
Mecanismos de reparación de lesiones en el...
47
Mecanismos de reparación de lesiones en el DNA
Transcript of Mecanismos de reparación de lesiones en el...

Mecanismos de reparación de lesiones en el DNA

Además, existen secuencias
naturales que adoptan
conformaciones diferentes
de la clásica doble hélice del
DNA (B-DNA):
Z-DNA, H-DNA, G-cuartetos,
cruciformes, horquillas, y
series de uno o mas
nucleótidos repetidos.
Lesiones espontáneas, causadas por actividades
metabólicas, o por agentes en el medio ambiente:
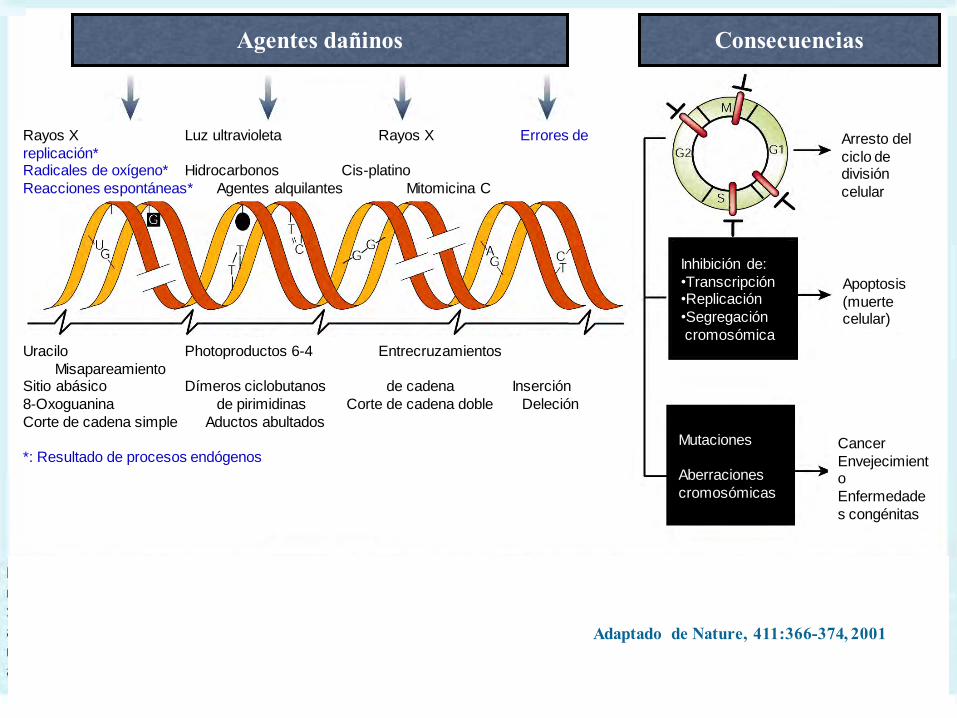
Adaptado de Nature, 411:366-374, 2001
Rayos X Luz ultravioleta Rayos X Errores de
replicación*Radicales de oxígeno* Hidrocarbonos Cis-platino
Reacciones espontáneas* Agentes alquilantes Mitomicina C
Agentes dañinos Consecuencias
Arresto del
ciclo de división
celular
Apoptosis
(muerte celular)
Cancer
Envejecimiento
Enfermedade
s congénitas
Inhibición de:
•Transcripción•Replicación
•Segregación
cromosómica
Mutaciones
Aberraciones
cromosómicas
Uracilo Photoproductos 6-4 Entrecruzamientos
MisapareamientoSitio abásico Dímeros ciclobutanos de cadena Inserción
8-Oxoguanina de pirimidinas Corte de cadena doble Deleción
Corte de cadena simple Aductos abultados
*: Resultado de procesos endógenos

Diversidad del sistema inmune: la conmutación
de clase de las inmunoglobulinas depende de la
actividad de enzimas que cortan y empalman el DNA
del gen de la cadena pesada, creando un isotipo
diferente.
Algunas mutaciones pueden resultar ventajosas y
permiten la evolución de las especies..
Terapias contra el cáncer basadas en agentes
genotóxicos.
G. Spivak©
No todas las lesiones en el DNA son dañinas

Mecanismos de reparación del DNA
Reversión enzimática
Reparación por escisión de bases (BER)
Reparación por escisión de nucleótidos (NER)
Reparación de desapareamientos (MMR)
Reparación de cortes de cadena simple
Reparación post-replicación
Reparación de cortes de cadena doble
G. Spivak©
Reparación de entrecruzamientos de cadenas

