
Manejo del Burrito de la vid y otros curculiónidos ...
81
Transcript of Manejo del Burrito de la vid y otros curculiónidos ...


1

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
2

3
Boletín INIA Nº 260 • ISSN 0717-4829
INSTITUTO DE INVESTIGACIONES AGROPECUARIAS
MINISTERIO DE AGRICULTURA
Autores:
Paola Luppichini B., Andrés France I., Irina Urtubia H.,Natalia Olivares P. y Fernando Rodríguez A.
Manejo del Burrito de la vid, Naupactus xanthographus (Germar)
y otros curculiónidos asociados a vides

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
4
Manejo del Burrito de la vid, Naupactus xanthographus (Germar) y otros curculiónidos asociados a vides
Autores: Paola Luppichini B., Andrés France I., Irina Urtubia H., Natalia Olivares P.
y Fernando Rodríguez A. Editores: Paola Luppichini B., Natalia Olivares P. y Ernesto Cisternas A.
© Instituto de Investigaciones Agropecuarias, INIA, 2013 Ministerio de Agricultura Centro Regional de Investigación La Cruz
Chorrillos 86 Fono/Fax (56-33) 32 17 80 Casilla 3 La Cruz, Región de Valparaíso (V) Chile
Registro de Propiedad Intelectual:
ISSN: 0717-4829
Permitida la reproducción parcial o total de esta obra sólo con permiso previo y por escrito del Instituto de Investigaciones Agropecuarias, INIA.
Diseño e Impresión:
Versión | producciones gráficas Ltda. [email protected]•(56-2)2690798
Impreso en ChIle | prInted In ChIle
Advertencia: INIA y los autores no se responsabilizan por los resultados que se obtengan del uso o aplicación de productos genéricos o comercia-les que son mencionados. El texto es una guía de apoyo a los agricultores y profesionales, quienes deberán determinar los procedimientos y productos más adecuados a su situación particular.

5
Índice
Agradecimientos Introducción 7 1. Principales curculiónidos de importancia agrícola asociados a vides ....................................... 9 Paola luppichini B., natalia olivares P. y Fernando rodríguez a.
Burrito de la vid 10 Capachito de los frutales 14 Gusano blanco del frejol 17 Gusano de la raíz de la alfalfa 19 Gorgojo de los invernaderos 19 Gorgojo de la frutilla 22
2. Estrategia de manejo de curculiónidos asociados a vides .............. 25 Paola luppichini B. y natalia olivares P.
3. Uso de nemátodos entomopatógenos para el control de insectos ................................................................ 35 andrés France i.
4. Nemátodos entomopatógenos para el control de curculiónidos en vides ....................................... 49
Paola luppichini B., andrés France i. e irina urtubia H.
5. Formulación de Nemátodos Entomopatógenos .............................. 63 irina urtubia H.
Glosario de términos 73
Bibliografía consultada 75

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
6
Agradecimientos
Al personal técnico de laboratorio y campo de INIA (La Cruz y Quilamapu), Patricia Véliz R., José Montenegro M., Viviana Guajardo T. (Q.E.P.D) y Claudio Fernández, por su gran dedicación y perseverancia en el desempeño de su trabajo.
A la Sra. Eliana San Martín, Periodista de INIA La Cruz, por su apoyo en la gestión de los procesos necesarios para realizar la publicación.
A las empresas socias del proyecto que a través de su apoyo contribuyeron a la in-vestigación que originó esta publicación: Agrícola El Ensueño, Agrícola El Maitenal, Xilema S.A., Agrícola Orrego Dos S.A. y Desarrollo Agrario S.A.
A INNOVA Chile de CORFO entidad que hizo posible el desarrollo y edición de este material técnico que permite la difusión y transferencia de la tecnología del control biológico a través de nemátodos entomopatógenos.

7
Las especies de la familia Curculionidae, como Naupactus xanthographus (Bu-rrito de la vid), Naupactus cervinus (Capachito de los frutales) y Otiorhynchus sulcatus (gorgojo de la frutilla o capacho de los invernaderos) entre otros, son plagas de importancia económica, que en su estado larval se encuentran en el suelo consumiendo raíces y raicillas de un gran número de cultivos, dañando principalmente a vides y frutales menores (Prado 1991; Artigas 1994). En las raíces se alimentan de la corteza de éstas dejando galerías su-perficiales y a medida que las larvas incrementan su tamaño, las galerías se hacen más profundas y comprometen incluso el sistema vascular (xilema). La forma de alimentarse de estos insectos tanto en estado adulto como larva conlleva a una pérdida de vigor de la planta, reducción de la masa radicular y disminución en la cantidad y calidad de la fruta a cosechar (Artigas 1994; González 1989, Pérez 1994, Prado 1991, Ripa 1992). En el caso de N. xantho-graphus y también otros curculiónidos los adultos al alimentarse ocasionan un daño característico que corresponde a pequeñas mordeduras marginales en las hojas y yemas, con la consiguiente caída de hojas y racimos florales de la temporada.
A pesar que el control químico puede ser efectivo en el estado adulto del insecto, éste se ve limitando por los problemas de registro de productos en la fruta de exportación. Además la detección de individuos en la fruta tie-ne connotación cuarentenaria en el proceso de exportación a los principales destinos. La alternativa más utilizada en los últimos tiempos ha sido la Banda INIA, la cual requiere de gran cantidad de mano de obra para su colocación, por lo que muchas veces se evita su uso.
Una alternativa viable al uso de productos químicos es la utilización de enemigos naturales de la plaga como agentes de control biológico. Los
Introducción

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
8
nemátodos entomopatógenos son una de las mejores alternativas de control biológico para este tipo de plaga, ya que poseen la capacidad de buscar a su huésped en lugares donde otros organismos no tienen alternativas de encon-trar a su presa (Griffin et al. 2005, Cambell et al. 2003). Entre todos los nemá-todos que parasitan insectos, especial dedicación se ha dado al estudio de las familias Steinernematidae y Heterorhabditidae (Stock y Koppenhöfer 2003) las cuales se caracterizan por tener bacterias simbiontes del género Xenorhabdus y Photorhabdus, responsables estas últimas de la muerte de los insectos parasi-tados (Akhurst y Boemare 2005, Kaya et. al. 1993, Woodring y Kaya, 1988). Estas especies de nemátodos reúnen la mayoría de los atributos que debe tener un efectivo agente de control biológico: matan su huésped rápidamente (24 a 48 horas), son específicos para cada insecto plaga (Wang et al. 2006 ), parasitan a su huésped en lugares donde los productos químicos no son capa-ces de llegar (Bai et. al. 2005 ), son inocuos para vertebrados y plantas, fáciles de propagar y almacenar (Shapiro-Ilan et al. 2003), son capaces de buscar su huésped (Darthel et al. 2004) por si mismos, compatibles con insecticidas y no contaminan el medio ambiente (Bedding et al. 1993; Ishibashi y Kondo 1993, Bilgrami et al. 2006,; Kaya et al., 1993; Wouts 1991).
En nuestro país, se han realizado diferentes evaluaciones experimen-tales desde la colección de entomopatógenos de INIA y se han seleccionado aislamientos para el control de importantes insectos plaga de la zona Cen-tro-Sur y Sur de Chile, destacándose el control de Caliroa cerasi (chape del cerezo) (Alvarez et al. 1998), Naupactus=Asynonychus cervinus (capachito de los frutales) (Sandoval y France 1998), Otiorhynchus sulcatus (gorgojo de la frutilla) (Gerding y France, 1998) y Listronotus bonariensis (capachito ar-gentino de las ballicas) (Cisternas et al., 1998). Además, se han controlado larvas de hábito subterráneo con nemátodos entomopatógenos: Aegorhinus superciliosus (cabrito de la frambuesa) (Espinoza y France 1999), Hylamorpha elegans (pololo verde), Asynonychus cervinus (capachito de los frutales), Tipu-la apterogyne (zancudo patón del trigo) (France y otros 1999), Cydia pomonella (polilla de la manzana), Cydia molesta (polilla oriental de la fruta), Dalaca pallens (cuncunilla negra de las praderas) y Eumerus sp.

9
Principales curculiónidosde importancia agrícola
asociados a vides
Paola luppichini B., natalia olivares P. y Fernando rodríguez a.
INIA La Cruz
* Cuarentenaria: plaga que presenta restricción de ingreso en un país de destino. Primaria: plaga que de importancia económica y que requiere control prácticamente todos los años. Secundaria: Plaga generalmente se presenta en el cultivo y eventualmente sobrepasa el umbral de daño,
requiriendo control. Ocasional: Plaga que se presenta en algunos años y eventualmente requiere control. Literatura: Plaga citada como insecto o ácaro presente en el cultivo.
Principales curculiónidos asociados a vides
Nombre común Nombre científico Importancia*
Burrito de la vid Naupactus xanthographus Primaria (Fruit tree weevil) (Germar) y cuarentenaria
Capachito de los frutales Naupactus (Pantomorus=Asynonychus) Ocasional (Fuller rose weevil) cervinus (Boheman)
Gusano blanco del frejol Naupactus (Graphognathus) Ocasional (White-fringed weevil) leucoloma (Boheman)
Gusano de la raíz de la alfalfa Naupactus (Pantomorus) ruizi Literatura (Alfalfa root weevil) (Brètes)
Capachito de los invernaderos Otiorhynchus sulcatus Ocasional (Black vine weevil) (Goeze)
Gorgojo de la frutilla Otiorhynchus rugosostriatus Ocasional (Rough strawberry weevil) (Goeze)
Gorgojo verde de las chacras Platyaspistes glaucus Literatura (Vegetable green weevil) (Fahraeus)
Capacho desfoliador del pino Genoiocremmus chiliensis Literatura (Tuberous pine weevil) (Boheman)
Capacho del manzano Strangaliodes mutuarius Kuschel Literatura (Apple weevil)
1.

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
10
A continuación se describen los curculiónidos que se encuentran más fre-cuentemente asociados a las vides: Naupactus xanthographus, Naupactus cervi-nus, Naupactus leucoloma, Otiorhynchus sulcatus y Otiorhynchus rugosostriatus.
Burrito de la vid, Fruit tree weevilNaupactus xanthographus (Germar)
Importancia agrícola Es considerada una plaga primaria en vid y en frutales de exportación,
especialmente en uva de mesa. Es cuarentenaria para mercados de exporta-ción, principalmente para E.E.U.U.
Distribución geográfica Originario del cono sudamericano primariamente Argentina, Uruguay,
Paraguay y sur de Brasil. En Chile se encuentra desde la Región de Arica y Parinacota (XV) a la Región de la Araucanía (IX) e Isla de Pascua.
Descripción morfológica Huevos: color amarillo, miden entre 1 a 1,2 mm de largo (Parrella., et al., no publicado), son dispuestos en grupos de alrededor de 60 unidades adheridos entre si con una sustancia pegajosa (Foto 1).
Larvas: ápodas, color blanco, alcanzan 20 mm de longitud en el último esta-dio larvario (Foto 2). Aparato bucal masticador (Foto 3). Poseen en el último segmento abdominal cuatro bandas delgadas quitinizadas (Foto 4).
Adultos: miden entre 11 y 14 mm, el cuerpo tiene forma alargada, de color marrón a gris (Foto 5). Los ojos son hemiesféricos y sobresalientes. Poseen élitros soldados con dos líneas dorsales amarillentas que cubren el abdomen, siendo más anchos en la hembra que en el macho.
Ciclo de vidaEste insecto es de reproducción sexual. La hembra deposita los huevos
en lugares protegidos, principalmente en las grietas de la corteza. El período de incubación de los huevos varía de 32 días en verano y 42 en el otoño. La larva recién eclosionada (Foto 6) cae al suelo, se entierra y comienza a alimentarse de raicillas de malezas y en la medida que alcanza mayor desa-

11
Foto 1. Huevos de N. xanthographus.
Foto 3. Aparato bucal de larva de N. xanthographus.
Foto 5. Adulto de N. xanthographus.
Foto 2. Larva de N. xanthographus.
Foto 4. Último segmento abdominal de larva de N. xanthographus.
Principales curculiónidos de importancia agrícola asociados a vides
N. O
livar
es
N. O
livar
esR
. Rip
aR
. Rip
a
R. R
ipa

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
12
rrollo, consume las raicillas y raíces del frutal. El período de desarrollo de la larva puede durar alrededor de nueve meses, luego pupa bajo la superficie del suelo. Al cabo de un mes al estado de pupa (Foto 7), emerge el adulto que permanece en el interior de la celdilla durante un mes. Los adultos recién emergidos están provistos de ganchos mandibulares que les permiten emerger a la superficie del suelo (Fotos 8 y 9), dejando orificios de salida característi-cos. La emergencia ocurre a una temperatura de 13,5 ºC, distinguiéndose dos períodos, el primero de mayor intensidad durante la primavera y el segundo en el verano. La hembra inicia la oviposición 15 a 20 días después de emerger del suelo. El ciclo total dura entre 12 y 14 meses.
Foto 6. Larvas neonatas de N. xanthographus.
Foto 7. Pupa de Naupactus xanthographus.
Foto 8. Adulto recién emergido de N. xanthographus.
N. O
livar
es
P. L
uppi
chin
i
R. R
ipa

13
DañoEs el estado de larva el que causa el daño más importante, debido a
que se alimentan del sistema radicular de las vides, afectan la absorción de agua y nutrientes causando un deterioro progresivo de la planta y raíces de mayor grosor, en las que se observan galerías superficiales que comprometen la corteza y el cambium.
Los adultos dañan brotes y hojas, este daño puede ser importante en plantaciones nuevas (Foto 10). Asimismo, la detección de individuos en la fruta tiene una connotación cuarentenaria en el proceso de exportación a los principales destinos.
Hospederos Frutales: vides, chirimoyo, ciruelo, duraznero, guindo, níspero, nogal, peral, kiwi, naranjo, limonero, palto, arándano, caqui, frambueso.
Foto 9. Cabeza de adulto recién emergido de N. xanthographus con ganchos mandibulares.
Principales curculiónidos de importancia agrícola asociados a vides
R. R
ipa

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
14
Malezas: maicillo, diente de león, achicoria, llantén, y Rumex sp.
Cultivos: alfalfa, remolacha, papa y poroto, entre otros.
Capachito de los frutales, Fuller rose weevil Naupactus (= Asynonychus = Pantomorus) cervinus (Boheman)
Importancia agrícola En vides está descrita como una plaga ocasional
Distribución geográficaCosmopolita, presente en EE.UU., Australia, Europa, Nueva Zelanda,
Sudamérica. En Chile se encuentra entre las Regiones de Atacama (III) y De Los Lagos (X) y en la Región de Arica y Parinacota (XV), Isla de Pascua y Juan Fernández.
Descripción morfológicaHuevos: son ovoides, aplanados y de color blanco cremoso miden 0,9 a 1
Foto 10. Daño del adulto de N. xanthographus en hojas de vid.
R. R
ipa

