
Estudio biótico para la delimitación del complejo de ...
127
Estudio biótico para la delimitación del complejo de Páramos de Pisba – Boyacá. Grupo de Estudios en Sistemas Andinos. Coordinador: Carlos Arturo Rocha. Instituto de Investigación de Recursos Biológicos Alexander von Humboldt Universidad pedagógica y tecnológica de colombia Tunja - Boyacá Noviembre 2013
Transcript of Estudio biótico para la delimitación del complejo de ...

Estudio biótico para la delimitación del complejo de Páramos de
Pisba – Boyacá.
Grupo de Estudios en Sistemas Andinos. Coordinador: Carlos Arturo Rocha.
Instituto de Investigación de Recursos Biológicos Alexander von Humboldt
Universidad pedagógica y tecnológica de colombia Tunja - Boyacá
Noviembre 2013

CONTENIDO
INTRODUCCIÓN ........................................................................................................................ 9
1.1. REVISIÓN DE INFORMACIÓN .................................................................................................. 12
1.2. INFORMACIÓN PRELIMINAR DEL SITIO ...................................................................................... 12
1.3. SELECCIÓN DE SITIOS Y UNIDADES DE MUESTREO ....................................................................... 13
2. VEGETACION .................................................................................................................. 16
2.1. METODOLOGÍA .............................................................................................................. 16
2.1.1. Perfiles de vegetación ........................................................................................... 17
2.1.2. Análisis de datos .................................................................................................... 17
2.2. RESULTADOS .................................................................................................................. 19
2.2.1. Gradiente altitudinal para Socotá ......................................................................... 19
2.2.2. Gradiente altitudinal de Socha .............................................................................. 27
2.2.3. Gradiente altitudinal de Tasco .............................................................................. 35
2.2.4. Análisis de datos .................................................................................................... 42
2.2.5. Índices de diversidad por franja altitudinal. .......................................................... 46
2.2.6. Diversidad Beta .................................................................................................... 48
2.2.7. Índice de valor de importancia por gradiente altitudinal ...................................... 51
2.2.8. Índice de valor de importancia por franjas altitudinales ....................................... 54
2.3. DISCUSIÓN ........................................................................................................................ 58
3. ANFIBIOS 61
3.1. INTRODUCCIÓN .............................................................................................................. 61
3.2. METODOLOGÍA .............................................................................................................. 62
3.2.1. Muestreos.............................................................................................................. 62
3.2.2. Análisis de datos. ................................................................................................... 63
3.3. RESULTADOS Y DISCUSION ............................................................................................. 64
3.3.1. Composición .......................................................................................................... 64
3.3.2. Esfuerzo de muestreo ............................................................................................ 64
3.3.3. Clasificación Taxonómica e información ecológica de las Especies de Anuros ..... 64
3.4. ESPECIES ENDÉMICAS........................................................................................................... 67
4. ARTROPODOS ................................................................................................................ 69
4.1. INTRODUCCIÓN .............................................................................................................. 69
4.2. MÉTODOS....................................................................................................................... 70
4.2.1. Muestreo ............................................................................................................... 70
4.3. ANÁLISIS DE DATOS ........................................................................................................ 71
4.3.1. Diversidad Alfa ...................................................................................................... 71
4.3.2. Diversidad Beta ..................................................................................................... 72
4.4. RESULTADOS: ................................................................................................................. 73
4.4.1. Diversidad Alfa ...................................................................................................... 73
4.4.2. Diversidad Beta ..................................................................................................... 86
4.5. DISCUSIÓN ........................................................................................................................ 91
5. AVES 95

5.1. AVES REGISTRADAS POR TRANSECTOS. ......................................................................... 96
5.1.1. AVES REGISTRADAS POR TRANSECTOS ............................................................... 100
5.1.2. SIMILARIDAD DE LAS ESPECIES REGISTRADAS EN LOS TRANSECTOS .................. 102
5.1.3. ESPECIES POTENCIALES EN EL ÁREA DE ESTUDIO. .............................................. 103
6. CONSIDERACIONES FINALES ......................................................................................... 111
6.1. ASPECTOS METODOLÓGICOS ............................................................................................... 111
6.2. ASPECTOS DE LA DELIMITACIÓN ............................................................................................ 111
8. LITERATURA CITADA ..................................................................................................... 118

TABLAS
UBICACIÓN ESTACIONES POR TRANSECTO. .............................................................. 15
REPRESENTATIVIDAD DE GÉNEROS Y ESPECIES DE LAS FAMILIAS REGISTRADAS. ....... 42
ÍNDICES DE DIVERSIDAD POR SITIO. ......................................................................... 46
INDICES DE DIVERSIDAD PARA SOCOTÁ Y SOCHA ..................................................... 47
INDICES DE DIVERSIDAD SEGÚN COTA ALTITUDINAL ................................................ 47
ÍNDICES DE DIVERSIDAD PARA EL GRADIENTE ALTITUDINAL DE SOCHA. ................... 79
ÍNDICES DE DIVERSIDAD PARA EL GRADIENTE ALTITUDINAL DE SOCOTA .................. 79
ÍNDICES DE DIVERSIDAD PARA EL GRADIENTE ALTITUDINAL DE TASCO ..................... 80
ÍNDICES DE DIVERSIDAD PARA LOS DATOS POR COTAS ALTITUDINALES RESUMIDOS.
81
MATRIZ DE ÍNDICES DE SIMILITUD PARA EL GRADIENTE ALTITUDINAL DE SOCHA. .. 88
MATRIZ DE ÍNDICES DE SIMILITUD PARA EL GRADIENTE ALTITUDINAL DE SOCOTÁ. 89
MATRIZ DE ÍNDICES DE SIMILITUD PARA EL GRADIENTE ALTITUDINAL DE TASCO. ... 90
MATRIZ DE ÍNDICES DE SIMILITUD PARA EL CONJUNTO TOTAL DE DATOS. ............. 91
RIQUEZA DE AVES (%) POTENCIALES, DISTRIBUIDAS EN LOS DIFERENTES ÓRDENES. 96
ESPECIES DE AVES ENCONTRADAS EN LOS PUNTOS DE MUESTREO ......................... 97
RIQUEZA DE AVES (%) EN LOS DIFERENTES ÓRDENES REGISTRADOS. ...................... 99
ALTURA A LA QUE FUERON UBICADOS LOS TRANSECTOS PARA EL MUESTREO. .... 100
ÍNDICE DE SIMILARIDAD DE JACCARD EN LOS TRANSECTOS DE MONITOREO. ....... 102
ESPECIES DE AVES OBSERVADASDAS EN LOS PUNTOS DE MUESTREO. .................... 110

FIGURAS
FIGURA 1. PERFIL VEGETACIÓN SOCOTÁ 3200 M. ........................................................... 22
FIGURA 2. PERFIL VEGETACIÓN SOCOTÁ 3100 M. ........................................................... 23
FIGURA 3. PERFIL VEGETACIÓN SOCOTÁ 3000 M. ........................................................... 24
FIGURA 4. PERFIL VEGETACIÓN SOCOTÁ 2900 M. ........................................................... 25
FIGURA 5. PERFIL VEGETACIÓN SOCOTÁ 2800 M. ........................................................... 26
FIGURA 6. PERFIL VEGETACIÓN SOCHA 3200 M. ............................................................. 30
FIGURA 7. PERFIL VEGETACIÓN SOCHA 3100 M. ............................................................. 31
FIGURA 8. PERFIL VEGETACIÓN SOCHA 3000 M. ............................................................. 32
FIGURA 9. PERFIL VEGETACIÓN SOCHA 2900 M. ............................................................. 33
FIGURA 10. PERFIL VEGETACIÓN SOCHA 2800 M. ............................................................. 34
FIGURA 11. PERFIL DE VEGETACIÓN TASCO TACION 3256 M ............................................. 37
FIGURA 12. PERFIL DE VEGETACIÓN TASCO TACION 3157 M ............................................. 38
FIGURA 13. PERFIL DE VEGETACIÓN TASCO TACION 3128 M ............................................. 39
FIGURA 14. PERFIL DE VEGETACIÓN TASCO 3036M ........................................................... 40
FIGURA 15. PERFIL DE VEGETACIÓN TASCO 2925 M .......................................................... 41
FIGURA 16. RIQUEZA DE ESPECIES POR SITIO. ................................................................... 44
FIGURA 17. RIQUEZA DE ESPECIES POR ESTACIÓN PARA CADA GRADIENTE ALTITUDINAL . 45
FIGURA 18. RIQUEZA DE ESPECIES POR COTA ALTITUDINAL. ............................................. 45
FIGURA 19. INDICES DE DIVERSIDAD SEGÚN LA ALTITUD DE LOS TRANSECTOS. EL ÍNDICE DE
SIMPSON INDICADO POR ROMBOS Y EL DE SHANNON-WIENER POR CUADRADOS. ................. 46
FIGURA 20. CLÚSTER DE SIMILITUD BRAY-CURTIS PARA LOS TRANSECTOS. ....................... 48
FIGURA 21. CLÚSTER DE SIMILITUD USANDO ÍNDICE DE JACCARD PARA LOS TRES SITIOS. 49
FIGURA 22. CLUSTER DE SIMILITUD ENTRE FRANJAS ALTITUDINALES USANDO ÍNDICE DE
BRAY – CURTIS. 50
FIGURA 23. CLÚSTER DE SIMILITUD ENTRE FRANJAS ALTITUDINALES USANDO ÍNDICE DE
JACCARD. 51
FIGURA 24. ÍNDICE DE VALOR DE IMPORTANCIA PARA EL GRADIENTE ALTITUDINAL
SOCOTÁ. 52
FIGURA 25. ÍNDICE DE VALOR DE IMPORTANCIA PARA EL GRADIENTE ALTITUDINAL DE
SOCHA. 53
FIGURA 26. ÍNDICE DE VALOR DE IMPORTANCIA PARA EL GRADIENTE ALTITUDINAL DE
TASCO. 54
FIGURA 27. ÍNDICE DE VALOR DE IMPORTANCIA PARA LA FRANJA DE 2875 M. ................. 55

FIGURA 28. ÍNDICE DE VALOR DE IMPORTANCIA PARA LA FRANJA DE 2950 M. ................. 55
FIGURA 29. ÍNDICE DE VALOR DE IMPORTANCIA PARA LA FRANJA DE 3025 M .................. 56
FIGURA 30. ÍNDICE DE VALOR DE IMPORTANCIA EN LA FRANJA 3100 M. .......................... 57
FIGURA 31. ÍNDICE DE VALOR DE IMPORTANCIA PARA LA FRANJA DE 3175 MSNM. .......... 58
FIGURA 32. COMPARACIÓN ENTRE GRADIENTES ALTITUDINALES EN CUANTO AL NÚMERO
DE MORFOTIPOS DENTRO DE CADA GRUPO TAXONÓMICO EN HEXAPODA. ............................ 73
FIGURA 33. COMPARACIÓN ENTRE GRADIENTES ALTITUDINALES PARA LAS ABUNDANCIAS
ABSOLUTAS DENTRO DE CADA GRUPO TAXONÓMICO EN HEXAPODA..................................... 74
FIGURA 34. COMPARACIÓN ENTRE GRADIENTES ALTITUDINALES PARA EL NÚMERO DE
MORFOTIPOS DENTRO DE CADA GRUPO TAXONÓMICO EN ARTRÓPODOS NO INSECTOS. ....... 75
FIGURA 35. COMPARACIÓN ENTRE GRADIENTES ALTITUDINALES PARA LAS ABUNDANCIAS
ABSOLUTAS DENTRO DE CADA GRUPO TAXONÓMICO EN ARTRÓPODOS NO INSECTOS. .......... 75
FIGURA 36. NÚMERO DE MORFOTIPOS PARA EL CONJUNTO TOTAL DE DATOS DENTRO DE
CADA GRUPO TAXONÓMICO EN HEXAPODA. .......................................................................... 76
FIGURA 37. ABUNDANCIAS RELATIVAS PARA EL CONJUNTO TOTAL DE DATOS DENTRO DE
CADA GRUPO TAXONÓMICO EN HEXAPODA. .......................................................................... 77
FIGURA 38. NÚMERO DE MORFOTIPOS PARA EL CONJUNTO TOTAL DENTRO DE CADA
GRUPO TAXONÓMICO EN ARTRÓPODOS NO INSECTOS. ......................................................... 78
FIGURA 39. ABUNDANCIAS RELATIVAS PARA EL CONJUNTO TOTAL DENTRO DE CADA
GRUPO TAXONÓMICO EN ARTRÓPODOS NO INSECTOS. ......................................................... 78
FIGURA 40. TENDENCIAS OBSERVADAS EN LA RIQUEZA DE ESPECIES PARA EL CONJUNTO
TOTAL DE DATOS, DE ACUERDO AL ÍNDICE DE DIVERSIDAD EMPLEADO (1/D, CORRESPONDE AL
EJE SECUNDARIO). .................................................................................................................. 81
FIGURA 41. PERFIL DE DIVERSIDAD DE RENYI PARA SOCHA............................................... 82
FIGURA 42. PERFIL DE DIVERSIDAD DE RENYI PARA SOCOTÁ............................................. 82
FIGURA 43. PERFIL DE DIVERSIDAD DE RENYI PARA TASCO ............................................... 83
FIGURA 44. PERFIL DE EQUITATIVIDAD DE RENYI PARA SOCHA. ........................................ 84
FIGURA 45. PERFIL DE EQUITATIVIDAD DE RENYI PARA SOCOTÁ. ...................................... 84
FIGURA 46. PERFIL DE EQUITATIVIDAD DE RENYI PARA TASCO. ........................................ 85
FIGURA 47. PERFILES DE DIVERSIDAD RENYI PARA LOS DATOS TOTALES POR COTAS
ALTITUDINALES. ..................................................................................................................... 85
FIGURA 48. PERFILES DE EQUITATIVIDAD RENYI PARA LOS DATOS TOTALES POR COTAS
ALTITUDINALES. ..................................................................................................................... 86
FIGURA 49. RIQUEZA DE AVES (%) POTENCIALES, DISTRIBUIDAS EN LOS DIFERENTES
ÓRDENES. 96
FIGURA 50. CURVA DE ACUMULACIÓN DE ESPECIES DE AVES REGISTRADAS. .................... 98

FIGURA 51. RIQUEZA DE AVES (%) EN LOS DIFERENTES ÓRDENES REGISTRADOS. .............. 99
FIGURA 52. DISTRIBUCIÓN PORCENTUAL DE ESPECIES DE AVES EN LOS DIFERENTES
TRANSECTOS. 101
FIGURA 53. ÍNDICE DE SIMILARIDAD DE JACCARD EN LOS TRANSECTOS DE MONITOREO. 102

FOTOS
FOTO 1. ESTACIÓN DE MUESTREO EN EL TRANSECTO SOCOTÁ. ............................................... 20
FOTO 2. VISTA DE UNA PARTE DEL TRANSECTO SOCOTÁ. ........................................................ 21
FOTO 3. ASPECTO DE LA COBERTURA VEGETAL EN UNA ESTACIÓN DEL TRANSECTO SOCHA. ... 28
FOTO 4. HERBAZAL CON PRESENCIA DE ESPELETIA ARGÉNTEA. ............................................... 29
FOTO 5. ASPECTO DE LA VEGETACIÓN EN UNA ESTACIÓN DE MUESTREO EN TASCO. .............. 36
FOTO 6. MATRIZ DEL PAISAJE EN UNA ZONA DEL TRANSECTO TASCO. .................................... 36
FOTO 7. DENDROPSOPHUS LABIALIS ....................................................................................... 65
FOTO 8. SCINAX RUBER ........................................................ ¡ERROR! MARCADOR NO DEFINIDO.

INTRODUCCIÓN
Conocer los límites de los páramos del país a una escala detallada es una tarea
que el Gobierno Nacional ha emprendido con el Plan Nacional de Desarrollo
2011 – 2014 (Ley 1450 de 2011), el cual establece en el artículo 202 que “los
ecosistemas de páramos deberán ser delimitados a escala 1:25.000 basados en
estudios técnicos, económicos, sociales y ambientales adoptados por el
Ministerio de Ambiente, Vivienda y Desarrollo Territorial o quien haga sus veces.
La delimitación será adoptada por dicha entidad mediante acto administrativo”.
El reconocimiento de estos límites involucra entre muchos otros aspectos, la
presencia de vegetación característica de los páramos, no obstante, tal como lo
han planteado algunos autores (Vargas y Pedraza 2004, Rangel-Ch. 2000,
Holtmeier y Broll 2005), los límites en los que empiezan y terminan las diferentes
franjas de vegetación no son fijos, dependen de las condiciones climáticas,
topografía, exposición a las corrientes eólicas, suelos, transformación antrópica y
de la altitud y tamaño de la formación montañosa en donde se encuentran, el

estado actual de transformación de la franja alto andina, el cual puede
distorsionar las interpretaciones al respecto (Rangel – Ch. 2000).
Bader (2007), usa el término “treeline” o “límite forestal montano” para denotar
la transición existente entre el bosque alto andino continuo y la vegetación de
páramo continua. Este autor, señala que el término más apropiado es “treeline
ecotone”, ya que en realidad este “límite forestal montano” es una zona de
transición (ecotono) y no una línea nítida. El límite superior de este ecotono está
definido por el límite inferior de la vegetación continua de páramo y el límite
inferior del ecotono está definido por el límite superior del bosque continuo.
Esto último es la llamada “línea de bosque” o “forest line” en la terminología de
Troll para montañas tropicales (Troll 1959, en Bader 2007).
En los Andes tropicales, varias investigaciones han identificado patrones claros
de cambio en la abundancia de las especies de plantas a lo largo de la zona de
transición entre el bosque y el páramo. Estos estudios florísticos se han basado
en la caracterización de los cambios en la estructura de las comunidades a partir
de una clasificación taxonómica de las especies (Arzac et al., 2011). En su estudio
para la detección de patrones de respuesta de las formas de vida en este
ecotono, (Arzac et al., 2011) resaltan el área de subpáramo (páramos bajos
inmediatamente por encima del límite continuo del bosque) como una zona
particularmente sensible a ser afectada por el cambio climático global, al
constituir la zona potencial de colonización o avance del bosque sobre el
páramo. Llambí et al. (En prensa)), ha encontrado que muestra una alta
diversidad de especies y una afinidad florística relativamente alta en las distintas
localidades estudiadas (al menos hasta el nivel de géneros). Es por ello, la
relevancia de definir esta zona de transición para su conservación
Colombia posee el 49% de los páramos del planeta con una superficie
aproximada de 1.932.987 hectáreas (Defensoría del pueblo 2010). El
departamento de Boyacá, tiene más de 600 mil hectáreas de páramos que

corresponden al 18.3 % del total de área de páramos del país, siendo este el
departamento con mayor representatividad (MMA, 2002). A su vez, las áreas de
páramo de Boyacá se encuentran en un alto grado de transformación debido a la
expansión de la frontera agrícola, actividades pecuarias y actividades mineras
principalmente. Dentro de las zonas protegidas de páramos en el departamento
de Boyacá, se destaca el parque nacional natural de Pisba que representa un 1.4
% de las áreas protegidas del país, abarca un área de 45.000 ha de superficie
aproximada distribuidas en diferentes municipios así: Socotá (79%), Socha (3%),
Tasco (4%) (Provincia Valderrama), Pisba (1,8%) (Provincia La Libertad) y Mongua
(12,2%) (Provincia Sugamuxi). Esta zona es fundamental para las comunidades
presentes como ecosistema estratégico en la generación de servicios
ambientales como almacenamiento y aporte hídrico no sólo para el
departamento sino para los Llanos Orientales y valles interandinos (GESA, 2005).
En este documento se presenta el Estudio Biótico para la delimitación del
complejo de páramos de Pisba – Boyacá, utilizando el componente de flora y
fauna (insectos, aves y anfibios) en tres transectos ubicados en las regiones de
páramo en los municipios de Socotá, Socha y Tasco, dentro de los cuales se
analizó la variación en aspectos ecológicos de las especies presentes a lo largo de
un gradiente altitudinal desde los 2800 a los 3200 m.
La información resultante de este componente, es parte del análisis de la
información procedente de otros criterios tenidos en cuenta para adelantar la
delimitación de las áreas de páramo en Colombia, como de otros sectores de
cordillera (Central y Occidental). No debe considerarse como un criterio único, ni
que posea un peso preliminar a dicho proceso, por lo cual, se debe analizarse en
conjunto con otras fuentes de información.
El efecto de las montañas sobre la distribución y riqueza de los organismos ha
constituido un factor importante para explicar patrones generales de riqueza de
especies ya sea desde el punto de vista biogeográfico (Lomolino 2001) hasta el

punto de vista adaptativo (Hodkinson et al. 2005), y explicado con base en
diferentes modelos que intentan evaluar los patrones observadas en la riqueza
con relación a la altitud (McCain y Grytnes 2010).
Sin embargo, debido al efecto del disturbio, generado en parte, por actividades
humanas, dichos patrones puede ser aparentes, por ejemplo, eventos de
disturbio a elevaciones bajas puede incrementar o concentrar la riqueza de
especies hacia elevaciones altas en donde las comunidades biológicas puedan
sufrir un reemplazo de especies especialistas por especies generalistas (McCain
& Grytnes 2010, Nogués-Bravo et al. 2008, Becker et al. 2007).
1.1. Revisión de información
Se realizó la revisión preliminar de la información secundaria de estudios de flora
y otros elementos de la vegetación para el área de estudio, sectores adyacentes
y cambios en gradientes altitudinales entre bosque alto andino y páramo. Se
identificó la información de planificación territorial, como Planes de
Ordenamiento Territorial, EOT, Planes de Ordenación y Manejo de Cuencas
Hidrográficas, cartografía del área de estudio de las diferentes entidades
nacionales y regionales.
Se examinó la cartografía existente, así como la cartografía del Atlas de Páramos
a escala 1:100.000 elaborado por el Instituto Alexander von Humboldt como un
insumo técnico para la delimitación de páramos a escala 1:25.000, con base en
esto se establecieron gradientes altitudinales y sus respectivas estaciones de
muestreo.
1.2. Información preliminar del sitio
Las regiones muestreadas, se caracterizan por tener niveles de inclinación altos;
desde el punto de vista ecológico, la unidad biogeográfica de las tres zonas
muestreadas (Socotá, Socha y Tasco) está representada por un 35% de áreas con

potreros, un 15% de áreas cultivadas, un 40% de áreas que son utilizadas para
procesos propios de las minas de carbón, donde se almacenan materiales y
desechos de las mismas y existe otro 10% de áreas utilizadas para la extracción
maderera en su mayoría con manejo de quema; por tal razón actualmente los
anfibios están sometidos a drásticos cambios en su hábitat, afectando sus
poblaciones. Estos cambios en el ambiente se deben al aumento de la frontera
ganadera y agrícola sumándose la minería como fuente de empleo y “desarrollo”
de las zonas, ocasionando fragmentación y pérdida del hábitat, sumado a esto,
las enfermedades, la contaminación y el calentamiento global (Rueda et ál.
2010).
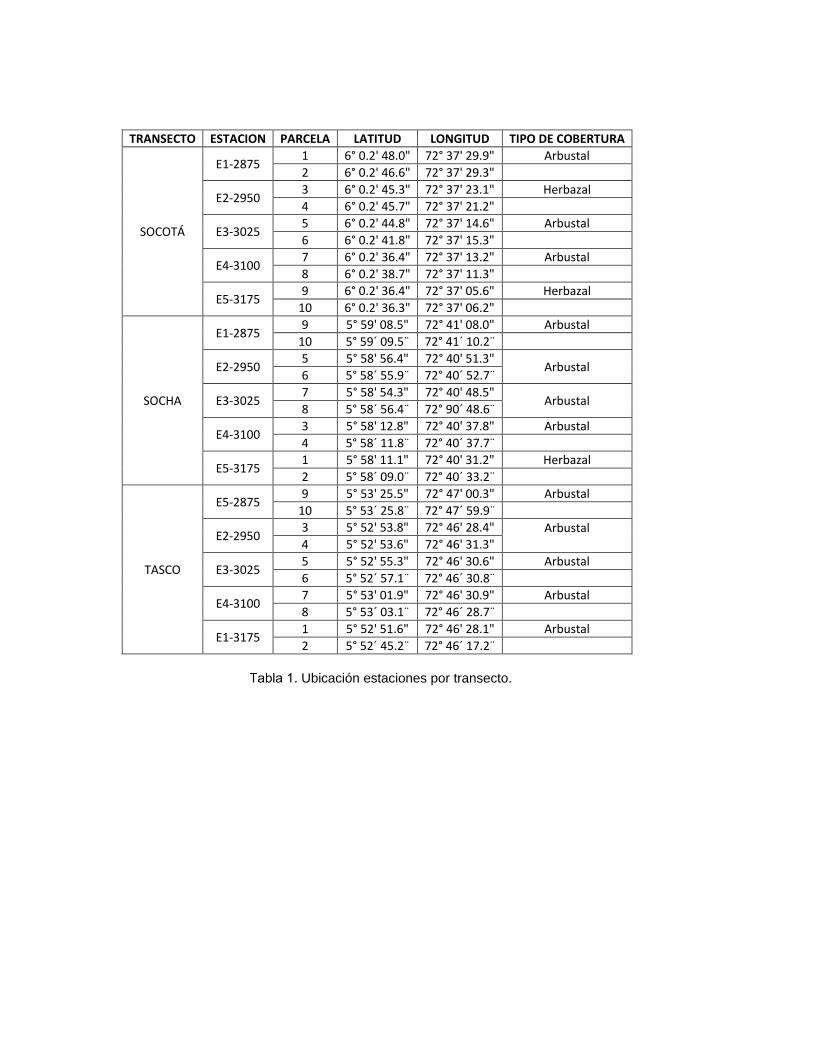
1.3. Selección de sitios y unidades de muestreo
Con base en imágenes satelitales se definieron tres gradientes altitudinales,
correspondientes a los municipios de Socotá, Socha y Tasco y definiendo a lo
largo de los mismos y cada 75 m estaciones de muestreo (Tabla 1), de acuerdo a
los siguientes criterios:
Tipo de cobertura de vegetación.
Tipo de uso de suelo, esto significa, áreas donde no se evidencia procesos
extractivos o actividades que ocasionen cambios de la cobertura vegetal.
Además de la presencia de caminos, vías, cañadas o quebradas que
favorecen el ascenso del bosque, lo que podría causar la toma de
decisiones sobre un límite de condiciones azonales.
En cada estación de muestreo se realizaron dos parcelas de tamaño según el tipo
de cobertura, separadas entre sí, a una distancia de 10 m y cada una dividida en
cinco subparcelas en relación a su tamaño. Para el levantamiento de las parcelas
al interior de las estaciones de muestreo se siguió la propuesta de Marín (2013)
quién sugiere para vegetación arbórea, con un área de 100 m2 parcelas de 4 m x
25 m divididas en 5 subparcelas de 4 m x 5 m y para vegetación tipo arbustal y

herbazal (hierbas, pajonales, frailejonales), con áreas de 50 m2 parcelas de 4 m x
12,5 m divididas en 5 subparcelas de 4 m x 2,5 m. Se registró en un formato de
levantamientos datos de la localidad (departamento, municipio, vereda,
quebrada, finca, etc.), coordenadas geográficas, altitud, inclinación y exposición
orográfica en cada una de las parcelas (León 2003).
TRANSECTO ESTACION PARCELA LATITUD LONGITUD TIPO DE COBERTURA
SOCOTÁ
E1-2875 1 6° 0.2' 48.0" 72° 37' 29.9" Arbustal
2 6° 0.2' 46.6" 72° 37' 29.3"
E2-2950 3 6° 0.2' 45.3" 72° 37' 23.1" Herbazal
4 6° 0.2' 45.7" 72° 37' 21.2"
E3-3025 5 6° 0.2' 44.8" 72° 37' 14.6" Arbustal
6 6° 0.2' 41.8" 72° 37' 15.3"
E4-3100 7 6° 0.2' 36.4" 72° 37' 13.2" Arbustal
8 6° 0.2' 38.7" 72° 37' 11.3"
E5-3175 9 6° 0.2' 36.4" 72° 37' 05.6" Herbazal
10 6° 0.2' 36.3" 72° 37' 06.2"
SOCHA
E1-2875 9 5° 59' 08.5" 72° 41' 08.0" Arbustal
10 5° 59´ 09.5¨ 72° 41´ 10.2¨
E2-2950 5 5° 58' 56.4" 72° 40' 51.3"
Arbustal 6 5° 58´ 55.9¨ 72° 40´ 52.7¨
E3-3025 7 5° 58' 54.3" 72° 40' 48.5"
Arbustal 8 5° 58´ 56.4¨ 72° 90´ 48.6¨
E4-3100 3 5° 58' 12.8" 72° 40' 37.8" Arbustal
4 5° 58´ 11.8¨ 72° 40´ 37.7¨
E5-3175 1 5° 58' 11.1" 72° 40' 31.2" Herbazal
2 5° 58´ 09.0¨ 72° 40´ 33.2¨
TASCO
E5-2875 9 5° 53' 25.5" 72° 47' 00.3" Arbustal
10 5° 53´ 25.8¨ 72° 47´ 59.9¨
E2-2950 3 5° 52' 53.8" 72° 46' 28.4" Arbustal
4 5° 52' 53.6" 72° 46' 31.3"
E3-3025 5 5° 52' 55.3" 72° 46' 30.6" Arbustal
6 5° 52´ 57.1¨ 72° 46´ 30.8¨
E4-3100 7 5° 53' 01.9" 72° 46' 30.9" Arbustal
8 5° 53´ 03.1¨ 72° 46´ 28.7¨
E1-3175 1 5° 52' 51.6" 72° 46' 28.1" Arbustal
2 5° 52´ 45.2¨ 72° 46´ 17.2¨
Ubicación estaciones por transecto.