Reparación de dímeros de pirimidinas por reversión directa:
fotoreactivación
timinas o
citosinas
fotodímero
luz
ultravioleta
240-260 nm
fotoliasa y
luz blanca
(300-500 nm)
Fotoliasas específicas para los dímeros ciclobutanos y los 6-4 fotoproductos.
Codificadas en fagos; activas en la mayoría de bacterias y algunos vertebrados
(marsupiales) pero no en mamíferos placentarios.
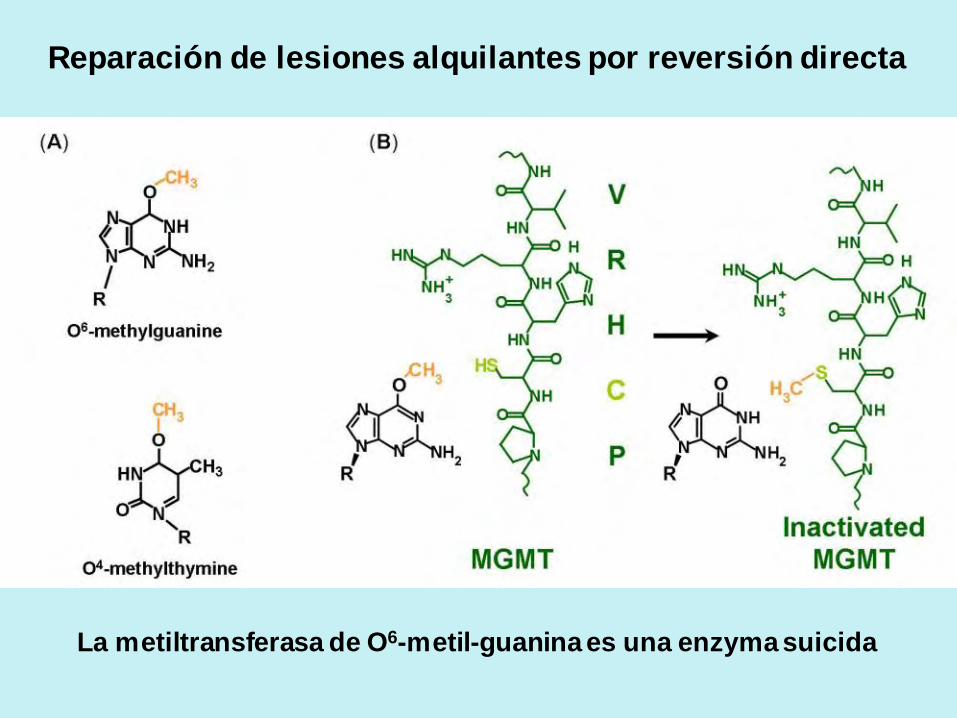
Reparación de lesiones alquilantes por reversión directa
La metiltransferasa de O6-metil-guanina es una enzyma suicida

Mecanismos de reparación del DNA
Reversión enzimática
Reparación por escisión de bases (BER)
Reparación por escisión de nucleótidos (NER)
Reparación de desapareamientos (MMR)
Reparación de cortes de cadena simple
Reparación post-replicación
Reparación de cortes de cadena doble
G. Spivak©
Reparación de entrecruzamientos de cadenas
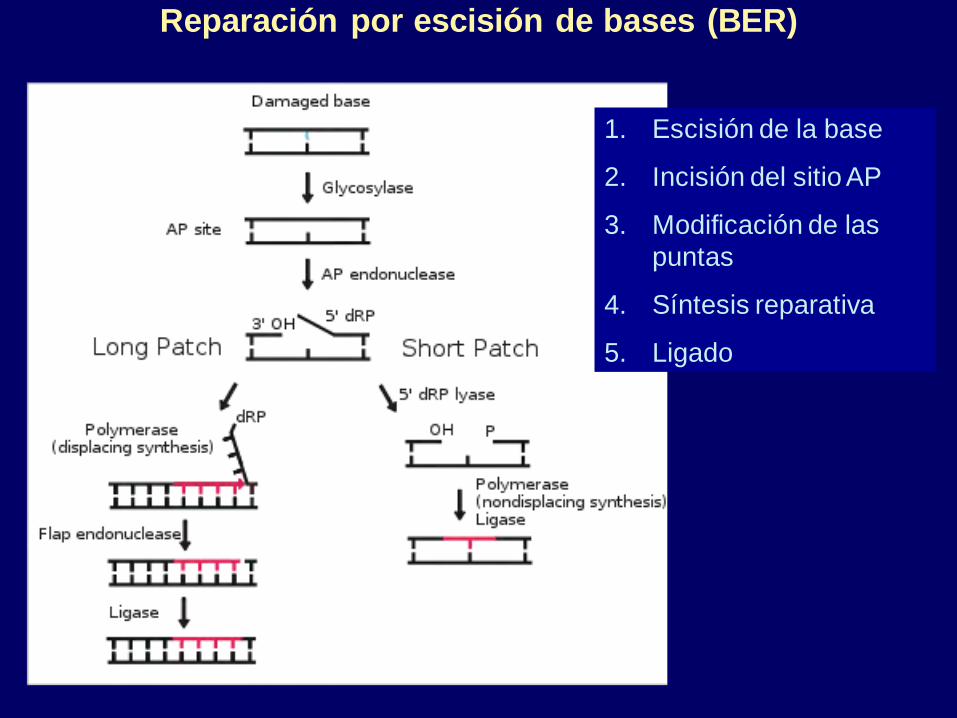
Reparación por escisión de bases (BER)
1. Escisión de la base
2. Incisión del sitio AP
3. Modificación de las
puntas
4. Síntesis reparativa
5. Ligado

8-oxoG
Sp
Gh
FapyG MeFapyG
8-oxoA
FapyA
Tg DHT 5OHMH
5OHC 5OHU DHU
Las Lesiones
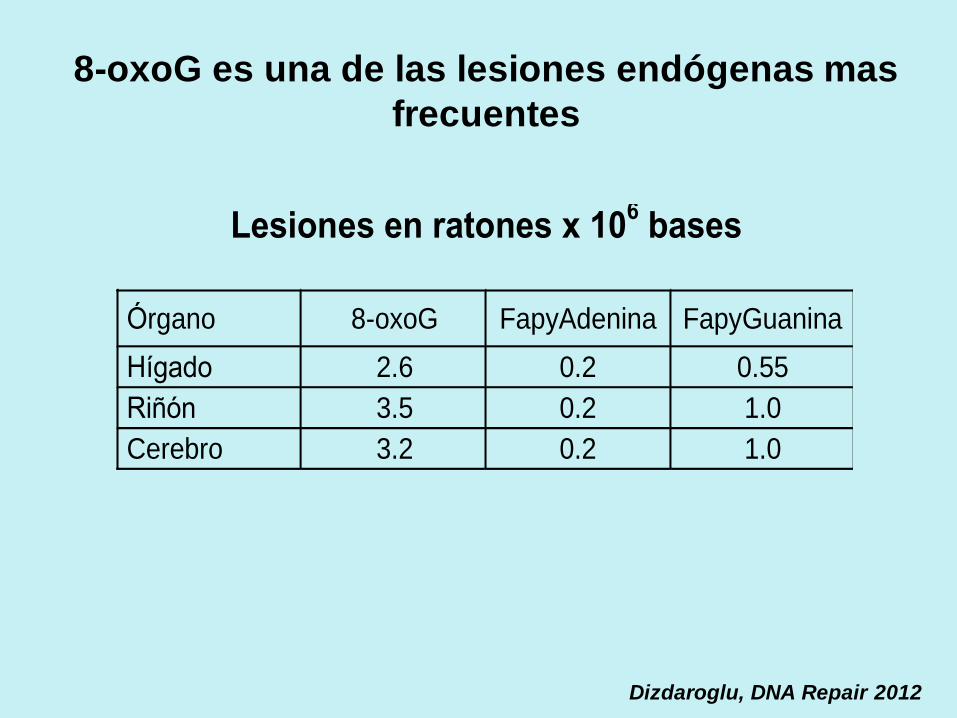
8-oxoG es una de las lesiones endógenas mas
frecuentes
Dizdaroglu, DNA Repair 2012
Lesiones en ratones x 106 bases
Órgano 8-oxoG FapyAdenina FapyGuanina
Hígado 2.6 0.2 0.55
Riñón 3.5 0.2 1.0
Cerebro 3.2 0.2 1.0

