
DETECCIÓN DE ELEMENTOS GENÉTICOS QUE LE CONFIEREN...
43
1 DETECCIÓN DE ELEMENTOS GENÉTICOS QUE LE CONFIEREN RESISTENCIA A ANTIBIÓTICOS A CEPAS DE Aeromonas spp. PROYECTO SIP 20070736 RESUMEN El presente trabajo tuvo como objetivo determinar la presencia de elementos genéticos relacionados con la adquisición de resistencias antimicrobianas en cepas de Aeromonas spp. aisladas de pacientes con cuadros diarreicos. Las especies identificadas fueron A. caviae, A. hydrophila, A. veronii y A. media, siendo la más frecuente A. caviae. Todas las cepas ensayadas fueron resistentes a ampicilina, característica de la mayoría de las especies de este género . Se encontraron cepas resistentes a cloranfenicol, tetraciclina, trimetoprim sulfametoxazol, ácido nalidíxico y estreptomicina, entre otros. Además de la resistencia a antibióticos todas las cepas soportaron una concentración de 10μg/mL HgCl2 , mientras que el 50% (22/41) de las cepas fueron capaces de crecer en presencia de 50 μg/mL. Se encontró DNA plasmídico en el 32% (13/41) de las cepas, A. caviae es la especie donde la frecuencia de plásmidos fue mayor. Se detectaron integrones clase 1, con diferentes genes cassette, que confieren resistencia a trimetoprim ( dhfr12, dhfr15, dfr2d ), estreptomicina/espectinomicina (aadA2, aadA 1), oxacilina (oxa2 ), cloranfenicol ( catB3, cmlA4 ) y un marco de lectura abierto ( orfD ). En este trabajo se detectaron los genes tet A, E, y B; la especie que presentó la mayor incidencia de genes tet es A. caviae , seguida de A. hydrophila y A. veronii. Adicionalmente se detectó el Tn1721 en 4 cepas. No se encontró Tn5393 completo, en ninguna de las cepas estudiadas, ya que sólo se detectó el gen de la transposasa, y no se amplificaron los genes strA-B comúnmente asociados a este transposón. Empleando el amplicón de los genes str A-B como sonda en una hibridación de alta astringencia, se detectó una cepa que contenía un gen homólogo, mientras que se encontraron dos cepas con genes de alta homología. La presencia de elementos de resistencia con características similares a los encontrados en una gran variedad de
Transcript of DETECCIÓN DE ELEMENTOS GENÉTICOS QUE LE CONFIEREN...

1
DETECCIÓN DE ELEMENTOS GENÉTICOS QUE LE CONFIEREN
RESISTENCIA A ANTIBIÓTICOS A CEPAS DE Aeromonas spp.
PROYECTO SIP 20070736
RESUMEN
El presente trabajo tuvo como objetivo determinar la presencia de elementos
genéticos relacionados con la adquisición de resistencias antimicrobianas en cepas
de Aeromonas spp. aisladas de pacientes con cuadros diarreicos. Las especies
identificadas fueron A. caviae, A. hydrophila, A. veronii y A. media, siendo la más
frecuente A. caviae. Todas las cepas ensayadas fueron resistentes a ampicilina,
característica de la mayoría de las especies de este género. Se encontraron cepas
resistentes a cloranfenicol, tetraciclina, trimetoprim sulfametoxazol, ácido
nalidíxico y estreptomicina, entre otros. Además de la resistencia a antibióticos
todas las cepas soportaron una concentración de 10µg/mL HgCl2, mientras que el
50% (22/41) de las cepas fueron capaces de crecer en presencia de 50 µg/mL. Se
encontró DNA plasmídico en el 32% (13/41) de las cepas, A. caviae es la especie
donde la frecuencia de plásmidos fue mayor. Se detectaron integrones clase 1, con
diferentes genes cassette, que confieren resistencia a trimetoprim (dhfr12, dhfr15,
dfr2d), estreptomicina/espectinomicina (aadA2, aadA1), oxacilina (oxa2),
cloranfenicol (catB3, cmlA4) y un marco de lectura abierto (orfD). En este trabajo se
detectaron los genes tet A, E, y B; la especie que presentó la mayor incidencia de
genes tet es A. caviae, seguida de A. hydrophila y A. veronii. Adicionalmente se
detectó el Tn1721 en 4 cepas. No se encontró Tn5393 completo, en ninguna de las
cepas estudiadas, ya que sólo se detectó el gen de la transposasa, y no se
amplificaron los genes strA-B comúnmente asociados a este transposón.
Empleando el amplicón de los genes strA-B como sonda en una hibridación de alta
astringencia, se detectó una cepa que contenía un gen homólogo, mientras que se
encontraron dos cepas con genes de alta homología. La presencia de elementos de
resistencia con características similares a los encontrados en una gran variedad de

2
bacterias de diferentes orígenes, sugiere la movilidad horizontal de estos
elementos.
INTRODUCCIÓN
Durante los últimos años ha aumentado la resistencia en diferentes especies
bacterianas a los antimicrobianos, y el género Aeromonas no es caso excepcional,
observándose un auge en su resistencia en diferentes partes del mundo. En
Aeromonas spp. algunos genes de resistencia están codificados en diferentes
elementos genéticos. Entre la resistencia a antibióticos relacionados con elementos
genéticos en Aeromonas spp. se encuentran: cloranfenicol,
trimetoprim/sulfametoxasol, tetraciclina, estreptomicina, ácido nalidíxico y sulfas.
Sin embargo, la resistencia a los diferentes antibióticos está claramente
influenciada por factores geográficos y socioeconómicos (Schmidt y cols., 2001a;
Goñi-Urriza y cols., 2000; Casas y cols., 2005; Sørum y cols., 2003; L´Abbed y
Sørum, 2000).
Las especies reportadas en diversos trabajos las cuales presentan elementos
genéticos son A. hydrophila, A. salmonicida, A. encheleia, A. icthiosmia, A. sobria, A.
media, A. veronii, A. bestiarum y A. caviae todas aisladas de muestras ambientales,
sin embargo no hay reportes de dichos elementos en cepas de origen clínico. Las
cepas de A. hydrophila, A. caviae, A. veronii y A. bestiarium causan mas infecciones en
el hombre lo cual sugiere que en cepas de origen clínico se podrían encontrar
también elementos genético. Aguilera-Arreola y cols., (2005) reporta que las cepas
de Aeromonas se pueden agrupar con base en su origen de aislamiento, lo cual
sugiere que las diferentes poblaciones pueden ocupar un nicho diferente y se
observa una distribución diferente de los genes de virulencia según el origen de la
bacteria, con ello podríamos encontrar diferencias en los elementos genéticos y los
genes de resistencia con respecto de las cepas de origen ambiental.

3
De hecho la transferencia “in vivo” e “in vitro” de marcadores de resistencia entre
diferentes especies de Aeromonas se ha demostrado. Trabajos realizados
recientemente en México demuestran la conjugación in vitro de A. hydrophila y A.
salmonicida aisladas de pescado (Vázquez-Ocampo, 2005). Bello-López, (2007)
demostró la conjugación in vivo de A. hydrophila y A. salmonicida, esta transferencia
de material genético se llevó a cabo en peces.
Por lo que hay suficiente evidencia para indicar que Aeromonas representa un
problema de salud pública: 1) la aparición de cepas resistentes a múltiples
antibióticos, que son capaces de transferir o capturar genes que le confieren
ventajas para el establecimiento en diferentes nichos, 2) la existencia de reservorios
en diferentes nichos donde pueda darse intercambio de material genético, 3)
diseminación para que estas bacterias resistentes se transmitan del humano vía el
medio ambiente; es necesario monitorear si las cepas aisladas de muestra clínicas
poseen elementos genéticos con genes de resistencia.
JUSTIFICACIÓN:
Las Aeromonas son consideradas actualmente patógenos emergentes y se han
aislado como agentes etiológicos de varios procesos infecciosos siendo el más
frecuente la gastroenteritis. Los cuadros diarreicos causados por Aeromonas son
autolimitados, por lo que de manera habitual el tratamiento no requiere el uso de
antibióticos. Sin embargo, durante los últimos años ha aumentado la resistencia en
diferentes especies bacterianas y el género Aeromonas no es caso excepcional,
observándose un auge en su resistencia en diferentes partes del mundo.
En Aeromonas algunos genes de resistencia pueden estar localizados en elementos
genéticos tales como plásmidos, transposones o integrones por ello la evolución y

4
diseminación tanto de las bacterias como de los elementos genéticos relacionados
con la transferencia de resistencia es un aspecto importante a estudiar,
especialmente porque no se tiene este tipo de información en cepas asociadas a
cuadros diarreicos en humanos.
HIPÓTESIS Si las especies del género Aeromonas spp. aisladas de origen clínico poseen
elementos genéticos que codifican para resistencias a diversos antibióticos se
explicaría la constante aparición de cepas resistentes a una amplia variedad de
antibióticos. Además, la transferencia de dichos elementos entre una misma
especie y entre especies diferentes facilitaría la diseminación de éstos y por
consiguiente las resistencias codificadas en los elementos
OBJETIVOS
• Objetivo general Detectar y caracterizar los elementos genéticos de resistencia a antibióticos
encontrados en cepas de Aeromonas spp. provenientes de pacientes con diarrea.
• Objetivos particulares
Identificar bioquímicamente las cepas a nivel de género.
Identificar genéticamente cepas de Aeromonas spp. provenientes de muestras
clínicas con base al protocolo del análisis de los patrones RFLPs del gen 16S
rDNA para determinar la especie.
Obtener el perfil plasmídico de las cepas.

5
Detectar elementos genéticos que codifican para resistencias a antibióticos
mediante la reacción en cadena de la polimerasa.
Determinar si existe correlación de los resultados obtenidos entre las
resistencias antimicrobianas y la presencia de los elementos genéticos.
MATERIAL Y MÉTODOS
Se analizaron 45 cepas de Aeromonas spp. de origen clínico aisladas de pacientes
que presentaban cuadros diarreicos en el estado de Hidalgo, México aisladas en el
transcurso de los meses julio-octubre del 2005.
En este trabajo se emplearon las siguientes cepas de referencia y cepas tipo:
A. hydrophila ATCC 7966 (839 CECT) a
A. salmonicida subsp. salmonicida 718 con el plásmido pRAS1 (integrón clase 1,
transposón 1721) b
E. coli C600 con tn5393 transposado en el plásmido pGEM-3ZF(+) c
E. coli K-12 NC 50078-02 [gen tet(A)] e
E. coli K-12 NC 50019 [gen tet(B)] e
E. coli K-12 NC 50270-01 [gen tet(C)] e
E. coli K-12 J53-1 RA1 NC 50073-02 [gen tet(D)] e
E. coli HB 101 pSL 1456 NC 50273-01 [gen tet(E)] e
E. coli V517 (diferentes plásmidos) f
E. coli con el plásmido pSa (integrón clase 1) f
Cepas proporcionadas por los doctores: a Graciela Castro Escarpulli, Escuela
Nacional de Ciencias Biológicas. b Glen Rhodes Centre for Ecology & Hydrology
Lancaster, Reino Unido. c Trine L´Abee-Lund, Noruegan School of Veterinary
Medicine . e Nahid Karami, Goteborg University, Suecia. f Everardo Curiel
Quesada, Escuela Nacional de Ciencias Biológicas.

