Diversidad de hongos endófitos asociados al frailejón ...
81
Universidad Andina Simón Bolívar Sede Ecuador Área de Ambiente y Sustentabilidad Maestría en Cambio Climático, Sustentabilidad y Desarrollo Diversidad de hongos endófitos asociados al frailejón (Espeletia hartwegiana) en un marco de cambio climático, páramo de Romerales, Quindío, Colombia Cristhian Aristizabal Ortiz Tutor: Jhon Alexander Osorio Romero Quito, 2021
Transcript of Diversidad de hongos endófitos asociados al frailejón ...

Universidad Andina Simoacuten Boliacutevar
Sede Ecuador
Aacuterea de Ambiente y Sustentabilidad
Maestriacutea en Cambio Climaacutetico Sustentabilidad y Desarrollo
Diversidad de hongos endoacutefitos asociados al frailejoacuten (Espeletia
hartwegiana) en un marco de cambio climaacutetico paacuteramo de Romerales
Quindiacuteo Colombia
Cristhian Aristizabal Ortiz
Tutor Jhon Alexander Osorio Romero
Quito 2021
2
3
Claacuteusula de cesioacuten de derecho de publicacioacuten
Yo Cristhian Aristizabal Ortiz autor del trabajo intitulado ldquoDiversidad de hongos
endoacutefitos asociados al frailejoacuten (Espeletia hartwegiana) en un marco de cambio climaacutetico en el
paacuteramo de Romerales Quindiacuteo Colombiardquo mediante el presente documento dejo constancia de
que la obra es de mi exclusiva autoriacutea y produccioacuten que la he elaborado para cumplir con uno de
los requisitos previos para la obtencioacuten del tiacutetulo de Magiacutester en Cambio Climaacutetico
Sustentabilidad y Desarrollo en la Universidad Andina Simoacuten Boliacutevar Sede Ecuador
1 Cedo a la Universidad Andina Simoacuten Boliacutevar Sede Ecuador los derechos exclusivos de
reproduccioacuten comunicacioacuten puacuteblica distribucioacuten y divulgacioacuten durante 36 meses a partir de mi
graduacioacuten pudiendo por lo tanto la Universidad utilizar y usar esta obra por cualquier medio
conocido o por conocer siempre y cuando no se lo haga para obtener beneficio econoacutemico Esta
autorizacioacuten incluye la reproduccioacuten total o parcial en los formatos virtual electroacutenico digital
oacuteptico como usos en red local y en internet
2 Declaro que en caso de presentarse cualquier reclamacioacuten de parte de terceros respecto de los
derechos de autora de la obra antes referida yo asumireacute toda responsabilidad frente a terceros y a
la Universidad
3 En esta fecha entrego a la Secretariacutea General el ejemplar respectivo y sus anexos en formato
impreso y digital o electroacutenico
Firma 23072021
Cristhian Aristizabal Ortiz
4
5
Resumen
Debido al aumento global de las temperaturas impulsado por el cambio climaacutetico de origen
antropogeacutenico y a la poca informacioacuten que se tiene sobre hongos endoacutefitos en especies de los
paacuteramos esta investigacioacuten buscoacute identificar y caracterizar la diversidad de hongos endoacutefitos del
frailejoacuten Espeletia hartwegiana Esta especie habita en los paacuteramos desde 3000 a 5000 metros de
lo que la hace una especie maacutes vulnerable a los aumentos de temperatura que se pronostican para
el antildeo 2100 de hasta 25degC La identificacioacuten y caracterizacioacuten de los hongos endoacutefitos se realizoacute
por medio de teacutecnicas filogenia molecular basado en datos de secuencias de genes muacuteltiples de
las regiones espaciadoras transcritas internas (ITS) en donde se encontraron un total de 15
geacuteneros de endoacutefitos como Nigrospora Trichoderma y Diaporthe con un total de 35 especies
Los hongos endoacutefitos encontrados podriacutean tener un rol vital en la supervivencia de esta especie
de frailejoacuten mediando la forma en coacutemo responde al estreacutes abioacutetico producido por el cambio
climaacutetico como aumento de la temperatura cambio en las precipitaciones y sequiacuteas ademaacutes del
estreacutes bioacutetico como virus bateriacuteas insectos y hongos fitopatoacutegenos
Palabras clave Cambio Climaacutetico Filogenia Molecular Hongos Endoacutefitos Paacuteramos
6
7
Con amor para mis padres Fernando Ivaacuten Aristizabal y mi madre Mariacutea Ligia
Ortiz
A Norberto Aristizabal y Natalia Aristizabal por creer en miacute
A mi hijo Joseacute David Aristizabal por representar las nuevas generaciones
A mi tutor Jhon Alexander Osorio por tenerme paciencia en este proceso de
aprendizaje y compartirme sus conocimientos
A Tatiana Tapias por darme su amor y llevarme por el camino de la virtud
siempre centrado
A la maestriacutea en Cambio Climaacutetico Sustentabilidad y Desarrollo por siempre
estar disponibles para miacute
A la Universidad Andina Simoacuten Boliacutevar por apoyar mi tesis de investigacioacuten
A la Repuacuteblica del Ecuador por abrirme sus puertas y darme un mejor futuro
8
9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
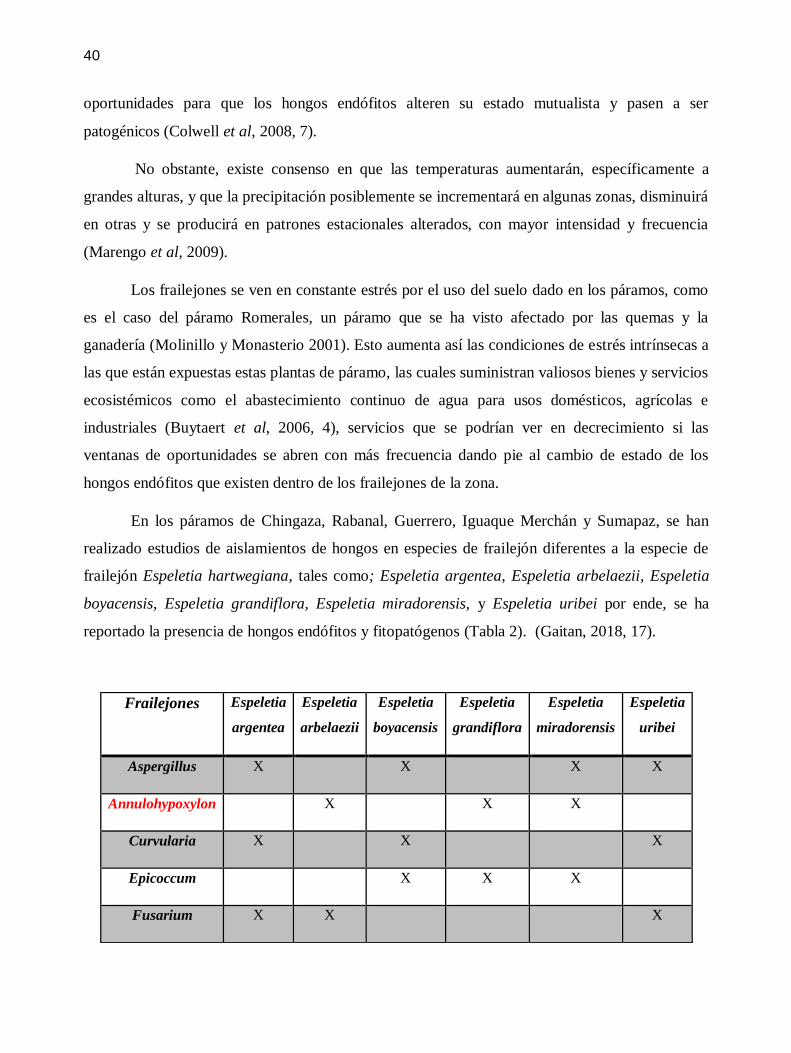
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

2
3
Claacuteusula de cesioacuten de derecho de publicacioacuten
Yo Cristhian Aristizabal Ortiz autor del trabajo intitulado ldquoDiversidad de hongos
endoacutefitos asociados al frailejoacuten (Espeletia hartwegiana) en un marco de cambio climaacutetico en el
paacuteramo de Romerales Quindiacuteo Colombiardquo mediante el presente documento dejo constancia de
que la obra es de mi exclusiva autoriacutea y produccioacuten que la he elaborado para cumplir con uno de
los requisitos previos para la obtencioacuten del tiacutetulo de Magiacutester en Cambio Climaacutetico
Sustentabilidad y Desarrollo en la Universidad Andina Simoacuten Boliacutevar Sede Ecuador
1 Cedo a la Universidad Andina Simoacuten Boliacutevar Sede Ecuador los derechos exclusivos de
reproduccioacuten comunicacioacuten puacuteblica distribucioacuten y divulgacioacuten durante 36 meses a partir de mi
graduacioacuten pudiendo por lo tanto la Universidad utilizar y usar esta obra por cualquier medio
conocido o por conocer siempre y cuando no se lo haga para obtener beneficio econoacutemico Esta
autorizacioacuten incluye la reproduccioacuten total o parcial en los formatos virtual electroacutenico digital
oacuteptico como usos en red local y en internet
2 Declaro que en caso de presentarse cualquier reclamacioacuten de parte de terceros respecto de los
derechos de autora de la obra antes referida yo asumireacute toda responsabilidad frente a terceros y a
la Universidad
3 En esta fecha entrego a la Secretariacutea General el ejemplar respectivo y sus anexos en formato
impreso y digital o electroacutenico
Firma 23072021
Cristhian Aristizabal Ortiz
4
5
Resumen
Debido al aumento global de las temperaturas impulsado por el cambio climaacutetico de origen
antropogeacutenico y a la poca informacioacuten que se tiene sobre hongos endoacutefitos en especies de los
paacuteramos esta investigacioacuten buscoacute identificar y caracterizar la diversidad de hongos endoacutefitos del
frailejoacuten Espeletia hartwegiana Esta especie habita en los paacuteramos desde 3000 a 5000 metros de
lo que la hace una especie maacutes vulnerable a los aumentos de temperatura que se pronostican para
el antildeo 2100 de hasta 25degC La identificacioacuten y caracterizacioacuten de los hongos endoacutefitos se realizoacute
por medio de teacutecnicas filogenia molecular basado en datos de secuencias de genes muacuteltiples de
las regiones espaciadoras transcritas internas (ITS) en donde se encontraron un total de 15
geacuteneros de endoacutefitos como Nigrospora Trichoderma y Diaporthe con un total de 35 especies
Los hongos endoacutefitos encontrados podriacutean tener un rol vital en la supervivencia de esta especie
de frailejoacuten mediando la forma en coacutemo responde al estreacutes abioacutetico producido por el cambio
climaacutetico como aumento de la temperatura cambio en las precipitaciones y sequiacuteas ademaacutes del
estreacutes bioacutetico como virus bateriacuteas insectos y hongos fitopatoacutegenos
Palabras clave Cambio Climaacutetico Filogenia Molecular Hongos Endoacutefitos Paacuteramos
6
7
Con amor para mis padres Fernando Ivaacuten Aristizabal y mi madre Mariacutea Ligia
Ortiz
A Norberto Aristizabal y Natalia Aristizabal por creer en miacute
A mi hijo Joseacute David Aristizabal por representar las nuevas generaciones
A mi tutor Jhon Alexander Osorio por tenerme paciencia en este proceso de
aprendizaje y compartirme sus conocimientos
A Tatiana Tapias por darme su amor y llevarme por el camino de la virtud
siempre centrado
A la maestriacutea en Cambio Climaacutetico Sustentabilidad y Desarrollo por siempre
estar disponibles para miacute
A la Universidad Andina Simoacuten Boliacutevar por apoyar mi tesis de investigacioacuten
A la Repuacuteblica del Ecuador por abrirme sus puertas y darme un mejor futuro
8
9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

3
Claacuteusula de cesioacuten de derecho de publicacioacuten
Yo Cristhian Aristizabal Ortiz autor del trabajo intitulado ldquoDiversidad de hongos
endoacutefitos asociados al frailejoacuten (Espeletia hartwegiana) en un marco de cambio climaacutetico en el
paacuteramo de Romerales Quindiacuteo Colombiardquo mediante el presente documento dejo constancia de
que la obra es de mi exclusiva autoriacutea y produccioacuten que la he elaborado para cumplir con uno de
los requisitos previos para la obtencioacuten del tiacutetulo de Magiacutester en Cambio Climaacutetico
Sustentabilidad y Desarrollo en la Universidad Andina Simoacuten Boliacutevar Sede Ecuador
1 Cedo a la Universidad Andina Simoacuten Boliacutevar Sede Ecuador los derechos exclusivos de
reproduccioacuten comunicacioacuten puacuteblica distribucioacuten y divulgacioacuten durante 36 meses a partir de mi
graduacioacuten pudiendo por lo tanto la Universidad utilizar y usar esta obra por cualquier medio
conocido o por conocer siempre y cuando no se lo haga para obtener beneficio econoacutemico Esta
autorizacioacuten incluye la reproduccioacuten total o parcial en los formatos virtual electroacutenico digital
oacuteptico como usos en red local y en internet
2 Declaro que en caso de presentarse cualquier reclamacioacuten de parte de terceros respecto de los
derechos de autora de la obra antes referida yo asumireacute toda responsabilidad frente a terceros y a
la Universidad
3 En esta fecha entrego a la Secretariacutea General el ejemplar respectivo y sus anexos en formato
impreso y digital o electroacutenico
Firma 23072021
Cristhian Aristizabal Ortiz
4
5
Resumen
Debido al aumento global de las temperaturas impulsado por el cambio climaacutetico de origen
antropogeacutenico y a la poca informacioacuten que se tiene sobre hongos endoacutefitos en especies de los
paacuteramos esta investigacioacuten buscoacute identificar y caracterizar la diversidad de hongos endoacutefitos del
frailejoacuten Espeletia hartwegiana Esta especie habita en los paacuteramos desde 3000 a 5000 metros de
lo que la hace una especie maacutes vulnerable a los aumentos de temperatura que se pronostican para
el antildeo 2100 de hasta 25degC La identificacioacuten y caracterizacioacuten de los hongos endoacutefitos se realizoacute
por medio de teacutecnicas filogenia molecular basado en datos de secuencias de genes muacuteltiples de
las regiones espaciadoras transcritas internas (ITS) en donde se encontraron un total de 15
geacuteneros de endoacutefitos como Nigrospora Trichoderma y Diaporthe con un total de 35 especies
Los hongos endoacutefitos encontrados podriacutean tener un rol vital en la supervivencia de esta especie
de frailejoacuten mediando la forma en coacutemo responde al estreacutes abioacutetico producido por el cambio
climaacutetico como aumento de la temperatura cambio en las precipitaciones y sequiacuteas ademaacutes del
estreacutes bioacutetico como virus bateriacuteas insectos y hongos fitopatoacutegenos
Palabras clave Cambio Climaacutetico Filogenia Molecular Hongos Endoacutefitos Paacuteramos
6
7
Con amor para mis padres Fernando Ivaacuten Aristizabal y mi madre Mariacutea Ligia
Ortiz
A Norberto Aristizabal y Natalia Aristizabal por creer en miacute
A mi hijo Joseacute David Aristizabal por representar las nuevas generaciones
A mi tutor Jhon Alexander Osorio por tenerme paciencia en este proceso de
aprendizaje y compartirme sus conocimientos
A Tatiana Tapias por darme su amor y llevarme por el camino de la virtud
siempre centrado
A la maestriacutea en Cambio Climaacutetico Sustentabilidad y Desarrollo por siempre
estar disponibles para miacute
A la Universidad Andina Simoacuten Boliacutevar por apoyar mi tesis de investigacioacuten
A la Repuacuteblica del Ecuador por abrirme sus puertas y darme un mejor futuro
8
9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

4
5
Resumen
Debido al aumento global de las temperaturas impulsado por el cambio climaacutetico de origen
antropogeacutenico y a la poca informacioacuten que se tiene sobre hongos endoacutefitos en especies de los
paacuteramos esta investigacioacuten buscoacute identificar y caracterizar la diversidad de hongos endoacutefitos del
frailejoacuten Espeletia hartwegiana Esta especie habita en los paacuteramos desde 3000 a 5000 metros de
lo que la hace una especie maacutes vulnerable a los aumentos de temperatura que se pronostican para
el antildeo 2100 de hasta 25degC La identificacioacuten y caracterizacioacuten de los hongos endoacutefitos se realizoacute
por medio de teacutecnicas filogenia molecular basado en datos de secuencias de genes muacuteltiples de
las regiones espaciadoras transcritas internas (ITS) en donde se encontraron un total de 15
geacuteneros de endoacutefitos como Nigrospora Trichoderma y Diaporthe con un total de 35 especies
Los hongos endoacutefitos encontrados podriacutean tener un rol vital en la supervivencia de esta especie
de frailejoacuten mediando la forma en coacutemo responde al estreacutes abioacutetico producido por el cambio
climaacutetico como aumento de la temperatura cambio en las precipitaciones y sequiacuteas ademaacutes del
estreacutes bioacutetico como virus bateriacuteas insectos y hongos fitopatoacutegenos
Palabras clave Cambio Climaacutetico Filogenia Molecular Hongos Endoacutefitos Paacuteramos
6
7
Con amor para mis padres Fernando Ivaacuten Aristizabal y mi madre Mariacutea Ligia
Ortiz
A Norberto Aristizabal y Natalia Aristizabal por creer en miacute
A mi hijo Joseacute David Aristizabal por representar las nuevas generaciones
A mi tutor Jhon Alexander Osorio por tenerme paciencia en este proceso de
aprendizaje y compartirme sus conocimientos
A Tatiana Tapias por darme su amor y llevarme por el camino de la virtud
siempre centrado
A la maestriacutea en Cambio Climaacutetico Sustentabilidad y Desarrollo por siempre
estar disponibles para miacute
A la Universidad Andina Simoacuten Boliacutevar por apoyar mi tesis de investigacioacuten
A la Repuacuteblica del Ecuador por abrirme sus puertas y darme un mejor futuro
8
9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

