Formación Reticular
30
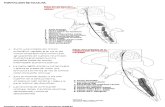
Como hemos visto anteriormente, el tegmento tronco encefálico está ocupado por núcleos específicos, por los núcleos de los nervios craneales y por los tractos nerviosos que transitan por él. Los núcleos de los nervios craneales están unidos unos a otros por un sistema de conexiones inter segmentarias la filogenia nos demuestra que, al relacionar los centros de mayor importancia, este sistema representa en todos los vertebrados, desde los más simple hasta los más evolucionados, una organización primitiva fundamenta que asegura la actividad de fondo de la totalidad del sistema nervioso central. Este sistema difuso, que recibe y envía influjos a cualquier parte, se denomina formación reticular, término que hace referencia al aspecto de malla o de red difusa con que aparece en los cortes histológicos teñidos con métodos de plata. Sin embargo, la impresión de retículo es engañosa. A pesar de su aparente desorganización, la formación reticular está altamente estructurada. De hecho, la formación reticular está constituida por grupos neuronales diferentes con funciones específicas y con neurotransmisores y conexiones particulares. La formación reticular se halla situada en una zona estratégica. Se extienden desde la médula espinal hasta el diencéfalo, en el que se continúa; ocupa, e n el tegmento o calota del tronco del encéfalo, la mayor parte del espacio que queda libre entre los núcleos de los nervios craneales y las grandes vías ascendentes y descendentes por lo que la formación reticular puede conectarse fácilmente a ellos, influenciarlos e interconectarlos. Su considerable número de sinapsis explica el carácter difuso de la actividad de la formación reticular y su importancia funcional en el sistema nervioso central. Forma
-
Upload
franciz-velasquez-cruz -
Category
Documents
-
view
21 -
download
3
description
Anatomía, vías y funciones de la formación reticular
Transcript of Formación Reticular

Como hemos visto anteriormente, el tegmento tronco encefálico está ocupado por núcleos
específicos, por los núcleos de los nervios craneales y por los tractos nerviosos que transitan
por él. Los núcleos de los nervios craneales están unidos unos a otros por un sistema de
conexiones inter segmentarias la filogenia nos demuestra que, al relacionar los centros de
mayor importancia, este sistema representa en todos los vertebrados, desde los más simple
hasta los más evolucionados, una organización primitiva fundamenta que asegura la actividad
de fondo de la totalidad del sistema nervioso central. Este sistema difuso, que recibe y envía
influjos a cualquier parte, se denomina formación reticular, término que hace referencia al
aspecto de malla o de red difusa con que aparece en los cortes histológicos teñidos con
métodos de plata. Sin embargo, la impresión de retículo es engañosa. A pesar de su aparente
desorganización, la formación reticular está altamente estructurada. De hecho, la formación
reticular está constituida por grupos neuronales diferentes con funciones específicas y con
neurotransmisores y conexiones particulares.
La formación reticular se halla situada en una zona estratégica. Se extienden desde la médula
espinal hasta el diencéfalo, en el que se continúa; ocupa, en el tegmento o calota del tronco del
encéfalo, la mayor parte del espacio que queda libre entre los núcleos de los nervios craneales
y las grandes vías ascendentes y descendentes por lo que la formación reticular puede
conectarse fácilmente a ellos, influenciarlos e interconectarlos.
Su considerable número de sinapsis explica el carácter difuso de la actividad de la formación
reticular y su importancia funcional en el sistema nervioso central.
En efecto, desempeña un papel particular, “no específico”. No transmite mensajes particulares,
ya sean sensitivos, motores o vegetativos. Recibe numerosas informaciones, las agrupa, las
asocia en una información general difusa que proporciona al sistema nervioso central lo que
podríamos denominar un estado de fondo, gracias al cual se ejercen sin dificultad las
actividades más precisas atribuidas a las estructuras segmentarias específicas o supra
segmentarias de recepción y de gobierno superior.
La formación reticular está implicada en diversas funciones y, en conjunto, debe considerarse el
centro del tronco encefálico regulador de las actividades motora, sensitiva, vegetativa y
niveles de conciencia, sobre cuyo fondo se realizan la mayor parte las funciones del SNC
Form
Característic

(O. 21-1) La estructura es el criterio clave para considerar que una zona específica del tronco
del encéfalo pertenece a la formación reticular. La cito arquitectura es el fundamento para
establecer su organización morfológica y funcional. El diseño general de las neuronas de la
formación reticular responde a la necesidad de difundir las señales nerviosas en todos los
sentidos y de crear gran convergencia y divergencia. Los rasgos básicos de las neuronas
reticulares son:
Tienen tamaños diferentes, pudiendo ser grandes, medianas y pequeñas. Cada uno de
estos tipos tiene a concentrarse, como veremos más adelante en ciertas zonas del tronco
del encéfalo.
Las neuronas de asociación típicas, del tipo Golgi II, con axón corto y profusamente
ramificado cerca del soma neuronal, son escasas.
Los axones suministran numerosas colaterales a lo largo de su trayecto, por lo que se
produce una gran dispersión del impulso nervioso.
Por lo general, las dendritas son largas y radiantes. Es decir, son neuronas del tipo
isodendrítico1, aunque también hay algunas neuronas con dendritas cortas y muy
ramificadas de tipo idiodentrítico2.
Gran parte de los árboles dendríticos se sitúan en planos perpendiculares al eje mayor del
tronco del encéfalo. Así, se disponen cráneo caudalmente, como una pila de monedad,
configurando cierta segmentación.
Por lo general, las ramas de bifurcación siguen trayectos muy largos
Bajo el punto de vista del neurotransmisor utilizado, las neuronas de la formación reticular
son heterogéneas.
(O 21-2) La formación reticular puede dividirse en tres bandas: medial, intermedia y lateral,
según las características de sus neuronas, sus conexiones y su función. Las bandas se sitúan
en cada mitad del tronco encefálico, de medial a lateral. A su vez, en cada banda pueden
distinguirse una serie de agrupaciones neuronales denominadas núcleos reticulares, que
difieren claramente del resto de los núcleos situados en el tronco del encéfalo. En su mayoría
no pueden observarse fácilmente con los métodos convencionales.
1 Isodendríticas.- Con las dendritas rectilíneas que se ramifican de modo que las ramas hijas son más largas que las madres. Características típicas de las dendritas.2 Idiodendríticas.- Con las dendritas organizadas dependiendo del tipo neuronal. Neuronas piramidales de la corteza cerebral: Soma triangular. Las dendritas en la porción final se ramifican profusamente (en forma de abanico)/ Células de Purkinje del Cerebelo: Somas piriforme (forma de pera)
Organizació

