Función gástrica
11
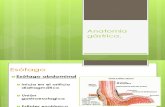
Sebastián Lavanderos B. 2do. Medicina UDP 1 FUNCIÓN GÁSTRICA El estómago tiene un papel importante en la nutrición humana, y tiene funciones secretoras, motoras y humorales. Esas actividades no están separadas ni son distintas, sino más bien representan una función integrada que se requiere como un todo para iniciar el proceso digestivo normal. El estómago tiene varios productos secretores específicos, entre los cuales encontramos, aparte del conocido ácido, al pepsinógeno, moco, bicarbonato, factor intrínseco y agua. Estas sustancias continúan la digestión que se inició en la boca, y además ayudan a proteger al estómago de daños. También tiene importantes funciones motoras, que regulan la ingesta de alimentos, su mezcla con las secreciones gástricas y reducción en tamaño, y la salida del material parcialmente digerido hacia el duodeno. Además, el estómago produce dos importantes factores humorales, que son la gastrina y la somatostatina, y tienen acciones endocrinas y paracrinas. Ambos son importantes en la regulación de la secreción gástrica. Aunque estas funciones son importantes en la mantención de una buena salud, el estómago no es un órgano necesario para sobrevivir. De hecho, hay personas que se remueven completamente su estómago por razones no neoplásicas (i.e. cáncer), y se pueden nutrir adecuadamente y alcanzar una excelente longevidad. ANATOMÍA FUNCIONAL DEL ESTÓMAGO LA MUCOSA ESTÁ COMPUESTA DE CÉLULAS EPITELIALES SUPERFICIALES Y GLÁNDULAS La estructura básica de la pared estomacal es similar a la que vemos en otras regiones del tracto GI; por lo cual decimos que la pared estomacal consiste de una capas mucosales y musculares. Como un todo, puede ser dividido en 3 regiones: 1. El cardias, justo distal a la unión gastroesofágica, lleno de células parietales (secretoras de ácido). 2. El cuerpo o corpus, que es la porción más grande, y a su vez su región más proximal es el fondo del estómago o fundus. 3. La porción distal o antro. El área de superficie de la mucosa gástrica se encuentra sustancialmente aumentada por la presencia de glándulas gástricas, que incluyen varios tipos celulares, como las mucosas, parietales, principales y endocrinas. Las células endocrinas se encuentran también presentes en el cuerpo y el antro. Las epiteliales superficiales tienen una estructura y función propia distinta, y secretan HCO 3 - y moco. Existe una heterogeneidad celular marcada no sólo en el mismo segmento, sino también entre segmentos del estómago, por ejemplo, como se discutirá después, la estructura y función de las células mucosales epiteliales en el antro y el cuerpo son muy distintas. De la misma manera, aunque el músculo liso de las porciones proximal y distal del estómago se ve similar estructuralmente, tiene función y propiedades farmacológicas diferentes. A MAYOR SECRECIÓN DE JUGO GÁSTRICO, MAYOR CONCENTRACIÓN DE H + Y MENOR DE NA + Las glándulas estomacales secretan ~2L/día de un fluido que es más menos isotónico con el plasma sanguíneo. La secreción gástrica tiene 2 componentes de secreción: uno que proviene de las células parietales y otro que no. De acuerdo a esto, la secreción gástrica consiste en: 1. Una secreción rica en Na + que viene de las células no parietales. 2. Una secreción rica en H + que viene de las células parietales. Este modelo ayuda a explicar la relación inversa que existe entre las concentraciones luminales de H + y Na + como función de la tasa de secreción gástrica. Por ejemplo, cuando la gastrina o histamina estimulan a las células parietales la [H + ] intraluminal es alta, mientras que la [Na + ] intraluminal es relativamente baja. A una secreción menor o en situaciones clínicas en donde la secreción máxima de ácido se ve reducida (como en la anemia perniciosa) la [H + ] intraluminal es baja pero la [Na + ] es alta. LA PORCIÓN PROXIMAL DEL ESTÓMAGO SECRETA ÁCIDO, PEPSINÓGENOS, FACTOR INTRÍNSECO, BICARBONATO Y MOCO, MIENTRAS QUE LA PORCIÓN DISTAL LIBERA GASTRINA Y SOMATOSTATINA CUERPO Los principales productos de secreción de la parte proximal del estómago son ácido, pepsinógenos y factor intrínseco. Cada uno de ellos es sintetizado por distintos tipos celulares que se agrupan en glándulas. Los dos tipos celulares principales en las glándulas gástricas del cuerpo del estómago son las células parietales y las principales. Las células parietales (u oxínticas) secretan ácido y factor intrínseco, este último es necesario para la absorción de la vitamina B12 en el íleon. Su morfología es grande, de forma triangular con un núcleo central y abundancia de mitocondrias.
-
Upload
sebastian-lavanderos-bunout -
Category
Documents
-
view
1.571 -
download
1
Transcript of Función gástrica

Sebastián Lavanderos B. 2do. Medicina UDP
1
FUNCIÓN GÁSTRICA
El estómago tiene un papel importante en la
nutrición humana, y tiene funciones secretoras,
motoras y humorales. Esas actividades no están
separadas ni son distintas, sino más bien representan
una función integrada que se requiere como un todo
para iniciar el proceso digestivo normal.
El estómago tiene varios productos secretores
específicos, entre los cuales encontramos, aparte del
conocido ácido, al pepsinógeno, moco, bicarbonato,
factor intrínseco y agua. Estas sustancias continúan la
digestión que se inició en la boca, y además ayudan a
proteger al estómago de daños.
También tiene importantes funciones motoras, que
regulan la ingesta de alimentos, su mezcla con las
secreciones gástricas y reducción en tamaño, y la
salida del material parcialmente digerido hacia el
duodeno.
Además, el estómago produce dos importantes
factores humorales, que son la gastrina y la
somatostatina, y tienen acciones endocrinas y
paracrinas. Ambos son importantes en la regulación
de la secreción gástrica.
Aunque estas funciones son importantes en la
mantención de una buena salud, el estómago no es
un órgano necesario para sobrevivir. De hecho, hay
personas que se remueven completamente su
estómago por razones no neoplásicas (i.e. cáncer), y
se pueden nutrir adecuadamente y alcanzar una
excelente longevidad.
ANATOMÍA FUNCIONAL DEL ESTÓMAGO
LA MUCOSA ESTÁ COMPUESTA DE CÉLULAS EPITELIALES
SUPERFICIALES Y GLÁNDULAS
La estructura básica de la pared estomacal es similar
a la que vemos en otras regiones del tracto GI; por lo
cual decimos que la pared estomacal consiste de una
capas mucosales y musculares. Como un todo, puede
ser dividido en 3 regiones:
1. El cardias, justo distal a la unión
gastroesofágica, lleno de células parietales
(secretoras de ácido).