5
Resumen
Debido al aumento global de las temperaturas impulsado por el cambio climaacutetico de origen
antropogeacutenico y a la poca informacioacuten que se tiene sobre hongos endoacutefitos en especies de los
paacuteramos esta investigacioacuten buscoacute identificar y caracterizar la diversidad de hongos endoacutefitos del
frailejoacuten Espeletia hartwegiana Esta especie habita en los paacuteramos desde 3000 a 5000 metros de
lo que la hace una especie maacutes vulnerable a los aumentos de temperatura que se pronostican para
el antildeo 2100 de hasta 25degC La identificacioacuten y caracterizacioacuten de los hongos endoacutefitos se realizoacute
por medio de teacutecnicas filogenia molecular basado en datos de secuencias de genes muacuteltiples de
las regiones espaciadoras transcritas internas (ITS) en donde se encontraron un total de 15
geacuteneros de endoacutefitos como Nigrospora Trichoderma y Diaporthe con un total de 35 especies
Los hongos endoacutefitos encontrados podriacutean tener un rol vital en la supervivencia de esta especie
de frailejoacuten mediando la forma en coacutemo responde al estreacutes abioacutetico producido por el cambio
climaacutetico como aumento de la temperatura cambio en las precipitaciones y sequiacuteas ademaacutes del
estreacutes bioacutetico como virus bateriacuteas insectos y hongos fitopatoacutegenos
Palabras clave Cambio Climaacutetico Filogenia Molecular Hongos Endoacutefitos Paacuteramos
6
7
Con amor para mis padres Fernando Ivaacuten Aristizabal y mi madre Mariacutea Ligia
Ortiz
A Norberto Aristizabal y Natalia Aristizabal por creer en miacute
A mi hijo Joseacute David Aristizabal por representar las nuevas generaciones
A mi tutor Jhon Alexander Osorio por tenerme paciencia en este proceso de
aprendizaje y compartirme sus conocimientos
A Tatiana Tapias por darme su amor y llevarme por el camino de la virtud
siempre centrado
A la maestriacutea en Cambio Climaacutetico Sustentabilidad y Desarrollo por siempre
estar disponibles para miacute
A la Universidad Andina Simoacuten Boliacutevar por apoyar mi tesis de investigacioacuten
A la Repuacuteblica del Ecuador por abrirme sus puertas y darme un mejor futuro
8
9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

6
7
Con amor para mis padres Fernando Ivaacuten Aristizabal y mi madre Mariacutea Ligia
Ortiz
A Norberto Aristizabal y Natalia Aristizabal por creer en miacute
A mi hijo Joseacute David Aristizabal por representar las nuevas generaciones
A mi tutor Jhon Alexander Osorio por tenerme paciencia en este proceso de
aprendizaje y compartirme sus conocimientos
A Tatiana Tapias por darme su amor y llevarme por el camino de la virtud
siempre centrado
A la maestriacutea en Cambio Climaacutetico Sustentabilidad y Desarrollo por siempre
estar disponibles para miacute
A la Universidad Andina Simoacuten Boliacutevar por apoyar mi tesis de investigacioacuten
A la Repuacuteblica del Ecuador por abrirme sus puertas y darme un mejor futuro
8
9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

7
Con amor para mis padres Fernando Ivaacuten Aristizabal y mi madre Mariacutea Ligia
Ortiz
A Norberto Aristizabal y Natalia Aristizabal por creer en miacute
A mi hijo Joseacute David Aristizabal por representar las nuevas generaciones
A mi tutor Jhon Alexander Osorio por tenerme paciencia en este proceso de
aprendizaje y compartirme sus conocimientos
A Tatiana Tapias por darme su amor y llevarme por el camino de la virtud
siempre centrado
A la maestriacutea en Cambio Climaacutetico Sustentabilidad y Desarrollo por siempre
estar disponibles para miacute
A la Universidad Andina Simoacuten Boliacutevar por apoyar mi tesis de investigacioacuten
A la Repuacuteblica del Ecuador por abrirme sus puertas y darme un mejor futuro
8
9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

8
9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

9
Agradecimientos
Estos agradecimientos van dedicados para Ecuador que me brindoacute la oportunidad
poder estudiar una maestriacutea a mi tutor por aceptarme y acompantildearme en este proceso a
mi familia por creer en miacute y en los sacrificios que hice para irme a vivir a otro paiacutes solo y
sin dinero para poder umlsalir adelanteuml como decimos en Colombia a mi compantildeera
sentimental por siempre guiarme hacia el camino correcto disciplina compromiso y
entrega a la universidad Andina Simoacuten Boliacutevar por aceptarme en sus aulas de clase y
por ultimo a la Universidad del Quindiacuteo por abrirle la puerta a uno de sus egresados y
acogerlo de buena forma con sus instalaciones de laboratorio
Gracias y mil gracias a todos de corazoacuten sin duda esta experiencia cambio mi
vida en todos los sentidos
Desde hoy hasta el final que podriacutea llamarse infinito
10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

10
11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

11
Tabla de contenidos
Figuras mapas y tablashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 13
Introduccioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 15
Capiacutetulo primero Cambio Climaacutetico Frailejones y Hongos Endoacutefitoshelliphelliphelliphelliphelliphelliphelliphelliphellip 21
1 Antecedenteshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 21
2 Justificacioacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 25
3 Hipoacutetesishelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 26
4 Objetivoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
41 Objetivo generalhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
42 Objetivos especiacuteficoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 27
Capiacutetulo segundo Materiales Meacutetodos y Resultadoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
1 Aacuterea de Estudiohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
2 Colecta de material bioloacutegicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 29
3 Aislamientos fuacutengicos a partir de muestras vegetaleshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 30
4 Crecimiento de cultivos microbioloacutegicos (Hongos
endoacutefitos)helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 32
5 Extraccioacuten de ADNhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip33
6 Bioinformaacutetica-Metagenoacutemica Fuacutengicahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
7 Secuenciamientohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
8 Anaacutelisis Filogeneacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 35
Capiacutetulo Tercero Discusioacuten sobre los hongos endoacutefitos encontrados y sus implicaciones
ecoloacutegicashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
1 Cambio Climaacutetico y Biodiversidadhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 39
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio
Climaacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 42
Conclusioneshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 57
Obras citadashelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 59
Anexoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 73
12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

12
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombiahelliphelliphellip 73
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia hartwegiana
Paacuteramo Romerales Quindiacuteo Colombia 73
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombiahelliphelliphelliphellip74
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana 75
Anexo 5 Hifas observadas a traveacutes de un
estereoscopio 75
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 76
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas y
enviadas 76
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 75
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a Alemania a la Universidad Ruhr de
Bochum 77
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de
parsimoniahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 78
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X 78
Anexo 12 Estadiacutesticos generados por el programa
JmoldelTest 79
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE
(AIC) 81
13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

13
Figuras mapas y tablas
Figura 1 Tipos de vegetacioacuten en los paacuteramoshelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 16
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo
Colombiahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip18
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras
N necromasa R raiacuteces EP epiacutefita DE endoacutefitohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 23
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y
abioacuteticohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 24
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en
medio de cultivo Agar papa Dextrosa primera semana de crecimiento micelialhelliphelliphellip 30
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la
teacutecnica de corte de hifa uacutenica helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip 31
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de
cultivo Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12)
aislados de las hojas de la especie de frailejoacuten Espeletia hartwegiana 31-32
Figura 8 Pasos esenciales de un proyecto metagenoacutemicohelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de
frailejoacuten Espeletia hartwegiana helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip34
Figura 10 Filograma obtenido a partir del anaacutelisis ML y MP del conjunto de datos ITS Aislados
de las hojas de frailejoacuten37
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip43
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e
Hypocreahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip46
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon48
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon
Biscogniauxia Nemania y Humiacutecolahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip49
14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

14
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies
Massarina Cryptocoryneum Cyphellophora y los grupos externos
helliphelliphelliphelliphelliphelliphellip52
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona
muestreadahelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip16
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia 29-30
Tabla 2 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos
endoacutefitos en la especie Espeletia hartwegiana
helliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip36
Tabla 3 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = presencia del genero
de hongo dentro de la especie de frailejoacutenhelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphelliphellip42-43
15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

15
Introduccioacuten
Hay un libro abierto siempre para
todos los ojos la naturaleza
(Jean-Jacques Rousseau 1712-1778)
El cambio climaacutetico corresponde a un fenoacutemeno que afecta el planeta de manera natural
permitiendo mantener las condiciones climaacuteticas como la temperatura la precipitacioacuten entre
otras en un rango aceptable para la vida (Jimeacutenez 2010) Sin embargo dado el aumento de las
emisiones de gases de efecto invernadero (GEI) principalmente asociado a las actividades
humanas y el alto consumo de combustibles foacutesiles los cambios en dichas condiciones han sido
alterados dando paso a un acelerado aumento de temperatura anual en las uacuteltimas deacutecadas con
precipitaciones y sequias maacutes frecuentes e intensas (IPCC 2020 5 Jimeacutenez 2010)
Acorde con el Panel Intergubernamental de Expertos en Cambio Climaacutetico (IPCC) para
fin de siglo el Cambio Climaacutetico se acrecentaraacute pronosticando un aumento de temperatura de 25
a 45 degC para el antildeo 2100 que tan solo para Colombia en los uacuteltimos 30 antildeos fue de 08degC
generando impactos notorios en las zonas de alta montantildea como los paacuteramos (Moreno y Palma
2016)
El cambio climaacutetico es un fenoacutemeno cuyos efectos alcanzaraacuten los paacuteramos los cuales son
ecosistemas de alta montantildea presente en elevaciones entre 3100 y 4700 m ubicados en el norte de
los Andes de Ameacuterica del Sur y el sur de Ameacuterica Central adyacente (Berdugo et al 2016) Estas
aacutereas se caracterizan por una vegetacioacuten principalmente endeacutemica compuesta por frailejones
matorrales y pastos (veacutease figura 1) (Garciacutea et al 2010)
Los ecosistemas de alta montantildea como los paacuteramos ofrecen diversos servicios
ambientales como la biodiversidad uacutenica que albergan los paisajes los suelos y su capacidad de
retener y almacenar agua (Morales et al 2019) Las formaciones vegetales juegan un papel
diferenciado en el mantenimiento de la humedad y las fuentes de agua en el ecosistema
presentando la capacidad de interceptar almacenar y regular los flujos hiacutedricos superficiales y
16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -

