TEMA 8. NUTRICIÓN Y METABOLISMO CELULAR€¦ · El metabolismo se puede definir como el conjunto...
25
Efi-Ciencia Red https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com https://www.facebook.com/eficiencia.red 1 TEMA 8. NUTRICIÓN Y METABOLISMO CELULAR ÍNDICE DE CONTENIDOS 1.- Introducción al metabolismo. 1.1. Concepto. Nutrición autótrofa y heterótrofa. 1.2. Excreción y secreción 1.3. Concepto de metabolismo y tipos. Ruta metabólica. 1.4. Intermediarios energéticos. 2.- Catabolismo celular. 2.1. Fases generales del catabolismo y su localización. 2.2. Glucólisis. 2.3. Fermentación. 2.4. Respiración Aerobia. Etapas: formación del acetil-CoA, ciclo de Krebs, cadena respiratoria y fosforilación oxidativa. 2.5. Balance general del catabolismo de la glucosa. Rendimiento energético. 2.6. β-oxidación de los ácidos grasos o hélice de Lynen. 3.- Anabolismo celular. 3.1. Concepto de fotosíntesis y fundamentos del proceso. 3.2. Importancia biológica de la fotosíntesis. 3.3. Pigmentos y fotosistemas. 3.4. Etapas de la fotosintesis. 3.5. Fase luminosa. Fase luminosa Acíclica. Fase luminosa Cíclica. 3.6. Fase oscura: Ciclo de Calvin 3.7. Resumen de la fotosíntesis. 3.8. Factores que influyen en la fotosíntesis. 3.9. Quimiosíntesis. 4.- Integración del Catabolismo y Anabolismo
Transcript of TEMA 8. NUTRICIÓN Y METABOLISMO CELULAR€¦ · El metabolismo se puede definir como el conjunto...
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed
https://mferminprofe.jimdo.com https://www.facebook.com/eficiencia.red
1
TEMA 8. NUTRICIÓN Y METABOLISMO CELULAR
ÍNDICE DE CONTENIDOS 1.- Introducción al metabolismo.
1.1. Concepto. Nutrición autótrofa y heterótrofa.
1.2. Excreción y secreción
1.3. Concepto de metabolismo y tipos. Ruta metabólica. 1.4. Intermediarios energéticos.
2.- Catabolismo celular.
2.1. Fases generales del catabolismo y su localización. 2.2. Glucólisis. 2.3. Fermentación. 2.4. Respiración Aerobia. Etapas: formación del acetil-CoA, ciclo de Krebs, cadena respiratoria y
fosforilación oxidativa.
2.5. Balance general del catabolismo de la glucosa. Rendimiento energético. 2.6. β-oxidación de los ácidos grasos o hélice de Lynen.
3.- Anabolismo celular.
3.1. Concepto de fotosíntesis y fundamentos del proceso.
3.2. Importancia biológica de la fotosíntesis. 3.3. Pigmentos y fotosistemas.
3.4. Etapas de la fotosintesis.
3.5. Fase luminosa. Fase luminosa Acíclica. Fase luminosa Cíclica. 3.6. Fase oscura: Ciclo de Calvin
3.7. Resumen de la fotosíntesis. 3.8. Factores que influyen en la fotosíntesis. 3.9. Quimiosíntesis.
4.- Integración del Catabolismo y Anabolismo
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed
https://mferminprofe.jimdo.com https://www.facebook.com/eficiencia.red
2
1. INTRODUCCION AL METABOLISMO
1.1. CONCEPTO DE NUTRICIÓN. NUTRICIÓN AUTÓTROFA Y HETERÓTROFA Conjunto de procesos que permiten la introducción de alimento en la célula y la posterior conversión de los nutrientes que contienen en energia y en las biomoléculas necesarias para el mantenimiento de las funciones vitales. Mediante las funciones de nutrición, la célula toma materia y energía del exterior, y las transforma con dos objetivos: fabricación de nuevos materiales celulares y obtención de energía para realizar trabajos de diversa índole (transporte, síntesis, mecánico, etc.). Según los medios utilizados para obtener la energía, se distinguen dos tipos de nutrición: a) Autótrofa: Cuando la fuente de carbono para fabricar sus biomoléculas es el CO2 (molécula inorgánica)
Dependiendo de la energía utilizada para la síntesis biomolecular se distinguen entre: - Fotosintéticas: la energía necesaria procede de la luz solar, absorvida gracias a diversos pigmentos
fotosintéticos. Son los autótrofos típicos, como las plantas verdes, las algas, las cianobacerias, las bacterias purpúreas sulfurosas y las bacterias verdes del azufre.
- Quimiosintéticas: la energía necesaria la obtienen de reacciones de oxidación de moléculas inorgánicas reducidas. Solo las pueden realizar algunas bacterias: las bacterias inodoras del azufre, las bacterias nitrificantes (Género Nitrosomonas), las bacterias del hidrógeno y las bacterias del hierro. Muchos de los compuestos reducidos que utilizan las bacterias, como el NH3 o el H2S son sustancias procedentes de la descomposición de la materia orgánica. Al oxidarlas, las transforman en sustancias minerales, NO3- y SO4
2-, respectivamente, que pueden ser absorbidas por las plantas. Estas bacterias cierran, por tanto, los ciclos biogeoquímicos, posibilitando la vida en el planeta.
b) Heterótrofa: Si la fuente de carbono para formar sus biomoléculas es la materia orgánica (biomoléculas orgánicas procedentes de otros organismos). De nuevo tenemos 2 tipos:
- Quimioheterótrofos: Son los heterótrofos típicos, que obtienen la energía necesaria para fabricar sus propias moléculas de la degradación de la propia materia orgánica de la que se han nutrido. Animales (metazoos), protozoos, hongos y la mayoría de las bacterias.
- Quimioautótrofos: Son muy raros, pues obtienen la energía necesaria para la fabricación de sus propias biomoléculas de la luz. El único representante conocido son las bacterias purpureas no sulfurosas.
1.2. EXCRECIÓN Y SECRECIÓN La excreción es la expulsión al exterior celular de las sustancias de desecho metabólicas ( en muchos casos tóxicas) y de las sustancias que no les sean útiles, como el exceso de agua en las células que viven en medios hipotónicos (los paramecios han desarrollado una vacuola pulsátil para eliminar el agua que les sobra). Ocurre, de modo general por exocitosis vesicular, salvo el CO2, que se elimina por difucion simple. En la secreción las sustancias expulsadas proceden del anabolismo y tienen utilidad, para la propia célula o para otras células. Ocurre por exocitosis.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
3
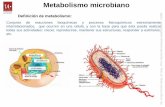
1.3. CONCEPTO DE METABOLISMO Los seres vivos son sistemas dinámicos que están continuamente intercambiando y transformando la materia del medio. Esto ocurre a través de innumerables reacciones llevadas a cabo en el medio acuoso del interior de los organismos. El metabolismo se puede definir como el conjunto de reacciones bioquímicas que procesan los cambios de la materia en los seres vivos. Hay 2 tipos de metabolismo:
A. CATABOLISMO Degradación de la materia orgánica en la que se produce un desprendimiento de energía y poder reductor. Parte de sustancias reducidas y se obtienen sustancias oxidadas.
