TESIS DOCTORAL - hera.ugr.es · tejido adyacente o del líquido articular. Cada placa o lámina de...
186
FACULTAD DE MEDICINA UNIVERSIDAD DE GRANADA IDENTIFICACIÓN DE PATRONES DE VIABILIDAD Y EXPRESIÓN GÉNICA EN CONDROCITOS ARTICULARES HUMANOS PARA SU UTILIZACIÓN EN INGENIERÍA TISULAR Álvaro Morales Villaescusa GRANADA 2010 TESIS DOCTORAL
Transcript of TESIS DOCTORAL - hera.ugr.es · tejido adyacente o del líquido articular. Cada placa o lámina de...

FACULTAD DE MEDICINA
FACULTAD DE MEDICINA
UNIVERSIDAD DE GRANADA
UNIVERSIDAD DE GRANADA
TESISDOCTORAL
IDENTIFICACIÓN DE PATRONES DE VIABILIDAD Y EXPRESIÓN GÉNICA EN CONDROCITOS ARTICULARES HUMANOS PARA SU UTILIZACIÓN EN INGENIERÍA TISULAR
Álvaro Morales Villaescusa GRANADA 2010
TESIS DOCTORAL
Álvaro Morales Villaescusa

Editor: Editorial de la Universidad de GranadaAutor: Álvaro Morales VillaescusaD.L.: GR 3800-2010ISBN: 978-84-693-6028-6

1
UNIVERSIDAD DE GRANADA
FACULTAD DE MEDICINA
IDENTIFICACIÓN DE PATRONES
DE VIABILIDAD Y EXPRESIÓN GÉNICA
EN CONDROCITOS ARTICULARES
HUMANOS PARA SU UTILIZACIÓN
EN INGENIERÍA TISULAR
TESIS DOCTORAL
ÁLVARO MORALES VILLAESCUSA
2010

2
UNIVERSIDAD DE GRANADA
FACULTAD DE MEDICINA
Departamento de Histología
Grupo de Investigación de Ingeniería Tisular CTS - 115
“IDENTIFICACIÓN DE PATRONES DE VIABILIDAD Y EXPRESIÓN GÉNICA
EN CONDROCITOS ARTICULARES HUMANOS PARA SU UTILIZACIÓN EN
INGENIERÍA TISULAR”
Esta Tesis Doctoral ha sido realizada en los laboratorios del Grupo de Ingeniería
Tisular del Departamento de Histología de la Universidad de Granada y financiada
por el Proyecto de Excelencia titulado “ELABORACIÓN DE ÓRGANOS Y TEJIDOS
ARTIFICIALES PARA SU APLICACIÓN TERAPEUTICA” – P06-CTS-2191 de la
Junta de Andalucía
Granada, a 1 de Junio de 2010.

3
IDENTIFICACIÓN DE PATRONES DE VIABILIDAD Y EXPRESIÓN GÉNICA
EN CONDROCITOS ARTICULARES HUMANOS PARA SU UTILIZACIÓN EN
INGENIERÍA TISULAR
Memoria que presenta el Licenciado en Medicina y Cirugía
Álvaro Morales Villaescusa
para aspirar al título de Doctor
Fdo.: Álvaro Morales Villaescusa
VºBº El Director de Tesis VºBº El Director de Tesis
Fdo.: Prof. Antonio Campos Muñoz Fdo.: Prof. Miguel Alaminos Mingorance
Doctor en Medicina y Cirugía Doctor en Medicina y Cirugía
Universidad de Granada Doctor en Ciencias Biológicas
Universidad de Granada
VºBº El Director de la Tesis
Fdo.: Prof.ª María del Carmen Sánchez Quevedo
Doctora en Ciencias Químicas
Universidad de Granada
Departamento de Histología
Universidad de Granada
2010

4
A toda mi familia.

5
“La habilidad es lo que permite hacer ciertas cosas.
La motivación determina lo que se hace. La actitud cuán bien se hace.”
Lou Holtz

6
AGRADECIMIENTOS
Suponen para mí estas líneas, una excelente oportunidad de poner de manifiesto mi
gratitud hacia el Profesor Campos. Alma incansable de este trabajo, representa desde
hace más de una década, una de las personas más influyentes en mi vida académica y
profesional. Ejemplo de iniciativa, capacidad de trabajo, dedicación y optimismo, ha sido
el ideólogo de casi todos los proyectos que he iniciado en los últimos años, que se han
apartado del convencionalismo que muchas veces seguimos los profesionales de la
medicina. Gracias Antonio, por el constante estímulo, por la ilusión transmitida, por el
cariño expresado, por la ausencia de desfallecimiento… en definitiva, gracias por estar
ahí una, tras otra, tras otra vez.
Ha sido para mí un auténtico privilegio, contar con la ayuda del Profesor Alaminos. Es
alentador comprobar que aún existe gente como él, con esa vocación por la
investigación y por la docencia. Gracias Miguel, por tus enseñanzas, por tu tiempo, por
tu paciencia y por tu sentido del humor, auténticamente exquisito, sin tí este trabajo no
habría sido posible.
Quiero además expresar mi más sincera gratitud a la Profesora Sánchez Quevedo, por
el tiempo y el esfuerzo con tanto cariño dedicados a este trabajo, así como a Ingrid y a
Renato por su incansable entrega.
Lugar privilegiado ocupa mi familia:
No puedo expresar con palabras la gratitud y admiración que siento por mis padres, a
quienes les debo mis bienes más preciados: mi educación y mi formación. Gracias, por
transmitirme los valores que hoy me sostienen: el sentido del amor, del trabajo, de la
lucha y de la honestidad. Lograsteis la difícil tarea de alcanzar el equilibrio entre la
libertad y la obligación. Lo habeis hecho muy bien.
A mi mujer, Magdalena, por constituir para mí un aliento de ánimo inesperado en la
consecución de este trabajo y por haber conseguido que cada día mire dentro de mí
para exigirme ser mejor persona. Gracias por sostener el proyecto más importante de mi
vida, a tu lado todo es muy fácil.
No quiero concluir sin mencionar a todas aquellas personas que de una forma u otra
han colaborado conmigo para que este trabajo salga adelante, tanto del Hospital de
Jerez de la Frontera, en especial del Servicio de Traumatología y del Centro Regional
de Transfusiones Sanguíneas, como del Hospital Universitario Virgen de las Nieves y
del Departamento de Histología de la Universidad de Granada.

INTRODUCCIÓN ………………………………………………………. 9
1.-Estructura histológica del tejido cartilaginoso ………………………………. 14
1.1.-Características histológicas cartílago articular ……………………..……… 17
1.2.-Histogénesis del cartílago …………………………………….……………… 24
1.3.-Metabolismo del cartílago articular …………………..…………...………… 25
1.4.-Evolución del cartílago con la edad …………………………….…………… 26
2.- Patología del cartílago articular ……………………………………………….. 27
3.- Tratamiento de la patología del cartílago articular ……………………….… 29
3.1.- Tratamiento sintomático ……………………………………………………... 29
3.2.- Tratamiento quirúrgico clásico ………….......……………………………… 29
3.3.- Inducción celular de la condrogénesis …………………………………….. 31
3.4.- Transplante osteocondral ……....………………………….…………………. 32
4.- Terapia celular con condrocitos ……………………………………………… 35
5.- Viabilidad celular ………………………………………………………………….. 42
5.1. Detección de alteraciones de la permeabilidad celular …………………… 44
5.2. Microanálisis por energía dispersiva de rayos X ………………..……...…... 45
5.3. Ensayos morfológicos …………………………………………….…………… 45
5.4. Determinación del perfil de expresión génica mediante microarrays …….. 46
OBJETIVOS ………………………………....…………………………… 48
ÍNDICE

MATERIALES Y MÉTODOS …………………………………………… 50
1.- Aislamiento de condrocitos ……………...……………………………………… 51
2.- Obtención de subcultivos celulares …………..........…………………………… 54
3.- Determinación de la viabilidad celular mediante ensayos
de exclusión del colorante vital azul tripán ………………………....…………… 55
4.- Análisis de la expresión génica mediante microarrays …………....……… 58
5.- Análisis estadístico ……….........……………………………………………… 59
RESULTADOS …………………………………………………………… 60
1.- Establecimiento de cultivos primarios
de condrocitos de cartílago articular humano ………………...……………….. 61
2.- Viabilidad celular determinada mediante
ensayos de exclusión de colorantes vitales ……………………………………. 63
3.- Patrón de expresión génica global de los condrocitos
articulares humanos determinado mediante microarray ……………………… 65
4.- Identificación de los genes cuya expresión
se relaciona con la evolución cronológica de los cultivos ……………………. 74
5.- Identificación del patrón de expresión de los genes relacionados
con la diferenciación condral en los ocho primeros subcultivos celulares ….... 75
6.- Identificación del patrón de expresión de genes vinculados con la
mortalidad celular (caspasas) en los ocho subcultivos celulares ……...…... 77
DISCUSIÓN ………………………….………………………………… 83
CONCLUSIONES ……………………..………………………………. 95
TABLAS SUPLEMENTARIAS ………………………………………… 98
REFERENCIAS BIBLIOGRÁFICAS ………………………………. 171

INTRODUCCIÓN
OBJETIVOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
TABLAS SUPLEMENTARIAS
REFERENCIAS BIBLIOGRÁFICAS

Introducción
10
El aparato locomotor permite al ser humano interactuar con el medio que le
rodea mediante el movimiento. Está constituido por huesos, músculos y articulaciones.
Se denomina articulación (Figura 1) a la unidad estructural y funcional existente entre
dos o más huesos, que posibilita su desplazamiento espacial, manteniendo
simultáneamente una estabilidad adecuada (Llusá et al., 2007). La articulación,
previene movimientos excesivos gracias a la congruencia entre las superficies
articulares (revestidas de cartílago), la cápsula, los ligamentos y las estructuras
músculo-tendinosas vecinas.
El cartílago articular es uno de los tres tipos de cartílago que podemos encontrar en el
organismo. Es la estructura que recubre los huesos en su porción epifisaria, facilitando
el deslizamiento de los mismos y amortiguando las presiones mecánicas. Tiene un
aspecto blanco-amarillento, brillante que resulta homogéneo al corte. Su grosor se
relaciona con la presión que se ejerce sobre su superficie, de forma que oscila entre
los 2-4 mm en cadera y rodilla y los 6-7 mm en la rótula.
Las lesiones del cartílago articular son de gran importancia dentro del campo de la
cirugía ortopédica y de la traumatología, ya que, cuando se producen, dan lugar a
cuadros degenerativos progresivos que culminan en la degradación del cartílago y
destrucción de la superficie articular donde se encuentra la lesión. Esto se debe, a que
el cartílago hialino, pese a ser un tejido metabólicamente activo posee una limitada
capacidad de reparación, por lo que estos cuadros suelen requerir de un

Introducción
11
reemplazamiento total o parcial de la articulación para eliminar el dolor y restaurar la
movilidad (artroplastia).
La incidencia y prevalencia real de las lesiones del cartílago hialino es desconocida.
Ello puede deberse, entre otros motivos, a que dichas lesiones pueden presentarse
directa o indirectamente a partir de otros daños producidos en la rodilla meses o años
después de ocurrir la lesión primaria.
Las lesiones condrales han sido históricamente un problema de muy difícil solución.
William Hunter afirmaba en 1743 que ―desde Hipócrates hasta nuestros días, se
acepta universalmente que el cartílago ulcerado es un asunto problemático y que una
vez destruido no se repara...‖. Un siglo después, en 1851, Sir James Paget decía que,
hasta donde él conocía, ―no existen ocasiones en las que un fragmento perdido del
cartílago haya sido repuesto o reparado con cartílago permanente nuevo y bien
desarrollado en el ser humano‖.
Desde entonces, numerosos estudios han confirmado que el cartílago tiene una
capacidad limitada de reparación directa (tiene capacidad para formar un tejido fibroso
o fibrocartilaginoso) y que su función mecánica no se recupera espontáneamente tras
una lesión importante (Martín, 2003).
Inicialmente, el tratamiento de las lesiones del cartílago articular, es sintomático:
reposo, cambios en el estilo de vida, rehabilitación o tratamientos médicos
farmacológicos. En los últimos años, se han desarrollado diversos tratamientos
quirúrgicos para la resolución de las lesiones del cartílago articular. El objetivo de
estos tratamientos es prevenir la expansión de las lesiones e intentar regenerar el
cartílago para evitar en un futuro la evolución a osteoartrosis obligando a un posible
recambio articular. Estas técnicas se pueden dividir en varias categorías: tratamiento
sintomático (lavado y/o desbridamiento artroscópico), tratamiento quirúrgico clásico
(perforaciones, micro fracturas, abrasiones superficiales…), inducción celular de la
condrogénesis (trasplante de periostio, trasplante de pericondrio y trasplante autólogo
de condrocitos), trasplante osteocondral (aloinjertos y autoinjertos osteocondrales,
etc.).
Las nuevas investigaciones buscan entender la biología, composición, metabolismo,
organización ultraestructural y molecular, y las propiedades biomecánicas del cartílago
articular con la esperanza de desarrollar un procedimiento de reparación biológico,
como una alternativa a las artroplastias en el tratamiento de las enfermedades
degenerativas de las articulaciones.

Introducción
12
El objetivo es intentar hallar un sustituto biológico viable, con las propiedades
bioquímicas y biomecánicas del cartílago articular normal. El cartílago articular al no
tener una capacidad de curación adecuada, generalmente y en las mejores
condiciones realiza esta curación con fibrocartílago, sin embargo, algunos estudios
experimentales han demostrado regeneración del cartílago hialino.
En la presente Tesis Doctoral vamos a considerar la viabilidad de los condrocitos
articulares humanos mantenidos en cultivo para su utilización en la terapia celular,
para ello describiremos y analizaremos en la presente introducción, como fundamento
de la investigación a realizar, los siguientes apartados:
Estructura histológica del cartílago.
Patología del cartílago articular.
Tratamiento de la patología del cartílago articular
Terapia celular con condrocitos.
Viabilidad celular.
A continuación enunciaremos los objetivos del presente trabajo de investigación y
describiremos el material y los métodos utilizados y los resultados obtenidos.
Finalmente enumeraremos las conclusiones alcanzadas en nuestro estudio.

Introducción
13
Figura 1.- Representación esquemática de la estructura básica de una articulación.

Introducción
14
1.- ESTRUCTURA HISTOLÓGICA DEL TEJIDO CARTILAGINOSO
El tejido cartilaginoso es un tipo de tejido conjuntivo especializado, constituido por
células y matriz intercelular.
Las células, denominadas condrocitos, se organizan en espacios llamados lagunas o
condroceles, rodeados por abundante matriz extracelular. Los condrocitos sintetizan y
secretan los componentes orgánicos de la matriz extracelular que son básicamente
colágeno, ácido hialurónico, proteoglucanos y glicoproteínas. Según las características
de la matriz se distingue cartílago hialino, elástico y fibroso. Los vasos sanguíneos no
penetran en la matriz cartilaginosa (el cartílago es avascular y aneuronal) y los
condrocitos se nutren con material que difunde desde de los capilares sanguíneos del
tejido adyacente o del líquido articular.
Cada placa o lámina de tejido cartilaginoso está rodeada por el pericondrio que
corresponde a tejido conjuntivo denso, en el cuál se distingue una capa externa fibrosa
y una capa interna celular, en la cual se ubican las células que pueden dar origen a los
condroblastos, que corresponden a precursores de los condrocitos y que difieren de
ellos sólo en su edad y en su mayor actividad de síntesis de componentes de la matriz
intercelular cartilaginosa. La superficie articular no está revestida por pericondrio.
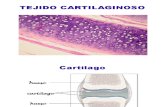
Existen tres tipos de tejido cartilaginoso (Finn, 2002): el hialino, que es el más
abundante en el cuerpo humano (Figura 2), el elástico y el fibroso (Figuras 3 y 4). El
cartílago hialino se encuentra en los adultos en las superficies articulares, en los
anillos traqueales, en la laringe y en los extremos anteriores de las costillas. El
cartílago elástico se localiza en diversas estructuras del aparato auditivo y en la
epiglotis. En general es más amarillento y elástico que el cartílago hialino. El cartílago
fibroso se encuentra en los discos intervertebrales, en la sínfisis pubiana, en el
ligamento redondo del fémur y en los lugares de inserción de algunos tendones.
Las propiedades especiales de los distintos tipos de cartílago, permiten que estos se
adapten a sus funciones específicas. Así, el cartílago articular permite el movimiento
de los extremos articulares de los huesos sin que apenas existan fricciones entre ellos,
a la vez que, por ser un gel rígido, absorbe la energía de los golpes a modo de
amortiguador. Se cree que esta última función se debe al elevado contenido de agua
en los dominios de los proteoglicanos. Ante la presión se eliminan las moléculas de
agua de los dominios, para volver cuando termina el efecto de la carga.