2. VEGETACION
2.1. METODOLOGÍA
Para cada especie vegetal, con un diámetro basal ≥ 2 cm a 30 cm del suelo,
dentro de cada parcela se tomaron los siguientes datos:
Altura total.
Diámetro del tallo a 30 cm de altura: para individuos con tallo ramificado
desde la base (arbustos), se midió el diámetro de cada una de las
ramificaciones y posteriormente se sumaron las áreas basales de cada
ramificación (Villáreal et al., 2006).
Cobertura de copa (m2): Se registró la medida de los diámetros mayores y
menor, asumiendo la forma ovalada en las copas de los árboles, arbolitos
y arbustos (Prieto 1994 citado en Rangel y Velásquez 1997) y se calculó la
cobertura así: 𝐶 =1
2(𝑑1 × 𝑑2), donde: d1: diámetro mayor y d2:
diámetro menor.
Hábito (árbol, arbusto, roseta, hierba, epífita).
Estrato: Herbáceo: 0,31 – 1,5 m; Arbustivo: 1,51 – 5 m y Arbóreo >5 m,
según lo propuesto por Rangel y Velásquez (1997).
En el caso de macollas, gramíneas y otras especies herbáceas cuya
individualización no fue posible se registró:
Altura total.
Cobertura de copa.
Porcentaje de cobertura con respecto a cada subparcela.
Hábito.

La colección de muestras de los ejemplares botánicos registrados en las planillas
de los levantamientos en campo se hicieron siguiendo lo propuesto por Villáreal
et al., (2004). Se tomó fotografía a cada ejemplar in situ y posterior al secado en
mufla, posteriormente se identificaron las especies vegetales.
Se colectaron entre dos y tres ejemplares, de los cuales uno de ellos en estado
fértil. La información adicional registrada en campo (nombres comunes, usos,
etc.) fue consignada en las fichas botánicas para los ejemplares de herbario.
La información tomada en campo se registró y organizo según el formato para
levantamientos en campo. En este caso, se tuvo en cuenta las recomendaciones
para el depósito de colecciones biológicas disponible en:
http://www.humboldt.org.co/iavh/documentos/colecciones_biologicas/Protocol
o_deposito_de_ejemplares.pdf.
2.1.1. Perfiles de vegetación
Para cada tipo de cobertura se elaboraron perfiles verticales de la vegetación a
escala, con base en los levantamientos realizados. Para ello se localizó un punto
de coordenada (X, Y) estimada en metros en cada subparcela, con el fin de ubicar
el o los individuos según la disposición observada en campo, teniendo en cuenta
la inclinación del terreno, morfología de las especies y altura.
2.1.2. Análisis de datos
2.1.2.1. Diversidad alfa
Se totalizó el número de especies para los tres gradientes altitudinales en
conjunto y de manera separada, el número de especies por cota altitudinal a lo
largo de los tres gradientes y el número de especies por estación de muestreo
dentro de cada gradiente. Para estimar la diversidad alfa, ya sea por gradiente
altitudinal, parcela y cota altitudinal a lo largo de cada gradiente, se empleó el
Índice de Shannon-Wiener, 𝑯′ = − ∑ 𝑷𝒊 × 𝒍𝒏 𝑷𝒊 , donde Pi es la abundancia

relativa y el Índice de Simpson,𝑺 = 𝟏 ∑(𝒏𝒊(𝒏𝒊−𝟏)
𝑵(𝑵−𝟏))⁄ , donde, ni es número de
individuos en la iésima especie y N el número total de individuos, dado que al
aumentar S la riqueza disminuye, se emplea al inverso de Simpson como 1/S. los
índices fueron calculados empleado el programa PAST (Hammer 2001).
2.1.2.2. Diversidad beta
Para evaluar la diversidad beta se analizó que tan semejantes son las
comunidades entre estaciones para cada uno de los gradientes altitudinales
empleando el Índice de similitud de Bray-Curtis (Krebs 1986), el cual tiene en
cuenta las abundancias, así,
𝐷𝐴−𝐵 = ∑(𝑥𝐴𝑖 − 𝑥𝐵𝑖)/ ∑(𝑥𝐴𝑖 + 𝑥𝐵𝑖), donde XAi es el número de individuos en el
sitio A y XBi es el N¡ número de individuos en el sitio B y el Índice de Jaccard el
cual se basa sobre la presencia/ausencia de un individuo en una muestra, así,
𝐼𝑗 = 𝑐/(𝑎 + 𝑏 + 𝑐), donde a es el número de especies presentes en la estación
A, b es el número de especies presentes en la estación B y c es el número de
especies presentes en ambas estaciones, A y B (Magurran 2005). Los índices de
similitud fueron calculados empleando el programa PAST (Hammer 2001).
Junto a los análisis de diversidad alfa y beta se estimó la densidad de especies,
como el número de individuos por parcela; frecuencia relativa como la relación
entre el número de subparcelas en la que se una especie y el número total de
subparcelas; áreas basales transformando los valores del diámetro o
circunferencia basal a 30 cm del suelo a valores de área con la fórmula del área
del círculo, así 𝑨 = 𝝅𝒓(𝒅
𝟐)𝟐, donde d es el diámetro del círculo, sabiendo que 𝑳 =
𝝅𝐝, donde L es la longitud de la circunferencia.
Por último, se calculó el Índice de valor de importancia (IVI), densidad, frecuencia
y dominancia relativa, así (Stiling 1999; Lamprecht 1990): IVI = DeR +DoR+FR

donde, DeR es la densidad relativa; DoR, dominancia relativa y FR la frecuencia
relativa. Para el cálculo de la densidad relativa se tuvo en cuenta la siguiente
fórmula: DeR= (Ei /SE) x 100, donde Ei es el número de ocurrencias de la especie
y SE el número total de individuos y la dominancia relativa se expresa como valor
relativo de la sumatoria de las áreas basales de la siguiente manera:
DR = (ABi /SAB) x 100, donde, SABi es la sumatoria de las áreas basales o
coberturas relativas de la especie i y SAB es la sumatoria de las áreas basales o
coberturas relativas de todas las especies en la muestra
La frecuencia relativa se calculó así: FR = (Fi /SF) x 100, donde, Fi es el número de
subparcelas donde la especie i ocurre y SF es la sumatoria total de ocurrencias de
todas las especies en todas las subparcelas.
2.2. RESULTADOS
Dentro de la zona se observó un paisaje totalmente heterogéneo, compuesto por
parches de vegetación que resguardan elementos vegetales asociados a
comunidades nativas conformando un paisaje con diferentes usos y coberturas,
corredores, fragmentos de matorrales y herbazales que constituyen un tipo de
vegetación con porte medio y dosel abierto. Las localidades presentan un alto
grado de intervención, principalmente por la expansión de la frontera agrícola,
minería y extracción de madera, por lo cual se consideran áreas con deterioro
ambiental y fragmentado. A continuación se da una descripción general de cada
uno de los sitios donde se establecieron las parcelas de muestreo.
2.2.1. Gradiente altitudinal para Socotá
Las estaciones se ubicaron en los sitios que muestra la foto 1. La zona se
caracteriza por tener un predominio de matorrales de dosel abierto y con
individuos que no sobrepasan los 2,5 m de altura. En el gradiente se observó una
matriz de vegetación continua, constituida por una mezcla entre parches de

matorral (la mayoría de las estaciones) y herbazales. Dominan las especies que
ocupan el estrato arbustivo, principalmente Miconia squamulosa, Cletha
fimbriata, Morella parvifola y Pentacalia corymbosa. En el estrato herbáceo se
destacan Cuphea sp., Agrostis sp., Cyperus sp. y Senecio sp. El estrato arbóreo o
presencia de cobertura boscosa no fue dominante en este gradiente (fotos 2 y 3).
Ubicación de las cinco estaciones de muestreo en el gradiente
altitudinal de Socotá.
Estación de muestreo en el gradiente altitudinal de Socotá.

Vista de una parte del gradiente altitudinal de Socotá.

Figura 1. Perfil Vegetación Socotá 3200 m.
1. Hypericum strictum
2. Calea pennellii
3. Pentacalia corymbosa
7. Clethra fimbriata
8. Miconia squamulosa
10. Ageratina tinifolia
17. Eryngium humboldtii
20. Baccharis tricuneata
33. Gaultheria bracteata
36. Miconia cundinamarcensis
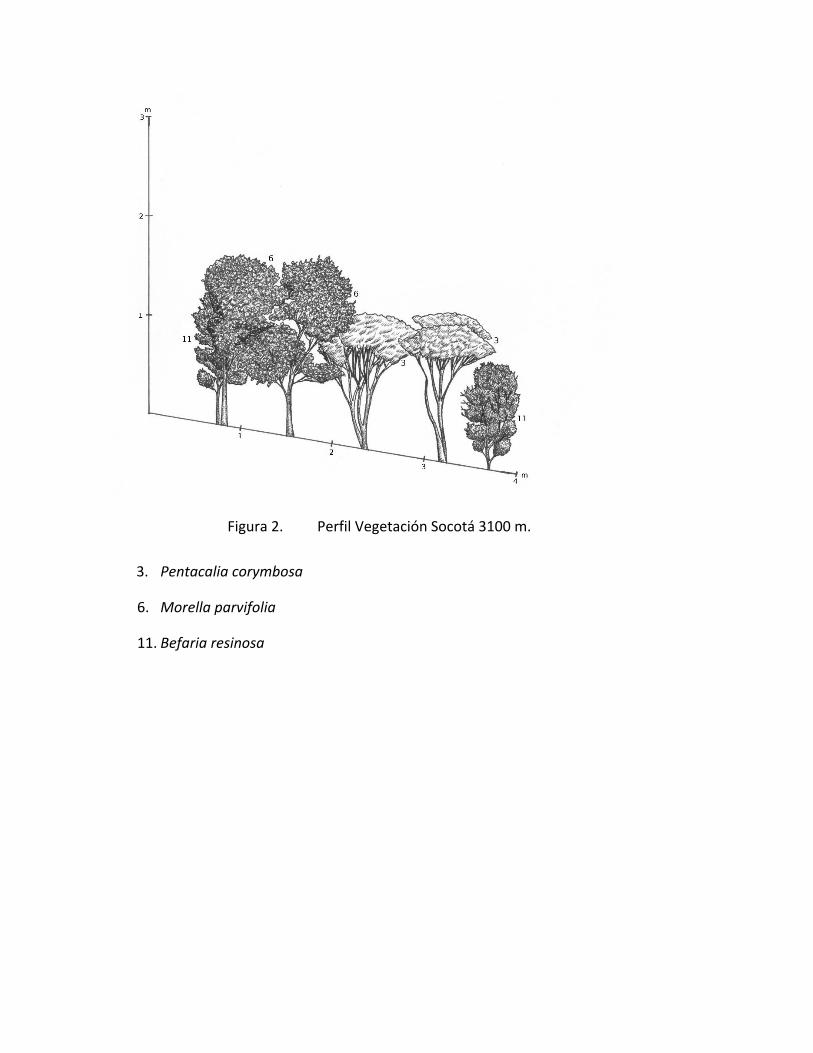
Figura 2. Perfil Vegetación Socotá 3100 m.
3. Pentacalia corymbosa
6. Morella parvifolia
11. Befaria resinosa

Figura 3. Perfil Vegetación Socotá 3000 m.
3. Pentacalia corymbosa
8. Miconia squamulosa
11. Befaria resinosa

Figura 4. Perfil Vegetación Socotá 2900 m.
1. Hypericum strictum
8. Miconia squamulosa
9. Baccharis floribundum
15. Dodonaea viscosa
17. Eryngium humboldtii
18. Hypericum mexicanum
19. Senecio sp
20. Baccharis tricuneata

Figura 5. Perfil Vegetación Socotá 2800 m.
6 Morella parvifolia
7. Clethra fimbriata
8. Miconia squamulosa
13. Myrcianthes leucoxyla
16. Berberis glauca

2.2.2. Gradiente altitudinal de Socha
La matriz del paisaje está formada por parches de vegetación muy pequeños,
factor que no permitió ubicar un área con vegetación continua lo
suficientemente grande para abarcar todo el gradiente, por esto las estaciones
se ubicaron en tres zonas distintas como se observa en la foto 4. Las estaciones
ubicadas a menor altitud (2875 y 2950 m) fueron en las que se observó
intervención humana con pastoreo para ganado. Las estaciones a mayor altitud
presentan un mejor estado de conservación, debido a que son de difícil acceso y
el terreno presenta mayor grado de inclinación.
En el gradiente altitudinal de Socha se observó el predominio de matorrales con
individuos de alturas promedio de 1.82 m, donde domina claramente el estrato
arbustivo, pero se observan algunos elementos arbóreos emergentes con alturas
de hasta 5 m. Las especies dominantes en los arbustales son Miconia
cundinamarcensis, Symplocos theiformis, Miconia squamulosa y Morella
parvifolia. Para las zonas de herbazal registradas las especies dominantes son
Calamagrostis effusa, Agrostis boyacensis y Chaetolepys microphyla. Este sitio
fue el que presento mayor riqueza diversidad y abundancia de especies y el
mayor número de especies exclusivas como Polypodium murorum, Juncus aff.
equadoriensis y Lycopodyum jusiiaei. Así mismo, fue la única zona donde se
registró una especie de frailejón Espeletia argentea, la cual hace parte de un área
que refleja el fenómeno de paramización. El estrato arbóreo o presencia de
cobertura boscosa no fue dominante en este gradiente (fotos 5 y 6).

Ubicación de las cinco estaciones de muestreo en el transecto Socha.
Aspecto de la cobertura vegetal en una estación del gradiente altitudinal
de Socha.

Herbazal con presencia de Espeletia argéntea.

Figura 6. Perfil Vegetación Socha 3200 m.
1 Hypericum strictum
3. Pentacalia corymbosa
17. Eryngium humboldtii
35. Bucquetia glutinosa
39. Vaccinium floribundum
42. Bucquetia glutinosa
44. Espeletia argentea
45. Habenaria sp
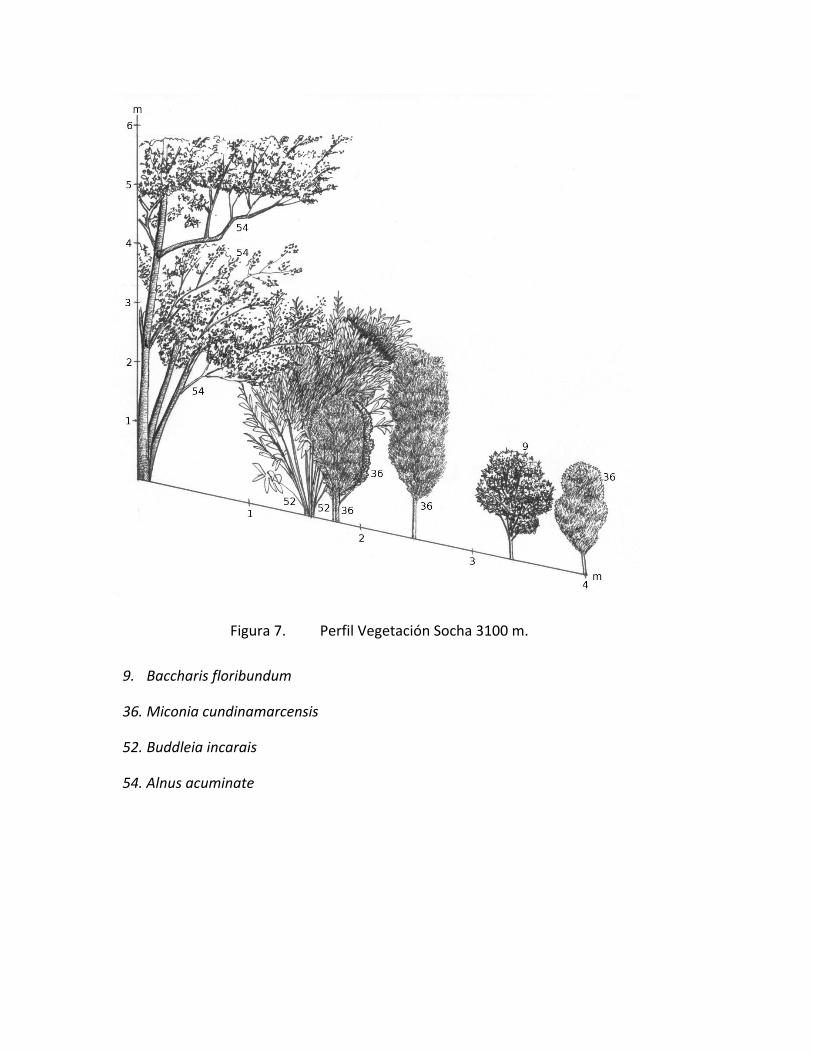
Figura 7. Perfil Vegetación Socha 3100 m.
9. Baccharis floribundum
36. Miconia cundinamarcensis
52. Buddleia incarais
54. Alnus acuminate

Figura 8. Perfil Vegetación Socha 3000 m.
7. Clethra fimbriata
8. Miconia squamulosa
9. Baccharis floribundum
57. Viburnum tinoides
61. Psycotria tatamana

Figura 9. Perfil Vegetación Socha 2900 m.
9. Baccharis floribundum
36. Miconia cundinamarcensis
52. Buddleja incana
54. Alnus acuminata

Figura 10. Perfil Vegetación Socha 2800 m.
4. Rapanea guianensis
6. Morella parvifolia
9. Baccharis floribundum
24. Lippia hirsuta
57. Viburnum tinoides
59. Symplocus theiformis
64. Phyllanthus salviifolius

2.2.3. Gradiente altitudinal de Tasco
En la foto 7 se muestra la ubicación de las estaciones de muestreo. El gradiente
para Tasco, presenta un conjunto de pequeños parches de matorrales, con
dominancia del estrato arbustivo en el cual se registraron individuos con alturas
promedio de 1.91 m. Son formaciones vegetales con dosel abierto, individuos
con tallos ramificados. Hay una dominancia marcada por Clethra fimbriata en la
zona, otras especies representativas en cuanto a dominancia, cobertura y
frecuencia son Pentacalia corymbosa, Miconia squamulosa y Bucquetia
glutinosa. Es el gradiente que presentó la menor diversidad dentro del área de
estudio. El estrato arbóreo o presencia de cobertura boscosa no fue dominante
en este gradiente (fotos 8 y 9).
Ubicación de las cinco estaciones de muestreo en el gradiente
altitudinal de Tasco.

Aspecto de la vegetación en una estación de muestreo en Tasco.
Matriz del paisaje en una zona del transecto Tasco.

Figura 11. Perfil de vegetación Tasco tacion 3256 m
3. Rapanea guianensis
8. Miconia squamulosa
9. Baccharis floribundum
15. Dodonaea viscosa
60. Baccharis bogotensis

Figura 12. Perfil de vegetación Tasco tacion 3157 m
1. Pentacalia corymbosa
8. Miconia squamosa
10. Ageratina tinifolia
36. Miconia cundinamarcensis
57. Viburnum tinoides

Figura 13. Perfil de vegetación Tasco tacion 3128 m
3 Pentacalia corymbosa
7. Clethra fimbriata
11. Befaria resinosa
62. Diplostephium rosmarinifolium

Figura 14. Perfil de vegetación Tasco 3036m
2. Pentacalia corymbosa
14. Verbesina centroboyacana
25 Myrcianthes sp.
29 Ilex kunthiana
30 Miconia cundinamarcensis

Figura 15. Perfil de vegetación Tasco 2925 m
5. Morella parvifolia
34. Arcythophylum nitidum
36. Miconia cundinamarcensis
39. Vacinium foribundum
55. Weinmannia tomenstosa

2.2.4. Análisis de datos
2.2.4.1. Diversidad Alfa
En los tres gradientes altitudinales se registraron un total de 62 especies
vegetales distribuidas en 38 familias y 55 géneros. (Anexo Tabla 2.), con
elementos característicos de zonas de páramo bajo (entre 3.200 y 3.500 (3.600)
m) (Pedraza-Peñaloza et al., 2004; Madriñan 2010), vegetación arbustiva
predominante, matorrales dominados por especies de Diplostephium,
Pentacalia, Hypericum, Pernettya, Vaccinium, Befaria y Gaultheria; y franja alto
andina (entre 3.000 y 3.200 m), la cual constituye una zona de ecotonía entre la
vegetación cerrada de bosques o selva de la media montaña y la vegetación
abierta de matorrales y pajonales de la parte alta. Según la literatura las
comunidades incluyen bosques altos dominados por especies de Miconia,
Weinmannia (encenillos) y Hesperomeles (mortiños), entre otros tipos de
vegetación, reflejando zonas de contacto con la vegetación de la región de la
media montaña conformando comunidades mixtas, en el estudio no se
encontraron, como se hab´pia ya mencionado (IAvH, 2011).
Familia No géneros No especies
Asteraceae 10 12
Melastomataceae 3 5
Poaceae 3 4
Ericaceae 3 3
Rubiaceae 3 3
Buddlejaceae 1 2
Myrtaceae 1 2
Otras familias 31 31
Representatividad de géneros y especies de las familias registradas.
Las familias más representativas en cuanto a número de géneros y especies
registradas en los tres sitios fueron: Asteraceae, Melasatomataceae, Poaceae,
ericaceae y Rubiaceae (Tabla 2), lo cual coincide con lo registrado por Rangel
(2003) y (Keating, 1999) para la región de vida paramuna, la cual se define como

una zona que comprende territorios que coronan las cordilleras entre el bosque
andino y el límite inferior de las nieves perpetuas. Está definida como región
natural por la relación entre el suelo, el clima, la biota y la influencia humana. Así
mismo se pudo observar la espesa capa de materia orgánica de los suelos, en
algunos casos mayor de 1 m de profundidad. Hay periodos contrastantes que se
alternan, noches frías, húmedas y días muy asoleados, en algunos casos con
radiación intensa. La temperatura media anual fluctúa entre 4° y 10°C (8°C). En la
franja baja (subpáramo) se alcanzan temperaturas ente 8° y 10°C (Sturm, 1998).
El anexo 2., muestra las especies registradas en cada altitud para cada uno de los
sitios. Socha es la localidad con un mayor número de especies exclusivas
(especies que solo se registraron en ese transecto) y también presenta el mayor
número de especies registradas en total. Socotá le sigue en el número de
especies de acuerdo a los atributos mencionados anteriormente y finalmente
Tasco el cual se presenta con tan solo una especie exclusiva y el menor número
de especies registradas.
Se registra un frailejón (Espeletia argentea), como elemento asociado a
comunidades de páramo en franjas altitudinales sobre los 3200 m; en Socha a los
3175 m hace parte de una zona de herbazal formando un parche aislado, esto
puede interpretarse como un indicador del fenómeno de paramización (Kok et
al., 1995; Hernández-Camacho 1997; Van der Hammen 1997; Cortez et al., 2003;
Velasco-Linares y Vargas 2008, en González et al.,2011) o Páramo azonal (IAvH,
2011) representado por Vegetación paramuna que se desarrolla fuera de las
condiciones climáticas y edáficas dominantes (IAvH, 2011) donde se evidencia una
tendencia a la sustitución de la vegetación del bosque por la del páramo en áreas
intervenidas.
El gradiente con mayor riqueza de especies fue Socha con 46 especies, seguido
de Socotá con 42 especies y finalmente tasco con 29 especies. (Figura 1).
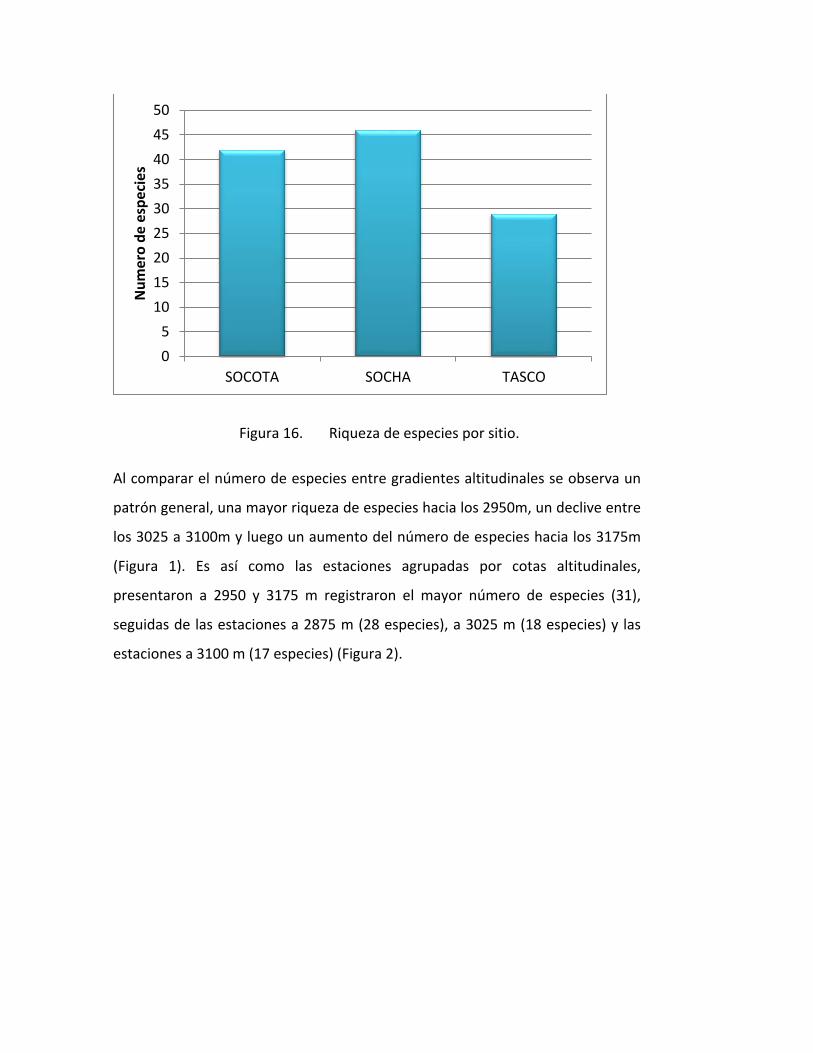
Figura 16. Riqueza de especies por sitio.
Al comparar el número de especies entre gradientes altitudinales se observa un
patrón general, una mayor riqueza de especies hacia los 2950m, un declive entre
los 3025 a 3100m y luego un aumento del número de especies hacia los 3175m
(Figura 1). Es así como las estaciones agrupadas por cotas altitudinales,
presentaron a 2950 y 3175 m registraron el mayor número de especies (31),
seguidas de las estaciones a 2875 m (28 especies), a 3025 m (18 especies) y las
estaciones a 3100 m (17 especies) (Figura 2).
0
5
10
15
20
25
30
35
40
45
50
SOCOTA SOCHA TASCO
Nu
me
ro d
e e
spe
cie
s
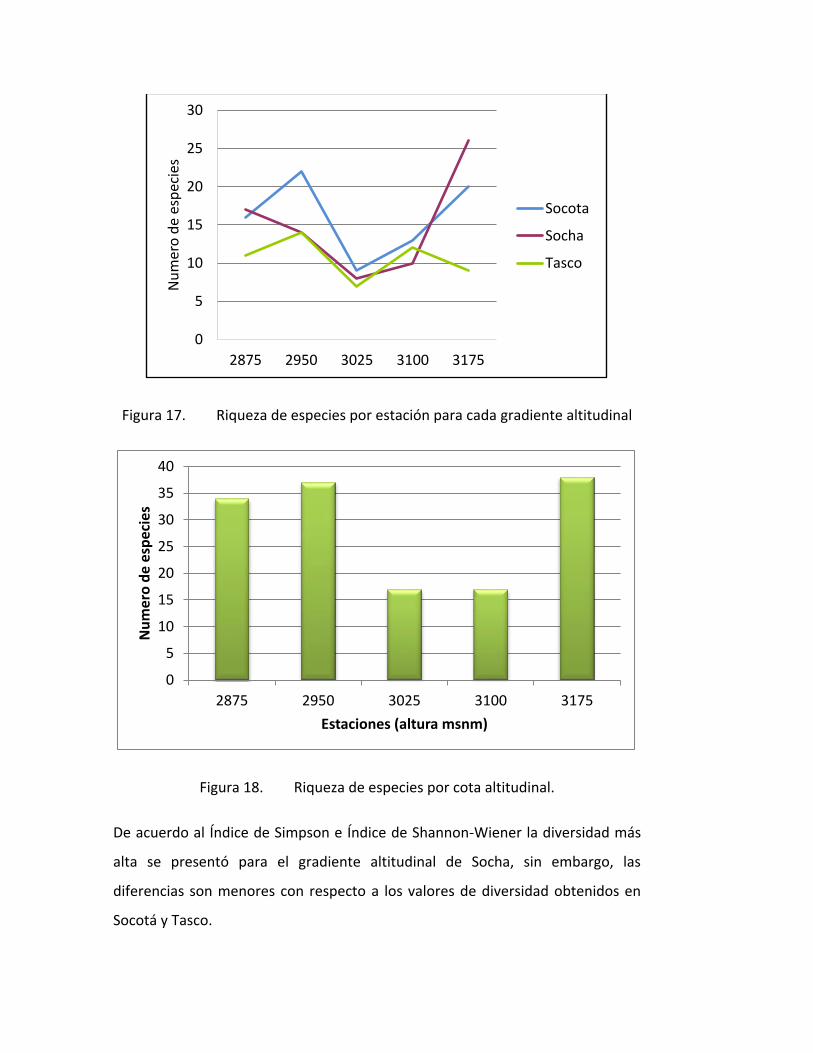
Figura 17. Riqueza de especies por estación para cada gradiente altitudinal
Figura 18. Riqueza de especies por cota altitudinal.
De acuerdo al Índice de Simpson e Índice de Shannon-Wiener la diversidad más
alta se presentó para el gradiente altitudinal de Socha, sin embargo, las
diferencias son menores con respecto a los valores de diversidad obtenidos en
Socotá y Tasco.
0
5
10
15
20
25
30
2875 2950 3025 3100 3175
Nu
mer
o d
e es
pec
ies
Socota
Socha
Tasco
0
5
10
15
20
25
30
35
40
2875 2950 3025 3100 3175
Nu
me
ro d
e e
spe
cie
s
Estaciones (altura msnm)

