Ciclo Celular Unidad Reproducción Celular: Ciclo Celular: Interfase y división Celular.

of 133
Upload
david-zepedaCategory
view
14download
0description
Ciclo Celular
MCs Carlos Espinoza Vzquez
Introduccin
Rudolf Virchow en el siglo XIX, las
clulas slo provienen de clulas.
Las clulas existentes se dividen a travs de una serie ordenada
de pasos denominados ciclo
celular.
La clula aumenta su tamao, el nmero de
componentes intracelulares (protenas y
organelos), duplica su material gentico y
finalmente se divide.
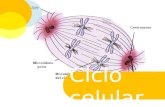
El ciclo celular se divide en dos
fases:
1. Interfase, que consta de:
Fase (S).
Fase (G1) y (G2).
2. Fase M: Mitosis.
Fase G1 y G2 (gap; intervalo)
Entre la fase S y M de cada ciclo hay 2 fases denominadas G, en las cuales la clula esta activa metabolicamente, lo cual le permite incrementar su tamao, de lo contrario las clulas se
haran ms pequeas con cada divisin.
En esta fase la clula est "quiescente" ( no est en divisin), por lo que se encuentra
fuera del ciclo celular.
Fase G0
Fase de sntesis (S)
La clula duplica su material gentico para pasarle una copia completa del genoma a cada
una de sus clulas hijas.
Fase M
Se reparte a las clulas hijas el
material gentico duplicado, a travs de la
segregacin de los cromosomas.
La fase M, para su estudio se divide
Profase: En esta etapa los cromosomas se condensan en el ncleo, mientras en el citoplasma se comienza a ensamblar el huso mittico entre los centrosomas.
Metafase: Comienza con el rompimiento de la
membrana nuclear, los cromosomas se pueden
unir al huso mittico (mediante los cinetocoros).
Una vez unidos los cromosomas estos, se
alinean en el ecuador de la clula.
Anafase: Se produce la separacin de las
cromtidas hermanas, las cuales dan lugar a
dos cromosomas hijos, los cuales
migran hacia polos opuestos de la clula.
Telofase: Ambos juegos de cromosomas llegan a los polos de la clula y adoptan una
estructura menos densa, posteriormente
se forma nuevamente la envoltura nuclear.
Citocinesis: Se divide la clula mediante el anillo contrctil de actina y miosina,
produciendo dos clulas hijas...
Cuando ya no se requieren ms clulas???, estas
entran en un estado
denominado G0, en el cual
abandonan el ciclo celular y entran en
un periodo de latencia.
Significa que entren en reposo ya que
stas clulas presentan un
metabolismo activo, pues si estas
clulas reciben el estmulo
adecuado abandonan el estado G0 y entran al
G1.
Algunas poblaciones
celulares altamente
especializadas como las
fibras musculares o
neuronas al entrar en
estado G0 abandonan
indefinidamente el ciclo
celular.
Las clulas de mamfero proliferan solo cuando son
estimuladas para hacerlo a travs de seales
intracelulares (TF) y extracelulares (GF, hormonas o
mitgenos).
Si se priva de seales
El CC se detendr en
un punto de control
G1 y la clula entrar
en el estado G0.
La clula puede
permanecer en G0
por das, semanas, o
incluso aos antes
que se divida otra vez.
Clasificacin de las clulas de acuerdo a su capacidad de proliferacin
Las clulas del organismo se agrupan segn su
capacidad proliferativa y su relacin con el ciclo
celular en:
Clulas en divisin contnua (clulas lbiles): Proliferan a lo largo de su vida
Clulas quiescentes (clulas estables):
ndice de replicacin bajo.
Pueden desarrollar una divisin rpida en respuesta a estmulos, por lo que son capaces de regenerar un
determinado tejido.
Clulas indivisibles o clulas permanentes:
Abandonan el ciclo celular y no pueden desarrollar orealizar una divisin mittica en la vida post-natal.
Regulacin del Ciclo Celular
Conjunto de procesos que ocurren durante el ciclo
celular; llevan un orden y supervisin estrictos.
Seales provenientes del medio y algunos controladores
internos, se encargan de dirigir el progreso de sta a
travs de las distintas fases del ciclo celular.
Regulacin Intracelular
Esta a cargo de
protenas, cuyas
acciones podran
resumirse en series de
activaciones e
inhibiciones de otras
protenas.
