
Fisiología de Iones Del Transporte
48
FISIOLOGÍA DE IONES DEL TRANSPORTE A TRAVES DEL TONOPLASTO DE PLANTAS SUPERIORES
Transcript of Fisiología de Iones Del Transporte

FISIOLOGÍA DE IONES DEL TRANSPORTEA TRAVES DEL TONOPLASTO DEPLANTAS SUPERIORES

INTRODUCCIÓN

¿QUÉ ES UN TONOPLASTO?
• El tonoplasto es la membrana que delimita la vacuola central en las células vegetales.
• Es selectivamente permeable• incorporar ciertos iones al interior de la vacuola.• responsable de la turgencia celular
• almacena agua con muy poco gasto de energía.• Regula la homeostasis y la osmorregulacion celular

TRANSPORTE DE TONOPLASTOS
Se a definido dos tipos de transportes: • Transporte primaria son los
que requieren metabolitos de alta energía para su funcionamiento han sido caracterizado a nivel funcional y molecular.
• Transporte secundario sólo se han definido en el nivel funcional, con la excepción de la proteína intrínseca tonoplasto

CONCLUSIÓN LLEGADA
• Estos estudios han a la comprensión de la función de la vacuola en la fisiología de células de la planta. • Se ha presentado evidencia de la
importancia de la transportadores tonoplasticos.• acumulación de sodio y tolerancia
a la sal vacuolar Ca2 + liberación y la transducción de señales, y para el almacenamiento de CO2 en forma de malato

• Las membranas lipo-proteicas de las vacuolas aíslan a las hidrolasas del resto de la célula las que no digieren a la membrana (no se sabe por qué razones). Esas membranas son permeables al agua y a pequeñas moléculas (aminoácidos, monosacáridos, ácidos grasos) y poseen bombas de protones (que aseguran el mantenimiento de un pH bajo en el interior) y proteínas canal de Cl -

TRANSLOCACIÓN DE LA BOMBA DE PROTONES

VACUOLAR H+-ATPASA

¿QUÉ SON LAS V-ATP ASA?
• V -ATPasas son enzimas que funcionan en la membrana plasmática son encimas especializadas.
• La función es la conversión de energía biológica ,generando la fuerza motora de protones ( PMF) por hidrólisis de ATP y por lo tanto proporcionar la fuerza motriz para una amplia gama de secundaria activa y pasiva procesos de transporte.
• Estas enzimas funcionan también en la homeostasis del pH celular así como en la acidificación del interior de varios orgánulos

V-ATPASA
• Es responsable de la acidificación de compartimientos en muchos organismos, lo que permite el correcto funcionamiento de proteasa y otras enzimas hidrolíticas. Por ejemplo, dentro de las vacuolas de las plantas superiores y los hongos el pH se mantiene muy por debajo del citoplasma circundante por acción de esta ATPasa (cuya denominación proviene de "vacuola"). Son también responsables de la acidificación de lisosomas, endosomas, complejo de Golgi y vesículas secretoras en células animales. No experimentan fosforilación ni desfosforilación, por lo cual tampoco son inhibidas por el vanadato. Aún no se conoce le mecanismo por el cual acoplan la hidrólisis del ATP al transporte de protones.

ESTRUCTURA DE LA V-H ATPASA
• Las ATPasas son enzimas muy complejas, conformadas por cadenas de aminoácidos muy largas, que pueden ir desde los 600 a los 1200 aminoácidos.
• con un peso molecular nativo de entre 400 y 650kDa
• Las subunidades que componen la enzima funcional se pueden agrupar en dos categorías basadas en su asociación con ya sea el periférico , hidrófilo

ISOFORMAS
• Se ha purificación complementos enzimaticos de papa , espinaca y Arabidopsis thaliana en las cuales se destaca la similitud alta con bacteria ,y mamífero y una menor similitud con bobino aunque su estructura es similar difiere en la función de las subunidades.• Todo esto debido a las familias multigénicas de
las v-atpasas.•

ISOFORMAS
V-ATPasa BACTERIANA V-ATPasa MAMIFERO

REVERSIBILIDAD DE LA V -ATPASA
• Se demostró el desacoplamiento parcial de la bomba a través de cambios en el pH en ambos lados de la tonoplasto y ayudar a explicar la capacidad de la V - ATPasa en el establecimiento de diferentes valores de pH vacuolar .
inhibidores
• Bajo pH.• Bafilomicina A1.• Pirofosfatasa vacuola.

