Citpkininas -Actividad y Traslocacion
22
Cytokinins: Activity, Biosynthesis, and Translocation Hitoshi Sakakibara RIKEN Plant Science Center, T surumi, Yo kohama 230-0045, Japan; email: [email protected] Annu. Rev. Plant Biol. 2006. 57:431–49 The Annual Review of Plant Biology is online at plant.annualreviews.org doi: 10.1146/ annurev.arplant.57.032905.105231 Copyright c 2006 by Annual Reviews. All rights reserved First published online as a Review in Advance on February 1, 2006 1543-5008/06/0602- 0431$20.00 Key Words adenosine phosphate-isopenten yltransferase, Agrobacterium tumefaciens , cytochrome P450 monooxygenase, plant hormones, plastid, root/shoot signaling, trans -zeatin Abstract Cytokinins (CKs) play a crucial role in various phases of plant growth and development, but the basic molecular mechanisms of their biosy nthes is and signaltransducti on only recen tly became clear. The progress was achieved by identifying a series of key genes en- coding enzymes and proteins controlling critical steps in biosynthe- sis, translocation, and signaling. Basic schemes for CK homeostasis and root/shoot communication at the whole-plant level can now be devised. This review summarizes recent findings on the relation- ship between CK structural variation and activity, distinct features in CKbio syn the sis bet weenhigherpla ntsand Agrobacterium infected plants, CK translocation at whole-plant and cellular levels, and CKs as signaling molecules for nutrient status via root-shoot communi- cation. 431 A n n u . R e v . P l a n t B i o l . 2 0 0 6 . 5 7 : 4 3 1 4 4 9 . D o w n l o a d e d f r o m w w w . a n n u a l r e v i e w s . o r g b y U n i v e r s i d a d d e l a S e r e n a o n 0 6 / 1 7 / 1 1 . F o r p e r s o n a l u s e o n l y .
-
Upload
jimmy-a-olivares-tapia -
Category
Documents
-
view
220 -
download
0
Transcript of Citpkininas -Actividad y Traslocacion

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 1/22
Cytokinins: Activity,Biosynthesis, and Translocation
Hitoshi Sakakibara
RIKEN Plant Science Center, Tsurumi, Yokohama 230-0045, Japan;email: [email protected]
Annu. Rev. Plant Biol.2006. 57:431–49
The Annual Review of Plant Biology is online atplant.annualreviews.org
doi: 10.1146/annurev.arplant.57.032905.105231
Copyright c 2006 by Annual Reviews. All rightsreserved
First published online as aReview in Advance onFebruary 1, 2006
1543-5008/06/0602-0431$20.00
Key Words
adenosine phosphate-isopentenyltransferase, Agrobacteriumtumefaciens , cytochrome P450 monooxygenase, plant hormones,
plastid, root/shoot signaling, trans -zeatin
Abstract
Cytokinins (CKs) play a crucial role in various phases of pla
growth and development, but the basic molecular mechanismstheirbiosynthesis andsignal transduction onlyrecently became cl
The progress was achieved by identifying a series of key genes e
coding enzymes and proteins controlling critical steps in biosyntsis, translocation, and signaling. Basic schemes for CK homeosta
and root/shoot communication at the whole-plant level can nowdevised. This review summarizes recent findings on the relatio
ship between CK structural variation and activity, distinct featuin CK biosynthesisbetween higherplantsand Agrobacterium infec
plants, CK translocation at whole-plant and cellular levels, and C
as signaling molecules for nutrient status via root-shoot commucation.
431

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 2/22
CK: cytokinin
tZ: trans -zeatin
Contents
INTRODUCTION.. . . . . . . . . . . . . . . . 432
STRUCTURAL VARIATION ANDBIOLOGICAL ACTIVITY . . . . . . 433
Structural Diversity of NaturalC y t o k i n i n s . . . . . . . . . . . . . . . . . . . . 4 3 3
Biological Activity . . . . . . . . . . . . . . . 433CYTOKININ BIOSYNTHESIS
AND METABOLISM. . . . . . . . . . . . 434
Basic Schemes of Cytokinin M etabol ism . . . . . . . . . . . . . . . . . . . 4 3 4
Cytokinin Biosynthesis . . . . . . . . . . . 436Regulation of Cytokinin
Biosyn th esis . . . . . . . . . . . . . . . . . . . 4 3 9
TRANSLOCATION. . . . . . . . . . . . . . . . 440Cytokinin Nucleobase Transport . . 440
Cytokinin Nucleoside Transport . . 441
Intracellular Traffic of Cytokinin . . 442
INTRODUCTION
Half a century has passed since the discovery of cytokinin (CK). Triggered by the isolation
of kinetin in autoclaved products of herringsperm DNA as a cell division promoting
factor in 1955 (4, 62, 63), a number of com-
pounds with CK activity have been identified,including trans -zeatin (tZ) as a naturally
occurring CK (54), diphenylurea as a syn-thetic compound (66, 84), and several natural
CKs with aromatic side chains (36, 93).Subsequent studies have clarified structural
requirements for CK activity. Our current
understanding is that naturally occur-ring CKs are adenine derivatives carrying
either an isoprene-derived or an aromatic sidechain at the N 6 terminus (66, 93); conven-
tionally, these families are called isoprenoidCKs and aromatic CKs, respectively. In both
groups, thereare small variations in side-chain
structure such as the absence or presence of hydroxyl groups and their stereoisomeric
position; the physiological significanceof these variations has not yet been fully
elucidated.
CK plays a crucial role in regulating theproliferation and differentiation of plant cells
and also controls various processes in plant
growth and development, such as delay osenescence (30, 67), control of shoot/root bal-
ance (112, 113), transduction of nutritionasignals (77, 80, 100), and increased crop pro-
ductivity (6). In spite of its biological andagricultural importance, only in the past few
years have the basic molecular mechanisms of
biosynthesis and signal transduction been elu-cidated. This progress was facilitated by the
identification of genes encoding enzymes andproteins controlling key steps in CK biosyn-
thesis and signaling, by in-depth analysis ofthebiochemicalproperties of theenzymes and
proteins, and by thorough determination o
the whole-plant and subcellular compartmen-
tation of CKs.Generally, CK biosynthesis andhomeostasis are finely controlled by internaand external factors such as other phytohor-
mones and inorganic nitrogen sources (77)Recent findings demonstrated that the CK
biosynthesis pathway in Agrobacterium in-
fected plants is distinct from that in higherplants in terms of substrate choice, and that
this difference is part of the surviving strategyof Agrobacterium in the host plant cells (78).
The previously widely accepted idea tha
CK and auxin are synthesized only in roottips and shoot apices, respectively, is now
overturned. Both hormones have coordinatedfunctions as long-distance messengers as wel
as local paracrine signals; they are synthesizedand act at various sites in a plant body, al-
though the physiological differentiation andthe mechanisms of the dual signaling system
have not been fully elucidated. In basipetalpo-
lar auxin transport, the AUX1/PIN-PGP sys-temtakes a major role, and themolecularbasis
of auxin translocation has been characterized(16, 70). The nature of CK translocation sys-
tems is less clear, but several lines of evidencepoint to a common translocation mechanism
for purines, nucleosides, and CKs.
In this review, I focus on activity, biosyn-thesis, translocation, and root-shoot commu-
nication, which complements recent excellen
432 Sakakibara

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 3/22
reviews on CK signaling (44) and metabolism(66).
STRUCTURAL VARIATION ANDBIOLOGICAL ACTIVITY
Structural Diversity of Natural
Cytokinins
Both isoprenoid and aromatic CKs are nat-urally occurring, with the former more fre-
quently found in plants and in greaterabundance than the latter. Common natural
isoprenoid CKs are N 6-(2-isopentenyl)-adenine (iP), tZ, cis -zeatin (cZ), and dihy-
drozeatin (DZ) (Figure 1). Among them, the
major derivatives generally are tZ and iP as well as their sugar conjugates, but there is a
lot of variation depending on plant species,tissue, and developmental stage. For instance,
tZ- and iP-type CKs are the major forms in Arabidopsis , whereas substantial amounts of
cZ-type CKs are found in maize (109), rice
(41), and chickpea (24). As for aromatic CKs,ortho-topolin (oT), meta-topolin (mT), their
methoxy-derivatives (meoT and memT, re-spectively), and benzyladenine (BA) are only
found in some plant species (93) (Figure 1).SeveralsyntheticderivativespossessCKactiv-
ity (40, 69, 86, 89) but have not been found innature so far. Human urine contains kinetin(8), but there is no evidence for its occur-
rence in plants. Usually, all natural CK nu-cleobaseshavethe corresponding nucleosides,
nucleotides, and glycosides (Figure 2).
Biological Activity
Results from classical bioassays such as thoseusing tobacco pith (81) and moss ( Funaria
hygrometrica) (92) suggested that CK nucle-obases are the active forms. In these assays,
tZ and iP generally exhibited higher activities
than cZ. tZ and iP generally exhibited higheractivity, but cZ had lower or no activity. How-
ever, the results between different bioassays were not always consistent (55). It is likely
that exogenous-applied CKs could be further
N
N
N
HN
NH
N 6 - (Δ2-isopentenyl)adenine
(iP)trans -zeatin
(tZ)
cis -zeatin
(cZ)
dihydrozeatin
(DZ)
OH
N
N
N
HN
NH
OH
N
N
N
HN
NH
OH
N
N
N
HN
NH
ortho -topolin
(oT)
meta -topolin
(mT)
N
N
N
HN
NH
OCH3
ortho -methoxytopolin(MeoT)
meta -methoxytopolin(MemT)
benzyladenine
(BA)
N
N
N
HN
NH
OH
N
N
N
HN
NH
OH
N
N
N
HN
NH
OCH3
N
N
N
HN
NH
Isoprenoid CKs
Aromatic CKs
Figure 1
Structures of representative active cytokinin (CK) species occurringnaturally. Only trivial names are given, with commonly used abbreviatioin parentheses.
www.annualreviews.org • Cytokinin Biosynthesis 433