La banda medial se localiza inmediatamente por fuera de la línea media del tronco del
encéfalo. L a línea media recibe el nombre de rafe dado que es una zona de
cruzamiento de fibras; de ahí que los núcleos reticulares situados en esta banda se
denominen núcleos del rafe. La mayor parte de estos núcleos utilizan la serotonina
como neurotransmisor.
La banda intermedia o paramedial se sitúa por fuera de la banda media. Está formada
por neuronas medianas y de gran tamaño, por lo que algunos de sus núcleos se
denominan giganto-celulares. Se extiende fundamentalmente por el bulbo y el puente,
aunque también ocupa la parte baja del mesencéfalo. Las neuronas de esta banda
constituyen el origen de la mayor parte de las proyecciones ascendentes y
descendentes de la formación reticular. Regulan los movimientos voluntarios y
posturales, la transmisión del dolor, las funciones vegetativas y el control de la
conciencia.
La banda lateral es muy prominente en el bulbo y en la porción baja del puente. Está
formada por neuronas pequeñas, por lo que algunos de sus núcleos se denominan
reticulares o reticulares. Unos núcleos son adrenérgicos y otros acetilcolinérgicos. Las
neuronas de esta banda intervienen fundamentalmente en la regulación de las funciones
viscerales relacionadas con el nervio vago y en el control de la actividad motora visceral.
NUCLEOS RETICULARES (R 177)
Se presentan como aglomerados celulares que se diferencian por su citología, sus conexiones
y su topografía; los núcleos reticulares de la médula oblongada prolongan la formación reticular
de la médula espinal; a su vez continúan mediante los núcleos reticulares del puente, a los que
siguen los núcleos reticulares mesencefálicos, que se extienden hasta la formación reticular del
diencéfalo.
En este conjunto difuso se reconoce una organización principal: los núcleos reticulares
centrales. Están comprendidos entre los núcleos reticulares laterales y los núcleos reticulares
medianos y para-medianos.
Núcleos Reticulares Centrales
Ocupan la mayor parte del tegmento del tronco encefálico; de ahí el nombre de núcleos
tegmentales que se da a algunos. Están atravesados por el tracto tegmental central.
Su masa está fragmentada en núcleos secundarios (núcleo reticular central en la médula
oblongada inferior, núcleo giganto nuclear en la M.O. superior, núcleos reticulares
Núcleos

caudal y oral del puente, núcleos reticulares en los pedúnculos cerebrales), pero cada
uno de ellos puede ser dividido en una porción lateral y una porción mediana, que son
funcionalmente diferentes,
La porción lateral, denominada sensorial por Brodal, presenta numerosas dendritas
transversales que la ponen en relación con las colaterales sensitivas del tracto espino-
talámico, del lemnisco medial, del núcleo mesencefálico del nervio trigémino, del tracto
solitario y de los núcleos cocleares y vestibulares; en suma, con todo el conjunto
sensitivo de la médula espinal y del tronco del encéfalo (R 178)
La porción mediana es la más voluminosa y está caracterizada por sus grandes
células. Se denomina porción motora o efectora; está comunicada con la parte
sensorial y se encuentra en el origen de los impulsos ascendentes y descendentes que
siguen las vías longitudinales.
Las corrientes ascendentes se dirigen hacia la parte alta del mesencéfalo, la formación
reticular talámico y el núcleo centro-mediano del tálamo, alcanzando, por medio de estas
formaciones, el hipotálamo y la corteza cerebral. Los impulsos de estas neuronas siguen
bien una vía larga, el tracto tegmental central, que forman en parte.
Este sistema ascendente, a pesar de la naturaleza de sus aferencias sensitivas y
sensoriales, no es, como ya se ha indicado, un conductor de impulsos específicos
sensitivos, sino un sistema inespecífico, transmisor de impulsos difusos susceptibles
bien de determinar reflejos a nivel del tronco del encéfalo, en razón de sus relaciones
con los núcleos de los nervios craneales vecinos, bien de activar los centros superiores
talámico, hipotalámicos y corticales, poniéndolos en estado de alerta o de vigilancia.
Los impulsos descendentes se encuentran en el origen de los dos tractos retículo-
espinal.
De los núcleos centrales de la M.O. parte el tracto bulbo-retículo-espinal, que ejerce una
cierta inhibición sobre los núcleos del asta anterior de la médula espinal, razón por la
cual se denomina inhibidor.
A partir de los núcleos pontinos y mesencefálicos se destaca el tracto ponto-retículo-
espinal, que facilita la acción de los núcleos del asta anterior de la médula espinal, por lo
que se denomina fascículo facilitador.
Estos núcleos centrales efectores están sometidos a la influencia de la corteza cerebral
(áreas motoras y cingulares) y de los núcleos estriados de los que reciben impulsos
descendentes por la vía de las fibras cortico-reticulares y estrío-reticulares (R 180)

