
Sexualidad humana y evolución
22
Los primeros primates, salvo los depredadores del suelo, tuvieron mucho éxito en los árboles. Muchos de ellos aumentaron de tamaño y se especializaron cada vez más a medida que descubrían modos diferentes de ganarse la vida en las alturas. Algunos se acomodaron a una dieta donde predominaban los frutos y las bayas. Otros acabaron comiendo solamente hojas. Desarrollaron sistemas diferentes para moverse entre las ramas. Algunos de los pequeños primates, como los tarseros y los gálagos continuaron saltando y agarrándose a las ramas; tenían miem- bros delanteros más bien cortos y miembros traseros largos, y renuncia- ron totalmente a correr. Un gran grupo se mantuvo cuadrúpedo a pesar de tener una mano de primate, e incluso un pie de primate con una planta plana y dedos prensiles. Es el grupo de los monos. Los monos prefirieron caminar sobre las ramas en lugar de agarrarse a ellas o colgar de ellas. Sus espinazos eran bastante largos. Conservaron una cierta capacidad para correr porque muchos de ellos estaban regresando al suelo y necesitaban disponer de una velocidad considerable para poder refugiarse en un árbol cuando corrían peligro. Los monos son distintos de los antropoides porque conservaron el cuadrupedismo. Los antropoides están hechos más bien para balancearse. En vez de caminar a lo largo de las ramas se cuelgan de ellas, se sientan encima, se yerguen sobre ellas, suben y bajan mano sobre mano. Tienen espi- nazos más cortos que los monos —tres o cuatro vértebras lumbares en lugar de siete— y en consecuencia no son en absoluto tan buenos corre- dores. De hecho algunos apenas pueden correr. Hay cinco tipos de simios antropoides en el mundo actual. Dos de ellos —los gibones y los siamang— son auténticos balanceadores. Se mueven por braquiación, oscilando como péndulos de rama en rama. Sus brazos son extraordinariamente largos, sus manos y sus dedos son alargados y especializados. Sus cuerpos son cortos y ligeros, sus piernas han disminuido de longitud. Gracias a este diseño —un mínimo de peso en la parte inferior del péndulo— pueden moverse con notable rapidez a través de los árboles, balanceándose de una rama a otra con una segu- ridad y una suavidad que hay que ver para poderla apreciar, salvando a menudo intervalos de más de tres metros. Los gibones cuando llegan al suelo, que es casi nunca, caminan erguidos anadeando sobre sus pier- nas cortas y débiles, manteniendo separados sus largos brazos a cada lado para tener mayor equilibrio. Un gibón paseándose por el suelo re- cuerda a un equilibrista sobre la cuerda floja. El orangután es tan arbóreo como el gibón, pero de un modo total-
description
Fragmento del libro El primer antepasado del hombre, de Donald Johanson y Maitland Edey. Ed. Plantea. Madrid, 1987, pp. 280-301
Transcript of Sexualidad humana y evolución

Los primeros primates, salvo los depredadores del suelo, tuvieron mucho éxito en los árboles. Muchos de ellos aumentaron de tamaño y se especializaron cada vez más a medida que descubrían modos diferentes de ganarse la vida en las alturas. Algunos se acomodaron a una dieta donde predominaban los frutos y las bayas. Otros acabaron comiendo solamente hojas. Desarrollaron sistemas diferentes para moverse entre las ramas. Algunos de los pequeños primates, como los tarseros y los gálagos continuaron saltando y agarrándose a las ramas; tenían miembros delanteros más bien cortos y miembros traseros largos, y renunciaron totalmente a correr. Un gran grupo se mantuvo cuadrúpedo a pesar de tener una mano de primate, e incluso un pie de primate con una planta plana y dedos prensiles. Es el grupo de los monos. Los monos prefirieron caminar sobre las ramas en lugar de agarrarse a ellas o colgar de ellas. Sus espinazos eran bastante largos. Conservaron una cierta capacidad para correr porque muchos de ellos estaban regresando al suelo y necesitaban disponer de una velocidad considerable para poder refugiarse en un árbol cuando corrían peligro. Los monos son distintos de los antropoides porque conservaron el cuadrupedismo.
Los antropoides están hechos más bien para balancearse. En vez de caminar a lo largo de las ramas se cuelgan de ellas, se sientan encima, se yerguen sobre ellas, suben y bajan mano sobre mano. Tienen espinazos más cortos que los monos —tres o cuatro vértebras lumbares en lugar de siete— y en consecuencia no son en absoluto tan buenos corredores. De hecho algunos apenas pueden correr.
Hay cinco tipos de simios antropoides en el mundo actual. Dos de ellos —los gibones y los siamang— son auténticos balanceadores. Se mueven por braquiación, oscilando como péndulos de rama en rama. Sus brazos son extraordinariamente largos, sus manos y sus dedos son alargados y especializados. Sus cuerpos son cortos y ligeros, sus piernas han disminuido de longitud. Gracias a este diseño —un mínimo de peso en la parte inferior del péndulo— pueden moverse con notable rapidez a través de los árboles, balanceándose de una rama a otra con una seguridad y una suavidad que hay que ver para poderla apreciar, salvando a menudo intervalos de más de tres metros. Los gibones cuando llegan al suelo, que es casi nunca, caminan erguidos anadeando sobre sus piernas cortas y débiles, manteniendo separados sus largos brazos a cada lado para tener mayor equilibrio. Un gibón paseándose por el suelo recuerda a un equilibrista sobre la cuerda floja.
El orangután es tan arbóreo como el gibón, pero de un modo total-

mente distinto. La articulación de la cadera es casi una articulación universal. Puede extender la pierna hacia abajo, hacia atrás, hacia adelante, en línea recta hacia el lado y en línea casi recta hacia arriba. Todas sus extremidades sirven para agarrar. Es en realidad un animal de cuatro brazos y cuatro manos. Extiende sus miembros entre las ramas como una enorme araña de color anaranjado-marrón, se agarra con cualquier pie o mano disponible y alarga la que sea para coger comida, principalmente fruta. En las raras ocasiones en que baja al suelo avanza a cuatro patas, de modo lento y deliberado como un anciano andando sobre dos bastones (así lo describió Sarel Eimerl); se apoya para ello sobre las plantas de los pies y los nudillos de las manos, aunque muy a menudo cierra las manos formando puños y camina sobre ellos. En consecuencia puede decirse que el orangután camina parcialmente sobre sus nudillos.
El gorila lo hace de modo total. Es el único antropoide que ha vuelto casi completamente al suelo. Allí encuentra lodo su alimento: materia vegetal basta, raíces, retoños de bambú, bayas. Ha renunciado a la carrera en favor de un gran tamaño y una fuerza enorme. Los gorilas para poder sobrevivir necesitan ser grandes y fuertes —un adulto macho pesa hasta doscientos kilos— y tener un aspecto terrible. Los gorilas jóvenes cuelgan de los árboles y juegan allí, pero los adultos son demasiado grandes y letárgicos para hacerlo. A veces se sientan en grupos sobre ramas muy bajas y muy gruesas, pero en realidad prefieren el suelo. Son muy sedentarios, se quedan mucho tiempo en los mismos lugares y casi nunca corren.
El chimpancé, que también camina sobre sus nudillos es el antropoide más adaptable. Es una especie de modelo universal que refleja un sistema de vida abierto a todo. Come mucha fruta, especialmente higos y le gusta encaramarse y buscarlos en la cubierta superior de los árboles. Sus brazos son largos y le permiten subir y bajar de todos los árboles, excepto de los más grandes. Sus piernas son cortas, pero de longitud suficiente para correr a una velocidad sorprendente. Es poco probable que si un hombre persigue a un chimpancé consiga darle alcance. El chimpancé, dotado de estas cualidades, se pasa gran parte del tiempo en el suelo, alimentándose de gusanos, termitas, bayas, insectos, vastagos y raíces, cuando los higos no están maduros. También caza de modo cooperativo, aunque a pequeña escala. En ocasiones, un grupo de chimpancés machos arrinconan a un mandril joven o a otro mono en un árbol aislado, lo cogen y se lo comen.
Todos los antropoides, a pesar de sus estilos de vida radicalmente distintos y de sus adaptaciones corporales tan divergentes, son en potencia animales erguidos. Todos viven en los bosques y se mantienen de un modo que no exige correr. En los inicios del mioceno, hace unos veinte millones de años, en un período en el que un einturón inmenso de bosques se extendía por toda la tierra en las regiones tropicales y subtropicales, los antropoides se multiplicaron y tuvieron éxito. En cambio los monos mucho menos. Actualmente la situación se ha invertido. Sin duda la decadencia de los bosques tropicales a fines del mioceno debida a cambios climáticos tuvo alguna relación con la decadencia de los antropoides, pero influyeron en ella otras fuerzas más sutiles. Lovejoy ha intentado analizarlas, y cree haber encontrado una explicación del origen del bipedismo en la interrelación de esta,s fuerzas.