6
MANEJO Y CONSERVACIÓN DE CEPAS Las cepas de trabajo se conservaron a largo plazo a -70°C en criotubos que
contenían glicerol al 40%. Para recuperar las cepas se tomó un inóculo de cada
criotubo, y se sembró por estría cruzada en cajas de medio Luria con ampicilina
(100 µg/mL) y se incubaron a 37ºC durante 24 h.
IDENTIFICACIÓN BIOQUÍMICA
Antes de llevar a cabo la identificación genética se confirmó su pertenencia al
género Aeromonas con base en las siguientes pruebas:
Gram, Oxidasa, Crecimiento a 3 y 6% de NaCl, producción de ácido del inositol y
utilización de la glucosa por vía oxidativa o fermentativa.
IDENTIFICACIÓN GENÉTICA
La identificación genética se realizó con base en el análisis de los patrones RFLPs
del gen 16S rDNA propuesto por Borrel y cols., 1997 y Figueras y cols., 2000 (Fig 4).
EXTRACCIÓN DE DNA TOTAL.
1. Se obtuvo un cultivo líquido de 18 a 24 h de la bacteria deseada.
2. En un tubo para microcentrífuga, se colocó 1.5 mL de cultivo, se centrifugó 1
min, se descartó el sobrenadante.
3. Se adicionaron 500 µl de TE (50/20), se resuspendieron las bacterias, se
centrifugó 1 min, se descartó el sobrenadante.
4. Se adicionaron 175 µl de TE (50/20), se resuspendieron las bacterias.
5. Se agregaron 5 µl de proteinasa K (20 mg/mL) y 20 µl de SDS al 10%, se
agitaron cuidadosamente para mezclar e incubar a 56º C por 1 ó 2 h.
6. Se adicionaron 500 µl de TE (10/1) y 20 µl de NaCl 5M, se agitó
cuidadosamente.

7
7. Se realizaron dos extracciones con fenol, agregando 500 µl al tubo, agitando
con cuidado (15 min de preferencia en un dispositivo rotatorio) para
mezclar lo mejor posible y centrifugar por 10 min.
8. Se realizó una extracción con 500 µl de cloroformo: alcohol isoamílico (24:1),
agitando cuidadosamente y centrifugando 2 min. Se separó la fase acuosa.
9. Se agregó un volumen de 500 µl de isopropanol, se agitó por inversión y se
centrifugó por 30 seg. Descartar el sobrenadante.
10. El DNA se enjuagó con 1mL de etanol al 70%, se centrifugó 30 segundos y se
descartó el sobrenadante para posteriormente secarlo a 65º C y disolverlo en 200 ó
400 µl de agua destilada, desionizada estéril.
AMPLIFICACIÓN MEDIANTE PCR DEL GEN 16S rDNA.
Para la amplificación mediante PCR del gen 16S rDNA, se sometieron a la
amplificación aproximadamente 200-300 ng del DNA genómico, en un volumen
final de 50 µl. La mezcla de reacción contenía: Tris-HCl 20 mM (pH8.4), KCl 50
mM, MgCl2 1.5 mM, desoxirribonucleósidos trifosfato dNTP´s (dATP, dCTP, dGTP
y dTTP ) 0.3 mM de cada uno, iniciadores 0.3 µM de cada uno y 2.5 U de Taq
polimerasa. Las condiciones para cada ciclo de reacción fueron: 96°C durante 5 min
(desnaturalización), 94°C durante 1 min (desnaturalización), 56°C durante 1 min
(hibridación), 72°C durante 1.30 min (extensión), estas condiciones se repitiero n
por 35 ciclos y al finalizar se efectuó una última extensión a 72°C durante 5 min.
Las secuencias de los iniciadores empleados para este propósito se muestran en el
siguiente cuadro.
Iniciadores empleados para la amplificación del gen 16S rDNA.
Tomado de Borrel y cols., 1997; Figueras y cols., 2000
Región Amplificada
Secuencia 5 -́3´ Tamaño del amplicón
16S rDNA FW: 5´ AGA GTT TGA TCA TGG CTC A 3 RV:5´ GGT TAC CTT GTT ACG ACT T 3 1502 pb

8
RESTRICCIÓN DEL AMPLICÓN DEL GEN 16S rDNA.
Previamente a la digestión se procedió a la purificación del amplificado empleando
el kit de purificación “High Pure PCR Product” (ROCHE). El producto resultante
de la limpieza se sometió a la digestión enzimática doble (AluI y MboI), se preparó
una mezcla la cual contiene 12 µl del fragmento amplificado, 1 µl del regulador de
digestión (10x), 5U Alu I y 5U de la enzima Mbo I, adicionando agua para un
volumen final de 20 µl, la mezcla se incubó a 37°C durante toda la noche.
Los fragmentos polimórficos, se separaron mediante electroforesis en geles de
poliacrilamida al 17% a 80 V durante 6 horas. Al término del tiempo se tiñeron los
geles durante 30 min con bromuro de etidio (0.5 µg/mL) y se visualizaron en un
documentador de geles con luz ultravioleta. Las fotografías de los PCR-RFLP
obtenidas de las digestiones dobles con las enzimas Alu I y Mbo I fueron
comparadas con los patrones para las cepas ti po de cada una de las especies
DETERMINACIÓN DE LA SENSIBILIDAD A ANTIMICROBIANOS
Se determinó el grado de resistencia de las cepas de Aeromonas spp. a diversos
agentes antimicrobianos, por el método de difusión en agar siguiendo las
indicaciones del CLSI 2007 ( antes NCCLS).
1. Al menos 3 a 5 colonias bien aisladas se seleccionaron de un medio de cultivo.
El crecimiento se transfirió a un tubo con 4 a 5 mL de caldo Mueller-Hinton. El
caldo de cultivo se incubó a 37ºC hasta que alcanzó la turbidez del estándar de
0.5 McFarland (usualmente 2 a 6 h). Esto resulta en una suspensión que
contiene aproximadamente 1 a 2 x 108 UCF/mL.
2. Se sembraron placas de agar Mueller-Hinton usando un hisopo a partir de la
suspensión ajustada en varias direcciones.

9
3. Se dejaron reposar las placas cinco min a temperatura ambiente para que secara
el inóculo.
4. Los sensidiscos se dispensaron sobre la superficie del agar. Cada disco debe ser
presionado para asegurar contacto pleno con la superficie del agar. Ya sea que
el disco se ponga individualmente o con un dispensador, deben ser distribuidos
en forma uniforme y no deben quedar a menos de 24 mm de distancia entre
centros. No más de 12 discos se deben poner por placa de 150 mm o no más de
5 en placas de 100 mm. Debido a que algunas drogas difunden casi
instantáneamente, un disco no debe ser relocalizado una vez que haya tomado
contacto con la superficie del agar.
5. Las placas se incubaron a 37ºC durante 24 h.
6. Se midieron los halos de inhibición en mm por el fondo de la placa y se
interpretaron los diámetros de las zonas de inhibición de las cepas para
determinar las categorías de susceptibles, intermedio y resistente según los
estándares de interpretación.
EXTRACCIÓN DE DNA PLASMÍDICO (TÉCNICA DE BIRNBOIM Y DOLY
MODIFICADA).
La técnica consiste en una lisis alcalina y se basa en el intervalo de pH estrecho que
existe de 12.0 a 12.5, dentro del cual hay ruptura y desnaturalización del DNA
cromosomal generándose al renaturalizarse, una masa de DNA lineal la cual se
puede separar por centrifugación (Birnboim y Doly, 1979). El procedimiento se
desarrolló como sigue (Sambrook, 1989):
1. De un cultivo puro de 24 h de crecimiento se seleccionaron de 4 ó 5 colonias,
se sembraron en 10 mL de caldo Luria y se incubaron a 37°C en agitación
durante 18 h.
2. Se pasaron 1.5 mL del cultivo a tubos de 2 mL y se centrifugó a 12 000 rpm
durante 5 min, posteriormente se eliminó el sobrenadante.

10
3. Se resuspendió el paquete celular con 100 µl de solución TEG (trisma - HCl
25 mM pH 8, EDTA 10 mM, glucosa 50 mM).
4. Se mezcló y se dejó 5 min a temperatura ambiente.
5. Se adicionó un volumen de 200 µl de la solución de lisis (NaOH 0.2N - SDS
1%) y se colocaron inmediatamente en hielo durante 5 min.
6. Se precipitó el DNA cromosomal y las proteínas con 150 µl de acetato de
potasio 3 M, pH 4.8 y se colocaron en hielo durante 5 min.
7. El tubo se centrifugó a 12 000 rpm durante 5 min.
8. Se tomaron 400 µl del sobrenadante y se transfirieron cuidadosamente a un
tubo limpio.
9. Se adicionaron 200 µl de cloroformo-fenol saturado con Tris pH 7.8.
10. Se centrifugó a 12 000 rpm durante 5 min, se separó la fase acuosa en un
tubo limpio.
11. Se precipitó el DNA de la fase acuosa con un volumen igual de isopropanol
y se mezcló en un Vortex.
12. Se centrifugó a 12 000 rpm por 5 min.
13. Se decantó e invirtió el tubo sobre un papel secante y se dejo secar a
temperatura ambiente.
14. El botón se resuspendió en 50 µl de solución reguladora TE (Tris-HCL 50
mM - EDTA 10 mM, pH 8).
DETECCIÓN DE GENES ASOCIADOS A INTEGRONES
La presencia de integrones de clase 1 se puso de manifiesto mediante la detección
de los genes que forman parte de dichos elementos por la técnica de PCR, bajo las
condiciones descritas previamente por Zhang y cols., 2004. Para la detección de la
región variable se empleó un par adicional de iniciadores, diseñados en este
trabajo.
Para la reacción de PCR se sometieron a la amplificación 5µl (200-300 ng) del DNA,
25 µl de Master Mix (eppendorff), 5 µl (10 pmol/µl) de cada iniciador, en un