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 4/22
R1
R6
β-D-glucose
β-D-ribose
alanine
(-SCH3)
methylthiol
β-D-glucose
β-D-glucose
N
N
N
HNR1
16
5
432
78
9R
2
β-D-ribose 5'-phosphate
β-D-glucoseβ-D-xyloseOR
6
β-D-glucose
OR6
β-D-glucoseβ-D-xyloseOR
6
OR6
β-D-glucose
R3
NR
5
R4
Figure 2
Cytokinin (CK) conjugates with sugars, sugar phosphates, and others.O-Glycosylation of side chain (colored in blue) is catalyzed by zeatinO-glucosyltransferase or O-xylosyltransferase. N -glucosylation of adeninemoiety (colored in red ) is catalyzed by cytokinin N -glucosyltransferase.
iP: N6-(2-isopentenyl)adenine
cZ: cis -zeatin
converted into other metabolites during the
experiments. The identification of CK recep-tors and the detailed analysis of their ligand
specificity helped us to better understand therelative activity of various CKs. CK receptors
in higher plants are encoded by a small gene
family (39, 97, 108, 115). Bioassays based onheterologous expression of CK receptors in
budding or fission yeast (39, 97) and in Es-
cherichia coli (91, 97, 114, 115) confirmed that
CK nucleobases are the primary ligands for
thereceptorswhereas thesugar conjugates areless active or inactive. In a heterologous assay
system, a CK receptor from maize (ZmHK1)
responded to cZ with a similar sensitivity totZ, and another one (ZmHK2) responded
to tZ riboside (tZR) as well as to tZ (115) The Arabidopsis CK receptor, AHK3, was as
sensitive to tZR and tZR 5
-monophosphate(tZRMP) as to tZ (91), suggesting that each
receptorhasaspecificspectrumofligandpref
erence (91, 114, 115). Thus, structural varia-tions of CK side chains and modifications of
the adenine moiety confer specificity of theCK-receptor interaction.
CYTOKININ BIOSYNTHESIS
AND METABOLISMBasic Schemes of Cytokinin
Metabolism
Cytokinin metabolism and homeostasis
CK metabolic pathways can be broadly clas-
sified into two types: the modification of theadenine moiety and that of the side chain. The
concomitant occurrence of CK nucleobases with the corresponding nucleosides and nu
cleotides in plant tissues suggests that impor-
tantmetabolicstepsaresharedwiththepurinemetabolic pathway [i.e., salvage pathway (22
66, 76)]. Thus, the metabolic flow from CKnucleotides to the active nucleobases is proba
bly not unidirectional but circular (Figure 3)In fact, exogenously applied CK nucleobases
are rapidly metabolized into the correspond-
ing nucleotides and nucleosides in plant tis-sues (56, 65, 88, 115). Enzymes of the purine
salvage pathway in plantsare encoded by smalmultigene families. It is now supposed that
some of the isoenzymes have broad substratespecificities, enabling them to act on CKs
as well as on authentic adenine. In this re-
gard, two isoenzymes of adenine phosphori-bosyltransferase from Arabidopsis , APT2 and
APT3, have a particularly strong preferencefor CK nucleobases as compared with other
isoenzymes (3, 83). Such preference is also
434 Sakakibara
8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 5/22
MEP pathway
DMAPP
MVA pathway
OH
prenyl-tRNA
cis -prenyl-tRNPPO
N
ONN
PO
N
OHHO
HN
N
ONN
PPO
N
OHHO
HN
N
ONN
PPPO
N
OHHO
HNOH
N
O
NN
HO
N
OHHO
HN
N
ONN
PO
N
OHHO
HN
N
ONN
PPO
N
OHHO
HN
N
ONN
PPPO
N
OHHO
HN
iPRMP
iP
iPR
iPRTP
iPRDP
tZRMP
tZ
tZR
tZRTP
tZRDP
Ade, Ado+
side chain
O -glucoside(tZOG, tZROG)
ZOGT
βGlc
N
ONN
PO
N
OHHO
HN OH
DZRMP
N
ONN
HO
N
OHHO
HN OH
DZ
DZR
N
ONN
PO
N
OHHO
HN
OH
cZRMP
cZ
cZR
CKX
Ade, Ado+
side chain
MVA pathwayMEP pathway
N -glucoside(iP7G, 9G)
OH
OH
OH
N -glucoside(Z7G, 9G)
CK-N-GTCK-N-GT
O -glucoside(cZOG, cZROG)
cZOGTβGlc
CKX
Ade, A+
side ch
βGlc
1 1
1 1
N
ONN
HO
N
OHHO
HN OH
2
5
5
7
HMBDP
6
6
AMP
ATP
ADP+
APRT
2
OH
tRNA-IPT
CKX
DMAPP+
tRNA
CYP735A
CYP735A
CYP735AIPT
IPT
IPT
AK AK AKAK
N
NH
N
N
HN
N
NH
N
N
HN
N
NH
N
N
HN
N
NH
N
N
HN
N
O
NN
HO
N
OHHO
HN
OH
3 4 3 3 4 4 3
2 2
4
6
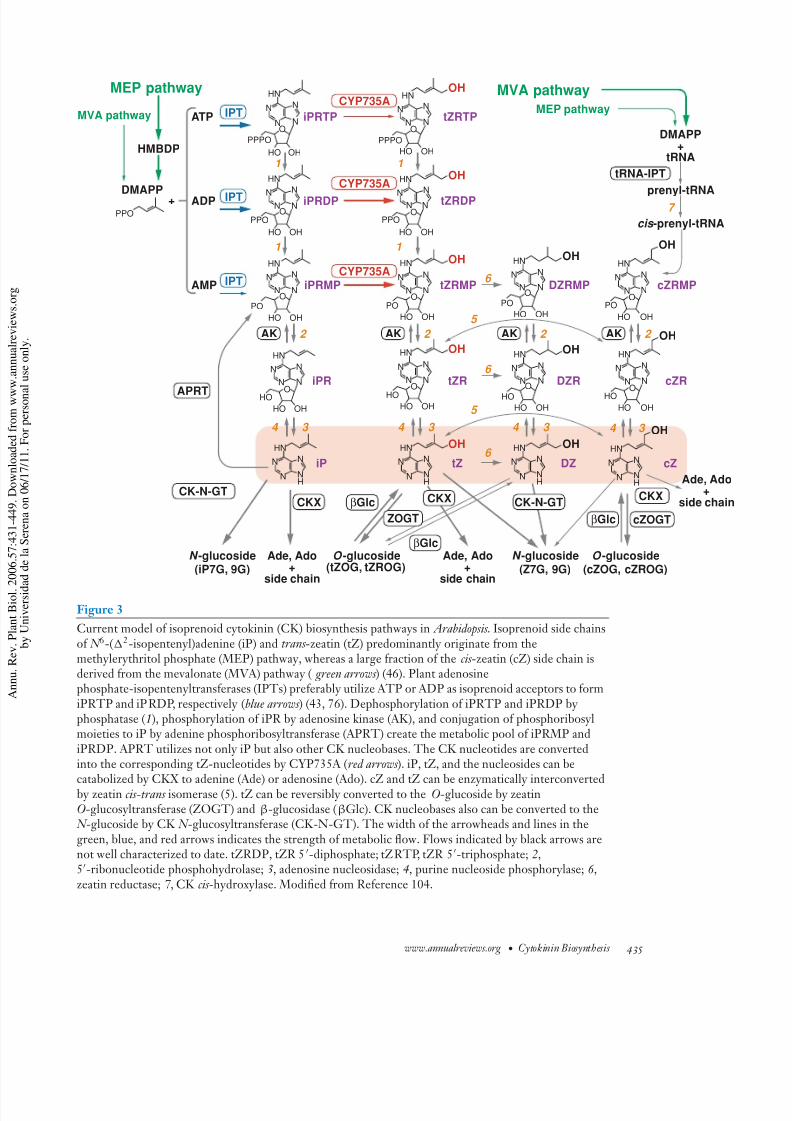
Figure 3
Current model of isoprenoid cytokinin (CK) biosynthesis pathways in Arabidopsis . Isoprenoid side chainsof N 6-(2-isopentenyl)adenine (iP) and trans -zeatin (tZ) predominantly originate from themethylerythritol phosphate (MEP) pathway, whereas a large fraction of the cis -zeatin (cZ) side chain isderived from the mevalonate (MVA) pathway ( green arrows ) (46). Plant adenosinephosphate-isopentenyltransferases (IPTs) preferably utilize ATP or ADP as isoprenoid acceptors to formiPRTP and iPRDP, respectively (blue arrows ) (43, 76). Dephosphorylation of iPRTP and iPRDP by phosphatase (1), phosphorylation of iPR by adenosine kinase (AK), and conjugation of phosphoribosylmoieties to iP by adenine phosphoribosyltransferase (APRT) create the metabolic pool of iPRMP andiPRDP. APRT utilizes not only iP but also other CK nucleobases. The CK nucleotides are convertedinto the corresponding tZ-nucleotides by CYP735A (red arrows ). iP, tZ, and the nucleosides can be
catabolized by CKX to adenine (Ade) or adenosine (Ado). cZ and tZ can be enzymatically interconvertedby zeatin cis-trans isomerase (5). tZ can be reversibly converted to the O-glucoside by zeatinO-glucosyltransferase (ZOGT) and β-glucosidase (βGlc). CK nucleobases also can be converted to the N -glucoside by CK N -glucosyltransferase (CK-N-GT). The width of the arrowheads and lines in thegreen, blue, and red arrows indicates the strength of metabolic flow. Flows indicated by black arrows arenot well characterized to date. tZRDP, tZR 5-diphosphate; tZRTP, tZR 5-triphosphate; 2,5-ribonucleotide phosphohydrolase; 3, adenosine nucleosidase; 4, purine nucleoside phosphorylase; 6 ,zeatin reductase; 7 , CK cis -hydroxylase. Modified from Reference 104.
www.annualreviews.org • Cytokinin Biosynthesis 435

