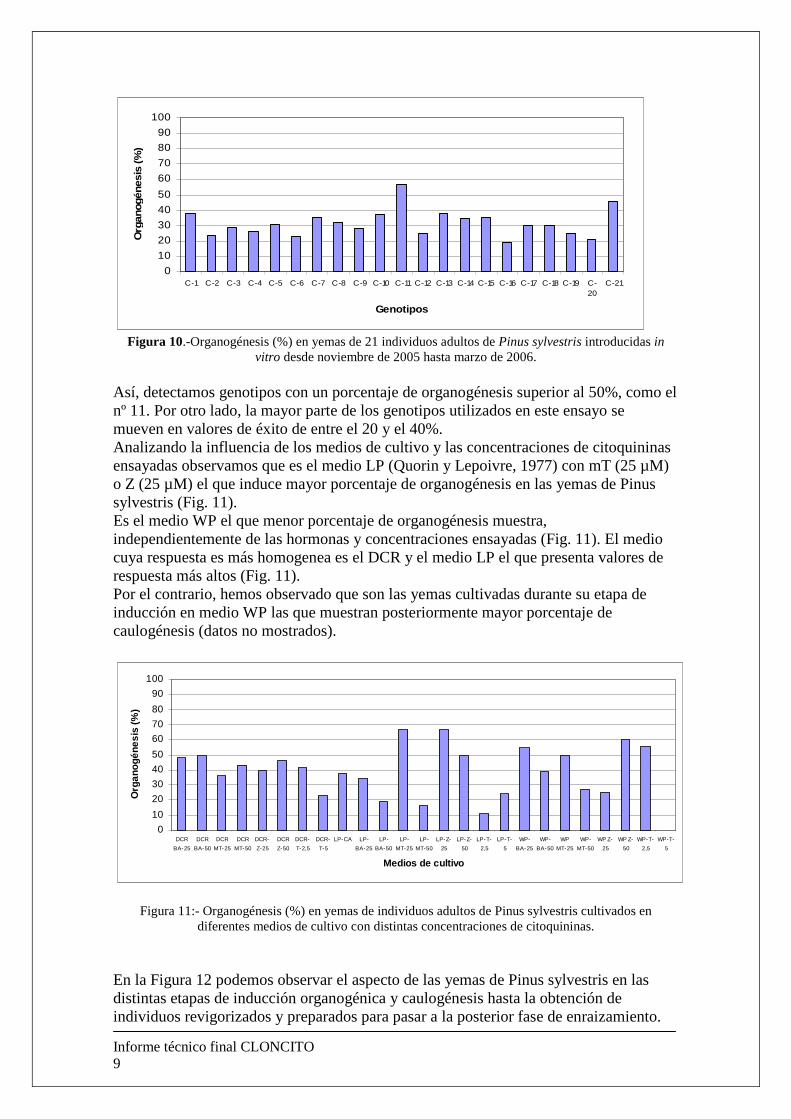
Cuando analizamos los resultados obtenidos en base a los diferentes genotipos introducidos (Fig. 10), detectamos que la influencia del genotipo afecta a la intensidad de la respuesta pero no a la posibilidad de obtenerla ya que todos los genotipos ensayados resultaron ser organogénicos.
Informe técnico final CLONCITO 9
0
10
20
30
40
50
60
70
80
90
100
C-1 C-2 C-3 C-4 C-5 C-6 C-7 C-8 C-9 C-10 C-11 C-12 C-13 C-14 C-15 C-16 C-17 C-18 C-19 C-20
C-21
Genotipos
Org
anog
énes
is (%
)
Figura 10.-Organogénesis (%) en yemas de 21 individuos adultos de Pinus sylvestris introducidas in
vitro desde noviembre de 2005 hasta marzo de 2006. Así, detectamos genotipos con un porcentaje de organogénesis superior al 50%, como el nº 11. Por otro lado, la mayor parte de los genotipos utilizados en este ensayo se mueven en valores de éxito de entre el 20 y el 40%. Analizando la influencia de los medios de cultivo y las concentraciones de citoquininas ensayadas observamos que es el medio LP (Quorin y Lepoivre, 1977) con mT (25 µM) o Z (25 µM) el que induce mayor porcentaje de organogénesis en las yemas de Pinus sylvestris (Fig. 11). Es el medio WP el que menor porcentaje de organogénesis muestra, independientemente de las hormonas y concentraciones ensayadas (Fig. 11). El medio cuya respuesta es más homogenea es el DCR y el medio LP el que presenta valores de respuesta más altos (Fig. 11). Por el contrario, hemos observado que son las yemas cultivadas durante su etapa de inducción en medio WP las que muestran posteriormente mayor porcentaje de caulogénesis (datos no mostrados).
