慰霊の日特集 - Naha...犯罪小説集 吉田 修一 この世界が消えたあとの科学文明 のつくりかた ルイス ダ ートネル モヤモヤするあの人 宮崎

1

2
大腸菌染色体と F プラスミドの分配機構の研究
平賀 壯太
はじめに
真核細胞の染色体が複製後に微小管でできた紡錘体の働きによって娘細胞に分配
されることは古くから知られていた。しかし、原核細胞では微小管は発見されておらず、
複製後に姉妹染色体 DNA が娘細胞へと分配される機構は不明であった。F. Jacob, S.
Brenner, and F. Cuzin (1963) は彼らの提唱したレプリコン仮説 (replicon theory) の
論文の中で大腸菌の染色体と F プラスミドの分配機構について彼らのモデルを提唱し
た。それは染色体や F プラスミドは細胞の中央の細胞膜に結合しており、そこで複製し
た姉妹 DNA 分子は細胞の中央部における膜の伸長によって左右に徐々に離れて行
くというモデルであった(図1)。しかしリン脂質からできた細胞膜には流動性があること
が明らかになると内膜と外膜の間にあるペプチドグリカン層に DNA が結合していると
彼らは考えた。しかし、大腸菌のペプチド層は細胞の中央部のみで合成伸長するの
ではなく、細胞全体で合成伸長することが分かるとこのモデルも事実と矛盾することと
なった。さらにこのモデルでは細胞分裂で新生細胞ができるのと同時に染色体と F プ
ラスミドが複製を開始しなければならないが、F プラスミドの複製は染色体の複製開始
とは必ずしも一致せず、ランダムに複製することが明らかになり、彼らのモデルでは説
明できなくなった。
このような当時の状況下で、私たちは F プラスミドと大腸菌の染色体の分配機構の解
明を研究課題に選んで研究を開始した。この研究課題にたどり着くまでに私がおこな
った研究についても述べて、それらの研究過程からどのようにして徐々に「分配機構」
が私の生涯を懸ける研究課題として浮かび上がって来たかという経過を紹介したい。
研究の節目節目でおこなう次の研究の進路選択は、その後の研究の発展にとって大
変重要なことであり、選択を誤らないようにしなくてはならない。

3
● トリプトファンオペロンの研究
大阪大学大学院理学研究科の院生のときは吉川秀男教授の遺伝学教室で学んだ。
1965 年の3月に博士課程を終了して理学博士の学位を取得し、4月から京都大学ウイ
ルス研究所の助手になった。当時、ウイルス研究所の化学部では由良隆・今井六雄・
岡本利雄さんたちがトリプトファンオペロンのリプレッサー蛋白質を生化学的手法によ
って精製しようとしていた。由良さんにはこの研究に加わるようにといわれたのだが、私
はトリプトファンオペロンの研究をするのならば遺伝学的方法でやりたいと申し出て研
究を始めた。
まず私が始めたのは、トリプトファンオペロンのプロモーター領域の変異株を分離す
ることだった。院生時代に Journal of Molecular Biology に掲載された F. Jacob and J.
Monod (1961) のラクトースオペロンのオペレーターの変異株を分離した論文を吉川

4
研究室の雑誌会で紹介したことがあった。彼らのオペロン説ではリプレッサーが作用
する領域「オペレーター」がオペロンの近くに存在していることが予想されており、その
予想のもとに実際にそういう変異株を分離した論文であった。私はこの恰好良い研究
のスタイルにすっかり魅了されてしまい、自分も将来このようなスタイルの研究をおこな
いたいと強く考えていた。
一方、由良さんたちがおこなおうとしていた研究のリプレッサーの検出方法は、トリプ
トファンオペロンの DNA からメッセンジャーRNA を in vitro で合成する RNA ポリメラー
ゼの系に大腸菌の細胞抽出液の分画を加えてメッセンジャーRNA 合成が阻害される
か否かという方法だった。当時、トリプトファンオペロンの DNA を大量に分離することは、
このオペロンを運ぶ80ファージが見つかっていたので可能であった。しかし、トリプト
ファンオペロンのリプレッサーの遺伝子 trpR はトリプトファンオペロンとは異なる染色体
座位に存在しており、TrpR 蛋白質を大腸菌に大量に産生させる方法はなかった。本
来細胞当たりのリプレッサー分子の数は尐ないと推測されるので、私にはこの研究が
成功するとはどうしても思われなかったのである。
大腸菌の細胞内でコリスミ酸とグルタミンからトリプトファンが合成される酵素反忚の
過程には5つの蛋白質が関与しており、それらの蛋白質をコードする5つの遺伝子
trpE・trpD・trpC・trpB・trpA によってトリプトファンオペロンが構成される(図2A)。培地
中にトリプトファンが存在するときには、TrpR リプレッサーによってこれらの遺伝子の発
現は抑制されているが、培地中のトリプトファンが或る濃度以下になるとその抑制が解
除されてトリプトファンオペロンの蛋白質群が産生され、その結果、細胞内でトリプトフ
ァンが効率良く合成されることになる。すなわち、トリプトファンはTrpRリプレッサーを活
性化する働きがある。
私はトリプトファンオペロンのオペレーターの変異株を分離するために、野生型株が
増殖できない高濃度の5-メチルトリプトファンに耐性の変異株を分離することから始め
た。5-メチルトリプトファンはトリプトファンの誘導体で蛋白質合成の阻害剤であり、か
つトリプトファンオペロンの発現を抑制する働きがある。オペレーターの変異株では、
培地に5-メチルトリプトファンが存在しても常にオペロンの蛋白質群が産生されて細胞
内でトリプトファン合成が盛んにおこなわれるため、細胞内のトリプトファン濃度が増加
して5-メチルトリプトファンによる蛋白合成阻害効果が減尐して、細胞は増殖して5-メ
チルトリプトファン耐性を示すはずである。

5
このようにして分離した多数の5-メチルトリプトファン耐性株について、変異遺伝子の
染色体上の座位を種々の Hfr 株との接合や P1 ファージを使った形質導入によって解
析した。一群の耐性株では、変異はトリプトファンオペロンの直ぐ上流に存在しており
オペレーター領域(trpO)の変異であることがわかった。これらの trpO の変異株は全て
培地にトリプトファンが存在している条件でもトリプトファンオペロンの蛋白質群を恒常
的に産生する性質を持っていた。これらの trpO の変異は cis の位置にあるトリプトプァ
ンオペロンを構成的(constitutive)に発現させるが、trans の位置にあるトリプトファンオ
ペロンを構成的に発現させることはなかった。第2のグループに属する5-メチルトリプト
ファン耐性変異株は trpR 遺伝子の変異であった。第3のグループに属する5-メチルト
リプトファン耐性変異株は argH に近接した染色体の位置にあり、トリプトファンオペロン

6
の発現調節は正常だった。これは新し い遺伝子であり mtr(methyltryptophan
resistant)遺伝子と名付けた。このトリプトファンオペロンのオペレーター領域の証明は、
Jacob らがおこなったラクトースオペロンのオペレーターの発見に継ぐ2番目の発見だ
った。さらに、私は他の変異株分離法によって, 新しいタイプのトリプトファンオペロン
の調節変異株を分離した。この変異は Hfr 接合実験から、str 遺伝子の近くに存在して
いることが明らかになり trpS と名づけた。そして院生の伊藤維昭君との共同研究で、こ
の変異がトリプトファンを tRNA に結合するトリプトファン-tRNA 合成酵素の遺伝子の変
異であることがわかった。
1970 年2月からアメリカ心臓協会から奨学金を受けてアメリカに留学することになっ
た。留学先はカリフォルニア州スタンフォード大学のトリプトファンオペロン研究の大御
所 Charles Yanofsky 博士の研究室である。妻と2才の娘と6ケ月の息子を連れての留
学だった。ユーカリの並木のある芝生の広大な敷地内にあったスタンフォード大学の
Yanofsky 博士の研究室を初めて訪問し博士に面会したときに、私は3つほど自分で考
えた研究プランを話し、Yanofsky 博士からは2つの研究テーマの提案があった。私の
案の方がより具体的で成功の可能性が大きいことに博士も同意したのでさっそくその
研究を始めた。当時日本では大学紛争の最中で落ち着いて研究ができる状態ではな
く相当嫌気がさしていたが、新天地に移り張り切って毎日研究に没頭することができ
た。
先ず手始めにやったことは、トリプトファンオペロンの一番目の遺伝子である trpE 遺
伝子だけを欠失した変異株を分離することである。trpE 遺伝子内のプロモーターに近
い位置のナンセンス変異株では、trpDCBA の発現が極端に低下する現象があった。
この現象はポーラー効果といって、このような変異株をポーラー変異株ともいう(図2A)。
このようなポーラー変異株はアントラニル酸が存在する最小培地では増殖できるが、ア
ントラニル酸と5-メチルトリプトファンが存在した培地では増殖できない性質がある。し
たがって、私の考えた trpE 欠失変異株の分離方法は、このようなポーラー変異株か
らアントラニル酸の存在下で 5-メチルトリプトファン耐性株を見つけることである。すな
わち、このポーラー変異の領域を含む欠失変異が起ったため、trpDCBA の発現が回
復してアントラニル酸から効率良くトリプトファンが合成されるはずである。私はこのよう
な変異株を多数分離して詳細な遺伝解析をした結果、tepE5欠失変異はそれまでに
分離されていた trpE 遺伝子の全ての点変異の DNA 領域を欠失していた。しかし、他
の遺伝子 trpDCBA は正常であった(図2B)。

7
当時トリプトファンオペロンの種々の DNA 断片を持つ80や80ファージが分離さ
れていたのでこれらのファージ DNA を使って DNA-RNA ハイブリッド法で野生型株や
欠失変異株から抽出した放射線同位元素の H3-ウリジンで標識したメッセンジャー
RNA を検出し定量することが可能だった。この方法で解析した結果、このtrpE5欠失
変異株はトリプトファンオペロンの転写調節は正常であったので、オペレーター/プロ
モーター領域は正常であることが確認された。すなわち、目的とする trpE 遺伝子だけ
を欠失した変異株であった。
増殖中の野生型株にリファンピシンを添加して RNA ポリメラーゼの RNA 合成開始能
を阻害したあと、経時的に H3-ウリジンで短時間標識した後、細胞から RNAを抽出して
DNA-RNA ハイブリッド法で解析すると、オペロン内の遺伝子の配列順に mRNA の合
成が阻害されて行くのが観察できた。リファンピシンと同時にトリプトファンも添加すると
trpE の mRNA 合成が直ちに著しく阻害され、そして幾分弱いながら trpD の mRNA に
もその阻害効果がみられた。このトリプトファンによる阻害効果は trpR 変異株でも見ら
れた。またtrpE5欠失変異株では trpD の mRNA 合成が直ちにトリプトファンによる著
しい阻害効果を受けた。これら結果は、この現象が TrpR 蛋白質や TrpE 蛋白質とは無
関係な未知の機構によるものであることを示している。
H3-ウリジンで短時間標識した後、リファンピシンで RNA 合成を阻害する実験によっ
て、野生型株のトリプトファンオペロンのメッセンジャーRNA は 175 秒の半減期で崩壊
することがわかった。一方、trpE 遺伝子の最もプロモーターに近い所にあるナンセンス
変異株(ポーラー変異)を H3-ウリジンで短時間標識した後、trpE 遺伝子に対忚する
mRNA の半減期を測定すると、著しくその半減期は短いことが分かった。すなわち、ナ
ンセンスコドンより後の領域の trpE の mRNA は異常に壊れ易いことを示していた
(1972)。またオペレーター/プロモーター領域からも RNA が作られていることが示唆
された。その後、この研究は Yanofsky 博士の研究室ではトリプトファンオペロンの発現
調節におけるアテニュエーター機構の発見へと発展した。この機構ではオペレーター
/プロモーター領域から作られた短い RNA がオペロンの発現調節に重要な働きをし
ているのである。

8
● チミジンキナーゼ遺伝子の発見
実は私はウイルス研究所の助手になったときトリプトファンオペロンの研究と併行して
他の研究も始めていた。大学院に進学した五十嵐貢一君が私たちの研究室にきて、
私が彼を指導することになったとき、私の提案で一緒に研究を始めたのは大腸菌の高
温感受性変異株の分離であった。それは増殖に必須な蛋白質の遺伝子の変異を解
析することを目標にした研究であり、まず初めに栄養豊富な培地において 30°C では
増殖できるが、42°C では増殖できない変異株を多数分離した。分離した変異株を大
まかに分類するために、42°C で保温したとき細胞の形態がどのように変化するかとい
うことと、蛋白質や DNA の合成能がどのように変化するかということを調べた。DNA 合
成能は放射同位元素 C14-チミジンの酸不溶性分画への取り込みで調べた。最初に変
異株10株を調べるようにといっておいたのだが、五十嵐君が持ってきたのは9株の実
験結果であった。それで、もう1株はどうしたのか訊ねると、「C14-チミジンを入れ忘れま
した」とのことで、もう一度実験をやり直してもらったところ、その変異株は、C14-チミジン
を 30°C でも 42°C でも全く DNA に取り込まない奇妙な変異株であることが明らかに
なった。そして、その変異株の細胞抽出液には、チミジンに ATP のリン酸基を結合して
デオキシチミジン1リン酸にするチミジンキナーゼの酵素活性がないことを明らかにし
た(図 3)。

9
この変異はトリプトファンオペロンに近接して存在していることが分かり、この遺伝子を
tdk (thymidinekinase)と名付けた。tdk 遺伝子を運ぶ 80 ファージも分離することがで
きた。チミジンキナーゼは菌の増殖には必須ではないから、この変異株はチミジンキナ
ーゼ遺伝子の変異と共に未知の必須遺伝子に高温感受性変異を持っている2重変
異株だと考えられる。
● F プラスミドの接合伝達
アメリカ留学中に学んだことは、相当オリジナリティーの高い研究をしないと世界的に
はなかなか認められないということであった。私は2年間のアメリカ留学を終えて帰国し
たとき、新しい分野の研究をしたいと考えていた。世界でまだ誰もやっていない研究分
野を自分で開拓したいといろいろ考えた末、留学前に分離したチミジンキナーゼの変
異株を使った或る実験を思いついた。
大腸菌の性を決定する F プラスミド(性決定因子)は他の細胞へ接合伝達する性質
がある。F プラスミドを持つ F+細胞と持たない F-細胞を接合させると、F+細胞から F プ
ラスミドが F−細胞に伝達され、その中で複製してその F-細胞が F+細胞になる。接合伝
達の際 F プラスミド上の特異的な 1 点で片側の DNA 鎖が切断されてその1本鎖のみ
が F-細胞に入って行き、そこで相補鎖が合成されて再び環状 2 本鎖 DNA となり複製
を始めることが分かっていた。接合伝達のとき F+細胞の中では環状1本鎖 DNA を鋳
型にしたローリングサークル DNA 合成によって環状2本鎖 DNA が作られる。したがっ
て接合後、両細胞ともに F プラスミドを持つ F+細胞になる。
一方、F プラスミドが染色体に組込まれている Hfr 細胞に F+細胞を接合させたときに
は、Hfr 細胞には F プラスミドの伝達が起らないことが知られていた。この現象は、F プ
ラスミド DNA がそもそも Hfr 細胞に入っていかないためなのか、あるいは Hfr 細胞に入
るがその後何らかのメカニズムで複製されないためなのか明らかではなかった。この現
象を解析するために私は次の実験をおこなった。
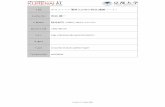
野生型の F+細胞とチミジンキナーゼを欠失した tdk 変異を持つ Hfr 細胞を H3-チミ
ジンを加えた培地中で接合させる。この Hfr 細胞は tdk 変異の他に T6 ファージに対す
る耐性変異と DNA 組み換え能を喪失した recA 変異を持っていた(図4)。野生型の F+細胞の染色体 DNA と F プラスミド DNA は H3-チミジンで標識されるが、tdk 変異を
持つ Hfr 細胞の DNA は標識されない。もし F+細胞内で標識された F プラスミド DNA
10
がこの Hfr細胞に伝達されたならば、Hfr 細胞からその標識された F プラスミドが検出さ
れるはずである。この接合は雄株同士の接合であるが、受容細胞にしたい方の雄株を
一晩栄養豊富な培地中で強く震盪させながら培養すると、受容細胞として働くようにな
る。接合後、細胞を遠心分離機で集めて、それに1細胞あたり 10 個以上の T6 ファー
ジを感染させて T6 ファージ感受性の F+細胞を全部溶かしてしまい(多数のファージの
吸着によって細胞壁が溶けて溶菌する現象)、残った T6 ファージ耐性の Hfr 細胞から
DNA を抽出してプラスミド DNA を蔗糖密度勾配遠心法で調べると、標識された環状 2
本鎖のプラスミド DNA が検出された。対照実験としては、接合させていなかった両株
の細胞を T6 ファージで処理した場合には標識された環状 2 本鎖のプラスミド DNA は
検出されなかった。これらの実験結果は、F+細胞から Hfr 細胞に F プラスミド DNA は
伝達されて環状2本鎖 DNA になるが、その後の複製が何らかのメカニズムで阻害され
ていることを示している。この実験は自分たちの分離した tdk 変異株を上手く利用して
おこなった実験であった。

11
● 染色体の複製開始に関与する RNA合成
アメリカ留学から帰った当時、大腸菌の染色体 DNA の複製開始のメカニズムはまだ
不明であり、私はその機構の解明に興味を持っていた。染色体 DNA の複製開始に関
与する遺伝子は数個知られていたが、それらがコードする蛋白質の機能については
全く分かっていなかったのである。大腸菌の染色体の複製開始には RNA 合成が関与
するということが、K. G. Lark (1972) によって報告されたので、まず私はその実験を院
生の斉藤綱男君と追試してみた。複製開始に関与する DnaA 蛋白質の高温感受性変
異株を使い、30°C で増殖中の細胞を 42°C で保温すると複製開始が阻害され、複
製中だった複製フォークはいずれ複製を終了する。この細胞を再び 30°C に戻すと
複製が同調して開始される。30°C に戻すときに蛋白合成の阻害剤であるクロラムフ
ェニコールを添加しておくと、1回だけ複製が開始される。このことは 42°C では不活
性だった DnaA 蛋白質が 30°C で再び活性化されて複製が開始するためである。
一方、30°C に戻すときにクロラムフェニコールと共にリファンピシンを加えて蛋白合
成と RNA 合成の両方を阻害すると複製は起らなかった(図5A)。このことから、複製開
始には DnaA 蛋白質の他に RNA 合成が必要であると結論され、Lark の実験結果の追
試ができた。
私たちは同じような実験を複製開始に必須な DnaC 蛋白質の高温感受性変異株を
使っておこなってみた。ところが、この株では、クロラムフェニコールと共にリファンピシ
ンを加えた場合にも1回複製が起った(図5B)。この現象を説明するために私たちは、
複製開始の過程を幾つかのステップに分けて考えて、DnaA が働くステップは RNA 合
成を必要とするステップより前であり、DnaC が働くステップは RNA 合成を必要とするス
テップより後であるという仮説をたてた (1974)。
しかし、こういうキネテックスによる実験だけでは蛋白質の機能を知ることは不可能な
ので、もっと生化学的な研究方法を模索しなければならないと考えていた。