11
volumen final de 50 µl. Las condiciones para cada ciclo de reacción programada
para el gen int1 fueron: 94°C durante 5 min, 94°C durante 30 seg
(desnaturalización), 50°C durante 30 seg (alineamiento), 72°C durante 1.5 min
(extensión), estas condiciones se repitieron por 30 ciclos y al finalizar se efectuó
una última extensión a 72°C durante 7 min. Para el gen variable I y qacEA1/Sul1 se
modificó la temperatura de alineación y el tiempo de la polimerización entre 52-
56ºC y 1-2 min respectivamente. Para la amplificación de la región variable II, la
temperatura de alineamiento fue de 68°C y se empleo la enzima Vent polimerasa
(New England).
Iniciadores utilizados para la detección de genes asociados a integrones.
Gen Iniciadores
(5´? 3´) No de Acceso
(GenBank) Amplicón (pb)
Int FW: GTTCGGTCAAGGTTCTG RV: GCCAACTTTCAGCACATG Y18050 923
Var I FW: GGCATACAAGCAGCAAGC RV: AAGCAGACTTGACCTGAT U12338 Variable
Var II FW: GCA ACT GGT CCA GAA CCT TG RV: GAT TAT GAC AAC GGC GGA AG AJ517790 Variable
qacEA1 Sul1
FW: ATC GCA ATA GTT GGC GAA GT RV: GCA AGG CGG AAA CCC GCG CC
X15370 X12869
800
CARACTERIZACIÓN DE INTEGRONES
Determinar el tamaño de la región variable y secuenciar los fragmentos de interés
para conocer que genes de resistencia se encuentran integrados.
Restricción del amplicón de la región variable
Previamente a la digestión se procedió a la purificación del amplificado empleando
el sistema de purificación “High Pure PCR Product” (ROCHE).
El producto resultante de la limpieza se sometió a la digestión enzimática con cinco
ezimas (AluI, MboI, HindIII, HinfI, MspI), se preparó una mezcla la cual contiene 10
µl del fragmento amplificado, 2.5 µl del regulador de digestión (10x), 5U de la
enzima, adicionando agua para un volumen final de 25 µl, la mezcla se incubó a
37°C durante toda la noche. Los fragmentos polimórficos, se separaron mediante

12
electroforesis en geles de poliacrilamida al 17% a 70 V durante 8 h. Al término del
tiempo se tiñeron los geles durante 30 min con bromuro de etidio (0.5 µg/mL) y se
visualizaron en un documentador de geles con luz ultravioleta.
Los patrones de los PCR-RFLP obtenidos de las digestiones fueron comparados
para saber si los fragmentos de la región variable son iguales. Se eligió un
amplificado de cada perfil de restricción para la secuenciación.
Secuenciación Los amplificados de la región variable se purificaron empleando la enzima agarasa
(Fermentans). El fragmento se corrió en un gel de agarosa de bajo punto de fusión,
se cortó la banda, la cual se incubó junto con la enzima ß-Agarasa I (1U/ 200 µl de
agarosa fundida) a 42°C, por 1 h. Posteriormente se centrifugó por 15 min a 12,000
rpm, para eliminar cualquier fragmento remanente, el sobrenadante se transfirió a
un tubo limpio y se precipitó con isopropanol. El DNA se lavó con etanol al 70%,
se centrifugó y se secó para luego resuspenderlo en 50 µl de agua.
El amplificado de la región variable se envió al servicio de secuenciación para
obtener dos diferentes secuencias una con el iniciador forward y otra con el reverse.
Las secuencias fueron visualizadas en el programa “Chromas”, cada una de las
secuencias obtenidas se sometió a una búsqueda de secuencias similares en la base
de datos del “GenBank” mediante un análisis BLAST
(http://www.ncbi.nlm.nih.gov/BLAST/), para encontrar las secuencias
emparentadas, posteriormente se realizó un alineamiento de secuencias en el
programa Vector NTI V9.
DETECCIÓN DE GENES ASOCIADOS A TRANSPOSONES
La detección de genes asociados a transposones se realizó empleando la técnica de
PCR. Los iniciadores empleados para la detección de estos elementos se diseñaron
en el presente estudio por medio de un análisis de secuencias empleando el

13
programa Vector NTI y utilizando iniciadores reportados en la literatura por Han y
cols. 2004.
Las condiciones de reacción programadas fueron: 94°C durante 2 min, 94°C
durante 30 seg (desnaturalización), 58°C durante 30 seg (alineamiento), 72°C
durante 30 seg (extensión), estas condiciones se repitieron por 30 ciclos y al
finalizar se efectuó una última extensión a 72°C durante 10 min. Para la reacción de
PCR se sometieron a la amplificación 5 µl (200-300 ng) del DNA, 25 µl de Master
Mix, 5µl (10 pmol/µl) de cada iniciador, en un volumen final de 50 µl.
Iniciadores utilizados para la detección de genes asociados a transposones.
tnp A= gen que codifica para la transposasa
DETECCIÓN DE DETERMINANTES tet (para las cepas resistentes a
tetraciclina).
Los genes que codifican la resistencia a tetraciclina (tet A-E) fueron detectados por
medio de la PCR utilizando condiciones e iniciadores establecidos por Schmidt y
cols., 2001; Guardabasi y cols. 2000. Para la reacción de PCR se sometieron a la
amplificación 5 µl (200-300 ng) del DNA, 25 µl de Master Mix (eppendorf), 5 µl (10
pmol/µl) de cada iniciador, en un volumen final de 50µl.
Iniciadores empleados en la detección de determinantes tet clases A, B, C, D y E.
Clase Iniciadores (5´? 3´)
Amplicón (pb)
A FW: GTA ATT CTG AGC ACT GCT GC RV: CTG CCT GGA CAA CAT TGC TT 937
Gen Iniciadores (5´? 3´) Amplicón (pb) Transposon
tnpA FW: GGA ACG ATG ACG GAT TTC AA RV: ATC CCT TGA TCG TGG CAT AG 607 Tn1721
tetA FW: GTA ATT CTG AGC ACT GCT GC RV: CTG CCT GGA CAA CAT TGC TT 937 Tn1721
tnpA
FW: CCAGAAGGTGTGGGTGTTTGT RV: GCTCGTCTCGCTGTGCCATT
339 Tn5393
str A-B FW: GGAACTGCGTGGGCTACA RV: GCTAGATCGCGTTGCTCCTCT 303 Tn5393

14
B FW: CTC AGT ATT CCA AGC CTT TG RV: CTA AGC ACT TGT CTC CTG TT 416
C FW: TCT ACC AAT GCG CTC ATC GT RV: GGT TGA AGG CTC TCA AGG GC 588
D FW: ATT ACA CTG CTG GAC GCG AT RV: CTG ATC AGC AGA CAG ATT GC 1123
E FW: GTG ATG ATG GCA CTG CTG AT RV: CTC TGC TGT ACA TCG CTC TT 1199
Schmidt y cols., 2001; Guardabasi y cols., 2000
Condiciones para la amplificación de los genes tet
tet A,B,C,D, E
Etapa Temperatura (oC)
Tiempo Ciclos
Desnaturalización inicial 95 5 min
Desnaturalización 95 30 seg Alineamiento 56-62 30 seg
Extensión 72 45 seg
30
Extensión final 72 7 min
Todos los productos de PCR obtenidos fueron analizados por electroforesis en
geles de agarosa teñidos posteriormente con bromuro de etidio.
HIBRIDACIÓN SOUTHERN PARA LA DETECCIÓN DE LOS GENES strA-B.
Se empleo la técnica de hibridación para descartar falsos negativos en la detección
de los genes strA-B mediante PCR. La hibridación se llevo acabo en cepas donde se
detectó la transposasa del Tn5393.
Preparación de la membrana de Nylon para la hibridación
La membrana de nylon (sin carga) fue sumergida en agua desionizada y
posteriormente en el regulador de transferencia (10X SSC) por 10 minutos. En la
membrana se colocaron 2 µl de la muestra del DNA previamente desnaturalizado.
Posteriormente se fijo con luz U.V por 2 minutos y se procedió a la prehibridación.
Hibridación
Para llevara acabo el marcaje de la sonda y la hibridación se empleo el Ki t Dig
High Prime DNA Labeling And Detection Starker Kit II (ROCHE):

15
1. El marcaje de la sonda (producto de PCR str A-B) fue con Digoxigenina-11-
dUTP (DIG-High Prime).
2. Se llevó acabo la prehibridación a 42º C por 30 min (DIG Easy Hyb).
3. Posteriormente se hibrido a 68º C por 4 h (DIG Easy Hyb + sonda).
4. Se realizaron 2 lavados de baja astringencia con SDS 0.1 % - SSC 2X por 5
min a temperatura ambiente y 2 de alta astringencia con SDS 0.1 % - SSC
0.5X por 15 min a 68º C.
5. La membrana se incubó por 30 min en solución de bloqueo y
posteriormente se adicionó el anticuerpo.
6. Se realizaron 2 lavados con el regulador de lavado
7. Se equilibro la membrana con el regulador de detección y se adicionó CSPD
para revelar, posteriormente la membrana fue expuesta a una placa
radiográfica para evidenciar la hibridación por quimioluminiscencia.

16
RESULTADOS Meta 1.- Demostración de la presencia de integrotes tipo I
La presencia de integrones de tipo 1 se puso de manifiesto mediante la detección
de los genes que forman parte de dichos elementos por la técnica de PCR. Todo el
elemento se amplificó en tres partes, primero se amplificó la región 5´del gen de la
integrasa, en segundo lugar se amplificó parte de los genes sul 1 y qacEA 1 que
forman la región 3´CS y por último se amplificó la región variable de aquellas
cepas que presentaron únicamente la región 5 y 3´CS.
El gen de la integrasa fue detectado en 16 cepas de las 41 cepas que fueron
sometidas a la búsqueda, el resultado de esta amplificación fue un fragmento de
923 pb como se observa en la siguiente figura.
De estas 16 cepas sólo 10 presentaron la región 3´CS donde se amplificó un
fragmento de 800 pb como lo muestra la figura siguiente.
1 2 3 4
923 pb 1000
500
100
Producto de amplificación del gen de la integrasa clase 1. Carril 1 Marcador de peso molecular Roche IX. Carril 2 A. hydrophila 839 testigo negativo Carril 3. A. salmonicida 718 (pRAS1) testigo positivo. Carril 4 E. coli (pSa) 185 testigo positivo. Carriles 5-7 cepas problema de Aeromonas spp.
1 2 3 4 5 6 7