Gradiente Altitudinal Simpson Shannon-Wiener
Socotá 0,9125 2,839
Socha 0,9324 2,983
Tasco 0,9154 2,786
Índices de diversidad por sitio.
Al contrastar la diversidad de especies entre cotas altitudinales, ya sea por medio
del Índice de Simpson o Índice de Shannon-Wiener, se obtuvieron valores altos
entre los 2875 a 2950 m, seguido por un declive en la diversidad y luego un
aumento de estos valores hacia la cota de 3175 (Figura 19)
2.2.5. Índices de diversidad por franja altitudinal.
Figura 19. Indices de diversidad según la altitud de los transectos. El índice
de Simpson indicado por rombos y el de Shannon-Wiener por cuadrados.
Los valores de abundancia, riqueza de especies e índices de diversidad tienen un
comportamiento similar a lo largo del gradiente, las estaciones a menor altura
(2875 y 2950) y la de mayor altura (3175) son las zonas con mayor número de
individuos, número de especies y diversidad. Mientras que las zonas intermedias
tienen valores más bajos. Esto muestra que no existen unos atributos ideales de
la vegetación a lo largo del gradiente, pues hay una discontinuidad que es
0,83
0,84
0,85
0,86
0,87
0,88
0,89
0,9
0,91
0,92
0,93
0
0,5
1
1,5
2
2,5
3
3,5
2875 2950 3025 3100 3175Ín
dic
e d
e S
imp
son
Índ
ice
de
Sh
ann
on
Altitud (msnm)
reflejada en los valores estudiados, indicando una heterogeneidad de individuos
y especies posiblemente debido a procesos de intervención.
Para las especies herbáceas, se calcularon los índices de diversidad teniendo en
cuenta el porcentaje de cobertura, según esto, Socotá presento valores altos de
diversidad de especies ya sea para el Índice de Simpson o Índice de Shannon-
Wiener, no obstante, las diferencias entre valores son escasas (tabla 4) lo cual
ocurre al comparar la diversidad de especies de herbáceas entre cotas
altitudianles, donde los valores obtenidos, son el resultado del efecto del tamaño
de las muestras (tabla 5)
Gradiente Altitudinal Simpson Shannon-Wiener
Socotá 0,810 1.76
Socha 0,733 1.64
Indices de diversidad para Socotá y Socha
Cota Altitudinal Simpson Shannon-Wiener
2950 0,813 1.769
3175 0,770 1.646
Indices de diversidad según cota altitudinal
2.2.6. Diversidad Beta
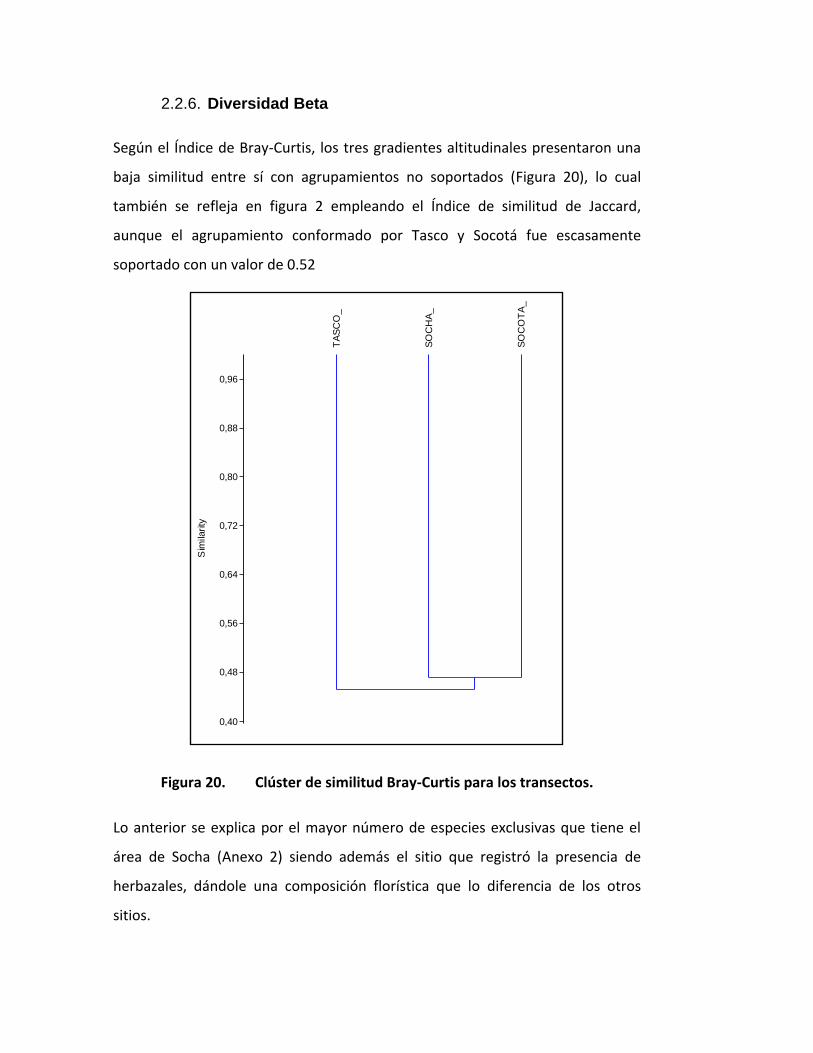
Según el Índice de Bray-Curtis, los tres gradientes altitudinales presentaron una
baja similitud entre sí con agrupamientos no soportados (Figura 20), lo cual
también se refleja en figura 2 empleando el Índice de similitud de Jaccard,
aunque el agrupamiento conformado por Tasco y Socotá fue escasamente
soportado con un valor de 0.52
Figura 20. Clúster de similitud Bray-Curtis para los transectos.
Lo anterior se explica por el mayor número de especies exclusivas que tiene el
área de Socha (Anexo 2) siendo además el sitio que registró la presencia de
herbazales, dándole una composición florística que lo diferencia de los otros
sitios.
0,40
0,48
0,56
0,64
0,72
0,80
0,88
0,96
Sim
ilarity
TA
SC
O_
SO
CH
A_
SO
CO
TA
_
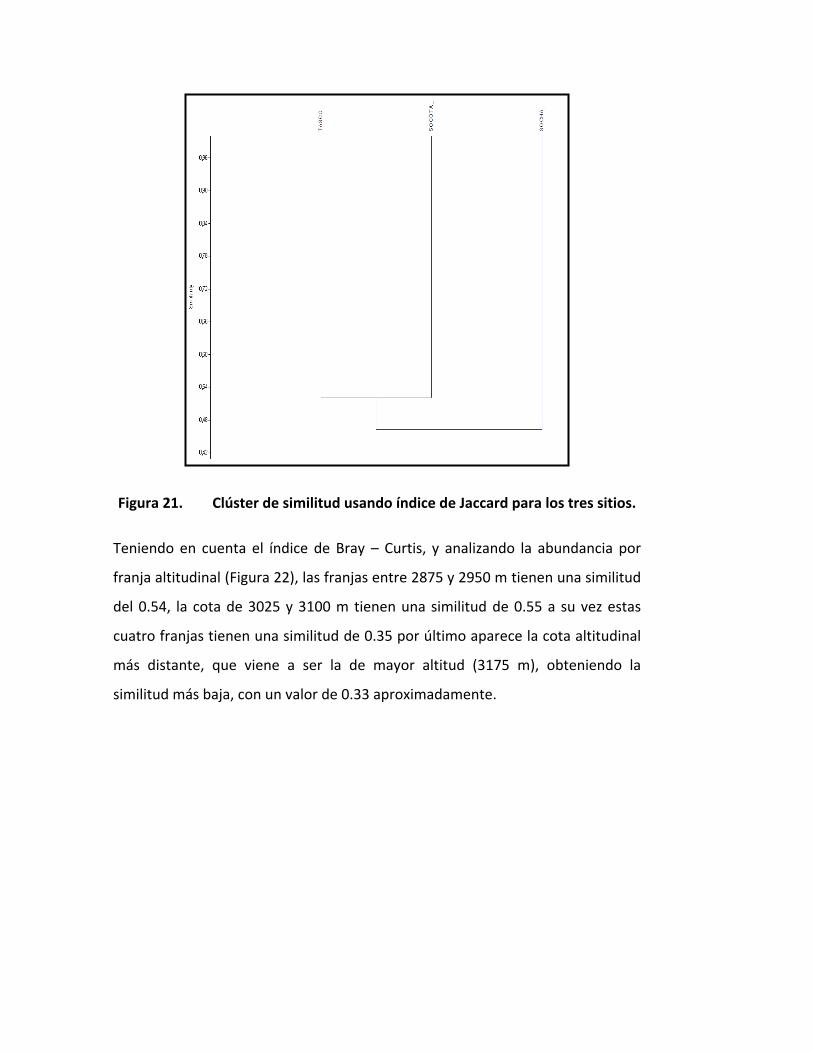
Figura 21. Clúster de similitud usando índice de Jaccard para los tres sitios.
Teniendo en cuenta el índice de Bray – Curtis, y analizando la abundancia por
franja altitudinal (Figura 22), las franjas entre 2875 y 2950 m tienen una similitud
del 0.54, la cota de 3025 y 3100 m tienen una similitud de 0.55 a su vez estas
cuatro franjas tienen una similitud de 0.35 por último aparece la cota altitudinal
más distante, que viene a ser la de mayor altitud (3175 m), obteniendo la
similitud más baja, con un valor de 0.33 aproximadamente.
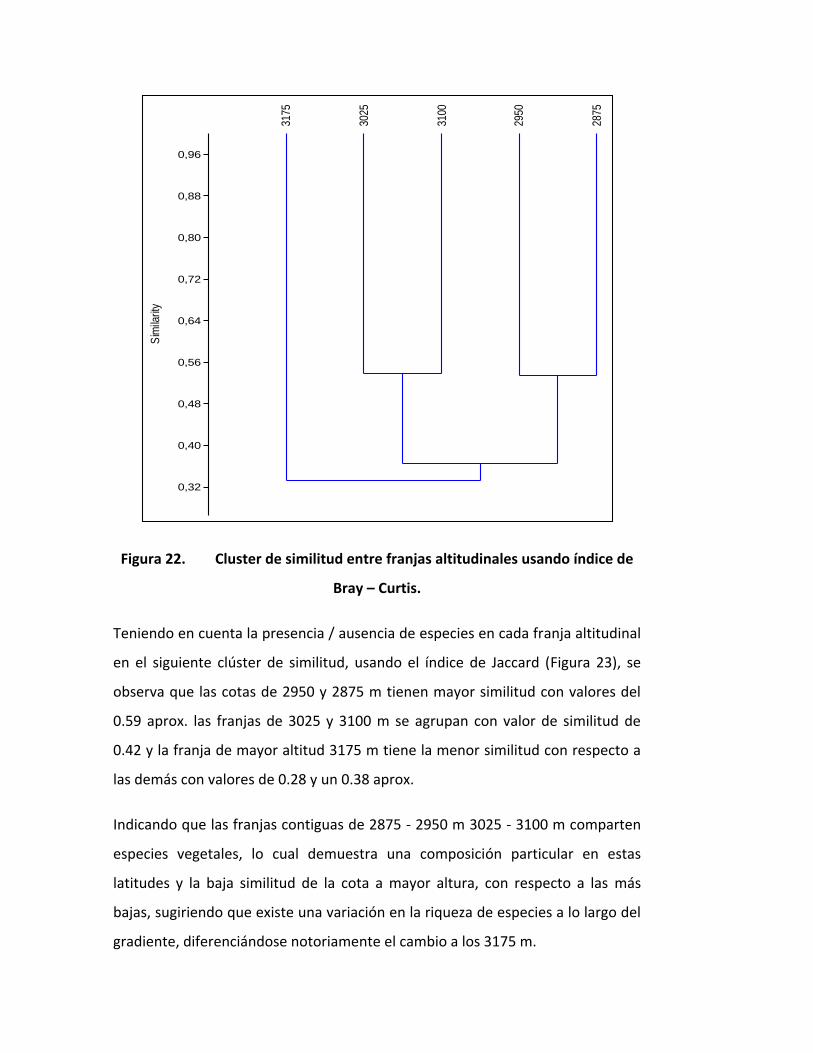
Figura 22. Cluster de similitud entre franjas altitudinales usando índice de
Bray – Curtis.
Teniendo en cuenta la presencia / ausencia de especies en cada franja altitudinal
en el siguiente clúster de similitud, usando el índice de Jaccard (Figura 23), se
observa que las cotas de 2950 y 2875 m tienen mayor similitud con valores del
0.59 aprox. las franjas de 3025 y 3100 m se agrupan con valor de similitud de
0.42 y la franja de mayor altitud 3175 m tiene la menor similitud con respecto a
las demás con valores de 0.28 y un 0.38 aprox.
Indicando que las franjas contiguas de 2875 - 2950 m 3025 - 3100 m comparten
especies vegetales, lo cual demuestra una composición particular en estas
latitudes y la baja similitud de la cota a mayor altura, con respecto a las más
bajas, sugiriendo que existe una variación en la riqueza de especies a lo largo del
gradiente, diferenciándose notoriamente el cambio a los 3175 m.
0,32
0,40
0,48
0,56
0,64
0,72
0,80
0,88
0,96S
imila
rity
3175
3025
3100
2950
2875
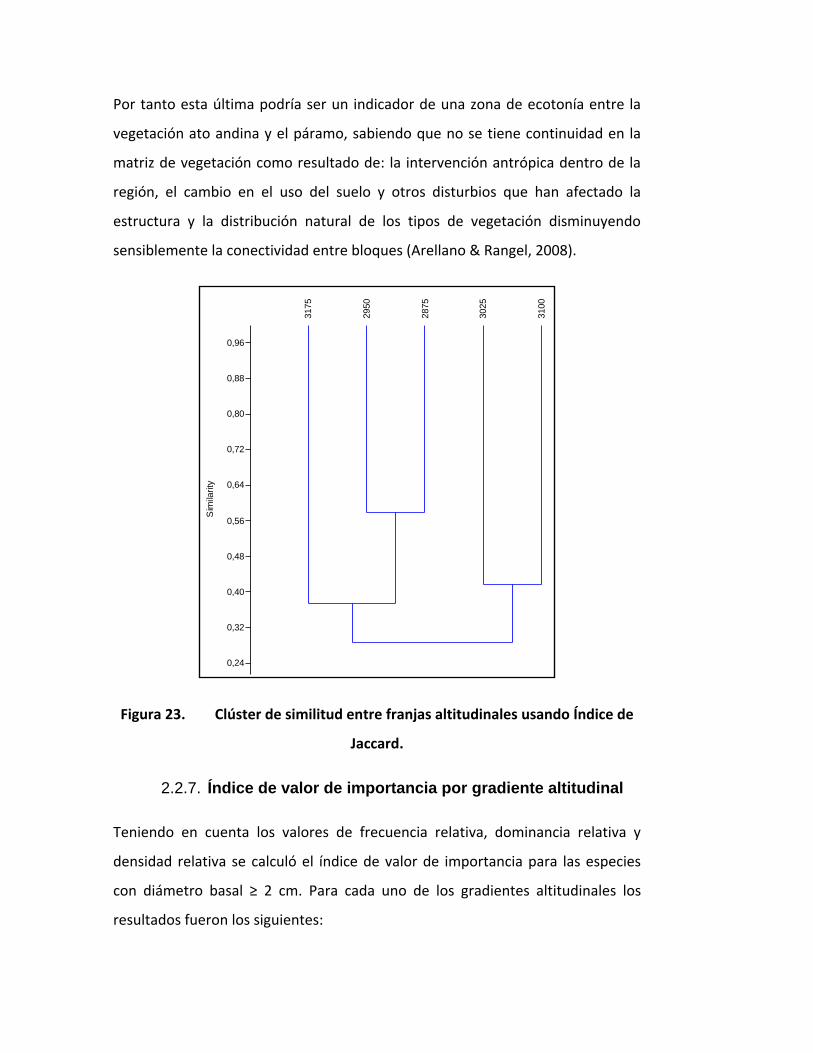
Por tanto esta última podría ser un indicador de una zona de ecotonía entre la
vegetación ato andina y el páramo, sabiendo que no se tiene continuidad en la
matriz de vegetación como resultado de: la intervención antrópica dentro de la
región, el cambio en el uso del suelo y otros disturbios que han afectado la
estructura y la distribución natural de los tipos de vegetación disminuyendo
sensiblemente la conectividad entre bloques (Arellano & Rangel, 2008).
Figura 23. Clúster de similitud entre franjas altitudinales usando Índice de
Jaccard.
2.2.7. Índice de valor de importancia por gradiente altitudinal
Teniendo en cuenta los valores de frecuencia relativa, dominancia relativa y
densidad relativa se calculó el índice de valor de importancia para las especies
con diámetro basal ≥ 2 cm. Para cada uno de los gradientes altitudinales los
resultados fueron los siguientes:
0,24
0,32
0,40
0,48
0,56
0,64
0,72
0,80
0,88
0,96
Sim
ilarity
3175
2950
2875
3025
3100
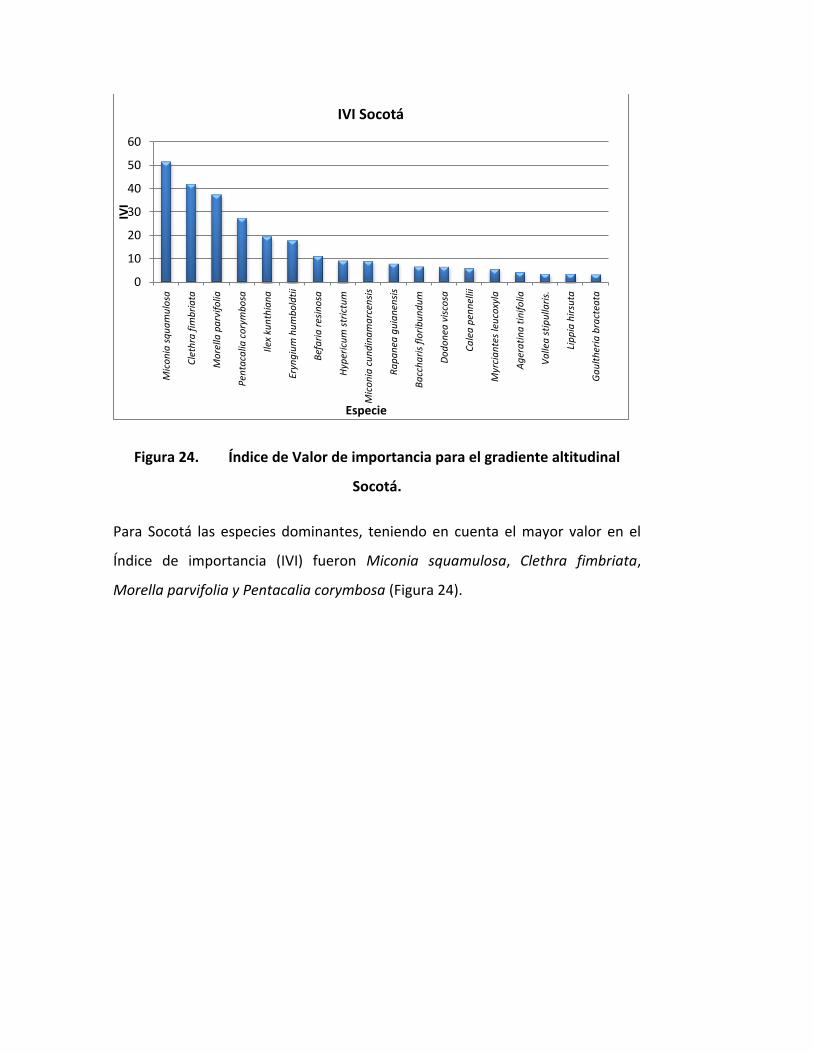
Figura 24. Índice de Valor de importancia para el gradiente altitudinal
Socotá.
Para Socotá las especies dominantes, teniendo en cuenta el mayor valor en el
Índice de importancia (IVI) fueron Miconia squamulosa, Clethra fimbriata,
Morella parvifolia y Pentacalia corymbosa (Figura 24).
0
10
20
30
40
50
60
Mic
on
ia s
qu
am
ulo
sa
Cle
thra
fim
bri
ata
Mo
rella
pa
rvif
olia
Pen
taca
lia c
ory
mb
osa
Ilex
kun
thia
na
Eryn
giu
m h
um
bo
ldti
i
Bef
ari
a r
esin
osa
Hyp
eric
um
str
ictu
m
Mic
on
ia c
un
din
am
arc
ensi
s
Ra
pa
nea
gu
ian
ensi
s
Ba
cch
ari
s fl
ori
bu
nd
um
Do
do
nea
vis
cosa
Ca
lea
pen
nel
lii
Myr
cia
nte
s le
uco
xyla
Ag
era
tin
a t
inif
olia
Va
llea
sti
pu
llari
s.
Lip
pia
hir
suta
Ga
ult
her
ia b
ract
eata
IVI
Especie
IVI Socotá

Figura 25. Índice de valor de importancia para el gradiente altitudinal de
Socha.
En Socha las especies dominantes para el IVI son: Miconia cundinamarcensis,
Symplocos theiformis, Miconia squamulosa y Morella parvifolia (Figura 25).
0
10
20
30
40
50
60Miconia…
Sym
plo
cos
thei
form
is
Mic
on
ia s
qu
am
ulo
sa
Mo
rella
pa
rvif
olia
Bu
cqu
etia
glu
tin
osa
Pen
taca
lia c
ory
mb
osa
Eryn
giu
m h
um
bo
ldti
i
Vib
urn
um
tin
oid
es
Esp
elet
ia a
rgen
tea
Va
llea
sti
pu
llari
s.
Cle
thra
fim
bri
ata
Ra
pa
nea
gu
ian
ensi
s
Ba
cch
ari
s fl
ori
bu
nd
um
Lip
pia
hir
suta
Aln
us
acu
min
ata
Bu
dd
leja
inca
na
Xilo
sma
sp
ecu
lifer
um
Ph
ylla
nth
us
salv
iifo
lius
IVI
Especie
IVI SOCHA
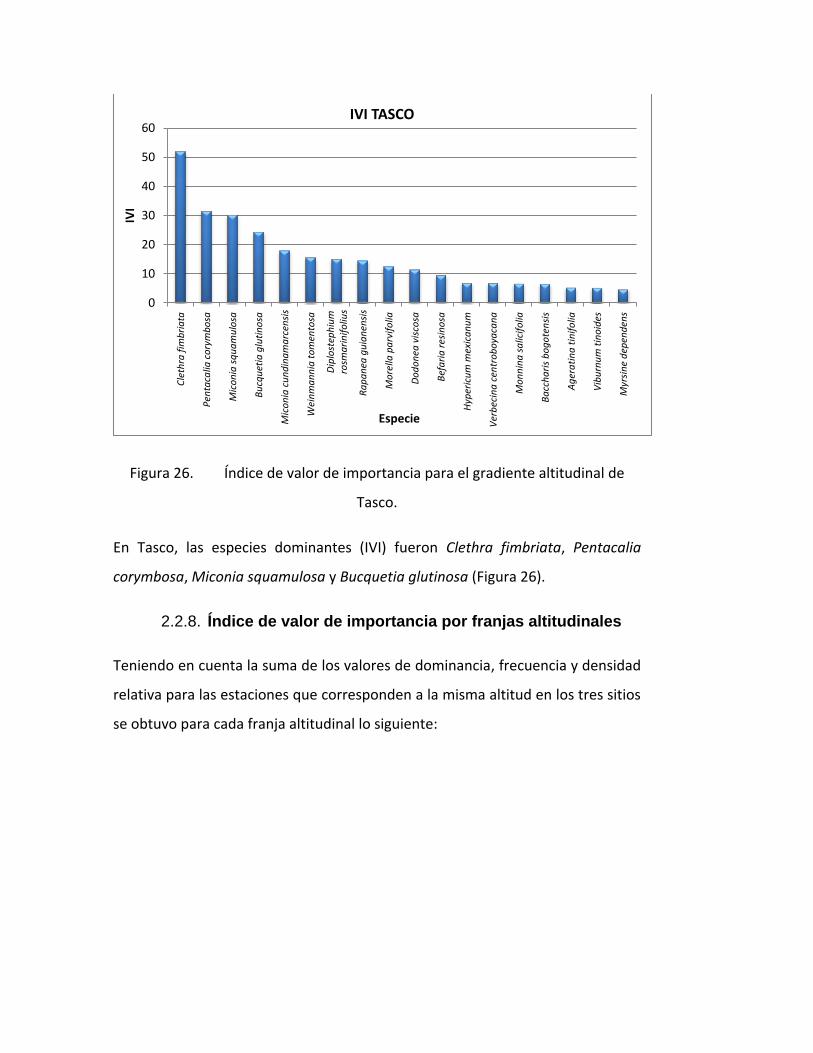
Figura 26. Índice de valor de importancia para el gradiente altitudinal de
Tasco.
En Tasco, las especies dominantes (IVI) fueron Clethra fimbriata, Pentacalia
corymbosa, Miconia squamulosa y Bucquetia glutinosa (Figura 26).
2.2.8. Índice de valor de importancia por franjas altitudinales
Teniendo en cuenta la suma de los valores de dominancia, frecuencia y densidad
relativa para las estaciones que corresponden a la misma altitud en los tres sitios
se obtuvo para cada franja altitudinal lo siguiente:
0
10
20
30
40
50
60
Cle
thra
fim
bri
ata
Pen
taca
lia c
ory
mb
osa
Mic
on
ia s
qu
am
ulo
sa
Bu
cqu
etia
glu
tin
osa
Mic
on
ia c
un
din
am
arc
ensi
s
Wei
nm
an
nia
to
men
tosa
Dip
lost
eph
ium
rosm
ari
nif
oliu
s
Ra
pa
nea
gu
ian
ensi
s
Mo
rella
pa
rvif
olia
Do
do
nea
vis
cosa
Bef
ari
a r
esin
osa
Hyp
eric
um
mex
ica
nu
m
Ver
bec
ina
cen
tro
bo
yaca
na
Mo
nn
ina
sa
licif
olia
Ba
cch
ari
s b
og
ote
nsi
s
Ag
era
tin
a t
inif
olia
Vib
urn
um
tin
oid
es
Myr
sin
e d
epen
den
s
IVI
Especie
IVI TASCO
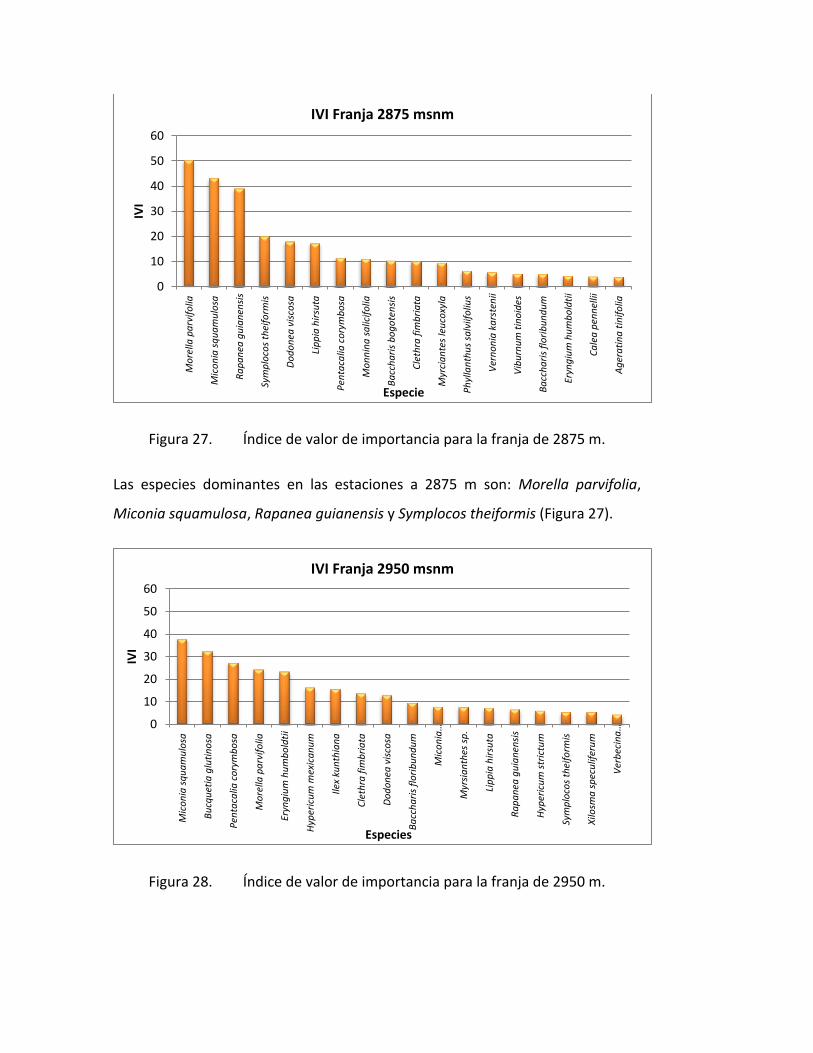
Figura 27. Índice de valor de importancia para la franja de 2875 m.
Las especies dominantes en las estaciones a 2875 m son: Morella parvifolia,
Miconia squamulosa, Rapanea guianensis y Symplocos theiformis (Figura 27).
Figura 28. Índice de valor de importancia para la franja de 2950 m.
0
10
20
30
40
50
60
Mo
rella
pa
rvif
olia
Mic
on
ia s
qu
am
ulo
sa
Ra
pa
nea
gu
ian
ensi
s
Sym
plo
cos
thei
form
is
Do
do
nea
vis
cosa
Lip
pia
hir
suta
Pen
taca
lia c
ory
mb
osa
Mo
nn
ina
sa
licif
olia
Ba
cch
ari
s b
og
ote
nsi
s
Cle
thra
fim
bri
ata
Myr
cia
nte
s le
uco
xyla
Ph
ylla
nth
us
salv
iifo
lius
Ver
no
nia
ka
rste
nii
Vib
urn
um
tin
oid
es
Ba
cch
ari
s fl
ori
bu
nd
um
Eryn
giu
m h
um
bo
ldti
i
Ca
lea
pen
nel
lii
Ag
era
tin
a t
inif
olia
IVI
Especie
IVI Franja 2875 msnm
0
10
20
30
40
50
60
Mic
on
ia s
qu
am
ulo
sa
Bu
cqu
etia
glu
tin
osa
Pen
taca
lia c
ory
mb
osa
Mo
rella
pa
rvif
olia
Eryn
giu
m h
um
bo
ldti
i
Hyp
eric
um
mex
ica
nu
m
Ilex
kun
thia
na
Cle
thra
fim
bri
ata
Do
do
nea
vis
cosa
Ba
cch
ari
s fl
ori
bu
nd
um
Miconia…
Myr
sia
nth
es s
p.
Lip
pia
hir
suta
Ra
pa
nea
gu
ian
ensi
s
Hyp
eric
um
str
ictu
m
Sym
plo
cos
thei
form
is
Xilo
sma
sp
ecu
lifer
um
Verbecina…
IVI
Especies
IVI Franja 2950 msnm