15
Foto 11. huevos de N. cervinus.
Foto 13. Aparato bucal de larva de N. cervinus.
Foto 15. Adulto de N. cervinus.
Foto 12. Larva de N. cervinus.
Foto 14. Último segmento abdominal de larva de N. cervinus.
Principales curculiónidos de importancia agrícola asociados a vides
R. R
ipa
N. O
livar
esR
. Rip
a
R. R
ipa
R. R
ipa

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
16
mm de largo (Parrella., et al., no publicado), dispuestos en masas de 30 a 60 unidades (Foto 11).
Larvas: ápodas y tiene forma curvada. Una vez que alcanzan mayor desarro-llo pueden llegar a medir hasta 1 cm de largo (Foto 12). Poseen un aparato bucal masticador prominente (Foto 13). No presentan bandas quitinizadas en el último segmento abdominal (Foto 14).
Adultos: miden entre 5,5 y 7,5 mm de largo (Foto 15). Cuerpo de forma ovalada de color pardo grisáceo, élitros soldados y con estrías cuyos bordes presentan franjas oblicuas cortas de escamas blancas. Ojos ovales y sobresa-lientes.
Ciclo de vidaEste insecto es partenogenético. Los huevos son depositados en masas
de hasta 60 unidades en grietas de la corteza en los troncos, en hojas secas enrolladas, bajo amarras y en cítricos bajo los sépalos de los frutos. Las larvas recién eclosadas penetran al suelo para alimentarse de raíces. El período lar-vario dura alrededor de 4 meses en condiciones de laboratorio (25±2 ºC y 60% HR). Posteriormente, pasan al estado de pupa (Foto 16) y luego emergen los adultos, estos salen desde el suelo y suben por el tronco para alimentarse del follaje y oviponer. El período de emergencia de los adultos ocurre finali-zando el verano hasta el otoño. El ciclo desde huevo a larva en condiciones de laboratorio dura alrededor de 5 meses.
Foto 16. Pupa de N. cervinus.
M. P
arre
lla

17
DañoEl principal daño en frutales está asociado a las larvas, las cuales con-
sumen las raíces y raicillas produciendo una reducción del volumen de raíces lo que ocasiona problemas de absorción de agua y nutrientes, lo que puede manifestarse en un menor crecimiento y vigor de la planta. Los adultos oca-sionan daño en el follaje consumiendo los bordes de las hojas.
En el caso de los cítricos además de los daños mencionados, la presen-cia de huevos bajo los sépalos del fruto es causal de rechazo en la fruta de ex-portación, debido a la apariencia similar con los huevos de N. xanthographus.
HospederosFrutales: naranjo, limonero, mandarino, palto, duraznero, damasco, manza-no, banano, palto, frambueso, frutilla, grosellero, vides, y zarzaparrilla.
Cultivos: hortalizas, papa, maíz, remolacha, poroto, feijoa, trébol rosado y blanco, alfalfa.
Flores: zinnia y rosal.
Gusano blanco del frejol, White-fringed weevil Naupactus leucoloma (Boheman)
Distribución geográfica Se encuentra en Argentina, Australia, Nueva Zelanda, Uruguay, E.E.
U. U. y Brasil. En Chile ha sido reportado desde la Región de Arica y Parina-cota (XV) hasta la Región De Los Lagos (X).
Descripción morfológica Huevos: son blancos, puestos en la superficie del suelo agrupados en masas de 20 a 60 huevos, cubiertos con partículas de tierra muy resistentes a las condiciones ambientales extremas, pudiendo permanecer como tales varios meses.
Larvas: miden 15 mm de largo en su estado maduro, color crema (Foto 17), ápodas, cabeza casi totalmente cubierta por el pronoto sólo sobresalen las mandíbulas de color café.
Principales curculiónidos de importancia agrícola asociados a vides

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
18
Adultos: Sólo existen hembras de 1 a 1,5 cm, las que se reproducen por par-tenogénesis. Ojos sobresalientes, ovalados, cabeza y frente con surco medial. Elitros con pilosidad erecta. Color gris con bandas claras (Foto 18).
Foto 17. Larva de Naupactus leucoloma. Foto 18. Adulto de N. leucoloma.
Ciclo de vida Esta especie desarrolla su ciclo en un período entre 12 y 22 meses. Los
adultos viven varios meses, oviponiendo sobre 1000 huevos dependiendo de su hospedero. La larva recién nacida se entierra unos 20 cm y permanece alimentándose de raíces entre 8 a 20 meses. La larva presenta gran movilidad vertical y horizontal en el suelo.
DañoRegularmente en el suelo se encuentran larvas de distintos tamaños.
La corteza de la corona de la planta y raíces principales son las más afectadas por las larvas. El adulto consume follaje. La mayor incidencia de este insecto se constata cuando el cultivo anterior ha sido leguminosas o existen cubiertas vegetales entre o sobre hileras.
E. C
iste
rnas
E. C
iste
rnas

19
Gusano de la raíz de la alfalfa, Alfalfa root weevilNaupactus ruizi (Brètes)
Distribución geográficaPresente en Argentina. En Chile se encuentra distribuido entre las
Regiones de Antofagasta (II) y De Los Lagos (X).
Descripción morfológicaAdultos: cuerpo largo, ovalado, cubierto de escamas grises. Miden entre 8,5 a 10 mm de largo. Los élitros cubren todo el abdomen formando en el ápice una punta.
Larvas: ápodas, ligeramente más pequeñas que las de N. xanthographus y muy similares en forma.
Ciclo de vidaSimilar a N. xanthographus.
DañoLas larvas consumen y perforan las raíces de los hospederos. En alfalfa
se ha observado a los adultos consumiendo hojas, tallos y brotes.
Gorgojo de los invernaderos, Black vine weevilOtiorhynchus sulcatus (Fabricius)
Importancia agrícola En vides es considerada una plaga ocasional, asociada a vivero.
Distribución geográficaPresente en Europa y Norteamérica. En Chile se encuentra entre las
Regiones de Valparaíso (V) y De Los Lagos (X).
Descripción morfológicaHuevos: inicialmente son blancos y en la medida que maduran se vuelven color marrón.
Principales curculiónidos de importancia agrícola asociados a vides

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
20
Larvas: son de color blanco (Foto 19), ápodas, con una forma similar a una “C”, cabeza color marrón (Foto 20). Miden 10 a 15 mm de largo en su estado maduro. En el último segmento abdominal se observan algunas setas cauda-les. (Foto 21).
Adultos: miden entre 8 y 11 mm de largo. Cuerpo color negro con pequeñas manchas amarillas, en los élitros (Foto 22). La parte posterior del cuerpo ter-mina en forma de punta redondeada. Ojos poco convexos. Hábito nocturno.
Foto 19. Larva de Otiorhyncus sulcatus.
Foto 21. Último segmento abdominal de larva de O. sulcatus.
Foto 20. Cápsula cefálica de larva de O. sulcatus.
Foto 22. Adulto de Otiorhyncus sulcatus.
R. R
ipa
R. R
ipa
R. R
ipa
R. R
ipa

21
Ciclo de vidaInsecto partenogenético. Los adultos depositan los huevos sobre el sue-
lo en forma aislada cerca de las raíces de las plantas (sobre 2000 huevos por hembra), de ellos eclosionan las larvas después de 15 a 30 días y comienzan a alimentarse de las raíces. Este insecto inverna como adulto bajo el suelo. El desarrollo de la pupa (Foto 23) ocurre cerca de la primavera y luego emergen los primeros adultos, los cuales comienzan a alimentarse de hojas para comen-zar a oviponer 21 a 28 días después. El ciclo de este insecto desde huevo a adulto dura aproximadamente 12 meses.
Foto 23. Pupa de Otiorhyncus sulcatus.
DañoEs de hábito polífago. Las larvas de este insecto son las que causan
el mayor daño, consumiendo raicillas y anillando las raíces principales. En huertos con presencia de abundantes poblaciones provoca la muerte de las plantas. Los adultos consumen follaje, produciendo muescas en el borde de la hoja, defoliación y enroscamiento en algunos brotes. En vides ha sido repor-tada atacando plantas en vivero.
HospederosFrutales: arándano, frambuesa, frutilla y vid.
Otros: leguminosas forrajeras, ornamentales.
Principales curculiónidos de importancia agrícola asociados a vides
R. R
ipa

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
22
Gorgojo de la frutilla, Rouge strawberry weevilOtiorhynchus rugosostriatus (Goeze)
Distribución geográficaEsta especie es originaria de Europa y se encuentra presente en Nue-
va Zelanda, Australia y América del Norte. En Chile se encuentra entre la Región de Coquimbo (IV) y De Los Lagos (X), además del Archipiélago de Juan Fernández.
Descripción morfológica Larvas: Color blanco casi traslúcido, ápodas, miden hasta 7 mm de largo, con la cabeza totalmente visible de color café rojizo (Foto 24), las mandíbulas son negras.
Adultos: La totalidad de los individuos son hembras. Miden entre 5,5 y 7 mm de largo, ojos muy poco convexos, casi aplanados. Cuerpo color marrón con apariencia granulosa (Foto 25).
Foto 24. Larva de Otiorhyncus rugosostriatus.
E. C
iste
rnas

23
Foto 25. Adulto de Otiorhyncus rugosostriatus.
Ciclo de vida El insecto bajo condiciones de campo pasa el invierno principalmente
como larva bajo el suelo y en menor medida como adulto hibernante bajo la hojarasca en el suelo. Desarrolla su ciclo en un año. Los huevos son puestos individualmente sobre el suelo entre la hojarasca. Los adultos se pueden de-tectar en la noche, pues en el día se esconden en los primeros centímetros del suelo y bajo las malezas.
Daño Las larvas pasan el invierno alimentándose de raicillas y la corteza de
raíces mayores, produciendo áreas superficiales irregulares sin corteza, lo que en raíces pequeñas se traduce en anillado y muerte de ellas y en raíces grue-sas sitios de entrada de enfermedades. Se alimentan del follaje, produciendo marcas características en los bordes, en forma de semicírculo.
Principales curculiónidos de importancia agrícola asociados a vides
E. C
iste
rnas

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
24

25
El manejo de los curculiónidos de importancia agrícola en frutales puede rea-lizarse utilizando una estrategia que puede incluir diversos métodos como químico, cultural y biológico, este último puede ser a través de artrópodos benéficos o microorganismos como los Hongos Entomopatógenos (HEP) y/o Nemátodos Entomopatógenos (NEP).
Control químicoDebido a la sincronización del ciclo de vida que presenta este insecto,
se han identificado 2 períodos de emergencia de los adultos. Claramente, la primera emergencia presenta una mayor abundancia y el daño en la vid se asocia al crecimiento de brotes. Es por ello que, el control tradicional de Naupactus xanthographus, considerado el curculiónido de mayor importancia en el cultivo de la vid, se ha focalizado en el control de los adultos, a través de dos métodos:
1. Barrera tóxica. Consiste en la colocación de una cinta plástica untada con una pasta insecticida (Pasta INIA 82.4) que causa mortalidad al insecto cuando este camina sobre ella. Se coloca una franja de polietileno de 15 a 20 cm. de ancho, enrollada en cada tronco y tutor (Foto 26) a una altura de 1 a 1,2 m sobre el nivel del suelo. Posteriormente se unta la mitad superior del plástico con una delgada capa de la pasta insecticida, empleando una brocha. En parronales se debe incluir la protección de los cabezales y tirantes. En ge-neral para frutales, si se usan tutores, éstos también deben usar barrera.
El momento de instalación de la barrera corresponde al período previo de la emergencia de los adultos desde el suelo (fines de agosto en la zona cen-tral, temperatura de suelo 13,5 ºC). La barrera pierde efectividad gradualmen-te, lo que se detecta al observar adultos en el follaje, lo cual indica la necesidad de reactivarla, aplicando pasta en la mitad inferior de la banda plástica.
Estrategia de manejo de curculiónidos asociados a vides
Paola luppichini B. y natalia olivares P.
INIA La Cruz
2.

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
26
Para el óptimo funcionamiento de este método, es necesario eliminar las malezas que sobrepasen la barrera, los chupones o ramas que estén en contacto con el suelo o tronco. Como en toda aplicación de insecticida, de-ben tomarse todas las medidas de protección para evitar intoxicación (guan-tes, mascarilla, traje, etc).
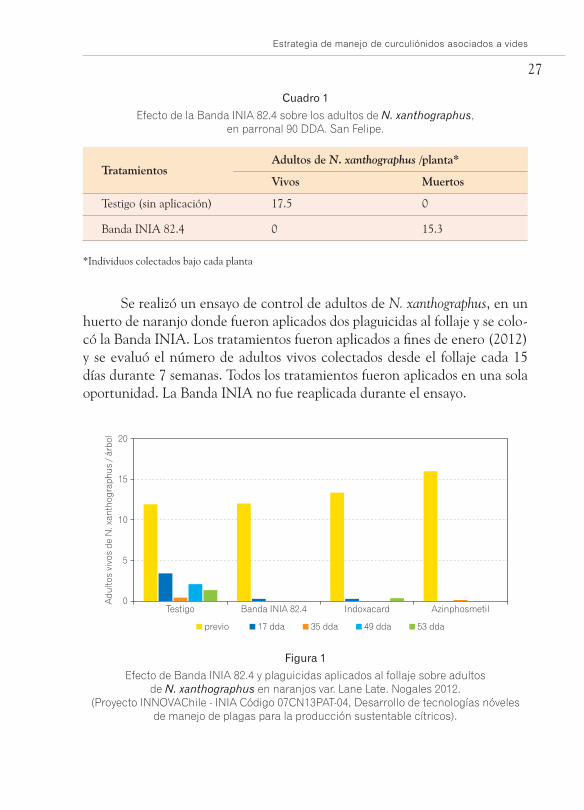
En el cuadro 1 se observa el efecto de la Banda INIA 82.4, sobre adul-tos de N. xanthographus presentes en un parronal de la zona de San Felipe. Se evaluaron los adultos vivos y muertos encontrados en cada planta. En las plantas tratadas con Banda INIA 82.4 se encontró una alta mortalidad de adultos de N. xanthographus. Después de 90 días de iniciado el tratamiento. Asimismo, es posible evidenciar que la población testigo (sin aplicaciones) continúa su desarrollo durante la temporada.
Foto 26. Vista de parronal con Banda INIA 82.4.
R. R
ipa
27
Figura 1
Efecto de Banda INIA 82.4 y plaguicidas aplicados al follaje sobre adultos de N. xanthographus en naranjos var. Lane Late. Nogales 2012.
(Proyecto INNOVAChile - INIA Código 07CN13PAT-04, Desarrollo de tecnologías nóveles de manejo de plagas para la producción sustentable cítricos).
estrategia de manejo de curculiónidos asociados a vides
Testigo0
5
10
15
20
previo
Banda INIA 82.4
Adu
ltos
vivo
s de
N. x
anth
ogra
phus
/ ár
bol
Indoxacard Azinphosmetil
17 dda 35 dda 49 dda 53 dda
Cuadro 1
Efecto de la Banda INIA 82.4 sobre los adultos de N. xanthographus, en parronal 90 DDA. San Felipe.
Tratamientos Adultos de N. xanthographus /planta*
Vivos Muertos
Testigo (sin aplicación) 17.5 0
Banda INIA 82.4 0 15.3
* Individuos colectados bajo cada planta
Se realizó un ensayo de control de adultos de N. xanthographus, en un huerto de naranjo donde fueron aplicados dos plaguicidas al follaje y se colo-có la Banda INIA. Los tratamientos fueron aplicados a fines de enero (2012) y se evaluó el número de adultos vivos colectados desde el follaje cada 15 días durante 7 semanas. Todos los tratamientos fueron aplicados en una sola oportunidad. La Banda INIA no fue reaplicada durante el ensayo.