0
10
20
30
40
50
60
70
80
90
100
DCR
BA-25
DCR
BA-50
DCR
MT-25
DCR
MT-50
DCR-
Z-25
DCR
Z-50
DCR-
T-2,5
DCR-
T-5
LP-CA LP-
BA-25
LP-
BA-50
LP-
MT-25
LP-
MT-50
LP-Z-
25
LP-Z-
50
LP-T-
2,5
LP-T-
5
WP-
BA-25
WP-
BA-50
WP
MT-25
WP-
MT-50
WP Z-
25
WP Z-
50
WP-T-
2,5
WP-T-
5
Medios de cultivo
Org
anog
énes
is (%
)
Figura 11:- Organogénesis (%) en yemas de individuos adultos de Pinus sylvestris cultivados en diferentes medios de cultivo con distintas concentraciones de citoquininas.
En la Figura 12 podemos observar el aspecto de las yemas de Pinus sylvestris en las distintas etapas de inducción organogénica y caulogénesis hasta la obtención de individuos revigorizados y preparados para pasar a la posterior fase de enraizamiento.

Informe técnico final CLONCITO 10
En la actualidad nos encontramos en la fase de elongación de los microtallos formados y desarrollando un protocolo de optimización de la multiplicación y enraizamiento de dichos microtallos
Figura 12.- Yemas de Pinus sylvestris procedente de un individuo adulto en su fase de inducción organogénica (A), tras su posterior etapa de manifestación de los primordios inducidos (B) y previo a su separación antes de pasar a su fase de elongación y enraizamiento (C).
Este objetivo se culminará a través de la ejecución del proyecto CLONCITO II financiado por el Departamento de Agricultura y Pesca del Gobierno Vasco.
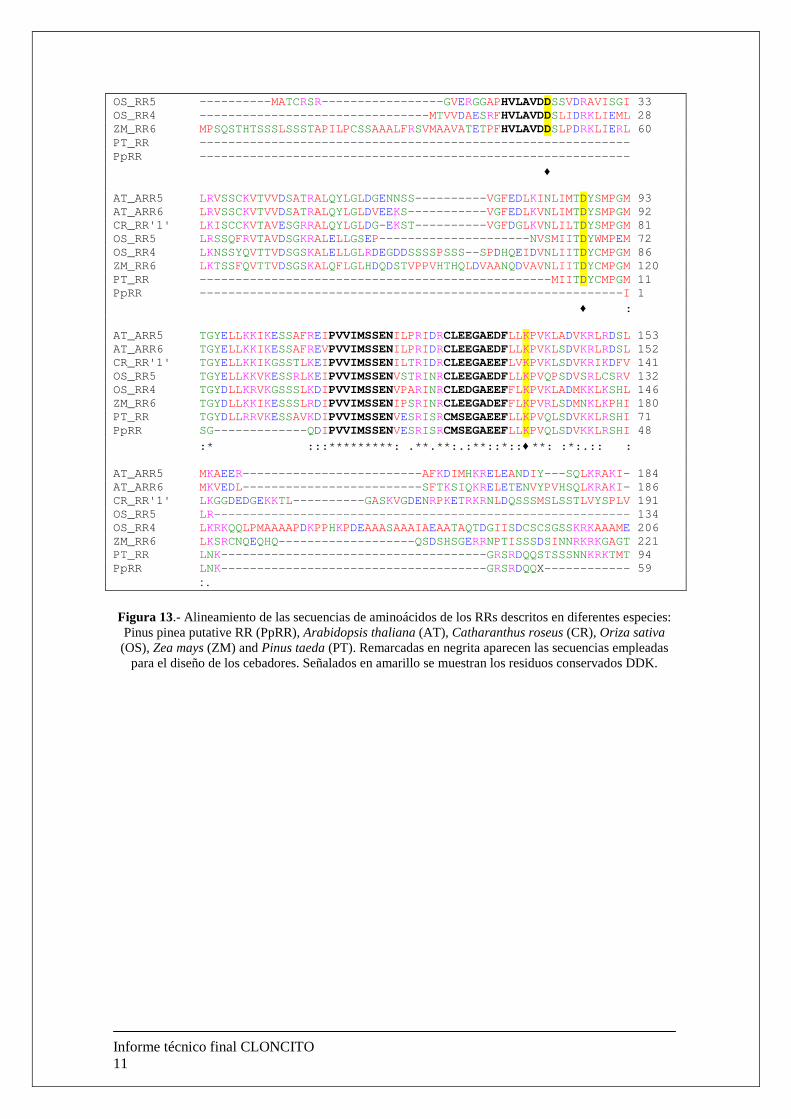
3.- Base molecular del mecanismo de acción de las citiquininas Teniendo en cuenta la reducción tanto del presupuesto solicitado como del tiempo de ejecución de las tareas del proyecto CLONCITO, hemos considerado maximizar los esfuerzos en las tareas relativas al objetivo 1 aumentando incluso las especies objeto de estudio y reducir las tareas descritas en el objetivo 2. No obstante, se ha iniciado un análisis de las secuencias descritas para los RRs tipo A y tipo B, proteínas implicadas en la transducción de señales en respuesta a citoquininas. Dentro de los RRs de tipo A identificados en Arabidopsis, se ha descrito tres residuos conservados DDK precediendo a una región rica en serina. A partir de diferentes secuencias obtenidas de la base de datos del NCBI, se procedió a realzar un análisis de homología ClustalW con diferentes especies: Pinus pinea, Pinus taeda, Populus alba, Catharanthus roseus, Zea maiz, Oryza sativa y Arabidopsis thaliana. El análisis de