12
● 染色体複製起点を持つプラスミドの分離
当時、DNA 複製機構についての研究は、Arthur Kornberg 博士らによって複製フォ
ークにおいて1本鎖 DNA を鋳型にして相補鎖が合成されるメカニズムは in vitro の酵
素反忚系によってほぼ解明されていた。すなわち、複製フォークの部分で岡崎フラグ
メントが合成されるメカニズムは1本鎖 DNA ファージの DNA を鋳型にした研究で解明
されていたのである。しかし当時、環状2本鎖の染色体 DNA が複製を開始するメカニ
ズムは不明であった。大腸菌の約 4700 キロベースの長い染色体 DNA が複製を開始
するのに必修な複製起点は染色体上に1ケ所しかないと推定されていたが、その染色
体起点の正確な位置は不明であった。大腸菌の細胞からフェノール法で DNA を抽出
すると染色体DNAは数百のDNA 断片に切断されてしまうので、複製起点から複製が
開始されるメカニズムを生化学的に研究するためには、何らかの方法で複製起点を含
んだ短い DNA 断片を大量に分離し、それを鋳型にして研究することが必須であると私
は考えていた。

13
そのために、私は染色体の複製起点を持つプラスミドが本来の染色体と共存してい
る大腸菌株を分離することはできないかと考えた。当時、自然界より種々のプラスミドを
持つ大腸菌株が分離されており、同じ複製システムを持ち2種類のプラスミドは非選択
培地の中では1つの細胞内に共存できないという不和合性 (incompatibility) の現象
が知られていたので、大腸菌の細胞の中で染色体の複製起点を持つプラスミドが本
来の染色体と共存できるなどと考える人はいなかったのである。
しかし、私はこのようなプラスミドが存在することを確信していた。そう考える根拠は、
大腸菌では染色体 DNA のマルチフォーク複製という現象が知られていたからである。
染色体が複製起点から2方向に開始した複製フォークは終了するまでに 37°C では
培地の条件には関係なく 40 分かかることが知られていた。栄養豊富な培地中では大
腸菌は 20 分ごとに分裂増殖を繰り返すので、このような条件下では 20 分ごとに複製
の開始が起こる。その結果、複製が終了する前に次々と新たに複製が開始して、染色
体 DNA は多数の複製フォークを持つことになる。この現象から推測されることは、尐な
くとも栄養豊富の条件下では複製に必要な蛋白質や基質は細胞内に十分存在して
おり、染色体の複製起点を持つプラスミドも本来の染色体と共に同一細胞内で複製が
可能だということである(図6)。
このことを証明するために、大腸菌染色体の複製起点を持つプラスミドが存在する菌
株を分離するにはどのようにしたら良いであろうか。先に述べた実験から F プラスミドは

14
接合で Hfr 細胞に入っても複製できないということが分かっていたが、もし F プラスミド
が染色体の複製起点を持っていたならば Hfr 細胞の中でもその染色体複製起点から
複製を開始することができるのではないかと私は考えた。すなわち、Hfr 細胞の中では
F プラスミドが本来持っている F の複製起点からの複製開始は起らなくても、染色体複
製起点を使って複製できるはずである。この仮説に従って染色体 DNA 断片を持つ F’
プラスミド(染色体 DNA 断片を持つ F プラスミドを F’プラスミドという)の中から Hfr 細
胞内で複製できるものを分離すれば証明になると考えた。この実験のアイデアが浮か
んだのはクリスマスイブにワインを飲んでいる最中だったが、さっそく翌日から実験を
始め、正月休みが明けて皆が研究室に出勤してきたときには、すでに目的の菌株を
多数分離することに成功していた。
実験方法はまず染色体の ilvD (isolucine and valine) 遺伝子を持つ F’プラスミドを多
数分離することだった。そのために、ilvD 遺伝子の近くに F プラスミドが挿入された Hfr
菌株をイソロイシンとバリンを要求する ilvD 変異を持つ F-株に接合させて、ストレプトマ

15

16
イシンを添加した最小培地(イソロイシンとバリンを欠如)において増殖してくるコロニー
を分離した。この F-株はストレプトマイシン耐性遺伝子 str と DNA 組み換えに関与する
recA 遺伝子の変異も持っていたで、この F−細胞に Hfr 株から ilvD 遺伝子を運ぶ F’
プラスミドが接合伝達した細胞を多数分離することができた(図7A)。
このようにして分離した株の F’プラスミドが染色体の複製起点も持っているか否かを
調べるために次の検定実験をおこなった。分離した F’プラスミドを持つ F’細胞を他の
Hfr 細胞に接合した場合、Hfr 細胞に伝達されたこのプラスミドがこの Hfr 細胞の中で
複製できるか否かを次の方法で検定した。受容細胞として使った Hfr 株は ilvD 変異・
リファンピシン耐性変異・recA 変異を持っていた。これらの菌株を接合させた後で、リ
ファンピシンを含んだ最小培地(イソロイシンとバリンを欠如)でコロニーが生えてくるか
否かを調べた(図7B)。するとコロニーが生える F’株と生えない F’株の2つのグルー
プに分類できた。すなわち、前者では Hfr 細胞の中でこの F’プラスミドが増殖しており、
これが目的の染色体の複製起点を持つ F’プラスミドである。このタイプのプラスミドは
Hfr 株でも複製できるので、この性質を Poh+ (permissive on Hfr)と名付けた。このグル
ープの F’プラスミドは ilvD 遺伝子の他に dnaA 遺伝子やその近傍の遺伝子も持って
いた。一方、コロニーが検出できなかったグループの F’プラスミドは dnaA 遺伝子近傍
を持たないことが分かった。このことは染色体の複製起点は dnaA 遺伝子の近傍に存
在していることを示唆している。この研究結果を報告した論文(Hiraga, 1967)で、私は
この染色体の複製起点をoriC (origin of chromosome)と命名した。また私はこの論文で、
当時開発された組み換え DNA 技術を使えば染色体の複製起点のみを持つプラスミド
も作製可能であることを予言した。一方、大腸菌と同じ方法で、サルモネラ菌の染色体
oriC を持つ F’ プラスミドを城圭一郎君(九州大学の院生)と一緒に分離することがで
きた。
上記の私の発見が起爆剤となり、世界のあちこちで大腸菌の染色体の複製起点をク
ローニングする試みがおこなわれた。そして数年後に3つの研究室、国立遺伝学研究
所の広田幸敬博士のグループ、ウイルス研究所の私たちのグループ、デンマーク工
科大学/微生物学大学研究所の K. von Meyenburg 博士のグループでほぼ同時に複
製起点 DNA 領域のみを持つプラスミドが分離された。私たちのグループの院生の三
木徹君は、dnaA 近傍に溶原化したファージを誘導して染色体 DNA 断片を運ぶファ
ージを多数分離して、その中から oriC を持つファージを検出し、oriC 近傍の遺伝子配
列を明らかにした。広田グループは複製起点領域の DNA 塩基配列を解析して、複製
起点として重要な配列は 245 塩基対であることを明らかにした。この大腸菌での成功

17
が刺激となり、他のバクテリアの染色体複製起点のクローニングや、酵母の染色体複
製起点 ARS (Autonomous Replicating Segment) のクローニングへと波及して行き、複
製開始のメカニズムについての生化学的研究の土台ができた。その後 A. Kornburg 博
士のグループは oriC プラスミドを使い、複製開始蛋白質 DnaA に依存した複製開始機
構を生化学的手法によって明らかにした。また、ARS により、酵母における複製開始機
構の研究も目覚ましく発展した。
染色体の複製開始頻度の調節に異常のある変異株を分離することを目標として、院
生の田中正史君は、oriC プラスミドのコピー数を増大させる染色体上の変異株を多数
分離して解析した。その結果、これらの変異株では、細胞当たりの染色体数と oriC プ
ラスミドのコピー数が増加していた。そして、これらの変異は RNA ポリメラーゼのまた
は’サブユニットの変異であることが分かった。
● F プラスミドの分配機構の発見
私たちが oriC DNA 断片とアンピシリン耐性遺伝子の DNA 断片を繋いで作った oriC
プラスミドを分離したときに、奇妙な現象に気付いた。アンピシリンを含まない培地でこ
のプラスミドを持つ菌株を増殖させていると高頻度でこのプラスミドを持たない細胞が
でてきた。すなわち、非選択培地ではこのプラスミドは不安定だった。一方、ミニ F プラ
スミドは非選択培地でも安定に子孫の細胞に受け継がれていく性質がある。ミニ Fプラ
スミドとは F プラスミド (約 94.5 キロベース)の複製起点領域を含む 9.2 キロベースの
DNA 断片とアンピシリン耐性遺伝子の DNA 断片を DNA 組み換え技術でつないでつ
くったプラスミドである。このプラスミドは接合伝達に関与する遺伝子群の領域や染色
体へ挿入するために使われる DNA 組み換えに必要な領域は失っているが、F プラスミ
ドと同様、細胞内のコピー数が尐ないにもかかわらず子孫の細胞に安定に受け継が
れて行く性質がある。ミニ F プラスミドの細胞内のコピー数は染色体数とほぼ同じであ
る。oriC プラスミドはミニ F プラスミドよりもコピー数が尐々多いにもかかわらず非選択
培地では不安定だった。
私はこの現象について、ミニ F プラスミドには細胞分裂のとき2つの娘細胞にプラスミ
ド DNA 分子を等分に分配する未知の機構が存在するが、oriC プラスミドはそのような
機構を持っていないので不安定なのではないかと推測した。そしてこのことを証明する
ために、院生の小椋光君にミニFプラスミドのあるDNA断片(約3.7キロベース)をoriC

18
プラスミドに挿入してみることを提案した。すると、作成したそのプラスミドは非選択培
地でも安定に保持された(図 8)。細胞当たりの oriC プラスミドのコピー数はこのミニ F
プラスミドの DNA 断片を挿入した後でも変化しなかった。一方、その DNA 断片を失っ
たミニ F プラスミドは不安定になった。この結果は、その DNA 断片にプラスミドを安定
に保持するために必要な未知の機構が存在していることを示している。その DNA 断片
は、F プラスミド固有の複製起点と複製開始に必要な蛋白質をコードする遺伝子に近
接する DNA 領域だった。この DNA 断片を詳しく解析した結果、そこには蛋白質をコー
ドする2つの遺伝子(sopA と sopB と名付けた。stable maintenance of plasmid)とシス位
置でのみで機能する特異な DNA 領域(sopC と名付けた)が存在することを発見した
(Ogura and Hiraga, 1983)。この3つの機能がプラスミドの安定保持には必要であり、こ
のシステムを「分配機構 (partition system)」と名付けた(図 9)。

19
sopB 遺伝子を他の多コピープラスミドに挿入し、このプラスミドをミニ F プラスミドと共
存させると、ミニ F プラスミドが不安定になる現象を不和合性(IncG)という。院生の楠
川典子さんはこの現象を解析し、この不和合性は SopB 蛋白質の過剰生産によって引
き起こされることを明らかにした。
F プラスミドの分配機構の研究は私が熊本大学医学研究科の教授に赴任してからも
続けて、森浩禎君がこの分配機構に必須な DNA 断片の全塩基配列を決定した
(Mori et al., 1986)。さらに大腸菌で大量生産させた SopA と SopB 蛋白質を精製して、
これらの蛋白質の性質をフットプリンティング法などの生化学的方法で解析した。その
結果、SopA 蛋白質は sopA 遺伝子と sopB 遺伝子で構成されたオペロンのプロモータ
ー領域に結合してオートリプレッサーとして働くことが分かった。SopB 蛋白質が共存す
ると、オペロンのプロモーター領域への SopA 蛋白質の結合はより強くなった。一方、
SopB 蛋白質は sopC 領域の DNA に結合した。sopC 領域には 42 塩基の直列反復配
列が 11.5 個並んでおり、それぞれの直列反復配列の中にある回文塩基配列に SopB
蛋白質は結合した (Mori et al., 1989)(図 10)。

20
永井和夫博士(東京大学)のグループは、SopA 蛋白質にはマグネシウムに依存した
ATPase 活性があり、その活性は DNA の存在下でより強くなることを報告した。SopA と
DNA に SopB 蛋白質を加えるとその ATPase 活性はさらに増加したが、SopB 蛋白質
自身には ATPase 活性は存在しなかった。
以前、外国の研究者によって、「F プラスミド DNA は染色体 DNA に結合しており、染
色体と一緒に娘細胞に分配される」という仮説が提唱されていたが、この仮説を検証
するために、院生の江崎文一君は mukB 欠失変異株を使い次の実験をおこなった。あ
とで詳しく述べるが、mukB 欠失変異株には、細胞分裂の際、染色体を持たない無核
細胞を放出する性質がある。この mukB 欠失変異株にミニ F プラスミドを入れた株を増
殖させて、放出された無核細胞の中にミニ F プラスミドが分配されるか否かを調べる実
験である。この実験結果から、ミニ F プラスミドは mukB 欠失変異株の中でも安定に保
持され、mukB 欠失変異株が放出する無核細胞の中にも正常に分配されることが明ら
かになった。したがって、外国の研究者によって提唱されていた F プラスミド DNA は染
色体 DNA に結合して娘細胞に分配されるという仮説は誤りである。江崎君はこの株の

21
培養液から無核細胞のみを分離精製して、この無核細胞の中に存在するミニ F プラス
ミドを DNA-DNA ハイブリダイゼーションで定量したのである。培養液から、無核細胞
のみを精製する方法は次の方法である。高温感受性の cI リプレッサーを持つファー
ジを染色体上に溶原化した mukB 変異株を使い、この菌株を 30°C で増殖したあとで
この培養液を 42°C に保温すると、染色体を持つ細胞ではファージが誘発され増殖
して細胞を溶かしてしまうが、染色体を持たない無核細胞はそのまま残るのである。放
出されたファージが無核細胞に感染しないようにするために、この株はファージが
外から感染できないように malA 変異(ファージが細胞に吸着できない)を持たせてお
いたのである。こうして精製した無核細胞が持つ F プラスミドを DNA-DNA ハイブリダイ
ゼーションで定量した結果、無核細胞も染色体を持つ細胞と同じようにミニ F プラスミド
を保持していることが証明されたのである。
F プラスミドの分配機構と似た分配機構を持つ他のプラスミドが現在では知られてい
る。P1 プラスミド(P1ファージが大腸菌内に溶原化した状態)の ParA と ParB や TP228
プラスミドの ParF と ParG は、それぞれ F プラスミドの SopA と SopB のホモログである。
しかし、sopC のように cis 位置で働く parC 領域の塩基配列はそれぞれのプラスミドに
特異的である。これらの分配機構は I 型 par システムと呼ばれている(図 11)。

22

23
● F プラスミドの ccd機構の発見
私たちは、ミニ F プラスミドをさらに解析して新規の機構を発見した。その機構には、
複製に必須な領域の左側にある2つの遺伝子によってコードされる蛋白質が関与して
いた。私たちはその遺伝子を ccdA と ccdB と名付け、その機構を ccd 機構と名付けた
(図 12)。ccdA と ccdB との両方を含む DNA 断片を他のプラスミドにクローニングする
ことはできるし、ccdA だけをクローニングすることもできるが、ccdB だけをクローニング
することは出来ない奇妙な現象があった。このことは CcdA 蛋白質が存在しないときに
は CcdB 蛋白質は細胞増殖に悪い影響を与えるが、CcdA 蛋白質が存在すると CcdB
蛋白質の悪い影響は打ち消されることを示していた (Ogura and Hiraga, 1983)。
複製が高温感受性の他のプラスミドに ccdA と ccdB を含む DNA 断片を挿入して、そ
のプラスミドを持つ大腸菌細胞を 30°C で増殖させて 42°C に移したときにどのような

24
ことが起るか Aline Jaffé 博士(パリ大学ジャックモノー研究所)と一緒に詳しく調べてみ
た。42°C ではこのプラスミドの複製が阻害されるので、数回の細胞分裂後にはこのプ
ラスミドを持たない娘細胞が細胞分裂のたびに出てくるが、それらの無核細胞はその
後死んでしまうことがわかった(Jaffé et al., 1988)。
これらの結果に基づいて私たちは次の仮説を考えた。このプラスミドから CcdA と
CcdB 蛋白質が産生され続けている間は CcdA-CcdB 複合体は細胞増殖に悪い影響
を与えないが、このプラスミドを持たなくなった細胞では細胞内に存在した CcdA 蛋白
質の方が CcdB 蛋白質よりも早く壊れ、残った CcdB 蛋白質が細胞増殖に悪い影響を
与えるという「post segregational killing system 仮説」である(図 13)。
この仮説を初めてアメリカでの研究集会で発表したときに、私は自然界から分離さ
れたコピー数の尐ない種々のプラスミドの多くは類似のシステムを持っているであろう
と予言した。このようなシステムは必ずしも CcdA や CcdB 蛋白質のホモログである必要
はないが、細胞増殖を阻害する蛋白質とその蛋白質の細胞阻害能を中和する活性を
持つ蛋白質のペアーから出来ていると推測した。その後、実際に種々のプラスミドから
2つの蛋白質のペアーによる post segregational killing system が他の研究者たちによ