Introducción
15
En lo que respecta a la regeneración, crecimiento y proliferación activa del cartílago,
este se limita a los primeros años de vida.
Figura 2.- Imagen de microscopía óptica de cartílago hialino. (Rosales R, 2008).
Figura 3.- Imágenes de microscopía óptica de cartílago elástico. http://escuela.med.puc.cl/paginas/cursos/segundo/histologia/HistologiaWeb/paginas/co25306.html

Introducción
16
Figura 4.- Imágenes de microscopía óptica de cartílago fibroso.
http://escuela.med.puc.cl/paginas/cursos/segundo/histologia/HistologiaWeb/paginas/co25505.html

Introducción
17
1.1.- Característica histológicas del cartílago articular
El término hialino, del griego hyalos, vidrio, hace referencia al aspecto blanco perlado y
translúcido, semejante a vidrio que presenta este tejido (Lesson et al., 1989). Es el
más abundante del organismo, por lo que suele tomarse como ejemplo general de
cartílago (Mow et al., 1984; Kuettner et al., 1987). En el individuo adulto, como ya se
ha comentado anteriormente, se encuentra en las superficies articulares, en los anillos
traqueales y bronquios, en la laringe, en la nariz y en los extremos anteriores de las
costillas. Como todos los tipos de cartílago, está constituido por agua, células
(condrocitos), y matriz extracelular (Figura 5).
a.-) Condrocitos.
Los condrocitos se localizan en las lagunas que existen en la matriz extracelular
(Figura 6). Constituyen el 2% de la masa del cartílago. Son grandes, con un diámetro
que puede alcanzar las 40 μm y van modificando su disposición en el espesor del
tejido, de la superficie a la profundidad, de forma que se diferencian cuatro zonas, que
van de la uno que es la más superficial, a la cuatro que es la más profunda. Así los
condrocitos más inmaduros, llamados condroblastos, y cercanos al pericondrio, se
ubican en lagunas ovales, aplanadas paralelas a la superficie, mientras que los
condrocitos más maduros, ubicados en la profundidad del cartílago se disponen en
lagunas más redondeadas. En el cartílago fetal las células son a menudo aplanadas y
rara vez se observan nidos celulares. Los condrocitos sufren un considerable grado de
retracción durante la preparación, por lo que rara vez se adaptan a la forma de las
lagunas.
Su núcleo es redondeado y se observan uno o varios nucleolos. Las organelas
citoplasmáticas de los condrocitos son semejantes a las que se encuentran en los
fibroblastos, que también sintetizan matriz extracelular. Hay un abundante retículo
endoplásmico rugoso y un notable complejo de Golgi. El tamaño del aparato de Golgi y
las características del retículo endoplásmico varían si el cartílago está o no en
crecimiento activo. Durante el crecimiento el aparato de Golgi se dilata y el retículo
endoplásmico presenta cisternas dilatadas. Cuando el cartílago no está en crecimiento
activo el retículo endoplásmico y el aparato de Golgi no están tan desarrollados. El
citoplasma de las células cartilaginosas es basófilo y puede ser vacuolado, también
encontramos gotitas lipídicas, glucógeno, mitocondrias y pigmentos. Las vesículas
secretoras se relacionan con la región del aparato de Golgi y secretan material a la
matriz circundante. Las células cartilaginosas maduras se encuentran en general en el

Introducción
18
centro del espesor del cartílago y presentan un retículo endoplásmico rugoso menos
abundante, un aparato de Golgi menos desarrollado y grandes acúmulos de glucógeno
citoplasmático (Lesson, 1989).
Durante el proceso de diferenciación a condrocito maduro, la basofilia se torna
gradualmente en acidofilia y se retrae el retículo endoplásmico rugoso.
Los condrocitos sintetizan, entre otras moléculas, la condronectina. Se trata de una
glucoproteína del mismo tipo que la fibronectina que fija los condrocitos a la matriz
extracelular gracias a la existencia de unos sitios de unión específicos para ello (Finn,
2002).
Cerca de la superficie articular los condrocitos presentan morfología alargada en
sección longitudinal y discoide en sección transversal; a este nivel, son menos activos
que los que se localizan en capas profundas. A mayor profundidad, los condrocitos
son menos numerosos, de mayor tamaño, más redondeados y con mayor actividad
metabólica.
Los condrocitos vivos aislados in Vitro exhiben un movimiento ameboide, y cambian
constantemente de forma mediante la emisión de pseudópodos.
b.-) Matriz extracelular.
La matriz cartilaginosa es la responsable de las características mecánicas del
cartílago. Es un material denso, que se sitúa entre las lagunas de condrocitos
constituido principalmente por agua, colágeno y sustancia fundamental
(proteoglucanos). Microscópicamente, la matriz extracelular parece carecer de
estructura, puesto que el colágeno aparece bajo la forma de finas fibrillas que tienen el
índice de refracción muy similar al de la sustancia fundamental, se dice que las fibrillas
están enmascaradas. Se pueden observar mediante microscopía electrónica.
En los preparados teñidos con hematoxilina-eosina se observa cómo la matriz es
acidófila en la zona superficial (condrocitos inmaduros) y cómo se va haciendo más
basófila en las regiones más profundas (condrocitos más maduros). Alrededor de cada
grupo isogénico, se observa una basofilia muy marcada, denominada matriz territorial.
La matriz interterritorial es la que se localiza entre las distintas zonas de matriz
territorial (Figura 7).
La sustancia fundamental adopta una coloración metacromática intensa. En su mayor
parte se compone de proteoglucanos, cuyos glucosaminoglucanos principales son el
condroitinsulfato y el queratansulfato. La basofilia se debe a los grupos sulfatos, que

Introducción
19
son muy ácidos, mientras que la basofilia de la matriz territorial se debe al mayor
contenido de proteoglucanos sulfatados en esta zona. Los proteoglucanos también
conforman agregados por unión con hialurónico (Finn, 2002). Los componentes más
importantes de la matriz extracelular son los siguientes:
Agua. Constituye el componente más abundante del cartílago (60-80%).
Permite la deformación del cartílago en respuesta al estrés, fluyendo
dentro y fuera del mismo. Su cantidad es mayor en superficie (80%) que
en profundidad (60%). Es muy importante para la nutrición del cartílago
y para la lubricación articular.
Proteoglucanos. Los proteoglucanos son complejos proteino-sacáridos
formados por cadenas de glucosaminoglucanos unidas a un filamento
proteico central mediante enlaces covalentes. Constituyen el segundo
componente más abundante del cartílago articular (5-10% del peso
total) (Maroudas, 1979; Muir, 1983; Poole, 1986). Son los responsables
de la resistencia a la compresión del cartílago. Los
glucosaminoglucanos representan el 95% de los proteoglucanos y son
básicamente el condroitínsulfato y el queratánsulfato. El
glucosaminoglucano más frecuente es el condroitínsulfato (del que hay
2 subtipos, el condroitín-4-sulfato y el condroitín-6-sulfato) (Fig. 8),
después el queratánsulfato y el dermatánsulfato. El condroitín-4-sulfato
es el más abundante y disminuye en cantidad con los años, el
condroitín-6-sulfato se mantiene constante y el queratansulfato aumenta
con la edad.
Se cree que los proteoglucanos se combinan con cationes y por tanto
desempeñan un papel importante en el transporte de agua y electrolitos
dentro de la matriz (Lesson, 1989). Los proteoglucanos son sintetizados
y excretados al exterior por los condrocitos, previamente a su excreción,
a los proteoglucanos se les acopla gran cantidad de polisacáridos por
glucosilación ligada al aparato de Golgi (Figura 9).
Colágeno. Las fibras de colágeno representan más del 40% del peso
seco de la matriz cartilaginosa. Mayoritariamente del tipo II (90-95%), lo
que confiere al cartílago una gran resistencia a la tensión. Las fibrillas
de colágeno tipo II son muy delgadas y forman un reticulado
especialmente denso alrededor de las lagunas de condrocitos. También

Introducción
20
se observan pequeñas cantidades de colágeno tipo I, V, VI, IX, X y XI.
El colágeno tipo VI se encuentra en fases precoces de osteoartritis. El
colágeno X se produce únicamente durante fases de osificación
endocondral (se asocia por lo tanto a calcificación del cartílago).
En el cartílago articular se pueden diferenciar las siguientes capas (Figura 10)
(Buckwalter JA, 2007):
a. Capa superficial o de deslizamiento (40 μm). Escasa actividad metabólica.
Presenta pocos proteoglucanos y una elevada concentración de fibras de colágeno
distribuidas perpendicularmente entre sí y paralelas a la superficie. Soporta las fuerzas
de cizallamiento. Los condrocitos presentes están aplanados y paralelos a la
superficie.
b. Capa intermedia o de transición (500 μm). Elevada actividad metabólica. Mayor
presencia de proteoglucanos y menos colágeno, dispuesto oblicuamente y al azar.
Soporta fuerzas de compresión y los condrocitos presentes son más redondeados.
c. Capa profunda o radial (1000 μm). Rica en proteoglucanos y fibras de colágeno.
Éste se distribuye radialmente formando arcos. Células redondeadas formando
columnas. Soporta fuerzas de compresión.
d. Capa calcificada (300 μm). No presenta proteoglucanos. Colágeno dispuesto
radialmente. Presenta cristales de hidroxiapatita y está adyacente al hueso
subcondral. Es la capa de anclaje del cartílago al hueso. La celularidad es escasa.
Está separada de la anterior por la ―línea de flujo‖ (tidemark), de 5 μm de grosor, que
es una barrera ondulada con disposición tangencial de sus fibras y que permite
soportar fuerzas de cizallamiento.

Introducción
21
Figura 5.- Corte esquemático de una porción de cartílago hialino (Buckwalter JA, 1995).
Figura 6.- Esquema que representa células cartilaginosas rodeadas de matriz extracelular.
http://www.bartleby.com/107/illus292.html

Introducción
22
Figura 7.- Representación gráfica de la matriz territorial e interterritorial.
http://escuela.med.puc.cl/paginas/cursos/segundo/histologia/HistologiaWeb/paginas/co24947.html
Figura 8.- Representación esquemática de un proteoglucano.

Introducción
23
Figura 9.- Los condrocitos sintetizan el colágeno y los proteoglicanos. Ambos interactúan
conformando la matriz extracelular del cartílago, capaz de retener grandes cantidades de agua
(Buckwalter JA, 1995).
Figura 10.- Capas del cartílago articular (Buckwalter JA, 1995).

Introducción
24
1.2.- Histogénesis del cartílago
El tejido cartilaginoso se origina en el mesénquima, a partir de células mesenquimales
que se redondean y agrupan en conglomerados con escaso material intercelular entre
ellas. Este conjunto de células precartilaginosas se denomina blastema. Las células
del blastema son inducidas a sintetizar matriz cartilaginosa y a partir de ese momento
se les llama condroblastos, ellas se separan progresivamente a medida que aumenta
la cantidad de matriz sintetizada y pasan a llamarse condrocitos. El tejido
mesenquimal que rodea a la masa condrogénica pasará a constituir el pericondrio.
En cuanto se forman las primeras células cartilaginosas a partir del mesénquima, se
produce el consiguiente crecimiento del cartílago de dos maneras:
- Crecimiento intersticial: En el centro de condrificación se produce el
crecimiento por divisiones mitóticas de las células cartilaginosas a partir de
condroblastos, e inmediatamente después de la mitosis, las células hijas
producen una delgada pared de matriz extracelular. Tras una nueva división de
las células hijas, se forma un pequeño grupo de cuatro células, que a su vez
se pueden dividir. Cada uno de los pequeños grupos formados, contiene
células derivadas de un único condroblasto, por lo que se denominan grupos
isogénicos.
- Crecimiento aposicional: Ocurre desde el pericondrio, en cuya capa celular se
localizan células indiferenciadas capaces de dividirse dando origen a células
que se diferenciaran a condroblastos y que producirán tejido cartilaginoso
sobre la superficie del cartílago preexistente, quedando los condroblastos
atrapados en la matriz que producen y pasando a ser condrocitos. Durante
toda la vida fetal, y a velocidad más reducida, durante la infancia y la pubertad,
se produce un crecimiento aposicional continuo desde el pericondrio. Las
lagunas ubicadas inmediatamente por debajo del pericondrio contienen células
cartilaginosas recién formadas. El crecimiento intersticial sólo tiene lugar en el
cartílago joven (Finn, 2002).

Introducción
25
1.3.- Metabolismo del cartílago articular
Como tejido avascular, el cartílago se nutre fundamentalmente a partir del líquido
sinovial, proceso en el que está muy implicado el mecanismo de lubricación articular,
aunque la capa más profunda (1/3 aproximadamente en cuanto a grosor) se puede
nutrir a partir de la vascularización epifisaria.
El cartílago articular es una estructura en constante actividad (Sopena JJ, 2004).
Destacan:
La síntesis del colágeno y de su catabolismo.
La síntesis y degradación de los proteoglicanos.
La acción de diversos factores de crecimiento que regulan la síntesis de
cartílago articular:
o Factor de Crecimiento Derivado de Plaquetas (PDGF); implicado
en la curación de laceraciones en el cartílago.
o Factor de Crecimiento Transformante beta (TGF-β); que
estimula la síntesis de PGs, si bien frena la del colágeno II. Así
mismo, estimula la formación de inhibidores de plasmina y
estromelisina.
o Factor de Crecimiento Fibroblástico (básico) (b-FGF); que
estimula la síntesis de ADN en condrocitos adultos.
o Factor de Crecimiento Análogo a la Insulina (IGF-I); que
estimula la síntesis de ADN y MEC en el cartílago adulto y en la
placa de crecimiento.

Introducción
26
1.4.- Evolución del cartílago con la edad
Con el paso del tiempo, y como consecuencia del desgaste, aparecen las lesiones
artrósicas, que son de tipo degenerativo. Son el resultado de fenómenos mecánicos
(sobrecarga) y biológicos (resistencia del cartílago) que desestabilizan el equilibrio
entre la síntesis y la degradación. Según la influencia preponderante de uno u otro
factor, se trata más bien de artrosis mecánica o de artrosis estructural. Unas
anomalías genéticas del colágeno tipo II pueden estar en el origen de artrosis
familiares excepcionales. Las lesiones artrósicas son lesiones de movimiento en
espejo sobre las dos vertientes articulares; son evolutivas y se extienden al conjunto
de las estructuras de la articulación. El envejecimiento cartilaginoso es un factor
predisponente.
Con el envejecimiento, se producen una serie de cambios en el cartílago que en líneas
generales son los siguientes:
Los condrocitos se agrandan, pierden capacidad de reproducción, es
decir, el cartílago se vuelve hipocelular.
Disminuyen los PTGs en cantidad y tamaño y cambian su proporción
(disminuye el condroitín-4-sulfato y aumenta el queratán-sulfato).
Aumenta el contenido en proteínas y disminuye el agua. Todo esto se
traduce en una disminución de la elasticidad y aumento de la rigidez del
cartílago articular.

Introducción
27
2.- PATOLOGÍA DEL CARTÍLAGO ARTICULAR
Hay que diferenciar entre las lesiones que afectan exclusivamente al cartílago
(lesiones parciales) que no suelen repararse espontáneamente, y las que alcanzan el
hueso subcondral hasta la cavidad medular del hueso (lesiones que afectan a todo el
espesor) que sí lo hacen, aunque esta reparación no restablece ni la morfología ni la
función normal del cartílago. Además de la importancia de la profundidad de las
lesiones, el diámetro de las mismas parece ser también un factor determinante en la
reparación.
El cartílago que se repara espontáneamente lo hace de manera anómala, con la
formación de un tipo de colágeno inapropiado (tipo I), un bajo contenido en
proteoglucanos y una disposición anormal de las células. Estas razones, entre otras,
pueden ser la causa de que el nuevo cartílago formado tenga unas propiedades
mecánicas inferiores al original (Messner, 2004).
Existen diversos mecanismos capaces de provocar una alteración en las propiedades
histoquímicas y por lo tanto mecánicas del cartílago, como son: la inmovilización, la
compresión, las sobrecargas de la articulación, los traumatismos directos e indirectos,
únicos o repetidos, la desorganización en los componentes articulares o las
inestabilidades provocadas por alteraciones en alguna de las estructuras
estabilizadoras de la rodilla. Entre las causas responsables de originar lesiones en el
cartílago articular, se reconoce en primer lugar el daño traumático seguido de la
osteocondritis disecante (Finn, 2002).
Hay varias clasificaciones para valorar la gravedad de las lesiones del cartílago
articular. Se basan principalmente en la visualización del defecto, por lo que puede
existir gran variabilidad interobservador. De las clasificaciones existentes en la
literatura, la establecida por Outerbridge en 1961 es la más empleada. En ella se
establecen cuatro grados (Figura 11):
Grado I: Reblandecimiento e inflamación (o edema) del cartílago.
Grado II: Fragmentación y fisuración en zonas ≤ 1,25 cm de diámetro.
Grado III: Fragmentación y fisuración en zonas > de 1,25 cm de diámetro.
Grado IV: Erosión del cartílago que llega hasta hueso subcondral.
Recientemente, la International Cartilage Repair Society (ICRS), ha propuesto
una nueva escala de valoración y descripción del daño de la superficie del cartílago

Introducción
28
articular, en la que se incluye la osteocondritis disecante, con el fin de crear un
lenguaje más universal para la comunicación y difusión de los avances en patología
del cartílago.
El cartílago articular es el único tejido sin aporte vascular, nervioso ni linfático,
propiedades que pueden condicionar su baja capacidad intrínseca de curación. No
existe respuesta inflamatoria al daño tisular y por lo tanto no se producirá la invasión
macrofágica para fagocitar y eliminar el tejido desvitalizado ni la migración de células
con capacidad reparadora dentro del área lesionada. Las lesiones condrales no se
solventan por sí mismas y con el tiempo pueden progresar a osteoartrosis.
La resolución más adecuada de una lesión condral debería implicar la regeneración
con un tejido idéntico al cartílago hialino. La simple reparación supone el relleno con
un tejido no idéntico que debería ser capaz de sellar el área defectuosa con una buena
adhesión al hueso subcondral y una completa integración con el cartílago circundante
así como de resistir el desgaste mecánico con el paso del tiempo e incluirse
gradualmente en el recambio natural del tejido normal.
Se observan también lesiones cartilaginosas secundarias en las necrosis epifisarias,
en los pacientes de más edad. La localización a nivel del cóndilo femoral, y más
raramente del platillo tibial, son las más clásicas. La osteocondritis con evolución
disecante, tipo König, se observa en el paciente joven, con dos localizaciones
preferenciales, el cóndilo femoral y el astrágalo. Su patogenia es, frecuentemente,
microtraumática y las lesiones cartilaginosas secundarias.
El tercer grupo de lesiones cartilaginosas es consecuencia de patología sinovial
(séptica, inflamatoria, metabólica). Las precipitaciones cálcicas intraarticulares
(hidroxiapatita, pirofosfato: condrocalcinosis) pueden también generar o agravar las
lesiones cartilaginosas. El ataque cartilaginoso es la regla en las formas tardías de la
osteocondromatosis sinovial. Las lesiones cartilaginosas secundarias a un ataque
sinovial tienen un tratamiento específico. Las sinovectomías químicas, físicas o
quirúrgicas pueden ser propuestas para proteger el cartílago.
Actualmente, existen diversas técnicas quirúrgicas desarrolladas en los últimos años,
para la resolución de las lesiones del cartílago articular. El objetivo de estas
intervenciones es prevenir la expansión de las lesiones e intentar regenerar el
cartílago para evitar en un futuro la evolución a osteoartrosis o un posible recambio
articular.