16
subterraacuteneos aportando los servicios de abastecimiento de agua para el 70 de la poblacioacuten de
Colombia (Vaacutesquez y Buitrago 2017)
Ademaacutes dada la caracteriacutestica de permanente humedad de su suelo la materia orgaacutenica
se descompone a menor velocidad logrando un efecto de retencioacuten y almacenamiento de carbono
(Anderson et al 2012) En Colombia se han realizado diferentes estudios sobre la distribucioacuten y
extensioacuten del ecosistema paramuno Rangel (2002 4) determinoacute que los paacuteramos colombianos
abarcaban aproximadamente el 26 de la superficie del paiacutes
Figura 1 Tipos de vegetacioacuten en los paacuteramos por Pedraza-Pentildealosa Betancur y Franco-Rosselli (2005)
En particular el paacuteramo Romerales se encuentra dentro del territorio adscrito a los
Parques Nacionales Naturales (PNN) especiacuteficamente el PNN de los Nevados en la cordillera
central colombiana hacia la zona norte del Departamento del Quindiacuteo con liacutemites fronterizos
entre los departamentos de Risaralda y Tolima Este paacuteramo es el hogar del frailejoacuten Espeletia
hartwegiana el cual estaacute afrontando los efectos del cambio climaacutetico (veacutease mapa 1)
Mapa 1 Parque Nacional Natural los Nevados ubicacioacuten paacuteramo Romerales zona muestreada
17
Existe evidencia cientiacutefica concreta del aumento de temperatura en los Andes Tropicales
tambieacuten cambios significativos en los patrones de precipitacioacuten sumado a la disminucioacuten en la
incidencia del clima nublado lo que conduce a las plantas a mayor cantidad de horas expuestas a
la radiacioacuten solar (Ruiz et al 2009) Otro efecto grave que trae consigo el cambio climaacutetico es el
desplazamiento de especies en el gradiente altitudinal Con un aumento de 3degC se espera que las
especies se muevan ~600 m en buacutesqueda de su optimo teacutermico esto afectariacutea gravemente las
especies de paacuteramo como los frailejones Cuesta Camacho (2007)
Los frailejones son emblemaacuteticas plantas de paacuteramo y ecosistemas de alta montantildea
(MINAMBIENTE 2019) Pertenecen a la familia Asteraceae contiene cerca de 142 especies
agrupadas en ocho geacuteneros Carramboa Coespeletia Espeletia Espeletiopsis Libanothamnus
Paramiflos Ruilopezia y Tamania de estos los maacutes importantes y con mayor diversidad de
especies son Espeletia Espeletiopsis y Libanothamnus (Rivera y Torres 2001)
Poseen adaptaciones que les permite captar y retener grandes cantidades de agua de su
entorno permitiendo un balance ecosistemico (Farias 2013) La morfologiacutea singular del genero
Espeletia le permite acumular las hojas antiguas en forma de ldquoabrigordquo brindando proteccioacuten
contra las bajas temperaturas nocturnas de hasta 0degC Los fuertes vientos y la radiacioacuten solar los
cuales seraacuten cada vez maacutes intensos y frecuentes por causa del cambio climaacutetico (Cortes y
Sarmiento 2013 34) (veacutease figura 2)
Estas plantas son especies claves para la conservacioacuten de los paacuteramos y la lucha contra el
cambio climaacutetico debido a que ofrecen una gran variedad de servicios ecosisteacutemicos (Diacuteaz et al
2007) El paacuteramo es referente sin igual de fuente hiacutedrica ciudades como Bogotaacute y Quito obtienen
aproximadamente el 80 de su agua dulce de los paacuteramos (Buytaert et al 2006) Los frailejones
en su mayoriacutea endeacutemicos se han visto afectados por actividades humanas como la ganaderiacutea
extensiva agricultura (cultivos de papa y cebolla) mineriacutea de oro y carboacuten turismo no
controlado y quemas intencionales (Hofstede 2000) Esta situacioacuten podriacutea comprometer a
mediano y largo plazo la funcioacuten de captacioacuten regulacioacuten y suministro del agua para el consumo
18
en las ciudades colombianas ubicadas en las regiones de influencia de estos ecosistemas (Varela
2015)
Figura 2 Fotografiacutea de la especie Espeletia hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
No obstante en asociacioacuten con especies como el frailejoacuten de los Andes Tropicales
encontramos a los hongos cuyos roles en las interacciones bioloacutegicas son de vital importancia
Los hongos pueden ser endoacutefitos simbiontes mutualistas paraacutesitos y comensales y son capaces
de responder a las amenazas que plantea el cambio climaacutetico en cuanto a perturbaciones
ambientales aumento de temperatura y estreacutes hiacutedrico adaptaacutendose a la disponibilidad de recursos
modificando sus actividades enzimaacuteticas e incluso su genoma a lo largo del tiempo (Moreno et
al 2013)
Los hongos son clasificados seguacuten su forma de reproduccioacuten sea sexual o asexual en
donde se observan las diferencias estructurales internas entre los diferentes grupos tales como
sus esporas e hifas como lo indican Blackwell y Spatafora (2004) estos organismos estaacuten
divididos en cuatro grandes grupos los cuales son Ascomycota Basidiomycota
Chytridiomycota Zygomycota es justo en el grupo de los Ascomycota donde se encuentra gran
diversidad de hongos conocidos como ldquohongos endoacutefitosrdquo en donde su nombre ldquoendoacutefitordquo
significa ldquodentro de la plantardquo (endon dentro phyton planta)
19
Estos microorganismos fuacutengicos tienen la facilidad de pasar gran parte o todo su ciclo de
vida invadiendo los tejidos de su hospedero (planta) sin tener un efecto negativo evidente
(Leacuteveilleacute 1846 3 de Bary 1866 8) Por lo tanto el concepto de endoacutefito indica una asociacioacuten de
costo-beneficio pero de una forma mutualista entre el hongo y la planta hospedera permitiendo
la estadiacutea en sus tejidos vivos (Clay 1998 8) Los hongos endoacutefitos se han encontrado en todas
las plantas (algas musgos pastos plantas y vasculares) desde los bosques templados hasta los
bosques tropicales hallaacutendose una gran diversidad de plantas y en asociacioacuten gran diversidad de
hongos endoacutefitos (Saacutenchez et al 2010)
Se espera que los hongos endoacutefitos jueguen un papel cada vez maacutes importante potenciado
por el cambio climaacutetico en las enfermedades producidas en plantas como pudricioacuten radicular
mal blanco entre otras con los aumentos pronosticados de la temperatura y el estreacutes por sequiacutea
asociados al cambio climaacutetico (Desprez-Loustau et al 2006 Slippers y Wingfield 2007 6) Los
cambios en la cantidad de dioacutexido de carbono (CO2) y la disponibilidad de nitroacutegeno en el suelo
afectan de forma similar tanto a las plantas como a los hongos por ende las alteraciones
generadas por parte del cambio climaacutetico pueden causar desbalances en sus relaciones (Ies et al
2013)
Algunos estudios realizados en el antildeo 1898 donde se registraron los primeros hongos
endoacutefitos como el de Bary (1866) brindo la iniciativa de investigaciones que lograron explicar
los cuadros toxicoloacutegicos observados en animales los cuales se daban por el consumo de
semillas o en aacutereas de pastos (Poaceae) que posiblemente albergaban hongos Clavicipitaacuteceos
(familia del ergot) los cuales poseiacutean actividades enzimaacuteticas con micotoxinas
Por diversas situaciones no trascendioacute la investigacioacuten hasta que Bacon (1977) hizo
relacioacuten del hongo endoacutefito Neotyphodium coenophialum debido a una intoxicacioacuten del ganado
llamado ldquosiacutendrome de veranordquo cuando estos animales consumiacutean pastos de la especie Festuca
arundinacea (Poaceae) Funk en el antildeo 80 caracterizoacute la resistencia de las Poaceae que
albergaban hongos endoacutefitos que frecuentemente eran atacados por insectos (proteccioacuten contra
herbiacutevoros) (Saacutenchez et al 2010)
Compant y colaboradores (2010) mencionaron que tras experimentar en el laboratorio
con una gama de temperaturas en los hongos endoacutefitos aislados obtuvieron resultados variables
respecto al crecimiento es decir positivos y neutrales llegando a la conclusioacuten de que los
microorganismos asociados a las plantas son un factor importante que influye en la respuesta de
20
las plantas acorde a los cambios de temperatura lo que compromete las respuestas del frailejoacuten y
los hongos endoacutefitos frente al calentamiento global causado por el cambio climaacutetico
Con relacioacuten a los estudios realizados sobre hongos endoacutefitos en frailejones se han
encontrado al menos 100 especies con capacidades de inhibir hongos fitopatoacutegenos o herbivoriacutea
(Cabezas et al 2012)
Los geacuteneros de hongos endoacutefitos encontrados en anteriores estudios son Acremonium
Aspergillus Aureobasidium Colletotrichum Fusarium Gliocadium Nigrospora Paecilomyces
Penicillium Trichothecium Trichoderma y Sordaria (Pineda 2000 Programa Nacional para la
Evaluacioacuten del Estado y Afectacioacuten de los Frailejones en los Paacuteramos de Andes del Norte 2009)
Teniendo en cuenta que para Colombia hay poca informacioacuten sobre hongos endoacutefitos en
ecosistemas paramunos se hace fundamental la ampliacioacuten de maacutes informacioacuten referente al tema
debido a que solo existen estudios significativos en cafeacute leguminosas y rosas los cuales maacutes allaacute
de la identificacioacuten de geacuteneros de hongos endoacutefitos no incluyen el cambio climaacutetico y sus
posibles consecuencias (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016) En
teacuterminos filogeneacuteticos los hongos son el reino de la naturaleza maacutes cercano a los animales
(Hibbett et al 2018)
Las relaciones que posean las diferentes especies de endoacutefitos dentro de la especie de
frailejoacuten es de importancia conocerla mediante el uso de la filogenia molecular (Judd et al 2007)
mediante secuencias de ADN ARN o proteiacutenas (Cooke et al 2000) Dependiendo de la presencia
de especies agresivas de endoacutefitos como Nigrospora se puede tener un mejor conocimiento
sobre las consecuencias que trae consigo los impactos del cambio climaacutetico sobre estas plantas de
los Andes Tropicales (Foden et al 2009)
21
Capiacutetulo primero
Cambio Climaacutetico Frailejones y Hongos Endoacutefitos
1 Antecedentes
Desde los estudios realizados por Leacuteveilleacute (1846) y Bary (1866) sobre hongos Endoacutefitos
en pastos de zonas templadas se ha logrado identificar que estos hongos son capaces de inducir
respuestas adaptativas a sus hospederos (Micotoxinas contra herbiacutevora adaptacioacuten a heladas o
climas extremos fijacioacuten de nutrientes) a cambio de alimento y hospedaje formando una relacioacuten
mutualista en la cual las dos especies involucradas son beneficiadas No obstante se ha reportado
que los hongos endoacutefitos ademaacutes de tener relaciones mutualistas con diferentes especies
vegetales se pueden tornar en especies patoacutegenas para su hospedero una vez la planta se
encuentra en condiciones desventajosas tornaacutendose asiacute el hongo endoacutefito en un hongo
fitopatoacutegeno (Herrera y Ulloa 1990 550)
Como se mencionoacute anteriormente los hongos endoacutefitos pueden pasar de un estado
mutualista a un estado de patogeacutenesis dada esta relacioacuten entre ambos organismos se producen
metabolitos secundarios potencialmente toacutexicos facilitando factores de virulencia dados por el
hongo endoacutefito como exoenzimas y metabolitos fitotoacutexicos a su vez la planta presenta defensas
tanto mecaacutenicas como bioquiacutemicas (Saacutenchez et al 2013 5)
Hasta la fecha los principales estudios de hongos endoacutefitos se han fijado en especies de
intereacutes econoacutemico y comercial tales como palma de aceite cafeacute leguminosas pastos de forrajeoacute
y diferentes tipos de rosas (Salgado y Cepero 2005 Vega et al 2010 Parsa et al 2016)
Las especies modelo para el estudio de los hongos endoacutefitos han sido las gramiacuteneas
forrajeras en donde se han reportado especies de endoacutefitos asociados a los geacuteneros Lolium
Festuca y Brachiaria (Siegel et al 1987) no obstante Otero y Bayman (2009) han reportado un
creciente nuacutemero de estudios en orquiacutedeas epifitas tropicales relaciones con hongos micorrizicos
22
Actualmente existen escasos reportes de hongos endoacutefitos asociados a ecosistemas con
climas extremos (temperaturas extremas) como aacuterticos desiertos paacuteramos y zonas alpinas
especialmente para zonas de temperaturas friacuteas como lo son los paacuteramos (Lopera et al 2008
Leung et al 2011 Cabezas et al 2012 Miles et al 2012 Zhang y Yao 2015)
Investigadores como Lopera Leung y colaboradores (2011) han llegado a la conclusioacuten
de que los hongos endoacutefitos juegan un papel importante en las plantas a la hora de adaptarse a
ecosistemas de clima templado (Kivlin et al 2013 Zhang y Yao 2015)
En Colombia los trabajos con hongos endoacutefitos son pocos por ende trabajos como el de
Lopera y colaboradores (2008) con especies del genero Espeletia de los paacuteramos Colombianos
son pioneros ademaacutes del trabajo de Cabezas y colaboradores (2012 4) en el cual aislaron y
caracterizaron celulasas (enzimas descomponedores de la celulosa) extraiacutedas de hongos endoacutefitos
que se hallaban en especies de Espeletia posteriormente Lopera y Miles (2012) unieron
esfuerzos para continuar caracterizando la diversidad de hongos endoacutefitos en los paacuteramos andinos
colombianos enfocados en Espeletia con el objetivo de encontrar especies con potencial
biocontrolador
Existen trabajos tesis en los repositorios de diversas universidades de los antildeos 2014 y
2015 en los cuales se aborda el tema de los hongos endoacutefitos paacuteramos y especies de Espeletia
(frailejones) y de la familia Poaceae en donde se caracterizan las diversas especies de hongos
endoacutefitos por medio taxonomiacutea y biologiacutea molecular ademaacutes de buscar metabolitos secundarios
con fines medicinales o de biocontrol
Los trabajos en filogenia molecular de hongos endoacutefitos realizados en frailejones se han
visto enfocados maacutes hacia la buacutesqueda de la diversidad funcional en comunidades
microbioloacutegicas como por ejemplo el estudio realizado por Ruiz y colaboradores (2015) en
donde buscaron mediante el anaacutelisis molecular de raiacuteces necromasa hojas maduras y joacutevenes de
diferentes especies de Espeletia (veacutease figura 3) ubicada en los Andes neotroacutepicales
colombianos secuencias geneacuteticas de las regiones V5-V6 de la unidad 16S del rARN
23
Figura 3 Distribucioacuten muestral por individuo recolectado Y hojas joacutevenes M hojas maduras N
necromasa R raiacuteces EP epiacutefita DE endoacutefito Tomado de Ruiz y colaboradores (2015)
Este estudio analizoacute la composicioacuten de comunidades microbioanas y su diversidad
funcional en un ecosistema de alta montantildea en donde las temperaturas los rayos UV y la altura
juegan papeles determinantes como filtros ecoloacutegicos severos en donde solo pueden existir
especies que han desarrollado capacidades para adaptarse sobrevivir y colonizar dichas aacutereas
por ende cuando se analizaron las comunidades existentes dentro de estas especies de Espeletia
Encontraron que algunas especies halladas podriacutean tener roles importantes como
productores de metabolitos secundarios enzimas o incluso como bioremediadores y que las
comunidades microbianas eran diferentes tanto en sus hojas como en sus raiacuteces y en la
necromasa
A medida que las deacutecadas avanzan y la evidencia sobre el cambio climaacutetico se hace maacutes
robusta y evidente los estudios en los hongos endoacutefitos han tomado vital importancia al estar
asociados con cultivos de especies comestibles escases de agua y de tierras productivas sumado
a los efectos negativos que trae consigo el cambio climaacutetico para las diferentes plantaciones y
cosechas a nivel global
Es por eso que Lu Yan y colaboradores (2019) analizaron los efectos positivos que teniacutean
la colonizacioacuten de hongos endoacutefitos en las plantas en donde ratificaron que los hongos endoacutefitos
dentro la planta generan metabolitos antimicrobianos estimulan las respuestas inmunoloacutegicas y
compiten por espacio dentro de la planta con diferentes patoacutegenos incrementando asiacute la
resistencia contra herbiacutevoros y la resistencia al estreacutes salino (factores bioacuteticos)
Ademaacutes de ayudar a la planta contra el estreacutes abioacutetico causado principalmente por
factores climaacuteticos como la falta de agua (estreacutes hiacutedrico) las temperaturas extremas la
intensidad de los rayos UV y los metales pesados como la muestra la figura 4 debido a esto se
24
concluyoacute que las fitohormonas cumplen un papel importante en la colonizacioacuten de hongos
endoacutefitos dentro de la planta
Aun que se ha reportado que los hongos endoacutefitos se pueden usar como biocontroladores
diferentes estudios reportan que el uso de hongos endoacutefitos puede tener efectos adversos como
un retraso significativo en el crecimiento de la planta con el uso de la especie Cryptosporiopsis
sp (Terhonen et al 2016) Otro reporte indica la fecundidad y supervivencia de enemigos
naturales como paraacutesitos esto es debido a que los hongos endoacutefitos transfieren sus toxinas y
alcaloides cuando son ingeridos por paraacutesitos u otros enemigos naturales de las plantas (Lopez y
Sword 2015)
Figura 4 La colonizacioacuten endoacutefita modula las defensas de las plantas contra el estreacutes bioacutetico y abioacutetico
Bajo estreacutes bioacutetico (herbiacutevoros y patoacutegenos) y abioacutetico (sequiacutea salinidad metales pesados y otros) (a) se
inducen respuestas de defensa de las plantas (b) los endoacutefitos producen metabolitos protectores y