1) Las protenas que permiten el progreso del ciclo,
complejos cdk-ciclina y
2) las protenas que las inhiben,
dos pequeas familias de protenas, las CIP y las INK4.
Los complejos cdk-ciclina estn
compuestos por 2 tipos de protenas,
las cdk (cinasa dependiente de
ciclina)
Ciclinas (que pasan por un ciclo de
sntesis y degradacin)
Se conocen 6 cdk pero slo se ha caracterizado la
funcin de 4 de ellas (cdk 1, 2, 4 y 6) mientras que de las
ciclinas slo se conocen 4 tipos (ciclinas A, B, D y E).
La cdk fosforila a.a especficos de algunas protenas, pero slo si esta unida a una ciclina.
Se conocen 6 distintas combinaciones de cdk-ciclina que actan en tiempos especficos durante el ciclo.
Las clulas sintetizan protenasinhibidoras de los complejos cdk-ciclinas, que colaboran al control del ciclo celular.
Estas protenas se han agrupado en
2:
Las protenas INK4 (inhibidoras de
cinasa 4) y las CIP (protenas
inhibidoras de cdks).
Las INK4, se unen e inhiben slo los
complejos cdk4-ciclina D y cdk6-ciclina D
(p16).
Las CIP se
unen e inhiben
a todos los
complejos que
tengan cdk 1, 2,
4 y 6,
actualmente se
conocen las:
p21, p27 y p53
Todas estas protenas y algunos TF (como el p53) tienen la funcin de impedir la proliferacin
celular.
Mutaciones de los genes que las codifican resulta
en la prdida de control sobre el ciclo
celular y la incapacidad para detenerlo,
(proliferacin celular con errores).
Por su accin
normal, a los
genes que
codifican estas
protenas se les
denominaron
genes supresores de
tumores.
Estas protenas actan a diferente tiempo, permitiendo o inhibiendo el progreso adecuado del ciclo celular.
Esto se debe, a que las protenas (ciclinas), que no se utilizan, son eliminadas por un complejo de degradacin
llamado ubiquitina-proteasoma.
Para el control del ciclo celular, se postularon 4 puntos en
los que se controla a la clula y al medio extracelular para
dar lugar o restringir las acciones propias de cada una de las
fases del ciclo.
Estos 4 puntos son: 1 punto de restriccin
y 3 puntos de control.
Punto de Restriccin
Se encuentra casi al final de G1 se
conoce as puesto que si la clula lo pasa se encuentra comprometida
irreversiblemente a entrar al ciclo
celular.
Es muy importante entender, que este
punto estprincipalmente
controlado por el medio y
depende de su capacidad de
induccin, el que la
clula se comprometaa completar el CC.
Los responsables intracelulares del paso a travs de
este punto, son los complejos cdk4 y cdk6 ciclinaD, que liberan al TF (E2F) de la pRb, las cdk tienen
que fosforilar a Rb para que libere a E2F.
El E2F estimula la sntesis de: cdk2 y ciclina E
(necesarios para el progreso de G1 a S),
protenas necesarias para la sntesis de ADN y de
l mismo, inactivando an ms Rbs y disminuyendo la concentracin de p27.
La inactivacin de Rb es mantenida a lo largo del ciclo por la concentracin de distintos complejos cdk-ciclina pero, una vez que las
ciclinas se degradan, el Rb es de nuevo activo, y
une al E2F.
La p16 inhibe la unin de la cdk4 y la ciclinaD, por lo
que son inactivos; el E2F no se puede liberar y en
consecuencia no se pasa el punto de restriccin.
La accin de p16 tiene que ver con el medio
extracelular, pues se sabe que si no existen
suficientes seales del exterior (mitgenos, GF,
nutrientes, etc.) p16 y p27 tienden a acumularse, por
lo que se hacen muy activos.
La fosfoprotena p27 es una CIP, y su importancia radica
en que no slo se encarga de suprimir la actividad de los
complejos cdk-ciclinas activos en los primeros dos
puntos de control, sino que adems, ayuda a retirar a la
clula del ciclo celular llevndola a G0.
Puntos de Control
Pequeos retenes
donde se revisan
distintas caractersticas
del medio y de la
clula, la cual debe
estar sana y el medio
debe ser lo
suficientemente bueno
para que se continu el
ciclo celular.