Introducción
29
3.- TRATAMIENTO DE LA PATOLOGÍA DEL CARTILAGO ARTICULAR
3.1.- Tratamiento sintomático:
El objetivo es disminuir los síntomas clínicos: se utiliza el lavado y el desbridamiento
artroscópico.
- El lavado artroscópico es una de las técnicas más básicas y más empleadas. Permite
el ―lavado‖ del líquido sinovial de restos articulares libres, enzimas y mediadores de la
inflamación. El efecto es sólo temporal y la patología subyacente no se resuelve.
- El desbridamiento artroscópico es un tratamiento dirigido a eliminar las estructuras
responsables de los síntomas mecánicos (plicas, cuerpos libres, ostofitos,...) que
interfieren con la función articular. Produce un alivio de los síntomas a corto plazo
pero no resuelve la causa que origina la lesión. El resultado favorable de esta técnica
es atribuible a la disminución o eliminación de las partículas libres y tejidos dañados,
que estimularían la inflamación del tejido sinovial y el incremento de los niveles de
enzimas proteolíticas y de actividad colagenolítica, con el consecuente aumento de
friabilidad del cartílago articular (Álvarez y Casanova, 2004). El desbridamiento de una
lesión condral es el tratamiento de primera elección en las lesiones parciales y totales
del cartílago articular (Peterson, 2004) y cuando existe destrucción masiva de la
articulación, endoprótesis o esté contraindicado otro tratamiento (Handogy y Sokosd,
1999). También puede ser utilizado como primer tiempo de otros procesos de
reparación. Los resultados más satisfactorios a corto plazo se observan en pacientes
jóvenes, delgados, con buena alineación de la articulación, con antecedente específico
de traumatismo y con sintomatología de menos de un año de evolución (Sgaglione y
Miniaci, 2002).
3.2.- Tratamiento quirúrgico clásico:
Se distingue la perforación múltiple, la microfractura múltiple, la abrasión superficial o
afeitado y la abrasión profunda o espongialización (Cebamanos, 1998).
- Perforación múltiple (Figura 12): esta técnica afecta a los huesos subcondral y
trabecular y suele ser complementaria al desbridamiento (Handogy y Sokosd, 1999).
Consiste en la realización de perforaciones con instrumental motorizado y se utiliza
para tratar áreas focales en las que hay desaparición del cartílago articular y
exposición del hueso subcondral con el fin de promover la formación de un
fibrocartílago de reparación (Canosa, 2002). Hay que tener en cuenta que los canales

Introducción
30
formados pueden taparse e impedir la formación de una superficie de cartílago
adecuada (Handogy y Sokosd, 1999).
- Microfractura: es una técnica similar a la perforación, pero con penetración sólo de la
placa de hueso subcondral. Consiste en la realización de pequeños orificios con un
instrumental mecánico (juego de punzones de diferentes angulaciones) para realizar,
de forma manual, múltiples agujeros en el hueso subcondral. No deben de
profundizarse más de 4 mm y no deben de practicarse más de 3 - 4 orificios por cm2.
Con esta técnica, a diferencia de la anterior, no se genera calor. Ambas técnicas se
basan en el acceso al hueso y a la cavidad medular, lo que daría como resultado la
liberación de sangre y células madre mesenquimales. Esto provocaría la formación de
un ―macro coágulo‖ de reparación que, en unas determinadas condiciones de carga,
movimiento, etc... prolifera y forma un fibrocartílago de reparación (Sgaglione y
Miniaci, 2002). El fibrocartílago formado carece de algunos componentes del cartílago
hialino normal, lo que lo hace más susceptible a la rotura, de ahí que los resultados de
esta técnica no se mantengan a lo largo del tiempo (Álvarez y Casanova, 2004). Para
el éxito de estos procedimientos van a ser factores determinantes, entre otros: la
precisión en la realización de la técnica, el contenido de oxígeno, la movilización de la
articulación o la ausencia de complicaciones locales (sobre la zona de reparación)
(Robert, 2002).
- Abrasión superficial o afeitado: mediante este procedimiento se lleva a cabo la
exéresis del hueso subcondral necrótico de forma manual con un escoplo y
generalmente mediante artrotomía (Cebamanos, 1998). Se alcanza la vascularización
intracortical, donde los fibroblastos y las células pluripotenciales pueden repoblar el
defecto desbridado. Estaría contraindicada en pacientes con procesos inflamatorios o
en rodillas con gran rigidez, deformidad o inestabilidad (Canosa, 2002).
- Abrasión profunda o espongialización: esta técnica implica la extirpación completa de
la placa de hueso subcondral, con una fresa mecánica eliminando de 1 a 2 milímetros
de la superficie ósea, dejando las superficies de hueso trabecular. Este procedimiento
se realiza generalmente en el transcurso de una artroscopia. No se emplea para
lesiones aisladas sino para daños más extendidos, especialmente de rótula y de la
tróclea (Peterson, 2004).

Introducción
31
3.3.- Inducción celular de la condrogénesis:
Trasplante de periostio, de pericondrio y trasplante autólogo de condrocitos.
Estos procedimientos se basan en la limitada capacidad intrínseca de reparación
espontánea del cartílago, lo que obliga a buscar y utilizar otros tejidos cuyo potencial
diferenciativo permite, teóricamente, su transformación en condrocitos y por tanto la
neoformación de cartílago (Cebamanos, 1998).
- Injerto perióstico: el estrato más profundo del periostio (el cambium) se enfrenta a la
articulación en combinación con un proceso de rehabilitación (movilización pasiva).
Algunos autores recomiendan practicar túneles en la base ósea de la zona del defecto
con el fin de tener un punto de fijación para el lado libre del periostio. Los resultados
de esta técnica son una superficie que se asemeja al cartílago hialino por su estructura
bioquímica, pero con importantes diferencias estructurales (Handogy y Sokosd, 1999).
Con el paso del tiempo se produce un deterioro del tejido neoformado, encontrándose
en la artroscopia un tejido deflecado y una estructura histológica alterada
(Cebamanos, 1998).
- Injerto de pericondrio: en esta técnica, el cartílago dañado se sustituye por
pericondrio. Una de las mayores limitaciones de este procedimiento, cuando se trata
de defectos grandes o múltiples, es el tamaño de la zona donante (que suelen ser las
costillas). Otras limitaciones de la eficacia de la técnica a largo plazo son la osificación
y la delaminación del cartílago de la placa subcondral (Minas y Nehrer, 1997).
El éxito de estas dos técnicas viene dado por la disponibilidad de obtener material a
injertar en suficientes cantidades para reparar los defectos grandes, por la pobre
integración del injerto o por la posibilidad de osificación endocondral del tejido
injertado.
- Trasplante autólogo de condrocitos: Se considerará en el apartado de terapia celular
con condrocitos.

Introducción
32
3.4.- Trasplante osteocondral:
Se utilizan los aloinjertos y los autoinjertos osteocondrales.
- Aloinjerto osteocondral: consiste en sustituir una parte de la articulación afectada por
una destrucción osteocondral severa, por un aloinjerto de tejido fresco o conservado
(Handogy y Sokosd, 1999). Su uso, debido al posible riesgo de reacciones
inmunológicas y/o de transmisión de enfermedades víricas, se limita a pacientes
jóvenes con lesiones focales condrales u osteocondrales. Las principales desventajas
son, la viabilidad celular, que se sitúa habitualmente entre el 10 y el 30% y la elevada
exigencia técnica del procedimiento.
- Autoinjerto osteocondral: entre las ventajas que presenta esta técnica destacan la
supervivencia del injerto al proceso de trasplante permitiendo la formación de una
superficie de cartílago hialino en el sitio del defecto, la posibilidad de realizar el
trasplante en bloque de cartílago hialino y de hueso subcondral, preservando así la
superficie de unión cartílago-hueso y que no hay riesgo de transmisión viral. Un
aspecto relevante es la congruencia del injerto, ya que los no congruentes tienen
tendencia a degenerar, lo que hace que la elección de los lugares de extracción sea
muy importante. Hay que tener en cuenta que es difícil obtener sitios donantes de más
de 4 cm2 sin tocar superficies importantes. Para eliminar los problemas ligados al sitio
de extracción y a la congruencia, se ideó una técnica de injerto en mosaico
(mosaicoplastia) (Handogy y Sokosd, 1999). Esta técnica consiste en la extracción de
pequeñas piezas cilíndricas de las zonas de mínimo apoyo y periferia de los cóndilos
femorales y trasplantarlos a las zonas de carga de peso. La combinación de distintos
tamaños permite rellenar el defecto en el 90 ó 100%. Como ventajas cabe señalar, por
una parte, que el uso de múltiples piezas permite una mayor cantidad de tejido a
trasplantar, al tiempo que se preserva la integridad del sitio donante y por otra parte,
su colocación permite dar contorno a la superficie articular. Como limitaciones
importantes destacar que el lugar de extracción del cartílago donante debe
corresponder a superficies articulares que no soporten carga, por lo que resulta muy
limitado y que el uso de grandes injertos puede causar incongruencia articular y alterar
así la biomecánica de la articulación (Alvares y Casanova, 2004).

Introducción
33
Figura 11.- Imágenes de artroscopia de lesiones del cartílago articular que ilustran la
clasificación de Outerbridge.
http://www.sofarthro.com/ANNALES/ANNALES_1999/CARTILAGE/classification.htm)

Introducción
34
Figura 12.- Imagen artroscópica de perforaciones realizadas en el cartílago articular.
http://www.sofarthro.com/fr/ANNALES/ANNALES_1999/CARTILAGE/debridement.htm
Figura 13.- Imagen artroscópica de una mosaicoplastia.
(http://www.sofarthro.com/ANNALES/ANNALES_1999/CARTILAGE/Mosaic.htm)

Introducción
35
4.- TERAPIA CELULAR CON CONDROCITOS
La degeneración del tejido cartilaginoso, con un origen bien congénito, bien
traumático, tiene una gran importancia clínica, dado el limitado potencial curativo
intrínseco que tiene este tejido. Como consecuencia del deficitario aporte sanguíneo y
en el contexto del intento reparativo, las lesiones condrales tienen como resultado una
incompleta reparación de la estructura cartilaginosa. Las lesiones de todo el espesor
del cartílago articular, dan lugar a una respuesta inflamatoria normal, pero con una
insuficiente formación de fibrocartílago. Para solucionar la degeneración articular
progresiva que ocurre en enfermedades como la artrosis, la intervención quirúrgica es
en ocasiones la única opción. Pese al éxito de la artroplastia, la mayoría de los
tratamientos orientados a reparar el cartílago articular son normalmente
insatisfactorios y raramente restauran completamente la función articular o devuelven
el tejido a su estado nativo.
La ingeniería tisular, constituye un emergente campo científico, que ofrece grandes
expectativas en la generación de sustitutos tisulares, intentando la ―fabricación‖ in vitro
de constructos para la posterior implantación in vivo.
El principio básico consiste en utilizar un ―andamiaje‖ compatible biológica, estructural
y mecánicamente, que será sembrado de células y cargado con moléculas bioactivas
para promover la maduración y/o diferenciación celular.
Aunque los últimos progresos en ingeniería de cartílago se han realizado en el campo
de la cosmética, son importantes los intentos para crear un tejido arquitecturalmente
complejo, multicapa, similar al cartílago articular, capaz de soportar carga. Ya se han
conseguido exitosas aproximaciones a la construcción artificial de un tejido
cartilaginoso por ingeniería tisular, incluyendo el uso de andamiajes con biomateriales
naturales y sintéticos, fuentes alogénicas y autogénicas de condrocitos maduros y de
células progenitoras condroides, factores condroinductores de crecimiento, como el
factor de crecimiento β-s transformado (TGF-βs), y combinaciones de ellos (Tuli y Li,
2003).
Durante las últimas décadas, son numerosas las técnicas y los métodos que se han
desarrollado para el cultivo de distintos tipos de células humanas y animales en
laboratorio. A continuación, presentaremos una breve reseña histórica de las
principales técnicas que se han propuesto para el cultivo y el implante de condrocitos
con vistas a su posible utilización terapéutica.

Introducción
36
El aislamiento y crecimiento de condrocitos en un medio de cultivo lo consiguió por
primera vez Smith (Peterson, 2004), en el año 1965, demostrando la supervivencia in
Vitro de los mismos a bajas temperaturas. En ese momento se inyectaron estas
células experimentalmente en la rodilla de conejo pero no se logró una reparación
significativa. Chesterman y Smith (Chesterman y Smith, 1968) lograron aislar en 1968
condrocitos del cartílago articular en conejos, retirando la matriz extracelular mediante
un proceso enzimático (papaina, colagenasa, pronasa). Posteriormente, estos autores
injertaron los condrocitos en un corte realizado en la superficie articular del húmero,
obteniendo como reparación un tejido fibroso.
Unos años más tarde, (1971 y 1974), Bentley y Greer (Bentley y Greer, 1971)
presentaron sus experiencias en el aislamiento de condrocitos procedentes de
cartílago articular, observando una escasa supervivencia de los condrocitos cuando
estos eran transplantados en defectos articulares en los platillos tibiales de conejos de
laboratorio.
Al mismo tiempo, Elves (Elves,1974) demostró en 1974 el papel protector de la matriz
cartilaginosa, identificando la presencia de antígenos del cartílago con un alto grado de
histocompatibilidad con los linfocitos, así como la disminución de estos antígenos
cuando el cartílago era tratado con papaína para el aislamiento de los condrocitos.
En 1977, Green (Green, 1977) realizó experimentos en los que sembró condrocitos
sobre matriz ósea estéril para luego ser implantados en ratones atímicos, siendo el
resultado la formación de tejido cartilaginoso.
En el año 1982, se diseñó e inició en el Instituto Ortopédico de Nueva York un modelo
experimental, con un grupo control, en conejos, utilizando condrocitos articulares
aislados y desarrollados en medios de cultivo. La idea consistía en utilizar condrocitos
autólogos como células encargadas de formar cartílago hialino. Los primeros
resultados de esta técnica se presentaron en 1984 y se demostró la aparición de un
cartílago similar al hialino con un relleno del 80% en el grupo tratado con células. En el
grupo control, en el que sólo se trató el defecto con recubrimiento perióstico pero no
con células, no se observó relleno de la lesión.
Desde ese año se han sucedido numerosos y extensos estudios en animales en la
Universidad de Goteborg, en Suecia. Peterson y cols. en 1984 y Grande y cols. en
1985 presentaron sus resultados con esta técnica en defectos condrales, en conejos,
que no afectaban hueso subcondral.

Introducción
37
En 1985, se planteó la posibilidad de realizar esta técnica en seres humanos y así, en
1987 se llevó a cabo en Suecia el primer trasplante autólogo de condrocitos en una
rodilla humana.
El procedimiento de cultivos celulares aplicado a los condrocitos ha permitido
aumentar el número de células y mantener su fenotipo. Así, durante los años ochenta
se incorporaron los geles de colágeno a los cultivos de condrocitos (Wakitani et al.,
1989), proporcionando una mejor fijación y un mejor medio para la síntesis celular de
macromoléculas de la matriz cartilaginosa.
Bujía y cols., (Bujía et al., 1993), demostraron que la rediferenciación de los
condrocitos en medios de cultivo tridimensionales (suspensión y gel de agarosa) fue
completa, adquiriendo de nuevo una morfología y funcionalidad equiparables a la de
los tejidos normales.
En octubre de 1994 Brittberg y cols. (Brittberg et al., 1994) publicaron en el New
England Journal of Medicine los resultados de un estudio piloto realizado con 23
pacientes. Era el primer estudio que publicaba los resultados de este procedimiento
aplicado a seres humanos.
Posteriormente, en 1995, Park (Park al., 1995) presentó una técnica para realizar
injertos articulares en conejos de laboratorio utilizando cultivos tridimensionales. En
este estudio, se demostró que el implante subcutáneo de este tipo de tejidos era bien
tolerado y presentaba adecuados niveles de diferenciación y biointegración.
El trasplante autólogo de condrocitos es una técnica que habitualmente se realiza en
dos tiempos (Kaigler et al., 2001; Jato, 2008). En un primer tiempo, mediante
artroscopia, se realiza una extracción de láminas de cartílago articular de una zona
que soporte baja carga, que se procesa en laboratorio, para aislar el mayor número de
condrocitos viables, a partir de los cuales se generarán cultivos primarios de este tipo
de células (Kaigler y Mooney, 2001), que se mantendrán durante 3-4 semanas. En una
segunda intervención, se realiza una artrotomía y se extirpa la lesión condral llegando
hasta hueso subcondral. El defecto se cubre con un injerto de periostio tomado de la
cara antero medial de la tibia, al que se deja una pequeña apertura para inyectar los
condrocitos cultivados. Una vez realizado este proceso, se cierra la abertura del
periostio con un par de suturas y se sella con un pegamento biológico de fibrina. Se
cierra la artrotomía y se aplica un vendaje elástico. De este modo, se pretende que
estas células puedan suplir la deficiencia estructural o funcional que se hubiera podido
producir en el tejido (Dove, 2002) (Figuras 14 y 15).

Introducción
38
El programa de rehabilitación posterior juega un papel clave en la recuperación de la
lesión, pues estimula la regeneración de los condrocitos y reduce la posibilidad de
adherencias intraarticulares. Aunque varía según la localización de la lesión, con
carácter general, consiste en una primera fase de la rehabilitación que comprendería
seis semanas y que abarcaría las siguientes fases:
1. Se inicia mediante movilización pasiva continua entre las seis y las cuarenta y ocho
horas posteriores a la intervención.
2. Al día siguiente de la intervención, además de la movilización pasiva continua,
iniciará cinesiterapia activa y asistida de recuperación del arco articular, de media hora
una o dos veces al día.
3. A partir del tercer día empezará a realizar ejercicios activos libres y autoasistidos de
flexoextensión completa de rodilla e isométricos de cuádriceps. En casos de poca
colaboración o escaso progreso puede considerarse asociar estimulación eléctrica del
cuádriceps. Así mismo empezará a ensayar equilibrio monopodal hasta conseguir la
independencia en la marcha en descarga. El paciente debe conseguir en el primer
mes 95º de flexión y extensión completa o faltando a ésta 10º.
Posteriormente se continuaría con:
1. Marcha bipodal, descargada con dos bastones de codo, salvo casos particulares e
indicación del cirujano, que se iniciará a las ocho semanas y será plena a las diez
semanas, prescindiendo de los bastones a las doce semanas.
2. Actividades como la bicicleta, natación, paseos largos etc., no están permitidas
hasta seis, nueve meses después de la intervención, durante una hora por día. La
carrera se iniciará gradualmente hacia los nueve, doce meses.
En el caso de la osteocondritis disecante el período de rehabilitación debe ser, como
mínimo, de dieciocho a veinticuatro meses.
Más tarde, se comprobó que la tasa real de reparación del cartílago dañado una vez
implantadas las células mediante transferencia celular, era menor de lo que se
esperaba (Wada y Wanatabe, 2003). En realidad, se cree que la tasa de reparación del
tejido dañado, inmediatamente después de la transferencia celular es muy baja, pero
que se va incrementando conforme pasan los días, llegando a una reparación casi
completa tras un periodo de nueve, doce meses.