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
28
Se observa una disminución consistente en el tiempo de los adultos de N. xanthographus en el follaje de los cítricos, destacando el tratamiento con la Banda donde se llegó a nivel cero. En los otros tratamientos se observó presencia de 0,3 individuos por árbol (Figura 1).
2. Tratamiento al follaje. Las aplicaciones químicas deben iniciarse en con-junto con la emergencia de los adultos, evitando el daño hacia los brotes. Dependiendo de la variedad de vid, en ocasiones el daño conlleva pérdidas importantes en rendimiento. En general, el umbral de acción estimado según juicio experto en vides es de 1 individuo por planta.
En vides, la efectividad de los plaguicidas en general es de corta du-ración, debido al crecimiento vegetativo de la temporada que genera nuevas hojas durante un largo período, lo cual provee al insecto de hojas libres de residuos del insecticida. Además los insecticidas van perdiendo el efecto de contacto en el tiempo, con la consiguiente disminución de efectividad.
Lo anterior es importante, considerando el extenso período de emer-gencia de los adultos desde el suelo (2 a 3 meses). Por esta razón las aplica-ciones deben ser realizadas para evitar el daño en el cultivo por efecto de la alimentación de los insectos. De la misma forma se evitará la oviposición de los adultos, que ocurre entre 15 a 20 días después de emergido desde el suelo, evitando el desarrollo de una nueva generación.
En la figura 2 se muestra el efecto de diferentes plaguicidas sobre adul-tos de burrito de la vid, en naranjos. Se observa, un efecto de control impor-tante sobre N. xanthographus con una sola aplicación de plaguicida. Tanto azinphosmetil como indoxacarb, controlan la plaga llegando a un nivel cer-cano a cero.
3. Larvas. El hábito subterráneo de las larvas y su ubicación estacional bajo 20 a 40 cm en el perfil del suelo, restringe la eficacia el efecto de los insecti-cidas aplicados con métodos tradicionales. En esas condiciones, los insecti-cidas no son efectivos principalmente por la fijación a los coloides y materia orgánica del suelo además de la degradación química y microbiológica. Bajo este escenario, el uso de microorganismos entomopatógenos como hongos y nemátodos, surgen como una alternativa que está siendo utilizada en diversos cultivos afectados por plagas que realizan parte de su ciclo biológico en el sue-lo (desarrollo de larvas, pupa y adulto). Tanto los hongos como los nemátodos

29
entomopatógenos (NEP) están ampliamente distribuidos en el mundo y son considerados inocuos para el medio ambiente.
El control de larvas de curculiónidos asociados a vides y otros frutales con NEP, será abordado con detalle en el capítulo 4.
Control cultural Recolección manual La colecta directa de los adultos en las vides se puede realizar fácilmen-
te en huertos de hasta dos años y en los mayores hasta brotes de 15 cm, debido a que en este estado de desarrollo aún es posible visualizar los adultos en el escaso follaje. En plantas más desarrolladas la recolección debe realizarse colo-cando sobre el suelo una lámina de polietileno de 3 x 1,5 m a ambos lados de la planta, luego con mazo de goma se golpea los brazos principales de las plantas y se colectan los adultos que caen al sobre una lámina de polietileno.
En viñedos principalmente orgánicos y biodinámicos, se realiza la co-lecta manual de los adultos y en algunos casos además se utilizan aves (galli-nas y gallinetas) como depredadores de los adultos que caen al suelo.
Figura 2
Efecto de plaguicidas aplicados al follaje sobre adultos de N. xanthographus en naranjos var. Lane Late. Nogales 2012.
(Proyecto INNOVA Chile - INIA Código 07CN13PAT-04, Desarrollo de tecnologías nóveles de manejo de plagas para la producción sustentable cítricos).
estrategia de manejo de curculiónidos asociados a vides
Phosmet Azinphosmethil Methidation Indoxacarb Testigo0,0
0,5
1,0
1,5
2,0
2,5
previo
Adu
ltos
vivo
s de
N. x
anth
ogra
phus
/ ár
bol
19 dda 33 dda 47 dda 60 dda

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
30
Control biológico Asociado a artrópodosPara los curculiónidos N. xanthographus y N. cervinus se han encontra-
do algunos enemigos naturales asociados, parasitando y/o depredando adul-tos y huevos.
ParasitoidesCentistes sp, (Hymenoptera: Braconidae), es una pequeña avispa. Para-
sitoide de adultos, muy poco frecuente (Foto 27).
Fidiobia asina (Hymenoptera: Platygastridae), parasitoide de huevos de N. xanthographus. En huertos con pocas aplicaciones de plaguicidas es fre-cuente observar huevos parasitados, bajo el ritidomo, los cuales se reconocen porque son de color marrón dorado (Foto 28).
Depredadores Megatoma sp., (Coleoptera: Dermestidae). La larva (Foto 29) depreda
los huevos de N. xanthographus.
Foto 27. Capullo de Centistes sp. Foto 28. Huevos de N. xanthographus parasitados por Fidiobia asina.
R. R
ipa
N. O
livar
es

31
Gryllus fulvipennis, (Orthoptera: Gryllidae). Insecto polífago (Foto 30). Depreda adultos de N. xanthographus y N. cervinus.
Foto 29. Larva de Dermestido.
Foto 30. Adulto de grillo de campo.
Asociado a microorganismos
Hongos Entomopatógenos (HEP)Los hongos entomopatógenos (HEP) son hongos que causan enferme-
dades en insectos y ácaros, lo que permite controlar plagas sin dejar residuos
estrategia de manejo de curculiónidos asociados a vides
R. R
ipa
R. R
ipa

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
32
y con un muy bajo impacto en el medioambiente y las personas. Los HEP afectan la capacidad de alimentarse de insectos y ácaros, por lo que finalmen-te mueren. Además su acción es muy específica ya que atacan a una especie o a grupos de especies muy relacionadas entre si, sin afectar a las especies benéficas. Los hongos entomopatógenos invaden a los insectos penetrando su cutícula mediante procesos enzimáticos. Una vez en el interior del insecto la colonia de hongos se desarrolla y produce toxinas que provocan la muerte del mismo. En la actualidad el banco de microorganismos del INIA contiene más de 1.800 cepas de hongos entomopatógenos (HEP), de los géneros Me-tarhizium y Beauveria, los cuales se utilizan en todo el mundo para el control de numerosas plagas.
En el caso de los HEP se formulan como esporas deshidratadas, pe-llets y líquidos emulsionables. La colección del INIA cuenta con cepas para el control de las siguientes especies plaga: Naupactus xanthographus (Foto 31), Naupactus (Asynonychus) cervinus, Naupactus (Graphognatus) leucoloma, Otiorhynchus sulcatus, Otiorhynchus rugosostriatus, Sericoides vi-ridis, Hylamorpha elegans, Phytoloema herrmanni y Aegorhinus supeciliosus, entre otros.
Foto 31. Larva de N. xanthographus afectada por Metarhizium sp.
R. R
ipa

33
Nemátodos Entomopatógenos (NEP)Los nematodos entomopatógenos (NEP) corresponden a gusanos mi-
croscópicos habitantes del suelo. Poseen la habilidad de alimentarse de insec-tos valiéndose para esto de una asociación con una bacteria simbionte de la cual se alimentan y que a su vez transportan e inoculan como una verdadera “inyección biológica”. Los géneros de nematodos entomopatógenos más utili-zados en el control biológico de insectos son Steinernema, del cual se conocen 24 especies, y Heterorhabditis, con ocho especies conocidas.
En el siguiente capítulo (3) se describe ampliamente el uso de los NEP como controladores biológicos de plagas.
estrategia de manejo de curculiónidos asociados a vides

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
34

35
El Control Biológico de plagas de importancia agrícola está basado en el control tradicional, esencialmente de plagas aéreas y con el uso de insectos benéficos. A pesar que esta disciplina tiene más de 100 años de investiga-ción y desarrollo, poco se ha explorado el uso de la patología de insectos como parte del manejo integrado de plagas y, menos aún, el uso de nemáto-dos entomopatógenos (NEP). La primera referencia en Chile del uso de un
Foto 32. Larva de gusano blanco parasitada por nemátodos entomopatógenos (Steinernema sp.). El recuadro muestra el detalle de los NEP emergiendo por la boca.
Uso de nemátodos entomopatógenos para el control de insectos
andrés France i.
INIA Quilamapu
A. F
ranc
e
3.

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
36
NEP corresponde al trabajo del Dr. Dutky (1957), quien fue responsable de la introducción del nemátodo Steinernema carpocapasae strain DD-136, que en aquellos años se denominaba Neoaplectana carpocapsae, para el control de gusanos blancos en la zona de Temuco (Foto 32). Este trabajo pionero en el mundo, fue completamente olvidado por el exitoso desarrollo del DDT.
Los nemátodos han convivido con insectos por millones de años, pro-duciéndose todo tipo de relaciones que van desde las beneficiosas para los insectos, hasta las perjudiciales como son los parasitismos y patogénesis. En efecto, el record más antiguo de detección de nemátodos corresponde a un Mermitido, parásito común en las larvas de gusanos blancos (Foto 33), que fue encontrado en un insecto atrapado en ámbar y fechado en la época del Cretáceo temprano, ¡140 millones de años atrás! (Poinar y Poinar, 2008). Durante todo este tiempo, el proceso evolutivo terminó adaptando a los NEP para utilizar los insectos como substratos de alimentación, dispersión y propa-gación, características que han permitido su uso para el control biológico de insectos plagas. El estudio de este tipo de control ha permitido la selección de cepas que pueden ejercer controles muy efectivos y en condiciones que otros sistemas de control no son posibles.
Foto 33. Emergencia de un nemátodo Mermitido desde larvas de gusano blanco.
A. F
ranc
e

37
Doce familias de nemátodos se asocian con los insectos, algunas utili-zan a estos artrópodos como un medio de transporte sin causar daños, otros pueden producir parasitismo y otros pueden transmitir enfermedades; estos últimos se les denomina entomopatógenos. Dos familias destacan como NEP: Steinernematidae y Heterorhabditidae, ambas con ejemplos notables de aso-ciación simbióntica y control de insectos. Todos los nemátodos corresponden a gusanos cilíndricos no segmentados y en el caso de los entomopatógenos, se caracterizan además por portar bacterias que les serán de utilidad durante la patogénesis del insecto. Estas bacterias pertenecen a la familia Enterobacte-riacea, son Gramm negativas y anaeróbicas facultativas (Griffin et al., 2005). Se han reconocido dos géneros de bacterias simbiontes: Xenorhabdus y Pho-torhabdus, las cuales se asocian a las familias Steinernematidae y Heterorha-bditidae, respectivamente (Sicard et al., 2004; Tailliez et al., 2006; Emelianoff et al., 2007).
Los NEP presentan diferentes medios de adaptación a las condiciones de suelo y clima, entre las que se destacan su habilidad de moverse en el perfil del suelo, en busca de condiciones óptimas de sobrevivencia, de acuerdo a temperatura y humedad, o para buscar un hospedero siguiendo gradientes de excreción, CO2 o movimiento (Alekseev et al., 2006). Básicamente existen dos estrategias que usan los NEP para encontrar su huésped, la cual se deno-mina forrajeo. La primera es seguir un estímulo o rastro que deja el insecto dentro de un substrato hasta encontrarlos, este hábito se denomina “perse-guidor” y es útil para insectos poco móviles, que viven en galerías o cavidades en el suelo o en el interior de raíces. El segundo, es mantenerse quietos o nictando (mantenerse erguidos en la punta de la cola y oscilando) a la espera de que un insecto pase lo suficientemente cerca, para que ellos puedan saltar sobre su cuerpo, este tipo de forrajeo se denomina “emboscador”. Además, pueden presentar una estrategia de forrajeo intermedia, esto le permite para-sitar a diferentes tipos de hospederos tanto móviles como menos móviles en el perfil del suelo (Lewis and Clarke, 2012).
El ciclo de vida de cualquier especie de NEP consta de huevo, cuatro estados juveniles (J1 a J4) y adulto. Los nemátodos infectivos corresponden al estado juvenil J3, pero que mantienen la cutícula del estado J2, y que son denominados “dauers”. Sólo los dauers son capaces de localizar a su hospe-dero e iniciar el proceso de parasitismo de forma activa. Los dauers entran al
uso de nemátodos entomopatógenos para el control de insectos

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
38
insecto a través de sus aberturas naturales (boca, ano y espiráculos), el insec-to se defiende barriendo de la boca los nemátodos, regurgitando, prolapsando el intestino o encapsulando los nemátodos a medida que estos logran entrar por los espiráculos. Si las medidas de defensa no son suficientes, los NEP terminan ingresando al hemocele, donde regurgita la bacteria simbionte que portan en una vesícula cercana a la boca, luego ésta se multiplica y destruye tejidos y hemolinfa, ocasionan la muerte del insecto por una septicemia gene-ralizada (Ramos-Rodriguez et al., 2007; Lewis and Clarke, 2012).
Foto 34. Masiva emergencia de estadíos juveniles de nemátodos desde una larva de gusano blanco parasitada por NEP.
A. F
ranc
e