Así, al ponerse en marcha el estado de vigilancia de la corteza cerebral por la formación
reticular ascendente, la corteza responde a su vez con la puesta en estado de vigilancia
de los centros reticulares efectores.
Núcleos Reticulares Lateral y Paramediano
Estos núcleos rodean, en la M.O. a los núcleos centrales; el núcleo reticular lateral se
extiende posterior a la oliva, mientras que el núcleo reticular lateral se extiende posterior
a la oliva, mientras que el núcleo reticular Paramediano se encuentra media al núcleo
reticular central (R 181)
Aseguran la mayor parte de la relaciones de la formación reticular con el cerebelo,
constituyendo un circuito corto retículo-cerebelo-reticular. Se ponen en acción a través
de las colaterales bulbares de los tractos espino-talámico y envían sus fibras eferentes
hacia el conjunto de la corteza cerebelosa, que los refleja hacia el núcleo reticular
Paramediano y, de ahí de nuevo hacia el cerebelo.
Destaquemos que, junto a este circuito retículo-cerebeloso corto, existe un circuito más
largo en el que están implicados el tracto tegmental central y la oliva, que se mantiene, a
su vez, por la actividad de los núcleos reticulares centrales.
Núcleos Reticulares Medianos
Están situados cerca de la línea media; son los núcleos del rafe de la médula oblongada
y del puente de la sustancia gris central alrededor del acueducto del mesencéfalo y,
además, del núcleo tegmental posterior
Estos núcleos reciben sus aferencias del hipotálamo y del rinencéfalo por medio del
fascículo telencefálico medial y están unidos a los núcleos vegetativos del tronco del
encéfalo. Desempeñan un papel importante en los mecanismos vegetativos del tronco
del encéfalo (R 182)
Así concluimos que la formación reticular del tronco del encéfalo asegura:
La coordinación de los núcleos de los nervios craneales por medio de sus colaterales de
vecindad
La vigilancia de los centros superiores; en respuesta a esta vigilancia, se ejerce un
control inhibidor o facilitador sobre los centros suprayacentes (N.R.C)
Las relaciones y el control del cerebelo (N.R.L y PM)
La unión entre los centros hipotalámicos o rinencefálicos y el tronco del encéfalo (N del
rafe y NR mesencefálicos)

NÚCLEO ROJO
Se sitúa en el tegmento del mesencéfalo (R 184). Al igual que el paleocerebelo se enriquece
con un neocereblo y el paleoencéfalo con un neoencéfalo, del mismo modo el núcleo rojo está
compuesto por dos porciones:
Porción magno celular o paleorrubro. Es reducida en el hombre.
Porción parvocelular o neorrubro.- es la más importante. Da origen al tracto
rubroespinal. Este tracto, después de entrecruzarse desciende hasta la porción cervical
de la médula espinal y termina en los centros motores periféricos que encuentra en su
recorrido.
La porción parvocelular será un centro de relevo para las vías reflejas cerebelosas y
estriadas que constituyen la parte esencial de las vías extrapiramidales corticales.
Enviará sus fibras a la formación reticular que la rodea y de la cual forma parte funcional.
Por todo esto, interviene en la sinergia de los diferentes movimientos del cuerpo, los
reflejos laberínticos, las actitudes y las posturas, fenómenos todos ellos autonómicos o
semiautomáticos.
SUSTANCIA NEGRA
Representa topográficamente el elemento nervioso que separa el tegmento del mesencéfalo de
a base del pedúnculo cerebral. Desde un punto de vista funcional es tan importante como el
núcleo rojo. Situada en el trayecto de las vías extrapiramidales corticales y subcorticales, sus
neuronas se comunican mediante sus fibras con las de la formación reticular. Este núcleo
desempeña el mismo papel y produce la misma sintomatología que el globo pálido: es un centro
de los movientes asociados que acompañan al movimiento voluntario.
COMPLEJO OLIVAR INFERIOR O NÚCLEOS OLIVARES INFERIORES
Recibe la mayor parte de sus aferencias de las formaciones reticulares del tronco del encéfalo
a través del tracto tegmental central. El núcleo olivar principal y los núcleos olivares accesorios
medial y posterior que lo acompañan proyectan sus fibras eferentes sobre el neo cerebelo, que
las reenvía al núcleo rojo y a los núcleos estriados y, a través de éstos, a las formaciones
reticulares; así se constituye un gran circuito de retorno extra piramidal en el que el complejo
olivar inferior es una pieza clave. Puede añadirse que este complejo se encuentra unido a la
médula espinal por fibras ascendentes y descendentes que reciben el nombre de fibras olivo
espinales y tracto espino olivar (R 186 y 190)

Aferencias
La formación reticular recibe aferencias de prácticamente todos los sistemas, tanto sensitivos
como motores. Ya hemos señalado que, al situarse en el tegmento tronco encefálico, puede
recibir fácilmente colaterales de los axones que transitan por esta zona de paso. Además, la
formación reticular recibe proyecciones procedentes de áreas extensas de la corteza cerebral,
en especial de la corteza cerebral, en especial de la corteza somato-sensitiva, los núcleos
basales, el núcleo rojo y la sustancia negra. Gran parte de estas proyecciones utilizan el tracto
tegmental central para alcanzar la formación reticular. Un importante contingente de
aferencias procede d neuronas asentadas en los núcleos de los nervios craneales, sobre todo
del núcleo sensitivo del trigémino. Estas aferencias tienen gran importancia funcional. No sólo
permiten establecer una serie de reflejos sino que, a través de los nervios craneales, hacen
posible la estimulación de toda la formación reticular, lo cual tiene importancia clínica.
Aunque los diferentes tipos de aferencias terminan preferentemente en ciertas zonas de la
formación reticular, no existen conexiones específicas. Por lo general una multitud de fibras
aferentes converge en una sola neurona produciendo respuestas no específicas.
Eferencias
Las eferencias de la formación reticular son a menudo difíciles de precisar dado que, en
muchos casos, se trata de tractos poli sinápticos formados tanto por fibras directas como
cruzadas.
Las proyecciones descendentes hacia la medula espinal están representadas por los tractos
retículo-espinales, de los cuales uno es medial y otro lateral.
Tracto retículo medial (O 21-3) Se origina en neuronas grandes ubicadas en la banda
intermedia a nivel del puente. Los axones son directos y descienden por fuera del tracto
cortico-espinal ventral y por detrás del tracto vestíbulo-espinal medial. Terminan
principalmente en la lámina VIII y en la porción adyacente de la lámina VII de Rexed
Tracto retículo-espinal lateral (O 21-4) Está formado por cilindroejes directos y cruzados
procedentes de neuronas localizadas en la banda intermedia a nivel bulbar. Descienden por
el cordón lateral de la médula espinal, situándose inmediatamente por delante del tracto
fascículo lateral. Este tracto termina más dorsalmente que el retículo-espinal medial, yq
que lo hace principalmente sobre la lámina VII y, en menor medida, sobre las láminas VIII y
IX. Es una vía glicinérgica y, por lo tanto, inhibidora.
Fibras