2. El cuerpo o corpus, que es la porción más
grande, y a su vez su región más proximal
es el fondo del estómago o fundus.
3. La porción distal o antro.
El área de superficie de la mucosa gástrica se
encuentra sustancialmente aumentada por la
presencia de glándulas gástricas, que incluyen varios
tipos celulares, como las mucosas, parietales,
principales y endocrinas. Las células endocrinas se
encuentran también presentes en el cuerpo y el
antro. Las epiteliales superficiales tienen una
estructura y función propia distinta, y secretan HCO3-
y moco.
Existe una heterogeneidad celular marcada no sólo
en el mismo segmento, sino también entre
segmentos del estómago, por ejemplo, como se
discutirá después, la estructura y función de las
células mucosales epiteliales en el antro y el cuerpo
son muy distintas. De la misma manera, aunque el
músculo liso de las porciones proximal y distal del
estómago se ve similar estructuralmente, tiene
función y propiedades farmacológicas diferentes.
A MAYOR SECRECIÓN DE JUGO GÁSTRICO, MAYOR
CONCENTRACIÓN DE H+ Y MENOR DE NA
+
Las glándulas estomacales secretan ~2L/día de un
fluido que es más menos isotónico con el plasma
sanguíneo. La secreción gástrica tiene 2
componentes de secreción: uno que proviene de las
células parietales y otro que no. De acuerdo a esto, la
secreción gástrica consiste en:
1. Una secreción rica en Na+ que viene de las
células no parietales.
2. Una secreción rica en H+ que viene de las
células parietales.
Este modelo ayuda a explicar la relación inversa que
existe entre las concentraciones luminales de H+ y
Na+ como función de la tasa de secreción gástrica.
Por ejemplo, cuando la gastrina o histamina
estimulan a las células parietales la [H+] intraluminal
es alta, mientras que la [Na+] intraluminal es
relativamente baja. A una secreción menor o en
situaciones clínicas en donde la secreción máxima de
ácido se ve reducida (como en la anemia perniciosa) la
[H+] intraluminal es baja pero la [Na+] es alta.
LA PORCIÓN PROXIMAL DEL ESTÓMAGO SECRETA ÁCIDO,
PEPSINÓGENOS, FACTOR INTRÍNSECO, BICARBONATO Y
MOCO, MIENTRAS QUE LA PORCIÓN DISTAL LIBERA
GASTRINA Y SOMATOSTATINA
CUERPO
Los principales productos de secreción de la parte
proximal del estómago son ácido, pepsinógenos y
factor intrínseco. Cada uno de ellos es sintetizado por
distintos tipos celulares que se agrupan en glándulas.
Los dos tipos celulares principales en las glándulas
gástricas del cuerpo del estómago son las células
parietales y las principales.
Las células parietales (u oxínticas) secretan ácido y
factor intrínseco, este último es necesario para la
absorción de la vitamina B12 en el íleon. Su
morfología es grande, de forma triangular con un
núcleo central y abundancia de mitocondrias.

Sebastián Lavanderos B. 2do. Medicina UDP
2
Las células principales (o pépticas) secretan
pepsinógenos, pero no ácido. Morfológicamente, son
más chicas que las parietales, y funcionalmente
existe una relación entre el pH y la secreción y
activación de la pepsina. La pepsina es una
endopeptidasa (hidroliza enlaces peptídicos
interiores) e inicia la digestión proteica mediante la
hidrólisis de enlaces peptídicos específicos. El pH
luminal basal del estómago oscila en un rango que va
de 4 a 6, pero, con estimulación, el pH de las
secreciones gástricas baja usualmente a menos de 2.
A valores de pH < 3, los pepsinógenos se activan
rápidamente a pepsinas. Un bajo pH gástrico también
ayuda a prevenir la colonización bacterial del
intestino delgado.
Además de las células parietales y principales, las
glándulas del cuerpo contienen también células
secretoras de moco que se ubican en el cuello de la
glándula, y 5 o 6 células endocrinas. Entre estas
últimas se encuentran las células similares a
enterocromafina, que liberan histamina.
ANTRO
Las glándulas del antro del estómago no tienen
células parietales. Entonces, el antro no secreta ni
ácido ni factor intrínseco. Las glándulas de la mucosa
antral contienen en cambio células principales y
células endocrinas. Las endocrinas incluyen a las
células G y D, que secretan gastrina y somatostatina,
respectivamente. La gastrina estimula la secreción de
ácido gástrico vía 2 mecanismos, y es un factor
trófico principal para la proliferación celular epitelial
del tracto GI. La somatostatina tiene hartas
funciones, pero su principal en la fisiología gástrica es
inhibir la liberación de gastrina y la secreción de ácido
de parte de las células parietales.
Además de las células que componen a las glándulas,
el estómago también contiene células epiteliales
superficiales que cubren los hoyos gástricos y las
superficies entre estos. Estas células secretan HCO3-.
EL ESTÓMAGO ACOMODA LA COMIDA, LA MEZCLA CON
LAS SECRECIONES GÁSTRICAS, LA MUELE Y VACÍA EL
QUIMO HACIA EL DUODENO
Además de sus funciones de secreción, el estómago
tiene múltiples funciones motoras. Estas son el
resultado de la actividad muscular lisa gástrica,
integrada por señales neurales y humorales, e
incluyen tanto movimientos propulsivos y
retrógrados de la comida y líquidos, así como un
movimiento no propulsivo que aumenta la presión
intragástrica.
Se observa una diversidad considerable en la
regulación y contractilidad del músculo liso gástrico.
El estómago tiene al menos 2 áreas de actividad

Sebastián Lavanderos B. 2do. Medicina UDP
3
motora; las porciones distal y proximal de este se
comportan como entidades distintas, pero
coordinadas. Se identifican al menos 4 eventos en el
proceso de llenado y vaciamiento gástrico:
1. Recepción y almacenamiento temporal de
la comida y líquidos de la dieta.
2. Mezcla de la comida y agua con los
productos de la secreción gástrica, incluidos
la pepsina y el ácido.
3. Molienda de la comida para que se reduzca
el tamaño de las partículas y de esta forma
estimular la digestión y permitir su paso por
el píloro.
4. Regulación de la salida del material retenido
desde el estómago hacia el duodeno en
respuesta a varios estímulos.
Los mecanismos mediante los cuales el estómago
recibe y vacía líquidos y sólidos son significativa-
mente distintos. El vaciamiento de los líquidos es
principalmente función del músculo liso de la parte
proximal del estómago, mientras que el vaciamiento
de los sólidos es regulado por el músculo liso antral.
