ESTRUCTURA FUNCION CELULAR OPTICA 2010 - Aula Virtual de … optica/ESTRUCTURA... · desarrollo de...
45
Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce BIOLOGÍA HUMANA Unidad 2: “Estructura y Función Celular” Texto Guía del Curso Biología General para Óptica Laboratorio de Antropología Física y Anatomía Humana – Laboratorio de Técnicas Anatómicas Pontificia Universidad Católica de Valparaíso Edición Primer Semestre 2010 “La mayoría de la imageneología utilizada en las guías y módulos, están tomados y modificados de los textos recomendados para la asignatura y solamente tienen un objetivo docente y bajo ninguna circunstancia lucrativo”
Transcript of ESTRUCTURA FUNCION CELULAR OPTICA 2010 - Aula Virtual de … optica/ESTRUCTURA... · desarrollo de...

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
BIOLOGÍA HUMANA Unidad 2: “Estructura y Función Celular”
Texto Guía del Curso Biología General para Óptica Laboratorio de Antropología Física y Anatomía Humana – Laboratorio de Técnicas
Anatómicas Pontificia Universidad Católica de Valparaíso
Edición Primer Semestre 2010 “La mayoría de la imageneología utilizada en las guías y módulos, están tomados y modificados de los textos recomendados para la asignatura y
solamente tienen un objetivo docente y bajo ninguna circunstancia lucrativo”

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce El primero en observar células fue el científico inglés Robert Hooke en 1665 sobre una preparación de corcho y con un microscopio muy rudimentario. El corcho, observado en aumento, parecía estar constituido por pequeñas celdillas rodeadas por una pared rígida. Fue Robert Hooke el primero en acuñar el término célula para referirse a estas estructuras (La imagen de la portada es uno de los dibujos de Robert Hooke de dos cortes de un trozo de corcho, reproducidos en su Micrographica, publicada en 1665). Sin embargo no llegó a reconocer la verdadera estructura de las células y, en realidad, lo que vio en el corcho no eran células sino sus paredes celulares muertas.
El primero que observó células vivas fue Antón van Leeuwenhoek entre finales del S.XVII y principios del S. XVIII, describió los glóbulos rojos y observó espermatozoides, protozoos e incluso bacterias, todo ello con microscopios muy rudimentarios (en realidad simples lupas) fabricados por él mismo.
Los progresos en el estudio de las células (Citología) dependen muy directamente del desarrollo de la microscopía; por ello hubo que esperar casi dos siglos hasta que se empezó a entender la verdadera significación de las células. En 1831 ya se conocía la existencia del núcleo celular; en 1839 se había descrito minuciosamente el proceso mitótico y en ese mismo año nacía la Teoría celular, atribuida a Schleiden y Schwann, que introducía el concepto fundamental de que la célula es la unidad básica de todos los seres vivos. Este principio fue ampliado finalmente por Virchow en 1859 con lo que la Teoría celular quedó definitivamente establecida. Se puede resumir en tres principios:
1. Todos los organismos vivos están constituidos por una o varias células; la célula es, por tanto, la unidad vital de los seres vivos. 2. Las células son capaces de una existencia independiente; las células son, por tanto, la unidad anatómica y fisiológica de los seres vivos. 3. Toda célula proviene de otra célula ya existente; la célula es, por tanto, la unidad genética de los seres vivos. Hasta finales del S.XIX se consideraba que el tejido nervioso constituía una excepción a la teoría celular ya que se creía que no estaba formado por células independientes sino por una red o retículo de fibras y células interconectadas en la que no era posible diferenciar los elementos celulares, las neuronas. Fue Santiago Ramón y Cajal quien resolvió este problema al enunciar su teoría de la neurona en la que se demostraba que las neuronas eran elementos independientes y constituían la unidad anatómica y funcional del sistema nervioso. De esta manera, la teoría celular quedó definitivamente generalizada a todas las células, tanto vegetales como animales.
No debe interpretarse la célula como un conjunto de partes que, encontrándose dispersas, se han agrupado para formarla. En realidad, las estructuras celulares son el resultado de un proceso de diferenciación en el seno de la propia célula. Cualquier estructura subcelular no puede gozar de vida propia aisladamente, de ahí que se considere a la célula como un sistema con una individualidad propia que las caracteriza como unidades vitales.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce La célula también presenta un aspecto dinámico que sería todo el conjunto de funciones que la célula realiza para perpetuarse y poder mantener esa unidad. Puesto que las células son las unidades vitales de los seres vivos, no es de extrañar que este conjunto de funciones que realizan sean, al mismo tiempo, las características de todo ser viviente:
Funciones de relación: en las células se manifiesta por su capacidad de percibir los estímulos que reciben y reaccionar frente a ellos. De esta manera ponen de manifiesto una excitabilidad o sensibilidad.
Funciones de nutrición: tienen como misión proporcionar alimento para transformarlo en su interior convirtiéndolo en sustancia propia que luego será desintegrada para liberar la energía que se necesita para vivir, expulsando los residuos que quedan después de esta transformación. En todo proceso de nutrición celular se distinguen tres etapas:
•Incorporación del alimento, sea éste sólido, líquido, gaseoso o disuelto. •Transformación del alimento en el interior de las células: el conjunto de estas transformaciones recibe el nombre de metabolismo y son la esencia íntima de la nutrición. Las reacciones químicas del metabolismo pueden ser de dos tipos: unas transforman el alimento en materia propia de la célula y constituyen el anabolismo o asimilación, y otras descomponen las sustancias en otras más sencillas liberando energía y constituyen el catabolismo o desasimilación. •Eliminación de residuos: como consecuencia de la destrucción de los alimentos durante el catabolismo, se generan residuos que son inútiles para la célula y deben ser eliminados. Funciones de reproducción: la reproducción celular se reduce siempre a una división mediante la cual la célula se parte en dos o más trozos, cada uno de los cuales aumenta de tamaño hasta alcanzar el propio de la que le ha dado origen. Durante la división celular desaparece como individuo la célula que se divide, llamada célula madre, y en su lugar aparecen dos o más células hijas. La célula como aparece en la actualidad es el resultado de un proceso de diferenciación que, a través de la evolución, ha conducido a una organización que ha ido adquiriendo complejidad. En consecuencia, según el grado de diferenciación estructural alcanzado, se han establecido dos niveles de organización celular:
Célula procariótica: se caracteriza por su gran simplificación pues en ella faltan muchas de las estructuras que poseen las otras células. El material hereditario se encuentra disperso en el citoplasma, no tienen por tanto núcleo. Este tipo de organización sólo lo presentan las Bacterias.
Célula eucariótica: es la propia de las células típicas. Poseen todas las estructuras como consecuencia de su gran diferenciación. La característica fundamental es que el material hereditario está encerrado en una membrana constituyendo un auténtico núcleo. Pertenecen a este tipo de organización las células que forman los organismos pluricelulares y la mayoría de los unicelulares.
Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce

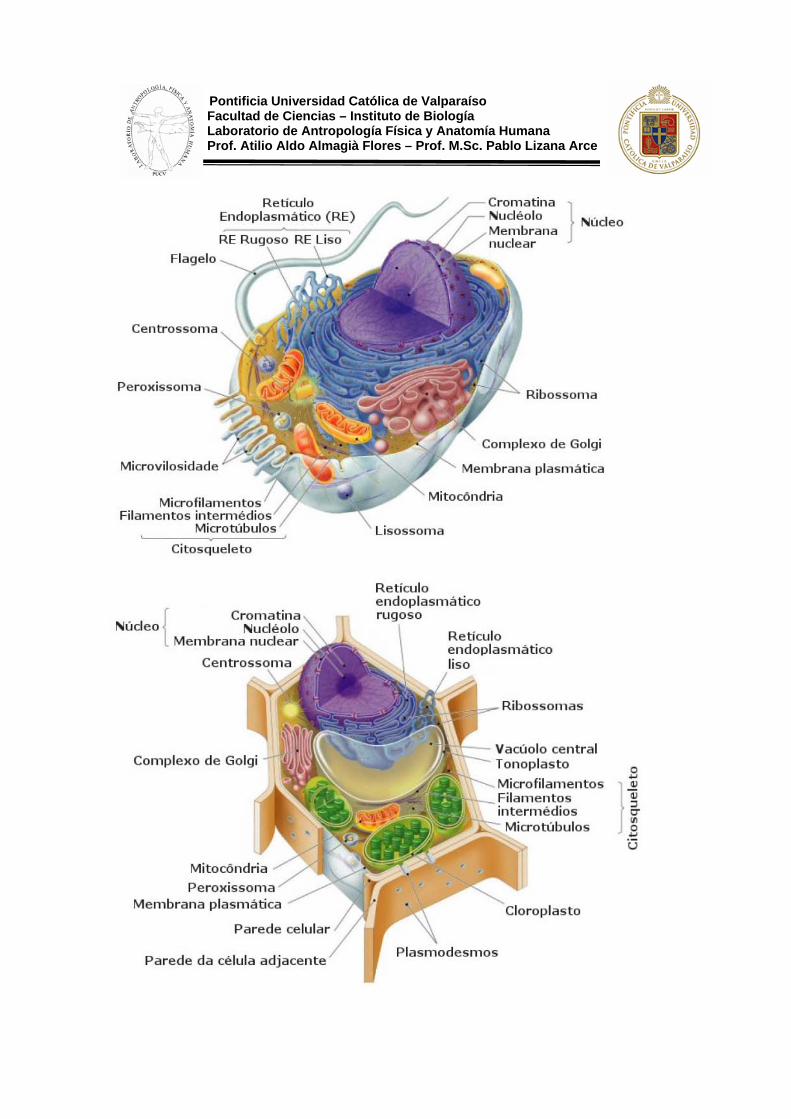
Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce La célula eucariótica se caracteriza porque manifiesta un alto grado de complejidad y de organización. El núcleo está separado del citoplasma mediante la membrana nuclear y contiene ADN asociado a proteínas del tipo de las histonas con el fin de permitir un mejor empaquetamiento.
El citoplasma contiene numerosos orgánulos y sistemas de membranas como las mitocondrias, cloroplastos, retículo endoplasmático, etc. que establecen una red de compartimentos celulares interrelacionados que se reparten el trabajo metabólico, lo que permite una mayor especialización y más eficacia en el desempeño de sus funciones.
Las células eucarióticas animales poseen unos orgánulos llamados centriolos de los que carecen las vegetales. Salvo los centriolos, las células eucarióticas vegetales contienen los mismos orgánulos que las animales, con la diferencia de que poseen una gruesa pared celulósica y tienen gran número de plastos y vacuolas de gran tamaño.
Las células eucarióticas pueden vivir aisladas, en forma de organismos unicelulares (Reino Protistas), o pueden formar parte de organismos pluricelulares. En este caso se organizan en tejidos especializados en ciertas funciones. Esta especialización les lleva a adoptar formas muy diversas de acuerdo con la función que realizan. Este proceso es la diferenciación celular.
Membrana Plasmática
Observada una célula con M.E. (microscopio electrónico) se aprecia una envoltura que, de modo continuo, delimita el territorio celular y actúa como frontera de la célula respecto al medio externo: es la membrana plasmática. Las células realizan el intercambio de sustancias con el medio externo a través de esta membrana en la que además tienen lugar muchas reacciones químicas esenciales para la supervivencia celular.
Se trata de una envoltura continua que por una de sus caras está en contacto con el medio externo y por la otra con el CITOPLASMA celular. En el M.E. se aprecia una estructura formada por tres capas con un espesor total de unos 7 nm.
Para estudiar su composición química hay que separarla del citoplasma. En la actualidad una de las membranas mejor estudiadas es la de los glóbulos rojos ya que es más fácil de aislar porque carecen de núcleo. La membrana de un glóbulo rojo está formada por un 60% de proteínas y un 40% de lípidos. En otros tipos de membranas los porcentajes pueden variar pero por lo general la masa total de proteínas suele ser mayor que la de lípidos.
Lípidos: Los más abundantes son los fosfolípidos, el colesterol y los glucolípidos. Debido a su carácter anfipático (poseen un extremo hidrófobo y uno hidrófilo), cuando se encuentran en medio acuoso se disponen formando una bicapa lipídica. La proporción que corresponde a cada lípido no es igual en cada una de las dos capas. La bicapa lipídica aporta la estructura básica a la membrana y, debido a su fluidez, son posibles muchas de las funciones que desempeñan las membranas celulares. Se dice que la