Los tractos retículo-espinales actúan, por medio de interneuronas, tanto sobre
motoneuronas α y γ como sobre neuronas sensitivas del asta posterior, Como veremos
más adelante los tractos retículo-espinales difieren funcionalmente.
Las proyecciones ascendentes de la formación reticular son muy numerosas y terminan
en el tálamo, especialmente en los núcleos intra-laminares, en el hipotálamo, en los
núcleos basales y, de manera difusa, prácticamente en toda la corteza cerebral. La
actividad de estas proyecciones es fundamental para el mantenimiento de la conciencia y
constituye el sustrato morfológico de lo que se ha denominado sistema de activación
reticular ascendente (SARA). Su activación produce el encendido de la corteza cerebral y,
por consiguiente, el estado de vigilia. Este término se emplea en sentido funcional y no
morfológico. Gran parte de las proyecciones ascendentes pertenecen a los sistemas mono-
aminonérgico y colinérgico.
CONEXIONES CON LA MÉDULA ESPINAL (B 7.9)
Las vías retículo-espinales se encuentran tanto en los cordones anteriores como en los
laterales. Brodal han señalado su rigen en el bulbo y en el puente, sobre todo en los núcleo
gigantonuclear, caudal del puente y región inferior del núcleo oral. Las fibras originadas en el
puente descienden por la parte media del cordón superior de la médula, por el mismo lado de
su origen, mientras que las bulbares ocupan los cordones laterales y son tanto directas como
cruzadas.
Las fibras espino reticulares tienen su origen en células de distintos niveles medulares y
terminan difusamente en todos los núcleos reticulares. Además, ellos reciben colaterales de
otros fascículos ascendentes, en especial de los espinotalámicos
CONEXIONES CON EL TRONCO ENCEFÁLICO
La organización de la formación reticular sugiere que una solo neurona reticular puede
transmitir impulsos en dirección caudal y rostral, puede ejercer sus influencias localmente y a
distancia y los impulsos pueden ser conducidos con rapidez (axón primario) y con lentitud
(mediante sinapsis colaterales)
Bulbo Raquídeo
La formación reticular del tronco del encéfalo comienza en el bulbo. Los núcleos reticulares
presentes en este nivel son el núcleo reticular lateral del bulbo y el núcleo reticular ventral
Conexiones

(C 5.6). En los niveles meso olivares, el núcleo reticular giganto celular ocupa el área dorsal
y medial al complejo olivar inferior. (C 5.12). Como lo implica su nombre, tiene grandes
células características, pero también posee muchas células de medianas dimensiones y
pequeñas. Este núcleo ocupa los dos tercios mediales de la formación reticular y se
extiende hasta la unión bulbo-protuberancial.
El núcleo reticular parvo-celular es una agrupación de pequeñas células localizadas
postero-lateralmente, mediales con respecto al núcleo trigémino-espinal y anterior en
relación con los núcleos vestibulares. Este núcleo, que ocupa aproximadamente el tercio
lateral de la formación reticular bulbar, ha sido mencionado como la parte “sensitiva” porque
recibe colaterales de vías sensitivas secundarias.
Los núcleos reticulares paramedianos consisten en varios grupos nucleares pequeños que
se encuentran próximos a las fibras del fascículo longitudinal medial y el lemnisco medial, y
a veces, están situados entre ellas. Estas neuronas reticulares proyectan la mayoría de sus
fibras hacia el vermis cerebeloso.
En esencia, la formación reticular del bulbo consiste en tres masaas nucleares principales:
un grupo nuclear reticular paremediano, un grupo central (los núcleos reticular ventral y
reticular GC) y un grupo nuclear lateral que consta de los núcleos reticular lateral y reticular
parvo celular.
Las fibras aferentes que se proyectan a la formación reticulat del bulbo se originan enn
lamédula espinal en los núcleos sensitivos secundarios de los nervis craneales, y en la
corteza cerebral y en los núcleos cerebelosos profundos. Las fibras espino reticulares
ascienden en el cordón antero lateral y terminanen las porciones caudal y lateral de la
formación reticular del bulbo. Un número considerable de estas fibras terminan en el
núcleos reticular GC, aunque algunas se proyectan havia partes más rstrales de la
formación reticular. Las fibras colaterales de las neuronas secundarias sensoriales de los
núcleos de los ervios craneales ( fibras auditivas, vestibulares, trigeminales y viscerales) se
proyectan el núcleo reticular parvo celular.
Las fibras cerebelo reticulares terminan principalmente en los núcleos reticulares par
medianos. Estas proyecciones reticulares se originan en los núcleos del techo y dentado.
Los núcleos reticulares para medianos forman parte de un sistema de retroalimentación, ya
que proyectan y reciben fibras del cerebelo.
Las fibras cortico reticulares se originan en amplias áreas de la corteza, pero el mayor
número de estas fibras proviene de pareas sensorio motoras. La mayor parte de estas
fibras terminan en porciones de la formación reticular de la protuberancia y el bulbo. Son