17
La búsqueda de la región variable se realizó con los iniciadores var I, en las 9 cepas
que presentaron ambas regiones conservadas. Sin embargo, sólo 3 cepas
presentaron un fragmento de tamaño variable, se obtuvieron bandas de
aproximadamente 800 y 1500.
En las cepas restantes se emplearon los iniciadores var II, que amplifican un
fragmento ligeramente mayor de la región variable, se logró amplificar a las 6
Producto de amplificación de la región 3´CS del integrón clase 1. Carril 1 Marcador de peso molecular Roche IX. Carril 2 A. hydrophila 839 testigo negativo Carril 3. A. salmonicida 718 (pRAS1) testigo positivo. Carril 4 E. coli 185 (pSa) testigo positivo. Carriles 5-14 cepas problema de Aeromonas spp.
1 2 3 4 5 6 7 8 9 10 11 12
1353 1078 872 603 310
800 pb
1 2 3 4 5 6 7 8 9 10 11 12 13 14
1 2 3 4 5 6 7 8 9
1353 1078 872 603 310 pb
Productos de amplificación de la región variable con los iniciadores var I. Carril 1 Marcador de peso molecular Roche IX. Carril 2 A. hydrophila 839 testigo negativo Carril 3. A. salmonicida 718 (pRAS1) testigo positivo. Carril 4 E. coli 185 (pSa) testigo positivo. Carriles 5-9 cepas problema de Aeromonas spp.
750 1510 1810 1147 1810 Tamaño de la banda (pb)
718 185 6479 3539 3430 Número de cepa
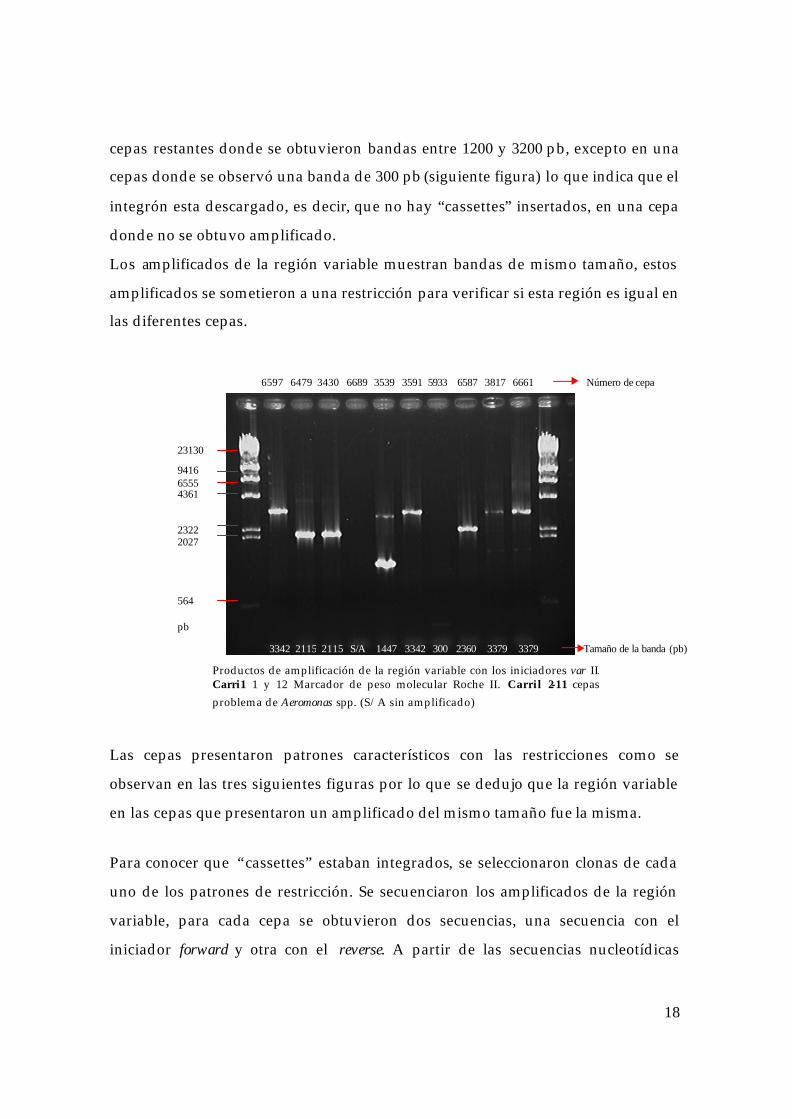
18
cepas restantes donde se obtuvieron bandas entre 1200 y 3200 pb, excepto en una
cepas donde se observó una banda de 300 pb (siguiente figura) lo que indica que el
integrón esta descargado, es decir, que no hay “cassettes” insertados, en una cepa
donde no se obtuvo amplificado.
Los amplificados de la región variable muestran bandas de mismo tamaño, estos
amplificados se sometieron a una restricción para verificar si esta región es igual en
las diferentes cepas.
Las cepas presentaron patrones característicos con las restricciones como se
observan en las tres siguientes figuras por lo que se dedujo que la región variable
en las cepas que presentaron un amplificado del mismo tamaño fue la misma.
Para conocer que “cassettes” estaban integrados, se seleccionaron clonas de cada
uno de los patrones de restricción. Se secuenciaron los amplificados de la región
variable, para cada cepa se obtuvieron dos secuencias, una secuencia con el
iniciador forward y otra con el reverse. A partir de las secuencias nucleotídicas
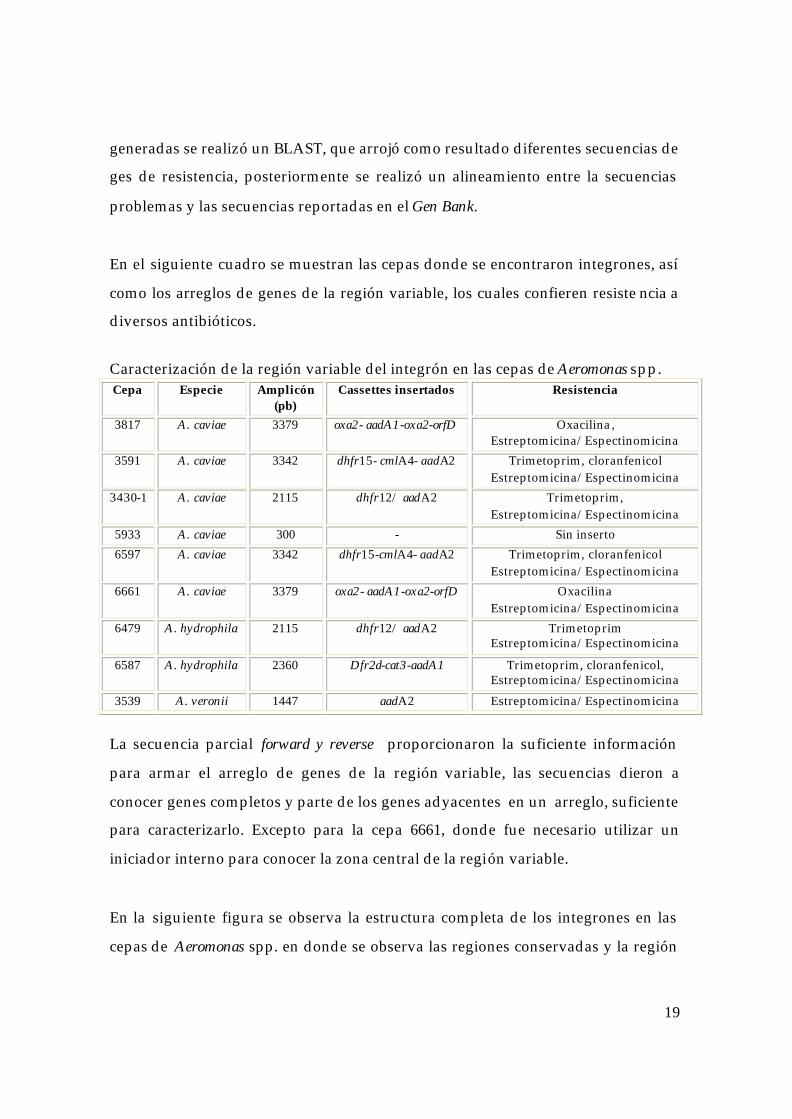
Productos de amplificación de la región variable con los iniciadores var II. Carri1 1 y 12 Marcador de peso molecular Roche II. Carril 2-11 cepas problema de Aeromonas spp. (S/A sin amplificado)
6597 6479 3430 6689 3539 3591 5933 6587 3817 6661 Número de cepa
23130 9416 6555 4361 2322 2027 564 pb
3342 2115 2115 S/A 1447 3342 300 2360 3379 3379 Tamaño de la banda (pb)
19
generadas se realizó un BLAST, que arrojó como resultado diferentes secuencias de
ges de resistencia, posteriormente se realizó un alineamiento entre la secuencias
problemas y las secuencias reportadas en el Gen Bank.
En el siguiente cuadro se muestran las cepas donde se encontraron integrones, así
como los arreglos de genes de la región variable, los cuales confieren resiste ncia a
diversos antibióticos.
Caracterización de la región variable del integrón en las cepas de Aeromonas spp. Cepa Especie Amplicón
(pb) Cassettes insertados Resistencia
3817 A. caviae 3379 oxa2- aadA1-oxa2-orfD Oxacilina, Estreptomicina/Espectinomicina
3591 A. caviae 3342 dhfr15- cmlA4- aadA2 Trimetoprim, cloranfenicol Estreptomicina/Espectinomicina
3430-1 A. caviae 2115 dhfr12/ aadA2 Trimetoprim, Estreptomicina/Espectinomicina
5933 A. caviae 300 - Sin inserto 6597 A. caviae 3342 dhfr15-cmlA4- aadA2 Trimetoprim, cloranfenicol
Estreptomicina/Espectinomicina 6661 A. caviae 3379 oxa2- aadA1-oxa2-orfD Oxacilina
Estreptomicina/Espectinomicina 6479 A. hydrophila 2115 dhfr12/ aadA2 Trimetoprim
Estreptomicina/Espectinomicina
6587 A. hydrophila 2360 Dfr2d-cat3-aadA1 Trimetoprim, cloranfenicol, Estreptomicina/Espectinomicina
3539 A. veronii 1447 aadA2 Estreptomicina/Espectinomicina
La secuencia parcial forward y reverse proporcionaron la suficiente información
para armar el arreglo de genes de la región variable, las secuencias dieron a
conocer genes completos y parte de los genes adyacentes en un arreglo, suficiente
para caracterizarlo. Excepto para la cepa 6661, donde fue necesario utilizar un
iniciador interno para conocer la zona central de la región variable.
En la siguiente figura se observa la estructura completa de los integrones en las
cepas de Aeromonas spp. en donde se observa las regiones conservadas y la región

20
variable formada por diferentes genes de resistencia, excepto en un integrón. En
todos los casos los arreglos encontrados correspondieron a los publicados en otras
bacterias.
DETECCIÓN DE GENES QUE CODIFICAN RESISTENCIA A TETRACICLINA
El criterio de selección para la detección de estos genes fueron aquellas cepas
resistentes a tetraciclina (n=16).
Se llevó a cabo la amplificación de los genes que codifican para la resistencia a
tetraciclina tet A, tet C, tet D y tet E. De las 16 cepas resistentes a tetraciclina en 5
cepas se detectó en gen tet A, en 5 cepas tet E y en 3 cepas se detectó el gen tet B
como se muestra en la siguiente figura. En ninguna cepa se detectaron los genes C
y D. Cabe mencionar que en tres cepas resistentes a tetraciclina no se detectó
ninguno de estos genes.
Productos de amplificación de los genes tet: tet A Carril 1 Marcador de peso molecular Roche IX. Carril 2 A. hydrophila 839 testigo negativo Carril 3. E. coli tet A testigo positivo. Carriles 4-8 cepas problema de Aeromonas spp. tet E Carriles 1-4 cepas problema de Aeromonas spp. Carril 5 E. coli tet E testigo positivo Carril 6. A. salmonicida 718 testigo negativo. Carril 7 Marcador de peso molecular Roche IX. tet B Carri1 1 Marcador de peso molecular Roche IX. Carril 2 E. coli tet B testigo positivo. Carriles 3-5 cepas problema de Aeromonas spp. Carril 6. A. salmonicida 718 testigo negativo.
1 2 3 4 5 6 7 8
1323 1078 872 603 310
937 pb
tet A
1 2 3 4 5 6 7
1199 pb
tet E
1323 1078 872 603 310