Introducción
39
Avalia (Jato y Ruano, 2005) en su trabajo de 2003 sobre la evolución clínica del
trasplante autólogo de condrocitos, describe la extracción de láminas de cartílago
articular de zona de no carga. La muestra se cultivó durante tres, cuatro semanas
hasta obtener un número suficiente de condrocitos, para proceder posteriormente a su
injerto. El número medio de condrocitos implantados fue de 19 millones y el mínimo de
12 millones. En este trabajo, sólo aparecieron complicaciones intraoperatorias en un
caso, que estuvieron relacionadas con un número bajo de células en el cultivo
primario. Los resultados globales después de dos años de evolución fueron
satisfactorios.
Al mismo tiempo, se llevó a cabo un estudio en el que se cultivaron los condrocitos,
inmersos en una matriz tridimensional de nanofibras de poli-E-caprolactona (Tuli y Li,
2003). De este modo, se pretendía generar un sustituto tridimensional del cartílago
mediante la elaboración de un constructo por Ingeniería Tisular. Los resultados de este
procedimiento demostraron una adecuada maduración de los condrocitos, los cuales
mantenían un estado funcional en este tipo de matriz, la cual favorecía la proliferación
celular y el mantenimiento del fenotipo condral.
En esta misma línea, Pineda y cols. (Pineda y Merentes, 2004) evaluaron en 2004 las
características morfológicas y bioquímicas de los condrocitos, humanos y de animales
de laboratorio, mantenidos en cultivo. Estos autores realizaron dos tipos de cultivos: un
cultivo celular monocapa y otro cultivo tridimensional. En el primer caso las células
adoptaron una morfología poliédrica, con espacios prominentes entre ellas. Estas
células sintetizaron glucosaminoglucanos en cultivo, pero la producción de
proteoglucanos fue escasa, difundiendo estos hacia el medio de cultivo en su mayor
parte, debido a una falta de matriz que los retuviera. En el segundo caso, cuando los
condrocitos se cultivaron tridimensionalmente en presencia de una matriz de colágeno
tipo I, mantuvieron su morfología esférica, sintetizando componentes característicos de
la matriz extracelular como los glucosaminoglucanos sulfatados y el colágeno tipo II.
Por otro lado, Darling y cols. (Darling et al., 2004), utilizaron TGF-B como factor de
crecimiento para la estimulación de la proliferación de condrocitos, concluyendo que el
TGF-B1 tiene mejor efecto sobre la proliferación celular después de ocho días de
cultivo y que el IGF-1 es capaz de incrementar la expresión del gen del colágeno tipo
II. Por tanto, recomendaron agregar factores de crecimiento para la rápida
construcción de cartílago en el reemplazo del tejido dañado para su uso clínico en
humanos.

Introducción
40
Glowacki (Glowacki et al., 2005) demostró que el uso de matrices extracelulares de
colágeno poroso favorece el cultivo de condrocitos procedentes de cartílago articular
bovino, mejorándose y promoviéndose la condroinducción y la condrogénesis al utilizar
factores de crecimiento en el cultivo.
Más recientemente, el desarrollo de las técnicas de cultivo tridimensional basadas en
nuevos biomateriales y de la ingeniería tisular en su conjunto, ha derivado en la
generación de diferentes modelos de cartílago artificial generados en laboratorio. Uno
de estos modelos es el que propusieron Oliveira y colaboradores (Oliveira et al., 2006),
utilizando matrices de hidroxiapatita y quitosán. Según estos autores, estos
biomateriales permiten el desarrollo de cartílago y hueso en el laboratorio a partir de
células de médula ósea.
De igual modo, Jiang y cols. (Jiang y Chiang, 2007) utilizaron biomateriales
compuestos de ácido DL-poli-lactólico-co-glicolico para la reparación de defectos del
cartílago articular con buenos resultados.
Finalmente, Saris y Vanlauwe (Saris y Vanlauwe, 2008), demostraron que el implante
de condrocitos autólogos era una técnica sencilla con buenos resultados. En concreto,
estos autores llevaron a cabo un ensayo clínico en el que demostraron que la
regeneración del cartílago es superior en los pacientes en los que se implantan los
condrocitos cultivados en comparación con los casos en los que se utiliza una técnica
quirúrgica convencional.
La conclusión es que la terapia celular con condrocitos autólogos es una técnica que
parece útil para la reparación de defectos sintomáticos del cartílago articular.

Introducción
41
Figura 14.- Proceso de ingeniería tisular del tejido cartilaginoso. (Según Freedet Van Jak
Novakovic, Principle of Tissue Engeneering, 1997).
Biopsia del cartílago sano
Digestión enzimática
Lesión
Se cultivan las células durante 11-21 días.
Tripsinización Suspensión celular tras centrifugación
Inyección del cultivo de condrocitos bajo en colgajo
Colgajo de periostio trasplantado a la
lesión
Colgajo del periostio de la
tibia medial
Figura 15.- Trasplante autólogo de condrocitos en el cóndilo femoral. Se muestra el fémur
distal y la tibia proximal (Brittberg et al., 1994).

Introducción
42
5.- VIABILIDAD CELULAR
El objetivo de la Ingeniería Tisular es regenerar y restablecer la función normal de un
tejido u órgano dañado, utilizando células que son cultivadas en el laboratorio o en
matrices artificiales, para posteriormente ser trasplantadas a un órgano receptor. De
esta forma, uno de los requisitos más importantes a destacar, antes de que los
condrocitos puedan ser implantados, es la determinación de la viabilidad de las células
mantenidas en cultivo.
Se han descrito diversos métodos y técnicas de laboratorio, útiles para la evaluación
de la viabilidad y de la funcionalidad celular (Tabla 1). Destacan entre ellas la
evaluación de la integridad de la membrana celular, los ensayos funcionales, los
ensayos con pruebas de fluorescencia, los estudios de morfología celular, el
microanálisis por energía dispersiva de rayos X y las técnicas de determinación de la
expresión génica mediante microarrays.

Introducción
43
Tabla 1: Tabla Resumen de los ensayos de evaluación de viabilidad más importantes.
Tabla Resumen de los ensayos de evaluación de viabilidad más importantes
Ensayos de evaluación de
la viabilidad
Métodos de evaluación de
la viabilidad celular Fundamento
Evaluación de la integridad de la
membrana
1. Métodos basados en la
exclusión o inclusión de
colorantes o sustancias
fluorescentes.
2. Métodos basados en la
utilización de tinciones
catiónicas.
3. Métodos basados en la
determinación de
liberación de moléculas.
Si una célula está dañada, la
función de la membrana celular
estará alterada. A su través pasarán
moléculas o no, que en condiciones
normales no lo harían.
Ensayos funcionales
1. Medición del ATP.
2. Tasa de ADN
3. Síntesis de proteínas
Evalúan los componentes
metabólicos que son necesarios
para el crecimiento celular
Ensayos con pruebas de
fluorescencia
Utilización de Biosensores de
fluorescencia
Miden la dinámica molecular de
macromoléculas, metabolitos e
iones en células vivas
Ensayos morfológicos Métodos basados en la observación
con el microscopio
Determinación del cambio
morfológico.
Microscopía electrónica analítica Microanálisis por energía dispersiva
de rayos X
Determinan la composición
elemental de una muestra en el
microscopio electrónico
Determinación de la expresión
génica global
Microarrays de ADNc
Microarrays de oligonucleótidos Evaluación de la expresión de
genes de un genoma completo

Introducción
44
5.1. Detección de alteraciones de la permeabilidad celular
Probablemente los más numerosos y utilizados, son los métodos basados en la
detección de alteraciones de la permeabilidad de la membrana, pudiendo distinguir
dentro de éstos dos variantes:
Métodos basados en la exclusión de colorantes vitales.
Métodos basados en la determinación de la liberación de moléculas,
fundamentalmente enzimas o ácidos nucleicos, en el medio extracelular.
Aquellos que se caracterizan por el empleo de colorantes, habitualmente consisten en
la utilización de un colorante que, en función de sus características, es capaz de
penetrar y colorear el interior, bien de las células vivas o bien de las células muertas.
La proporción relativa de las células coloreadas o no, refleja el número exacto de las
células vivas o muertas y, en consecuencia, la viabilidad del conjunto de la población
celular. El contaje de las diferentes poblaciones celulares puede ser efectuado por
métodos microscópicos, citometría de flujo, espectrofotometría o espectrofluometría
automatizada. Con frecuencia, se han utilizado colorantes orgánicos como el azul
tripán (Hoskins et al., 1956) (Phillips, 1973) (Patterson, 1979), la eosina (Hoskins et al.,
1956), el rojo Congo (Geschickter, 1930), la eritrosina B (Phillips et al., 1957) (Bhuyan
et al., 1976), el ioduro de propidio o el bromuro de etidio (Edidin, 1970) (Krishan,
1975) (Hamilton et al., 1980), los cuales son capaces de teñir y colorear únicamente
las células muertas, siendo expulsados activamente del citoplasma celular cuando las
células están vivas. Por ese motivo, este tipo de ensayos se denominan con
frecuencia ensayos de exclusión de colorantes orgánicos.
De otra forma, los que consisten en la detección y medición de determinadas
moléculas intracelulares liberadas al medio de cultivo, están fundamentados en las
alteraciones de la permeabilidad de la membrana celular bajo la acción de
determinados tóxicos o bien por envejecimiento celular espontáneo. Dichos métodos
comprenden dos tipos de ensayos, uno enzimático y otro radiactivo, siendo más
utilizados los primeros. Uno de los ensayos enzimáticos más comunes es la
determinación del enzima citosólico lactato deshidrogenasa (LDH) en el medio de
cultivo, enzima que es liberada al medio extracelular únicamente cuando la membrana
plasmática ha sido gravemente alterada. Otras moléculas de interés a la hora de
evaluar la viabilidad celular son las enzimas mitocondriales o incluso el ADN nuclear.

Introducción
45
Niveles elevados de cualquiera de estas moléculas en el medio de cultivo son claros
indicativos de que un porcentaje significativo de células ha sido destruido y, por tanto,
de que la viabilidad celular es baja (Coco-Martin et al., 1992; da Costa et al., 1999;
Posadas et al., 2007; kegami et al., 2007; Park et al., 2008).
De entre los métodos más utilizados, cabe destacar aquel que determina la viabilidad
celular mediante la prueba del bromuro de 3-(4,5-dimetiltiazol-2-yl)-2,5-difeniltetrazolio
(MTT). Este compuesto, de color amarillento, es capaz de reducirse y adquirir color
morado por acción de las enzimas intramitocondriales de las células vivas. Por este
motivo, la adición de MTT al medio de cultivo y la posterior cuantificación colorimétrica
del producto nos puede dar una idea bastante fiable del número de células vivas y
muertas que existen en el cultivo celular (Brink et al., 2008; Meriç et al., 2008).
Para terminar, aquellos que se basan en la radioactividad tienen su fundamento
científico en la medición de la liberación del Cr51, isótopo radiactivo que se une de
manera no covalente a los aminoácidos básicos de las proteínas intracelulares. Las
células inertes liberan el Cr51 en el medio extracelular, el cual puede ser cuantificado
por un contador gamma (Rinaldi et al., 1998).
5.2. Ensayos morfológicos
Tienen su fundamento en la observación con el microscopio. Los cambios
morfológicos que se producen en la superficie celular o en el citoesqueleto pueden
estar relacionados con la viabilidad celular (Emilson et al., 1978; Wiesel et al., 1983;
Beattie et al., 1994; Amato y Lozzi, 1995; Debbage, 1998). De esta forma, los cambios
de volumen irreversibles pueden ser utilizados para indicar la muerte celular. Si existe
una gran disminución en el volumen celular, este puede ser secundario a la pérdida de
proteínas o iones intracelulares, o debido a una alteración de la permeabilidad para el
sodio o el potasio (Allen, 1988).
5.3. Microanálisis por energía dispersiva de rayos X
Una de las técnicas más sensibles para determinar la viabilidad de las células en
cultivo es la cuantificación del contenido iónico, especialmente del potasio y del sodio
(Rodríguez-Morata et al., 2008). La concentración intracelular de estos iones se
correlaciona bastante bien con el estado vital celular y es un excelente marcador de la
fisiología y de la viabilidad celular (Fernández-Segura et al., 1999; Roomans, 2002;

Introducción
46
Zierold, 1997). El microanálisis por energía dispersiva de rayos X asociado a la
microscopía electrónica supone una potente herramienta para cuantificar los
elementos de una muestra, al mismo tiempo que se consigue determinar la
concentración de los mismos y la ultraestructura de las células (Buja et al., 1985; Hall,
1988; Vanthanouvong et al., 2003; Warley, 1997).
Es una técnica que, utilizando un haz de electrones, permite estudiar la composición
química de la muestra de forma simultánea a su observación microscópica (Carini et
al., 1995; Carini et al., 1997; Carini et al., 2000).
El microanálisis por energía dispersiva de rayos X permite el análisis simultáneo de
todos los cationes y aniones (Z 11) (Warley et al., 1994; Rodríguez-Morata et al.,
2008), presentes en el espécimen irradiado por el haz de electrones y requiere un
pequeño número de células.
El estudio de muestras biológicas con Microscopía Electrónica Analítica permite
asimismo la cuantificación de los elementos objeto de estudio, lo cual exige el
desarrollo de protocolos específicos (Campos et al., 1992; Campos et al., 1994;
Crespo et al., 1993; Fernández-Segura et al., 1999a y 1999b; López-Escámez et al.,
1992 y 1993; López-Escámez y Campos, 1994; Warley et al., 1994; Warley, 1997).
Pero estos cambios son más difíciles de medir, y como consecuencia, tienen menos
utilidad que la evaluación de integridad de la membrana o los ensayos funcionales.
5.4. Determinación del perfil de expresión génica mediante microarrays
El microarray es una técnica que permite la evaluación simultánea de un gran número
de genes o incluso de un genoma completo, en un único experimento (Friemert et al.,
1989; Gress et al., 1992). Recientemente, esta técnica se ha convertido en una nueva
herramienta para estudiar algunas propiedades específicas de las células susceptibles
de utilización en Terapia Celular y en Ingeniería Tisular (Schena et al., 1995; Gill,
2003; Jaluria et al., 2007 y 2008). Por ejemplo, una de las propiedades celulares que
se puede evaluar con el microarray podría ser la viabilidad celular mediante la
identificación de genes relacionados con la mortalidad celular (por ejemplo, genes de
apoptosis y anti-apoptosis) (Wong et al., 2006) y, de esta manera, se podría realizar
una selección de las células, escogiendo las que tienen un mayor grado de viabilidad.
Atendiendo a su función, podemos distinguir tres tipos fundamentales de microarrays:

Introducción
47
1.- Microarrays de expresión génica (Bowtell, 1999). Esta técnica se basa en
la detección de ARN mensajeros específicos que están presentes en una
muestra biológica en un momento dado. Para ello, se extrae el ARN total de
dicha muestra, el cual se marca con un pigmento y se hibrida frente a un chip o
matriz en la que existen copias de ADN complementario (ADNc) a los genes
que se pretenden cuantificar. Por lo general, este tipo de microarrays son los
más utilizados y los mejor conocidos.
2.- Microarrays de ADN (Bier et al., 2008; Wiltgen y Tilz, 2008). En este tipo
de microarray, el ADN total procedente de una muestra se marca y se hibrida
frente a un chip en el que existen copias de los genes a identificar. De este
modo, se puede evaluar el número de copias de cada gen existente en cada
célula, así como la presencia de delecciones, mutaciones o ganancias génicas.
Por ello, este tipo de microarrays permite realizar un análisis completo del
genoma de la célula (genotipificación).
Un tipo especial de microarrays de ADN es aquél en el que se evalúa la
presencia de modificaciones epigenéticas tipo metilación a nivel del promotor
de ciertos genes (Zilberman y Henikoff, 2007).
3.- Microarrays de proteínas (Tao et al., 2007). Consisten en un chip en el
que se coloca cierto número de anticuerpos específicos de origen conocido,
frente al cual se hibrida un extracto proteico previamente marcado.
En la presenta tesis doctoral, para los estudios de viabilidad de los condrocitos, se
utiliza el método de exclusión del colorante azul tripán, que detecta las alteraciones de
la permeabilidad celular y la determinación del perfil de expresión génica mediante
microarray, para identificar el patrón de expresión de los genes implicados en la
diferenciación y en la mortalidad celular y por tanto en la viabilidad de los condrocitos.

INTRODUCCIÓN
OBJETIVOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
TABLAS SUPLEMENTARIAS
REFERENCIAS BIBLIOGRÁFICAS

Objetivos
49
1.- Aislar y generar cultivos primarios de condrocitos del cartílago hialino articular
humano.
2.- Identificar el patrón de viabilidad celular de los condrocitos articulares humanos
mediante el método de exclusión de colorantes vitales en los ocho primeros
subcultivos celulares.
3.- Identificar el patrón de expresión génica global de los condrocitos articulares
humanos en cada uno de los ocho primeros subcultivos celulares mediante microarray
e identificar los genes cuya expresión se relaciona con la evolución cronológica de los
mismos.
4.- Identificar el patrón de expresión de los genes relacionados con la diferenciación
condral en los ocho primeros subcultivos celulares.
5.- Identificar el patrón de expresión de genes vinculados con la mortalidad celular en
los ocho subcultivos celulares.
6.- Correlacionar la viabilidad y la expresión génica de mortalidad y diferenciación para
seleccionar la población de condrocitos susceptible de ser utilizada en terapia celular
cartilaginosa.