25
って発見された。そして、これらの現象は「プラスミド中毒症」とも呼ばれることになった。
プラスミドを持つ細胞の方が環境内で増殖に優位になるというプラスミドが持つ自己中
心的(selfish)な機構であるといえる。
三木健良博士(九州大学)のグループは、ccdB 遺伝子だけを運ぶプラスミドが存在
しても増殖できる大腸菌の変異株を分離して、その変異が DNA ジャイレースのサブユ
ニット B の遺伝子の変異であることを突き止めた。さらに、CcdA と CcdB とは複合体を
作るが、CcdA の存在しない条件下では CcdB は DNA ジャイレースの B サブユニット
に特異的に結合して DNA ジャイレースを阻害することを生化学的な手法で明らかにし
た。
アメリカの NIH 研究所の Michael Yarmolinsky 博士(NIH)のグループはミニ F プラス
ミドの resD 遺伝子でコードされる蛋白質は rtsF 部位で働く部位特異的な DNA リゾル
ベースであることを発見した(図 12 参照)。細胞内で2つのミニ F プラスミド DNA 分子
どうしが相同組み換えを起こして大きな2量体分子になってしまったときに、rtsF 部位
で効率良く部位特異的組み換えを起こして2つの1量体分子にする機構である。
以上の研究をまとめると、ミニ F プラスミドには複製開始機構、分配機構、ccd 機構
(post segregational killing system)、部位特異的組み換え機構の4つの機構が存在し
ており、これらの機構はコピー数の尐ないミニ F プラスミドを安定に保持するために必
要な基本的な機構である(図 12 参照)。これらの研究成果は、現在 DNA 断片のクロー
ニング用のベクタープラスミドにも忚用されている。分配機構はベクタープラスミドの安
定保持に使われているし、ターゲット DNA 断片を挿入するベクター上の部位として
ccdB遺伝子が使われている。ターゲット DNA断片が挿入されて ccdB遺伝子が破壊さ
れたときのみ、そのプラスミドを持つ大腸菌は増殖できる。したがって、このような市販
のベクタープラスミドを使うと効率よく目的の遺伝子をクローニングすることができるの
である。このベクタープラスミドを増やすためには、CcdB 耐性の変異を持つ DNA ジャ
イレースの変異株を使えば良い。
● ミニ Fプラスミドの安定保持に関与する宿主遺伝子の解析
ミニ F プラスミドの分配機構に働く大腸菌染色体の遺伝子が存在するか否かというこ
とを調べるために、ミニ F プラスミドを安定に保持できなくなった大腸菌の変異株を多
数分離した。遺伝子解析の結果、それらは既知の遺伝子の変異株 (たとえば DNA ジ

26
ャイレースの変異など)であることがわかったが、それらの変異は分配機構に特異的に
関与しているのではなく、他の要因でプラスミドが不安定になったものと考えられる。こ
の研究には時間と労力をかけたが不発に終わってしまった。このことはミニ F プラスミド
の分配機構に直接関与する大腸菌の遺伝子は存在しないことを暗示しているのかも
知れない。
● ミニ Fプラスミドの細胞内位置の解析
行き詰まっていたミニ F プラスミドの分配機構の研究に突破口が開かれたのは、ミニ
F プラスミド DNA の細胞内位置を直接観察する技術を開発したことだった。仁木宏典
君が in situ ハイブリダイゼーションの方法で特異的 DNA 断片の細胞内の位置を調べ
ることにバクテリアで初めて成功した。それは、分離精製したミニ F プラスミドの DNA を
蛍光色素に結合した ATP で標識し、それを DNA-DNA ハイブリッド法の蛍光プローブ
として使って、スライドグラス上にメタノール/酢酸で固定した大腸菌と反忚させてミニ
Fプラスミドを蛍光顕微鏡下で可視化する方法である。ミニFプラスミドを持つ大腸菌を
対数増殖させて、それをこの方法で解析すると強い蛍光焦点が観察された(図 14)。

27
蛍光焦点が1個の細胞と2個の細胞に分けたあと、蛍光焦点の細胞内位置をプロット
した図を作成した。すると、蛍光焦点が1個の細胞ではその蛍光焦点は細胞の中心に
存在しており、2個の蛍光焦点を持つ細胞では細胞長の 1/4 と 3/4 の位置に存在して
いることが明らかになった。一方、分配機構の領域を欠失したミニ F プラスミドの細胞
内分布を調べると、多くの蛍光焦点が細胞の端(端には核様体はない)、あるいは細
胞の中心部(2つの核様体の間)に存在していた。このことは、ミニ F プラスミドの分配
機構はプラスミド DNA の細胞内位置を決定する機構であることを示唆している (Niki
and Hiraga, 1997)。細胞分裂時までに複製したミニ F プラスミドの2つの DNA 分子はそ
れぞれ細胞長の 1/4 と 3/4 の位置に存在しており、細胞の中央で細胞分裂が起ると2
つの娘細胞はそれぞれミニ F プラスミド DNA 分子を受け取ることになる。一方、分配機
構を持たないミニ F プラスミドは染色体 DNA が存在しない細胞端にランダムに存在す
るので細胞分裂の際、プラスミドを持たない娘細胞が出現するのである。

28
● F プラスミドの SopA・SopB蛋白質の細胞内分布
私は技官の一ノ瀬千代女さんと一緒に、特異的な蛋白質の細胞内分布を調べるた
めに新しい免疫蛍光顕微鏡法を開発した。それまでの古い免疫蛍光顕微鏡法ではバ
クテリアの細胞の表面抗原しか染めることができなかったが、この新しい方法によって
細胞内の蛋白質の分布を観察することができるようになった。古い免疫蛍光顕微鏡法
では抗体がバクテリアの膜のペプチドグリカン層を通過できなかったが、この新しい方
法ではリゾチーム処理によってペプチドグリカン層に穴を開けて抗体が通過できるよう
に改良したのである。この方法を使って院生の平野真君が、細胞内の SopA と SopB 蛋
白質の分布を調べ、SopA や SopB が蛍光焦点として局在していることを明らかにした。
その後私が熊本大学教授を定年退官し京都大学の医学研究科で研究を続けること
になったとき、京都大学の院生の足立隼君とこの研究をもう尐し詳しく続けることにした。
ミニ F プラスミドを持った大腸菌を種々の培地で増殖させて SopA と SopB の細胞内分
布を改良した免疫蛍光顕微鏡法で解析すると、SopA 蛋白質は細胞内に均等に分布
しているのではなく、細胞の一端から他の端の間にらせん状に分布していた(図 15A)。
そのらせんのピッチは短い細胞でも長い細胞でもほぼ一定だった。SopB 蛋白質はら
せん状分布と共に何個かの強い蛍光焦点として観察された(図 15A)。これらの SopB
蛋白質の強い蛍光焦点の数は細胞長に比例して増加しており、それぞれの蛍光焦点
の間の距離はほぼ一定だった(図 15C)。SopB 蛋白質とミニ F プラスミド DNA を異なる
色の蛍光物質で染色すると、SopB の強い蛍光焦点とプラスミド DNA の蛍光焦点の多
くは重なっていた(図 15B)。

29

30
国立遺伝学研究所に移った仁木宏典君のグループの研究では、異なる色の蛍光蛋
白質で標識した SopA 蛋白質とミニ F プラスミド DNA を最小培地で増殖中の生きた細
胞で経時的に観察すると、SopA の蛍光は細胞の一端から他の一端にゆっくりと移動し、
プラスミド DNA はその SopA の動きを追いかけて移動していることが分かった。
これらの実験結果と生化学的な研究結果をもとにして、私たちは次のようなモデルを
考えた(図 16A)。SopA-ATP 型はポリマー(フィラメント)を作る。DNA に結合した SopB
は SopA の ATPase 活性を促進するので、SopB-DNA の結合によって SopA-ATP が
SopA-ADP に変換されて SopA ポリマーが2量体になる。2量体になった SopA-ADP は
SopB 濃度の低い他の場所で ATP と結合して再び SopA-ATP ポリマーを形成すること
になる。その結果ミニ F プラスミド DNA に結合している SopB は再びその SopA-ATP
ポリマーに近付くので, SopA 焦点の移動を SopB が結合したプラスミド DNA が追いか
けて行くように見えるのだと推論した。すなわち、SopA ポリマーによって SopB が結合し
たミニ F プラスミド DNA が引き寄せられるというモデルである。
そして私たちは、細胞内に SopB の蛍光焦点がほぼ等間隔で並び、その数が細胞の
伸長とともに増加する現象を、SopA と SopB の反忚拡散系としてとらえる仮説を提唱し
た (Adachi et al., 2006)。反忚拡散系は天才的な数学者 Alan M. Turing 博士が動物
の皮膚の模様形成を説明するために考えた数理理論であり、現在、発生学の分野で
トピックスになっている理論である。SopAはこの反忚拡散系のアクチベーターで、SopB
はインヒビターであると推測される。このシミュレーションの結果は図 16B に示した。こ
の仮説では、F プラスミドの分配機構には大腸菌の染色体上の遺伝子がコードする蛋
白質は必要ない。このことは、以前私たちが F プラスミドの分配に関与する大腸菌染
色体の遺伝子の変異株を分離しようと試みたがそのような変異株を分離できなかった
ことと矛盾しない。
大腸菌における細胞分裂面決定に関与する Min 蛋白質群でも同様な現象が知られ
ていたため、私はこの系を Min システムと同様の数式で表せるのではないかと足立君
に相談したら、彼はこの系を「反忚拡散系」として計算式をたててコンピューターシミュ
レーションをやってくれた。この結果は蛍光焦点の数が細胞の長さに比例することを上
手く再現した。「反忚拡散系」は上に述べたように、脊椎動物の縞模様形成の原理とし
ても有名である。私は複雑な数学は苦手だが、足立君の御蔭でこのシミュレーションの
結果を論文に記載することができた。

31

32
● R1 プラスミドの分配機構
R1 プラスミドは F プラスミドの分配機構と異なる分配機構を持つことが Kenn Gerdes
博士のグループの研究で明らかになった。その分配機構は2つのタンパク質 ParM と
ParR、それにセントロメアとして働く特異的な DNA 領域 (parC)で 構成されている(図
17)。ParR は、parC 領域に結合する性質と、parM-parR オペロンのリプレッサーといて
働く性質とを兼ね備えている。parC には、11 塩基対からなる繰り返し配列が par プロモ
ーターの両側に5個ずつ、計 10 個が存在する。プラスミドが最も安定に保持されるた
めには、この 10 個の繰り返し配列すべてが必要である。ParR タンパク質と parC を持
つ環状らせんプラスミド DNA とを混合すると、DNA の長さは短くなり、parC 領域がら
せんを巻いていることが示唆されている。in vitro において、ParR は parC を持つ 2 つ
のプラスミド DNA 分子を対合させる性質がある。そして、ParM と ATP の存在下では、
このペアリングの頻度が増大する。

33
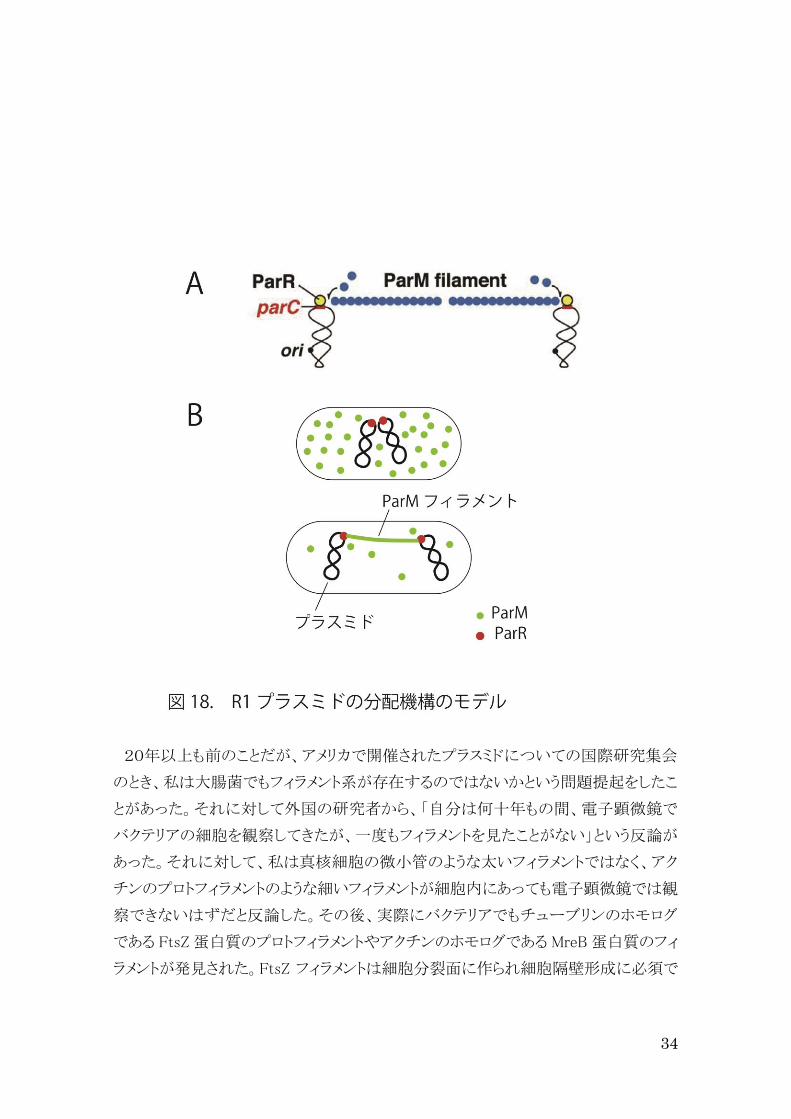
ParM はアクチン様蛋白質で、 ATP 存在下で重合しフィラメントを形成する。ParM は
par プロモーターの転写調節には関係しない。 ParM の ATPase 活性は、ParR が parC
に結合することによって促進する。免疫蛍光顕微鏡法による観察から、ParM は細胞内
に長くカーブしたフィラメントを形成することが明らかになった、このフィラメントは、或る
細胞では細胞端から他の細胞端へと伸び、また或る細胞では細胞の中心から2方向
に伸びていた。さらに ParM は細胞内で蛍光焦点として存在したり、分散したりして存
在していた (図 18)。
ParM は或る ATPase スパーファミリーに属している。このファミリーには、真核細胞の
アクチンや熱ショック蛋白質(Hsc70)・糖キナーゼ・原核細胞の細胞周期蛋白質(FtsA
や MreB)も含まれ、ATP に結合して加水分解する3次元構造をしている。さらに、ParM
にはアクチンと同様、互いにゆるくねじれ合った二重のプロトフィラメントを形成する性
質がある。一定の時間がたつとこの伸長はとまり、脱重合は1方向に急速に進行する。
この ParM フィラメントの崩壊は、ParM の ATP 加水分解と関連している。ADP-ParM フ
ィラメントは著しく不安定であり、一方、ATP--S-ParM は著しく安定である。
アクチンフィラメントの場合は、重合反忚には脱重合反忚と同様、他の因子が関係し
ている。しかし、ParM の場合は他の因子は必要でない。また、ParM フィラメントの
ATP 加水分解によってフィラメントの脱重合が起こる。重合中のフィラメントが2分子の
ParR-parC 複合体と遭遇すると、ParM フィラメントの両端が安定化すると考えられる。
ATP 結合型 ParM が ParM-ParR 結合部位に挿入されることによってフィラメントが伸
長し、それに伴いフィラメントの両端に結合したプラスミドが細胞の両端に向かって 2
方向に離れていくことになる。in vitro と in vivo の実験結果から得られた R1プラスミド
の分配モデルを図 18A に示した。このように、R1 プラスミドでは、ParR の結合した DNA
分子間にできた ParM フィラメントの伸長によって両 DNA 分子が細胞の両端へと押し
やられ、細胞の中央に出来た隔壁によって仕切られてそれぞれ娘細胞に分配される
のである。実際、GFPで標識したParM フィラメントが2つのParRの蛍光焦点の間に伸
長していくことが観察された(図 18B)。R1 プラスミドの持つこの分配機構は、II 型 par
システムと名付けられた(図 11 参照)。一方、F プラスミドの分配機構では、先に述べた
ように SopA ポリマー(フィラメント)がプラスミド分子を引き寄せるのである。
34
20年以上も前のことだが、アメリカで開催されたプラスミドについての国際研究集会
のとき、私は大腸菌でもフィラメント系が存在するのではないかという問題提起をしたこ
とがあった。それに対して外国の研究者から、「自分は何十年もの間、電子顕微鏡で
バクテリアの細胞を観察してきたが、一度もフィラメントを見たことがない」という反論が
あった。それに対して、私は真核細胞の微小管のような太いフィラメントではなく、アク
チンのプロトフィラメントのような細いフィラメントが細胞内にあっても電子顕微鏡では観
察できないはずだと反論した。その後、実際にバクテリアでもチューブリンのホモログ
である FtsZ 蛋白質のプロトフィラメントやアクチンのホモログである MreB 蛋白質のフィ
ラメントが発見された。FtsZ フィラメントは細胞分裂面に作られ細胞隔壁形成に必須で

35
あり、MreB フィラメントは細胞の形を桿状に保つ働きがある。そして、F プラスミドと R1
プラスミドの分配機構にも上に述べたようにフィラメント系が関係しており、私の提起し
た「バクテリアにおけるフィラメント系」という予測は当たっていたことになる。
● 染色体分配に関与した遺伝子の解析
私は F プラスミドの分配機構を研究する一方、大腸菌の染色体の分配機構にも興味
を持っていた。大腸菌の染色体は約 4700 キロベースの長い環状2本鎖 DNA であり、
この DNAが複製しながら2つの姉妹染色体分子となり、細胞分裂時に娘細胞へとそれ
ぞれ分配されるメカニズムは不明であった。栄養豊富な培地では先に述べたように多
数の複製フォークによって複製が進行しているにもかかわらずに、いかにしてこの複製
中の姉妹染色体分子が正確に娘細胞へと分配されるのであろうか。2つの姉妹染色
体分子を細胞分裂時に分裂面の両側に位置するようにする機構(positioning 機構)が
あるに違いないと考えた。ちなみに、私は 1989 年に、細胞内の染色体やプラスミドの
位置を問題にするときには「positioning」という言葉を使い、分裂した姉妹細胞が染色
体やプラスミドを受け取ったか否かを問題にするときには「partitioning」という言葉を使
おうと提案した (Hiraga et al., 1989)。この positioning メカニズムを解析するために、そ
の過程に関与するであろう未知の蛋白質をコードする遺伝子の変異株の分離を試み
た。
この機構に変異を持つ大腸菌ではどのようなことが起るのであろうか。このような変異
株では、細胞分裂時に高い頻度で娘細胞の片方に2つの姉妹染色体 DNA 分子が入
ってしまい他の娘細胞は染色体 DNA を持たない無核細胞になることが起るに違いな
いと考えた(図 19A)。また細胞分裂の際、細胞隔壁形成によって染色体 DNA が切断
されてしまうことも起るかも知れない(「ギロチン効果」と命名)。私たちはこのような細胞
分裂に際して無核細胞放出変異株を放出する変異株を分離する方法を考えた
(Hiraga et al., 1989)。
無核細胞放出変異株を分離するために使うために、次に述べる親株を作製した(図
20A)。この親株は、染色体上には trpE 変異と his 変異があり野生型のファージが溶
原化しており、pXX747 と名付けたプラスミドを持っていた(747 はジャンボジェット機の
番号に由来)。このプラスミドは、或るプラスミドの複製起点とその複製開始蛋白質をコ
ードする repA 遺伝子を持ち、この repA 遺伝子の発現はファージの cI リプレッサーに