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce bicapa lipídica es fluida porque se comporta del mismo modo en que lo haría un líquido, es decir, las moléculas pueden desplazarse girando sobre sí mismas o intercambiar su posición con la de otras moléculas situadas dentro de la misma monocapa. Es poco frecuente el intercambio entre moléculas situadas en monocapas distintas.
Proteínas: Las proteínas se sitúan en la bicapa lipídica en función de su mayor o menor afinidad por el agua. Debido a ello se asocian con los lípidos de la membrana de diversas formas:
• Proteínas que atraviesan la membrana. Se llaman proteínas transmembrana. • Proteínas que se introducen en parte dentro de la membrana. • Proteínas situadas en el medio externo a uno u otro lado de la bicapa y unidas a proteínas transmembrana o a lípidos. El lugar que ocupan las proteínas y su mayor o menor grado de unión con los lípidos influyen en la facilidad con que pueden ser separadas del resto de los componentes de la membrana. Según esto se clasifican en dos grupos:
Proteínas Integrales o intrínsecas: están íntimamente asociadas a los lípidos y son difíciles de separar. Constituyen aproximadamente el 70% del total y son insolubles en disoluciones acuosas.
Proteínas periféricas o extrínsecas: están poco asociadas a los lípidos, se aíslan con facilidad y son solubles en disoluciones acuosas.
Al igual que los lípidos, las moléculas de proteína pueden desplazarse por la membrana aunque su difusión es más lenta debido a su mayor masa molecular.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce Glúcidos: Se asocian a los lípidos formando glucolípidos o a las proteínas formando glucoproteínas. Están situados en la cara de la membrana que da al medio extracelular y forma la cubierta celular o glucocálix. Esta disposición de los glúcidos y el hecho de que los lípidos de las dos monocapas sean distintos, da a la membrana plasmática un claro carácter asimétrico.
Representación tridimensional del Modelo de Mosaico fluido
Hoy día, el modelo de membrana que se acepta integra los conocimientos que se poseen sobre la disposición de sus componentes. Dicho modelo fue propuesto por Singer y Nicholson en 1972 y se denomina "Modelo de Mosaico Fluido". Este modelo se basa en 3 premisas:
1.- Los lípidos y las proteínas integrales que forman la membrana constituyen un mosaico molecular. 2.- Los lípidos y las proteínas pueden desplazarse en el plano de la bicapa lipídica. Por ello las membranas son fluidas. 3.- Las membranas son asimétricas en cuanto a la disposición de sus componentes moleculares.
Funciones de la membrana plasmática
En general se encarga de relacionar a los organismos unicelulares con su medio externo o a unas células con otras en el caso de los organismos pluricelulares. No es tan sólo una estructura que sirva para mantener encerrada a la célula e impedir que se escape el contenido de su citoplasma. También está dotada de una gran actividad y desempeña numerosas funciones, como por ejemplo:

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce 1. Recibir y transmitir señales, es decir, controlar el flujo de información entre las células y su entorno. Esto es posible gracias a que la membrana contiene receptores específicos para los estímulos externos. A su vez, algunas membranas generan señales, que pueden ser químicas o eléctricas (ejemplo: las neuronas). 2. Proporcionar un medio óptimo para el funcionamiento de las proteínas de membrana (enzimas, receptores y proteínas transportadoras). Los enzimas de membrana catalizan reacciones que difícilmente tendrían lugar en un medio acuoso. 3. Controlar el desarrollo de la célula y la división celular. 4. Permitir una disposición adecuada de moléculas funcionalmente activas (antígenos, anticuerpos, etc.). 5. Delimitar compartimentos intracelulares. 6. Mantener una permeabilidad selectiva mediante el control del paso de sustancias entre el exterior y el interior de la célula. Es el denominado transporte celular. Transporte Celular
Las células precisan para su subsistencia de un continuo intercambio de sustancias con el exterior que se realiza a través de su membrana plasmática. Deben tomar del medio extracelular las moléculas necesarias, en ocasiones en concentraciones muy bajas, al tiempo que tienen que impedir el paso a las que no les convienen. Además, a través de la membrana, la célula debe liberar al exterior las moléculas del catabolismo celular y los productos de secreción. Para ello la membrana plasmática debe actuar como una barrera semipermeable muy selectiva, tanto frente a los iones como a las sustancias de alta y baja masa molecular.
A.- Permeabilidad Selectiva
El transporte a través de la membrana puede ser pasivo cuando se produce de un modo espontáneo o activo, cuando requiere un aporte de energía. Al mismo tiempo, la membrana ejerce un severo control del tipo de sustancias que la pueden atravesar.
Transporte Pasivo
Los solutos, movidos por un gradiente de concentración, se desplazan espontáneamente a través de la membrana desde la zona mas concentrada a la más diluida; si el soluto posee cargas eléctricas, además de este gradiente químico, influye también un gradiente eléctrico. Ambos constituyen el gradiente electroquímico. El transporte pasivo, por tanto, no es más que un proceso de difusión a través de la membrana, que puede ser de dos tipos:
Difusión simple: cuando los solutos atraviesan la membrana por sí mismos, ya sea a través de la bicapa lipídica o a través de canales específicos formados por determinadas proteínas de la membrana. En el primer caso, a través de la bicapa lipídica, sólo es posible la difusión en el caso de las moléculas lipófilas que se disuelven en la membrana

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce y la atraviesan. El Oxígeno y el Nitrógeno son gases que atraviesan la membrana de esta forma, así como muchas sustancias químicas lipófilas, entre ellas algunos fármacos, anestésicos como el cloroformo y el éter, disolventes como el benceno, hormonas esteróidicas, insecticidas organofosforados, etc. La mayoría de estas sustancias, por su capacidad de atravesar la membrana plasmática con facilidad, pueden producir graves intoxicaciones. También pueden atravesar la bicapa lipídica moléculas polares de pequeño tamaño que no posean cargas eléctricas, como el agua, urea, etanol, glicerina y el anhídrido carbónico. No pueden atravesarla en cambio los azúcares, los aminoácidos, las macromoléculas y los iones, sustancias que, sin embargo, son nutrientes indispensables para la vida de la célula por lo que su transporte se realiza a través de un conjunto de proteínas que atraviesan en su totalidad la bicapa lipídica delimitando en su interior un orificio o canal que permite el paso de algunos solutos de pequeño tamaño, sobre todo los iones sodio, potasio, calcio y cloro. Estos canales constituyen puertas de entrada a la célula y pueden estar permanentemente abiertos o tener apertura regulada. Difusión facilitada: Se denomina así debido a la existencia de unas proteínas transportadoras que se unen a las moléculas de los solutos y facilitan su transporte de una parte a otra de la membrana. Este transporte es específico ya que a cada molécula de soluto se une exclusivamente con su correspondiente transportador. De esta forma se transportan azúcares, aminoácidos y macromoléculas.
Transporte Pasivo: difusión, difusión mediada por canal de ion y difusión mediada por transportador.
Transporte Activo
Los solutos atraviesan la membrana con ayuda de proteínas transportadoras, pero en este caso, el transporte se realiza en contra de un gradiente electroquímico, es decir, pasan de una zona más diluida a otra más concentrada. Este proceso, por esta razón, no
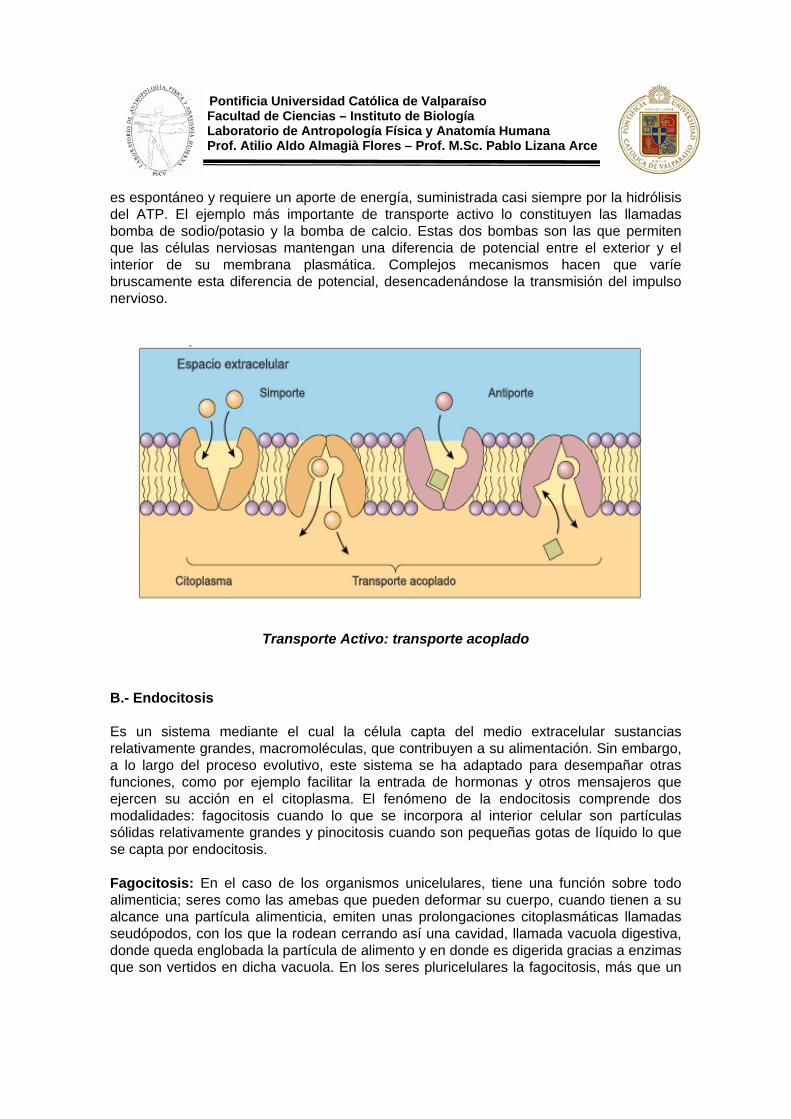
Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce es espontáneo y requiere un aporte de energía, suministrada casi siempre por la hidrólisis del ATP. El ejemplo más importante de transporte activo lo constituyen las llamadas bomba de sodio/potasio y la bomba de calcio. Estas dos bombas son las que permiten que las células nerviosas mantengan una diferencia de potencial entre el exterior y el interior de su membrana plasmática. Complejos mecanismos hacen que varíe bruscamente esta diferencia de potencial, desencadenándose la transmisión del impulso nervioso.
Transporte Activo: transporte acoplado
B.- Endocitosis
Es un sistema mediante el cual la célula capta del medio extracelular sustancias relativamente grandes, macromoléculas, que contribuyen a su alimentación. Sin embargo, a lo largo del proceso evolutivo, este sistema se ha adaptado para desempañar otras funciones, como por ejemplo facilitar la entrada de hormonas y otros mensajeros que ejercen su acción en el citoplasma. El fenómeno de la endocitosis comprende dos modalidades: fagocitosis cuando lo que se incorpora al interior celular son partículas sólidas relativamente grandes y pinocitosis cuando son pequeñas gotas de líquido lo que se capta por endocitosis.
Fagocitosis: En el caso de los organismos unicelulares, tiene una función sobre todo alimenticia; seres como las amebas que pueden deformar su cuerpo, cuando tienen a su alcance una partícula alimenticia, emiten unas prolongaciones citoplasmáticas llamadas seudópodos, con los que la rodean cerrando así una cavidad, llamada vacuola digestiva, donde queda englobada la partícula de alimento y en donde es digerida gracias a enzimas que son vertidos en dicha vacuola. En los seres pluricelulares la fagocitosis, más que un