C
G
C
GO
A
GO
GO
C
GO
C
C
G
C
GO
MutM (Fpg)
reparación
reparación
MutY
replicación
oxidación
Tres mecanismos para procesar oxidación de guanina en E. coli
dGTP 8-oxodGTP 8-oxodGMPoxidación MutT
(hMYH)
(MTH1)
(hNeil1,
hNeil2,
hNeil3)
hOGG1

Aunque los productos de oxidación del DNA
son los daños mas frecuentes, hay otras
lesiones y sustratos de BER
Alquilación (1 o 2 carbonos)
Pérdida espontánea de la base
Desaminación de citosina uracilo
Incorporación de ribonucleótidos en DNA
Roturas de cadena
Total ≥ 20.000 lesiones / célula / dia

Glicosilasas que reconocen y remueven bases modificadas en el DNA de humanos
hNTH1 Tg, hoU, hoC, urea, FapyG
hOGG1 8-oxoG, FapyG
hMYH A:8-oxoG
hNEIL1 Tg, hoU, hoC, urea, FapyG, FapyA
hNEIL2 sitio AP, hoU, 8-oxoG en cadena simple
hNEIL3 Tg, hoU, hoC, urea, FapyG, FapyA
UNG U
hSMUG1 U, hoU, hmU, fU
TDG T:G desapareamiento
MBD4 T:G desapareamiento
Aag 3-meA, 7-meG, hipoxantina, etenoA
MPG 3-meA, hipoxantina
AlkB reversión de bases metiladas en cadena simple
& productos de superoxidaciónde guanina

Mecanismos de reparación del DNA
Reversión enzimática
Reparación por escisión de bases (BER)
Reparación por escisión de nucleótidos (NER)
Reparación de desapareamientos (MMR)
Reparación de cortes de cadena simple
Reparación post-replicación
Reparación de cortes de cadena doble
G. Spivak©
Reparación de entrecruzamientos de cadenas
Reparación por escisión de ribonucleótidos (RER)
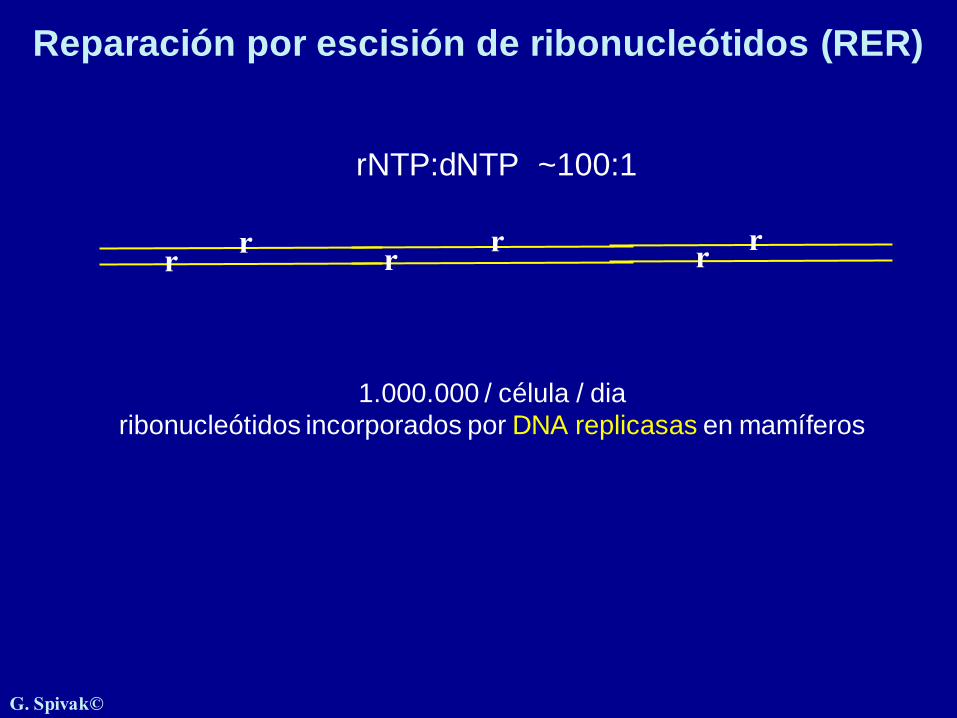
Reparación por escisión de ribonucleótidos (RER)
G. Spivak©
r
1.000.000 / célula / dia
ribonucleótidos incorporados por DNA replicasas en mamíferos
rr
rNTP:dNTP ~100:1
rr r