INTRODUCCIÓN
OBJETIVOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
TABLAS SUPLEMENTARIAS
REFERENCIAS BIBLIOGRÁFICAS

Materiales y Métodos
51
1.- AISLAMIENTO DE CONDROCITOS
Para la realización de este trabajo, hemos utilizado condrocitos articulares humanos
obtenidos a partir de cuatro diferentes biopsias de cartílago articular de rodilla humana.
Dichas biopsias se llevaron a cabo en el transcurso de intervenciones quirúrgicas
consistentes en el reemplazo articular completo de la rodilla (artroplastia total de
rodilla).
Los pacientes fueron escogidos al azar entre aquellos que figuraban en la lista de
espera quirúrgica para someterse a una artroplastia total de rodilla, en el Servicio de
Cirugía Ortopédica y Traumatología del Hospital del SAS de Jerez de la Frontera
(Cádiz), tras la obtención del consentimiento informado.
Durante el procedimiento quirúrgico se obtuvieron fragmentos de cóndilo femoral,
procedentes de las osteotomías realizadas necesarias para la colocación de la
prótesis. De estos fragmentos se aislaron las biopsias de cartílago articular que fueron
conservadas en medio de transporte tisular a 4ºC, donde se mantuvieron hasta el
momento de su procesamiento. Este medio de transporte consistió en un medio básico
DMEM (medio de Eagle modificado por Dulbecco, Sigma-Aldrich ref. D5796, St. Louis,
Missouri, EEUU; ver composición en Tabla 2) suplementados con antibióticos y
antimitóticos (500 U/ml de anfotericina B, Sigma-Aldrich ref.A5955) pero sin suero
bovino fetal.
Con el objetivo de conseguir disgregar la matriz extracelular del cartílago y conseguir
la separación de los condrocitos de dicha matriz, las muestras se incubaron en una

Materiales y Métodos
52
disolución de colagenasa I y II de Clostridium hystoliticum (Gibco BRL Life
Technologies Ref. 17100-017, Karlsruhe, Alemania) al 2% a 37ºC durante 12 horas.
Esta solución es capaz de digerir el colágeno y la matriz y liberar los condrocitos
existentes en el cartílago.
Las muestras digeridas en colagenasa se centrifugaron a 1000 revoluciones por
minuto durante 10 minutos, con el fin de obtener las células disgregadas de la matriz
extracelular, recogiéndose el precipitado pellet celular correspondiente.
Posteriormente, este precipitado celular se cultivó en frascos de cultivo de 25 cm2 de
superficie tipo Falcon® con medio de cultivo QN. El medio QN fue inicialmente descrito
por De Diego (De Diego et al., 2004) y Llames (Llames et al., 2004) para el cultivo de
queratinocitos de la piel, habiendo demostrado su utilidad en el cultivo de diversos
tipos de células humanas y animales (Alaminos et al., 2006 y 2007; Sánchez Quevedo
et al., 2007). La composición de este medio de cultivo es la siguiente:
- Medio de Eagle modificado por Dulbecco (DMEM): 300 ml (Sigma-Aldrich ref. D5796)
(Ver tabla 2).
- Medio HAM-F12: 150 ml (Sigma-Aldrich Ref. N6658) (Ver tabla 2).
- Toxina colérica: 8 ng/ml (Sigma-Aldrich Ref. C3012).
- Adenina: 24 µg/ml (Sigma-Aldrich Ref. A9795).
- Hidrocortisona: 0, 4 mg/ml (Sigma-Aldrich Ref. H0888).
- Insulina: 5 mg/ml (Sigma-Aldrich Ref. I2767).
- Triyodotironina: 1, 3 ng/ml (Sigma-Aldrich Ref. T5516).
- Suero bovino fetal (SBF): 50 ml (Sigma-Aldrich ref. F9665).
- Antibióticos y antimicóticos: 100 U/ml de penicilina G, 100 g/ml de estreptomicina y
0,25 g/ml de anfotericina B (Sigma-Aldrich Ref. A5955).
Todos los cultivos celulares se incubaron a 37ºC con un 5% de dióxido de carbono, en
condiciones estándar de cultivo. Los medios de cultivo se renovaron cada tres días.

Materiales y Métodos
53
Tabla 2: Composición de los medios DMEM y HAM-F12. Todos los valores están expresados
en gramos por litro de medio de cultivo.
COMPONENTE Medio de Eagle Modificado
por Dulbecco (DMEM) Medio de cultivo HAM-F12
SALES INORGÁNICAS CaCl2•2H2O 0.265 0.0441 CuSO4•5H2O — 0.0000025
Fe(NO3)3•9H2O 0.0001 — FeSO4•7H2O — 0.000834
KCl 0.4 0.224 MgCl•6H2O — 0.123
MgSO4 0.09767 — Na2HPO4 — 0.14204
NaCl 6.4 7.599 NaH2PO4 0.109 — NaHCO3 3.7 1.176
ZnSO4•7H2O — 0.000863 AMINOÁCIDOS
Glicina 0.03 0.00751 L-Alanina — 0.009
L-Arginina•HCl 0.084 0.211 L-Asparagina•H2O — 0.01501 L- Ácido Aspártico — 0.0133 L-Cisteína•2HCl 0.0626 0.035
L- Ácido Glutámico — 0.0147 L-Glutamina 0.584 0.146
L-Histidina•HCl•H2O 0.042 0.02096 L-Isoleucina 0.105 0.00394 L-Leucina 0.105 0.0131
L-Lisina•HCl 0.146 0.0365 L-Metionina 0.03 0.00448
L-Fenilalanina 0.066 0.00496 L-Prolina — 0.0345 L-Serina 0.042 0.0105
L-Treonina 0.095 0.0119 L-Triptófano
0.016 0.00204 L-Tirosina•2Na•2H2O 0.10379 0.00778
L-Valina 0.094 0.0117 VITAMINAS
Colina 0.004 — D- Ácido Pantoténico •½Ca 0.004 0.00048
Ácido Fólico 0.004 0.00132 Hipoxantina — 0.00408
Ácido Linoleico — 0.0000084 Mio-Inositol 0.0072 0.018
Ácido nicotínico 0.004 0.000037 Piridoxina•HCl 0.004 0.000062
Riboflavina 0.0004 0.000038 Tiamina•HCl 0.004 0.00034
Ácido Tióctico — 0.00021 Timidina — 0.00073
Vitamina B-12 — 0.00136 OTROS
D-Glucosa 4.5 1.802 Fenol rojo•Na 0.0159 0.0013 Ácido pirúvico — 0.11 Putrescina•HCl — 0.000161

Materiales y Métodos
54
2.- OBTENCIÓN DE SUBCULTIVOS CELULARES
Toda vez que los condrocitos articulares humanos, en su disposición de monocapa
alcanzaron la semiconfluencia, las células se lavaron con 5 ml de PBS (tampón
fosfato, Sigma-Aldrich ref. P4417) para eliminar cualquier resto del medio de cultivo y
las células muertas. A continuación, se cubrió toda la superficie de cultivo con
aproximadamente 2 ml de solución de disociación celular (Sigma-Aldrich ref. C5789)
para disgregar los mecanismos de adhesión celular y obtener células individualizadas
y no adheridas a la superficie del frasco de cultivo, incubándose durante cinco, diez
minutos a 37 ºC.
Pasado este tiempo, y una vez quedó comprobada la separación de las células del
frasco, se añadieron 5 ml de medio QN para inactivar el efecto de la solución de
disociación, recogiéndose ambos en un tubo cónico que se centrifugó a 1000 rpm
durante 10 minutos. El precipitado o pellet celular que se obtuvo se resuspendió en
medio de cultivo QN y se sembró en nuevos frascos de cultivo con medio QN.
Este procedimiento se llevó a cabo un total de ocho veces, obteniéndose así un total
de ocho subcultivos celulares.

Materiales y Métodos
55
3.- DETERMINACIÓN DE LA VIABILIDAD CELULAR MEDIANTE ENSAYOS DE
EXCLUSIÓN DEL COLORANTE VITAL AZUL TRIPÁN
El método elegido para la determinación de la viabilidad celular mediante ensayos de
exclusión de colorantes vitales, fue el de tinción con azul tripán y contaje celular en
cámara de Neubauer.
Dicha cámara, consiste en un sistema de contaje adaptado al microscopio óptico. Se
trata de un portaobjetos con una depresión en el centro, en el fondo de la cual se ha
marcado, con la ayuda de un diamante, una cuadrícula de 3 x 3 mm2, con una
separación entre dos líneas consecutivas de 0,25 mm. La depresión central del
cubreobjetos está hundida 0,1 mm respecto a la superficie, de forma que cuando la
cámara se cubre con un cubreobjetos, éste dista de la superficie marcada 0,1
milímetros, siendo el volumen comprendido entre la superficie y el cubreobjetos de 0,1
milímetro cúbico, es decir 0,1 microlitro. Esto permite hacer un cálculo del número de
células existente en un volumen fijo. Dentro de cada cuadrícula existen áreas de
contaje de 1 mm2.
La tinción con azul tripán utiliza un colorante soluble en agua, altamente tóxico, que
posee grupos amino y sulfato cargados electricamente. La utilidad de este colorante
en biología es muy importante, habiéndose utilizado este método para determinar la
viabilidad de distintos tipos de células (Alaminos et al., 2007; Hu et al., 2007). En
general, el azul tripán es capaz de teñir de color azul solamente las células muertas o
aquéllas en las que la membrana celular ha sido fragmentada. Sin embargo, las
células vivas son capaces de excluir activamente este colorante y, por tanto,
mantienen su color original translúcido o blanco.
El protocolo de trabajo que se realizó, consistió, en primer lugar, en tripsinizar los
cultivos celulares para separar las células del frasco de cultivo, centrifugándose la
mezcla para obtener un precipitado o pellet celular, el cual se diluyó en una pequeña
cantidad de medio de cultivo. A continuación, se tomaron 50 l de la suspensión
celular obtenida, la cual se incubó en un tubo tipo Eppendorf de 1,5 ml con un volumen
equivalente (50 l) de solución de azul tripán al 0,4% (Sigma-Aldrich ref. T8154)
durante 5 minutos a temperatura ambiente. Posteriormente, se tomó una muestra de
10-15 l de la mezcla que se colocó en una cámara de Neubauer (Figuras 16 y 17),
procediéndose al recuento de las células vivas (blancas, birrefringentes) y muertas
(azules) utilizando un microscopio óptico Leica Laborlux 12 (Leica, Barcelona,
España). Para el cálculo del porcentaje de células vivas se utilizó la siguiente fórmula:

Materiales y Métodos
56
Todo este proceso se llevó a cabo un total de ocho veces, de forma que en cada
subcultivo se realizó un control de la viabilidad celular con este método.
Cámara de Neubauer
Cámara de Neubauer
Canales longitudinales
Canal transversal
Figura 16.- Cámara de Neubauer. En el puente central (fondo de la cámara) están grabadas
las redes de conteo. Cuando se coloca un cubreobjetos sobre los puentes exteriores, entre la
cara inferior del cubreobjetos y el puente central de la cámara se forma una ranura capilar.
Viabilidad = [Nº células vivas/(Nº células vivas + Nº células muertas)] x 100

Materiales y Métodos
57
1. Colocar cubreobjetos en la cámara de Neubauer.
2. Adicionar 10–15 µl de la solución (suspensión celular en azul tripán).
3. Recuento de las células vivas (blancas) y de las células muertas (azules).
Figura 17.- Representación esquemática de la cámara de Neubauer utilizada para el conteo de
células viables mediante la técnica del azul tripán.

Materiales y Métodos
58
4.-ANALISIS DE LA EXPRESIÓN GÉNICA MEDIANTE MICROARRAYS
Para el análisis de expresión génica se extrajo el ARN total de los ocho primeros
subcultivos celulares de los condrocitos mantenidos en cultivo utilizando el sistema
comercial Qiagen RNeasy System (Qiagen, Mississauga, Ontario, Canadá). Una vez
extraído el ARN, se comprobó su integridad y su calidad mediante visualización directa
del ARN ribosómico de 28 y 18S en ARN total separado mediante electroforesis en
geles de agarosa al 1,2% y tinción con bromuro de etidio, así como mediante el
sistema de análisis Agilent (Agilent Technologies, Inc., Santa Clara, California, EEUU)
A continuación, todos los ARNm fueron transformados en ADNc mediante una
transcriptasa inversa (Superscript II, Life Technologies, Inc., Carlsbad, California,
EEUU) con un oligonucleótido rico en colas de timina (T7-polyT primer), el cual
permitió la amplificación de cualquier ARN mensajero presente en la célula.
Posteriormente, se sintetizaron los ARNc correspondientes a todos los ADNc mediante
transcripción in vitro, utilizando para ello UTP y CTP marcados con biotina (Enzo
diagnostics, Farmingdale, New York, EEUU). Una vez sintetizados, y para favorecer la
hibridación, estos ARNc se fragmentaron quimicamente añadiendo una concentración
elevada de sales y aplicando altas temperaturas.
Finalmente, los ARNc marcados y fragmentados se hibridaron frente a los chips que
constituyen el sistema de microarray Affymetrix Human Genome U133 plus 2.0 de la
casa comercial Affymetrix, durante 16 horas a 45ºC. Este sistema incluye 54675 genes
y secuencias génicas expresadas (EST), lo cual permite realizar un estudio global de
todos los genes y funciones génicas existentes en una célula en un momento dado.
Tras un proceso totalmente estandarizado y automatizado, los chips se lavaron y se
escanearon para obtener valores absolutos de expresión génica expresados como
unidades fluorescentes, en una escala arbitraria.

Materiales y Métodos
59
5.- ANÁLISIS ESTADÍSTICO
En la presente Tesis Doctoral se llevaron a cabo los siguientes análisis estadísticos:
1.- Análisis de la viabilidad celular determinada mediante ensayos de exclusión
de colorantes vitales. Para la comparación de los niveles de viabilidad celular
obtenidos mediante ensayos de exclusión de azul tripán, se utilizó el test no
paramétrico de Mann-Whitney, comparándose en cada caso los valores
correspondientes a dos subcultivos consecutivos (p. ej., el tercer subcultivo frente al
cuarto subcultivo)
2.- Identificación de patrones de expresión génica determinados mediante
microarray. Para determinar el perfil genético global existente en cada uno de los
subcultivos analizados, se utilizaron los programas informáticos BiNGO® y
Cytoscape® (Boyer et al., 2006). Estos programas permiten identificar aquellas
funciones génicas que son especialmente abundantes en el conjunto de genes/EST
seleccionados, calculando un valor estadístico p de acuerdo con una distribución
hipergeométrica. De este modo, el valor p calculado indica la posibilidad de que tal
enriquecimiento génico funcional ocurra por simple azar. Para ello, en primer lugar se
seleccionaron todos los genes cuya expresión superaba las 5000 unidades
fluorescentes (UF), analizándose las funciones génicas que se encontraban
sobrerrepresentadas en estos genes mediante los mencionados programas
informáticos BiNGO® y Cytoscape®.
3.- Identificación de genes cuya expresión se relaciona con la evolución
cronológica de los cultivos celulares. Para la identificación de genes cuyos niveles
de expresión se correlacionaban de forma directa o inversa con el subcultivo celular
considerado, se utilizó el test de correlación de Pearson. De este modo, se calculó el
nivel de correlación de cada uno de los genes analizados mediante microarray
respecto a los sucesivos subcultivos celulares (1 a 8), seleccionándose todos los
genes cuyos niveles de correlación R superaban un valor de 0,800 (correlación directa
con el subcultivo celular) o eran inferiores a -0,800 (correlación inversa con el
subcultivo celular).

INTRODUCCIÓN
OBJETIVOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
TABLAS SUPLEMENTARIAS
REFERENCIAS BIBLIOGRÁFICAS

Resultados
61
1.- Establecimiento de cultivos primarios de condrocitos de cartílago articular
humano
Utilizando los métodos descritos en el apartado anterior, las células cartilaginosas
crecieron de forma adecuada en los ocho subcultivos analizados en el trabajo,
alcanzando en cada uno de ellos el nivel de confluencia (Figs. 18, 19 y 20) en un
periodo de 5.5 días de media ± 2.29 (desviación típica).
En cultivo, las células cartilaginosas humanas, presentaron morfología estrellada,
poligonal o fusiforme.
Figura 18.- Imagen de microscopio óptico de contraste de fases en la que se aprecian cultivos
no confluentes de células del tejido cartilaginoso. Magnificación × 200.

Resultados
62
Figura 19.- Imagen de microscopía correspondiente a células subconfluentes del tejido
cartilaginoso.
Figura 20.- Imagen de microscopía correspondiente a células subconfluentes del tejido
cartilaginoso.

Resultados
63
2.- Viabilidad celular determinada mediante ensayos de exclusión de colorantes
vitales
Los resultados obtenidos con la técnica del azul tripán en los ocho sucesivos
subcultivos de cartílago articular humano se indican en la Tabla 3 y en la figura 21. En
dicha Tabla se muestran los valores medios en forma de porcentaje de células viables
respecto de aquellas que han incorporado el colorante.
Como se puede observar en la figura 21 existe variabilidad en cuanto a la viabilidad
celular de los diversos subcultivos celulares. Así, inicialmente podemos observar cómo
existe una tendencia a la disminución de la viabilidad celular desde el primer subcultivo
hasta el tercer subcultivo. A continuación observamos cómo a partir del subcultivo, la
tendencia se invierte hacia el aumento de la viabilidad, de forma marcada entre el
cuarto y el quinto subcultivo y de forma más moderada a partir de este hasta el octavo.
En cualquier caso, las diferencias de viabilidad no son estadísticamente significativas
(p> 0.05).
Tabla 3.- Valores medios en forma de porcentaje de células viables respecto de aquellas que
han incorporado el colorante.
PRIMER
Subcult.
SEGUNDO
Subcult.
TERCER
Subcult.
CUARTO
Subcult.
QUINTO
Subcult.
SEXTO
Subcult.
SÉPTIMO
Subcult.
OCTAVO
Subcult.
%
Super-
vivencia
95
93,16
93,18
91,39
98,09
96,70
97,22
94,78
95,23
94,65
93,75
92,60
96,07
97,97
100
100
94,44
93,73
90,69
91,47
96,57
94,58
97,22
99,23
96,51
94,51
85,78
93,97
95,80
96,77
97,50
98,80
Superv.
media
Total
95,29
94,01
89,35
92,36
96,63
96,50
97,98
98,20

Resultados
64
Fig. 21.- Porcentaje de viabilidad celular de cada uno de los subcultivos, con la técnica de azul
tripán.