36
よって抑制されるように工夫してあった。さらにこのプラスミドは−ガラクトシダーゼの
lacZ 遺伝子(リプレッサー遺伝子 lacI は持たない)と高温感受性変異の cI リプレッサー、
そしてアンピシリン耐性遺伝子 bla を持っていた。
この親株を X−Gal を含む培地中で 42°C でコロニーを作らせると−ガラクトースの
発現は染色体上の lacI 遺伝子がコードする LacI リプレッサーによって抑制されるので、
尐し青味のある白いコロニー(真っ白ではないことに注意!)を作るはずである。そして、
もしこの株に無核細胞を放出する変異が起るとその変異株は同じ条件下で親株よりも
青味の強いコロニーを作るはずである。

37

38
その理由は次にあげる。コロニーの中に出現した無核細胞の中では野生型の cI リプ
レッサーが新たに産生されず、プラスミドがコードする高温感受性 cI リプレサーは
42°C で失活するために、プラスミド上の repA 遺伝子が発現されて RepA 蛋白質が大
量に作られてプラスミドの複製が盛んに起り、プラスミドのコピー数が増加する。すると
プラスミド上の lacZ 遺伝子が発現して−ガラクトシダーゼが大量につくられて培地中
の X-Gal を青色色素にするためコロニーの青味が強くなることが期待される(図 20B)。
この分離方法を考えたのは院生だった仁木宏典君である。
実際にこのやり方で無核細胞を放出する変異株は青味の強いコロニーを作るか否
かを調べるために、私は min 変異をこの親株の染色体上に形質導入で入れてみた。
min 変異とは細胞分裂が細胞の端の近くでも起る結果、染色体を持たないミニ細胞を
放出する変異である。min 変異を入れた株は予想通り青味の強いコロニーをつくり、こ
の分離方法の有効性が確認された。ただし、この無核細胞放出変異株の判定は慎重
におこなわなければならない。なぜならば、親株は lacZ-ではないのであるから真っ白
なコロニーを作るのではなく、尐し青味のあるコロニーを作るのである。すなわち、親株
と変異株の違いは青味の強さの微妙な比較であるので、その差が最も大きくなる時間
帯に色の判定をするためには、コロニーの保温時間が重要である。
私はこの親株を突然変異誘起剤で処理した後で、青味の強いコロニーを作る多数
の変異株を分離した。実際に無核細胞を放出しているか否かを簡単に調べるために、

39
私は新しい顕微鏡法(Hiraga’s fluo-phase combined method)を開発した。それは、
DNA に特異的に結合する青白い蛍光物質 (DAPI) で細胞を染色して、それを蛍光
顕微鏡法と位相差顕微鏡法の両方で同時に観察する方法である(Hiraga et al.,
1989)。この顕微鏡法では青白く輝く染色体を持つ細胞と黒色の無核細胞をはっきりと
区別することができる。そして細胞内の染色体 DNA の形や位置を明瞭に観察すること
もできる。この技術は「コロンブスの卵」的な発見で、その後、多くの研究者によってバ
クテリアの研究で使われることになった。
● mukB 遺伝子の発見
分離した変異株の中には、pXX747 が存在しないときにも青いコロニーを作るラクトー
スリプレッサーlacI の変異と思われる株もあったが、無核細胞を放出する min、rodA、
tolA などの既知の遺伝子の変異も見つかった。しかし、私たちは新しい遺伝子の変異
を発見してそれを mukB 遺伝子と命名した(Niki et al., 1991)。この遺伝子名は日本語
の「MUKAKU」(無核)に由来する。この遺伝子の欠失変異株を作製すると22°Cでは
コロニーを作るが、30°C 以上の温度ではコロニーを作れないという奇妙な性質を持
っており、22°C で増殖中に高頻度で正常の長さの無核細胞を放出し、またギロチン
効果で染色体 DNA の一部しか持たない細胞も放出した(図 19B)。これは最初に予想
した形態を示す変異株である。
この変異を相補する野生型の mukB 遺伝子をクローニングし、その DNA 塩基配列を
仁木君が解析した。それから演繹したアミノ酸配列からこの MukB 蛋白質は 170 キロダ
ルトンの大きな蛋白質であり、コイルドコイルの構造を取りうる長い紐状の構造をしてお
り、N 端と C 端は球状をしていることを示していた。そしてその紐状の中央部にはヒンジ
(蝶番、ちょうつがい)構造を取りうる配列があることが分かった。精製した MukB 蛋白
質は DNA 結合能とマグネシウム依存性の ATPase/GTPase 活性を持っていた(Niki et
al., 1992)。バングラデッシュから来た院生の Abu Z. M. Saleh 君は MukB の変異株を
分離して、C端の球状ドメインの多数の変異株の中には DNA結合能を失ったものがあ
ることを明らかにした。
中国から来た院生の Jin Feng 君が mukB 遺伝子の上流をさらにクローニングして塩
基配列を決定したら、mukB の上流には3つの遺伝子が存在していることが分かった。
助手の山中俊邦君がそれらの遺伝子を1つ1つ欠失した菌株を作製したところ、最上

40
流の遺伝子 (smtA、S-adenosylmethionine-dependent methytransferase) は増殖には
必要でない遺伝子であることが分かったが、2番目と3番目の遺伝子の欠失変異株は
mukB 欠失変異株と同様に 22°C で増殖中に無核細胞を放出し、30°C 以上の温度
ではコロニーを作ることはできない性質を持っていた。したがって、この2つの遺伝子を
mukE、mukF と名付けた(Yamanaka et al., 1996)(図 21A)。Feng 君がおこなったプロ
モーターの解析結果は、smtA-mukE-mukF-mukB は 1 つのオペロンを形成しているこ
とを示していた。
● MukB-MukE-MukF複合体の研究
私たちのグループに加わった助手の山添光芳君が MukE・MukF・MukB をそれぞれ
大量生産する菌株から、これらの蛋白質を精製して種々の性質を生化学的に解析し
た(Yamazoe et al., 1999)。これらの蛋白質はカルシウム依存的に MukB-MukE-MukF
複合体(MukBEF 複合体)を作ることが明らかになった。MukF に強いカルシウム結合
能があった。 MukB の C 端球状ドメインの点突然変異株の中には、DNA 結合能を失
ったものや、MukF との結合能を失ったものがあった。MukBEF 複合体の構成は、
MukB が2分子、MukE が4分子、MukF が4分子であった。またこれらの3種の蛋白質
を大量生産する菌株からは MukBEF 複合体を精製することができた。
森川耿右博士の研究室(大阪大学)の的場京子さんらとの共同研究によって、精製
した MukBEF 複合体と MukB2量体をネガティブ染色して電子顕微鏡で観察したところ、
MukB2量体は予想したように長い細い紐状ドメインはヒンジ部分で折れ曲がり、両端
には球状ドメインが観察された。MukBの2量体のサンプルの中には尐数だがMukBの
1量体も観察され、ヒンジ部位で折れ曲がって紐状ドメインは互いにからまりコイルドコ
イル構造をとり、N 端の球状ドメインと C 端のドメインは接近して存在していることを示し
ていた。このことは、Muk 蛋白質の N 端の球状ドメインと C 端のドメインの複合体は、
或る種の ATPase と3次元構造が似ているという Löwe 博士のグループ(ケンブリッジ大
学)とOh博士のグループ(浦項科学技術大学)によるX線結晶回析の実験結果と矛盾
しない。したがって、このような形態の MukB1量体が2つ、ヒンジ部位で結合して2量体
を作っていると考えられる(図 21B)。

41

42
私たちの MukB 蛋白質の発見から数年後に両生類のアフリカツメガエルの SMC
(XCAP-E/C) 蛋白質や分裂酵母の SMC ホモログが発見されたが、この SMC の形態
と MukB の形態は酷似している。一方、MukBEF 複合体の両端の球状ドメインは MukB
2量体の球状ドメインに比べて大きい(図 21B)。それは予想通り、MukB2量体の両端
の球状ドメインに MukE と MukF が結合しているためである。MukBEF 複合体は連結し
て長い鎖を作り、その鎖は環状になりロゼット型(花びら型)をとりやすい性質があった
(図 21C)。MukB の N 端の球状ドメインの X 線回析の結果が van den Ent ら (1999) に
よって報告され、N 端の球状ドメインと C 端の球状ドメインが結合してある種の ATPase
に類似した構造になることが示唆された。
MukB のアミノ酸配列から演繹した蛋白質の形は、当時発見された真核生物のキネ
シンの形に似ていたのでモーター蛋白質の可能性を初めは考えていたが、Weitao ら
(1999) によって、mukB 欠失変異株は DNA ジャイレースの阻害剤ノボビオシンに対し
て超感受性を示すことが分かり、MukB は染色体 DNA の折りたたみなどの構造に関与
していることが示唆された。院生の小野木君はこのことを追試実験で確かめ、私たちも
MukB は DNA ジャイレースとともに染色体 DNA の3次元的構造に何らかの働きをして
いるという考えに至った。また、dam や seqA の欠失変異は mukB・mukE・mukF 欠失変
異が示す高温感受性を抑制するが、ノボビオシンに対する超感受性や無核細胞の放
出を抑制することはできなかった。
熊本大学を定年退官後に、京都大学において院生の足立隼君とこの問題をさらに
解析するために、mukB 欠失変異の示すノボビオシン超感受性を抑制するノボビオシ
ン耐性変異株を分離したら、それらは全て DNA ジャイレースのサブユニットの gyrB
遺伝子の変異であった。DNA 塩基配列の解析によって gyrB 遺伝子の変異であること
を確認した。これらのノボビオシン耐性株は親株である mukB 欠失変異株と同様に
37°C でコロニーを作ることができないし、22°C で増殖中に無核細胞を放出する。
Rebenkov のグループ(オクラホマ大学)は、環状2本鎖プラスミド DNA と精製した
MukB2量体を混合したあとトポイソメラーゼ処理してから、DNA を抽出し DNA の超ら
せん構造を調べ、環状 DNA の捻れが増加したことを報告した。しかし、この解析方法
では、MukB2量体が結合したときに DNA に生じるねじれを測定できるが、MukB の細
胞内の真の働きを測定しているのでないと思われる。
私たちのグループと吉川研一博士(京都大学)のグループとの共同研究で、Chen
Ning さんは新しい測定法を使って MukBEF 複合体の機能を解析した (Chen et al.,
2008)。それは蛍光化合物 YOYO で染色した T4ファージの長い直鎖2本鎖 DNA(約

43
166,000 塩其対、57 m)をビデオカメラで撮影する方法である。この方法によって
DNA 分子は蛍光を発する塊として液体中に浮遊しているのが観察される(図 22A)。こ
の塊の長軸の平均長は 4 m であったが、ATP 存在下で MukBEF 複合体を加えたと
きには 3 m に縮み(図 22B)、DNA の塊の中に高い蛍光強度を示す部分が現れた
(図 22A 参照)。このような効果は ATP の代わりに ADP や ATP-S を存在させたとき
には観察されなかった。これらの結果は MukBEF 複合体の ATPase 活性が DNA を縮
ませることに関与していることを示している。また、ATP 存在下で MukB の2量体を加え
たときは DNA の縮みは小さかった。このことは、MukB2量体の ATPase 活性は
MukBEF 複合体の ATPase 活性よりも低いことと矛盾しない。
カバーグラスをかける時の水流の力で DNA 分子を引き延ばして観察すると、
MukBEF 複合体と ATP との存在下では DNA 繊維上に小さな DNA の塊があることが
分かった。MukBEF 複合体が DNA 分子に結合して、DNA 分子の一部に塊を作ってい
るものと推測される。
MukBEF 複合体が細胞内で染色体 DNA の構造にどのような影響を与えているの
かを知るために、足立君が野生型株と mukB 欠失変異株から抽出した RNA 分画を大
島拓博士(奈良先端科学技術大学院大学)に送り、大腸菌ゲノムのDNAアレーを使っ
て全遺伝子の発現量の比較を調べてもらった。両株の間には遺伝子の発現に大きな

44
違いはなく、MukBEF 複合体は DNA のらせん構造に大きな影響を与えないことが明ら
かになった(未発表)。
ポスドクの大住克史君は、GFP が結合した MukB 蛋白質をコードする組み換え遺伝
子 mukB-gfp を染色体上の本来の mukB 遺伝子と置き換えた菌株を作成した。この菌
株の細胞を生きたまま蛍光顕微鏡で観察し、MukB-GFP が蛍光焦点として存在するこ
とを見いだした(図 23)。蛍光焦点が1つの細胞では、その焦点は細胞中心に存在し
ており、蛍光焦点が2つの細胞ではそれぞれ細胞長の 1/4 と 3/4 の位置に存在してい
た。MukB-GFP と SeqA の蛍光焦点の位置は同じではない(Ohsumi et al., 2001)。

45
● mukB 変異株およびその抑制変異株の解析から派生した種々の研究
院生の今村龍君は無核放出変異株の解析過程で、染色体地図の 66.2 分に位置す
る新しい遺伝子 (cpdA と命名)を見つけて、この遺伝子 3’-5’-adenosine-
monophosphate phosphodiesterase をコードすることを発見した。
助手の山中君は、MukB 蛋白質の機能を別の角度から明らかにするため、mukB 欠
失変異の示す高温感受性を抑制するサプレッサー変異株を多数分離した。遺伝子解
析の結果、全ての変異が単一の遺伝子の変異であり、その遺伝子を smbA と名付けた。
その後、smbA は UNP キナーゼの遺伝子(pyrH)である事が明らかになった。また
smbA 変異を抑制するマルチコピー遺伝子 mssA と mssB を見つけたが、その後これら
はそれぞれ CMP/dCMP キナーゼ (cmk)、ATP 依存性 RNA ヘリカーゼ (deaD) をコ
ードすることが明らかになった。
院生の城戸麻喜子さんは、mukB106 ミスセンス変異(Ser33→Phe)の示す高温感受
性を抑制する変異株のうち、ある連関群に属する変異は全てRNase Eをコードする rne
遺伝子の変異である事を発見した。これらの変異株は RNase E の C 端側の半分が欠
失した蛋白質を産生していた。これらの欠失蛋白質はエンドリボヌクレアーゼ活性を保
持しているが、ポリヌクレオチドフォスファターゼ (PNPase) との結合能を失っているこ
とを免疫共沈殿法により証明した。RNase E と PNPase は複合体を作り、mRNA や他の
RNA のプロセッシングや崩壊に協同して働いているものと考えられる。実際にこの rne
変異株では MukB 蛋白質の量が増加していた。したがって、これらの rne 変異による
mukB106 変異の示す高温感受性の抑制は MukB106 変異蛋白質の量的増加に起因
すると結論した。
一方、山中君とチリから来た JAICA 留学生の三谷忠男君は、mukB106 変異を抑制
するマルチコピー抑制遺伝子の解析からは、cspC・cspE 遺伝子を発見した。これらの
遺伝子がコードする蛋白質はコールドショック蛋白質のアナログであった。残念ながら、
これらの抑制変異株の解析結果からは MukB の機能を知る手掛かりは得られなかっ
た。
私たちが開発した無核放出変異株を検出する方法は、他の研究者の興味をひき、
親株を何カ所かに送ったが、コロニーの色の判定が微妙なので困惑する研究者が多
かった。しかし、和地正明博士(東京大学)と第1製薬株式会社の山岸純一博士らは、
この方法を MukB 質の機能を阻害する薬剤のスクリーニングに使い、無核細胞を放出
させる薬剤 A22 [S-(3,4-dichlorobenzyl)isothiourea]を発見した。その薬剤に耐性の変

46
異株を分離して、その変異を解析したところ、それは mukB 遺伝子ではなく、アクチン
のホモログである mreB 遺伝子であった。MreB を阻害すると細胞の形が丸くなるととも
に、丸い無核細胞が放出されたのである。足立君はこの薬剤を添加後に、細胞が丸く
なる寸前までの oriC の細胞内の位置を詳細に調べたが、異常は見られなかった。大
腸菌では、MreB は染色体の positioning には関与していないと考えられる。
● DNA トポイソメラーゼ IVの発見
私たちが MukB 遺伝子の塩基配列を解析しているとき、この遺伝子の下流に未知の
ORF の一部があることが分かった。この ORF は偶然にも加藤潤一博士(東京大学)が
解析中の新しい DNA トポイソメラーゼのサブユニットの遺伝子(parC) の一部であるこ
とに気付いた。加藤博士もこの遺伝子の塩基配列を半分解析中だったので、両グル
ープのデータを合体して ORF を完全なものとし、この DNA トポイソメラーゼ IV の発見
の論文を両グループの共同研究として発表した。その後、DNA ジャイレースは主に
DNA のらせん構造に関与するが、DNA トポイソメラーゼ IV は2分子の環状2本鎖 DNA
が連鎖しているときにその連鎖を切断・再結合によって解除する働きがあることが
Nicholas R. Cozzarelli 博士(カリフォルニア大学)のグループの研究から明らかになっ
た。
● 染色体 DNAの細胞内位置の解析
染色体 DNA の細胞内の位置を調べるために、仁木宏典君は大腸菌染色体の複製
起点または複製終点近傍の DNA 断片を持つファージ DNA をプローブとして、最小
グルコース培地で対数増殖していた大腸菌細胞を in situ ハイブリダイゼーションによ
って解析した。複製起点の蛍光焦点を1つ持つ細胞では、その焦点は細胞長さの 1/4
の位置に存在しており、蛍光焦点を2つ持つ細胞では 1/6 と 5/6 の位置に存在してい
た。一方、複製終点近傍の蛍光焦点を1つ持つ細胞では、その焦点は 1/6 または 1/2
近傍に存在していた。また複製終点の蛍光焦点を2つ持つ細胞では、1/2 近傍に互
いに接近して存在していた。これらの結果は、染色体の複製起点と複製終点は規則
性を持って細胞内に存在していることを示している。