SECRECIÓN DE ÁCIDO
LA CÉLULA PARIETAL TIENE UNA ESTRUCTURA TÚBULO-
VESICULAR ESPECIALIZADA QUE AUMENTA EL ÁREA
APICAL DE MEMBRANA CUANDO LA CÉLULA ES
ESTIMULADA PARA SECRETAR ÁCIDO
En estado basal, se secreta poco ácido. Esta
secreción ocurre gracias a una bomba H-K (o H,K-
ATPasa) que se encuentra en las membranas
tubulovesiculares de la porción apical de las células
parietales en reposo. Al ser estimuladas, un re arreglo
del citoesqueleto hace que las membranas
tubulovesiculares que contienen a la bomba H-K se
fusionen con la membrana canalicular, lo que hace
que el área de superficie de la membrana apical de la
célula parietal aumente de 50 a 100 veces, y que
aparezcan microvellosidades. Además, las bombas H-
K y canales de K+ y Cl- se insertan en la membrana
canalicular. La excreción de ácido, además, requiere
ATP, por lo cual existe un alto número de
mitocondrias en la célula parietal.
UNA BOMBA H-K ES RESPONSABLE DE LA SECRECIÓN DE
ÁCIDO GÁSTRICO POR LAS CÉLULAS PARIETALES
La bomba H-K de la célula parietal es miembro de la
familia de ATPasas tipo P, que incluyen a la bomba
Na-K presente en la membrana basolateral de
virtualmente todas las células epiteliales de un
mamífero y en la membrana plasmática de células no
polarizadas. Esta bomba requiere 2 subunidades para
funcionar. Una α, en donde reside su actividad
catalítica, y una β, que la ancla a la membrana apical.
La actividad de esta ATPasa es afectada por
inhibidores con importancia clínica en el control de la
secreción de ácido gástrico:
1. Los benzimidazoles, como el omeprazol,
que se unen covalentemente a cisteínas en
la superficie extracitoplasmática.
2. Sustancias que actúan como inhibidores
competitivos del sitio de unión del K+.
Cuando la célula parietal es estimulada, bombas H-K
(alimentadas por la hidrólisis de ATP), mandan H+
hacia el lumen de la glándula gástrica haciendo entrar
K+ en el proceso. El K+ se recicla y vuelve al lumen por
canales de K+. Sale Cl- a través de canales en la
membrana luminal, completando de este modo la
exceción neta de HCl. El H+ requerido para el
funcionamiento de la bomba H-K es provisto por la
entrada de CO2 y H2O, que son convertidos en H+ y
HCO3- por la anhidridasa carbónica. El HCO3
- sale a
través de la membrana basolateral a través de un
intercambiador Cl-HCO3. El intercambiador Na-H
basolateral puede participar en la regulación
intracelular del pH, especialmente en estado basal.
3 SECRETAGOGOS (ACH, GASTRINA E HISTAMINA)
INDUCEN, TANTO DIRECTA COMO INDIRECTAMENTE, LA
SECRECIÓN DE ÁCIDO POR LAS CÉLULAS PARIETALES
La acción de los secretagogos en la secreción de
ácido gástrico ocurre a través de al menos 2
mecanismos paralelos y quizás redundantes:

Sebastián Lavanderos B. 2do. Medicina UDP
4
1. La ACh, gastrina e histamina se unen
directamente a sus receptores de
membrana respectivos en la célula parietal,
y estimulan la secreción de ácido gástrico
sinérgicamente.
La ACh es liberada desde las terminaciones
nerviosas del vago, la gastrina de las células
G. La histamina se sintetiza a partir de la
histidina.
2. La ACh y la gastrina inducen la secreción de
ácido gástrico indirectamente estimulando
a las células ECL de la lámina propia para
que liberen histamina, y esto último induce
directamente el aumento de secreción.
Existen antagonistas del receptor H2 (de histamina)
como la cimetidina y la ranitidina, que no sólo
bloquean la acción directa de la histamina en las
células parietales, sino que además inhiben a la
secreción de ácido gástrico estimulada por la ACh y
gastrina, lo que los hace un tratamiento efectivo para
tratar úlceras pépticas activas.
LOS 3 SECRETAGOGOS PARA ÁCIDO ACTÚAN A TRAVÉS DE
CA+2/DIACILGLICEROL O CAMP
La estimulación de la secreción de ácido gástrico por
la ACh, gastrina e histamina es mediada por una serie
de procesos de transducción de señal intracelulares.
Los 3 se unen a GPCRs (Receptores acoplados a
proteína G) en la membrana de la célula parietal.
La ACh se une al receptor muscarínico M3, que se
asocia a una proteína Gαq, que activa a la PLC, que
convierte PIP2 en IP3 y DAG. El IP3 hace que se libere
Ca+2 desde los reservorios internos celulares que
probablemente actúa luego vía una quinasa de
proteínas dependiente de calmodulina y el DAG
activa a la PKC. Además, el receptor M3 activa un
canal de Ca+2 de membrana.
La gastrina se une a un receptor específico de la
célula parietal identificado como receptor de
gastrina-colecistoquinina B (CCKB). 2 receptores
relacionados de CCK han sido identificados: CCKA y
CCKB. Sus secuencias de aminoácidos son idénticas en
un ~50%, y ambos son GPCRs. El receptor CCKB tiene
la misma afinidad por la gastrina y la CCK, mientras
que el receptor CCKA tiene una afinidad 3 órdenes de
magnitud mayor por la CCK que por la gastrina. Esto
explica los efectos paralelos pero a veces opuestos
de la CCK y la gastrina en la función GI. El receptor
CCKB se une a Gαq y activa la vía de la PLC,
produciendo un [Ca+2]i y activación de la PKC.
El receptor de histamina en las células parietales es
un receptor H2, que se encuentra unido a una
proteína Gas de unión a GTP. La activación de este
receptor por parte de la histamina estimula a la
enzima adenil ciclasa, lo que genera cAMP,
aumentando su concentración intracelular. Esto hace
que se active la PKA, que lleva a la fosforilación de
ciertas proteínas específicas de la célula parietal,
incluyendo a la bomba H-K.
LA GASTRINA ES LIBERADA POR LAS CÉLULAS G
ANTRALES Y DUODENALES, Y LA HISTAMINA ES LIBERADA
POR LAS CÉLULAS SIMILARES A ENTEROCROMAFINA EN EL
CUERPO DEL ESTÓMAGO
La gastrina tiene 3 efectos principales en las céls. GI:
1. Estimula la secreción de ácido gástrico por
parte de las células parietales.
2. Hace que las células ECL liberen histamina.
3. Regula el crecimiento mucosal en el cuerpo
del estómago y en el intestino delgado y
grueso.