21
Meta 2.- Detección de transposonones tipo Tn5393
Las cepas (n=41) que fueron seleccionadas para la búsqueda de este elemento
fueron cepas resistentes a estreptomicina.
El gen de la transposasa del elemento Tn5393 se detectó en 27 cepas (ver figura),
posteriormente se llevó a cabo la detección del gen de estreptomicina el cual no se
detectó en ninguna.
Meta 2 .- Detección de transposones tipo Tn1721
El criterio de selección para la detección de este transposón fueron aquellas cepas
resistentes a tetraciclina con el gen tet A (n=5).
Se amplificó el gen que codifica la transposasa (600 pb) y esté se presentó en 4
cepas (ver figura). Por lo tanto el elemento Tn1721 se encontró en 4 cepas de
Aeromonas spp.
Fig. 24 Producto de amplificación del gen de la transposasa del Tn1721 (600 pb). Carril 1 A. hydrophila 839 testigo negativo Carril 2 E. coli C600 Tn5393 testigo negativo. Carril 3 A. salmonicida 718 (pRAS1) testigo positivo. Carriles 4-8 cepas problema de Aeromonas spp. Carril 9 Marcador de peso molecular Roche IX.
1323 1078 872 603 310
Fig. 25 Producto de amplificación del gen de la transposasa del Tn5393. Carri1 1 Marcador de peso molecular Roche IX. Carril 2 A. hydrophila 839 testigo negativo Carril 3. E. coli C600 Tn5393 testigo positivo. Carriles 4-15 cepas problema de Aeromonas spp.
1353 1078 872 603 310
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15
340 pb

22
Meta 3 Obtención de amplicones de la región variable del integron. Los resultados obtenidos de la aplicación de la sonda derivada de los genes str A-B
EN cepas de Aeromonas spp. las cuales contenían el gen del la transposasa del
elemento Tn5393 y cepas resistentes a estreptomicina..
Se detectaron 3 cepas que hibridaron con la sonda del gen strA-B, una cepa
muestra una señal intensa en la hibridación (carril 15, cepa 6661) lo que indica que
esta cepa contiene el gen strA-B, por lo que podemos decir que el transposón
Tn5393 fue detectado en una cepa de origen clínico. Cabe mencionar que dos cepas
mostraron una señal menor (carriles 3 y 11 cepas 6336 y 6376 respectivamente),
esto se pudo deber que son genes similares u homólogos a dicho gen.
Mediante esta técnica se comprobó que la mayoría de las cepas no contienen el gen
(strA-B) asociado al transposón Tn5393, además de que se evidenciaron falsos
negativos mediante la técnica de PCR.
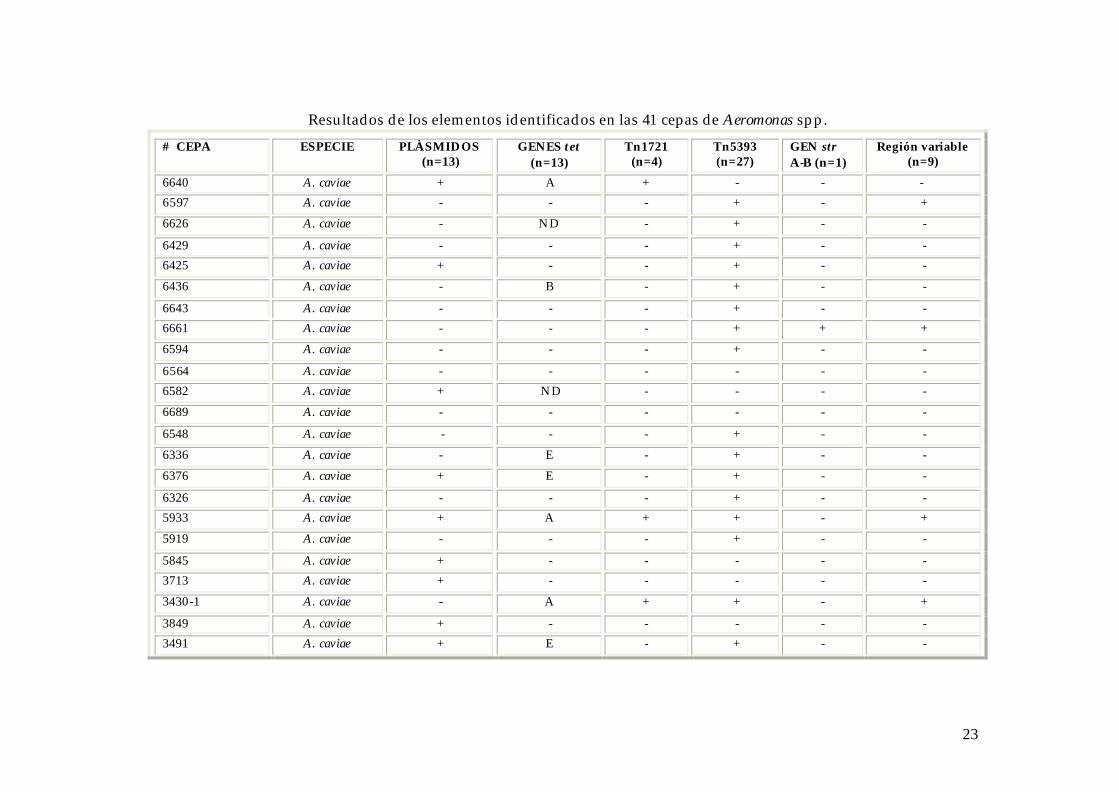
En el siguiente cuadro se muestran el número de genes y elementos genéticos de
resistencia encontrados en las cepas de Aeromonas spp. aisladas de heces diarreicas.
23
Resultados de los elementos identificados en las 41 cepas de Aeromonas spp.
# CEPA ESPECIE PLÀSMIDOS (n=13)
GENES tet (n=13)
Tn1721 (n=4)
Tn5393 (n=27)
GEN str A-B (n=1)
Región variable (n=9)
6640 A. caviae + A + - - - 6597 A. caviae - - - + - +
6626 A. caviae - ND - + - -
6429 A. caviae - - - + - - 6425 A. caviae + - - + - -
6436 A. caviae - B - + - -
6643 A. caviae - - - + - - 6661 A. caviae - - - + + +
6594 A. caviae - - - + - -
6564 A. caviae - - - - - - 6582 A. caviae + ND - - - -
6689 A. caviae - - - - - -
6548 A. caviae - - - + - -
6336 A. caviae - E - + - -
6376 A. caviae + E - + - -
6326 A. caviae - - - + - - 5933 A. caviae + A + + - +
5919 A. caviae - - - + - -
5845 A. caviae + - - - - - 3713 A. caviae + - - - - -
3430-1 A. caviae - A + + - +
3849 A. caviae + - - - - - 3491 A. caviae + E - + - -
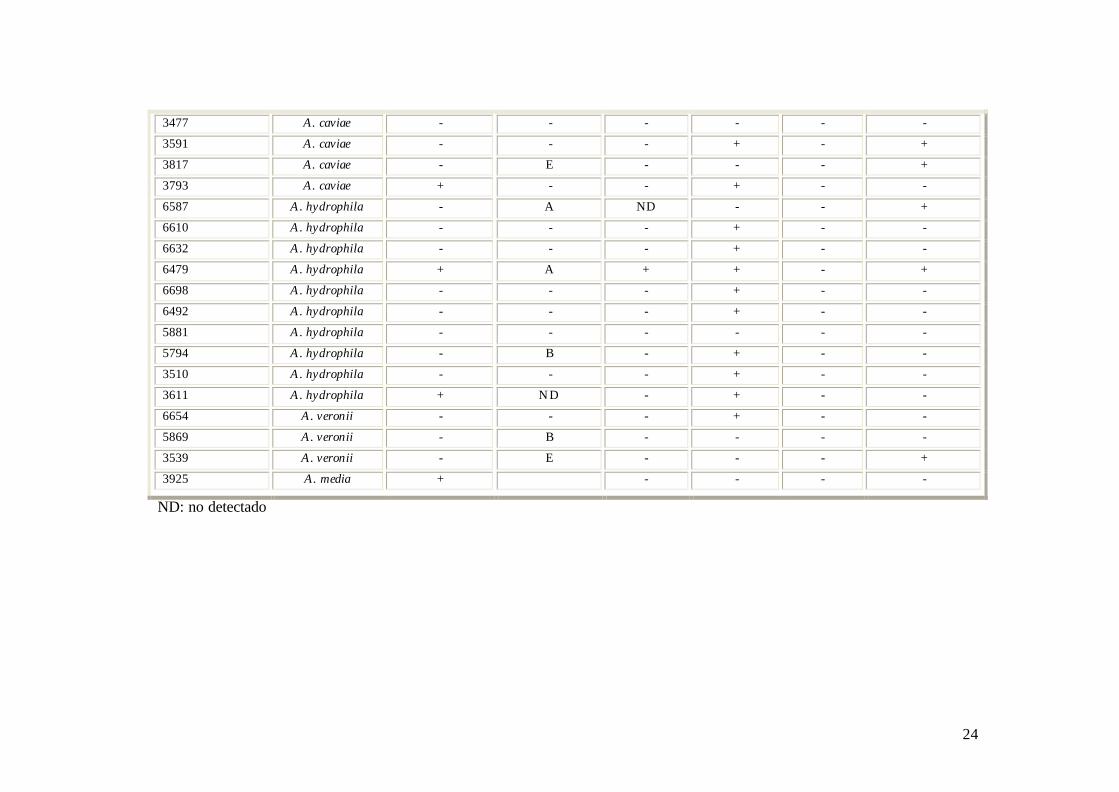
24
3477 A. caviae - - - - - -
3591 A. caviae - - - + - +
3817 A. caviae - E - - - +
3793 A. caviae + - - + - -
6587 A. hydrophila - A ND - - +
6610 A. hydrophila - - - + - -
6632 A. hydrophila - - - + - -
6479 A. hydrophila + A + + - +
6698 A. hydrophila - - - + - -
6492 A. hydrophila - - - + - -
5881 A. hydrophila - - - - - -
5794 A. hydrophila - B - + - -
3510 A. hydrophila - - - + - -
3611 A. hydrophila + ND - + - -
6654 A. veronii - - - + - -
5869 A. veronii - B - - - -
3539 A. veronii - E - - - +
3925 A. media + - - - -
ND: no detectado

25
Meta 4.- Análisis de las secuencias y establecimiento del número de copias de
elementos genéticos mediante Southern blot.
Placa autoradiográfica de la hibridación Southern. Cepas de Aeromonas spp. las cuales contenían el gen del la transposasa del Tn5393 y cepas resistentes a estreptomicina. Carril 1 E. coli C600 Tn5393 testigo positivo. Carril 2 A. hydrophila 839 testigo negativo. Carriles 3 – 34 cepas problema de Aeromonas spp. CORRELACIÓN ENTRE GENES DE RESISTENCIA Y RESISTENCIAS POR ESPECIES. Como se mencionó anteriormente en diferentes cepas de Aeromonas spp. se
detectaron diferentes genes de resistencia (relacionados a integrones y
transposones) y el fenótipo de resistencia. Sin embargo en algunas cepas se
detectaron genes de resistencia pero no el fenotipo
Correlación entre genes de resistencia y fenotipo de resistencia.
# CEPA ESPECIE GENES DE RESISTENCIA PERFIL DE RESISTENCIA
3591 A. caviae dhfr15, cmlA, aadA2 TMR , CS, ETS
6597 A. caviae dhfr1, cmlA, aadA2 TMR, CS, ETS
6661 A. caviae oxa2, aadA1, oxa2 OXR, ETS,
3817 A. caviae oxa2, aadA1, oxa2 OXR, ETI,
5933 A. caviae tet A TER