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 6/22
CKX: cytokinin oxi-dase/dehydrogenase
DMAPP:dimethylallyldiphosphate
HMBDP: hydroxy-methylbutenyldiphosphate
IPT: adenosinephosphate-isopentenyl-transferase
reported from an isoform of tobacco adeno-sine kinase, ADK2S (50).
On the other hand, the enzymes for the N -glucosylation of the adenine moiety (37)and those for the hydroxylation (104), O-
glucosylation, and O-xylosylation of the sidechain (59–61) are specific for particular CK
substrates.
Glycosylation of Cytokinins. Glucosyla-
tion of CK has been observed at the N3, N7 , and N9 position of the purine moiety
as N -glucosides, and at the hydroxyl groupof the side chains of tZ, DZ, and cZ asO-glucosides or O-xylosides (Figure 2). O-glucosylation is reversible; the deglycosyla-
tion is catalyzed by β-glucosidase (18). On
the other hand, N -glucoconjugates are not
efficiently cleaved by β-glucosidase (18); asa result, N -glucosylation is practically irre- versible. The physiological consequences of
the differences in stability of N -glucosides
and O-glucosides are not fully understood todate. However, it has been suggested that the
readily cleaved O-glucosides represent inac-tive, stable storage forms of CKs.
Although genes involved in the O-glycosylation of CKs have been well char-
acterized (59–61, 68, 109), information on
the molecular biology of N -glucosylationis scarce. In Arabidopsis , two enzymes
(UGT76C1 and UGT76C2) glucosylate a se-ries of CK nucleobases at the N7 - or N9-
positions, and also tZ-O-glucoside (tZOG)at N7 (37). Both enzymes prefer glucosyla-
tion at N7 to that at N9, which corresponds
well with the concentrations of various CK N -glucosylates in Arabidopsis tissues (78).
Determinants of the levels of active Cy-
tokinins. Steady-state levels of active CK inplanta are determined by the rate of release
of CK nucleobase from the conjugates and
that of CK degradation and inactivating con- jugation. CK oxidase/dehydrogenase (CKX)
catalyzes the irreversible degradation by cleavage of the side chain (5, 28, 82), and gly-
cosyltransferases, described above, inactivate
CKs by glycosylation (Figure 3). Phosphori-bosylation of CK nucleobase by adenine phos-
phoribosyltransferase also reduces the biolog-ical activity (Figure 3). The deribosylation
of CK nucleoside to release the nucleobase
might be an important step to regulate thelevel of active CKs, but the enzymes catalyz-
ing it and their genes have not been identified yet.
Stability among CKs also has an effect on
biological activity and is determined by itsaffinity to metabolic enzymes. For instance
tZ and iP are readily catalyzed by CKXs from various plant species (13, 29), whereas cZ is
generally less amenable (13). Because CKXsrecognize the double bond of the isoprenoid
side chain, DZ and aromatic CKs are resistan
to CKX (5).
Cytokinin Biosynthesis
Primary reaction of de novo cytokinin
biosynthesis. The first step in theisoprenoidCK biosynthesis is N -prenylation of adeno-
sine 5-phosphates (AMP, ADP, or ATP) at the N 6 -terminus with dimethylallyl diphosphate
(DMAPP) or hydroxymethylbutenyl diphos-phate (HMBDP); this reaction is catalyzed by
adenosine phosphate-isopentenyltransferase
(IPT; EC 2.5.1.27) (Figure 3). It had longbeen thought that DMAPP and AMP were
the only substrates for CK biosynthesis, butnow it seems clear that substrate specificities
of IPTs vary depending on the origin and thespecies.
The first identification of substrates fo
the isoprenoid CK biosynthesis was achievedin the slime mold, Dictyostelium discoideum
which produces discadenine [3-(3-amino-3carboxypropyl)- N 6-(2-isopentenyl)adenine]
an inhibitor of spore germination (1). Thefirst step of discadenine biosynthesis is
the IPT-catalyzed prenylation of AMP. D
discoideum IPT utilizes AMP or ADP as aprenyl side-chain acceptor, but not ATP or
cyclic AMP (38, 107). The K m value for AMPis 100 nM and for DMAPP it is 2.2 μ M
(38).
436 Sakakibara

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 7/22
The first characterization of an IPT gene was carried out in Agrobacterium tumefaciens , a
crown gall-forming bacterium (2, 9). A. tume-
faciens has two IPT genes, Tmr and Tzs . Tmr
is encoded on the T-DNA region of the Ti-
plasmid and Tzs is located on the virulence re-gionof nopaline-typeTi-plasmids,whose role
is to promote T-DNA transfer efficiency (42,73). After infection, Tmr is integrated into the
host nuclear genome and functions in the host
cell,whereas Tzs actswithin the bacterialcells.Both recombinant enzymes have been puri-
fied and their kinetic parameters character-ized: The K m value of Tmr for AMP is 85 nM
(14) and those for DMAPP and HMBDPare 10.1 μ M and 13.6 μ M, respectively (78).
Tzs also utilizes HMBDP as an isoprene
donor (48), and the affinities for HMBDP
and DMAPP are similar (H. Sakakibara, un-published results). Both Tmr and Tzs donot utilize adenosine phosphates other than
AMP.
The reaction pathway of iP riboside 5-moophosphate (iPRMP) synthesis in higher
plants has been assumed to resemble thatfound in D. discoideum and A. tumefaciens . Al-
though many attempts have been made to pu-rify andcharacterize plant IPT, only a fewbio-
chemical properties have been reported (15,
21), probably due to low contents or enzymeinstability. Higher plant IPT genes have been
identifiedin Arabidopsis (43,99),petunia(116),and hop (79). In Arabidopsis , seven IPT genes
( AtIPT1 and AtIPT3 to AtIPT8) are involvedin CK biosynthesis (43, 95, 99). Biochemical
studies strongly suggest that plant IPTs pre-
dominantlyuseADPorATPratherthanAMPas prenyl acceptors, resulting in the produc-
tion of iP riboside 5-diphosphate (iPRDP)or iP riboside 5-triphosphate (iPRTP) (43,
76, 79, 103). The K m values of AtIPT1 for AMP, ADP, and ATP were 185 μ M, 14.6 μ M,
and 11.4 μ M, respectively, and for DMAPP
it was 8.3 μ M (99, 103). AtIPT4 had K m val-ues of 9.1 μ M for ADP, 3.4 μ M for ATP, and
11.6 μ M for DMAPP (43, 76). The predom-inant accumulation of iP-type CKs in trans-
genic lines of Arabidopsis that overexpress any
MEP: pathway:methylerythritolphosphate pathwa
MVA: pathway:mevalonate pathw
of the AtIPTs suggested that all AtIPTs havesimilar substrate preferences in terms of the
isoprenoid donor (78).
Although some Arabidopsis IPTs could uti-lize HMBDP as a prenyl donor in vitro, the
affinities were low and there is little evidencethat tZ-type species are formed via this reac-
tion in vivo (78, 98).
Metabolic origin of the isoprenoid side
chain. HMBDP is a metabolic intermedi-ate of the methylerythritol phosphate (MEP)
pathway, which occurs in bacteria and plastids(33). Thus, in higher plants, it is believed that
HMBDP is formed only in plastids. DMAPPis synthesized via the MEP pathway and the
mevalonate (MVA) pathway, which is com-
monly found in the cytosol of eukaryotes (57,
75). When DMAPP is used as a substrate forCK biosynthesis, the primary product is iPnucleotide (Figure 3). tZ nucleotide can be
formed directly when IPT acts on HMBDP.Before the characterization of the MEP path-
way (57, 75), the MVA pathway was the only
reaction sequence known to produce the iso-prenoid precursors DMAPP and isopentenyl
diphosphate in higher plants. In cultured to-bacco BY-2 cells, lovastatin, an inhibitor of
the MVA pathway, significantly decreases CK
accumulation (51, 52), corroborating the ideathat the MVA pathway is the metabolic ori-
gin of isoprenoid CK side chains. However,in Arabidopsis , a large number of the AtIPTs
(AtIPT1, AtIPT3, AtIPT5, and AtIPT8) arelocated in the plastids (46), and AtIPT3 and
AtIPT5 are the dominant isoforms in young
and mature plants under normal conditions(64, 102). Thus, it seems that plastids are the
major subcellular compartment for iP-typeCK biosynthesis in higher plants. Selective
labeling experiments using 13C-labeled pre-cursors specific for either the MEP or MVA
pathway demonstrate that the isoprenoid side
chain of iP- and tZ-type CKs predominantly originates from the MEP pathway (46), which
is consistent with the IPT subcellular loca-tion. On the other hand, AtIPT4 and AtIPT7
are localized in the cytosol and mitochondria,
www.annualreviews.org • Cytokinin Biosynthesis 437

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 8/22
P450: CytochromeP450monooxygenase is aheme-containingenzyme thatcatalyzes the
oxidative reaction of a wide variety of organic compoundsby utilizingatmospheric O2
respectively (46). Therefore, a major role of the MEP pathway in the biosynthesis of tZ-
and iP-type CK does not rule out a greater
contribution of the MVA pathway to the syn-thesis of these CKs under different growth
or environmental conditions, if the relativeabundance of IPT isoenzymes is modulated.
The hydroxylation step in tZ biosynthe-
sis. In higher plants, there are two pos-
sible pathways for tZ biosynthesis, the iPnucleotide-dependent and the iP nucleotide-
independent one (7, 72, 104) (Figure 3).In the iP nucleotide-dependent pathway, tZ
synthesis is catalyzed by a cytochrome P450
monooxygenase (P450); two such enzymes,CYP735A1 and CYP735A2, were recently
identified in Arabidopsis (104). Although pre-
vious studies on microsomal fractions isolatedfrom cauliflower showed trans -hydroxylationof iP and iPR only (20), CYP735A1 and
CYP735A2, which lack cis -hydroxylation ac-
tivity (104), utilize iP nucleotides but not thenucleoside and free-base forms (104). Com-
Nucleus
AgrobacteriumTi plasmid
IAA
Tms1Tms2
Cytosol
T-DNA
MEP pathway
HMBDP
DMAPP
Plastids
tZ-CK
Tumorigenesis
TmrTmr
Figure 4
Proposed mechanism for the modification of cytokinin (CK) biosynthesisby Tmr upon Agrobacterium infection. On infection, the T-DNA region istransferred to the host plant cells and integrated into the nuclear genome.
A series of genes including biosynthesis of CK (Tmr) and auxin (Tms1and Tms2) are expressed in the host plants. Among them, Tmr is targetedinto the plastids and directly produces trans -zeatin (tZ)-type CK by using1-hydroxy-2-methyl-2( E )-butenyl 4-diphosphate (HMBDP).
parison of the specificity constants (kcat / K m)for iP-nucleotides suggests that CYP735As
predominantly acts on iPRMP or iPRDP
rather than on iPRTP (104). Although thephysiological role of the CK nucleotides
has not been fully clarified, the nucleotide-specific hydroxylation indicates thattheyform
a metabolic pool for side-chain modificationsIf the CK nucleobases iP and tZ serve distinct
physiological functions that are determined
by side-chain structure, the metabolic com-partmentalization of the corresponding nu-
cleotides would be important to maintain thephysiological division of tasks at the nucle-
obase level.In the iP nucleotide-independent pathway
tZ nucleotides are assumed to be produced
directly by IPT using an unknown hydrox-
ylated side-chain precursor (7). This precur-sor probably is derived from the MVA path- way, because mevastatin, an inhibitor of tha
pathway, reduces the rate of tZ biosynthe-sis (7). Although the biochemical nature o
the iP nucleotide-independent pathway re-
mains obscure, one possible explanation isthat tZ biosynthesis via the iP nucleotide-
independent pathway might be mediated bycis-trans isomerization of cZ derivatives, in
which the prenyl side chain is primarily de-
rived from the MVA pathway (46) (Figure 3)It will be necessary to identify the postulated
isomerase (10) to understand the fabric of thepathway.
The strategy of Agrobacterium to mod
ify cytokinin biosynthesis. In vitro stud-
ies on the substrate preference of Tmr (78)and measurements of CK contents in Tmr -
overexpressing transgenic plants (7, 25, 78)and crown galls (71, 94) suggested that Tmr
utilizes HMBDP as the primary substrateand produces tZRMP. However, because Tmr
lacks any apparent sequences for subcellular
localization, it was believed that Tmr func-tions in the cytosol. However, recent work
demonstrated that Tmr is targeted to andfunctions in the plastids of infected host plant
cells (78) (Figure 4). In the stroma of the
438 Sakakibara