47
大腸菌染色体の複製起点を持つ oriC プラスミド DNA は細胞端に近い部位に存在し
ていたが、F プラスミドの sopABC を含む DNA 断片を結合した oriC プラスミド DNA は、
その蛍光焦点が1つのときには細胞の中心部に存在しており、焦点が2つの場合は細
胞長の 1/4 と 3/4 の位置に存在していた。これは先に述べたミニ F プラスミドでの研究
結果から予想さる結果であるが、sopABC がプラスミド DNA の細胞内位置を決定する
システムであることを支持している。またこの実験結果は、oriC 自身には細胞内の位置
を決定する機能はないことを示しており、染色体上のoriCが規則正しい細胞内の位置
に存在するためには oriC に近い座位に未知の塩基配列が存在することを暗示してい
る。この研究はその後、仁木君と院生の山市嘉治君によって oriC から 211 キロベース
離れたところに存在する migS と名づけた塩基配列の発見へと発展した。
最小グルコース培地で対数増殖していた大腸菌細胞を種々な染色体 DNA 断片を
もつファージ DNA をプローブとして使って in situ ハイブリダイゼーションによって解析
した。蛍光焦点を2つ持つ細胞では、複製起点は細胞長さの 1/4 と 3/4 の位置に存在
しており、複製終点は細胞の中央部に互いに接近して存在しており、他の部位の
DNA は複製起点と複製終点の位置の間に染色体の座位の順に位置しているように見
えた。このことから、染色体 DNA は長いままに無規則に絡まった状態で細胞内に存在
するのではなく、太い環状な状態で存在していることが明らかになった。DNA に特異
的に結合する蛍光物質 DAPI で染色すると、太い環状の染色体が観察された。
● SeqA蛋白質の細胞内分布
大腸菌 K-12 株の染色体 DNA 上には GATC 配列が約 19130 箇所存在するが、こ
の GATC 配列のアデニンは DNA アデニンメチルトランスフェラーゼ(Dam メチラーゼ)
の働きでメチル化されている。 DNA が複製した直後は古い DNA 鎖の GATC のアデ
ニンはメチル化されているが新たに合成された相補鎖上の CTAG 配列のアデニンが
未 だ メ チ ル 化 さ れ て い な い と き が あ る は ず で あ る 。 す な わ ち 、 半 メ チ ル 化
(hemimethylation)の状態である。
アンピシリン耐性遺伝子を持つoriCプラスミドを野生型dam+ 株の細胞で増殖させて
そのプラスミド DNA を dam 欠失株に形質転換で入れた場合、アンピシリン耐性の形質
転換体の出現効率が dam+細胞に入れた場合と比べて著しく低くなる現象がある。一
方、dam 欠失変異株の細胞で増殖させた oriC プラスミドを dam 欠失株に形質転換で

48
入れた場合には dam+細胞に入れた時と同じように効率良く、アンピシリン耐性の形質
転換体が得られる。野生型 dam+ 株の細胞で増殖させた oriC プラスミドを dam 欠失株
に形質転換で入れたときには、1回複製した段階で半メチル化 DNA になるがこの状態
の DNA は複製できないことを暗示している。
このように野生型 dam+ 株の細胞で増殖させた oriC プラスミド DNA を dam 欠失株に
形質転換に入れた場合には複製できないが、このプラスミドが複製できる第2の変異
を持つ大腸菌の変異株を Nancy E. Kleckner 博士(ハーバード大学)のグループは分
離し、遺伝解析の結果 seqA (sequestration A) 遺伝子を発見した。SeqA 蛋白質は半
メチル化した GATC 配列に特異的に強く結合することが分かった。oriC 領域には
GATC 配列が 11 個所存在する。dam 欠失変異株では染色体の複製開始の調節が異
常になることが知られている。複製開始の同調性が崩れているのである。
私たちは SeqA 蛋白質の抗体を使って蛍光抗体顕微鏡法で細胞内の SeqA 蛋白質
の分布を調べたところ、SeqA は強い蛍光焦点として観察されることを見いだした(図
24)。指数増殖中の大腸菌を調べると、SeqA 焦点を1個だけ持っている細胞ではその
焦点は細胞の中央に存在し、SeqA 焦点を2個持っている細胞ではその焦点は細胞長
の 1/4 と 3/4 の位置に存在していた。一方、DNA アデニンメチラーゼを欠く dam 欠失
変異株では、SeqAは細胞に広く分散して分布しており、蛍光焦点は観察されなかった。
すなわち、SeqA の蛍光焦点は DNA のメチル化と関連がある現象である。免疫蛍光法
と in situ ハイブリダイゼーションとを組み合わせた方法によって、SeqA 蛋白質と oriC
DNA を異なる色の蛍光物質で染色すると、大部分の SeqA 蛍光焦点は oriC 蛍光焦点
とは異なる位置に存在していた。oriC 欠失変異株 (oriC rhn)でも SeqA 蛍光焦点
は観察されるので、oriC 領域に結合した SeqA 分子の塊を見ているのではなく、oriC
以外の染色体領域にある GATC 配列がこの現象に関与しているものと考えられる。ま
た、複製開始蛋白 DnaA の欠失変異株 (dnaA rhn)でも SeqA 蛍光焦点は観察され
るので、SeqA 蛍光焦点の形成には DnaA は必要ない (Hiraga et al., 1998)。

49
この SeqA の蛍光焦点が、複製フォークに存在する半メチル化 DNA に結合した多数
の SeqA 分子の塊であることを証明するために次の実験をおこなった。高温感受性の
複製開始蛋白質をコードする dnaA 変異株を 30°C で増殖させて 42°C で保温する
と染色体 DNA の複製開始が阻害されて暫くすると複製中の複製は終了するが、その
細胞を再び 30°C に短時間さらし複製が開始したあとで直ちに 42°C に移すと次の
複製が阻害されるために、複製が同調して1回だけ起る系を作ることができる。この系
を使って SeqA の細胞内分布を調べたところ、複製が終了した細胞では SeqA は細胞
内に広く分散して強い蛍光焦点は観察できなかった。しかし、染色体が複製を開始す
ると直ちに細胞中心部に蛍光焦点が1個現れた。複製が 25%ほど進行したときにこの
蛍光焦点は2つになり細胞長の 1/4 と 3/4 の位置に存在していた (Hiraga et al. 2000)。
院生の小野木利成君が緑色蛍光タンパク質(GFP)を結合した SeqA 蛋白質を産生する
菌株を作成して生きた細胞を経時的に蛍光顕微鏡で観察すると、1つだった SeqA の
蛍光焦点が2つに分離していく過程が観察できた。SeqA 蛍光焦点野移動速度は1分
当り約1m であった。この生きた細胞での SeqA-GFP の観察は、免疫蛍光抗体法で

50
観察した SeqA の局在が細胞固定によるアーティファクトではないことを示している。
さらに複製装置(レプリゾーム)の細胞内位置を調べるために、同じ複製同調系にお
いて複製装置で働く DnaB ヘリカーゼを観察した。DnaB ヘリカーゼは複製が完了した
細胞では、細胞の端の染色体の存在しないに所に分布しているが、複製が開始する
と1個の強い蛍光焦点として細胞中央部に現れて、複製が 25%程進行したときには細
胞長の 1/4 と 3/4 の位置に2つの蛍光焦点として存在していた。これらの結果は、
SeqA の蛍光焦点は複製フォークにおいて合成された直後の半メチル化 DNA に結合
した多数の SeqA 蛋白質分子の塊であることを示している。またこの結果は、複製起点
から2方向に進行する2つの複製装置は初め細胞中心部に接近して存在するが、複
製が 25%程進行した時点で2カ所に分かれることを示している。
さらに複製フォークで合成された直後のDNA断片の細胞内位置を調べるために、複
製同調をした細胞をチミヂンのホモログである5-ブロモデオキシウリジンで短時間標
識し、その細胞を 5-ブロモデオキシウリジンのモノクローナル抗体を使って免疫蛍光
染色をした。その結果、複製開始直後に合成された DNA 断片は1個の蛍光焦点とし
て細胞中心部に存在しているが、複製が 25%程進行したときに合成された DNA は細
胞長の 1/4 と 3/4 の位置に存在した。この実験結果は SeqA や DnaB の結果と同様
に、複製起点から2方向の複製に働く2つの複製装置は初め細胞中央に接近して存
在するが、複製が 25%程進行したときに2カ所に分かれることを強く示している。すなわ
ち、複製装置は細胞内の1箇所で動かずに複製をしているのではない。
私は高温感受性 dnaC 変異株を使った複製開始の同調系の実験をおこない、複製
した2つの oriC コピーは暫くの間、細胞内に接近して存在するために1つの蛍光焦点
として観察されるコヒージョン (cohesion) の現象を発見した。
● 複製開始における SeqA蛋白質の阻害機構
先に述べたように半メチル化した oriC プラスミド DNA は複製できないことが分かって
いる。このことは大腸菌の染色体複製起点 oriC で複製が起ったのちに、しばらくは再
び複製は開始されないという現象と関連がある。すなわち、複製起点領域に存在する
GATC 塩基配列が半メチル化されている期間 SeqA 蛋白質はこの配列に強く結合して
おり、複製開始を抑制しているものと推則される。

51
山添君は大量の SeqA 蛋白質を効率良く精製するために His タグを C 端に結合した
SeqA 蛋白質をコードするプラスミドを作成した。この His タグを付けた SeqA 蛋白質は
seqA 欠失変異が示す複製開始異常を相補するので、正常の活性を持っていると結
論できる。
SeqA による複製開始阻害のメカニズムを生化学的手法で明らかにするために私た
ちは Masamichi Kohiyama 博士(パリ大学ジャックモノー研究所)のグループと共同研
究をおこなった。精製した SeqA 蛋白質と DnaA 蛋白質を使って oriC プラスミド DNA
への結合パターンを DNaseI フットプリント法で解析した。oriC 領域には5箇所に DnaA
蛋白質が結合する塩基配列 DnaA ボックスがあり、SeqA 蛋白質が結合する GATC 配
列が11個所あるが(図25A)、半メチル化oriC DNAを使ったときにはSeqA蛋白質は、
oriC 領域の AT クラスター、13 マー、R1 DnaA ボックス, M DnaA ボックスを含む領域

52
に効率良く結合した。DnaA もこの oriC DNA の R1 DnaA ボックスから 13 マーの領域
にかけて結合した。
A. Kornberg らによって開発された oriC プラスミド DNA の in vitro の複製系において
精製した SeqA 蛋白質の阻害効果を調べたところ、半メチル化状態の oriC プラスミド
DNA の複製は SeqA によって著しく阻害されたが、一方メチル化されていない oriC プ
ラスミド DNA の複製は全く阻害されなかった。半メチル化した oriC プラスミド DNA に
DnaA は結合するが、そこに SeqA を加えると DNA から DnaA が遊離し、代わり SeqA
が結合することがわかった(図 25B)。このことは SeqA が複製開始反忚を阻害するメカ
ニズムを示している。また、Dam メチラーゼ・SeqA・oriC DNA の相互関係を解析した結
果、半メチル化 oriC プラスミドは SeqA 存在下では Dam メチラーゼによってメチル化さ
れないことが明らかになった。
● SeqA蛋白質の結晶構造解析
SeqA 蛋白質を大量発現するプラスミドを横山茂之博士・胡桃坂仁志博士(理研ゲノ
ムサイエンスセンター/東京大学)に送り、SeqA 蛋白質の結晶構造解析を頼んだ。
SeqA 蛋白質は 181 個のアミノ酸よりできているが、N 端のドメイン(1-59 アミノ酸残基)
は DNA 結合能を持たず、2量体をつくるための自己結合能を持っていることがわかっ
た。C 端のドメイン(71-181 アミノ酸残基)は自己結合能を欠き、半メチル化した
G-mA-T-C 配列に特異的に結合した (2003)。この最小の DNA 結合ドメインである
SeqA71-181 は、mA:G のミスマッチ塩基配列を持つ半メチル化 DNA [G-mA(:G)-T-C]
にも正常な半メチル化 DNA [G-mA(:T)-T-C] と同様に特異的に結合することが分か
った。これらのミスマッチ塩基配列や正常塩基配列と結合した SeqA71-181 を結晶化し
てその構造を解析した。両者ともに同じ構造をしていたが、電子密度はメチル化されて
いないアデニンの近くに観察された。この電子密度はおそらく水分子または金属イオ
ンによるものと考えられる。GATC 配列の両 DNA 鎖のアデニンがメチル化されていると
きには、このような電子密度は観察されなかった (Kurumizaka et al., 2004)。

53
● 大腸菌のセックス現場の視覚化
SeqA 蛋白質が半メチル化 DNA に特異的に結合することを利用して、ちょっと面白い
実験をした。大腸菌のセックスの現場を可視化して観察しようという実験である。すな
わち、「ピーピングトム」である。接合の雄株(供与株)には野生型の F+細胞または Hfr
細胞を使った。雌株(受容株)には DNA アデニンメチラーゼを欠く dam 欠失変異を持
ち緑色蛍光蛋白質(GFP)を産生する F-細胞を使った。両者を接合後に細胞を固定し、
SeqA を赤色蛍光色素で染め、GFP を緑色蛍光色素で染めて蛍光顕微鏡によって観
察した。接合させない F+細胞や Hfr 細胞は赤色の蛍光焦点を持っていた。これは先に
述べた複製フォークの半メチル化 DNA に結合した SeqA の塊である。一方、接合しな
い F- dam 変異細胞は細胞全体が緑色の蛍光を発していたが赤色蛍光焦点は観察さ
れなかった。しかし、接合した後では高頻度で、赤色の蛍光焦点を持つ緑色の細胞が
観察された(図 26)。このタイプの細胞内の赤色の蛍光焦点は F-細胞に伝達された
DNA に多数の SeqA 分子が結合したものである。供与細胞から F- dam 変異細胞に伝
達されたメチル化された1本鎖 DNA に対して合成された相補鎖はメチル化されないの
で半メチル化のままで存在して,それに SeqA 分子が多数結合しているのである。この
接合伝達された DNA の可視化の研究結果は Kohiyama, Hiraga, Matic, and Radman
(2003)「Bacterial Sex: Playing Voyeurs 50 Years Later」(バクテリアのセックス:50年後
の覗き視)というタイトルの論文として Science に掲載された。それは Joshua Lederberg
博士による「大腸菌における性の発見」(ノーベル賞受賞)からちょうど50年後のことで
あった.。

54
● 大腸菌とその近縁な種に特異的に存在している遺伝子群
細胞の増殖にとって重要な遺伝子はバクテリアに普遍的に存在していると一般的に
は考えられているが、果たしてそうであろうか。ゲノムプロジェクトの世界的な発展によ
って多くのバクテリア種の全ゲノム塩基配列が明らかになって来たので、この問題につ
いてコンピューター検索をおこなってみた。
DNA アデニンメチルトランスフェラーゼをコードする dam 遺伝子はプロテオバクテリア
のサブデビションに分類される種の中の大腸菌 Escherichia coli および近縁種である
Salmonella typhi ・ Salmonella paratyphi ・ Salmonella typhimurium ・ Klebsiella
pneumoniae・Yersinia pestis・Vibrio cholerae・Actinobacillus actinomycetemcomitans・
Haemophilus influenzae・Pasteurella multocida などにのみ存在しており、同じくサブデ
ビションに分類される Pseudomonas aeruginosa や Legionella pneumophilania には存在
しないことがわかった。dam 遺伝子はプロテオバクテリアの他のサブディビジョンの種
やグラム陽性の種にも存在しなかった(Hiraga et al., 2000)(図 27)。
また半メチル化 GATC 配列に結合する SeqA 蛋白質をコードする seqA 遺伝子や、
同じく半メチル化 GATC 配列を識別するミスマッチ修復に関与する MutH 蛋白質をコ

55
ードする mutH 遺伝子も dam 遺伝子と同様にこれらの大腸菌の近縁種にのみ存在す
る。さらに mukB・mukE・mukF 遺伝子も大腸菌の近縁種にのみ存在していた。 興味あ
ることに、MukB 蛋白質の機能的ホモログである Smc 蛋白質をコードする smc 遺伝子
は、mukB 遺伝子を持たない種に広く存在していた。Smc 遺伝子はグラム陽性菌の枯
草菌 Bacillus subtilis にも存在している。16S リボゾーム RNA の塩基配列の比較から演
繹された系統樹からは、枯草菌は祖先型の種に近いことが分かっている。大腸菌とそ
の近縁種がサブディビジョン内の他の種と分岐したときに dam 遺伝子を獲得し、それ
に伴ってこのような大きな変化が起ったものと推測される。これらの大腸菌の近縁種は
哺乳類の進化に伴って進化してきた新しいタイプのバクテリアであると推測される。
F プラスミドの sopA・sopB 遺伝子のホモログ (parA・parB など) は、大腸菌やインフ
ルエンザ菌のゲノムには存在しないが、コレラ菌や多くのグラム陽性菌のゲノムに存在
している(図28A)。枯草菌では、SopBのホモログである Spo0Jが胞子形成時の染色体
分配に必須である。染色体とプラスミド間では、遺伝子のやり取りが長い進化の過程で
頻繁に起ってきたのであろう。

56
ParA 蛋白質のホモローグの分子進化の系統樹を図 28B に示した。細菌のゲノム上
の ParA ホモローグよりも、プラスミドの ParA ホモローグの進化スピードが速い。細胞壁
を細胞の中央に形成するために働く MinD 蛋白質も SopA 蛋白質に似ており、SopA 蛋
白質と同じ祖先蛋白質から進化してきたと考えられる。MinD 蛋白質は、MinC 蛋白質
と MinE 蛋白質と一緒に反忚拡散系を構成しているという仮説が de Boer ら(ケース・ウ
ェスタン・リザーブ大学)によって提唱され、実際に Loose ら(ドレスデン大学/マック
ス・プランク研究所)によって in vitroで証明された。MinDはこの系のアクチベーターで
ある。先にのべた私たちの SopA・SopB の反忚拡散系仮説でも、SopA はアクチベータ
ーであることは興味深い。
大腸菌染色体には sopA・sopB は存在しないが、代わりに、先に述べた oriC に近い
染色体部位に存在する migS 領域が細胞内の oriC の位置を決めることに関与している
のである。