La gastrina existe en muchas formas, pero las 2
principales son la G-17 o “gastrina pequeña” y la G-34
o “gastrina grande”. Tiene un residuo de tirosina, el
cual puede estar sulfatado (la llamada gastrina II) o
sin sulfatar (la llamada gastrina I). Ambas formas son
igualmente activas y se encuentran en cantidades
iguales en condiciones normales. La gastrina y la CCK,
una hormona relacionada a esta tiene secuencias C-
terminales idénticas. La G-17 es más activa que la G-
34, pero la última se degrada a una velocidad menor
que la G-17, por lo tanto, una infusión de cantidades
iguales de G-17 o G-34 produce aumentos similares en
la secreción de ácido gástrico.
Células endocrinas especializadas (células G) en el
antro y el duodeno sintetizan ambas gastrinas. Las
células G antrales son la fuente principal de G-17,
mientras que las células G duodenales lo son de G-34.
Las células G antrales responden a estímulos tanto
luminales como basolaterales, y tienen
microvellosidades en su superficie apical, por lo que
nos referimos a ellas como células endocrinas de tipo
abierto. Éstas liberan gastrina en respuesta a
péptidos y aminoácidos luminales, así como a

Sebastián Lavanderos B. 2do. Medicina UDP
5
péptido liberador de gastrina (GRP), que es liberado
por las terminaciones vagales. La liberación de la
gastrina es inhibida por la somatostatina, que es
liberada de las células D adyacentes.
LA SOMATOSTATINA, LIBERADA POR LAS CÉLULAS D
GÁSTRICAS, ES EL MECANISMO PRINCIPAL PARA LA
INHIBICIÓN DE LA SECRECIÓN DE ÁCIDO
La secreción de ácido gástrico es controlada por vías
estimulantes y vías inhibitorias. La inhibitoria
principal es a través de somatostatina, que es
sintetizada por las células D del antro y el cuerpo del
estómago. También es sintetizada por las células δ de
los islotes pancreáticos y por neuronas
hipotalámicas. Existe en 2 formas, SS-28 y SS-14, y la
SS-28 es la predominante en el tracto GI.
La somatostatina inhibe la secreción de ácido
gástrico a través de 2 mecanismos: uno directo y 2
indirectos:
En la vía directa la somatostatina se une a un
receptor Gai (SST) en la membrana basolateral de la
célula parietal e inhibe a la adenil ciclasa. El efecto
neto es antagonizar la acción de la histamina y de
esta manera inhibir la secreción de ácido gástrico. La
somatostatina puede ser paracrina (céls. D del
cuerpo) o endocrina (céls. D del antro). Mecanismos
neurales y hormonales estimulan a las células D del
cuerpo (que no sensan pH), mientras que las células
D del antro son estimuladas por un pH intraluminal
bajo.
La somatostatina actúa también a través de 2 vías
indirectas, ambas son paracrinas. En el cuerpo del
estómago, las células D liberan somatostatina que
inhibe la liberación de histamina de las células ECL, y
como la histamina es secretagogo de ácido gástrico
esto inhibe su secreción. La otra vía consiste en la
liberación de somatostatina en el antro del estómago
por las células D, lo que inhibe la liberación de
gastrina de las células G. Como la gastrina también es
un secretagogo de ácido gástrico, esto también
inhibe su secreción. La gastrina liberada por las
células G se hace feed back a si misma estimulando
las células D para que liberen somatostatina.
La presencia de múltiples mecanismos mediante los
cuales la somatostatina inhibe la secreción de ácido
es otro ejemplo de las vías regulatorias redundantes
que controlan este proceso. Aunque es un campo
aún en investigación, parece ser que la gastrina
estimula la liberación de somatostatina, mientras que
agonistas colinérgicos la inhiben.
MUCHAS HORMONAS ENTÉRICAS “ENTEROGASTRONAS”
Y PROSTAGLANDINAS INHIBEN LA SECRECIÓN DE ÁCIDO
GÁSTRICO
Múltiples procesos en el duodeno y yeyuno
participan en el feedback negativo que inhibe la
secreción de ácido gástrico. La grasa, ácido y
soluciones hiperosmolares en el duodeno son
potentes inhibidores se la secreción de ácido
gástrico. De ellos, los lípidos son el más potente, pero
el ácido también es muy importante. Se han
propuesto varias hormonas que podrían mediar esta
inhibición, como la CCK, secretina, péptido YY, VIP,
GIP y la neurotensina. Ninguno de ellos ha podido ser
identificado como la “enterogastrona” única, todos
inhiben la secreción de ácido después de su
administración.
Hay evidencia que sugiere que la secretina, que es
liberada por las células S duodenales, tendría un rol
fundamental en inhibir la secreción de ácido gástrico
luego de la entrada de grasa y ácido al duodeno. Esto
lo haría a través de al menos 3 mecanismos:
1. Inhibición de la liberación de gastrina antral.
2. Estimulación de la liberación de SS.
3. Downregulation directa del proceso
secretor de H+ de las céls. parietales.
La presencia de ácidos grasos en el lumen hace que
las células enteroendocrinas del duodeno liberen GIP
y CCK. El GIP reduce la secreción de ácido gástrico
directamente inhibiendo la secreción de las células
parietales e inhibiendo la liberación antral de
gastrina. La CCK participa en la inhibición por
feedback de la secreción de ácido reduciendo
directamente la secreción de ácido de las células
parietales.
La Prostaglandina E2 (PGE2) inhibe la secreción de
ácido gástrico desde las células parietales,
probablemente inhibiendo la activación por
histamina de las células parietales distal a su
receptor. La PGE2 parece unirse a un receptor EP3 en
la membrana basolateral de la célula parietal lo que
estimula una proteína Gai, que inhibe la adenil ciclasa.
También inhiben la secreción indirectamente a través
de la reducción de la liberación de histamina de las
células ECL y la liberación de gastrina de las células G
antrales.
COMER GATILLA 3 FASES DE SECRECIÓN DE ÁCIDO
ESTADO BASAL
La secreción de ácido gástrico ocurre día y noche.
Después de comer aumenta, y entre comidas es baja
(en la fase intedigestiva). Este periodo interdigestivo
sigue un ritmo circadiano; la secreción de ácido es la
más baja en la mañana antes de despertar y es la más
alta en la noche. La secreción es, también, función del
número de células parietales, lo que se ve
influenciado por el peso. Entonces, los hombres
tienen tasas de secreción basal de ácido mayores que
las de las mujeres. El pH intragástrico varía de 3 a 7 en
descanso, y depende no sólo de la secreción de
ácido, sino también del poder de tamponamiento

Sebastián Lavanderos B. 2do. Medicina UDP
6
que tiene la comida que se ingiere y de la velocidad
en que se vacíe el ácido y material parcialmente
digerido al duodeno.
La regulación de la secreción de ácido durante una
comida puede ser mejor caracterizada si la
separamos en 3 fases separadas pero
interrelacionadas: la cefálica, la gástrica y la intestinal.