26
3430-1 A. caviae dhfr12, aadA2, tetA TMR, ETS TER
6640 A. caviae tet A TER 6436 A. caviae tet B TER
3817 A. caviae tet E TER
3491 A. caviae tet E TER 6376 A. caviae tet E TER
6336 A. hydrophila tet E TER
5794 A. hydrophila tet B TER 6479 A. hydrophila dhfr12, aadA2, tet A TMR, ETS, TER
6587 A. hydrophila dhfr2d, catB3, aadA1, tet A TMR, CS, ETR, TER
5869 A. veronii tet B TER 3539 A. veronii aadA2, tet E ETR, TER
TM= trimetoprim, C= cloranfenicol, ET= estreptomicina, TE tetraciclina, OX= oxacilina, S sensible, R
resistente.
Con respecto a la relación entre los elementos genéticos encontrados, no se
observó relación entre la presencia de plásmidos con integrones y transposones
ya que se observaron cepas con plásmidos que no tienen integrón o transposones
y viceversa.
DISCUSIÓN
Por mucho tiempo en Aeromonas se ha estudiado la presencia de plásmidos y su
relación con factores de virulencia y resistencia a antibióticos, demostrándose
que Aeromonas contiene plásmidos de resistencia a diversos antibióticos, no
obstante su relación con factores de virulencia no es lo suficientemente clara.
Algunos autores concluyen que no hay evidencias sustentables de que la
presencia de algún plásmido influya en la virulencia de Aeromonas, aunado a ésto
Stuber y cols., en el 2003 reportan que en A. salmonicida genes del sistema de
secreción tipo III están localizados en un plásmido termolábil, recientemente se
ha relacionado la presencia de un plásmidos de 21 Kb con la virulencia en cepas

27
de A. hydrophila patógena para peces. Está cepas causan síndrome ulcerativo en
peces y éste plásmido parece regular la virulencia (Majumdar y cols., 2006).
La importancia en este trabajo de determinar la presencia de plásmidos radicó en
saber si estos elementos influyen en el perfil de resistencia de las cepas o si sólo
son los vehículos para la diseminación de otros elementos. Se buscó la presencia
de plásmidos en 41 cepas de 4 diferentes especies aisladas de heces diarreicas,
todas ellas aisladas de la misma región geográfica. Los plásmidos sólo se
obtuvieron de 3 especies: A. hydrophila, A. caviae y A. media. La especie en donde
se observó una mayor incidencia de plásmidos fue A. caviae (9/27), siguiéndole
A. hydrophila (3/10). En general la incidencia de estos elementos en este trabajo
fue del 32%, lo que coincide con los resultados de otros autores donde reportan
que hay una incidencia entre el 15 y 30 % (Chang y Bolton, 1987; Pérez-
Valdespino, 2004). En contraste, algunos otros reportan una incidencia menor a
15% o mayor al 70% (Huddleston y cols., 2006; Peres y cols., 2006; Borrego y cols.,
1991.). Además de las especies antes mencionadas se obtuvo DNA plasmídico en
la cepa de A. media (1/1) especie que se considera como patógeno emergente por
su bajo aislamiento de cuadros clínicos en nuestro país (Castro -Escarpulli y cols.,
2002).
En los perfiles plasmídicos obtenidos para cada cepa se observó que algunas
compartían el mismo perfil o alguna banda de un mismo peso molecular, lo que
puede sugerir que sea un plásmido que esté diseminándose entre la población,
sin embargo no podemos asegurar que cada banda corresponda a un plásmido,
para ello se tendría que realizar un estudio particular como someter a ese DNA a
una restricción.
Algunas cepas con plásmidos presentaron un perfil de resistencia diferente, estas
cepas presentaron resistencia a tetraciclina, ácido nalidíxico, trimetoprim,

28
eritromicina, cloranfenicol, azitromocina y ciprofloxacina; estos antibióticos
pudieran estar relacionados con la presencia de plásmidos, sin embargo se
necesitarían hacer otras pruebas, como curar al plásmido o realizar
conjugaciones, ya que algunas cepas presentaron resistencia a alguno de los
antibióticos antes mencionados pero no se les detectó DNA plasmídico. Además,
es importante tomar en cuenta que se han reportado plásmidos crípticos en A.
salmonicida, esto quiere decir que estos elementos no le confieren a la bacteria
alguna característica selectiva, por lo que no se descarta la posibilidad de que
alguno de los plásmidos encontrados sea un elemento críptico (Boyd y cols.,
2003).
Recientemente en cepas de A. hydrophila y A. caviae aisladas de vegetales en Brasil
se determinó la presencia de un plásmido relacionado con la resistencia a
tetraciclina (Peres y cols., 2006). Otros trabajos, han reportado plásmidos R
conjugativos los cuales contienen otros elementos genéticos como transposones e
integrones como es el caso de los plásmidos pFABOT, pRAS1, pASOT (Sørum y
cols., 2003; Rhodes y cols., 2000).
Otros elementos que contribuyen de forma importante en la adquisición de
nuevas resistencias son los integrones, es substancial remarcar que la búsqueda
de estos elementos no se ha llevado a cabo en cepas de Aeromonas de origen
clínico, solo en cepas de origen ambiental, por lo que los resultados obtenidos
son importantes para corroborar la transferencia horizontal entre cepas del
género Aeromonas, ya que hay la posibilidad de que presenten genes similares
cepas ambientales y patógenas.
La detección de integrones se realizó en las 41 cepas, no se tomó criterio de
selección con respecto a las resistencias que presentaban, ya que en la literatura

29
se han descrito más de 70 genes “cassette” de resistencia, además de que se han
descrito integrones descargados es decir no hay genes cassette insertados en la
región variable o los genes pueden estar interrumpidos por otros elementos
como transposones, por estas razones se decidió llevar a cabo la búsqueda de
estos elementos en todas la cepas.
La clasificación de los integrones se basa en la secuencia de la integrasa, en el
presente trabajo sólo se detectaron integrones de clase 1, que son los que se
encuentran con mayor frecuencia en las cepas aisladas de casos clínicos (Sabate y
Prats, 2002). Se caracterizan por tener la secuencia 5´CS que contiene el gen
codificante de la integrasa, la región variable y la mayoría de ellos incluye una
secuencia 3´ CS (Rowe-Magnus y Mazel., 2002).
El integrón se amplificó en tres partes, primero las 2 regiones conservadas y
después la región variable, sólo en 16 de 41 cepas se encontró el gen de la
integrasa (región 5´CS) de las cuales sólo en 10 se amplificó el extremo 3´ CS; en
la literatura se ha reportado que en esta región se puede presentar diferentes
estructuras (Gaze y cols., 2005). El primer tipo de arreglo esta formado por el
Tn402 like, el cual consta de un módulo de transposición (tni) conformado por 3
genes y el gen de una resolvasa. El segundo tipo In5 consiste de qacE∆1, sul1, orf
5, orf 6 y un módulo parcial de tni. El tercer tipo In4 contiene qacE∆1, sul1, orf 5 y
orf 6. En algunas ocasiones se han detectado integrones que carecen de los genes
qacE∆1 y sul1 en el extremo 3´ conservado (Martínez-Freijo y cols., 1998; Dalsgard
y cols., 2000; Gaze y cols., 2005). Por consiguiente las cepas que presentaron la
región 5´CS y no la región 3´CS podrían presentar una estructura diferente. Cabe
destacar que en una cepa el amplificado de la región 3´CS fue de menor tamaño
que el esperado, por lo que podría presentar una estructura diferente.

30
Para la detección de la región variable sólo se seleccionaron aquellas cepas que
presentaron ambas regiones conservadas (n= 10), para la detección de esta región
se emplearon 2 pares de iniciadores, unos reportados en la literatura (var I) y los
otros fueron diseñados en este trabajo (var II). Los iniciadores reportados en la
literatura amplificaron la región variable de 3 cepas, con tamaños entre 800 y
1500 pb.
Las cepas en donde no se obtuvo amplificado en la región variable (n=6) se
emplearon los iniciadores diseñados en este trabajo (Variable II) que hibridan
dentro de la región 5´CS y el extremo 3´CS , se amplificaron 5 de las 6 cepas
restantes, con amplificados entre 1400 y 3300 pb. En una cepa se observó una
banda menor a 300 pb, que indica que no hubo “cassette” insertado y en una
cepa no se obtuvo amplificado, sumado a esto las condiciones para amplificar
esta región fueron estrictas, la temperatura de alineamiento fue 10 grados más
arriba de la temperatura a la cual se amplificaron los testigos positivos.
Es importante el hecho de que con los iniciadores reportados en la literatura no
amplificaran la región variable de estas cepas, esto a pesar de que las secuencias
flanqueadoras están conservadas, lo que podría indicar una diferencia entre la
región variable y las regiones conservadas de nuestras cepas.
Se realizaron combinaciones de iniciadores, para la amplificación de la región
variable y parte de las regiones conservadas (resultados no mostrados), esto
sirvió para comprobar que el tamaño del amplificado era del tamaño esperado.
La literatura reporta que cuando hay sólo un “cassette” insertado el tamaño es
aproximadamente de 500 pb y nuestros amplificados fueron mayores, se sabe
que el número de cassettes insertados es variable y suele oscilar entre 1 y 4, como

31
en el caso de las cepas provenientes del estado de Hidalgo (Schmidt y cols., 2001;
García, 1999; Martínez- Freijo y cols., 1998).
Se sabe que puede haber integrones descargados (In0), esto es sin genes cassette
insertados (García, 1999), sin embargo la frecuencia de integrones descargados es
baja, como en el presente trabajo donde sólo en una cepa se encontró un integrón
descargado.
Se observaron amplificados de la región variable con el mismo tamaño y
regularmente presentaban un perfil de resistencia similar, dentro de los
antibióticos relacionados con la presencia de estos elementos encontramos a
tetraciclina, polimixina, azitromicina, eritromocina, ácido nalidíxico,
trimetoprim, cloranfenicol, estreptomicina, kanamicina, oxacilina y piperacilina.
Para corroborar que los amplificados del mismo tamaño correspondían al mismo
arreglo de genes, se sometieron a una restricción, y se secuenció una cepa de
cada patrón.
La secuencia de los amplificados de la región variable revelo que, los genes
cassette insertados correspondieron a dhfr12, dhfr15 y dfrB4, que codifican para
distintas dihidrofolato reductasas que confieren resistencia a trimetoprim.
Además se encontraron aadA1, aadA2, que confieren resistencia a
estreptomicina/espectinomicina, oxa2 resistencia a oxacilina, catB3 y cmlA4 que
determinan resistencia a cloranfenicol, y un marco de lectura abierto (orfD) cuya
función es desconocida.
En el género Aeromonas se ha descrito la presencia de genes dhfr 1, 2, 12 y16, en
cepas aisladas de peces, agua y heces de ganado (Schmidt y cols., 2001; Sørum y
cols., 2003; Casas y cols., 2004; Barlow y cols., 2004; Jacobs y Chenia, 2006; Poole y
cols., 2006). Sin embargo en cepas de Aeromonas spp. de origen clínico no se