57

58
● 細胞周期における染色体の動態
上に述べて来たように、種々の蛋白質や DNA の細胞内の位置は細胞周期に伴って
規則的に変化するが、このことをもっとシステマテックに解析することを足立君と試みた
(Adachi, Fukusima, and Hiraga, 2008) 。培地の条件により大腸菌の染色体複製開始
頻度は異なるので、3つの培地の条件下で対数増殖中の大腸菌における染色体や蛋
白質の動態を調べた。培地は最小グリセロール培地とそれに 20 種のアミノ酸を加えた
培地と、栄養豊富の LB 培地である。温度が 22°C のときに1世代の長さは、それぞれ
270、180、90 分であった。調べたのは複製起点 (oriC)、複製終点 (ter)、SeqA 蛋白
質、MukB 蛋白質、FtsZ 蛋白質、短時間 DNA に取り込ませたブロモデオキウリジン
(BrdU) の細胞内の位置、DAPI で染色した染色体 DNA(核様体)の形態である。そし
てフローサイトメーターを使って細胞当たりの複製起点の数も測定した。SeqA 蛋白質
と BrdU の蛍光焦点は複製フォークの位置を示す。また BrdU の蛍光焦点は染色体
DNA の複製を表している。FtsZ リングは細胞分裂をする位置に作られる。
最小グリセロール培地での結果を図 29 に示す。この実験条件では、cell age の 0.3
から 0.7 に染色体 DNA の複製が起った(cell age は細胞分裂から次の細胞分裂までを
0 から 1 として表す)。細胞中心にあった複製起点は複製後しばらく互いに同じ場所に
存在した後で 1/4 と 3/4 の位置に移動する。いわゆるコヒージョン (cohesion) の現象
が観察された。一方、複製終点は初め細胞端に近い所に存在するが 0.6 cell age のこ
ろ細胞中心に移動し、そこで複製するが複製したあと互いに接近して存在しているの
で1つの蛍光焦点として見え、その後分かれて2つの焦点となる。すなわち、複製焦点
にもコヒージョンの現象が観察された。SeqA や BrdU の焦点として観察される複製フォ
ークは複製開始と同時に細胞中心に1つ存在するが、複製が相当進行した後で2つ
に分かれて 1/4 と 3/4 の位置に移動する。興味あることは、MukB の蛍光焦点は複製
開始前にすでに 1/4 と 3/4 の位置に2つ存在することである。複製起点の複製後のコ
ヒージョンの現象は20種のアミノ酸を添加した培地でも、栄養豊富のLB培地でも観察
された。LB 培地ではマルチフォーク複製が起っていた。
2001 年の論文で、院生の砂子由美さんがおこなった染色体複製の同調系において
は mukB 欠失変異株で複製起点のコヒージョンが観察されなかったことを報告した。し
かし、その後の研究からは増殖条件によっては mukB 欠失変異株でも複製起点のコヒ
ージョンが観察され、複製起点のコヒージョンは必ずしも MukB 蛋白質の機能によるも
のではないことが分かった。

59
このような細胞周期に伴う規則正しい染色体の動態が、どのようなメカニズムで起る
のかはまだ明らかでは無く、今後に残された課題である。MukB のクラスターが染色体
DNA の複製前には既に 1/4 と 3/4 の位置に存在している現象は、MukB の真の機能
の解明と共に興味ある問題である。

60
おわりに
大腸菌の染色体 DNA やプラスミドの分配機構の研究を始めたときには、この分野に
はほとんど研究者がいなかった。私はオリジナリティーのある研究がやりたくてこのテ
ーマを選んだのであるが、このテーマには DNA の細胞内位置の問題が含まれている
にもかかわらず、当時バクテリアでは DNA や蛋白質の細胞内分布を調べる方法がな
かった。したがって、私はこの研究テーマを選ぶことに尐なからず不安を感じていた。
しかし、私はいずれ将来新しい実験方法が見つかり解決するだろうと楽観して、自分
が得意とする遺伝的研究方法で研究を始めた。そして最初のうちはほぼ独走体勢で
研究を続けることができた。しかし、いくらオリジナリティーの高い研究だと言っても、だ
れも興味を示さない研究なら寂しいものだが、幸いのことに二十年三十年と経つうちに
世界の研究者が多数参入してきてバクテリアにおける分子細胞生物学ともいえる一大
研究分野に発展した。
2004 年にアメリカのニューメキシコ州サンタフィで開催されたキ−ストーンミーテング
「Bacterial Chromosome」では、集会の最初に1時間の基調講演をする栄誉に浴した。
オルガナイザーのアメリカの Nancy Kleckner 博士たちが私のために定年退官に伴う
「引退の花道」を作ってくれたのだった。この講演の最初に私は次のようなジョークをい
った。
「10年前の第1回目のこのミーテングの時には、私の講演は最後の日の、一番最後で
した。そのとき、オルガナイザーの Nancy Kleckner 博士が『参加者が集会の途中で帰
ってしまわないように Sotaの講演を最後(トリ)にした』といいました。今回は私が集会の
最初に話しますが、皆さん、明朝、家に帰ってしまわないでください」(笑)
「oriC プラスミドを分離したときに、なぜ複製開始の研究をしないで、分配の研究を始
めたのかとよく人に尋ねられます。分配機構についての論文は当時ほとんどありませ
んでしたが、複製機構についての論文は既に何百もありました。私は英語で自分の論
文を書くのは大好きですが、人の書いた英語の論文を読むことは嫌いなのです」(大
笑)
引退するころになってやっと国際集会においてこのようなジョークを英語で言えるよう
になった。
この講演で「英語の論文を読むのが嫌いだ」とはいったのが、実際には Annual
Review of Biochemistry や Annual Review of Genetics などにいくつか総説を英文で書
いたときには、たくさんの英文論文を読まなければならなかった。総説を執筆するとき

61
には、1年ほど前から関連論文を集めて数百の論文を読んだが、現在のようにネットで
関連論文を検索することができなかったので論文を集めるだけでも大変な作業であっ
た。そして全ての論文を網羅して、それぞれを公平に評価して紹介することはなかな
か難しいことだった。そのために自分が研究をする時間を相当圧迫されてしまった。し
かし、総説の執筆はこの研究分野の全体像を知るためには良い勉強になったし、世界
の多くの研究者と友達になることにも役立った。
一人の人間が一生に出来ることはそう多くはないので、自分の持ち味を発揮できる
研究テーマを探り当てて、そこに特化していく覚悟がいるのだと私は常々考え研究を
続けてきた。そしてそのようにして一忚目的は達成したのではないかと現在考えてい
る。

62
アペンディクス——昆虫の研究
オオゴマシジミの生態
実は私は中学生・高校生のときには熱狂的な昆虫尐年で、昆虫採集のために郷里
の新潟県南魚沼郡の野山を走り回っていた。採集した昆虫の標本を作るだけではなく、
野外から採集してきたチョウの卵や幼虫を飼育して観察することが好きだった。そして
それらが生育して行く段階を正確な細密画に描き記録した。私は、当時その生活史が
不明だった珍蝶のオオゴマシジミ Maculinea arionides takamukui Matsumura の生活史
を是非自分で明らかにしたいと思っていた。中学生のとき私はアリの人工巣を使って、
このチョウの幼虫が特殊なアリの巣の中でアリの幼虫を食べて生育することを発見した。
このチョウは亜高山帯の渓谷に群生しているカメバヒキオコシやクロバナヒキオコシの
花穂に産卵して、孵化した幼虫はこれらの花を食べているが、4齢になったときに特殊
のアリが自分の巣に運び込んでしまい、アリの巣の中でこのチョウの幼虫は肉食に変
化してアリの幼虫を食べて越冬することがわかった。この研究結果は2編の論文として
昆虫雑誌「新昆虫」(1952, 1955)で発表した。一方、当時まだ生活史が良く分かって
いなかったスギタニルリシジミ(Celastrina sugitanii)やベニヒカゲ (Erebia niphonica
niphonica)の生活史も調べて新潟県の「越佐昆虫同好会」の会誌 (1952,1952) で発
表した。
大学生になってからは、チョウの世界からは遠のいていたが、熊本大学教授を定年
退職する寸前には、50 年間も大切に保存しておいた中学時代にオオゴマシジミの飼
育に使ったアリの乾燥標本をアリの分類の専門家緒方和夫博士(九州大学)に送って
種名を同定してもらった。現代の分類ではシワクシケアリ Myrmica kotokui Forel である
ことを教えていただき、その同定結果を論文 (2001, 2003) にして発表した。
一方、多くの昆虫図鑑や単行本には「オオゴマシジミの宿主アリはヤマアシナガアリ」
と書いてあるが、それは山口進著の写真集「五麗蝶譜」(講談社)の「オオゴマシジミの
宿主アリはヤマアシナガアリ Aphaenogaster japonica (Worker)である」という記載を引
用したものである。しかし、この山口氏のアリの同定は誤同定であることを、私は私の
論文(2001、2003)の中で指摘した。私の鑑定では、「五麗蝶譜」に載っている生態写
真に写っているアリは全てシワクシケアリであり、ヤマアシナガアリではないのである。
「五麗蝶譜」に載っている乾燥標本のアリだけがヤマアシナガアリである。この私の鑑
定が正しいことは、2002 年にアリ分類の専門家である久保田政雄博士(山口氏のアリ
の同定者である)に国立遺伝学研究所でお会いしたとき確認していただいた。私が研

63
究したオオゴマシジミの生息地にはシワクシケアリとヤマアシナガアリの両方が住んで
いたが、私はヤマアシナガアリをオオゴマシジミの飼育には使わなかった。おそらく山
口氏は誤って採集したヤマアシナガアリを久保田博士に送って同定してもらったので
あろう。その後、白水隆博士は昆虫雑誌「むし」(2003)の報文でこの私の論文を引用
して、白水博士自身が「五麗蝶譜」の監修者であったにもかかわらずこの宿主アリの誤
同定に気が付かなかったことを謝罪した。
私はオオゴマシジミの生態研究時に使ったアリの標本を一度 55 年前に安松京三博
士(九州大学)に同定してもらったことがある。「Myrmica rubra (Linne)の1亜種である」
と同定していただき、その同定結果を私は論文 (1955) に記載した。緒方博士による
と、日本産のアリの分類学は当時未発達な状態で、シワクシケアリの学名はいろいろ
変転しており、Myrmica rubra の1亜種とされていたときもあったということである。この
一件からも分かるように、分類学は時代とともに進化するので、証拠になる標本を長期
間保存しておくことは大切なことである。
サナギの保護色
熊本大学教授を定年退職してからは、京都大学で研究員として大腸菌の研究を続
けていたが、夜は自宅で趣味のチョウの研究を再開した。研究のテーマは、中学時代
に一度研究を試みたが満足できる成果が上げられなかったアゲハチョウ(ナミアゲハ、
Papilio xuthus Linne)のサナギの保護色についての疑問の解明である。アゲハチョウ
のサナギには緑色のものと褐色のものがある。自然界ではミカンやカラタチの生きた緑
色の枝では緑色のサナギが見つかり、枯れた褐色の枝では褐色のサナギが見つかる。
これは鳥等の捕食者に対する保護色となっていると考えられる。アゲハチョウのサナギ
の色については先人の研究があり、日高敏隆博士は生きた植物の匂いで緑色のサナ
ギになると述べている。また本田計一博士やA. G. Smith博士は、サナギになる場所の
素材や幅や曲面および湿度等が複雑に関与していることを報告していた。しかし、自
然界で実際に保護色を決める最も重要な環境要因は何かという問題は未解決であっ
た。
私は食草のカラタチ (Poncirus trifoliate) から多数のアゲハチョウの卵や幼虫を採
集して来て、プラスチック容器のなかでカラタチの葉を食べさせて飼育し、蛹化の過程
で種々の環境要因について実験を繰り返し、植物の匂いやサナギになる場所の色は
サナギの色彩決定には関係しないことを証明した。そして強い照度 (100 ルクス) の光
や 100%湿度は褐色のサナギの出現頻度を抑制させ、一方、サナギになる場所からの

64
触覚刺激は褐色のサナギの出現頻度を増加させることを明らかにした。光沢紙で作っ
た台紙の上では、台紙の色彩とは無関係に緑色のサナギになり、サンドペーパーで作
った台紙の上では全て褐色のサナギになった。最後の脱糞から幼虫の皮を脱いでサ
ナギになるまでの過程を、帯糸(胸部の回りにかける絹糸)形成の前と後に分けて、環
境要因の影響を調べた結果、帯糸前ばかりではなく帯糸後も環境の影響を受けて色
彩決定が行われることが明らかになった。
自然光の元で飼育したアゲハチョウの幼虫を多数観察した結果、日暮れに最後の脱
糞をして、真夜中に帯糸をかけて前蛹になりそのまま次の昼間の自然光を浴びて夜に
なってから脱皮してサナギになることがわかった。すなわち、自然界では幼虫の全ての
個体がサナギになる過程で等しく昼間の自然光を充分受けているので、サナギになる
場所から受ける触覚刺激量の違いが色彩決定に最も重要な環境要因であるということ
が私の結論である。そして実際に自然光の下で光沢紙とサンドペーパーの台紙の実
験をおこない、このことを証明した。頭部に生えている機械感覚毛や触角から入った
触覚刺激が脳に伝えられて、触覚刺激が或る閾値を越えるとメラニン色素誘導のため
ホルモン分泌が誘起されるものと考えられる。光の照度や湿度はこの閾値を変化させ
る要因である。生きた枝は緑色でツルツルしており、枯れた枝は褐色でザラザラしてい
るので、このメカニズによって自ずと保護色のサナギになれるのである。
日高敏隆博士が生きた植物の匂いがサナギの色彩決定に関与していると考えた暗
箱内の実験は、生きた植物から発散された水蒸気の効果を誤解したものであろう。ま
た、暗箱での実験は、自然界の条件を反映した実験とはいえないのである。一方、本
田計一博士がおこなったサナギになる場所の棒の幅や曲面が影響するという実験結
果は、幼虫が棒から受ける触覚刺激の量の大小によって説明することができる。すな
わち、5齢幼虫の頭部幅は約 5 mm であるから、直径 1.3 mm の針金上では頭部の機
械感覚毛やアンテナ(触角)が針金に触る頻度が著しく低く、触覚刺激が尐ないので
緑色のサナギが多くなるのである。
一方、アオスジアゲハ (Graphium sarpedon nipponum Fruhstorfer) はアゲハチョウと
同じくアゲハチョウ科に属しているが、アゲハチョウと全く異なる環境要因によってサナ
ギの色が決定されることを発見した。アオスジアゲハの幼虫は白い台紙(光沢紙)の上
では黄緑色のサナギになり、黒い台紙の上では赤褐色のサナギになり、灰色の台紙
の上では中間色(緑がかった灰色)になることが分かった。一方、暗箱の中では、黄緑
色のサナギになる。また、側単眼を全て黒いアクリル絵の具で塗りつぶすと、黒い台紙
の上でも緑色のサナギになった。種々の実験結果から、アオスジアゲハでは幼虫の背

65
面から受ける光の照度と腹側から受けるサナギになる場所からの反射光の照度との
「照度差」が色彩決定に関与していることが分かった。背後からの照度が 100 ルクスの
条件下では、台紙から腹側に受ける反射光との照度差が40ルクス以上のときに、赤褐
色のサナギになる。
アオスジアゲハは野外では食草のクスノキ (Cinnamomum camphora) の葉裏でサナ
ギになることが観察されるが、背側から受ける空からの散乱光の照度(約 10,000 ルク
ス)と腹側から受ける葉からの反射光と透過光の照度の和がほぼ等しくなるために黄
緑色のサナギになるものと推測される。すなわち、強い自然光のもとでは、背側からと
腹側からとの光の「照度比」が重要な働きをしているものと推測される。そしてアオスジ
アゲハではサナギになる場所の素材からの触覚刺激は色彩決定に関係しないし、植
物の匂いも関係しないことが分かった。
自然光下で飼育した幼虫を使って、帯糸形成直後の明所と暗所の切り替え実験を
おこなうと、アオスジアゲハの幼虫は最後の脱糞から帯糸形成までの間に受ける光に
よってサナギの色彩決定をおこなうことが分かった。このことは、朝にアオスジアゲハの
幼虫が最後の脱糞をして、昼のうちに帯糸形成をおこなうという生態と矛盾しない。
アオスジアゲハの赤褐色のサナギは自然界では珍しく、それがどのような環境で見
つかったのか不明である。私はこの問題を解決するために、食草のクスノキの紅葉し
た赤い葉(クスノキは常緑樹であるが、散発的に古い葉が紅葉して落下する性質があ
る)で実験をおこなったところ、全て赤褐色のサナギになった。幼虫は頭部にある6対
の側単眼で光を感受するが、側単眼の中には3種の視細胞が存在することが知られて
いる。それはそれぞれ紫外線、青、緑の光を感じる視細胞であるが、赤を感じる視細
胞は存在しない。したがって、赤い葉からの赤色の反射光を幼虫は感知できず、黒い
台紙のときと同じように赤褐色のサナギになるものと考えられる。幼虫はサナギになると
き葉と小枝の間に糸を張って固定する性質があるので、その葉が枯れても落葉しな
い。
この私のアオスジアゲハにおける色彩決定機構の発見は世界で初めてのものであり、
アオスジアゲハが含まれる Graphium 属の中の他の種における研究は報告されていな
い。
ミトコンドリアの DNA の塩基配列の比較から、アゲハチョウとアオスジアゲハは約
8000 万年前に祖先種から分岐して進化してきたことが知られている。アゲハチョウ科
の中で比較的に原始的な性質を残しているギフチョウ(Luehdorfia japonica)やウスバ
シロチョウ(Parnassius glacialis)は落葉の下で褐色のサナギになる。アゲハチョウやア

