
RELACIONES HORMONALES Y DINAMICA … · expresión de buena eficiencia reproductiva en individuos...
102
RELACIONES HORMONALES Y DINAMICA FOLICULAR DURANTE EL PERIODO POSPARTO EN VACAS SANMARTINERO GIOVANNI MAURICIO BAEZ SANDOVAL UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE MEDICINA VETERINARIA Y DE ZOOTECNIA PROGRAMA DE POSGRADO MAESTRIA EN PRODUCCION ANIMAL BOGOTA D.C. 2010
-
Upload
trinhduong -
Category
Documents
-
view
222 -
download
0
Transcript of RELACIONES HORMONALES Y DINAMICA … · expresión de buena eficiencia reproductiva en individuos...

1
RELACIONES HORMONALES Y DINAMICA FOLICULAR DURANTE EL
PERIODO POSPARTO EN VACAS SANMARTINERO
GIOVANNI MAURICIO BAEZ SANDOVAL
UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE MEDICINA VETERINARIA Y DE ZOOTECNIA
PROGRAMA DE POSGRADO MAESTRIA EN PRODUCCION ANIMAL
BOGOTA D.C. 2010

2
RELACIONES HORMONALES Y DINAMICA FOLICULAR DURANTE EL
PERIODO POSPARTO EN VACAS SANMARTINERO
GIOVANNI MAURICIO BAEZ SANDOVAL Código: 780163
Trabajo de grado presentado para optar al título de:
Maestro en Ciencias – Producción Animal
Director: Henry Alberto Grajales Lombana. PhD. Profesor Asociado. UN
Codirector: José Henry Velásquez Penagos. MSc. Investigador. CORPOICA
UNIVERSIDAD NACIONAL DE COLOMBIA FACULTAD DE MEDICINA VETERINARIA Y DE ZOOTECNIA PROGRAMA DE POSGRADO
MAESTRIA EN PRODUCCION ANIMAL BOGOTA D.C.
2010

3
Nota de aceptación:
___________________________________ ___________________________________ ___________________________________ ___________________________________ ___________________________________ ___________________________________
___________________________________ Presidente del Jurado
___________________________________ Jurado
___________________________________ Jurado
___________________________________ Jurado
Bogotá D.C. 29 de Enero de 2010

4
Dedico este trabajo a mi hija Camila y a Ingrid, y se los ofrezco a cambio del
tiempo que deje de compartir con ellas por estar dedicado a mis estudios de
maestría.
A mis padres y mi hermana.
A todas las personas que me quieren y me estiman.

5
AGRADECIMIENTOS
Expreso mis agradecimientos al Dr. Henry Grajales, por su dirección y por la
oportunidad que me dio al reclutarme para la ejecución de este proyecto de
investigación.
A la Universidad Nacional de Colombia, por brindarme la oportunidad de realizar
mis estudios de pregrado y posgrado, con los beneficios que me fueron otorgados.
A los doctores Agustín Góngora, de la Universidad de los llanos, Guillermo
Velásquez, del C.I. CORPOICA La Libertad, y Harvey Lozano, de la Universidad
Nacional de Colombia, por las recomendaciones y sugerencias para esta tesis.
Al Dr. Aureliano Hernández, por sus aportes y por permitirme pertenecer a su
grupo de investigación.
Al Dr. Henry Velásquez, del C.I. CORPOICA La Libertad, por hacer parte de toda
la gestión financiera, logística y técnica del proyecto.
Al Dr. Marcel Amstalden, de Texas A&M University, por el entrenamiento que me
brindo en técnicas de radioinmunoanálisis.
A todos mis compañeros y amigos del posgrado, que en algún momento me
colaboraron.
Al personal del centro de investigaciones CORPOICA La Libertad.
A todas las personas que me brindaron su apoyo y motivación.

6
DECLARATORIA DE ORIGINALIDAD Y RECONOCIMIENTO
Declaro que el trabajo reportado en esta tesis fue llevado a cabo con recursos de
la Universidad Nacional de Colombia y CORPOICA.
El trabajo de toma de muestras sanguíneas fue realizado por el autor, con la
colaboración del personal del centro de investigaciones CORPOICA La Libertad.
Las determinaciones de progesterona y estradiol fueron realizadas por el autor en
el Laboratorio de Hormonas de la Facultad de Medicina Veterinaria y de Zootecnia
de la Universidad Nacional de Colombia.
____________________________.
Giovanni M. Báez S.

7
CONTENIDO
pág.
INTRODUCCION 15
BIBLIOGRAFIA 19
1. REVISION DE LITERATURA 20
1.1 ANESTRO POSPARTO EN EL TROPICO 20
1.2 FISIOLOGIA DEL POSPARTO 20
1.2.1 Nutrición 21
1.2.1.1 Balance energético 21
1.2.1.2 Hormonas metabólicas 22
1.2.2 Amamantamiento 25
1.2.3 Otros factores 26
1.2.3.1 Estrés calórico 27
1.2.3.2 Involución uterina 28
1.2.3.3 Edad (Número de partos) 28
1.2.3.4 Raza 29
1.2.3.5 Factores sociales 30
1.2.4 Dinámica folicular durante el posparto 30
1.3 CICLOS ESTRALES DE CORTA DURACION 33
1.4 COMPORTAMIENTO DE LOS NIVELES DE PROGESTERONA
DURANTE EL PERIODO POSPARTO 33
1.4.1 Normal alta (tipo 1-a) 34
1.4.2 Normal media (Tipo 1-b) 34
1.4.3 Normal baja (Tipo 1-c) 34
1.4.4 Media (Tipo 2) 34
1.4.5 Cesación de la ciclicidad (Tipo 3) 35
1.4.6 Anovulación prolongada (Tipo 4) 35

8
1.5 NIVELES SUPRABASALES DE PROGESTERONA 36
BIBLIOGRAFIA 37
2. PERFILES HORMONALES Y DINAMICA FOLICULAR DURANTE
EL PERIODO POSPARTO EN VACAS SANMARTINERO 43
2.1 RESUMEN 43
2.2 INTRODUCCION 44
2.3 HIPOTESIS 46
2.4 OBJETIVO 46
2.5 OBJETIVOS ESPECIFICOS 46
2.6 MATERIALES Y METODOS 47
2.6.1 Tipo de estudio 47
2.6.2 Localización 47
2.6.3 Animales 47
2.6.4 Aval del comité de ética para la experimentación animal 48
2.6.5 Muestreos sanguíneos 48
2.6.6 Condición corporal 49
2.7 ANALISIS DE DATOS 49
2.8 RESULTADOS Y DISCUSION 49
2.8.1 Reactivación ovárica posparto 49
2.8.2 Ciclos de corta duración 55
2.8.3 Perfiles de estradiol durante el posparto 57
2.8.4 Reactivación ovárica posparto y condición corporal 61
2.9 CONCLUSIONES 68
BIBLIOGRAFIA 69
3. CICLICIDAD ESTRAL POSPARTO, MOMENTO DEL
INCREMENTO POST-OVULATORIO DE PROGESTERONA
Y LUTEOLISIS EN VACAS SANMARTINERO 73
3.1 RESUMEN 73
3.2 INTRODUCCION 74
3.3 HIPOTESIS 76

9
3.4 OBJETIVO 76
3.5 OBJETIVOS ESPECIFICOS 76
3.6 MATERIALES Y METODOS 77
3.6.1 Tipo de estudio 77
3.6.2 Localización 77
3.6.3 Aval del comité de ética para la experimentación animal 77
3.6.4 Animales 78
3.6.5 Muestreos sanguíneos 78
3.6.6 Determinación de preñez 78
3.7 Análisis de datos 79
3.8 RESULTADOS Y DISCUSION
3.8.1 Restablecimiento de la ciclicidad posparto 79
3.8.2 Agrupamiento de animales en celo 80
3.8.3 Duración del ciclo estral 82
3.8.4 Niveles de progesterona durante el ciclo estral 82
3.8.5 Variación en los niveles de progesterona 86
3.8.6 Preñez temprana 92
3.9 CONCLUSIONES 96
BIBLIOGRAFIA 97
4. CONCLUSIONES GENERALES 101
5. RECOMENDACIONES 102

10
LISTA DE TABLAS
pág.
Tabla 1. Cuadro resumen. Intervalos a la primera ovulación,
segunda ovulación, duración del ciclo breve y nadir
de condición corporal 67
Tabla 2. Niveles de progesterona durante los días del ciclo estral 83

11
LISTA DE FIGURAS
pág.
Figura 1. Perfiles individuales de progesterona durante el posparto 52
Figura 2. Duración de los ciclos breves en el posparto 57
Figura 3. Perfiles de progesterona y estradiol durante el posparto 58
Figura 4. Variación promedio de la condición corporal durante el posparto 63
Figura 5. Perfiles individuales de variacion de la condicion corporal,
en relacion con la reactivacion de la actividad ovarica
medida por perfiles de progesterona 63
Figura 6. Perfiles de progesterona durante el posparto 80
Figura 7. Incremento de la agrupación de celos a través del periodo
posparto 81
Figura 8. Curva promedio de progesterona durante el ciclo estral 83
Figura 9. Perfiles individuales de progesterona para animales que
presentaron ciclos estrales consecutivos completos 85
Figura 10. Variación promedio de los niveles de progesterona entre
muestreos consecutivos. 87
Figura 11. Variación individual de los niveles de progesterona
durante el ciclo estral 88
Figura 12. Porcentaje de variación de los niveles de progesterona
entre muestreos consecutivos durante el ciclo de preñez
temprana y el ciclo estral previo 93

12
RELACIONES HORMONALES Y DINAMICA FOLICULAR DURANTE EL
PERIODO POSPARTO EN VACAS SANMARTINERO
RESUMEN
El objetivo de este estudio fue realizar una descripción básica acerca de los
fenómenos hormonales y foliculares durante el posparto en un grupo de vacas de
la raza Sanmartinero, identificando posibles factores determinantes de la
expresión de buena eficiencia reproductiva en individuos de esta raza.
Se siguieron 18 vacas desde el día 7 posparto, tomando muestras de sangre tres
veces por semana para determinar los niveles de progesterona y estradiol durante
la reanudación de la actividad estral, haciendo seguimiento de los primeros ciclos
estrales y la preñez temprana. Adicionalmente, se evaluó la condición corporal.
Se evidenciaron ciclos de corta duración que preceden al establecimiento de la
ciclicidad normal. Igualmente, se describieron los niveles de progesterona durante
los primeros ciclos estrales y se establecieron algunas relaciones progesterona-
estradiol en el periodo periovulatorio. Los perfiles de estradiol permitieron
confirmar la existencia de ondas de desarrollo folicular desde el inicio del periodo
posparto, y el incremento de los niveles de estradiol hasta la primera ovulación.
En conclusión, este estudio permite contrastar los reportes existentes en ganado
Bos taurus de otras latitudes, con la raza criolla Sanmartinero, resaltando algunos

13
patrones comunes que se presentan en esta raza no obstante las diferencias
ambientales presentes en el medio colombiano, que afectarían a los ganados
introducidos. Esta adaptación puede representar la oportunidad de explorar más a
fondo los mecanismos que permiten tener una mejor eficiencia reproductiva.
Palabras clave: Postparto, progesterona, estradiol, condición corporal

14
ABSTRACT
HORMONE RELATIONSHIP AND FOLLICULAR DYNAMICS DURING
POSTPARTUM PERIOD IN SANMARTINERO BREED COWS
The aim of this study was to describe hormone and follicular phenomena during
postpartum period in Sanmartinero breed cows, to determine decisive factors in
expression of high reproductive efficiency in some individuals of this breed.
18 cows were studied since day 7 postpartum, blood samples were taken three
times a week and progesterone and estradiol levels were measured during the
onset of ovarian activity. First estrous cycles until early pregnancy were
characterized. Body condition was evaluated.
Short estrous cycles were found preceding the onset of normal ciclicity.
Progesterone levels were described during the first cycles as well as some
progesterone-estradiol relationships during oestrus. Estradiol profiles showed
follicular waves early in the lactation, and increasing levels until ovulation.
This study allows to compare current reports regarding Bos taurus cattle in
template regions versus Colombian Sanmartinero breed, standing out some
physiological patterns presented despite the environmental differences that would
affect introduced breeds of cattle. These adaptation phenomena could represent a
quite interesting opportunity to explore in a comprehensive way some mechanisms
involved in high reproductive efficiency.
Key words: post-partum, progesterone, estradiol, body condition

15
INTRODUCCION
Muchos de los hatos bovinos del mundo están localizados en regiones tropicales,
caracterizadas por altos índices pluviométricos y altas temperaturas, que proveen
condiciones favorables para el crecimiento de forraje y la producción de carne a
bajo costo. En los trópicos predominan los animales Bos indicus debido a su
adaptación a las altas temperaturas y a la humedad (Baruselli et al., 2004).
En Colombia, se estima que la ganadería participa con cerca del 3,6% del
producto interno bruto (PIB) nacional. Ya dentro del sector agropecuario, su
importancia relativa es indiscutible, con una participación del 27% del PIB
agropecuario y del 64% del PIB pecuario (Fedegan, 2006). El inventario ganadero
de Colombia es de 24’800.000 bovinos, de los cuales cerca de 16 millones están
representados por hembras. Un 85% de la actividad se lleva a cabo en zonas de
trópico bajo, en donde un 97% de los individuos tiene sangre cebuina en su
composición genética (Fedegan, 2006) De acuerdo con cifras reportadas por la
Federación Nacional de Ganaderos (Fedegan, 2006), 60% del hato se destina a la
producción de carne (cría, levante, ceba), el 38% al doble propósito y el resto (2%)
a la lechería especializada. Dentro del inventario que está destinado a la
producción de leche, se estima que existen alrededor de 4.337.837 hembras de
más de dos años en sistemas doble propósito y 345.431 en sistemas de leche

16
especializada. Según Fedegan (2006), para el año 2019 la meta es tener 48
millones de cabezas en 28 millones de hectáreas para 48 millones de habitantes.
Uno de los principales problemas para los productores de carne bovina es el
intervalo que transcurre desde el momento del parto hasta la aparición de un estro
con ovulación, en especial en el trópico (Galina, 2005). En Colombia el porcentaje
de natalidad es del 54% (Minagricultura, 2005), indicando que el promedio de
intervalo entre partos (IEP) es de 676 días y el intervalo parto-concepción cercano
a los 400 días, cuando los parámetros ideales son 365 días de intervalo posparto y
85 días de intervalo parto concepción (Basurto, 2005).
Esta diferencia representa pérdidas económicas importantes, teniendo en cuenta
lo que se deja de ganar en términos de incremento de peso de las crías, y sin
tener en cuenta las lactancias en los sistemas con ordeño, así como las pérdidas
por costos de operación y mantenimiento de animales no productivos. Teniendo
en cuenta estos factores el costo de un día abierto se estima oscila entre US$ 3-5
en el trópico (Grajales, 2001).
El anestro posparto es un periodo de transición durante el cual se recupera la
funcionalidad del eje hipotálamo-hipófisis-ovarios-útero de la preñez previa. El
anestro en el trópico se manifiesta por largos intervalos parto-primer servicio y
parto-concepción, superiores a los 120 y 170 días, respectivamente (Basurto,
2005).
Numerosos autores han puesto de manifiesto que el estado energético del animal
en el periparto afecta la reanudación de la actividad ovárica (Galina, 2005).

17
Durante las 3 primeras semanas ocurre la involución uterina, el reestablecimiento
de los depósitos de hormona luteinizante (LH) y el inicio del desarrollo de ondas
foliculares. Sin embargo, la percepción del ternero por parte de la vaca durante el
amamantamiento inhibe los pulsos de liberación de GnRH debido a la secreción
de opioides endógenos en el hipotálamo, dando como resultado supresión de la
liberación pulsátil de LH y falla en la ovulación (Yavas y Walton, 2000)
Mucho se ha especulado acerca de los grupos bovinos criollos, descendientes de
los ganados traídos por los españoles hace cerca de 500 años, acerca de los
cuales se indica, aunque tienen relativamente poca capacidad productiva, pueden
ofrecer características de desempeño reproductivo más eficientes que las que
muestran los grupos bovinos foráneos que han ingresado a los trópicos, ello
debido a factores de adaptación que aún no son claramente entendidos o
estudiados (Grajales, 2001). En la raza criolla Costeño con cuernos se
determinaron porcentajes de natalidad del 84% e intervalos entre partos de 436
días (Báez, 2007). En el centro de investigaciones Corpoica La Libertad, en el
Departamento del Meta, piedemonte llanero Colombiano, existe un núcleo de
conservación de la raza Sanmartinero; numerosos estudios se han hecho allí para
caracterizar y valorar tanto en su comportamiento social, como productiva y
reproductivamente, este importante recurso genético (Velásquez et al., 2005;
Góngora y Hernández, 2006)
Se tiene claramente identificada la ineficiencia reproductiva como uno de los
factores determinantes de la disminución en la productividad ganadera, y dentro

18
de esa ineficiencia el anestro posparto participa de manera importante (Crowe,
2008; Peter et al., 2009). Sin embargo, se han identificado algunos individuos, o
grupos de individuos que se caracterizan por presentar un comportamiento
reproductivo superior, este es el caso de las razas criollas bovinas Colombianas, y
de algunos individuos particulares de otras razas. Se han logrado establecer
algunos fenómenos fisiológicos particulares, como los niveles basales de
progesterona durante la fase folicular y su dinámica de cambio durante la fase
luteal (Grajales, 2001; Báez, 2007) que podrían estar vinculados a esta eficiencia
reproductiva, lo que lleva a la necesidad de continuar avanzando en la
investigación básica y tener mayores elementos de juicio para identificar, analizar,
comprender o explicar los mecanismos fisiológicos de adaptación.
El objetivo del presente trabajo es generar información básica acerca de los
fenómenos fisiológicos durante el posparto, utilizando como modelo animal vacas
criollas Sanmartinero, para identificar patrones ocurrentes en este genotipo en las
condiciones particulares del trópico.