B. ANABOLISMO Síntesis de materia orgánica que precisa de un aporte energético o poder reductor. Parte de sustancias oxidadas y se obtienen sustancias reducidas.
Cada sucesión o conjunto de reacciones que conducen a la producción de una o varias sustancias determinadas se conoce como ruta metabólica. Las rutas o vías metabólicas se caracterizan porque:
1. Están encadenadas, es decir los metabolitos de una pueden ser el inicio de otra. Esto crea una red metabólica enorme y compleja en la que se pueden relacionar todas las rutas entre sí.
2. Están reguladas por biocatalizadores o enzimas específicas.
3. Están acopladas energéticamente. Esto significa que la energía liberada en unas reacciones (exotérmicas), gracias a los transportadores energéticos como el ATP, es aprovechada en otras reacciones que precisan energía (endotérmicas).
Se muestra: - Varias reacciones formando rutas metabólicas (R1, R2 y R3). - Cada reacción catalizada por su enzima específica. - Varias rutas formando una red metabólica encadenada. - Una representación del acople energético entre 2 rutas metabólicas.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
4
Además, también es muy útil saber, de las rutas metabólicas, que:
4. Los productos de las rutas metabólicas se llaman metabolitos. 5. Cuanto más reducido está un compuesto, mayor cantidad de energía contiene, cuanto más oxidado se
halla, menor cantidad de energia tiene.
6. Las reacciones que constituyen el metabolismo son prácticamente iguales en todos los seres vivos.
1.4. INTERMEDIARIOS ENERGÉTICOS
En la célula, ya hemos visto que la energía puede ser producida en unos lugares y requerida en otros, pero… ¿cómo se transporta? Gracias a unas pocas moléculas, derivadas de ribonucleótidos (por lo que ya debemos de conocerlas) llamadas intermediarios energéticos.
A. Intermediarios transportadores de ENLACES ALTO-ENERGÉTICOS. La energía química está contenida en uno o varios enlaces de la molécula y puede ser liberada y utilizada directamente a nivel de sustrato por hidrólisis. El ATP (adenosina trifosfato) es el más común, pero hay otros como el GTP o el UDP.
B. Intermediarios transportadores de PODER REDUCTOR. Son sustancias oxidantes que le quitan electrones a otras moléculas de modo que al reducirse se “cargan” de electrones y protones. Los más comunes son el NAD+, NADP+ y FAD. Como vemos no son más que reacciones de oxidación-reducción (REDOX, donde el que se oxida pierde electrones, que gana el que se reduce). Dichos electrones se llevarán a una cadena de transporte de electrones de la membrana plasmática para que transforme el poder reductor en enlaces alto-energéticos (ATP).
La célula produce ATP medíante dos procedimentos básicos:
Por fosforilación a nivel de sustrato. Consiste en que algunos compuestos fosfatados pueden ceder grupos fosfato directamente al ADP, es decir, el proceso no está ligado a un transporte de electrones.
Por fosforilación oxidativa gracias a la ya comentada cadena de transporte de electrones. A esta cadena entran electrones de alta energía, donados por los transportadores de poder reductor, permitiendo una liberación progresiva de su energía, que será utilizada para obtener ATP gracias a ATP-sintasas.
Que el transporte de energía útil metabólica resida en un número tan escaso de moléculas que, además, son muy parecidas entre sí (derivadas de ribonucleóticos) y son universales (se encuentran en todos los seres vivos) ha hecho hipotetizar a la comunidad científica que su origen se sitúa en el comienzo mismo de la vida, o al poco tiempo en su evolución. Su eficacia y versatilidad los ha mantenido como principios dinámicos fundamentales prácticamente sin cambios a lo largo de la historia de la vida.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
5
2.- CATABOLISMO CELULAR 2.1. FASES GENERALES DEL CATABOLISMO Y SU LOCALIZACIÓN Aunque existen otros combustibles en las células (ácidos grasos y aminoácidos), vamos a describir las fases del catabolismo refiriéndonos a la glucosa como principal combustible.
La oxidación completa de una molécula de glucosa puede resumirse en la siguiente reacción general: Esta oxidación se realiza en la célula en dos fases bien diferenciadas, glucólisis y respiracion, existiendo la posibilidad de fermentación en condiciones anaerobias (ausencia de oxígeno): 1.- Glucólisis: Consiste en una serie de reacciones, mediante las cuales una molécula de glucosa (de 6 átomos de carbono) se transforma en dos moléculas de ácido pirúvico (de 3 átomos de carbono). Durante esta serie de reacciones se obtienen 2 ATP, 2 NADH + H. Se realiza en el citosol de las células, sin intervención del oxígeno (es anoxigénica). Es una ruta universal que realizan todos los seres vivos, tanto autótrofos como heterótrofos. 2.- Fermentación. Es una alternativa anaerobia (sin necesidad de oxígeno) a la respiración celular, que comento más abajo). Tiene un rendimiento energético mucho menor, ya que no se produce la oxidación total de la glucosa, resultanto un compuesto orgánico (aún bastante reducido, es decir, con mucha energía) como producto final. Dependiendo del producto final hay diferentes tipos: alcohólica, láctica, acetica, ácido mixta y butírica son las más importantes.
3.- Respiración celular: Son el conjunto de reacciones bioquímicas por las cuales determinados compuestos orgánicos son degradados completamente, por oxidación, hasta convertirse en sustancias inorgánicas. El resultado es la obtención de una enorme cantidad de energía (ATP). Puede ser tanto aerobia u oxibiótica (aceptor final de electrones es el oxígeno, formandose H2O) como anaerobia o anoxibiótica (el aceptor final de electrones es una molécula inorgánica diferente al oxígeno. Si es NO3
- se formará NO2-; si es SO4
2- se formará S2-; si es CO2 se formará CH4 ). Consta de dos procesos:
A. Ciclo de Krebs: En esta fase se termina de oxidar la materia orgánica. El ácido pirúvico (de 3 átomos de carbono) debe perder un átomo de carbono y convertirse en acetil-CoA (de 2 átomo de carbono), que ya puede entrar en el ciclo. Mediante una serie de reacciones, estos dos carbonos se transformarán en dos moléculas de C02 (producto final de la reacción general citada anteriormente). También se obtienen: 3 NADH + 3 H, 1 FADH2 y 1 GTP. Esta fase se lleva a cabo en la matriz mitocondrial.