modulan la viacutea de las fitohormonas para mejorar la inmunidad de las plantas (c) tomada de Lu yan y
colaboradores (2019 7)
25
2 Justificacioacuten
La presente investigacioacuten pretende identificar y caracterizar que especies de hongos
endoacutefitos estaacuten asociadas al frailejoacuten Espeletia hartwegiana y que posibles implicaciones podriacutea
tener que estas especies se encuentren dentro de esta especie de alta montantildea La importancia de
conocer esta informacioacuten reside en que los hongos endoacutefitos pueden beneficiar o perjudicar a la
adaptacioacuten y resistencia de especies vegetales en los distintos escenarios climaacuteticos propuestos
por la ciencia (IPCC 2020)
El calentamiento global podriacutea afectar el equilibrio de los hongos endoacutefitos en especies
vegetales generando desbalances en la planta o la peacuterdida de especies endoacutefitas de importancia
biotecnoloacutegica Por ejemplo se podriacutean perder especies endoacutefitas con potencial para control
bioloacutegico de plagas y enfermedades de plantas como productores de enzimas y metabolitos
secundarios degradacioacuten de plaacutesticos o biorremediacioacuten entre otros (Schulz et al 2002) Por
ello se hace fundamental estudiar los hongos endoacutefitos en ecosistemas de paacuteramo ya que en
Colombia y en los paacuteramos en general han sido poco estudiados y su efecto sobre las plantas de
alta montantildea podriacutea ser clave en el mantenimiento de sus poblaciones
26
3 Hipoacutetesis
H1 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute diferente en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo diferentes los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten tambieacuten seria
modificada
H0 La identificacioacuten y caracterizacioacuten molecular de los hongos endoacutefitos encontrados en
la especie de frailejoacuten Espeletia hartwegiana ubicada en el paacuteramo romerales Quindiacuteo
Colombia seraacute similar en geacuteneros y especies de otras identificaciones de hongos endoacutefitos
realizadas en especies de frailejoacuten siendo similares los geacuteneros y especies encontrados en
Espeletia hartwegiana las consecuencias del cambio climaacutetico sobre el frailejoacuten continuariacutean
como se ha registrado con anterioridad
27
4 Objetivos
41 Objetivo general
Caracterizar la diversidad de hongos endoacutefitos presentes en los frailejones
de la especie Espeletia hartwegiana en el paacuteramo de Romerales tomando en cuenta su
relacioacuten con el cambio climaacutetico
42 Objetivos especiacuteficos
Identificar los hongos endoacutefitos asociados en Espeletia hartwegiana del
paacuteramo de Romerales
Hallar relaciones filogeneacuteticas entre las diferentes especies de hongos
Endoacutefitos encontradas en Espeletia hartwegiana
Advertir las repercusiones del cambio climaacutetico sobre Espeletia
hartwegiana en base a su diversidad de hongos endoacutefitos
28
29
Capiacutetulo segundo
Materiales Meacutetodos y Resultados
1 Aacuterea de Estudio
El trabajo se realizoacute en el paacuteramo de Romerales ubicado en la zona alta del municipio de
Salento Quindiacuteo (4deg39202N 75deg25506W) con una extensioacuten aproximada de 6794 ha (Cano
et al 2015)
2 Colecta de material bioloacutegico
La colecta del material bioloacutegico se realizoacute mediante muestreos aleatorios lo largo del
paacuteramo tomando coordenadas de cada punto de colecta (veacutease anexo 3) las hojas se colectaron
por medio de tijeras podadoras siendo empacadas cada una en bolsas Ziploc diferentes se
colectoacute un total de 15 hojas cada una correspondiente a una planta de frailejoacuten para un total de
15 plantas muestreadas de E hartwegiana
Adicionalmente las bolsas Ziploc fueron marcadas con los datos de colecta en campo y
transportadas a los laboratorios del Programa de Biologiacutea de la Universidad del Quindiacuteo para su
procesamiento
Tabla 1 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten Espeletia
hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
3 Aislamientos fuacutengicos a partir de muestras vegetales
Las muestras obtenidas fueron lavadas con agua para eliminar el exceso de residuos se
cortaron 4 trozos de cada hoja con un tamantildeo aproximado de 05 a 10 cm (veacutease figura 5) luego
cada trozo de hoja se sometioacute a una esterilizacioacuten la cual se completoacute con algunas modificaciones
acorde al procedimiento descrito por (Osorio et al 2017) especiacutefico para el aislamiento de
30
hongos endoacutefitos que consiste en sumergir perioacutedicamente el material vegetal en etanol al 90
durante un minuto seguido de hipoclorito de sodio (NaClO) al 4 por un minuto continuando
con etanol al 70 durante un minuto y por uacuteltimo se lavaraacuten con agua destilada todo dentro de la
campana de flujo
Las cuatro secciones de hoja se transfirieron a una caja de Petri para un total de 15 cajas
de Petri con Agar Papa Dextrosa (PDA) (una caja de Petri por cada 4 cortes de hoja)
Posteriormente las muestras fueron incubadas a temperatura ambiente (25degC aproximadamente)
Figura 5 Cortes de las hojas de frailejoacuten Espeletia hartwegiana esterilizadas y sembradas en medio de
cultivo Agar papa Dextrosa primera semana de crecimiento micelial caja de Petri de 20ml de capacidad
Pasadas 3 semanas se revisaron los cultivos fuacutengicos con el fin de revisar el crecimiento
del micelio y hacer un repique a un cultivo puro una vez maduros se procedioacute a utilizar la teacutecnica
de corte de hifa uacutenica (Osorio et al 2017) esta teacutecnica consistioacute en tomar la punta de la hifa con
ayuda de un estereoscopio y un bisturiacute de grado quiruacutergico agua destilada alcohol al 70 y un
mechero para mantener el bisturiacute esterilizado una vez identificada la hifa ideal para cultivar de
forma pura (veacutease figura 6) con el bisturiacute se procedioacute a cortar el agar PDA en forma de cuadro
con una medida aproximada de 3 miliacutemetros cuadrados
31
Figura 6 Hifa uacutenica observada desde el estereoscopio lista para ser cortada por medio de la teacutecnica de
corte de hifa uacutenica
Se esperoacute entre 3 a 5 semanas a que crecieran los cultivos puros aislados de las hifas uacutenicas en
nuevas cajas de Petri con agar PDA
4 Crecimiento de cultivos microbioloacutegicos (Hongos endoacutefitos)
Los hongos cultivados de forma pura consiguieron un crecimiento micelial significativo a partir
de los 15 diacuteas a temperatura ambiente como lo demuestra la figura 7 que contiene doce de los
cultivos maacutes representativos con su respectivos micelios aeacutereos desarrollados donde variacutean sus
colores formas y consistencias esto indicando que pertenecen a diferentes especies
2
1
3
32
Figura 7 Ejemplos de las diferencias morfoloacutegicas en micelios generados por los medios de cultivo
Axeacutenicos (Puros) de hongos endoacutefitos en agar PDA (1 2 3 4 5 6 7 8 9 10 11 12) aislados de las
hojas de la especie de frailejoacuten Espeletia hartwegiana
5 Extraccioacuten de ADN
4
5
6
7
8
9
10
11
12
33
Una vez el micelio de los hongos se encontraba desarrollado en la cantidad necesaria se
procedioacute a extraer el ADN genoacutemico siguiendo el protocolo de extraccioacuten de Raeder y Broda
(1985) por medio de un barrido superficial del micelio evitando tomar el agar este micelio
posteriormente se colocoacute en tubos Eppendorf esteacuteriles de 2 ml y se pulverizoacute mediante el uso de
un asa microbioloacutegica y 1ml de buffer de extraccioacuten compuesto de Tris-HCL 1 M pH 78 EDTA
05 M Ph 8 SDS 10 wv NaCL y agua destilada para cada tubo a continuacioacuten se agregoacute 800
ul de etanol al 70 500 ul de fenol y se mezcloacute
Se adicionoacute 300 ul de cloroformo y se mezcloacute de nuevo de alliacute se llevoacute a la centrifuga a
10000 rpm por 60 minutos luego de centrifugar se forma un sobrenadante el cual es transferido
en su totalidad a un nuevo tuvo Eppendorf de 15 ml en el cual se agregaron 400 ul de cloroformo
y se llevoacute a la centrifuga a 10000 rpm por 10 minutos en donde nuevamente se forma un
sobrenadante aunque esta vez solo se toman 500 ul del tubo para ser transferidos otro tubo
Eppendorf de 15 ml ya en el nuevo tubo se adicionaron 50 ul de NaAC y se invirtioacute varias veces
para homogenizar la mezcla a continuacioacuten se guardoacute en el refrigerador a 4degC toda la noche
Al diacutea siguiente se tomaron las muestras en los tubos Eppendorf y se centrifugaron a
10000 rpm por 30 minutos (Formacioacuten del Pellet de ADN) cuando la centrifuga terminoacute se
procedioacute a remover el etanol ya sin etanol se agregoacute de nuevo 1 ml de etanol al 70 a cada tubo
y se centrifugoacute a 10000 rpm por 5 minutos en este paso se removioacute el etanol y se dejoacute secar el
pellet de ADN una vez el etanol se evaporoacute se re suspendioacute el pellet en 100 ul de agua grado
molecular o Sabax con 5 ul de RNAsa con el fin de evitar la presencia de moleacuteculas de ARN
posteriormente se incubaron a 37degC por 30 minutos y se guardaron para su envioacute a -20degC
(Raeder y Broda 1985)
6 Bioinformaacutetica-Metagenoacutemica Fuacutengica
Una vez se secuenciaron las muestras de ADN enviadas a la Universidad Ruhr de
Bochum Alemania se hizo el ensamblaje de las secuencias en contigs y scaffolds en el
programa CLC Main Workbench 8 (Nicola et al 2013) Como lo muestra la figura 8 donde se
muestran los pasos esenciales de una teacutecnica de Metagenoacutemica donde posteriormente se
analizaron con maacutes detalle mediante teacutecnicas filogeneacuteticas
34
Figura 8 Pasos esenciales de un proyecto metagenoacutemico Fuente Thomas y colaboradores (2012)
7 Secuenciamiento
Se extrajoacute con eacutexito un total de 35 muestras de hongos endoacutefitos encontrados en las hojas de
Espeletia hartwegiana (veacutease figura 9) que posteriormente fueron secuenciadas por la
Universidad Ruhr de Bochum (Alemania) logrando un total de 70 secuencias incluyendo
aquellas obtenidas en Genk Bank
Figura 9 ADN extraiacutedo de cultivos puros de hongos endoacutefitos aislados de hojas de la especie de frailejoacuten
Espeletia hartwegiana
35
8 Anaacutelisis Filogeneacutetico
Se aislaron 35 secuencias basadas en la especie Espeletia hartwegiana fueron
comparadas posteriormente con 35 secuencias de especies publicados en GenBank
(httpblastncbinihgovblastcgi) se obtuvo un total de 70 secuencias Las secuencias de
consenso ITS 58S DNA obtenidas se usaron para realizar buacutesquedas BLAST en GenBank
usando BLASTn (Altschul et al 1990) con el fin de identificar mediante las secuencias aisladas
los geacuteneros o las especies maacutes cercanas La matriz de datos fue alineada en MAFFT v7 (Katoh y
Standley 2013) Se utilizaron las especies Coprinellus xanthothrix y Coprinellus domesticus
como grupos externos
Los anaacutelisis de las secuencias para construir los arboles filogeneacuteticos de Maacutexima
Parsimonia (MP) fueron realizados usando PAUP v40b10 (Swofford 2003) Para el anaacutelisis de
Maacutexima Verosimilitud o ML por sus siglas en ingleacutes (Maximum Likelihood) se utilizoacute
JmodelTest (Darriba et al 2012) con el cual se infirieron los criterios del modelo de sustitucioacuten
apropiado utilizando los criterios de informacioacuten de Akaike (AIC) (Akaike 1974)
Los anaacutelisis de ML se realizaron con el programa PhyML v 30 (Guindon y Gascuel
2003) teniendo en cuenta la proporcioacuten de sitios invariables Los valores de soporte de confianza
para los nodos se estimaron mediante 1000 anaacutelisis de arranque de replicacioacuten La alineacioacuten de
los datos de la secuencias de ITS para 70 aislamientos determinoacute quince geacuteneros siendo los
siguientes Annulohypoxylon Biscogniauxia Colletotrichum Crassiparies Cryptocoryneum
Cyphellophora Diaporthe Daldinia Humiacutecola Hypoxylon Hypocrea Nemania Nigrospora
Massarina y Trichoderma de los cuales se obtuvieron 11 clados principales (veacutease figura 10)
Con base en estos resultados todos los aislamientos se analizaron maacutes a fondo dentro de los
geacuteneros hasta llegar a especie (veacutease tabla 1)
36
Geacuteneros Especie Especie Especie Especie Especie Especie Especie
Annulohypoxylon A nitens A bahnphaden
gense
Biscogniauxia B petrensis
Colletotrichum C gloeosporioides
C gigasporum
Cyphellophora C
reptans
Crassiparies C quadrisporus
Cryptocoryneum C
longicondensatum
Daldinia D dennisii D
eschscholtzi
D
conceacutentri
ca
Diaporthe D
longicolla
D
foeniculina
D
anacardi
i
D
macada
miae
Humiacutecola H
fuscoatra
Hypoxylon H
trugodes
H
pulicicidum
Hypocrea H
koningii
Massarina M albocarnis
Nemania N
primolutea
Nigrospora N sacchari N rubi N sphaeric
a
N hainanen
sis
N oryzae
N gorlenko
ana
N aurantia
ca
Trichoderma T trixiae T
olivascens
T
paraviridescens
T
viridescens
T
samuelsii
T viride
Grupo Externo Coprinellus C
domesticus
C
xanthothrix
Tabla 1 Geacuteneros y especies encontrados en los aislamientos e identificaciones de hongos endoacutefitos en la
especie Espeletia hartwegiana
37
38
Figura 10 Filograma consenso obtenido con el programa MEGA ndashX a partir del anaacutelisis ML y MP del
conjunto de datos ITS Aislados de las hojas de frailejoacuten Los coacutedigos en Verde son las secuencias que se
aislaron (aislados) en este estudio los coacutedigos y nombres en Negro son las secuencias encontradas por
identidad en GenBank Los valores de soporte de bootstrapgt 70 se indican cerca de los nodos como MP
ML = valores de soporte de bootstrap lt70
El nuacutemero de caracteres los modelos de sustitucioacuten usada asiacute como otros resultados
estadiacutesticos de las pruebas de Maacutexima Parsimonia y Maacutexima Verosimilitud generados a partir de
los anaacutelisis filogeneacuteticos de los conjuntos de datos se muestran en los anexos
39
Capiacutetulo Tercero
Discusioacuten sobre los hongos endofitos encontrados y sus implicaciones
ecoloacutegicas
1 Cambio Climaacutetico y Biodiversidad de Endoacutefitos
Tanto la temperatura la estacionalidad y los tipos de precipitacioacuten son factores de vital
importancia para la composicioacuten de las diferentes comunidades y la arquitectura vegetal Estos
factores limitan muchos de los procesos ecosisteacutemicos ya que existe una estrecha relacioacuten entre
las variaciones climaacuteticas y las variaciones dentro de los ecosistemas Por lo tanto se espera que
el cambio climaacutetico transforme los ecosistemas Andinos tal como los conocemos hoy (Colwell et
al 2008) modificando las relaciones mutualistas entre plantas y hospederos
Posiblemente la resiliencia y resistencia de muchos ecosistemas como el paacuteramo sean
superadas en este siglo por una combinacioacuten sin precedentes de disturbios tales como el cambio
climaacutetico perturbaciones asociadas a este (sequiacuteas fuegos invasioacuten de especies exoacuteticas
inundaciones) y otros factores como organismos oportunistas (hongos endoacutefitos) cambios en el
uso del suelo contaminacioacuten y sobreexplotacioacuten de recursos (IPCC 2020)
Considerando que el frailejoacuten Espeletia hartwegiana es una especie de alta montantildea
(paacuteramos) endeacutemica de la cordillera Central de los Andes (Schultz y Carl 1856) se encuentra
dentro de los ecosistemas andinos en donde a su vez se ha comprobado la existencia de los
hongos endoacutefitos es de tener en cuenta el papel de estos organismos endoacutefitos ya que los
paacuteramos son ecosistemas particularmente sensibles a los efectos adversos del cambio climaacutetico
en especial al calentamiento global al estar ubicados en los troacutepicos generando asiacute ventanas de
40
oportunidades para que los hongos endoacutefitos alteren su estado mutualista y pasen a ser
patogeacutenicos (Colwell et al 2008 7)
No obstante existe consenso en que las temperaturas aumentaraacuten especiacuteficamente a
grandes alturas y que la precipitacioacuten posiblemente se incrementaraacute en algunas zonas disminuiraacute
en otras y se produciraacute en patrones estacionales alterados con mayor intensidad y frecuencia
(Marengo et al 2009)
Los frailejones se ven en constante estreacutes por el uso del suelo dado en los paacuteramos como
es el caso del paacuteramo Romerales un paacuteramo que se ha visto afectado por las quemas y la
ganaderiacutea (Molinillo y Monasterio 2001) Esto aumenta asiacute las condiciones de estreacutes intriacutensecas a
las que estaacuten expuestas estas plantas de paacuteramo las cuales suministran valiosos bienes y servicios
ecosisteacutemicos como el abastecimiento continuo de agua para usos domeacutesticos agriacutecolas e
industriales (Buytaert et al 2006 4) servicios que se podriacutean ver en decrecimiento si las
ventanas de oportunidades se abren con maacutes frecuencia dando pie al cambio de estado de los
hongos endoacutefitos que existen dentro de los frailejones de la zona
En los paacuteramos de Chingaza Rabanal Guerrero Iguaque Merchaacuten y Sumapaz se han
realizado estudios de aislamientos de hongos en especies de frailejoacuten diferentes a la especie de
frailejoacuten Espeletia hartwegiana tales como Espeletia argentea Espeletia arbelaezii Espeletia
boyacensis Espeletia grandiflora Espeletia miradorensis y Espeletia uribei por ende se ha
reportado la presencia de hongos endoacutefitos y fitopatoacutegenos (Tabla 2) (Gaitan 2018 17)
Frailejones Espeletia
argentea
Espeletia
arbelaezii
Espeletia
boyacensis
Espeletia
grandiflora
Espeletia
miradorensis
Espeletia
uribei
Aspergillus X X X X
Annulohypoxylon X X X
Curvularia X X X
Epicoccum X X X
Fusarium X X X
41
Tabla 2 Geacuteneros de hongos encontrados en otras especies de frailejoacuten X = a presencia del genero de
hongo dentro de la especie de frailejoacuten
Los geacuteneros de hongos en rojo son los geacuteneros que se encontraron en comuacuten con los