Los primates presentan toda una variedad de modos de locomoción. El tarsero (primer plano) es un tipo primitivo, nocturno, exclusivamente arbóreo que avanza saltando y agarrándose. El gibón (arriba a la izquierda) también arbóreo, es un braquiante, que se balancea de un miembro a otro como un péndulo. El orangután (centro izquierda) es un cuadrumano, capaz de agarrarse tan bien con las manos como con los pies. El gorila (abajo a la izquierda) es un auténtico caminante sobre nudillos. Es un animal semierguido que puede ponerse erguido con facilidad y que a menudo lo hace. El mono (arriba a la derecha) es un auténtico cuadrúpedo. Carece del potencial para la postura erguida que tienen los antropoides en diversos grados. En cambio ha conservado la opción de correr rápidamente, lo cual consigue con piernas largas, un espinazo largo y una mano que aplica plana sobre el suelo en cada paso. Sólo los homínidos se han aventurado a convertise en bípedos completos.
—Menos mal que volvemos al bipedismo —murmuró alguien—. Me estaba aburriendo tanto antropoide.
—Aburridos o no, teníais que entender sus orígenes: eran depredadores que se subieron a los árboles y consiguieron ser arbóreos y al mismo tiempo depredadores mediante el sistema de convertirse en animales balanceadores semierguidos con manos, espinazos cortos, visión binocular y cerebros grandes. No se había visto nunca nada semejante sobre la tierra. Cuando algunos de los animales más grandes, como los gorilas y los chimpancés, regresaron al suelo estuvieron capacitados para
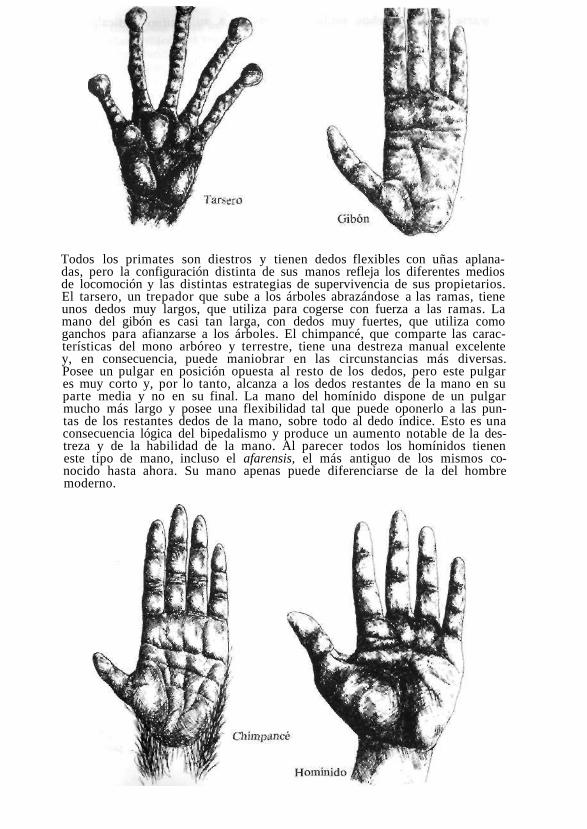
Todos los primates son diestros y tienen dedos flexibles con uñas aplanadas, pero la configuración distinta de sus manos refleja los diferentes medios de locomoción y las distintas estrategias de supervivencia de sus propietarios. El tarsero, un trepador que sube a los árboles abrazándose a las ramas, tiene unos dedos muy largos, que utiliza para cogerse con fuerza a las ramas. La mano del gibón es casi tan larga, con dedos muy fuertes, que utiliza como ganchos para afianzarse a los árboles. El chimpancé, que comparte las características del mono arbóreo y terrestre, tiene una destreza manual excelente y, en consecuencia, puede maniobrar en las circunstancias más diversas. Posee un pulgar en posición opuesta al resto de los dedos, pero este pulgar es muy corto y, por lo tanto, alcanza a los dedos restantes de la mano en su parte media y no en su final. La mano del homínido dispone de un pulgar mucho más largo y posee una flexibilidad tal que puede oponerlo a las puntas de los restantes dedos de la mano, sobre todo al dedo índice. Esto es una consecuencia lógica del bipedalismo y produce un aumento notable de la destreza y de la habilidad de la mano. Al parecer todos los homínidos tienen este tipo de mano, incluso el afarensis, el más antiguo de los mismos conocido hasta ahora. Su mano apenas puede diferenciarse de la del hombre moderno.

labrarse nuevos nichos ecológicos gracias a su equipo radicalmente nuevo.
—Pero no se convirtieron en bípedos. —Algunos sí. Los homínidos: nuestros antepasados. —Ya lo sabemos, pero ¿por qué? —¿Queréis que hablemos de sexo? —Prefiero hablar de bipedismo. —De acuerdo, hablaremos de sexo.
Hablar de sexo equivale sin duda a remontarse de nuevo a los orígenes. Recuerda la astuta definición formulada hace casi un siglo y atribuida a Herbert Spencer: «Una gallina es el sistema que utiliza un huevo para hacer otro huevo.» Dicho de otro modo: consideremos la supervivencia de la especie no desde el punto de vista del individuo, sino desde el punto de vista humilde pero esencial del gen. Consideremos que lo importante a largo plazo es la preservación de los genes, la fabricación de otro individuo igual a uno mismo.
Lovejoy opina en esto como Spencer. Cualquier individuo viviente de cualquier especie no es más que una envoltura protectora que contiene las semillas de la propagación, empaquetadas hábilmente para la probabilidad de que viva el tiempo suficiente para que la producción de individuos de esta especie sea máxima. Para ello tiene que comer. Para comer, si es un mamífero, necesita moverse. Por lo tanto, la adaptación locomotriz de un animal no puede entenderse totalmente si antes no se entiende su estrategia sexual.
Un animal puede funcionar sexualmente de ój°s maneras fundamentalmente distintas. Puede producir una gran cantidad de huevos, invir-tiendo muy poca energía en cada huevo. O bien puede producir muy pocos huevos, pero invirtiendo en ellos mucha energía. La ciencia llama a estos sistemas la estrategia «r» y la estrategia «K», respectivamente.
Un ejemplo extremo de «r» sería el de una ostra, que puede producir hasta quinientos millones de huevos al año. Los ejemplos más extremos de «K» son los grandes antropoides —el gorila, el chimpancé y el orangután—, que producen sólo un hijo cada cinco o seis años. Entre estos extremos hay una confusa mezcla de «r» y dé «K» entre animales
Las estrategias reproductoras en el mundo animal recorren toda la gama desde una «r» extrema, la estrategia basada en una producción máxima de huevos sin cuidados paternos, hasta la «K» extrema, en la que la importancia