cruzadas y directas. Las regiones de la formación reticular del tronco del encéfalo que
reciben fibras cortico reticulares dan origen a fibras retículo espinal. En los núcleos
reticulares convergen impulsos de una amplia variedad de procedencias. La máxima
superposición de fibras en la formación reticular bulbar, que da origen a un gran número de
axones largos ascendentes y descendentes.
Las fibras eferentes proviene de los dos tercios mediales de la FR del bulbo, que
corresponde al núcleo reticular GC; esta masa nuclear da rigen a fibras largas ascendentes
y descendentes. Las fibras ascendentes de esta masa nuclear ascienden en el haz central
de la calota, son directas y termina principalmente en porciones de los núcleos intra
laminares talámicos. Este sistema desempeña una importante función en el mecanismo del
despertar del sueño. Las fibras descendentes de esta región medial de la FR dan origen a
los haces retículo espinales que son bilaterales pero de predominio directo (C 4.14). Los
axones de las neuronas reticulares del bulbo forman proyecciones retículo espinales
descendentes largas que proporcionan colaterales a todos los niveles espinales y
proyecciones más cortas sólo a los segmentos espinales cervicales. La estimulación de la
parte rostral y dorsal del núcleo reticular GC y regiones de la FR caudal de la protuberancia
producen potenciales post sinápticos excitadores mono-sinápticos en las neuronas motoras
que inervan los músculos axiales del cuello y la espalda. La estimulación de la FR caudal y
medial del bulbo produce potenciales post sinápticos en las neuronas motoras de todos los
niveles. De esta manera, la FR del bulbo se puede dividir en una región facilitadora dorso
rostral y una zona caudal inhibidora. Las neuronas de esta área “efectora” de la formación
reticular del bulbo reciben proyecciones de células del núcleo reticular PC. Se considera
que esta última región reticular tiene fx de recepción o asociación, mientras que la región
reticular más medial, de mayor tamaño, está organizada para ejercer influencias efectoras a
nivel medular y a niveles superiores del tronco del encéfalo.
Núcleos del rafe (C 5.13)
Las fibras que se originan en las células de los núcleo del rafe tienen un amplia distribución
en el neuroeje y liberan serotonina en sus terminaciones. En el bulbo, las neuronas
serotoninérgicas se encuentran en los núcleos magno oscuro y pálido del rafe, que dan
origen a proyecciones espinales descendentes. Las células del núcleo magno del rafe dan
origen a proyecciones espinales bilaterales que descienden en el cordón dorso-lateral y
terminan en su mayoría en las láminas I y II. La estimulación del núcleo magno del rafe
produce un efecto analgésico mediante una acción inhibidora sobre las neuronas de las
láminas I y II en las partes caudales del núcleo trigémino espinal y en el asta espina dorsal.

Los NT que intervienen en la modulación del dolor son probablemente múltiples y parecen
actuar sobre interneuronas nociceptivas y neuronas que dan origen a vías del dolor
(neuronas del haz espino-talámico).
Puente
Dos masas nucleares, el núcleo reticular protuberancial caudal y el oral, componen el
volumen de la formación reticular protuberancial. La parte caudal, que sustituye al NR GC
del bulbo, se extiende en dirección rostral hasta el nivel del núcleo motor del trigémino. La
parte oral se extiende en dirección rostral al mesencéfalo caudal, donde carece de límites
definidos. Las células de la FRP dan origen a fibras retículo-espinal directo, las cuales
descienden en el tronco encefálico como componente del fascículo longitudinal medial.
Otras células dan origen a fibras que ascienden como parte del haz central de la calota.
Muchas células poseen axones que se bifurcan en ramas que se proyectan en dirección
rostral y caudal. Las fibras ascendentes de la formación reticular protuberancial se
proyectan a través del fascículo central de la calota hacia los núcleos talámicos intra-
laminares. Los impulsos que pasan a estos núcleos talámicos influyen de manera
acentuada sobre la actividad eléctrica de amplias áreas de la corteza cerebral.
Otros núcleos reticulares presentes en los niveles pontinos son el retículo-tegmentario y el
central superior. El núcleo retículo-tegmentario se encuentra próximo al rafe medio dorsal
en relación con el lemnisco medial y se lo considera un grupo nuclear protuberancial
desplazado hacia la calota. El núcleo central superior se desenvuelve en el rafe medio, se
agranda a nivel del istmo, y se le conoce como núcleo medio del rafe.
Núcleos del rafe
La protuberancia y el bulbo contienen grupos celulares en el rafe medio que pertenecen a
la formación reticular. El núcleo central inferior parece en el rafe medio, en la unión de la
protuberancia con el bulbo y representa la parte rostral del núcleo magno del rafe. El núcleo
protuberancial del rafe consiste en varios grupos celulares de localización dorsal y rostral
con respecto al núcleo central inferior. La extensión rostral de los núcleos protuberanciales
del rafe es el núcleo central superior, también conocido como núcleo medio del rafe (C
6.27). Las fibras cruzadas del pedúnculo cerebeloso superior atraviesan porciones del
núcleo central superior. Los núcleos dorsales del rafe son pares y se encuentran a cada
lado del rafe, en la sustancia gris pero acueductal anterior, en situación dorsal con respecto
a los fascículos longitudinales mediales.

Las principales fibras ascendentes se originan en cuerpos celulares que contienen
serotonina, localizados en los núcleos dorsal del rafe y central superior. La principal vía
ascendente de los núcleos rostrales del rafe pasa a través del área tegmentaria ventral y se
une con el fascículo prosencefálico medial en el hipotálamo lateral. Las fibras que salen de
este haz ascendente principal ingresan a la sustancia negra, los núcleos talámicos intra-
laminares, la estría terminal, el septum y la cápsula interna. Las proyecciones más rostrales
terminan principalmente en el lóbulo frontal, aunque algunas fibras se distribuyen por toda
la neo-corteza. El núcleo dorsal del rafe inerva de manera selectiva la sustancia negra, el
cuerpo geniculado lateral, el neo-estriado, el lóbulo piriforme, el bulbo olfatorio y el complejo
nuclear amigdalino. El núcleo central superior está asociado en particular con fibras
serotoninérgicas que se proyectan al núcleo inter-peduncular, los cuerpos mamilares y la
formación del hipocampo. Las proyecciones ascendentes de los núcleos caudales del rafe
son menos numerosas y se distribuyen en el tubérculo cuadragémino superior, el pretectum
y los núcleos de la comisura posterior. Las vías serotoninérgicas ascendentes del núcleo
central superior se proyectan principalmente a las estructuras mesolímbicas, como el
hipocampo y los núcleos septales, mientras que el núcleo dorsal del rafe envía sus
proyecciones importantes hacia el neoestriado y la sustancia negra. Las proyecciones
ascendentes de las células de los núcleos del rafe son principalmente ipsilaterales.
El núcleo central superior da origen a proyecciones descendnetes hacia el cerebelo a
través del pedcúnculo cerebelo medio, el locus ceruleus y grandes rehiones de la FRP. Las
eestructuras que reciben eferentes del núcleo magno del rafe comprenden los núcleos
motor dorsal del vago, solitario y trigémino-espinal. La serotonina y los núcleos del rafe ha
sido implicados en la regulación del sueño, el humor, la conducta agresiva y diversas
funciones neuro-endocrinas. Cuando dormimos, las neuronas del rafe descargan de
manera lenta, y dejan de descargar durante e sueño de movimiento oculares rápidos.
Se ha considerado que las neuronas que contienen serotonina y las que contienen
noradrenalina en la FR desempeñan funciones activas en los mecanismos que controlan
los estados de sueño. La inhibición de la síntesis de serotonina o la destrucción de las
neuronas que contienen serotonina en el sistema del rafe que conduce al insomnio, La
serótina puede estar implicada en el mecanismo neural relacionado con el denominado
sueño lento, un estado caracterizado por un electroencefalograma de ondas lentas y picos.
Además, la serotonina puede tener efectos sobre células del locus ceruleus que
desencadenan el llamado sueño paradójico. El sueño paradójico se caracteriza por: la
abolición del tono muscular antigravitatorio. La disminución de la presión arterial,