19
BIBLIOGRAFIA
Báez GM, Grajales HA, Pérez JE. Caracterización del ciclo estral mediante perfiles de esteroides
(progesterona, 17 β-estradiol) en la raza Costeño con cuernos (Bos taurus) en el trópico
Colombiano. Livestock Research for Rural Development 2007; Volume 19, Article #132.
http://www.cipav.org.co/lrrd/lrrd19/9/baez19132.htm
Baruselli PS, Reis EL, Marques MO, Nasser LF, Bó GA. The use of hormonal treatments to improve
reproductive performance of anestrous beef cattle in tropical climates. Anim. Reprod. Sci. 2004;
82-83: 479-486.
Basurto HB. El anestro posparto en la ganadería bovina del trópico. Memorias Congreso
internacional de reproducción bovina, Intervet, Bogotá 2005.
Crowe MA. Resumption of Ovarian Cyclicity in Post-partum Beef and Dairy Cows Reprod. Dom.
Anim. 2008; 43 (Suppl. 5), 20–28.
Fedegan, Federación Colombiana de Ganaderos, FNG. Plan estratégico de la ganadería
Colombiana 2019. ISBN 978-958-98018-1-9 2006.
Galina CS, Maquivar M. Sistemas de separación de terneros de vacas lactantes y su efecto en la
producción de un celo fértil. Memorias Congreso internacional de reproducción bovina. Intervet,
Bogotá. 2005.
Góngora A y Hernández A. Comportamiento sexual, duración del estro y del ciclo estral en novillas
criollas sanmartineras y brahman del piedemonte llanero colombiano. Livestock Research for
Rural Development 2006. Volume 18, Article #1.
Grajales, H.A. Comportamiento reproductivo de grupos raciales bovinos en el trópico calido-
humedo Colombiano: Pubertad, ciclo estral, preñez temprana, posparto. Niveles de hormonas
esteroides y su relación con la eficiencia reproductiva. 2001. Tesis PhD. Facultad de Medicina
Veterinaria y de Zootecnia. Universidad Nacional de Colombia. Bogota D.C.
Peter AT, Vos PLAM, Ambrose DJ. Postpartum anestrus in dairy cattle Theriogenology 2009; 71:
1333–1342
Velásquez JG, Aristizabal J, Caicedo R, Lesmes N, Mendoza D, Velásquez H y González G
Comportamiento social y productivo del bovino criollo Sanmartinero en la Orinoquía
Colombiana. Corpoica 2005; Plegable divulgativo No 41
Yavas Y, Walton JS. Postpartum aciclicity in suckled beef cows: a review. Theriogenology 2000; 54:
25-55

20
1. REVISION DE LITERATURA
1.1 ANESTRO POSPARTO EN EL TROPICO
La ganadería en el trópico se caracteriza por presentar una baja producción por
animal, así como por fluctuaciones entre temporadas, ya que durante la
temporada de lluvias los animales se encuentran relativamente bien alimentados,
produciendo a su máxima capacidad; pero en la temporada seca la alimentación
es pobre, reduciendo notablemente la producción (Kanuya et al., 2006).
La productividad de la ganadería bovina, en términos de carne y leche, es
dependiente del desempeño reproductivo, y este a su vez se encuentra
íntimamente relacionado con la fertilidad. De esta manera un intervalo entre partos
normal incrementa tanto el número de terneros nacidos como la cantidad de leche
producida por vaca durante la vida útil, y consecuentemente influencia la tasa de
reemplazo (Salisbury et al., 1978)
1.2 FISIOLOGIA DEL POSPARTO
El anestro postparto es el período después del parto durante el cual las vacas no
muestran señales conductuales de estro (Montiel y Ahuja, 2005). La primera
ovulación posparto se presenta cuando existe un folículo dominante y pulsos de

21
LH (Hormona Luteinizante) cada hora (Duffy et al., 2000). Durante el posparto
temprano los pulsos de LH son secretados cada 3 a 6 h (Nett et al., 1988).
Los principales factores que afectan la duración del anestro postparto son el
estado nutricional (condición corporal) y la presencia del ternero (Montiel y Ahuja,
2005). Algunos otros factores como la raza, edad, número de partos, producción
de leche, temporada de parto, presencia del toro, involución uterina, distocias y
estado de salud general influencian la duración del anestro (Peters, 1984; Galina
et al., 1989). No obstante, Wettemann (2003) afirma que cualquier factor
considerado como una causa probable de anestro, además de la nutrición y
presencia del ternero, sólo modula los efectos provocados por estos dos factores
principales.
1.2.1 Nutrición
1.2.1.1 Balance energético. El balance energético negativo reduce la
disponibilidad de glucosa e incrementa la movilización de reservas corporales. El
metabolismo basal, la actividad, el crecimiento y la lactancia tienen prioridad sobre
los procesos reproductivos, como el reinicio de la ciclicidad y el establecimiento y
mantenimiento de una nueva preñez (Short et al., 1990).
Un inadecuado consumo de nutrientes en relación con las demandas metabólicas
es un factor que contribuye a prolongar el anestro posparto y aunque la naturaleza
de estos factores y sus interacciones es compleja y poco conocida, muchos de
ellos parecen actuar por vía de mecanismos hormonales (Jolly et al., 1995). La

22
transición de un balance energético negativo a uno positivo durante la lactancia
está asociada al incremento en la frecuencia de pulsos de LH, lo cual sugiere que
la secreción pulsátil de LH puede ser inhibida hasta tanto no se alcance el valor
más bajo de balance energético, denominado como “nadir” (Canfield y Butler,
1991).
En vacas de leche se ha relacionado el intervalo entre partos y la primera
ovulación con el estatus metabólico (Zurek et al., 1995), encontrando una relación
inversamente proporcional entre balance energético y la reanudación de actividad
ovárica (Butler y Smith, 1989). Canfield y Butler (1991), encontraron que la primera
ovulación ocurrió aproximadamente a los 14 días después del máximo balance
energético negativo. En vacas de la raza costeño con cuernos se reportó el nadir
entre los días 50-60 posparto (Báez et al., 2007). Corro (1993) observó en vacas
Bos indicus en condiciones normales de explotación valores mínimos de condición
corporal entre los 90 y 120 días posparto y una ligera recuperación hasta los 180
días.
En regiones tropicales, vacas que paren con condición corporal aceptable y que
mantienen esta condición durante el posparto, tienen mayores tasas reproductivas
que vacas con baja condición (Galina y Arthur, 1989).
1.2.1.2 Hormonas metabólicas. En los primeros estudios relacionados con la
interacción nutrición-reproducción (Short y Adams, 1988), se establecía que las
concentraciones de glucosa en sangre relacionan el estado nutricional con la
función reproductiva en el hipotálamo. Vacas con concentración de glucosa

23
reducida disminuyen su cantidad de progesterona en plasma (Villa-Godoy et al.,
1988). Aunque las concentraciones de glucosa en bovinos son muy constantes en
comparación con monogástricos, la insulina regula la utilización de glucosa por
parte de las células. La insulina estimula la liberación de GnRH de fragmentos
hipotalámicos in vitro, cuando existe glucosa disponible (Arias et al, 1992).
También estimula la producción de esteroides en las células ováricas (Spicer y
Echterkamp, 1995).
El sistema IGF (Factor Insulínico de Crecimiento), compuesto por IGF-I, IGF-II, sus
receptores y proteínas ligantes, tienen un rol importante en la modulación del nivel
nutricional en el desarrollo folicular (Constant et al., 2000). El IGF-I es producido
en el hígado y tiene efectos reguladores en muchos tipos de células. Las
concentraciones de IGF-I en sangre disminuyen durante la restricción alimenticia
(Richards et al, 1995). Por lo menos seis proteínas de unión de alta afinidad
(IGFBP) pueden influenciar las funciones del IGF-I (Jones y Clemmons, 1995).
Los folículos sintetizan IGF-I y este influye en la función ovárica (Spicer y
Echternkamp, 1995). Específicamente, la proliferación celular ovárica y la
esteroidogenesis son estimuladas por IGF-I (Spicer y Chamberlain, 1998). Los
folículos dominantes presentan menor actividad IGFBP que los subordinados
(Yuan et al, 1998), esta disminución puede incrementar la disponibilidad de IGF-I
para las células foliculares. Adicionalmente, las concentraciones de IGFBP-4
parecen determinar cual folículo se vuelve dominante durante la selección (Mihm
et al, 2000).

24
Las concentraciones plasmáticas de estradiol están altamente correlacionadas con
los niveles de IGF-I (Butler, 2003). El IGFBP-3 parece ser importante en el
transporte de IGF-I hacia tejidos blanco y se ha encontrado en mayor cantidad en
plasma de vacas con un folículo ovulatorio durante las tres primeras semanas
posparto (Frajblat, 2000)
La importancia de la insulina y el IGF-I en la regulación del desarrollo folicular
posparto demuestra que la capacidad funcional del eje somatotrópico está
entrelazada con la actividad ovárica durante el balance energético negativo (Webb
et al, 1999)
Desde el descubrimiento de la leptina, ha existido mucho interés acerca de su
función como señal de información acerca de las reservas energéticas (Spicer,
2001; Smith et al, 2002). La restricción nutricional incrementa la expresión de
receptores de leptina en el núcleo hipotalámico (Dyer et al, 1997). El consumo de
nutrientes influye sobre las cantidades de mRNA para leptina en grasa (Amstalden
et al, 2000) y sobre las concentraciones de leptina en plasma (Ehrhardt et al,
2000). Las concentraciones de leptina en plasma disminuyen durante el balance
energético negativo.
Dentro del sistema nervioso central, el hipotálamo es el sitio de mayor acción de la
leptina con respecto al control de consumo de alimento y gasto de energía.
Numerosos estudios han evaluado la localización del receptor de RNA mensajero
(mRNA) para leptina dentro del hipotálamo (Dyer et al, 1997). El hipotálamo
convierte las señales de la leptina en respuestas neuronales sobre el consumo de
alimento. El Neuropéptido Y (NPY) parece ser importante en la regulación del

25
consumo. La leptina inhibe las señales del NPY inhibiendo el consumo de alimento
(Jang et al, 2000).
La administración de leptina también estimula la producción de gonadotropinas en
la hipófisis por medio de estimulación de GnRH en el hipotálamo (Zieba et al,
2003). En rumiantes, la administración de leptina recombinante ovina en vacas de
carne adultas sometidas a ayuno estimuló la secreción de LH (Amstalden et al,
2002). En vacas de leche ovariectomizadas afectó la secreción de LH en relación
directa con la dosis aplicada (Zieba et al, 2003).
En un estudio realizado por Liefers et al. (2003), no se encontró relación entre los
niveles de leptina y la primera actividad luteal posparto, aunque la leptina tiende a
incrementarse durante la primera ovulación, indicando la existencia de
mecanismos reguladores adicionales en el hipotálamo, hipófisis y ovarios.
Durante la preñez los niveles de leptina son altos y disminuyen rápidamente luego
del parto. Esta caída es debida a los costos energéticos de la producción de leche
(Block et al, 2001). El estimulo de la lactancia por sí mismo no parece influenciar la
disminución de las concentraciones de leptina (Brogan et al, 1999).
1.2.2 Amamantamiento. El amamantamiento afecta la actividad del hipotálamo,
hipófisis y ovarios, mediante la reducción de liberación de GnRH, la cual conduce
a insuficientes pulsos de LH (Williams et al., 1996). Debido a esto los folículos son
incapaces de madurar y por lo tanto de ovular, ya que existe una incorrecta
síntesis de estrógenos a nivel folicular. Adicionalmente el amamantamiento genera
la secreción a nivel hipotalámico de -endorfina, en respuesta al estimulo de

26
succión, y los estrógenos producidos en la placenta durante el último tercio de la
gestación provocan la inhabilitación de la secreción de LH a través de la inhibición
en el hipotálamo (Day, 2004).
En el parto, las concentraciones de LH en la hipófisis anterior se agotan debido al
efecto inhibitorio de los estrógenos placentarios. El efecto inhibitorio del estradiol
acaba con el parto, y al cabo de 2-3 semanas las reservas de LH son similares a
las de vacas cíclicas (Nett et al. 1988). El amamantamiento del ternero prolonga la
demora en la aparición de la secreción pulsátil de LH, en vacas que pierden su
ternero al nacimiento comúnmente muestran estro más temprano (Wettemann,
1994)
La inhibición de la secreción de LH debida al amamantamiento puede estar
influenciada por la secreción del opioide hipotalámico β-endorfina, en respuesta al
estímulo de succión (Boland, 1990). La administración de antagonistas de opioides
incrementa la secreción de LH en vacas lactantes (Williams y Griffith, 1995).
El efecto del amamantamiento es de mayor magnitud en vacas primíparas y en
vacas en baja condición (Short et al., 1990), las vacas desarrollan un vínculo con
su cría, y el efecto del ternero propio es mayor que el de un ternero ajeno
(Stevenson et al., 1997)
Estudios iniciales con mastectomía (Short et al, 1972) o destete (Williams et al.,
1987), sugieren que el amamantamiento inhibe la reanudación de los pulsos de LH
a través de la estimulación táctil, mientras que Griffith y Williams sugieren
estímulos visuales y olfativos (1996).

27
En ganado Costeño con cuernos (Báez, 2007) se observó, en cuanto al
comportamiento de amamantamiento, que las vacas amamantaban a sus crías en
determinadas horas del día, mientras que la mayor parte del tiempo permanecían
solas alimentándose y las crías permanecían en grupo, con presencia de una
hembra nodriza, similar a lo encontrado por Velásquez et al (2005) en ganado
criollo Sanmartinero. En un estudio hecho por Sanz et al (2003) en España, el
anestro posparto fue marcadamente afectado por la interacción entre la frecuencia
de amamantamiento y la raza.
1.2.3 Otros factores
1.2.3.1 Estrés calórico. Con respecto al patrón de secreción de LH en vacas bajo
estrés de calor se han reportado disminuciones en la amplitud (Gilad et al., 1993) y
frecuencia (Wise et al., 1988) de los pulsos. La disminución de los niveles de LH
durante el pico preovulatorio ha sido controversial y se ha relacionado con los
niveles preovulatorios de estradiol.
La poca información que existe disponible acerca de el efecto del estrés calórico
sobre las concentraciones sanguíneas de FSH e inhibina sugieren que la FSH se
incrementa y que esto puede ser debido a la disminución en la producción de
inhibina (De Rensis y Scaramuzzi, 2003).
Wilson et al. (1998) encontraron que el estrés calórico no tuvo efectos sobre la
producción de progesterona pero sí retrasó la luteolisis. Otros reportes han sido
controversiales (Wilson et al., 1998); estas diferencias probablemente se deban a

28
otros factores no controlados como el tipo de estrés calórico (agudo o crónico) y
las diferencias en el consumo de materia seca (De Rensis y Scaramuzzi, 2003).
Los mecanismos mediante los cuales el estrés calórico afecta las concentraciones
de hormonas reproductivas son desconocidos. Se ha sugerido como causa el
incremento en la secreción de corticosteroides porque estos pueden inhibir la
secreción de GnRH y por lo tanto de LH (Gilad et al., 1993). En este mismo
estudio se encontró que la secreción de gonadotropinas fue inhibida en mayor
grado en vacas con bajas concentraciones plasmáticas de estradiol, sugiriendo
que las altas concentraciones de estradiol pueden neutralizar el efecto del estrés
calórico.
1.2.3.2 Involución Uterina. La involución uterina no está relacionada con la
duración del anestro posparto (Kiracofe, 1980), sin embargo, la involución es una
barrera física en el posparto temprano tanto para el transporte del esperma como
para la implantación (Short, 1990), muy pocas vacas pueden presentar estro tan
temprano como para que la involución uterina interfiera con la concepción, a
menos que condiciones patológicas prolonguen la involución normal.
La involución uterina se completa dentro de las 4 a 6 semanas posparto y ocurre
más rápidamente en el cuerno no grávido de la preñez anterior (Spicer et al.,
1986)
1.2.3.3 Edad (Número de partos). Debido a que las vacas de primer parto
continúan en crecimiento, La frecuencia de pulsos de LH en el posparto es menor,

29
y el anestro posparto se prolonga de 1 a 4 semanas más que en vacas multíparas
(Yavas y Walton, 2001).
1.2.3.4 Raza. En vacas de lechería especializada, los folículos medianos (5-10
mm) son detectables a partir del día 5 posparto. El primer folículo dominante que
se desarrolla logra la ovulación entre los días 15 a 27 posparto en la mayoría de
vacas de leche (Beam et al., 1997).
En vacas de carne lactantes, folículos medianos se encuentran desde los días 5-7
posparto, y su número y tamaño se incrementa a medida que transcurre el
posparto. Los folículos dominantes son detectables en los días 10-21 posparto, sin
embargo, la mayoría de estos folículos no llegan a ovular, debido a que sufren
atresia antes de obtener el tamaño optimo de ovulación, posiblemente debido a la
sensibilidad del eje hipotálamo-hipofisiario al efecto de retroalimentación negativa
del 17 β estradiol ovárico que retrasa el restablecimiento de los pulsos de LH
(Yavas y Walton, 2001).
En vacas Bos indicus en el trópico, bajo sistema de manejo extensivo con
amamantamiento, la actividad ovárica se ha caracterizado por el crecimiento y
regresión de folículos de 4-6 mm con dominancia esporádica a partir de los 26
días posparto, restableciendo la ciclicidad a los 217-278 días (Ruíz-Cortes y
Olivera-Angel, 1999).
En ganado criollo Costeño con cuernos, manejado en pastoreo continuo con
amamantamiento en el Valle del Sinú, se ha observado estro con ovulación y
reanudación de la ciclicidad ovárica a partir del día 51 posparto (Báez, 2007).