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 9/22
plastids, Tmr creates a CK biosynthesis by-pass using HMBDP, an intermediate of the
MEP pathway, without the requirement for
CYP735A-mediated hydroxylation (78). Thisbypass enables A. tumefaciens to produce high
amounts of tZ in order to induce tumori-genesis while the host’s CYP735A-mediated
CK hydroxylation activity could be repressedby auxin (104). Moreover, HMBDP reduc-
tase produces isopentenyl diphosphate and
DMAPP at a ratio of 5:1 (74), and these com-pounds are used as common building blocks
for all isoprenoids. Therefore, the pool sizeof HMBDP available for Tmr may be larger
than that of DMAPP in the plastids.
Aromatic cytokinins. Aromatic CKs, BA,
andtopolins (Figure 1) were identified in sev-
eral plant species including poplar (93) and Arabidopsis (106),butitisnotyetclearwhetherthey are ubiquitous in plants. Although they
exhibit strong CK activity, their biosynthesisand degradation pathways remain to be elu-
cidated. The mechanisms of glycosylation of
aromatic CKs andof their interaction with thecellular signaling system appear to be shared
with isoprenoid CKs because the enzymesand receptors involved recognize members of
both groups (39, 66, 68, 114). In tomato peri-
carp tissue, the modification of BA is greatly inhibited by metyrapone, a potent inhibitor of
P450 enzymes (58). Although details are un-known, CYP735A or some other P450s may
be involved in the biosynthesis of topolins.
A neglected pathway: cytokinins derived
from tRNA degradation. Shortly after thediscovery of CKs, it was assumed that tRNA
is a major source of CKs because isoprenoidCKs were identified in the hydrolysates of
tRNAs (45, 90, 110, 111). Several of thetRNA species with anticodons complemen-
tary to codons beginning with uridine, such
as tRNA Leu and tRNA Ser, carry a prenylatedadenosine adjacent to the anticodon. Thus,
tRNA prenylation could contribute, at leastto some extent, to CK production. The first
stepofthepathwayleadingtoCKsiscatalyzed
by tRNA-isopentenyltransferase (tRNA-IPT;EC 2.5.1.8) (32) (Figure 3). Because the
prenyl-moiety of the tRNA contains a cis -
hydroxylated group (111), tRNA-degradationis a source of cZ-type CKs; the biochemical
natureofthe cis -hydroxylating enzyme has notbeen characterized yet. A large fraction of the
cZ side chain in Arabidopsis is derived from the MVA pathway, suggesting that plants are able
toindependentlymodulatethelevelsoftZand
cZ (46). Early calculations of turnover rates of tRNA led to the conclusion that tRNA degra-
dation was not a major pathway of CK synthe-sis (47). However, tRNA-derived CKs should
not be neglected as some plant species such asmaize and rice contain substantial amounts of
cZ-type CKs.
Regulation of Cytokinin BiosynthesisSpatial expression of AtIPTs. Analyses of
spatial expression patterns of AtIPTs us-
ing their promoter::reporter genes revealedtissue- and organ-specific patterns of CK syn-
thesis by IPT (64, 102). AtIPT1 is expressedin xylem precursor cell files in root tips, leaf
axils, ovules, and immature seeds; AtIPT3 isexpressed in phloem companion cells; AtIPT4
and AtIPT8 are expressed in immature seeds
with highest expression in the chalazal en-dosperm; AtIPT5 is expressed in lateral root
primordia, columella root caps, upper partsof young inflorescences, and fruit abscission
zones; AtIPT6 is expressed in siliques; AtIPT7
is expressed in phloem companion cells, theendodermis of the root elongation zones, tri-
chomes on young leaves, and occasionally inpollen tubes. CK biosynthesis in aerial organs
was also confirmed by in vivo deuterium la-beling methods (72).
Regulation by plant hormones. The ex-
pression of key genes for CK biosynthe-
sis and homeostasis such as IPT , CKX , andCYP735A is regulated by phytohormones in-
cluding CKs, auxin, and abscisic acid (ABA).In Arabidopsis , the accumulation of the tran-
scripts of AtIPT5 and AtIPT7 is promoted by
www.annualreviews.org • Cytokinin Biosynthesis 439

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 10/22
auxin in roots, whereas the transcript levelsof AtIPT1, AtIPT3, AtIPT5 , and AtIPT7 are
negatively regulated by CK (64). On the other
hand, the expression of both CYP735A1 andCYP735A2 is upregulated by CKs in roots but
downregulated by auxin or ABA (104). Genesfor CKX in maize are upregulated by CK and
ABA (17). These regulation patterns suggestthatthe enzymes antagonistically regulate cel-
lular CK levels and the balance between iP
andtZ, which interact with auxin and/or ABA. The interdependent regulation of phytohor-
mones might provide a basis for the variablemorphogenetic responses of plants to envi-
ronmental factors.
Cytokinins as a local signal. In the con-
trol of outgrowth and dormancy of axillary
buds, the mutual regulation of auxin, ABA,and CKs has been proposed to play a cen-tral role (85). In pea, the expression of two
IPT genes, PsIPT1 and PsIPT2, which are ex-pressed in nodes, is negatively regulated by
auxin (105). Although an antagonistic role of
auxin andCKs in theregulation of axillary budoutgrowth has been postulated for a consid-
erable time, little is known about the under-lying molecular mechanisms. Recent studies
revealed that one role of apex-derived auxin
in apical dominance is to repress CK biosyn-thesis in the nodes and that after decapitation
CKs are locally synthesized in the stem ratherthan being transported to the stem from the
roots (105).
Regulation by nitrogen supply. Inorganic
nitrogen strongly affects plant growth and de- velopment. Plants constantly sense the nutri-
ent status and modulate their metabolic ac-tivities and developmental program to adapt
efficientlyto the nutritional environment. CK is a pivotal signaling substance communicat-
ing the nitrogen nutrient status from root to
shoot via the xylem vessels (87, 100, 101).Recent studies reveal a molecular mecha-
nism of nitrogen-dependent CK biosynthe-sis and regulation (Figure 5); AtIPT3 and
AtIPT5 are regulated differentially depend-
ing on the nitrogen sources available (64102). AtIPT3 rapidly and specifically responds
to NO3− under nitrogen-limited conditions
whereas AtIPT5 responds to both NO3− and
NH4+ under long-term treatment (102). This
dual-response system might be importanfor plants coping with unpredictably chang-
ing nitrogen availability. AtIPT3 and AtIPT5are the dominant IPTs in Arabidopsis under
normal conditions (64, 102); they are lo
calized in plastids (46), strongly suggestingthat CK biosynthesis is affected by nitrogen
sources. In a Ds transposon-insertion mutanof AtIPT3, NO3
−-dependent CK accumula-
tion was greatly reduced (102), indicating that AtIPT3 is a key determinant of CK biosynthe-
sis in response to rapid changes in NO3− in
the soil.
Cytokinins as a long-range signal. Expres-
sion of CYP735A2 in roots is strongly upregu-lated by CK application (104). In this contex
the fact that AtIPT3 is upregulated by NO3−
seems to explain the observation that apply-
ing NO3− induces tZ-nucleotide accumula-
tioninroots(100)(Figure 5).tZRisthemajor
species of CKs in the xylem (53, 80, 100). In-
terestingly, its translocation rate in xylem ves-sels is controlled by NO3
− in the root medium
(100). Thus, xylem-mediated signaling by tZ-type CKs in the transpiration stream appears
important for the communication of nutri-ent signals on the whole-plant level. How-
ever, leaf exudates contain mainly iP-type
CKs(23).NotethatNO3−-dependentexpres
sion of AtIPT3 occurs in phloem tissue (64
102). One possible explanation is that trans -hydroxylation controls the differential com-
partmentalization of CK species and thus alsocontrols the direction of CK translocation, al
though the detailed mechanisms remain to beelucidated.
TRANSLOCATION
Cytokinin Nucleobase Transport
Because de novo CK biosynthesiscatalyzed by
IPTs is tissue- and cell-specific, the CKs must
440 Sakakibara
8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 11/22
Leaf
Root
Vascular System
Phloem
AtIPT3/ AtIPT5 CYP735AiP-nuc.tZ-nuc.tZR
NO3-, (NH
4+)NO
3-
NO3-
tZR
N-assimilatory genes& others (e.g. AtIPT3 )
tZ His-Asp P-relay system
Xylem
iP-nuc.iPRAtIPT3
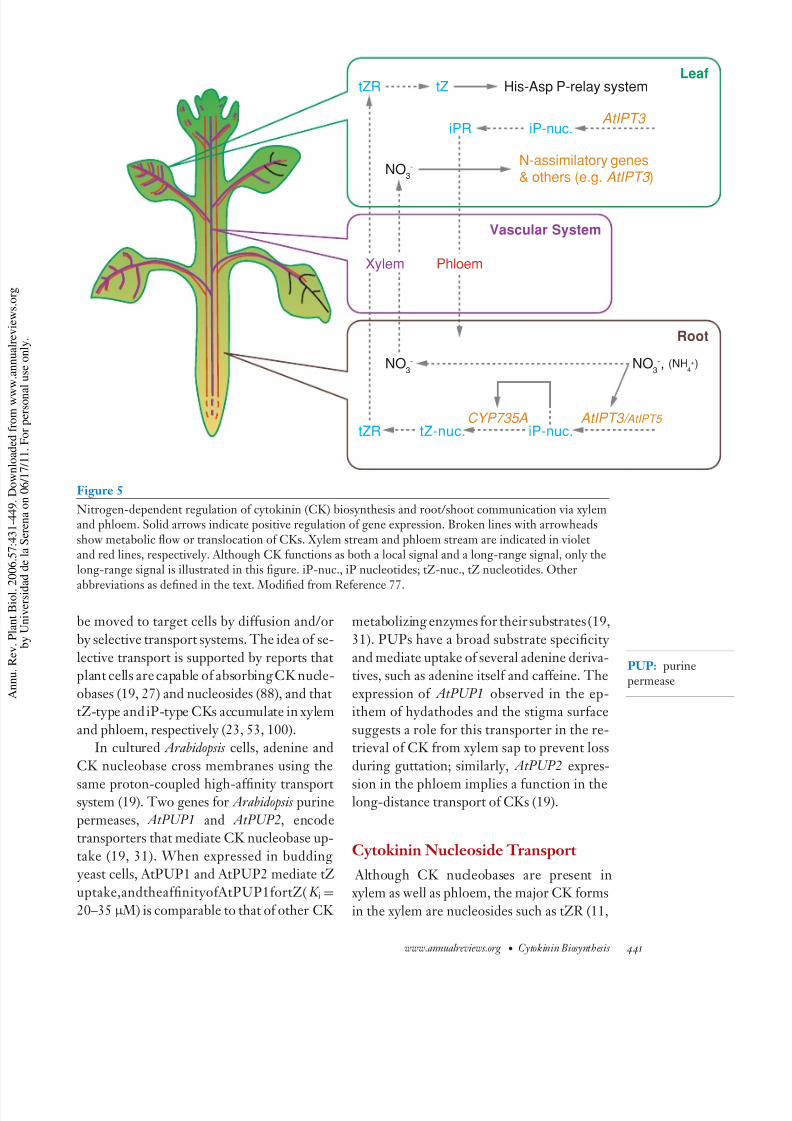
Figure 5
Nitrogen-dependent regulation of cytokinin (CK) biosynthesis and root/shoot communication via xylemand phloem. Solid arrows indicate positive regulation of gene expression. Broken lines with arrowheadsshow metabolic flow or translocation of CKs. Xylem stream and phloem stream are indicated in violetand red lines, respectively. Although CK functions as both a local signal and a long-range signal, only thelong-range signal is illustrated in this figure. iP-nuc., iP nucleotides; tZ-nuc., tZ nucleotides. Other
abbreviations as defined in the text. Modified from Reference 77.
be moved to target cells by diffusion and/or
by selective transport systems. The idea of se-
lective transport is supported by reports thatplant cells are capable of absorbing CK nucle-
obases (19, 27) and nucleosides (88), and thattZ-type and iP-type CKs accumulate in xylem
and phloem, respectively (23, 53, 100).In cultured Arabidopsis cells, adenine and
CK nucleobase cross membranes using the
same proton-coupled high-affinity transportsystem (19). Two genes for Arabidopsis purine
permeases, AtPUP1 and AtPUP2, encodetransporters that mediate CK nucleobase up-
take (19, 31). When expressed in budding yeast cells, AtPUP1 and AtPUP2 mediate tZ
uptake,andtheaffinityofAtPUP1fortZ( K i =
20–35 μ M) is comparable to that of other CK
PUP: purinepermease
metabolizing enzymes for their substrates(19,
31). PUPs have a broad substrate specificity
and mediate uptake of several adenine deriva-tives, such as adenine itself and caffeine. The
expression of AtPUP1 observed in the ep-ithem of hydathodes and the stigma surface
suggests a role for this transporter in the re-trieval of CK from xylem sap to prevent loss
during guttation; similarly, AtPUP2 expres-
sion in the phloem implies a function in thelong-distance transport of CKs (19).
Cytokinin Nucleoside Transport
Although CK nucleobases are present in xylem as well as phloem, the major CK forms
in the xylem are nucleosides such as tZR (11,
www.annualreviews.org • Cytokinin Biosynthesis 441