32
tienen reportes, por lo que este trabajo es el primero en detectar el gen dhfr 12.
Este gen se ha detectado en Vibrio, Pseudomonas y coliformes todas ellas de origen
ambiental (Rosser y cols., 1999) y cepas patógenas como V. cholerae O1 implicado
en una pandemia en 1997 en Guinea-Bissau (Dalsgaard y cols. 2000), en E. coli
uropatogénica y E. coli aislada de heces diarreicas en cerdos (Solberg y cols.,
2006, Kang y cols., 2005) y en bacterias Gram positivas (Shi y cols., 2006).
El gen dhfr15 se reportó por primera vez en 1998, en cepas de E. coli aislada de
heces, en África (Adrian y cols., 1998). Este gen se ha encontrado en cepas
clínicas de Klebsiella pneumoniae y en Salmonella enterica (Fonseca y cols., 2006).
El gen dfrB4 (anteriormente dfr2d) se reportó en cepas de E. coli aisladas de orina
en Suecia (Grape y cols., 2003; Levings y cols., 2006).). Es importante destacar que
son pocos los reportes de este gen dfrB4 a pesar de que los genes dhfr están
ampliamente distribuidos y son los genes que se encuentran más frecuentemente
en los integrones y se han descrito múltiples variantes de este gen (Solberg y
cols., 2006).
El trimetoprim es una droga sintética que fue introducida para uso clínico en
Europa en 1960 y el primer reporte de resistencia a trimetoprim mediada por
plásmidos fue en 1972. Esta droga tiene una estructura similar al dihidrofolato,
por lo tanto es un inhibidor competitivo de la enzima dihidrofolato reductasa de
bacterias, hongos y protozoarios. Este antibiótico inhibe la vía de síntesis de
ácido fólico y la síntesis de bases nitrogenadas. El gen dhfr codifica para la
enzima dihidrofolato reductasa (DHFR), lo que implica la producción de más
enzima para que ésta no sea inhibida, se ha reportado la síntesis de DHFR con
menor afinidad por el trimetoprim (Alonso y Grady, 2006).

33
La resistencia a aminoglucósidos esta mediada por la producción de diferentes
enzimas como adeniltransferasa, acetiltransferasa y fosfotransferasa, los genes
que codifican para estas enzimas se encuentran distribuidos en integrones y
plásmidos, hay distintas variantes de genes que codifican para estas enzimas
(Fluit y Schmitz., 2004). Los genes aadA1 y aadA2 (anteriormente ant(3”)1a y
ant(3”)1b respectivamente) confieren resistencia a estreptomicina-
espectinomicina, han sido descritos en un amplio número de bacterias Gram
negativas aisladas del cuadros clínicos y muestras ambientales (Rosser y cols.,
1999; Levings y cols., 2006; Adrian y cols., 2000).
El gen aadA2 se ha identificado en A. hydrophila aislada de heces diarreicas de
cerdos (Poole y cols., 2006) y en A. salmonicida aislada de peces (Sørum y cols.,
2003). El gen aadA1 se ha encontrado en cepas de A. hydrophila, A. salmonicida, A.
sobria, A. icthiosomia y A. encheleia todas ellas aisladas de sistemas de acuacultura
en África (Jacobs y Chenia, 2006).
El gen aadA, al igual que el gen dhfr se encuentran distribuidos entre un gran
número de bacterias en diferentes áreas geográficas, es común encontrar estos
genes en un mismo arreglo (dhfr-aadA) y justamente este arreglo fue el más
común en nuestras cepas. Esto sugiere que hay flujo genético entre células de
diferentes comunidades microbianas (Adrian y cols., 2000; Gestal y cols., 2005).
El gen aadA se encontró formando parte de todos los arreglos de los integrones
encontrados (n=8), y este ha sido reportado como un gen predominante en E. coli
de origen clínico y ambiental, Vibrio cholerae O139 y S. enterica, la predominancia
de este gen sugiere que es un gen muy estable dentro del integrón, y esto pudiera
estar relacionado al uso extensivo de la estreptomicina en el control de
enfermedades de animales, plantas y el hombre (Kang y cols., 2005).

34
Otro de los genes detectados fue el gen catB3, que confiere resistencia a
cloranfenicol, el cual es un antibiótico de amplio espectro, actúa sobre un gran
número de bacterias Gram positivas y Gram negativas inhibiendo la síntesis de
proteínas. El gene catB3 codifica para una cloranfenicol acetiltransferasa (CAT),
miembro de una familia de proteínas denominadas CAT, se han descrito 3
variantes de genes catB y se describieron por primera vez en Agrobacetrium
tumefaciens (Bunny y cols., 1995).
Este gene se ha encontrado en una cepa de A. encheleia de origen ambiental (datos
no publicados). Esta cepa de A. encheleia presenta el mismo arreglo de genes
cassette, que la cepa 6587 A. hydrophila, igualmente Barlow y cols., (2004)
reportaron 2 cepas de A. veronii con el arreglo de la cepa 6587.
El gen cmlA4 es un gen que confiere resistencia a cloranfenicol, pero el
mecanismo de resistencia es diferente al codificado por el gen catB3, el gen cmlA4
codifica para una proteína de membrana externa encargada de expulsar al
antibiótico.
Las cepas de Aeromonas que contienen este gen presentan un arreglo dhfr15-
cmlA4-aadA2, este mismo arreglo se presenta en cepas de Salmonella enterica y
Klebsiella pneumoniae (Michael y cols., 2005).
Las cepas de Aeromonas se caracterizan porque producen una gran variedad de ß-
lactamasas de diferentes clases. En el presente trabajo se detectaron dos cepas se
detectó el gen oxa2 que codifica para una ß-lactamasa clase D y confiere
resistencia a oxacilina este gen se encuentra duplicado en el mismo arreglo .
Además dicho gen forma parte de un gran número de integrones, incluyendo
una cepa de A. sobria de origen ambiental (Jacobs y Chenia, 2006).
En este trabajo se identificaron dos cepas de A. caviae con el arreglo oxa2-aadA2-
oxa2-orfD, dicho arreglo se encontró formando parte del plásmido R46, aislado

35
en Europa en la década de los 70´s en S. typhimurium (Stokes y Hall, 1992; Recchia
y Hall, 1995).
Los integrones confieren ventajas selectivas y raramente capturan genes
indispensables para la bacteria, se caracterizan por tener una gran versatilidad en
el intercambio y sobre todo en la reserva de cassettes, por ello han contribuido a
una rápida adaptación al cambio que se presenta en los nichos ecológicos (Sabate
y Prats, 2002). En el presente estudio las cepas que se estudiaron contienen
integrones y estos poseen genes de resistencia que se presentan en arreglos
conservados los cuales se encuentran distribuidos en una gran variedad de
bacterias de diferentes orígenes, ratificando la movilidad horizontal de genes.
En algunas cepas con integrón se detectaron genes de resistencia, pero no el
fenotipo, esto se puede deber a la ubicación del gen con respecto al promotor, ya
que entre mas alejado de esté no se expresa, otro factor que podría influir es que
el gen mas cercano al promotor presente secuencias terminadoras que eviten la
transcripción de los genes subsecuentes o que los genes no sean funcionales. Es
importante tomar en cuenta que para determinar los perfiles de resistencia se
empleo la técnica de difusión en disco la cual es una prueba con diversas
desventajas para conocer si la cepa es sensible o resistente, por lo que seria
importante determinar la concentración mínima inhibitoria para los antibióticos
de interés.
Rowe-Magnus en el 2001 demostró la presencia de estos integrones en
microorganismos ancestrales de diferentes géneros, corroborando así que los
integrones son estructuras antiguas que han ido evolucionando conjuntamente
con el genoma bacteriano. Estas estructuras muestran alta estabilidad genética,
incluyendo los arreglos de genes, por periodos prolongados de tiempo (Dubois y

36
cols., 2007); en nuestro trabajo los integrones muestran arreglos descritos en otras
bacterias de diferentes nichos, lo cual se podría estar relacionado a su estabilidad,
sin embargo seria importante estudiar si la integrasa es funcional y por esta
razón no hay recambio de genes “cassettes” y por ellos son conservados.
En este trabajo se observó que las cepas con plásmidos no tienen integrón y
viceversa, por lo que estos elementos se podrían encontrar insertados en el
cromosoma, tal como se ha descrito para cepas de Salmonella (Zhang y cols.,
2004).
Otros genes distribuidos en un gran número de bacterias son los genes tet, que
confieren resistencia a tetraciclina. Las tetraciclinas inhiben la síntesis de
proteínas y son antimicrobianos de amplio espectro, actúan sobre un gran
número de bacterias Gram negativas, Gram positivas, microorganismos atípicos
como clamidias, micoplasmas, rickettsias y protozoarios. Hasta el momento en la
literatura se han descrito 35 genes de resistencia a tetraciclina en bacterias, estos
genes codifican para tres diferentes mecanismos de resistencia: proteínas de
protección para el ribosoma, proteínas de membrana que regulan en flujo del
antibiótico e inactivación del antibiótico vía enzimática (Chopra, 2001; Waturangi
y cols., 2003).
En Aeromonas spp. se han descrito los genes tet A, B, D, E, H y tet 31, estos genes
codifican para proteínas de membrana que regulan en flujo del antibiótico, no se
ha descrito algún gen que codifique para un mecanismo distinto de resistencia
(Chopra, 2001). En este trabajo se detectaron los genes tet A, E, y B; la especie que
presentó una notable incidencia de genes tet es A. caviae, seguida de A. hydrophila
y finalmente A. veronii. Los genes tet A y E se encuentran distribuidos entre las

37
especies antes mencionadas, sin embargo los genes tetB no se presentaron en
cepas de A. caviae.
Jacobs y Chenia, (2006) reporta cepas de Aeromonas spp. aisladas de agua de
granjas de peces, donde el 75 % de las cepas contienen genes tet A, B, E, D y H; la
especie que presenta un mayor número de estos genes es A. sobria, seguida de A.
hydrophila, no se reporta ninguna cepa de A. caviae.
Otro trabajo indica que el gen tet E esta ampliamente diseminado en cepas de A.
hydrophila y A. salmonicida aisladas de granjas de peces, y este gen esta presente
en un plásmido conjugativo (Agersø y cols., 2006). Cepas aisladas de aguas
domésticas y hospitalarias, presentan genes tet A, la especie de mayor incidencia
es A. hydrophila, seguida de A. veronii y A. salmonicida, sólo se reporta una cepa de
A. caviae (Rhodes y cols., 2000). La incidencia de genes tet en nuestro trabajo es
menor que la reportada en Aeromonas spp. de origen ambiental donde se
presentan con mayor frecuencia posiblemente debido al uso
indiscriminado de la tetraciclina en agricultura y acuicultura donde se utiliza
como promotor de crecimiento (Schmidt y cols., 2001; Miranda y cols., 2003;
Jacobs y Chenia, 2006).
Los genes tet se encuentran distribuidos en una amplia variedad de bacterias
aisladas de humanos, animales y del ambiente, la mayoría de estos genes se
encuentran asociados a plásmidos o transposones, lo cual explica su amplia
distribución entre diversas especies bacterianas.
En el 2004 Pezzella y cols., reportaron alta incidencia de genes tet en cepas de S.
enterica aislada de animales en Italia. El reporte de genes tet en cepas aisladas de
cuadros clínicos es variable, en cepas de Acinetobacter el 94% de las cepas