Para la franja de 2950 m las especies con mayor índice de importancia son
Miconia squamulosa, Bucquetia glutinosa, Pentacalia corymbosa y Morella
parvifolia (Figura 28).
Figura 29. Índice de Valor de importancia para la franja de 3025 m
En las estaciones de los 3025 m las especies dominantes son: Clethra fimbriata,
Miconia squamulosa, Symplocos theiformis e Ilex kunthiana (Figura 29).
0
10
20
30
40
50
60
70
80
90
Cle
thra
fim
bri
ata
Mic
on
ia s
qu
am
ulo
sa
Sym
plo
cos
thei
form
is
Ilex
kun
thia
na
Pen
taca
lia c
ory
mb
osa
Bef
ari
a r
esin
osa
Diplostep
hium…
Vib
urn
um
tin
oid
es
Ba
cch
ari
s fl
ori
bu
nd
um
Mo
rella
pa
rvif
olia
Miconia…
Myr
sin
e d
epen
den
s
Verbecina…
Ba
cch
ari
s b
og
ote
nsi
s
Mo
nn
ina
sa
licif
olia
Psy
cho
tria
ta
tam
an
a
IVI
Especie
IVI Franja 3025 msnm

Figura 30. Índice de valor de importancia en la franja 3100 m.
Clethra fimbriata, Miconia cundinamarcensis, Miconia squamulosa y Morella
parvifolia son las especies que se presentan como dominantes en la franja de
3100 m. (Figura 30)
0
10
20
30
40
50
60
Cle
thra
fim
bri
ata
Miconia…
Mic
on
ia s
qu
am
ulo
sa
Mo
rella
pa
rvif
olia
Pen
taca
lia c
ory
mb
osa
Va
llea
sti
pu
llari
s.
Aln
us
acu
min
ata
Ra
pa
nea
gu
ian
ensi
s
Vib
urn
um
tin
oid
es
Bu
dd
leja
inca
na
Bef
ari
a r
esin
osa
Bu
cqu
etia
glu
tin
osa
Ag
era
tin
a t
inif
olia
Ilex
kun
thia
na
Bu
dd
leja
sp
.
Psy
cho
tria
ta
tam
an
a
Weinmannia…
Lip
pia
hir
suta
IVI
Especie
IVI Franja 3100 msnm
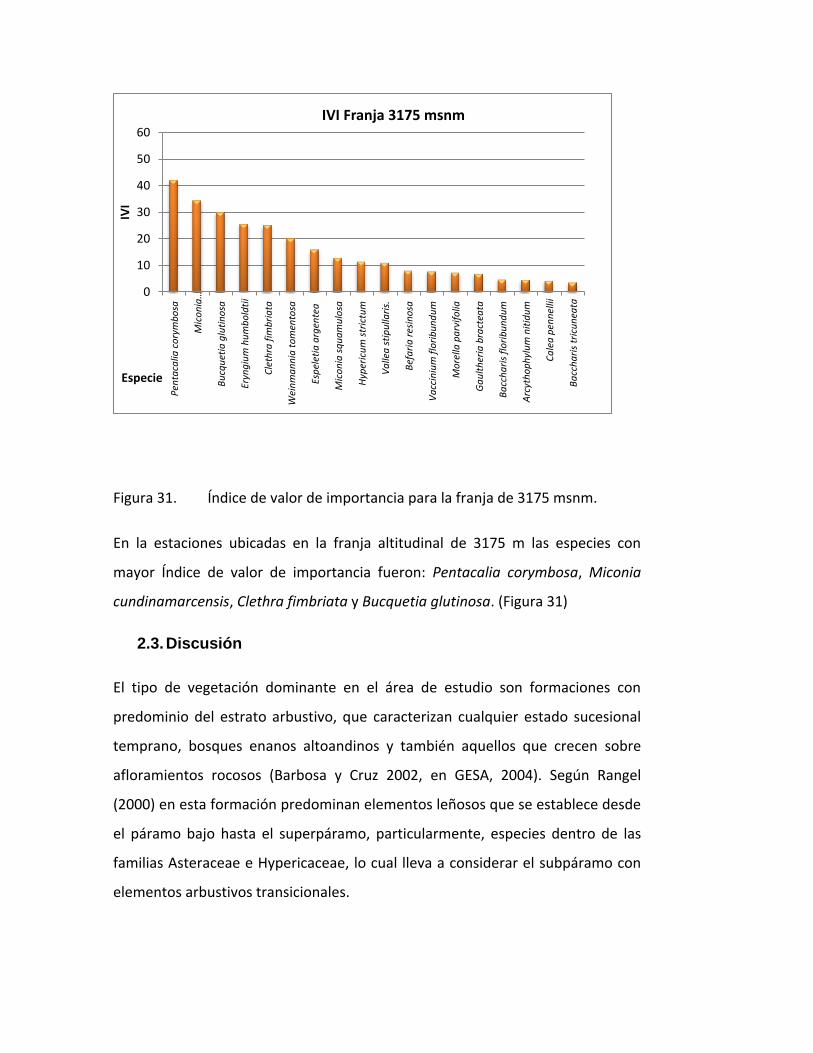
Figura 31. Índice de valor de importancia para la franja de 3175 msnm.
En la estaciones ubicadas en la franja altitudinal de 3175 m las especies con
mayor Índice de valor de importancia fueron: Pentacalia corymbosa, Miconia
cundinamarcensis, Clethra fimbriata y Bucquetia glutinosa. (Figura 31)
2.3. Discusión
El tipo de vegetación dominante en el área de estudio son formaciones con
predominio del estrato arbustivo, que caracterizan cualquier estado sucesional
temprano, bosques enanos altoandinos y también aquellos que crecen sobre
afloramientos rocosos (Barbosa y Cruz 2002, en GESA, 2004). Según Rangel
(2000) en esta formación predominan elementos leñosos que se establece desde
el páramo bajo hasta el superpáramo, particularmente, especies dentro de las
familias Asteraceae e Hypericaceae, lo cual lleva a considerar el subpáramo con
elementos arbustivos transicionales.
0
10
20
30
40
50
60
Pen
taca
lia c
ory
mb
osa
Miconia…
Bu
cqu
etia
glu
tin
osa
Eryn
giu
m h
um
bo
ldti
i
Cle
thra
fim
bri
ata
Wei
nm
an
nia
to
men
tosa
Esp
elet
ia a
rgen
tea
Mic
on
ia s
qu
am
ulo
sa
Hyp
eric
um
str
ictu
m
Va
llea
sti
pu
llari
s.
Bef
ari
a r
esin
osa
Va
ccin
ium
flo
rib
un
du
m
Mo
rella
pa
rvif
olia
Ga
ult
her
ia b
ract
eata
Ba
cch
ari
s fl
ori
bu
nd
um
Arc
yth
op
hyl
um
nit
idu
m
Ca
lea
pen
nel
lii
Ba
cch
ari
s tr
icu
nea
ta
IVI
Especie
IVI Franja 3175 msnm

De acuerdo a los resultados obtenidos no se distingue claramente una zona de
transición entre la vegetación característica de bosque alto andino y la del
subpáramo, lo cual, puede explicarse en gran parte a la discontinuidad en las
unidades de vegetación analizadas, como resultado de distintas actividades
antrópicas, aunque un efecto del recambio de especies a lo largo de los
gradientes altitudinales, procesos invasivos por especies de plantas (e. g.
paramización) e incluso una zona de transición amplia (franja de ecotono),
estado sucecional puede dar lugar a las observaciones hechas. Otra explicación a
este fenómeno puede entenderse como una zona de transición en altitudes
intermedias. Esto implicaría una conjunción de especies tanto de alturas bajas y
altas disminuyendo la riqueza de especies y por tanto su diversidad.
La diversidad encontrada puede calificarse como baja y dominante teniendo en
cuenta los índices calculados. Por una parte el índice de Simpson, entendido
como la probabilidad de encontrar al azar dos individuos de la misma especie,
tiene valores altos (>0.7) es tanto por altitud como por localidad alto. Lo cual
implica una moderada-alta dominancia de las pocas especies que se presentan.
Adicionalmente el índice de Shannon revela valores cercanos a 3 (2.8) lo cual
está entre los valores marginales de diversidad, teniendo en cuenta que
localidades con alta diversidad registran índices por encima de 3, como lo son las
selvas tropicales con cifras superiores a 4.5 (Villarreal et al., 2006, Zarco-Espinosa
et al., 2010).
Si bien, la diversidad de especies tiende a disminuir a medida que se incrementa
la altitud, en este momento no se pude identificar algún patrón, debido en gran
parte al rango altitudinal estrecho considerado en este estudio, esto significa que
picos altos de diversidad de especies para algunas cotas altitudinales, pueden
explicarse por otros factores que involucran por ejemplo, tipo de suelo o
sucesión. Sin embargo, se sugiere realizar aproximaciones multivariadas para

evaluar qué factores explican la diversidad de especies observada en cada
gradiente altitudinal.
Los tres sitios comparten las especies con mayor valor de importancia, es decir,
las que son dominantes; esto indica que son especies con una distribución
amplia para el área de estudio. Se puede observar que Socha tiene valores de IVI
con una distribución más homogénea entre las especies, esto sugiere que el sitio
se caracteriza por formaciones vegetales que no tienen una dominancia marcada
de especies lo cual puede relacionarse con su significativa abundancia, riqueza y
diversidad sobre los otros sitios. A diferencia de lo anterior, Socotá y Tasco
muestran una discrepancia de valores más amplia entre las especies dominantes
y las de menores IVI, interpretándose como los sitios donde las formaciones
vegetales pueden caracterizarse por la dominancia marcada de 2 o 3 especies,
siendo esto más evidente en el gradiente de Tasco.
De acuerdo a Cuatrecasas (1958, 1968) para el área de estudio, la zona de
transición corresponde a subpáramo, con un cambio de la vegetación de bosque
montano hacia gramíneas, un mosaico de arbustos y pequeños arboles dispersos
que gradualmente reducen su tamaño, dando origen a un matorral y una
vegetación baja de arbustos bajos. La delimitación de zona de páramo, resulta
complejo dada la influencia local de la topografía, variables climáticas sobre la
vegetación junto al efecto de la intervención humana (Laegaard, 1992).

3. ANFIBIOS
3.1. INTRODUCCIÓN
En Colombia se estima que la abundancia de anfibios es de 785 especies (Frost,
2013), representando el segundo país con el mayor número de especies en este
grupo (Rueda-A et al., 2004, Young et al. 2004), lo que ubica al País en el tercer
lugar de abundancia en este grupo a nivel mundial (Mittermeier et al., 1997). El
área de influencia del proyecto Socotá, Socha y Tasco en el PNN Pisba, la fauna
se encuentra asociada a la jurisdicción de la Unidad biogeográfica de la Cordillera
Oriental, en la cuenca seca.
Colombia alberga más de 754 especies de anfibios de las cuales alrededor de 359
(47.6%) son endémicas, 25 (3.3%) están listadas en CITES por presiones de tráfico
ilegal, y 227 (30.1%) se encuentran en alguna categoría de amenaza a nivel global
(IUCN 2010) y 49 (6.5%) a nivel nacional (Rueda-Almonacid et al., 2004).
La gran mayoría de especies amenazadas en Colombia se distribuyen en la región
Andina, y probablemente por la contribución de un sinnúmero de factores

intrínsecos y extrínsecos, que actúan de manera sinérgica (Cooper et al., 2008;
Sodhi et al., 2008; Laurence y Useche 2009).
De acuerdo con Hoffmann y colaboradores (2010) el 30% de las especies de
anfibios se encuentran en alguna categoría de amenaza (En peligro crítico [CR],
En peligro [EN] y Vulnerable [VU]) y el 25% carecen de información de tamaños
poblacionales, extensión de la distribución y amenazas para ser categorizadas
(Datos deficientes [DD]).
Actualmente la existencia de los anfibios se encuentra amenazada por factores
directos (e.g. enfermedades emergentes, plaguicidas, tráfico ilegal, pérdida de
hábitat), indirectos (e.g. cambio climático, especies invasoras, efectos de borde,
degradación del hábitat) y efectos sinérgicos que reducen la viabilidad de las
poblaciones e incrementan su vulnerabilidad a la extinción (Crump 2003).
A pesar de que la principal causa de disminución histórica de poblaciones y
especies amenazadas a nivel mundial es la pérdida y fragmentación de hábitat,
recientemente enfermedades emergentes como la quitridiomicosis en posible
sinergia con el cambio climático y la expansión del rango geográfico de especies
invasoras, han complicado mucho más el panorama de conservación de los
anfibios (Gardner et al. 2007, Urbina-Cardona 2008). III Congreso Colombiano de
Zoología
3.2. METODOLOGÍA
3.2.1. Muestreos
Los anfibios en campo se estudiaron por medio de prospecciones al azar usando
la técnica de relevamiento por encuentros visuales descrita en Crump y Scott
(2001), desde las 0600 h a las 1000 h y nocturnas desde las 1800 h a las 2030 h. y
en cada sitio (Socha, Socota, Tasco), durante 5 días consecutivos, explorando
todos los micro hábitats disponibles dentro de los tipos de hábitats presentes en

la zona, como bajo y sobre rocas, troncos caídos, colchones de hepáticas,
macollas, bajo-entre y sobre necromasa de frailejones y puya, troncos de árboles
y arbustos, sobre el suelo cubierto por hojarasca o desnudo, entre vegetación
rasante, esto sin incluir fuentes de agua, ya que según lo descrito en la
metodología para el componente vegetación, no era apropiado muestrear estos
sitios debido a la variabilidad de la vegetación.
Teniendo en cuenta que los sitios de muestreo debían tener ciertas
características para la toma adecuada de datos, como conservación y tipo de
alteración antrópica se consideró, en caso tal, ubicarlos en un sitio cercano al
muestreo de vegetación, pero con las mismas condiciones de conservación.
Se relevaron tres zonas de muestreo aleatorizado (Socotá, Socha y Tasco) a lo
largo de un transecto altitudinal desde los 2875 m hasta los 3175 m,
obteniéndose 9 zonas de muestreo para los tres sitios
Los ejemplares fueron capturados de forma manual, y depositados
individualmente en bolsas de tela humedecida. Se tomaron datos ecológicos
como ubicación del individuo, hora de la actividad, comportamiento y registro
fotográfico para la debida identificación.
3.2.2. Análisis de datos.
Según los parámetros e índices establecidos, se realizó un análisis de los datos de
anfibios colectados en campo para establecer si dicha información es
representativa en relación a los parámetros de riqueza y abundancia. Se
analizaron los datos de las especies identificadas por los métodos de captura y
avistamiento, y demás estimadores propuestos por el Instituto Alexander von
Humboldt (acumulación y distribución).

3.3. RESULTADOS Y DISCUSION
3.3.1. Composición
Basados en los registros obtenidos en campo, para el área, se encontró un total
de 2 especies de anfibios, de la familia: Hylidae y Craugastoridae.
Este resultado en parte se debe a la alta presión que ejerce la ganadería, la
agricultura y la minería, en el cambio de uso del suelo, lo cual conlleva que las
condiciones apropiadas para la permanencia de anfibios en este ambiente sean
totalmente diferentes a las de los hábitats naturales
3.3.2. Esfuerzo de muestreo
Cada uno de los sitios se visitó durante 5 días consecutivos. Se muestreo de
06:00 a 09:00 y de 18:00 a 20:30 horas, con un esfuerzo de captura de 94.5 horas
/ hombre
3.3.3. Clasificación Taxonómica e información ecológica de las Especies
de Anuros
3.3.3.1. Dendropsophus labialis (Peters, 1863)
Rana sabanera o rana andina, esta especie se conoce desde los andes centrales
hasta el noreste de Colombia, en los departamentos de Boyacá, Cundinamarca,
Santander y Norte de Santander. Se registra desde los 1600 – 3600 m.
Catalogado como Preocupación Menor a la vista de su amplia distribución y a la
tolerancia de una amplia gama de hábitats, presenta una gran población, y es
poco probable que se disminuya con la rapidez suficiente como para calificar
para su inclusión en una categoría más amenazada.

Dendropsophus labialis
No hay amenazas conocidas para esta especie. Se utiliza para los experimentos
en las universidades con fines de enseñanza, pero esto no se considera una
amenaza. (IUCN. 2004)
3.3.3.2. Pristimantis elegans (Anura: Leptodactylidae) (Peters,
1863)
Craugastoridae es un clado netamente Americano desde el sur de los Estados
Unidos hasta el norte de Argentina. La especie es endémica de la Cordillera
Oriental (Colombia), entre 2600-3300 m de altitud, en zonas de páramo y en el
bosque nuboso. Catalogado como Vulnerable (B1ab(iii)) debido a su extensión de
presencia es menor de 20.000 km2, su distribución está muy fragmentada, y no
hay tendencia a la disminución en el alcance y la calidad de su hábitat forestal en
la Cordillera Oriental de los Andes colombianos. La especie se puede encontrar

en la vertiente oriental de la Cordillera Oriental de los páramos de Chingaza,
Sumapaz y La Rusia, en los departamentos de Cundinamarca y Boyacá, Colombia,
a partir de 2600-3300 m. Las principales amenazas a la población son la pérdida
de hábitat, y la degradación del hábitat, debido a la agricultura y la ganadería. Se
reportó la presencia de los dendrobatidis quítrido Batrachochytrium hongo
(Chytridiomycota: Chytridiales) en Pristimantis elegans (Ruiz y Rueda -Almonacid,
2008). Esta especie se encuentra en zonas de páramo, y también en el bosque
nuboso. Se asocia con vegetación herbácea, y muy pequeños arbustos. El 40 %
de los anuros conocidos en Colombia son del género Eleutherodactylus cuyas
especies no poseen etapa larval (Lynch 1998) aunque necesitan cierto grado de
humedad, la presencia de agua no es condición para reproducirse.
Eleutherodactylus, ha desarrollado gran capacidad de habitar lejos del ambiente
acuático y colonizar nuevos hábitat, es el grupo de mayor diversidad y
abundancia en el neotrópico (Duellman, 1992). En realidad es el género de
mayor diversidad de todos los vertebrados conocidos
Pristimantis elegans (Antiguamente Eleutherodactylus, con un diseño
similar a P.affinis)

3.4. Especies Endémicas
Según los reportes de distribución de la UICN (2013), el Plan de Manejo del
Parque Nacional Natural Pisba y los POT’s de las localidades Socotá, Socha y
Tasco, solo se destacan algunas especies pero sólo por su abundancia como
Dendrosophus labilis, Atelopus sp., Colostethus sp., pero esto no significa que
sean endémicas
Relación de las especies con las coberturas identificadas en los sitios de
muestreo Socotá, Socha y Tasco.
Según los resultados obtenidos las coberturas en la que se obtuvo presencia de
especies de Anfibios fue en las de bosque alto andino con presencia de
herbazales (altura 2875 m, Socotá) y “páramo” en zonas con hojarasca (altura de
3100 m, Tasco), lo que nos permite corroborar la capacidad de adaptación de los
anfibios, teniendo en cuenta que las especies encontradas Dendropsophus
labialis y Pristimantis elegans, son especies abundantes y muy comunes en
distintos tipos de hábitat.
Por tal razón, al excluir las áreas paramizadas por intervención antrópica, se
puede concluir que el grupo de anfibios propios de páramo es más pequeño que
lo reconocido en la actualidad. Igualmente se debe tener en cuenta que el filtro
ecológico considerado en este caso la altitud, para caracterizar la fauna
paramuna no es el suficiente, ya que se deben considerar otros aspectos
característicos del hábitat.
Teniendo en cuenta que la diversidad se compone de dos elementos básicos: el
número de especies (riqueza) y la abundancia proporcional de especies
(equitatividad), los datos obtenidos en este trabajo no permiten dilucidar de
estos índices para poder conocer y valorar las comunidades de anfibios en las
diferentes unidades o niveles; por lo tanto podemos decir que es importante

estudiar más a fondo este grupo, para los sitios del complejo de páramo de
Pisba.
Diversos trabajos han encontrado que, debido a sus características fisiológicas y
etológicas, la riqueza, abundancia y distribución de anuros son influenciados
significativamente por la heterogeneidad espacial, respondiendo en mayor grado
a la estructura del hábitat que a la presencia o ausencia de especies vegetales
particulares; de igual forma para algunas especies simpátricas de una comunidad
es importante cierto tipo de fisonomía vegetal y éste a su vez está
correlacionado con el cuerpo y el tamaño del organismo, incluso el espacio del
hábitat puede ser usado por otras especies con diferentes horas de actividad y
estas a su vez habitan condicionadas por los efectos de la actividad humana
(Meza-Ramos et al., 2008; Ramírez, 2008; Cortez-Fernandez, 2006; García et al.,
2005; Venegas, 2005; Suárez-Badillo & Ramírez-Pinilla, 2004; Herrera et al 2004;
Arroyo et al 2003; Contreras et al 2001; Guerrero et al 1999; Osorno-Muñoz,
1999; Lynch, 1999; Lynch, 1997 & Lynch, 1979b. citado en Ramírez et al., 2009).
Los gradientes altitudinales también abarcan cambios en la calidad, la diversidad
y densidad de depredadores y presas, con el último mencionado se sabe que
afectan a las tasas de crecimiento y rasgos de historia de vida relacionados con
los anfibios (Bernardo y Agosta, 2003 citado Navas 2003).
Todas estas variables físicas y bióticas pueden cambiar de forma simultánea, por
lo que los efectos sinérgicos podrían aumentar aún más el reto ecofisiológica
para ectotermos vertebrados que viven en altitudes elevadas. (Navas 2003)

4. ARTROPODOS
4.1. INTRODUCCIÓN
Estudios sobre la riqueza de especies a lo largo de gradientes altitudinales para la
región neotropical han abordado diferentes taxones dentro de los artrópodos,
pudiendo citar: insectos en general (Janzen et al. 1976; Wolda 1987), polillas
(Brehm y Fiedler 2004), mariposas (Pyrcz et al. 2009), escarabajos (Escobar et al.
2007, Jones et al. 2012), hormigas (Wilkie et al. 2010), avispas formadoras de
agallas (Lara et al. 2002), Opiliones (Almeida-Neto et al. 2006), pero siendo los
trabajos de Olson (1994) y un estudio realizado por Van der Hammen (1984)
citado por el mismos autor, donde se hace una exploración acerca de la riqueza,
abundancia y biomasa de organismos de suelo.
Este trabajo busca examinar, de manera preliminar, diversidad Alfa y diversidad
Beta para las comunidades artrópodos a lo largo de tres gradientes altitudinales
en los municipios de Socha, Tasco y Socotá como parte del complejo de paramos
de Pisba y con la información obtenida evaluar la existencia de una zonificación
altitudinal, empleando artrópodos de suelo.