39
Una vez que el insecto muere, el nemátodo puede alimentarse de esta solución de hemolinfa y tejidos degradados ricos en células bacterianas, per-mitiendo que los juveniles muden a adultos y se reproduzcan en forma masiva (Foto 34). Xenorhabdus y Photorhabdus se caracterizan por destruir los tejidos internos del insecto, para crear un medio favorable para la alimentación y re-producción de los NEP, sin embargo estas bacterias no pueden vivir de forma libre, dependiendo totalmente de los NEP para ser transportadas e inocula-das en el substrato correcto, es decir el insecto (Griffin et al., 2005).
No solo se ha demostrado la efectividad de los NEP como parásitos obligados y portadores de enfermedades en una amplia variedad de insectos que habitan en el suelo, pero además el que estos organismos presenten la mayoría de los atributos que debe tener un efectivo agente de control bio-lógico: matan su huésped rápidamente, son específicos contra numerosos insectos plagas, inocuos para vertebrados y plantas, fáciles de propagar y al-macenar, capaces de buscar a su huésped por sí mismos, no contaminan el medio ambiente y se adaptan bien a un control integrado de plagas, (Griffin et al., 2005; Koppenhöfer, 2007; Lewis and Clarke, 2012). La característica de buscar por sus propios medios al insecto huésped, lo convierte en el único tipo de microorganismo que puede cumplir este tipo de función, permitiendo el control de plagas que se encuentran en sitios crípticos como el suelo o al interior de raíces o coronas de plantas (Foto 35).
Foto 35. Larvas de Curculiónidos alimentándose del interior de una corona de arándano.
uso de nemátodos entomopatógenos para el control de insectos
A. F
ranc
e

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
40
En Chile, el Programa de Patología de Insectos del Instituto de Inves-tigaciones Agropecuarias (INIA), ha estado prospectando en forma sistemá-tica el país en busca de nemátodos entomopatógenos, demostrándose la pre-sencia de los géneros Steinernema y Heterorhabditis y de especies únicas en el mundo (Edgington et al., 2010), constituyéndose en una base biológica para ser usados en el control biológico de insectos plagas de importancia econó-mica. La tendencia de estos organismos en el país es a presentarse con mayor abundancia hacia el extremo sur (Figura 3), indicando ciertas adaptaciones o preferencias climas más fríos.
Figura 3
Distribución de aislamientos de NEP a lo largo de Chile.
Ensayos de control con NEP chilenos han sido realizados principal-mente en Curculiónidos, debido a que varias especies de estos insectos afec-tan de manera importante diferentes frutales de la zona centro sur y sur de Chile. Entre ellos se destacan el cabrito o marinerito (Aegorhinus supercilio-sus), capachito de los frutales (Naupactus cervinus) y el gorgojo de los inver-naderos (Otiorhynchus sulcatus). Para este tipo de plagas los NEP son una de las mejores alternativas de control biológico, ya que poseen la capacidad de
Arica Antofagasta Valparaíso Concepción Temuco Los Choros Patagonia0
5
10
15
20
25
30
35
Prop
orci
ón d
e N
EP (%
)

41
buscar a su huésped, en lugares donde otros organismos no tienen alternati-vas de encontrar a su presa.
Los resultados han permitido demostrar niveles de control de hasta un 75% en la emergencia de adultos. El Gorgojo de los invernaderos es una plaga que consume raicillas y anilla raíces más gruesas de diferentes especies frutales, particularmente es importante en arándanos donde las cantidades de larvas en el suelo pueden alcanzar grandes números, dado que todos los in-dividuos son hembras y muy prolíficas. Ensayos realizados en huertos comer-ciales de arándanos muestran que los NEP logran disminuir la emergencia de adultos hasta 75% (Figura 4). Se ha demostrado que la presencia de NEP en el suelo disminuye sostenidamente la emergencia de adultos, como este in-secto tiene un período en el cual la emergencia es mayor, si esta no ocurre en la fecha apropiada existen menos posibilidades para que la plaga sea exitosa.
Figura 4
Efecto de las aplicaciones al suelo de nemátodos entomopatógenos (NEP) en la emergencia de adultos Otiorhynchus sulcatus Huerto de arándanos
ubicado en Purranque.
uso de nemátodos entomopatógenos para el control de insectos
0 20 40 60
Días
80 1000
10
20
30
40
50
60
70
80
N° a
dulto
s/m
2
Con NEP Testigo

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
42
En las plagas de suelo no siempre es factible evaluar directamente lo que está ocurriendo con la mortalidad de larvas (Foto 36), debido a que ésta ocurre bajo el suelo, pero si es posible en forma indirecta, mediante la emer-gencia de adultos u otro estadío que sea visible sobre el suelo. También es importante señalar que el remanente de adultos que emerge del suelo puede también estar contaminado con nemátodos, dado que estos insectos mudan a adultos en el suelo, todavía hay una última oportunidad para que sean pa-rasitados por los NEP y terminen muriendo en la parte aérea. Los adultos que escapen al control pueden volver a repoblar el huerto, sin embargo los NEP que logran matar insectos en el suelo se reproducen y pueden repetir el ciclo de parasitismo y patogénesis.
Foto 36. Larva de Otiorhynchus sulcatus parasitada por NEP, las diferencias de tamaño que se observa en los nemátodos corresponde a los adultos y juveniles.
A. F
ranc
e
43
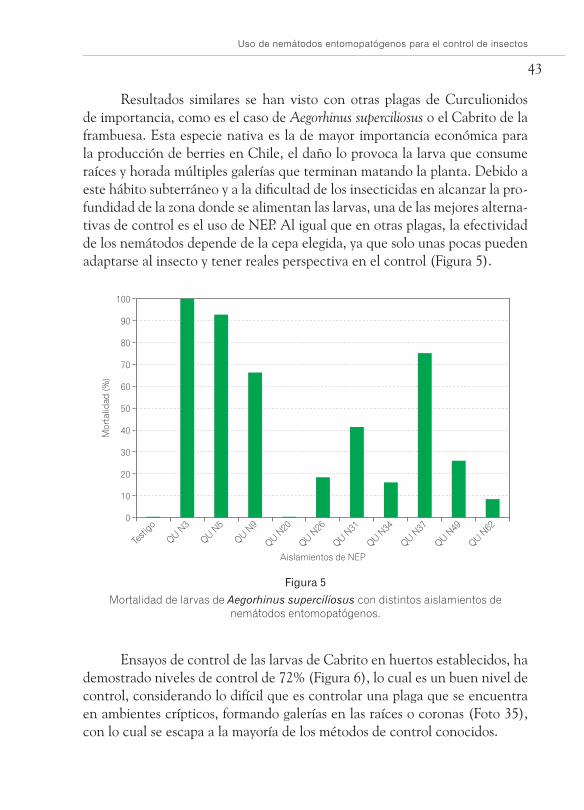
Resultados similares se han visto con otras plagas de Curculionidos de importancia, como es el caso de Aegorhinus superciliosus o el Cabrito de la frambuesa. Esta especie nativa es la de mayor importancia económica para la producción de berries en Chile, el daño lo provoca la larva que consume raíces y horada múltiples galerías que terminan matando la planta. Debido a este hábito subterráneo y a la dificultad de los insecticidas en alcanzar la pro-fundidad de la zona donde se alimentan las larvas, una de las mejores alterna-tivas de control es el uso de NEP. Al igual que en otras plagas, la efectividad de los nemátodos depende de la cepa elegida, ya que solo unas pocas pueden adaptarse al insecto y tener reales perspectiva en el control (Figura 5).
Figura 5
Mortalidad de larvas de Aegorhinus superciliosus con distintos aislamientos de nemátodos entomopatógenos.
Ensayos de control de las larvas de Cabrito en huertos establecidos, ha demostrado niveles de control de 72% (Figura 6), lo cual es un buen nivel de control, considerando lo difícil que es controlar una plaga que se encuentra en ambientes crípticos, formando galerías en las raíces o coronas (Foto 35), con lo cual se escapa a la mayoría de los métodos de control conocidos.
uso de nemátodos entomopatógenos para el control de insectos
Testi
go
QU N3
QU N5
QU N9
QU N20
QU N26
QU N31
QU N34
QU N37
QU N49
QU N62
Aislamientos de NEP
0
10
20
30
40
50
60
70
80
90
100
Mor
talid
ad (%
)

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
44
Figura 6
Efecto de las aplicaciones al suelo de nemátodos entomopatógenos (NEP) en la emergencia de adultos de Aegorhinus superciliosus Huerto de frambuesas
ubicado en La Unión.
Por último, se puede señalar como ejemplo de control de Cur-culionidos las evaluaciones realizadas con el Capachito de los frutales (Naupactus=Asynonychus cervinus), insecto que también solo presenta hem-bras, por lo cual es suficiente que ingrese un solo individuo a un huerto, para que se produzca una población cada vez mayor hasta convertirse en plaga. Las larvas de esta especie viven en el suelo y se alimenta de las raicillas de árboles o arbustos, por lo cual es un buen candidato para ser controlados por NEP. Ensayos realizados en arándanos afectados por esta plaga han demostra-do que los NEP reducen en 68% la emergencia de adultos (Figura 7).
0 50 100 150
Días
0
10
20
30
40
50
60
70
80
90
N° a
dulto
s/m
2
Con NEP Testigo

45
Figura 7
Efecto de las aplicaciones al suelo de nemátodos entomopatógenos (NEP) en la emergencia de adultos de Naupactus cervinus Huerto de arándano ubicado en Freire.
Es importante señalar que los NEP tienen especificidad por especies de insectos, y aunque se ve parasitismo y mortalidad con diferentes especies em-parentadas, siempre hay unas pocas cepas que pueden ejercer una alta mor-talidad (Figura 8). Esto es importante para evitar el uso de una sola especie o cepa para múltiples plagas. Lo anterior puede considerarse una desventaja, al requerir más de una cepa para el control de plagas de un cultivo, pero la especificidad permite que no se afecte otros insectos benéficos y que puedan estar ayudando al control de la plaga.
El principal uso que se le dan a los NEP es en el control de plagas que viven o pasan la mayor de su ciclo en el suelo, debido a que una de las limitantes a su uso es la susceptibilidad a la radiación solar o deshidratación por viento. Sin embargo, también es factible su uso en plagas aéreas bajo situaciones especiales, como son aplicaciones en épocas del año en que el clima no daño a los nemátodos. Por ejemplo, las aplicaciones en meses de invierno es una ventana que se puede utilizar en el caso de plagas que pupan o invernan en grietas de cortezas o galerías, donde los nemátodos
uso de nemátodos entomopatógenos para el control de insectos
0 20 40 60 80 100 120
Días
0
1
2
3
4
5
6
7
8
9
10N
° adu
ltos/
m2
Con NEP Testigo

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
46
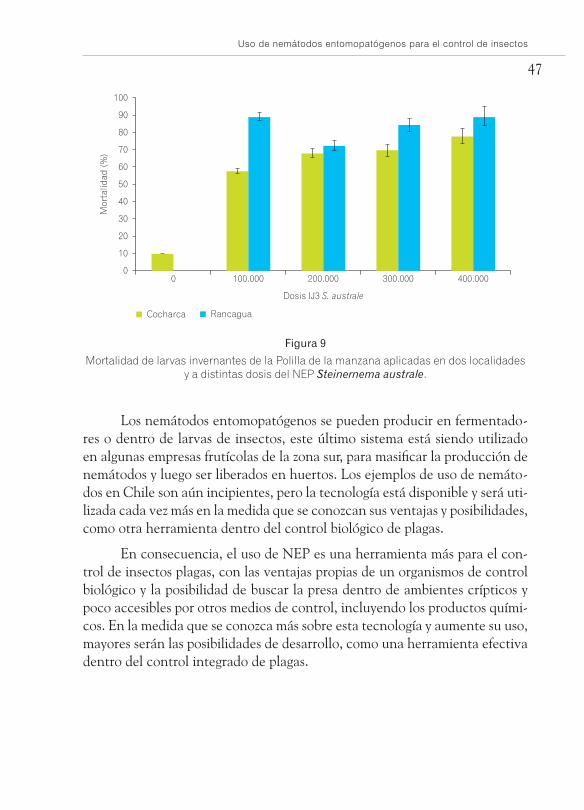
pueden acceder gracias a que se mueven por sí mismo en busca de su presa. Un buen ejemplo es el control de las larvas invernantes de la Polilla de la manzana (Foto 37), la cual pasa el invierno en las grietas de la corteza del manzano, con lo cual se pueden hacer tratamientos dirigidos al tronco. Tra-tamientos realizados en huertos de este frutal han mostrado que se puede lograr controles de hasta 90% de las larvas presentes en los troncos (Figura 9), lo cual es un medio efectivo para disminuir la presión de la plaga a la temporada siguiente.
Figura 8
Selección de cepas de NEP contra larvas de la Polilla de la manzan. Nótese la variabilidad de respuesta con respecto a la mortalidad que se puede alcanzar.
Foto 37. Adulto, larva y daño de la Polilla de la manzana.
QU
N22
a
QU
N3
QU
N4
QU
N50
QU
N96
QU
N10
QU
N16
QU
N59
QU
N65
QU
N34
QU
N76
QU
N19
QU
N11
QU
N61
QU
N31
QU
N30
QU
N25
QU
N66
QU
N47
QU
N51
QU
N36
QU
N27
QU
N8
QU
N55
QU
N53
QU
N23
QU
N29
QU
N48
QU
N17
QU
N14
QU
N46
QU
N20
QU
N60
QU
N9
QU
N28
QU
N6
QU
N33
QU
N39
QU
1Q
U N
24Q
U N
21Q
U N
63Q
U N
89Q
U N
32Q
U N
62TE
STI
GO
QU
N5
QU
N18
QU
N41
QU
N56
QU
N58
QU
N94
QU
N95
0
20
40
60
80
100
Mor
talid
ad (%
)
Aislamientos
a
ab ab ab abab ab
ab ab abb b b b b b b b
bc bc bc bc bc bc bc bc bc bc bc bc bcbc bc bc bc
c ccd cd cd cd cd
cd cd cd
d d d d d d d
A. F
ranc
e
47
Figura 9
Mortalidad de larvas invernantes de la Polilla de la manzana aplicadas en dos localidades y a distintas dosis del NEP Steinernema australe.
Los nemátodos entomopatógenos se pueden producir en fermentado-res o dentro de larvas de insectos, este último sistema está siendo utilizado en algunas empresas frutícolas de la zona sur, para masificar la producción de nemátodos y luego ser liberados en huertos. Los ejemplos de uso de nemáto-dos en Chile son aún incipientes, pero la tecnología está disponible y será uti-lizada cada vez más en la medida que se conozcan sus ventajas y posibilidades, como otra herramienta dentro del control biológico de plagas.
En consecuencia, el uso de NEP es una herramienta más para el con-trol de insectos plagas, con las ventajas propias de un organismos de control biológico y la posibilidad de buscar la presa dentro de ambientes crípticos y poco accesibles por otros medios de control, incluyendo los productos quími-cos. En la medida que se conozca más sobre esta tecnología y aumente su uso, mayores serán las posibilidades de desarrollo, como una herramienta efectiva dentro del control integrado de plagas.
uso de nemátodos entomopatógenos para el control de insectos
Mor
talid
ad (%
)
00
10
20
30
40
50
60
70
80
90
100
100.000 200.000
Dosis IJ3 S. australe
300.000 400.000
Cocharca Rancagua