Las fases cefálica y gástrica son de mayor
importancia. La regulación de la secreción de ácido
involucra tanto estimulación como inhibición. La ACh,
gastrina e histamina promueven la secreción de
ácido, mientras que la SS inhibe la secreción de ácido
gástrico. Existe superposición de las fases.
FASE CEFÁLICA
La fase cefálica se inicia al oler, sentir, probar, pensar
y tragar comida. Es mediada principalmente por el
nervio vago, que al sensar los estímulos, activa el
núcleo dorsal motor del vago en la médula, y activa
los nervios pregangliónicos parasimpáticos
eferentes. Hipoglicemia inducida por insulina también
estimula el vago, y al hacer esto promueve la
secreción de ácido. La estimulación del vago resulta
en 4 eventos fisiológicos distintos:
1. En el cuerpo del estómago, los nervios
muscarínicos vagales postgangliónicos
liberan ACh que estimula directamente la
secreción de H+ en las células parietales.
2. La ACh liberada por las terminaciones
nerviosas vagales induce la liberación de
histamina por parte de las células ECL de la
lámina propia del cuerpo del estómago, lo
que estimula la secreción de ácido.
3. En el antro, neuronas peptidérgicas
postgangliónicas parasimpáticas vagales
junto a otras neuronas del SNE inducen la
liberación de GRP, que induce la liberación
de gastrina desde las células G antrales, lo
cual hace que se estimule la secreción de
ácido tanto directamente como
indirectamente a través de histamina (ECL).
4. En el antro y el cuerpo del estómago, el
nervio vago inhibe a las células D, haciendo
que liberen menos SS.
Entonces, la fase cefálica estimula la secreción de
ácido directa e indirectamente actuando sobre la
célula parietal. En la fase cefálica se secreta un ~30%
de la secreción total de ácido, y ocurre antes de la
entrada de comida al estómago.
Uno de los enfoques quirúrgicos para el tratamiento
de la úlcera péptica es cortar los nervios vagos (una
vagotomía), para inhibir la secreción de ácido. Es muy
poco frecuente, y poco preferida, debido al alto
número de opciones farmacológicas poco invasivas
que existen para el tratamiento de la enfermedad.
Además, produciría un retraso en el vaciamiento
gástrico y diarrea. Para minimizar estos efectos
colaterales, se han realizado vagotomías selectivas,
en donde se cortan sólo las fibras que van hacia las
células parietales.
FASE GÁSTRICA
La entrada de comida al estómago inicia los 2
estímulos principales para la fase gástrica de
secreción de ácido:
1. La comida distiende la mucosa gástrica, lo
que activa un reflejo vagovagal y reflejos
locales del SNE.
2. Proteínas parcialmente digeridas estimulan
las células G antrales.
La distensión de la pared gástrica (en el cuerpo y en
el antro) luego de la entrada de comida hace que el
estómago invoque 2 vías mediadas neuralmente:
1. Activación de un reflejo vagovagal, en que
la distensión de la pared gástrica activa una
vía aferente vagal, que, a su vez, estimula
una respuesta vagal eferente en el núcleo
dorsal del vago. La estimulación de la
secreción de ácido ocurre a través de las 4
vías paralelas que se activan con el vago.
2. La distensión de la pared gástrica activa
también una vía local del SNE que libera
ACh, que, a su vez, estimula la secreción de
ácido gástrico por las células parietales.
La presencia de proteínas parcialmente digeridas o
aminoácidos en el antro estimula directamente a las
células G para que liberen gastrina. Proteínas intactas
no tienen efecto.
La secreción de ácido y la activación de pepsinógeno
se encuentran relacionados por un loop de feedback
positivo, en donde un pH bajo estimula la conversión
de pepsinógeno a pepsina. La pepsina digiere
proteínas a péptidos, que promueven la liberación de
gastrina. Finalmente, la gastrina promueve la
secreción de ácido, lo que cierra el loop.
Además de las 2 vías estimulatorias que actúan
durante la fase gástrica, hay una tercera vía que
inhibe la secreción de ácido gástrico vía un
mecanismo de feedback negativo clásico. Un pH
intragástrico bajo estimula a las células D antrales
para que liberen somatostatina. Ya que la
somatostatina inhibe la liberación de gastrina por las
células G, el efecto neto es una reducción en la
secreción de ácido gástrico. La efectividad de un pH
bajo en la inhibición de la liberación de gastrina se
enfatiza en la siguiente observación: Aunque
péptidos normalmente son un estímulo potente para
liberar gastrina, ellos no logran estimular la liberación
de gastrina ni cuando el pH intraluminal del antro se

Sebastián Lavanderos B. 2do. Medicina UDP
7
mantiene en 1,0 o cuando se infunde SS. La fase
gástrica de la secreción de ácido, que ocurre
principalmente como resultado de la liberación de
gastrina, cuenta por el 50-60% de la secreción total de
ácido gástrico.
FASE INTESTINAL
La presencia de aminoácidos y péptidos parcialmente
digeridos en la porción proximal del intestino
delgado estimula la secreción de ácido por 3
mecanismos:
1. Los péptidos estimulan las células G
duodenales para secretar gastrina, justo
como estimulan a las células G antrales en la
fase gástrica.
2. Los péptidos estimulan una célula
endocrina desconocida para que libere una
señal humoral adicional que ha sido referida
como entero-oxintina.
3. Aminoácidos absorbidos por la parte
proximal del intestino delgado estimulan la
secreción de ácido por mecanismos sin
definir.
La secreción de ácido gástrico mediada por la fase
intestinal es estimulada después de un shunt
portacaval. Dicho shunt, usado en el tratamiento de
la hipertensión portal causada por enfermedad
crónica del hígado, desvía la sangre portal que drena
el intestino delgado alrededor del hígado en su vuelta
al corazón, entonces, la señal liberada por el intestino
delgado durante la fase intestinal, en individuos
normales, es probablemente removida, en parte, por
el hígado antes de alcanzar su blanco, el cuerpo del
estómago. Aproximadamente de un 5 a un 10% de la
secreción total de ácido gástrico ocurre como
resultado de la fase intestinal.
SECRECIÓN DE PEPSINÓGENO
LAS CÉLULAS PRINCIPALES, GATILLADAS POR LAS VÍAS
DEL CAMP Y CA+2, SECRETAN MÚLTIPLES
PEPSINÓGENOS, QUE INICIAN LA DIGESTIÓN PROTEICA
Las células principales de las glándulas gástricas, así
como las células mucosas secretan pepsinógenos,
que son proenzimas proteolíticas (o zimógenos) que
pertenecen a la familia de proteasas aspárticas. Estos
se activan a pepsinas cuando se les rompe un péptido
N-terminal. Las pepsinas son endopeptidasas que
inician la hidrólisis de las proteínas ingeridas en el
estómago.