Los controladores
implicados en estos
puntos tienen la
capacidad de
llamar a otros a reparar, cuando por
ejemplo el material
gentico est
daado, o a terminar
distintos procesos.
Primer Punto de Control
Se encuentra justo despus del punto de restriccin, an en G1.
Este se encarga de:
1) revisar las condiciones del
medio,
buscando factoresexternos que induzcan
el progreso del CC.
2) revisar que la
clula haya
crecido lo
suficiente
y
3) que el material
gentico est
intacto.
La bsqueda de factores externos es muy importante, pues
stos estimulan la sntesis de protenascomo algunas cdks y ciclinas, y sin estas, la
continuacin y el control del ciclo celular
seran imposibles.
Participan en este punto, el complejo cdk2 - ciclina E, que como los implicados en el punto de restriccin,
tambin se encarga de inactivar a Rb y de favorecer el trabajo de E2F para que estn listas las enzimas para
comenzar la sntesis de ADN en la fase S.
Los encargados de la
inhibicin en este
punto de control son
un factor de
transcripcin y una
CIP.
La p53 y la p21, en
ese orden.
La p53 es uno de las ms conocidos supresores de tumores, usualmente se encuentra en la clula pero es
muy inestable en condiciones normales porque se encuentra unido a otra protena llamada Mdm2, que
funciona como un marcador para que la p53 se degrade.
Si existe una lesin en el ADN, distintas enzimas se
activan, ayudando a separar a p53 de su marcador, por lo que una mayor concentracin de p53 estimula
la sntesis de p21 (CIP) que se une a cdk2 y ciclina E,
inhibiendo la accin del complejo.
La clula entonces no puede entrar a S.
Fase sin punto de Control
La fase S no tiene como tal un punto de control,
aun cuando algunos autores los consideran; sin
embargo es indispensable la presencia del
complejo cdk2-ciclina A para que la sntesis de
ADN se lleve a cabo.
Fase S el DNA se replica, para ello es
necesario que se arme la maquinaria especfica.
Dentro de ellas hay un conjunto de protenas
conocido como complejo de
reconocimiento del origen (ORC);
reconocensecuencias bien
definidas de bases en el ADN llamadas
orgenes de replicacin.
Durante la fase G1, se
forma el ORC, tras el
cual otras protenas
(como cdc6 y mcm) se
unen para formar el
complejo de
prereplicacin
(pre-RC).
El complejo cdk2-ciclina A se encarga de
deshacerse de las protenas del pre-RC y de unir
las enzimas necesarias para la replicacin (como
la ADN polimerasa).
Segundo Punto de Control
Se
encuentra al
final de G2.
Los complejos cdk1- ciclina A y ciclina B permiten el paso a travs de este punto.
En conjunto la actividad de estos dos complejos se denomin Factor Promotor de la Mitosis (MPF).
El segundo punto
de control se
encarga de revisar:
1) que el material
gentico se haya
duplicado
completamente.
2) que el material
gentico no tenga
errores y
3)que el medio
extracelular sea
adecuado.
Por lo tanto: los complejos cdk1-ciclina A y
ciclina B, se encargan de inducir el ensamble del
huso mittico y en parte de asegurarse de que los
cromosomas se unan a ste.
Tambin inicia la condensacin del materialgentico, activando un grupo de protenas conocidas
como condensinas, desensambla la envolturanuclear
fosforilando las lminas nucleares, arma el citoesqueleto celular y reorganiza el A.Golgi y RE.
Separacin de las Cromtidas Hermanas
Las cohesinasson protenas
requeridas para mantener unidasa las cromtidas
hermanas.
Es durante la anafase cuando las cromtides se
separan.
Para que esto suceda es necesaria la actividad de
varios complejos proteicos.
El principal de stos es el complejo promotor de la
anafase (APC).
Este complejo es activado por la unin de una protena
semejante a una cdk, llamada cdc20
(cdc= ciclo de divisin celular).
Una vez activado, el APC se encarga de marcar a diversas protenas para que se degraden, una
de ellas es la securina, que inactiva a la separasa.
Esta separasa es la
protena encargada
de inactivar a las
cohesinas
eliminando las
uniones entre las
cromtides
hermanas.
Tercer Punto de Control
Este ltimo punto
de control se
encuentra en la
fase M, entre la
metafase y la
anafase.