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce proceso de nutrición celular, representa un mecanismo defensivo realizado por células especializadas llamadas fagocitos. De esta forma son eliminados microorganismos siguiendo un mecanismo parecido al de las vacuolas digestivas.
Microfotografía donde puedes visualizar el proceso de endocitosis en un capilar
Pinocitosis: Es la ingestión de líquidos mediante la formación de invaginaciones que engloban los líquidos y se estrangulan formando vacuolas digestivas. Se presenta en organismos unicelulares y en diversas células de los pluricelulares, especialmente las que tapizan las cavidades digestivas. La pinocitosis no siempre va dirigida a la captura de nutrientes líquidos para su posterior digestión. A veces es un mecanismo destinado a introducir sustancias de reserva en las células, para después incorporarse a las cavidades del retículo endoplasmático donde son almacenados. Otras veces la pinocitosis tiene como objeto el transporte de sustancias extracelulares de un lado a otro de la célula sin que queden retenidas en ella. Este es el caso de las células epiteliales que tapizan el intestino, que capturan gotas de grasa del alimento en uno de sus extremos trasladándolas al otro dentro de una vesícula pinocítica. De ahí pasan a los capilares linfáticos siendo así absorbida.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce C.- EXOCITOSIS
Existe también un mecanismo contrario a la endocitosis que recibe el nombre de exocitosis: consiste en la liberación al exterior de la célula de productos elaborados por ella, es por tanto un fenómeno de excreción. La exocitosis está precedida de una fase de migración de los gránulos que contienen las sustancias que se van a liberar; se van aproximando a la membrana plasmática hasta adherirse a su cara interna. La exocitosis propiamente dicha consistiría en la rotura de la membrana del gránulo, con lo que se abre al exterior y libera el contenido.
Esquema que ilustra las vías de endocitosis. Obsérvese la acción y reciclaje de clatrina.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
Citoplasma: Citosol y Organelos
El citoplasma es aquella parte de la célula contenida entre la membrana y el núcleo. Está constituido por una sustancia semilíquida de aspecto viscoso sin estructura aparente, denominada citosol, en la cual se hallan inmersos una serie de estructuras o formaciones que constituyen los denominados orgánulos.
CITOSOL
El citosol, también llamado hialoplasma, es la fracción soluble del citoplasma. Esta formado por una masa gelatinosa que ocupa todo el espacio desde el citoplasma externo hasta los orgánulos celulares. Constituye el verdadero jugo celular aunque no se trata de una simple disolución dispersa al azar pues posee una compleja organización interna denominada citoesqueleto, muy difícil de estudiar ya que se desintegra con mucha facilidad, sobre todo cuando se someten las células a procesos de fijación y tinción necesarios para que puedan ser observados al microscopio.
El citosol contiene los sistemas enzimáticos responsables de gran parte de las reacciones del metabolismo, como la glucólisis, glucogénesis, gluconeogénesis, síntesis de ácidos grasos, nucleótidos y aminoácidos. También se sintetizan en el citosol algunas proteínas mediante los ribosomas que se encuentran libres en él. En el citosol se almacenan algunos productos de la biosíntesis, sobre todo sustancias de reserva, como el glucógeno y las grasas que, en forma de gotas dispersas, pueden llegar a ocupar todo el volumen celular, como es el caso de los adipocitos.
ORGANELOS NO MEMBRANOSOS
CITOESQUELETO
El citosol no es una masa amorfa ya que el funcionamiento ordenado y armónico de todos

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce los componentes celulares no podría llevarse a cabo. Por el contrario, el citosol posee una compleja organización interna formada por redes de microfilamentos y microtúbulos denominada citoesqueleto. Este término no se refiere a una estructura rígida, ni a un armazón articulado como otros esqueletos, sino que se trata de una matriz organizada que cambia de aspecto con gran rapidez y proporciona a la célula, además de consistencia morfológica, una disposición ordenada y dinámica del citoplasma. Este citoesqueleto dinámico es responsable de la forma, el desplazamiento y la división de la célula, así como del transporte de vesículas y sustancias de una parte a otra del citoplasma. Incluso es posible que aporte un marco tridimensional adecuado para la concentración en diferentes zonas del citosol de los enzimas responsables de una determinada ruta metabólica; de esta manera se consigue una mayor compartimentación celular.
Hasta ahora se han descrito 3 clases de sistemas de filamentos proteicos integrantes del citoesqueleto:
1.- Microfilamentos: son las fibras más delgadas y son un conjunto de filamentos proteicos relacionados con la arquitectura y el movimiento de la célula. La proteína más característica es la actina, que se asocia con otros componentes proteicos según el tipo de célula y la función que desempeñe: en los eritrocitos la actina se asocia con la espectrina para formar una red en la cara interna de la membrana plasmática que confiere a estas células la flexibilidad necesaria para atravesar los capilares sanguíneos. En las fibras musculares estriadas, la actina se asocia con la miosina para formar los filamentos responsables de la contracción muscular.
2.- Microtúbulos: son los más gruesos y huecos. Son estructuras filamentosas huecas formadas a partir de subunidades proteicas que se asocian por un proceso de polimerización en el que intervienen 2 tipos de monómeros ligeramente distintos: tubulina a y tubulina b; ambas son proteínas globulares unidas para constituir las estructuras cilíndricas de los microtúbulos.
3.- Filamentos intermedios: su diámetro se sitúa entre el de los dos anteriores. Constituye un grupo de filamentos proteicos de distinta naturaleza que no siempre son los mismos en todas las células e incluso pueden faltar en algún tejido. Reciben distintos nombres: en las células epiteliales queratinas, en las neuronas neurofilamentos, en las fibras musculares filamentos de desmina, etc. En todos los casos forman un entramado interno relacionado sobre todo con otros componentes del citoesqueleto.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
Esquema de los elementos del citoesqueleto y el centríolo.
CILIOS Y FLAGELOS
Son prolongaciones móviles localizadas en la superficie de muchas células que permiten a éstas desplazar el medio que les rodea. A su vez, el desplazamiento del medio da

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce origen al movimiento de las células si viven aisladas.
Los cilios, de 0,2 µ de diámetro y entre 2 y 10 µ de longitud, aparecen en gran número sobre la misma célula. Los flagelos, de diámetro semejante, alcanzan hasta 200 µ de longitud. Las células flageladas sólo poseen uno o, a lo sumo, algunos flagelos. La estructura interna de cilios y flagelos es muy similar.
Estructura: poseen en su interior una estructura formada por un haz de microtúbulos orientados según el eje principal del cilio. A esta estructura se le denomina tallo o axonema y es continuación de otra, anclada en el hialoplasma, llamada corpúsculo basal. El axonema está rodeado de la membrana plasmática y tiene en su interior dos microtúbulos centrales rodeados de una delgada vaina. En la periferia hay 9 pares de microtúbulos periféricos; de cada par uno es completo (a) y el otro no (b). Del microtúbulo a salen dos brazos formados por una proteína, la dineína, que se dirigen hacia el microtúbulo b de la pareja vecina. Otras fibras proteicas unen cada par de microtúbulos entre sí y con la vaina central. El corpúsculo basal es un cilindro de 0,2 a 0,5 µ de longitud, colocado en la base del cilio. Carece del par de microtúbulos centrales, mientras que los dobletes de microtúbulos periféricos se han transformado en tripletes.
La función más importante de cilios y flagelos consiste en mover el líquido que rodea la célula. Si la célula vive aislada, este movimiento la propulsa a través del medio. Cilios y flagelos se mueven de forma distinta: los primeros tienen movimiento pendular y el flagelo ondulante. La causa del movimiento es el deslizamiento de unos dobletes periféricos respecto a otros. Como los dobletes están anclados en el corpúsculo basal, el deslizamiento provoca la flexión del cilio o del flagelo.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
CENTRÍOLOS Y CENTROSOMAS
Para que los microtúbulos intervengan eficazmente en la trama estructural y en los movimientos celulares, deben estar unidos a otras regiones de la célula. Los microtúbulos que forman el axonema de cilios y flagelos están anclados en los corpúsculos basales, mientras que los microtúbulos citoplasmáticos terminan en una región de la célula próxima al núcleo, llamada centro celular, citocentro o centrosoma. El corpúsculo basal y el centrosoma son centros organizadores de los microtúbulos, es decir, los microtúbulos crecen y se organizan a partir de ellos. En las células eucarióticas, el centrosoma se localiza cerca del núcleo y frecuentemente está rodeado por los dictiosomas del aparato de Golgi. El centrosoma de una célula animal está formado por:
Centríolos con su particular conformación 9 + 0
1.- Un par de centríolos o diplosoma. Los centríolos son estructuras idénticas a los corpúsculos basales de los cilios. Se sitúan ocupando el centro del centrosoma dispuestos perpendicularmente entre sí. En la división celular, cada uno de ellos da origen, por duplicación, a su pareja. De este modo se obtienen dos diplosomas, uno para cada célula hija. Las células de los vegetales superiores carecen de centriolos.
2.- Las fibras del áster, formadas por microtúbulos que crecen y se organizan en forma de radios a partir de los centriolos. Durante la división celular, las fibras del áster dan origen a los microtúbulos del huso acromático.
RIBOSOMAS
Son orgánulos visibles solamente con M.E. Son partículas globulares de 15-30 nm. de diámetro. Cada ribosoma está formado por dos subunidades, una mayor y otra menor, que se disocian reversiblemente después de cada ciclo de síntesis proteica. Poseen una estructura compleja y son muy porosos. Están formados por ARNr y docenas de proteína. Todas las células, ya sean procariotas o eucariotas poseen ribosomas. En las eucariotas se encuentran en el hialoplasma o adosados a las paredes del retículo endoplasmático rugoso. Es posible encontrar ribosomas incluso en el interior de mitocondrias y cloroplastos. Los ribosomas procarióticos son más pequeños que los eucarióticos. Los