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
48

49
Control de larvas de N. xanthographus con NEP, evaluación de campo
El control químico de N. xanthographus es difícil debido al extenso pe-ríodo de emergencia del adulto y los hábitos subterráneos de la larva la que queda fuera del alcance de los insecticidas convencionales, ya que éstos no pueden ser incorporados a la profundidad necesaria. Siendo también su uso restrictivo para la producción orgánica. Por lo anterior, el uso de nemátodos entomopatógenos (NEP) nativos surge como una alternativa viable para el control de estas plagas.
En Chile el uso de los nemátodos entomopatógenos (NEP), como es-trategia de control de larvas de insectos es poco conocida. A pesar de ello, nuestro país posee un alto potencial como fuente de NEP, debido a la am-plia variabilidad de condiciones edafoclimáticas, donde existen sectores con condiciones extremas; esta diversidad natural tiene como resultado un alto endemismo y microorganismos entomopatógenos adaptados a una amplia variedad de condiciones como, altas y bajas temperaturas, salinidad, alca-linidad, humedad, altitud etc., (Figura 3, Capítulo 3), lo que les otorga un interesante potencial como agentes biocontroladores. INIA ha colectado, de norte a sur, aislaciones de NEP, los cuales pueden ser masificados y aplicados en los cultivos para el control de plagas.
Los nemátodos, constituyen uno de los filum de invertebrados más importante, por su potencial como entomopatógenos, ya que son parásitos obligados de insectos, de allí su importancia como agentes de de control de plagas agrícolas Además son capaces de atacar a un amplio rango de insectos plaga, que habitan en el suelo en cultivos de importancia agrícola.
Nemátodos entomopatógenos para el control de
curculiónidos en vides
Paola luppichini B.*, andrés France i.** e irina urtubia H.**
INIA La Cruz *INIA Quilamapu**
4.

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
50
La efectividad del control con NEP ha sido comprobada ampliamente. La manera en que estos nemátodos ejercen el control es más parecida a la acción de un insecticida químico que al de uno biológico. Sin embargo, un insecto muerto por NEP es a su vez una nueva fuente de inóculo ampliando así su efectividad total en el tiempo. Otra de sus cualidades es que pueden ser multiplicados artificialmente a gran escala en medio líquido o sólido, pueden ser almacenados por largos períodos y ser aplicados por métodos convencio-nales. Poseen habilidad para buscar su hospedero y tienen compatibilidad con plaguicidas para persistir en el ambiente natural, y finalmente son inocuos para plantas y mamíferos.
En este contexto fueron realizados ensayos de laboratorio y campo de control de larvas de N. xanthographus, utilizando los NEP nativos de la colec-ción del INIA. Los principales resultados se muestran a continuación.
Selección de aislamientos de NEP sobre larvas de N. xanthographus
Para las pruebas de patogenicidad de los aislamientos de NEP fueron utilizadas larvas de N. xanthographus, obtenidas de las crianzas de laboratorio (22 ± 2 ºC y ±50% HR) (Foto 38). De los aislamientos de NEP, provenientes del Banco de Recursos Microbiológicos del INIA, fueron evaluados 72 con el fin de determinar la efectividad de control sobre larvas de esta especie de curculiónido. Se evaluó la mortalidad de las larvas a las 24 horas post inocu-lación y se extendió durante 10 días. Se consideraron larvas muertas, aquellas que no tenían respuesta (movimiento) frente al estímulo físico con un pincel (Foto 39). Las larvas muertas fueron llevadas a cámara húmeda para observar la aparición de nemátodos.
De los aislamientos de NEP evaluados, 63 mostraron tener algún grado de patogenicidad sobre las larvas de N. xanthographus, variando desde cinco a 90% de mortalidad (Figura 3). En nueve de los aislamientos no se observó patogenicidad (0%). Del total de aislamientos evaluados con cinco se obtuvo mortalidad del 60%, uno del 70% (QU N 21) y uno del 90% (QU N85) como se observa en la figura 10. Estas diferencias en cuanto a mortalidad podrían deberse a la respuesta de inmunidad que presentan los insectos en contra de la infección del nemátodo (Li et al., 2007), una vez que éstos han llegado al hemocele del huésped. Para contrarrestar las células bacterianas, el insecto

51
puede usar proteínas antibacteriales y/o fagocitosis, seguido de la formación de nódulos. Para inactivar a los nemátodos, los hemocitos de los insectos pueden encapsularlos y luego melanizarlos (Hazir et al. 2003; Kaya y Koppen-höffer 2004). Otra teoría para explicar la diferencia en la mortalidad de larvas es la hipótesis de la fase infectiva (Hominick y Reid 1990) que postula que una efectiva estrategia de sobrevivencia podría ser que la infectividad pueda ser sincronizada en el tiempo. De esta forma durante la emergencia desde un hospedero (Foto 40), algunos individuos (NEP) pueden ser inmediatamente infecciosos, mientras otros no lo son y estos últimos pueden ser no-infeccio-sos permanentemente o sólo por un tiempo.
En el estudio realizado con los aislamientos de la colección INIA, se logró seleccionar dos aislamientos que resultaron ser los más patogénicos para larvas de N. xanthographus, estos corresponden a QU N85 y QU N21 de las especie Steinernema unicornium (90%) especie nativa determinada por Edgington et. al. (2009) y S. feltiae (70%), respectivamente. Estos dos aisla-mientos QU N85 y QU N21 fueron los que mostraron el mejor potencial de control sobre las larvas de N. xanthographus, en todas las pruebas realizadas en laboratorio.
Foto 38. Larvas de N. xanthographus de crianza de laboratorio.
Foto 39. Larvas de N. xanthographus, sana (arriba) afectada por NEP (abajo).
nemátodos entomopatógenos para el control de curculiónidos en vides
P. L
uppi
chin
i
P. L
uppi
chin
i

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
52
Figura 10
Mortalidad de larvas de N. xanthographus inoculadas con juveniles infestantes de aislamientos de nemátodos entomopatógenos de la Colección INIA,
en condiciones de laboratorio.
Foto 40. Larva de N. xanthographus con NEP emergen desde cuerpo.
(%) M
orta
lidad
de
larv
as d
e N
. xan
thog
raph
us
Aislamientos de NEPs (Colección INIA)TE
STIG
O
0
10
20
30
40
50
60
70
80
90
100
N2
N4
N5
N8
N9
N10
N11
N13
N14
N15
N16
N17
N18
N20
N21
N22
N23
N24
N26
N27
N28
N29
N30
N31
N32
N35
N34
N37
N38
N39
N40
N41
N42
N44
N45
N47
N49
N50
N52
N55
N58
N59
N60
N62
N63
N64
N65
N68
N69
N71
N72
N73
N75
N76
N77
N78
N79
N84
N85
N86
N89
N90
N92
N95
N96
N97
N98
N99
N100
N101
N102
P. L
uppi
chin
i

53
Dosis y tiempo letalComo parámetro de comparación de patogenicidad de los aislamientos
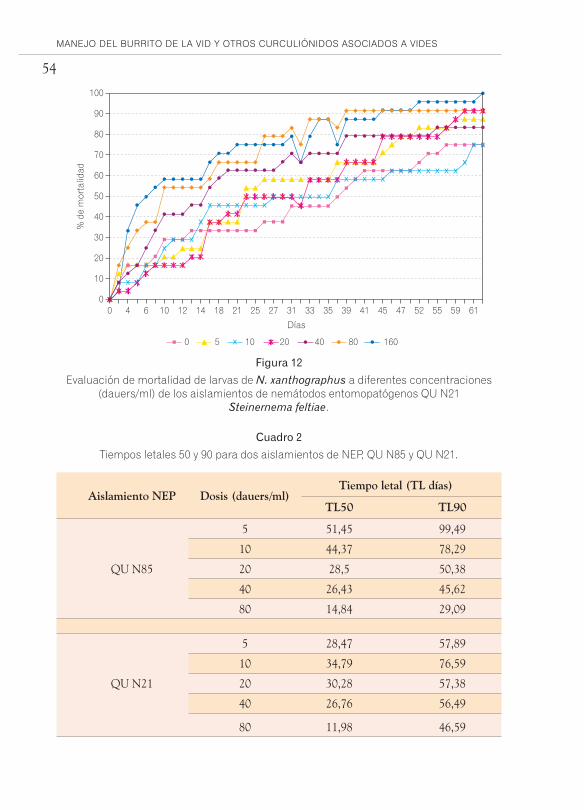
que alcanzaron el mayor porcentaje de mortalidad, se determinó Dosis Letal 50 y 90 (DL 50 y DL90) y Tiempo Letal 50 y 90 (TL50 y TL90), utilizando diferentes concentraciones de IJ (Infectivos juveniles o dauers).
Larvas de N. xanthographus fueron inoculadas con NEP en concentra-ciones de 0, 5, 10 20, 40, 80 y 160 IJ/larva, incubados en oscuridad a 15±2 ºC, evaluando diariamente la mortalidad hasta alcanzar el 100%.
Con los datos obtenidos, que se muestran en las figuras 11 y 12 y mediante análisis Probit se determinaron las dosis letales, para QU N85: LD 50 es de 12,8 dauers/ml, la LD 90 es de 152,5 dauers/ml. Para el aisla-miento QU N21 las dosis LD 50 y LD 90 fueron 3,17 y 106,8 dauers/ml, respectivamente.
Figura 11
Evaluación de mortalidad de larvas de N. xanthographus a diferentes concentraciones (dauers/ml) de los aislamientos de nemátodos entomopatógenos QU N85
Steinernema unicornium.
0
10
0 4 5 6 7 8 11 12 13 15 18 19
Días
0 5 10 20 40 80 160
20 22 25 26 27 28 29 33 34 35 36
20
30
40
% d
e m
orta
lidad
50
60
70
80
90
100
nemátodos entomopatógenos para el control de curculiónidos en vides
Manejo del Burrito de la vid y otros curculiónidos asociados a vides
54
0
10
0 4 6 10 12 14 18 21 25 27 31 33 35 39 41 45 47 52 55 59 61
Días
0 5 10 20 40 80 160
20
30
40
% d
e m
orta
lidad
50
60
70
80
90
100
Figura 12
Evaluación de mortalidad de larvas de N. xanthographus a diferentes concentraciones (dauers/ml) de los aislamientos de nemátodos entomopatógenos QU N21
Steinernema feltiae.
Cuadro 2
Tiempos letales 50 y 90 para dos aislamientos de NEP, QU N85 y QU N21.
Aislamiento NEP Dosis (dauers/ml) Tiempo letal (TL días)
TL50 TL90
5 51,45 99,49
10 44,37 78,29
QU N85 20 28,5 50,38
40 26,43 45,62
80 14,84 29,09
5 28,47 57,89
10 34,79 76,59
QU N21 20 30,28 57,38
40 26,76 56,49
80 11,98 46,59

55
A partir del análisis de regresión se determinaron los tiempos letales, los cuales se resumen en el cuadro 2. Cabe destacar que en el caso del aisla-miento QU N85 las dosis letales son mayores que para QU N21 y los tiempos letales son menores para las concentraciones más efectivas.
Resultados de campoA partir de los resultados obtenidos en laboratorio se continuó reali-
zando pruebas de efectividad de los dos aislamientos de NEP, en condiciones de campo en un huerto de arándano (Foto 41), un viñedo y en macetas.
Ensayo en ArándanosFueron evaluaron cuatro de los aislamientos de NEP, considerados en
la primera etapa de selección como los más promisorios para el control de N. xanthographus: QU N85, QU N21, QU N23 y QU N37 todos del género Steinernema.
Foto 41. Vista general huerto arándanos.
En la figura 13 se muestra la mortalidad de las larvas de N. xanthographus recuperadas del campo, 15 días post inoculación. De los aislamientos aplicados, con QU N85 (S. unicornium) se obtuvo un 61,9% de mortalidad y un 52,9% con QU N21 (S. feltiae). Los otros dos aislamientos probados, QU N23 y QU N37, no superaron el 40 % de mortalidad de larvas. Los resultados de campo son consistentes con los obtenidos en laboratorio, donde se observó que QU N85 tiene mayor potencial de control sobre las larvas de N. xanthographus (90 % mortalidad) que QU N21, QU N23 y QU N37 (Figura 10).
nemátodos entomopatógenos para el control de curculiónidos en vides

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
56
Figura 13
Mortalidad de larvas de N. xanthographus con inoculaciones de diferentes aislamientos de NEP, en aplicaciones de campo. Los Angeles 2011.
Ensayo en ParronalFueron evaluados 2 aislamientos de NEP QU N85 y QU N21 de las
especies Steinernema unicornium y S. feltiae, respectivamente. La aplicación se realizó a través de larvas de Galleria melonella inoculadas con los NEP, las cuales fueron ubicadas en cada planta en dos puntos únicos de inoculación (Fotos 42 y 43). Se utilizó larvas de N. xanthographus como centinelas (2 larvas/planta a 20 cm de la base de la planta y a 20 cm de profundidad) para evaluar la efectividad de los NEP. Estas larvas fueron colocadas en pequeñas bolsas de malla y sujetas con un alambre galvanizado (Foto 44).
En cuanto a la mortalidad de larvas de N. xanthographus utilizadas como centinelas evaluadas para diferentes tiempos post inoculación (1, 2, 3 y 4 semanas), se observó un 55% de mortalidad de larvas con el aislamiento QU N21 (S. feltiae) durante las semanas 1 y 2. Para las condiciones del en-sayo con el aislamiento QU N85 de la especie S. unicornium, se observó mor-talidad superior al 80% en las semanas 3 y 4 posteriores a la inoculación con NEP. En estas evaluaciones también se observó larvas parasitadas por hongos asociados al suelo (Figura 14).
0
20
Testigo
40
60
80
100
% m
orta
lidad
de
larv
as d
e N
. xan
thog
raph
us
QU N85 QU N21
Tratamientos NEP
QU N23 QU N37