Se encarga de
revisar que todos
los cromosomas
se hayan unido al
huso mittico.
Si detecta que uno de los cinetocoros no se
encuentra unido, se manda una seal negativa
al sistema de control bloqueando la activacin
de protenas implicadas en la separacin de las
cromtidas hermanas.
Se inactiva al conjunto APC-cdc20, lo que
inhibe la liberacin de la separasa, impidiendo
que las cromtides hermanas se separen hasta
que la seal desaparezca.
Control Extracelular del Ciclo Celular
La forma y el tamao de un
organismo estn definidos por los
tres procesos fundamentales:
el crecimiento celular,
proliferacincelular y la muerte
celular.
La mayora de los mitgenos controlan la tasa de
divisin celular actuando en la fase G1; liberan el
control negativo del CC permitiendo
la entrada a la fase S.
Actan unindose a receptores de mem. con actividad de Tyr-K los cuales activan a la prot G monomrica Ras
cambindola de su estado unido a GDP-GTP; esta activacin desencadena una cascada de fosforilaciones
a travs de las protenas MAPK.
A su vez estas
protenas MAPK
transmiten el
estimulo a diversas
molculas
efectoras (cinasas
de protenas o TF).
Ocasionando la
trascripcin de genes
tempranos (entre los
que destacan los que
codifican a las ciclinas
de G1), algunos de
estos genes a su vez
activan la
trascripcin de
genes tardios.
De esta manera la
va de sealizacin
Ras-MAPK
transmite seales
extracelulares al
ncleo activando la
maquinaria del
ciclo celular.
Muchos tipos celulares como los fibroblastos o las
clulas epiteliales, requieren de adhesin a sustratos de
la matriz extracelular (fibronectina o laminina), para
crecer y proliferar en adicin de las seales
y medio adecuados.
Este requerimiento se debe a que la unin de molculas de la MEC a integrinas, activa otras vas de sealizacin requeridas para entrar al
ciclo celular, mediadas por la activacin de otras cinasas (FAK, cinasa de adhesin focal).
Las clulas de mamfero no se dividen infinitamente, muchas clulas se dividen un nmero limitado de
veces antes de diferenciarse en clulas especializadas.
P/e: Fibroblastos en medio de cultivo se dividen entre 25 y 50 veces, hacia el final la proliferacin disminuye su velocidad y finalmente se detiene a esto se le ha
denominado senescencia replicativa.
Apoptosis
La apoptosis a diferencia de la necrosis es un proceso ordenado, las clula muere limpiamente sin daar a sus clulas vecinas con el contenido de su citoplasma, la clula
se condensa y reduce su tamao, (cuerpo apopttico o cuerpo residual) se colapsa el citoesqueleto, la membrana nuclear se destruye, el DNA se fragmenta y finalmente la superficie de la clula cambia de manera que puede ser reconocida por clulas vecinas o macrfagos para ser
fagocitada.
La maquinara intracelular de la apoptosis depende de una familia de proteasas llamadas caspasas (cortan a la
protena blanco en residuos de aspartato).
Las caspasas se encuentran en forma de procaspasaslas cuales son activadas por un corte proteoltico,
estas a su vez activan otras procaspasas en una cascada de amplificacin.
Las caspasas cortan
protenas clave en la
clula como la
laminina que al
romperse desintegra
la mem nuclear y
degradan a la
enzima que inactiva
a la DNAsa
ocasionando la
degradacin del
material gentico.
Las seales de muerte pueden originarse a 2 niveles: en
algunas clulas se puede inducir apoptosis presentando el
ligando de Fas, el cual se une a un receptor de muerte
(Fas); el agregado de Fas y su ligando recluta a los
adaptadores que unen y activan a la procaspasa8.
La clula tambin en respuesta a dao o estrs puede
activar la apoptosis, p/e: un dao severo al DNA
puede inducirla mediante la p53, la cual activa la
trascripcin de genes que codifica para protenas que
promueven la liberacin del cit C de la mitocondria, en
el citoplasma estas se unen a Apaf1, el cual agrega
y activa a la pro-caspasa 9.
La cascada apopttica de membrana se dispara a travs de receptores de la familia de TNF tipo I (p55) que
poseen dominios de muerte.