que han intentado todas las combinaciones posibles. Tanto «r» como «K» dan resultado mientras no se lleven al límite. Imaginemos que una ostra dobla su producción de huevos y llega a mil millones; si la energía a invertir por huevo es fija, la energía disponible por huevo quedará reducida a la mitad. Esto puede disminuir la capacidad reproductora de todos los huevos, de modo que si bien habrá más huevos, llegará a la madurez un número de ellos inferior incluso al de los muy escasos huevos que ahora sobreviven y llegan a la madurez. Es evidente que una «r» pura es una imposibilidad, y que ahora la están explotando hasta su límite seres tan humildes como las ostras. Una «r» extrema puede considerarse como un sistema de reproducción ineficaz. ¿Qué sentido tiene producir quinientos millones de huevos si sólo llegan a la madurez media docena?
El sistema tiene sentido para la ostra, porque no tiene otra opción. Sus órganos de los sentidos son rudimentarios, carece de cerebro, no puede moverse. Es incapaz de cuidar de sus huevos; ni siquiera puede dejarlos en un lugar seguro. Lo único que puede hacer es verterlos al exterior, y por ello hace máximas sus posibilidades de supervivencia como especie vertiendo la cantidad máxima que puede alcanzar razonablemente.
Aunque «r» da resultado con las ostras (todavía están entre nosotros) es un sistema claramente derrochador. Cuando evolucionaron animales de desarrollo superior, se dispuso de otras opciones más frugales. Cuando entraron en escena seres con espinazos y cerebros, se hicieron posibles unos primeros y rudimentarios cuidados paternos. Algunos peces construyen nidos para sus huevos. Otros los guardan en sus bocas hasta que el animal sale del huevo y puede nadar por sí solo. Algunos reptiles lo hacen mejor todavía. El caimán construye un gran nido en forma de montículo para sus huevos, los guarda allí durante la incubación y cuando los hijos salen del cascarón y empiezan a chillar los oye y les ayuda a salir abriendo el nido. Pero no hay muchos reptiles tan atentos, y ninguno protege o enseña a sus hijos cuando crecen. Esto quedó para los mamíferos más inteligentes y más orientados a la «K», que sucedieron a los reptiles.
Nadie sabe cómo se comportaban los dinosaurios cuando eran padres,
se centra casi exclusivamente en el cuidado de los hijos y la tasa de natalidad queda reducida a un mínimo.

aunque algunos de ellos según demuestran las nidadas de huevos fósiles encontradas en el desierto de Gobi producían huevos con mucha parsimonia; algunos «nidos» sólo contienen una docena o dos de huevos. Por lo tanto, si los dinosaurios no eran mejores padres que la mayoría de los reptiles, salieron perdiendo de dos maneras: tendían a la *r» porque probablemente eran incapaces de cuidar intensivamente de sus hijos, y tendían a la «K» porque ponían un número reducido de huevos. Esto podría explicar su extinción en un mundo que empezaba a llenarse con mamíferos que tendían a la «K» y que eran mejores padres.
«No hay que recurrir a manchas solares, catástrofes climáticas o cualquier explicación misteriosa para justificar la desaparición de los dinosaurios —dijo Lovejoy—. Las cosas les fueron bien mientras el mundo estuvo a su disposición, mientras no hubo una estrategia reproductora mejor. Duraron más de cien millones de años; también los hombres tendrían que durar. Pero cuando se produjo una adaptación radicalmente nueva, cuando los dinosaurios tuvieron que enfrentarse con animales que podían reproducirse con éxito a una velocidad tres o cuatro veces superior a la suya, sonó su hora.»
Es evidente que «K» es mucho más eficiente que «r», pero también tiene sus límites. Los accidentes, la depredación, la escasez estacional de alimentos, la enfermedad: todo esto se cobra sus víctimas entre los animales. Perder un hijo por uno de estos azares después de una inversión de cinco o de seis años supone un coste terrible en comparación con la pérdida de un huevo por una ostra. Dos o tres accidentes seguidos de este tipo pueden suponer la extinción de un surtido concreto de genes en posesión de una determinada madre antropoide, porque quizá hayan pasado ya sus años de fertilidad. Un ritmo reproductivo lento es tan peligroso para una población entera de antropoides —para una reserva entera de genes— como lo es para un solo individuo. Una serie de desastres puede destruirla para siempre.
De hecho el nivel de «K» que se observa en los antropoides modernos es ya peligrosamente elevado. Todos ellos existen actualmente en cantidades peligrosamente reducidas y sólo en los medios ambientes más favorables. Su gran vulnerabilidad a la extinción ha quedado oscurecida por el papel que ha jugado el hombre al acelerar su desaparición en épocas recientes. Se nos dice constantemente que estamos exterminando a los simios antropoides. Es cierto. Pero esto es sólo la traca final de lo que los antropoides han estado haciendo consigo mismos desde hace unos millones de años. Si los dejáramos a su aire probablemente también se extinguirían. En cambio los monos han prosperado y se han multiplicado. La diferencia en los resultados es tan espectacular que vale la pena comparar los dos casos y ver si existe alguna diferencia básica que pueda explicarlo.
Resulta que hay dos diferencias de este tipo. Los monos, como se ha dicho, son cuadrúpedos. Los antropoides son bípedos en potencia. Los monos tienden menos a la «K» que los antropoides. Tienen hijos cada dos años y no cada cinco o seis. No son tan inteligentes como los antropoides, ni mucho menos, ni tan buenos padres, pero lo son en grado suficiente para compensar esta deficiencia con su superior ritmo reproductor. El éxito moderno de los monos de mayor tendencia «r» hace pensar fundadamente que el antropoide, de tendencia «K» más acentuada, ha llevado demasiado lejos esta estrategia.

¿Por qué se hace algo que no resulta? Esta pregunta se ha planteado ya en otra ocasión. Primero dijimos: ¿Por qué hacerse bípedo, si es ineficaz? Ahora decimos: ¿Por qué tender tanto a la «K», si es peligroso? Las dos preguntas están relacionadas y hay que considerarlas conjuntamente.
En primer lugar hay que dejar bien sentado, según la anterior discusión de la evolución de los primates, que los simios antropoídes que viven en los bosques y pasan su tiempo escalando árboles y balanceándose en ellos o lo pasan bajo los árboles sentados y paseándose, no son ineficaces. No necesitan realmente ser buenos corredores. Los espinazos cortos y las posturas semierguidas que desarrollaron anteriormente en los árboles no les perjudicaron en nada cuando algunos de ellos bajaron al suelo, siempre que los bosques fueran extensos y produjeran el tipo de alimentos que consumían.
La lección que puede extraerse de todo esto es que el cuadrupedis-mo, al parecer ventajoso, puede dejarse de lado —y de hecho se deja de lado— si se dispone de otras adaptaciones más útiles. Repitamos lo que Lovejoy dijo al empezar este capítulo: la capacidad de correr bien parece tan obviamente eficiente que su abandono podría considerarse caprichoso y estúpido. Pero si se estudia la estrategia total de supervivencia del animal, esta ventaja desaparece. Esto hace posible la aparición de antropoides potencialmente erguidos sobre el suelo en los bosques del mioceno. Si queremos entender por qué motivo algunos de estos antropoides llegaron hasta el final de esta evolución tenemos que estudiar otras necesidades y otras estrategias.
El siguiente factor a considerar es que durante todo el cenozoico, desde hace unos setenta millones de años hasta el mismo mioceno, hubo entre los primates una tendencia cada vez mayor hacia la «K» en su estrategia reproductora. Esta tendencia continuó acentuándose porque daba resultado. A medida que las madres de los antropoides evolucionaban y podían ofrecer cuidados paternos mejores, el porcentaje de supervivencia de sus hijos en relación al de los hijos de madres menos solícitas tenía que aumentar.
Hay que examinar en este punto la mecánica de un complejo circuito de realimentación, dentro del cual varios elementos interaccionan reforzándose mutuamente. Si los cuidados paternos son beneficiosos serán un elemento seleccionado, porque aumentará la probabilidad de que las madres mejores criarán mejor a sus hijos, y de este modo se intensificará cualquier tendencia genética preexistente en la población hacia la mejora de las madres. Pero unos mayores cuidados paternos exigen también otras cosas. Exigen un coeficiente de inteligencia mayor en la madre, que no podrá aumentar sus cuidados si no está capacitada inte-lectualmente para hacerlo. Esto supone un desarrollo cerebral, no sólo de la madre sino también de la hija que algún día será madre. Dar a luz un niño de cerebro grande exige mucho oxígeno y el paso de una carga considerable de energía a través de la placenta, es decir una gran inversión por parte de la madre. Esto se debe a que un hijo de cerebro grande no puede desarrollarse fuera del útero. Y por varios motivos: uno de ellos es que el sistema neurológico ha de ser bastante adulto en el momento del parto. El niño no puede aprender si no tiene de entrada un buen cerebro; si nace con un cerebro poco desarrollado, le será imposible luego ponerse a tono. Por lo tanto la madre ha de asumir gran