30
1.2.3.5 Factores sociales. La presencia del macho (Short, 1990), así como las
interacciones entre hembras, la presencia de otras hembras en celo y el
encadenamiento de conductas sexuales en grupos de hembras (Galina, 2005),
afectan la duración del intervalo posparto. Góngora y Hernández (2006) reportan
diferencias en la actividad sexual de novillas Sanmartinero y Cebú, debidas a la
jerarquía de los machos con los que se encuentran, así como la conformación de
grupos de actividad sexual.
1.2.4 Dinámica folicular durante el posparto
Aunque la función de las gonadotropinas y otros reguladores endocrinos de la
actividad folicular durante el postparto temprano no están claros, la ultrasonografía
hace posible comparar los cambios en el desarrollo folicular ovárico con variables
endocrinas y metabólicas para encontrar el origen endocrino del anestro
prolongado (Stagg et al., 1995).
El crecimiento folicular, y la habilidad de los folículos para producir cantidades
considerables de estradiol dependen de un adecuado aporte de FSH (Hormona
Folículo Estimulante) y LH (Driancourt, 1991). La FSH controla el crecimiento
folicular, mientras que la LH está asociada con el mantenimiento de los folículos
dominantes y la producción de estradiol necesaria para la inducción del estro, el
pico preovulatorio de LH y la ovulación (Fortune, 1994). La síntesis de estradiol
por las células de la granulosa dependen del suministro de substrato aromatizable

31
(predominantemente androstenediona) de las células de la teca interna
adyacentes gobernadas por LH (McNatty et al., 1984).
La reanudación del crecimiento folicular luego del parto se debe al incremento de
la secreción de FSH a los 3-4 días posparto en vacas bien alimentadas (Lamming
et al., 1981), pero la habilidad de los folículos para desarrollarse y ser dominantes
depende de la secreción pulsátil de LH, que a su vez refleja la secreción de GnRH
hipotalámico (Jolly et al., 1995). La falla de los mecanismos que controlan la
maduración folicular incluye la inhibición de la liberación pulsátil de mayor
frecuencia de LH necesaria para estimular la mayor secreción de estradiol, o la
inhibición central de liberación de GnRH en respuesta al estradiol (Jolly et al.,
1995).
Los patrones de crecimiento y desarrollo folicular son inconsistentes luego del
parto. Algunas vacas presentan ovarios relativamente inactivos (sin crecimiento
folicular mayor a 10 mm), mientras otras muestran folículos de diámetro mayor a
10 mm dentro de los 10 a 15 días después del parto (Lucy et al., 1992). La
inactividad ovárica puede deberse a insuficiente secreción de LH asociada con un
inadecuado consumo de energía. Este tipo de anestro es común en vacas
primíparas en las cuales el balance energético negativo es más evidente ( Lucy et
al., 1992)
El desarrollo de ondas foliculares ocurre temprano en el posparto tanto en ganado
de carne como de leche. Posteriormente al parto existe un incremento dramático
en la FSH que es que es seguido por la emergencia de la primera onda folicular
(2-7 días posparto) (Wiltbank et al., 2002). Sin embargo, la ovulación de el folículo

32
dominante solamente ocurre cuando este es expuesto a una adecuada frecuencia
de pulsos de LH (aproximadamente 1 pulso/h), que permita el incremento en el
tamaño del folículo y la producción de estradiol, la cual eventualmente puede
conllevar a un pico de LH y una ovulación (Roche et al., 1992). Se ha determinado
que el folículo dominante de la primera onda folicular ovula en el 42% de vacas
holstein de alta producción (Beam y Butler, 1997) y solamente en el 11% de vacas
de carne lactantes (Murphy et al, 1990).
Los patrones de secreción de gonadotropinas en el periodo posparto también
difieren entre B. taurus y B. indicus. Bajo las mismas condiciones, vacas B. taurus
presentaron mayores concentraciones plasmáticas de LH que vacas brahman,
(D’occhio et al., 1990). Adicionalmente, una alta proporción de vacas B. taurus
tienen una secreción pulsátil de LH mayor que vacas B. indicus (D’occhio et al.,
1990).
Periodos posparto prolongados (mas de 150 días) son característicos de ganado
Bos indicus y cruces B. taurus/B. indicus en regiones tropicales, originando
pérdidas económicas debido al fracaso en alcanzar el intervalo entre partos
deseable de 12 meses (Vaccaro, 1990). Ruiz-Cortes y Olivera-Angel (1999)
reportan reanudación de la ciclicidad estral de los 217 a los 278 días posparto en
vacas B. indicus lactantes mantenidas en praderas naturales en Colombia,
resultando en un intervalo entre partos de 17-19 meses; en este mismo estudio se
observó que durante los primeros seis meses después del parto muchas de las
vacas tuvieron periodos en los cuales los folículos crecían hasta menos de 6 mm
de diámetro. En ganado criollo costeño con cuernos se reporta estro con ovulación

33
a partir de los 51 días posparto (Báez, 2007), resultados similares a los
encontrados por Grajales (2001) en ganado Romosinuano y por Sinclair et al.
(2002) en ganado de carne lactante en zona templada.
1.3 CICLOS ESTRALES DE CORTA DURACION
Los ciclos de corta duración ocurren normalmente en la primera ovulación
espontánea posparto en la mayoría de vacas (Day et al 1990); La liberación
prematura de PGF2α por el útero (Cooper et al 1991), como resultado de los
efectos de la baja progesterona (Zollers et al 1993) y estradiol (Mann y Lamming
2000) antes de la ovulación en los receptores de oxitocina y progesterona
endometriales parecen ser la causa de la temprana regresión del cuerpo lúteo. La
progesterona producida durante la fase luteal corta puede ser necesaria para el
establecimiento de una función luteal normal en el siguiente ciclo estral (Short et al
1990; Mihm 1999).
1.4 COMPORTAMIENTO DE LOS NIVELES DE PROGESTERONA DURANTE
EL PERIODO POSPARTO
Opsomer et al. (1998) clasificaron la actividad ovárica posparto de acuerdo al
modelo de secreción de progesterona así:

34
1.4.1 Normal alta (Tipo 1-a). El primer incremento de progesterona se presenta
antes del día 50 posparto, dándose una actividad luteal normal, la cual es
producida luego de un calor manifiesto y la respectiva ovulación. En más del 90%
de los casos el servicio que recibe la hembra en el primer calor manifiesto es
efectivo, se produce la concepción y se tiene la capacidad de mantener la preñez.
1.4.2 Normal media (Tipo 1-b). El primer incremento de progesterona se
presenta antes del día 50 posparto, pudiendo obedecer a un incremento transitorio
de progesterona (ITP4) o a un “calor silencioso”, aparentemente sin ovulación, no
conducente a una actividad luteal normal. Las hembras reciben uno o dos
servicios antes de lograr una concepción efectiva (entre el día 50 a 100 posparto)
con capacidad para el mantenimiento de la preñez.
1.4.3 Normal baja (Tipo 1-c). El primer incremento de progesterona se presenta
hacia el día 50 posparto, pudiendo obedecer a un ITP4 o a un “calor silencioso”,
aparentemente sin ovulación, no conducente a una actividad luteal normal. Las
hembras reciben dos, tres o más servicios antes de lograr una concepción efectiva
(después del día 100 posparto) con capacidad para el mantenimiento de la preñez.
1.4.4 Media (Tipo 2). El primer incremento de progesterona se presenta hacia el
día 100 posparto, pudiendo obedecer a un ITP4 o a un “calor silencioso”,

35
aparentemente sin ovulación, no conducente a una actividad luteal normal. Las
hembras reciben dos, tres o más servicios antes de lograr una concepción efectiva
(después del día 150 posparto) con capacidad para el mantenimiento de la preñez.
1.4.5 Cesación de la ciclicidad (Tipo 3). Los primeros incrementos de
progesterona se presentan generalmente hacia el día 50 posparto, pudiendo
obedecer a un ITP4, a un “calor silencioso”, aparentemente sin ovulación, no
conducente a una actividad luteal normal o a una actividad luteal normal, la cual es
producida luego de un calor manifiesto y la respectiva ovulación. Hay interrupción
de la actividad ovárica por un periodo indeterminado. Las hembras al reiniciar la
ciclicidad pueden recibir dos o más servicios antes de lograr una concepción
efectiva (después del día 100 posparto) con capacidad para el mantenimiento de
la preñez.
1.4.6 Anovulación prolongada (Tipo 4). Puede presentar esporádicamente o no
presentar ITP4 o un “calor silencioso”, aparentemente sin ovulación, no conducente
a una actividad luteal normal, la cual es seguida por una interrupción de la
actividad ovárica hasta después del día 150 posparto, cuando se presenta el
reinicio de la ciclicidad. Las hembras pueden recibir dos o más servicios antes de
lograr una concepción efectiva (después del día 150 posparto) con capacidad para
el mantenimiento de la preñez.

36
1.5 NIVELES SUPRABASALES DE PROGESTERONA
En estudios controlados se ha observado como las hembras que se denominan
repetidoras de servicio o de calor presentan una asincronía hormonal durante el
estro caracterizada por niveles de progesterona que significativamente exceden
los niveles normales, a los que se ha llamado “niveles suprabasales de
progesterona” (Gustafsson, 1986; Duchens et al., 1995; Bage, 1997). Ello ha
estado relacionado con un retardo en el pico de LH y la presencia de un folículo de
mayor tamaño y de vida media más larga, lo que pudiera estar alterando la estricta
secuencia de cambios en el crecimiento y desarrollo del folículo preovulatorio y
causando efectos detrimentales sobre el oocito que resultan en una disminución
de la fertilidad.
De otra parte, ha sido demostrado que un incremento de los niveles de
progesterona en la fase luteal temprana, de alguna manera controlan o determinan
el tiempo en que se produce la luteólisis, alterando la duración del ciclo estral
(Mann et al., 1998). Por el contrario, cuando se presenta un retardo en el
incremento de los niveles de progesterona puede darse como resultado una más
tardía luteólisis y un ciclo estral de mayor duración.

37
BIBLIOGRAFIA
Amstalden, M, Garcia MR, Williams SW, Stanko RL, Nizielski SE, Morrison SD et al. Leptin gene
expression, circulating leptin, and luteinizing hormone pulsatility are acutely responsive to short-
term fasting in prepubertal heifers: Relationships to circulating insulin and insulin-like growth factor
I. Biol. Reprod 2000.63:127–133.
Arias P, Rodriguez M, Szwarcfarb B, Sinay IR, Moguilevsky JA. Effect of insulin on LHRH release by
perifused hypothalamic fragments. Neuroendocrinology 1992.56:415–418.
Báez GM, Grajales HA, Pérez JE. Caracterización del ciclo estral mediante perfiles de esteroides
(progesterona, 17 β-estradiol) en la raza Costeño con cuernos (Bos taurus) en el trópico
Colombiano. Livestock Research for Rural Development 2007.. Volume 19, Article #132.
http://www.cipav.org.co/lrrd/lrrd19/9/baez19132.htm
Bage. R. Gustafsson. H, Forsberg. B, Larsson. B y Rodríguez-Martínez. H. Suprabasal progesterone
levels in repeat breeder heifers during the pro and oestrous period. Theriogenology. 1997.47: 141 –
147
Beam SW, Butler WR. Energy balance and ovarian follicle development prior to the first ovulation
postpartum in dairy cows receiving three levels of dietary fat. Biol Reprod 1997;56: 133-142.
Block SS, Butler WR, Ehrhardt RA, Bell AW, Van Amburgh ME, Boisclair YR. Decreased concentration
of plasma leptin in periparturient dairy cows is caused by negative energy balance. J Endocrinol
2001;171(2):339–48.
Brogan RS, Mitchell SE, Trayhurn P, Smith MS. Suppression of leptin during lactation: contribution of
the suckling stimulus versus milk production. Endocrinology 1999;140(6):2621–7.
Butler WR. Energy balance relationships with follicular development, ovulation and fertility in
postpartum dairy cows. Livestock Production Science 2003..83 211–218
Butler WR, Smith RD, Interrelationships between energy balance and postpartum reproductive function
in dairy cows. J. Dairy Sci. 1989. 72, 767–783.
Canfield RW, Butler WR, Energy balance, first ovulation and the effects of naloxone on LH secretion in
early postpartum dairy cows. J. Anim. Sci. 1991. 69, 740–746.
Constant F, Ponter AA, Ponsart C, Fontaine JJ, Mialot JP, Grimard B, Chastant S, Effect of
underfeeding on plasmatic and intrafollicular IGFBPs in postpartum anoestrus beef cows. J.
Reprod. Fertil.Abstr. Ser 2000.26, 15 (abstract).
Corro, MMD. Efecto de la suplementario mineral preparto sobre el comportamiento reproductivo y
productivo posparto en vacas Holstein X Cebú en trópico húmedo 1992. Tesis de maestría. FMVZ,
UNAM, México., D.F.

38
Day ML, Dyer RM, Wilson GW, Pope WF, Influence of estradiol on duration of anestrus and incidence
of short estrous cycles in postpartum cows. Domest. Anim. Endocrinol 1990: 7, 19–25.
Day ML. Hormonal induction of estrous cycles in anestrous bos taurus beef cows. Animal
Reproduction Science 2004 82-83: 487-494
De Rensis F, Scaramuzzi RJ Heat stress and seasonal effects on reproduction in the dairy cow—a
review Theriogenology 2003 60 1139–1151
D’Occhio MJ, Neish A, Broadhurst L, Differences in gonadotrophin secretion postpartum between Zebu
and European breed cattle. Anim. Reprod. Sci. 1990. 22, 311–317.
Driancourt, MA, Follicular dynamics in sheep and cattle. Theriogenology 1991.35, 55–79.
Duchens. M, Forsberg. M, Gustafsson. H, Edqvist. LE, y Rodríguez-Martínez. H. Reproductive
performance of heifers induced to oestrous asynchrony by suprabasal plasma progesterone levels.
Anim. Reprod. Sci. 1995 39: 171 – 182:
Duffy P, Crowe MA, Boland MP, Roche JF. Effect of exogenous LH pulses on the fate of the first
dominant follicle in postpartum beef cows nursing calves. J. Reprod. Fertil. 2000. 118:9–17.
Dyer CJ, Simmons JM, Matteri RL, Keisler DH. Leptin receptor mRNA is expressed in ewe anterior
pituitary and adipose tissues and is differentially expressed in hypothalamic regions of well-fed and
feed-restricted ewes. Domest. Anim. Endocrinol. 1997. 14:119–128.
Ehrhardt, RA, Slepetis RM, Siegal-Willott J, Van Amburgh JE, Bell AW, Boisclair YR. Development of
a specific radioimmunoassay to measure physiological changes of circulating leptin in cattle and
sheep. J. Endocrinol. 2000. 166:519–528.
Fortune JE, Ovarian follicular growth and development in mammals. Biol. Reprod. 1994. 50, 225–232.
Frajblat M. Metabolic state and follicular development the postpartum lactating dairy cow. PhD
dissertation, Cornell University, 250 p. 2000.
Galina CS, Arthur GH, Review of cattle reproduction in the tropics. 3. Puerperium. Anim. Breed. Abstr.
1989. 57, 889–910.
Galina, CS, Maquivar M. Sistemas de separación de terneros de vacas lactantes y su efecto en la
producción de un celo fértil. Congreso internacional de reproduccion bovina. 2005. Intervet. Bogota.
Gilad E, Meidan R, Berman A, Graber Y, Wolfenson D. Effect of heat stress on tonic and GnRH-
induced gonadotrophin secretion in relation to concentration of oestradiol in plasma of cyclic cows.
J Reprod Fertil 1993; 99:315–21.
Grajales HA. Comportamiento reproductivo de grupos raciales bovinos en el trópico calido-humedo
Colombiano: Pubertad, ciclo estral, preñez temprana, posparto. Niveles de hormonas esteroides y
su relación con la eficiencia reproductiva. 2001. Tesis PhD. Facultad de Medicina Veterinaria y de
Zootecnia. Universidad Nacional de Colombia. Bogotá D.C.

39
Griffith MK, Williams GL. Roles of maternal vision and olfaction in suckling-mediated inhibition of
luteinizing hormone secretion, expression of maternal selectivity, and lactational performance of
beef cows. Biol Reprod 1996;54:761-768.
Gustafsson. H, Larsson. K, Kindahl. H y Madej. A. Sequential endocrine changes and behaviour
during oestrus and metoestrus in repeat breeder and virgin heifers. Anim. Reprod. Sci. 1986: 10:
261-273.
Jang M, Mistry A, Swick AG, Romsos DR. Leptin rapidly inhibits hypothalamic neuropeptide Y
secretion and stimulates corticotropin-releasing hormone secretion in adrenalectomized mice. J
Nutr 2000; 130(11):2813–20.
Jolly PD, McDougall S, Fitzpatrick LA, Macmillan KL, Entwistle KW, Physiological effects of
undernutrition on postpartum anoestrus in cows. J. Reprod. Fert. 1995. 49 (Suppl.), 477–492.
Jones JI, Clemmons DR. Insulin-like growth factors and their binding proteins: Biological actions.
Endocr. Rev. 1995. 16:3–34.
Kanuya NL, Matiko MK, Nkya R, Bittegeko SBP, Mgasa MN, Reksen O et al. Seasonal changes in
nutritional status and reproductive performance of Zebu cows kept under a traditional agro-pastoral
system in Tanzania Trop Anim Health Prod 2006. 38:511–519
Kiracofe GH. Uterine involution: Its role in regulating postpartum intervals. J. Anim. Sci. 1980. Sl
(Supp1. II):l6
Lamming, GE, Wathes DC, Peters AR, Endocrine patterns in the post-partum cow. J. Reprod. Fert.
(Suppl.) 1981.155–170.
Liefers SC. Veerkamp RF, Te Pas MFW, Chilliard Y, Van der Lende T. Genetics and physiology of
leptin in periparturient dairy cows. Domest Anim Endocrinol. 2005 Jul;29(1):227-38.
Lucy MC, Savio JD, Badinga L, De La Sota RL, Thatcher WW. Factors That Affect Ovarian Follicular
Dynamics in Cattle J. Anim. Sci. 1992. 70:3615-3626
Mann. GE, Lamming. GE y Payne. JH. Role of early luteal phase progesterone in control of the timing
of the luteolytic signal in cows. J. Reprod. Fertil. 1998. 113: 47 – 51
Mann GE, Lamming GE. The role of sub-optimal preovulatory oestradiol secretion in the aetiology of
premature luteolysis during the short oestrous cycle in the cow.Anim Reprod Sci. 2000 Dec
29;64(3-4):171-80.
McNatty, KP, Heath DA, Henderson KM, Lun S, Hurst PR, Ellis LM, Montgomery GW, Morrison L,
Some aspects of thecal and granulosa cell function during follicular development in the bovine
ovary. J. Reprod. Fertil. 1984. 72, 39–53.
Mihm. M. Delayed resumption of cyclicity in postpartum dairy and beef cows Reprod. Dom. Anim.
1999: 34: 276-284

40
Mihm M, Austin EJ, Good TEM, Ireland JLH, Knight PG, Roche JF, Ireland JJ, Identification of potential
intrafollicular markers involved in selection of the dominant ollicle in heifers. Biol Reprod 2000 63,
811–819.
Montiel F, Ahuja C. Body condition and suckling as factors influencing the duration of postpartum
anestrous in cattle: a review. Animal Reproduction Science 2005. 85, 1-26.
Murphy MG, MP Boland, and JF Roche. Pattern of follicular growth and resumption of ovarian activity
in post-partum beef suckler cows. J. Reprod. Fertil 1990.. 90:523.
Nett TM, Cermak D, Braden T, Manns J, Niswender G, Pituitary receptors for GnRH and estradiol and
pituitary content of gonadotropins in beef cows. II. Changes during the postpartum period. Dom.
Anim. Endocrinol. 1988. 5, 81–89.
Opsomer. G, Coryn. M, Deluyker. H y Kruif. A. An analysis of ovarian dysfunction in high yielding dairy
cows after calving, based on progesterone profiles. Reprod. Dom. Anim. 1998. 33: 193 – 204:
Peters AR, Reproductive activity of the cow in the post-partum period. I. Factors affecting the length of
the post-partum acyclic period. Br. Vet. J. 1984. 140, 76–84.
Richards MW, Wettemann RP, Spicer LJ, Morgan GL. Nutritional anestrus in beef cows: Effects of
body condition and ovariectomy on serum luteinizing hormone and insulin-like growth factor-I. Biol.
Reprod. 1991. 44:961–966.
Roche JF, Crowe MA, Boland MP. Postpartum anestrus in dairy and beef cows. Anim. Reprod. Sci.
1992. 28, 371–378.
Ruiz-Cortez ZT, Olivera-Angel M, Ovarian follicular dynamics in suckled zebu (Bos indicus) cows
monitored by real time ultrasonography. Anim. Reprod. Sci. 1999. 54, 211–220.
Salisbury GW, Van Dermark NL, Lodge JR, Physiology of Reproduction and Artificial Insemination of
Cattle, (W.H. Freeman, San Fransisco) 1978.
Short RE, Bellows RA, Moody EL, Howland BE. Effects of suckling and mastectomy on bovine
postpartum reproduction. J Anim Sci 1972;34:70-74.
Short RE, Bellow RA, Staigmiller RB, Berardinelli JG, Custer EE. Physiological mechanisms controlling
anestrous and fertility in postpartum beef cattle. J. Anim. Sci. 1990: 68: 799-816
Short RE, Adams DC, Nutritional and hormonal interrelationships in beef cattle reproduction. Can. J.
Anim. Sci. 1988 68, 29.
Sinclair KD, Molle G, Revilla R, Roche JF, Quintans G, Marongiu L, Sanz A, Mackey DR, Diskin MG.
Ovulation of the first dominant follicle arising after day 21 postpartum in suckling beef cows. Animal
Science 2002 75: 115-126.
Smith GD, Jackson LM, Foster DL. Leptin regulation of reproductive function and fertility.
Theriogenology 2002. 57:73–86.