4.2. MÉTODOS
4.2.1. Muestreo
Se definieron tres gradientes altitudinales de acuerdo a cobertura de vegetación
o uso de suelo y topografía correspondientes a los municipios de Tasco entre
2900 a 3195m, Socha entre 2800 a 3200m y Socotá entre 2850 a 3230m. A lo
largo de cada gradiente se ubicaron cinco transectos cada 100m y
perpendiculares a la pendiente. Dentro de cada transecto se instalaron cinco
trampas de caída distanciadas entre si cada 5 metros (Olson, 1994), para
completar transectos de 20 m lineales; si bien, puede existir un efecto de la
distancia entre trampas, cuando se realizan muestreos intensivos para grupos
taxonómicos como escarabajos coprófagos o carábidos, la distancia aquí
empleada no presenta diferencias en la riqueza y/o abundancias cuando se
compara con distancias de 10 o 20m entre trampas (Ward et al., 2001), además
se buscó reducir el efecto de la estructura de la vegetación, dada la
discontinuidad de la misma en los sitios de estudio y que puede tener una mayor
afectación potencial sobre las trampas (Melbourne, 1999).
Para todas las trampas se empleó una solución preservante con etanol al 10% y
solución jabonosa. Como complemento al método anterior se empleó un
método de cuadrados, paralelo a cada transecto dentro de cada gradiente. Este
método consistió en definir una parcela de 5m2 y luego delimitar subparcelas de
1m2, posteriormente se seleccionaron 4 subparcelas de manera aleatoria, por
medio de cuadrado latino y se realizó en cada una colecta manual de los
artrópodos presentes, por medio de la remoción de materia orgánica, durante 30
minutos.
El material coleccionado fue preservado en alcohol al 70% y etiquetado con sus
respectivos datos de colecta. Se realizó la identificación taxonómica hasta el
máximo nivel posible, no obstante, debido a la complejidad en la taxonomía de

los organismos del suelo como Collembola, Acari, Araneae y Opiliones, se
definieron morfotipos.
4.3. ANÁLISIS DE DATOS
4.3.1. Diversidad Alfa
La diversidad de especies, fue evaluada para cada cota altitudinal dentro de cada
gradiente altitudinal Socha, Socotá y Tasco y luego para el total de los datos por
franja altitudinal, esto debido a las diferencias existentes en cotas altitudinales
entre gradientes, así: Alt 1 (2800-2900 m), Alt 2 (2900-3000 m), Alt 3 (3000-
3100), Alt 4 (3000-3150 m) y Alt 5 (3195-3230 m). Para cada gradiente y el total
de datos los muestreos por medio de trampas de caídas y método de cuadrados
fueron unificados. Se emplearon los siguientes índices:
1. Índice de Berger & Parker (dominancia).El cual es adecuado para describir
comunidades donde existen pocas especies dominantes (Caruso et al. 2007), en
este caso, por la mayor proporción de la riqueza observada en los órdenes Acari
y Collembola.
2. Índices de Simpson (dominancia) e Índice de Shannon-Wiener (equidad). Estos
índices, si bien, son muy empleados, necesitan de varios requerimientos. En el
caso de Shannon, todas las especies de la comunidad necesitan ser
coleccionadas, por lo que es sensible al tamaño de las muestras y sus resultado
son difíciles de interpretar, mientras la estimación hecha por el Índice de
Simpson depende del tipo de distribución de las abundancias, aunque no es
necesario describir esta distribución (Magurran 2004). En este estudio el índice
de Shannon-Wiener es empleado con propósitos comparativos.
Perfiles de diversidad Renyi
Los perfiles de diversidad Renyi son un método que busca ordenar las
comunidades desde una baja a una alta diversidad (Tóthmérész 1995). Los

valores Hα son calculados desde la frecuencia de cada especie dentro de la
comunidad y desde un parámetro alfa con un rango entre 0 e infinito (Kindt et
al., 2005); a una escala de 0, 1, 2 e infinito; está relacionado respectivamente al
Índice de diversidad de Shannon (riqueza), Índice de diversidad de Simpson e
Índice de diversidad de Berger-Parker, basados sobre dominancia y que a partir
de los cuales es posible extraer perfiles de equitatividad (Legendre y Legendre
1998), por ultimo este método es adecuado cuando se comparan comunidades
con distintos tamaños de muestra (Tóthmérész 1995).
4.3.2. Diversidad Beta
Gran parte de los estudios que involucran la descripción de la diversidad beta
emplean índices de similitud basados sobre la incidencia de especies
(presencia/ausencia), como son los Índices de Jaccard y Sørensen
(Magurran2004), pero los cuales al no considerar las abundancias, no considera
el efecto de las especies raras o dominantes dentro de una comunidad (Chao et
al. 2005, con excepción del Índice de Bray-Curtis, pero el cual puede ser afectado
por muestreos desiguales (Chao et al. 2005).
Por lo anterior, se emplearan dos índices de similitud basados sobre las
abundancias de especies y que considera el efecto de las especies compartidas
no observadas, el índice de similitud “Chao-Jaccard-Raw Abundance-based” el
cual estima las especies compartidas en una de dos muestras, pero no en ambas
y el índice de similitud de “Chao-Jaccard-Est Abundance-based” que estima las
especies compartidas no observadas en ambas muestras (Chao et al., 2005).
Además se empleó el índice Chao-Sørensen (especies no observadas no
corregido) y Chao-Sørensen (especies no observadas corregido), que estiman la
probabilidad que dos individuos seleccionados aleatoriamente uno de cada sitio,
pertenezcan a especies compartidas en ambos sitios (Chao et al. 2005).

Los índices de diversidad, perfiles de diversidad y equitatividad Renyi
(permutaciones=1000) e Índice de Bray-Curtis fueron calculados por medio del
paquete BiodiversityR, el cual emplea los siguientes paquetes: vegan, Rcmdr,
Permute, MASS, knitr bajo la plataforma R (R Team Core 2013). Los estimadores
de riqueza compartida fueron calculados por medio del programa EstimateS
versión 9.0.1. (Colwell, 2013).
4.4. RESULTADOS:
4.4.1. Diversidad Alfa
Para Hexapoda, los grupos que presentaron la mayor riqueza de especies en
orden descendente fueron en Socha Collembola, Hemiptera, Diptera, Coleoptera
y Blattaria (Dyctioptera); en Socotá Hemiptera, Collembola, Coleoptera, Diptera e
Hymenoptera y en Tasco Collembola, Diptera, Hemiptera, Hymenoptera y
Coleoptera (Figura 32)
Figura 32. Comparación entre gradientes altitudinales en cuanto al número
de morfotipos dentro de cada grupo taxonómico en Hexapoda.
0
20
40
60
80
100
120
Socha Socota Tasco
Collembola Hemiptera Diptera Coleoptera Blataria
Orthoptera Hymenoptera Formicidae Psocoptera Thysanoptera
Diplura Archaeognata Lepidoptera Mantodea Phasmida
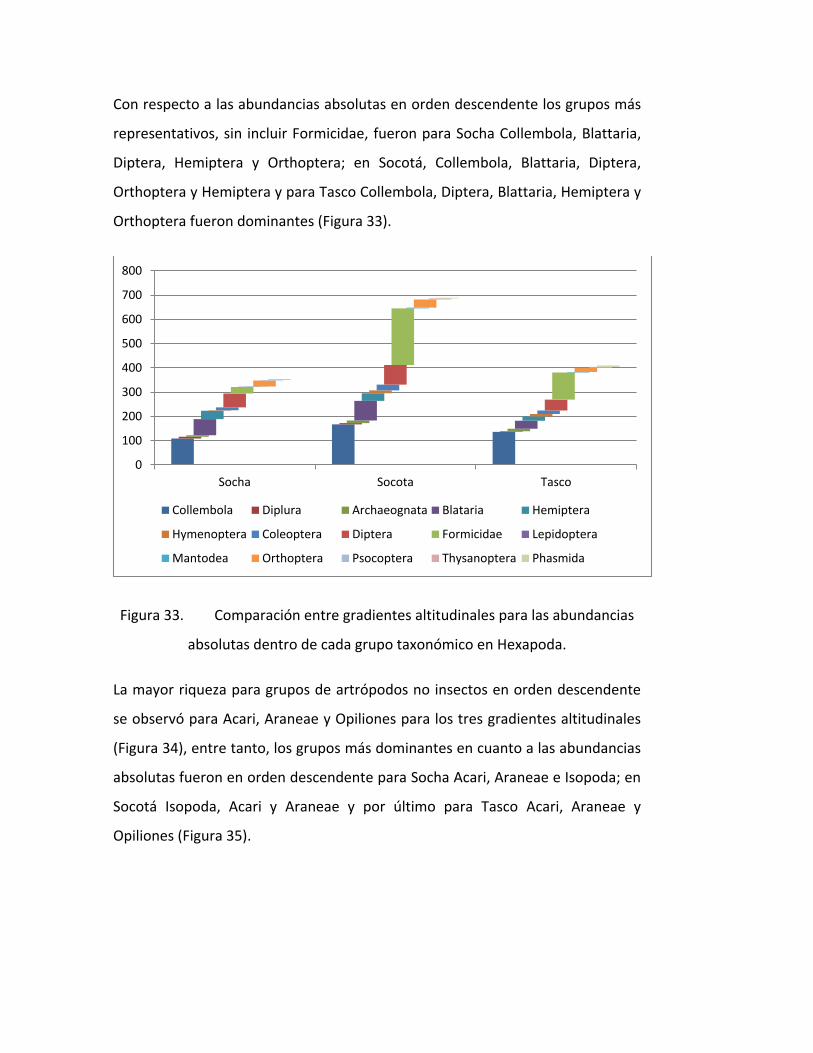
Con respecto a las abundancias absolutas en orden descendente los grupos más
representativos, sin incluir Formicidae, fueron para Socha Collembola, Blattaria,
Diptera, Hemiptera y Orthoptera; en Socotá, Collembola, Blattaria, Diptera,
Orthoptera y Hemiptera y para Tasco Collembola, Diptera, Blattaria, Hemiptera y
Orthoptera fueron dominantes (Figura 33).
Figura 33. Comparación entre gradientes altitudinales para las abundancias
absolutas dentro de cada grupo taxonómico en Hexapoda.
La mayor riqueza para grupos de artrópodos no insectos en orden descendente
se observó para Acari, Araneae y Opiliones para los tres gradientes altitudinales
(Figura 34), entre tanto, los grupos más dominantes en cuanto a las abundancias
absolutas fueron en orden descendente para Socha Acari, Araneae e Isopoda; en
Socotá Isopoda, Acari y Araneae y por último para Tasco Acari, Araneae y
Opiliones (Figura 35).
0
100
200
300
400
500
600
700
800
Socha Socota Tasco
Collembola Diplura Archaeognata Blataria Hemiptera
Hymenoptera Coleoptera Diptera Formicidae Lepidoptera
Mantodea Orthoptera Psocoptera Thysanoptera Phasmida

Figura 34. Comparación entre gradientes altitudinales para el número de
morfotipos dentro de cada grupo taxonómico en artrópodos no insectos.
Figura 35. Comparación entre gradientes altitudinales para las abundancias
absolutas dentro de cada grupo taxonómico en artrópodos no insectos.
Al analizar la riqueza taxonómica para los tres gradientes en conjunto dentro de
Hexapoda, los grupos más representativos fueron Collembola con 41 morfotipos,
0
10
20
30
40
50
60
70
Socha Socota Tasco
Diplopoda
Chilopoda
Scorpiones
Pseudoscorpiones
Opiliones
Araneae
Acari
Isopoda
0
50
100
150
200
250
300
Socha Socota Tasco
Diplopoda
Chilopoda
Scorpiones
Pseudoscorpiones
Opiliones
Araneae
Acari
Isopoda
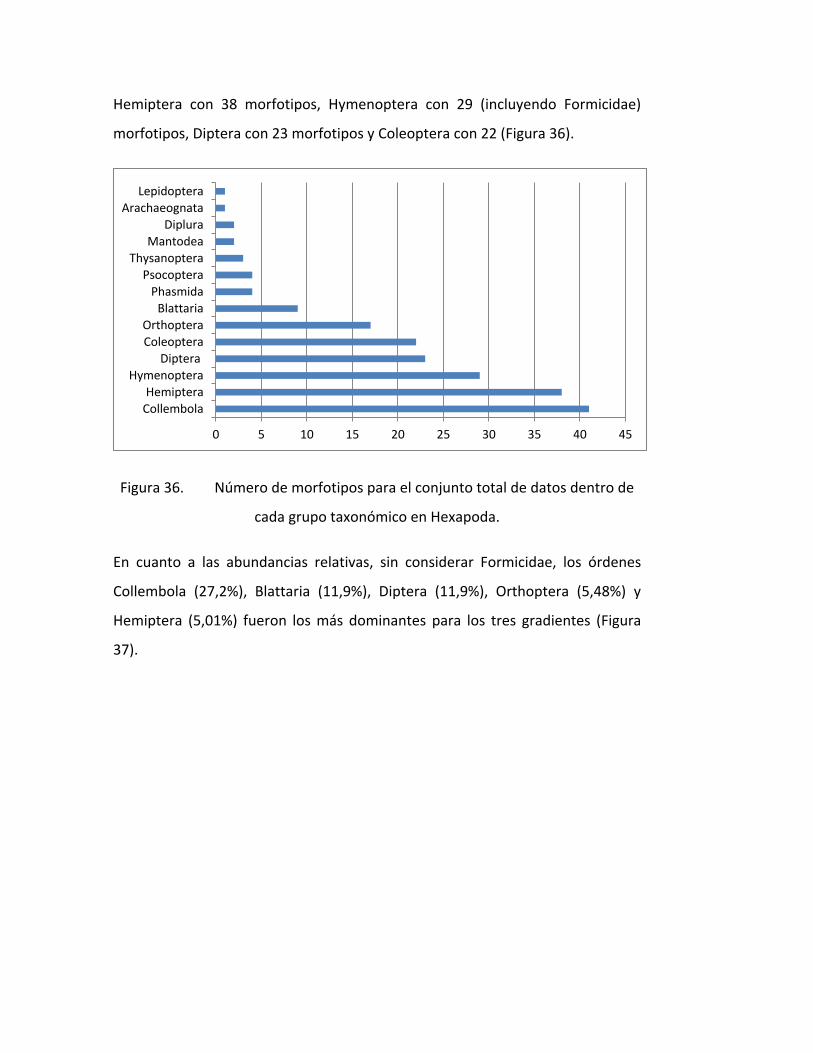
Hemiptera con 38 morfotipos, Hymenoptera con 29 (incluyendo Formicidae)
morfotipos, Diptera con 23 morfotipos y Coleoptera con 22 (Figura 36).
Figura 36. Número de morfotipos para el conjunto total de datos dentro de
cada grupo taxonómico en Hexapoda.
En cuanto a las abundancias relativas, sin considerar Formicidae, los órdenes
Collembola (27,2%), Blattaria (11,9%), Diptera (11,9%), Orthoptera (5,48%) y
Hemiptera (5,01%) fueron los más dominantes para los tres gradientes (Figura
37).
0 5 10 15 20 25 30 35 40 45
Collembola
Hemiptera
Hymenoptera
Diptera
Coleoptera
Orthoptera
Blattaria
Phasmida
Psocoptera
Thysanoptera
Mantodea
Diplura
Arachaeognata
Lepidoptera
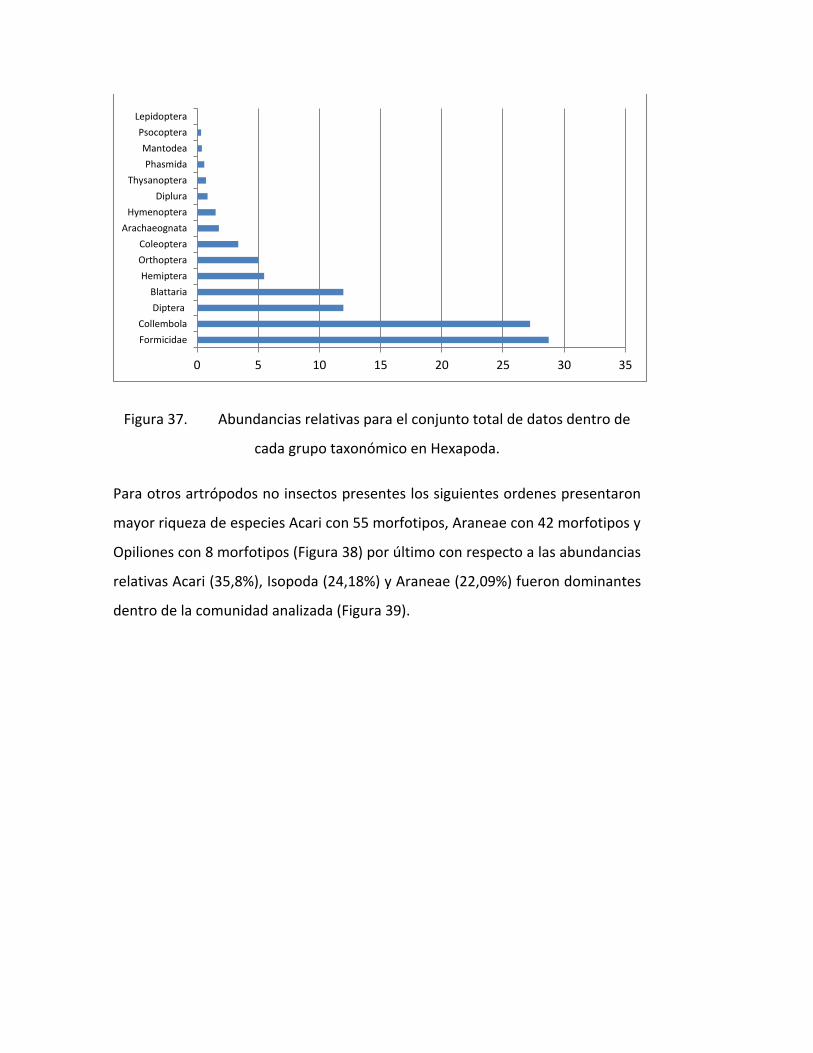
Figura 37. Abundancias relativas para el conjunto total de datos dentro de
cada grupo taxonómico en Hexapoda.
Para otros artrópodos no insectos presentes los siguientes ordenes presentaron
mayor riqueza de especies Acari con 55 morfotipos, Araneae con 42 morfotipos y
Opiliones con 8 morfotipos (Figura 38) por último con respecto a las abundancias
relativas Acari (35,8%), Isopoda (24,18%) y Araneae (22,09%) fueron dominantes
dentro de la comunidad analizada (Figura 39).
0 5 10 15 20 25 30 35
Formicidae
Collembola
Diptera
Blattaria
Hemiptera
Orthoptera
Coleoptera
Arachaeognata
Hymenoptera
Diplura
Thysanoptera
Phasmida
Mantodea
Psocoptera
Lepidoptera
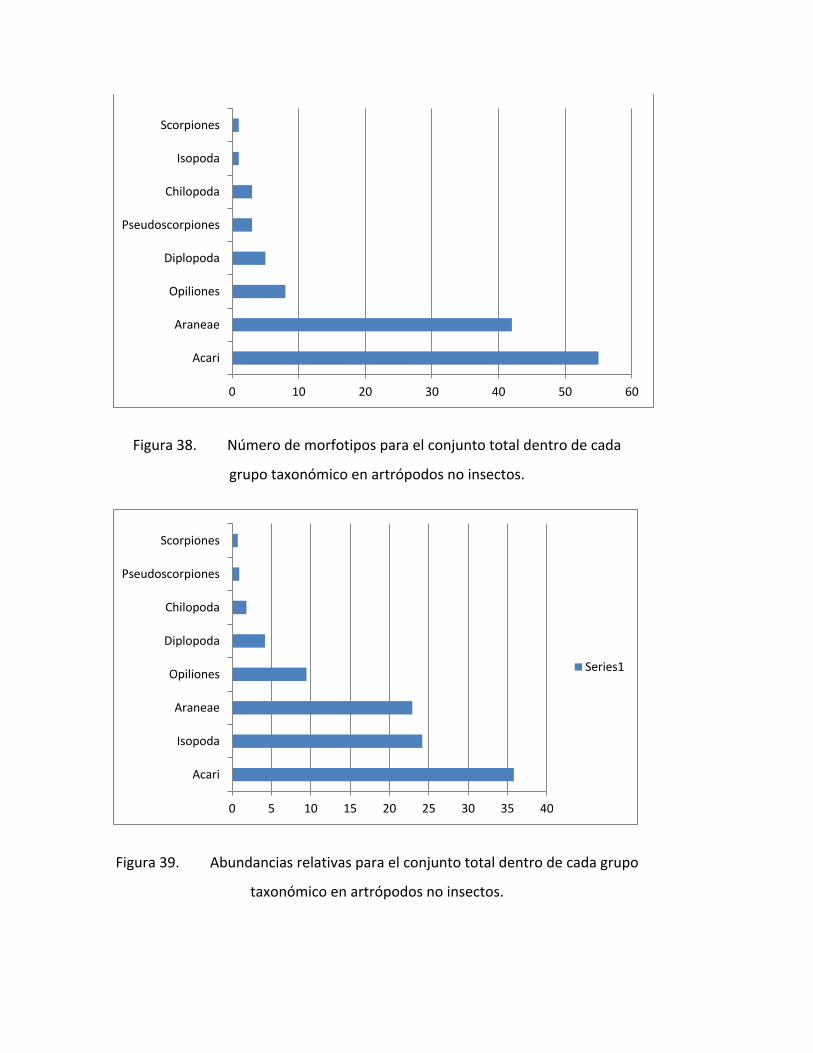
Figura 38. Número de morfotipos para el conjunto total dentro de cada
grupo taxonómico en artrópodos no insectos.
Figura 39. Abundancias relativas para el conjunto total dentro de cada grupo
taxonómico en artrópodos no insectos.
0 10 20 30 40 50 60
Acari
Araneae
Opiliones
Diplopoda
Pseudoscorpiones
Chilopoda
Isopoda
Scorpiones
0 5 10 15 20 25 30 35 40
Acari
Isopoda
Araneae
Opiliones
Diplopoda
Chilopoda
Pseudoscorpiones
Scorpiones
Series1

Al considerar las distintas propiedades de los índices empleados basados ya sea
sobre la riqueza (Shannon-Wiener) o sobre la dominancia (Inverso de Simpson y
Berger-Parker) permiten realizar distintas interpretaciones acerca de los
patrones observados en los gradientes altitudinales de la diversidad de especies.
En el gradiente para Socha según los índices Shannon-Wiener y el inverso de
Simpson la mayor riqueza de especies se observó sobre los 3100m, como
consecuencia de una reducción en la dominancia y que se expresa bajo el índice
de Berger-Parker (Tabla 6).
ÍNDICE/ALTITUD 2800 2900 3000 3100 3200
Riqueza 41 21 27 63 43
Abundancias 94 43 68 169 110
1/d 17,67 22,76 23,99 24,04 24,11
Berger-Parker 0,223 0,14 0,176 0,089 0,145
Shannon-wiener 2,97 2,83 2,79 3,68 3,33
Índices de diversidad para el gradiente altitudinal de Socha.
En Socotá, los índices empleados arrojan resultados dispares en cuanto a las
franjas con mayor riqueza de especies, de acuerdo a Shannon-Wiener sobre los
3000m se registraron valores altos de riqueza de especies, contrastando en parte
con el Inverso de Simpson (1/D) donde la mayor riqueza se obtuvo para los
extremos del gradiente y con Berger-Parker, donde las cotas 2850 y 2915m
presentaron la mayor riqueza o menor (Tabla 7).
ÍNDICE/ALTITUD 2850 2915 3000 3100 3230
Riqueza 64 76 52 41 54
Abundancias 346 316 102 86 177
1/d 12,16 11,25 10,56 11,69 12,93
Berger-Parker 0,63 0,332 0,147 0,151 0,266
Shannon-wiener 2,02 2,81 3,56 3,36 3,14
Índices de diversidad para el gradiente altitudinal de Socota

Por último, en Tasco las cotas 2900, 3000 y 3150m presentaron la mayor riqueza
de especies con respecto a Shannon-Wiener, resultado que nuevamente
contrasta con los demás índices , en este caso los máximos valores de riqueza, de
acuerdo al Inverso de Simpson (1/D) y Berger-Parker se obtuvieron sobre los
3100m para este gradiente (Tabla 8)
ÍNDICE/ALTITUD 2900 3000 3100 3150 3195
Riqueza 44 64 34 53 41
Abundancias 72 151 73 121 139
1/d 7,66 11,61 14,53 16,33 17,67
Berger-Parker 0,083 0,093 0,205 0,107 0,195
Shannon-wiener 3,59 3,82 3,12 3,62 3,05
Índices de diversidad para el gradiente altitudinal de Tasco
Un patrón con respecto a un incremento, disminución o ningún efecto de la
altitud sobre la riqueza de especies se puede observar en la figura al comparar
los índices de diversidad promediados para cada altitud se observa que Shannon-
Wiener y el Inverso de Simpson (1/D) presentan un incremento en la riqueza con
respecto a la altitud, mientras el Índice de Berger-Parker, no presento, en
general ningún cambio importante con respecto a la altitud. Lo anterior se
traduce en un aumento de la equitatividad en la riqueza y una reducción de la
dominancia de las especies hacia las cotas altitudinales superiores (Figura 40).

Figura 40. Tendencias observadas en la riqueza de especies para el conjunto
total de datos, de acuerdo al índice de diversidad empleado (1/D, corresponde al
eje secundario).
En la Tabla 9 se presenta los índices calculados para el total de los datos dentro
de cada cota o franja altitudinal, nuevamente se observa un incremento en la
riqueza de especies con respecto a la altitud
ÍNDICE/ALTITUD alt1 alt2 alt3 alt4 alt5
Riqueza 93 98 65 98 87
Abundancias 511 513 241 374 429
1/d 4,09 19,03 25,11 36,72 16,64
Berger-Parker 0,486 0,163 0,084 0,082 0,191
Shannon-Wiener 2,76 3,76 3,63 4,06 3,57
Índices de diversidad para los datos por cotas altitudinales resumidos.
Con el fin de contrastar el análisis anterior se ordenó la diversidad y equitatividad
(evenness) de las comunidades de suelo a lo largo de cada gradiente altitudinal.
En Socha, la cota 3100 seguido de la cota 3000 presentaron las comunidades con
la mayor riqueza de especies (Figura 41), lo cual se obtuvo para Socotá, aunque
en este caso los perfiles Renyi no discriminan claramente cuál fue la cota más
importante (Figura 42). Por último , las cotas altitudinales dentro del gradiente
0
2
4
6
8
10
12
14
16
18
20
0
0,5
1
1,5
2
2,5
3
3,5
4
Alt 1 Alt 2 Alt 3 Alt 4 Alt 5
Berger-Parker
Shanon-Wiener
1/D

altitudinal de Tasco fueron similares en cuanto a la riqueza de especies, sin
determinarse una comunidad dominante siendo las cotas 3150, 300 y 2900m
presentaron las riquezas más altas (Figura 43)
Figura 41. Perfil de diversidad de Renyi para Socha.
Figura 42. Perfil de diversidad de Renyi para Socotá

Figura 43. Perfil de diversidad de Renyi para Tasco
Con respecto a los perfiles de equitatividad (equitatividad), lo cual muestra la
distribución de las especies con respecto a sus abundancias, ofrece los siguientes
resultados. En Socha, las comunidades para las cotas 2900 y 2800 m presentaron
la mayor y menor equitatividad, respectivamente (Figura 44). Entre tanto, Socotá
la cota 3100 m presento la mayor equitatividad el rango más bajo se observó
para la cota 2850m (Figura 45) y para Tasco, la comunidad con mayor rango se
presentó para la cota de 2900 m, pero si existir un ordenamiento claro para las
demás cotas altitudianles dentro de este gradiente (Figura 46). A partir de los
resultados anteriores se destaca un efecto de importante de las especies más
dominantes dentro de la comunidad y en general la baja equitatividad para todas
las comunidades en cada gradiente altitudinal, dado que en ningún caso las
curvas reflejan una forma plana.
Figura 44. Perfil de Equitatividad de Renyi para Socha.
Figura 45. Perfil de Equitatividad de Renyi para Socotá.

Figura 46. Perfil de Equitatividad de Renyi para Tasco.
Para los gradientes altitudinales en conjunto, las franja entre los 3000 y 3150 m
presenta las comunidades con mayor riqueza de especies (Figura 47), mientras la
franja entre los 2800 y 2900 m presento la riqueza de especies más baja, de igual
forma, la franja entre 3000 y 3150 m y la franja entre 2800 y 2900 m presentaron
las comunidades con mayor y menor equitatividad respectivamente. (Figura 48).
Figura 47. Perfiles de diversidad Renyi para los datos totales por cotas
altitudinales.
Figura 48. Perfiles de equitatividad Renyi para los datos totales por cotas
altitudinales.
4.4.2. Diversidad Beta
Los índices de similitud empleados en este estudio arrojan de manera preliminar
una baja similitud entre cotas altitudinales para cada gradientes altitudinal, es así
como Socha se obtuvo un máximo valor de similitud (Chao-Sørensen-Raw) de
0,52 entre las cotas 3100 y 3200. En Socotá se obtuvo una escaza similitud entre
las cotas 2915 y 3000 m de 0,50 según Chao-Jaccard-Raw, mientras por medio de
Chao-Sørensen-Raw las cotas 2915-3000 m con 0,66; 3100 – 3230 m con 0,63 y
2850-3230 con 0,53, presentaron los máximos valores de similitud y para Tasco
por medio de Chao-Sørensen-Raw las cotas 2900-3000 m y 3100-3195 m
presentaron los máximos valores de similitud (Tabla 10).
De acuerdo, a los índices de similitud basados sobre las especies no observadas,
lo cual “maximiza” la similitud entre cotas asocia en el caso de Socha las cotas
3100-3200m (Chao-Jaccard-Est) y las cotas 3000-3100 m; 3000-3200 m y 3100-
3200 m a través de Chao-Sørensen-Est. En Socotá, Chao-Jaccard-Est asocio las
cotas 2915-3000m y 3000-3100m y por medio de Chao-Sørensen-Est las cotas
2915-3000 m; 3000-3100 m; 3100-3230 m y 2850-3230m (Tabla 11) y por último
para Tasco las distintas cotas presentaron asociación entre sí de acuerdo a los

índices Chao-Jaccard-Est y Chao-Sørensen-Est (Tabla 12). Los anteriores
resultados, evidencian que no existe una zonificación altitudinal clara empleando
comunidades de artrópodos de suelo.
Para el conjunto total de datos, se obtuvo una similitud escasa entre la franja (Alt
1) y la franja (Alt 5), con un valor de 0,574 y entre la franja (Alt 4) y la franja Alt 5
con un valor de 0,531, lo anterior basado sobre Chao-Jaccard-Raw y al emplear
Chao-Sørensen-Raw y Bray-Curtis no se identificaron de manera clara
agrupamientos entre franjas. Con base en los índices Chao-Jaccard-Est y Chao-
Sørensen-Est se obtuvo un solo agrupamiento donde se incluyen todas las franjas
altitudinales (Tabla 13).

CHAO-JACCARD-RAW 2800 2900 3000 3100 3200
2800
0,2 0,013 0,179 0,168
2900
0,013 0,147 0,109
3000
0,228 0,202
3100
0,351
CHAO-JACCARD-EST 2800 2900 3000 3100 3200
2800
0,279 0,025 0,278 0,268
2900
0,015 0,171 0,116
3000
0,43 0,482
3100
0,582
CHAO-SØRENSEN-RAW 2800 2900 3000 3100 3200
2800
0,333 0,025 0,304 0,287
2900
0,026 0,256 0,197
3000
0,372 0,336
3100
0,52
CHAO-SØRENSEN-EST 2800 2900 3000 3100 3200
2800
0,437 0,049 0,435 0,422
2900
0,03 0,292 0,208
3000
0,602 0,65
3100
0,735
BRAY-CURTIS 2800 2900 3000 3100 3200
2800 0,117 0,025 0,152 0,225
2900 0,018 0,151 0,105
3000 0,186 0,202
3100 0,28
Matriz de índices de similitud para el gradiente altitudinal de Socha.