parte de este desarrollo dentro de su propio cuerpo. Pero su capacidad para transferir energía a un feto es siempre limitada (como también lo es la capacidad de una ostra para invertir en huevos), y el resultado de tener un cerebro más grande es tener menos hijos.
Expresemos lo dicho a la inversa: si uno ha de tener menos hijos, es mejor tener un cerebro grande y poder cuidar de ellos. Aquí actúa el principio de realimentación. Cada tendencia actúa junto con las demás, depende de ellas y las refuerza.
En el caso de la evolución de los primates, la realimentación no es un simple estímulo A-B, con un movimiento de vaivén entre dos polos. Es un circuito multipolar y circular y no con dos, sino con muchos rasgos que se refuerzan mutuamente. Por ejemplo, si un hijo ha de tener un cerebro grande, hay que darle tiempo para que utilice este cerebro antes de que tenga que enfrentarse por sí solo con el mundo. Esto supone una niñez prolongada. La mejor manera de aprender en la niñez es jugando. Esto supone tener compañeros de juego, lo cual a su vez supone un sistema de grupos sociales que proporcionan estos compañeros. Pero para poder vivir en un grupo así hay que aprender un comportamiento social aceptable. Este comportamiento sólo puede aprenderse si se es inteligente. Por lo tanto el comportamiento social queda relacionado en definitiva con el coeficiente de inteligencia (un circuito de realimentación), con una niñez prolongada (otro circuito), y finalmente con la inversión de energía y con el sistema de cuidados paternos que proporcionan un cerebro capaz de tener este coeficiente de inteligencia, lo cual cierra todo el circuito de realimentación.
Todas las partes de este sistema de realimentación están interconec-tadas. Por ejemplo, si uno vive en un grupo, el tiempo dedicado a buscar comida, a vigilar los depredadores y a encontrar pareja puede reducirse por el hecho mismo de estar viviendo en un grupo. En consecuencia queda más tiempo para el cuidado paterno (un circuito), el juego (otro) y la actividad social (otro), todo lo cual estimula la inteligencia (otro) y al final da por resultado tener menos hijos (otro circuito más). El circuito completo muestra todos los polos conectados con los restantes polos.
«Consideremos esto desde otro punto de vista —dijo Lovejoy—. Imaginemos el circuito de realimentación sin uno de sus componentes esenciales y veamos qué pasa. Imaginemos un bebé de chimpancé criado en una incubadora de laboratorio sin que le sea posible aprender jugando con sus amigos. Hagámosle pasar allí unos cuantos años críticos y luego soltémosle para que participe en la sociedad normal de los chimpancés. Le será imposible participar, no sabrá qué hacer, cómo comportarse. No podrá saltar ni encaramarse a los árboles tan bien como los demás. No sabrá cómo pelear ni cómo evitar las peleas, cómo respetar a sus mayores. Un gorila joven criado en el aislamiento no sabe ni aparejarse. Hay que pasarle películas.»
Por lo tanto, continuó diciendo Lovejoy, hay unas cuantas razones muy convincentes que explican la aparición de «K». Esta tendencia coincidió con una mejora y una especialización crecientes de los vertebrados a través del tiempo. Las ranas son más listas que las ostras. Los caimanes más listos que las ranas. Los conejos más listos que los caimanes. Los monos más listos que los conejos. Los antropoides más listos que los monos. Más cerebro, menos huevos, más «K».

«No llegaré al extremo de afirmar que "K" produce inteligencia, pero no hay duda que los dos factores están relacionados. Puesto que la inteligencia es lógicamente adaptativa, no cuesta mucho ver que si las circunstancias son adecuadas se intensificará. Al parecer estas circunstancias fueron adecuadas para algunos primates. Podemos especular, aunque sólo tengamos las pruebas indirectas de los fósiles, que al llegar al mioceno la inteligencia de los simios antropoides había llegado bastante lejos. Los fósiles nos dan una imagen de animales no muy distintos de los chimpancés modernos. Eran semierguidos, manualmente diestros, y tenían ya cerebros más grandes que los monos. Podemos también especular que tendían fuertemente a la "K". El rodeo necesario para llegar aquí ha sido largo, pero en este momento ya podemos examinar los posibles factores que condujeron en una línea de antropoides al bipedis-mo auténtico. En otras palabras: ¿qué relación tuvo "K" con la locomoción erguida?»
El bipedismo ha torturado a los antropólogos durante casi un siglo. Poco después de descubrir Dubois en 1891 el hombre simio de Java, descubrió también un hueso de pierna que indicaba que aquel ser caminaba erguido. La gente estaba tan poco dispuesta a creer que un cráneo tan primitivo estuviera asociado con un caminar erguido que la reacción general fue negar cualquier conexión entre las dos partes fósiles: La pierna pertenecía a un ser posterior parecido al hombre que

quedó enterrada accidentalmente cerca del cráneo; era imposible que algo tan primitivo como el hombre antropoide de Java pudiese caminar erguido.
Tanto si la asociación era accidental como si no lo era, la reacción refleja un prejuicio antiguo y arraigado. Este prejuicio revivió con fuerza cuando Raymond Dart aseguró que el Niño de Taung caminaba erguido. Incluso en los años 1960, cuando desde hacía tiempo el mundo se había visto obligado a aceptar el porte erguido de los australopitecus, el prejuicio continuaba vivo, pero en forma modificada: los antropólogos aseguraban que los australopitecus caminaban con dificultad, que arrastraban los pies de modo poco eficiente.
Se había creado un consenso general según el cual el bipedismo, el desarrollo del cerebro y el uso de herramientas tenían un origen conjunto. Se construyó para explicar esto un circuito de realimentación donde cada rasgo actuaba sobre los demás intensificándolos. De este modo, un animal que empezaba a utilizar herramientas, que tenía un cerebro ampliado capaz de fabricarlas, se dedicaría a transportarlas, y el incentivo para hacerlo impulsaría el modo erguido de caminar. A medida que este porte erguido liberaba la mano para transportar cosas, iría aumentando la tendencia al uso de herramientas y al desarrollo del cerebro. Se pensaba que hace dos millones de años las tres tendencias habían alcanzado un estadio de semidesarrollo: el cerebro había crecido pero era todavía pequeño; había herramientas (porque se habían encontrado en Olduvai), pero eran las más primitivas que podían imaginarse y merecer aún el nombre de herramientas; probablemente estaba emergiendo también la locomoción erecta (ya que de acuerdo con la teoría tenía que desarrollarse conjuntamente con los otros dos rasgos emergentes), pero todavía no se había perfeccionado.
A principios de los años I960, Sherwood Washburn, de la Universidad de California, en la mitad de una carrera larga y eminente como estudioso de la evolución y el comportamiento de los primates, formuló la hipótesis de que la locomoción sobre nudillos propia de chimpancés y de gorilas podía constituir una clave del caminar erguido, un paso intermedio en el camino hacia el bipedismo auténtico. Señaló que podían encontrarse todavía rastros de este sistema en el comportamiento humano: en la postura con tres dedos en el suelo de un jugador de rugby, o en la manera de inclinarse una persona sobre una mesa y sostenerse sobre sus nudillos y su pulgar. Washburn calificó de significativo el papel de las herramientas en la elevación gradual desde la locomoción apoyada en los nudillos hasta el bipedismo auténtico. En 1974 escribió: «La locomoción y el uso de herramientas fueron simultáneamente causa y efecto uno del otro.»
Se supusieron también otras ventajas. Cuando un mejor conocimiento de la geología y del clima sugirió que los australopitecus vivían probablemente en campo abierto, pareció razonable derivar otro beneficio del bipedismo: la postura erguida aumentaba la estatura de los individuos y les permitía descubrir mejor a los depredadores en la hierba alta.
Todas estas ideas resultaban en conjunto poco convincentes para Lovejoy, que se había licenciado no hacía mucho y que empezaba ya a interesarse en todos los aspectos de la locomoción. No podía apartar de su cabeza la idea de que ningún homínido caminando de modo imperfecto se hubiera atrevido a salir a la sabana dando traspiés para apren-