Reparación por escisión de ribonucleótidos (RER)
G. Spivak©
r
Ribonucleasa H2
DNA pol β
Flap endonucleasa FEN1
ligasa
Thomas Kunkel

Mecanismos de reparación del DNA
Reversión enzimática
Reparación por escisión de bases (BER)
Reparación por escisión de nucleótidos (NER)
Reparación de desapareamientos (MMR)
Reparación de cortes de cadena simple
Reparación post-replicación
Reparación de cortes de cadena doble
G. Spivak©
Reparación de entrecruzamientos de cadenas

Medio ambiente + estilo de vida: cientos de lesiones en DNA
NER no necesita glicosilasas específicas
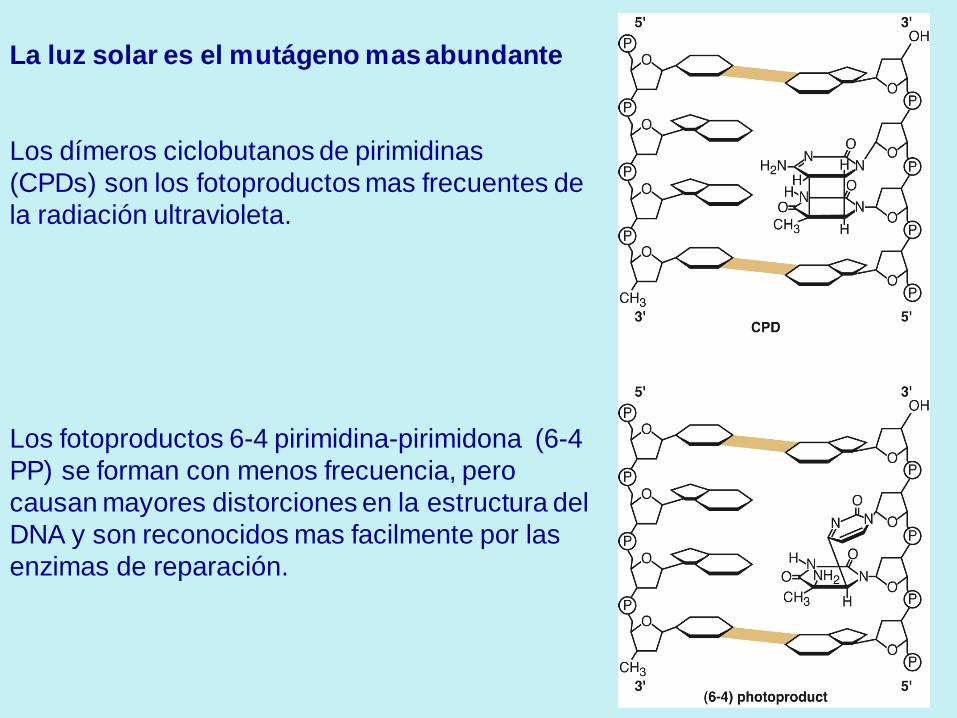
La luz solar es el mutágeno mas abundante
Los dímeros ciclobutanos de pirimidinas
(CPDs) son los fotoproductos mas frecuentes de
la radiación ultravioleta.
Los fotoproductos 6-4 pirimidina-pirimidona (6-4
PP) se forman con menos frecuencia, pero
causan mayores distorciones en la estructura del
DNA y son reconocidos mas facilmente por las
enzimas de reparación.

Benzo(a)pireno Benzo(g)criseno
Los hidrocarbonos policíclicos aromáticos (PAH) son
convertidos en metabolitos reactivos que forman aductos
de purinas.
Benzopireno diol
epóxido (BPDE) B(g)CDE

Ejemplos de lesiones “abultadas” reparadas por NER
Luz UV Dímeros de pirimidinas
Tabaco, humo Hidrocarbonos aromáticos policíclicos
Oxidación Ciclopurinas, peroxidación de lípidos
Alquilación Grupos alquilos de >2 carbonos
Entrecruzamientos con proteínas
Quimioterapia Ligado de bases vecinas por cisplatino: GG, GNG
Mostazas de Aductos alquilantes de purinas
nitrógeno
⌃ ⌃

Reconocimiento de la lesión por UvrA
UvrB es reclutado por UvrA
Complejo de pre-incisión
UvrC es reclutado por UvrBIncisión doble por UvrC
Escisión por UvrD
Síntesis de reparación y ligado
UvrC
UvrD
LigasePol I
(UvrA)2
(UvrB)2
(UvrB)2
Reparación por escisión de nucleótidos (NER) en E. coli
(UvrA)2

XPE-DDB2 ?centrin1
centrin1
centrin1
XPF-ERCC1
Ligase I
NER global en seres humanos
x
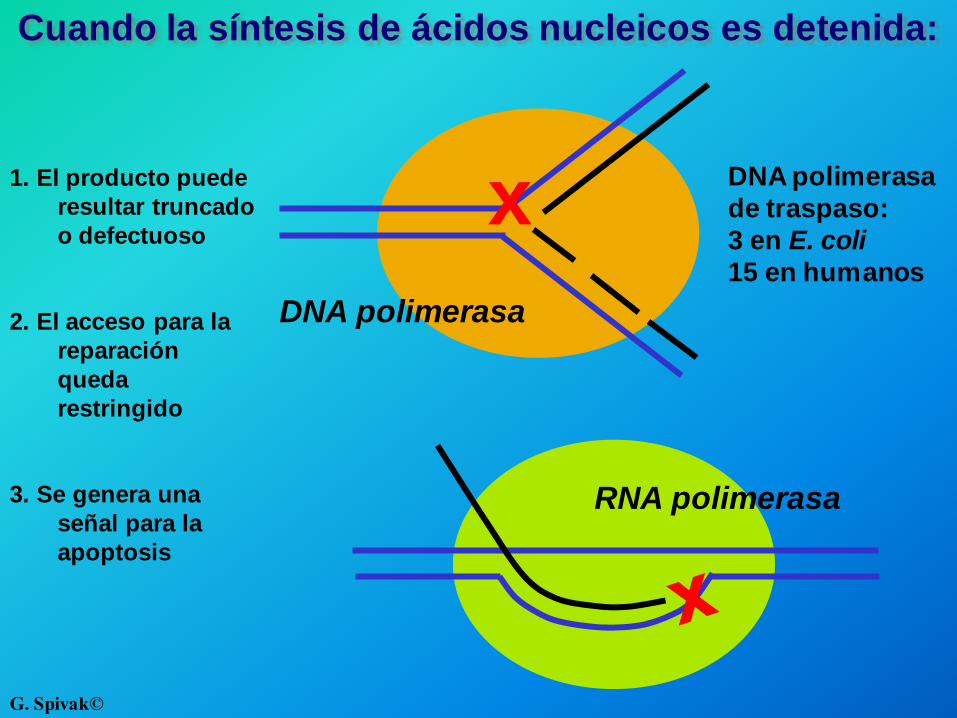
Cuando la síntesis de ácidos nucleicos es detenida:
RNA polimerasa
DNA polimerasa
1. El producto puede
resultar truncado
o defectuoso
2. El acceso para la
reparación
queda
restringido
3. Se genera una
señal para la
apoptosis
G. Spivak©
DNA polimerasa
de traspaso:
3 en E. coli
15 en humanos