B. Fosforilación Oxidativa (cadena de transporte de electrones): es la síntesis ATP que se produce a
expensas de la energía generada en el transporte de electrones a través de la cadena respiratoria. Durante esta fase el hidrógeno (poder reductor) acumulado en las coenzimas se utiliza para sintetizar ATP (síntesis quimiosmótica). Simultáneamente, las coenzimas se oxidan de nuevo, siendo el aceptor final de electrones, en la aerobia, que es la que estudiaremos, el 02, que se transforma en H20. Esta fase tiene lugar en las crestas mitocondriales (membrana mitocondrial interna).
C6H1206 + 6 02 C02 + 6 H20 + 38 ATP
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
6
2.2. GLUCÓLISIS Es la ruta inicial en la degradación de la glucosa, aunque todos los glúcidos pueden acceder a la misma, o bien directamente o tras sufrir algunas transformaciones. Es una ruta universal (que realizan todos los seres vivos), que ocurre en el citoplasma celular (citosol), de carácter anerobio (no necesita oxígeno para realizarse) y el ATP obtenido es a nivel de sustrato. Estas características sugieren que la glucólisis es una ruta muy primitiva, que ya apareció en los mismos orígenes de la vida y que se ha mantenido prácticamente sin modificaciones hasta la actualidad.
La vía más conocida de la glucólisis es la de Embden-Meyerhof, que es la que estudiamos aquí. Hay algunas alternativas que varían ligeramente de esta, pero son mucho menos comunes. En la glucólisis ocurre una secuencia de reacciones en la que una molécula de glucosa de (6 átomos de carbono) se transforma en dos moléculas de ácido pirúvico (3 átomos de carbono cada una), produciéndose 2 (NADH + H+) y 2 ATP de ganancia neta (se producen 4, pero se consumen 2 en la misma ruta). Ecuación general: El NADH + H+ producido podrá ser utilizado posteriormente como moléculas con poder reductor en la cadena de transporte electrónico para obtener más ATP por fosforilación oxidativa.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
7
La glucólisis se divide en tres etapas para una mejor comprensión del proceso:
A. FOSFORILACIÓN. De activación energética en la que se utilizan 2 moléculas de ATP para activar la glucosa a glucosa 6-fosfato primero, y a fructosa 1,6-difosfato después.
B. OXIDACIÓN. En esta fase se oxidan los gliceraldehído-fosfato para rendir energía (2ATP) y poder reductor (2NADH + H+).
C. RESTITUCIÓN del ATP consumido. Una última enzima quinasa actuará para transformar los fosfoenolpiruvato en 2 piruvatos, rindiendo 2 ATP, que compensan los 2 ATP utilizados en la fase de activación.
A continuación os dejo la ruta metabólica al completo, paso a paso, con los nombres de los metabolitos y las enzimas que intervienen. No es necesario que conozcáis las formulas de los compuestos.
2.3. FERMENTACIÓN Concepto. La fermentación es un proceso de oxidación de la materia orgánica en el que el aceptor final de electrones no es una molécula inorgánica, como el oxígeno, sino una molécula orgánica, que varía según el tipo de fermentación. Finalidad. Regenerar el NAD+ consumido en la glucólisis y que fueron reducido. Su rendimiento energético es mucho menor que en la respiración (solo 2 ATP), ya que los productos finales orgánicos retienen parte de la energía libre original de la glucosa, es decir, no han sufrido una oxidación total. ¿En qué células ocurre? En células anaerobias o anaerobias facultativas (realizan la respiración o no dependiendo de la presencia de oxígeno) que viven en ambientes sin oxígeno; o en algunas que puedan encontrarse circunstancialmente con falta de oxígeno, como les ocurre a las células musculares durante el ejercicio intenso, cuando la sangre no aporta suficiente oxígeno a la célula y su única fuente de ATP es la glucólisis. Tipos de fermentación. Aunque ya hemos comentado que hay varios, nosotros solo estudiaremos 2.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
8
A. Fermentación láctica. En esta fermentación el pirúvico se reduce gracias a la enzima lactato deshidrogenasa y se convierte en lactato (el ácido láctico en disolución se llama lactato). Esta fermentación se lleva a cabo en algunas bacterias (Lactobacillus y Streptococcus), responsables de la obtención de productos derivados de la leche (yogur, queso, kéfir, etc.). También se puede producir esta fermentación en las células musculares de animales o en los súper especializados glóbulos rojos de mamiferos, que han perdido las mitocondrias.
B. Fermentación alcohólica. En ella los productos finales son etanol y CO2. Las enzimas protagonistas son la piruvato descarboxilasa y la alcohol deshidrogenasa, encargándose esta última de recuperar los NAD+, imprescindibles para que la glucólisis no se bloquee y pueda seguir funcionando. Es común en microorganismos facultativos como levaduras (Gen. Saccharomyces) y bacterias (Gen. Zymomonas). En la industria química es una ruta de suma importancia, pues es el modo de obtener alcohol etílico. También es de destacar su utilidad en la industria alimenticia, para la obtención de bebidas alcohólicas (vino, cerveza, sidra, destilados…) y en bollería, donde el producto más útil es el CO2, el cual permite obtener masas esponjosas.
2.4. RESPIRACIÓN AEROBIA (La anaerobia no la estudiaremos, basta con conocerla)
Concepto. Proceso catabólico aerobio en el que las moléculas orgánicas se oxidan totalmente. El oxígeno es el último aceptor de electrones. El rendimiento energético es mucho mayor que en las fermentaciones. (38 ATP). Localización. Tiene lugar en las mitocondrias de las células eucariotas, y en el citosol y la membrana plasmática en los organismos procariotas. Importancia biológica. Es la forma más rentable de obtener energía (ATP).
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
9
Etapas de la respiración aerobia: I. Formación de Acetil-CoA por oxidación y descarboxilación del piruvato, de los ácidos grasos (beta-
oxidación) y de los aminoácidos. Se produce CO2 y se obtiene poder reductor (1 NADH + H+).
II. Ciclo de Krebs. Degradación del Acetil Co-A con la producción de CO2 como los restos oxidados y la obtención de energía a nivel de sustrato (1 GTP) y poder reductor (3 NADH + 3H+ y 1 FADH2).
y de átomos de hidrógeno. III. Cadena respiratoria y fosforilación oxidativa. Uso del poder reductor creado para liberar, poco a
poco y gracias a la cadena de transporte electrónico de la membrana interna mitocondrial, la energía contenida en electrones de orbitales alto energéticos. Tiene acoplada la fosforilación del ADP a ATP por el complejo ATP sintasa (fosforilación oxidativa).