CHAO-JACCARD-RAW 2850 2915 3000 3100 3230
2850
0,179 0,208 0,108 0,367
2915
0,501 0,214 0,25
3000
0,284 0,307
3100
0,468
CHAO-JACCARD-EST 2850 2915 3000 3100 3230
2850
0,257 0,305 0,189 0,497
2915
0,793 0,332 0,313
3000
0,546 0,46
3100
0,544
CHAO-SØRENSEN-RAW 2850 2915 3000 3100 3230
2850
0,304 0,344 0,195 0,537
2915
0,668 0,353 0,401
3000
0,442 0,469
3100
0,638
CHAO-SØRENSEN-EST 2850 2915 3000 3100 3230
2850
0,409 0,468 0,318 0,664
2915
0,885 0,498 0,477
3000
0,706 0,63
3100
0,705
BRAY-CURTIS 2850 2915 3000 3100 3230
2850 0,184 0,205 0,088 0,199
2915 0,206 0,114 0,203
3000 0,277 0,251
3100 0,319
Matriz de índices de similitud para el gradiente altitudinal de Socotá.

CHAO-JACCARD-RAW 2900 3000 3100 3150 3195
2900
0,346 0,244 0,18 0,295
3000
0,251 0,167 0,281
3100
0,242 0,384
3150
0,317
CHAO-JACCARD-EST 2900 3000 3100 3150 3195
2900
0,562 0,525 0,222 0,342
3000
0,292 0,198 0,392
3100
0,34 0,501
3150
0,623
CHAO-SØRENSEN-RAW 2900 3000 3100 3150 3195
2900
0,515 0,392 0,305 0,456
3000
0,401 0,286 0,439
3100
0,39 0,555
3150
0,481
CHAO-SØRENSEN-EST 2900 3000 3100 3150 3195
2900
0,72 0,688 0,363 0,51
3000
0,451 0,33 0,563
3100
0,508 0,668
3150
0,768
BRAY-CURTIS 2900 3000 3100 3150 3195
2900 0,287 0,221 0,249 0,227
3000 0,277 0,206 0,241
3100 0,227 0,368
3150 0,2
Matriz de índices de similitud para el gradiente altitudinal de Tasco.

CHAO-JACCARD-RAW Alt 1 Alt 2 Alt 3 Alt 4 Alt 5
Alt 1
0,352 0,28 0,496 0,574
Alt 2
0,468 0,489 0,37
Alt 3
0,462 0,473
Alt4
0,531
CHAO-JACCARD-EST Alt 1 Alt 2 Alt 3 Alt 4 Alt 5
Alt 1
0,397 0,416 0,668 0,7
Alt 2
0,638 0,622 0,524
Alt 3
0,82 0,745
Alt4
0,756
CHAO-SØRENSEN-RAW Alt 1 Alt 2 Alt 3 Alt 4 Alt 5
Alt 1
0,52 0,438 0,663 0,73
Alt 2
0,638 0,657 0,54
Alt 3
0,632 0,642
Alt4
0,693
CHAO-SØRENSEN-EST Alt 1 Alt 2 Alt 3 Alt 4 Alt 5
Alt 1
0,569 0,588 0,801 0,824
Alt 2
0,779 0,767 0,688
Alt 3
0,901 0,854
Alt4
0,861
BRAY-CURTIS 2800 2900 3000 3100 3200
Alt 1 0,33 0,263 0,231 0,328
Alt 2 0,297 0,293 0,282
Alt 3 0,371 0,376
Alt4 0,421
Matriz de índices de similitud para el conjunto total de datos.
4.5. Discusión
Las comunidades de artrópodos de suelo examinadas para los tres gradientes
altitudinales, reflejan una composición similar a la observada en otros estudios
similares, esto es, un dominio en cuanto a número de especies e individuos por

parte de Acari, Collembola, Araneae y Diptera, y otros grupos que presentaron
una riqueza no esperada de especies Hemiptera y Opiliones.
Sin embargo, otros grupos típicos la fauna epigea del suelo, como Staphylinae,
Scarabaeidae y Carabidae, presentaron una riqueza de especies baja o fueron
ausentes en los muestros, es posible que sea un efecto del diseño de muestreo
utilizado en este estudio, por ejemplo, distancia entre trampas pitfall, que fueron
menores, según lo recomendado en la literatura (Ward et al., 2001), aunque
otros factores no evaluados como disturbio, suelo o estructura de la vegetación
pueden igualmente afectar el muestro de estos grupos.
No obstante, el principal limitante para evaluar la riqueza de especies de suelo,
son las dificultades taxonómicas dentro de grupos que hacen parte de la
microfauna e.g. bacterias, mesofauna e.g. ácaros, colémbolos, y macrofauna e.g.
Anelida, que permitan obtener una imagen real de la diversidad biológica en el
suelo (Wardle, 2002). En este sentido Opiliones, constituye un ejemplo, que
nueves especies para la región de estudio constituye un estimativo alto, si se
compara con cuatro especies reportadas para el departamento de Boyacá (Kury,
2003).
Destaca la importancia de los Collembola y Acari, los cuales son dominantes en
comunidades de suelo (Giller 1996; Heneghan et al. 1998) dentro de la
comunidad, generalmente no abordados en estudios de suelo, en principio, por
la dificultad que subyace a su taxonomía. No obstante, tienen una relevancia
especial, dada las respuestas ecológicas que hacen sus comunidades frente a
distintos factores vegetación, uso de suelo e incluso perturbación (Ponge et al.
2003; Caruso et al. 2007), las cuales, tentativamente pueden influir estas
comunidades para los gradientes altitudinales evaluados y en parte se relacionan
de manera preliminar a los patrones de riqueza de especies de plantas
observados en cada gradiente (ver Figura 1 de este documento); es necesario

entrar a explicar que factores pueden modular las comunidades de colémbolos y
ácaros estudiadas.
Segú Wardle (2002), la vegetación, la entrada de nutrientes al suelo, disturbio y
las propiedades físicas del suelo modulan fuertemente la diversidad de especies
en el suelo, afectando así, los distintos organismos que lo integran, incluso,
grupos no descomponedores como son los Hemiptera (Hartley et al. 2003). Cabe
mencionar que estos factores, pueden actuar de manera directa, sin embargo
muchos otros son indirectos, como es el efecto de la calidad y cantidad de
hojarasca que entra al suelo (Stork y Egleton 1992; Meier y Bowman 2008)
Existen muy pocos estudios comparativos para la fauna de artrópodos del suelo,
y que expliquen la riqueza de especies a lo largo de gradientes altitudinales,
debido a diferencias en los taxones analizados, rangos altitudinales y diferencias
en los protocolos de muestreo. Un rango estrecho considerado en este estudio
tiene como consecuencia que no hace factible evaluar qué tipo de modelo
explica mejor la relación entre riqueza y altitud, al no poderse detectar el efecto
de esta variable.
Con respecto a las tendencias en la riqueza de especies observadas en este
estudio ya sea dentro de cada gradiente altitudinal o para el conjunto total de
datos, sugieren que es la estructura de la vegetación, el principal factor
modulador de la riqueza, además de eventos de disturbio por actividades
humanas como es perdida de la cobertura de la vegetación, lo cual crea “islas de
vegetación” hacia las franjas superiores, asociado a una concentración de la
riqueza (McCain y Grytnes 2010).
La diversidad beta, para cada gradiente altitudinal y el conjunto total de datos
fue en general baja, lo cual refleja un alto recambio de especies. Las
comunidades de suelo, se caracterizan por una alta diversidad alfa y una baja
diversidad beta, lo cual se observa para Acari, como respuesta a condiciones de

suelo o vegetación (Maraun y Scheu 2000; Fagan et al., 2006;Caruso et al. 2007;),
no obstante, en un estudio para Collembola donde se analizaron distintos
factores que controlan la riqueza de especies, incluyendo altitud y hábitat,
realizado por Deharveng y Bedos, (1993) la diversidad beta alta o baja depende
en muchos casos del tipo de hábitat analizado.
El análisis aquí realizado sobre la diversidad beta es preliminar, en el sentido, que
se deben considerar otros factores, como es el efecto de las especies
generalistas en la comunidad, que puedan generar una tendencia en
“homogeneizar” las comunidades de suelo a lo largo del gradiente altitudinal; el
papel del suelo, sus propiedades físicas y químicas (Hansen y Coleman 1998) y el
efecto del uso del hábitat, aprovechamiento de recursos y dispersión.
El estudio realizado acá, está lejos de ser completo, siendo necesario seguir
monitoreando las comunidades de artrópodos de suelo, con el fin comprender y
reconocer patrones generales de riqueza de especies a lo largo de gradientes
altitudinales, particularmente en sistemas fuertemente afectados por actividades
humanas; así mismo, es necesario superar los límites que impone la taxonomía
de taxones claramente dominantes en las comunidades analizadas como son
ácaros y colémbolos.
Si bien los invertebrados del suelo presentan un alto potencial para evaluar la
calidad del suelo y de manera indirecta cambios ocurridos en la estructura de la
vegetación, no son aconsejables para realizar zonificaciones altitudinales, debido
en gran parta a su diversidad beta baja o en otras palabras un recambio de
especies importantes a los largo de gradientes altitudinales, más aun cuando el
rango altitudinal aquí considerado fue estrecho (~400 m), lo cual se pude
evidenciar a partir de otros componentes bióticos como la vegetación.

5. Aves
Dentro del orden Passeriformes, el suborden Oscines es el más representativo
con el 63% (14 especies) y está conformado por aves caracterizadas por su
desarrollo vocal para el canto, el cual es aprendido de padres y vecinos y les
permite generar dialectos locales, el suborden Suboscines agrupa el 37% ( 82
especies del total de las especies registradas), aun cuando es muy abundante en
el Neotrópico y agrupa aves que ocupan gran variedad de hábitats, mostró una
menor riqueza en el área del proyecto (Tabla 14); esta se caracteriza por incluir
especies con una siringe algo primitiva, como es el caso de las especies de la
familia Thamnophilidae (hormigueros) y Furnariidae (trepatroncos y
horneros).(Laverde et al., 2002).
Las aves No-Passeriformes agrupan 156 especies (41 %) del total de las especies
reportadas como probables para el proyecto, entre estas el orden más

representativo es el Apodiformes (colibríes y vencejos) con el 10%, el resto de
orden agrupan menos del 3% del total de las especies reportadas
Las aves No-Passeriformes, se caracterizan por ser especies más antiguas que
presentan una menor riqueza de especies debido a que los integrantes de los
diferentes órdenes muestran una dieta especifica como es el caso de los
Cathartiformes (gallinazos), (Haddad 2008), dentro de este grupo se caracteriza
por su riqueza el orden Apodiforme (vencejos y colibrís) agrupando cerca del
10% de las especies registradas.
Figura 49. Riqueza de aves (%) potenciales, distribuidas en los diferentes
órdenes.
5.1. AVES REGISTRADAS POR TRANSECTOS.
Durante los recorridos efectuados en los transeptos de monitoreo en los meses
de Septiembre y Octubre del 2013, en los municipios de Socota, Socha y Tasco.
Se pudieron registrar un total de 53 especies de aves, distribuidas en ocho
órdenes y 21 familias (Tabla 15), lo que corresponde al 14% del total de las
especies potenciales para el área de estudio.
2%2%
1%3%
1%3%
1%2%3%2%2%2%2%2%
10%1%1%
2%59%
Anseriformes
Galliformes
Podicipediformes
Pelecaniformes
Cathartiformes
Accipitriformes
Falconiformes
Gruiformes
Charadriiformes
Columbiformes
Psittaciformes
Cuculiformes
Strigiformes
Caprimulgiformes
Apodiformes
Trogoniformes
Coraciiformes
Piciformes
Passeriformes
Porcentaje de especies
Ord
en
es

Orden Familia Genero Especies Nombre Común
Cathartiformes Cathartidae Cathartes aura Gallinazo
Coragyps atratus Chulo
Accipitriformes Accipitridae
Accipiter striatus Gavilán
Elanus leucurus Milano
Circus cinereus Águila
Geranoaetus melanoleucus Águila de paramo
Rupornis magnirostris Gavilán
Columbiformes Columbidae Zenaida auriculata Paloma
Patagioenas fasciata Paloma
Strigiformes Strigidae Megascops albogularis Murrucu
Megascops choliba Currucu
Apodiformes
Apodidae Aeronautes montivagus Vencejo
Trochilidae
Chlorostilbon poortmani Colibrí
Colibri thalassinus Colibrí
Colibri coruscans Colibrí
Ensifera ensifera Colibrí pico de lanza
Eriocnemis vestita Colibrí
Lesbia victoriae Colibrí
Metallura tyrianthina Colibrí
Ramphomicron microrhynchum Colibrí
Piciformes Picidae Colaptes rivolii Carpintero
Falconiformes Falconidae Falco sparverius Cernícalo
Passeriformes
Furnariidae Synallaxis subpudica Chamicero
Rhinocryptidae Scytalopus griseicollis Tapa culo
Grallariidae Grallaria ruficapilla Dame pan
Tyrannidae
Elaenia frantzii Copetón
Mecocerculus leucophrys -
Myiotheretes striaticollis -
Ochthoeca fumicolor -
Tyrannus melancholicus Suirirí
Tyrannus savana Tijereta
Hirundinidae
Riparia riparia Golondrina
Hirundo rustica Golondrina
Orochelidon murina Golondrina
Pygochelidon cyanoleuca Golondrina
Turdidae Turdus fuscater Mirlo
Turdus serranus Mirlo
Troglodytidae Troglodytes aedon Cucarachero
Thraupidae
Anisognathus igniventris -
Conirostrum rufum -
Diglossa humeralis -
Catamenia inornata -
Cardinalidae Pheucticus aureoventris -
Parulidae
Setophaga ruticilla Abanico
Basileuterus nigrocristatus -
Myioborus miniatus -
Myioborus ornatus -
Icteridae Sturnella magna Chirlobirlo
Icterus chrysater Toche
Emberizidae Zonotrichia capensis Copetón
Atlapetes pallidinucha -
Fringillidae Sporagra spinescens -
Astragalinus psaltria -
Especies de aves encontradas en los puntos de muestreo
Para comprobar la eficiencia del muestreo en campo y su representatividad se
realizó una curva de acumulación de especies (Figura 50). La cual relaciona a las
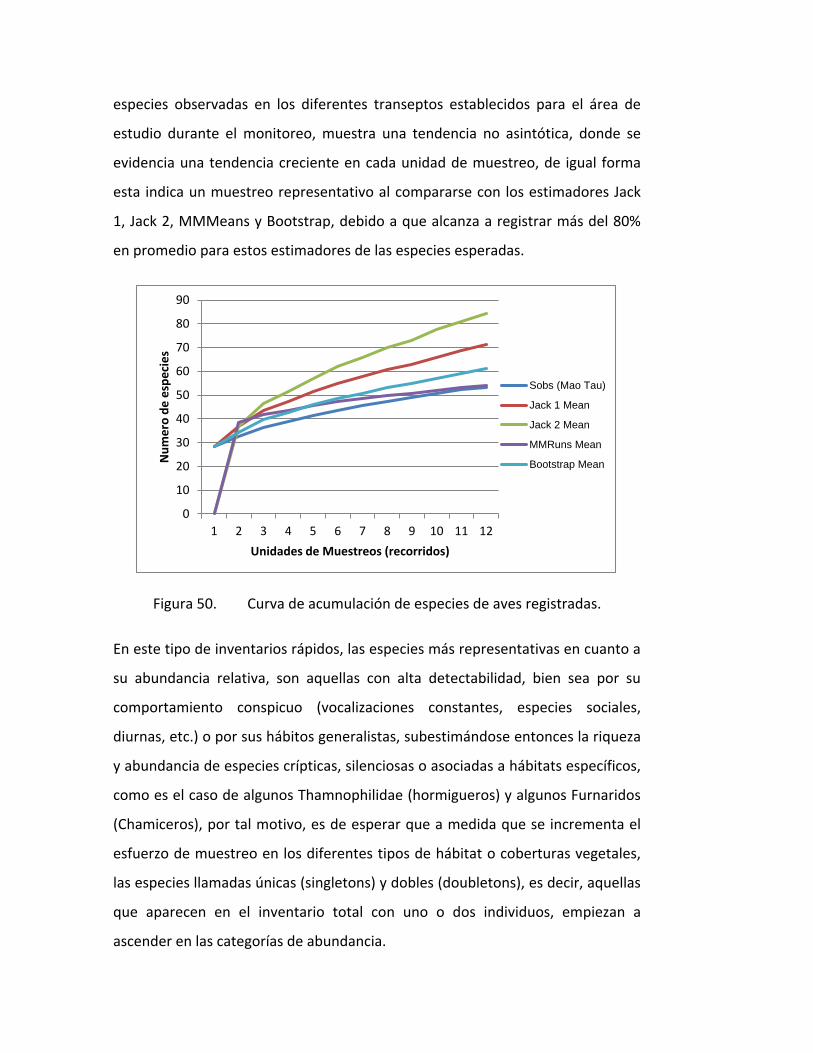
especies observadas en los diferentes transeptos establecidos para el área de
estudio durante el monitoreo, muestra una tendencia no asintótica, donde se
evidencia una tendencia creciente en cada unidad de muestreo, de igual forma
esta indica un muestreo representativo al compararse con los estimadores Jack
1, Jack 2, MMMeans y Bootstrap, debido a que alcanza a registrar más del 80%
en promedio para estos estimadores de las especies esperadas.
Figura 50. Curva de acumulación de especies de aves registradas.
En este tipo de inventarios rápidos, las especies más representativas en cuanto a
su abundancia relativa, son aquellas con alta detectabilidad, bien sea por su
comportamiento conspicuo (vocalizaciones constantes, especies sociales,
diurnas, etc.) o por sus hábitos generalistas, subestimándose entonces la riqueza
y abundancia de especies crípticas, silenciosas o asociadas a hábitats específicos,
como es el caso de algunos Thamnophilidae (hormigueros) y algunos Furnaridos
(Chamiceros), por tal motivo, es de esperar que a medida que se incrementa el
esfuerzo de muestreo en los diferentes tipos de hábitat o coberturas vegetales,
las especies llamadas únicas (singletons) y dobles (doubletons), es decir, aquellas
que aparecen en el inventario total con uno o dos individuos, empiezan a
ascender en las categorías de abundancia.
0
10
20
30
40
50
60
70
80
90
1 2 3 4 5 6 7 8 9 10 11 12
Nu
me
ro d
e e
spe
cie
s
Unidades de Muestreos (recorridos)
Sobs (Mao Tau)
Jack 1 Mean
Jack 2 Mean
MMRuns Mean
Bootstrap Mean

De los órdenes registrados, el más representativo es el Passeriformes (pájaros
cantores) con el 58% (31 especies), del total de las especies registradas, el grupo
de aves No-Passeriformes que agrupa a los órdenes restantes conforma el 42%
(22 especies) del total de las especies registradas (Figura 51), el gran porcentaje
de especies registrada para el Orden Paseriformes concuerda con lo registrado
por Remsem et al. (2011) y Salaman et al. (2008), donde exponen que este orden
es el más diverso en el neotrópico, donde logran ocupar gran variedad de
habitas, debido a su gran diversidad de hábitos alimenticios, además que la
mayoría de sus especies son residente permanentes en Colombia
Dentro del orden Passeriformes, el suborden Oscines es más representativo y
está conformado por aves caracterizadas por su desarrollo vocal para el canto, el
cual es aprendido de padres y vecinos y les permite generar dialectos locales, El
suborden Suboscines, aun cuando es muy abundante en el Neotrópico y agrupa
aves que ocupan gran variedad de hábitats, mostró una menor riqueza en el área
de estudio; se caracteriza por incluir especies con una siringe algo primitiva como
es el caso de las especies de la familia Thamnophilidae (hormigueros) y
Furnariidae (trepatroncos y horneros).(Laverde et al. 2002)
Figura 51. Riqueza de aves (%) en los diferentes órdenes registrados.
0% 10% 20% 30% 40% 50% 60% 70%
Catartiformes
Accipitriformes
Columbiformes
Strigiformes
Apodiformes
Piciformes
Falconiformes
Passeriformes
Porcentaje de aves
Ord
en
es
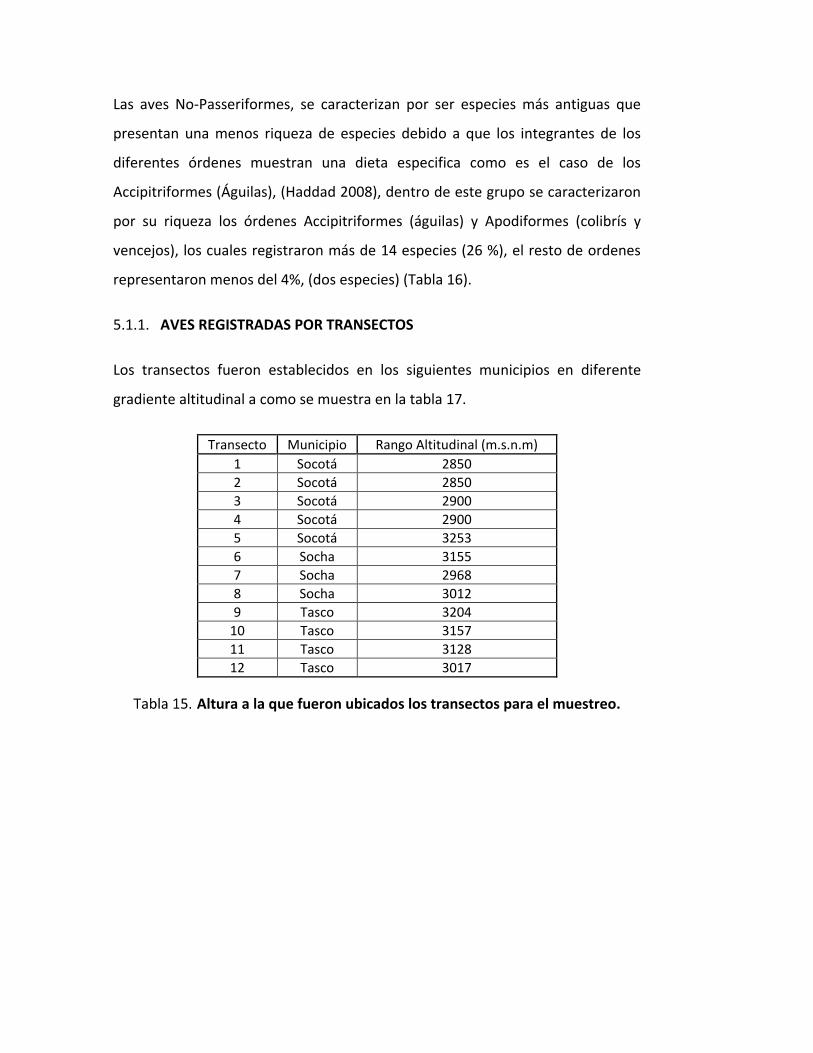
Las aves No-Passeriformes, se caracterizan por ser especies más antiguas que
presentan una menos riqueza de especies debido a que los integrantes de los
diferentes órdenes muestran una dieta especifica como es el caso de los
Accipitriformes (Águilas), (Haddad 2008), dentro de este grupo se caracterizaron
por su riqueza los órdenes Accipitriformes (águilas) y Apodiformes (colibrís y
vencejos), los cuales registraron más de 14 especies (26 %), el resto de ordenes
representaron menos del 4%, (dos especies) (Tabla 16).
5.1.1. AVES REGISTRADAS POR TRANSECTOS
Los transectos fueron establecidos en los siguientes municipios en diferente
gradiente altitudinal a como se muestra en la tabla 17.
Transecto Municipio Rango Altitudinal (m.s.n.m)
1 Socotá 2850
2 Socotá 2850
3 Socotá 2900
4 Socotá 2900
5 Socotá 3253
6 Socha 3155
7 Socha 2968
8 Socha 3012
9 Tasco 3204
10 Tasco 3157
11 Tasco 3128
12 Tasco 3017
Altura a la que fueron ubicados los transectos para el muestreo.

Las especies registrada no mostraron una especificad para cada uno de los
transectos al registrarse la gran mayoría en cada uno de ellos, como se muestra
en la figura 52.
Figura 52. Distribución porcentual de especies de aves en los diferentes
transectos.
Aunque algunas especies se registraron en un solo transepto como lo fueron el
gavilán (Accipiter striatus), colibrí (Eriocnemis vestita), cernícalo (Falco
sparverius), el atrapamoscas (Myiotheretes striaticollis), abaniquito (Setophaga
ruticilla) y el Suirirí (Tyrannus melancholicus), no quiere decir que estas especies
tengan una especificad para un recurso único en el transepto de registro, si no
que su única observación se debió a que estas especies se describen como
cripticas y de poca detectabilidad ya sea por su comportamiento conspicuo o
poca vocalización y actividad (Hilty y Brow 2008).
TRA 1 8%
TRA 29%
TRA 38%
TRA 48%
TRA 59%TRA 6
8%TRA 7
8%
TRA 88%
TRA 98%
TRA 109%
TRA 118%
TRA 129%

5.1.2. SIMILARIDAD DE LAS ESPECIES REGISTRADAS EN LOS
TRANSECTOS
De acuerdo al análisis de similaridad de Jaccard, los transectos establecido
mostraron similaridad de especies al acercarse la mayoría a uno (1) , siendo los
que presentan mayor similitud los transeptos TRA12 - TRA 7 y TRA4 - TRA3 con
0.95 y 0.94 respectivamente para cada uno (Figura 53) con respecto a la
composición de especies, la similaridad entre estos transectos se debe a que las
cotas altitudinales son similares y las especies registradas se distribuyen entre
estas de una manera uniforme (Hilty y Brow 2008).
Figura 53. Índice de similaridad de Jaccard en los transectos de monitoreo.
Se registra una especie endémica el chamicero (Synallaxis subpudica), una
especie endémica para el grupo faunístico de las aves, está restringida a un área
de distribución menor a 50.000 km (Stattersfield et al., 1998). Cuando se llevan a
cabo cambios o afectaciones dentro de las áreas de ocupación de especies con
rangos de distribución restringidos, o cuando estos taxones dependen de
condiciones específicas de hábitat o recursos, la probabilidad de estar en
circunstancias de vulnerabilidad se incrementa.

Las aves de distribución restringida son importantes para la definición de áreas
de conservación, pues tienden a ser especialistas y generalmente presentan
sensibilidad alta a las perturbaciones (IAvH, 1998). La presencia de ciertas
especies con alguna característica especial (algún grado de endemismo,
vulnerabilidad a los disturbios) en determinados lugares, puede servir para
priorizar áreas de interés y diseñar estrategias para su conservación.
5.1.3. ESPECIES POTENCIALES EN EL ÁREA DE ESTUDIO.
Dentro del área de estudio se registran como potenciales 378 especies de aves
estas se encuentran distribuidas en 19 órdenes y 51 familias (Tabla 16), dentro
de estas especies potenciales los órdenes el que cuenta con el mayor número de
especies es el Passeriformes (Pájaros cantores) con un total de 222 especies
(59%), lo cual concuerda con lo reportado por (Remsen et al. 2013) donde
describen que este orden contiene a más de la mitad de las especies registradas
para el territorio nacional y para el neotrópico, esto también se puede explicar
por la variedad de hábitats que pueden ocupar, además de las diferentes dietas
que van desde insectos, frutas, semillas, hasta carroña, además también porque
la mayoría de sus especies residen permanentemente en la región (Correa et al.
2005).