a. Pausa y continuación;
mutagénesis transcripcional
b. Retroceso, corte del RNA naciente,
1. probar de nuevo
2. reparar y continuar
Cuando la transcripción se detiene: Opciones?
c. Remoción y destrucción de la
polimerasa y del mRNA, reparación.
Ub
señal para apoptosis
muerte celular
G. Spivak©

TC
RG
GR
(UvrA)2
(UvrB)2
XMfd
RNAP
RNA
(UvrA)2
UvrC
UvrD
LigasePol I
UvrA
Mfd
(UvrB)2
(UvrB)2
Reparación por escisión de nucleótidos (NER) en E. coli
La RNA polimerasa se detiene al
llegar a una lesiónMfd se une a la RNAP y recluta UvrA
RNAP y el RNA son liberados
UvrB es reclutado por UvrA
Reconocimiento de la lesión por UvrA y verificación por UvrB
Complejo de pre-incisión
UvrC es reclutado por UvrBIncisión doble por UvrC
Escisión por UvrD
Síntesis de reparación y ligado

Reparación del gen lacZ en E. coli irradiadas con UV

3’ 5’
RPA
XPF-ERCC1
XPC-HR23B-Centrin
XPE-DDB2
TFIIH
XPA
XPG
RNA
RNAPII
CSACSBUVSSA
XAB2TFIIS
GGR TCR
XPDTFIIHXPB
Reparación por escisión de nucleótidos (NER) en humanos
HMGN1p300
USP7
remoción del 30-mero con la lesión, síntesis, ligado

RNAPII
nuclear membrane
nuclear
pore RNA export
complex
RNA biogenesis
complexes elongation factors
UVSS
A
USP7
CSB
reassembled
nucleosomes
disassembled
nucleosome
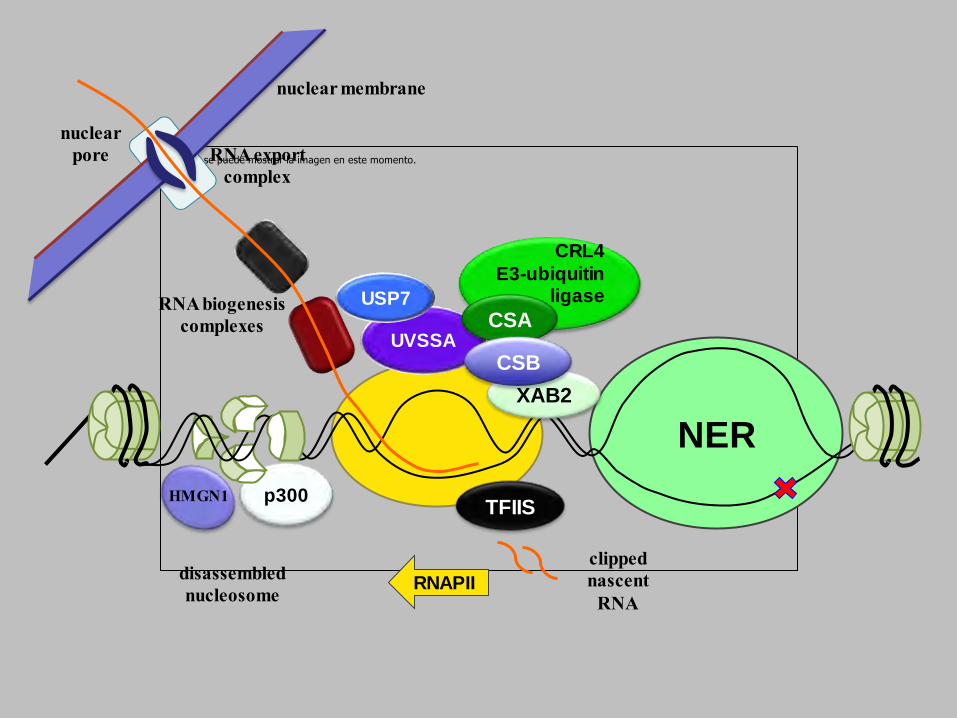
HMGN1 p300
CRL4
E3-ubiquitin ligase
NER
RNAPII
nuclear membrane
nuclear
pore RNA export
complex
RNA biogenesis
complexesUVSSA
USP7
disassembled
nucleosome
CSA
TFIIS
XAB2
clipped
nascent
RNA
CSB

Reparación por excisión de nucleótidos: dos caminos
Reparación global del genomio (GGR)
• Lesiones en todo el genomio
• La eficiencia depende del grado de distorsión creado por la lesión,
de la secuencia local, de la estructura cromatínica y de otros
factores
Reparación acoplada a la transcripción (TCR)
• Lesiones en cadenas transcritas de genes activos
• Requiere que la RNA polimerasa esté configurada para elongación
• La RNA polimerasa detenida recluta enzimas de la reparación
G. Spivak©

¿Por qué existe la reparación acoplada a la transcripción?
TCR es importante para la sobrevivencia celular porque:
• Remueve RNA polimerasas bloqueadas que podrían interferir con
la replicación.
• Remueve lesiones que interrumpen la transcripción de genes
esenciales.
• Elimina una señal muy fuerte para iniciar apoptosis.