41
Spicer IJ, Leung K, Convey EM, Gunther J, Short RE, Tucker HA. Anovulation in postpartum suckled
beef cows. I. Associations among size and numbers of ovarian follicles, uterine involution, and
hormones in serum and follicular fluid. J Anim Sci 1986;62.734-741.
Spicer LJ. Leptin: A possible metabolic signal affecting reproduction. Domest. Anim. Endocrinol. 2001.
21:251–270.
Spicer LJ, Chamberlain CS. Influence of cortisol and insulin- and insulin-like growth factor I (IGF-I)-
induced steroid production and on IGF-I receptors in cultured bovine granulosa cells and thecal
cells. Endocrine 1998. 9:153–161.
Spicer LJ, Echternkamp SE. The ovarian insulin and insulin-like growth factor system with an emphasis
on domestic animals. Domest. Anim. Endocrinol. 1995. 12:223–245.
Stagg K, Diskin MG, Sreenan JM, Roche JF. Follicular development in long-term anoestrus suckler
beef cows fed two levels of energy postpartum. Anim. Reprod. Sci. 1995. 38, 49–61.
Stevenson JS, Lamb GC, Hoffmann DP, Minton JE. Interrelationships of lactating and postpartum
anovulation in suckled and milked cows. Livestock Prod. Sci. 1997. 50:57–74.
Vaccaro R. Comportamiento de bovinos para doble propósito en el trópico, Seminario Internacional
sobre lechería Tropical. Villahernosa, Tabasco, México, pp. 14–35. 1990.
Velásquez JG, Aristizabal J, Caicedo R, Lesmes N, Mendoza D, González G. Comportamiento social
y productivo del bovino criollo Sanmartinero en la Orinoquía Colombiana. Plegable divulgativo No
41. Corpoica. 2005
Villa-Godoy A, Hughes, TL, Emery RS, Chapin LT, Fogwell RL. Association between energy balance
and luteal function in lactating dairy cows. J. Dairy Sci. 1988. 71, 1063–1072.
Webb R, Garnsworthy PC, Gong JG, Robinson RS, Wathes DC. Consequences for reproductive
function of metabolic adaption to load. Anim. Sci. 1999. Occas. Publ. 24, 99–112.
Wettemann RP, Lents CA, Ciccioli NH, White, FJ, Rubio I. Nutritional- and suckling-mediated
anovulation in beef cows J. Anim. Sci. 2003. 81(E. Suppl. 2):E48–E59
Wettemann RP, Precalving nutrition/birth weight interaction and rebreeding efficiency. Oklahoma State
University (Ed.), Anim. Sci. Res. Report. 1994.
Williams GL, Koziorowski M, Osbom RG, Kirsch JD, Slanger WD. The postweaning rise of tonic
luteinizing hormone secretion in anestrous cows is not prevented by chronic milking or the physical
presence of the calf. Biol Reprod 1987 36:1079-1084.
Williams GL, Gazal, OS, Guzman Vega GA, Stanko RL. Mechanisms regulatin suckling-mediated
anovulation in the cow. Animal Reproduction Science 1996 42: 289-297.
Wilson SJ, Marion RS, Spain JN, Spiers DE, Keisler DH, Lucy MC. Effect of controlled heat stress on
ovarian function in dairy cattle: I. Lactating cows. J Dairy Sci 1998;1:2124–31.
Wiltbank MC, Gumen A, Sartori R, Physiological classification of anovulatory conditions in cattle.
Theriogenology 2002. 57, 21–52.

42
Wise ME, Armstrong DV, Huber JT, Hunter R,Wiersma F. Hormonal alterations in the lactating dairy
cow in response to thermal stress. J Dairy Sci 1988;71:2480–5.
Yavas Y, Walton JS. Postpartum aciclicity in suckled beef cows: a review. Theriogenology 2000. 54:
25-55
Yuan W, Bao B, Garverick HA, Youngquist RS, Lucy MC. Follicular dominance in cattle is associated
with divergent patterns of ovarian gene expression for insulin-like growth factor (IGF)-I, IGF-II, and
IGF binding protein-2 in dominant and subordinate follicles. Domest. Anim. Endocrinol. 1998.
15:55–63.
Zieba DA, Amstalden M, Morton S, Gallino JL, Edwards JF, Harms PG, et al. Effects of leptin on basal
and GHRH-stimulated GH secretion from the bovine adenohypophysis are dependent upon
nutritional status. J Endocrinol 2003; 178(1):83–9.
Zollers WG, Garverick HA, Smith MF, Moffatt RJ, Salfen BE, Youngquist RS, Concentrations of
progesterone and oxytocin receptors in endometrium of post partum cows expected to have a short
or normal oestrous cycle. J. Reprod. Fertil. 1993. 97, 329–337.
Zurek E, Foxcroft OR, Kennelly JJ, Metabolic status and interval to first ovulation in postpartum dairy
cows. J. Dairy Sci. 1995. 78, 1909–1920.

43
2. PERFILES HORMONALES Y DINAMICA FOLICULAR DURANTE EL
PERIODO POSPARTO EN VACAS DE RAZA SANMARTINERO
2.1 RESUMEN
El objetivo de este estudio fue determinar los niveles de progesterona y estradiol y
su relación con la función ovárica durante el periodo posparto en vacas de la raza
Sanmartinero. 18 vacas lactantes, paridas en la misma temporada, fueron
estudiadas desde el día 7 posparto hasta la reactivación de la ciclicidad ovárica.
Se realizaron muestreos sanguíneos tres veces por semana para determinar los
niveles de progesterona y estradiol en suero sanguíneo mediante
radioinmunoanálisis. Se evaluó la condición corporal, en una escala de 1 a 5
(Edmonson et al., 1989), durante el periodo posparto.
El punto mínimo de condición corporal (nadir) promedio fue el día 49.47±21.55
posparto, y el intervalo parto a primera ovulación fue de 63.82±25.02 días. La
mayoría de las vacas (16/17) presentaron una fase luteal corta luego de la primera
ovulación, con una duración promedio 6.69±1.54 días. Los signos de estro fueron
de menor intensidad o estuvieron ausentes antes de la primera ovulación. Los
niveles promedio de Estradiol mostraron una tendencia creciente a través de la
lactancia, desde 26.43±5.6 pg/ml en las dos primeras semanas después del parto,
hasta 43.76±6.5 pg/ml hacia el momento de la primera ovulación (R2=0.436) en un
patrón consistente. Este estudio permite confirmar hallazgos previos con respecto
a los patrones endocrinos así como su relación con el estatus nutricional.

44
2.2 INTRODUCCION
Uno de los principales problemas para los productores de carne bovina es el
intervalo que transcurre desde el momento del parto hasta la aparición de un estro
con ovulación, en especial en el trópico (Galina, 2005). En Colombia el porcentaje
de natalidad es del 54% (Minagricultura, 2005), indicando que el promedio de
intervalo entre partos (IEP) es de 676 días y el intervalo parto-concepción cercano
a los 400 días, cuando los parámetros ideales son 365 días de intervalo posparto y
85 días de intervalo parto concepción (Basurto, 2005).
El anestro posparto es un periodo de transición durante el cual se recupera la
funcionalidad del eje hipotálamo-hipófisis-ovarios-útero de la preñez previa. El
anestro en el trópico se manifiesta por largos intervalos parto-primer servicio y
parto-concepción, superiores a los 120 y 170 días, respectivamente (Basurto,
2005).
Numerosos autores han puesto de manifiesto que el estado de balance energético
del animal en el periparto afecta la reanudación de la actividad ovárica (Galina,
2005). Durante las 3 primeras semanas ocurre la involución uterina, el
reestablecimiento de los depósitos de LH y el inicio del desarrollo de ondas
foliculares. Sin embargo, la presencia del ternero al lado de la vaca durante el
amamantamiento inhibe los pulsos de liberación de GnRH debido a la secreción
de opioides endógenos en el hipotálamo, dando como resultado supresión de la
liberación pulsátil de LH y falla en la ovulación (Yavas y Walton, 2000)

45
Se han realizado estudios previos acerca de la función luteal y dinámica folicular
asociada con el reinicio de la actividad cíclica en vacas durante el periodo
posparto. En un estudio realizado por Perry et al. (1991), se demostró que la
presentación de fases luteales cortas, o ciclos de corta duración fue un evento
común en vacas de carne lactantes durante la reactivación posparto. En ese
mismo estudio se reporto que las concentraciones de 17β estradiol presentaron
una tendencia creciente durante el periodo posparto, hasta el momento de la
primera ovulación.
Se tiene claramente identificada la ineficiencia reproductiva como uno de los
factores determinantes de la disminución en la productividad ganadera, y dentro
de esa ineficiencia el anestro posparto participa de manera importante. Sin
embargo, se han identificado algunos individuos, o grupos de individuos que se
caracterizan por presentar un comportamiento reproductivo superior, este es el
caso de las razas criollas bovinas Colombianas, y de algunos individuos
particulares de otras razas.
Existen al menos siete razas criollas en Colombia distribuidas en el país. Se han
realizado varios estudios relacionados con aspectos reproductivos en Costeño
Con Cuernos (Báez et al., 2007) y Romosinuano (Grajales et al, 2006). Estos
genotipos han mostrado adaptación a las condiciones ambientales de los trópicos
en términos de desempeño reproductivo.
La generación de información básica acerca de los fenómenos fisiológicos
durante el posparto permitirá en el largo plazo, hacer selección temprana de

46
individuos superiores incrementando la productividad en la ganadería bovina del
país.
2.3 HIPOTESIS
En hembras bovinas de la raza criolla Sanmartinero, las relaciones hormonales y
foliculares se encuentran vinculadas a la expresión de fenómenos reproductivos
que posiblemente determinan su eficiencia.
2.4 OBJETIVO
Generar información básica acerca de los fenómenos fisiológicos en vacas criollas
Sanmartinero durante el posparto, identificando posibles patrones ocurrentes en
este genotipo en las condiciones particulares del trópico que puedan estar
determinando una expresión en la eficiencia reproductiva.
2.5 OBJETIVOS ESPECIFICOS
Determinar los niveles de progesterona durante el periodo posparto y el
restablecimiento de la ciclicidad estral.

47
Generar información referencial acerca de la dinámica folicular durante el
periodo posparto, la reactivación ovárica y el ciclo estral, con base en la
expresión de hormona 17β estradiol.
Relacionar la condición corporal de los animales con la reactivación de la
ciclicidad ovárica
2.6 MATERIALES Y METODOS
2.6.1 Tipo de Estudio. El presente estudio tiene un componente descriptivo de
las determinaciones hormonales (progesterona, estradiol) y foliculares; y un
análisis desde el punto de vista fisiológico de estas variables, para integrar las
descripciones y obtener una información básica valiosa.
2.6.2 Localización. El estudio se realizó en el Centro de Investigaciones La
Libertad, CORPOICA, ubicado en el Departamento de Meta, zona de piedemonte
llanero, a 330 msnm, con una temperatura promedio de 26C, 2800 mm/año de
precipitación, distribuida entre los meses de abril y diciembre, y una humedad
relativa de 85% en época lluviosa y 65% en época seca, iniciando en la temporada
de parición (marzo-abril).
2.6.3 Animales. Se evaluaron 18 vacas Sanmartinero entre tercer y quinto parto,
con edades, pesos y registros productivos y reproductivos similares. Se

48
seleccionaron animales con condición corporal de 3.0 a 3.5 (escala 1-5) al
momento del parto. Se tomaron datos de condición corporal de las vacas para
construir un perfil de variación durante la lactancia. Los animales se manejaron en
pastoreo alterno en sistema de cría libre, en praderas compuestas principalmente
por Brachiaria decumbens. Las vacas estuvieron en presencia permanente de dos
machos receladores con pene desviado.
2.6.4 Aval del comité de ética para la experimentación animal. El trabajo se
realizó bajo el aval del Comité de Bioética de la Facultad de Medicina Veterinaria y
de Zootecnia – UNAL Bogotá; igualmente, bajo los parámetros de manejo animal
del Centro de Investigaciones Corpoica La Libertad.
2.6.5 Muestreos sanguíneos. Se tomaron muestras de sangre tres veces por
semana mediante punción de la vena coccígea, en tubos de vacío sin
anticoagulante de 10 ml (Becton Dickinson and Company, Franklin Lakes NJ,
USA), iniciando 7 días después del parto y durante la reanudación de la actividad
reproductiva, monta y hasta la confirmación de preñez. Las muestras de sangre
fueron centrifugadas a 2500 rpm durante 15 minutos y el suero mantenido a -20º C
hasta la cuantificación de los niveles de progesterona y estradiol por la técnica del
radioinmunoanálisis (RIA) en fase sólida (MP Biomedicals, ICN Inc, Costa Mesa,
CA), sin realizar ningún tipo de extracción. El coeficiente de variación intraensayo
fue de 1.85% para progesterona y 3.53% para estradiol, y el coeficiente de
variación interensayo fue de 4.61% para progesterona y 5.99% para estradiol.

49
2.6.6 Condición corporal. Se evaluó condición corporal, en una escala de 1 a 5,
(Edmonson et al., 1989), con el fin de determinar de manera indirecta el balance
energético. El evaluador fue el mismo durante todas las observaciones y tuvo un
entrenamiento previo al inicio de estas.
2.7 ANALISIS DE DATOS
Este estudio es un análisis exploratorio de datos acerca de la fisiología
reproductiva posparto en hembras bovinas de la raza Sanmartinero, se describen
las variables hormonales y de condición corporal mediante estadística descriptiva,
medidas de tendencia central y de dispersión, y con base en el conocimiento
fisiológico se integran, observando patrones comunes de comportamiento. Se
presentan graficas de los perfiles endocrinos de cada individuo.
2.8 RESULTADOS Y DISCUSION
2.8.1 Reactivación ovárica posparto. Se considero el reinicio de la actividad
ovárica cuando al menos tres muestreos consecutivos presentaron niveles de P4
superiores a 1 ng/ml (Koniggson et al., 2008). El intervalo parto – primera
ovulación fue de 63.8±25 días, con un rango entre 31 a 106 días (Tabla 1), esta
primera ovulación estuvo seguida en la mayoría de animales de una fase luteal de
corta duración, y una segunda ovulación, generalmente acompañada de signos de
estro a los 70.12±25.07 días. En la figura 1 se observan los perfiles individuales de

50
de progesterona que muestran el momento de la reactivación ovárica. De acuerdo
con la clasificación hecha por Opsomer et al. (1998) sobre actividad ovárica
posparto, el 61% de las vacas (11/18) en este estudio presento actividad normal
tipo 1(reinicio de la actividad ovárica antes del día 80 posparto), el 28% (5/18)
actividad tipo 2 (reactivación alrededor del día 100 posparto), uno de los animales
(5.5%) mostro el tipo 3 (interrupción de la ciclicidad) así como otra vaca (5.5%)
mostro anestro prolongado (tipo 4).
En un estudio realizado en vacas Costeño con Cuernos (Báez et al., 2007) se
observó estro con ovulación a los 51 días posparto en 20% de las vacas (2/10),
mientras que el 70% (7/10) presento ovulación hacia los 70-80 días posparto, y un
10% (1/10) el día 90 posparto. Grajales (2001) observo 87.6% de perfiles Tipo 1
en vacas Romosinuano, 63.6% en vacas Simmental X Cebú y 41.7% en vacas
Holstein X Cebú.
Domínguez et al. (2004) encontraron en vacas doble propósito en el trópico
venezolano que el 50% de las vacas reiniciaron actividad luteal alrededor de los
90 días postparto y el 74 % aproximadamente a los 120 días post-parto. Un 26%
de las vacas se mantuvieron aciclicas durante este período. En ganado de doble
propósito los promedios en el inicio de la actividad luteal estuvieron entre los
valores comprendidos de 70 y 100 días post-parto: 103 ± 54 días (Rubio et al.,
1999); 71,7 días (Martínez et al., 1996); 73,2 días (Domínguez et al., 1996). En
vacas mestizas cebú el reinicio de actividad luteal fue a los 91,89 ± 6 días
(Hernández-Fernández et al., 1998).