geacuteneros hallados en la especie de frailejoacuten Espeletia hartwegiana de un total de 15 geacuteneros 3
han sido reportados anteriormente en estudios similares ( 20 de geacuteneros identificados)
Dentro de un marco de cambio climaacutetico es importante la metodologiacutea de muestreo
como lo fue el formato de muestreo en puntos aleatorios realizado en el presente estudio se llevoacute
acabo principalmente por la homogeneidad de los frailejones en el paacuteramo como lo indica el
manual de meacutetodos baacutesicos de muestreo y anaacutelisis de ecologiacutea vegetal (BOLFOR 2000 9) donde
se menciona que el muestreo aleatorio se utiliza cuando se dispone de poca informacioacuten previa de
las caracteriacutesticas de las poblaciones a medirse en este caso la poblacioacuten a identificarse fueron
los hongos endoacutefitos que se hallaban dentro de las formaciones homogeacuteneas de los frailejones
No obstante el muestreo aleatorio tambieacuten presenta limitaciones a la hora de comparar los
hongos endoacutefitos a lo largo del gradiente altitudinal (muestreo aleatorio estratificado) dejando de
percibir informacioacuten proveniente de los distintos microclimas que se encuentran y de las
incidencias que podriacutean causar a la hora de identificar los geacuteneros y especies de hongos endoacutefitos
dentro de la especie de frailejoacuten ademaacutes de posibles micro efectos del cambio climaacutetico a lo
largo del gradiente altitudinal (Caacutemara et al 2008)
2 Identificacioacuten y Filogenia de Hongos Endoacutefitos en el Marco del Cambio Climaacutetico
Nigrospora X X X X X
Penicillium X X X
Pleospora X X X X
Trichoderma X X X X
42
Las teacutecnicas empleadas en este estudio para lograr aislar e identificar los hongos endoacutefitos
dentro de la especie de frailejoacuten Espeletia hartwegiana se hacen importantes dentro del cambio
climaacutetico debido a que el margen de error en la identificacioacuten molecular es reducido (Bakermans
et al 2014) Al ser reducido el margen de error se pudo conocer con mayor certeza que especies
y geacuteneros de hongos endoacutefitos se encuentra dentro de la planta
Al conocer sus identidades es posible conocer las capacidades que tienen cada una de
estas especies de hongos endoacutefitos de producir metabolitos secundarios de infiltrarse y proteger
los tejidos de la planta de condiciones bioacuteticas y abioacuteticas competir contra herbiacutevoros y
fitopatoacutegenos (Schulz et al 2002) Con sus capacidades reconocidas es posible vincular los
efectos del cambio climaacutetico como estreacutes hiacutedrico aumento de la temperatura migraciones
tempranas enfermedades emergentes aumentos de la precipitacioacuten y heladas severas con las
reacciones por parte de los hongos endoacutefitos Estas reacciones podriacutean desencadenar aumentos de
la patogenicidad en las poblaciones de frailejones debido a los constantes disturbios y estreacutes al
que se ve sometido la planta generando asiacute notables declives poblacionales (Mendoza 2010)
Las relaciones filogeneacuteticas entre las especies halladas toman relevancia debido a las
sinergias positivas o negativas que se pueden presentar dependiendo de las especies que se
encuentren asociadas dentro de la planta (Lamichhane y Venturi 2015) como por ejemplo
Nigrospora y Colletotrichum pueden activar una sinergia negativa aumentando la virulencia y
afectando la especie Espeletia hartwegiana
Las especies de hongos endoacutefitos se identificaron por medio de secuencias obtenidas de
las regiones ITS (regioacuten de espaciadores internos transcritos) las cuales son regiones altamente
conservadas con tasas de evolucioacuten muy lentas en el tiempo permitiendo hacer inferencias
filogeneacuteticas (Baldwin et al 1995 Hamby y Zimmer 1992) es por ello que se usaron para
generar las diferentes identificaciones de los hongos endoacutefitos logrando asiacute identificar al geacutenero
Nigrospora con un total de 7 especies N sacchari N rubi N sphaerica N hainanensis N
gorlenkoana N oryzae y N aurantiaca pertenecientes a la familia Apiosporaceae orden
Xylariales Divisioacuten Ascomicota (veacutease figura 11) (Wang et al 2017)
Dentro del marco del cambio climaacutetico Nigrospora se caracteriza por poseer una
distribucioacuten cosmopolita y una amplia gama de hueacutespedes como lo es en este caso la especie E
hartwegiana que habita en los Andes Tropicales que se veraacuten alterados por los efectos del
cambio climaacutetico este geacutenero se ha logrado aislar en su forma endoacutefita en tallos y hojas de
43
diversas plantas tambieacuten como saprofitos en detritos hojarasca y larvas muertas (Uzor et al
2015 1)
Figura 11 Cladograma de las relaciones filogeneacuteticas del genero Nigrospora y Collectotrichum
Los hongos endoacutefitos hallados en esta investigacioacuten se encontraron dentro de la especie
de frailejoacuten E hartwegiana la cual pertenece a los Andes Tropicales ubicados maacutes
especiacuteficamente en los paacuteramos Estos ecosistemas son sensibles a los efectos del cambio
climaacutetico por su altura su alto endemismo y su sensibilidad al efecto de borde Cuando las
especies lentildeosas empiecen a migrar a lo largo del gradiente altitudinal es posible que los
frailejones se encuentren con suelos agrestes en donde sus semillas no podraacuten fijarse y los efectos
adversos de hongos como Nigrospora N oryzae y N sphaerica seraacuten maacutes notables
aprovechando las ventanas de oportunidad que generen los desbalances climaacuteticos (Sharma et al
2013)
Los cambios en los patrones climaacuteticos potenciaran la especie N sphaerica dentro del
frailejoacuten la cual tiene la capacidad de causar tizoacuten foliar en la especie Camellia sinensis siendo
una planta de teacute perteneciente a la divisioacuten de las magnolifiotas como el geacutenero Espeletia (Liu et
al 2015) Nigrospora fue el geacutenero maacutes ampliamente aislado en las identificaciones generadas a
partir de las hojas de frailejoacuten esto se veriacutea explicado debido a que N sphaerica tiene un
mecanismo de descarga de esporas efectivo que puede proyectar a la fuerza sus esporas a una
distancia de hasta 2 cm en vertical y 68 cm en horizontal como lo demostroacute Webster (1952)
ademaacutes el estudio de Wu y colaboradores (2004) tambieacuten evidencioacute que las esporas de
44
Nigrospora son uno de los grupos maacutes dominantes en la atmoacutesfera y estaacuten asociadas con
tormentas de polvo
La gran capacidad colonizadora del geacutenero Nigrospora le permite adaptarse a las
condiciones climaacuteticas cambiantes con la estrategia de dispersioacuten de sus esporas logrando
distanciarse del frailejoacuten hospedero para encontrar nuevos hospederos producto de las
migraciones tempranas a lo largo del gradiente altitudinal (Riveros 2010)
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
por encima del 80 lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen menos
mutaciones en los nodos internos del aacuterbol basados en la distribucioacuten de probabilidad asignadas
a los posibles arboles resultantes dando lugar a una hipoacutetesis evolutiva maacutes confiable (Schmidt y
Haeseler 2009) y con una parsimonia del 100 es decir el cladograma obtenido es el aacuterbol maacutes
corto (menos cambios) y el cual se ajusta mejor a los datos (Swofford 1997)
Teniendo asiacute una inferencia filogeneacuteticamente robusta los geacuteneros Collectotrichum y
Nigrospora pertenecen a la Divisioacuten Ascomycota ambos geacuteneros son particularmente agresivos
con las plantas en especial la especie Colletotrichum gloeosporioides la cual es la fase asexual
de Glomerella cingulata (Raj et al 2013) Se caracteriza por tener un amplio rango de
hospederos entre ellos el frailejoacuten a los cuales les causa una enfermedad conocida pudricioacuten
amarga antracnosis o chancros siendo esta la necrosis de frutos flores nervios y tallos
(Gonzaacutelez et al 2006 4)
Uno de los primeros reportes de afectaciones a especies de frailejoacuten se obtuvo en el 2009
en el Parque Natural Nacional Chingaza Colombia en donde encontraron las especies de
frailejoacuten Espeletia grandiflora y Espeletia uribei afectadas debido a insectos y una especie de
hongo del genero Colletotrichum (Medina 2009- 9 Hartung y Burton 1981)
Por ende que estos dos geacuteneros de hongos se encuentren juntos podriacutea tener
implicaciones en el aumento de virulencia potenciado a su vez por los desbalances abioacuteticos
causados por el cambio climaacutetico como lo han mencionado Belisario y colaboradores (2002) es
posible que las afectaciones no sean producidas por un uacutenico hongo si no por un complejo de
varios generando sinergismos entre diferentes especies que solas no hariacutean dantildeo a la planta pero
juntas podriacutean ser causantes de enfermedad (Lamichhane y Venturi 2015 4) aun mas en un
45
marco de cambio climaacutetico que gradualmente aumenta sus efectos nocivos sobre los paacuteramos
aumentando la temperatura cambios en la precipitacioacuten heladas intensas cambios en la
nubosidad mayor exposicioacuten a la radiacioacuten solar teniendo por resultado un iacutendice de estreacutes
mayor en las plantas de paacuteramo generando ventanas de oportunidades para que los hongos
endoacutefitos modifiquen sus roles de protector a patoacutegeno
En frailejones tambieacuten se ha reportado la presencia de Nigrospora y Trichoderma por
Bonilla y Argenis (2005) maacutes especiacuteficamente en la especie Espeletia grandiflora la cual es una
especie que habita en los paacuteramos de Colombia Ecuador y Venezuela en conjunto con Espeletia
hartwegiana (Luteyn 1999 3) El hecho que existan estos dos geacuteneros presentes en los frailejones
advierte de un equilibrio entre hongos endoacutefitos que se puede romper bajo las tensiones del
cambiante clima maacutes aun en los sensibles paacuteramos alto andinos
En cuanto a N sacchari se tiene reporte que es un productor de metabolitos con una
notable actividad herbicida demostrado en plantas cultivadas en invernadero (Fukushima et al
1998 2) debido a que existe evidencia que sugiere que los hongos endoacutefitos han evolucionado
directamente de hongos patoacutegenos (Schulz et al 1999 4) sumado al estreacutes del cambio climaacutetico
en cualquier momento de extrema tensioacuten hiacutedrica caloacuterica o heladas en estos ecosistemas de alta
montantildea Estos anomaliacuteas ambientales podriacutea disparar mecanismos de accioacuten fitopatoacutegenos
dentro de los frailejones generando un declive notable en sus poblaciones y por ende un declive
de la cantidad recurso hiacutedrico aguas abajo y su regulacioacuten (Diacuteaz et al 2002 5)
Como se mencionoacute anteriormente los medios de propagacioacuten de estos hongos endoacutefitos
pueden ser por medio de esporas que estaacuten en el ambiente aunque tambieacuten los vectores que
juegan un rol clave en su dispersioacuten son los insectos ya que estos pueden transportar esporas o
partes del hongo en su cuerpo patas o aparato bucal (Eigenbrode et al 2018 5)
Considerando la dispersioacuten homogeacutenea de los frailejones dentro del paacuteramo la carga
esporas en el ambiente y la herbivoriacutea de insectos es probable que se puedan ver infectados la
mayoriacutea de individuos (Varela 2016) es por ello que en el futuro puede que las afectaciones en
frailejones no sean producidas por hongos fitopatoacutegenos si no por hongos endoacutefitos que tuvieron
la ventana de oportunidad gracias al cambio climaacutetico aumento de la temperatura aumento en
las concentraciones de CO2 y alteraciones de precipitacioacuten que pueden hacer susceptible la planta
al ataque de estos hongos (Chakraborty 2011 Elad y Pertot 2014)
46
Para que se genere la enfermedad esta depende de la interaccioacuten de tres o cuatro factores
1) hospedero susceptible 2) agente causal (virulento) 3) condiciones ambientales favorables y 4)
tiempo (Francl 2001 2) la falta de alguno de ellos no permite que el patoacutegeno produzca la
enfermedad en la planta debido a ello algunas especies de frailejoacuten pueden tener una o dos cepas
que puedan actuar como fitopatoacutegenos pero no estaacuten activas si no estaacute completa su asociacioacuten
con otro patoacutegeno u otro factor
Ademaacutes existe un estudio que evaluoacute el efecto de la temperatura sobre la capacidad de
Fusarium verticillioides para generar infeccioacuten en plantas de maiacutez se probaron bajo 3 escenarios
de temperatura como resultados obtuvieron que la enfermedad de este hongo se veiacutea potenciada
por el aumento de temperatura en temperaturas por encima de 35degC (Murillo et al 2008) este
estudio da soporte a la hipoacutetesis de que existen factores que influyen significativamente a la
infeccioacuten en la planta tal vez el paacuteramo difiacutecilmente alcance 35degC no obstante la mayoriacutea de
especies son sensibles a cambios de temperatura de hasta de 1degC en donde modifican su
comportamiento rutas metaboacutelicas interacciones ecoloacutegicas y por ende sus defensas se ven
alteradas
Figura 12 Cladograma de las relaciones filogeneacuteticas del genero Trichoderma e Hypocrea
El geacutenero Trichoderma con seis especies diferentes (veacutease figura 12) fue el segundo
geacutenero maacutes numeroso despueacutes de Nigrospora que contoacute con siete especies aunque las primeras
especies que se analizaron fueron especies que principalmente actuaban generando enfermedades
en las plantas en las que se encontraban ante disturbios el geacutenero Trichoderma pertenece a la
47
familia Hypocreaceae y se caracteriza por tener una gran capacidad biocontroladora sobre
fitopatoacutegenos como Fusarium oxysporum los cuales podriacutean llegar masivamente con las
migraciones tempranas de insectos y plantas lentildeosas (Miles et al 2012 7)
No solo actuacutean repeliendo patoacutegenos internos si no tambieacuten patoacutegenos externos como
insectos que desean consumir u aprovechar la planta ya que estos pueden secretar diferentes
metabolitos secundarios y enzimas que actuacutean inhibiendo su desarrollo (Boughalleb-M et al
2018 8)
Es bien conocido que el geacutenero Trichoderma produce amplio rango de sustancias
antibioacuteticas ademaacutes de poseer la caracteriacutestica de parasitar otros hongos (Howell 2002 6)
Debido a la competencia que generan las especies del genero Trichoderma por espacio y
nutrientes inhiben o degradan pectinasas y otras enzimas que son esenciales para que los hongos
fitopatoacutegenos como Botrytis cinerea penetre en la superficie de las hojas logrando proteger de
esta forma al frailejoacuten que se veraacute expuesto a nuevas amenazas microbioloacutegicas auspiciadas por
los cambios en los patrones climaacutetico (Zimand et al 1996)
Ademaacutes de la capacidad del Trichoderma sp para atacar o inhibir el crecimiento de
patoacutegenos de plantas directamente descubrimientos recientes indican que tambieacuten pueden inducir
resistencia sisteacutemica y localizada a una variedad de patoacutegenos de plantas tales como el frijol es
protegido por Trichoderma harzianum de especies fitopatoacutegenas como Colletotrichum
lindemuthianum Botrytis cineacuterea las cuales producen lesiones en las hojas de la planta (Harman
y Howell 2004) Incluso se descubrioacute que ciertas especies de Trichoderma tienen una influencia
sustancial en el buen crecimiento y desarrollo de las plantas sin y con condiciones de estreacutes
climaacutetico (Lorito 1998)
Como lo demostraron Bonilla y Argenis (2005) en su estudio llevado a cabo en el
frailejoacuten Espeletia grandiflora diferentes cepas pertenecientes a Trichoderma pueden proteger
esta especie de paacuteramo de organismos fitopatoacutegenos emergentes por el cambio climaacutetico por
ende si puede proteger a E grandiflora tambieacuten puede proteger la especie E hartwegiana lo
que le permitiraacute sobrellevar diferentes ataques fortuitos de patoacutegenos e insectos que nunca han
logrado alcanzar las altitudes de paacuteramo (3200 - 5000 m)
Cuando el cambio climaacutetico aumente la temperatura en 1degC o 2degC traeraacute consigo sus
efectos de modificacioacuten en cuanto a la estructura y composicioacuten de las especies de plantas que
48
existen en zonas altas para este escenario basado en los caacutelculos de Thuiller (2007) las especies
se habriacutean desplazado en el gradiente altitudinal aproximadamente 450 metros para el antildeo 2070
Las consecuencias de este desplazamiento serian uacutenicas debido a que gran cantidad de
insectos y patoacutegenos tendriacutean migraciones tempranas en el gradiente altitudinal en buacutesqueda de
su optimo teacutermico llevando a cabo un proceso conocido como Termofilizacioacuten (Cuesta et al
2019) es ahiacute donde entra en juego la proteccioacuten que le pueden brindar especies endoacutefitos como
las que pertenecen al geacutenero Trichoderma en el marco de cambio climaacutetico
En concordancia con las cepas de Trichoderma y el geacutenero Hypocrea que tambieacuten
pertenece a la familia Hypocreaceae se conocen maacutes de 700 secuencias de peptaiboles (peacuteptidos
bioloacutegicamente activos) de ambos geacuteneros donde algunos de eacutestos son ecoloacutegica y
comercialmente relevantes debido a su potencial antimicrobiano actividad enzimaacutetica uacutetil para
competir por espacio y alimento de los nuevos patoacutegenos emergentes procedentes de las
alteraciones climaacuteticas y las migraciones tempranas (Degenkolb et al 2008 6)
En cuanto al geacutenero Hypocrea tambieacuten se ha reportado actividad antimicrobiana y anti
fuacutengica contra Staphylococcus aureus y Fusarium oxysporum (Gogoi et al 2008 2) habitando