Resultados
65
3.- Patrón de expresión génica global de los condrocitos articulares humanos
determinado mediante microarray
El análisis de expresión génica global ha demostrado que el número de genes cuya
expresión era superior a 5000 U.F. se relaciona con el subcultivo celular considerado,
existiendo 242, 239, 227, 312, 319, 300, 289 y 252 genes/EST (secuencias génicas
expresadas) sobreexprasados en los subcultivos celulares uno a ocho
respectivamente (figura 22). Los valores de expresión correspondientes a cada
subcultivo, para todos los genes cuya expresión era superior a 5000 UF, se muestran
en la tabla 4 (Tablas Suplementarias).
Cuando se analizaron las funciones génicas correspondientes a los genes/EST
sobreexpresados en cada subcultivo celular se comprobó la existencia de 242
funciones génicas representadas en el primer subcultivo (fig. 23 y Tabla 5), 239 en el
segundo (fig. 24 y Tabla 6), 227 en el tercero (fig. 25 y Tabla 7), 312 en el cuarto (fig.
26 y Tabla 8), 319 en el quinto (fig. 27 y Tabla 9), 300 en el sexto (fig. 28 y Tabla 10),
289 en el séptimo (fig. 29 y Tabla 11) y 252 en el octavo (fig. 30 y Tabla 12).
Fig. 22.- Número de genes/EST sobreexpresados en los ocho subcultivos celulares.

Resultados
66
Figura 23.- Representación gráfica de las 242 funciones génicas correspondientes al primer
subcultivo, que se encontraron sobreexpresadas utilizando el programa Cytoscape-BiNGO®.
Las funciones génicas que se encontraron sobrerrepresentadas con mayor nivel de
significación estadística se muestran en color anaranjado, mientras que las que no resultaron
significativas aparecen en color blanco. El amarillo indica niveles de significación intermedia.

Resultados
67
Figura 24.- Representación gráfica de las 239 funciones génicas correspondientes al segundo
subcultivo, que se encontraron sobreexpresadas utilizando el programa Cytoscape-BiNGO®.
Las funciones génicas que se encontraron sobrerrepresentadas con mayor nivel de
significación estadística se muestran en color anaranjado, mientras que las que no resultaron
significativas aparecen en color blanco. El amarillo indica niveles de significación intermedia.

Resultados
68
Figura 25.- Representación gráfica de las 227 funciones génicas correspondientes al tercer
subcultivo, que se encontraron sobreexpresadas utilizando el programa Cytoscape-BiNGO®.
Las funciones génicas que se encontraron sobrerrepresentadas con mayor nivel de
significación estadística se muestran en color anaranjado, mientras que las que no resultaron
significativas aparecen en color blanco. El amarillo indica niveles de significación intermedia.

Resultados
69
Figura 26.- Representación gráfica de las 312 funciones génicas correspondientes al cuarto
subcultivo, que se encontraron sobreexpresadas utilizando el programa Cytoscape-BiNGO®.
Las funciones génicas que se encontraron sobrerrepresentadas con mayor nivel de
significación estadística se muestran en color anaranjado, mientras que las que no resultaron
significativas aparecen en color blanco. El amarillo indica niveles de significación intermedia.

Resultados
70
Figura 27.- Representación gráfica de las 319 funciones génicas correspondientes al quinto
subcultivo que se encontraron sobreexpresadas utilizando el programa Cytoscape-BiNGO®.
Las funciones génicas que se encontraron sobrerrepresentadas con mayor nivel de
significación estadística se muestran en color anaranjado, mientras que las que no resultaron
significativas aparecen en color blanco. El amarillo indica niveles de significación intermedia.

Resultados
71
Figura 28.- Representación gráfica de las 300 funciones génicas correspondientes al sexto
subcultivo, que se encontraron sobreexpresadas utilizando el programa Cytoscape-BiNGO®.
Las funciones génicas que se encontraron sobrerrepresentadas con mayor nivel de
significación estadística se muestran en color anaranjado, mientras que las que no resultaron
significativas aparecen en color blanco. El amarillo indica niveles de significación intermedia.

Resultados
72
Figura 29.- Representación gráfica de las 289 funciones génicas correspondientes al séptimo
subcultivo, que se encontraron sobreexpresadas utilizando el programa Cytoscape-BiNGO®.
Las funciones génicas que se encontraron sobrerrepresentadas con mayor nivel de
significación estadística se muestran en color anaranjado, mientras que las que no resultaron
significativas aparecen en color blanco. El amarillo indica niveles de significación intermedia.

Resultados
73
Figura 30.- Representación gráfica de las 252 funciones génicas correspondientes al octavo
subcultivo, que se encontraron sobreexpresadas utilizando el programa Cytoscape-BiNGO®.
Las funciones génicas que se encontraron sobrerrepresentadas con mayor nivel de
significación estadística se muestran en color anaranjado, mientras que las que no resultaron
significativas aparecen en color blanco. El amarillo indica niveles de significación intermedia.

Resultados
74
4.- Identificación de los genes cuya expresión se relaciona con la evolución
cronológica de los cultivos
Como se muestra en la Tabla 13 (Tablas Suplementarias), el análisis estadístico
mediante correlación lineal reveló la existencia de 489 genes/secuencias expresadas
(EST) que se correlacionaban de forma directa con el subcultivo celular (R> 0.8),
existiendo 192 genes/EST cuya correlación con el subcultivo celular era inversa
(R < -0.8).

Resultados
75
5.- Identificación del patrón de expresión de los genes relacionados con la
diferenciación condral en los ocho primeros subcultivos celulares
Como se muestra en la Tabla 14 nuestro análisis permitió identificar los niveles de
expresión de 21 genes vinculados a la diferenciación o la actividad funcional de los
elementos celulares del cartílago articular. Dichos genes se relacionan
fundamentalmente con la síntesis de matriz extracelular cartilaginosa.
1er
Subc.
2º
Subc.
3er
Subc.
4º
Subc.
5º
Subc.
6º
Subc.
7º
Subc.
8º
Subc.
Símbolo
Gen Nombre Gen
331,5 26,5 262,5 147,6 126,3 212,1 174,2 127,7 AGC1
Agrecán 1 (condroitín sulfato
proteoglicano 1, proteoglicano de
agregación grande, antígeno
identificado por anticuerpo
monoclonal A0122)
4269,2 4495,4 6399,8 7074,5 11561,0 8356,5 5767,9 2850,4 CHI3L1 Kitinasa 3-like 1 (glicoproteína de
cartílago 39)
10,9 1,6 20,9 10,3 16,4 6,6 4,4 12,1 CHST11 Carbohidrato (condroitín 4)
sulfotransferasa 11
231,1 422,9 173,8 211,0 239,5 210,3 297,8 269,6 CHST12 Carbohidrato (condroitín 4)
sulfotransferasa 12
1,4 1,6 16,8 7,8 6,5 4,8 1,6 2,7 CHST13 Carbohidrato (condroitín 4)
sulfotransferasa 13
184,7 165,4 189,2 197,5 206,5 300,5 246,7 202,4 CHST3 Carbohidrato (condroitín 6)
sulfotransferasa 3
1009,6 732,4 276,8 350,7 489,9 296,9 802,3 722,4 CHSY1 Carbohidrato (condroitín)
sintetasa 1
77,8 50,9 216,6 148,3 145,9 159,5 46,6 57,4 CILP
Proteina de cartílago de capa
intermedia, nucleótido
pirofosfohidrolasa
47,9 6,0 43,4 60,9 64,3 80,1 32,2 25,4 CILP2 Proteina de cartílago de capa
intermedia 2
542,3 18,7 85,0 5,5 13,1 46,3 7,1 9,0 COL2A1
Colágeno tipo II, alfa 1
(osteoartritis primaria, displasia
espondiloepifisaria congénita)
3596,2 915,9 1762,3 1378,2 814,6 443,3 3649,7 690,2 COMP Proteína oligomérica de matriz
cartilaginosa
57,3 13,5 105,6 22,3 35,2 32,9 7,1 8,7 CRTAC1 Proteína acídica cartilaginosa 1
793,8 880,9 823,1 838,1 1026,1 1091,3 882,3 778,6 CRTAP Proteína asociada a cartílago
260,8 223,4 223,6 307,8 256,5 271,3 241,4 247,2 CSGlcA-T Condroitín sulfato
glucuroniltransferasa

Resultados
76
2625,3 2539,1 1536,5 2627,5 4283,6 1982,6 2871,0 1239,5 CSPG2 Condroitín sulfato proteoglicano
2 (Versicán)
9,8 4,5 9,6 5,6 12,2 36,3 9,2 7,0 CSPG3 Condroitín sulfato proteoglicano
3 (Neurocán)
55,4 70,4 68,1 93,9 29,4 35,8 71,0 92,7 CSPG4 Condroitín sulfato proteoglicano
4
62,4 12,4 21,3 24,8 48,8 33,8 44,5 57,4 CSPG6 Condroitín sulfato proteoglicano
6 (Bamacán)
227,4 199,0 112,5 102,9 204,9 114,9 247,7 164,1 CSS3 Condroitín sulfato sintetasa 3
477,8 410,0 204,2 243,0 281,6 172,8 405,3 331,2 GALNACT-
2 Condroitín sulfato GalNAcT-2
7,1 9,2 32,2 37,1 16,6 37,6 19,0 17,7 MATN1 Matrilín 1, proteína de matriz
cartilaginosa
Tabla 14.- Niveles de expresión de 21 genes vinculados a la diferenciación o a la actividad
funcional de los elementos celulares del cartílago articular.

Resultados
77
6.- Identificación del patrón de expresión de genes vinculados con la mortalidad
celular (caspasas) en los ocho subcultivos celulares
El análisis de expresión génica mediante la técnica de microarray permitió identificar
los niveles de expresión de los genes/EST relacionados con la mortalidad celular. A tal
efecto, se consideraron los genes/EST de las caspasas (familia de proteasas Cisteil
Aspartato proteasas, que median fenómenos de apoptosis) como indicadores de
mortalidad celular (Perales S, 2007), para cualquier valor de expresión, identificándose
de esta forma 34 EST relacionadas con la mortalidad celular (Tabla 15).
Agrupando las EST por el gen del que se habían obtenido, se elaboró una Tabla que
refleja las medias de expresión relacionadas con la mortalidad celular (Tabla 16).
Considerando las medias de los niveles de expresión de los genes relacionados con la
mortalidad en cada subcultivo celular, podemos diferenciar tres grupos diferentes: un
primer grupo que engloba a los genes Casp1, Casp3, Casp6, Casp8AP2 y Casp8 en el
que los niveles de expresión siguen un modelo en ―V‖ ; un segundo grupo con los
genes Casp9, Casp10, Casp12 y Casp14 en el que los niveles de expresión siguen un
modelo en ―V invertida‖; y un tercer grupo inespecífico con los genes Casp2, Casp4,
Casp5, Casp7 y Casp11. A modo de ejemplo en las figuras 31, 32 y 33 se representa
la distribución de los genes 3, 9 y 5, representativos de cada uno de los grupos.

Resultados
78
Tabla 15.- Niveles de expresión de los 34 EST vinculados a la mortalidad de los condrocitos
en los diferentes subcultivos celulares.
MEDIA SUBC1
MEDIA SUBC 2
MEDIA SUBC 3
MEDIA SUBC 4
MEDIA SUBC 5
MEDIA SUBC 6
MEDIA SUBC 7
MEDIA SUBC 8
Probe Set ID
Símbolo Gen
Nombre Gen
66,5 49,4 47,7 43,3 28,2 56,4 48,7 72,4 211367_s_at
CASP1
caspase 1, apoptosis-related cysteine
peptidase (interleukin 1, beta, convertase)
214,3 119,0 243,1 124,5 124,8 148,6 124,4 177,7 211366_x_at
CASP1
caspase 1, apoptosis-related cysteine
peptidase (interleukin 1, beta, convertase)
132,8 112,3 54,5 41,2 70,3 52,5 81,9 165,4 211368_s_at
CASP1
caspase 1, apoptosis-related cysteine
peptidase (interleukin 1, beta, convertase)
209,5 191,6 85,7 92,2 131,8 155,9 146,7 277,4 209970_x_at
CASP1
caspase 1, apoptosis-related cysteine
peptidase (interleukin 1, beta, convertase)
28,8 44,1 33,3 27,9 50,5 47,4 48,5 49,1 206011
_at CASP1
caspase 1, apoptosis-related cysteine
peptidase (interleukin 1, beta, convertase)
15,6 7,5 413,5 161,3 47,9 22,7 5,3 13,4 210708_x_at
CASP10 caspase 10,
apoptosis-related cysteine peptidase
7,5 9,7 65,0 38,1 17,1 13,4 10,6 14,6 205467
_at CASP10
caspase 10, apoptosis-related
cysteine peptidase
6,5 6,2 157,5 138,2 50,7 125,1 5,2 4,2 211888_x_at
CASP10 caspase 10,
apoptosis-related cysteine peptidase
8,7 8,1 65,6 58,5 52,7 41,4 12,8 17,1 210955
_at CASP10
caspase 10, apoptosis-related
cysteine peptidase
3,7 0,3 15,0 20,1 12,9 12,6 3,4 6,4 1564736_a_at
CASP12 caspase 12
(gene/pseudogene)
1,8 1,1 37,6 18,7 5,6 13,1 7,4 3,8 231722
_at CASP14
caspase 14, apoptosis-related
cysteine peptidase
21,9 12,3 16,8 18,3 10,5 33,3 19,1 23,6 208050_s_at
CASP2 caspase 2, apoptosis-
related cysteine peptidase
259,8 257,0 178,9 192,7 206,1 227,6 240,3 268,7 226032
_at CASP2
caspase 2, apoptosis-related cysteine
peptidase
10,0 9,0 17,6 23,2 9,7 9,3 9,7 9,6 34449_
at CASP2
caspase 2, apoptosis-related cysteine
peptidase
1,8 1,0 39,2 10,7 11,9 15,5 2,9 2,5 211140_s_at
CASP2 caspase 2, apoptosis-
related cysteine peptidase
72,1 22,3 153,3 119,2 101,9 131,8 39,5 38,9 226036_x_at
CASP2 caspase 2, apoptosis-
related cysteine peptidase
9,1 13,7 39,8 21,6 28,6 36,2 21,5 24,4 209811
_at CASP2
caspase 2, apoptosis-related cysteine
peptidase
1,8 1,1 13,4 11,6 11,4 23,3 2,1 1,6 209812_x_at
CASP2 caspase 2, apoptosis-
related cysteine peptidase
262,4 258,8 130,0 160,2 192,3 212,7 192,6 186,3 202763
_at CASP3
caspase 3, apoptosis-related cysteine
peptidase
5,3 5,3 15,4 36,2 11,9 15,8 9,4 1,6 236729
_at CASP3
Cysteine protease Yama
12,9 23,5 23,5 6,7 26,7 29,3 30,3 31,0 213596
_at CASP4
caspase 4, apoptosis-related cysteine

Resultados
79
peptidase
149,4 218,1 196,4 238,9 231,0 253,1 143,1 176,0 209310_s_at
CASP4 caspase 4, apoptosis-
related cysteine peptidase
4,8 5,0 20,5 11,2 15,2 15,9 10,6 16,8 207500
_at CASP5
caspase 5, apoptosis-related cysteine
peptidase
152,7 125,8 66,8 99,4 79,0 107,7 108,0 137,2 209790_s_at
CASP6 caspase 6, apoptosis-
related cysteine peptidase
112,8 136,9 92,4 83,9 120,7 106,0 126,7 163,9 211464_x_at
CASP6 caspase 6, apoptosis-
related cysteine peptidase
173,2 150,7 193,0 237,3 171,3 234,6 194,6 201,0 207181_s_at
CASP7 caspase 7, apoptosis-
related cysteine peptidase
218,4 167,1 129,1 112,9 149,7 142,7 165,4 222,2 213373_s_at
CASP8 caspase 8, apoptosis-
related cysteine peptidase
62,1 10,1 47,7 47,3 44,4 61,5 10,8 28,5 207686_s_at
CASP8 caspase 8, apoptosis-
related cysteine peptidase
4,5 1,9 14,2 6,1 4,7 7,4 2,2 5,6 155330
6_at CASP8
caspase 8, apoptosis-related cysteine
peptidase
169,8 102,9 58,1 56,3 95,2 56,6 106,3 112,4 222201_s_at
CASP8AP2
caspase 8 associated protein 2
5,1 3,4 40,5 27,5 11,5 12,5 6,5 6,2 157000
1_at CASP8A
P2 caspase 8 associated
protein 2
16,0 12,7 29,1 22,8 14,1 3,4 5,3 5,3 240437
_at CASP9
caspase 9, apoptosis-related cysteine
peptidase
90,0 98,9 222,9 106,8 125,5 95,2 89,6 102,0 203984_s_at
CASP9 caspase 9, apoptosis-
related cysteine peptidase
70,0 56,6 107,5 133,8 110,7 132,3 65,8 69,6 210775_x_at
CASP9 caspase 9, apoptosis-
related cysteine peptidase

Resultados
80
Tabla 16.- Medias de los niveles de expresión por genes de los EST vinculados a la mortalidad
de los condrocitos en los diferentes subcultivos celulares.
MEDIA SUBC1
MEDIA SUBC 2
MEDIA SUBC 3
MEDIA SUBC 4
MEDIA SUBC 5
MEDIA SUBC 6
MEDIA SUBC 7
MEDIA SUBC 8
Probe Set ID
Símbolo Gen
Nombre Gen
130,4 103,3 92,8 65,8 81,1 92,1 90,0 148,4 MEDIA CASP1
Caspasa 1, Cistein peptidasa
relacionada con la apoptosis
53,8 45,2 65,5 56,7 54,3 68,1 47,8 52,7 MEDIA CASP2
Caspasa 2, Cistein peptidasa
relacionada con la apoptosis
133,9 132,1 72,7 98,2 102,1 114,3 101,0 93,9 MEDIA CASP3
Caspasa 3, Cistein peptidasa
relacionada con la apoptosis
81,1 120,8 109,9 122,8 128,9 141,2 86,7 103,5 MEDIA CASP4
Caspasa 4, Cistein peptidasa
relacionada con la apoptosis
4,8 5,0 20,5 11,2 15,2 15,9 10,6 16,8 207500
_at CASP5
caspase 5, apoptosis-related
cysteine peptidase
132,7 131,4 79,6 91,6 99,9 106,8 117,3 150,6 MEDIA CASP6
Caspasa 6, Cistein peptidasa
relacionada con la apoptosis
173,2 150,7 193,0 237,3 171,3 234,6 194,6 201,0 207181_s_at
CASP7 caspase 7,
apoptosis-related cysteine peptidase
95,0 59,7 63,6 55,4 66,3 70,5 59,5 85,4 MEDIA CASP8
Caspasa 8, Cistein peptidasa
relacionada con la apoptosis
87,5 53,2 49,3 41,9 53,3 34,5 56,4 59,3 MEDIA CASP8A
P2 Proteina asociada a
Caspasa 8
80,0 77,8 165,2 120,3 118,1 113,7 77,7 85,8 MEDIA CASP9
Caspasa 9, Cistein peptidasa
relacionada con la apoptosis
9,6 7,9 175,4 99,0 42,1 50,6 8,5 12,3 MEDIA CASP10
Caspasa 10, Cistein peptidasa
relacionada con la apoptosis
3,7 0,3 15,0 20,1 12,9 12,6 3,4 6,4 1564736_a_at
CASP12 caspase 12
(gene/pseudogene)
1,8 1,1 37,6 18,7 5,6 13,1 7,4 3,8 231722
_at CASP14
caspase 14, apoptosis-related
cysteine peptidase

Resultados
81
Fig. 31.- Gen CASP3 cuyos niveles de expresión por subcultivo celular sigue una distribución
en ―V‖.
Fig. 32.- Gen CASP9 cuyos niveles de expresión por subcultivo celular sigue una distribución
en ―V invertida‖.