57
En el recuento de larvas en terreno previo al ensayo fueron colectadas en promedio 2 larvas de N. xanthographus por calicata (Foto 45), que corres-ponde a un nivel ligero de infestación (Ripa y Luppichini 2010). Durante las evaluaciones semanales se realizaron 3 calicatas por tratamiento, obteniendo un promedio de 6,8 larvas/calicata.
Al evaluar la mortalidad por NEP de las larvas de N. xanthographus presentes naturalmente en el suelo (Figura 15), se observó superior al 50% de mortalidad por NEP de las larvas recuperadas desde el tratamiento con el aislamiento QU N85 (Foto 46) hasta la semana 3 post inoculación. En el
Foto 42. Puntos únicos de inoculación en cada planta.
Foto 43. Aplicación de larvas de G. melonella inoculadas con los NEP,
Foto 44. Bolsa de malla con un alambre galvanizado con larva centinela.
P. L
uppi
chin
i
P. L
uppi
chin
i
N. O
livar
es
nemátodos entomopatógenos para el control de curculiónidos en vides

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
58
tratamiento con QU N21 se observó efecto sobre larvas (Foto 47) durante las dos primeras semanas post aplicación, con un promedio de 58% de mor-talidad asociada a los NEP. Al igual que en las otras evaluaciones se observó mortalidad de larvas por hongos asociados al suelo.
Figura 14
Mortalidad de larvas centinelas de N. xanthographus, con dos aislamientos de NEP. San Felipe, 2011.
Figura 15
Mortalidad de larvas de N. xanthographus, provenientes de la infestación natural, con dos aislamientos de NEP. San Felipe, 2011.
Semana 1
% m
orta
lidad
de
larv
as d
e N
. xan
thog
raph
us
Semanas post inoculación con NEP
Semana 2 Semana 3
QU N21 QU N85
0
20
40
60
80
100
0
20
40
60
80
100
Semana 1
% m
orta
lidad
de
larv
as d
e ce
ntin
elas
N. x
anth
ogra
phus
Semanas post inoculación
Semana 2
Testigo
Semana 3 Semana 4
QU N21 QU N85

59
Fotos 45. Realización de calicata y extracción de larvas de N. xanthographus de la infestación natural.
Fotos 46. Larva de N. xanthographus muerta por NEP aislamiento QU N85.
Foto 47. Larva de N. xanthographus muerta por aislamiento NEP QU N21.
P. L
uppi
chin
iA
. Fra
nce
A. F
ranc
e
P. L
uppi
chin
i
nemátodos entomopatógenos para el control de curculiónidos en vides

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
60
Ensayo en ViñedoEn este ensayo se utilizó la misma metodología del ensayo descrito en
los párrafos anteriores, donde La aplicación de los NEP se realizó a través de larvas de Galleria melonella inoculadas, las cuales fueron ubicadas en dos puntos únicos.
En la figura 16, se muestran los resultados de la evaluación de la mor-talidad de las larvas centinelas, se observó un promedio cercano al 60% en promedio para los dos aislamientos evaluados, en el viñedo ubicado en Casablanca.
Figura 16
Mortalidad de larvas centinelas de N. xanthographus,con dos aislamientos de NEP. Casablanca, 2012.
Fueron realizadas siete evaluaciones post inoculación con los NEP, me-diante 5 calicatas por tratamiento. Al evaluar la mortalidad por NEP de estas larvas de la infestación natural del viñedo, el aislamiento QU N85 mostró una mortalidad que varió entre el 50 y 66% de las larvas hasta los 33 días post inoculación para las condiciones del ensayo. Desde el día 43 post inoculación la mortalidad fue disminuyendo desde el 28% hasta un 12,5% a los 71 DDI.
0
20
12
40
60
80
100
% m
orta
lidad
de
larv
as c
entin
elas
de
N. x
anth
ogra
phus
22 33 43
Días después de la inoculación (DDI) con NEP
Testigo
50 61 71
QU N21 QU N85
61
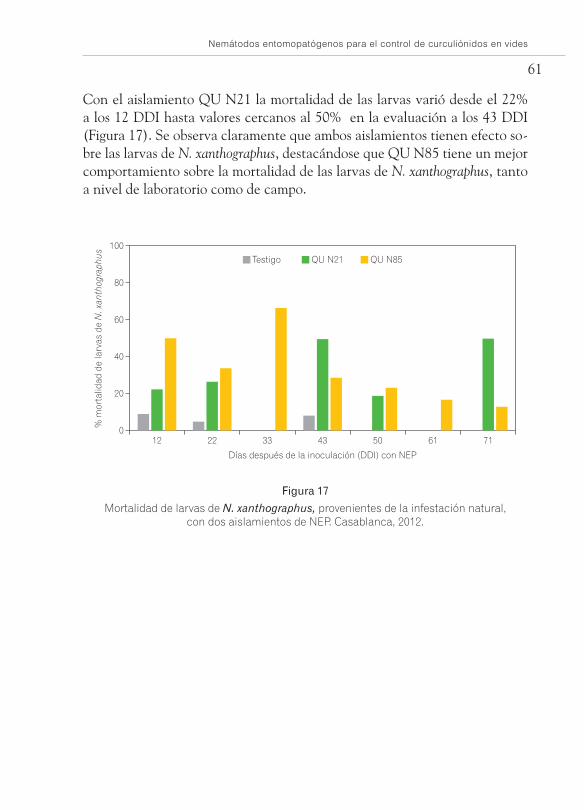
Con el aislamiento QU N21 la mortalidad de las larvas varió desde el 22% a los 12 DDI hasta valores cercanos al 50% en la evaluación a los 43 DDI (Figura 17). Se observa claramente que ambos aislamientos tienen efecto so-bre las larvas de N. xanthographus, destacándose que QU N85 tiene un mejor comportamiento sobre la mortalidad de las larvas de N. xanthographus, tanto a nivel de laboratorio como de campo.
Figura 17
Mortalidad de larvas de N. xanthographus, provenientes de la infestación natural, con dos aislamientos de NEP. Casablanca, 2012.
0
20
40
60
80
100
12
% m
orta
lidad
de
larv
as d
e N
. xan
thog
raph
us
22 33 43
Días después de la inoculación (DDI) con NEP
Testigo
50 61 71
QU N21 QU N85
nemátodos entomopatógenos para el control de curculiónidos en vides

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
62

63
Los nemátodos entomopatógenos (Rhabditida: Steinernematidae y Heterorha-bditidae) son una excelente alternativa para el control de plagas agrícolas. El año 1923 Gotthold Steiner descubrió la primera especie de Steinernema (S. kraussei), iniciándose la investigación respecto a los nemátodos entomopató-genos y sus aplicaciones, especialmente en el área agrícola. Sin embargo, el desarrollo de productos comerciales de nemátodos formulados ha sido reali-zado sólo en las últimas décadas.
Para la comercialización de los nemátodos es necesario producir a gran escala, éstos pueden ser reproducidos masivamente a través de métodos in vivo e in vitro (Bedding 1981).
En los procesos in vivo, los nemátodos son criados en hospederos vivos, los cuales funcionan como un biorreactor. Generalmente se utilizan larvas de la pollilla de la cera Galleria mellonella. Los nemátodos son fácilmente co-sechados al permitirles nadar fuera del cadáver del hospedero dentro de un sistema de recolección conocido como trampa White modificada. Los rendi-mientos que se obtienen oscilan alrededor de 1*10 4 IJ/larva, (IJ= infectivo juvenil) pero estos pueden variar dependiendo la especie de nemátodo.
El cultivo de nemátodos in vitro fue iniciado por Glaser en 1940. Estos métodos se basan en la utilización de medios de cultivo inertes y sintéticos como fuente de Nitrógeno y Carbono.
Formulación de Nemátodos Entomopatógenos
irina urtubia H.
INIA Quilamapu
5.

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
64
Uno de los principales pasos hacia una producción comercial fue la incorporación de la bacteria simbionte del nemátodo (Photorhabdus spp y Xe-norhabdus spp) a los medios artificiales. Inicialmente se desarrollaron cultivos en medios sólidos, utilizando trozos de esponja, que brindan una gran área superficial. Los rendimientos obtenidos fueron 6*10 5-10*10 5 IJ/g de medio aproximadamente.
Los retos hacia una masificación más eficiente de nemátodos se pue-den resumir en:
1. identificar nutrientes de bajo costo 2. determinar parámetros fisicoquímicos de cultivo que permitan aumen-
tar los rendimientos 3. utilizar medios de cultivo líquidos en lugar de sólidos.
En la actualidad la utilización de fermentaciones líquidas han permiti-do disminuir los costos de producción, facilitando la cosecha de nemátodos y aumentando los rendimientos, hasta alrededor de 1*10 5 IJ/cm 3 en el caso de S. carpocapsae y S. feltiae en fermentadores de 15.000-80.000 L. Cabe men-cionar que en el caso de Heterorhabditis los resultados en medios líquidos no han sido tan promisorios como con Steinernema, por lo que generalmente se siguen produciendo en matrices sólidas. El siguiente paso tras la masifica-ción es la formulación de los nemátodos, de manera de obtener un producto comercial que garantice la sobrevivencia y que incremente la estabilidad de ellos durante un mayor período de tiempo de almacenamiento, además de facilitar su aplicación y por ende mejorar su eficacia en el campo.
Las formulaciones de nemátodos se basan fundamentalmente en la restricción de movilidad (inmovilidad física), permitiendo de esta manera prolongar sus reservas energéticas (lípidos) o en la reducción del consumo de oxígeno, induciéndoles un estado parcial de anhidrobiosis (inmovilización fisiológica) (Grewal y Georgis 1999).
Para seleccionar el tipo de formulación más adecuada es fundamental que se realice un estudio de preformulación para la cepa que se desee utilizar, definiendo parámetros fisicoquímicos óptimos como temperatura, humedad, requerimiento de oxígeno, compatibilidad con excipientes, entre otros. Se debe tener en cuenta que entre las especies Steinernema y Heterorhabditis exis-ten diferencias fisiológicas, que deben tomarse en cuenta para seleccionar la

65
formulación más adecuada (ingredientes, dimensiones, condiciones de alma-cenamiento, forma de aplicación (Georgis, 1992).
Las investigaciones de los últimos años se han enfocado en la búsque-da de sustancias con propiedades que faciliten la manipulación, aplicación y sobrevivencia de los nemátodos durante el tiempo de almacenamiento. Estas sustancias inertes deben inmovilizar o deshidratar parcialmente los nemátodos para reducir su metabolismo, asegurando de esta forma una mayor estabilidad en condiciones específicas de almacenamiento. Se ha determinado que en esta-do anhidrobiótico se logra incrementar la resistencia a temperaturas por sobre el óptimo, al vacío, radiación y a ciertas sustancias tóxicas (Grewal 1998). Es así como preparaciones deshidratadas podrían formar parte de formulaciones más ligeras, estables, de menor volumen, fáciles de manipular y aplicar.
También los nemátodos pueden inmovilizarse o reducir su movilidad llegando a un estado metabólico reducido, para preservar sus energías lipídi-cas. En este caso se pueden utilizar matrices de materiales arcillosos (Bedding, 1984), fáciles de mezclar, disolverse y aplicar, pero su estabilidad no sobrepasa los 3 meses, especialmente en el caso de Heterorhabditis (Connick et. al. 1993 a; Connick et. al. 1993 b). Otras matrices basadas en el mismo principio son aquellas en forma de gel o encapsulados (Soler et. al. 2003), aunque también se han formulado nemátodos en otro tipo de soportes inertes, como esponja y vermiculita (1999), pero su limitante ha sido la falta de estabilidad a tempe-ratura ambiente (Grewal 1998).
Formulaciones de NEP desarrolladas por INIAActualmente en INIA se están desarrollando formulaciones de nemá-
todos entomopatógenos nativos, de dos cepas de Steinernema para el control del Burrito de la vid (Naupactus xanthographus) pertenecientes a la colección del Banco de Recursos Microbiológicos de INIA, Steinernema unicornium (QU N85) y S. feltiae (QU N21).
La finalidad de estas formulaciones es permitir mayor sobrevivencia de los nemátodos, de manera de asegurar un mayor porcentaje de viabilidad con respecto al tiempo, pudiendo ser almacenados por un mayor número de meses asegurando la calidad y capacidad de parasitismo de los NEP. Por otra parte, otro objetivo es facilitar la aplicación en terreno, ya sea directamente al suelo o vía riego, disminuyendo las dificultades de manipulación del producto.
Formulación de nemátodos entomopatógenos

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
66
Una de las formulaciones desarrolladas es de tipo granular (Foto 48), para aplicación directa en el campo, ya sea al momento de preparar el suelo o en cultivos ya establecidos. En esta formulación los NEP estarían encap-sulados en una matriz polimérica, con un grado tal de humedad que asegura su viabilidad y con la porosidad adecuada para permitir su salida. Este último punto es de vital importancia ya que una vez aplicados al suelo, y con la hú-medad necesaria, en una primera etapa, los NEP deben pasar a través de los poros del gránulo. Luego en una segunda etapa, el gránulo es biodegradado, permitiendo de esta manera la liberación total de los NEP contenidos en él.
Foto 48: Formulado granular de NEP. A: Gránulos com 60% de humedad. B: Gránulos suspendidos en agua por unos minutos.
Esta formulación está compuesta de una matriz polimérica, estabiliza-dor de NEP, preservante y antifúngico, que aseguran la calidad de los NEP e impiden contaminación por agentes microbianos. La concentración de NEP es de 10 5 IJ ml -1, con una humedad del 60%.
Estos formulados han sido almacenados en sala de frío a 15-18 ºC du-rante 6 meses, período en el cual se ha determinado la viabilidad de los NEP, expresada como porcentaje de individuos infectivos viables. Como se observa en la figura 18, los NEP formulados presentan un mayor porcentaje de via-bilidad que aquellos no formulados y almacenados en suspensión acuosa en iguales condiciones de temperatura.
I. U
rtub
ia
I. U
rtub
ia