I. OXIDACIÓN DEL PIRÚVICO A ACETIL-COA En condiciones aerobías, el piruvato se convierte en Acetil-CoA mediante una descarboxilación oxidativa. Esta reacción la realiza un gran complejo multienzimático de la matriz mitocondrial (en procariotas se encuentra en el citosol) llamado piruvato deshidrogenasa. En primer lugar el piruvato debe de ser transportado desde el citoplasma a la matriz mitocondrial. La membrana externa la atraviesa sin problemas, como la mayoría de metabolitos pequeños; pero la interna en mucho más restrictiva. En el caso del piruvato ocurre un cotransporte por simporte de protones. Posteriormente, ya en la matriz, actúa la piruvato deshidrogenasa.
II. CICLO DE KREBS El Ciclo de Krebs (también llamado Ciclo del Ácido Cítrico o Ciclo de los Ácidos Tricarboxólicos) recibe su nombre en honor a Hans Adolf Krebs, bioquímico alemán que lo describió en 1937 y que recibió el premio nobel de medicina en 1953 por sus aportaciones al metabolismo energético. Se trata de una ruta cíclica que oxida completamente a CO2 el acetil-CoA, liberando energía en forma de GTP (molécula similar al ATP en estructura y rendimiento), NADH y FADH2. Estas dos últimas moléculas son, como ya sabemos, transportadoras de poder reductor, de electrones altamente energéticos que serán utilizados en la cadena respiratoria transportadora de electrones.
Simporte del piruvato Acción del complejo piruvato deshidrogenasa
+
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
10
La molécula de acetil-CoA de la matriz mitocondrial será la que ingrese al Ciclo de Krebs. Las reacciones que lo componen son ocho, todas ellas catalizadas por sus correspondientes enzimas, y las tenéis expuestas en el esquema inferior que, como suele, se trata del resumen del vídeo sobre el mismo tema que podéis ver en mi canal de Youtube. Para una mejor comprensión y para recordarlo con mayor facilidad, he dividido el proceso en 2 fases: - FASE I. En esta primera fase se produce la total oxidación del acetil-CoA a CO2.
- FASE II. Ocurre la regeneración del oxalacetato para que pueda volver a entrar otra molécula de acetil-CoA.
III. CADENA RESPIRATORIA Y FOSFORILACIÓN OXIDATIVA
Ambos procesos ocurren en la membrana interna de la mitocondria, en las crestas mitocondriales, donde la ATP-sintasa es muy abundante. Veamos cómo sucede: 1. En la cadena respiratoria las moléculas NADH y FADH2 ceden sus electrones alto-energéticos a unas enzimas de membrana que, en una cadena de electrones y sucesivas oxidaciones-reducciones liberan progresivamente su energía. El NADH los cede al complejo-I (activarán los 3 niveles de acoplamiento) y el FADH2 al complejo-II (solo activará dos niveles de acoplamiento; es por ello que por cada NADH se obtendrán 3 ATP mientras que por cada FADH2 solo se obtienen 2 ATP. Al final de la cadena hay una última molécula que será reducida (aceptor último de electrones, que en la respiración aerobia es el oxígeno, produciendo una molécula de agua).
2. Dicha cadena electrónica produce, en tres bombas de protones (“lugares de acoplamiento”) la salida de protones desde la matriz mitocondrial al espacio intermembranal.
3. Esto genera una gran inestabilidad por gradiente de potencial electroquímico (mayor carga eléctrica positiva y con mayor concentración de protones en el espacio intermembranal) que se resuelve con la salida de los protones de vuelta hacia la matriz mitocondrial a través de la ATP-sintasa. Este proceso se conoce como teoría quimiosmótica de Mitchell.
4. La ATP-sintasa aprovecha la “fuerza protón-motriz” para sintetizar ATP a partir de ADP y un fosfato inorgánico. Dicha síntesis de ATP se conoce como fosforilación oxidativa y ocurre gracias a la cadena respiratoria, quedando claramente diferenciada de la fosforilación a nivel de sustrato que ocurría directamente en las reacciones metabólicas de Glucólisis o Ciclo de Krebs.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
11
2.5. BALANCE GENERAL DEL CATABOLISMO DE LA GLUCOSA. RENDIMIENTO ENERGÉTICO.
Dos de los NADH son formados en el citoplasma durante la glucólisis. Para ser transportados a la matriz mitocondrial para que posteriormente sean oxidados por la cadena transportadora de electrones, tienen que pasar por medio de transporte activo al interior de la mitocondria , esto supone 1 ATP por NADH. Por lo tanto, el balance final, resulta en 36 ATP por glucosa y no 38 ATP en células eucariotas (los procariotas no tienen mitocondrias a los que introducir los NADH.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
12
2.6. β-OXIDACIÓN DE LOS ÁCIDOS GRASOS
Los ácidos grasos, provenientes en su mayor parte de la digestión de triacilglicéridos (grasas y aceites), aportan una enorme cantidad de energía gracias al proceso de ß-oxidación, también conocido como Hélice de Lynen en honor a su descubridor, el bioquímico alemán Feodor Lynen. Ocurre en la matriz mitocondrial, a la que deberán de penetrar. Recordemos que los que tienen colas demasiado largas deben primero de ser escindidos en el peroxisoma. Definición: proceso degradativo de los ácidos grasos por sucesivas oxidaciones en las que van escindiéndose moléculas de Acetil-CoA hasta su completa oxidación. Como es común en bioquímica, vamos a dividir el proceso en varias fases para una mejor comprensión del mismo (ya que en la célula ocurre de un modo fluido y continuo). 1. FASE DE ACTIVACIÓN. Ocurre en el citosol celular, y consiste en la adición del Coencima-A a una molécula de ácido graso, formando un Acil-CoA. Como vemos se invierte 1 ATP, pero como se utiliza el enlace éster fosfórico entre el primer y segundo fosfato (más energético), resultando una molécula de AMP, es equivalente al gasto de 2 ATP. 2. ENTRADA A LA MITOCONDRIA. El Acil-CoA no puede atravesar las membranas mitocondriales, de modo que necesita de un sistema de transporte transmembrana, basado en la lanzadera de carnitina que realizan los complejos enzimáticos Carnitina Aciltransferasa-I y Carnitina Aciltransferasa-II, en la membrana externa e interna respectivamente. 3. ß-OXIDACIÓN O HÉLICE DE LYNEN. Ocurre en la matriz mitocondrial. A su vez subsividida en varias fases:
3.1. Deshidrogenación del Acil-CoA (Acil-CoA es el nombre genérico del ácido graso unido al coenzima A, independientemente de su número de carbonos) y obtención de FADH2. 3.2. Hidratación con adición de una molécula de agua. 3.3. Segunda dehidrogenación y obtención de más poder reductor, en este caso en forma de NADH. 3.4. Fragmentación del 3-Cetoacil con liberación de una molécula de Acetil-CoA y una molécula de Acil-CoA con 2 carbonos menos.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
13
A continuación tenéis en detalle la ruta de un ciclo, para la obtención de un aceil-CoA. Es evidente que, dependiendo de la longtud de la cadena del ácido graso se necesitarán más o menos ciclos de oxidación. En el caso de palmatoil-CoA, de 16 carbonos, se necesitan 7 vueltas de ciclo para escindir sus 8 acetil-CoA
Estas reacciones seguirían repitiéndose hasta la completa degradación del Acil-CoA. El rendimiento es muy elevado. Así, para un ácido graso de 16 carbonos, como el Palmatoil-CoA obtendríamos: - 8 Acetil-CoA, que en ciclo de Krebs + Cadena Respitatoria rinden 96 ATPs. - 7 FADH2, que en cadena respiratoria rinden 14 ATPs. - 7 NADH, que en cadena respiratoria rinden 21 ATPs. - TOTAL de 129 ATPs !!! (Si le restamos el equivalente a ATP gastados en la activación del ácido graso 131-2).