66
オスジアゲハは木の枝や葉の上でサナギを作るようになり、緑色や褐色の保護色にな
る性質を獲得したものと考えられる。そして、その進化の過程で両者はそれぞれ上に
述べたような異なる保護色決定機構を進化させてきたのであろう。これらの研究結果
は 2 編の英文論文にして昆虫生理の専門誌 Journal of Insect Physiology (2005, 2006)
に発表した。また、この研究の成果を「蝶・サナギの謎」(トンボ出版)という日本語の単
行本にして出版した。この本で私の説の根拠となった実験結果を沢山のカラー写真や
イラストを使って中学生にも分かるように平易に書いた。アマチュアでも普通種を使っ
てこのように面白い自由研究ができることを読者に伝えたいと考えて作成した本であ
る。
油絵
未知のことを解明する醍醐味を知ったのは、中学生時代のオオゴマシジミの生態の
解明であった。このことが科学の道に進む私の原点になったのだと今では考えている。
ついでに、昆虫研究の趣味の他に、もう一つ私の趣味を紹介しておく。それは油絵を
描くことである。油絵の描き方は7才のころ父から教わった。父は開業医であったが、
沢山の趣味を持っていて油絵も描いていたのである。小学生のころから私の描いた油
絵はいろいろな展覧会で賞を頂き、知事賞を頂いたこともあった。私は絵の才能を昆
虫研究においても、昆虫の彩色細密画や点描画に発揮していた。
中学生時代に私は父に将来のことについて相談したことがある。私が「動物図鑑等
の絵を描く挿絵画家になりたい」と言うと、父は「それはだめだ」といった。「壯太は挿絵
画家にはなれるが、挿絵画家は中途半端だ。挿絵画家は科学者の下働きをすること
になる。なるのならば、科学者になれ。絵を描きたいのならば、ピカソのような大芸術家
になれ」と父は言った。いくら何でもピカソのような大芸術家にはなれないが、科学者
の方ならなれるかもしれないと私はそのとき思った。幸いなことには、父は「アインシュ
タインのような大科学者になれ」とは言っていなかったのだ。そういう訳で、私は「普通
の科学者」になったのだった。
閑話休題。分子生物学の研究をやっている間は忙しくてとても油絵を描いているよう
な暇はなく、油絵を描きたいという欲望が蓄積していた。2008 年に京都大学の研究員
を退職してからこの欲望が爆発して、自宅で油絵を描きまくっている。1 年間に 70 枚
も描いた。芸術はバクハツなのだ。2009 年には京都市繁華街のギャラリーマロニエで
油絵の個展を開き、その後は、京都市美術館で開催される種々の美術団体の展覧会
に忚募して入選している。2011 年には郷里の新潟県南魚沼市の美術館で、父との親

67
子展「平賀洗一・壯太の昆虫と油絵展」を開催する。市役所に寄付した父と私が採集
した昆虫標本や当時の昆虫研究ノートも展示する。子供時代の夢が実現しそうであ
る。
(2011 年3月記)

68
英文論文リスト
1. Mutations and linkage groups in Japanese strains of the housefly. M. Tukamoto, Y. Baba,
and S. Hiraga. Japanese J. Genetics 36: 168-174 (1961).
2. Tryptophan metabolism in eye-color mutants of the housefly. S. Hiraga. Japanese J.
Genetics 39: 240-253 (1964).
3. Regulation of synthesis of alkaline phosphatase by deoxyribonucleic acid synthesis in a
constitutive mutant of Bacillus subtilis. S. Hiraga. J. Bacteriol. 91: 2192-2199 (1966).
4. Nucleoside monophosphokinases of Escherichia coli. S. Hiraga, and Y. Sugino. Biochem.
Biophys. Acta 114: 416-418 (1966).
5. A deoxythymidine kinase-deficient mutant of Escherichia coli. I. Isolation and some
properties. S. Hiraga, K. Igarashi, and T. Yura. Biochem. Biophys. Acta. 145: 41-51
(1967).
6. A deoxythymidine kinase deficient mutant of Escherichia coli. II. Mapping and
transduction studies with phage 80. K. Igarashi, S. Hiraga, and T. Yura. Genetics 57:
643-654 (1967).
7. A new regulatory gene for the tryptophan operon of Escherichia coli. S. Hiraga, K. Ito, K.
Hamada, and T. Yura. Biochem. Biophys. Res. Comm. 26(5): 522-527 (1967).
8. Transcription of the tryptophan operon of Escherichia coli in vitro. I. Detection and
quantitative determination of specific RNA. T. Yura, M. Imai, T. Okamoto, and S.
Hiraga. Biochem. Biophys. Acta 159: 494-510 (1968).
9. 5-Methyltryptophan-resistant mutants linked with the arginine G marker in Escherichia
coli. S. Hiraga, K. Ito, T. Matsuyama, H. Ozaki, and T. Yura. J. Bacteriol. 96(5):
1880-1881 (1968).
10. Operator mutants of the tryptophan operon in Escherichia coli. S. Hiraga. J. Mol. Biol.
39: 159-179 (1969). S. Hiraga. J. Mol. Biol. 39: 159-179 (1969).
11. Temperature-sensitive repression of the tryptophan operon in Escherichia coli. K. Ito, S.
Hiraga, and T. Yura. J. Bacteriol. 99(1): 279-286 (1969).
12. Tryptophanyl transfer RNA synthetase and expression of the tryptophan operon in the
trpS mutants of Escherichia coli. K. Ito, S. Hiraga, and T. Yura. Genetics 61: 521-538
(1969).
13. Hyper-labile messenger RNA in polar mutants of the tryptophan operon of Escherichia
coli. S. Hiraga, and C. Yanofsky. J. Mol. Biol. 72:103-110 (1972).

69
14. Normal repression in a deletion mutant lacking almost the entire operator-proximal gene
of the tryptophan operon of E. coli. S. Hiraga, and C. Yanofsky. Nature New Biol. 237:
47- 49 (1972).
15. Inhibition of the progress of transcription on the tryptophan operon of Escherichia coli.
S. Hiraga, and C. Yanofsky, J. Mol. Biol. 79: 339-349 (1973).
16. Initiation of DNA replication in Escherichia coli: I. characteristics of the initiation
process in dnaA mutants. S. Hiraga, and T. Saitoh. Mol. Gen. Gent. 132: 49-62 (1974).
17. Initiation of DNA replication in Escherichia coli: II. Effect of rifampicin on the
resumption of replication of F episome from a non-permissive to a permissive
temperature. S. Hiraga, and T. Saitoh. Mol. Gen. Genet. 137: 239-248 (1975).
18. Initiation of DNA replication in Escherichia coli: III. Genetic analysis of the dna mutant
s Mol. Gen. Gent. 137: 249-261 (1975).
19. F deoxyribonucleic acid transferred to recipient cells in the presence of rifampicin. S.
Hiraga, and T. Saitoh. J. Bacteriol. 121(3):1000-1006 (1975).
20. F deoxyribonucleic acid superinfected into phenocopies of donor strains. T. Saitoh, and
S. Hiraga. J. Bacteriol. 121(3): 1007-1013 (1975).
21. Novel F prime factors able to replicate in Escherichia coli Hfr strains. S. Hiraga. Proc.
Natl. Acad. Sci. USA 73(1): 198-202 (1976).
22. A mutant of Escherichia coli incapable of supporting vegetative replication of F-like
plasmids. C. Wada, S. Hiraga, and T. Yura. J. Mol. Biol. 108: 25-41 (1976).
23. Replication of Fpoh+ plasmid in mafA mutants of Escherichia coli defective in plasmid
maintenance. C. Wada, T. Yura, and S. Hiraga. Molec. Gen. Genet. 152: 211-217 (1977).
24. Plasmid carrying the replication origin of the Escherichia coli chromosome. S. Hiraga.
In Microbiology. pp. 65-67, American Society for Microbiology.
25. Bacteriophage carrying the Escherichia coli chromosomal region of the replication
origin. T. Miki, S. Hiraga, T. Nagata, and T. Yura. Proc. Natl. Acad. Sci. USA 75(10):
5099-5103 (1978).
26. Genetic mapping of the chromosomal replication origin of Salmonella typhimurium. K.
Joh, and S. Hiraga. J. Bacteriol. 138(2): 297-304 (1979).
27. Cloning and physical mapping of the dnaA region of the Escherichia coli chromosome.
T. Miki, M. Kimura, S. Hiraga, T. Nagata, and T. Yura. J. Bacteriol. 140(3): 817-824
(1979).
28. Conditionally lethal amber mutants in the dnaA region of the Escherichia coli

70
chromosome that affect chromosome replication. M. Kimura, T. Miki, S. Hiraga, T.
Nagata, T. Yura. J. Bacteriol. 140(3): 825-834 (1979).
29. Copy-number mutants of the plasmid carrying the replication origin of the Escherichia
coli chromosome: evidence for a control region of replication. T. Ogura, T. Miki, and S.
Hiraga. Proc. Natl. Acad. Sci. USA 77(7): 3993-3997 (1980).
30. Isolation and characterization of Escherichia coli dnaA amber mutants. T. Kimura, T.
Miki, and S. Hiraga. J. Bacteriol. 144: 649-655 (1980).
31. In vitro replication of recombinant plasmids carrying chromosomal segments of
Xenopus laevis. S. Hiraga, T. Sudo, M. Yashida, H. Kubota, and H. Ueyama. Proc. Natl.
Acad. Sci. USA 79: 3697-3701 (1982).
32. Partition mechanism of F plasmid: two plasmid gene-encoded products and a cis-acting
region are involved in partition. T. Ogura, and S. Hiraga. Cell 32: 351-360 (1983).
33. Mini-F plasmid genes that couple host cell division to plasmid proliferation. T. Ogura,
and S. Hiraga. Proc. Natl. Acad. Sci. USA 80: 4784-4788 (1983).
34. A novel type of E. coli mutants with increased chromosomal copy number. M. Tanaka,
H. Ohmori, and S. Hiraga. Mol. Gen. Genet. 192: 51-60 (1983).
35. Initiation site of deoxyribonucleotide polymerization at the replication origin of the
Escherichia coli chromosome. S. Hirose, S. Hiraga, and T. Okazaki. Mol. Gen. Genet.
189: 422-431(1983).
36. Novel form of DNA polymerase associated with DNA primase activity of vertebrates. T.
Yagura, T. Kozu, T. Seno, M. Saneyoshi, S. Hiraga, and H. Nagano. J. Biol. Chem.
258(21): 13070-13075 (1983).
37. Molecular cloning of the phosphoenolpyruvate gene, ppc, of Escherichia coli. H. Sabe,
T. Miwa, T. Kodaki, K. Izui, S. Hiraga, and H. Katsuki. Gene 31: 279-283 (1984).
38. Prophage induction caused by mini-F plasmid genes. H. Mori, T. Ogura, and S. Hiraga.
Mol. Gen. Genet. 196: 185-193 (1984).
39. Mechanisms essential for inheritance of mini-F plasmid. S. Hiraga, T. Ogura, H. Mori,
and M. Tanaka. In Plasmids in Bacteria. pp. 469-487, Plenum Publishing Corporation
(1985).
40. Negative control of oriC plasmid replication by transcription of the oriC region. M
Tanaka, and S. Hiraga. Mol. Gen. Genet. 200:21-26 (1985).
41. Effects of the ccd function of the F plasmid on bacterial growth. A. Jaffé, T. Ogura, and
S. Hiraga. J. Bacteriol. 163(3): 841-849 (1985).

71
42. F plasmid ccd mechanism in Escherichia coli. S. Hiraga, A. Jaffé, T. Ogura, H. Mori,
and H. Takahashi. J. Bacteriol. 166(1): 100-104 (1986).
43. Mechanisms of stable plasmid inheritance. S. Hiraga. Adv. Biophys. 21: 91-103 (1986).
44. Structure and function of the F plasmid genes essential for partitioning. H. Mori, A.
Kondo, A. Ohshima, T. Ogura, and S. Hiraga. J. Mol. Biol. 192: 1-15 (1986).
45. Partitioning of the F plasmid: overproduction of an essential protein for partition
inhibits plasmid maintenance. N. Kusukawa, H. Niki, A. Kondo, and S. Hiraga. Mol.
Gen. Genet. 208: 365-372 (1987).
46. Molecular cloning of a human apoC-III variant: Thr74-Ala74 mutation prevents
o-glycosylation. H. Maeda, R. R. K. Hashimoto, T. Ogura, S. Hiraga, and H. Uzawa. J.
Lipid Res. 28: 1405-1409 (1987).
47. Minicell-forming mutants of Escherichia coli: production of minicells and anucleate
rods. A. Jaffé, R. D’Ari, and S. Hiraga. J. Bacteriol. 170(7): n3094-3101 (1988).
48. Chromosomal genes essential for stable maintenance of the mini-F plasmid in
Escherichia coli. H. Niki, C. Ichinose, T. Ogura, H. Mori, M. Morita, M. Hasegawa, N.
Kusukawa, and S. Hiraga. J. Bacteriol. 170(11): 5272-5278 (1988).
49. Penicillin-binding protein 2 is essential in wild-type Escherichia coli but not in lov or
cya mutants. T. Ogura, P. Bouloc, H. Niki, R. D’Ari, S. Hiraga, and J. Jaffé. J. Bacteriol.
171(6): 3025-3030 (1989).
50. Purification and characterization of SopA and SopB proteins essential for F plasmid
partitioning. H. Mori, Y. Mori, C. Ichinose, H. Niki, T. Ogura, A. Kato, and S. Hiraga. J.
Biol. Chem. 264(26): 15535-1554 (1989).
51. Identification of human apolipoprotein E variant gene: apolipoprotein
E7(Glu244,245-Lus244,245). H. Maeda, H. Nakamura, S. Kobori, M. Okada, H. Mori,
H. Niki, T. Ogura, and S. Hiraga. J. Biochem. 105: 51-54 (1989).
52. Molecular cloning of a human apolipoprotein E variant: E5(Glu3-Lys3). H. Maeda, H.
Nakamura, S. Kobori, M. Okada, H. Niki, T. Ogura, and S. Hiraga. J. Biochem. 105:
491-493 (1989).
53. Purification and characterization of the sopB gene product which is responsible for
stable maintenance of mini-F plasmid. E. Watanabe, S. Inamoto, M.-H. Lee, S. U. Kim,
T. Ogura, H. Mori, S. Hiraga, M. Yamasaki, and K. Nagai. Mol. Gen. Genet. 218:
431-436 (1989).
54. Chromosome partitioning in Escherichia coli: novel mutants producing anucleate cells.

72
S. Hiraga, H. Niki, T. Ogura, C. Ichinose, H. Mori, B. Ezaki, and A. Jaffé. J. J.
Bacteriol. 171(3): 1496-1505 (1989).
55. Involvement of DnaK protein in mini-F plasmid replication: temperature-sensitive seg
mutants are located in the dnaK gene. B. Ezaki, T. Ogura, H. Mori, H. Niki, nad S.
Hiraga. Mol. Gen. Genet. 218:183-189 (1989).
56. The complete cDNA sequence for the premature form of growth hormone of the
flounder Paralichthys olivaceus. H. Mori, T. Kimura, T. Tsunenari, T. Ogura, H. Niki, B.
Ezaki, K. Shigesada, and S. Hiraga. Nucl. Acids Res. 17: 3977 (1989).
57. Nucleotide sequence of the tolC gene of Escherichia coli. H. Niki, R. Imamura, and T.
Ogura, and S. Hiraga. Nucl. Acids Res. 18(18): 5547.
58. Possible involvement of the ugpA gene product in the stable maintenance of mini-F
plasmid in Escherichia coli. B. Ezaki, H. Mori, and S, Hiraga. Mol. Gen. Genet.
223:361-368 (1990).
59. New topoisomerase essential for chromosome segregation in E. coli. J. Kato, Y.
Nishimura, R. Imamura, H. Niki, S. Hiraga, and H. Suzuki. Cell 63: 393-404 (1990).
60. Maintenance of plasmids in HU and IHF mutants of Escherichia coli. T. Ogura, H. Niki,
Y. Kano, F. Imamoto, and S. Hiraga. Mol. Gen. Genet. 220: 197-203 (1990).
61. Linear multimer formation of plasmid DNA in Escherichia coli. H. Niki, and S. Hiraga.
Mol. Gen. Gent. 224: 1-9 (1990).
62. Identification and characterization of gyrB mutants of Escherichia coli that are defective
in partitioning of mini-F plasmids. T. Ogura, H. Niki, H. Mori, M. Morita, M.
Hasegawa, C. Ichinose, and S. Hiraga. J. Bacteriol. 172(3): 1562-1568 (1990).
63. Positioning of replicated chromosomes in Escherichia coli. S. Hiraga, T. Ogura, H. Niki,
C. Ichinose, and H. Mori. J. Bacteriol. 172(1): 31-39 (1990).
64. Partitioning of nucleoids. S. Hiraga. Res. Microbiol. 141: 50-56 (1990).
65. Partitioning of a mini-F plasmid into anuclete cells of the mukB null mutant. B. Ezaki, T.
Ogura, H. Niki, and S. Hiraga. J. Bacteriol. 173(20): 6643-6646 (1991).
66. Control of chromosome partition in bacteria. S. Hiraga. pp. 47-60. In Control of Cell
Growth and Division, Japn. Sci. Soc. Press.
67. Structure and function of the ftsH gene in Escherichia coli. T. Ogura, T. Tomoyasu, T.
Yuki, S. Morimura, K. J. Begg, W. D. Donachie, H. Mori, H. Niki, and S. Hiraga. Res.
Microbiol. 142: 279-282 (1991).
68. Mutants defective in chromosome partitioning in E. coli. S. Hiraga, H. Niki, R.