67
Figura 18
Viabilidad de NEP cepa QU N21 y QU N85 en formulados granulares.
Otro tipo de formulación desarrollada es un gel hidrosoluble y una mezcla arcillosa, productos que al disolverlos y/o resuspenderlos en agua, per-mitirían la liberación de los NEP, aplicándolos a través del riego.
El Gel hidrosoluble, que se aprecia en la foto 49, está compuesto de un polímero absorbente inerte, además de agentes preservantes, estabilizan-tes de NEP y antifúngicos. La concentración de NEP es del orden de 10 5 IJ ml -1 y el almacenamiento se realiza en las mismas condiciones del formulado anterior.
Foto 49: Formulación tipo Gel hidrosoluble.
Formulación de nemátodos entomopatógenos
01
Tiempo de almacenamiento (meses)
Gránulos NQ21
10
20
30
40
50
60
70
80
90
100Po
rcen
taje
de
viab
ilida
d
2 3 4 5 6
Suspensión NQ21
Gránulos NQ85
Suspensión NQ21
I. U
rtub
ia
I. U
rtub
ia
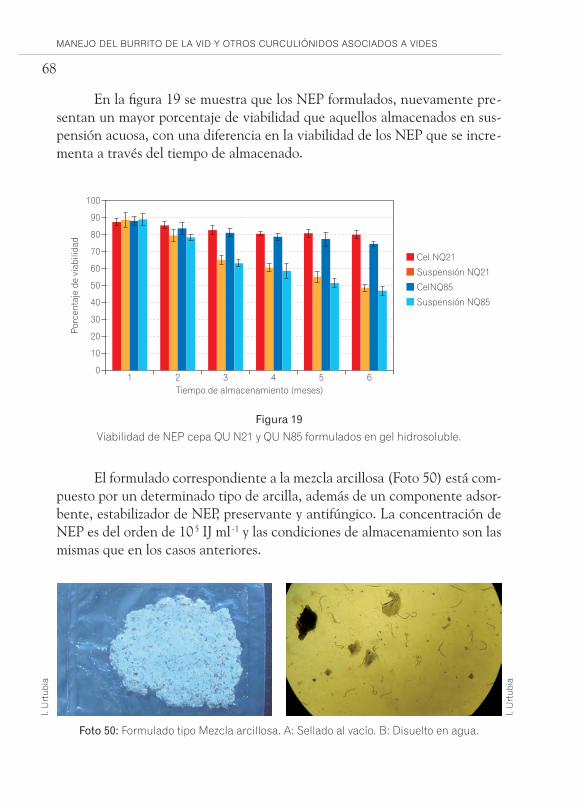
Manejo del Burrito de la vid y otros curculiónidos asociados a vides
68
En la figura 19 se muestra que los NEP formulados, nuevamente pre-sentan un mayor porcentaje de viabilidad que aquellos almacenados en sus-pensión acuosa, con una diferencia en la viabilidad de los NEP que se incre-menta a través del tiempo de almacenado.
Figura 19
Viabilidad de NEP cepa QU N21 y QU N85 formulados en gel hidrosoluble.
El formulado correspondiente a la mezcla arcillosa (Foto 50) está com-puesto por un determinado tipo de arcilla, además de un componente adsor-bente, estabilizador de NEP, preservante y antifúngico. La concentración de NEP es del orden de 10 5 IJ ml -1 y las condiciones de almacenamiento son las mismas que en los casos anteriores.
Foto 50: Formulado tipo Mezcla arcillosa. A: Sellado al vacío. B: Disuelto en agua.
01
Tiempo de almacenamiento (meses)
Cel NQ21
10
20
30
40
50
60
70
80
90
100
Porc
enta
je d
e vi
abili
dad
2 3 4 5 6
Suspensión NQ21
CelNQ85
Suspensión NQ85
I. U
rtub
ia
I. U
rtub
ia

69
En la figura 20 se muestra que la tendencia es similar a los casos anterio-res, lográndose un mayor porcentaje de viabilidad en los NEP formulados.
Figura 20
Viabilidad de NEP cepa QU N21 y QU N85 formulados en mezcla arcillosa.
El cuadro 3 muestra un resumen del comportamiento de ambos NEP formulados en cuanto a viabilidad después de 1 mes y 6 meses de almacena-miento. Se puede apreciar claramente la diferencia entre los distintos tipos de formulados con respecto a la suspensión acuosa, lográndose aumentar la sobrevivencia de éstos con respecto al tiempo.
Además de medir la viabilidad de los NEP en los distintos tipos de formulados, también se debe determinar su capacidad de parasitismo, para asegurar la calidad de ellos en el producto. Para este fin se prepararon tram-pas de suelo con larvas de Galleria mellonella, adicionando los formulados y manteniendo la concentración de NEP aplicados. La mortalidad se determi-nó a los 7 días de inoculación.
En la figura 21 se puede apreciar que con todos los formulados con un tiempo de almacenamiento de 5 meses se obtuvo un 100% de mortalidad de las larvas, no así en el caso de la suspensión acuosa.
Formulación de nemátodos entomopatógenos
01
Tiempo de almacenamiento (meses)
Mezcla arcillosa NQ21
10
20
30
40
50
60
70
80
90
100
Porc
enta
je d
e vi
abili
dad
2 3 4 5 6
Suspensión NQ21
Mezcla arcillosa NQ85
Suspensión NQ85
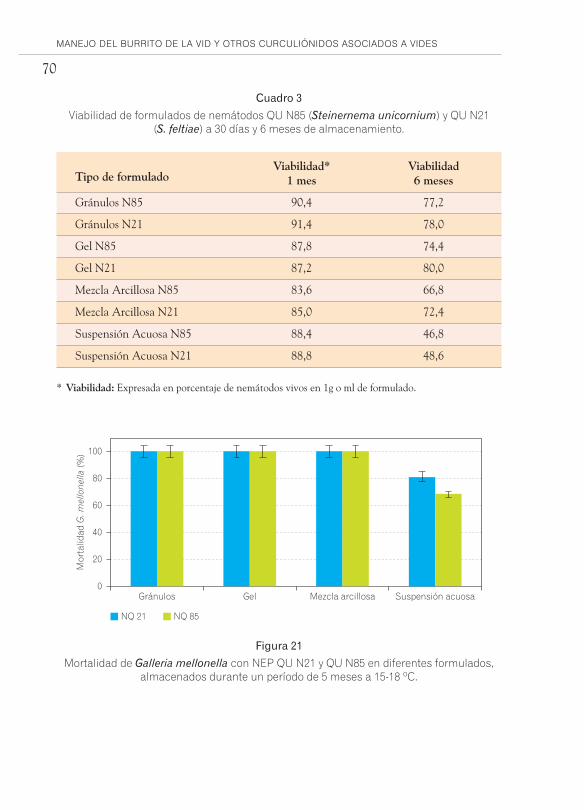
Manejo del Burrito de la vid y otros curculiónidos asociados a vides
70
Figura 21
Mortalidad de Galleria mellonella con NEP QU N21 y QU N85 en diferentes formulados, almacenados durante un período de 5 meses a 15-18 ºC.
0
20
40
60
80
100
Gránulos
Mor
talid
ad G
. mel
lone
lla (%
)
NQ 21 NQ 85
Gel Mezcla arcillosa Suspensión acuosa
Cuadro 3
Viabilidad de formulados de nemátodos QU N85 (Steinernema unicornium) y QU N21 (S. feltiae) a 30 días y 6 meses de almacenamiento.
Tipo de formulado Viabilidad* Viabilidad
1 mes 6 meses
Gránulos N85 90,4 77,2
Gránulos N21 91,4 78,0
Gel N85 87,8 74,4
Gel N21 87,2 80,0
Mezcla Arcillosa N85 83,6 66,8
Mezcla Arcillosa N21 85,0 72,4
Suspensión Acuosa N85 88,4 46,8
Suspensión Acuosa N21 88,8 48,6
* Viabilidad: Expresada en porcentaje de nemátodos vivos en 1g o ml de formulado.

71
Finalmente se realizó la evaluación de la efectividad de control de los NEP sobre las larvas de N. xanthographus. La mortalidad fue determinada hasta 1 mes de post aplicación de los NEP formulados almacenados durante 5 meses. El proceso de las larvas se muestra en la foto 51.
En la figura 22, se muestra la efectividad de los NEP con todos los formulados. Se aprecia un porcentaje de mortalidad superior al obtenido con la suspensión acuosa, lográndose diferencias de hasta un 20% de mortalidad aproximado, en el caso de gránulos y gel hidrosoluble.
Foto 51: Larvas de N. xanthographus. A: en trampa de suelo. B: parasitada por nemátodo. C. emergencia de nemátodos desde el interior de la larva.
Figura 22
Mortalidad de Naupactus xanthographus con NEP QU N21 y QU N85 en diferentes formulados, almacenados durante un período de 5 meses a 15-18 ºC.
Formulación de nemátodos entomopatógenosI.
Urt
ubia
0
10
20
30
40
50
60
Gránulos
NQ 21
Mor
talid
ad N
. xan
thog
raph
us (%
)
NQ 85
Gel Mezcla arcillosa Suspensión acuosa

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
72
Estos resultados, sumados a los porcentajes de viabilidad, ratifican la calidad de los NEP en el producto formulado y abren una muy buena posibilidad de aplicabilidad de un producto formulado para el control de esta y otras plagas de suelo.

73
Glosario de términos
Áptero: Sin alas.
Ciclo de vida 1. En la biología, se entiende por ciclo de vida a la alternancia de generaciones. 2. Conjunto de fenómenos transcurridos entre un momento dado de una generación de un organismo y el mismo de la generación siguiente.
Control biológico: la acción de depredadores, parasitoides y patógenos en la regulación de po-blaciones de otros organismos (enemigos naturales). El control biológico puede ocurrir en forma natural en el campo o como resultado de la manipulación o introducción por el hombre de agentes de control biológico.
Cuarentenario: cualquier organismo (ácaro, insecto, enfermedad, etc), que presente restric-ciones de ingreso en un país.
Dauer: tercer estado juvenil de los nemátodos.
Depredador: organismo que se alimenta de otros causándoles la muerte, normalmente consu-me varios individuos durante su desarrollo.
Élitros: Son las alas anteriores, modificadas por endurecimiento (esclerotización), de ciertos órdenes de insectos (Coleóptero y Hemíptero). Sirven como protección para las alas posteriores que están inmediatamente debajo y que sirven para ejecutar el vuelo.
Generación: período comprendido entre un estado determinado en el ciclo de vida al mismo estado en la descendencia. Ejemplo, de huevo a huevo.
Larva: estado inmaduro entre el huevo y pupa de los insectos que poseen metamorfosis com-pleta, donde los estados inmaduros difieren del adulto. Ej. orugas, gusanos.
Hemocele: cavidad que forma parte del aparato circulatorio de los artrópodos, por donde circula la hemolinfa.
Hemolinfa: líquido circulatorio de los artrópodos, moluscos, etc., análogo a la sangre de los vertebrados. Puede ser de diferentes colores o incluso incolora y contiene células san-guíneas de diferentes tipos y funciones.
Parasitoide: organismo que vive, parte de su ciclo de vida a expensas de otro causándole la muerte, siendo el adulto generalmente de vida libre. Los parasitoides se diferencian de los verdaderos parásitos en que matan a su huésped.
Partenogénesis: de reproducción asexuada se distinguen especies arrenotóquicas y telitóqui-cas. Las primeras aquellas en donde los machos son producto de huevos no fecundados y las hembras son resultantes de una reproducción sexuada corriente. Las especies telitóquicas son aquellas en donde los huevos no fecundados dan origen a hembras.
Simbiosis: vida en común, que puede ser permanente, de dos o más organismos distintos mediante una asociación estrecha, la cual es ventajosa tanto para el hospedador como para el huésped. A los organismos participantes se les llama simbiontes.
Vector: organismo capaz de transmitir un agente infeccioso a otros individuos que no lo portan.

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
74

75
Bibliografía
Álvarez, M., M. Gerding y A. France. 1998. Control de Caliroa cerasi (L.) con hongos entomopató-genos. Tesis de Ingeniero Agrónomo, Universidad Adventista de Chile, Chillán. 35 p.
Alekseev, E., I. Glazer and M. Samish. 2006. Effect of soil texture and moisture on the activity of entomopathogenic nematodes against female Boophilus annulatus ticks. BioControl. 51(4): 507-518.
Akhurst, R. J. and M. E. Boemare. 2005. Biology and taxonomy of Xenorhabdus. Pp. 75-90, in: Entomopathogenic nematodes in biological control. R. Gaugler and H. K. Kaya, eds. CRC Press, Florida, USA. 365 p.
Alves, S., J. Almeida, A. Moino y L. Alves. 1998. Técnicas de laboratorio. Pp. 637-712, in: S. Alves (Ed.). Controle microbiano de insetos (2° ed.). FEALQ, Piracicaba, Brasil. 1163 p.
Artigas, J. 1994. Entomología económica. Insectos de interés agrícola, forestal, médico y veterinario (nativos, introducidos, susceptibles de ser introducidos). Vol. 1. Ediciones Universidad de Concepción, Concepción. 1126 p.
Bai, Shapiro-Ilan, Gaugler & Hopper, 2005. Biological control whit entomopathogenic nematodes. Biol. Control. 32 (4): 121-130.
Barattini, P. 2008. Estudio de aspectos biológicos de capachito de los frutales Pantomorus cervinus (Boheman) (Coleoptera: Curculionidae) en realción a parámetros naturales. 25 p. Tesis Biólogo Universidad Católica de Valparaíso. Valparaíso- Chile.
Bedding, R.A. 1981: Low cost in vitro mass producction of Neoaplectana and Heterorhabditis spe-cies (nematoda) for field control of insect pests. Nematologica, 27:109-114.
Bedding, R.A. 1984. Storage of entomopathogenic nematodes. US. Patent. US 5042427 910827.
Bedding, R. A., R. J. Akhurst, and H. K. Kaya. 1993. Future prospects for entomogenous and ento-mopathogenic nematodes. Pp. 157-170, in: Nematodes and the biological control of insect pests. R. Bedding, R. Akhurst, and H. Kaya, eds. Csiro, Victoria, Australia. 178 p.
Bilgrami, Gaugler, Shapiro-Ilan, Adams, 2006 Entomopathogenic nematodes. Nematology 41 (2) 227-301.
Cambell J., Orza G., Poder F. y Gaugler R. 2003. Evolution of host search stages in entomopathogenic nemátodos (Nematoda Steinernematidae). Journal of Nematology 35, 142 – 145.
Casals, P. y J. Rodríguez. 2012. Control de adultos de curculiónidos en frutales y vid. Revista Re-dagrícola. Nº 42. p. 70 - 71.
Cisternas, E., A. France y M. Gerding. 1998. Control de Listronotus bonariensis con Beauveria bas-siana en praderas en el sur de Chile. Pp. 145, in: Anais do VI SICONBIOL. Río de Janeiro, Brasil. 226 p.
Cisternas, E. y A. France. 2009. Manual de campo. Plagas, enfermedades y desódenes fisiológicos del arándano en Chile. Boletín INIA Nº 189. 127 p.
Bibliografía