38
presentaron genes tet y en cepas de E. coli solo el 12% de las cepas presentó genes
de resistencia a tetraciclina (Karami y cols., 2006; Guardabassi y cols., 2000).
Es importante tomar en cuenta que la resistencia a tetraciclina puede estar
influenciada por factores geográficos (Chopra, 2001).
Como se menciona inicialmente otros elementos de gran importancia en la
transferencia de genes de resistencia son los transposones. El transposón Tn1721
se describió desde 1981 en un plásmido conjugativo en E. coli, se encuentra
distribuido ampliamente, y es el responsable de la amplia diseminación de
resistencia a tetraciclina en bacterias Gram negativas (Schmitt y cols., 1979).
Diversos estudios muestran que los genes de resistencia a tetraciclina se
encuentran en transposones como: tet A en Tn1721, tet B en Tn10, tet D el cual
esta flanqueado por dos secuencias de inserción (IS26), tet G en un integrón y tet
H está en Tn5706 (Schnabel y Jones, 1999).
En cepas clínicas se ha observado un incremento en las cepas resistentes a
tetraciclina debido a que forma parte de los antibióticos recomendados para el
tratamiento de infecciones intestinales, aunado a la contaminación ambiental.
En nuestro trabajo se detecto el Tn1721 en 4 cepas de origen clínico, con lo que
podemos decir que este elemento es de amplia distribución ya que éste se ha
reportado en cepas de Aeromonas spp. de origen ambiental (Rhodes y cols., 2000;
Sørum y cols., 2003).
El Tn1721 el cual se presentó en cepas de A. caviae (3/27) y A. hydrophila (1/10),
se reporta en la literatura mayor incidencia de este elemento en cepas de A.
hydrophila, seguida de A. veronii y A. caviae en estas cepas el transposón se
encontró en un plásmido conjugativo (pFABOT) el cual también contiene un

39
integrón (Rhodes y cols., 2000). Este transposón se ha descrito también en cepas
de A. salmonicida formando parte del plásmido conjugativo pRAS1 y del
plásmido pASOT, ambos plásmidos contienen un integrón clase 1 (Sørum y cols.,
2003).
L´Abee-Lund y Sørum en el 2000 realizaron un estudio donde ponen de
manifiesto la presencia del transposón Tn5393 el cual contiene genes que
codifican la resistencia a estreptomicina (strA-strB ). Dicho transposón se
encuentra en un plásmido conjugativo (pRAS2) en una cepa de A. salmonicida
subsp. salmonicida patógena de peces. Este plásmido también confiere resistencia
a tetraciclina y a sulfas. Cabe mencionar que este elemento es de alta incidencia
en bacterias fitopatógenas como Erwinia amilovora, Pseudomonas syringae y
Xantomonas campestre (Han y cols., 2004).
Los genes strA-strB se han aislado de bacterias que infectan animales y humanos
asociados al Tn5393 y a plásmidos (Han y cols., 2004; Matengoli y Rossolini,
2005). Se sabe que los transposones son capaces de formar diversas
combinaciones por la integración de diversos elementos y este transposón no es
excepción, ya que este elemento se relaciona con genes de resistencia a metales
pesados (Rice, 2002).
Para la detección de este elemento en nuestras cepas de origen clínico se
seleccionaron aquellas cepas resistentes a estreptomicina, en estas cepas sólo se
detectó el gen de la transposasa del Tn5393. A pesar de la aparente presencia de
la transposasa de Tn5393 no se pudo amplificar los genes strA-strB característicos
de este transposón, por lo que probablemente se trate de genes atípicos o exista
en estas cepas una variante del transposón.

40
Para descartar la presencia de falsos negativos por la técnica de PCR, se empleo
la hibridación Southern, utilizando como sonda el amplificado de los genes strA-
B, con esta técnica se encontraron tres cepas que dieron señal en la placa
autoradigráfica, una de ella con señal intensa, lo cual indica que esta cepa
presenta los genes strA-B, y que pudieran presentar una región diferente donde
pegan los iniciadores por ello no se detectaron mediante PCR.
Con este resultado podemos decir que el Tn5393 completo fue encontrado en una
cepa de Aeromonas de origen clínico.
La señal débil que presentaron dos cepas de Aeromonas spp. se pudo deber a que
son genes con alta homología a los genes str A-B, es importante resaltar que las
condiciones de hibridación fueron a una elevada astringencia. Por lo anterior es
importante llevar acabo la caracterización de estos genes.
Otra resistencia asociada a este elemento es a metales pesados, sin embargo no
parece haber una relación entre esta resistencia y la presencia de la transposasa
de Tn5393 en nuestras cepas, ya que cepas resistentes no amplificaron el gen de
la transposasa y viceversa.
Los transposones que se estudian en este trabajo pertenecen a la familia Tn3, del
subgrupo Tn21, se ha descrito que estos elementos tienen una diseminación
global y generalmente están relacionados con integrones clase 1, los integrones
no son móviles por si mismos, por ello se hallan con frecuencia en transposones
que a su vez se encuentran en plásmidos conjugativos o movilizables, por lo que
su movilidad horizontal y vertical está asegurada.
En este trabajo se detectaron elementos genéticos como plásmidos, trasnposones
e integrones en cepas de A. caviae, A. hydrophila, y A. veronii de origen clínico,
cabe mencionar que la literatura solo muestra reportes de la detección de estos

41
elementos en cepas de origen ambiental, por lo que este es el primer trabajo en
cepas aisladas de origen clínico.
La cepa más frecuentemente aislada en este trabajo fue A. caviae. Es importante
recordar que esta es la especie que se aísla en mayor número de cuadros
diarreicos (Castro-Escarpulli y cols., 2002), sin embargo el aislamiento de esta
especie en agua y pescado es menos frecuente (Castro-Escarpulli y cols., 2003,
Vásquez-Ocampo 2005). Aunado a lo anterior, se ha demostrado que cepas de
Aeromonas de la misma especie se pueden agrupar con base en su origen de
aislamiento, lo cual indica que las diferentes poblaciones, pueden ocupar un
nicho diferente (Aguilera y cols., 2005). Con base en estos antecedentes es posible
especular que existan diferencias en los elementos genéticos y los genes de
resistencia de estas cepas con respecto a los de las cepas de origen ambiental. Por
lo anterior, es importante realizar un estudio en cepas aisladas de muestras
ambientales, para comparar la incidencia de estos elementos con la observada en
cepas clínicas. Lo anterior permitiría abundar en el conocimiento de la
diversidad de los genes de resistencia en nuestro país.
CONCLUSIONES
• Las especies que se aislaron con mayor frecuencia de cuadros clínicos
intestinales son A. hydrophila A. caviae y A. veronii. Siendo de estas tres A.
caviae la que se presentó en mayor número de aislamientos.
• El 32 % (13/41) de las cepas estudiadas presentaron plásmidos entre 3 y 53
Kb.
• Se detectaron integrones clase 1 (n=9), con genes de resistencia que se
encuentran distribuidos en un gran número de bacterias,
independientemente del origen y el área geográfica, los genes “cassette”
insertados corresponden a dhfr12, dhfr15, dfrB4, que confieren resistencia a

42
trimetoprim; aadA1, aadA2, que confieren resistencia a
estreptomicina/espectinomicina, oxa2 que confieren resistencia a
oxacilina, catB3 y cmlA4 que determinan resistencia a cloranfenicol, y un
marco de lectura abierto (orfD).
• Las cepas de origen clínico resistentes a tetraciclina poseen los genes tet A,
tet E y tet B, genes ampliamente distribuidos en cepas de Aeromonas spp.
de origen ambiental.
• Se detectó el transposón Tn1721 en 4 de las 5 cepas resistentes a
tetraciclina con el gen tet A.
• Se detectó el Tn5393 completo en una cepa y en el 63% (26/41) de las
cepas de Aeromonas solo se amplificó el gen de la transposasa lo que
sugiere la presencia de un transposón diferente.
IMPACTO
Los antibióticos y otros agentes antimicrobianos se usan ampliamente en
diversas actividades, entre los cuales encontramos terapia y profilaxis en
medicina humana y veterinaria e incluso en la agricultura. El uso indiscriminado
de antibióticos repercute en el aumento de infecciones por bacterias resistentes a
los antibióticos en las poblaciones humanas y animales, alteraciones importantes
de las relaciones ecológicas y en la contaminación del medio ambiente (Cabello,
2003).
En la práctica médica las infecciones causadas por Aeromonas son tratadas, con
fluoroquinolonas, trimetoprim-sulfametoxazol, aminoglucósidos, imipenem,
meropenem, cefalosporinas y tetraciclinas, algunos de éstos son empleados
particularmente para el tratamiento de infecciones gastrointestinales (Soler, 2003;
Coria y cols., 2001).
Algunas cepas con plásmidos presentaron un perfil de resistencia diferente, estas
cepas presentaron resistencia a tetraciclina, ácido nalidíxico, trimetoprim,
eritromicina, cloranfenicol, azitromocina y ciprofloxacina; estos antibióticos

43
pudieran estar relacionados con la presencia de plásmidos, sin embargo se
necesitarían hacer otras pruebas, como curar al plásmido o realizar
conjugaciones, ya que algunas cepas presentaron resistencia a alguno de los
antibióticos antes mencionados pero no se les detectó DNA plasmídico. Además,
es importante tomar en cuenta que se han reportado plásmidos crípticos en A.
salmonicida, esto quiere decir que estos elementos no le confieren a la bacteria
alguna característica selectiva, por lo que no se descarta la posibilidad de que
alguno de los plásmidos encontrados sea un elemento críptico (Boyd y cols.,
2003).
El impacto ambiental que coadyuva en la investigación del ambiente es porque
los centros de atención en salud, especialmente los hospitales, constituyen
importantes puntos de origen de descargas de antibióticos hacia el ambiente,
produciendo un fuerte impacto en la composición física, química y biológica de
los cuerpos receptores.
Aeromonas adquiere resistencia por la presencia de elementos genéticos en los
aislados de origen ambiental; sin embargo las cepas descritas en este trabajo
provenientes de cuadros clínicos en especial de cuadros diarreicos en las cuales
la presencia de elementos genéticos (plásmidos, integrones y transposones) se
pudieron asociar a la resistencia antimicrobiana que pueden transferirse a otras
cepas.
El conocimiento de la presencia de estos elementos genéticos en cepas aisladas de
cuadros diarreicos en nuestro país destaca la importancia de llamar la atención
de las autoridades sanitarias en el control de uso indiscriminado de antibióticos
en prácticas médicas, agrícolas y veterinarias por la diseminación de cepas de
Aeromonas resistentes a antimicrobianos presentes en el medio ambiente
complicando de manera importante la efectividad de los tratamientos
empleados comúnmente.


