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce ribosomas suelen presentarse en grupos de 5 a 20 unidos por filamentos de ARNm; a estos grupos se les denomina polisomas.
ORGANELOS MEMBRANOSOS
RETÍCULO ENDOPLASMÁTICO
Entre el complejo sistema de membranas internas que caracteriza a las células eucarióticas se encuentra el R.E. Se trata de un conjunto de cavidades cerradas de formas muy variables: láminas aplanadas, vesículas globulares o tubos de aspecto sinuoso. Estas cavidades constituyen el 10% del volumen celular, se comunican entre sí y forman una red continua, separada del hialoplasma por la membrana del propio R.E.
Retículo Endoplasmático Liso y Rugoso
Se pueden distinguir dos tipos de R.E.: el R.E.rugoso (R.E.R.) con ribosomas adosados en el lado de la membrana que da al hialoplasma, y el R.E.liso (R.E.L.) que no tiene ribosomas.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce La membrana del R.E. es más delgada que la membrana plasmática, entre 5-6 nm. pero su estructura es análoga. La composición química de ambas es similar aunque en el R.E. la proporción de lípidos es menor (aprox. 30%) y la de proteína mayor. Estas proteínas son en su mayoría enzimas encargados del transporte de electrones. Las cavidades contienen en su interior una solución acuosa rica en holoproteinas, glucoproteínas y lipoproteínas. Dentro de las funciones del R.E. destacan las siguientes:
1.-Síntesis de proteína: los ribosomas unidos a las membranas del R.E.R. son los responsables de esta síntesis. Las proteínas obtenidas pueden tener dos destinos: si forman parte de los productos de secreción celular son transferidas al interior de cavidades por las que circular n por la célula. Si forman parte de las membranas celulares, quedar n ancladas a la membrana del R.E.
2.- Síntesis de lípidos: Los fosfolípidos y el colesterol se sintetizan en las membranas del R.E. Sólo los ácidos grasos se sintetizan en el hialoplasma. Es por tanto en el R.E. donde se fabrican los componentes de las membranas celulares. Estos son exportados en vesículas que se dirigen hacia la membrana de un orgánulo concreto o hacia la membrana plasmática en la cual se integran.
3.- Detoxificación: en la membrana del R.E.L. existen enzimas capaces de eliminar la toxicidad de aquellas sustancias que resultan perjudiciales para la célula, ya sean producidas por ella misma como consecuencia de su actividad vital o provengan del medio externo. La pérdida de toxicidad se consigue transformando estas sustancias en otras solubles que puedan abandonar la célula y ser secretadas por la orina. Esta función la realizan principalmente las células de los riñones, los pulmones, el intestino y la piel.
APARATO DE GOLGI (A.G.)
Está formado por un apilamiento de sacos de forma discoidal (cisternas), rodeados por un enjambre de pequeñas vesículas. Está situado cerca del núcleo de la célula y en las células animales suele rodear a los centriolos.
Cada pila de sacos recibe el nombre de dictiosoma. Cada uno mide alrededor de 1 micra de diámetro y agrupa unas 6 cisternas, aunque en los eucariotas inferiores su número puede llegar a 30. Las células eucarióticas poseen un A.G. más o menos desarrollado según la función que desempeñen. En cada caso el número de dictiosomas varia de unos pocos hasta cientos de ellos.
Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
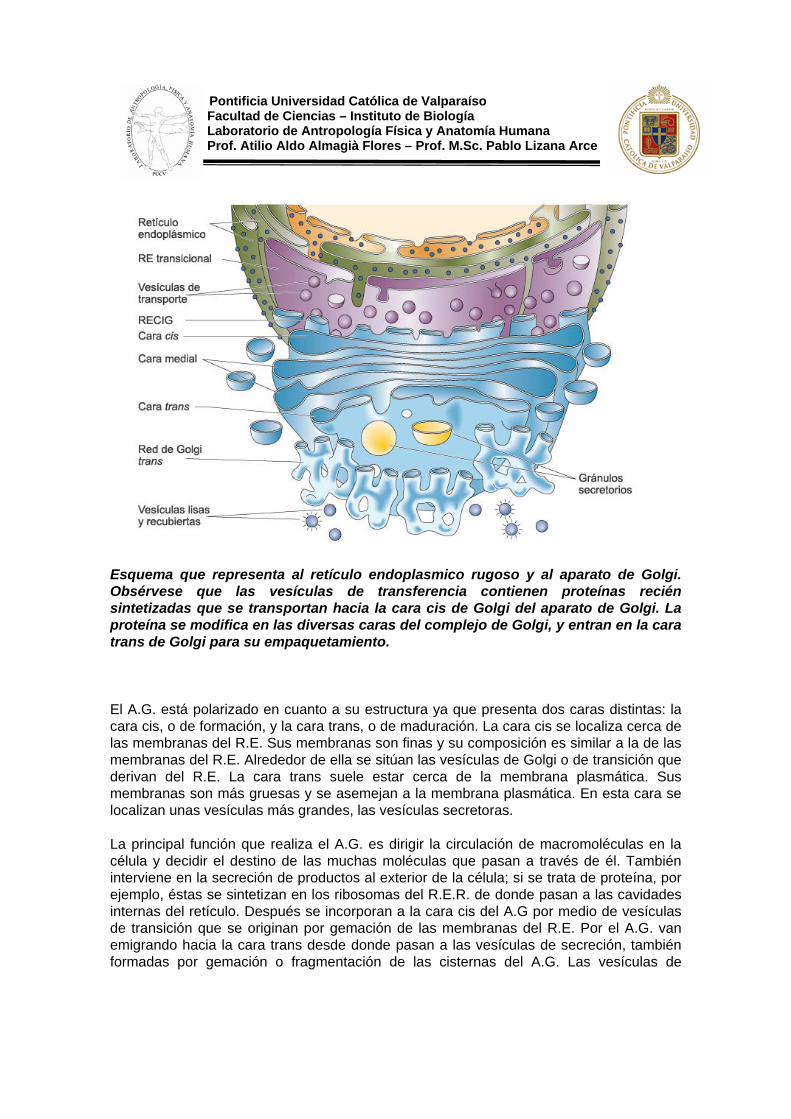
Esquema que representa al retículo endoplasmico rugoso y al aparato de Golgi. Obsérvese que las vesículas de transferencia contienen proteínas recién sintetizadas que se transportan hacia la cara cis de Golgi del aparato de Golgi. La proteína se modifica en las diversas caras del complejo de Golgi, y entran en la cara trans de Golgi para su empaquetamiento.
El A.G. está polarizado en cuanto a su estructura ya que presenta dos caras distintas: la cara cis, o de formación, y la cara trans, o de maduración. La cara cis se localiza cerca de las membranas del R.E. Sus membranas son finas y su composición es similar a la de las membranas del R.E. Alrededor de ella se sitúan las vesículas de Golgi o de transición que derivan del R.E. La cara trans suele estar cerca de la membrana plasmática. Sus membranas son más gruesas y se asemejan a la membrana plasmática. En esta cara se localizan unas vesículas más grandes, las vesículas secretoras.
La principal función que realiza el A.G. es dirigir la circulación de macromoléculas en la célula y decidir el destino de las muchas moléculas que pasan a través de él. También interviene en la secreción de productos al exterior de la célula; si se trata de proteína, por ejemplo, éstas se sintetizan en los ribosomas del R.E.R. de donde pasan a las cavidades internas del retículo. Después se incorporan a la cara cis del A.G por medio de vesículas de transición que se originan por gemación de las membranas del R.E. Por el A.G. van emigrando hacia la cara trans desde donde pasan a las vesículas de secreción, también formadas por gemación o fragmentación de las cisternas del A.G. Las vesículas de