los geacuteneros Trichoderma e Hypocrea dentro de especies de frailejoacuten como E hartwegiana se
espera que puedan generar sinergias de proteccioacuten que las ayude a combatir los fuertes y
extremos eventos climaacuteticos
Figura 13 Cladograma de las relaciones filogeneacuteticas del genero Annulohypoxylon
Este cladograma se encuentra soportado por valores estadiacutesticos de Maacutexima Verosimilitud
del 100 (veacutease figura 13) lo que indica que entre maacutes verosiacutemil (maacutes cercano a 100) existen
menos mutaciones en los nodos internos del aacuterbol por ende este aacuterbol contiene la menor
cantidad de mutaciones o cambios dentro sus nodos internos teniendo como resultado una
hipoacutetesis evolutiva confiable (Schmidt y Haeseler 2009) y con una parsimonia del 100 que
indica que el cladograma obtenido es el aacuterbol maacutes corto (menos cambios) y el cual se ajusta
mejor a los datos (Swofford 1997)
49
El geacutenero Annulohypoxylon es de importancia dentro del marco del cambio climaacutetico ya
que pertenece a la familia Xylariaceae y se caracteriza por poseer metabolitos secundarios
antitumorales-citotoacutexicos como la anuloescuamulina que ayudan a combatir el estreacutes abioacutetico
(Cheng et al 2012) estas cepas de hongos son principalmente saprofitos y oportunistas han sido
reportadas en hojas sanas y afectadas de frailejones en otros estudios (Gonzaacutelez 2012 Nonsoque
2012) siendo un geacutenero resiliente a las perturbaciones ambientales como los cambios en los
patrones climaacuteticos su relacioacuten como mutualista dentro del frailejoacuten se mantendraacute hasta que el
frailejoacuten se adapte o colapse ante el estreacutes abioacutetico
Aunque no se conocen como fitopatoacutegenos al contrario pueden actuar como mutualistas
no se descarta que puedan estar involucrados en el desarrollo de diferentes enfermedades en
plantas cuando las condiciones ecosistemicas se alteran a puntos extremos (Gonzalez et al 2017)
Figura 14 Cladograma de las relaciones filogeneacuteticas del genero Daldinia Hypoxylon Biscogniauxia
Nemania y Humiacutecola
El geacutenero Daldinia posee unos valores de MPML correspondientes a 10094 con lo cual
se encuentra bien soportado estadiacutesticamente el geacutenero Daldinia se encuentra ubicado en la
familia Xylariaceae al igual que el geacutenero Hypoxylon Biscogniauxia y Nemania que en su
respectivo orden contienen un soporte estadiacutestico MPML de 10082 10099 y 100100 (veacutease
figura 14) los cuales pertenecen a la divisioacuten Ascomycota las cepas de Daldinia se caracterizan
principalmente por ser especies endoacutefitas teniendo la capacidad de estar dentro de su hueacutesped
mucho tiempo sin causarle dantildeo o al menos un dantildeo evidente por medio de su estado asexual
50
No obstante su estado se puede modificar cuando las condiciones externas se modifican
abruptamente en poco tiempo como es el caso de las alteraciones en la temperatura
precipitacioacuten y concentraciones de dioacutexido de carbono (CO2) auspiciados por el cambio
climaacutetico (Saacutenchez et al 2013)
Un estudio realizado por Prieto (2017) en la especie de frailejoacuten E grandiflora encontroacute
que la especie Daldinia eschscholtzii estaba asociado en el siacutentoma de clorosis (falta de clorofila
en las hojas) Tambieacuten se reportoacute que especies de Daldinia como Daldinia conceacutentrica tienen la
capacidad de controlar nematodos fitopatoacutegenos (Silva et al 2020 104) los cuales pueden invadir
y romper los tejidos de la planta dantildeando sus estructuras los nematodos tambieacuten seraacuten sujetos a
migraciones tempranas por el cambio climaacutetico que trae consigo presiones de seleccioacuten por ende
nuevas amenazas para los frailejones (Nakashima et al 2007)
Como hemos visto hasta este punto las especies de hongos endoacutefitos se muestran en
muchos casos como benefactores y protectores de las diferentes plantas en las que habitan
incluyendo diferentes especies de frailejones
Los hongos endoacutefitos protegen tanto de factores bioacuteticos como virus hongos
fitopatoacutegenos bacterias insectos y factores abioacuteticos como las intemperancias del clima no
obstante en su relacioacuten endoacutefita geacuteneros como Hypoxylon que pertenece igual que Daldinia a la
familia Xylariaceae pueden tener roles amenazadores en conjunto con el cambio climaacutetico
seguacuten Gonzales y colaboradores (2019) inocularon cepas pertenecientes al geacutenero Hypoxylon en
hojas de Theobroma cacao y encontraron que en 48 horas se comenzoacute a notar las marcas de
clorosis y deshidratacioacuten foliar esto demuestra que cepas de este geacutenero pueden tener efectos
fitopatoacutegenos ocasionando muerte descendente pudricioacuten del tronco negro y resinosis de ramas
de mezquite (De la Torre-Almaraacutez et al 2003 De la Torre-Almaraacutez et al 2009)
La presencia de este geacutenero dentro del frailejoacuten en un marco del cambio climaacutetico lo
convierte en una amenaza latente ya que las condiciones seraacuten cada vez maacutes agrestes en los
paacuteramos de los Andes Tropicales Puede favorecer el desarrollo de este geacutenero que tiene las
enzimas necesarias para producir la pudricioacuten del tronco negro entre otras enfermedades
relevantes
51
Tambieacuten las especies del geacutenero Hypoxylon se han reportado como endoacutefitos en tejidos de
plantas lentildeosas o herbaacuteceas (Petrini y Petrini 1985 3) asiacute como saprofitos o fitopatoacutegenos
deacutebiles en madera de aacuterboles de zonas templadas o tropicales (Miller 1961) como lo menciona
Miller (1961)
Los geacuteneros Biscogniauxia Nemania tambieacuten estaacuten dentro de la familia Xylariaceae y
Divisioacuten Ascomycota un aspecto interesante de las especies de Biscogniauxia es que son
paraacutesitos exclusivos de las angiospermas dicotiledoacuteneas en donde concluyeron que todas las
especies pertenecientes a este geacutenero son patoacutegenos umllevesuml que se aprovechan de situaciones de
sequiacutea y estreacutes hiacutedrico para aumentar su patogenicidad (Ju et al 1998)
La especie de frailejoacuten E hartwegiana es una Dicotiledoacutenea eso explica porque
podemos encontrar esta especie de hongo endoacutefito en el frailejoacuten del presente estudio siendo de
vital importancia denotar que las especies de Biscogniauxia se aprovechan especialmente de las
situacioacuten de sequiacutea y estreacutes hiacutedrico difiacutecilmente las especies de este geacutenero podriacutean afectar a los
frailejones debido a que se caracterizan por ser reguladores de los ciclos hiacutedricos el agua es el
recurso que maacutes tiende a existir en un paacuteramo por la estructura de sus suelos compuesta por
andosoles porosos y necromasa acumulada teniendo asiacute un alto iacutendice de retencioacuten hiacutedrica
(Hernandez y Muntildeoz 2015) de modo que el cambio climaacutetico no favorece al geacutenero
Biscogniauxia en su estado patogeacutenico dentro del ecosistema de paacuteramo
En cuanto al geacutenero Nemania Da Silva y colaboradores (2018) llevaron a cabo una
investigacioacuten en la cual identificaron cepas pertenecientes a los geacuteneros Colletotrichum
Phyllosticta Xylaria Nemania y Alternaria en la especie ornamental Pachystachys lutea en
donde las mismas cepas fueron ensayadas para inhibir el crecimiento de hongos fitopatoacutegenos
como Colletotrichum sp en donde encontraron que Nemania era capaz de inhibir por medio de la
secrecioacuten de distintas enzimas el crecimiento de dichas especies gracias a su actividad
enzimaacutetica el geacutenero Nemania puede compensar los desequilibrios causados por el cambio
climaacutetico en teacuterminos de hongos fitopatoacutegenos
Hay que tener en cuenta que los elementos que dan forma a las interacciones planta-
endoacutefito incluyen el modo de transmisioacuten el patroacuten de infeccioacuten la edad de la planta las
condiciones ambientales potenciadas por el cambio climaacutetico y los antecedentes geneacuteticos (Aly et
al 2011 6) Los frailejones de la especie E hartwegiana que se muestrearon para el estudio se
52
tomoacute las hojas sanas lo que indica que poseiacutean estas cepas de endoacutefitos en equilibrio siendo este
uno de los pocos reportes de este geacutenero en especies de alta montantildea en los Andes Tropicales
El geacutenero Humiacutecola se encuentra soportado por un MPML de 10076 siendo una
hipoacutetesis evolutiva considerable es el uacutenico de la figura 14 que no estaacute dentro de la familia
Xylariaceae ya que pertenece a la familia Chaetomiaceae aunque se halla en la divisioacuten
Ascomycota se podriacutea considerar a la especie Humiacutecola fuscoatra como una de las especies de
endoacutefitos maacutes umlinofensivasuml debido a trabajos de investigacioacuten reportados como el de Menzies y
colaboradores (1998) y el de Naik y colaboradores (2009)
Humiacutecola fuscoatra no se considera un peligro latente para la especie E hartwegiana ya
que existe en equilibrio con su entorno Estaacute comprobado por medio de caracterizaciones
quiacutemicas que Humiacutecola fuscoatra es un importante productor de moleacuteculas de estimacioacuten
medica como el resorciacuteclico y lactonas aacutecidas (Mejia et al 2014) Por lo tanto es una especie
prometedora con un reservorio de metabolitos secundarios de importancia medicinal e industrial
que de seguir los efectos nocivos del cambio climaacutetico en los Andes Tropicales maacutes
especiacuteficamente en los paacuteramos esta especie podriacutea desaparecer
Figura 15 Cladogramas de las relaciones filogeneacuteticas del genero Diaporthe Crassiparies Massarina
Cryptocoryneum Cyphellophora y los grupos externos
Uno de los grupos mejores soportados en este cladograma es el grupo de Diaporthe en el
cual se encontraron cuatro especies siendo uno con maacutes abundancia de especies del estudio y que
posee un MPML de 10085 10089 y 10095 acorde a sus 3 nodos (veacutease figura 15) teniendo asiacute
una hipoacutetesis evolutiva confiable de estas relaciones las especies de Diaporthe pertenecen a la
familia Diaporthaceae de la divisioacuten Ascomycota y son especies de endoacutefitos que se distinguen
por inhibir el crecimiento de hongos patoacutegenos en el hospedero que se encuentren El estudio de
53
Da Silva y colaboradores (2018) mostroacute que cepas de Diaporthe sp Inhibieron el crecimiento de
Fusarium oxysporum y Colletotrichum sp hongos fitopatoacutegenos que aumentaran su presencia
aprovechando las alteraciones producidas por el cambio climaacutetico en los paacuteramos
Las especies de este geacutenero estaacuten en continuo cambio entre endoacutefitos y patoacutegenos
funcionando en la mayoriacutea de los casos en una relacioacuten mutualista en donde le brindan a la planta
mayor tolerancia a las sequias proteccioacuten de patoacutegenos y pestes caracteriacutestica uacutetil frente a los
retos ecoloacutegicos que plantea el cambio climaacutetico en los Andes Tropicales (Hubbard et al 2014)
se conocen cerca de 800 especies de este geacutenero asociadas a una amplia gama de plantas
vasculares a nivel mundial (Rossman et al 2015)
Esto explica porque las especies de Diaporthe fueron una de las maacutes numerosas
encontradas en la especie de alta montantildea E hartwegiana teniendo en cuenta que esta especie
pasa raacutepidamente de un estado endoacutefito a un estado patogeacutenico y dada su faacutecil y alta distribucioacuten
representa un peligro en conjunto con el cambio climaacutetico para las especies de alta montantildea como
los frailejones y supone una amenaza para el balance hiacutedrico y ecosistemico en el futuro
El geacutenero Crassiparies de la familia Neohendersoniaceae y el geacutenero Massarina de la
familia Massarinaceae pertenecen al orden Pleosporales aunque se discute debido a las
similaridades morfoloacutegicas y genoacutemicas con Massarina si Crassiparies deberiacutea estar en la
familia Massarinaceae (Hyde et al 2013 4) Se tiene poca informacioacuten sobre la especie
Crassiparies quadrisporus por ende este puede ser uno de los primeros reportes de esta especie
en una especie de alta montantildea como lo es el frailejoacuten especiacuteficamente en la especie E
hartwegiana en un marco de cambio climaacutetico el geacutenero Crassiparies necesita mayores estudios
a nivel ecoloacutegico y microbioloacutegico para entender su papel dentro de esta crisis climaacutetica
Mientras tanto el geacutenero Massarina ha logrado ser aislado de agua dulce con el fin de
obtener fuentes de productos naturales bioloacutegicamente activos por medio de las investigaciones
quiacutemicas de Shearer (1999 2001) en una especie de Massarina obtuvieron como resultado el
aislamiento de tres nuevos sequiterpenoides y dos lactonas derivadas de policeacutetidos con nuevos
sistemas de anillos lo que indica que esta especie tambieacuten puede tener un rol importante
mediante la produccioacuten de enzimas y metabolitos secundarios en el marco de las invasiones
bioloacutegicas impulsadas por el cambio climaacutetico
54
El geacutenero Cryptocoryneum de la familia Cryptocoryneaceae existe poca informacioacuten de
sus mecanismos de accioacuten como endoacutefito o patoacutegeno maacutes aun en especies de los Andes
Tropicales este podriacutea ser uno de los primeros reportes sobre esta especie de hongo en una
especie de frailejoacuten especiacuteficamente E hartwegiana en el 2015 se reportoacute en Brasil el
descubrimiento de una nueva especie del genero Cryptocoryneum trataacutendose de Cryptocoryneum
parvulum hallada en las hojas de la especie Araucaria angustifolia (Gimnosperma) (Silva et al
2015 1) la informacioacuten sobre el geacutenero Cryptocoryneum en relacioacuten al cambio climaacutetico y sus
efectos dentro de especies de alta montantildea como el frailejoacuten auacuten son poco conocidas
Uno de los geacuteneros maacutes cercanos a los grupos externos del cladograma es Cyphellophora
perteneciente a la familia Cyphellophoraceae el cual posee un MPML de 10091 logrando una
hipoacutetesis evolutiva confiable Diversas especies de Cyphellophora han logrado ser aisladas de
infecciones superficiales de la piel o las untildeas en humanos y de materiales vegetales (Decock et
al 2003 Cheewangkoon et al 2009) Esto indica su amplia distribucioacuten y su capacidad para ser
patogeacutenica tanto en plantas como en animales maacutes aun en condiciones desfavorables para la
planta como son las condiciones de cambio climaacutetico
De la especie Cyphellophora reptans se tiene reporte que su morfologiacutea es muy similar a
la de una levadura por lo que es faacutecilmente confundida con una (Reacuteblovaacute et al 2013) Es posible
que este sea el uacutenico reporte de Cyphellophora reptans en las hojas de un frailejoacuten o especie de
alta montantildea que habite en los troacutepicos como E hartwegiana debido a ello sus mecanismos de
accioacuten en especies de paramo dentro de un marco de cambio climaacutetico es poco conocido Se
necesitan mayores estudios enfocados en esta especie para entender su ecologiacutea y dinamismo
ecoloacutegico respecto a los cambios en las variables ambientales
Se eligieron las especies Coprinellus domesticus y Coprinellus xanthothrix como grupos
externos con el fin de enraizar el aacuterbol debido a que pertenecen a la Divisioacuten Basidiomycota la
cual difiere en grado de la divisioacuten Ascomycota aunque conserva semejanzas conocidos como
caracteres ancestrales o plesiomorficos que avanzan a medida la filogenia se construye con los
datos de las diferentes secuencias de especies evidenciando novedades evolutivas conocidas
como apomorfias que cuando se dan en un grupo en particular pasan a ser sinapomorfias es decir
novedades evolutivas dentro de un grupo particular (Lehtonen 2020) Se optoacute por enraizar con
55
estas dos especies con el objetivo de dar robustez al aacuterbol filogeneacutetico (Barriel y 1998) lo cual se
consiguioacute como se nota en los estadiacutesticos del aacuterbol filogeneacutetico
56
57
Conclusiones
Se rechaza la hipoacutetesis nula (H0) y se concluye que la identificacioacuten y caracterizacioacuten
molecular de los hongos endoacutefitos encontrados en la especie de frailejoacuten Espeletia hartwegiana
ubicada en el paacuteramo romerales Quindiacuteo Colombia es diferente en 80 de sus geacuteneros y
especies de otras identificaciones de hongos endoacutefitos realizadas en especies de frailejoacuten
Por lo tanto los nuevos geacuteneros y especies de hongos endoacutefitos encontrados en el frailejoacuten
plantean nuevos desafiacuteos respecto al cambio climaacutetico ya que tienen un rol vital en la
supervivencia de estas especies de alta montantildea como los frailejones debido a su relacioacuten
mutualista o patoacutegena maacutes aun en los paacuteramos que son altamente sensibles a los efectos del
cambio climaacutetico incluso tan sensibles como las zonas polares (Bradley et al 2006)
La mayoriacutea de especies encontradas en el presente estudio tienen potencial para ayudar a
combatir el estreacutes abioacutetico producido por el cambio climaacutetico en los Andes Tropicales
Especiacuteficamente pueden ayudar a combatir el estreacutes bioacutetico producido por virus hongos
bacterias e insectos en un marco de cambio climaacutetico aumentos de temperatura cambios en la
concentracioacuten de CO2 cambios en la precipitacioacuten migraciones tempranas cambios en la
nubosidad disponibilidad hiacutedrica alteraciones en la estructura y composicioacuten de las especies
endeacutemicas del paacuteramo
58
59
Obras citadas
Anderson EP Marengo JA Villalba R Halloy SR Young B Cordero D Gast F