Orden Familia Especie Nombre Local Altitud (m.s.n.m)
Tianmiformes Tinamidae Nothocercus julius Gallineta 1800-3500
Nothocercus bonapartei Gallineta 600-2300
Anseriformes Anatidae
Dendrocygna bicolor Pato <2600
Dendrocygna viduata Pato <2600
Dendrocygna autumnalis Pato <2600
Anas flavirostris Pato 2600-4300
Anas discors Pato <3600
Merganetta armata Pato 1500-3500
Netta erythroptalma Pato <2600
Sarkidiornis melanotos Pato <3500
Oxyura jamaicensis Pato 2500-4000
Galliformes Cracidae
Ortalis guttata Guacharca 540-2000
Penelope argyrotis Pava 900-2300
Penelope montagnii Pava 2200-3400
Aburria aburri Pavón 600-2500
Chamaepetes goudotii - 1500-2500
Odontophoridae Colinus cristatus Perdiz 540-1900
Podicipediformes Podicipedidae Tachybaptus dominicus Cerceta <2600
Podilymbus podiceps Cerceta 3100
Pelecaniformes Ardeidae
Ardea alba Garza <2800
Bubulcus ibis Garza <2200
Egretta caerulea Garza <2800
Egretta thula Garza <2800
Nycticorax nycticorax Garza <2600
Butorides striata Garza <3000
Philherodius pileatus Garza <2600
Agamia agami Garza <2600
Ixobrychus exilis Garza <3000
Threskiornithidae Phimosus infuscatus - 630-1400
Cathartiformes Cathartidae
Cathartes aura Guara 2500
Coragyps atratus Chulo 3600
Vultur gryphus Condor de los andes 2000-4800
Accipitriformes
Pandionidae Pandion haliaetus Águila <3000
Accipitridae
Elanus leucurus Águila <2800
Ictinia mississippiensis - <2800
Circus cinereus - 1800-3000
Accipiter striatus - 1000-3500
Leucopternis princeps - 500-2000
Geranoaetus melanoleucus - 1800-3500
Rupornis magnirostris Gavilán caminero <2500
Buteo platypterus Águila <2500
Buteo leucorrhous Águila 1800-3000
Accipitriformes Accipitridae
Buteo albicaudatus Águila <2400
Buteo swainsoni Águila <2600
Buteo brachyurus Águila <3150
Falconiformes Falconidae
Micrastur ruficollis - <2500
Caracara cheriway Halcón 750-2500
Falco sparverius Halcón 750-2600
Milvago chimachima - <3500
Falco peregrinus Halcón <3500
Gruiformes Rallidae
Porzana carolina - <3000
Neocrex erythrops - <2500
Gallinula melanops - 2300-3000
Gallinula galeta - <3000
Porphyrio martinica - <2600

Orden Familia Especie Nombre Local Altitud (m.s.n.m)
Gruiformes Rallidae Fulica americana - <3500
Charadriiformes
Charadriidae
Vanellus chilensis - <3000
Vanellus resplendens - <3800
Charadrius vociferus - <2600
Scolopacidae
Gallinago galeata - 2800-4000
Gallinago nobilis - <4000
Gallinago jamesoni - 3000-4300
Batramia longicauda - <3500
Actitus macularius - <4000
Tringa flavipes - <3000
Tringa melanoleuca - <3500
Tringa solitaria - <3500
Columbiformes Columbidae
Columbina passerina Palomita <2500
Columbina talpacoti Palomita <2500
Claravis mondetoura - 540-950
Columba livia - <2700
Patagioenas fasciata - 600-3300
Patagioenas subvinacea Torcaza <3150
Zenaida auriculata - <3000
Leptotila verreauxi - <2800
Geotrygon linearis - 1000-2500
Psittaciformes Psittacidae
Pyrrhura calliptera Perico 1800-3500
Forpus conspicillatus Cascabelito <2300
Bolborhynchus lineola - 540-2600
Hapalopsittaca amazonina Perico 1500-3500
Pionus tumultosus - 1500-2800
Pionus chalcopterus - 500-2500
Amazona mercenaria - 1500-3800
Cuculiformes Cuculidae
Piaya cayana - <2800
Coccyzus americanus - <3000
Crotophaga ani Guañu <2600
Crotophaga major - <2500
Tapera naevia - <2000
Dromococcyx pavoninus - 1500-1800
Strigiformes
Tytonidae Tyto alba Buho <4000
Strigidae
Megascops choliba Buho <2800
Megascops albogularis Buho 2000-3500
Cicccaba albitarsis Buho 1800-3000
Glaucidium jardinii Buho 2000-3500
Asio stygius Buho 1500-3000
Asio flammeus Buho <3500
Caprimulgiformes
Steatornithidae Staetornis caripensis - <3000
Caprimulgidae
Lurocalis semitorquatus Lechuza <2600
Lurocalis rufiventris Lechuza 1200-3000
Chordeiles minor Lechuza <2600
Nyctidromus albicollis Lechuza <2600
Caprimulgus carolinensis Lechuza 1000-2600
Caprimulgus longirostris Lechuza 1600-3500
Uropsalis lyra Lechuza 750-950
Uropsalis segmentata Lechuza 2200-3500
Apodiformes
Apodidae
Streptoprocne zonaris - 1000-2500
Streptoprocne rutila - 1000-3000
Chaetura cinereiventris - <2000
Aeronautes montivagus - 500-2500
Trochilidae
Eutoxeres aquila Pico de hoz <2400
Phaethornis guy Colibrí Ermitaño 800-2700
Phaethornis syrmatophotus Colibrí ermitaño 1000-2400
Doryfera ludoviciae Colibrí 500-3200

Orden Familia Especie Nombre Local Altitud (m.s.n.m)
Apodiformes Trochilidae
Schistes geoffroyi Colibrí 800-2200
Colibri delphinae Colibrí 1100-2000
Colibri thalassinus Colibrí 1200-3200
Colibri coruscans Colibrí 1400-3500
Lophornis delatreii Colibrí 600-2200
Adelomyia melanogenys Colibrí 1100-2900
Aglaiocercus kingi Colibrí 1500-2900
Heliangelus amethysticollis Colibrí 2000-3000
Lesbia victoriae Colibrí 2400-3800
Lesbia nuna Colibrí 2200-3300
Ramphomicron microrhynchus Colibrí 2500-3500
Chalcostigma heteropogon Colibrí 2800-3500
Oxypogon guerinii Colibrí 3200-5000
Metallura tyrianthina Colibrí 2000-3600
Haplophaedia aureliae Colibrí 1500-2200
Eriocnemis vestita Colibrí 2300-3700
Eriocnemis cupreoventris Colibrí 2000-3000
Eriocnemis alinae Colibrí 1000-2700
Aglaeactis cupripennis Colibrí 2900-3600
Coeligena coeligena Colibrí 400-2500
Coeligena torquata Colibrí 2000-3500
Coeligena bonapartei Colibrí 2000-3000
Coeligena helianthea Colibrí 2000-3000
Lafresnaya lafresnayi Colibrí 2000-3300
Ensifera ensifera Colibrí 2400-3200
Pterophanes cyanopterus Colibrí 2800-3600
Boissonneaua flavescens Colibrí 1500-3200
Ocreatus underwoodii Cola de raqueta 1200-2600
Heliodoxa leadbeateri Colibrí 1300-2400
Chaetocercus mulsant Colibrí 1500-3200
Chalybura buffonii Colibrí <2000
Chlorostilbon poortmani Colibrí 600-2500
Campylopterus falcatus Colibrí 400-2500
Trogoniformes Trogonidae
Pharomachrus auriceps - 1100-2800
Pharomachrus antisianus - 1400-2500
Trogon personatus - 1000-3500
Trogon collaris - <2400
Coraciiformes
Alcedinidae Megaceryle torquata - <2500
Momotidae Momotus aequatorialis - 1500-3000
Capitonidae Eubucco bourcierii - 1200-2500
Ramphastidae
Aulacorhynchus prasinus Tucaneta esmeralda 1500-3200
Aulancorhynchus haematopygus Tucaneta esmeralda 500-2300
Andigena nigrirostris - 1200-3200
Pteroglosus castanotis Yataro <2000
Picidae
Melanerpes formicivorus Carpintero 1400-2700
Picoides fumigatus Carpintero 700-2700
Colaptes puctigula Carpintero <2000
Colaptes rubiginosus Carpintero <3000
Colaptes rivolii Carpintero 1500-3400
Dryocopus lineatus Carpintero Real <2200
Campephilus pollens Carpintero 1300-3300
Passeriformes Furnariidae
Sclerurus mexicanus - <2100
Cinclodes albidiventris - 3400-4500
Leptasthenura andicola - 3000-4200
Asthenes fuliginosa - 2400-3600
Asthenes flammulata - 2700-4200
Synallaxis subpudica - 1300-3200
Synallaxis azarae Arrurador 1200-3300
Synallaxis unirufa - 2000-3300

Orden Familia Especie Nombre Local Altitud (m.s.n.m)
Passeriformes
Furnariidae
Hellmayrea gularis - 2500-3500
Premnornis guttligera - 1400-2700
Premnoplex brunnescens - 800-3100
Margarornis squamiger - 1500-3500
Pseudocolaptes boissonneautii - 1700-3200
Anabacerthia striaticollis - 600-2800
Syndactyla subalaris - 1000-2800
Thripadectes holostictus - 700-3150
Thripadectes virgaticeps - 1200-2500
Thripadectes flammulatus - 1500-3200
Dendrocincla tyrannina - 1500-3000
Xiphocolaptes promeropirhynchus - 100-3000
Dendrocolaptes picumnus - <2700
Xiphorhynchus triangularis - 1500-2600
Lepidocolaptes lacrymiger - 1700-3500
Drymotoxeres pucheranii - 1000-3000
Thamnophilidae
Thamnophilus doliatus Carcajada <1800
Dysithamnus mentalis - 300-2400
Drymophila caudata - 1200-3150
Myrmotherula schistocolor - 1000-2300
Conophagidae Conoppophaga castaneiceps - 1000-1800
Grallaridae
Grallaria squamigera - 2200-3700
Grallaria ruficapilla - 1200-3000
Grallaria rufula - 1800-3500
Grallaricula nana - 2000-3300
Rhynocriptidae
Acropternis orthonyx - 2300-3200
Myornis senilis - 2300-3700
Scytalopus latrans - 2000-3700
Scytalopus atratus - 800-1700
Scytalopus spillmanni - 1900-3200
Scytalopus griseicollis - 2000-3300
Tyrannidae
Phyllomyias nigrocapillus - 1600-3400
Phyllomyias cinereiceps - 1800-2800
Phyllomyias uropygialis - 1800-2800
Elaenia frantzii - 600-3000
Mecocerculus poecilocercus - 1800-2800
Mecocerculus stictopterus - 2000-3600
Mecocerculus leucophrys - 2500-3600
Mecocerculus minor - 1100-2500
Anairetes agilis - 1800-3400
Serpophaga cinerea - 500-3200
Zimmerius chrysops - <2300
Mionectes striaticollis - 1500-2700
Mionectes olivaceus <2000
Leptopogon superciliaris - 500-2100
Leptopogon rufipectus - 1600-2800
Myiotriccus ornatus - 600-2400
Lophotriccus pileatus - <2900
Hemitriccus granadensis - 1500-3500
Poecilotriccus ruficepis - 1500-2700
Platyrinchus mystaceus - 400-2400
Myiophobus flavicans - 1500-2800
Myiophobus fasciatus - 640-1000
Nephelomyias pulcher - 1800-2500
Pyrrhomyias cinnamomeus - 1500-3000
Empidonax traillii - <1000
Empidonax alnorum - <1000
Contopus virens - <2600
Contopus cooperi - <3000
Orden Familia Especie Nombre Local Altitud (m.s.n.m)
Passeriformes
Tyrannidae
Contopus cinereus - <1500
Contopus fumigatus - 600-2500
Sayornis nigricans - 100-2800
Pyrocephalus rubinus - <2800
Knipolegus poecilurus - 1500-2500
Myiotheretes striaticollis - 2400-3400
Myiotheretes fumigatus - 2000-3600
Ochtoeca frontalis - 2200-3600
Ochtoeca diadema - 1800-3200
Ochtoeca cinnamomeiventris - 1600-3000
Ochtoeca rufipectoralis - 2600-3200
Myiodynastes chrysocephalus - 1000-2900
Myiodynastes luteiventris - <2500
Tyrannus melancholicus Pechiamarillo 540-2600
Tyrannus savana - 630-2000
Tyrannus tyrannus - 750-1800
Myiarchus cephalotes - 1500-2500
Cotingidae
Pipreola riefferi - 1500-3200
Pipreola arcuata - 2000-3400
Ampelion rubrocristatus - 2300-3800
Rupicola peruvianus - 350-2400
Pyroderus scutatus - 1000-2800
Lipaugus fuscocinereus - 1800-3100
Pipridae Masius chrysopterus - 600-2400
Tityridae Pachyramphus versicolor - 1500-2800
Pachyramphus polychopterus - <2000
Vireonidae
Cyclarhis nigrirostris - 1500-2500
Vireo flavifrons - <2000
Vireo olivaceus - <3400
Vireo leucophrys - 540-2700
Corvidae Cyanocorax yncas - 540-1000
Alaudidae Eremophila alpestris peregrina Alomdra 2500-3000
Hirundinidae
Pygochelidon cyanoleuca - <3000
Orochelidon murina - 2000-3600
Stelgidopteryx ruficollis - <2200
Progne tapera - <2500
Progne subis - <3100
Riparia riparia - <3000
Hirundo rustica - <2800
Petrochelidon pyrrhonota - <3000
Troglodytidae
Troglodytes aedon - <3200
Troglodytes solstitialis - 1500-3600
Cistothorus platensis - 2500-4000
Cistothorus apolinari Cucarachero 2500-3800
Pheugopedius mystacalis - 1200-2500
Cinnycerthia unirufa - 2300-3700
Henicorhina leucophrys - 1000-3000
Cinclidae Cinclus leucocephalus - 500-4000
Turdidae
Myadestes ralloides - 350-2800
Catharus fuscater - 800-2800
Catharus aurantirostris - 600-2300
Catharus ustulatus - 780-2000
Catharus fuscescens - <2300
Catharus minimus - -
Turdus ignobilis - 540-2400
Turdus fuscater Ciote 1800-4000
Turdus serranus Ciote 1400-3000
Turdus leucomelas Embarradora <2000
Mimidae Mimus gilvus Mirla Blanca <2600
Orden Familia Especie Nombre Local Altitud (m.s.n.m)
Passeriformes
Thraupidae
Cissopis leverianus Urraca <2000
Schistochlamys melanopis Sabanera <2000
Sericossypha albocristata - 2000-3300
Hemispingus atropileus - 2000-3600
Hemispingus superciliaris - 2200-3400
Hemispingus frontalis - 1500-3200
Hemispingus melanotis - 2000-3000
Hemispingus verticalis - 3000-3600
Cnemoscopus rubrirostris - 2200-3200
Tachyphonus rufus - <1800
Thraupis episcopus Azulejo 540-2500
Thraupis palmarum Azulejo <2000
Thraupis cyanocephala Azulejo 1800-3300
Buthraupis montana - 2200-3700
Buthraupis eximia - 2500-3800
Anisognathus igniventris - 2200-3400
Anisognathus lacrymosus - 2200-3800
Anisognathus sumptuosus - 1300-2900
Chlorornis riefferi - 2000-3400
Dubusia taeniata - 2200-3400
Iridosornis rufivertex - 2500-3600
Pipraidea melanonota - 1300-3000
Tangara rufivertex - 2500-3600
Tangara ruficervix - 1500-2800
Tangara heinei - 500-2300
Tangara vitriolina - 600-2600
Tangara cyanicollis - 700-2500
Tangara guttata - 700-2000
Tangara vassorii - 2000-3400
Tangara nigroviridis - 800-2800
Tangara labradorides - 1300-2700
Tangara gyrola - 640-2000
Tangara xanthocephala - 1300-2200
Tangara arthus - 800-2500
Dacnis hartlaubi Azulejo 1000-2200
Chlorophanes spiza - <2200
Conirostrum sitticolor - 2700-3700
Conirostrum rufum - 2500-3400
Conirostrum albifrons - 1500-3000
Diglossa lafresnayii - 2500-3800
Diglossa humeralis - 2200-3800
Diglossa caerulescens - 1600-3000
Diglossa cyanea - 1400-3500
Diglossa sittoides - 1500-3400
Diglossa albilatera - 1200-3500
Tiaris olivaceus - 600-2400
Chlorospingus ophthalmicus - 1000-3200
Chlorospingus canigularis - 1200-2000
Incertae sedis Saltator striatipectus - 540-2400
Saltator maximus - <2000
Emberizidae
Zonotrichia capensis Copetón 800-3600
Phrygilus unicolor - 2900-4300
Sicalis flaveola - <2600
Volatina jacarina - 540-2400
Sporophila luctuosa - 1300-2600
Sporophila nigricollis - 540-2000
Sporophila minuta - 540-1300
Catamenia analis - 2600-4000
Catamenia homochroa - 2200-3700

Orden Familia Especie Nombre Local Altitud (m.s.n.m)
Passeriformes
Emberizidae
Catamenia inornata - 2200-3700
Arremon brunneinucha - 800-2600
Arremon torquatus - 2000-3600
Atlapetes albofrenatus - 1400-2500
Atlapetes pallidinucha - 2400-3300
Atlapetes latinuchus - 1600-3600
Atlapetes schistaceus - 2000-3600
Cardinalidae
Piranga olivacea - <3000
Piranga rubra - 540-2600
Pheucticus aureoventris Bábaro 1700-3100
Pheucticus ludovicianus Degollado <3500
Parulidae
Vermivora chrysoptera - <2400
Leiothlypis peregrina - <2800
Setophaga pitiayumi - <2600
Setophaga petechia - <2600
Setophaga striata - <3000
Setophaga fusca - <3200
Setophaga cerúlea - 500-2200
Setophaga ruticilla - <3000
Mniotilla varia - <2500
Oporornis philadelphia - <3000
Wilsonia canadensis - <3000
Myioborus miniatus - 600-2800
Myioborus ornatus - 2000-3400
Basileuterus luteoviridis - 2200-3400
Basileuterus nigrocristatus - 2300-3400
Basileuterus cinereicollis - 600-2000
Basileuterus tristriatus - 500-2600
Icteriadae
Psarocolius angustifrons Mochilero <2400
Cacicus chrysonotus - 1700-3500
Cacicus uropygialis - 1500-2450
Amblycercus holosericeus - <2100
Icterus chrysater Toche <2800
Macroagelaius subalaris Toche negro 2000-3200
Chrysomus icterocephalus - 580-2000
Molothrus bonariensis Chamón 580-2400
Molothrus oryzivorus - <2600
Sturnella magna Chirlobirlo <3700
Dolichonyx oryzivorus - <2600
Fringillidae
Sporaga spinescens Torvellino 1500-4000
Astragalinus psaltria Torvellino 540-2400
Sporaga xanthogasta Torvellino 1000-3000
Euphonia cyanocephala Fifi 1200-3000
Euphonia laniirostris Fifi 540-2000
Especies de aves observadasdas en los puntos de muestreo.
En la tabla 19 se usan las siguientes convenciones: El nombre escrito en verde:
Amenazadas, rojo: Endémicas (hay un par amenazadas), azul: Casi endémicas.
Endemismos: Pyrrhura calliptera (Endémico), Coeligena bonapartei (Casi
endémico), Coeligena helianthea (Casi endémico), Tangara vitriolina (Casi
endémica), Conirostrum rufum (Casi endémica), Myioborus miniatus (Casi

endémica), Cistothorus apolinari (Endémico). Grado de amenaza: Aburria aburri
(NT) Casi Amenazado, Vultur gryphus (NT) Casi Amenazado, Pyrrhura calliptera
(VU) Vulnerable, Gallinago nobilis (NT) Casi amenazada, Hapalopsittaca
amazonina (VU) Vulnerable, Cisthothorus apolinari (EN) En Peligro.
6. CONSIDERACIONES FINALES
6.1. Aspectos Metodológicos
Los grupos taxonómicos escogidos para el trabajo de campo son los adecuados,
la vegetación da respuesta a los tratamientos estadísticos y de índices, de igual
forma anfibios, aunque no se presentan datos, esto no quiere decir que no sea
apropiado, lo que nos muestra es la sensibilidad de los organismos a la
conformación y estructura del ecosistema en este periodo de tiempo, en general
disturbado, con poca cobertura, con vegetación muy dispersa, etc. los insectos al
igual que la vegetación dan respuesta, pero consideramos que se debe
intensificar el muestreo en este grupo, por ultimo aves por ser móviles en un
transecto corto para este grupo, lo separa de manera muy gruesa en dos grupos.
Consideramos que los grupos son buenos pero se debe aumentar mínimo 100
metros en los dos extremos y aumentar los transectos para aumentar el volumen
de información, para obtener una muestra más significativa y por ello una
respuesta más acorde.
6.2. Aspectos de la delimitación
Es de tener en cuenta el pasado tanto a corto plazo (ocupación y uso por parte
de pobladores), como el evolutivo, decimos esto, porque los espacios
muestreados han sido seriamente afectados por los diferentes efectos de uso
por el hombre, que ha moldeado espacialmente estos paisajes hasta el grado de
fragmentación que observamos hoy. Teniendo en cuenta las anteriores
consideraciones y conociendo de antemano la importancia de estos ecosistemas
(agua, evolutivamente, escénico, etc.), como también que existen páramos a lo

largo del país de bajo a medianamente conservados, es por ello con toma vital
importancia de atender y conservar estos ecosistemas. Teniendo elementos
teóricos y basados en los resultados biológicos, podemos decir que existen dos
franjas, donde se ve la variación. La primera e inferior es de 2800 y 2900 y en la
franja superior entre 3000 y 3100, (ver capitulo vegetación e insectos).

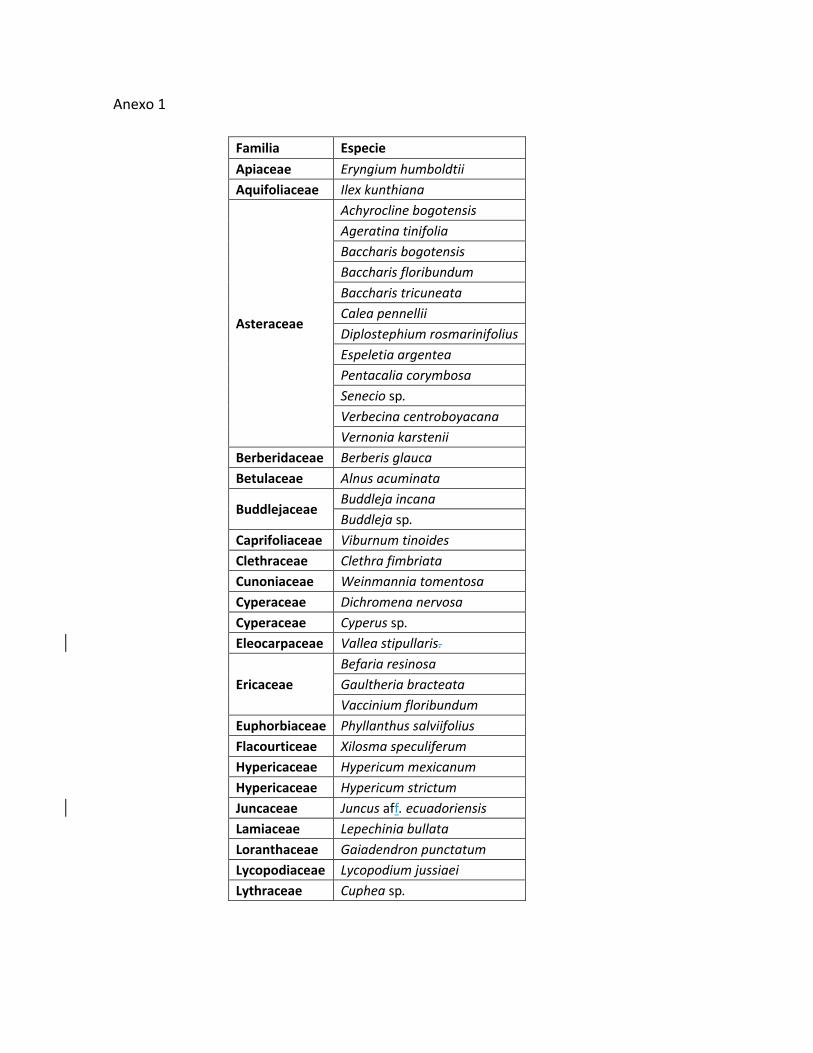
Anexo 1
Familia Especie
Apiaceae Eryngium humboldtii
Aquifoliaceae Ilex kunthiana
Asteraceae
Achyrocline bogotensis
Ageratina tinifolia
Baccharis bogotensis
Baccharis floribundum
Baccharis tricuneata
Calea pennellii
Diplostephium rosmarinifolius
Espeletia argentea
Pentacalia corymbosa
Senecio sp.
Verbecina centroboyacana
Vernonia karstenii
Berberidaceae Berberis glauca
Betulaceae Alnus acuminata
Buddlejaceae Buddleja incana
Buddleja sp.
Caprifoliaceae Viburnum tinoides
Clethraceae Clethra fimbriata
Cunoniaceae Weinmannia tomentosa
Cyperaceae Dichromena nervosa
Cyperaceae Cyperus sp.
Eleocarpaceae Vallea stipullaris.
Ericaceae
Befaria resinosa
Gaultheria bracteata
Vaccinium floribundum
Euphorbiaceae Phyllanthus salviifolius
Flacourticeae Xilosma speculiferum
Hypericaceae Hypericum mexicanum
Hypericaceae Hypericum strictum
Juncaceae Juncus aff. ecuadoriensis
Lamiaceae Lepechinia bullata
Loranthaceae Gaiadendron punctatum
Lycopodiaceae Lycopodium jussiaei
Lythraceae Cuphea sp.
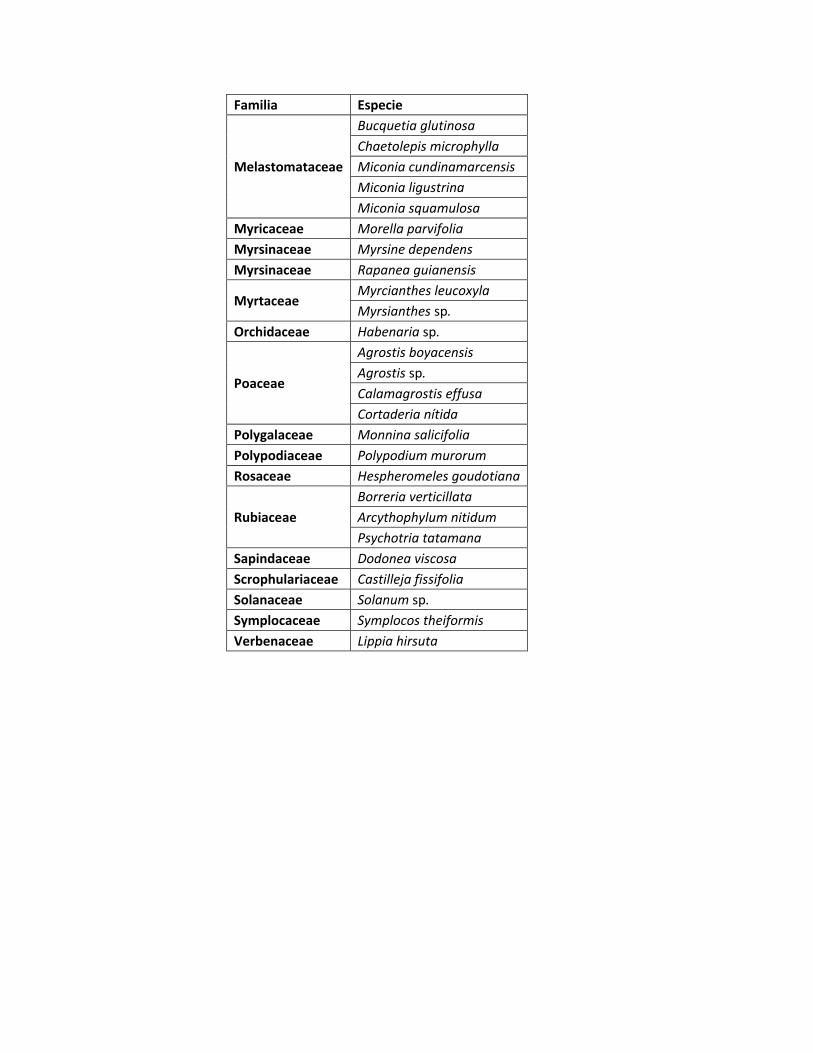
Familia Especie
Melastomataceae
Bucquetia glutinosa
Chaetolepis microphylla
Miconia cundinamarcensis
Miconia ligustrina
Miconia squamulosa
Myricaceae Morella parvifolia
Myrsinaceae Myrsine dependens
Myrsinaceae Rapanea guianensis
Myrtaceae Myrcianthes leucoxyla
Myrsianthes sp.
Orchidaceae Habenaria sp.
Poaceae
Agrostis boyacensis
Agrostis sp.
Calamagrostis effusa
Cortaderia nítida
Polygalaceae Monnina salicifolia
Polypodiaceae Polypodium murorum
Rosaceae Hespheromeles goudotiana
Rubiaceae
Borreria verticillata
Arcythophylum nitidum
Psychotria tatamana
Sapindaceae Dodonea viscosa
Scrophulariaceae Castilleja fissifolia
Solanaceae Solanum sp.
Symplocaceae Symplocos theiformis
Verbenaceae Lippia hirsuta