Mecanismos de reparación del DNA
Reversión enzimática
Reparación por escisión de bases (BER)
Reparación por escisión de nucleótidos (NER)
Reparación de desapareamientos (MMR)
Reparación de cortes de cadena simple
Reparación post-replicación
Reparación de cortes de cadena doble
G. Spivak©
Reparación de entrecruzamientos de cadenas

Mecanismo conservado en organismos de unicelulares a
humanos. Corrige errores durante la replicacion del DNA.
Asociado con el replisoma, repara únicamente la cadena
hija para conservar la secuencia original.
Requiere detección de nucleótidos normales, apareados
incorrectamente.
Sustratos incluyen los 8 desapareamientos de bases
posibles, ademas de horquillas causadas por inserciones
o deleciones.
Reparación de desapareamientos
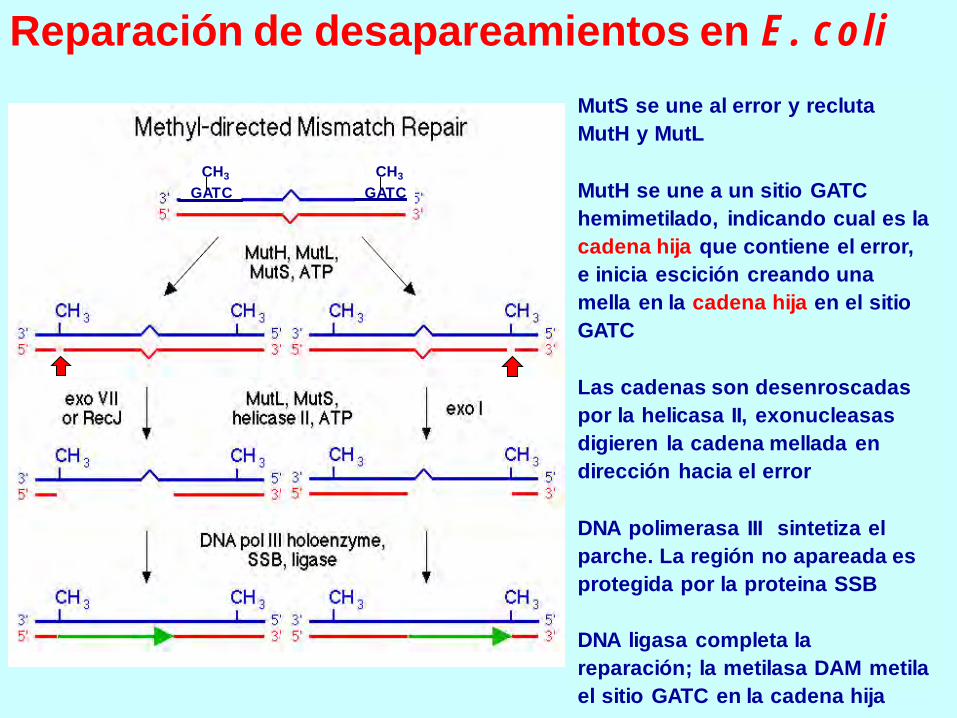
MutS se une al error y recluta
MutH y MutL
MutH se une a un sitio GATC
hemimetilado, indicando cual es la
cadena hija que contiene el error,
e inicia escición creando una
mella en la cadena hija en el sitio
GATC
Las cadenas son desenroscadas
por la helicasa II, exonucleasas
digieren la cadena mellada en
dirección hacia el error
DNA polimerasa III sintetiza el
parche. La región no apareada es
protegida por la proteina SSB
DNA ligasa completa la
reparación; la metilasa DAM metila
el sitio GATC en la cadena hija
Reparación de desapareamientos en E. coli
GATC GATC
CH3
GATC
CH3

Reparación de desapareamientos en humanos
El mecanismo y la maquinaria son similares pero no hay metilación de GATC.
La cadena hija se distingue porque
• es discontinua?
• contiene ribonucleótidos?
E. coli Homólogos humanos
MutS heterodímeros Msh2 / Msh6 = MutSα
Msh2 / Msh3 = MutSβ
MutL heterodímeros 2x Mlh1 / Pms1 = MutLα
Mlh1 / Pms1 = MutLβ
Mlh1 / Mlh3 = MutLγ
MutH no homólogos; la función de endonucleasa es
llevada a cabo por los homólogos de MutL

Mecanismos de reparación del DNA
Reversión enzimática
Reparación por escisión de bases (BER)
Reparación por escisión de nucleótidos (NER)
Reparación de desapareamientos (MMR)
Reparación de cortes de cadena simple
Reparación post-replicación
Reparación de cortes de cadena doble
G. Spivak©
Reparación de entrecruzamientos de cadenas
ligado
BER
NER

Mecanismos de reparación del DNA
Reversión enzimática
Reparación por escisión de bases (BER)
Reparación por escisión de nucleótidos (NER)
Reparación de desapareamientos (MMR)
Reparación de cortes de cadena simple
Reparación post-replicación
Reparación de cortes de cadena doble
G. Spivak©
Reparación de entrecruzamientos de cadenas