homología ha permitido identificar una región de aminoácidos altamente conservada en la mayoría de las especies analizadas, tanto en gimnospermas como en angiospermas. La región HVLAVDD situada en el extremo amino terminal, se ha utilizado para el diseño de oligonucleótidos específicos que podrán ser empleados para la identificación y aislamiento de RRs de tipo A en proyectos futuros. La segunda región identificada y comúnmente empleada en el diseño de cebadores específicos para la identificación y aislamiento de los RRs de tipo A, corresponde con la región CLEEGAED. En el análisis de homología realizado, se ha podido observar que dicha región se conserva en la mayor parte de las especies de angiospermas, no ocurre lo mismo con las gimnospermas. Teniendo en cuenta que la especie modelo objeto de este estudio es pinus radiata, parece conveniente emplear la región PVVIMSSEN para el diseño de cebadores que amplifiquen secuencias ortólogas a los RRs tipo A. AT_ARR5 MAEVLRPEMLDI SNDTS----------------- SLASPKLLHVLAVDDSMVDRKFI ERL 43 AT_ARR6 MAEVMLPRKMEIL NHSS----------------- KFGSPDPLHVLAVDDSHVDRKFI ERL 43 CR_RR'1' ----------- MASQCC----------------- SSGSAKELHVLAVDDSLVDRKVI ERL 32
A B C
Informe técnico final CLONCITO 11
OS_RR5 ---------- MATCRSR----------------- GVERGGAPHVLAVDDSSVDRAVI SGI 33 OS_RR4 -------------------------------- MTVVDAESRFHVLAVDDSLI DRKLI EML 28 ZM_RR6 MPSQSTHTSSSLSSSTAPILP CSSAAALFRSVMAAVATETPFHVLAVDDSLPDRKLI ERL 60 PT_RR --------------------------------------- --------------------- PpRR --------------------------------------- --------------------- ♦ AT_ARR5 LRVSSCKVTVVDSATRALQYLGLDGENNSS---------- VGFEDLKI NLIMTDYSMPGM 93 AT_ARR6 LRVSSCKVTVVDSATRALQYLGLDVEEKS----------- VGFEDLKVNLIMTDYSMPGM 92 CR_RR'1' LKI SCCKVTAVESGRRALQYLGLDG- EKST---------- VGFDGLKVNLIL TDYSMPGM 81 OS_RR5 LRSSQFRVTAVDSGKRALELLGSEP--------------------- NVSMII TDYWMPEM 72 OS_RR4 LKNSSYQVTTVDSGSKALELLGLRDEGDDSSSSPSSS-- SPDHQEI DVNLII TDYCMPGM 86 ZM_RR6 LKTSSFQVTTVDSGSKALQFLGLHDQDSTVPPVHTHQLDVAANQDVAVNLII TDYCMPGM 120 PT_RR --------------------------------------- ---------- MII TDYCMPGM 11 PpRR --------------------------------------- -------------------- I 1 ♦ : AT_ARR5 TGYELLKKI KESSAFREI PVVIMSSENILP RI DRCLEEGAEDFLLKPVKLADVKRLRDSL 153 AT_ARR6 TGYELLKKI KESSAFREVPVVIMSSENILP RI DRCLEEGAEDFLLKPVKLSDVKRLRDSL 152 CR_RR'1' TGYELLKKI KGSSTLKEI PVVIMSSENIL TRI DRCLEEGAEEFLVKPVKLSDVKRI