bradicardia y respiración irregular. Accesos de movimientos oculares rápidos y EEG
semejante al del estado de vigilia. Las lesiones bilaterales del locus ceruleus en los
animales causan una supresión selectiva del sueño paradójico durante alrededor de dos
semanas, luego de las cuales retornan a un estado próximo a la normalidad.
Mesencéfalo
La FR del mesencéfalo es menos extensa que la de la protuberancia. Aunque el núcleo rjo
forma parte de la FR. Se reconocen tres núcleos reticulares principales: cuneiforme,
subcuneiforme, y núcleo protuberancial de la calota.
El núcleo pedúnculo protuberancial se encuentra en la parte lateral de la calota, en relación
ventral con respecto el tubérculo cuadragémino inferior. Las fibras del pedúnculo
cerebeloso superior atraviesan este núcleo en su trayecto en dirección ventro medial hacia
su decusación. El núcleo pedúnculo protuberancial recibe impulsos de múltiples
procedencias que comprenden: la corteza cerebral, el segmento medial del pálido y la parte
reticular de la sustancia negra. Las células de la parte compacta del núcleo pedúnculo
protuberancial son fuertemente colinérgicas, pero las células adyacentes tienen otros NT.
Este núcleo se encuentra en una región que al ser estimulada da lugar a movimientos de
archa. Esta región se denomina “centro locomotor”.
Los núcleos cuneiforme y subcuneiforme se encuentran en situación ventral con respecto a
la región tectal y dorsal en relación al núcleo pedúnculo protuberancial, y se extienden en
dirección rostral. Las fibras del haz central de la calota se encuentran en localización medial
con respecto a estos núcleos reticulares. El núcleo interpeduncular, compuesto por células
pequeñas, se encuentra en la línea media, dorsal a la fosa inter peduncular. Este núcleo
recibe fibras de los núcleos de la habénula a través del fascículo a través del fascículo
retroflexo.
SISTEMA RETICULAR ASCENDENTE
Se conoce con este nombre a un conjunto de fibras que ascienden desde distintos núcleos
reticulares del tallo cerebral hasta el diencéfalo y la corteza. En el tallo cerebral forman parte
del fascículo tegmental central; a nivel del diencéfalo, unas terminan en los núcleos
intralaminares y otras en distintos núcleos del hipotálamo; otras ascienden desde el área
hipotalámica lateral hasta la corteza. El sistema reticular ascendente ejerce su acción sobre
amplias áreas de la corteza y se manifiesta tanto en los estados de conciencia como en la
actividad eléctrica cortical.

CONEXIONES CON EL CEREBELO
Son tanto aferentes como eferentes. Según Brodal, las fibras retículo cerebelosas se originan,
en especial, en los núcleos lateral y Paramediano del bulbo y del núcleo tegmento póntico de
la protuberancia. A su vez, las fibras cerebelo-reticulares provenientes de los núcleos
centrales, y principalmente del núcleo fastigial, se distribuyen ampliamente en los distintos
núcleos reticulares.
Tracto retículo-cerebeloso.- se origina en núcleos de la formación reticular bulbar y póntica.
Está formado por fibras directas y cruzadas que penetran en el cerebelo por el pedúnculo
cerebeloso inferior. Aporta información compleja, tan cutánea del tronco y de las extremidades
como procedente de la corteza cerebral y de otros puntos del SNC. Alcanza el espino-
cerebelo y también envía aferencias al cerebro cerebelo.
CONEXIONES CORTICO RETICULARES
Se originan en distintas áreas de la corteza, pero principalmente en las regiones centrales
sensitivas y motoras. Se distribuyen bilateralmente y terminan sobre todo en los núcleos del
bulbo y del puente. Especial mención merecen las conexiones que recibe la corteza entorrinal,
el hipocampo y la región septal. Algunas de estas fibras llegan al tallo cerebral por la porción
ventral del diencéfalo (área hipotalámica lateral), mientras que otras forman parte de la estría
medular del tálamo y del fascículo retroreflexo.
OTRAS CONEXIONES DE LOS NÚCLEOS RETICULARES
La formación reticular tiene además conexiones con los sistemas sensoriales, como en el
visual-a través del colículo superior, con los núcleos cocleares y vestibulares, con los núcleos
del sistema extrapiramidal y con las vías propio y exteroreceptivas.
Ya hemos señalado que la formación reticular interviene en la regulación y control de diferentes
funciones, las más importantes de las cuales son las funciones motoras, formación reticular y
movimientos oculares, funciones visuales, funciones sensitivas y conciencia.
Formación Reticular: Vigilia Y Sueño
Funciones