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
76
Connick, W.J., Jr.; Nickle, W.R.; Vinyard, B.J. 1993 (a). “Pesta”: New granular formulations for Steinernema carpocapsae. J. Nematol. 25:198-203.
Connick, W.J., Jr.; Nickle, W.R.; Williams, K.S., Vinyard, B.T. 1993 (b). Granular formulations for Steinernema carpocapsae (strain All) (Nematoda:Rhabditida) with improved shelf life. J. Ne-matol. 26:352-359.
Darthel S. Espinoza D y Morrison F. 2004 Entomopathogenic nematodes: Mass production in Inte-grated pest management: Potential, constraints and challenges. Koul.
Dutky, S.R. 1957. Report on white grub control Project in Chile. Agricultura Técnica (Chile) 17(2):92-105.
Edgington, S., A. G. Buddie, D. Moore, A. France, L. Merino, L. Tymo and D. J. Hunt. 2010. Di-versity and distribution of entomopathogenic nematodes in Chile. Nematology 12(6):915-928.
Edgington, S., A. Buddie, L. Tymo, D. Hunt, K. Nguyen, A. France, L. Merino and D. Moore. 2009. Steinernema unicornium n. sp. (Panagrolaimomorpha: Steinernematidae), a new entomo-pathogenic nematode from Tierra del fuego, Chile. J. Nematode Morphol. Syst., 12 (2): 113-131.
Elgueta, M. y A.E. Marvaldi. 2006. Lista sistemática de las especies de Curculionoidea (Insecta: Coleoptera) presentes en Chile, con su sinonímia. Boletín del Museo Nacional de Historia Natural, Chile, 55: 113-153
Elgueta, M. 1993. Las especies Curculionoidea (Insecta: Coleoptera) de interés agrícola en Chile. Publicación ocasional Nº 48. Museo Nacional de Historia Natural. 79 p.
Emelianoff, V., M. Sicard, N. Le Braun, C. Moulia and J-B. Ferdy. 2007. Effect of bacterial symbionts Xenorhabdus on mortality of infective juveniles of two Steinernema species. Parasitol. Res. 100(3): 657-659.
Escalante, L. S. 1982. Control de Naupactus xanthographus (Germar) mediante tratamiento al suelo. Tesis (Ing. Agr.), Universidad de Chile. Esc. de Agronomía, Santiago. 72 p.
Espinoza, S. y A. France. 1999. Uso de nemátodos entomopatógenos nativos en el control de Ae-gorhinus superciliosus Pp. 776, in: Resúmenes 50° Congreso Agronómico de la Sociedad Agronómica de Chile. Universidad de la Frontera, Pucón. 835 p.
France, A., S. Espinoza, M. Gerding y E. Cisternas. 1999. Determinación de nemátodos entomo-patógenos Chilenos y su efectividad en plagas seleccionadas. Pp. 778, in: Resúmenes 50º Congreso Agronómico de la Sociedad Agronómica de Chile. Universidad de la Frontera, Pucón. 835 p.
Georgis, R. 1992. Present and future prospects for entomopathogenic nematode products. Biocon-trol Science and technology. 2:83-99.
Gerding G., M.; A. France y E. Cisternas.2000. Evaluación de cepas nativas de Metarhizium aniso-pliae var. anisopliae sobre Otiorhynchus sulcatus Fab. (Coleoptera: Curculionidae). Agricul-tura Técnica. vol. 60, n. 3 pp. 216-223.
Gómez, C. y A. Lanteri. 2006. Primer registro de Naupactus ruizi (Coleoptera: Curculionoidea) asociado con Pinus ponderosa (Gymnospermae: Pinaceae) en Patagonia. Rev. Soc. Entomol. Argent. [online]. Vol. 65, n. 1-2, pp. 107-109.
http://www.scielo.org.ar/pdf/rsea/v65n1-2/v65n1-2a12.pdf

77
González, H. 1989. Insectos y Acaros de importancia agrícola y cuarentenaria en Chile. Universidad de Chile y BASF S.A. Chile. 310 p.
González, R.H. 1973. Catálogo de las plagas agrícolas en Chile. Universidad de Chile, Facultad de agronomía, Santiago, Chile. 68 p.
González, R.H. 1983. Manejo de plagas de la vid. Universidad de Chile. Facultad de Ciencias Agra-rias, Veterinarias y Forestales. Santiago, Chile. Publicaciones en Ciencias Agrícolas Nº 13. 115 p.
Grewal, P.S. 1998. Formulations of entomopathogenic nematodes for storage and application. Jpn. J. Nematol. 28:68-74
Grewal, P.S.; Georgis, R. 1999. Entoopathogenic nematodes. En: Methods in biotechnology. Vol. 5.
Griffin, C., N. Boemare and E. Lewis. 2005. Biology and behaviour. pp. 47-59 In: P. Grewal, R.-U. Ehlers and D. Shapiro-Ilan (Eds.). Nematode as Biocontrol Agents. CABI Publishing. Wallingford, U.K.
Hazir, S., H. Kaya, S.P. Stock and N. Keskin. 2003. Entomopathogenic nematodes (Steinernematidae and Heterorhabditidae) for biological control of soil pests. Turk. J. Biol. 27(4): 181-202.
Hominick, W. and A. Reid. 1990. Perspectives on entomopathogenic nematology. pp: 327-345. In: R. Gaugler and H. Kaya (Eds.). 23 Entomopathogenic nematodes in biological control. CRC Press. Florida, USA.
Ishibashi, N. and E. Kondo. 1993. Behavior of infective juveniles. Pp. 139-150, in: Entomopathoge-nic nematodes in biological control. R. Gaugler and H. K. Kaya, eds. CRC Press, Florida, USA. 365 p.
Kaya, H.; R. Bedding, and R. Akhurst. 1993. An overview of insect parasit and entomopathogenic nematodes. Pp: 1-10, in: Nematodes and the biological control of insect pests. R. Bedding, R. Akhurst and H. Kaya, eds. Csiro, Victoria, Australia. 178 p.
Kaya, H. and A. Koppenhöfer. 2004. Biological control of insects and other invertebrates with ne-matodes. pp: 1083-1132. In: Z. Chen, S. Chen and D. Dickson (Eds.). Nematology: advan-ces and perspectives. Volume 2. Nematode management and utilization. CABI Publishing. Wallingford, UK.
Klein, C. and D. Waterhouse. 2000. The distribution and importance of arthropods associated with agriculture and forestry in Chile (Distribución e importancia de los artrópodos asociados a la agricultura y silvicultura en Chile). ACIAR Monograph Nº 68. 231 pp.
Koppenhöfer, A., H. Kaya. 1999. Ecological characterization of Steinernema rarum. J. Invertebr. Pa-thol. 73(1): 120-128.
Lecuona, R. 1996. Microorganismos patógenos empleados en el control microbiano de insectos plagas. Talleres gráficos Mariano Mas, Buenos Aires, Argentina. 338 p.
Li, X.-Y., R. Cowles, E. Cowles, R. Gaugler and D. Cox-Foster. 2007. Relationship between the succesful infection by entomopathogenic nematodes and the host inmune response. Int. J. Parasitol. 37(3-4): 365-374.
Luppichini, P., N. Olivares y A. France. 2012. Control biológico de burrito de la vid con nemátodos entomopatógenos. Revista Redagrícola. Nº 49. p. 82-84.
Luppichini, P. y N. Olivares. 2012. Monitoring and Management of Naupactus (= Asynonychus)
Bibliografía

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
78
cervinus in Chilean citrus orchards. XII Citrus Congress. Valencia, España. 19 al 23 de noviembre de 2012. Libro de resumenes p. 296.
Lewis, E. and D. Clarke, 2012. Nematode Parasites and Entomopathogens. Pp. 395-426, in: F.Vega and H. Kaya. Insect Pathology, 2nd ed. Academic Press, CA, USA. 508 p.
Merino L. y A. France. 2009. Nemátodos entomopatógenos: Control biológico de insectos plaga de importancia económica. Revista Tierra Adentro. Mayo–junio 2009. p. 24-25.
Olivares, N., P. Luppichini y J. Montenegro. 2012. Reconocimiento, monitoreo y manejo de Naupac-tus cervinus (Boheman) en cítricos. Ficha técnica INIA Nº 7. Disponible on line en: http://www.inia.cl/medios/lacruz/Pdf/ficha7.pdf
Olivares N., P. Luppichini, J. E. Ortúzar y J. Montenegro. 2012. Manejo de Naupactus cervinus (Bo-heman) en cítricos. Revista Redagrícola. Nº 48. p. 82-83.
Pérez, H. 1994. Descripción de aspectos morfológicos, biológicos y de comportamiento de Aegorhi-nus superciliosus (Coleoptera: Curculionidae). Tesis de Licenciado en Agronomía, Universi-dad Austral de Chile, Valdivia. 102 p.
Prado, E. 1991. Artrópodos y sus enemigos naturales asociados a plantas cultivadas en Chile. Bole-tín Técnico Nº 169. INIA, CRI La Platina. Santiago. Chile. 203 p.
Prado, E., R. Ripa y F. Rodríguez. 2000. Insectos y ácaros. In: Uva de Mesa en Chile. (J. Valenzuela Ed.). Colección libros INIA Nº 5. Centro Regional de Investigación La Platina. Santiago. Chile. 234-250 p.
Poinar, G. and R. Poinar. 2008. What bugged the dinosaurs? Insects, diseas, and death in the Creta-ceous. Princeton University Press, NJ, USA. 264 p.
Ramos-Rodríguez, O., J. Campbell, E. Lewis, D. Shapiro-Ilan and S. Ramaswamy. 2007. Dynamics of carbon dioxide release from insects infected with entomopathogenic nematodes. J. In-vertebr. Pathol. 94(1): 64-69.
Ripa, R. 1992. Burrito de los frutales Naupactus xanthographus (German). Boletín Técnico Nº 192. Estación experimental La Platina. Subestación Experimental La Cruz. Instituto de Investi-gaciones Agropecuarias. 74 p.
Ripa, R. y P. Luppichini. 2008. Capachito de los frutales. p. 158-162. En: Capítulo 8, Ripa, R.; P. Larral (eds). Manejo de Plagas en Paltos y Cítricos. Colección Libros INIA Nº 23. INIA La Cruz. La Cruz, Chile.
Ripa, R. 2008. Burrito de la vid o Burritos de los frutales. p. 155-158. EN: Capítulo 8, Ripa, R.; P. Larral (ed). Manejo de Plagas en Paltos y Cítricos. Colección Libros INIA Nº 23. INIA La Cruz. La Cruz, Chile.
Ripa, R. y P. Luppichini. 2010. Burrito de los frutales (Naupactus xanthographus). Ficha Técnica Nº 6. INIA / CODESSER. Disponible en: http://www.inia.cl/medios/lacruz/Pdf/Ficha6Burrito-delosfrutales.pdf
Ripa R. y P. Luppichini. 2010. Manejo de plagas de la Vid. Libro Colección INIA Nº 26. 145 p.
Rozas A. 2009. Nemátodos Entomopatógenos Nativos para el control de Aegorhinus superciliosus (Guérin) (Coleoptera: Curculionidae). Tesis de Ingeniero Agrónomo, Universidad Con-cepción, Chillán. 33 p.

79
Sandoval A. y A. France.1998. Evaluación del hongo Beauveria spp. Como potencial agente con-trolador del coleóptero Asynonychus cervinus (Boheman). Tesis de Ingeniero Agrónomo, Universidad Adventista de Chile, Chillán. 42 p.
Sazo, L. 1996. Manejo de curculiónidos de importancia económica asociados a frutales de hoja ca-duca y vid. In: Avances en sanidad vegetal de frutales y vides. 1996. Universidad de Chile. Fac. de Ciencias Agrarias y Forestales, Santiago. 63-65 p.
Shapiro-Ilan, Lewis, Son, & Tedders. 2003. Survey of Entomopathogenic Nematodes and Fungi Endemic to Pecan Orchards of the Southeastern United States and Their Virulence to the Pecan Weevil (Coleoptera: Curculionidae). J. Invertebr. Pathol. 187-19.
Sicard, M., J-B. Ferdy, S. Pagés, N. Le Brun, B. Godelle, N. Boemare and C. Moulia. 2004. When mutualists are pathogens: an experimental study of the symbioses between Steinernema (entomopathogenic nematodes) and Xenorhbdus (bacteria). J. Evol. Biol. 17(5): 985-993.
Soler, D.M., L. Gómez, L. Sánchez. 2003. Formulación de nemátodos entomopatógenos. Rev. Pro-tección Veg. 18:1: 7-14.
Stoc S. y A. Koppenhöfer. 2003. Steirneirnema scarabei n. ap. (Rhabditida: Steinernematidae), a natural pathogen of scarab beetle larvae (Coleoptera Scarabeidae) from New Jersey, USA. Nematology 5, 191-204.
Tailliez, P., S. Pagés, N. Ginibre and N. Boemare. 2006. New insight into diversity in the genus Xenorhabdus, including the description of ten novel species. Int. J. Syst. Evol. Microbiol. 56(12): 2805-2818.
Urtubia I., A. France y P. Luppichini. 2012. Selection of native isolates of entomopathogenic nema-todes to control the Chilean grape weevil (Naupactus xanthographus). Congreso Internacio-nal: Invertebrate Pathology and Microbial Control & 45th Annual Meeting of the Society for Invertebrate Pathology (SIP). Buenos Aires, Argentina 05 al 09 de agosto de 2012.
Vázquez, J. 1977. Antagonistas de larvas Scarabeidae presentes en las praderas de la Provincia de Valdivia. Tesis de Ingeniero Agrónomo. Universidad Austral, Valdivia, Chile. 50 p.
Wang, Bilgrami, Shapiro-Ilan & Gaugler, 2006. Stability of entomopathogenic bacteria, Xenorhab-dus nematophila and Photorhabdus luminescens, during in vitro culture J. Ind. Microbiol. and Biotechno.
Woodring, J. L., and H. K. Kaya. 1988. Steinernema and Heterorhabditid nematodes: a handbook of biology and techniques. Southern Cooperative Series Bulletin 331. Arkansas Agric. Exp. Station, Fayetteville, Arkansas, USA. 30 p.
Wouts, W. M. 1991. Steinernema (Neoaplectana) and Heterorhabditis species. Pp. 855-897, in: Manual of agricultural nematology. W. R. Nickle, ed. Marcel Dekker, New York, USA. 1035 p.
Bibliografía

Manejo del Burrito de la vid y otros curculiónidos asociados a vides
80


