51
Ramírez-Iglesia et al. (2008) reportan para vacas criollas de la raza Carora en el
trópico venezolano, un intervalo parto-primer celo de 73±2 días. En un estudio de
Soto-Belloso et al. (1997) realizado en 49 hembras brahman de edad adulta en
trópico húmedo, con presencia de machos receladores, el intervalo parto-primer
estro fue de 76.3±8.8 días, mientras que en animales no expuestos a toro el
intervalo fue de 95±10.1 días. En el presente estudio las vacas estuvieron en
presencia de machos receladores.
Corro et al. (1999) encontraron una alta variación en los valores de intervalos
parto-primer estro entre diferentes granjas de doble propósito en el trópico
mexicano, igualmente, hallaron diferencias entre animales que parieron en época
seca (111±8.7 días) y época de lluvias (145±6.9 días).
Con respecto al sistema de amamantamiento, Pérez et al. (2001) reportan un
intervalo de días a primer estro de 69.9±24.7 días en amamantamiento tradicional,
frente a 58.4±23.6 días en sistemas que iniciaron el amamantamiento 8 horas
después del ordeño, durante 1 hora, en explotaciones doble propósito con cruces
Bos indicus x Bos taurus en el trópico mexicano. En el presente estudio el
amamantamiento fue de tipo tradicional, con la cría permanentemente junto a su
madre, y sin ordeño.
Los resultados de este estudio son similares a los encontrados por Sinclair et al
(2002) en ganado de carne lactante en zona templada y superiores a los

52
observados por Ruiz-Cortez y Olivera-Angel (1999) en vacas Bos Indicus lactantes
mantenidas en praderas naturales en Colombia, las cuales reanudan su ciclicidad
de los 217 a los 278 días posparto, resultando en un intervalo entre partos de 17-
19 meses.
Cordova y Pérez (2002) recopilaron una serie de datos reproductivos en el trópico
mexicano, y reportaron que el intervalo parto-primer estro es de 70±34 días, con
un rango variando entre 44 y 112 días. En este mismo estudio se presentan
diferencias entre diferentes climas tropicales, reportando intervalos de 83 días en
trópico húmedo, 76 días en trópico seco, 60 días en trópico subhumedo y 43 días
en clima subtropical húmedo.
Figura 1. Perfiles individuales de progesterona durante el posparto.
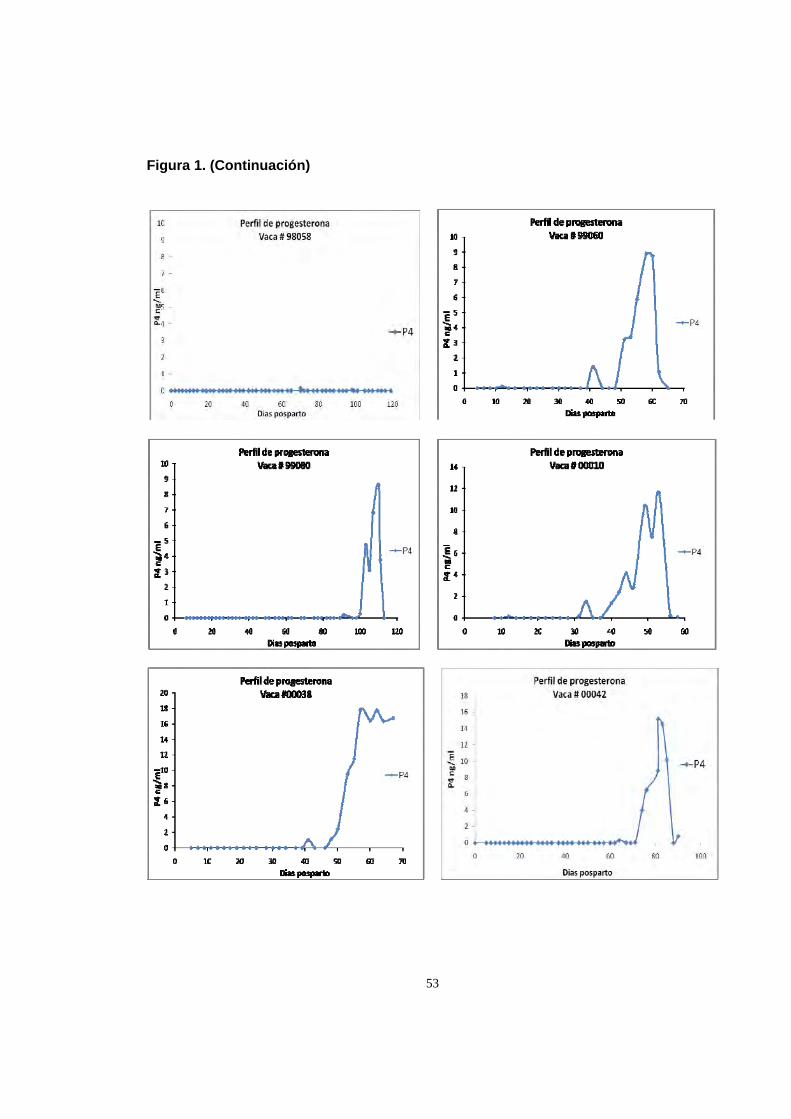
53
Figura 1. (Continuación)
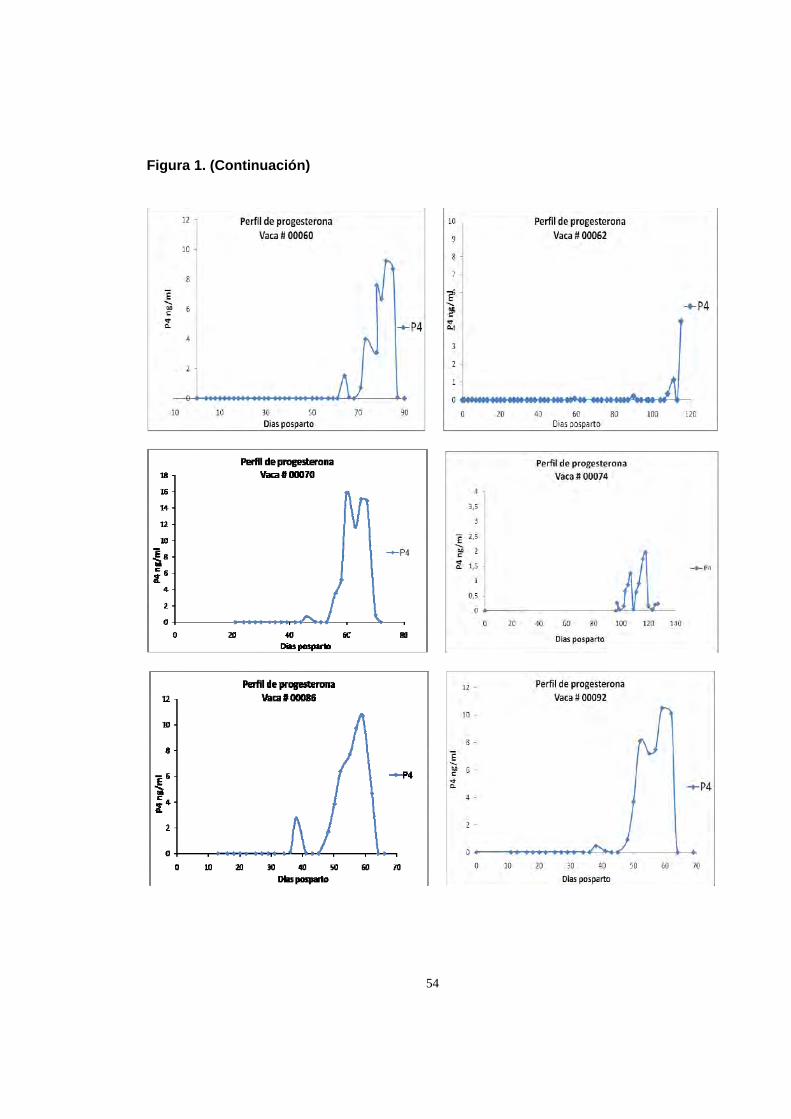
54
Figura 1. (Continuación)

55
Figura 1. (Continuación)
2.8.2 Ciclos de corta duración. La ovulacion se determino como la presentacion
de niveles de progesterona menores a 0.5 ng/ml seguida de por lo menos dos
muestras con niveles superiores a 1 ng/ml. El intervalo parto – segunda ovulacion
fue de 70.12±25.07 dias (Tabla 1.). 94% de las vacas que mostraron reanudación
de la ciclicidad durante el estudio (16/17) presentaron incrementos de corta
duracion en los niveles de progesterona previos al primer ciclo estral normal
(Figura 1), estos ciclos breves tuvieron una duración promedio de 6.69±1.54 dias

56
(Tabla 1), siendo 7 dias la duracion mas común (7 de 16 vacas) (Figura 2.). En un
estudio realizado por Perry et al. (1991) en 13 vacas de carne, todas las vacas
exhibieron una fase luteal corta luego de la primera ovulacion, con un intervalo
entre la primera y la segunda ovulacion de 8.5±0.2 días. Los ciclos cortos ocurren
normalmente en la primera ovulación espontánea posparto en la mayoría de vacas
de carne (Day, 1990); Taponen et al. (2003) demostraron que la regresión luteal
del cuerpo lúteo durante los ciclos estrales cortos es debida a la liberación
prematura de PGF2α. La progesterona producida durante la fase luteal corta
puede ser necesaria para el establecimiento de una función luteal normal en el
siguiente ciclo estral (Short et al., 1990; Mihm, 1999).
En un estudio de Ramírez-Iglesia et al. (1992) realizado en trópico seco en
Venezuela, y utilizando animales Brahman y cruces con holstein y pardo suizo, se
encontró que previo al primer estro se presentaron descargas de progesterona
que duraron en promedio 19,11 y 10 días para las razas brahmán y los cruces con
pardo suizo y holstein respectivamente, sugiriendo que en el ganado brahman la
corta duración de este primer ciclo estral no es tan evidente. Igualmente, en este
estudio se reporta que esas primeras descargas de progesterona solo fueron
evidentes en 50.8% de los animales.
En Tanzania, Lyimo et al. (2004) encontraron que, en vacas doble propósito cebú
x holstein mantenidas en pastoreo en la costa tropical sub-húmeda, con destete
temprano (3-4 semanas post parto) y ordeñadas dos veces al día, el primer

57
incremento de progesterona sucedió a los 66.1±8.1 días (rango 21-146) y se
evidenciaron ciclos de corta duración (< 14 días) en 45% de los animales.
Figura 2. Duración de los ciclos cortos en el posparto
2.8.3 Perfiles de estradiol durante el posparto. A partir de la primera semana
de lactancia se evidenciaron variaciones en los niveles de estradiol. Estos niveles
mostraron una tendencia lineal creciente a través de la lactancia, desde 26.43±5.6
pg/ml promedio en la segunda semana después del parto, hasta 43.76±6.5 pg/ml
hacia el momento de la primera ovulación (R2=0.436) en un patrón consistente
(Figura 3). Este coeficiente de determinación es aceptable si se tiene en cuenta
que se trata de una recta ajustada a unos niveles de estradiol variantes entre cada
muestreo, lo cual es lógico si se tiene en cuenta la dinámica de crecimiento y
atresia folicular y que es correspondiente con incrementos y disminuciones en los

58
niveles de secreción de 17β estradiol. En un estudio realizado por Perry et al.
(1991) en vacas de carne amamantando se reporto que las concentraciones de
17β estradiol presentaron una tendencia creciente durante el periodo posparto,
hasta el momento de la primera ovulación.
Los valores de estradiol deben ser mirados con cautela ya que no indican
directamente un comportamiento de las estructuras ováricas, sino que a partir de
las variaciones entre muestreos observadas en los perfiles de estradiol se sugiere
una dinámica folicular entendida como el posible crecimiento y atresia de folículos,
y el consiguiente incremento y descenso de los niveles de secreción en las células
de la granulosa, y se pueden inducir algunos fenómenos fisiológicos relacionados
como los incrementos transitorios de FSH previos al reclutamiento (Ireland et al.,
2000).
Figura 3. Perfiles de progesterona y estradiol durante el posparto

59
Figura 3. (continuación)
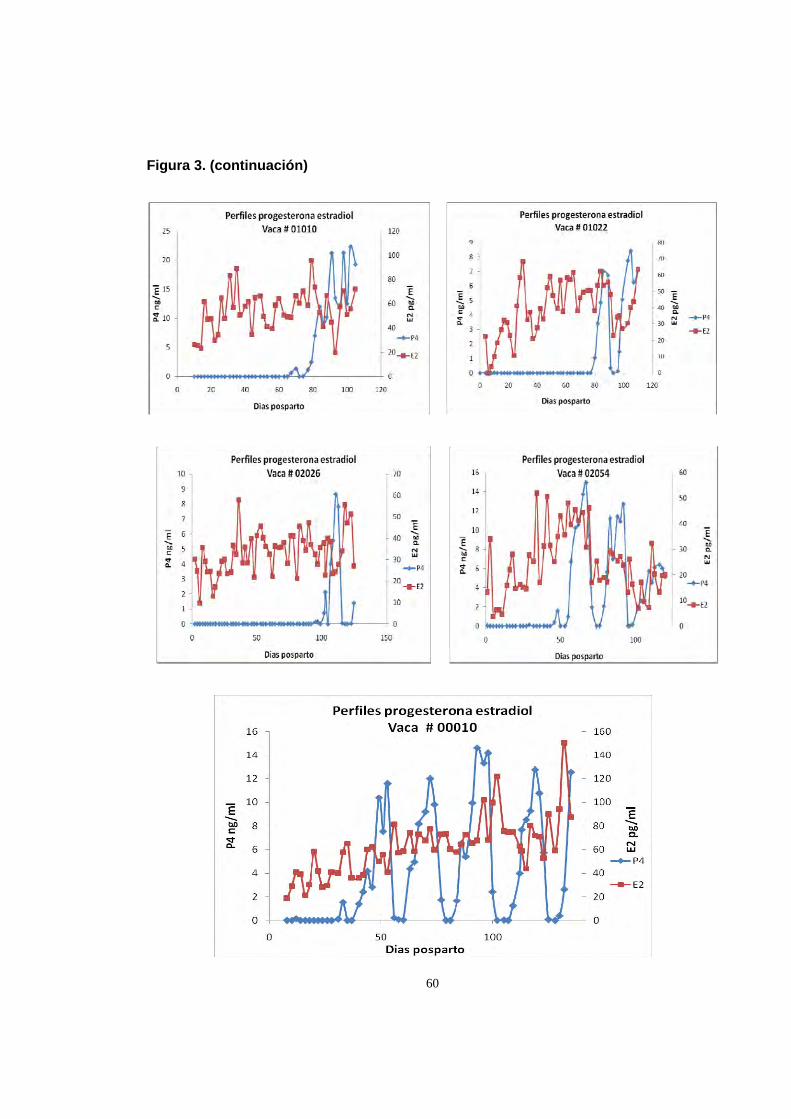
60
Figura 3. (continuación)

61
2.8.4 Reactivación ovárica posparto y condición corporal. Los valores medios
de condicion corporal estuvieron entre 2.6 y 3.6 en la mayoria de los animales, con
extremos individuales de 2.0 y 4.0 en dos de los animales. El punto mínimo de
condición corporal (nadir) promedio se presento el día 49.4±21. (Figura 4), en la
Tabla 1 se presentan los datos individuales del momento en que cada animal
alcanza el nadir de condición corporal, y se puede observar como en la mayoría
de los casos este se presenta unos días antes de la primera ovulación (Figura 5).
Estos resultados coinciden con los encontrados en vacas de la raza costeño con
cuernos (Báez et al., 2007). Corro (1992) observó en vacas Bos indicus en
condiciones normales de explotación valores mínimos entre los 90 y 120 días
posparto y una ligera recuperación hasta los 180 días. Según Galina y Maquivar
(2005), existe la necesidad de superar la etapa de nadir para poder esperar una
reactivación de la función reproductiva. Una vez que los animales dejan de perder
condición corporal y/o peso, el balance energético se equilibra, por lo que la
retroalimentación negativa cesa y la secreción de GnRH se reanuda normalmente.
De acuerdo a lo anterior, el momento del nadir de condición corporal estuvo
relacionado con la presentación de los primeros signos de estro. Sinclair et al
(2002) observaron que la condición corporal al parto es de suma importancia, ya
que las vacas que paren con una buena condición presentan el estro con
ovulación primero que vacas con bajas puntuaciones.
Corro (1999) encontró, para vacas doble propósito en trópico húmedo mexicano,
valores medios de condición corporal variando en un rango entre 2.9 y 3.9; con

62
una disminución en las primeras 4 semanas entre 3.5 y 2.6, y se presentaron
diferencias significativas en la condición corporal en el momento del parto entre
vacas que parieron en época de lluvias (3.7) y época seca (3.3). En el presente
estudio las vacas parieron al final de la época seca e inicios de la época de lluvias.
Ramírez-Iglesia et al., (1994) Observaron condición corporal al momento del parto
de 3.0±0.2 y de 2.4±0.2 al momento del primer celo, en 27 vacas mestizas
producto del cruce indiscriminado de razas lecheras, criollo Carora y cebú, con
predominio de la raza Carora, en trópico seco del piedemonte andino venezolano.
En un estudio mas reciente (Ramírez-Iglesia et al., 2008) este mismo grupo
estudio 89 vacas en similares condiciones, y observaron una relación entre la
condición corporal y el primer celo posparto. En el citado estudio los animales con
condición corporal al parto <3.0 presentaron el primer celo posparto a los 77±4
días, con condición corporal igual a 3.0 el primer celo se presento a los 71±3 días
y animales con condición superior a 3.0 presentaron primer celo posparto a los
67±3 días.

63
Figura 4. variación promedio de la condición corporal durante el posparto
Figura 5. Perfiles individuales de variacion de la condicion corporal, en relacion con
la reactivacion de la actividad ovarica medida por perfiles de progesterona.