Fases G1 / S: fusión no homóloga de extremos (NHEJ)
Fases S / G2: recombinación homóloga (HR)
G. Spivak©
Causas naturales:
o recombinación meiótica y mitótica
o reordenación V (D) J de inmunoglobulinas
o replicación del DNA
o ROS productos del metabolismo
Agentes ambientales:
o radiaciones ionizantes
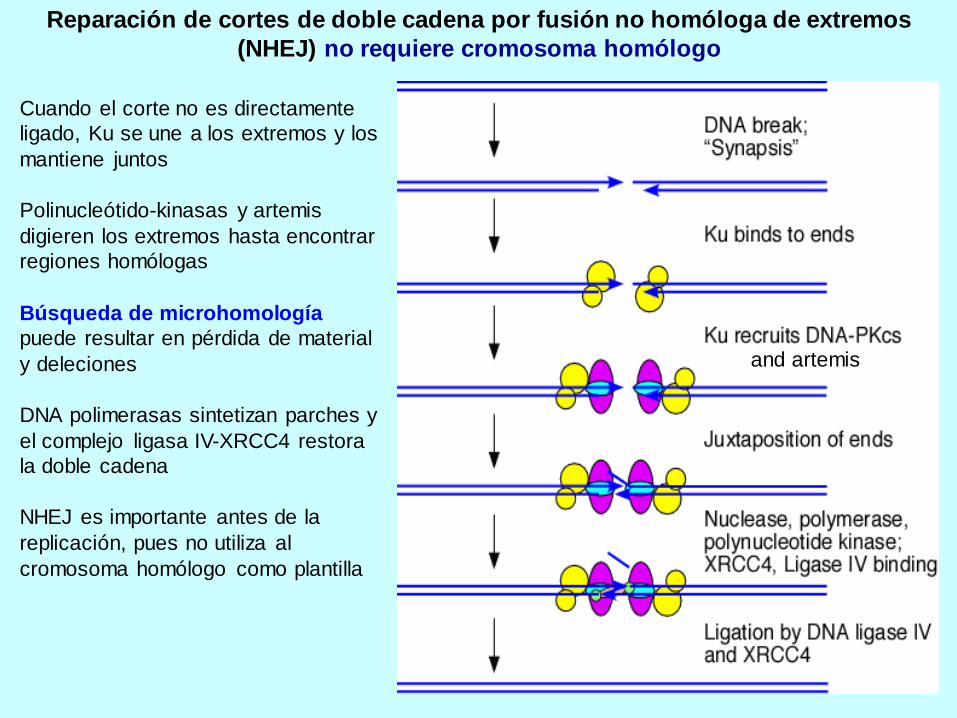
Reparación de cortes de doble cadena por fusión no homóloga de extremos
(NHEJ) no requiere cromosoma homólogo
Cuando el corte no es directamente
ligado, Ku se une a los extremos y los
mantiene juntos
Polinucleótido-kinasas y artemis
digieren los extremos hasta encontrar
regiones homólogas
Búsqueda de microhomología
puede resultar en pérdida de material
y deleciones
DNA polimerasas sintetizan parches y
el complejo ligasa IV-XRCC4 restora
la doble cadena
NHEJ es importante antes de la
replicación, pues no utiliza al
cromosoma homólogo como plantilla
and artemis

Reparación de cortes de doble cadena por recombinación homóloga requiere grandes regiones de homología
nucleasa, helicasa
invasión del cromosoma homólogo
síntesis copiando el cromosoma homólogo
síntesis reparando el cromosoma dañado
ligado
resolvasa, ligasa
unión Holliday de hebras cruzadas
5’5’
restitución de la secuencia original