Hemos visto que ciertas formaciones reticulares del tronco encefálico constituyen el
centro activador del sistema nervioso central, manteniéndolo en estado de vigilancia y
vigilia.
La vigilia se caracteriza porque el electroencefalograma (EEG) está desincronizado; es
decir, muestra un patrón de actividad eléctrica rápida (mayor de 20 Hz) y de bajo voltaje.
Además, el individuo es capaz de captar información del medio ambiente y de responder
de forma adecuada a dicha información. Durante el sueño, el EEG muestra un trazado
sincrónico que se caracteriza por una actividad eléctrica lenta de alto voltaje. Durante el
sueño, hay períodos en que el EEF se desincroniza. En estas fases se producen
movimientos oculares rápidos. Durante los períodos de desincronización, denominados
de sueño REM, se producen las imágenes oníricas que conocemos como ensoñación.
La vigilia alterna con el sueño y los trabajos de estos últimos años han demostrado que
estas mismas formaciones reticulares son también el origen de los impulsos inhibidores
que provocan el sueño; precisemos que existen dos clases de sueño sucesivas; el
sueño lento y el sueño paradójico. Mientras que la actividad eléctrica del cerebro
disminuya en el transcurso del primero, aumenta en el curso del segundo como durante
el estado de vigilia, lo que justifica la denominación de sueño paradójica.
En el mecanismo del sueño están implicados numerosos núcleos reticulares (R 183).
Los núcleos del rafe del puente, la parte superior de los núcleos pontinos (núcleo
reticular rostral del puente o núcleo reticular oral del puente) inhiben los mecanismos del
despertar, el diencéfalo y la corteza cerebral. Estos núcleos serían el origen del
adormecimiento, del sueño lento, relativamente superficial y en el que no se sueña.
Los núcleos del rafe de la médula oblongada y los núcleos pontinos inferiores (núcleo
reticular caudal del pente) ehercen más tarde su acción, activando los núcleos motores
del globo ocular (se observan entonces bajo los parapados los movimientos del globo
ocular). Un pqueño núcleo del puente, el locus cerúleo, participaría en la producción de
los sueños que acompañan al sueño paradójico.
Em el mecanismo del sueño, una priera oleada de impulsos parte de los centros del
sueño lento, una segunda oleada parte de los centros del sueño paradójico. Cuando
estos impulsos agotan sus efectos, los estímulos sensoriales son capaces de determinar
de nuevo un efecto activador que alcanza por medio de la formación reticular
diencefálica, que la devuelva a la vida activa consciente: el despertar.
Estas formaciones no conducen impulsos específicos, es decir, mensajes sensitivos o
motores; su papel es otro: dirigen a los centros vegetativos y finalmente regulan la

actividad de los centros motores vecinos o subyacentes. Este papel múltiple y complejo
no puede ser reservado a un único engranaje, pero sí a un conjunto de ellos, cuyo papel
cnsiste en asociar y cobinar los datos procedentesde todaspartes. Aspi, el cerebeo, el
cuerpo estriado, el núcleo rojo, los núcleos del techo del mesencéfalo, los núcleos
vestibulares, la corteza cerebral y las colaterales de las grandes vías proporcionan las
informaciones a la formación reticular. Ësta las traduce en una información única para
los pisos de rececpción y ejecución.
El nivel de consciencia representa la suma de toda la actividad de la corteza cerebral y
de las interacciones dela corteza con el tálamo. La formación reticular es, por medio del
SARA, responsable de la regulación y modulación de la ritmicidad de las interacciones
entre el tálamo y la corteza cerebral y, por lo tanto, desempeña un papel decisivo en la
oscilación vigilia-sueño. La lesión del SARA, que se produce en las secciones
transversales del tronco del encéfalo por encima del colículo inferior, produce el coma
que es un estado de pérdida de la conciencia con incapacidad para responder a los
estímulos externos.
Funciones motoras
La formación reticular regula la actividad de otros sistemas motores. Además, es capaz
de originar respuestas motoras estereotipadas y diversos reflejos.
Los tractos retículo-espinales (bandas intermedia) constituyen una especia de vía
alternativa al tracto cortico-espinal. Los impulsos motores, originados en la corteza
motora como en otras zonas cerebrales, que llegan a la formación reticular, pueden
descargarse hacia las motoneuronas de la médula espinal. El tracto retículo-espinal
medial está implicado en el control de movimientos posturales, facilitando las
motoneuronas que inervan los músculos axiales y produciendo respuestas extensores
en los músculos de las extremidades inferiores. Así, su estimulación produce respuestas
que intentan mantener la postura erguida. También es capaz de provocar movimientos
estereotipados de las extremidades. La estimulación del tracto retículo-espinal lateral
provoca atonía y parece desempeñar un papel importante en el sueño REM.
Las neuronas de la banda lateral, que se extienden desde los núcleos del hipogloso y
ambiguo hasta el núcleo del facial y el núcleo motor del trigémino, participan en la
coordinación de las respuestas motoras realizadas por las neuronas de estos núcleos.
Las más importantes son los movimientos de masticación, los de la lengua y los labios

que se realizan durante la ingestión de alimentos, y las expresiones emocionales
faciales, en especial la sonrisa y el llanto.
Además de las funciones específicas en la regulación motora, la formación reticular
interviene de manera indirecta en el movimiento, ya que las actividades motoras
voluntarias sólo pueden realizarse n estado consciente. La formación reticular
desempeña un papel decisivo en el mantenimiento de la conciencia. Por último,
debemos recordar que la formación reticular y el cerebelo establecen conexiones entre
sí, participando de esta manera en el control motor cerebeloso.
Formación reticular y movimientos oculares (O 21-5)
Algunas zonas de la formación reticular son fundamentales para el control de los
movimientos oculares Básicamente existen dos tipos de movimientos oculares que
periten mantener enfocados los objetos en ambas fóveas: los movimientos de
convergencia y los movimientos conjugados de los ojos. En los movimientos de
convergencia los ojos se aproximan a la línea media, de tal manera que sus ejes
convergen. Este movimiento tiene lugar cuando observamos objetos próximos (durante
la lectura). En los movimientos conjugados, los ojos se desplazan en la misma dirección
y sus ejes son paralelos. El desplazamiento puede realizarse horizontalmente o
verticalmente. En unos casos se realizan lentamente: son los movimientos de rastreo o
seguimiento, destinados a mantener enfocado un objeto móvil sobre las fóveas.
Los movimientos horizontales de los ojos son realizados por los músculos rectos medial
y lateral (inervados respectivamente por los nervios oculomotor y abducen). En los
movimientos verticales participan los músculos recto superior e inferior (inervados por el
nervio oculomotor) y los músculos oblicuos superior e inferior (inervados
respectivamente por los nervios troclear y oculomotor). Son elevadores el recto superior
y el oblicuo inferior, y descensores el recto inferior y el oblicuo superior,
Los movimientos conjugados de los ojos están controlados por neuronas de la formación
reticular, agrupadas en dos “centros” denominados de la “mirada horizontal” y de la
“mirada vertical”. Cada uno de ellos es capaz de generar movimientos oculares a lo largo
de una de estas dos direcciones. El centro de la mirada horizontal se sitúa cerca del
núcleo del nervio abducens y corresponde a la banda paramedial de las porciones
pontina y bulbar alta de la formación reticular. El centro de la mirada vertical el cual se
localiza en la formación reticular mesencefálica próxima al núcleo oculomotor. Los dos
centros de la mirada se conectan de manera similar con los núcleos de los nervios que