64
Figura 5. (Continuación)

65
Figura 5. (Continuación)
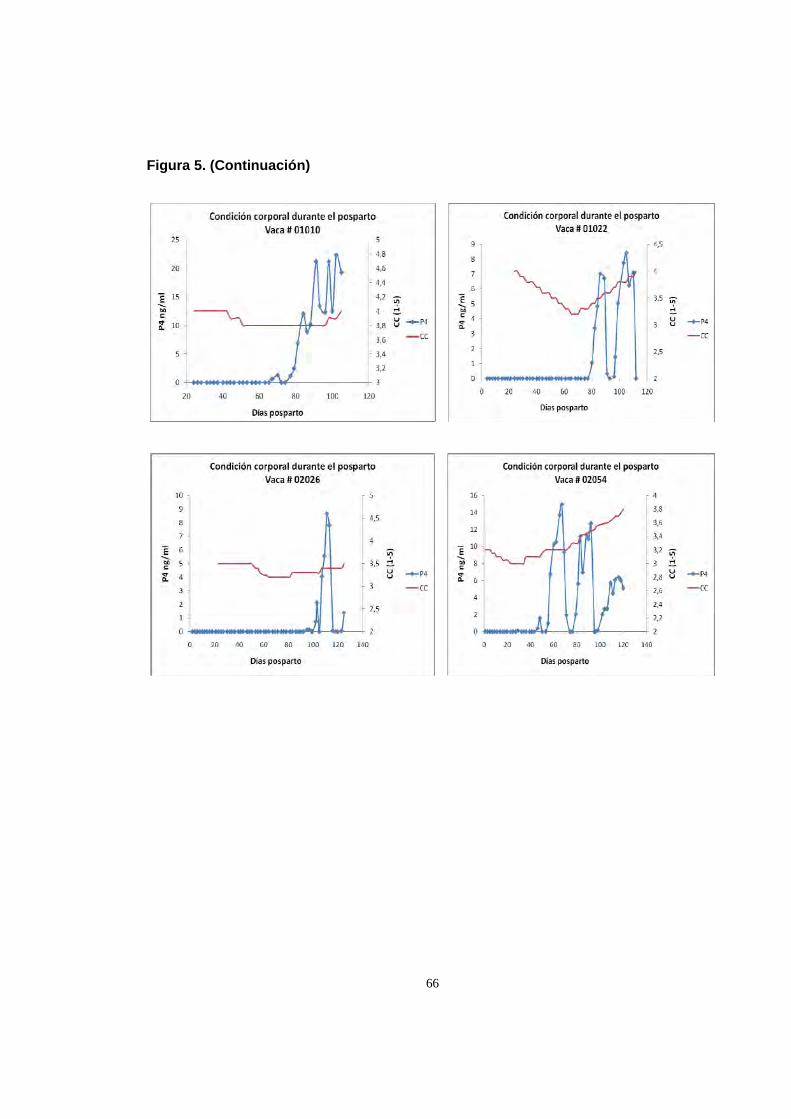
66
Figura 5. (Continuación)

67
Tabla 1. Cuadro resumen. Intervalos a la primera ovulación, segunda ovulación,
duración del ciclo breve y nadir de condición corporal
Vaca PPO PSO DCB P4CB NCC
(días) (ng/ml) días 98036 75 82 7 0,79 98056 86 95 9 1,26 50 99058 anestro anestro 51 99060 39 46 7 1,43 24 99080 89 98 9 0,19 65 00010 31 36 5 1,52 10 00038 39 44 5 1,02 16 00042 62 69 7 0,32 50 00060 64 68 4 1,51 50 00062 106 113 7 1,14 100 00070 44 51 7 0,68 56 00074 95 99 4 0,26 64 00086 36 43 7 2,75 50 00092 36 43 7 0,47 50 01010 65 73 8 1,31 51 01022 75 65 02026 99 105 6 2,12 64 02054 44 52 8 1,6 25
Promedio 63,82 70,12 6,69 1,15 49,47 Desviación 25,02 25,07 1,54 0,70 21,55
Mediana 64 69 7 1,2 50 Mínimo 31 36 4 0,19 10 Máximo 106 113 9 2,75 100
PPO: Intervalo parto – primera ovulación
PSO: Intervalo parto – segunda ovulación
DCB: Duración del ciclo corto
P4CB: Nivel máximo de P4 durante el ciclo corto
NCC: Nadir de condición corporal

68
2.9 CONCLUSIONES
La primera ovulación post parto ocurrio a los 63.82±25.02 días. En la mayoria de
vacas de este estudio (16/17), se evidencio un ciclo de corta duracion posterior a
esta ovulacion, con una duracion de 6.69±1.54 días. Posterior a la presentación de
este primer ciclo corto, los animales estabilizaron su ciclicidad estral.
Los niveles de secrecion de estradiol mostraron un incremento a lo largo del
posparto, desde 26.43±5.6 pg/ml en la segunda semana después del parto, hasta
43.76±6.5 pg/ml en el momento de la primera ovulación (R2=0.436); posterior a
esta, los niveles promedio de estradiol tendieron a estabilizarse.
La condicion corporal en los animales de este estudio disminuyo luego del parto,
alcanzando un nadir el día 49.47±21.55 que estuvo relacionado con el reinicio de
la actividad reproductiva.
La identificación de estos patrones fisiológicos en este genotipo particular, es una
base de información importante que sirve como punto de partida para nuevas
investigaciones que permitan ampliar el conocimiento de los factores que afectan
el periodo de restablecimiento de la actividad ovarica posparto en el medio
tropical.

69
BIBLIOGRAFIA
Báez GM, Grajales HA y Pérez JE Caracterización del ciclo estral mediante perfiles de esteroides
(progesterona, 17 β-estradiol) en la raza Costeño con cuernos (Bos taurus) en el trópico
Colombiano. Livestock Research for Rural Development 2007 Volume 19, Article #132.
http://www.cipav.org.co/lrrd/lrrd19/9/baez19132.htm
Basurto, HB El anestro posparto en la ganadería bovina del trópico. Congreso internacional de
reproducción bovina. Intervet. 2005. Bogotá.
Butler WR Energy balance relationships with follicular development, ovulation and fertility in
postpartum dairy cows. Livestock Production Science 2003: 83 211–218
Córdova A, Pérez JF. Indicadores reproductivos de bovinos en el trópico mexicano y factores que
lo determinan . Med Vet 2002 vol. 19 (3): 47-56.
Corro M, Rubio I, Castillo E, Galindo L, Aluja A, Galina CS, Murcia C. Effect of blood metabolites,
body condition and pasture management on milk yield and postpartum intervals in dual-purpose
cattle farms in the tropics of the State of Veracruz, Mexico. Preventive Veterinary Medicine 1999
38: 101-117
Corro, M.M.D. Efecto de la suplementario mineral preparto sobre el comportamiento reproductivo y
productivo posparto en vacas Holstein X Cebú en trópico húmedo . Tesis de maestría. FMVZ,
UNAM, México., D.F. 1992
Day ML, Dyer RM, Wilson GW, Pope WF Influence of estradiol on duration of anestrus and
incidence of short estrous cycles in postpartum cows. Domest. Anim. Endocrinol 1990. 7, 19–25.
Domínguez C, Martínez N y Colmenares O. Características reproductivas de rebaños bovinos
doble propósito en los llanos centrales de Venezuela. Zootecnia Tropical 2004 22(2):133-145.
Domínguez C, Martínez N, Labrador C, Risso J, López S. Effect of strategic feed supplementation
with multinutrient block on productive and reproductive performance in dual purpose cows.. En:
Development of feed supplementation strategies for improving ruminant productivity on small –
holder farms in Latin-America through the use of immunoassay techniques. IAEA-TELDOC-877.
IAEA. 1996. Viena, .pp. 97-105.
Edmonson AJ, Lean IJ, Weaver LD, Farver T, Webster G A body condition chart for Holstein dairy
cows. J Dairy Sci 1989 72, 68–78.
Galina CS, Arthur GH. Review on cattle reproduction in the tropics. Part 4. Oestrous cycles. Animal
Breeding Abstracts 1990 58: 697-707.
Galina CS, Maquivar M. Sistemas de separación de terneros de vacas lactantes y su efecto en la
producción de un celo fértil. Congreso internacional de reproducción bovina. Intervet. Bogota.
2005.

70
Grajales HA. Comportamiento reproductivo de grupos raciales bovinos en el trópico calido-humedo
Colombiano: Pubertad, ciclo estral, preñez temprana, posparto. Niveles de hormonas esteroides
y su relación con la eficiencia reproductiva. 2001. Tesis PhD. Facultad de Medicina Veterinaria y
de Zootecnia. Universidad Nacional de Colombia. Bogota D.C.
Grajales H, Hernández A y Prieto E Determinación de parámetros reproductivos basado en los
niveles de progesterona en novillas doble propósito en el trópico Colombiano. Livestock
Research for Rural Development 2006. Volume 18, Article #144.
http://www.cipav.org.co/lrrd/lrrd18/10/graj18144.htm
Henao GM, Olivera-Ángel M, Maldonado-Estrada JG. Follicular dynamics during postpartum
anestrus and the first estrous cycle in suckled or non-suckled Brahman (Bos indicus) cows.
Animal Reproduction Science 2000 63 127–136
Hernández-Fernández A, Soto Belloso A, Portillo G, Rincón R, Cahuao N. Efecto del destete
temporal y progestágenos sobre la eficiencia reproductiva en vacas mestizas cebú en anestro:
Intervalos reproductivos. Rev. Facultad. Agronomía (LUZ) 1998 15:350-358. .
Ireland JJ, Mihm M, Austin E, Diskin MG, Roche JF. Historical perspective of turnover of dominant
follicles during the bovine estrous cycle: key concepts, studies, advancements, and terms. J
Dairy Sci. 2000 Jul;83(7):1648-58.
Konigsson K, Savoini G, Govoni N, Invernizzi G, Prandi A, Kindahl H and Veronesi GM. Energy
balance, leptin, NEFA and IGF-I plasma concentrations and resumption of post partum ovarian
activity in swedish red and white breed cows. Acta Veterinaria Scandinavica 2008, 50:3
Lyimo ZC, Nkya R, Schoonman L and van Eerdenburg FJCM. Post-partum reproductive
performance of crosbred dairy cattle on smallholder farms in sub-humid coastal Tanzania.
Tropical Animal Health and Production, 36(3), 269-279
Mann GE, Lamming GE. The role of sub-optimal preovulatory oestradiol secretion in the aetiology
of premature luteolysis during the short oestrous cycle in the cow. Anim Reprod Sci. 2000 Dec
29;64(3-4):171-80.
Mihm. M. Delayed resumption of cyclicity in postpartum dairy and beef cows Reprod. Dom. Anim.
1999: 34: 276-284
Minagricultura. 2005. Programa desarrollo ganadero 2005-2019. Ministerio de Agricultura y
desarrollo Rural. Republica de Colombia.
Opsomer. G, Coryn. M, Deluyker. H y Kruif. A. An analysis of ovarian dysfunction in high yielding
dairy cows after calving, based on progesterone profiles. Reprod. Dom. Anim 1998 33: 193 –
204:

71
Pérez H, Solaris M, García-Winder, Osorio-Arce M y Gallegos-Sánchez J. Comportamiento
productivo y reproductivo de vacas de doble propósito en dos sistemas de amamantamiento en
el trópico. Arch. Latinoam. Prod. Anim. 2001. 9(2): 79-85
Perez-Marın CC, Espana F. Oestrus Expression and Ovarian Function in Repeat Breeder Cows,
Monitored by Ultrasonography and Progesterone Assay Reprod Dom Anim 2007 42, 449–456
Perry RC, Corah LR, Kiracofe G H, Stevenson JS and Beal WE. Endocrine changes and
ultrasonography of ovaries in suckled beef cows during resumption of postpartum estrous
cycles. J Anim Sci 1991. 69:2548-2555.
Pierson, R.A. and Ginther, O.J. Ultrasonography of the bovine ovary. Theriogenology 1984.21: 495-
507
Ramírez-Iglesia LN, Soto-Belloso E, González-Stagnaro C, Rojas A, Díaz de Ramírez A. Anestro y
días vacios en ganado lechero en el trópico. Mundo Pecuario 2008, IV, Nº 1, 22-27
Ramírez-Iglesia L, Gonzalez-Stangnaro C, Soto-Belloso E, Soto Castillo G and Rincon Urdaneta E
Factor affecting postpartum ovarían activity in crossbred primiparous tropical heifers.
Theriogenology 1992 38:449-460
Ramírez-Iglesia LN, Soto-Belloso E, González-Stagnaro C. Ciclicidad posparto en vacas mestizas
lecheras del piedemonte andino venezolano. Revista Cientifica FCV-LUZ 1994 Vol IV No 2 107-
112
Rhodes FM, De Ath G, Entwistle KW Animal and temporal effects on ovarian follicular dynamics in
Brahman heifers. Anim. Reprod. Sci. 1995.38, 265–277.
Rubio I, Corro M, Castillo E, Galindo L, Aluja A, Galina CS y Murcia C. Factors related to the onset
of postpartum ovarian activity in dual purpose cattle in the tropics. Rev. Facultad de Agronomía
(LUZ) 1999 16:637-650.
Martínez N, Escobar A, López A, Combellas J y Gabaldón L. Effect of strategic feed
supplementation on productive and reproductive performance in dual purpose cows. En:
Development of feed supplementation strategies for improving ruminant productivity on small –
holder farms in Latin-America through the use of immunoassay techniques. IAEA-TELDOC-877.
IAEA. Vienna. (AUSTRIA). 1996. pp-135-144.
Ruiz-Cortez ZT, Olivera-Angel M. Ovarian follicular dynamics in suckled zebu (Bos indicus) cows
monitored by real time ultrasonography. Anim. Reprod. Sci. 1999. 54, 211–220.
Savio JD, Keenan L, Boland MP and Roche JF. Pattern of growth of dominant follicles during the
oestrous cycle of heifers. J. Reprod. Fertil. 1988 83: 663- 671
Short RE, Bellow RA, Staigmiller RB, Berardinelli JG, Custer EE. Physiological mechanisms
controlling anestrous and fertility in postpartum beef cattle. J. Anim. Sci. 1990: 68: 799-816

72
Sinclair KD, Molle G, Revilla R, Roche JF, Quintans G, Marongiu L, Sanz A, Mackey DR, Diskin MG
Ovulation of the first dominant follicle arising after day 21 postpartum in suckling beef cows.
Animal Science 2002 75: 115-126.
Sirois J and Fortune JE Ovarian follicular dynamics during the estrous cycle in heifers monitored by
real time ultrasonography. Biol. Reprod. 1988 39: 308- 317
Soto Belloso E, Ramirez Iglesia L, Guevara L. and Soto Castillo G. Bull effect on the reproductive
performance of mature and first calf-suckled zebu cows in the tropics. Theriogenology 1997 48
1185-1190,
Wettemann RP, Lents CA, Ciccioli NH, White FJ and Rubio I Nutritional- and suckling-mediated
anovulation in beef cowsJ. Anim. Sci. 2003 81(E. Suppl. 2):E48–E59
Yavas Y, Walton JS. Postpartum aciclicity in suckled beef cows: a review. Theriogenology 2000 54:
25-55

73
3. CICLICIDAD ESTRAL POSPARTO, MOMENTO DEL INCREMENTO
POST- OVULATORIO DE PROGESTERONA Y LUTEOLISIS
EN VACAS SANMARTINERO
3.1 RESUMEN
El objetivo de este estudio fue describir el incremento post-ovulatorio de los
niveles de progesterona, así como su posterior descenso en la fase de luteolisis,
durante el ciclo estral, en vacas Sanmartinero. Se estudiaron los ciclos estrales de
15 vacas lactantes luego del restablecimiento posparto de la ciclicidad ovárica. Se
realizaron muestreos sanguíneos tres veces a la semana mediante venopunción
coccígea, y se determinaron los niveles de progesterona mediante técnica de
radioinmunoanálisis. La duración del ciclo estral fue de 20.95±1.99 días, con
niveles máximos de progesterona de 10.23±2.45 ng/ml el día 15. Se encontraron
niveles basales de progesterona por debajo de 0.5 ng/ml hasta el día 3 después
de la ovulación, similar a lo encontrado en otras razas criollas (Báez et al., 2007;
Grajales et al 2006, Grajales et al 2008), posterior al día 3 luego de la ovulación,
los niveles de progesterona mostraron un crecimiento consistente a una tasa
constante hasta alcanzar su máximo nivel, que fue de 10.23 ng/ml en promedio.
Por otra parte, se evidencio un fuerte descenso en los niveles promedio de
progesterona (49.44%) entre los días 17 y 19 del ciclo estral, durante la luteolisis.

74
3.2 INTRODUCCION
Las más altas perdidas reproductivas en ganado se concentran en el periodo
embrionario, especialmente durante los primeros 42 días luego de la fertilización
(Inskeep, 2004). La secreción luteal de progesterona es esencial para la ovulación
de un oocito saludable y la sobrevivencia embrionaria. Durante la fase luteal que
antecede al estro, e inmediatamente después de este, la progesterona regula el
momento de la siguiente luteolisis, en el caso que no exista preñez (Garrett et al.,
1988). Igualmente, la progesterona prepara al útero para el reconocimiento de la
gestación (Vincent et al., 1986). Adicionalmente, las concentraciones de
progesterona regulan el desarrollo folicular ejerciendo un control negativo sobre la
secreción pulsátil de LH (Kinder et al., 1996).
Las concentraciones de progesterona han sido relacionadas con mortalidad
embrionaria durante el periodo post-ovulatorio temprano (antes del día 6 post-
ovulación) en vacas que desarrollaron folículos persistentes bajo niveles bajos de
progesterona durante el ciclo previo a la fertilización. Igualmente, durante el
periodo de reconocimiento materno de la gestación (día 14 al 17), bajas tasas de
preñez han estado asociadas a niveles bajos de progesterona (Inskeep, 2004).
Una inadecuada función luteal representa problemas reproductivos. La
concentración de progesterona circulante durante la preñez temprana es uno de
los principales determinantes del desarrollo y sobrevivencia embrionarios
(Robinson et al., 2005). Existe evidencia de que es el momento del incremento de

75
los niveles de progesterona, más que las concentraciones finales de esta, el factor
clave que influencia el desarrollo embrionario (Robinson et al., 2005).
Las hembras que se denominan repetidoras de calor presentan una asincronía
hormonal durante el estro caracterizada por niveles suprabasales de progesterona
(Duchens et al 1995; Bage et al., 2002). Ello ha estado relacionado con un retardo
en el pico de LH y la presencia de un folículo de mayor tamaño y de vida media
más larga, el excesivo crecimiento del folículo puede causar una alteración en la
maduración del oocito o alterar el medio ambiente uterino oviductal debido a la
prolongada liberación de estradiol (Gustafsson 1999).
Las razas de ganado criollo Colombiano han mostrado tener un alto grado de
adaptación a las condiciones medioambientales que se presentan en el trópico, en
términos de buen desempeño reproductivo (Báez et al., 2007; Grajales et al.,
2006). Se han logrado establecer algunos fenómenos fisiológicos particulares,
como los niveles basales de progesterona durante la fase folicular y su dinámica
de cambio durante la fase luteal (Grajales, 2001; Báez et al., 2007) que podrían
estar vinculados a esta eficiencia reproductiva, lo que lleva a la necesidad de
continuar avanzando en la investigación básica y tener mayores elementos de
juicio para identificar, analizar, comprender o explicar los mecanismos fisiológicos
de adaptación.
El objetivo de este estudio es describir el incremento post-ovulatorio de los niveles
de progesterona, así como su posterior descenso en la fase de luteolisis, durante
el ciclo estral, en vacas Sanmartinero.