Mecanismos de reparación del DNA
Reversión enzimática
Reparación por escisión de bases (BER)
Reparación por escisión de nucleótidos (NER)
Reparación de desapareamientos (MMR)
Reparación de cortes de cadena simple
Reparación post-replicación
Reparación de cortes de cadena doble
G. Spivak©
Reparación de entrecruzamientos de cadenas
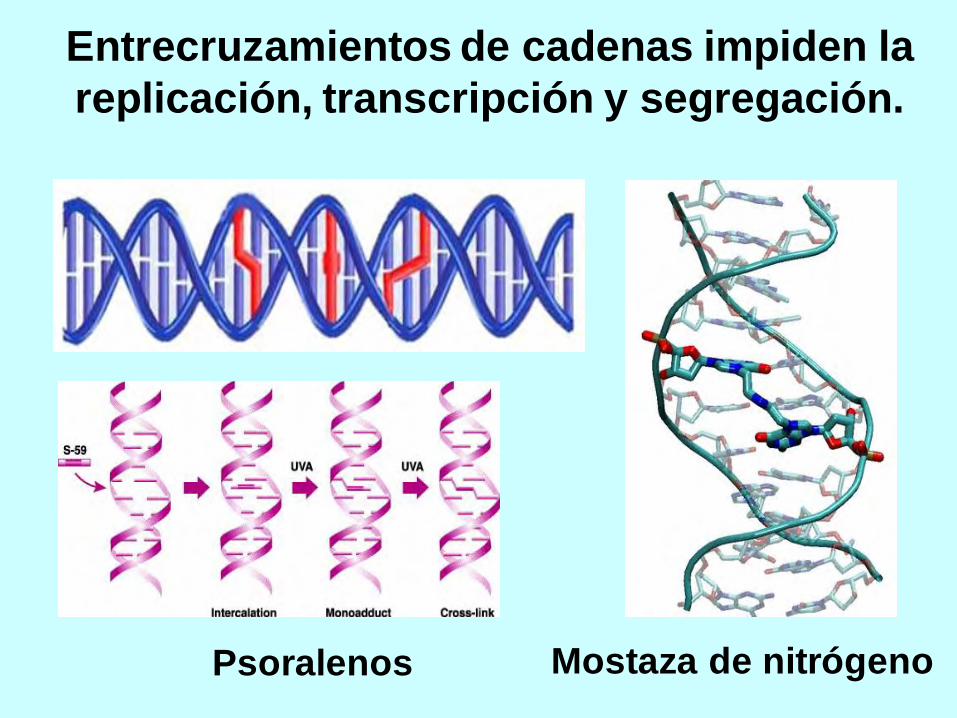
Entrecruzamientos de cadenas impiden la
replicación, transcripción y segregación.
Psoralenos Mostaza de nitrógeno
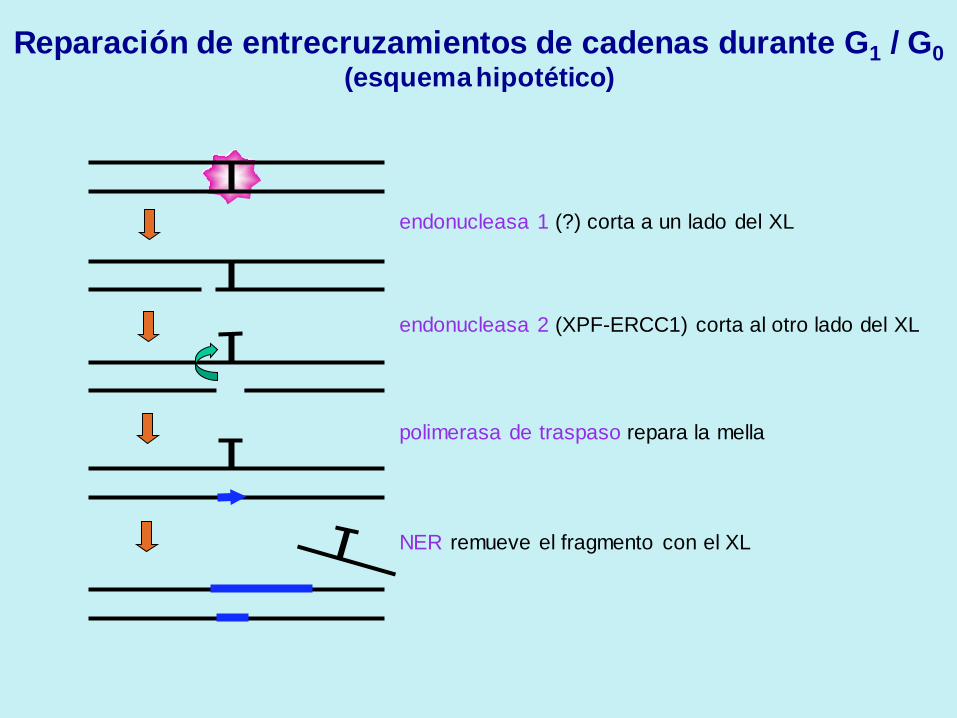
endonucleasa 1 (?) corta a un lado del XL
endonucleasa 2 (XPF-ERCC1) corta al otro lado del XL
polimerasa de traspaso repara la mella
NER remueve el fragmento con el XL
Reparación de entrecruzamientos de cadenas durante G1 / G0
(esquema hipotético)
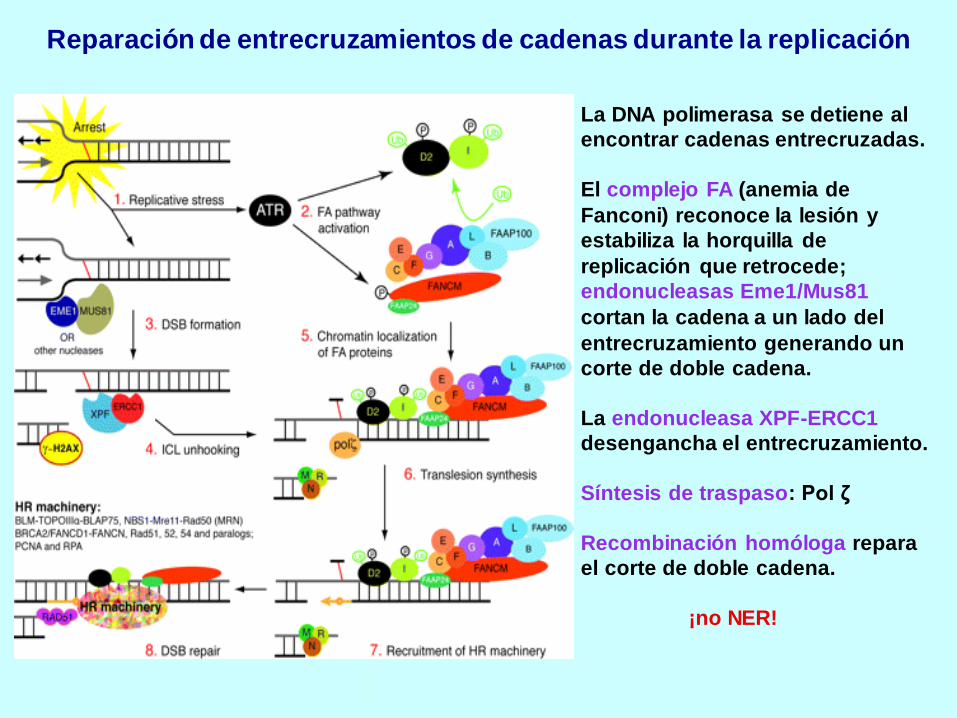
Reparación de entrecruzamientos de cadenas durante la replicación
La DNA polimerasa se detiene al
encontrar cadenas entrecruzadas.
El complejo FA (anemia de
Fanconi) reconoce la lesión y
estabiliza la horquilla de
replicación que retrocede;
endonucleasas Eme1/Mus81
cortan la cadena a un lado del
entrecruzamiento generando un
corte de doble cadena.
La endonucleasa XPF-ERCC1
desengancha el entrecruzamiento.
Síntesis de traspaso: Pol ζ
Recombinación homóloga repara
el corte de doble cadena.
¡no NER!



