derlo luego a hacerlo mejor en la sabana misma. Si no estaba ya dotado para pasearse erguido por la sabana, no se habría aventurado en ella, y de haberlo hecho no habría sobrevivido a la experiencia.
Llegó pues a la conclusión de que los homínidos ya caminaban erguí-dos cuando se desplazaron a la sabana, y que perfeccionaron este porte extravagante por motivos que no tenían ninguna relación con su establecimiento en la sabana, y que quizá tampoco se relacionaban con el uso de herramientas. Si como él sospechaba, el bipedismo auténtico se había perfeccionado antes de que las herramientas empezaran a aparecer en el registro geológico, no podían ser un factor coadyuvante.
Por desgracia Lovejoy no pudo encontrar pruebas fósiles que avalaran estas ideas. Las discutió con sus iguales basándose en motivos lógicos, pero sin mucho éxito. Se dedicó entonces al análisis de la mecánica de la locomoción, y consiguió una comprensión de ella superior a la de la mayoría de anatomistas. Pero la «escopeta humeante», es decir los fósiles reales que probaran sus afirmaciones, no quería salir a la luz. El australopiteco bien fechado más antiguo tenía sólo unos dos millones de años de edad, y ya podía caminar bien. O eso creía él, porque otros no opinaban lo mismo. Era muy frustrante para Lovejoy que los huesos de pierna y pies de australopitecus no estuvieran representados en las colecciones de modo suficiente para confirmar este punto. Quedó pues muy asombrado cuando a principios de 1974 entré en su despacho llevando una articulación que tenía tres millones de años.
En un día la locomoción erecta se había hecho de repente un millón de años más antigua.
—Quizá sí —dijo Lovejoy al grupo reunido en mi laboratorio—. Sólo teníamos aquella pequeña articulación de rodilla. Le dije a Don que volviera allí y me encontrara el animal entero. Respondió a mis deseos descubriendo a Lucy. Le dije: «Muy bien, ahora consigue alguna variedad», y al año siguiente encontró la Primera Familia. Después de estudiar estos huesos quedó bien claro que podían caminar erguidos sin ningún problema. Esto me dio la confianza suficiente para continuar avanzando e intentar descubrir el porqué de aquel cambio. No podían ser las herramientas. Los fósiles de Don eran más antiguos que cualquier herramienta conocida. Me centré entonces en la estrategia reproductora. Esto me hizo pensar en «K».
Lovejoy preguntó a uno de los estudiantes: —Veamos —dijo— tú eres una especie con demasiado «K»; ¿qué pue
des hacer para mejorar la situación? —Pues reducir algo esta «K». Los monos tienen más éxito y yo trataré
de imitarlos. —¿Es decir que piensas retroceder? —dijo Lovejoy—. ¿Piensas volver
al cuadrupedismo? ¿Quieres un cerebro más pequeño? —Algo así. —Esto es imposible. Estás metido en aquel circuito de realimenta
ción y no puedes salir. Todo te empuja en el sentido opuesto. —-Entonces, ¿qué hago? —¿Que te parece actuar sobre el punto más evidente del circuito: una
tasa baja de natalidad? ¿Dedicarse a tener más hijos? —Pero la «K» extrema lo impide —protestó el estudiante—. Usted
mismo lo ha dicho, y ha dado un argumento muy convincente basado en el circuito de realimentación.

—Vale. Supongamos que cambiamos algo para poder tener más hijos. —¿Por ejemplo? —Por ejemplo que nos convertimos en bípedo. Este salto cuántico, este aparente «non sequitur», confundió tanto al
grupo, que Lovejoy tuvo que hacer una pausa para repasar la situación. Volvió al mioceno, período en el cual los antropoides abundaban y estaban en disposición de explorar varios modos de vida y de locomoción. De aquellos predecesores miocénicos salieron algunos antropoides bra-quiantes, otros cuadrumanos, otros que caminaban sobre los nudillos y otros bípedos. Todos practicaban mucho la «K», y con excepción de los bípedos se han de considerar todos hoy en día como experimentos evolutivos fracasados.
Si el bípedo triunfó, ¿qué hizo diferente de los demás? Respuesta: consiguió evitar una trampa en la que cayeron los demás antropoides de tendencia «K»: a saber, el espaciamiento de los nacimientos típico de los primates, la estrategia que sentencia a todos los antropoides a una tasa baja de natalidad. Esos antropoides tienen por costumbre criar un hijo cada vez. Aunque esto dure mucho, aunque la madre tenga que dedicar mucho tiempo a llevar consigo a su hijo, a alimentarlo, a cuidarlo, deberá transcurrir todo este tiempo antes de que pueda tener otro hijo. Una madre chimpancé no volverá a ser sexualmente receptiva hasta que su hijo tenga cinco años, por el motivo práctico de que tiene ya sus manos ocupadas con el primero y no puede cuidar del segundo. La razón biológica es que el amamantamiento y el cuidado del hijo inhiben el inicio del estro.
—Una solución al problema sería un sistema —dijo Lovejoy— que permitiera acelerar este ritmo de modo que los niños se superpusieran más. ¿Cómo lo haríamos?
—Desprendiéndose más pronto del primer hijo. —No es este el sistema. El sistema consiste en que la madre cuide
de dos hijos a la vez; que divida su energía primero entre dos, luego entre tres y cuatro hijos.
—¿Cómo lo hace? —Se traslada menos de lugar. De este modo gasta menos energía.
Además —explicó—, cuanto más se cambia de lugar a los hijos, más peligroso resulta hacerlo. Es más fácil que se pierdan, que se caigan del árbol o que los cacen los depredadores. Cuando la madre transporta un hijo, o bien el hijo se agarra a ella, o bien ella sostiene al hijo. Si se sube a un árbol para coger higos y alarga el brazo, el niño puede caérsele. En cambio si se queda en el suelo y no se aleja de un lugar, es menos probable que dé con él un depredador, porque tanto la madre como el hijo tienen la protección del grupo. También el sistema de alarma es mejor: treinta pares de ojos en lugar de un solo par. Los demás animales exploran también el terreno en busca de alimentos. Cuando los encuentran, la madre puede acudir directamente a la comida con una pérdida de tiempo mínima y una exposición mínima a los depredadores. Pasa su vida en un pequeño territorio que conoce íntimamente. Sabe dónde están los árboles a los que puede encaramarse. Conoce la distancia más corta a cada árbol en caso de peligro.
Hay una regla simple para esto, continuó diciendo Lovejoy: —Una menor movilidad constituye una adaptación. Si se consigue,
se podrá tener hijos con mayor frecuencia. En cualquier caso la ten-