La secreción de pepsinógeno en estado basal es un
~20% de su secreción máxima luego de estimulación.
La liberación de pepsinógeno a través de la
membrana apical es resultado de exocitosis
compuesta, en donde gránulos secretores se
fusionan con la membrana plasmática y otros
gránulos. Esto permite la secreción rápida y sostenida
de pepsinógeno. Después de la estimulación, el peak
inicial en la secreción de pepsinógeno es seguido de
una tasa de secreción menor, lo que nos indica que
primero se libera el pepsinógeno preformado seguido
de la liberación del pepsinógeno que está siendo
sintetizado.
2 grupos de agonistas estimulan a las células
principales para que secreten pepsinógeno. Un grupo
actúa a través de la adenil ciclasa y cAMP, mientras
que el otro grupo actúa a través de [Ca+2]i.

Sebastián Lavanderos B. 2do. Medicina UDP
8
AGONISTAS QUE ACTÚAN A TRAVÉS DE CAMP
Las células principales tienen receptores para
secretina/VIP, adrenérgicos β2 y EP2 para PGE2. Todos
estos receptores activan la adenil ciclasa. A
concentraciones más bajas que las requeridas para
estimular la secreción de pepsinógeno, la PGE2
también puede inhibir la secreción de pepsinógeno,
presumiblemente al unirse a otro subtipo de
receptor.
AGONISTAS QUE ACTÚAN A TRAVÉS DEL CA+2
Las células principales también tienen receptores
muscarínicos M3 para la ACh, así como receptores
para la familia de péptidos gastrina/CCK. La
secreción de pepsinógeno es estimulada por el
receptor CCKA, que tiene una afinidad mucho mayor
por la CCK que por la gastrina. La activación de
ambos receptores hace que se libere Ca+2 de los
reservorios intracelulares vía IP3 y por tanto produce
un aumento en la [Ca+2]i.
De estos agonistas, el más importante para la
secreción de pepsinógeno es la ACh liberada en
respuesta a la estimulación vagal. La ACh no sólo
estimula a las células parietales para que liberen
pepsinógeno, sino que también a las parietales para
que liberen ácido. El ácido gástrico produce una
secreción aumentada de pepsinógeno vía 2
mecanismos distintos. Primero, en el estómago, una
caída en el pH invoca un reflejo colinérgico local que
resulta en la estimulación incrementada de las células
principales para que liberen pepsinógeno. Entonces,
la ACh que estimula a las células principales puede
venir del vago o del reflejo local. Segundo, en el
duodeno, el ácido gatilla la liberación de secretina
desde las células S. Esta estimula por vía endocrina a
las células principales para que liberen más
pepsinógeno. El rol exacto de la histamina y la
gastrina en la secreción de pepsinógeno permanece
poco claro.
SE REQUIERE UN BAJO PH PARA LA ACTIVACIÓN DEL
PEPSINÓGENO Y LA ACTIVIDAD DE LA PEPSINA
El pepsinógeno es inactivo, y requiere su activación a
pepsina, su estado de proteasa para iniciar la
digestión de proteínas. Esta activación ocurre por el
clivaje espontáneo de un pequeño fragmento
peptídico N-terminal, pero sólo a un pH menor a 5.
Entre un pH 3 y 5, la activación es lenta, pero se
vuelve extremadamente rápida a pH <3. Además, el
pepsinógeno se autoactiva, esto es, la pepsina
formada cliva pepsinógenos a pepsina.
Una vez que la pepsina se forma, su actividad
también es pH-dependiente. Su pH óptimo se
encuentra entre 1,8 y 3,5. Valores de pH > a 3,2
inactivan reversiblemente a la pepsina, y valores de
pH > a 7,2 inactivan irreversiblemente a la enzima.
Los péptidos generados por la pepsina estimulan la
secreción de ácido gástrico requerida para la
activación y acción de la pepsina. Entonces, los
péptidos que libera la pepsina son importantes para
iniciar una respuesta coordinada a una comida.
También se encuentran en el estómago productos
digestivos de carbohidratos y lípidos, aunque la
secreción de sus enzimas digestivas respectivas no
ocurre o no es función principal de las células
epiteliales gástricas. La digestión de carbohidratos es
iniciada en la boca por la amilasa salival. Sin
embargo, luego de que nos tragamos esta enzima, el
estómago se vuelve un sitio más importante para la
hidrólisis del almidón que la boca. De la misma
manera, aunque la digestión de lípidos también se
inicia en la boca por la lipasa lingual, ocurre una
digestión bastante significativa de lípidos en el
estómago como resultado de la lipasa lingual
tragada y de la lipasa gástrica, ambas con pH de
acción óptimos ácidos.
PROTECCIÓN DEL EPITELIO DE LA SUPERFICIE
GÁSTRICA Y NEUTRALIZACIÓN DEL ÁCIDO EN EL
DUODENO Cuando las células parietales secretan H+ a su
capacidad máxima, el pH intraluminal del estómago
puede llegar a valores tan bajos como 1 o incluso
menos por largos periodos de tiempo. El epitelio
gástrico debe mantener gradientes de concentración
de H+ hasta un millón de veces mayores que su
contraparte intracelular, ya que este pH en las células
epiteliales es de 7,2 y el del plasma es de 7,4. Lo
mismo se observa con el Na+. ¿Cómo el estómago es
capaz de mantener estos gradientes?, ¿Cómo las
células epiteliales no se destruyen con esta acidez?, y
¿Cómo las pepsinas no digieren a las mismas células
epiteliales si están en el lumen gástrico? Todo se
responde gracias a la barrera de difusión gástrica.
Aunque definirla es complejo, se sabe que esta
barrera es tanto anatómica como fisiológica, y que
tiene 3 componentes:
1. La membrana apical y las tight junctions
epiteliales en las glándulas gástricas, que
proporcionan una impermeabilidad relativa
al ácido de la membrana apical.
2. Una capa de gel mucosa de grosor variable
(50-200 µm) sobre las células epiteliales
superficiales.
3. Un microclima con HCO3- adyacente a las
células epiteliales superficiales que
mantiene un pH local relativamente alto.
LA ESTIMULACIÓN VAGAL Y LA IRRITACIÓN ESTIMULAN A
LAS CÉLULAS MUCOSAS GÁSTRICAS PARA QUE SECRETEN
MUCINA, UNA GLICOPROTEÍNA PARTE DE LA BARRERA
La capa mucosa está compuesta de mucina,
fosfolípidos, electrolitos y agua. La mucina es la

Sebastián Lavanderos B. 2do. Medicina UDP
9
glicoproteína de alto peso molecular que contribuye
a formar la capa de protección sobre la mucosa
gástrica. La mucina gástrica es un tetrámero de 4
péptidos iguales, a los cuales se les unen
polisacáridos largos resultando en un carbohidrato, al
cual se debe la viscosidad del moco, lo que explica,
en su mayoría, su rol de protección en fisiología
gástrica.