REVERSIBILIDAD DE LA V -ATPASA

REGULACIÓN DE LA V-ATPASA
• En las células vegetales que acumulan de NaCl, la conducción actual para el transporte activo secundario Na + en la vacuola es provista por la fuerza motora de protones que genera a través del tonoplasto dos bombas + H, la V-ATPasa y V-PPasa. Esto indica que la V-ATPasa puede desempeñar un papel fundamental en la activación de Na + / H + antiporte actividad en las células que acumulan cantidades considerables de NaCl.

VACUOLA PIROFOSFATASA

Pirofosfatasa es el nombre genérico de un grupo de enzimas con actividad hidrolasa que catalizan la ruptura del enlace de alta energía entre grupos fosfatos del compuesto químico pirofosfato. Dicha actividad suele estar asociada a rutas metabólicas con termodinámicamente desfavorables, pues la mencionada hidrólisis es muy exergónica.
Suele hacerse referencia a una pirofosfatasa inorgánica como aquélla que degrada moléculas de pirofosfato. Sin embargo, existen otras enzimas con actividad ligeramente distinta, como la tiamín pirofosfatasa (que degrada tiamín difosfato). Existen pirofosfatsas, como las de la vacuola (H+ PPiasa), que intervienen en el transporte de membrana del tonoplasto.
Pirofosfatasa

Sistemas de transporte en vacuolas
Poseen en su membrana una V-ATPasa y una pirofosfatasa (H+-PPasa) vacuolar que impulsan la acumulación de protones Esto genera un gradiente de potencial electroquímico de H+'s a través del tonoplasto (pH 3-6 interior y un 20 mV positivo en el interior). Este gradiente de potencial electroquímico impulsa el transporte y acumulación de diversos solutos desde el citosol: Cl- and NO3-, Na+, Ca2+, Mg2+ y azúcares como sacarosa. En transporte de agua se hace a través de acuaporinas (TIPs). En el tonoplasto existen diversos canales de iones y contratransportadores proton/X a través de los que se acumulan Na+, Ca2+ y sacarosa.

En la actualidad la sequia afecta más del 10% de la
tierra cultivable esta condición se está incrementando
debido al cambio climático. Estas condiciones reducen
la productividad del cultivo de frijol y en casos
extremos provoca la muerte de la plantas. Sin
embargo, existen alternativas de mejoramiento
genético como la transferencia directa de genes que
confieren tolerancia a sequía, tales como el gen H+-
Pirofosfatasa Vacuolar de Arabidopsis thaliana (AVP1)
que aumenta el gradiente vacuolar de protones
resultando en un incremento de acumulación de
solutos y retención de agua.


Cotransportadores iónicosAntiporte Na+/H+
Antiport. El soluto es transportado en dirección opuesta al H+/Na+.

El Sodio internado en la célula puede ser
almacenado en la vacuola. La vacuola es una
estrategia eficiente para el ajuste osmotico
producto de un estrés salino.
Se han clonado antiporters Vacuolar de Na+/H+ .
Los antiporter Na+/H+ funcionan en la membrana
plasmática exportando Na+ desde la célula.
El sodio en el citosol puede ser exportado
nuevamente al apoplasto. Se espera que los
antiporters Na+/H+ de la membrana plasmatica
realicen eficientemnente esta función.
CA 2+/H+ ANTIPORT

CA 2+/H+ ANTIPORT
• La acumulación vacuolar de Ca 2+ en plantas es explicada por la actividad del tonoplasto Ca 2+/H+ antiporte.• . Estudios farmacológicos han demostrado que el
sistema de Ca 2+/H+ antiporte es inhabilitado por varios agentes que afectan otros transportadores de calcio incluyendo: ruthenium, verapamil, La 3+, Cd2+, y otros cationes divalentes.• Existen resultados sobre la presencia de Cd 2+/H+
antiporte de la raíz de avena.