inervan los músculos que realizan el movimiento en cuestión. En ambos casos, los
axones de las neuronas reticulares establecen sinapsis con las neuronas del núcleo más
próximo del mismo lado, es decir, el núcleo abducens en el caso del centro de la mirada
horizontal y los núcleos del oculomotor y del troclear en el del centro de la mirada
vertical. Todos estos núcleos tienen motoneuronas cuyos axones inerva las fibras de los
músculos del globo ocular, e interneuronas, cuyos axones, después de cruzarse,
terminan en los núcleos homólogos del otro lado. Estas conexiones son excitadoras para
las motoneuronas destinadas a los músculos que realizan el movimiento e inhibidoras
para los que se oponen a éste. Además, en el caso del centro de la mirada horizontal,
parte de los axones cruzados se incorporan al fascículo longitudinal medial y ascienden
para alcanzar el núcleo oculomotor que inerva el músculo recto medial contralateral.
Todas estas conexiones explican que la estimulación unilateral del centro de la mirada
produzca un desplazamiento horizontal de los ojos hacia el mismo lado.
Los centros de la mirada reciben aferencias de la corteza cerebral y del colículo superior.
Estas conexiones son muy importantes para iniciar y dirigir con precisión los
movimientos oculares. Las aferencias corticales proceden fundamentalmente del campo
motor ocular frontal, situado en la circunvolución frontal media, que controla los
movimientos voluntarios de los ojos independientemente de los estímulos visuales. La
estimulación produce movimientos sacádicos contralaterales. Las eferencias también
provienen del campo motor ocular occipital, que controla los movimientos de los ojos
ocasionados por estímulos visuales (movimientos de seguimiento). Los campos motores
oculares también se proyectan sobre el colículo superior.
Como ya sabemos, el colículo superior está formado por seis capas celulares y, además
de las aferencias corticales, recibe aferencias visuales, acústicas y somatoestésicas. Por
otro lado, el colículo superior envía eferencias cruzadas a los centros de la mirada. En el
colículo superior se integra toda la información, de tal manera que contiene mapas
topográficos superpuestos del espacio visual, sonoro y de la superficie corporal. Así, la
estimulación e un punto concreto del colículo superior producen un movimiento ocular
conjugado con una dirección, de amplitud determinada y precisa.
La parálisis de los movimientos conjugados de los ojos es siempre supranuclear y no
se acompaña de diplopía (visión doble). Existen dos zonas clave de posibles lesiones: el
fascículo longitudinal medial y el colículo superior. Las lesiones de este último son con
frecuencia debidas a tumores del cuerpo pineal. La lesión unilateral del campo ocular
frontal ocasiona incapacidad de realizar movimientos sacádicos hacia el lado opuesto.

Un hecho característico es que la mirada conjugada horizontal se dirige hacia el lado de
la lesión. Por lo general, la incapacidad es transitoria. La lesión del colículo superior
modifica la precisión, frecuencia y velocidad de los movimientos sacádicos. Sin
embargo, esta deficiencia suele mejorar con el tiempo. Se conoce con el nombre de
Síndrome de Parinaud la parálisis de la convergencia. A menudo se debe a tumores
del cuerpo pineal que comprimen la porción del mesencéfalo donde se sitúa el centro de
estos movimientos. La lesión unilateral del fascículo longitudinal medial da lugar a la
llamada oftalmoplejía internuclear, caracterizada por la incapacidad de dirigir
voluntariamente el ojo del lado afectado hacia a línea media. Sin embargo, se mantiene
el movimiento de convergencia ocular.
Funciones viscerales
La formación reticular está involucrada en respuestas viscerales diversas y en la
regulación del sistema nervioso vegetativo. Ya hemos comentado las proyecciones
ascendentes hacia el hipotálamo, centro vegetativo por excelencia. En este contexto
tiene gran importancia las neuronas de la banda externa situadas en torno a os núcleos
de origen del nervio vago. Estas neuronas participan en la coordinación de diversos
patrones de compartimento motor estereotipado en relación con las funciones viscerales
del nervio vago. Es el caso de las respuestas digestivas representadas por la deglución
y el vómito, de actividades respiratorias (regulación del ritmo respiratorio, el estornudo, la
tos o el hipo) y de respuestas cardiovasculares (reflejos barorreceptores y control del
ritmo cardiaco)
Es frecuente designar estas neuronas como centros vitales (respiratorio, cardiaco, etc.).
Sin embargo, n pueden considerarse como tales; se trata simplemente de neuronas de
la formación reticular con funciones específicas.
Funciones sensitivas
La formación reticular regula la transmisión de la información sensitiva por los tractos
ascendentes, modulando la actividad de los relés neuronales. Ya hemos señalado que
los tractos retículo-espinales terminan por medio de interneuronas sobre las neuronas
del asta posterior. Esto parece de gran importancia de la regulación de la percepción del
dolor. En este sentido, el tracto retículo-espinal lateral desempeñaría un papel especial,
ya que desempeñaría un papel especial, ya que su estimulación produce la inhibición de
a información dolorosa.