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce secreción se adosan a la membrana plasmática y descargan su contenido por exocitosis.
LISOSOMAS
Son vesículas rodeadas por una membrana en cuyo interior tiene lugar la digestión controlada de macromoléculas. Se encuentran en todas las células eucarióticas. Contiene al menos 40 enzimas del tipo hidrolasas ácidas. Estos enzimas son capaces de romper las macromoléculas y su funcionamiento es óptimo a pH ácido. La membrana del lisosoma impide que sea digerido ‚l mismo por estos enzimas y, además, es la que se encarga de mantener en el interior un pH ácido. El enzima más característico de los lisosomas es la fosfatasa ácida, que hidroliza los enlaces éster fosfórico y libera grupos fosfato.
Aunque todos los lisosomas contienen enzimas hidrolíticos, el resto de su contenido puede ser muy distinto. Debido a ello se distinguen dos tipos:
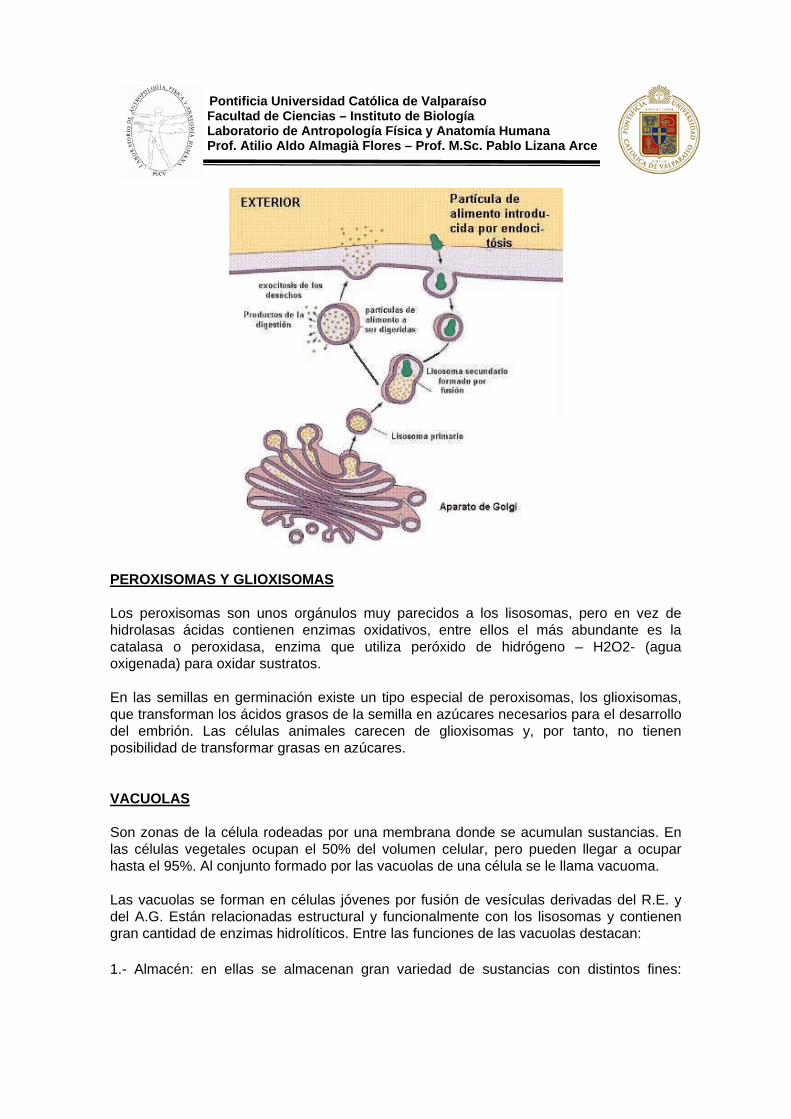
1.- Lisosomas primarios: sólo contienen enzimas hidrolíticos; se trata de vesículas de secreción, recién formadas por gemación a partir del A.G.
2.- Lisosomas secundarios: contienen, además de las hidrolasas, sustratos en vía de digestión. Se trata de lisosomas primarios que se han fusionado con otras sustancias; si éstas tienen origen externo se llaman vacuolas heterofágicas o digestivas, y tiene origen interno de la célula se denominan vacuolas autofágicas.
Los lisosomas pueden realizar la digestión celular de dos formas distintas: Digestión extracelular, se produce cuando los lisosomas vierten su contenido al exterior de la célula, donde tiene lugar la digestión. Es muy corriente en los Hongos. Digestión intracelular, se produce cuando el lisosoma permanece en el interior de la célula, pero según de donde provenga el sustrato se distinguen dos tipos: Autofagia, cuando el sustrato es un constituyente celular como porciones del A.G., del R.E., etc. Desempeña un importante papel en la vida de las células, ya que destruye zonas dañadas o innecesarias de las mismas; Heterofagia, cuando el sustrato es de origen externo. Su finalidad es doble, nutrir y defender a la célula. Los sustratos son capturados por endocitosis y se forma una vesícula que se fusiona a un lisosoma primario dando origen a una vacuola digestiva o heterofágica. En su interior se produce la digestión de los sustratos pasando los productos de la digestión al hialoplasma donde son utilizados. Los desechos no digeribles son expulsados al exterior de la célula por exocitosis. En las semillas existe un tipo especial de lisosomas, los granos de aleurona, que almacenan sustancias de reserva. En ellos no tiene lugar la digestión celular hasta que no llegue el momento de la germinación de la semilla.
Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
PEROXISOMAS Y GLIOXISOMAS
Los peroxisomas son unos orgánulos muy parecidos a los lisosomas, pero en vez de hidrolasas ácidas contienen enzimas oxidativos, entre ellos el más abundante es la catalasa o peroxidasa, enzima que utiliza peróxido de hidrógeno – H2O2- (agua oxigenada) para oxidar sustratos.
En las semillas en germinación existe un tipo especial de peroxisomas, los glioxisomas, que transforman los ácidos grasos de la semilla en azúcares necesarios para el desarrollo del embrión. Las células animales carecen de glioxisomas y, por tanto, no tienen posibilidad de transformar grasas en azúcares.
VACUOLAS
Son zonas de la célula rodeadas por una membrana donde se acumulan sustancias. En las células vegetales ocupan el 50% del volumen celular, pero pueden llegar a ocupar hasta el 95%. Al conjunto formado por las vacuolas de una célula se le llama vacuoma.
Las vacuolas se forman en células jóvenes por fusión de vesículas derivadas del R.E. y del A.G. Están relacionadas estructural y funcionalmente con los lisosomas y contienen gran cantidad de enzimas hidrolíticos. Entre las funciones de las vacuolas destacan: 1.- Almacén: en ellas se almacenan gran variedad de sustancias con distintos fines:
Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce productos de desecho que resultarían perjudiciales para la célula si se almacenaran en el citoplasma. Las vacuolas de ciertas células acumulan sustancias tan especiales como el caucho o el opio. También pueden acumular sustancias que la planta utiliza en su relación con otras plantas o animales, como colorantes para los pétalos o alcaloides venenosos. Por último las vacuolas también acumulan sustancias de reserva, como ocurre en las semillas.
2.- Permiten que aumente de tamaño la célula vegetal sin que ello suponga un gasto de energía. Las células vegetales crecen, en gran medida, por acumulación de agua en sus vacuolas.
3.-En las células animales existe un tipo especial de vacuolas denominadas pulsátiles que presentan las células que viven en ambientes hipotónicos y que son utilizadas para bombear el exceso de agua que penetra hacia el exterior.
MITOCONDRIAS
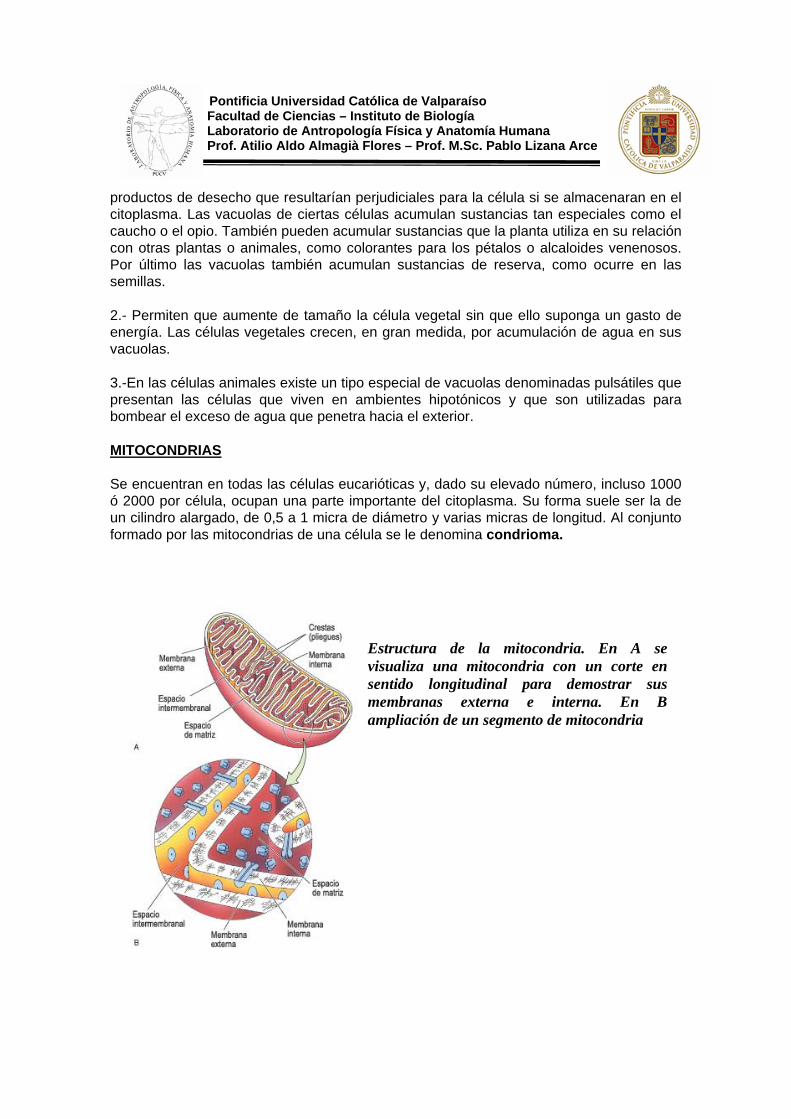
Se encuentran en todas las células eucarióticas y, dado su elevado número, incluso 1000 ó 2000 por célula, ocupan una parte importante del citoplasma. Su forma suele ser la de un cilindro alargado, de 0,5 a 1 micra de diámetro y varias micras de longitud. Al conjunto formado por las mitocondrias de una célula se le denomina condrioma.
Estructura de la mitocondria. En A se visualiza una mitocondria con un corte en sentido longitudinal para demostrar sus membranas externa e interna. En B ampliación de un segmento de mitocondria