El moco es secretado por 3 células mucosas distintas:
las de superficie, las mucosas del cuello y las mucosas
glandulares, que secretan distintos tipos de moco. El
sintetizado y secretado en las células glandulares es
una glicoproteína neutra, mientras que las células
mucosas de la superficie y del cuello secretan
glicoproteínas neutras y ácidas. La mucina forma la
capa de gel mucosa junto a fosfolípidos, electrolitos y
agua. Ésta provee protección contra lesiones. La
mucina también lubrica a la mucosa gástrica, para
minimizar los efectos abrasivos de la comida.
La barrera mucosa no es estática, y abrasiones
pueden remover pedazos de moco. Cuando el moco
toca soluciones con pH muy bajo, el moco precipita y
se muda, entonces, las células mucosas están
constantemente secretando moco. Hay 2 estímulos
que inducen la secreción de moco: la estimulación
vagal y la irritación (física o química) de la mucosa
gástrica por la comida ingerida. El modelo actual
sugiere que la estimulación vagal induce la liberación
de ACh, que produce un [Ca+2]i y estimula la
secreción. El cAMP no parece estar involucrado.
LAS CÉLULAS DE LA SUPERFICIE GÁSTRICA SECRETAN
HCO3- AL SER ESTIMULADAS POR ACH, ÁCIDOS Y PGS
Las células epiteliales superficiales del cuerpo y el
antro del estómago secretan HCO3-. Aunque la
secreción es poca, es muy importante, ya que es
parte del mecanismo de defensa mucosal gástrico. La
capa de gel mucosa es una capa sin movimiento, en
donde el HCO3- secretado permanece ahí y mantiene
un pH local de ~7,0 vs. Un pH intraluminal de 1 a 3. Un
cotransportador electrogénico Na/HCO3- media el
ingreso de HCO3- a través de la membrana basolateral
de las células epiteliales superficiales. Cómo sale el
HCO3- fuera de la célula hacia la capa mucosa apical
no es sabido, pero se cree que es vía un canal.
Al igual que con la secreción de moco, poco se sabe
sobre cómo se regula la secreción de HCO3-. El
modelo actual postula que la estimulación vagal
mediada por ACh hace que [Ca+2]i que a su vez
estimula la secreción de HCO3-. La alimentación en
falso es un estímulo potente de secreción de HCO3-
por esta vía. Un segundo estímulo potente para la
secreción de HCO3- es el ácido intraluminal.
Finalmente, hay evidencias que sugieren que un
factor humoral se encontraría implicado en la
inducción de la secreción de HCO3- por ácido.
EL MOCO PROTEGE AL EPITELIO DE LA SUPERFICIE
GÁSTRICA ATRAPANDO UN FLUIDO RICO EN HCO3- CERCA
DEL BORDE APICAL DE ESTAS CÉLULAS
Las células mucosas de la superficie estomacal y de
las glándulas gástricas secretan moco y HCO3-. Pero,
¿Qué es lo que hace a esta barrera tan efectiva?
Primero, el moco secretado forma una capa mucosa
de gel que es relativamente impermeable a la
difusión de H+ desde el lumen gástrico hacia las
células. Segundo, bajo esta capa de moco hay un
microclima que contiene fluido con alto pH y alta
[HCO3-], que neutraliza a la mayoría del ácido que
difunde por la capa de moco. La integridad mucosal
es mantenida además por la PGE2, que estimula la
secreción de HCO3-.
En la profundidad de la glándula gástrica, donde no
hay una capa mucosa protegiendo las células
parietales, principales ni ECL, la impermeabilidad de
la misma barrera apical celular parece impedir el paso
de H+ aún incluso a valores de pH < 1. La paradoja de
cómo el HCl secretado por las células parietales
emerge de la glándula hacia el lumen puede
explicarse por un proceso conocido como digitación
viscosa. Ya que el líquido que secretan las glándulas
es extremadamente ácido y presumiblemente se
encuentra bajo presión, puede atravesar como un
túnel a través de la capa mucosa que cubre el orificio
de la glándula gástrica hacia la superficie del
estómago. Sin embargo, este chorro de líquido no
parece distribuirse a lateral, sino que emerge hacia la
superficie como un “dedo”, sin neutralizar entonces
el HCO3- en el microambiente entre la superficie
epitelial y la capa mucosa.
La capa de gel mucosa y la solución alcalina rica en
HCO3- atrapada protegen a las células epiteliales
superficiales no sólo del H+ sino también de la
pepsina. El moco per sé actúa como una barrera de
difusión de la pepsina. La alcalinidad de la solución
rica en HCO3- inactiva cualquier pepsina que logre
penetrar el moco. Recordemos que la pepsina se
inactiva reversiblemente a pH > ~3,5 e
irreversiblemente a pH > ~7,2, entonces, la capa
mucosa HCO3- tiene un rol importante en prevenir la
autodigestión de la mucosa gástrica.
LA ENTRADA DE ÁCIDO AL DUODENO INDUCE LA
LIBERACIÓN SE SECRETINA DESDE LAS CÉLULAS S,
GATILLANDO ENTONCES LA SECRECIÓN DE HCO3- POR EL
PÁNCREAS Y DUODENO, QUE, A SU VEZ, NEUTRALIZA EL
ÁCIDO GÁSTRICO
La regulación de la secreción de ácido gástrico
involucra procesos estimulantes e inhibitorios, y
también de neutralización del ácido gástrico que pasa
desde el estómago hacia el duodeno. La cantidad de
ácido gástrico secretada se refleja en un descenso en
el pH intragástrico. Esto hace que las células D

Sebastián Lavanderos B. 2do. Medicina UDP
10
antrales liberen SS gatilladas por esta baja en el pH
para inhibir la secreción de más ácido, completando
un loop de feedback negativo. De la misma manera,
un bajo pH en el duodeno sirve como señal para la
secreción de álcalis para neutralizar esto.
El factor clave en este proceso de neutralización es la
secretina (la misma que inhibe la secreción de ácido
gástrico y promueve la secreción de pepsinógeno por
las células principales). Un pH duodenal bajo, con un
umbral de 4,5; gatilla la liberación de secretina desde
las células S duodenales. Sin embargo, estas células
no son sensibles a pH, pero responden a señales
emitidas por células que sí lo son. Altas tasas de
secreción de ácido gástrico gatillan la liberación de
altas cantidades de secretina, que estimula al
páncreas para que libere HCO3-, para que, a su vez,
neutralice la carga ácida en el duodeno.