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
14
3.- ANABOLISMO CELULAR
Hemos visto que el metabolismo lo podemos dividir en dos grandes grupos: anabolismo y catabolismo. Ya hemos visto el catabolismo, pasemos ahora al anabolismo. Entendemos por anabolismo como todos aquellos procesos metabólicos que tienen como finalidad la construcción o síntesis de materia orgánica. Dependiendo del tipo de materia prima utilizada en los procesos anabólicos, hay dos líneas generales:
1. Uso de moléculas inorgánicas para obtener moléculas orgánicas sencillas (Ejs: Ácido-3
fosofglicérico o Acetil-Co-A). A través de unas importantísimas rutas metabólicas, base de la vida en la Tierra, que realizan los organismos autótrofos.
2. Uso de moléculas orgánicas sencillas para sintetizar moléculas orgánicas más complejas y macro-moléculas orgánicas (Ejs: colesterol, aminoácidos, nucleótidos, proteínas o ácidos nucleicos). La llevan a cabo todos los seres vivos, tanto organismos autótrofos como heterótrofos.
En nuestro estudio revisaremos solamente las vías anabólicas que llevan a la creación de materia orgánica a partir de inorgánica. Los seres vivos que son capaces de realizar esta proeza son los autótrofos y, como os podéis imaginar, son la base de todas las cadenas tróficas en cualquier ecosistema… ¡¡sin ellos la vida no sería posible!! Hay dos tipos de organismos autótrofos:
A. QUIMIOAUTÓTROFOS. También llamados quimiolitotrofos, utilizan la energía química que, mediante el proceso de quimiosíntesis, obtienen de la oxidación de moléculas inorgánicas. Solo lo realizan algunas bacterias, como las bacterias nitrificantes del suelo. B. FOTOAUTÓTROFOS. También llamados fotolitotrofos, utilizan energía lumínica para, gracias al proceso de fotosíntesis, obtener materia orgánica a partir de inorgánica. Hay dos tipos:
B.1. Anoxigénica o bacteriana. No libera oxígeno, la llevan a cabo solo algunas bacterias, como las bacterias púrpura del azufre y verdes del azufre. B.2. Oxigénica o vegetal. Típica de las plantas, se produce la liberación de oxígeno. Hay tres rutas ligeramente diferentes entre sí: la C3, la C4 (o de Hatch-Stack) y la CAM (Crassulacean Acid Metabolism o Mecanism), aunque nosotros solo vamos a estudiar la de tipo C3.
3.1. CONCEPTO DE FOTOSÍNTESIS Y FUNDAMENTOS DEL PROCESO
La fotosíntesis es considerada, por muchos, como el proceso biológico más importante que haya existido. Esta afirmación puede en un principio parecer exagerada, pero si tenemos en cuenta que no solo ha permitido la vida en la Tierra tal y como la conocemos, sino que también ha cambiado la composición atmosférica de todo el planeta va ganando sentido. Fotosíntesis puede ser definida como el proceso de transformación de energía luminosa en energía química, que en última instancia es ATP, la moneda energética biológica por excelencia y poder reductor (NADPH + H+). Dichos ATP y poder reductor serán posteriormente utilizados en la síntesis de materia orgánica a partir de inorgánica.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
15
Los protagonistas de este proceso son los pigmentos fotosintéticos y las reacciones REDOX (oxidación-reducción) que ocurren en una cadena de electrones de fundamento similar a la ya conocida de las mitocondrias para obtener ATP por fosforilación oxidativa. Dichos pigmentos (clorofilas, carotenos y xantofilas) son moléculas que tienen la peculiaridad de excitarse con la luz, quedando sus electrones de última capa tan activos que se pierden con relativa facilidad al entrar en contacto con otra molécula, el aceptor primario de electrones, que los toma (se reduce y oxida al pigmento). Estos electrones altamente energéticos son los que van a ir poco a poco, y en una cadena de electrones, soltando la energía que habían adquirido gracias a la luz. La energía liberada en la cadena de electrones será utilizada para formar ATP y poder reductor, los cuales, posteriormente, aportarán la energía necesaria para crear materia orgánica a partir de inorgánica en el Ciclo de Calvin. Los electrones perdidos por el pigmento serán repuestos por otra molécula, el dador primario de electrones.
A. Si este dador primario es el agua se desprenderá oxígeno, y el tipo de fotosíntesis se conoce como oxigénica o vegetal. La llevan a cabo plantas, algas y cianobacterias.
B. Si el dador primario no es el agua no se desprende oxígeno, y el tipo de fotosíntesis se llama anoxigénica o bacteriana. La llevan a cabo algunas bacterias como las purpúreas y verdes del azufre.
3.2. IMPORTANCIA BIOLÓGICA DE LA FOTOSÍNTESIS
Insistamos en la importancia de la fotosíntesis como proceso biológico, y desgranemos los motivos:
La síntesis de materia orgánica a partir de la inorgánica se realiza fundamentalmente mediante la fotosíntesis; luego irá pasando de unos seres vivos a otros mediante las cadenas tróficas, para ser transformada en materia propia por los diferentes seres vivos.
Produce la transformación de energía luminosa, no utilizable por los seres vivos, en energía química, necesaria y utilizada por todos los seres vivos.
En la fotosíntesis se libera oxígeno como subproducto, que es necesario para ser utilizado en la respiración aerobia como oxidante.
La fotosíntesis fue causante del cambio producido en la atmósfera primitiva, que era anaerobia y reductora.
De la fotosíntesis depende también la energía almacenada en combustibles fósiles como el carbón, el petróleo y gas natural.
Se puede concluir que la diversidad de vida existente en la Tierra depende principalmente de la fotosíntesis.
Acción de la fotosíntesis
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
16
3.3. PIGMENTOS Y FOTOSISTEMAS La fotosíntesis implica la absorción de energía lumínica, lo cual ocurre gracias a los pigmentos fotosintéticos, así que veamos primero qué es un pigmento. Pigmento: toda sustancia que absorbe unas determinadas longitudes de onda (λ) del espectro visible, reflejando el resto. Es por ello que los diferentes pigmentos son observados con distintos colores. Así, por ejemplo un pigmento rojo lo es porque absorbe longitudes de onda de alta frecuencia (por debajo de 650 nm) y refleja el resto (de baja frecuencia o rojas). Será negro si absorbe todas las λ del espectro visible, y blanco si las refleja todas. Los pigmentos fotosintéticos son aquellos que participan en la fotosíntesis, y se encuentran asociados a proteínas y lípidos, en unos complejos llamados fotosistemas.