73
Imamura, T. Ogura, K. Yamanaka, J. Feng, B. Ezaki, and J. Jaffé. Res. Microbiol. 142:
189-194 (1991).
69. The new gene mukB codes for a 177 kd protein with coiled-coil domains involved in
chromosome partitioning of E. coli. H. Niki, A. Jaffé, R. Imamura, T. Ogura, and S.
Hiraga. EMBO J. 10(1): 183-193 (1991).
70. Chromosome and plasmid partition in Escherichia coli. S. Hiraga. Annu. Rev. Biochem.
61: 283-306 (1992).
71. Escherichia coli mutant Y16 is a double mutant carrying thermosensitive ftsH and ftsI
mutations. K. Begg, T. Tomoyasu, W. D. Donachie, M. Khattar, H. Niki, K. Yamanaka,
S. Hiraga, and T. Ogura. J. Bacteriol. 174: 2416-2417 (1992).
72. E. coli MukB protein involved in chromosome partition forms a homodimer with a
rod-and-hinge structure having DNA binding and ATP/GTP binding activities. EMBO J.
11(13): 5101-5109 (1992).
73. Characterization of high molecular weights of complexes and polymers of cytoplasmic
proteins in Escherichia coli. R. Imamura, H. Niki, M. Kitaoka, K. Yamanaka, T. Ogura,
and S. Hiraga. Res. Microbiol. 143: 741-753 (1992).
74. Chromosome partition in Escherichia coli. S. Hiraga. Curr. Opin. Genet. Dev. 5:
789-801 (1993).
75. Identification and characterization of the smbA gene, a suppressor of the mukB null
mutant of Escherichia coli. K. Yasmanaka, T. Ogura, H. Niki, and S. Hiraga. J.
Bacteriol. 174(23): 7517-7526 (1992).
76. The Escherichia coli FtsH protein is a prokaryotic member of a protein family of
putative ATPase involved in membrane functions, cell cycle control, and gene
expression. T. Tomoyasu, T. Yuki, S. Morimura, H. Mori, K. Yamaanaka, H. Niki, S.
Hiraga, and T. Ogura. J. Bacteriol. 175(5): 1344-1351 (1993).
77. Cell growth and l development controlled by the same essential Escherichia coli gene,
ftsH/hftB. C. Herman, T. Ogura, T. Tomoyasu, S. Hiraga, Y. Akiyama, K. Ito, R. Thomas,
R. D’Ari, and P. Bouloc. Proc. Natl. Acad. Sci. USA 90: 10861-10865 (1993).
78. Topology and subcellular localization of FtsH protein in Escherichia coli. T. Tomoyasu,
K. Yamanaka, K. Murata, T. Suzuki, P. Bouloc, A. Kato, H. Niki, S. Hiraga, and T.
Ogura. J. Bacteriol. 175(5): 1352-1357 (1993).
79. Two mutant alleles of mukB, a gene essential for chromosome partition in Escherichia
coli. K. Yamanaka, T. Mitani, J. Feng, T. Ogura, H. Niki, and S. Hiraga. FEMS

74
Microbiology Letters 123: 27-32 (1994).
80. Multicopy suppressors, mssA and mssB, of an smbA mutation of Escherichia coli. K.
Yamanaka, T. Ogura, E. V. Koonin, H. Niki, and S. Hiraga. Mol. Gen. Gent. 243: 9-16
(1994).
81. Characterization of translucent segments observed in an smbA mutant of Escherichia
coli. K. Yamanaka, T. Ogura, K. Murata, T. Suzaki, H. Niki, and S. Hiraga. FEMS
Microbiology Letters 116: 61-66 (1994).
82. Cloning, sequencing, and characterization of multicopy suppressors of a mukB mutation
in Escherichia coli. K. Yamanaka, T. Mitani, T. Ogura, H. Niki, and S. Hiraga. Mol.
Microbiol. 13(2) 301-312 (1994).
83. New killing system controlled by two genes located immediately upstream of the mukB
gene in Escherichia coli. J. Feng, K. Yamanaka, H. Niki, T. Ogura, and S, Hiraga. Mol.
Gen. Genet. 243: 136-147 (1994).
84. Escherichia coli FtsH is a membrane-bound, ATP-dependent protease which degrades
the heat-shock transcription factor 32. T. Tomoyasu, J. Gamer, B. Buau M. Kanemori,
H. Mori, A. J. Rutman, A. B. Oppenheim, T. Yura, K. Yamanaka, H. Niki, S. Hiraga,
and T. Ogura. EMBO J. 14(11): 2551-2560 (1995).
85. Characterization of the smtA gene encoding an S-adenosylmethionine-dependent
methyltransferase of Escherichia coli. K. Yamanaka, T. Ogura, H. Niki, and S, Hiraga.
FEMS Microbiology Letters 133: 59-63 (1995).
86. RNase E polypeptides lacking a carboxyl-terminal half suppress a mukB mutation in
Escherichia coli. M. Kido, K. Yamanaka, T. Mitani, H. Niki, T. Ogura, and S. Hiraga. J.
Bacteriol. 178(13): 3917-3925 (1996).
87. Identification of the cpdA gene encording cyclic 3’, 5’-adenosine monophosphate
phosphodiesterase in Escherichia coli. R. Imamura, K. Yamanaka, T. Ogura, S. Hiraga,
N. Fujita, A. Ishihama, and H. Niki. J. Biol. Chem. 271(41): 25423-25429 (1996).
88. Carboxyl terminal region of the MukB protein in Escherichia coli is essential for DNA
binding activity. A. Z. Saleh, K. Yamanaka, H. Niki, T. Ogura, M. Yamazoe, and S.
Hiraga. FEMS Microbiology Letters 143: 211-216 (1996).
89. Identification of two new genes, mukE and mukF, involved in chromosome partitioning
in Escherichia coli. K. Yamanaka, T. Ogura, H. Niki, and S. Hiraga. Mol. Gen. Gent.
250: 241-251 (1996).
90. Subcellular distribution of actively partitioning F plasmid during the cell division cycle

75
in E. coli. H. Niki, and S. Hiraga. Cell 90: 951-957 (1997).
91. Cell cycle-dependent duplication and bidirectional migration of SeqA-associated
DNA-protein complexes in E. coli. S. Hiraga, C. Ichinose, H. Niki, and M. Yamazoe.
Mol. Cell 1: 381-387 (1998).
92. Polar localization of the replication origin and terminus in Escherichia coli nucleoids
during chromosome partitioning. H. Niki, and S. Hiraga. Genes Dev. 12: 1036-1045
(1998).
93. Autoregulation of the partition genes of the mini-F plasmid and the intercellular
localization of the their products in Escherichia coli. M. Hirano, H. Mori, T. Onogi, M.
Yamazoe, H. Niki, T. Ogura, and S. Hiraga. Mol. Gen. Genet. 257: 392-403 (1998).
94. Subcellular localization of plasmids containing the oriC region of the Escherichia coli
chromosome, with or without the sopABC partitioning system. H. Niki, and S. Hiraga.
Mol. Microbiol. 34 (3): 498-503 (1999).
95. The assembly and migration of SeqA-Gfp function in living cells of Escherichia coli. T.
Onogi, H. Niki, M. Yamazoe, and S. Hiraga. Mol. Microbiol. 31 (6): 1775-1782 (1999).
96. Complex formation of MukB, MukE and MukF proteins involved in chromosome
partitioning in Escherichia coli. M. Yamazoe, T. Onogi, Y. Sunako, H. Niki, K.
Yamanaka, T. Ichimura, and S. Hiraga. EMBO J. 18 (21): 5873-5884 (1999).
97. Bidirectional migration of SeqA-bound hemimethylated DNA clusters and pairing of
oriC copies in Escherichia coli. S. Hiraga, C. Ichinose, T. Ogura, H. Niki, and M.
Yamazoe. Genes Cells 5: 327-341 (2000).
98. Two types of localization of the DNA-binding proteins within the Escherichia coli
nucleoid. T. A. Azam, S. Hiraga, and A. Ishihama. Genes Cells 5: 613-626 (2000).
99. Competition between the replication initiator DnaA and the sequestration factor SeqA
for binding to the hemimethylated chromosomal origin of E. coli in vitro. A. Taghbalout,
A. Landoulsi, R. Kern, M. Yamazoe, S. Hiraga, B. Holland, M. Kohiyama, and A.
Malki. Genes Cells 5: 873-884 (2000).
100. Null mutation of the dam or seqA gene suppresses temperature-sensitive lethality but
not hypersensitivity to novobiocin of muk null mutations. Onogi, M. Yamazoe, C.
Ichinose, H. Niki, and S. Hiraga. J. Bacteriol. 182 (20): 5898-5901 (2000).
101. Dynamic organization of chromosomal DNA in Escherichia coli. H. Niki, Y. Yamaichi,
and S. Hiraga. Genes Dev. 14: 212-223 (2000).
102. Dynamic localization of bacterial and plasmid chromosomes. S. Hiraga. Annu. Rev.

76
Genet. 34: 21-59 (2000).
103. Different location of SeqA-bound nascent DNA clusters and MukF-MukE-MukB
complex in Escherichia coli cells. K. Ohsumi, M. Yamazoe, and S. Hiraga. Mol.
Microbiol. 40 (4): 835-845 (2001).
104. Sister chromosome cohesion of Escherichia coli. Y. Sunako, T. Onogi, and S. Hiraga.
Mol. Microbiol. 42 (5): 1233-1241 (2001).
105. Replication-dependent recruitment of the b-subunit of DNA polymerase III from
cytosolic spaces to replication forks in Escherichia coli. T. Onogi, K. Ohsumi, T.
Katayama, and S. Hiraga. J. Bacteriol. 184 (3): 867-870 (2002).
106. Behavior of sister copies of mini-F plasmid after synchronized plasmid replication in
Escherichia coli cells. T. Onogi, T. Miki, and S. Hiraga. J. Bacteriol. 184 (11):
3142-3145 (2002).
107. Proteolytic activity of YibP protein in Escherichia coli. T. Ichimura, M. Yamazoe, M.
Maeda, C. Wada, and S. Hiraga. J. Bacteriol. 184 (10): 2595-2602 (2002).
108. Amplification of hot DNA segments in Escherichia coli. K. Kodama, T. Kobayashi, H.
Niki, S. Hiraga, T. Oshima, H. Mori, and T. Horiuchi. Mol. Microbiol. 45 (6):
1575-1588 (2002).
109. Genome-wide analysis of deoxyadenosine methyltransferase-mediated control of gene
expression in Escherichia coli. T. Oshima, C. Wada, Y. Kawagoe, T. Ara, M. Maeda, Y.
Masuda, S, Hiraga, and H. Mori. Mol. Microbiol. 45 (3): 673-695 (2002).
110. Cohesion of sister chromosomes in Escherichia coli. Sota Hiraga. Microbiology 148:
3755-3756 (2002).
111. Mutants suppressing novobiocin hypersensitivity of a mukB null mutation. S. Adachi
and S. Hiraga. J. Bacteriol. 185(13): 3690-3695 (2003).
112. Bacterial sex: playing voyeurs 50 years later. M. Kohiyama, S. Hiraga, I. Matic, and M.
Radman. Science 301 (5634): 802-803 (2003).
113. Identification of functional domains of the Escherichia coli SeqA protein. N. Fujikawa,
H. Kurumizaka, M. Yamazoe, S. Hiraga, and S. Yokoyama. Biochem. Biophys. Res.
Commun. 300: 699-705 (2003).
114. Structural and biochemical analysis of hemimethylated DNA binding by the SeqA
protein. N. Fujikawa, H. Kurumizaka, O. Nureki, Y. Tanaka, M. Yamzoe, S. Hiraga, and
S. Yokoyama. Nucleic Acid Res. 32 (1): 82-92 (2004).
115. Sequential binding of SeqA protein to nascent DNA segments at replication forks in

77
Synchronized cultures of Escherichia coli. M. Yamazoe, S. Adachi, S. Kanaya, K.
Ohsumi, and S. Hiraga. Mol. Microbiol. 55: 289-298 (2005).
116. A comparison of MukB homodomer versus MukBEF complex molecular architectures
by electron microscopy reveals a higher-order multimerization. K. Matoba, M. Yamazoe,
K. Mayanagi, K. Morikawa, and S. Hiraga. Biochem. Biophy. Res. Commun. 333:
694-702 (2005).
117. The GTP binding protein Obg homologue ObgE is involved in ribosome maturation. A.
Sato, G. Kobayashi, H. Hayashi, H. Yoshida, A. Wada, M. Maeda, S. Hiraga, K.
Takeyasu, and C. Wada. Genes Cells. 10: 393-408 (2005).
118. Localization of replication forks in wild-type and mukB mutant cells of Escherichia
coli. S. Adachi, M. Kohiyama, T. Onogi, and S. Hiraga. Mol. Genetics Genomics, 274:
264-271 (2005).
119. Subcellular positioning of F plasmid DNA molecules mediated by dynamic
localization of SopA and SopB. S. Adachi, K. Hori, and S. Hiraga. J. Mol. Biol. 356:
850-863 (2006).
120. Dynamic events of sister chromosomes in the cell cycle of Escherichia coli. S. Adachi,
T. Fukushima, and S. Hiraga. Genes Cells 13(2): 181-197 (2008).
121. ATP-induced shrinkage of DNA with MukB protein and MukBEF complex of
Escherichia coli. N. Chen, A. A. Zinchenko, Y. Yoshikawa, S. Araki, S. Adachi, M.
Yamazoe, S. Hiraga, and K. Yoshikawa. J. Bacteriol. 190: 3731-3737 (2008).
その他の論文リスト
DNA複製におけるRNAの関与
平賀 壮太
蛋白質核酸酵素 20(1) p11-25 1975 年 1 月
プラスミド・ウイルスの起源--染色体からのプラスミド新生機構
平賀 壮太
科学 46(12) p730-736 1976 年 12 月
細菌染色体の複製起点をもつプラスミド (DNA複製<特集>) -- (DNA複製の最小単位)
平賀 壮太, 三木 徹
生物物理 18(4) p174-178 1978 年 8 月
複製開始領域の機能的構成--大腸菌染色体の複製開始点 (DNA複製開始<特集>)

78
小椋 光, 平賀 壮太
蛋白質核酸酵素 27(1) p9-19 1982 年 1 月
核DNA (真核生物のDNA合成<特集>) -- (複製開始および組換え部位)
平賀 壮太
蛋白質核酸酵素 28(9) p1033-1043 1983 年 8 月
細菌におけるプラスミドの安定保持機構
平賀 壮太
蛋白質核酸酵素 29(6) p430-443 1984 年 6 月
細菌におけるプラスミドの安定保持機構 (セミナー「凍結・乾燥とその忚用II」,故根井 外喜男
先生追悼号)
平賀 壮太
凍結及び乾燥研究会会誌 31 88-89 1985 年 9 月
大腸菌の染色体分配機構
平賀 壯太, 仁木 宏典
生物物理 30(6) 296-304 1990 年 11 月
細菌における染色体分配と細胞分裂の機構
平賀 壮太
学術月報 44(9) p866-871 1991 年 9
「遺伝子」第5版
平賀 壮太
化学と生物 35(1) 1997 年 1 月
目で見る大腸菌の染色体分配 細胞内の蛋白質やDNAの局在性を調べる新しい手法を開発
平賀 壯太
化学と生物 35(12) 820-822 1997 年 12 月
細菌における分子細胞生物学の新展開--染色体分配と細胞分裂
仁木 宏典, 守家 成紀, 平賀 壯太
蛋白質核酸酵素 45(2) 153-163 2000 年 2 月
大腸菌における染色体動態: MukBタンパク質の細胞内局在
平賀 壯太, 足立 隼
Genes & genetic systems 79(5) 2004 年 10 月
大腸菌染色体動態に関わる局在振動タンパク質群の相互作用ネットワーク
足立隼, 平賀壯太
Genes & genetic systems 81(5) 2006 年 10 月

79
細菌におけるプラスミドと染色体の分配機構 (特集 原核細胞の細胞骨格)
平賀 壯太, 足立 隼
蛋白質核酸酵素 53(13) 1732-1738 2008 年 10 月
趣味の昆虫研究の論文
ベニヒカゲの生態: I. 野外観察と室内観察. 平賀壯太. 越佐昆虫同好会会報 6(1): 1-5
(1952)
カラスシジミ上越国境に産す. 平賀壯太. 越佐昆虫同好会会報 6(3): 89 (1952)
オオゴマシジミの生態:第1報. 平賀壯太. 新昆虫 8(3) : 7-14 (1952).
オオゴマシジミの生態:第2報. 平賀壯太. 新昆虫 8(3) : 7-14 (1955).
オオゴマシジミの生態:第3報. 平賀壯太. 越佐昆虫同好会会報 85: 3-6 (2001).
オオゴマシジミの宿主アリの再同定について. 平賀壯太. やどりが 196: 31-34 (2003).
アリ人工巣によるチョウの飼育法. 平賀壯太. やどりが 197: 31-33 (2003).
Two different sensory mechanisms for the control of pupal protective coloration in
butterflies. S. Hiraga. J. Insect Physiol. 51: 1033-1040 (2005).
Interactions of environmental factors influencing pupal coloration in swallowtail
butterfly Papilio xuthus. S. Hiraga. J. Insect Physiol. 52: 826-838 (2006).
新潟県で採集されたマークオサムシの記録. 平賀壯太・島田久隆. 越佐昆虫同好会会報
99: 83-91 (2008).

80
一般向け著書
南魚沼のファーブル 昆虫少年物語
六日町地区理科教育センター編 (平賀壯太分担執筆)
六日町地区理科教育センター 2001 年 3 月
当時、生活史が不明だったオオゴマシジの生態を、シワクシケアリの人工巣を使って
中学生時代に明らかにした経緯は生き生きと再現されている。 熱血昆虫尐年の活躍
ぶりを書いたこの本は郷里の生徒たちの間で人気がある。
オオゴマシジミ 迎えて舞う
六日町地区理科教育センター編(平賀壯太分担執筆)
六日町地区理科教育センター 2001 年
オオゴマシジミの生態についての平賀の論文が3編収録されており、中学校の先生た
ちの自然観察の記録ももりだくさん。

81
生物の惑星
平賀壯太
太陽書房 2003 年 7 月 ISBN:4901351532
「南魚沼のファーブル 昆虫尐年物語」の発刊記念講演会における著者の講演をまと
めた本で、講演に使った沢山のカラースライドの絵や写真が挿絵として使われている。
講演会での昆虫放談に会場は爆笑。
透明なノート
ひらがそうた
新風舎 2004 年 3 月 ISBN:479744067
著者の描いたイラストと文による詩的なユーモア世界がひろがる大人のための絵本。
「ひらがワールド」にどうぞ。

82
蝶・サナギの謎
平賀 壯太
トンボ出版 2007 年 3 月 ISBN:488716158
著者が子供のころから疑問に思っていたアゲハチョウやアオスジアゲハのサナギの
保護色について、大学教授を定年退官してあと詳細な実験によって、保護色決定の環
境因子を明らかにした驚くべき実験結果を中学生でも分かる平易な文で書いた本。著
者の描いたイラストやカラー写真が豊富に使われた楽しい自由研究の解説書。
カステラのかけら
ひらが そうた
牧歌舎 2008 年 6 月 ISBN:4434118617
著者の描いたイラストと文によってかもしだされるエスプリのきいたユニークなユ
ーモアイッセイ。ユーモアのない人生なんて、泡のでないビールのようなものです。

83
平賀壯太略年譜
1936 新潟県南魚沼郡六日町(現在:南魚沼市)に生まれる
1960 金沢大学理学部生物学科卒業
1965 大阪大学大学院博士課程修了(理学博士)
1965-1985 京都大学ウイルス研究所助手
1970 - 1972
米国スタンフォード大学客員研究員
1985-2002 熊本大学医学研究科教授
2002 - 2008
京都大学生命科学/医学研究科研究員
所属学会
米国微生物学会(アカデミー会員)、日本遺伝学会(名誉会員)、日本分子生物学会、日本
生化学会、日本鱗翅学会、昆虫 DNA 研究会
受賞
日本遺伝学会 木原賞 「大腸菌染色体およびプラスミドの分配機構」(2000 年 11 月)