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 12/22
ENT: Theequilibratednucleosidetransporter (ENT)catalyzes thetransport of various
kinds of nucleosidesdown theirconcentration
12, 100). In addition, the mobility of kinetinriboside in the xylem exceeds that of BA and
kinetin (34). Therefore, nucleosides are con-
sidered the major translocation form of CKs.In higher plants, some members of the equili-
brative nucleoside transporter (ENT) family appear to mediate the selective translocation
of CK nucleosides. The rice genome containsfour ENT genes (OsENT1 - OsENT4); one
of the gene products, OsENT2, mediates the
uptake of CK nucleosides as well as that of adenosine (35). OsENT2 prefers iPR ( K m =
32 μ M) over tZR ( K m = 660 μ M), suggest-ing that it may be responsible in part for the
selective transport of CK nucleosides in the vascular tissues (35). OsENT2 expression was
detected in the scutellum during germina-
tion and in the vascular tissues in germinated
seedlings, suggesting that OsENT2 partici-pates in the retrieval of endosperm-derivednucleosides in the germinating embryo and in
the long-distance transport of nucleosides in
growing plants (35). In Arabidopsis , an ENTgene, SOI33/AtENT8, was also suggested to
function in CK nucleoside transport (96). Insummary, our current understanding of CK
translocation in planta is that CK transportis achieved by the translocation systems that
also mediate the transport of purine deriva-
tives and nucleosides.Compared with the highly elaborated po-
lar transportation system for auxin, CK trans-
port appears somewhat unspecific. Howeverdifferential loading into xylem or phloem
might be sufficient for acropetal and sys-
temic transport of CKs, respectively. It is in-teresting, though, that CK transport seems
to occur in parallel with that of fundamen-tally important building blocks of the ge
netic apparatus and the energy transductionmachinery, such as purine and nucleosides
which may be related to the CKs’ function
as a signal in the coordination of growth anddevelopment.
Intracellular Traffic of Cytokinin
Plastids are the major subcellular compart-
ments of de novo CK biosynthesis throughIPT (46, 102). This leads us to speculate on a
transport system of CKs from plastids to thecytosol. At present, we have no evidence as
to which form of CKs is transported across
the plastid membranes. There is a report that AtIPT7 is localized in mitochondria (46). CK
glycosides accumulate in the vacuole (26), andthe deglucosylating enzyme,β-glucosidase, is
localized in the plastids (49). Such complex-ity in the subcellular location of CK biosyn-
thesis and metabolism points to an intricate
intracellular CK transport network. It is stil
unknown whether the intracellular traffic oCK is also based on a transporting system forstructurally related compounds.
SUMMARY POINTS
1. Structural variations at the side chain of isoprenoid and aromatic CKs affect the
interaction with CK receptors, perhaps signifying functional specificity.
2. The initial step of CK biosynthesis (iP nucleotide synthesis) is catalyzed by adenosine
phosphate-isopentenyltransferase; plastids are the major subcellular compartment for
this initial step.3. trans -Hydroxylation in tZ biosynthesis is catalyzed by a P450 enzyme, CYP735A.
The hydroxylation predominantly occurs at the nucleotide step.
4. Tmr of Agrobacterium is targeted to the plastids of infected cells despite lacking a
typical plastid-targeting sequence, and creates a CK biosynthesis bypass using anintermediate in the plastid-specific MEP pathway to synthesize tZ.
442 Sakakibara

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 13/22
5. CK translocation shares the same transport systems moving purines and nucleosidesat the whole-plant and perhaps at the cellular level.
6. Root/shoot signaling may be related to movement of tZ and iP derivatives, respec-tively, in the xylem and phloem streams.
7. CK biosynthesis and homeostasis are fine-tuned by internal and external factors such
as phytohormones and inorganic nitrogen sources, which appears importantin linkingnutrient signals and morphogenetic responses.
FUTURE ISSUES TO BE RESOLVED
1. Identification of genes involved in cZbiosynthesisand cZ-tZ isomerization: It is essen-
tial to understand the physiological function of cZ and the physiological significanceof tRNA-derived CK production.
2. Identification and characterization of genes encoding CK nucleosidases: Because CK nucleobase is the active form, their release from the nucleobases is one of the critical
steps to control active CK level.
3. Analysis of knockout mutants of genes involved in side-chain modification:It is impor-tant to understand the possible unique properties and physiological roles of different
isoprenoid and aromatic CKs.
4. Determine the molecular mechanism of Tmr import into the plastid: It would be of
interest to know if the Toc-Tic system is involved in the translocation.
ACKNOWLEDGMENTS
The author acknowledges Dr. David W. S. Mok for his critical reading of the manuscript
and for giving helpful comments. The writing of this review and research conducted in theauthor’s laboratory are supported by the Ministry of Education, Culture, Sports, Science, and
Technology and the Ministry of Agriculture, Forestry, and Fisheries, Japan.
LITERATURE CITED
1. Abe H, Uchiyama M, Tanaka Y, Saito H. 1976. Structure of discadenine, a spore ger-mination inhibitor from cellular slime-mold, Dictyostelium discoideum. Tetrahedron Lett.
42:3807–10
2. Akiyoshi DE, Klee H, Amasino RM, Nester EW, Gordon MP. 1984. T-DNA of Agrobac-terium tumefaciens encodes an enzyme of cytokinin biosynthesis. Proc. Natl. Acad. Sci. USA
81:5994–98
3. AllenM,QinW,MoreauF,MoffattB.2002.Adeninephosphoribosyltransferaseisoformsof Arabidopsis and their potential contributions to adenine and cytokinin metabolism.
Physiol. Plant. 115:56–68
4. Amasino R. 2005. 1955: kinetin arrives. The 50th anniversary of a new plant hormone. Plant Physiol. 138:1177–84
www.annualreviews.org • Cytokinin Biosynthesis 443