Los fotosistemas están constituidos por una antena, en la que hay cientos de moléculas de pigmentos, tanto principales como accesorios. Funciona como un colector de energía luminosa de diferentes longitudes de onda, que transmiten por resonancia, de unos a otros, la energía absorbida por uno de ellos. Esta energía es finalmente dirigida hacia una molécula de clorofila a, conocida como clorofila diana, la cual se encuentra en el centro de reacción del fotosistema. Esta clorofila diana pierde un par de electrones al excitarse (se oxida), dándoselos al aceptor primario de electrones (se reduce), que comenzará con una cadena electrónica en la que se va extrayendo poco a poco la energía a los electrones excitados. La clorofila diana recupera sus electrones gracias al dador primario de electrones. El fotosistema I (PSI) capta la luz cuya longitud de onda es menor o igual a 700 nm. El fotosistema II (PSII) capta la luz cuya longitud de onda es menor o igual a 680 nm.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
17
3.4. ETAPAS DE LA FOTOSÍNTESIS Y SU LOCALIZACIÓN
La fotosíntesis se divide en dos fases bien diferenciadas:
- Fase luminosa. Requiere luz para que ocurra y ocurre en la membrana del tilacoide. Suceden tres procesos sumamente importantes, la fotólisis del agua (y liberación de O2), la fotofosforilación y la fotorreducción, necesarios para obtener la energía y el poder reductor que serán utilizados en la siguiente fase.
- Fase oscura. Llamada así de modo tradicional porque la luz no es imprescindible, sin embargo personalmente prefiero la denominación “fase biosintética”, ya que no lleva al error de que esta fase ocurre solo cuando no hay luz (en realidad ocurre constantemente, con y sin luz). Ocurre en el estroma del cloroplasto y en ella se sintetizan las moléculas orgánicas básicas para el organismo gracias a la fijación del CO2 con el ATP y NADPH obtenidos en la fase luminosa.
También se les puede llamar fases dependiente e independiente de la luz, respectivamente.
3.5. FASE LUMINOSA (O DEPENDIENTE DE LA LUZ) La fase luminosa tiene lugar en la membrana de los tilacoides, ya que en ella se encuentra la maquinaria molecular necesaria para llevarla a cabo: los dos fotosistemas (PSII y PSI), la cadena transportadora de electrones y enzimas ATP-sintetasas. Gracias a los fotosistemas se capta la energía lumínica que permitirá la salida de un par de electrones excitados de los fotosistemas. Dichos electrones irán pasando por determinados aceptores a lo largo de una cadena electrónica. Los procesos de óxido reducción producidos permitirán que la energía contenida en los electrones excitados sea, poco a poco, convertida en energía química. Mediante los procesos de fotofosforilación y fotorreducción. Dependiendo de que participen uno o dos fotosistemas, la cadena de electrones puede ser de dos tipos: acíclica (participan los dos fotosistemas) y cíclica (solo el fotosistema I). En la cadena acíclica los electrones son repuestos por otros cedidos por una molécula del agua en la fotosíntesis oxigénica, por el proceso conocido como fotólisis del agua, con el consiguiente desprendimiento de oxígeno y acumulación de protones en el interior del tilacoide, lo que ayuda a la formación del gradiente electroquímico necesario para la fotofosforilación*. Como podemos deducir, en la cadena cíclica no se produce la fotólisis del agua, por lo que no hay desprendimiento de oxígeno ni fotoreducción, pues la ferredoxina no cede los electrones a la NADP reductasa, sino al citocromo b6f. * La fotofosforilación se explica según la teoría quimiosmótica de Mitchell, propuesta en 1961 por el bioquímico inglés Peter Mitchell, la cual postula que la diferencia en concentración de protones entre el interior y el exterior del ti lacoide genera un gradiente electroquímico que se resuelve con la salida de protones a través de la ATP-sintasa, que aprovecha esta energía cinética de los protones en la salida (fuerza protón-motriz) para sintetizar ATP.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
18
A. FASE LUMINOSA ACÍCLICA - Definición: Flujo de electrones que proceden de los fotosistemas, al excitarse por la acción de la luz, y que son conducidos a través de los diferentes aceptores hasta el NADP+ a la vez que hay un gradiente de protones cuya energía es utilizada para la síntesís de ATP por la enzima ATP-sintasa de membrana. - Localización: Ocurre en la membrana de los tilacolides, ya que en ella se encuentra la maquinaria molecular necesaria para llevarse a cabo: los dos fotosistemas (PSI y PSII), la cadena transportadora de electrones y la ATP- sintasa.
- Procesos que tienen lugar: .Fotolisis del agua: H2O
1/2 02 + 2 H+ + 2e-
.Fotorreducción (síntesis de poder reductor): NADP+ + 2e- + 2H+ NADPH + H+
.Fotofosforilación (formación de ATP): nADP + nPi nATP .
- Desarrollo del proceso: El proceso se inicia con la llegada de fotones al fotosistema II. Esto provoca la excitación de su pigmento diana, la clorofila P680, que pierde 2 electrones. Los electrones son captados por un aceptor, el cual los cede a una cadena transportadora de electrones. Los electrones perdidos por la clorofila P680 son repuestos por los liberados de la fotolisis del agua. Los protones procedentes de la fotolisis se acumulan en el interior del tilacoíde. Al incidir la luz sobre el fotosistema I, la clorofila P700 pierde 2 electrones que son captados por moléculas transportadoras, que los conducen hasta el NADP que se reduce a NADPH (fotorreducción). Los electrones perdidos por la clorofila P700 son repuestos por los electrones que proceden del PSII, desde donde han pasado por el transporte electrónico que vemos en el esquema de la cadena de la página siguiente. La energía desprendida en algunos pasos del transporte electrónico se aprovecha para bombear H+ desde el estroma hasta el interior del tilacoide. Estos, sumados a los procedentes de la fotólisis del agua, crean un gradiente electroquímico a ambos lados de la membrana. La salida de H+ a favor de gradiente desde el tilacoide hasta el estroma, activa a una ATP-sintasa, con la consiguiente formación de ATP o fotofosforilación (según la hipótesis quimioosmótica de Mitchell).