dencia a tenerlos más a menudo está siempre presente. Lo único que la reprime es la incapacidad de la madre para cuidar adecuadamente de ellos. La madre antes de tener otro hijo ha de estar segura de que ha criado al anterior lo suficiente para que pueda valerse por sí mismo. La permanencia en un lugar aumenta esta probabilidad y le permite tener un segundo hijo algo más pronto.
Había un fallo en la lógica de Lovejoy y mi grupo lo captó inmediatamente:
—Un menor espaciamiento entre nacimientos plantea un problema. Las necesidades alimentarias se hacen mayores —la madre tiene más bocas que alimentar— precisamente cuando una menor movilidad hace más difícil conseguir comida.
—Me lo esperaba —dijo Lovejoy—. Lo que afirmáis es que la madre ha de disponer de más ayuda. Tenéis razón. Alguien tendrá que traerle comida. La gente dirá que esto es imposible citando como ejemplo el caso de los mandriles: «Viven en la sabana como los homínidos y no comparten la comida.» 0 el de los chimpancés: «Son como nosotros y tampoco comparten la comida.» En realidad la mejor manera de estudiar el problema es considerar las estrategias reproductivas de todos los primates. Estas estrategias varían, y se observan también en grados diversos casos en que se comparte la comida y se cuida de los hijos. Entre los titís, un tipo de mono del Nuevo Mundo, el macho es el principal encargado de cuidar de los hijos. Se trata de una adaptación ecológica. Los titís son animales pequeños y activos que han elaborado un sistema, consistente en tener mellizos, para responder al problema del espaciamiento de los nacimientos en los primates. Debido a su pequeño tamaño y a su actividad, tienen que comer mucho y con frecuencia. La madre podría soportar este régimen con un hijo, pero no con mellizos. Por lo tanto el padre tiene que ocuparse de ellos. La madre sale a buscar comida para ella y luego vuelve a amamantarlos. Sucede lo mismo con los monos lechuza y algunos más. Los primates tienen un comportamiento muy plástico, que recorre toda la gama posible, desde el padre como cuidador principal de los hijos hasta la madre como cuidadora exclusiva.
—Inyectemos pues algo de comportamiento masculino en el ciclo de crianza del hijo —continuó diciendo—. Pero al hacerlo se plantean otros problemas. Para que la hembra pueda disponer de ayuda ha de haber un nivel importante de cooperación en el grupo. Esto es evidente. Para que un puñado de simios antropoides puedan vivir juntos como una unidad social inteligente ayudándose mutuamente con todas las ventajas que ya hemos citado, estos animales han de poder convivir bien. No puede haber peleas continuas dentro del grupo. Por desgracia, cuando uno mezcla machos y hembras —lo que llamamos S-E-X-0— es cuando más probables son las peleas. En los grupos de lémures, de mandriles (veo que lo sabéis) cuando una hembra se pone caliente [en estro] es cuando los machos se muestran más agresivos. Es natural. Tu obligación consiste en meter tus genes en esta hembra antes de que lo haga otro compañero.
Hay sistemas para desactivar esta agresión, explicó Lovejoy. Una consiste en disminuir la competencia por el sexo. Esto puede conseguirse desarrollando un sistema de consolidación de la pareja. Si cada macho tiene su hembra, su receptáculo privado de genes, no tendrá que pelearse

Estos cuatro diagramas, preparados por Owen Lovejoy, ilustran algunas de las condiciones que según él actuaron conjuntamente para producir un homínido totalmente erguido. La sección 1 de cada diagrama describe el probable progreso del porte erguido. La sección 2 se refiere a la diferenciación epigámica: los símbolos de macho y hembra se hacen cada vez más individualistas a medida que pasa el tiempo y provocan una mayor selectividad en el emparejamiento sexual. Esto se refleja en las formas de aparejamiento de la sección 3, donde los machos se aparejan al principio indiscriminadamente con las hembras pero más tarde se estabilizan como pareja de hembras concretas. La sección 4 trata del espaciamiento de los hijos. En el diagrama A no hay solapamiento, en el diagrama B empieza a haberlo, y finalmente en el diagrama D hay familias de tres y cuatro niños.
Estos cambios van acompañados por cambios en la actividad sexual (sección 5). El ciclo menstrual de la hembra está representado por la barra larga, con una porción central que indica el período concreto de receptividad sexual. En el diagrama A casi toda la actividad sexual tiene lugar durante el estro. En los diagramas siguientes el periodo de receptividad se va alargando, hasta tener en el diagrama D una situación de receptividad sexual continua por parte de la hembra que es enteramente independiente de su ciclo de estro. Mientras tanto las relaciones de parentesco están cambiando (sección 6). En el diagrama A, sólo hay relaciones de parentesco entre una madre y su hijos. En el diagrama B se inicia una relación macho-hembra, pero no hay autén-
con los res tan tes machos p a r a es ta r r ep resen tado en las generaciones subsiguientes .
—¿Cómo se consigue es to? Un sis tema serla el iminar todo el montaje del apare jamien to abier to a todos. Renuncia r a algunas de las señales visuales de contenido sexual, las tumefacciones y los olores excitantes que dicen «Voy caliente», y que vuelven locos a los machos . Si sólo se exci tan an te u n a cier ta h e m b r a la mi t ad de los machos , e l potencial de pelea queda reduc ido de golpe a la mi tad . Es to puede lograrse haciendo

tica unión. En el diagrama C el macho queda confirmado como padre y se forma un grupo familiar. En el diagrama D estos grupos son ya numerosos.
Todas estas actividades tienen lugar en un medio ambiente cambiante (y probablemente están influidas por él: sección 7). La selva tropical, donde los antropoides cuadrúpedos se emparejaban de modo casual y no formaban familias, se convierte paulatinamente en terreno boscoso abierto y en sabana. Los círculos en los centros de los cuatro diagramas están relacionados con la búsqueda de comida y la movilidad. El círculo interior marca la zona nuclear del grupo, donde las hembras y los niños pasan su tiempo. El círculo mayor representa el límite de la zona donde los machos buscan comida. En el dia-gama A los dos círculos tienen casi el mismo tamaño, porque el macho no trae comida a la hembra y ella tiene que buscarla por sí sola. En el diagrama B el macho se aleja más en busca de comida para apartarse de la zona nuclear y dejar más comida a la hembra, que tiene necesidad de ella porque empieza a tener más hijos que alimentar. En el diagrama C este comportamiento se intensifica. El macho empieza a traer comida a su pareja. Se aleja todavía más en busca de comida, con lo que ella puede permitirse menores movimientos y exponerse así a menores peligros. El diagrama D muestra el establecimiento de un hogar-base auténtico, donde la madre puede dejar a sus hijos al cuidado de tías y de parientes mayores. Ahora está en disposición también ella de alejarse más en busca de comida.
los s ímbolos sexuales m á s individualistas. Si nos concen t ramos cada vez m á s en la individualización de las respues tas sexuales, al cabo de un t iempo vemos que una de te rminada h e m b r a sólo resul ta sexualmente excitante pa ra unos cuantos machos , quizá sólo p a r a uno . Ha dejado de ser atract iva pa ra los demás ; los demás la ignoran. El desarrol lo de s is temas es t imulantes específicos p a r a individuos se l l ama diferenciación epigámica. Yo lo l lamo enamora r se .
—Los mandr i les no se enamoran , sólo quieren sexo t sexo con cual-