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce Una mitocondria está limitada por una doble membrana, la membrana mitocondrial externa (mme), que la separa del hialoplasma, y la membrana mitocondrial interna (mmi), que forma unos repliegues hacia el interior, las crestas mitocondriales. Estas membranas definen dos compartimentos separados: el espacio intermembranario, limitado por ambas, y la matriz, espacio interno limitado por la membrana mitocondrial interna.
El compartimiento más especial, desde el punto de vista químico, es la matriz, junto con la membrana mitocondrial interna que la rodea. El contenido de la matriz incluye: moléculas de ADN (ADN-mitocondrial) que contiene la información para sintetizar las proteínas mitocondriales, ribosomas (mitoribosomas) que pueden estar libres en la matriz o adosados a la membrana mitocondrial interna y son semejantes a los bacterianos, y gran cantidad de enzimas y moléculas de ATP.
La membrana mitocondrial interna posee una gran superficie gracias a las crestas. Es más rica en proteína (80%) que otras membranas celulares y entre sus lípidos no se encuentra el colesterol, lo cual también es típico de las membranas bacterianas. Entre las proteínas de la mmi destacan las que forman la cadena de transporte electrónico y el complejo enzimático llamado ATP-sintetasa que cataliza la síntesis de ATP. Por su parte, la mme se asemeja más a otras membranas celulares, en particular a la del R.E. y es muy permeable debido a la gran cantidad de canales proteicos que posee. La composición del espacio intermembranario es similar a la del hialoplasma debido a la permeabilidad de la mme.
TEORIA DE LA ENDOSIMBIOSIS
Es una teoría que explica satisfactoriamente la evolución celular con los datos que se tienen actualmente. También se denomina teoría de las comunidades microbianas coevolucionadas. Propone que el origen de la célula eucariótica podría deberse a la simbiosis permanente entre diferentes tipos de procariotas, las cuales habrían sido víctimas de la captura por fagocitosis por un precursor ancestral anaerobio. Posteriormente lograron sobrevivir en su interior y establecieron una relación simbiótica con su cazador. De esta manera los antepasados de las mitocondrias fueron bacterias aerobias Gram negativas y los cloroplastos procederían de primitivas bacterias fotosintéticas.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
Núcleo Celular
Es un corpúsculo que destaca con claridad inmerso en el citoplasma de todas las células eucarióticas. Normalmente su posición es central pero puede hallarse desplazado por los constituyentes del citoplasma, como es el caso de las vacuolas en las células vegetales.
Su importancia es trascendental porque es el elemento que rige todas las funciones celulares y es además el portador de los factores hereditarios. Entre el núcleo y el citoplasma existe una relación muy estrecha y depende el uno del otro de tal manera que ninguna de las dos partes puede mantenerse viva mucho tiempo separada de la otra. Si se secciona una célula en dos porciones de manera que una de ellas quede con el núcleo y la otra no, sólo la primera sería capaz de regenerar el fragmento perdido y seguir viviendo.
Sus funciones son:
• Almacenar los genes en los cromosomas • Organizar los genes en los cromosomas y permitir la división celular • Transporte de los factores regulatorios y los productos de los genes vía los poros nucleares. • Producir mensajes (RNA mensajero o mRNA) que codifican para las proteínas. • Producir ribosomas en el nucléolo • Organizar el desenrrollamiento del ADN para replicar genes claves. En cuanto a su forma, la más frecuente es la esférica, pero existen muchos casos de núcleos elipsoidales, arriñonados e incluso lobulados, como en muchos glóbulos blancos. Para cada tipo de células, la relación entre el volumen nuclear y el volumen citoplasmático es constante.
La mayor parte de las células, tanto animales como vegetales, sólo poseen un núcleo, pero existen casos de células plurinucleadas. En estos casos los núcleos supernumerarios pueden tener dos orígenes distintos: • Por fusión de varias células uninucleadas. En este caso la célula plurinucleada resultante se denomina sincitio. • Por división repetida del núcleo sin que se vea seguida por una división del citoplasma. En este caso la célula plurinucleada se llama plasmodio. En todos los núcleos se pueden distinguir cuatro partes: membrana nuclear (o envoltura nuclear), nucleoplasma, nucleolo y cromosomas.
ENVOLTURA NUCLEAR: La envoltura nuclear (E.N.) es una doble membrana formada por dos finísimas hojas entre las cuales queda un espacio de unos 10-15 nm. de ancho que se continúa con el R.E. En realidad, toda la E.N. no es más que una prolongación del propio R.E. que contornea el núcleo envolviéndolo. La E.N. no forma una cubierta continua ya que en varios puntos de ella existen interrupciones por la presencia de poros nucleares que ponen en comunicación el contenido del núcleo con el citoplasma.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce Alrededor de los bordes de estos poros nucleares, ambas membranas se continúan. Los poros tienen un diámetro de 80 nm. y la superficie ocupada por ellos respecto a la superficie total del núcleo es un 10%, aunque en ciertas células vegetales puede llegar al 36%. En general, cuanto más activa es una célula, mayor es el número de poros que posee su núcleo.
El esquema representa la estructura del poro nuclear, demostrando que está constituida por varias combinaciones de ocho unidades cada una.
NUCLEOPLASMA: Es el contenido interno del núcleo y es similar al citosol. Está formado por una disolución compuesta por gran variedad de principios inmediatos, especialmente nucleótidos y enzimas implicados en la transcripción y replicación del ADN. Inmersos en el nucleoplasma se encuentran los cromosomas y el/los nucleolo/s, pero no se encuentran libres, sino que están sostenidos por una red proteica tridimensional similar al citoesqueleto que se extiende por todo el núcleo y sirve de anclaje a las demás partes.
NUCLEOLO: Es un corpúsculo esférico que, a pesar de no estar delimitado por una membrana, suele ser muy visible dado que su viscosidad es mayor que la del resto del núcleo. Es frecuente que exista más de un nucleolo; el caso más extremo es el de los óvulos de los Anfibios que poseen más de un millar.
Existe una relación entre el tamaño del nucleolo y la actividad sintética de la célula. Los nucleolos de las células que exhiben poca síntesis proteica son muy pequeños o no se

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce encuentran, mientras que en las células cuya síntesis proteica es alta los nucleolos son muy grandes. El nucleolo contiene el aparato enzimático encargado de sintetizar los diferentes tipos de ARNr. Su función es precisamente la de formar y almacenar ARNr con destino a la organización de los ribosomas. Son también indispensables para el desarrollo normal de la Mitosis.
CROMATINA Y CROMOSOMAS: La cromatina es la sustancia fundamental del núcleo y recibe este nombre por su capacidad de teñirse con colorantes básicos. Aunque con el M.E. se observa una masa grumosa aparentemente amorfa, es una de las estructuras celulares dotadas de mayor complejidad en su organización. Las fibras de cromatina constan de diferentes niveles de organización que son el nucleosoma, el collar de perlas y la fibra de cromatina, de 30 nm de diámetro. Estos niveles de organización permiten empaquetar grandes cantidades de ADN, asociado a las histonas, en el reducido volumen nuclear. En un momento dado, no toda la cromatina se encuentra en el mismo grado de condensación. Según esto, se distinguen dos tipos de cromatina:
En la figura, se visualiza elNÚCLEO. Donde se observa la membrana externa tapizada de ribosomas sobre su superficie citoplasmática, la que se continúacon el retículo endoplásmico rugoso RER. Observe que ambas membranas están unidas a nivel de poros nucleares.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce Eucromatina: comprende el conjunto de zonas donde la cromatina está poco condensada con el fin de que los bucles de ADN se encuentren suficientemente distendidos para que se pueda llevar a cabo sobre ellos la transcripción. La eucromatina, junto con el nucleolo, son las zonas donde los genes se están transcribiendo.
Heterocromatina: es la parte de la cromatina que presenta mayor grado de empaquetamiento con el fin de que el ADN que contiene no se transcriba y permanezca funcionalmente inactivo. Existen dos clases de heterocromatina: constitutiva y facultativa. La 1ª es el conjunto de zonas que se encuentran condensadas en todas las células y, por tanto, su ADN no se transcribe nunca en ninguna de ellas. Todavía se sabe poco sobre su función. La heterocromatina facultativa comprende zonas distintas en diferentes células, ya que representa el conjunto de genes que se inactivan de manera específica en cada tipo de célula durante la diferenciación celular. En los tejidos embrionarios es muy escasa la heterocromatina facultativa y aumenta cada vez más conforme se especializan las células de los diferentes tejidos pues se inactivan determinados genes y para ello se empaquetan de forma condensada de manera que ya no pueden transcribirse.
Cromosomas: En los periodos de división celular (Mitosis o Meiosis), la cromatina da lugar a unas estructuras denominadas cromosomas visibles con M.O. Tienen forma de bastoncillos más o menos alargados. Dentro de la misma especie la forma de cada cromosoma es constante, de tal manera que puede ser identificado cada uno de ellos. En cada cromosoma se distingue un estrangulamiento denominado constricción primaria en cuyo interior se encuentra un gránulo llamado centrómero. La constricción primaria divide al cromosoma en dos partes o brazos cuyos extremos redondeados reciben el nombre de telómeros. Según el tamaño relativo de estos brazos se distinguen 3 tipos de cromosomas: 1.- Telocéntricos: sólo es visible un brazo porque el centrómero se encuentra en un

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce extremo. 2.- Acrocéntricos: los dos brazos son de distinta longitud. 3.- Metacéntricos: los dos brazos tienen, aproximadamente, la misma longitud porque el centrómero se encuentra en la mitad del cromosoma.
Algunos cromosomas presentan otros estrangulamientos llamados constricciones secundarias. Cuando una de ellas se encuentra cerca del extremo de un brazo delimitan un pequeño segmento esferoidal llamado satélite.
El tamaño de los cromosomas es variable. Los más largos tienen unas 30nm de longitud. En la especie humana su longitud varía entre 1,5 y 5nm. Muchas especies poseen cromosomas puntiformes, cuyo tamaño se reduce a las 0,2nm. En las células de las glándulas salivares de los Insectos Dípteros se presentan cromosomas gigantes que miden cerca de 500nm.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
Imagen que representa el cariotipo humano normal El número de cromosomas de cada especie es constante. El conjunto formado por los cromosomas de una especie constituye su cariotipo. Las especies llamadas haploides poseen un número n de cromosomas distintos. Sin embargo las llamadas diploides poseen 2n cromosomas, es decir, n parejas de cromosomas homólogos (idénticos). En cada pareja, uno de los cromosomas procede del padre y otro de la madre. En la especie humana, las células poseen 46 cromosomas en 23 parejas de homólogos. Es lo que se denomina dotación cromosómica de la especie humana.
Ciclo Celular
El axioma de Virchow, según el cual “toda célula procede de otra célula”, lleva implícito el concepto de división celular. Esto significa que, una vez terminado su desarrollo, toda célula tiene dos posibilidades: dividirse al alcanzar un cierto tamaño o, por el contrario, morir si ha perdido la capacidad de división. En el primer caso se habla de ciclo celular para describir el proceso que se inicia al término de una división celular y acaba con el final de la siguiente división, en la que se habrán formado dos nuevas células. Dicho de otro modo, el ciclo celular es el tiempo que transcurre desde que una célula se forma por división de una preexistente hasta que se divide y da origen a dos células hijas.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
En el esquema se ilustra el ciclo celular en células que se dividen de manera activa. Las células que no se dividen dejan el ciclo para entrar a la fase G0 o etapa de reposo.
La duración del ciclo celular es muy variable, dependiendo del tipo de célula, de los nutrientes y de la temperatura. Como ejemplo, en condiciones óptimas de cultivo, una ameba tiene un ciclo celular de unas 10 horas, un cicliado de 2-3 horas y células de mamífero unas 20 horas.
El ciclo vital de una célula se divide en dos fases muy definidas:
• Interfase o intermitosis • División, mitosis, cariocinesis o fase M.
INTERFASE
Es la fase más larga del ciclo celular, 94% del mismo, y transcurre entre dos mitosis. En ella se produce una intensa actividad biosintética a todos los niveles lo que, junto al mantenimiento y crecimiento celular, prepara la fase de división. Durante esta fase la