Resultados
82
Fig. 33.- Gen CASP5 cuyos niveles de expresión por subcultivo celular no sigue un modelo de
distribución concreto.

INTRODUCCIÓN
OBJETIVOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
TABLAS SUPLEMENTARIAS
REFERENCIAS BIBLIOGRÁFICAS

Discusión
84
El cartílago articular aporta a las articulaciones sinoviales la capacidad de
movilidad sin dolor y con baja fricción. El grosor, densidad celular, composición de la
matriz y propiedades mecánicas difieren en una misma articulación y entre
articulaciones diferentes. Aún así, en todas las articulaciones sinoviales, el cartílago
tiene los mismos componentes, la misma estructura general y realiza las mismas
funciones. Con un grosor de pocos milímetros, tiene una sorprendente resistencia a la
compresión y una excepcional capacidad para distribuir las cargas, reduciendo las
presiones máximas sobre el hueso subcondral. Quizás la característica más llamativa
del cartílago articular, sea su gran durabilidad, que permite en muchas personas una
función articular normal durante 80 o más años. (Norman Scott W, 2007).
El cartílago articular carece de suministro sanguíneo o linfático. No tiene elementos
nerviosos y está aislado del sistema inmunológico. Así las cosas, la capacidad de
curación de este tejido está limitada por su escasa capacidad regenerativa (Hannallah
et al., 2003; O’Driscoll, 1998; Sellars et al., 1997).
Las opciones terapéuticas de las alteraciones del cartílago articular pueden resumirse
en: restablecer, reemplazar, liberar y resecar. En el pasado, el tratamiento consistía en
la resección. Avances recientes en trasplante de cartílago articular (O’Driscoll, 1998),
así como nuevas modalidades esperanzadoras como la terapia génica (Hannallah et
al., 2003; O’Driscoll, 1998; Sellars et al., 1997), nos llevan a pensar que es posible
superar este problema de ausencia de respuesta reparadora (Buckwalter, 2003).

Discusión
85
Para solucionar los problemas de la reparación del cartílago articular es necesario
incorporar la ciencia básica a la práctica clínica (Buckwalter, 2003). El conocimiento de
la ciencia básica del cartílago articular, que abarca la anatomía, histología, la función
biomecánica y la respuesta reparadora, es el sustrato para desarrollar un tratamiento
efectivo de las lesiones y trastornos de este tejido.
La Ingeniería Tisular, constituye una nueva disciplina dentro del campo de la medicina,
estrechamente ligada al conocimiento y desarrollo de la histología, que está centrada
en la creación de tejidos artificiales para su utilización como herramientas terapéuticas
(Campos A, 2004; Alaminos et al., 2007b). Para conseguir este objetivo se precisa una
fuente de células que sean funcionales y viables (Alaminos et al., 2007a), así como
biomateriales y señales moleculares de diversa naturaleza. De esta forma, para crear
tejidos artificiales, necesitamos aislar células con capacidad proliferativa, que se
obtienen habitualmente de biopsias de tejido adulto.
La Ingeniería Tisular aplicada al tejido cartilaginoso, considerando la alta prevalencia
de la patología del cartílago articular, constituye en estos momentos una de las
herramientas terapéuticas más prometedoras de cara al futuro. En este contexto, se
han venido desarrollando en los últimos años diferentes modelos para la formación de
nuevo tejido cartilaginoso, que van desde la terapia celular con condrocitos aislados
hasta la elaboración de constructos cartilaginosos tridimensionales que incorporan
células cartilaginosas a distintos biomateriales, añadiéndose en algunos protocolos
distintos factores de crecimiento (factor de transformación beta o TGF- βs) (Tuli et al.,
2003; Steinert et al., 2007). La terapia celular y los distintos constructos de cartílago
han generado resultados muy diversos, habiéndose descrito, entre los problemas más
significativos, la insuficiente diferenciación celular, la pérdida del número de células
implantadas, la degeneración de las matrices utilizadas como biomateriales y la falta
de integración del nuevo tejido en el sujeto que lo recibe (Steinert et al., 2007). En este
contexto, Goldstein afirma que el éxito de cualquier protocolo de Ingeniería Tisular
exige una adecuada combinación de cuatro factores: las células, los factores de
crecimiento, los biomateriales y las condiciones ambientales.
Como se expuso en la introducción de esta Tesis Doctoral, durante las últimas
décadas se han desarrollado numerosas técnicas para el cultivo de distintos tipos de
células humanas y animales en laboratorio.
Destacan en este sentido, como hemos indicado en la introducción de la presente
Tesis, los trabajos pioneros de Chesterman y Smith en los años sesenta del pasado

Discusión
86
siglo (Chesterman y Smith, 1968), los de Bentley y Green en los años setenta
(Bentley, 1971; Green, 1977), los de Peterson en los ochenta (Peterson y cols., 1984)
y el trabajo pionero de Brittberg en los noventa (Brittberg et al., 1995) al realizar un
estudio piloto con veintitrés pacientes a los que inyecta condrocitos autólogos aislados,
trabajo que se publica en el New England Journal of Medecine y que inaugura la
aplicación generalizada de la terapia celular con condrocitos.
Numerosos han sido los autores, que en los últimos años han seguido trabajando en el
campo de la Ingeniería Tisular relacionada con el cartílago articular, mostrando
importantes avances: Pineda (Pineda y Merentes, 2004), Darling (Darling et al., 2004),
Richmond (Richmon et al., 2005), Glowacki (Glowacki et al., 2005), Oliveira (Oliveira et
al., en 2006), Jiang (Jiang y Chiang, 2007), Saris (Saris y Vanlauwe, 2008).
Aunque la terapia celular con condrocitos parece útil, los resultados de los estudios
son heterogéneos. Uno de los estudios más recientes sobre la terapia celular en las
lesiones del cartílago articular de la rodilla de Norimasa Nakamura (Norimasa
Nakamura et al., 2009) afirma que no existe suficiente evidencia en los estudios
actuales para determinar cúando la terapia celular es superior a otros tratamientos en
la patología del cartílago articular de la rodilla. Algunas de las limitaciones de la terapia
celular con condrocitos son la alta diferenciación y la baja capacidad proliferativa de
los cultivos.
Estudios recientes, utilizando técnicas clásicas de evaluación de la viabilidad celular,
como el ensayo de exclusión del colorante vital azul tripán y técnicas de microscopía
analítica, parecen demostrar la heterogeneicidad, en lo que a viabilidad se refiere, de
las células procedentes de los distintos subcultivos utilizados en los protocolos de
ingeniería tisular. En efecto, Rodríguez-Morata en su trabajo con células endoteliales
procedentes de vena umbilical humana (Rodríguez Morata et al., 2008), demuestra
que la máxima viabilidad celular corresponde al tercer subcultivo. Alaminos, en su
trabajo con células endoteliales de cornea animal (Alaminos et al., 2007) demostró que
las células del cuarto subcultivo eran las que presentaban mejores indicadores de
viabilidad. Montalvo, en su Tesis Doctoral sobre la utilización de células madre
procedentes de la gelatina de Wharton en Ingeniería Tisular (Montalvo, 2008a),
concluye que los subcultivos celulares más viables son el cuarto y el quinto.
En relación con el cartílago existen pocos estudios relativos a la evaluación de la
viabilidad. Montalvo, en un trabajo sobre la viabilidad de condrocitos procedentes de

Discusión
87
cartílago fibroso animal (Montalvo et al., 2008), concluye que el subcultivo más viable
es el tercero.
De acuerdo con los objetivos establecidos en la presente Tesis Doctoral, planteamos
la discusión analizando en primer lugar el aislamiento de condrocitos y la obtención de
subcultivos celulares a partir de biopsias de cartílago articular humano. En segundo
lugar, analizamos el estudio clásico de evaluación de viabilidad celular, el azul tripán,
que se realizó en cada uno de los ocho subcultivos celulares del estudio. Finalmente
procedemos al análisis de los diferentes patrones genéticos identificados, en relación
con cada uno de los subcultivos celulares, examinando sucesivamente los conjuntos
de genes implicados tanto en la diferenciación condrogénica como en la mortalidad
celular.
Como ya se expuso en el apartado de Materiales y Métodos de la presente Tesis
Doctoral, los condrocitos articulares se obtuvieron de biopsias de rodilla humana
durante el acto quirúrgico consistente en la sustitución protésica de dicha articulación.
Tras la incubación de las biopsias en una disolución de colagenasa, se consiguieron
aislar los condrocitos articulares humanos. Estos condrocitos fueron cultivados en
frascos con medio QN (De Diego et al., 2004; Llames et al., 2004) hasta que
alcanzaron la semiconfluencia, momento en el que los condrocitos se separaron del
frasco y se volvieron a sembrar en medio QN. Esta sistemática se repitió un total de
ocho veces, obteniéndose ocho subcultivos celulares.
Con el objetivo de determinar la viabilidad de los condrocitos mantenidos en cultivo, se
estudió el fenotipo morfológico, mediante técnicas de microscopía óptica y el fenotipo
genético, a través de la identificación de patrones de expresión génica.
En primer lugar, se utilizó la técnica clásica de exclusión de colorantes vitales, es
decir, el azul tripán, con el objeto de sistematizar el patrón de viabilidad que se utiliza
básicamente en cualquier laboratorio. El proceso de muerte celular programada se
caracteriza, a nivel morfológico, por la sucesión de una serie de signos concretos,
como son: las alteraciones de la membrana plasmática, las cuales conllevan la pérdida
de contacto con las células vecinas y la adquisición de una forma redondeada; la
condensación y fragmentación de la cromatina nuclear sin que se pierda la envoltura
nuclear; la disgregación del núcleo en masas pequeñas de cromatina, con una
disminución paralela del volumen citoplasmático secundario a una pérdida de agua y a
la condensación de las proteínas, permaneciendo intacta la mayoría de los orgánulos
celulares y, finalmente, la rotura de la célula en vesículas rodeadas de membrana

Discusión
88
llamadas cuerpos apoptóticos que serán fagocitados por los macrófagos o por células
vecinas (Choy et al., 2001).
Tal y como defienden la mayor parte de autores, la técnica del azul tripán sólo es
capaz de identificar aquellas células que se encuentran en una fase avanzada de
apoptosis, puesto que durante las distintas fases se mantiene la integridad de la
membrana celular (Alaminos et al., 2007).
Una vez realizada la técnica del azul tripán en los diferentes subcultivos celulares
analizados en la presente Tesis Doctoral, se pudo constatar la variabilidad existente en
relación con la viabilidad celular en dichos subcultivos celulares. De esta forma se
observa una tendencia a la disminución de la viabilidad entre el primer subcultivo
celular (correspondiente al cultivo primario) y el tercero, tendencia esta, que se invierte
a partir del cuarto subcultivo celular, con un aumento de la viabilidad moderado entre
el tercer y el cuarto subcultivo y más significativa a partir de este último.
De estos resultados se puede inferir que los condrocitos articulares humanos
presentan al inicio, un periodo de adaptación al cultivo en el que un importante número
de ellos no sobrevive a las condiciones del medio. A partir del cuarto subcultivo
celular, los condrocitos se recuperan, observándose un incremento del nivel de
viabilidad hasta el octavo subcultivo celular.
No obstante, la tasa de viabilidad celular en cultivo primario de los condrocitos
articulares humanos es alta, aún habiendo sufrido un enérgico procesamiento
mecánico y enzimático durante el aislamiento. A este respecto podemos destacar la
diferencia con las células condrales animales, tal y como demostró Rosales en su
Tesis Doctoral (Rosales, 2008). De esta forma, Rosales evidenció, que los cultivos
primarios de condrocitos articulares animales, presentan un nivel de viabilidad celular
medio más bajo que el de los condrocitos articulares humanos, cuando se utiliza la
técnica del azul tripán.
Continuando con el estudio de la viabilidad celular de los condrocitos articulares
humanos, el segundo método utilizado en la presente Tesis Doctoral para este
propósito fue el análisis de la expresión génica mediante microarrays de
oligonucleótidos en las condrocitos de todos los subcultivos celulares.
Actualmente, la utilización del microarrays de oligonucleótidos, se presenta como una
magnífica herramienta para identificar los genes que se expresan en las diferentes
etapas funcionales de la célula como manifestación de la actividad celular.

Discusión
89
En la presente Tesis Doctoral, se ha insistido en la identificación de los patrones de
expresión génica de los diferentes subcultivos celulares, prestando especial atención a
los patrones relacionados con la diferenciación condral y la mortalidad y estableciendo
una correlación entre el número de subcultivo y su patrón de expresión génica.
Debido al elevado número de genes/secuencias expresadas (EST) en el estudio ha
sido necesaria la utilización de programas informáticos capaces de integrar en
conjunto toda la información. Gracias a ello hemos podido clasificar los resultados
atendiendo a las dos premisas anteriormente citadas, capacidad de diferenciación y
mortalidad.
Inicialmente se analizó el patrón de expresión génica global, observándose cómo el
número de genes/EST sobreexpresados, varía en función del número de subcultivo
celular. Así podemos comprobar cómo inicialmente, el número de genes/EST
sobreexpresados, desciende levemente hasta el tercer subcultivo celular, momento en
el que aumenta, alcanzando su máximo valor en el quinto subcultivo celular, para
luego volver a disminuir progresivamente.
Por otra parte, se observó la existencia de 489 genes/secuencias expresadas (EST)
que se correlacionaban de forma directa con el número de subcultivo celular y de 192
genes/EST cuya correlación con el número de subcultivo celular era inversa, lo que
demuestra la heterogeneidad de los subcultivos y la importancia de evaluar, a efectos
de la terapia celular, el subcultivo que reúne mayor viabilidad y mayor potencial
condrogénico.
Entre los genes cuya expresión se relaciona de forma directa con el subcultivo celular,
vamos a considerar cuatro tipos: los relacionados con el metabolismo celular, con las
moléculas de adhesión, con la proliferación y división celular y con la apoptosis.
En relación con los genes vinculados al metabolismo celular, cabe destacar tres
secuencias relacionadas con el gen ADAMTS 5, también conocido como agrecanasa
2. Este enzima pertenece a la familia ADAMTS (desintegrinas y metaloproteasas con
dominios tromboespondina) y se encarga de la degradación del Agrecan, que como
sabemos es el proteoglucano más abundante en el cartílago articular (Stanton H,
2005). Los niveles de expresión de ADAMTS 5 ya han sido relacionados en el pasado
con la capacidad condrogénica, de forma que los condrocitos con baja capacidad
condrogénica, expresan altos niveles de agrecanasa 2 (Grogan SP et al., 2007). De
esta forma, analizando los niveles de expresión de ADAMTS 5 en nuestro trabajo,
podemos afirmar que, aunque la tendencia es al aumento de la expresión con el

Discusión
90
subcultivo celular, en el segundo y tercer subcultivo los niveles de expresión son
mínimos, lo que posiblemente indica que son estos los subcultivos con mayor
capacidad condrogénica. En cualquier caso, el gen ADAMTS 5, resulta un marcador
de capacidad condrogénica, cuya utilidad debe ser tenida en cuenta en la terapia
celular con cartílago.
Otra de las secuencias/genes que aumentan su expresión con el subcultivo celular es
CADM3, que codifica la molécula de adhesión celular 3. En nuestro trabajo
observamos cómo su expresión disminuye en el segundo subcultivo para aumentar
luego progresivamente y disminuir de nuevo en el último subcultivo.
En relación con la proliferación y la división celular, destacamos el gen CDC42EP3
(CDC42 proteína efectora 3). En nuestro estudio, este gen sigue un comportamiento
idéntico a CADM3, disminuye su expresión en el segundo subcultivo y luego aumenta
progresivamente hasta el séptimo, disminuyendo en el octavo.
El perfil de expresión de estos dos genes pone de relieve que en los sucesivos
subcultivos a partir del segundo, existe un proceso de proliferación y división celular
activo, así como una tendencia a la adhesión intercelular que habría que relacionar
con la formación de grupos isogénicos en el desarrollo tisular del tejido cartilaginoso.
Por último, en relación con los genes cuya expresión se incrementa con el subcultivo
celular, encontramos genes de apoptosis, como por ejemplo el BAG4, cuya expresión,
sigue un aumento constante, salvo una leve disminución en el subcultivo 6. En
relación con los genes de apoptosis discutiremos más tarde su incidencia en la
viabilidad celular.
De entre los 192 genes/EST cuyos niveles de expresión disminuyen en relación con el
subcultivo celular (relación inversa) consideraremos a continuación algunos genes
relacionados así mismo con la adhesión y con la proliferación celular.
La molécula de adhesión CADM1, va disminuyendo sus niveles de expresión
progresivamente, con un leve repunte en el séptimo subcultivo. Este dato hay que
relacionarlo con el equilibrio entre las distintas moléculas de adhesión que participan
en el proceso de configuración y conformación de una estructura tisular.
Entre los genes relacionados con la proliferación celular, destacamos G0S2, FGF13 y
MAPK10. La expresión de G0S2, posibilita que la célula avance en el ciclo celular,
entrando en fase G1. Su máxima expresión en nuestro trabajo es en el primer y tercer
subcultivo, aunque en conjunto va disminuyendo con el número de subcultivos. El gen