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 14/22
5. Armstrong DJ. 1994. Cytokinin oxidase and the regulation of cytokinin degradation. InCytokinins: Chemistry, Activity, and Function, ed. DWS Mok, MC Mok, pp. 139–54. Boca
Raton, Florida: CRC Press
6. Ashikari M, Sakakibara H, Lin S, Yamamoto T, Takashi T, et al. 2005. Cytokinin oxidase
regulates rice grain production. Science 309:741–45
7. ˚ AstotC,DolezalK,Nordstr omA,WangQ,KunkelT,etal.2000.Analternativecytokininbiosynthesis pathway. Proc. Natl. Acad. Sci. USA 97:14778–83
8. Barciszewski J, Mielcarek M, Stobiecki M, Siboska G, Clark BF. 2000. Identification o6-furfuryladenine (kinetin) in human urine. Biochem. Biophys. Res. Commun. 279:69–73
9. Barry GF, Rogers SG, Fraley RT, Brand L. 1984. Identification of a cloned cytokininbiosynthetic gene. Proc. Natl. Acad. Sci. USA 81:4776–80
10. Bassil NV, Mok D, Mok MC. 1993. Partial purification of a cis-trans -isomerase of zeatinfrom immature seed of Phaseolus vulgaris L. Plant Physiol. 102:867–72
11. Beck E, Wagner BM. 1994. Quantification of the daily cytokinin transport from the rootto the shoot of Urtica dioica L. Bot. Acta 107:342–48
12. Beveridge CA, Murfet IC, Kerhoas L, Sotta B, Miginiac E, Rameau C. 1997. The shoo
controls zeatin riboside export from pea roots. Evidence from the branching mutantrms4
Plant J. 11:339–45
13. Bilyeu KD, Cole JL, Laskey JG, Riekhof WR, Esparza TJ, et al. 2001. Molecular andbiochemical characterization of a cytokinin oxidase from maize. Plant Physiol. 125:378–86
14. Blackwell JR, Horgan R. 1993. Cloned Agrobacterium tumefaciens ipt1 gene productDMAPP:AMP isopentenyl transferase. Phytochemistry 34:1477–81
15. Blackwell JR, Horgan R. 1994. Cytokinin biosynthesis by extracts of Zea mays . Phyto-
chemistry 35:339–42
16. Blakeslee JJ, Peer WA, Murphy AS. 2005. Auxin transport. Curr. Opin. Plant Biol. 8:494–500
17. Brugiere N, Jiao S, Hantke S, Zinselmeier C, Roessler JA, et al. 2003. Cytokinin oxidase
gene expression in maize is localized to the vasculature, and is induced by cytokininsabscisic acid, and abiotic stress. Plant Physiol. 132:1228–40
18. Brzobohaty B, Moore I, Kristoffersen P, Bako L, Campos N, et al. 1993. Release of activecytokinin by a beta-glucosidase localized to the maize root meristem. Science 262:1051–54
This articlereported the first characterization of purine permeasesinvolved in transport of CK nucleobases.
19. Burkle L, Cedzich A, Dopke C, Stransky H, Okumoto S, et al. 2003. Transport of
cytokinins mediated by purine transporters of the PUP family expressed in phloem
hydathodes, and pollen of Arabidopsis . Plant J. 34:13–26
20. Chen C-M, Leisner SM. 1984. Modification of cytokinins by cauliflower microsomal
enzymes. Plant Physiol. 75:442–46
21. Chen C-M, MelitzDK. 1979.Cytokinin biosynthesis in a cell-freesystemfrom cytokinin-
autotrophic tobacco tissue cultures. FEBS Lett. 107:15–20
22. Chen CM. 1997. Cytokinin biosynthesis and interconversion. Physiol. Plant. 101:665–73
23. Corbesier L, Prinsen E, Jacqmard A, Lejeune P, Van Onckelen H, et al. 2003. Cytokininlevels in leaves, leaf exudate and shoot apical meristem of Arabidopsis thaliana during floratransition. J. Exp. Bot. 54:2511–17
24. Emery RJN, Leport L, Barton JE, Turner NC, Atkins A. 1998. cis -Isomers of cytokininspredominate in chickpea seeds throughout their development. Plant Physiol. 117:1515–23
25. Faiss M, Zalubilov a J, Strnad M, Schm ulling T. 1997. Conditional transgenic expressionof the ipt gene indicates a function for cytokinins in paracrine signaling in whole tobacco
plants. Plant J. 12:401–15
444 Sakakibara

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 15/22
26. Fusseder A, Ziegler P. 1988. Metabolism and compartmentation of dihydrozeatin exoge-nously supplied to photoautotrophic suspension-cultures of Chenopodium rubrum. Planta
173:104–9
27. Fusseder A, Ziegler P, Peters W, Beck E. 1989. Turnover of O-glucosides of dihy-drozeatin and dihydrozeatin-9-riboside during the cell-growth cycle of photoautotrophic
cell-suspension cultures of Chenopodium rubrum. Bot. Acta 102:335–4028. Galuszka P, Frebort I, Sebela M, Sauer P, Jacobsen S, Pec P. 2001. Cytokinin oxidase or
dehydrogenase? Mechanism of cytokinin degradation in cereals. Eur. J. Biochem. 268:450–61
29. Galuszka P, Frebortova J, Werner T, Yamada M, Strnad M, et al. 2004. Cytokinin oxi-
dase/dehydrogenase genes in barley and wheat. Eur. J. Biochem. 271:3990–400230. Gan S, Amasino RM. 1995. Inhibition of leaf senescence by autoregulated production of
cytokinin. Science 270:1986–8831. Gillissen B, Burkle L, Andre B, Kuhn C, Rentsch D, et al. 2000. A new family of high-
affinity transporters for adenine, cytosine, and purine derivatives in Arabidopsis . Plant Cell
12:291–300
32. Golovko A, Sitbon F, Tillberg E, Nicander B. 2002. Identification of a tRNA isopen-
tenyltransferase gene from Arabidopsis thaliana. Plant Mol. Biol. 49:161–69
33. Hecht S, Eisenreich W, Adam P, Amslinger S, Kis K, et al. 2001. Studies on the non-mevalonate pathway to terpenes: the role of the GcpE (IspG) protein. Proc. Natl. Acad.
Sci. USA 98:14837–42
34. Hill-Cottingham DG, Lloyd-Jones CP. 1968. Relative mobility of some organic nitroge-nous compounds in the xylem of apple shoots. Nature 220:389–90
This articlereported the firsidentification ancharacterization rice ENT genesinvolved in transport of CK
nucleosides.
35. Hirose N, Makita N, Yamaya T, Sakakibara H. 2005. Functional characterization
and expression analysis of a gene, OsENT2, encoding an equilibrative nucleoside
transporter in rice suggest a function in cytokinin transport. Plant Physiol. 138:196–
206
36. Horgan R, Hewett EW, Purse JG, Wareing PF. 1973. A new cytokinin from Populus
robusta. Tetrahedron Lett. 14:2827–28
37. Hou B, Lim EK, Higgins GS, Bowles DJ. 2004. N -Glucosylation of cytokinins by gly-cosyltransferases of Arabidopsis thaliana. J. Biol. Chem. 279:47822–32
38. Ihara M, Taya Y, Nishimura S, Tanaka Y. 1984. Purification and some properties of delta 2-isopentenylpyrophosphate:5 AMP delta 2-isopentenyltransferase from the cellu-
lar slime mold Dictyostelium discoideum. Arch. Biochem. Biophys. 230:652–6039. Inoue T, Higuchi M, Hashimoto Y, Seki M, Kobayashi M, et al. 2001. Identification of
CRE1 as a cytokinin receptor from Arabidopsis . Nature 409:1060–63
40. Iwamura H. 1994. Cytokinin antagonists: synthesis and biological activity. In Cytokinins:
Chemistry, Activity, and Function, ed. DWS Mok, MC Mok, pp. 43–55. Boca Raton,
Florida: CRC Press41. Izumi K, Nakagawa S, Kobayashi M, Oshio H, Sakurai A, Takahashi N. 1988. Levels of
IAA, cytokinins, ABA and ethylene in rice plants as affected by a gibberellin biosynthesisinhibitor, uniconazole-P. Plant Cell Physiol. 29:97–104
42. John MC, Amasino RM. 1988. Expression of an Agrobacterium Ti plasmid gene involved
in cytokinin biosynthesis is regulated by virulence loci and induced by plant phenoliccompounds. J. Bacteriol. 170:790–95
This articlereported the firsidentification ancharacterization Arabidopsis IPT genes.
43. Kakimoto T. 2001. Identification of plant cytokinin biosynthetic enzymes as
dimethylallyl diphosphate:ATP/ADP isopentenyltransferases. Plant Cell Physiol.
42:677–85
www.annualreviews.org • Cytokinin Biosynthesis 445

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 16/22
44. Kakimoto T. 2003. Perception and signal transduction of cytokinins. Annu. Rev. Plan
Biol. 54:605–2745. Kaminek M. 1982. Mechanisms preventing the interference of tRNA cytokinins in
hormonal regulation. In Plant Growth Substances 1982, ed. PF Wareing, pp. 215–23
New York: Academic
This article first reported that the
side chain of iP-and tZ-type CKs isderived from the MEP pathway.
46. Kasahara H, Takei K, Ueda N, Hishiyama S, Yamaya T, et al. 2004. Distinct iso-
prenoid origins of cis - and trans -zeatin biosyntheses in Arabidopsis . J. Biol. Chem
279:14049–5447. Kl ambt D. 1992. The biogenesis of cytokinins in higher plants: our present knowl-
edge. In Physiology and Biochemistry of Cytokinins in Plants , ed. M Kaminek, DWS Mok
E Zazımalov a, pp. 25–27. The Hague: SPB Academic48. Krall L, Raschke M, Zenk MH, Baron C. 2002. The Tzs protein from Agrobacterium
tumefaciens C58 produces zeatin riboside 5-phosphate from 4-hydroxy-3-methyl-2-( E )-butenyl diphosphate and AMP. FEBS Lett. 527:315–18
49. Kristoffersen P, Brzobohaty B, Hohfeld I, Bako L, Melkonian M, Palme K. 2000. De- velopmental regulation of the maize Zm-p60.1 gene encoding a beta-glucosidase located
to plastids. Planta 210:407–1550. Kwade Z, Swiatek A, Azmi A, Goossens A, Inze D, et al. 2005. Identification of four
adenosine kinase isoforms in tobacco BY-2 cells and their putative role in the cell cycle-
regulated cytokinin metabolism. J. Biol. Chem. 280:17512–1951. Laureys F, Dewitte W, Witters E, Van Montagu M, Inze D, Van Onckelen H. 1998
Zeatin is indispensable for the G2-M transition in tobacco BY-2 cells. FEBS Lett. 426:29–
3252. Laureys F, Smets R, Lenjou M, Van Bockstaele D, Inze D, Van Onckelen H. 1999. A low
content in zeatin type cytokinins is not restrictive for the occurrence of G1 /S transitionin tobacco BY-2 cells. FEBS Lett. 460:123–28
53. Lejeune P, Bernier G, Requier M-C, Kinet J-M. 1994. Cytokinins in phloem and xylemsaps of Sinapis alba during floral induction. Physiol. Plant. 90:522–28
54. Letham DS. 1963. Zeatin, a factor inducing cell division from Zea mays . Life Sci 8:569–7355. Letham DS, Palni LMS, Tao G-Q, Gollnow BI, Bates CM. 1983. Regulators of cell divi-
sion in plant tissues XXIX. The activities of cytokinin glucosides and alanine conjugatesin cytokinin bioassays. J. Plant Growth Regul. 2:103–15
56. Letham DS, Zhang R. 1989. Cytokinin translocation and metabolism in lupin species. IINew nucleotide metabolites of cytokinins. Plant Sci. 64:161–65
57. Lichtenthaler HK. 1999. The 1-deoxy-D-xylulose-5-phosphate pathway of isoprenoidbiosynthesis in plants. Annu. Rev. Plant Physiol. Plant Mol. Biol. 50:47–65
58. Long AR, Chism GW 3rd. 1987. The effect of metyrapone on cytokinin([8-14C]benzylaminopurine) metabolism in mature green tomato pericarp. Biochem
Biophys. Res. Commun. 144:109–1459. Martin RC, Mok MC, Habben JE, Mok DW. 2001. A maize cytokinin gene encoding an
O-glucosyltransferase specific to cis -zeatin. Proc. Natl. Acad. Sci. USA 98:5922–26
60. Martin RC, Mok MC, Mok DW. 1999. A gene encoding the cytokinin enzyme zeatinO-xylosyltransferase of Phaseolus vulgaris . Plant Physiol. 120:553–58
61. Martin RC, Mok MC, Mok DW. 1999. Isolation of a cytokinin gene, ZOG1, encoding
zeatin O-glucosyltransferase from Phaseolus lunatus . Proc. Natl. Acad. Sci. USA 96:284–8962. Miller CO, Skoog F, Okumura FS, von Saltza MH, Strong FM. 1955. Structure and
synthesis of kinetin. J. Am. Chem. Soc. 78:2662–6363. Miller CO, Skoog F, Saltza vNH, Strong M. 1955. Kinetin, a cell division factor from
deoxyribonucleic acid. J. Am. Chem. Soc. 77:1329–34
446 Sakakibara