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce célula duplica sus componentes y especialmente su material genético, el ADN, lo cual ocurre en un periodo muy concreto de la interfase llamado fase S. Los periodos anterior y posterior a esta duplicación del ADN se denominan respectivamente fase G1 y G2 (del inglés “gap”, intervalo o separación).
Fase G1: Duración variable, aproximadamente 5 horas. Transcurre entre el final de una mitosis y el inicio de la síntesis del ADN. En ella la célula aumenta de volumen hasta alcanzar el tamaño de su forma celular típica, al tiempo que se produce la acumulación de las sustancias necesarias para la división celular. En cierto momento de esta fase se alcanza el llamado punto R o de no retorno, pasado el cual la célula ya está obligada a completar la totalidad del ciclo celular. Si la célula no sobrepasa ese punto R, puede permanecer indefinidamente en un estado de reposo, llamado fase G0, en el que no se sintetizan proteínas cromosómicas ni el ARNm de las mismas. Las células que no se dividen nunca, como las neuronas, permanecen de por vida en esta fase G0 y se denominan quiescentes. Cuando una célula de este tipo sale de este estado a destiempo y de forma descontrolada se produce cáncer.
Cerca del final de la fase G1 se produce una activa síntesis proteica, en especial las histonas de la cromatina, que son transportadas al núcleo. Igualmente se producen los desoxirribonucleótidos y los enzimas necesarios para la replicación, al tiempo que aumenta el número de microtúbulos y el tamaño de los centriolos.
Fase S: Duración de 6 a 8 horas. En este periodo se replica el ADN del núcleo, en primer lugar la eucromatina y la heterocromatina, más condensada, lo hace al final de la fase. No hay crecimiento celular y disminuye la actividad metabólica de la célula, aunque persiste la síntesis de ARNm y de histonas. Se produce, además, la duplicación de los centríolos, pero los dos pares permanecen incluidos en un único centrosoma hasta el inicio de la mitosis.
Fase G2: Duración 3-5 horas. Es un periodo de latencia que se extiende desde el final de la replicación hasta el comienzo de la división nuclear. Puede haber en ella un cierto crecimiento celular y la célula adopta procesos de regulación para evitar el exceso de productos génicos (la cantidad de ADN está duplicada). La fase G2 acaba cuando los cromosomas empiezan a condensarse al inicio de la mitosis.
MITOSIS
El crecimiento de los organismos exige que sus células lleven a cabo algún tipo de división celular y ésta debe producirse de forma que los componentes celulares esenciales sean distribuidos correctamente entre las células hijas. Una distribución desigual de algo tan esencial como el material genético, el portador de la información biológica, pondría en peligro el futuro de la especie. Para que esto no ocurra existe un mecanismo que produce la división regular de los componentes hereditarios, los cromosomas, llamado mitosis.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce La división celular exige por un lado que se divida el núcleo mediante un mecanismo complejo encaminado a mantener constante el número de cromosomas de las células hijas, pero, por otro lado, se produce también una división del citoplasma, lo que implica un reparto más o menos equitativo de los orgánulos celulares, como las mitocondrias o los cloroplastos. Por esta razón es preciso distinguir entre la mitosis o división del núcleo, también llamada cariocinesis, y la división del citoplasma que logra la separación total de las dos células hijas, proceso llamado citocinesis.
Con el fin de describir la mitosis, se suelen distinguir 4 fases o etapas: profase, metafase, anafase y telofase, pero en realidad es un proceso continuo y las 4 se suceden sin que exista un límite claro entre ellas.
Profase: Cuando finaliza la fase G2, la célula presenta en su núcleo una red enmarañada de cromatina, pero al iniciarse la mitosis se produce un ligero aumento del tamaño del núcleo y dentro de él se empiezan a diferenciar una serie de filamentos claramente visibles que son los cromosomas. Al final de la profase, cada cromosoma es perfectamente identificable, pero, debido a la replicación del ADN que se produjo durante la fase S del ciclo celular, cada uno se halla partido longitudinalmente en dos mitades llamadas cromátidas unidas entre sí únicamente por la zona correspondiente al centrómero. Al mismo tiempo que ocurre esto, la membrana nuclear se va disolviendo y llega a desaparecer por completo quedando los cromosomas libres en el citoplasma. Los nucleolos también desaparecen.
Paralelamente también ocurren cambios en el citoplasma, sobre todo en el centrosoma. Los centriolos empiezan a separarse y cada uno se dirige hacia un polo de la célula. Entre ellos se desarrolla un haz de finísimas fibras denominado huso acromático. Esta estructura está perfectamente desarrollada al final de la profase. En las células vegetales no existen centríolos y sin embargo también se forma el huso acromático, pero se desarrolla a partir de dos zonas que se diferencian en los polos de la célula con forma de casquete. Por ello se llaman casquetes polares y a partir de ellos surgen los filamentos

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce del huso.
Metafase: Durante ella, los cromosomas, que estaban dispersos en el citoplasma, se empiezan a agrupar y acaban todos colocados en un plano ecuatorial de la célula doblados por su centrómero, que mira hacia la parte central del huso, mientras que los brazos se dirigen hacia fuera. Esta disposición recibe el nombre de estrella madre. En este momento, el centrómero se duplica con lo que las dos cromátidas de cada cromosoma quedan completamente independientes. En cada centrómero se inserta un filamento del huso acromático.
Anafase: Los filamentos del huso empiezan a acortarse progresivamente por lo que cada pareja de cromátidas se va separando entre sí a partir del vértice ocupado por el centrómero. A medida que se separan se van desplazando hacia los polos de la célula de

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce forma que, al final de la anafase, la mitad de las cromátidas acaba situada en uno de los polos y la otra mitad en el otro. Paralelamente los filamentos del huso empiezan a palidecer.
Telofase: Se caracteriza por la reconstrucción de los núcleos de las células hijas. Las cromátidas, que ya se pueden considerar cromosomas completos, empiezan a difuminarse hasta adquirir el aspecto que tienen en una célula en reposo, es decir, filamentos de cromatina. Se forma una nueva membrana nuclear y el huso desaparece totalmente.
Citocinesis: Durante la telofase ocurre también la división del citoplasma o citocinesis, aunque de forma distinta según sean células vegetales o animales.
En la vegetales se empiezan a condensar unos gránulos en la zona ecuatorial de la célula hasta que llegan a formar un tabique llamado placa celular que crece de dentro hacia fuera hasta llegar a la periferia celular; en ese momento quedan independientes las dos células hijas, aunque unidas por la placa celular, que pasará a constituir la lámina media de la pared celular de ambas. En las células animales se empieza a formar un surco en la periferia celular que poco a poco va estrangulando la zona ecuatorial hasta que acaba dividiendo a la célula madre en dos células hijas.
La duración de todo el proceso mitótico es variable según las condiciones y el tipo de célula, pero normalmente suele durar entre 30 minutos y 3 horas. La profase es la de mayor duración.
La mitosis es un proceso común a todas las células eucarióticas que garantiza que las células hijas tengan los mismos cromosomas que la célula madre y, por tanto, la misma información genética. En los organismos pluricelulares es necesario que, durante su crecimiento y desarrollo, las nuevas células que se forman tengan la misma información genética (los mismos cromosomas) que el resto de las células del organismo. De igual forma, cuando se reparan los tejidos dañados las nuevas células deben ser idénticas a las

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce que se reemplazan. La mitosis asegura que esto sea así. En la reproducción asexual, los hijos son idénticos a los padres puesto que son producidos como resultado de la división celular por mitosis. Como consecuencia, la descendencia tiene las mismas ventajas y desventajas que los padres para sobrevivir en el medio. Los descendientes producidos por reproducción asexual forman un clon. MEIOSIS
Ya en 1883 se había descubierto que una especie de gusano tenía 4 cromosomas, en tanto que el huevo (óvulo) y el espermatozoide respectivos contenían sólo dos cromosomas cada uno. La teoría de la herencia se encargó posteriormente de aclarar la aparente paradoja de que dos células de tamaños tan distintos podían contribuir por igual a la información genética del nuevo individuo. Este hallazgo también implicaba que las células germinales (sexuales) debían ser formadas por una clase especial de división nuclear en la cual el número de cromosomas debía reducirse a la mitad. Este tipo de división fue llamada meiosis (del griego: “disminución”), pero no fue sino hasta 1930 que las características esenciales de la meiosis fueron establecidas.
La meiosis involucra dos divisiones nucleares
Con la excepción de los cromosomas que determinan el sexo (los cromosomas sexuales), un núcleo diploide contiene dos versiones muy similares de cada uno de los otros cromosomas (los autosomas), uno que proviene del padre (cromosoma paterno) y otro de la madre (cromosoma materno). Las dos versiones cromosómicas se denominan cromosomas homólogos y en la mayoría de las células mantienen una existencia completamente separada, constituyendo cromosomas independientes. Cuando cada cromosoma es duplicado por replicación del ADN (período S) las copias gemelas de cada cromosoma se mantienen al principio estrechamente asociadas y son denominadas cromátidas hermanas. En una división celular ordinaria (mitosis), cada una de las cromátidas hermanas se separa en la anafase y se convierte en un cromosoma individual, por lo que cada una de las dos células hijas recibirá una copia de cada cromosoma paterno y otra de cada cromosoma materno.
En contraste, una gameta haploide producida por la división de una célula diploide durante la meiosis debe contener la mitad del número original de cromosomas (sólo un cromosoma en lugar de cada par de cromosomas homólogos). Este mecanismo que ha producido la evolución requiere que cada par de homólogos se reconozcan y se apareen físicamente antes de que se ubiquen en el huso mitótico, constituyendo una característica distintiva de la meiosis.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
Este hecho (el reconocimiento de ambos homólogos y su íntimo contacto hasta la metafase en que se separan) es esencial para asegurar la distribución equitativa de los cromosomas homólogos. De este modo uno de los homólogos (el paterno o el materno, al azar) irá a una de las células hijas y el otro a la segunda. De no ser así los cromosomas se distribuirían al azar y una célula hija podría recibir un par de homólogos y carecer en absoluto de una copia de otros cromosomas, distribución que sería letal para ellas.
Tal como se ha planteado, y existiendo un mecanismo por el cual los cromosomas homólogos se reconocen, la meiosis pudo haberse resuelto simplemente suprimiendo la fase de duplicación del ADN (fase S de la mitosis) y separando los homólogos durante la anafase. Por alguna razón aún no clara, el mecanismo meiótico es un poco más complejo. Antes de que los homólogos se apareen, cada uno de los cromosomas individuales produce la replicación de su ADN como en una mitosis normal, formando dos cromátidas hijas, que se conservarán como una estructura única bastante más tiempo que en la mitosis.
Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
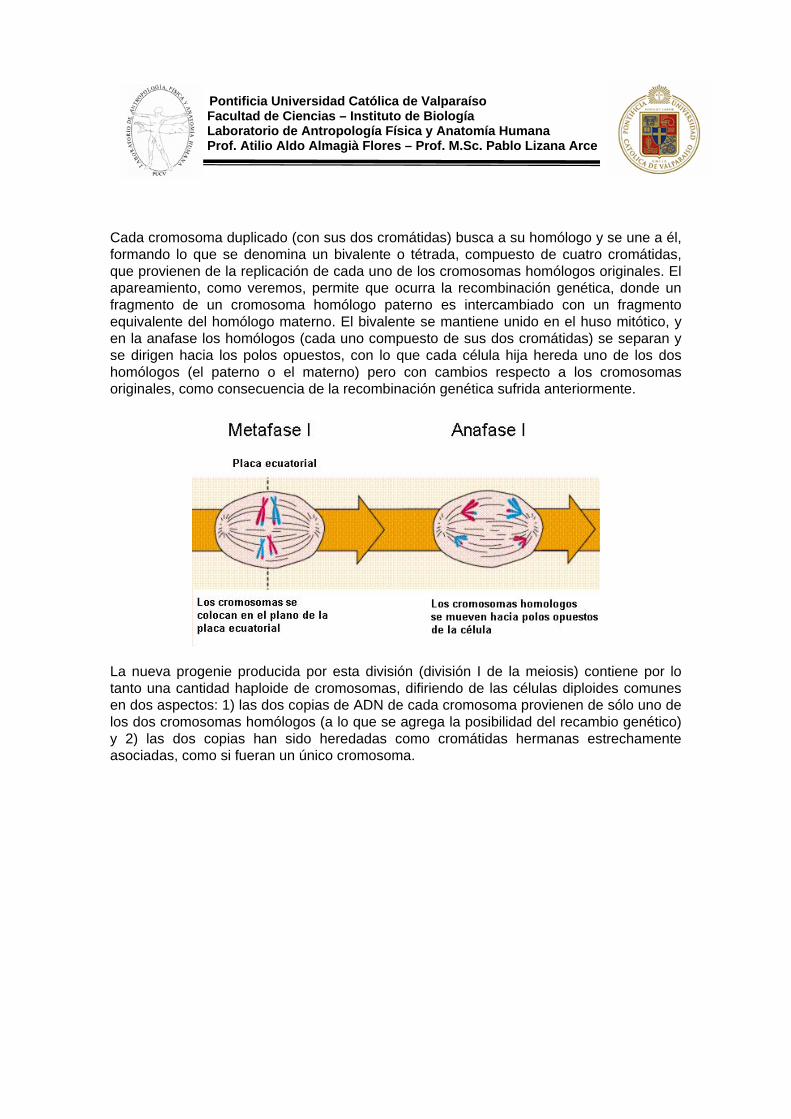
Cada cromosoma duplicado (con sus dos cromátidas) busca a su homólogo y se une a él, formando lo que se denomina un bivalente o tétrada, compuesto de cuatro cromátidas, que provienen de la replicación de cada uno de los cromosomas homólogos originales. El apareamiento, como veremos, permite que ocurra la recombinación genética, donde un fragmento de un cromosoma homólogo paterno es intercambiado con un fragmento equivalente del homólogo materno. El bivalente se mantiene unido en el huso mitótico, y en la anafase los homólogos (cada uno compuesto de sus dos cromátidas) se separan y se dirigen hacia los polos opuestos, con lo que cada célula hija hereda uno de los dos homólogos (el paterno o el materno) pero con cambios respecto a los cromosomas originales, como consecuencia de la recombinación genética sufrida anteriormente.
La nueva progenie producida por esta división (división I de la meiosis) contiene por lo tanto una cantidad haploide de cromosomas, difiriendo de las células diploides comunes en dos aspectos: 1) las dos copias de ADN de cada cromosoma provienen de sólo uno de los dos cromosomas homólogos (a lo que se agrega la posibilidad del recambio genético) y 2) las dos copias han sido heredadas como cromátidas hermanas estrechamente asociadas, como si fueran un único cromosoma.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
Tras una interfase muy corta (intercinesis), y a veces inexistente, tiene lugar la formación de los gametos, que ocurre a través de una segunda división, la división II de la meiosis, sin previa replicación del ADN. Los cromosomas se alinean en un segundo huso y las cromátidas hijas se separan, como en una mitosis normal, para producir células con un contenido haploide de ADN. La meiosis consiste así en dos divisiones celulares con un simple paso de replicación del ADN, como resultado de lo cual se generan cuatro células haploides por cada célula diploide que entra en meiosis.
La recomposición genética está potenciada por el entrecruzamiento entre cromátidas no hermanas de cromosomas homólogos a menos que se trate de gemelos, que se desarrollan a partir de un mismo cigoto, ninguno de los hijos de una pareja de progenitores es igual a otro. Esto es así porque, antes de que se produzca la fusión de los