quier hembra que presente los signos del estro. Entre los mandriles todas las chicas cuando van calientes atraen a todos los chicos. Las sociedades de mandriles resuelven esta situación explosiva elaborando una jerarquía de dominancia masculina. El macho jefe, o alfa, ha alcanzado la cumbre luchando con los demás machos y se mantiene en su lugar amenazando con desencadenar nuevas luchas. Es el animal m á s fuerte e intimidante del grupo, y el que puede copular de entrada con las hembras sin discusión posible. Los demás machos le ceden el lugar siguiendo un orden claro de rango descendente. El problema con este sistema es que la autoridad del macho alfa sólo se impone si él está presente. Si baja al río a beber un trago, la pierde. Siempre hay algún compañero, ojo avizor, a punto de aprovecharse. Cuando el macho alfa vuelve del río su probabilidad de tener descendencia puede haber desaparecido.
El grupo de estudiantes estaba empezando a entender que el nivel de cooperación social entre animales cuya agresividad sexual es elevada y cuya discriminación sexual es baja tiene que ser limitado. Si introducimos parejas en una sociedad así la armonía .social puede aumentar. Los machos pueden abandonar el grupo durante breves períodos de tiempo sin perder su oportunidad de estar representados sexualmente en la siguiente generación. Se hace posible el cuidado paterno a cargo de los machos y la comida puede compartirse. En consecuencia, las hembras pueden permitirse perder algo de movilidad.
—Estamos ahora en otro circuito de realimentación —dijo Lovejoy—. Podemos estudiarlo empezando por la movilidad. Si uno se hace menos móvil, puede hacerse más bípedo. ¿Por qué? Porque si uno no tiene que correr mucho, puede permitirse ser menos eficiente en la carrera y hacer otras cosas que ahora empiezan a tener más valor de supervivencia, como coger y transportar la comida adicional que uno necesita a medida que aumenta el número de hijos que está criando. Acabamos de hablar de la formación de parejas. Si tu compañero camina ahora erguido estará mejor equipado para transportar comida, y será más fácil que te la traiga. Mientras tanto tú también has mejorado. Cuando eras cuadrúpeda sólo tenías una mano libre. Una mano servía para caminar y la otra para llevar cosas. Ahora dispones de dos manos, una para aprovisionarte, la otra para aguantar al hijo. Y cada vez es más importante que lo sostengas, porque está perdiendo la capacidad de agarrarse a ti. Su pie se está convirtiendo en un mecanismo bípedo y está perdiendo la capacidad de agarrarse. Un bebé homínido no puede agarrarse a su madre con los pies porque ya no tienen la forma que permite hacerlo. De hecho apenas puede sujetarse con sus manos; es un niño demasiado desvalido. Un bebé chimpancé hace todo esto mucho mejor pero, incluso así, su madre le ha de ayudar bastante. Si un bebé de mono se suelta y cae del árbol, se acabó todo. Por lo tanto, cuando vi que los fósiles que Don encontró en Hadar eran bípedos supe algo sobre las estrategias sociales y reproductoras de estos seres. Supe que los bebés no podían agarrarse a sus madres. Supe que los padres tenían que cuidar mucho de sus hijos.
—Todo esto está muy bien —dijo un miembro del grupo—, pero hay algo que no queda claro en estos sistemas de realimentación. Además de todo esto, ¿qué pone en marcha el proceso?
—No hay puesta en marcha —dijo Lovejoy—. Sólo un flujo muy suave. Hay mucho tiempo disponible: quizá centenares de miles de

años. Uno dispone aquí y allí de medios ambientes ligeramente distintos, donde se acentúan ligeramente comportamientos distintos. Uno dispone también de unos animales bastante inteligentes. Esto es importante, porque pueden observarse e imitarse mutuamente. Pueden intentar hacer cosas. No me refiero a que un día tengan que exclamar: «Magnífico, somos bípedos, tenemos una mano libre, llevaremos algo a la chica para que desayune.» No me refiero a esto. Ningún animal ha sido nunca ni remotamente consciente de los procesos evolutivos que está experimentando. Estos procesos tienen lugar mediante diminutos incrementos. Cualquier ligera irregularidad en el circuito, si es adaptativa, ejercerá un efecto en esta parte del circuito y desde allí se difundirá a las demás partes.
El análisis todavía no les convencía del todo. —Muy bien, pero se ha dejado algo importante. ¿Por qué motivo
tiene que traer el macho comida a la hembra? Si todo depende tanto de este punto y si no hay indicios de que lo haga ningún antropoide más, ¿por qué supone que lo hizo precisamente este bípedo?
—Dije que hablaríamos de sexo. —Sí. —Continuemos hablando de sexo. Todo el sistema es una estrategia
reproductora. Empezamos ya a comprender que la locomoción no es un simple expediente para desplazarse, sino que forma parte del mecanismo total de supervivencia de la especie, el cual incluye tanto el apareamiento y la socialización, como la carrera, la comida y el juego infantil. Hemos visto el posible efecto de la formación de parejas que disminuye la agresión y mejora las posibilidades de cooperación entre los machos. Tú estás preguntando algo que ya apuntabas anteriormente: ¿cómo se decide el macho a traer por primera vez comida? ¿De acuerdo?
—Exactamente. —Quizá le guste la hembra. —Por favor, dejemos esto. —Lo digo en serio. Hablamos hace un rato de la diferenciación epi-
gámica, del enamoramiento. El único objetivo de todas las señales sexuales que una hembra exhibe cuando está en estro es atraer a machos, generar un grado máximo de actividad sexual de modo que el óvulo que está produciendo en aquel mismo momento quede fertilizado. Si no agita con la fuerza necesaria estas banderas, puede perder la ocasión. Si la pierde tendrá que esperar hasta su siguiente estro para probar de nuevo. Todos los mamíferos saben exhibir con eficacia su disponibilidad sexual. El estro sirve para esto: para garantizar la actividad sexual. Si falta esta actividad la hembra habrá ovulado en vano. Esto para un antropoide que quizá ha exagerado demasiado la «K», constituiría un esfuerzo terrible para su ya baja tasa de reproducción. La antropoide que está en celo ha de ser impregnada inmediatamente para el bien de todos los antropoides.
—Pero —dijo un estudiante— si uno aumenta la discriminación sexual disminuye el número de machos interesados en esta hembra, y disminuye la probabilidad de que quede impregnada.
—No necesariamente —respondió Love joy—. Uno puede aumentar el interés que siente por ella un macho concreto. Incluso entre los chimpancés, que se aparejan siempre de modo indiscriminado, se observan en ocasiones parejas en luna de miel que van a lo suyo durante varios

días mientras la hembra está en celo. Son pequeños casos de diferenciación epigámica.
—De todos modos... —¿Qué pasaría si uno ampliara el período de atractividad sexual de
la hembra para un macho determinado? Supongamos que ella emite sus destellos sexuales algo más de tiempo. Supongamos que prescinde paulatinamente de señales como los olores y las tumefacciones, que son elementos del ciclo del estro, y empieza a recurrir a algunos rasgos permanentes de su cuerpo: su pelo, su piel, su forma. Esto se parece más a lo que hacen las personas. Pensemos un momento en esto. Los machos y las hembras de la especie humana se atraen sexualmente los unos a los otros en todo momento, con independencia de la época o de la estación. Esta atracción no tiene nada que ver con el ciclo del estro. Ningún otro mamífero actúa así. ¿Por qué han de ser precisamente los hombres?
Nadie pudo responder. —Vamos ya. ¿De qué hablamos todo el rato? Estaba emergiendo
entre aquellos primitivos homínidos y prehomínidos un sistema de formación de parejas destinado a que un macho se sintiera atraído por una hembra permanentemente y quedara asegurada su impregnación mediante la estrategia de un aparejamiento bastante continuo en lugar de un frenesí comunal en el punto culminante de su ciclo ovultario. Es imposible que haya tremendas peleas y cópulas indiscriminadas y al mismo tiempo se formen parejas y se comparta comida. Es simplemente imposible. ¿Qué sustituto mejor para ello que un sistema que devuelva el macho a su hembra? Y si trae comida, el valor selectivo de este comportamiento empezará a notarse con bastante rapidez por la presencia de un número superior de sus genes en la reserva de genes.
—Por lo tanto no hay un punto de arranque —dije yo. —Ninguno —dijo Lovejoy—. Creo que es un fenómeno paulatino.
Podemos especular que tardó mucho tiempo en imponerse. Se inició cuando una hembra mantuvo su atractividad durante una semana por ejemplo, después de finalizado su estro. Este período se amplió lentamente. Al final los indicadores del estro ya no contaron; otros indicadores permanentes mantuvieron a su hombre (a su homínido) interesado en ella todo el tiempo.
»Y le interesa que él continúe interesado por ella, porque sólo es fértil durante unos tres días. Si copulan una vez cada dos semanas, la probabilidad de quedar embarazada es muy baja. Tan baja que no puede permitírsela. Su tarea consiste en quedar embarazada rápidamente, tan pronto como pueda hacerse cargo del siguiente hijo.
—Usted nos dice —preguntó un estudiante—, que el bipedismo provoca la formación de parejas y que provoca la distribución de comida y el nacimiento de más hijos.
—No hablo de provocar nada —dijo Lovejoy—. Esta palabra es incorrecta. En un circuito de realimentación no hay causas. Hay un reforzamiento mutuo. Yo sólo digo que todo esto sucedió y que cada factor ejerció un efecto sobre todos los demás. No digo cuál fue el primer factor. Lo ignoro. Dudo que alguien llegue a saberlo. Sospecho que no hubo una primera causa. Pero todo esto sucedió. Lo sabemos porque tú y yo estamos sentados aquí hablando del tema. Nosotros somos el resultado. Y si paso revista a las principales diferencias de comporta-, miento entre los antropoides y los homínidos, y te pido que encuentres