• Salt y Wagner demostraron la absorción de Cd 2+ por el tonoplasto enriquecido con vesículas de las raíces de avena energizadas por ATP o por K/nigericim.• Se sugiere la labor de detoxificación por el Cd/H
antiporte por el cual Cd es sacado por el citoplasma para prevenir el potencial tóxico de la reacción conteniendo una mezcla de SH. • Es posible que la actividad del sistema antiporte
de Cd/H quizá sea el mismo que el sistema Ca/H, como es indicado en los resultados de Chanson, quien demostró la inhibición de Ca/H por Cd2+.

L a t e r c e r a c a t e g o r í a d e t r a n s p o r t e d e i o n e s p r e s e n t e e n e l t o n o p l a s t o c o r r e s p o n d e a l o s c a n a l e s d e i o n e s q u e e s t á n m e d i a d o s p o r e l
m o v i m i e n t o d e i o n e s c o n b a j o g r a d i e n t e e l e c t r o q u í m i c o .
CANALES IÓNICOS

SLOW-ACTIVATING CHANNELS (LENTA ACTIVACIÓN DE CANALES)
• Fue el primer canal descrito en el tonoplasto de las células vegetales y fue presentado en todas las especies estudiadas. • Sus principales características son:La lenta y constante activación del canal.Una marcada salida de potenciales positivos en el
tonoplasto. Activación citoplasmática de Ca 2+ de altos
niveles de 10-6 M.Una mínima especificidad por cationes
monovalentes.

• La disminución del pH de 7.2 a 5.5 no tuvo efecto en la actividad de SV en las células de guardia de la cebolla. Sin embargo, el pH ácido redujo la actividad de SV en los canales de las células de guardia Vicia faba.
• La regulación de los canales de SV por calmodulina ha sido demostrada y apareció para ser dependiente de Ca 2+.• La calmodulina estimuló la corriente SV para
almacenar las proteinas vacuolares de la aleurona de cebada, pero solo con Ca 2+ debajo de 10-5M.
REGULACIÓN DE CANALES
CALMODULINA

• Los antagonistas de calmodulina como trifluoperazine y calmidazolium son directamente inhibidores del canal SV por afectar su mecanismo.• La estimulación vacuolar de los canales SV es a
través del tonoplasto asociada a calmodulina activada por calcio.
Estas indicaciones pueden ser reguladas por la asociación de calcio y calmodulina.
• Cambios en los niveles citoplasmáticos de cloruro también son reportados como reguladores de los canales, reduciendo su actividad.
CLORURO

• Regulación en raíz de remolacha de los canales SV por Mg2+ tuvo también reportes. Si la concentración de magnesio cambia de 0.4 Mm a 2.4 Mm en vacuolas aisladas resulta un fuerte incremento de la magnitud de las corrientes de SV .
IONES DE MAGNESIO

CANALES DE ACTIVACIÓN RÁPIDA

CANALES DE ACTIVACION RAPIDA
• Los canales de rapida activacion (FV) de las vacuolas Son canales que están activos en niveles fisiológicos de Ca2 + C, son independiente de tensión, y tienen una selectividad entre K + y Cl-.• Características similares se han reportado en las
vacuolas de cultivos en células de la remolacha azucarera, células de V. faba guardia y el tejido de almacenamiento de la remolacha roja. Los canales de FV a partir de remolacha son tan selectivos para K + como para Na +, con una relación de permeabilidad.