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce gametos, se han producido dos clases de recomposición genética aleatoria durante la meiosis.
Una clase de recomposición es consecuencia de la distribución aleatoria (al azar) de los cromosomas homólogos maternos y paternos entre las células hijas de la división I de la meiosis, como resultado de la cual cada célula obtiene una mezcla diferente de cromosomas maternos y paternos. Simplemente por este proceso de separación al azar de los homólogos uno puede obtener 2n distribuciones distintas, siendo n el número haploide de cromosomas: así, para la especie humana cada individuo podrá producir 223 = 8,4 x 106 gametos diferentes.
Pero este número de variaciones puede aumentar significativamente como consecuencia de un nuevo tipo de recomposición genética, denominado entrecruzamiento o crossing-over cromosómico, que ocurre durante la división I de la meiosis, en la que se intercambian partes de los cromosomas homólogos. En promedio, ocurren dos a tres entrecruzamientos en cada par de cromosomas humanos durante la meiosis. El proceso de entrecruzamiento implica la ruptura del ADN de una cromátida paterna y lo propio en la zona equivalente de una cromátida materna, con el consiguiente intercambio de fragmentos de una cromátida a otra. Las consecuencias del entrecruzamiento pueden ser observadas citológicamente ya en la profase de la primera división meiótica: las cromátidas hijas están estrechamente unidas en toda su extensión formando los bivalentes y los cromosomas homólogos duplicados (materno y paterno) pueden verse físicamente conectados en puntos específicos denominados quiasmas, correspondientes al sitio en el que se produjo el entrecruzamiento de dos cromátidas no hermanas, cada una perteneciente a cada cromosoma homólogo. En este estadio de la meiosis, cada par de cromosomas homólogos duplicados, o bivalente, posee al menos un quiasma. Muchos bivalentes contienen más de un quiasma, indicando que pueden ocurrir múltiples entrecruzamientos entre homólogos.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce Ocurren elaborados cambios morfológicos en los cromosomas homólogos durante el apareamiento (sinapsis) y desapareamiento (desinapsis) durante la profase I de la meiosis. Esta profase es tradicionalmente dividida en cinco estadios secuenciales: leptoteno, zigoteno, paquiteno, diploteno y diacinesis. Durante el leptoteno, que sobreviene a la interfase, se produce el ensamblado del elemento central del complejo sinaptonémico, que veremos luego, y finaliza la estrecha asociación entre las cromátidas hijas de cada cromosoma homólogo. El evento más importante es la íntima sinapsis cromosómica, que comienza en el zigoteno y que concluye con la formación de una estructura denominada complejo sinaptonémico que implica el contacto de los dos juegos de cromátidas hermanas de cada bivalente. El paquiteno empieza cuando la sinapsis ya es completa y generalmente persiste durante varios días, hasta que comienza la desinapsis en el estadio de diploteno, donde ya pueden verse los quiasmas que se han formado.
El complejo sinaptonémico mantiene a los cromosomas homólogos de cada bivalente totalmente en contacto lineal y consiste en un núcleo proteico (el elemento central) en forma de escalera de mano, a cuyos costados se alinean los dos homólogos. Las cromátidas hermanas de cada homólogo están mantenidas muy juntas a uno de los dos lados del eje proteico y su ADN se encuentra parcialmente extendido formando una serie de bucles.
Si bien el complejo sinaptonémico puede proveer el marco estructural para que ocurra el proceso de recombinación, éste parece mediado por grandes complejos proteicos, denominados nódulos de recombinación, de un diámetro de hasta 90 nm. Los nódulos de recombinación se disponen a intervalos en el eje central del complejo sinaptonémico, que se piensa que funciona como una gran máquina de recombinación enzimática que acerca regiones equivalentes de ADN materno y paterno para que el entrecruzamiento tenga lugar. La evidencia que existe en este sentido es indirecta y se basa en las siguientes

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce observaciones:
1) El número total de nódulos formados es igual al número total de quiasmas observados.
2) La distribución de los nódulos es equivalente a la posición que ocupan los quiasmas; una observación adicional es que los nódulos vecinos nunca se encuentran demasiado cercanos.
3) En mutantes de Drosophila en los que se disminuye la frecuencia de recombinación se observa una correspondiente disminución del número de nódulos.
4) Se piensa que la recombinación genética implica una limitada síntesis de ADN en el lugar del entrecruzamiento. La incorporación de precursores (nucleótidos) marcados revela que los mismos se incorporan preferentemente en la zona de los nódulos o en sus vecindades
Resumen Etapas de Meiosis
Profase I: La replicación del ADN precede el comienzo de la meiosis I. Durante la profase I, los cromosomas homólogos se aparean y forman sinapsis, un paso que es único a la meiosis. Los cromosomas apareados se llaman bivalentes, y la formación de quiasmas causada por recombinación genética se vuelve aparente. La condensación de los cromosomas permite que estos sean vistos en el microscopio. Note que el bivalente tiene dos cromosomas y cuatro cromátidas, con un cromosoma de cada padre.
Prometafase I: La membrana nuclear desaparece. Un cinetocoro se forma por cada cromosoma, no uno por cada cromátida, y los cromosomas adosados a fibras del huso comienzan a moverse.
Metafase I: Bivalentes, cada uno compuesto de dos cromosomas (cuatro cromatidas) se alinean en el plato de metafase. La orientación es al azar, con cada homólogo paterno en un lado. Esto quiere decir que hay un 50% de posibilidad de que las células hijas reciban el homólogo del padre o de la madre por cada cromosoma.
Anafase I: Los quiasmas se separan. Los cromosomas, cada uno con dos cromátidas, se mueven a polos opuestos. Cada una de las células hijas ahora es haploide (23 cromosomas), pero cada cromosoma tiene dos cromátidas.
Telofase I: Las envolturas nucleares se pueden reformar, o la célula puede comenzar rápidamente meiosis II.
Citocinesis: Análoga a la mitosis dónde dos células hijas completas se forman.
La meiosis II es similar a la mitosis. Sin embargo no hay fase "S". Las cromátidas de cada cromosoma ya no son idénticas en razón de la recombinación. La meiosis II separa las cromátidas produciendo dos células hijas, cada una con 23 cromosomas (haploide), y cada cromosoma tiene solamente una cromatida.

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
COMPARACIÓN DE MEIOSIS Y MITOSIS
Comportamiento de los cromosomas
Mitosis: Cromosomas homólogos independientes
Meiosis: Cromosomas homólogos se aparean formando bivalentes hasta anafase I
Número de cromosomas - reducción en meiosis
mitosis- células hijas idénticas
meiosis- células hijas haploides
Identidad genética de la progenie:
Mitosis: células hijas idénticas Meiosis: Las células hijas tienen una nueva variedad de cromosomas paternos. Meiosis: Cromátidas no idénticas, intercambio de segmentos

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce

Pontificia Universidad Católica de Valparaíso Facultad de Ciencias – Instituto de Biología Laboratorio de Antropología Física y Anatomía Humana Prof. Atilio Aldo Almagià Flores – Prof. M.Sc. Pablo Lizana Arce
Bibliografía
Audesirk, T. Audesirk, G. Byers ,B.E.(2003). Biología. La vida en la Tierra. Prentice Hall.
6ª Edición.
Curtis, H. & Barnes, S. (2000).Biología. Ed. Medica Panamericana. 6ª Edición.
Drake, R. Vogl, W. Mitchell, A. (2005). Gray anatomía para estudiantes. Primera edición.
Elsevier España.
Chatain, I. Bustamante, J. (1986). Anatomía Macroscópica Funcional y Clínica.
Addison – Wesley Iberoamericana. Sistemas Técnicos de Edición, S.A. de C.V.
Netter, F.H. (1999). Atlas de Anatomía Humana. Novartis-Masson.
Moore, K.L. (2002). Anatomía con orientación clínica. Médica Panamericana. Cuarta
edición.
Williams, P. (1998). Anatomía de Gray. Churchill Livingstone. 38 edición. Tomo I y Tomo
II.