Además del HCO3- pancreático, la carga ácida
duodenal se ve parcialmente neutralizada por la
secreción de HCO3- duodenal. Esta secreción ocurre
en la parte proximal pero no distal del duodeno, bajo
la influencia de prostaglandinas. Esta secreción
involucra un intercambiador Cl- HCO3 y el canal CFTR
apical. Pacientes con úlceras duodenales tienden a
tener secreción de ácido gástrico aumentada y
secreción duodenal de HCO3- disminuida. Entonces, la
carga ácida aumentada en el duodeno es sólo
parcialmente neutralizada, por lo que la mucosa
duodenal se ve expuesta en demasía a una solución
de bajo pH.
LLENADO Y VACIAMIENTO DEL ESTÓMAGO
LA ACTIVIDAD MOTORA GÁSTRICA PARTICIPA EN EL
LLENADO, MEZCLA Y VACIAMIENTO
La actividad motora gástrica tiene 3 funciones.
Primero, al recibir el material ingerido se representa
la función de reservorio del estómago, y ocurre
cuando se relaja el músculo liso. Esto ocurre
principalmente en la porción proximal del estómago
(en el fundus). Segundo, el material ingerido es
mezclado, y modificado entonces a una forma que le
permite salir rápidamente del estómago a través del
píloro hacia el duodeno, y facilita también la
digestión y absorción yeyunal normal. Entonces,
junto a las enzimas y ácido gástrico, la actividad
motora del estómago ayuda a iniciar la digestión.
Tercero, el antro pilórico, el píloro y la parte proximal
del duodeno funcionan como unidad para vaciar el
quimo (consistente en los contenidos gástricos
modificados, que son la comida parcialmente
digerida y las secreciones gástricas) hacia el
duodeno. El llenado y vaciamiento gástrico son
alcanzados por la actividad coordinada del músculo
liso en el esófago, LES y porciones proximal y distal
del estómago, así como en el píloro y duodeno.
El patrón de actividad muscular lisa es distinto en los
estados de ayuno y alimentación. Durante el ayuno se
conoce como MMC, conectado al intestino delgado.
Este patrón se termina al comer, donde se reemplaza
por el llamado patrón alimentado. Justo como las
regiones proximal y distal del estómago difieren en
su función secretora, también difieren en su función
motora, responsable de almacenar, procesar y vaciar
líquidos y sólidos. La parte proximal del estómago es
el lugar principal donde se almacenan líquidos y
sólidos. La parte distal es el lugar donde se agitan los
sólidos y se genera un material más pequeño y fluido,
que sale del estómago como los líquidos. Entonces, el
vaciamiento gástrico de líquidos y sólidos se
encuentra muy integrado.
EL LLENADO DEL ESTÓMAGO SE FACILITA POR LA
RELAJACIÓN RECEPTIVA Y LA ACOMODACIÓN GÁSTRICA
Incluso el tragar sin tener nada en la boca relaja el
LES y la parte proximal del estómago. Obviamente lo
mismo sucede cuando tragamos comida. La
relajación en el fundus se regula principalmente vía
un reflejo vagovagal y se le ha llamado relajación
receptiva. El volumen intragástrico aumenta
entonces sin que aumente la presión intragástrica.
Además de la relajación receptiva del estómago, que
lo prepara para la llegada de comida después de
tragar, el estómago se puede relajar también en
respuesta al llenado gástrico per sé. Entonces, el
aumento del volumen intragástrico por comida o

Sebastián Lavanderos B. 2do. Medicina UDP
11
secreción gástrica no produce entonces un aumento
marcado en la presión intragástrica. En su lugar,
pequeños aumentos en el volumen no causan
aumentos en la presión intragástrica hasta que se
alcanza un umbral, luego del cual la presión
intragástrica aumenta marcadamente. Esta dilatación
activa del fundus ha sido llamada acomodación
gástrica. Una vagotomía abole este fenómeno en su
mayoría, por lo que aumentos en el volumen
intragástrico producen mayores aumentos en la
presión intragástrica. Sin embargo, el rol del vago es
de modulación, el SNE es el regulador principal que
permite el almacenamiento de los sólidos y líquidos
en la parte proximal del estómago sin que aumente la
presión intragástrica.
EL ESTÓMAGO AGITA SUS CONTENIDOS HASTA QUE LAS
PARTÍCULAS SON LO SUFICIENTEMENTE PEQUEÑAS PARA
SER VACIADAS GRADUALMENTE AL DUODENO
La solución salina isotónica y el agua son las
sustancias vaciadas más rápido por el estómago. El
vaciamiento de estos líquidos ocurre casi sin retraso y
es más rápido a mayor volumen de fluido. Los fluidos
ácidos y los con alto contenido calórico dejan el
estómago más lentamente, mientras que los
materiales grasosos lo dejan aún más lentamente.
Los sólidos no salen del estómago (en su mayoría).
Primero tienen que ser reducidos en su tamaño a
menos de 2 mm, ya que partículas de más de 2 mm
no pueden pasar a través del píloro en el periodo
digestivo postprandial inmediato. Al ser reducidos de
tamaño, dejan el estómago como los líquidos.
El movimiento de sólidos a través del antro se
alcanza por la interacción entre contracciones
gástricas propulsivas y la oclusión del píloro, proceso
llamado propulsión. Las contracciones gástricas son
iniciadas por el marcapasos gástrico, que se
encuentra en la curvatura mayor. Estas contracciones
propulsan los contenidos luminales a través del
píloro, que se encuentra parcialmente cerrado por la
contracción de la musculatura pilórica antes de la
llegada del bolo. Una vez que el bolo llega cerca del
antro, es agitado para ayudar a que se reduzca el
tamaño de sus partículas, proceso llamado molienda.
Sólo una pequeña porción del material gástrico, que
contiene partículas más pequeñas que 2 mm, pasa a
través del píloro hacia el duodeno. De esta manera, la
mayoría de los contenidos gástricos se devuelven al
cuerpo del estómago para la pulverización y esquileo
de sus partículas sólidas, proceso conocido como
retropulsión. Estos 3 procesos se repiten hasta que el
estómago se vacía. Las partículas más grandes de 2
mm que no son digeridas se quedan en el estómago
pero son vaciadas al duodeno por los MMCs en el
periodo interdigestivo que empiezan ~2 horas
después de comer.
A medida que pequeños chorros de fluido gástrico
dejan el estómago, quimio y mecanorreceptores,
ubicados principalmente en la parte proximal pero
también distal del intestino delgado, sensan bajo pH,
alto contenido calórico, lipídico o algunos aás (como
el triptófano), o cambios en la osmolaridad. Todas
estas señales bajan la velocidad de vaciamiento
gástrico mediante una combinación de señales
neurales y hormonales, que incluyen al vago,
secretina, CCK y GIP liberado de la mucosa duodenal.