Una mayor selectividad se informó para la FV canales de células oclusivas con una secuencia de permeabilidad K +> Rb +> NH4 + >> Cs + »de Na +» Li + (117). Los canales de FV de vacuolas celulares de oculcion también fueron estimulados por los pequeños aumentos en Ca2 + c. Registros de la actividad del canal FV en toda la vacuola y el nivel de un solo canal sugieren que estos canales pueden función en la liberación y absorción de K + durante la osmorregulación celular, y en el caso particular de células oclusivas durante la apertura y cierre de los estomas (117). La comprensión de la función de los canales de FV se ha obtenido mediante el estudio la regulación de estos canales por factores citoplasmáticos.

CANALES SELECTIVOS DE CA2 +-

• Se estableció que el Ca2 + juega un papel importante en el estímulo-respuesta de acoplamiento en las plantas. Algunos de los procesos de acoplamiento estímulo-respuesta donde los cambios en el Ca2 +.
• los canales selectivos se han descrito hasta la fecha: canales dependientes de voltaje y canales agonista-activado

CANALES DEPENDIENTES DE VOLTAJE
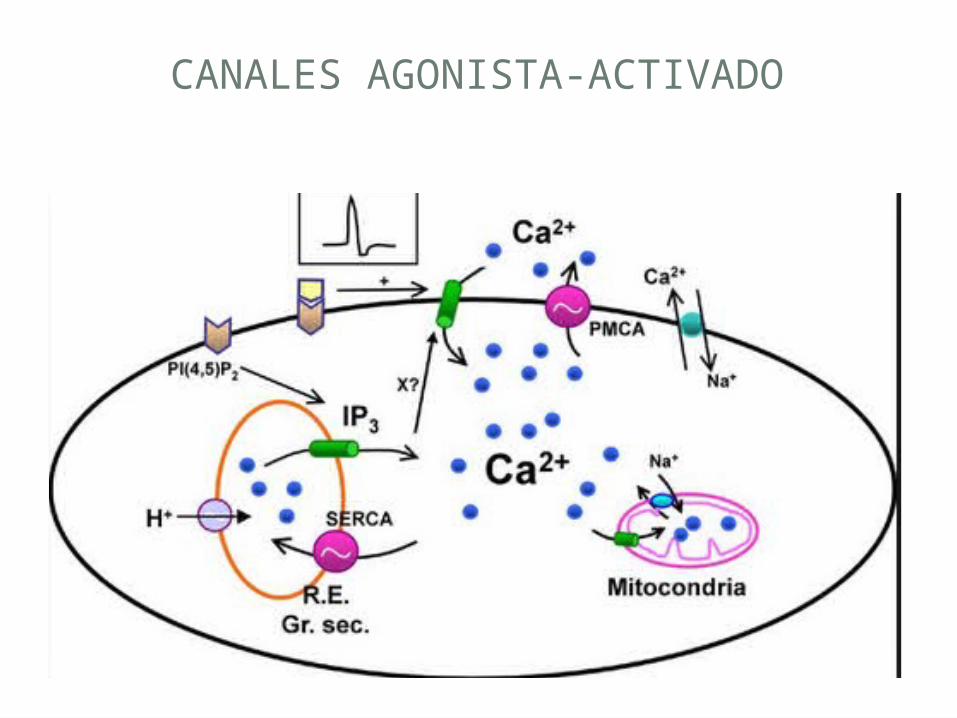
CANALES AGONISTA-ACTIVADO

CANALES SELECTIVOS DE MALATO

FUNCION DEL MALATO
El papel de malato en la fisiología de las células vegetales es múltiple: -Almacenamiento de CO2. - Osmolito implicada de la turgencia celular.- Regulación del pH citoplásmico .
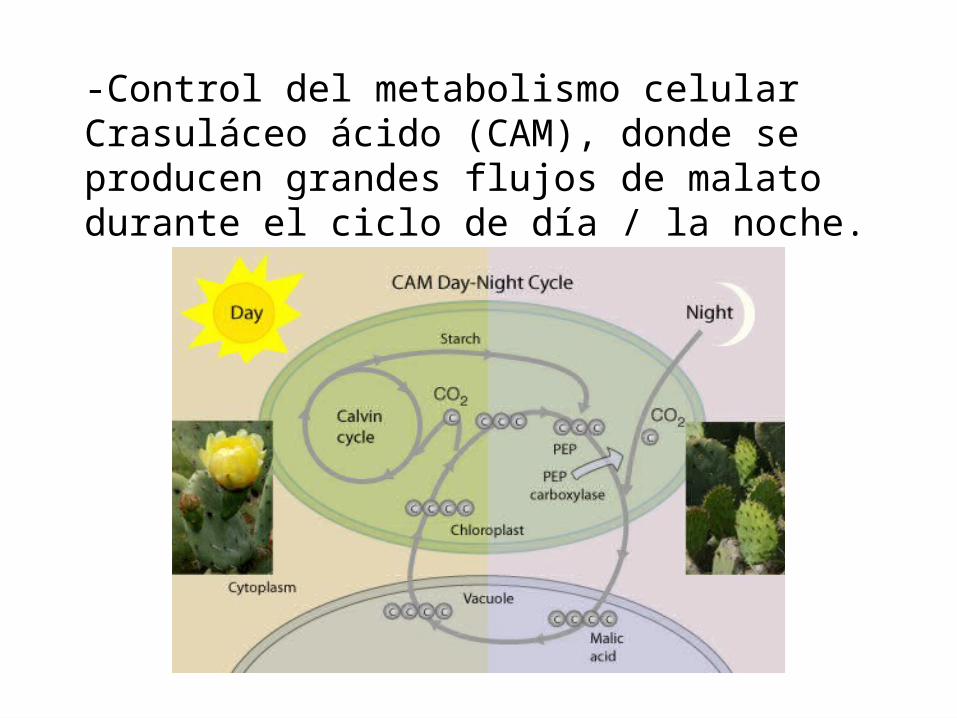
-Control del metabolismo celular Crasuláceo ácido (CAM), donde se producen grandes flujos de malato durante el ciclo de día / la noche.