Anexo 2
ESPECIE
SOCOTÁ SOCHA TASCO
28
75
29
50
30
25
31
00
31
75
28
75
29
50
30
25
31
00
31
75
28
75
29
50
30
25
31
00
31
75
Achyrocline
bogotensis
X
Ageratina
tinifolia
X
X X
X
X
Agrostis sp.
X
X
Alnus acuminata
X
Arcythophylum
nitidum
X
X
Baccharis
bogotensis
X
X
X
Baccharis
floribundum
X X
X X X X
X
Baccharis
tricuneata
X
X
X
Befaria resinosa X
X X
X
X
Berberis glauca X
Borreria
verticillata
X
X
Bucquetia
glutinosa
X X
X X
X
X X
Buddleja incana
X
Buddleja sp.
X
X
X
Calamagrostis
effusa
X
X
X
Calea pennellii X X
X
X
Castilleja
fissifolia
X
Chaetolepis
spmicrophylla
X
Clethra fimbriata X X X X X
X X X X
X X X X
Cortaderia nítida
X
X
Cuphea sp.
X
X
X
Cyperus sp.
X
X
Dichromena
nervosa
X
Diplostephium
rosmarinifolius
X X
X
Dodonea viscosa X X
X
Eryngium
humboldtii
X X
X
X
Espeletia
argentea
X
Gaiadendron
punctatum
X
Gaultheria
bracteata
X
Habenaria sp.
X
Hespheromeles
goudotiana
X
Hypericum
mexicanum
X
X X X
Hypericum
strictum
X
X
X X
X
Ilex kunthiana
X X X X X
X
X
Juncus af.
ecuadoriensis
X
Lepechinia
bullata
X
Lippia hirsuta
X
X
X
Lycopodium
jussiaei
X
Miconia
cundinamarcensis
X X
X X
X X X

ESPECIE
SOCOTÁ SOCHA TASCO
28
75
29
50
30
25
31
00
31
75
28
75
29
50
30
25
31
00
31
75
28
75
29
50
30
25
31
00
31
75
Miconia
ligustrina
X
X
Miconia
squamulosa
X X X X X X X X X
X X X X
Monnina
salicifolia
X
X X
X X
Morella
parvifolia
X X X X X X X X
X X Myrcianthes
leucoxyla
X
Myrsianthes sp.
X
X X
X
Myrsine
dependens
X
X
Pentacalia
corymbosa
X X X X X
X
X X
X
X X Phyllanthus
salviifolius
X
Polypodium
murorum
X
Psychotria
tatamana
X
X
X
Rapanea
guianensis
X X X X
X X
X
X X
X
Solanum sp.
X
Symplocos
theiformis
X X X
Vaccinium
floribundum
X
X
X
Vallea stipullaris.
X
X
X
Verbecina
centroboyacana
X
X X
Vernonia
karstenii
X
X
X
X
Viburnum
tinoides
X
X X X
X
Weinmannia
tomentosa
X
X
X
Xilosma
speculiferum
X
X
TOTAL Sp POR
ESTACION
16 22 9 13 20 17 14 8 10 26 11 14 7 12 9 TOTAL Sp POR
TRANSECTO
80 75 53 SP EXCLUSIVAS POR TRANSECTO
8 14 1

8. LITERATURA CITADA
ACOSTA-GALVIS, A.R. 2000. Ranas, Salamandras y Caecilias (Tetrapoda: Amphibia) de
Colombia. Biota Colombiana: 289-319.
ALLAIRE, J., HORNER, J., MARTI, V., PORTE, N. 2013. Markdown: Markdown rendering for
R. R package version 0.6.3.
ALMEIDA‐NETO, M., MACHADO, G., PINTO‐DA‐ROCHA, R., & GIARETTA, A. A. (2006).
Harvestman (Arachnida: Opiliones) species distribution along three Neotropical
elevational gradients: an alternative rescue effect to explain Rapoport's rule?.
Journal of Biogeography, 33(2), 361-375.
ARELLANO, H., RANGEL, O. 2008. Patrones en la distribución de la vegetación en áreas
de páramo de Colombia: heterogeneidad y dependencia espacial. Caldasia
30(2):355-411.
ARELLANO, H., RANGEL, O. 2008. Patrones en la distribución de la vegetación en áreas
de páramo de Colombia: heterogeneidad y dependencia espacial. Caldasia
30(2):355-411.
ARZAC, A. J., CHACON-MORENO, E., LLAMBI, L. D., y R. DULHOSTE. Distribución de
formas de vida de plantas en el límite superior del ecotono bosque – páramo en los
Andes Tropicales. Ecotrópicos.
ATKIN, L. AND PROCTOR, J. 1988. Invertebrates in the Litter and Soil on Volcan Barva,
Costa Rica. Journal of Tropical Ecology, 4(3):307-310.
BADER, M.Y. 2007. Tropical alpine treelines; How ecological processes control
vegetation patterning and dynamics. PhD Thesis, Wageningen University,
Wageningen, The Netherlands
BECKER, A., KÖRNER, C., BRUN, J., GUISAN, N. TAPPEINER, U. 2007. Ecological and Land
Use Studies Along Elevational Gradients. Mountain Research and Development
27(1):58-65.
BECKER, A., Körner, C., Brun, J., Guisan, N. Tappeiner, U. 2007. Ecological and Land Use
Studies Along Elevational Gradients. Mountain Research and Development
27(1):58-65.
BENNETT, A. 2010. The role of soil community biodiversity in insect biodiversity. Insect
Conservation and Diversity, 3(3): 157-171.
BENNETT, A. 2010. The role of soil community biodiversity in insect biodiversity. Insect
Conservation and Diversity, 3(3): 157-171.

BREHM, G., & Fiedler, K. 2004. Bergmann's rule does not apply to geometrid moths
along an elevational gradient in an Andean montane rain forest. Global Ecology and
Biogeography, 13(1), 7-14.
CADAVID JUAN-G, CESAR ROMÁN- VALENCIA & A. F. 2005. Composición y estructura de
anfibios anuros en un transecto altitudinal de los Andes Centrales de Colombia.
Rev. Mus. Argentino Cienc. Nat., 7(2): 103-118, Buenos Aires.
CARUSO, T., PIGINO, G., BERNINI, F., BARGAGLI, R., & MIGLIORINI, M. 2008. The Berger–
Parker index as an effective tool for monitoring the biodiversity of disturbed soils: a
case study on Mediterranean oribatid (Acari: Oribatida) assemblages. In Biodiversity
and Conservation in Europe (pp. 35-43). Springer Netherlands
CASTAÑO-MORA, O. V, G. CÁRDENAS Y E. HERNÁNDEZ. 2000. Herpetofauna reptiliana
en el bioma páramo. Pp. 58-62.En B. Bermúdez, D. Bernal y H. Vélez. (ed.). Paramos
y bosques de niebla. IV conferencia latinoamericana de paramos y bosques
altoandinos. Málaga, Santander, Colombia. 196 pp;
CHAO, A., R. L. Chazdon, R. K. Colwell, and T.-J. Shen. 2005. A new statistical approach
for assessing compositional similarity based on incidence and abundance
data. Ecology Letters 8:148-159.
COLWELL, R. K. 2013. Estimates: Statistical estimation of species richness and shared
species from samples. Version 9. User's Guide and application published at:
http://purl.oclc.org/estimates.
CORREA, A; J. ARMESTO, R. SCHLATTER, RODRIGUEZ & J TORREZ – MURRA. 1990. La
Dieta del Chucao (Scelorchilus rubecula), un Passeriforme terrícola endémico del
bosque templado húmedo de Sudamérica Austral. Rev. Chilena H y T Nat. 63:197-
202.
CRUMP, M.L. 2003. Conservation of amphibians in the New World tropics. En: Semlitsch,
R.D. (Ed.), Amphibian Conservation. Smithsonian Institution, USA, pp. 53–69.Crump
y Scott 2001
CUATRECASAS, J. 1958. Aspectos de la vegetación natural de Colombia. Rev. Acad. Col.
Cs. Ex. Fis. Nat. 10 (40):221268.
CUATRECASAS, J. 1968. Páramo vegetation and its life forms. En: Troll, C. (ed.),
Geoecology of the montains Regions of the Tropical. Coll. Geogr (Bonn) 9:163-186
DEFENSORÍA DEL PUEBLO, 2010. La minería de hecho en Colombia. Defensoría delegada
para los derechos y del ambiente. 285 pgn.

DEHARVENG, L., & BEDOS, A. 1993. Factors influencing diversity of soil Collembola in a
tropical mountain forest (Doi Inthanon, Northern Thailand). Paoletti MG Foissner W
& Coleman D (Eds) Soil Biota Nutrient Cycling and Farming Systems, 91-111.
DUELLMAN, W. E. 1992. Eleutherodactylus bearsei new species (Anura: Leptodactylidae)
from northeastern Peru. Occasional Papers of the Museum of Natural History,
University of Kansas 150: 1–7.
EOT 2000. Esquema de Ordenamiento Territorial Tasco Boyacá, Alcaldía Municipal de
Tasco, Boyacá. 450.
EOT 2001. Esquema de Ordenamiento Territorial Socotá, Boyacá. Alcaldía Municipal de
Socotá, Boyacá. 623.
ESCOBAR, F., HALFFTER, G., & ARELLANO, L. 2007. From forest to pasture: an evaluation
of the influence of environment and biogeography on the structure of beetle
(Scarabaeinae) assemblages along three altitudinal gradients in the Neotropical
region. Ecography, 30(2), 193-208.
FAGAN, L. L., DIDHAM, R. K., WINCHESTER, N. N., BEHAN-PELLETIER, V., CLAYTON, M.,
LINDQUIST, E., & RING, R. A. (2006). An experimental assessment of biodiversity
and species turnover in terrestrial vs canopy leaf litter. Oecologia, 147(2), 335-347.
FOX, J. 2005. The R Commander: A Basic Statistics Graphical User Interface to R. Journal
of Statistical Software, 14(9): 1-42.
FOX, J. and Weisberg, S. 2011. An R Companion to Applied Regression, Second Edition.
Thousand Oaks CA: Sage.
GARDNER, T.A., BARLOW, J. Y C.A. PERES. 2007. Paradox, presumption and pitfalls in
conservation biology: The importance of habitat change for amphibians and
reptiles. Biological Conservation 138:166-179.
GAVIN L. 2012. Permute: Functions for generating restricted permutations of data. R
package version 0.7-0.
GAVIN, L. 2012. Permute: Functions for generating restricted permutations of data.
Rpackage version 0.7-0.
GESA - GRUPO DE ESTUDIOS DE SISTEMAS ANDINOS. 2004. Caracterización ambiental
del PNN de Pisba y su montaje en un SIG. Universidad Pedagógica y Tecnológica de
Colombia.
GILLER, P. S. 1996. The diversity of soil communities, the ‘poor man's tropical rainforest’.
Biodiversity & Conservation, 5(2), 135-168.

GONZALEZ, W., Llambi, L., SMITH, J., GAMEZ, L. 2011. Dinamica sucesional del
componente arbóreo en la zona de transición bosque - páramo en los andes
tropicales. ECOTRÓPICOS 24(1):60-79.
HAMMER, O., HARPER, D.A.T., AND P. D. RYAN, 2001. PAST: Paleontological Statistics
Software Package for Education and Data Analysis. Paleontología Electronica 4(1):
9pp.
HANSEN, R. A., COLEMAN, D. C. 1998. Litter complexity and composition are
determinants of the diversity and species composition of oribatid mites (Acari:
Oribatida) in litterbags. Applied Soil Ecology 9, 17–23.
HARTLEY, S. E., GARDNER, S. M., & MITCHELL, R. J. 2003. Indirect effects of grazing and
nutrient addition on the hemipteran community of heather moorlands. Journal of
Applied Ecology, 40(5), 793-803.
HENEGHAN, L., COLEMAN, D. C., ZOU, X., CROSSLEY, D. A., & HAINES, B. L. 1998. Soil
microarthropod community structure and litter decomposition dynamics: a study of
tropical and temperate sites. Applied Soil Ecology, 9(1), 33-38.
HERNANDEZ-CAMACHO, J.I. 1997. Comentarios preliminares sobre la paramización en
los Andes de Colombia. Premio a la vida y obra 1997. Fondo Fen, Bogotá.
HILTY, S.L. & BROWN, W.L. 2008. Guía de las aves de Colombia. (Traducción de
Humberto Álvarez López). Cali, Colombia. American BirdConservancy, Universidad
del Valle, Sociedad Antioqueña de Ornitología. Princeton University Press.
HODKINSON, I. D. 2005. Terrestrial insects along elevation gradients: species and
community responses to altitude. Biological Reviews, 80(3), 489-513.
HOFFMANN M, HILTON-TAYLOR C, ANGULO A, BÖHM M, BROOKS TM, BUTCHART SHM,
et al. 2010. The Impact of Conservation on the Status of the World's Vertebrates.
Science 330:1503-1509
IAvH, 2011. Guía divulgativa de criterios para la delimitación de páramos de Colombia.
Instituto de investigaciones biológicas Alexander von Humboldt. Rivera Ospina, D.,
Rodríguez C.
IGAC 1996. Diccionario Geográfico de Colombia, 4 tomos
IUCN 2013. Fernando Castro, María Isabel Herrera, Adolfo Amézquita, Jose Vicente
Rueda 2010. Pristimantis elegans. <www.iucnredlist.org>.
IUCN. 2004. 2004 IUCN Red List of Threatened Species. www.iucnredlist.org.

IUCN. 2010. IUCN Red List of Threatened Species. Version 2010.4.
<www.iucnredlist.org>.
JANZEN, D. H., ATAROFF, M., FARINAS, M., REYES, S., RINCON, N., SOLER, A., & VERA, M.
1976. Changes in the arthropod community along an elevational transect in the
Venezuelan Andes. Biotropica, 193-203.
JONES, R. W., NIÑO-MALDONADO, S., & LUNA-COZAR, J. 2012. Diversidad y afinidades
biogeográficas de Apionidae (Coleoptera: Curculionoidea) a lo largo de un gradiente
altitudinal en la Reserva de la Biosfera El Cielo en el noreste de México. Revista
mexicana de biodiversidad, 83(1), 100-109.
KEATING, P. 1999. Changes in páramo vegetation along an elevation gradient in
southern Ecuador.Journal of the Torrey Botanical Society, Vol 126, No2. (Apr – Jun,
1999), pp. 159-175.
KINDT, R. & Coe, R. 2005 Tree diversity analysis. A manual and software for common
statistical methods for ecological and biodiversity studies. World Agroforestry
Centre (ICRAF), Nairobi.
KOK, K., P. VERWEIJ y H. BEUKEMA. 1995. Effects of cutting and grazing on Andean
treeline Vegetation. En: Churchill, S.P., Balslev, H., Forero, E., y J. L. Luteyn (eds.):
Biodiversity and Conservation of Neotropical Montane Forest. The New York
Botanica Garden. New York. 527-539 p.
KREBS, C. J. 1985. Ecology: The Experimental Analysis of Distribution and Abundance.
Harper and Row. New York.
KRICHER, P. J. 2006. Un Compañero Neotropical. Una introducción a los animales,
plantas, y ecosistemas del trópico del nuevo mundo. Segunda Edición, Modificada y
Expandida. American Birding Association.
KURY, A. B. 2003. Annotated catalogue of the Laniatores of the new World (Arachnida,
opiliones). Revista Ibérica de Aracnología, (especial monográfico nº1), 5-337
LAEGAARD, S. 1192. Influence of fire in the grass páramo vegetation in the jalca of
northeastern Peru. Pp. 151-170 in H. Balslev y J.l. Luteyn (eds): Paramo An Andean
Ecosystem under Human Influence Academic Press.
LEGENDRE, P. and Legendre L. 1998. Numerical ecology. Elsevier. 843p.
LEÓN, O. 2003. Distribución espacial y caracterización de comunidades vegetales de
paramo en un gradiente altitudinal con vegetación natural y en proceso de

paramización (Reserva Forestal del Municipio de Cogua, Cundinamarca). Tesis
Ecóloga. Pontificia Universidad Javeriana.
LLAMBÍ, L. D., FARIÑAS, M., SMITH, J. K., CASTAÑEDA, M. Y BRICEÑO, B. en prensa.
Diversidad de la vegetación en dos páramos de Venezuela: un enfoque multiescalar
con fines de conservación. Proyecto Páramo Andino, Quito, Ecuador.
LOMOLINO, M. A. R. K. 2001. Elevation gradients of species‐density: historical and
prospective views. Global Ecology and Biogeography, 10(1), 3-13.
LYNCH, J. D. & A. M. SUÁREZ-MAYORGA. 2002. Análisis Biogeográfico de los anfibios
paramunos. Caldasia 24 (2): 471-480.
LYNCH, J. D. 1979. Leptodactylid frogs of the genus Eleutherodactylus from the Andes of
Southern Ecuador. The University of Kansas, Museum of Natural History,
Miscellaneous Publications 66:1-62.
LYNCH, J. D. 1997. Intrageneric relationships of mainland Eleutherodactylus II. A review
of the Eleutherodactylus sulcatus group.Revista Acad. Colomb. Cienc. 21 (80):353-
372
LYNCH, J. D. 1998. A new frog (genus Eleutherodactylus) from the cloud forests of
southern Boyacá. Revista Acad. Colomb. Cienc. 22(84): 429-432.
LYNCH, J. D. 1999. Ranas pequeñas, la geometría de evolución, y la especiación en los
Andes colombianos. Revista Acad. Colomb. Cienc. 23 (86): 143-159.
LYNCH, J. D. Y J. M. RENJIFO. 2001. Guía de los anfibios y reptiles de Bogotá y sus
alrededores. Impreso Ediciones Ltda. Bogotá. 78 pp;
MAGURRAN, A. 2004. Measuring biological diversity. Blackwell Publishing. 256p.
MAGURRAN, A. E. 1988. Ecological diversity and its measurement. Princenton Univ.
Londres. 179 pp.
MARÍN, C. 2013. Propuesta metodológica para caracterizar las coberturas vegetales en
los páramos de Santurbán y Rabanal. Proyecto Páramos y Sistemas de Vida.
Instituto de Investigación en Recursos Biológicos Alexander von Humboldt.
Documento interno.
MCCAIN, C. M; AND GRYTNES, J. ARVID. 2010. Elevational Gradients in Species Richness.
In: Encyclopedia of Life Sciences (ELS). John Wiley & Sons, Ltd: Chichester.
MCCAIN, CHRISTY M; AND GRYTNES, J. ARVID. 2010. Elevational Gradients in Species
Richness. In: Encyclopedia of Life Sciences (ELS). John Wiley & Sons, Ltd: Chichester.

MEIER, C. L., & BOWMAN, W. D. 2008. Links between plant litter chemistry, species
diversity, and below-ground ecosystem function. Proceedings of the National
Academy of Sciences, 105(50), 19780-19785.
MELBOURNE, B. A. 1999. Bias in the effect of habitat structure on pitfall traps: an
experimental evaluation. Australian Journal of Ecology, 24(3), 228-239
MENDOZA, H. 2012. Patrones de riqueza específica de las familias Melastomataceae y
Rubiaceae en la cordillera oriental, Colombia, norte de los andes y consideraciones
para la conservación. Colombia Forestal Vol. 15(1) Colombia Forestal Vol. 15(1): 5 -
54 / Enero – Junio.
MMA - MINISTERIO DEL MEDIO AMBIENTE. 2002. Páramos: Programa para el Manejo
Sostenible y Restauración de Ecosistemas de la Alta Montaña Colombiana. Bogotá
D.C.
MORALES M, OTERO J., VAN DER HAMMEN T., TORRES A., CADENA C., PEDRAZA C.,
RODRÍGUEZ N., FRANCO C, BETANCOURTH J.C., OLAYA E., POSADA E. Y CÁRDENAS
L. 2007. Atlas de páramos de Colombia. Instituto de Recursos Biológicos Alexander
von Humboldt. Bogotá, D.C. 208 p
MÚNERA, C. & O. LAVERDE-R. 2002. Uso de hábitat por Capito hypoleucus, una especie
endémica y amenazada de Colombia (Aves: Capitonidae). Tesis de pregrado.
Departamento de Biología. Universidad Nacional de Colombia, Bogotá D. C.
NAVAS, C. A. 2003. Herpetological diversity along Andean elevational gradients: Links
with physiological ecology and evolutionary physiology. Comp. Biochem. Physiol.
133:469–485.
NAVAS, C. A. 2005. Patterns of distribution of anurans in high Andean Tropical
elevations: Insights from integrating biogeography and evolutionary physiology en:
Integrative and Comparative Biology, volume 46, number 1, pp. 82–91
NOGUÉS-BRAVO, D., ARAÚJO, M., ROMDAL, T., RAHBEK, C. 2008. Scale effects and
human impact on the elevational species richness gradients. Nature 453(8): 216-
219.
OKSANEN, F., BLANCHET, G., KINDT, R., LEGENDRE, P., MINCHIN, P., O'HARA, R.,
SIMPSON, G., SOLYMOS, P., STEVENS M., WAGNER, H. 2013. Vegan: Community
Ecology Package. R package version 2.0-8.
OKSANEN, F., Blanchet, G., Kindt, R., Legendre, P., Minchin, P., O'Hara, R., Simpson, G.,
Solymos, P., Stevens M., Wagner, H. 2013. Vegan:Community Ecology Package. R
package version 2.0-8.

OLSON, D. 1994. The distribution of leaf litter invertebrates along a neotropical
altitudinal gradient. Journal of Tropical Ecology, 10(2): 129-150.
OLSON, D. 1994. The distribution of leaf litter invertebrates along a neotropical
altitudinal gradient. Journal of Tropical Ecology, 10(2): 129-150.
PONGE, J. F., GILLET, S., DUBS, F., FÉDOROFF, E., HAESE, L., SOUSA, J. P., & LAVELLE, P.
2003. Collembolan communities as bioindicators of land use intensification. Soil
Biology and Biochemistry, 35(6), 813-826.
POT 1995- 2005. Plan de Ordenamiento Territorial Socha Boyacá 1995 – 2005. 15 doc.
PYRCZ, T. W., WOJTUSIAK, J., & GARLACZ, R. 2009. Diversity and distribution patterns of
Pronophilina butterflies (Lepidoptera: Nymphalidae: Satyrinae) along an altitudinal
transect in north-western Ecuador. Neotropical Entomology, 38(6), 716-726.
R CORE TEAM (2013). R: A language and environment for statistical computing. R
Foundation for Statistical Computing, Vienna, Austria.http://www.R-project.org/.
RAMÍREZ S.J., P. MEZA-RAMOS, M. YÁNEZ-MUÑOZ & J. REYES. 2009. Asociaciones
interespecíficas de anuros en cuatro gradientes altitudinales de la Reserva Biológica
Tapichalaca, Zamora-Chinchipe, Ecuador División de Herpetología, Sección
Vertebrados del Museo Ecuatoriano de Ciencias Naturales, Boletín Técnico 8, Serie
Zoológica 4-5: 35-49
RAMIREZ. S., P. MEZA-RAMOS, M. YAÑES-MUÑOZ & J REYES. 2009. Asociaciones
interespecíficas de anuros en cuatro gradientes altitudinales de la Reserva Biológica
Tapichalaca, Zamora-Chinchipe, Ecuador. Boletín Técnico 8,
RANGEL -CH., J. O. 2000. La región paramuna y franja aledaña en Colombia. Colombia
Diversidad biótica III. La región de vida paramuna. Universidad Nacional de
Colombia - Instituto de Ciencias Naturales, Instituto de Investigación en Recursos
Biológicos Alexander von Humboldt. Bogotá.
RANGEL-CH, J. O. 2003. Biodiversidad en la región del páramo: con especial referencia a
Colombia. Evento: Congreso Mundial de Páramos Ponencia: Libro: Congreso
Mundial De Páramos, Gente Nueva Editorial, p.168 - 200, v.1.
RANGEL-CH., J.O., & A. VELÁZQUEZ. 1997. Métodos de estudio de la vegetación. Pp. 59-
87. En: J.O.Rangel-Ch (ed.), Diversidad Biótica II. Instituto de Ciencias Naturales.
Universidad Nacional de Colombia. Bogotá.
REMSEM, J. V., JR., A. JARAMILLO, M. NORES, J. F. PACHECO, M. B. ROBBINS, T. S.
SCHULENBERG, F. G. STILES, J. M. C. DA SILVA, D. F. STOTZ& K. J. ZIMMER. Versión
[May 2013]. A classification of the bird species of South America. American
Ornithologists'Union.http://www.museum.lsu.edu/~Remsen/SACCBaseline.html

RUEDA-ALMONACID J.V., LYNCH., J.D. Y AMEZQUITA, A. (eds). 2004. Libro rojo de
anfibios de Colombia. Serie de Libros Rojos de Especies Amenazadas de Colombia.
Conservación Internacional Colombia, Instituto de Ciencias Naturales – Universidad
Nacional de Colombia, Ministerio del Medio Ambiente, Bogotá, D. C. Colombia. 384
pp.
RUIZ, A Y RUEDA - ALMONACID, J.V. 2008. Batrachochytrium dendrobatidis and
Chytridiomycosis in Anuran Amphibians of Colombia. Ecohealth5:27-33.
RUSEK, J. 1998. Biodiversity of Collembola and their functional role in the ecosystem.
Biodiversity & Conservation, 7(9): 1207-1219.
SALAMAN, P., DONEGAN, T. & CARO, D. 2008. Listado de las Aves de Colombia
2008.Conservación Colombiana 5: 1-85. Mayo 2008.
SODHI, N. S., BICKFORD, D., DIESMOS, A. C., LEE, T. M., KOH, L. P., BROOK, B. W.,
SEKERCIOGLU, C. H. Y BRADSHAW, C.J.A. 2008. Measuring the Meltdown: Drivers of
Global Amphibian Extinction and Decline. PLoS ONE 3: e1636
STATTERSFIELD, A., CROSBY, M.J., LARGAS, A.J. Y WEGE, D. C. 1998 Áreas de Aves
Endémicas del mundo: las prioridades para la conservación de la biodiversidad.
Cambridge, Reino Unido: BirdLife International.
STILES, F.G. & C.I., BOHÓRQUEZ. (2000). Evaluando el estado de la Biodiversidad: El caso
de la avifauna de la serranía de las Quinchas, Boyacá, Colombia. Caldasia: 22(1): 61-
92.
STORK, N. E. and Eggleton, P. 1992. Invertebrates as determinants and indicators of soil
quality. American Journal of Alternative Agriculture, 7(1): 38-47.
STURM, H. 1998. The Ecology of the Páramo Region in Tropical High Mountains. Verlag
Franzbecker: 286 pp. Hildesheim-Berlín
TÓTHMÉRÉSZ, B. 1995 Comparison of Different Methods for Diversity Ordering. Journal
of Vegetation Science, 6(2): pp. 283-290
URBINA-CARDONA, J.N. 2008. Conservation of Neotropical herpetofauna: research
trends and challenges. Tropical Conservation Sciences Vol.1 (4):359-375.
VAN DER HAMMEN, T. 1997. Plan Ambiental de la Sabana de Bogotá. CAR, Bogotá.
VELÁSQUEZ, B.E., CASTRO-HERRERA, F., BOLIVAR-G, W. Y HERRERA-MONTES, M.I. 2008.
Infección por el hongo quitrido Batrachochytrium dendrobatidis en anuros de la
Cordillera Occidental de Colombia. Herpetotropicos - Journal on Tropical
Amphibians and Reptiles 4: 65 – 70.

VENABLES, W. N. & Ripley, B. D. (2002) Modern Applied Statistics with S. Fourth Edition.
Springer, New York.
VILLAREAL, H., ALVAREZ, M., CORDOBA, S., ESCOBAR, F., FAGUA, G., GAST, F.,
MENDOZA, H. 2004. Manual de métodos para el desarrollo de inventarios de
biodiversidad. Instituto de Investigación de Recursos Biológicos Alexander von
Humboldt, Programa de Inventarios de Biodiversidad, Grupo de Exploración y
Monitoreo Ambiental (GEMA). 235 pág.
VOYLES J, YOUNG S, BERGER L, CAMPBELL C, VOYLES WF, DINUDOM A. 2009.
Pathogenesis of chytridiomycosis, a cause of catastrophic amphibian declines.
Science. 326:582–585
VREDENBURG V.T., KNAPP R.A., TUNSTALL T.S. Y BRIGGS C.J. 2010. Dynamics of an
emerging disease drive large-scale amphibian population extinctions. Proceedings
of the National Academy of Sciences 107:9689-9694.
WALKER S.F., JAIME B., GOMEZ V., GARNER T.W.J., CUNNINGHAM A.A., SCHMELLER D.S.,
NINYEROLA, M. HENK, D.A. GINESTET, C. ARTHUR, C-P. Y FISHER, M.C. 2010. Factors
driving pathogenicity vs. prevalence of amphibian panzootic chytridiomycosis in
Iberia. Ecology Letters 13:372-382.
WARD, D. F., NEW, T. R., & YEN, A. L. 2001. Effects of pitfall trap spacing on the
abundance, richness and composition of invertebrate catches. Journal of Insect
Conservation, 5(1), 47-53.
Wardle, D. A. 2002. Communities and ecosystems: linking the aboveground and
belowground components (Vol. 34). Princeton University Press.
Wolda, H. 1987. Altitude, habitat and tropical insect diversity. Biological Journal of the
Linnean Society 30:313–323.
WOLDA, H. 1987. Altitude, habitat and tropical insect diversity. Biological Journal of the
Linnean Society 30:313–323.
XIE, Y. 2013. Knitr: A Comprehensive Tool for Reproducible Research in R. In Victoria
Stodden, Friedrich Leisch and Roger D. Peng, editors, Implementing Reproducible
Computational Research. Chapman and Hall.


