B. FASE LUMINOSA CÍCLICA En la fase luminosa cíclica interviene sólo el fotosistema I, creándose un flujo de electrones que en cada vuelta da lugar a la síntesis de ATP. Como no interviene el PSII, no hay fotolisis del agua y, consecuentemente, no se desprende oxígeno ni hay reducción del NADP+. Sólo se obtiene ATP. Se denomina cíclica porque en ella los propios electrones perdidos por la clorofila diana vuelven a ella en circuito cerrado. La finalidad que tiene esta fase cíclica es subsanar el déficit de ATP obtenido en la fase acíclica (algo más de una molécula de ATP por cada par de electrones que va desde el agua hasta el NADP+ para formar NADPH) para poder realizar la fase oscura posterior, ya que para la reducción de una molécula de CO2 se necesitan dos moléculas de NADPH y tres de ATP.
Luz
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
19
3.6. FASE OSCURA: Ciclo de Calvin (O BIOSINTÉTICA O INDEPENDIENTE DE LA LUZ) En la fase oscura, también llamada biosintética, se utiliza la energía y el poder reductor obtenidos en la fase luminosa para sintetizar materia orgánica a partir de materia inorgánica. Esto ocurre mediante una serie de reacciones que tienen lugar en el estroma del cloroplasto en las células fotosintéticas eucariotas. La vía que estudiaremos fue descrita por Melvin Calvin en los años cuarenta del s. XX (también se conoce como Ciclo de Calvin, en su honor). El ciclo, que tenéis con detalle en la imagen de la siguiente página, lo he dividido en cuatro fases para un mejor estudio: A. FASE DE CARBOXILACIÓN (o fijación del carbono). La ribulosa 1-5 bifosfato es carboxilada (se le añaden moléculas de dióxido de carbono atmosférico) por acción de la enzima rubisco (ribulosa bifosfato* carboxilasa/oxidasa). Cada ribulosa será transformada en dos moléculas de 1,3 bifosfoglicerato. Se necesitan seis moléculas de ribulosa para obtener, al final del ciclo, una molécula de glucosa.
B. FASE DE REDUCCIÓN. Se utilizan los ATP y NADPH obtenidos en la fase luminosa para reducir los gliceratos, aumentando su energía química. 12 gliceratos son transformados en 7 moléculas de gliceraldehído 3 fosfato y 5 moléculas de dihidroxiacetona fosfato.
C. FASE DE FORMACIÓN DE HEXOSAS. Tres moléculas de gliceraldehído y tres de dihidroxiacetona serán utilizadas para sintetizar 3 moléculas de fructosa 6 fosfato. De ellas solo una sale del ciclo para ser transformada en glucosa 6 fosfato y almacenada como almidón. Las otras dos moléculas de fructosa 6 fosfato vuelven al ciclo, entrando en la siguiente fase.
D. FASE DE REGENERACIÓN. Las dos moléculas de fructosa 6 fosfato comentadas, más las restantes de gliceraldehído (4 moléculas) y de dihidroxiacetona (2 moléculas), sufrirán una serie de reordenamientos de sus átomos de carbono para ser finalmente transformadas en 6 moléculas de ribulosa 6 fosfato. Éstas, podrán recomenzar el ciclo tras ser activadas por ATP y transformadas en la ribulosa 1-5 bifosfato del inicio de la ruta. * Es común utilizar el prefijo “bis” en lugar de “bi”: Ej. Ribulosa bisfosfato.
Esquema de las fases acíclica y cíclica
3 FOTORREDUCCIÓN
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
20
El ciclo puede resumirse, sin nombre de enzimas, de esta otra manera:
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
21
3.7. RESUMEN DE LA FOTOSÍNTESIS
La ecuación global de la fotosíntesis quedaría así:
En el proceso se han utilizado 12 moléculas de NADPH (fotorreducción) y 18 ATP (fotofosforilación) obtenidos en la fase luminosa a partir de energía lumínica. Dicho proceso ha involucrado, en las membranas fotosintéticas, a un par de cadenas de transporte electrónico (cíclica y acíclica) en las que, gracias a sucesivas reacciones de oxidación-reducción, se ha ido extrayendo la energía a electrones de pigmentos fotosintéticos, energizados la luz visible; así como la fotólisis del agua (en la fotosíntesis oxigénica) con el consecuente desprendimiento de oxígeno. La síntesis de glucosa ha ocurrido en una serie de reacciones realizadas en el estroma del cloroplasto (o en el citoplasma si se trata de bacterias fotosintéticas), conocidas como Ciclo de Calvin, en la que la importantísima enzima RUBISCO ha realizado la carboxilación o fijación de carbono atmosférico. El listado de procesos fotosintéticos es el siguiente: 1. Absorción de energía lumínica.
2. Fotólisis del agua.
3. Desprendimiento de oxígeno.
4. Fotofosforilación.
5. Fotorreducción.
6. Carboxilación
Cuadro resumen
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
22
3.8. FACTORES QUE INFLUYEN EN LA FOTOSÍNTESIS
Cuando estudiamos la ecuación general de la fotosíntesis, vimos que, para sintetizar una molécula de glucosa hacen falta 18 ATP y 12 NADPH obtenidos en la fase lumínica. Estas moléculas contienen energía química almacenada en sus enlaces, la cual procede de energía luminosa que ha sido colectada por los pigmentos. En concreto, vimos que:
- Por cada vuelta de la fase acíclica se necesitaban 4 fotones de luz (dos por fotosistema). Se necesitan 12 vueltas para obtener los 12 NADPH necesarios, y también se obtienen 12 moléculas de ATP. En total 48 fotones.
- Por cada vuelta de la fase cíclica se necesitan solo 2 fotones (si recordáis, solo actúa el fotosistema I). Como hay que completar con 6 ATP se necesitan 6 vueltas. En total 12 fotones.
- La suma de los fotones necesarios para obtener una molécula de glucosa es, por tanto, de 60 fotones.
Sin embargo, estudios experimentales demuestran que el consumo real de energía es bastante mayor, correspondiendo con 180-240 fotones… ¡¡de 3 a 4 veces más de lo esperado!! Este rendimiento menor al esperado se debe a que, como en todo proceso natural, en la fotosíntesis hay numerosos factores que afectan al mismo. Veamos a continuación cuáles son los más importantes: 1. TEMPERATURA. Cada organismo tiene una fisiología adaptada al medio en el que vive, y esta adaptación llega al nivel enzimático. De esta manera, plantas que viven en ambientes cálidos tendrán complejos enzimáticos que trabajan mejor en rangos de temperaturas altas que otras que viven en el ártico, y su rendimiento fotosintético en ambientes más fríos al suyo, será menor.
2. FOTOPERIODO. Toda ruta metabólica puede sobrecargarse y necesita periodos de descanso para que se recupere su maquinaria enzimática y de metabolitos, y la fotosíntesis no es una excepción. Según la especie, se necesitan periodos de oscuridad más o menos largos para recuperarse y seguir produciendo materia orgánica con normalidad.