TRANSPORTE DE MALATO
• Uso de Ca2+ C por debajo de 10-7 M para eliminar la actividad del canal SV .• Empleo de malato de potasio como electrolito para
la activación de canales por voltaje.• En ambos casos pH era 7,5, lo que indica que la
forma divalente del anión era la especie que se movía a través de canales.• La inhibición por la acidificación citoplasmática
dentro del intervalo fisiológico de pH 7,0 a 6,5 se observó en K. daigremontiana.

CONCLUSION
• Se demostró la presencia de varios tipos de canales iónicos en el tonoplasto, Este canal permite la acumulación de ácidos di y tricarboxilicos (malato, fumarato, citrato, etc.) dentro de la vacuola de células vegetales y juega un papel central en la regulación osmótica y de pH celular.

TONOPLAST PROTEÍNA INTRÍNSECA
• oocitos inyectados con γ TIP ARNm mostraron hinchazón rápida y estallan dentro de 6 minutos después de la exposición a soluciones hipo osmóticas. Oocitos no inyectados o con inyección de agua se hincharon muy lentamente sobre el mismo tratamiento.

TONOPLAST PROTEÍNA INTRÍNSECA
• γ TIP causó una de seis a ocho veces aumento en la permeabilidad al agua de los ovocitos, similar a los resultados obtenidos con CHIP28, bien caracterizados una canal de agua en los eritrocitos humanos (90a).• γ TIP, por otro lado, no afectó a la tasa de
absorción de glicerol del ovocito • la actividad de γ TIP como un canal de malato
sólo sería registrada si este anión estaban presentes en el interior del oocito, una condición que no ha sido probado.

ACUAPORINAS

GRACIAS!!


