Cuando estos receptores unen sus ligandos especficos reclutan molculas adaptadoras como FADD (Fas
Associated Death Domain) involucrada en la activacin de la Caspasa 8. En el caso de TNF, a travs de otras
molculas adaptadoras como TRAF2 (Tumor Necrosis Factor Receptor Associated Factor 2), se dispara
simultneamente una cascada protectora de apoptosis, con la activacin de NF-B, va la degradacin de su factor
inhibidor IkB y translocacin del factor activo al ncleo.
La va mitocondrial se dispara en forma directa, una vez en el citosol, el Citocromo c se asocia con
protenas adaptadoras y la pro-Caspasa 9, para formar el apoptosoma, que lleva a la activacin de
esta caspasa.
Tanto la va de membrana como la mitocondrial a travs de las caspasas iniciadoras, convergen en la activacin de
la Caspasa 3, que es una caspasa efectora que acta sobre otros substratos especficos para manifestar el
fenotipo apopttico.
Gliomas Ciclo y Muerte Celular
Los 3 tipos de
gliomas que afectan
a los adultos son:
1. Astrocitomas
2. Oligodendogliomas
3. Oligoastricitomas
El grado ms
maligno del glioma,
que puede formarse
a partir de los 3
antes mencionados;
es el glioblastoma
multiforme,
denominado por la
heterogenicidad de
sus caractersticas
anato-pato.
Las clulas malas curan a las enfermas
Lo genes supresores
demostraron, que el
crecimiento neoplsico,
era inhibido por la
formacin de un hbrido
con clulas normales.
En el material gentico
de las clulas sanas,
existan ciertos genes
capaces de suprimir el
crecimiento no
controlado celular
El guardan del Genoma y los Gliomas
1. Permite a las clulas reparar
su DNA en presencia de mutaciones
2. Destruye aquellas clulas
con dao irreparable.
En los gliomas la
mutacin de p53,
esta presente en un
40%.
Se encuentra en
astrocitomas de
grado bajo.
Se han hecho
estudios con
adenovirus para
observar el papel
de p53 en los
gliomas.
Adenovirus de
replicacin
deficiente.
Este adenoV,
transporta el ADN
complementario
del gen p53, en la
regin deletada
E1.
Las clulas del
glioma que
poseen un p53
mutado,
experimentan
La induccin de la
apoptosis,
mediada por p53;
parace
relacionada con la
elevacin de bax;
mientras que la
detencin en ciclo
celular puede esta
mediada por p21
(inh CDKs).
El gen p21 no se
ha encontrado
mutado ni en los
gliomas ni en otros
tumores.
La inactivacin de
gen p16 (del,
transloca, meti) se
ha encontrado en
el 50% de los
gliomas.
La inactivacin del
gen p16, se ha
encontrado aprox
en el 19% de los
gliomas.
La va p16-Rb, es
en donde se
encuentran las
anormalidades
genticas en los
tumores cerebrales.
La transferencia
del gen p16 o su
sustrato funcional
Rb, a clulas de
glioma que no los
expresan.
Impiden su
progresin a
travs de la fase
G1 del CC.
Este bloqueo en el
CC tiene traduccin
antitumoral, ya que
las clulas tratadas
en cultivo con p16 y
Rb, no pueden
formar colonias en
cultivos sin sustrato
para las clulas ni
tumores.
Altas [] de Rb y p16,
no se asocia con
apoptosis, pero si
puede actuar como
activador de otras
protenas
adaptadoras para la
activacin de
factores de
transcripcin que
iniciene con la
apotposis.
Estas protenas E2F pueden comportarse como genes
supresores tumorales en ciertas circunstancias.
Ratones transgnicos sin E2F pueden presentar tumores
a los 6 meses de vida (no hay apoptosis).
Tratamiento experimental en gliomas
Utiliza la
transferencia de
E2F en clulas de
glioma como una
nueva arma
teraputica para la
activacin de la
apoptosis y la
disminucin del
cncer.
Vehculos Gnicos para el Tratamiento de Cancer
Los dos ms
utilizados son:
1. Los vectores virales
2. Transfeccin gnica
En el caso de los
vactores virales los
retrovirus estn
siendo sustituidos
por los adenovirus;
ya que estos
pueden perder una
secuencia E1 que
es fcilmente
manipulable, y en
este punto se
insertar el gen a
monitorear.
Epidemiologa de la Terapia Clnica
Por indicacin
Por Vector
Por Tipo de Gen
Transfeccin Gnica Lneas Germinales