3. ESCASEZ DE AGUA. Cuando la sequedad o la temperatura son demasiado elevadas se cierran los estomas para evitar la deshidratación de la planta. Esto la protege de la pérdida de agua, pero produce una disminución de CO2 (que no puede entrar) y un aumento de O2 (que no puede salir), que la expone al fenómeno de la fotorrespiración, muy perjudicial para su rendimiento.
4. INTENSIDAD LUMINOSA. En principio a mayor intensidad luminosa, mayor rendimiento fotosintético… pero todo tiene su límite y en este caso se trata de la fotooxidación, la cual oxida de modo irreversible los pigmentos, que pierden su actividad y afectan enormemente al rendimiento de la planta.
5. COLOR DE LA LUZ. Según el medio en el que vivan, los organismos fotosintéticos tendrán diferentes proporciones de pigmentos fotosintéticos, permitiendo una más eficaz absorción de determinadas longitudes de onda (colores).
6. FOTORRESPIRACIÓN. Una elevada concentración de O2 respecto de la de CO2 desata una actividad oxidasa en la rubisco, que deja de fijar carbono, lo que disminuye enormemente el rendimiento fotosintético.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
23
3.9. QUIMIOSÍNTESIS
Llamamos quimiosíntesis al proceso que permite obtener energía química (ATP) a partir de la oxidación de sustancias inorgánicas. Los organismos que la realizan se llaman “quimioautótrofos”, y son todos bacterianos.
Al igual que en la fotosíntesis, el proceso quimiosintético se divide en DOS FASES:
Como la fase de carboxilación es prácticamente igual a la ya estudiada en fotosíntesis, vamos a decir algo más respecto a la cadena de electrones de la primera fase. El fundamento es el mismo que el visto en la fotofosforilación, ocurriendo una serie de oxidaciones-reducciones a lo largo de varios complejos enzimáticos de membrana que permiten acoplar un bombeo de protones desde el citosol hacia el espacio periplasmático. La diferencia capital es que los electrones que allí se excitaban en un complejo multienzimático capaz de absorber energía luminosa, aquí son tomados directamente, como electrones alto-energéticos, de determinadas moléculas donadoras inorgánicas y reducidas. Si tomamos como ejemplo el ácido sulfhídrico (SH2), este cede un par electrónico al complejo NADH óxido-reductasa de la membrana bacteriana y comienza la cadena de electrones, muy parecida a la de la respiración oxidativa de mitocondrias. El aceptor último de la cadena de electrones es el oxígeno, el cual queda reducido a agua.
Resumen de los factores que afectan a la fotosíntesis
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
24
El gradiente quimiosmótico generado con el bombeo de propones se resuelve con su paso a través de la (ya debe de ser famosísima ;) ATP-sintasa. Aprovecha la fuerza protón motriz para sintetizar ATP (fosforilación oxidativa). El NADH se obtiene gracias a un transporte inverso de electrones. En este, gastando parte del ATP producido en la fosforilación oxidativa, se fuerzan a los electrones a “subir” por la cadena electrónica anterior y llegar a la NADH óxido-reductasa. Esta enzima, en su actividad reductasa es capaz de ceder el par electrónico alto-energético al NAD+ sintetizando NADH.
TIPOS DE BACTERIAS QUIMIOSINTETICAS: Reciben un nombre u otro según el tipo de sustrato que utilizan para realizar la quimiosíntesis. Hay de 4 tipos: -Bacterias Nitrificantes. Producen ATP para fijar CO2 gracias a la energía desprendida en la oxidación de compuestos de nitrógeno. Ejemplos: Nitrosomas y Nitrobacter. - Nitrosomas. Aprovechan la energía desprendida en la reacción de oxidación del amoniaco a ácido nítroso.
2 NH3 + 3 02 2 N02H + 2 H20 + Energia - Nitrobacter. Usan la energía desprendida en la oxidación de nitroso a nítrico. 2 N02 + 02 2 N03H + Energía -Ferrobacterias. Utilizan la energía desprendida en la oxidación de carbonatos y sulfatos de hierro, es decir, oxidan compuestos que contienen hierro ferroso (Fe++), transformándolo en férrico (Fe+++). 4 FeC03 + 6 H20 + 02 4 Fe (OH)3 + 4 C02 + Energía -Sulfobacterias incoloras. Utilizan la energía desprendida en la oxidación del H2S que se transforma en ácido sulfúrico. H2S + 2 02 SO4H2 + Energía O bien oxidan el S hasta sulfato S + 2 O2 SO4
2- + Energía -Bacterias del hidrógeno. Obtienen energia directamente de la oxidación del hidrógeno atmosférico, prosuciendo agua. Absolutamente increíble. H2 + 1/2O2 H2O + Energía
3.8 INTEGRACIÓN DEL CATABOLISMO Y ANABOLISMO.
El anabolismo es la parte constructiva o de síntesis del metabolismo. La materia y energía obtenidos en los procesos catabólicos se utilizan en los procesos biosintéticos, es decir, en los procesos anabólicos.
En el catabolismo aerobio existen tres fases principales:
Durante la fase I, las macromoléculas se hidrolizan, hasta sus moléculas sillares (monómeros).
En la fase II, los distintos monómeros son transformados en el grupo acetilo del Acetil-CoA, con desprendimiento de ATP y NADH. Además, en la degradación de los aminoácidos se origina NH3, que es un producto final del catabolismo.
En la fase III el grupo acetilo del Acetil-CoA se incorpora al ciclo de Krebs, que es el camino terminal común, en él se oxida totalmente, dando finalmente CO2 y H20, produciéndose gran cantidad de NADH, que proporciona mucho ATP a través de la cadena de transporte electrónico o cadena respiratoria.
-
Efi-Ciencia Red
https://www.youtube.com/user/EfiCienciaRed https://mferminprofe.jimdo.com
https://www.facebook.com/eficiencia.red
25
Las rutas catabólicas son convergentes, pues confluyen hacia unos pocos productos finales. El anabolismo también sucede en tres fases, comenzando con moléculas precursoras pequeñas. Comienza en la fase III a partir de los pequeños compuestos originados en la tercera fase del catabolismo.
En la fase II se forman los monómeros que se reunirán en la fase I para originar polímeros. Las rutas anabólicas son divergentes, ya que a partir de unas pocas moléculas precursoras sencillas se sintetizan una gran variedad de macromoléculas. El catabolismo y el anabolismo suceden simultáneamente y son interdependientes; pero las etapas que originan la degradación de un producto y su síntesis no suelen ser idénticas, a causa de diferentes razones:
Algunas reacciones catabólicas son irreversibles.
Las rutas catabólicas y anabólicas pueden estar localizadas en diferentes orgánulos.
La regulación de las rutas catabólicas y anabólicas puede ser diferente.
CICLO DE
CALVIN
FASE I
FASE II
FASE III
ESQUEMA RESUMEN DEL METABOLISMO
ANABOLISMO (Divergente)
CATABOLISMO (Convergente)