un sistema mejor para explicar cómo llegamos donde estamos ahora, me gustaría enterarme. —Empezó a hacer una lista sobre un trozo de papel:
—Fijaos en esto. Todo sucedió hace más de dos millones de años. Es necesario que todas estas cosas estuviesen interrelacionadas:
Homínido
Vive exclusivamente en el suelo.
Bípedo.
Forma parejas, tiende a establecer familias nucleares.
Inmovilidad cada vez mayor de las hembras y los jóvenes. Posibilidad de un hogar base.
Se comparte la comida.
Inicio del uso y la fabricación de herramientas.
El cerebro continúa creciendo.
Sexualidad continua.
Cuidados múltiples al niño.
Póngido (antropoide)
Algunos predominantemente en arboles. Algunos predominantemente en el suelo. Ninguno es exclusivamente terrestre.
No bípedo.
Sin parejas. Sin familias nucleares excepto en los gíbones.
Las hembras se mueven para conseguir comida y se llevan a los hijos consigo. No hay hogar base.
No se comparte la comida.
Falta el uso de herramientas o es inconsecuente.
El cerebro no crece más.
La sexualidad está limitada al estro.
Cuidado individual del hijo.
—Estoy tratando de explicar —continuó diciendo Lovejoy—, que uno no pretende de pronto adoptar un sistema de locomoción, al parecer estúpido, sin ningún motivo. Para entender por qué Lucy iba erguida hay que tener en cuenta muchos más elementos, aparte de la manera que tenía de moverse. Simplificando mucho, podríamos afirmar que Lucy se puso erguida para poder tener hijos con mayor frecuencia. Pero esto sería aducir una causa directa, y nosotros no queremos causas directas. Si queremos expresarnos con mayor precisión podríamos decir que en un animal cuyo complejo de especializaciones le condujo demasiado lejos en el sentido de «K», el bipedismo es una salida, un sistema para reducir «K» de nuevo.
—O bien —dije yo—, en un animal social muy inteligente que precisa de una niñez prolongada y de muchos cuidados maternos, un animal que dispone ya del potencial para caminar erguido, un animal tan orientado hacia la «K» que sólo puede vivir en los ambientes más favorables, un animal que comparte su comida, que...
—Desde luego, desde luego —dijo Lovejoy—. Esto estaba yo diciendo. —Lo resumía —dije—, porque creo que te dejaste un punto. —¿Qué punto? —Has demostrado que los antropoides están prácticamente extingui-

dos debido a sus problemas «K». Pero olvidaste decir que los homínidos, más orientados hacia la «r», están ahora en todas partes.
—Lo reconozco, pero se trata de otra historia. Es un fenómeno posterior. Introduce el tema de la importancia de las herramientas y de la cultura, que favorecen la supervivencia y la diseminación de las poblaciones. Sin ellas estaríamos todavía atascados en la sabana tropical, en una situación algo mejor que los simios antropoides, pero no mucho mejor. Nuestro tema era hoy la locomoción bípeda, algo que le sucedió al hombre antes de convertirse en hombre. Las herramientas y la cultura tienen relación con él después de hacerse hombre.
«Permitidme resumir. Creo que los homínidos aprendieron a desplazarse bípedamente en la selva, no en la sabana, aunque sea allí donde después los encontremos viviendo. Se fueron a la sabana siendo ya bípedos. Lo hicieron hace casi cuatro millones de años, según sabemos gracias a tus fósiles, Don, o quizá mucho antes, y lo hicieron sin la ayuda de herramientas. Creo que el motivo fue principalmente de índole sexual y social. Una vez perfeccionada la locomoción erguida, pudieron irse andando donde les apeteció, porque la locomoción erguida no es de por sí ineficiente. Uno puede andar todo el día tan bien como un perro. El perro sólo le ganará cuando intente correr. Por lo tanto si no hay necesidad de correr por la sabana, uno puede muy bien trasladarse y vivir allí. Y es probable que lo haga, porque en la selva empiezan a notarse las apreturas. La reproducción da bastante buen resultado, y en la selva, con tantos antropoides, no queda mucho sitio. Sabemos que esto sucedió así porque hace uno o dos millones de años los homínidos se habían esparcido ya por todas partes.
»Bastaba cubrir solamente siete metros al año, que no es mucho, para en dos millones de años alcanzar los siete mil kilómetros. Probablemente las herramientas ayudaron a llevar a cabo esta dispersión. ¿Se utilizaron las herramientas desde el principio? Es probable que los homínidos no las usaran más que los chimpancés. Pero eran ya bípedos. Caminaban erguidos y podían aspirar a algo diferente. Disfrutaban de una capacidad muy superior para coger, transportar y lanzar cosas. Hubo un período, situado en algún punto entre los cuatro millones y por ejemplo un millón y medio de años, en que el uso de herramientas pasó de ser algo totalmente insignificante a ser un elemento crítico en el desarrollo de los homínidos. El impulso inicial no se debió a que eran antropoides que usaban herramientas, como se ha dicho tanto tiempo. Los homínidos fueron antes antropoides innovadores, social y sexualmente, que se hicieron bípedos y que de este modo pudieron propagar su especie mejor o más que otros antropoides. Fue un caso de pura buena suerte que la capacidad de mantenerse erguido y de utilizar las manos, condujera a un desarrollo posterior de las herramientas y a una cultura, y a un cerebro más grande todavía, y finalmente a cuatro mil millones de seres como nosotros. Todas estas cosas llegaron como subproductos de lo que realmente importaba: una mejor estrategia reproductora.»
Lovejoy se puso el gabán: —Puede decirse que estas innovaciones «r» hicieron posible al hom
bre. Luego podríamos añadir que las herramientas le refinaron. Las herramientas crearon a Homo erectus, no a Homo en sí.

Cuando Lovejoy se hubo ido, me preguntaron si yo creía todo lo que él había dicho.
E,n general sí —dije—, aunque el mismo Owen no os pediría que le «creyerais». Esta palabra es tan inadecuada como la palabra «causa». ¿Cómo puede creer uno algo que no puede demostrar? Se acepta como algo probable si satisface los datos y si parece lógico.
—Pero usted cree en la evolución. —Sí, creo en ella, porque se trata de una teoría que ha estado mucho
tiempo circulando y recibiendo palos. Ha resistido este trato violento y sin embargo continúa siendo lógica. De hecho cuanto más se estudia (cuanto más se aprende sobre ella) más lógica se hace. Sometamos la teoría de Owen a la misma prueba del tiempo; si lo resiste la creeré. De momento la acepto como la mejor explicación del bipedismo formulada hasta el momento.
Preguntaron a Tim White su opinión. —Ya me conocéis. No creo en nada que no pueda medir, y el compor
tamiento sexual no deja fósiles. Pero en general estoy de acuerdo con Don; es lo mejorcito de que disponemos. Creo que Owen está totalmente en lo cierto cuando dijo que el bipedismo se desarrolló en la misma selva. Es imposible hacerse bípedo dando traspiés por la sabana. Es absurdo imaginar que alguien pueda correr y escapar cuando apenas está aprendiendo a hacerlo.
—¿Por qué motivo salieron los homínidos a la sabana? —le pregunté.
—Por lo que dijo Owen —contestó Tim—. En la selva empezaba a faltar espacio. No sólo porque había más homínidos, sino porque la selva estaba retrocediendo. Hacia el plioceno (la época en que debía de estar pasando todo esto) aquella enorme selva miocénica se había encogido mucho. Lo que no acabo de aceptar totalmente es esta teoría de traer comida a las hembras y a las crías. No es necesaria. Creo que la posibilidad de transportar objetos, de transportar niños, de transportar comida fue un incentivo suficiente para el bipedismo.
—¿Dejas de lado el argumento del celo? —le pregunté. —Sí, no he visto nunca un celo fósil.