Jaimes E y Ruiz D 2012 Consecuencias del cambio climaacutetico en los ecosistemas y
servicios ecosisteacutemicos de los Andes Tropicales Cambio climaacutetico y Biodiversidad en
los Andes Tropicales
Akaike H 1974 A new look at the statistical model identification IEEE Transactions on
Automatic Control 19 716ndash723
Altschul SF Gish W Miller W Myers EW Lipman DJ 1990 Basic local alignment search tool
Journal of Molecular Biology 215 403ndash410
Aly AH Debbab A Proksch P 2011 Fungal endophytes unique plant inhabitants with great
promises Appl Microbiol Biotechnol 90(6)1829ndash1845 12 de noviembre
httpsdoiorg101007s00253-011-3270-y
Barriel V and Tassy P 1998 Rooting with Multiple Outgroups Consensus Versus Parsimony
Cladistics 14 193-200 httpsdoiorg101111j1096-00311998tb00332x
Baldwin BG Sanderson MJ Poner JM Wojciechowski MF Campbell CS y Donoghue MJ
1995 The ITS region of nuclear ribosomal DNA a valuable source of evidence on
angiosperm phylogeny Annals of the Missouri Botanical Carden 82 247-277
Bakermans C Skidmore ML Douglas S McKay CP 2014 Molecular characterization of
bacteria from permafrost of the Taylor Valley Antarctica FEMS Microbiology Ecology
89 331ʹ346
Belisario A Maccaroni M Corazza L Balmas V Valier A 2002 Occurrence and Etiology of
Brown Apical Necrosis on Persian Walnut Fruit Plant Disease 86(6)599-602 12 de
noviembre 101094pdis2002866599
Berdugo-Lattke M L Goacutenzalez F Rangel-Ch J O amp Goacutemez F 2016 P-type based
dimensionality reduction for open contours of Colombian Paacuteramo plant species
Ecological informatics 36 1-7
Bonilla Goacutemez MA 2005 Estrategias adaptativas de plantas del paacuteramo y del bosque altoandino
en la cordillera oriental de Colombia Universidad Nacional de Colombia Facultad de
Ciencias Departamento de Biologiacutea
60
Boughalleb-MrsquoHamdi N Salem I MrsquoHamdi M 2018 Evaluation of the efficiency of
Trichoderma Penicillium and Aspergillus species as biological control agents against
four soil-borne fungi of melon and watermelon Egyptian Journal of Biological Pest
Control 28(1) 12 de noviembre 101186s41938-017-0010-3
BOLFOR Mostacedo Bonifacio Fredericksen Todd S 2000 Manual de Meacutetodos Baacutesicos de
Muestreo y Anaacutelisis en Ecologiacutea Vegetal Santa Cruz Bolivia
Buytaert W R Ceacutelleri B De Biegravevre F Cisneros G Wyseure J Deckers y R Hofstede 2006
Human impact on the hydrology of the Andean paacuteramos EarthminusScience Reviews 79
53minus72
Blackwell M y JW Spatafora 2004 ldquoFungi and their alliesrdquo En Biodiversity of Fungi
Inventory and Monitoring Methods editado por Mueller G M Bills G F amp Foster
MS 7-2 Elsevier Academic Press
Bradley R S M Vuille H F Diacuteaz y W Vergara 2006 Threats to water supplies in the
tropical Andes Science 3121755minus1756col
Cabezas L Calderon C Medina LM Bahamon I Cardenas M Bernal AJ Gonzalez A
Restrepo S 2012 Characterization of cellulases of fungal endophytes isolated from
Espeletia spp Journal of Microbiology 50 (6) 1009-1013
Cano Botero G D Maya L C Hernaacutendez A M amp Agudelo Quintana D 2015 Estudio
Teacutecnico Econoacutemico Social Ambiental (ET-ESA) del Complejo de Paacuteramos Los Nevados
en jurisdiccioacuten de la Corporacioacuten Autoacutenoma Regional del Quindiacuteo (CRQ)
Catalogo Virtual de Alta Montantildea 2020 Fotografiacutea de la especie Espeletia hartwegiana
Universidad EIA 17 de julio httpscatalogofloraaltamontanaeiaeducospecies13
Colwell R K G Brehm C L Cardeluacutes A C Gilman y J T Longino 2008 Global warming
elevational range shifts and lowland biotic attrition in the wet tropics Science 322 258minus
261
Compant S Cleacutement C amp Sessitsch A 2010 Colonization of plant growth‐promoting bacteria in
the rhizo‐ and endosphere of plants importance mechanisms involved and future
prospects Soil Biol Biochem 42 669ndash678
61
Cooke D Drenth A Duncan J M Wagels G Brasier CM 2000 A molecular phylogeny of
Phytophthora and related Oomycetes Fungal Genet Biol 30 17-32
Cortes J y Sarmiento E 2013 Visioacuten socioecosisteacutemica de los paacuteramos y la alta montantildea
colombiana memorias del proceso de definicioacuten de criterios para la delimitacioacuten del
paacuteramo Bogotaacute Instituto Humbolt Ministerio de Medio Ambiente
Cuesta F Llambi L Huggel C Drenkhan F Cosling W Muriel P Tovar C 2019 New
land in the Neotropics a review of biotic community ecosystem and landscape
transformations in the face of climate and glacier change
Cuesta Camacho F 2007 Efectos del cambio climaacutetico en el rango de distribucioacuten de especies
en los Andes del Norte La Paz Curso GLORIA Bolivia
Chakraborty S Climate change and plant diseases Plant Pathology 2011 60(1)1-1 11 de
noviembre 101111j1365-3059201002415
Cheewangkoon R Groenewald JZ Summerell BA Hyde KD To- Anun C Crous PW 2009
Myrtaceae a cache of fungal biodiversity Persoonia 2355ndash85
Clay K 1998 Fungal Endophytes of Grasses A Defensive Mutualism Between Plants and Fungi
Ecology 691016
Darriba D Taboada GL Doallo R Posada D 2012 jModelTest 2 more models new heuristics
and parallel computing Nature Methods 9 (8) 772
Da Silva Ribeiro A Polonio JC Costa AT 2018 Bioprospection of Culturable Endophytic
Fungi Associated with the Ornamental Plant Pachystachys lutea Curr Microbiol 75 588ndash
596 15 de noviembre httpsdoiorg101007s00284-017-1421-9
David S Hibbett Meredith Blackwell Timothy Y James Joseph W Spatafora John W Taylor
et Rytas Vilgalys 2018 laquoPhylogenetic taxon definitions for Fungi Dikarya Ascomycota
and Basidiomycotaraquo IMA Fungus 9 (5) 291-298 14 de noviembre
105598imafungus2018090205
De Bary A 1866 Morphologie und Physiologie der Pilze Flechten und Myxomyceten
Engelann Leipzig Alemania
Decock C Delgado-Rodriguez G Buchet S Seng JM 2003 A new species and three new
combinations in Cyphellophora with a note on the taxonomic affinities of the genus and
62
its relation to Kumbhamaya and Pseudomicrodochium Antonie Van Leeuwenhoek
84209ndash216
Degenkolb T H von Dohren KF Nielsen GJ Samuels y H Bruckner 2008 Recent
advances and future prospects in peptaibiotics hydrophobin and mycotoxin research
and their importance for chemotaxonomy of Trichoderma and Hypocrea Chemical
Biodiversity 5 671-680
De La Torre A R Cota T F M and San M F 2003 First report of Hypoxylon diatrypeoides
inducing Dieback and Black Trunk Rot on Mesquite (Prosopis laevigata) in Mexico
Plant Dis 87(4)447-453
mdashmdashmdash A R Cota T F M Garciacutea R J L Campos J E y San M F 2009 Etiologiacutea de la
muerte descendente del mezquite (Prosopis laevigata L) en la reserva de la biosfera del
Valle de Zapotitlaacuten Meacutexico Agrociencia 43(2)197-208
Diacuteaz MA Navarrete JD Suaacuterez T 2002 Paacuteramos Hidrosistemas Sensibles Revista de
Ingenieriacutea Universidad de Los Andes 1 (22) 66ndash73
Diacuteaz S Lavorel S de Bello F Queacutetier F Grigulis K y Matthew Robson T 2007
Incorporating plant functional diversity effects in ecosystem service assessments 14
20684ndash20689
Eigenbrode S Forest-Perez N Davis T 2018 Insect-Borne Plant Pathogens and Their Vectors
Ecology Evolution and Complex Interactions 63 (1) 169-191 15 de noviembre
101146annurev-ento-020117-043119
Elad Y Pertot I Climate Change Impacts on Plant Pathogens and Plant Diseases 2014 Journal
of Crop Improvement 28(1)99-139 11 de noviembre 101080154275282014865412
Fadrique Belen amp Baez Selene amp Duque Alvaro amp Malizia Agustina amp Blundo Cecilia amp
Carilla Julieta amp Osinaga Oriana amp Malizia Lucio amp Silman Miles amp Farfan-Rios
William amp Malhi Yadvinder amp Young Kenneth amp Cuesta Francisco amp Homeier
Juergen amp Peralvo Manuel amp Pin|to Esteban amp Jadaacuten Oswaldo amp Aguirre Nikolay amp
Aguirre Zhofre amp Feeley Kenneth 2018 Widespread but heterogeneous responses of
Andean forests to climate change Nature 564 16 de noviembre 101038s41586-018-
0715-9
Farias Chiquillo A d 2013 Elementos para una propuesta de educacioacuten ambiental desde el
enfoque criacutetico social como alternativa para el acercamiento de poblacioacuten escolar
urbana al ecosistema de paacuteramo
63
Font Quer P (comp) 1953 Diccionario de Botaacutenica Labor Barcelona
Fukushima T Tanaka M Gohbara M et al 1998 Phytotoxicity of three lactones from
Nigrospora sacchari Phytochemistry 48 625ndash630
Francl L 2001 The Disease Triangle A Plant Pathological Paradigm Revisited The Plant Health
Instructor 51(1)1-5 12 de noviembre 101094phi-t-2001-0517-01
Foden W B G M Mace JminusC Vie A Angulo S H M Butchart L DeVantier H T Dublin
A Gutsche S Stuart y E Turak 2009 Species susceptibility to climate change impacts
Pp 77minus88 en Wildlife in a changing world an analysis of the 2008 IUCN Red List of
threatened
Gaitaacuten Naranjo P 2018 Identificacioacuten de hongos asociados a lesiones de frailejones (Espeletia
spp) en paramos de Cundinamarca Pontificia Universidad Javeriana 17
Garciacutea VL Rios Q Aamp Molina LJ 2010 Structure Plant Composition And Leaf Litter
Decomposition In Soil At Two Sites Of An Andean Cloud Forest (Reforested And In
Spontaneous Succession) In Pentildeas Blancas Calarcaacute (Quindiacuteo) Colombia
ActualidadesBiologicas32 (93) 147-164
Gogoi DK S Mazumder R Saikia y TC Bora 2008 Impact of submerged culture conditions
on growth and bioactive metabolite produced by endophyte Hypocrea spp NSF-08
isolated from Dillenia indica Linn in North-East India World Journal of Microbiology
and Biotechnology 24 79-87
Gonzaacutelez D 2012 Estudio preliminar de la composicioacuten de hongos endoacutefitos en Espeletia
argentea en la cuenca de la quebrada Calostros (PNN Chingaza) Pontificia Universidad
Javeriana 26
Gonzaacutelez E Sutton TB Correll JC2006 Clarification of the etiology of Glomerella leaf spot and
bitter rot of apple caused by Colletotrichum spp based on morphology and genetic
molecular and pathogenicity tests Phytopathology 96 982ndash992
Gonzalez V Martin J Siles J Gonzalez-Tejero M Reyes F Platas G 2017 Biodiversity and
chemotaxonomy of Preussia isolates from the Iberian Peninsula Mycological Progress
16(7)713-728 12 de noviembre 101007s11557-017-1305-1
Guindon S y Gascuel O 2003 A simple fast and accurate method to estimate large phylogenies
by maximum-likelihood Systematic Biology 52 696-704
64
Cheng MJ Wu MD Yuan GF Chen YL SU YS Hsieh MT Chen ES 2012 Metabolitos
secundarios y actividades citotoacutexicas del hongo endofiacutetico Squam Annulohypoxylon
Phytochem Lett 5 (1) 219-223
Hamby RK y Zimmer EA 1992 Ribosomal RNA as a phylogenetic tool in plant systematics
En Soltis PS Soltis DE ancl Doyle JJ Eclrs Molecular systematics of plants New
York Chapman amp Hall 50-9 1
Hernandez M amp Muntildeoz J 2015 Influencia de las curvas de retencioacuten de humedad en la
estimacioacuten de la evaporacioacuten de acuiacuteferos someros Departamento de Ingenieriacutea
Hidraacuteulica y Ambiental Escuela de Ingenieriacutea Pontificia Universidad Catoacutelica de Chile
Herrera T Ulloa M 1990 El reino de los hongos micologiacutea baacutesica y aplicada Meacutexico DF
Universidad Nacional Autoacutenoma de Meacutexico p 550
Hofstede R 2000 Aspectos teacutecnicos ambientales de la forestacioacuten en los paacuteramos En La
forestacioacuten en los paacuteramos Serie Paacuteramo 6 editado por G Medina C Josse y P Mena 41-
66 Quito
Howell C R 2002 Cotton seedling preemergence damping-off incited by Rhizopus oryzae and
Pythium spp and its biological control with Trichoderma spp Phytopathology 92 177ndash
180
Hubbard M Germida J Vujanovic V 2014 Fungal endophytes enhance wheat heat and drought
tolerance in terms of grain yield and second-generation seed viability Journal of applied
microbiology 116 109-122
Hyde KD Jones EBG Liu JK Ariyawansa H Boehm E Boonmee S Braun U Chomnunti P
Crous PW Dai DQ Diederich P Dissanayake A Doilom M Doveri F Hongsanan S
Jayawardena R Lawrey JD Li YM Liu Y-X Lucking R Monkai J Muggia L Nelsen
MP Pang KL Phookamsak R Senanayake IC Shearer CA Suetrong S Tanaka K
Thambugala KM Wijayawardene NN Wikee S Wu HX Zhang Y Aguirre-Hudson B
Alias SA Aptroot A Bahkali AH Bezerra JL Bhat DJ Camporesi E Chukeatirote E
Gueidan C Hawksworth DL Hirayama K De Hoog S Kang JC Knudsen K Li WJ Li
XH Liu ZY Mapook A McKenzie EHC Miller AN Mortimer PE Phillips AJL Raja
HA Scheuer C Schumm F Taylor JE Tian Q Tibpromma S Wanasinghe DN Wang Y
Xu JC Yacharoen S Yan JY Zhang M 2013 Families of Dothideomycetes Fungal
Divers 63 1ndash313
65
Ies JM James TY Mcmunn M amp I Ibantildeez 2013 Predecting species-specific responses of
fungi to climatic variation using historical records Global Change Biology 19 3145-
3154
Intergovernmental Panel on Climate Change 2000 umlLand use Land-use Change and Forestryuml
Cambridge U K Cambridge University Press 12 de noviembre
httpwwwipccchipccreportssresland_useindexphpidp=0
IPCC 2020 Cambio Climaacutetico Resumen para responsables de poliacuteticas editado por Valeacuterie
Masson-Delmotte Hans-Otto Poumlrtner Jim Skea Panmao Zhai Debra Roberts
Priyadarshi R Shukla Cambridge Cambridge University Press
Jaramillo CA Congreso Mundial de Paacuteramos Bogotaacute Ministerio del Medio Ambiente 2003
Jimeacutenez M A Chain y B Locatelli 2010 umlEfectos del cambio climaacutetico en la distribucioacuten de
zonas de vida en Centroameacutericauml Comunicacioacuten Teacutecnica en Recursos Naturales y
Ambiente 59 32-40
Ju Y-M Rogers JD San Martin Gonzales F amp Granmo A 1998 The genus Biscogniauxia
Mycotaxon 66 1-98
Judd W S C S Campbell E A Kellogg P F Stevens M J Donoghue 2007 Plant
Systematics A Phylogenetic Approach 3rd edition Sunderland Massachusetts Sinauer
Associates ISBN 0-87893-407-3 ISBN 978-0-87893-407-2
Katoh K Standley DM 2013 MAFFT Multiple sequence alignment software version 7
improvements in performance and usability Molecular Biology and Evolution 30 772ndash
780
Kivlin SN Emery SM Rudgers JA 2013 Fungal symbionts alter plant responses to global
change American Journal of Botany 100 (7) 1445-1457
Lamichhane J Venturi V 2015 Synergisms between microbial pathogens in plant disease
complexes a growing trend Frontiers in Plant Science 11 de noviembre
103389fpls201500385
Lehtonen S 2020 Phenotypic characters of static homology increase phylogenetic stability
under direct optimization of otherwise dynamic homology characters Cladistics
httpsdoiorg101111cla12438
66
Leung G Robson GD Robinson CH 2011 Characterisation of cold-tolerant fungi from a
decomposing High Arctic moss Soil Biology and Biochemistry 43(9)
Leacuteveilleacute JH 1846 Consideacuterations mycologiques suivis drsquoune nouvelle classification des
champignons Paris
Liu YJ Tang Q Fang L 2015 First report of Nigrospora sphaerica causing leaf blight on
Camellia sinensis in China Plant Disease 100 221
Lopera C Avila L Rojas A Franco-Molano AE Cepero MC Restrepo S 2008 Fungal
endophytes from the Colombian Andean Paacuteramo ecosystem inhibit the growth of plant
pathogens in vitro Phytopathology 98 (6) 94
Lopez DC Sword GA 2015 The endophytic fungal entomopathogens Beauveria bassiana and
Purpureocillium lilacinum enhance the growth of cultivated cotton (Gossypium hirsutum)
and negatively affect survival of the cotton bollworm (Helicoverpa zea) Biol Control
8953ndash60 11 de noviembre httpsdoiorg101016jbiocontrol201503010
Lorito M 1998 Genes from mycoparasitic fungi as a source for improving plant resistance to
fungal pathogens Proc Natl Acad Sci USA 95 7860ndash7865
Marengo J A R Jones L Alves y M Valverde 2009 Future change of temperature and
precipitation extremes in South America as derived from the PRECIS regional climate
modeling system Internatiexclonal Journal of Climatology 292241minus2255
Marie-Laure Desprez-Loustau Benoit Marccedilais Louis-Michel Nageleisen Dominique Piou
Andrea Vannini 2006 Interactive effects of drought and pathogens in forest trees
Annals of Forest Science Springer VerlagEDP Sciences 63 (6) pp597-612
Mejia EJ Loveridge ST Stepan G Tsai A Jones GS Barnes T White KN Drasˇkovicacute M
Tenney K Tsiang M Geleziunas R Cihlar T Pagratis N Tian Y Yu H Crews P (2014)
Study of marine natural products including resorcyclic acid lactones from Humiacutecola
fuscoatra that reactivate latent HIV-1 expression in an in vitro model of central memory
CD4 T cells J Nat Prod 77618ndash624
Mendoza S 2010 Aspectos ecoloacutegicos de los patoacutegenos asociados a Espeletia grandiflora en el
Parque Nacional Natural Chingaza Universidad Jorge Tadeo Lozano Bogotaacute Colombia
67
Menzies James amp Ehret David amp Koch Carol amp Bogdanoff Carl 1998 Humicola fuscoatra
infects tomato roots but is not pathogenic European Journal of Plant Pathology 104
769-775 15 de noviembre 101023A1008680018677
Miller J H 1961 A monograph of the world species of Hypoxylon University of Georgia Press
USA 158
Miles LA Lopera C Gonzaacutelez S de Garciacutea MCC Franco AE Restrepo S 2012 Exploring
the biocontrol potential of fungal endophytes from an Andean Colombian Paacuteramo
ecosystem BioControl 57 (5) 697-710 11 de noviembre
httpsdoiorg101007s10526-012-9442-6
MINAMBIENTE 2019 Ecosistemas de Alta Montantildea y Paacuteramo Obtenido de Ecosistemas de
Alta Montantildea y Paacuteramo httpwwwminambientegovcoindexphpbosques-
biodiversidad-yservicios-ecosistematicosecosistemas-estrategicosparamos
Mohali S Slippers B and Wingfield MJ 2007 Identification of Botryosphaeriaceae from
Eucalyptus Acacia and Pinus in Venezuela Fungal Diversity 25 103-125
Molinillo m Y m Monasterio 2001 Uso del espacio en sistemas pastorales andinos una
comparacioacuten mediante sistemas de informacioacuten geograacutefica y evaluacioacuten multicriterio IV
Simposio Internacional de Desarrollo Sustentable en Los Andes
Morales-Rivas M Otero Garcia J Hammen T V D Torres Perdigoacuten A Cadena Vargas C