76
3.3 HIPOTESIS
Luego del restablecimiento de la ciclicidad posparto, existen patrones de secreción
de progesterona durante el periodo periovulatorio, así como variación de los
niveles durante el ciclo estral, que se encuentran vinculados a fenómenos de
estructuración del cuerpo lúteo, sobrevivencia embrionaria y luteolisis, que a su
vez, afectan a los ciclos subsiguientes previos al establecimiento de una nueva
gestación.
3.4 OBJETIVO
Generar información básica acerca del comportamiento de los niveles de
progesterona durante los primeros ciclos estrales posteriores al restablecimiento
de la actividad reproductiva luego del parto en vacas criollas Sanmartinero,
identificando posibles patrones ocurrentes en este genotipo en las condiciones
particulares del trópico que puedan estar determinando una expresión en la
eficiencia reproductiva.
3.5 OBJETIVOS ESPECIFICOS
• Describir algunos eventos alrededor el restablecimiento de la ciclicidad
estral mediante los perfiles de progesterona

77
• Describir el fenómeno de agrupamiento, dentro de un periodo determinado,
de los celos para el grupo de animales en estudio
• Generar información referencial relacionada con la duración del ciclo estral
y los niveles de progesterona durante sus etapas
• Describir la dinámica de variación de la hormona progesterona durante el
ciclo estral y la preñez temprana.
3.6 MATERIALES Y METODOS
3.6.1 Tipo de Estudio. El presente estudio es un análisis exploratorio de datos
individuales de niveles de progesterona, que busca identificar patrones de
presentación de los perfiles hormonales en esta raza en particular.
3.6.2 Localización. El estudio se realizó en el Centro de Investigaciones La
Libertad, CORPOICA, ubicado en el Departamento de Meta, zona de piedemonte
llanero, a 330 msnm, con una temperatura promedio de 26C, 2800 mm/año de
precipitación, distribuida entre los meses de abril y diciembre, y una humedad
relativa de 85% en época lluviosa y 65% en época seca, iniciando en la temporada
de parición (marzo-abril).
3.6.3 Aval del comité de ética para la experimentación animal. El trabajo se
realizó bajo el aval del Comité de Bioética de la Facultad de Medicina Veterinaria y

78
de Zootecnia – UNAL Bogotá, igualmente, bajo los parámetros de manejo animal
del Centro de Investigaciones La Libertad.
3.6.4 Animales. Se evaluaron 15 vacas Sanmartinero entre tercer y quinto parto,
con edades, pesos y registros productivos y reproductivos similares. Se
seleccionaron animales con condición corporal de 2.5 a 3.0 (escala 1-5) al
momento del parto. Los animales se manejaron en pastoreo alterno en sistema de
cría libre, en praderas compuestas principalmente de Brachiaria decumbens.
3.6.5 Muestreos sanguíneos. Se tomaron muestras de sangre tres veces por
semana mediante punción de la vena coccígea, en tubos de vacío sin
anticoagulante (10 ml), durante la reanudación de la actividad reproductiva, ciclos
estrales consecutivos, monta y hasta la confirmación de preñez. Las muestras de
sangre fueron centrifugadas a 2500 rpm durante 15 minutos y el suero mantenido
en congelación a -20º C hasta la cuantificación de los niveles de progesterona
mediante radioinmunoanálisis (RIA) en fase sólida (MP Biomedicals, ICN Inc,
Costa Mesa, CA). El coeficiente de variación intraensayo fue de 1.85%, y el
coeficiente de variación interensayo fue de 4.61% para progesterona.
3.6.6 Determinación de preñez. Al final del periodo de estudio se realizo
ecografía para determinar preñez. Se utilizo un ecografo Piemedical con
transductor lineal de 7.5 MHz. Se debe aclarar que solamente unos pocos
animales fueron montados por condiciones administrativas del experimento, de

79
manera que los animales que no resultaron preñados no fue debido a fallas
fisiológicas o repetición sino posiblemente porque nunca fueron montados.
3.7 ANALISIS DE DATOS
Este estudio es un análisis exploratorio de datos acerca de la fisiología
reproductiva posparto en hembras bovinas de la raza Sanmartinero, se describen
Los niveles de progesterona mediante estadística descriptiva, medidas de
tendencia central y de dispersión, observando patrones comunes de
comportamiento. Se presentan graficas de los perfiles endocrinos de cada
individuo y diagramas de variación de los niveles de progesterona a lo largo del
ciclo estral.
3.8 RESULTADOS Y DISCUSION
3.8.1 Restablecimiento de la ciclicidad posparto. En la figura 6 se muestran los
perfiles de progesterona del grupo de animales en estudio, en donde se puede
observar la actividad cíclica durante el periodo posparto. El restablecimiento de la
ciclicidad estral luego del parto se presento antes del dia 80 en el 61% de los
animales del estudio (11/18), y para el dia 100 posparto, el 88.8% (16/18) de las
vacas ya habían estabilizado sus ciclos. Resultados similares fueron hallados en
vacas costeño con cuernos en el valle del Sinú, donde hacia el día 90 se
presentaba el restablecimiento de la ciclicidad (Báez et al., 2007)
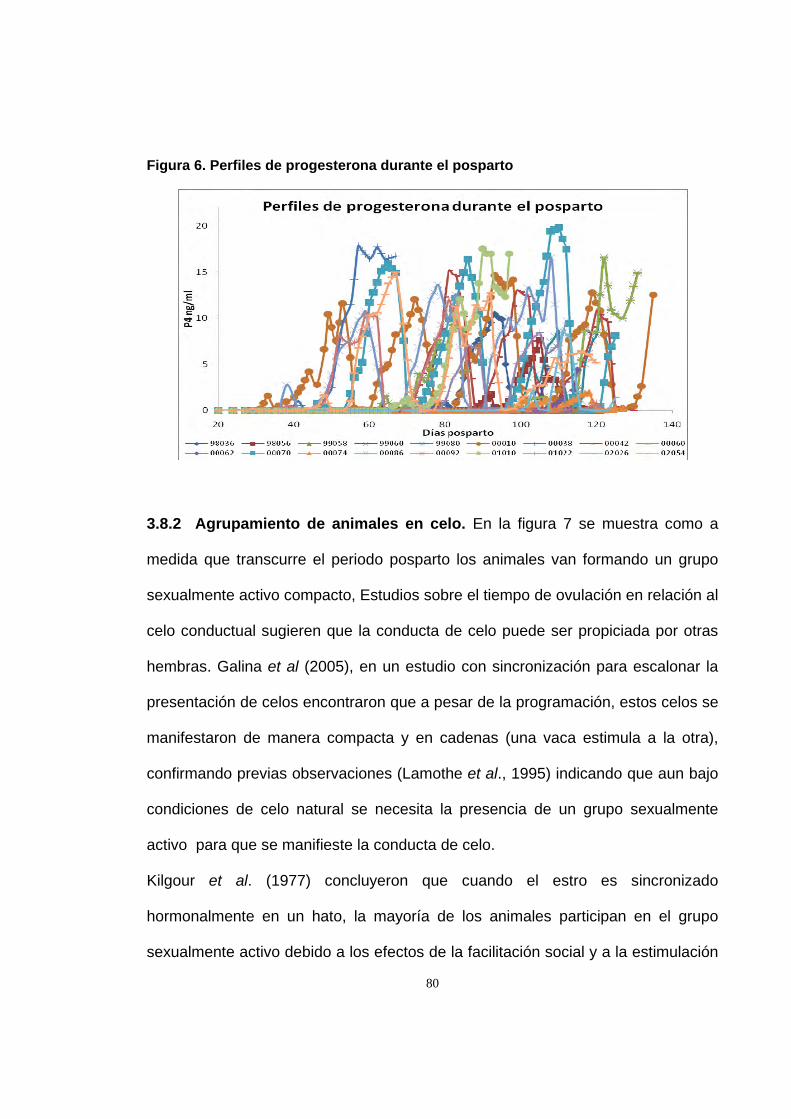
80
Figura 6. Perfiles de progesterona durante el posparto
3.8.2 Agrupamiento de animales en celo. En la figura 7 se muestra como a
medida que transcurre el periodo posparto los animales van formando un grupo
sexualmente activo compacto, Estudios sobre el tiempo de ovulación en relación al
celo conductual sugieren que la conducta de celo puede ser propiciada por otras
hembras. Galina et al (2005), en un estudio con sincronización para escalonar la
presentación de celos encontraron que a pesar de la programación, estos celos se
manifestaron de manera compacta y en cadenas (una vaca estimula a la otra),
confirmando previas observaciones (Lamothe et al., 1995) indicando que aun bajo
condiciones de celo natural se necesita la presencia de un grupo sexualmente
activo para que se manifieste la conducta de celo.
Kilgour et al. (1977) concluyeron que cuando el estro es sincronizado
hormonalmente en un hato, la mayoría de los animales participan en el grupo
sexualmente activo debido a los efectos de la facilitación social y a la estimulación
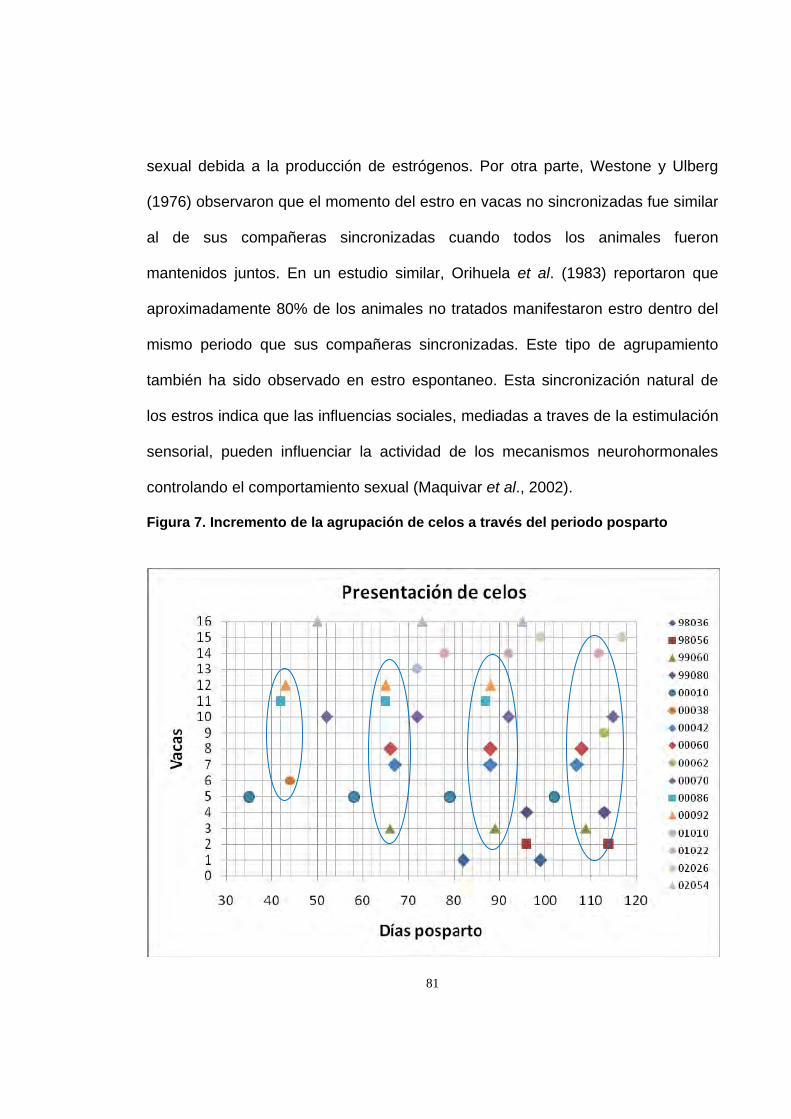
81
sexual debida a la producción de estrógenos. Por otra parte, Westone y Ulberg
(1976) observaron que el momento del estro en vacas no sincronizadas fue similar
al de sus compañeras sincronizadas cuando todos los animales fueron
mantenidos juntos. En un estudio similar, Orihuela et al. (1983) reportaron que
aproximadamente 80% de los animales no tratados manifestaron estro dentro del
mismo periodo que sus compañeras sincronizadas. Este tipo de agrupamiento
también ha sido observado en estro espontaneo. Esta sincronización natural de
los estros indica que las influencias sociales, mediadas a traves de la estimulación
sensorial, pueden influenciar la actividad de los mecanismos neurohormonales
controlando el comportamiento sexual (Maquivar et al., 2002).
Figura 7. Incremento de la agrupación de celos a través del periodo posparto

82
3.8.3 Duración del ciclo estral. Para la determinación de la duración del ciclo
estral, se considero ciclicidad regular el comportamiento estral, con o sin
manifestación de calor observado, respaldado por una función luteal basada en el
mantenimiento de los niveles de progesterona por encima de 1 ng/ml, durante al
menos tres muestreos consecutivos (Wherman et al 1996, Hommeida et al, 2005).
El promedio de duración del ciclo estral fue de 20.95±1.99 días. Grajales (2001)
reportó una duración de 18.84 +/- 3.29 días, para cuatro genotipos en trópico bajo;
en ganado Costeño con Cuernos la duración del ciclo estral fue de 21.14 días
(Báez et al., 2007). Cardozo (1993) de 21.1 +/- 1.37 días; Prieto (1993) de 20.06
+/- 2.51 para novillas Holstein y Castro y Gómez (1979) de entre 22 y 23 días para
cruces de razas europeas por cebú. En novillas Sanmartinero, Góngora y
Hernández (2006) reportaron una duración del ciclo estral de 21.2±0.3 días (rango
18-28 días) sin diferencias en función de la época seca o de lluvias
3.8.4 Niveles de progesterona durante el ciclo estral. Los perfiles de
progesterona se destacaron por presentar niveles basales sostenidos durante la
fase folicular y hasta el tercer día del ciclo estral (Tabla 2, Figura 2). Se
presentaron niveles de progesterona durante la fase luteal que alcanzaron niveles
de hasta 15 ng/ml. El promedio de nivel máximo de progesterona fue de 10.23
ng/ml, hacia el día 15 del ciclo estral (Tabla 2.). Randel (1989) observo que el CL
de hembras cebú puras y cruzadas contiene menos progesterona por gramo de
tejido luteal que el CL de hembras Bos taurus.

83
Tabla 2. Niveles de progesterona durante los días del ciclo estral
Día PromedioDesviación Estándar
% Error Estándar
0 0,0049 0,018 0,018 3 0,2261 0,416 0,416 5 2,4214 1,601 1,601 7 4,8799 1,464 1,464 9 7,2752 2,341 2,341 12 9,5466 2,470 2,470 15 10,2348 2,449 2,449 17 8,9033 3,920 3,920 19 3,8427 5,249 5,249 21 0,7840 1,693 1,693 23 0,0089 0,021 0,021
Figura 8. Curva promedio de progesterona durante el ciclo estral

84
No solo las concentraciones de progesterona son importantes para la preñez, sino
que la demora en el incremento post ovulatorio de progesterona compromete el
desarrollo del conceptus y su habilidad para producir Interferon tau, dando como
resultado tasas de concepción reducidas (Darwash y Lamming 1998). Mann y
Lamming (1999) observaron que suplementando progesterona antes del día 6 post
IA se incrementan las tasas de concepción, Sin embargo, la suplementación de
progesterona inmediatamente después de la IA causa la ocurrencia de ciclos
cortos (Van Cleef et al 1996; Garret et al 1988; Lynch et al 1999).Se ha reportado
(Duchens et al 1995) un aumento en la velocidad de transporte del oocito en el
oviducto en novillas que han sido tratadas con progesterona durante el estro, y
esto se ha asociado a una sobre maduración del folículo ovulatorio y su
consiguiente mayor liberación de estradiol. La exposición prematura a altos niveles
de progesterona puede estar avanzando el momento en el cual la progesterona
pierde su habilidad para estimular la expresión de sus propios receptores e inhibir
la expresión de los receptores para estrógenos y oxitocina, adelantando el
momento en que debe producirse la luteolisis (Mann et al 1998). En la figura 9 se
muestran los perfiles de progesterona de animales que presentaron ciclos estrales
consecutivos

85
Figura 9. Perfiles individuales de progesterona para animales que presentaron
ciclos estrales consecutivos completos

86
3.8.5 Variación en los niveles de progesterona. En las Figuras 10 y 11 se
muestran los porcentajes de variación, en promedio e individuales,
respectivamente, de los niveles de progesterona de cada fecha de muestreo con
respecto al muestreo inmediatamente anterior. En general, los niveles de
progesterona se mantuvieron a niveles basales durante la fase folicular. Los
niveles suprabasales de progesterona en el periodo periovulatorio se han
relacionado con un excesivo desarrollo y sobre maduración de los folículos
preovulatorios en hembras repetidoras (Bage et al., 2002). Por otra parte, se
evidencio un fuerte descenso en los niveles de progesterona (49.44%) entre los
días 17 y 19 del ciclo estral, durante la luteolisis. Existe evidencia con respecto a
que un periodo prolongado entre la luteolisis y la ovulación está asociado con el
retraso en el incremento de los niveles de progesterona durante el ciclo
subsiguiente (Robinson et al., 2002).
Burke et al. (1998) encontraron que aunque la concentración de progesterona
plasmática fue mayor en vacas con condición corporal alta durante la fase luteal
respecto a vacas con baja condición, durante la fase folicular los niveles de
progesterona fueron menores en las vacas con condición alta con respecto a las
de baja condición. Estas bajas concentraciones de progesterona pueden alterar la
secreción de LH y la dinámica folicular, dando como resultado un pico
preovulatorio de LH y una ovulación más temprana. Estos resultados sugieren que
existen alteraciones en el metabolismo y el estatus endocrino asociadas a la

87
condición corporal que pueden contribuir a diferencias en la dinámica folicular
entre animales con alta y baja condición corporal.
Esta variación puede estar contribuyendo a que se dé una mejor relación de los
esteroides ováricos en cuanto al momento, tiempo y duración de su acción y de
los efectos coordinados que deben regular la expresión de receptores tanto de la
misma progesterona (bloqueo de la progesterona), como de los receptores para
estrógenos y oxitocina en estas mismas fases, los cuales modulan los pulsos
luteolíticos de PGF2α por el endometrio (Spencer et al 1995; Asselin et al 1996;
Psychoyos et al 1995; Del Vecchio et al 1996). Estas variaciones en la
concentración de progesterona son determinantes en la modulación de la
expresión y secreción de factores de crecimiento, citoquinas y proteínas, que
condicionan el medio uterino en los procesos de receptividad endometrial y con
ello de viabilidad embrionaria (Poland et al 1995; Kerbler et al 1997).
Figura 10. Variación promedio de los niveles de progesterona entre muestreos
consecutivos.