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 17/22
This articlereported the firscharacterization expression patteof AtIPT genes.
64. Miyawaki K, Matsumoto-Kitano M, Kakimoto T. 2004. Expression of cytokinin
biosynthetic isopentenyltransferase genes in Arabidopsis : tissue specificity and reg-
ulation by auxin, cytokinin, and nitrate. Plant J. 37:128–38
65. Moffatt B, Pethe C, Laloue M. 1991. Metabolism of benzyladenine is impaired in a
mutant of Arabidopsis thaliana lacking adenine phosphoribosyltransferase activity. Plant
Physiol. 95:900–8
66. Mok DW, Mok MC. 2001. Cytokinin metabolism and action. Annu. Rev. Plant Physiol.
Plant Mol. Biol. 52:89–11867. Mok MC. 1994. Cytokinins andplant development–An overview.In Cytokinins: Chemistry,
Activity, and Function, ed. DWS Mok, MC Mok, pp. 155–66. Boca Raton, Florida: CRC
Press
68. Mok MC, Martin RC, Dobrev PI, Vankova R, Ho PS, et al. 2005. Topolins and hy-
droxylated thidiazuron derivatives are substrates of cytokinin O-glucosyltransferase withposition specificity related to receptor recognition. Plant Physiol. 137:1057–66
69. Mok MC, Mok DWS, Armstrong DJ, Shudo K, Isogai Y, Okamoto T. 1982. Cytokininactivityof N -phenyl- N’ -1,2,3-thiadiazol-5-ylurea (Thidiazuron). Phytochemistry 21:1509–
11
70. Morris DA, Friml J, Zazimalova E. 2005. The transport of auxin. In Plant Hormones:
Biosynthesis, Signal Transduction, Action! , ed. PJ Davies, pp. 437–70. Dordrecht: Kluwer Academic
71. Morris RO. 1986. Genes specifying auxin and cytokinin biosynthesis in phytopathogens. Annu. Rev. Plant Physiol. Plant Mol. Biol. 37:509–38
72. Nordstr om A, Tarkowski P, Tarkowska D, Norbaek R, ˚ Astot C, et al. 2004. Auxin reg-
ulation of cytokinin biosynthesis in Arabidopsis thaliana: A factor of potential importance
for auxin-cytokinin-regulated development. Proc. Natl. Acad. Sci. USA 101:8039–44
73. Powell GK, Hommes NG, Kuo J, Castle LA, Morris RO. 1988. Inducible expression of cytokinin biosynthesis in Agrobacterium tumefaciens by plant phenolics. Mol. Plant Microbe
Interact. 1:235–42
74. Rohdich F, Hecht S, Gartner K, Adam P, Krieger C, et al. 2002. Studies on the non-
mevalonate terpene biosynthetic pathway: metabolic role of IspH (LytB) protein. Proc.
Natl. Acad. Sci. USA 99:1158–63
75. Rohmer M. 1999. The discovery of a mevalonate-independent pathway for isoprenoidbiosynthesis in bacteria, algae and higher plants. Nat. Prod. Rep. 16:565–74
76. Sakakibara H. 2004. Cytokinin biosynthesisandmetabolism. In PlantHormones: Biosynthe-
sis, Signal Transduction, Action! , ed. PJ Davies, pp. 95–114. Dordrecht: Kluwer Academic
77. Sakakibara H. 2005. Cytokinin biosynthesis and regulation. Vit. Hor. 72:271–87
This articlereported the firsidentification of
plastid-location Tmr in the host plant cell.
78. Sakakibara H, Kasahara H, Ueda N, Kojima M, Takei K, et al. 2005. Agrobac-
terium tumefaciens increases cytokinin production in plastids by modifying the
biosynthetic pathway in the host plant. Proc. Natl. Acad. Sci. USA 102:9972–77
79. Sakano Y, Okada Y, Matsunaga A, Suwama T, Kaneko T, et al. 2004. Molecular cloning,expression, and characterization of adenylate isopentenyltransferase from hop ( Humulus
lupulus L.). Phytochemistry 65:2439–46
80. Samuelson ME, Larsson C-M. 1993. Nitrate regulation of zeatin riboside levels in barley roots: effects of inhibitors of N assimilation and comparison with ammonium. Plant Sci.
93:77–84
81. Schmitz RY, Skoog F. 1972. Cytokinins: synthesis and biological activity of geometric
and position isomers of zeatin. Plant Physiol. 50:702–5
www.annualreviews.org • Cytokinin Biosynthesis 447

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 18/22
82. Schm ulling T, Werner T, Riefler M, Krupkova E, Bartrina y Manns I. 2003. Structureand function of cytokinin oxidase/dehydrogenase genes of maize, rice, Arabidopsis and
other species. J. Plant Res. 116:241–5283. Schnorr KM, Gaillard C, Biget E, Nygaard P, Laloue M. 1996. A second form of ade-
nine phosphoribosyltransferase in Arabidopsis thaliana with relative specificity towards
cytokinins. Plant J. 9:891–9884. Shantz EM, Steward FC. 1955. The identification of compound A from coconut milk as
1,3-diphenylurea. J. Am. Chem. Soc. 77:6351–5385. Shimizu-Sato S, Mori H. 2001. Control of outgrowth and dormancy in axillary buds.
Plant Physiol. 127:1405–1386. Shudo K. 1994. Chemistry of phenylurea cytokinins. In Cytokinins: Chemistry, Activity
and Function, ed. DWS Mok, MC Mok, pp. 35–42. Boca Raton, Florida: CRC Press87. Simpson RJ, Lambers H, Dalling MJ. 1982. Kinetin application to roots and its effec
on uptake, translocation and distribution of nitrogen in wheat (Triticum aestivum) grown
with a split root system. Physiol. Plant. 56:430–3588. Singh S, Letham DS, Jameson PE, Zhang R, Parker CW, et al. 1988. Cytokinin bio-
chemistry in relation to leaf senescence. IV. Cytokinin metabolism in soybean explants Plant Physiol. 88:788–94
89. Skoog F, Armstrong DJ. 1970. Cytokinins. Annu. Rev. Plant Physiol. 21:359–84
90. Skoog F, Armstrong DJ, Cherayil JD, Hampel AE, Bock RM. 1966. Cytokinin activitylocalization in transfer RNA preparations. Science 154:1354–56
91. Spichal L, Rakova NY, Riefler M, Mizuno T, Romanov GA, et al. 2004. Two cytokinin
receptors of Arabidopsis thaliana, CRE1/AHK4 andAHK3, differ in their ligand specificityin a bacterial assay. Plant Cell Physiol . 45:1299–305
92. Spiess LD. 1975. Comparative activity of isomers of zeatin and ribosyl-zeatin on Funaria
hygrometrica. Plant Physiol. 55:583–8593. Strnad M. 1997. The aromatic cytokinins. Physiol. Plant. 101:674–8894. Stuchbury T, Palni LMS, Horgan R, Wareing PF. 1979. The biosynthesis of cytokinin
in crown-gall tissue of Vinca rosea. Planta 147:97–10295. Sun J, Niu QW, Tarkowski P, Zheng B, Tarkowska D, et al. 2003. The Arabidopsis
AtIPT8/PGA22 gene encodes an isopentenyl transferase that is involved in de novo cy-tokinin biosynthesis. Plant Physiol. 131:167–76
96. Sun JP, Hirose N, Wang XC, Wen P, Xue L, et al. 2005. Arabidopsis SOI33/AtENT8
gene encodes a putative equilibrative nucleoside transporter that is involved in cytokinin
transport in planta. J. Integr. Plant Biol. 47:588–60397. Suzuki T, Miwa K, Ishikawa K, Yamada H, Aiba H, Mizuno T. 2001. The Arabidopsis
sensor His-kinase, AHK4, can respond to cytokinins. Plant Cell Physiol. 42:107–1398. Takei K, Dekishima Y, Eguchi T, Yamaya T, Sakakibara H. 2003. A new method for
enzymatic preparation of isopentenyladenine-type and trans -zeatin-type cytokinins withradioisotope-labeling. J. Plant Res. 116:259–63
This articlereported the first identification andcharacterization of Arabidopsis IPT genes.
99. Takei K, Sakakibara H, Sugiyama T. 2001. Identification of genes encoding
adenylate isopentenyltransferase, a cytokinin biosynthesis enzyme, in Arabidop- sis thaliana. J. Biol. Chem. 276:26405–10
100. Takei K, Sakakibara H, Taniguchi M, Sugiyama T. 2001. Nitrogen-dependent accumu-lation of cytokinins in root and the translocation to leaf: implication of cytokinin species
that induces gene expression of maize response regulator. Plant Cell Physiol. 42:85–93101. Takei K, Takahashi T, Sugiyama T, Yamaya T, Sakakibara H. 2002. Multiple routes
communicating nitrogen availability from roots to shoots: a signal transduction pathway
mediated by cytokinin. J. Exp. Bot. 53:971–77
448 Sakakibara