E Pedraza Pentildealoza C A amp Posada Gilede E 2019 Atlas de paacuteramos de Colombia
Moreno C D y Palma J D 2016 Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-
Barragaacuten A Los Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Obtenido de Vulnerabilidad Ecoloacutegica Del Complejo De Paacuteramos Chiliacute-Barragaacuten A Los
Incrementos De Temperatura En Un Escenario De Cambio Climaacutetico
Moreno G Manjon JL y J Aacutelvarez-Jimenez 2013 Los hongos y el cambio climaacutetico En
KAGAN-ZUR V SITRIT Y ROTH-BEJERANO NA MORTE (eds) Desert
truffle Springer Verlag Berliacuten 129-135
Murillo A Munkvold G 2008 Systemic Infection by Fusarium verticillioides in Maize Plants
Grown Under Three Temperature Regimes Plant Disease 92(12)1695-1700 15 de
noviembre 101094phi-t-2001-0517-01
68
Naik BS Shashikala J Krishnamurthy Y 2009 Study on the diversity of endophytic
communities from rice (Oryza sativa L) and their antagonistic activities in vitro
Microbiol
Nakashima K Tran L-S P Van Nguyen D Fujita M Maruyama K Todaka
DYamaguchi- Shinozaki K 2007 Functional analysis of a NAC-type transcription
factor OsNAC6 involved in abiotic and biotic stress-responsive gene expression in rice
The Plant Journal 51(4) 617-630 15 de noviembre 101111j1365-313X200703168
Nicola Segata Daniela Boernigen Timothy L Tickle Xochitl C Morgan Wendy S Garrett
Curtis Huttenhower 2013 ldquoComputational metarsquoomics for microbial community
studiesrdquo Molecular Systems Biology 9 (666) 666 11 de noviembre
101038msb201322
Nonsoque Y 2012 Comparacioacuten de hongos endoacutefitos en individuos sanos y enfermos de
Espeletia grandiflora (microcuenca de la quebrada Calostros ndash Parque Nacional Natural
Chingaza) Pontificia Universidad Javeriana 26
Oh H Shearer C A Gloer J B J 1999 Nat Prod 62 497-501
Olave L F C 2011 Characterization of Cellulases and Antagonistic Activities in Fungal
Endophytes Isolated from Espeletia Spp (Doctoral dissertation Uniandes)
Osorio JA Crous CJ de Beer ZW Roux J 2017 Endophytic Botryosphaeriaceae including five
new species associated with mangrove trees in South Africa Fungal Biol 121 361ndash393
Otero J T y Bayman P 2009 Germinacioacuten simbioacutetica y asimbioacutetica en semillas de orquiacutedeas
epiacutefitas Acta Agronoacutemica 58 (4) 270-276
Parsa S Garciacutea-Lemos AM Castillo K Ortiz V Loacutepez-Lavalle LAB Braun J Vega FE 2016
umlFungal endophytes in germinated seeds of the common bean Phaseolus vulgarisuml
Fungal biology 120(5) 783-790
Pedraza-Pentildealosa P Betancur J y Franco-Rosselli P 2005 Chisacaacute un recorrido por los
paacuteramos andinos Segunda edicioacuten Instituto de ciencias Naturales e Instituto de
Investigacioacuten de recursos Bioloacutegicos Alexander von Humbolt Bogotaacute Colombia
Petrini L E and Petrini O 1985 Xylariaceous fungi as endophytes Sydowia 38 216-234
69
Pontificia Universidad Javeriana Universidad Jorge Tadeo Lozano Universidad de la Amazoniacutea
Parques Nacionales Naturales Instituto Alexander von Humboldt Sociedad Colombiana
de Entomologiacutea 2009 Programa Nacional para la Evaluacioacuten del Estado y Afectacioacuten de
los Frailejones en los Paacuteramos de Andes del Norte Bogotaacute Colombia 1-9
Prieto M 2017 Lesiones de frailejones (Espeletia sp) y potenciales hongos fitopatoacutegenos
asociados en los paacuteramos de Chingaza y Cruz Verde Pontificia Universidad Javeriana
Raeder U and Broda P 1985 Rapid preparation of DNA from filamentous fungi Letters in
Applied Microbiology 1 17-20 httpsdoiorg101111j1472-765X1985tb01479x
Raj Mithun Hegde Vinayaka Jeeva M L Senthil M Nath Vishnu S Vidyadharan Pravi
Archana P V 2013 Molecular diagnosis of Colletotrichum gloeosporioides causing
AnthracnoseDieback disease in Greater Yam (Dioscorea alata L) Archives of
Phytopathology and Plant Protection 46 (8) 927ndash936 15 de noviembre
101080032354082012755336
mdashmdashmdash 2002 Colombia diversidad bioacutetica III La regioacuten de vida paramuna Bogotaacute
Universidad Nacional de Colombia
Reacuteblovaacute M Untereiner WA Reacuteblovaacute K (2013) Novel Evolutionary Lineages Revealed in the
Chaetothyriales (Fungi) Based on Multigene Phylogenetic Analyses and Comparison of
ITS Secondary Structure PLOS ONE 8(5) e63547 15 de noviembre
httpsdoiorg101371journalpone0063547
Rivera OD Torres RD 2001 Paacuteramos de Colombia Bogotaacute DC Banco de Occidente 233
Riveros A 2010 Induccioacuten de resistencia en plantas Interaccioacuten Planta-Patoacutegeno Instituto
interamericano de cooperacioacuten para la Agricultura (IICA) 155p
Rossman AY Adams GC Cannon PF Castlebury LA Crous PW Gryzenhout M Jaklitsch WM
Mejia LCStoykov D Udayanga D 2015 Recommendations of generic names in
Diaporthales competing for protection or use IMA Fungus 6 145-154
Ruiz-Peacuterez CA Restrepo S Zambrano MM 2016 Microbial and functional diversity within the
phyllosphere of Espeletia species in an Andean high-mountain ecosystem Appl Environ
Microbiol 82 1807ndash1817 12 de noviembre 101128AEM02781-15
70
Ruiz D M P Arroyave A M Molina J F Barros M E Gutieacuterrez y P A Zapata 2009
Signals of climate variabilitychange in surface water supply of highminusmountain
watersheds minus case study Claro River high mountain basin Los Nevados Natural Park
Andean Central Mountain Range Colombia World Bank Group 207 pp
Salgado-Salazar C Caridad Cepero 2005 umlEndophytic fungi in rose (Rosa hybrida) in Bogotaacute
Colombiauml Revista iberoamericana de micologiacutea 22 99-101
Saacutenchez R Saacutenchez B Monserrat Y Ulloa A Armendaacuteriz B Garciacutea M Maciacuteas M
2013 Hongos endoacutefitos fuente potencial de metabolitos secundarios bioactivos con
utilidad en agricultura y medicina Revista Especializada en Ciencias Quiacutemico-
Bioloacutegicas 16 (2)132-146 15 de noviembre 101016s1405-888x(13)72084-9
Siegel MR GCM Latch y MC Johnson 1987 Fungal endophytes of grasses Ann Rev
Phytopath 25 293-315
Silva-Valenzuela M R I Rojas-Martiacutenez R H Manzanilla-Loacutepez M L Maciacuteas-Ruvalcaba
S ArandaOcampo y E Zavaleta-Mejiacutea 2020 Hongos endoacutefitos Una alternativa
bioloacutegica para el manejo de nematodos fitoparaacutesitos Nematropica 50101-117
Silva S Gusmatildeo LF Castantildeeda RF 2015 Cryptocoryneum parvulum a new species on
Araucaria angustifolia (Brazilian pine) Mycotaxon Volume 130 (2) April-June 2015
pp 465-469(5) 15 de noviembre httpsdoiorg105248130465
Schmidt HA von Haeseler A 2009 Phylogenetic inference using maacuteximum likelihood methods
En The phylogenetic handbook a practical approach to phylogenetic anaysis and
hypothesis testing Lemey P Slemi M Vandamme AM (eds) Published by Cambridge
Univrsity Press
Schultz Carl 1856 Espeletia hartwegiana Hugh Algernon Weddell (ed) Chloris Andina 1
62
Schulz B Roumlmmert A Dammann U Aust H Strack D 1999 The endophyte-host interaction a
balanced antagonism Mycological Research 103 (10)1275-1283 15 de noviembre
101017s0953756299008540
Sharma P Meena PD Chauhan JS 2013 First report of Nigrospora oryzae (Berk amp Broome)
Petch causing stem blight on Brassica juncea in India Journal of Phytopathology 161
439ndash441
71
Swofford D L 1997 Phylogenetic analysis using parsimony Sinauer Associates Sunderland
Massachusetts USA
Swofford D L 2003 Phylogenetic Analysis Using Parsimony (and Other Methods) Sinauer
Associates Sunderland Massachusetts
Terhonen E Sipari N Asiegbu FO 2016 Inhibition of phytopathogens by fungal root
endophytes of Norway spruce Biol Control 99 53ndash63
httpsdoiorg101016jbiocontrol201604006
Thomas T Gilbert J Meyer F 2012 laquoMetagenomics - a guide from sampling to data
analysisraquo Microbial Informatics and Experimentation 2 (1) 3 15 de noviembre
1011862042-5783-2-3
Thuiller Wilfried 2007 Biodiversity climate change and the ecologist Nature 448 550-552
12 de noviembre 101038448550a
Uzor PF Ebrahim W Osadebe PO 2015 Metabolites from Combretum dolichopetalum and its
associated endophytic fungus Nigrospora oryzae ndashevidence for a metabolic partnership
Fitoterapia 105 147ndash150
Varela A 2015 ldquoAnaacutelisis de la relacioacuten hongos patoacutegenos ndash afectacioacuten de frailejones en el
Parque Nacional Natural Chingazardquo Informe final actividades proyecto Bogotaacute DC
Colombia Pontificia Universidad Javeriana
mdashmdashmdash 2016 Aunar esfuerzos teacutecnicos y cientiacuteficos para desarrollar una investigacioacuten
participativa sobre el efecto de la transformacioacuten antroacutepica del paacuteramo en las
interacciones planta- insecto-hongo en los E argentea y E grandiflora)
Vaacutesquez Ceroacuten A Buitrago Castro A C 2017 El gran libro de los paacuteramos Instituto de
Investigacioacuten de Recursos Bioloacutegicos Alexander von Humboldt
Vega FE Simpkins A Aime CM Posada F Peterson SW Rehner SA Infante F Castillo A
Arnold E 2010 umlFungal endophyte diversity in coffee plants from Colombia Hawairsquoi
Mexico and Puerto Ricouml Fungal Ecology 3 (3) 122-138
72
Wang M y Liu Fang y Crous Pedro y Cai Lei 2017 Phylogenetic reassessment of Nigrospora
Ubiquitous endophytes plant and human pathogens Persoonia - Molecular Phylogeny
and Evolution of Fungi 39 11 de noviembre 103767persoonia20173906
Webster J 1952 Spore projection in the hyphomycete Nigrospora sphaerica New Phytologist
51 229ndash235
Wood S N 2001 umlmgcv GAMs and generalized ridge regression for R R newsuml 1 (2) 20-25
Wu PC Tsai JC Li FC 2004 Increased levels of ambient fungal spores in Taiwan are associated
with dust events from China Atmospheric Environment 38 4879ndash4886
Yan L Zhu J Zhao X Shi J Jiang C Shao D 2019 Beneficial effects of endophytic fungi
colonization on plants Appl Microbiol Biotechnol 103 (8) 3327-3340 12 de
noviembre 101007s00253-019-09713-2
Zimand G Elad Y amp Chet I 1996 Effect of Trichoderma harzianum on Botrytis cinerea
pathogenicity Phytopathology 86 1255ndash1260
Zhang T Yao YF 2015 Endophytic Fungal Communities Associated with Vascular Plants in
the High Arctic Zone Are Highly Diverse and Host-Plant Specific Plos One 10 (6)
73
Anexos
Anexo 1 Tesista Cristhian Aristizabal junto al Frailejoacuten Espeletia hartwegiana Paacuteramo
Romerales Quindiacuteo Colombia
Anexo 2 Tesista Cristhian Aristizabal Colectando Hojas del Frailejoacuten Espeletia
hartwegiana Paacuteramo Romerales Quindiacuteo Colombia
74
Anexo 3 Coordenadas geograacuteficas tomadas de cada punto de colecta de las hojas del frailejoacuten
Espeletia hartwegiana ubicadas en el paacuteramo de romerales Quindiacuteo Colombia
Coordenadas Geograacuteficas
(Latitud)
De la Colecta del
Material Bioloacutegico total de 15
muestras (Longitud)
LAT N 4deg 37 5871717 LONG W 75deg 25
3292611
LAT N 4deg 37 5885220 LONG W 75deg 25
3271217
LAT N 4deg 37 5884485 LONG W 75deg 25
3312103
LAT N 4deg 36 5995487 LONG W 75deg 27
3045854
LAT N 4deg 37 6195784 LONG W 75deg 30
3292611
LAT N 4deg 40 159171 LONG W 75deg 26
380142
LAT N 4deg 40 363762 LONG W 75deg 26
209072
LAT N 4deg 40 399832 LONG W 75deg 27
173554
LAT N 4deg 40 383965 LONG W 75deg 27
464325
LAT N 4deg 39 486438 LONG W 75deg 27
173559
LAT N 4deg 40 56894 LONG W 75deg 26
568287
LAT N 4deg 40 159174 LONG W 75deg 27
190654
LAT N 4deg 40 125083 LONG W 75deg 26
345912
LAT N 4deg 40 116691 LONG W 75deg 27
208264
LAT N 4deg 40 314698 LONG W 75deg 27
140791
75
Anexo 4 Hoja del Frailejoacuten Espeletia hartwegiana
Anexo 5 Hifas observadas a traveacutes de un estereoscopio con el propoacutesito de seleccionar la
maacutes indicada y luego sembrar un cultivo puro a partir de ese micelio
76
Anexo 6 Backups o copias de cultivos axeacutenicos de los hongos endoacutefitos encontrados las
copias se conservaron con glicerol a -80degC en un ultra congelador de Thermo Scientifictrade
Anexo 7 Caja de laboratorio con las muestras de ADN extraiacutedas listas para ser empacadas
y enviadas
77
Anexo 8 Campana de flujo laminar umlBIOBASEuml en donde se trabajoacute con los cultivos
microbioloacutegicos de forma segura y esteacuteril propiedad de la Universidad del Quindiacuteo
Anexo 9 Muestras de ADN extraiacutedos de los Hongos endoacutefitos encontrados en las hojas de
frailejoacuten enviadas a la Universidad Ruhr de Bochum (Alemania)
78
Anexo 10 Estadiacutesticos resultantes del programa PAUP producto del aacuterbol de parsimonia
Anexo 11 Alineamiento de las secuencias ITS en el programa MEGA-X en donde se
observa en amarillo la secuencia umlTCATTAuml la cual es una secuencia altamente conservada
que se usa para marcar el inicio de la misma en hongos endoacutefitos
79
Anexo 12 Estadiacutesticos generados por el programa JmoldelTest usados para generar el
aacuterbol de relaciones filogeneacuteticas a partir de la regioacuten ITS de los hongos endoacutefitos aislados
total de modelos evaluados 88
ID Nombre Particioacuten lnL P AIC detaAIC Peso Cumpeso Udelta
1 JC 0 100820157 138 204400314 1341348 0 1 -
2 JC+I 0 98912511 139 200605022 9618188 0 1 -
3 JC+G 0 95470113 139 193720225 2733392 0 1 -
4 JC+I+G 0 95467507 140 193735014 274818 0 1 -
5 F81 0 100658095 141 204136189 13149356 0 1 -
6 F81+I 0 98761481 142 200362962 9376128 0 1 -
7 F81+G 0 95314814 142 193469628 2482794 0 1 -
8 F81+I+G 0 95324364 143 193508729 2521895 0 1 -
9 K80 10010 99734495 139 20224899 11262157 0 1 -
10 K80+I 10010 97835191 140 198470382 7483549 0 1 -
11 K80+G 10010 94227821 140 191255643 268809 0 1 -
12 K80+I+G 10010 94209963 141 191239926 253092 0 1 -
13 HKY 10010 99648091 142 202136181 11149348 0 1 -
14 HKY+I 10010 97761512 143 198383024 7396191 0 1 -
15 HKY+G 10010 94170799 143 191201599 214765 0 1 -
16 HKY+I+G 10010 94118678 144 191117357 130523 600E-04 0999 -
17 TrNef 10020 99564653 140 201929306 10942472 0 1 -
18 TrNef+I 10020 97712623 141 198245245 7258412 0 1 -
19 TrNef+G 10020 94150977 141 191121955 135121 400E-04 09994 -
20 TrNef+I+G 10020 94132131 142 191104263 117429 00011 09959 -
21 TrN 10020 99563809 143 201987618 11000784 0 1 -
22 TrN+I 10020 97702964 144 198285929 7299095 0 1 -
23 TrN+G 10020 94094235 144 191068469 81635 00065 09719 -
24 TrN+I+G 10020 94101378 145 191102757 115923 00012 09948 -
25 TPM1 12210 99713879 140 202227759 11240925 0 1 -
26 TPM1+I 12210 97826641 141 198473282 7486449 0 1 -
27 TPM1+G 12210 94249837 141 191319674 33284 0 1 -
28 TPM1+I+G 12210 942022 142 1912444 257566 0 1 -
29 TPM1uf 12210 99663473 143 202186945 11200112 0 1 -
30 TPM1uf+I 12210 97879458 144 198638917 7652083 0 1 -
31 TPM1uf+G 12210 94137349 144 191154699 167865 100E-04 09999 -
32 TPM1uf+I+G 12210 94141897 145 191183794 196961 0 1 -
80
33 TPM2 10212 99722544 140 202245087 11258254 0 1 -
34 TPM2+I 10212 97828213 141 198476426 7489592 0 1 -
35 TPM2+G 10212 94185859 141 191191718 204885 0 1 -
36 TPM2+I+G 10212 94153667 142 191147335 160501 100E-04 09998 -
37 TPM2uf 10212 99648091 143 202156182 11169348 0 1 -
38 TPM2uf+I 10212 97757018 144 198394035 7407202 0 1 -
39 TPM2uf+G 10212 941052 144 1910904 103566 00022 09904 -
40 TPM2uf+I+G 10212 9408801 145 19107602 89187 00044 0982 -
41 TPM3 12012 99768005 140 20233601 11349176 0 1 -
42 TPM3+I 12012 97958099 141 198736198 7749364 0 1 -
43 TPM3+G 12012 94227963 141 191275926 289093 0 1 -
44 TPM3+I+G 12012 94323885 142 191487769 500936 0 1 -
45 TPM3uf 12012 99625421 143 202110843 11124009 0 1 -
46 TPM3uf+I 12012 97770469 144 198420939 7434105 0 1 -
47 TPM3uf+G 12012 94165576 144 191211152 224319 0 1 -
48 TPM3uf+I+G 12012 94150861 145 191201721 214888 0 1 -
49 TIM1ef 12230 99592239 141 202004478 11017644 0 1 -
50 TIM1ef+I 12230 97822787 142 198485574 749874 0 1 -
51 TIM1ef+G 12230 94121043 142 191082087 95253 00033 09853 -
52 TIM1ef+I+G 12230 94122386 143 191104772 117939 00011 09969 -
53 TIM1 12230 99518653 144 201917306 10930473 0 1 -
54 TIM1+I 12230 9781156 145 19852312 7536286 0 1 -
55 TIM1+G 12230 94105198 145 191110396 123562 800E-04 09977 -
56 TIM1+I+G 12230 94087158 146 191094317 107483 00018 09922 -
57 TIM2ef 10232 99550617 141 201921233 109344 0 1 -
58 TIM2ef+I 10232 97704602 142 198249204 726237 0 1 -
59 TIM2ef+G 10232 94093484 142 191026968 40135 00516 0852 -
60 TIM2ef+I+G 10232 94073864 143 191007729 20895 0135 07169 -
61 TIM2 10232 99530022 144 201940044 1095321 0 1 -
62 TIM2+I 10232 97820158 145 198540315 7553482 0 1 -
63 TIM2+G 10232 94058678 145 191017356 30523 00834 08004 -
64 TIM2+I+G 10232 94060837 146 191041674 54841 00247 09248 -
65 TIM3ef 12032 99546286 141 201912572 10925739 0 1 -
66 TIM3ef+I 12032 97713931 142 198267862 7281028 0 1 -
67 TIM3ef+G 12032 94129376 142 191098753 111919 00014 09936 -
68 TIM3ef+I+G 12032 94112005 143 191084009 97176 0003 09883 -
69 TIM3 12032 99514376 144 201908751 10921918 0 1 -
70 TIM3+I 12032 97700023 145 198300047 7313213 0 1 -
71 TIM3+G 12032 94115323 145 191130646 143812 300E-04 09997 -
72 TIM3+I+G 12032 94096941 146 191113882 127048 700E-04 09984 -
73 TVMef 12314 9970418 142 20224836 11261526 0 1 -
74 TVMef+I 12314 97940598 143 198741196 7754363 0 1 -
81
Anexo 13 Estadiacutesticos generados a partir de correr los 88 modelos para los datos en el
programa jModelTest corriendo el criterio de informacioacuten AKAIKE (AIC)
75 TVMef+G 12314 94159829 143 191179657 192824 0 09999 -
76 TVMef+I+G 12314 94160993 144 191201987 215153 0 1 -
77 TVM 12314 99615991 145 202131982 11145148 0 1 -
78 TVM+I 12314 97746676 146 198413353 7426519 0 1 -
79 TVM+G 12314 94075503 146 191071005 84171 00057 09776 -
80 TVM+I+G 12314 94052588 147 191045176 58342 00208 09456 -
81 SYM 12345 99543166 143 201946331 10959498 0 1 -
82 SYM+I 12345 97757103 144 198394207 7407373 0 1 -
83 SYM+G 12345 94083056 144 191046112 59278 00198 09654 -
84 SYM+I+G 12345 94050035 145 19100007 13236 0198 05819 -
85 GTR 12345 99529525 146 20197905 10992217 0 1 -
86 GTR+I 12345 97686178 147 198312356 7325522 0 1 -
87 GTR+G 12345 94044194 147 191028387 41553 00481 09001 -
88 GTR+I+G 12345 94013417 148 190986834 0 03839 03839 -