KDFV 141 OS_RR5 TGYELLKKVKESSRLKEI PVVIMSSENVSTRI NRCLEEGAEDFLLKPVQPSDVSRLCSRV 132 OS_RR4 TGYDLLKRVKGSSSLKDI PVVIMSSENVPARI NRCLEDGAEEFFLKPVKLADMKKLKSHL 146 ZM_RR6 TGYDLLKKI KESSSLRDI PVVIMSSENIP SRI NRCLEEGADEFFLKPVRLSDMNKLKPHI 180 PT_RR TGYDLLRRVKESSAVKDI PVVIMSSENVESRI SRCMSEGAEEFLLKPVQLSDVKKLRSHI 71 PpRR SG------------- QDI PVVIMSSENVESRI SRCMSEGAEEFLLKPVQLSDVKKLRSHI 48
:* :::*********: .**.**:.:* *::*:: ♦**: :*:.:: : AT_ARR5 MKAEER------------------------- AFKDIMHKRELEANDI Y--- SQLKRAKI - 184 AT_ARR6 MKVEDL------------------------- SFTKSI QKRELETENVYPVHSQLKRAKI - 186 CR_RR'1' LKGGDEDGEKKTL---------- GASKVGDENRPKETRKRNLDQSSSMSLSSTLVYSPLV 191 OS_RR5 LR--------------------------------------------------- ------- 134 OS_RR4 LKRKQQLPMAAAAPDKPPHKPDEAAASAAAIAEAATAQTDGII SDCSCSGSSKRKAAAME 206 ZM_RR6 LKSRCNQEQHQ------------------- QSDSHSGERRNPTI SSSDSI NNRKRKGAGT 221 PT_RR LNK------------------------------------- GRSRDQQSTSSSNNKRKTMT 94 PpRR LNK------------------------------------- GRSRDQQX------------ 59 : .
Figura 13.- Alineamiento de las secuencias de aminoácidos de los RRs descritos en diferentes especies: Pinus pinea putative RR (PpRR), Arabidopsis thaliana (AT), Catharanthus roseus (CR), Oriza sativa (OS), Zea mays (ZM) and Pinus taeda (PT). Remarcadas en negrita aparecen las secuencias empleadas
para el diseño de los cebadores. Señalados en amarillo se muestran los residuos conservados DDK.

Informe técnico final CLONCITO 12
4.- Información científica generada
� Publicaciones Científicas Internacionales
1.- In vitro regeneration of plants from isolated cotyledons of stone pine (Pinus pinea L.). 2006. Alonso P., Moncaleán P., Fernández B., Rodríguez A., Centeno M.L., Ordás R.J. Annals of Forest Science 63:879-885. 2.-Actinidia deliciosa leaf stomatal behaviour in relation to benzyladenine incubation periods in micropropagated explants. 2006. Moncaleán P., Fernández B., Rodríguez A. New Zealand Journal of Crop and Horticultural Science (en prensa). 3.-In vitro regeneration of Pinus pinaster adult buds. 2006. De Diego N., Fernández de Larrinoa E., Moncaleán P. En preparación.
� Publicaciones Científicas Nacionales
1.-Clonación de individuos adultos del género Pinus mediante organogénesis de yemas. Revista de la Sociedad Española de cultivo in vitro de tejidos vegetales. SG. Atienza; F. Barro; J. Ballesteros; A. Martín (Eds.); Córdoba (España). pp.65.