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 19/22
This articlereported theidentification of AtIPT3 as a key determinant for nitrogen-dependent CK biosynthesis.
102. Takei K, Ueda N, Aoki K, Kuromori T, Hirayama T, et al. 2004. AtIPT3, an
Arabidopsis isopentenyltransferase gene, is a key determinant of macronutrient-
responsive cytokinin biosynthesis. Plant Cell Physiol. 45:1053–62
103. Takei K, Yamaya T, Sakakibara H. 2003. A method for separation and determination of cytokinin nucleotides from plant tissues. J. Plant Res. 116:265–69
This articlereported the firsidentification ancharacterization CYP735A as CKtrans -hydroxylas
104. Takei K, Yamaya T, Sakakibara H. 2004. Arabidopsis CYP735A1 and CYP735A2
encode cytokinin hydroxylases that catalyze the biosynthesis of trans -zeatin. J.
Biol. Chem. 279:41866–72
105. Tanaka M, Takei K, Kojima M, Sakakibara H, Mori H. 2006. Auxin controls local cy-tokinin biosynthesis in the nodal stem in apical dominance. Plant J. doi: 10.1111/j.1365-
313x.2006.02656.x106. Tarkowska D, Dolezal K, Tarkowski P, ˚ Astot C, Holub J, et al. 2003. Identification of new
aromatic cytokinins in Arabidopsis thaliana and Populus x canadensis leaves by LC-(+)ESI-
MS and capillaryliquid chromatography/frit-fast atom bombardment mass spectrometry. Physiol. Plant. 117:579–90
107. Taya Y, Tanaka Y, Nishimura S. 1978. 5-AMP is a direct precursor of cytokinin inDictyostelium discoideum. Nature 271:545–47
108. Ueguchi C, Koizumi H, Suzuki T, Mizuno T. 2001. Novel family of sensor histidine
kinase genes in Arabidopsis thaliana. Plant Cell Physiol. 42:231–35109. Veach YK, Martin RC, Mok DW, Malbeck J, Vankova R, Mok MC. 2003.O-Glucosylation of cis -zeatin in maize. Characterization of genes, enzymes, and endoge-nous cytokinins. Plant Physiol. 131:1374–80
110. Vreman HJ, Skoog F. 1972. Cytokinins in Pisum transfer ribonucleic acid. Plant Physiol.
49:848–51
111. Vreman HJ,Thomas R, Corse J. 1978.Cytokinins in tRNA obtained from Spinacia oleracea
L. leaves and isolated chloroplasts. Plant Physiol. 61:296–306112. Werner T, Motyka V, Laucou V, Smets R, Van Onckelen H, Schm ulling T. 2003.
Cytokinin-deficient transgenic Arabidopsis plants showmultiple developmental alterationsindicating opposite functions of cytokinins in the regulation of shoot and root meristem
activity. Plant Cell 15:2532–50113. Werner T, Motyka V, Strnad M, Schm ulling T. 2001. Regulation of plant growth by
cytokinin. Proc. Natl. Acad. Sci. USA 98:10487–92
This articlereported the firsevidence for direbinding of CK tothe receptor in vitro.
114. Yamada H, Suzuki T, Terada K, Takei K, Ishikawa K, et al. 2001. The Arabidopsis
AHK4 histidine kinase is a cytokinin-binding receptor that transduces cytokinin
signals across the membrane. Plant Cell Physiol. 42:1017–23
115. Yonekura-Sakakibara K, Kojima M, Yamaya T, Sakakibara H. 2004. Molecular character-
ization of cytokinin-responsive histidine kinases in maize: differential ligand preferencesand response to cis -zeatin. Plant Physiol. 134:1654–61
116. Zubko E, Adams CJ, Machaekova I, Malbeck J, Scollan C, Meyer P. 2002. Activation
tagging identifies a gene from Petunia hybrida responsible for the production of active
cytokinins in plants. Plant J. 29:797–808
RELATED RESOURCE
Forde BG.2002. Local and long-range signaling pathways regulating plant responsesto nitrate. Annu. Rev. Plant Physiol. Plant Mol. Biol. 53:203–24
www.annualreviews.org • Cytokinin Biosynthesis 449

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 20/22
Annual Revie
of Plant Biolo
Volume 57, 20
Contents
Looking at Life: From Binoculars to the Electron Microscope
Sarah P. Gibbs p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 1
MicroRNAs and Their Regulatory Roles in Plants
Matthew W. Jones-Rhoades, David P. Bartel, and Bonnie Bartel p p p p p p p p p p p p p p p p p p p p p p p p p p 19
Chlorophyll Degradation During Senescence
S. H¨ ortensteiner p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 55
Quantitative Fluorescence Microscopy: From Art to Science Mark Fricker, John Runions, and Ian Moore p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 79
Control of the Actin Cytoskeleton in Plant Cell Growth
Patrick J. Hussey, Tijs Ketelaar, and Michael J. Deeks p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 109
Responding to Color: The Regulation of Complementary Chromatic
Adaptation
David M. Kehoe and Andrian Gutu p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 127
Seasonal Control of Tuberization in Potato: Conserved Elements with
the Flowering Response
Mariana Rodríguez-Falcón, Jordi Bou, and Salomé Prat p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p
151
Laser Microdissection of Plant Tissue: What You See Is What You Get
Timothy Nelson, S. Lori Tausta, Neeru Gandotra, and Tie Liu p p p p p p p p p p p p p p p p p p p p p p p p p p 181
Integrative Plant Biology: Role of Phloem Long-Distance
Macromolecular Trafficking
Tony J. Lough and William J. Lucas p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 203
The Role of Root Exudates in Rhizosphere Interactions with Plants
and Other Organisms
Harsh P. Bais, Tiffany L. Weir, Laura G. Perry, Simon Gilroy,
and Jorge M. Vivancop p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p
233
Genetics of Meiotic Prophase I in Plants
Olivier Hamant, Hong Ma, and W. Zacheus Cande p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 267
Biology and Biochemistry of Glucosinolates
Barbara Ann Halkier and Jonathan Gershenzon p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 303
v

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 21/22
Bioinformatics and Its Applications in Plant Biology
Seung Yon Rhee, Julie Dickerson, and Dong Xu p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 335
Leaf Hydraulics
Lawren Sack and N. Michele Holbrook p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 361
Plant Uncoupling Mitochondrial Proteins
An´ ıbal Eugˆ enio Vercesi, Jiri Boreck´ y, Ivan de Godoy Maia, Paulo Arruda,
Iolanda Midea Cuccovia, and Hernan Chaimovichp p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p
383
Genetics and Biochemistry of Seed Flavonoids
Lo¨ ıc Lepiniec, Isabelle Debeaujon, Jean-Marc Routaboul, Antoine Baudry,
Lucille Pourcel, Nathalie Nesi, and Michel Caboche p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 405
Cytokinins: Activity, Biosynthesis, and Translocation
Hitoshi Sakakibara p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 431
Global Studies of Cell Type-Specific Gene Expression in Plants
David W. Galbraith and Kenneth Birnbaum p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 451
Mechanism of Leaf-Shape Determination Hirokazu Tsukaya p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 477
Mosses as Model Systems for the Study of Metabolism and
Development
David Cove, Magdalena Bezanilla, Phillip Harries, and Ralph Quatrano p p p p p p p p p p p p p p 497
Structure and Function of Photosystems I and II
Nathan Nelson and Charles F. Yocum p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 521
Glycosyltransferases of Lipophilic Small Molecules
Dianna Bowles, Eng-Kiat Lim, Brigitte Poppenberger, and Fabi´ an E. Vaistij p p p p p p p p p p p 567
Protein Degradation Machineries in Plastids
Wataru Sakamoto p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 599
Molybdenum Cofactor Biosynthesis and Molybdenum Enzymes
G ¨ unter Schwarz and Ralf R. Mendel p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 623
Peptide Hormones in Plants
Yoshikatsu Matsubayashi and Youji Sakagami p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 649
Sugar Sensing and Signaling in Plants: Conserved and Novel
Mechanisms
Filip Rolland, Elena Baena-Gonzalez, and Jen Sheenp p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p
675
Vitamin Synthesis in Plants: Tocopherols and Carotenoids
Dean DellaPenna and Barry J. Pogson p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 711
Plastid-to-Nucleus Retrograde Signaling
Ajit Nott, Hou-Sung Jung, Shai Koussevitzky, and Joanne Chory p p p p p p p p p p p p p p p p p p p p p p 739
v i Con te nt s

8/3/2019 Citpkininas -Actividad y Traslocacion
http://slidepdf.com/reader/full/citpkininas-actividad-y-traslocacion 22/22
The Genetics and Biochemistry of Floral Pigments
Erich Grotewold p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 761
Transcriptional Regulatory Networks in Cellular Responses and
Tolerance to Dehydration and Cold Stresses
Kazuko Yamaguchi-Shinozaki and Kazuo Shinozaki p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 781
Pyrimidine and Purine Biosynthesis and Degradation in Plants
Rita Zrenner, Mark Stitt, Uwe Sonnewald, and Ralf Boldt p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p
805
Phytochrome Structure and Signaling Mechanisms
Nathan C. Rockwell, Yi-Shin Su, and J. Clark Lagarias p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 837
Microtubule Dynamics and Organization in the Plant Cortical Array
David W. Ehrhardt and Sidney L. Shaw p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 859
INDEXES
Subject Index p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 877
Cumulative Index of Contributing Authors, Volumes 47–57 p p p p p p p p p p p p p p p p p p p p p p p p p p p 915
Cumulative Index of Chapter Titles, Volumes 47–57 p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p p 920
ERRATA
An online log of corrections to Annual Review of Plant Biology chapters (if any, 1977 to
the present) may be found at http://plant.annualreviews.org/