88
Figura 11. Variacion individual de los niveles de progesterona durante el ciclo estral

89
Figura 11. (Continuación)

90
Figura 11. (Continuación)

91
Figura 11. (Continuación)

92
3.8.6 Preñez temprana
Por condiciones logísticas del estudio, no todos los animales que mostraban
signos de celo fueron sometidos a monta, de manera tal que animales que no
resultaron preñados posiblemente fue debido a que no fueron montados, y no a
repeticiones o ausencias de estro.
En la figura 12 se muestran las variaciones en los niveles de progesterona durante
la preñez temprana y el ciclo estral inmediatamente anterior a la concepción. Tres
de los animales lograron la gestación en la segunda ovulación posparto, posterior
al ciclo de corta duración. La secreción de progesterona es esencial para el
establecimiento exitoso del embrión y el mantenimiento de la preñez. Según
(Diskin et al., 2006) las concentraciones de progesterona tanto en el ciclo
precedente como en el ciclo posterior a la inseminación afectan la sobrevivencia
embrionaria, evidenciando que tanto niveles insuficientes como niveles excesivos
de progesterona son detrimentales. Reportes previos encontraron que una más
rápida tasa de crecimiento en las concentraciones post-ovulatorias de
progesterona incrementan la sobrevivencia embrionaria (Ahmad et al., 1996).
Igualmente, existen estudios que observaron que bajas concentraciones de
progesterona durante los días 5-7 de preñez estuvieron asociadas con
disminución de la fertilidad (Stonge et al., 2005), así como también se reporto que
la aplicación de progesterona suplementaria mejora el desarrollo de los conceptus
en vacas de carne (Garrett et al., 1988). Se ha especulado que la relación entre el
desarrollo embrionario exitoso y los niveles de progesterona está probablemente

93
mediada por los efectos benéficos de la progesterona sobre el ambiente uterino,
así como también se ha concluido que un reconocimiento de la gestación exitoso
requiere un adecuado grado de desarrollo embrionario y un apropiado patrón de
secreción después de la ovulación (Mann y Lamming, 2001).
Figura 12. Porcentaje de variación de los niveles de progesterona entre muestreos
consecutivos durante el ciclo de preñez temprana y el ciclo estral previo.

94
Figura 12. (Continuación)
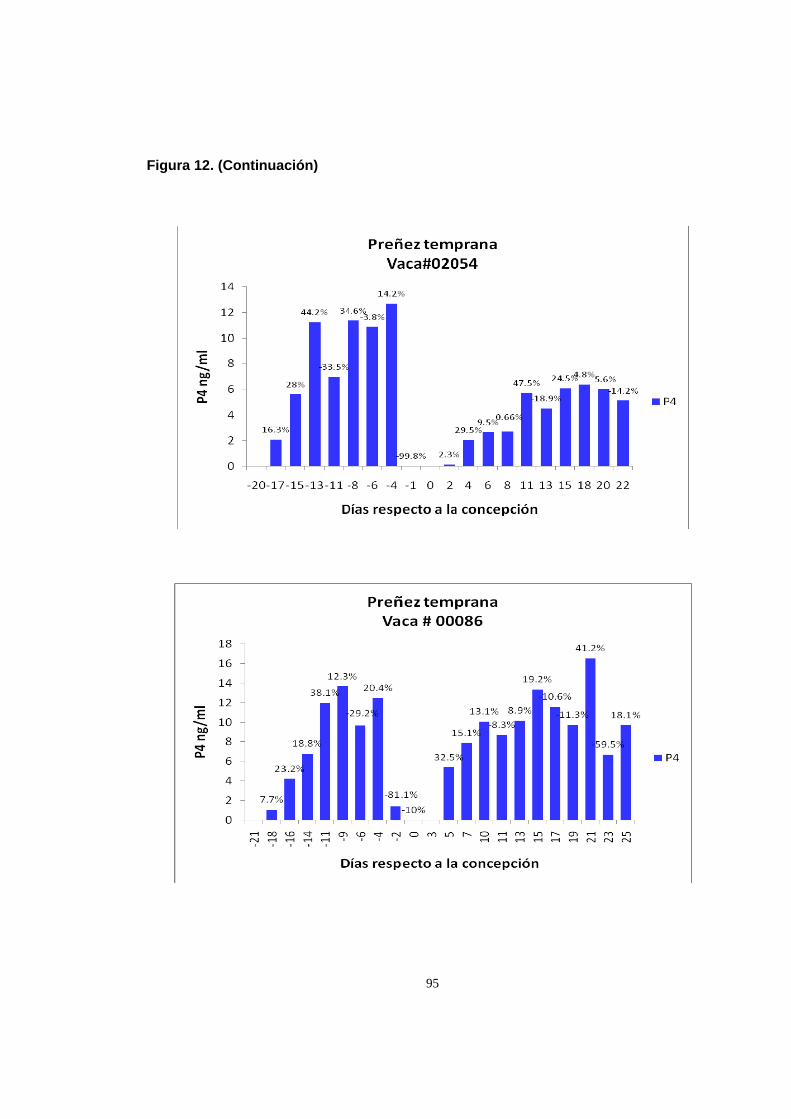
95
Figura 12. (Continuación)

96
3.9 CONCLUSIONES
El restablecimiento de la ciclicidad estral luego del parto se presento antes del dia
80 en el 61% de los animales del estudio (11/18), y para el dia 100 posparto, el
88.8% (16/18) de las vacas ya habían estabilizado sus ciclos.
Se observó un fenomeno social de agrupamiento de los celos en la medida que
transcurría el periodo posparto, esto es favorable desde la perspectiva de la
inducción a la ciclicidad y su implicación en el mejoramiento del comportamiento
de los indicadores reproductivos.
El promedio de duración del ciclo estral fue de 20.95±1.99 días, El nivel máximo
de progesterona fue en promedio de de 10. 23 ng/ml, hacia el día 15 del ciclo
estral.
Los niveles de progesterona presentaron niveles basales sostenidos durante la
fase folicular. Durante la fase luteal, Se observó un patrón consistente de
incremento de los niveles de progesterona hasta el día 12 del ciclo, un posterior
descenso leve (aparente luteolisis funcional) , seguido de un descenso fuerte
(luteolisis estructural).

97
BIBLIOGRAFIA
Ahmad N, Beam SW, Butler WR, Deaver DR, Duby RT, Elder DR, et al. Relationship of fertility to
pattern of ovarian follicular development and associated hormonal profiles in dairy cows and
heifers. J Anim Sci 1996;74:1943–52.
Asselin E, Goff AK, Bergeron H and Fortier M. Influence of sex steroids on the production of
prostaglandins F2 alfa, E2 and response to oxytocin in cultured epithelial and stromal cells of the
bovine endometrium. Biology of Reproduction. 1996: 54: 371-379
Báez GM, Grajales HA y Pérez JE Caracterización del ciclo estral mediante perfiles de esteroides
(progesterona, 17 β-estradiol) en la raza Costeño con cuernos (Bos taurus) en el trópico
Colombiano. Livestock Research for Rural Development 2007. Volume 19, Article #132.
http://www.cipav.org.co/lrrd/lrrd19/9/baez19132.htm
Bage R, Gustafsson H, Larsson B, Forsberg M, Rodriguez-Martinez H. Repeat breeding in dairy
heifers: Follicular dynamics and estrous cycle characteristics in relation with sexual hormone
patterns. Theriogenology 2002, 57:2257-2269.
Burke JM, Hampton JH, Staples CR, Thatcher WW. Body condition influences maintenance of a
persistent first wave dominant follicle in dairy cattle. Theriogenology 1998. 49:751-760
Cardozo C J Relación entre factores climáticos y algunas características del ciclo estral en novillas
Holstein de la sabana de Bogotá. Tesis de Maestría. 1993 FMVZ. Universidad Nacional de
Colombia.
Castro A y Gómez R Duración del ciclo estral, del estro y tiempo de ovulación en ganado de leche
en climas cálidos de Colombia. Revista ICA. 1979: 3: 171-176
Darwash AO and Lamming GE. The importance of milk progesterone concentrations during early
pregnancy in the cow. Journal of Animal Breeding 1998. 2, 41-43.
Del Vecchio RP, Sutherland WD and Sasser RG Bovine luteal cell production in vitro of
prostaglandin E2, oxytocin and progesterone in response to pregnancy-specific protein B and
prostaglandin F2 alfa. Journal of Reproduction and Fertility 1996. 107: 131-136
Diskin MG, Murphy JJ, Sreenan JM. Embryo survival in dairy cows managed under pastoral
conditions. Anim Reprod Sci 2006 96:297–311.
Duchens M, Forsberg M, Gustafsson H, Edqvist LE, y Rodríguez-Martínez. H. Reproductive
performance of heifers induced to oestrous asynchrony by suprabasal plasma progesterone
levels. Anim. Reprod. Sci. 1995. 39: 171 – 182:
Galina CS, Maquivar M. Presentación del estro y su sincronía con el momento de ovulación.
Congreso internacional de reproducción bovina. Intervet. Bogotá. 2005.

98
Garrett JE, Geisert RD, Zavy MT and Morgan GL. Evidence for maternal regulation of early
conceptus growth and development in beef cattle. Journal of Reproduction and Fertility 1988.
84, 437-446.
Góngora A y Hernández A Comportamiento sexual, duración del estro y del ciclo estral en novillas
criollas sanmartineras y brahman del piedemonte llanero colombiano. Livestock Research for
Rural Development 2006:. Volume 18, Article #1.
Grajales, H.A. Comportamiento reproductivo de grupos raciales bovinos en el trópico calido-
humedo Colombiano: Pubertad, ciclo estral, preñez temprana, posparto. Niveles de hormonas
esteroides y su relación con la eficiencia reproductiva. 2001. Tesis PhD. Facultad de Medicina
Veterinaria y de Zootecnia. Universidad Nacional de Colombia. Bogota D.C.
Grajales H, Hernández A y Prieto E Determinación de parámetros reproductivos basado en los
niveles de progesterona en novillas doble propósito en el trópico Colombiano. Livestock
Research for Rural Development. 2006 Volume 18, Article #144.
http://www.cipav.org.co/lrrd/lrrd18/10/graj18144.htm
Gustaffson H 1999 CL function and early embryonic development - some introductory notes.
Reproduction of Domestic Animals: 34: 201-202
Henao G, Olivera-Ángel M, Maldonado-Estrada JG. Follicular dynamics during postpartum anestrus
and the first estrous cycle in suckled or non-suckled Brahman (Bos indicus) cows. Animal
Reproduction Science 2000, 63 127–136
Hommeida A, Nakao T, Kubota H. Onset and duration of luteal activity postpartum and their effect
on first insemination conception rate in lactating dairy cows.J Vet Med Sci. 2005
Oct;67(10):1031-5.
Inskeep EK. Preovulatory, postovulatory, and postmaternal recognition effects of concentrations of
progesterone on embryonic survival in the cow J. Anim. Sci. 2004. 82(E. Suppl.):E24–E39
Kerbler TL, Buhr MM, Jordan LT, Leslie KE and Walton JS Relationship between maternal plasma
progesterone concentration and interferon-tau synthesis by the conceptus in cattle.
Theriogenology1997 47: 703-714
Kilgour R, Skarsholt BH, Smith JF, Bremner KJ, Morrison MCL. Observations on the behaviour and
factors influencing the sexually active group in cattle. Proc NZ Soc Anim Prod 1977;128–35.
Kinder JE, Kojima FN, Bergfeld EGM, Wehrman ME, and Fike KE. Progestin and estrogen
regulation of pulsatile LH release and development of persistent ovarian follicles in cattle. J.
Anim. Sci. 1996 74:1424–1440.
Lamothe C, Montiel F, Fredricksson G, Galina, CS. Reproductive performance of Zebu cattle in
Mexico. Influence of season and social interaction on the timing of expressed estrous. Tropical
agriculture1995 72: 1-5.

99
Lynch PR, Macmillan KL and Taufa VK Treating cattle with progesterone as well as GnRH
analogue affects oestrous cycle length and fertility. Animal Reproduction Science. 1999 56, 189-
200.
Mann GE, Lamming GE. Relationship between maternal endocrine environment, early embryo
evelopment and inhibition of the luteolytic mechanism in cows. Reproduction 2001. 121: 175–80.
Mann GE, Lamming GE. The influence of progesterone during early pregnancy in cattle. Reprod.
Domest. Anim. 1999 34, 269–274.
Mann. GE, Lamming GE y Payne. JH. Role of early luteal phase progesterone in control of the
timing of the luteolytic signal in cows. J. Reprod. Fertil. 1998. 113: 47 – 51
Maquivar M, Galina CS, Orihuela A. Cows treated with Synchromate B may cluster their sexual
behavior independent of follicular growth at the time of oestrus. Physiology & Behavior 2002 76
199– 203
Orihuela A, Galina CS, Escobar J, Riquelme E. Oestrous behaviour following prostaglandin f2a
injection in Zebu cattle under continuous observation. Theriogenology 1983;19:795– 809.
Poland ML, Simon C, Frances A, Lee BY and Prichard LE Role of embrionic factors in human
implantation. Human Reproduction. 1995: 10: Supl. 2: 22-28
Prieto ME Edad y peso al inicio de la pubertad y seguimiento de los tres primeros ciclos estrales en
novillas Holstein. Tesis de maestría 1993 FMVZ. Universidad Nacional de Colombia.
Psychoyos A, Nikas G and Gravanis The role of prostaglandins in blastocyst implantation. Human
Reproduction 199510: 2: 30
Putney DJ, Mullins S, Thatcher WW, Drost M, Gross TS. Embryonic development in superovulated
dairy cattle exposed to elevated ambient temperature between the onset of estrus and
insemination. Anim Reprod Sci 1989;19:37–51.
Randel RD LH and ovulation in Brahman, Brahman × Hereford and Hereford heifers. Journal of
Animal Science 1989. 43, 300 (abstract).
Robinson JJ Nutrition in the reproduction of farm animals. Nutr. Res. Rev. 1990 3, 267–268.
Robinson RS, Hammond AJ, Hunter MG, Mann GE The induction of a delayed post-ovulatory
progesterone rise in dairy cows: a novel model. Domestic Animal Endocrinology 2005 28 285–
295
Spencer TE, Becker WC, George P, Mirando MA, Ogle TF and Bazer FW Ovine interferon-tau
inhibits estrogen receptor up-regulation and estrogen-induced luteolysis in cyclic ewes.
Endocrinology 1995 136: 11: 4932-4944
Stronge AJH, Sreenan JM, Diskin MG, Mee JF, Kenny DA, Morris DG. Post-insemination milk
progesterone concentration and embryo survival in dairy cows. Theriogenology 2005;64: 1212–
24.

100
Vincent DL, Meredith S and Inskeep EK. Advancement of uterine secretion of prostaglandin E2 by
treatment with progesterone and transfer of asynchronous embryos. Endocrinology 1986
119:527–529.
Westone JS, Ulberg LC. Responses of dairy cows to the behavior of treated herdmates during the
postpartum period. J Dairy Sci 1976; 59:1985– 8.
Wherman ME, Kojima FN, Sanchez T, Mariscal DV and Kinder JE Incidence of precocious puberty
in developing beef heifers. Journal of Animal Science 1996 74: 2462-2467

101
4. CONCLUSIONES GENERALES
Este trabajo se constituye en el primer estudio exploratorio sobre el
comportamiento hormonal durante el periodo de posparto de vacas criollas
Sanmartinero, abarcando la reactivación ovárica, la ciclicidad estral y el
establecimiento de la preñez, y será un pilar fundamental para futuras
investigaciones sobre la fisiología reproductiva de este tipo de razas.
Se generó información referencial acerca de los niveles de progesterona y
estradiol durante el periodo de reactivación ovárica posparto, confirmando la
presencia de ciclos de corta duración previos al restablecimiento de la ciclicidad,
así como un incremento gradual de los niveles de secreción de estradiol desde el
parto hasta la primera ovulación. Igualmente, se relacionó el momento de minima
condicion corporal con la posterior reactivacion ovarica.
Se describieron los fenomenos relacionados con la dinamica de comportamiento
de la hormona progesterona durante las diferentes etapas de los ciclos estrales
posteriores a la reactivacion posparto, incluyendo la caracterizacion de los niveles
basales sostenidos durante la fase folicular, asi como los niveles de incremento y
caida de los niveles de progesterona durante la fase luteal. Se describio el
fenómeno de agrupación de las manifestaciones de celo dentro del grupo de
animales en un periodo de tiempo determinado.

102
5. RECOMENDACIONES
Se sugiere para futuras investigaciones continuar ampliando el marco referencial
de patrones fisiológicos que afectan las diferentes etapas reproductivas,
profundizando en las relaciones hormonales y su dinámica de cambio a lo largo de
estas, involucrando factores adicionales como la función de los receptores de
estas hormonas y sus posibles interacciones.
Por otra parte, es necesario diseñar estudios que permitan comparar diferentes
genotipos dentro de un mismo ambiente, con el fin de corroborar las hipótesis
acerca de los posibles mecanismos de adaptación que pueden favorecer o no a
una determinada raza en particular; esto con el fin de determinar con certeza los
factores comunes que determinan la eficiencia reproductiva y que pueden llegar a
ser una base de selección de individuos adaptados que van a permitir mejorar el
comportamiento de los indicadores reproductivos y la productividad del hato
bovino nacional.


