Discusión
91
FGF13, codifica el factor de crecimiento fibroblástico 13 y al igual que el anterior tiene
dos picos de expresión, correspondientes en este caso al primer y cuarto subcultivo.
Por último MAPK10 codifica una proteína quinasa activadora de la mitogénesis. Sus
niveles de expresión, aunque siguen un descenso progresivo, se mantienen más
constantes que en los casos de los genes anteriores.
En relación con el potencial diferenciativo de los distintos subcultivos celulares, se
identificaron los niveles de expresión de 21 genes/EST vinculados a la diferenciación o
a la actividad funcional de los condrocitos articulares. Dichos genes se relacionan
fundamentalmente con la síntesis de matriz extracelular cartilaginosa, correspondiendo
más de la mitad de ellos a la síntesis de condroitín sulfato.
El gen para el Agrecán 1, el más común de los proteoglucanos del cartílago articular,
tiene su máximo nivel de expresión en el primer subcultivo, disminuyendo
posteriormente, aunque existe un importante pico de expresión en el subcultivo 3.
El grupo de genes relacionado con la síntesis de condroitín sulfato,
glucosaminoglucano más frecuente en el cartílago articular, agrupa doce genes que
siguen una distribución con niveles de expresión similares, excepto para el condroitín
sulfato 2 y el condroitín sintetasa, cuyos niveles son muy superiores al resto. El
modelo de expresión de todos ellos se caracteriza por un descenso inicial en la
expresión génica en los primeros subcultivos, y una cierta tendencia al aumento en los
últimos. Destaca sin embargo, el incremento de expresión de condroitín sulfato 4
sulfotransferasa 11.
En cuanto al gen del colágeno tipo II, hemos constatado cómo sufre una drástica
disminución en su expresión desde el primer subcultivo. De forma que los valores de
expresión en los subcultivos 2 al 8 son sensiblemente inferiores a los del cultivo inicial,
si bien existe un pico de elevación en el subcultivo número tres.
Por último, el gen encargado de la síntesis de la proteína de matriz cartilaginosa 1,
Matrilín 1, presenta unos niveles de expresión que van aumentando progresivamente
con el subcultivo celular, y aumentando significativamente a partir del subcultivo
número tres y disminuyendo posteriormente desde el subcultivo número seis.
En relación con los genes vinculados al potencial diferenciativo, nuestro estudio pone
de relieve que en torno al subcultivo número tres, se incrementa la expresión de
algunos genes vinculados a la síntesis de matriz cartilaginosa (agrecán, condroitín
Sulfato-4 sulfotransferasa11, colágeno II y matrilín), circunstancia esta, que coincide a

Discusión
92
la vez con la disminución en la expresión de los niveles de agrecanasa 2, en el
subcultivo número tres, como describimos con anterioridad. Es evidente la relación
que existe entre baja expresión de agrecanasa 2 y mayor potencial de diferenciación
condral en torno al subcultivo número tres.
Por último, se observó en todos los subcultivos celulares la expresión de genes
responsables de la síntesis del colágeno tipo I y tipo VI. Resulta llamativa la expresión
de estos dos tipos de colágeno, que en condiciones normales no son abundantes en el
cartílago articular en estado de salud (Couceiro et al., 2003), si bien su incremento
está descrito en determinadas circunstancias (edad, sobrecarga…) (So et al., 2001). Si
el incremento de la expresión de los genes que sintetizan el colágeno tipo I y VI está o
no relacionado con la presencia de alguna de esas circunstancias (edad, etc.) es algo
que habrá que determinar en el futuro cuando estudios semejantes a los desarrollados
en la presente Tesis se realicen en otras series de edades y situaciones diferentes.
En cuanto a la expresión de genes relacionados con la mortalidad celular, se
consideraron los genes/EST de las caspasas (familia de proteasas Cisteil Aspartato
proteasas, que median fenómenos de apoptosis) como indicadores de este proceso
celular (Perales S, 2007). De esta forma, mediante la técnica de microarray, se
identificaron 34 EST relacionadas con la mortalidad celular.
Se conocen 14 tipos de caspasas (Thornberry N, 1998), divididas en dos grupos:
iniciadoras y efectoras que actúan en cascada (Shi, 2002). De entre las iniciadoras (1,
2, 4, 5, 8, 9, 10, 12, 13) la más importante es la caspasa 9 (Li P, 1997) y de entre las
efectoras (3, 6, 7, 14) la 3 (Stennicke HR, 1998).
Las caspasas se sintetizan como zimógenos y necesitan de una activación proteolítica
(Rotonda J, 1996; Wilsonm KP, 1994). Tridimensionalmente, las caspasas constan de
un predominio, una subunidad larga y otra pequeña. La activación consiste en la
separación de la subunidad larga de la pequeña y la eliminación del predominio. Es el
tipo de predominio lo que determina que la caspasa sea efectora (no tiene predominio
o es muy corto) o iniciadora (predominio largo). (Chang Hi, 2000; Nicholson D et al.,
1995; Thornberry NA, 1992).
Una vez llega la señal apoptótica, se activan muchas caspasas mediante una cascada
de activación secuencial, lo cual sirve para amplificar la señal apoptótica y además
permite la regulación a varios niveles. Los mecanismos de activación pueden ser:
transactivaciones o autoactivaciones. (Chang Hi, 2000; Ashkenazi A, 1998; Hueber
AO, 1997; Li P, 1997).

Discusión
93
La transactivación se produce de forma secuencial y son las caspasas iniciadoras las
que activan a las efectoras, estas amplifican la señal y ejecutan la muerte celular
actuando sobre proteínas estructurales y reguladoras, lo que lleva al colapso celular. A
modo de ejemplo, caspasa 8, 9 y 10 activan a caspasa 3 y 7. Caspasa 3 puede
activar a caspasa 9, con lo que se produce un bucle de amplificación (Slee EA, 1999;
Srinivasula SM, 1998). La autoactivación se produce principalmente por agregación
(Colossi PA, 1998).
Entre las acciones de las caspasas hay que destacar la inactivación de proteínas del
citoesqueleto, la acción sobre proteínas asociadas al DNA, la liberación de moléculas
apoptóticas mitocondriales, la producción de moléculas proapoptóticas y el bloqueo de
la transcripción de genes de supervivencia, etc.
La inhibición de las caspasas puede darse de dos formas: mediante inactivación por
interacción de proteínas (Roy N, 1997; Deveraux QL, 1997; Deveraux QL, 1998; Irmle
M, 1997; Koseki T 1998) o por inactivación por modificación proteica (Hermann C,
2000; Manick JB, 1999).
En el estudio realizado en nuestra Tesis, agrupando las EST por genes y obteniendo
las medias de expresión para cada subcultivo celular, identificamos tres grupos
diferentes, según el modelo de expresión que siguen. Un primer grupo que engloba a
los genes Casp1, Casp3, Casp6, Casp8AP2 y Casp8, en el que la expresión
disminuye y luego aumenta conforme va avanzando el subcultivo celular (modelo en
―V‖); un segundo grupo con los genes Casp9, Casp10, Casp12 y Casp14 en el que los
niveles de expresión aumentan y luego disminuyen, siguiendo un modelo en ―V
invertida‖; y un tercer grupo inespecífico con los genes Casp2, Casp4, Casp5, Casp7 y
Casp11.
Siguiendo con el análisis de los niveles de expresión de las caspasas, podemos
observar como caspasa 9, ejemplo fundamental de caspasa iniciadora, alcanza su
máximo nivel de expresión en el subcultivo 3. Considerando, como ya hemos dicho
anteriormente, que caspasa 9 es iniciadora, podemos afirmar que es en el subcultivo
3, cuando el estímulo para desencadenar el proceso de mortalidad celular, alcanza su
máximo nivel.
Por otra parte, podemos observar cómo los niveles de expresión de caspasa 3,
ejemplo de efectora, son mínimos en el subcultivo 3, y van aumentando
progresivamente conforme se aumenta de subcultivo.

Discusión
94
Analizando la expresión de las caspasas globalmente, existe un comportamiento
inverso entre el grupo de las caspasas iniciadoras y el de las efectoras, que queda de
manifiesto en la representación gráfica de ambos grupos, en la que se observa como
uno es el equivalente especular del otro. De esta forma, es en el tercer subcultivo
celular cuando el estímulo para el inicio del proceso de mortalidad celular alcanza su
pico máximo desencadenando así una cascada que lleva al aumento de las moléculas
efectoras y posteriormente al proceso de muerte celular.
En suma, en esta Tesis Doctoral se ha determinado la viabilidad celular utilizando dos
métodos diferentes: azul tripán y microarray, con los cuales podemos establecer
modelos de viabilidad divergentes en relación con los subcultivos celulares. Aunque
con el método del azul tripán la determinación de la viabilidad parece aceptable, la
determinación de la expresión génica, nos demuestra importantes cambios en los
subcultivos celulares en relación tanto con el potencial proliferativo, como con la
capacidad de diferenciación y el desarrollo de la muerte por apoptosis. De estudios
conjuntos como los desarrollados en la presente Tesis Doctoral y de las conclusiones
de la misma, debe inferirse la selección de la población celular condrocítica más apta
para su utilización como terapia celular en la patología del cartílago.

INTRODUCCIÓN
OBJETIVOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
TABLAS SUPLEMENTARIAS
REFERENCIAS BIBLIOGRÁFICAS

Conclusiones
96
1.- Los métodos y técnicas desarrollados y optimizados en la presente Tesis Doctoral,
permiten el aislamiento de los condrocitos del cartílago hialino articular y su
mantenimiento hasta el octavo subcultivo sin que dicho aislamiento y mantenimiento
hayan generado alteraciones artefactuales significativas.
2.- La evaluación de la viabilidad de los condrocitos articulares humanos utilizando la
técnica de exclusión del colorante azul tripán pone de relieve la existencia de un nivel
elevado de viabilidad en todos los subcultivos celulares analizados, si bien existe una
disminución moderada de la misma a partir del primer subcultivo, que se recupera a
partir del cuarto y que posiblemente puede relacionarse con la adaptación de los
condrocitos al cultivo.
3.- El análisis de la expresión génica mediante microarrays de los condrocitos
articulares humanos, pone de relieve que el número de genes/EST sobreexpresados,
varía en función del subcultivo celular, observándose que el número de genes que
muestra una correlación directa con el subcultivo celular duplica el número de genes
cuya correlación con el subcultivo celular es inversa. La correlación directa e inversa
está relacionada con funciones génicas vinculadas al metabolismo celular, la
proliferación, la adhesión celular y la apoptosis.
4.- El análisis de la expresión génica mediante microarrays de los condrocitos
articulares humanos, pone de relieve que la expresión de los genes relacionados con
la diferenciación condral, varía en función del subcultivo celular observándose que en
el subcultivo tres existe un pico de expresión significativo de los genes vinculados a la
síntesis de la matriz extracelular cartilaginosa, concretamente a la síntesis de las

Conclusiones
97
moléculas agrecán, condroitín sulfato, colágeno tipo II y proteína de matriz
cartilaginosa tipo I. El pico de expresión del gen que codifica el agrecán coincide en el
tercer subcultivo con niveles mínimos de expresión del gen que codifica la agrecanasa
2, que es la enzima responsable de la degradación de dicha molécula.
5.- El análisis de la expresión génica mediante microarrays de los condrocitos
articulares humanos, pone de relieve que la expresión de los genes relacionados con
la mortalidad celular, varía en función del subcultivo celular, observándose que el
máximo nivel de expresión de la caspasa 9, caspasa iniciadora del proceso de
apoptosis, tiene lugar en el tercer subcultivo, mientras que los niveles de expresión de
caspasa 3, caspasa efectora del proceso de apoptosis, son mínimos en dicho
subcultivo y aumentan progresivamente a partir del mismo.
6.- El estudio comparado de la viabilidad celular determinada conjuntamente mediante
azul tripán y microarray, nos permite establecer la disociación existente entre ambos
métodos y postular, sustentados en el análisis de expresión génica, la mayor
idoneidad del tercer subcultivo, en el que convergen una mayor capacidad de
diferenciación condral y una menor disposición para el inicio del proceso de apoptosis.
Todo ello nos permite postular la utilización de dicho subcultivo en los protocolos de
terapia celular con condrocitos.

INTRODUCCIÓN
OBJETIVOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
TABLAS SUPLEMENTARIAS
REFERENCIAS BIBLIOGRÁFICAS

Tablas Suplementarias
99
Tabla 4.- Se muestran los valores de expresión correspondientes a cada subcultivo para todos
los genes cuya expresión era superior a 5000 UF.
Probe Set ID
1er
Sub-
cultivo
2º Sub-
cultivo
3er
Sub-
cultivo
4º Sub-
cultivo
5º Sub-
cultivo
6º Sub-
cultivo
7º Sub-
cultivo
8º Sub-
cultivo
Símbologen
Nombre gen
221798_x_at 6760,4 7031,0 17491,6 25030,0 19971,4 21245,1 9399,9 8733,3 --- ---
226237_at 1762,4 5974,7 4364,4 4988,1 2585,6 3715,0 4378,2 4349,7 --- ---
1555623_at 2,1 5,5 21,2 5651,1 10,8 6,6 1,8 3,1 --- ---
200801_x_at 7181,9 7832,5 13428,6 18978,0 13137,7 16714,3 9265,1 9561,2 ACTB actin, beta
213867_x_at 6077,4 6144,5 12069,0 17981,8 12321,1 15732,3 7622,5 7619,4 ACTB actin, beta
224594_x_at 6212,4 6196,3 12479,6 18205,6 12733,4 15754,3 7711,0 7607,5 ACTB actin, beta
201550_x_at 5655,0 5524,1 8784,2 12546,3 10084,9 12602,2 7634,3 7532,6 ACTG1 actin, gamma 1
211970_x_at 5840,8 5673,2 8124,3 11717,9 9509,8 12183,7 7471,0 7450,7 ACTG1 actin, gamma 1
211983_x_at 6452,0 6327,7 10657,6 14516,7 11622,7 14408,9 8397,5 7925,3 ACTG1 actin, gamma 1
211995_x_at 5895,2 5023,3 11046,6 13939,6 11296,2 14457,8 7439,5 7147,1 ACTG1 actin, gamma 1
212988_x_at 5877,7 5722,4 9958,7 13119,9 10805,1 13109,4 7494,3 7268,7 ACTG1 actin, gamma 1
213214_x_at 6279,9 6301,7 8608,4 13032,5 10453,5 13651,7 8407,6 8136,2 ACTG1 actin, gamma 1
221607_x_at 6603,6 6007,7 12302,8 16020,7 12182,9 15404,5 8366,6 8145,0 ACTG1 actin, gamma 1
224585_x_at 6600,8 5897,8 12305,1 15922,6 12196,9 15636,9 8441,8 8000,9 ACTG1 actin, gamma 1
212363_x_at 5508,1 4900,6 9129,6 12077,1 9674,9 12214,0 6865,6 6769,1 ACTG1 actin, gamma 1
204151_x_at 5341,0 5596,1 7129,2 9055,4 7534,3 7257,7 5487,6 4377,5 AKR1C1
aldo-keto reductase family 1, member C1
(dihydrodiol dehydrogenase 1;
20-alpha (3-alpha)-hydroxysteroid
dehydrogenase)
216594_x_at 4704,5 4634,8 6064,0 7664,0 6234,2 5950,7 4622,4 3942,3 AKR1C1
aldo-keto reductase family 1, member C1
(dihydrodiol dehydrogenase 1;
20-alpha (3-alpha)-hydroxysteroid
dehydrogenase)
209699_x_at 5443,0 6029,2 5664,5 7615,7 6389,2 5372,8 4491,4 3828,9 AKR1C2
aldo-keto reductase family 1, member C2
(dihydrodiol dehydrogenase 2; bile acid binding protein; 3-alpha hydroxysteroid
dehydrogenase, type III)
211653_x_at 5205,5 5105,3 6366,7 8447,8 6724,3 5656,9 4536,6 3885,4 AKR1C2
aldo-keto reductase family 1, member C2
(dihydrodiol dehydrogenase 2; bile acid binding protein; 3-alpha hydroxysteroid
dehydrogenase, type III)
214687_x_at 4543,7 5045,5 4815,5 6410,0 5344,0 5415,7 5598,0 4817,7 ALDOA aldolase A, fructose-
bisphosphate
200966_x_at 4526,1 4432,5 3639,1 6085,3 4686,4 5046,6 5411,9 4576,7 ALDOA aldolase A, fructose-
bisphosphate
201590_x_at 5553,4 5378,8 11211,3 13900,3 9700,0 11447,9 6719,9 6408,9 ANXA2 annexin A2

Tablas Suplementarias
100
210427_x_at 5442,2 5380,9 11581,8 13649,3 9853,9 11656,4 6800,3 6409,2 ANXA2 annexin A2
213503_x_at 5596,8 5622,7 11283,3 13511,6 9532,9 11274,4 7027,4 6856,5 ANXA2 annexin A2
208816_x_at 3867,7 4925,1 6906,8 10159,6 4068,5 8242,9 5051,6 5860,1 ANXA2P
2 annexin A2
pseudogene 2
213738_s_at 4506,2 5207,6 4513,2 5993,4 5527,1 5617,1 4666,5 4928,6 ATP5A1
ATP synthase, H+ transporting,
mitochondrial F1 complex, alpha
subunit 1, cardiac muscle
201891_s_at 6321,7 6495,9 7506,7 10287,4 10564,8 10718,3 7340,1 6056,0 B2M beta-2-microglobulin
216231_s_at 5838,3 4865,3 6210,6 7930,3 9347,1 8474,4 6030,7 5443,1 B2M beta-2-microglobulin
226227_x_at 2614,7 2275,9 2531,2 4226,4 5604,9 5107,4 3777,3 3303,9 C20orf1
99
chromosome 20 open reading frame
199
224915_x_at 2611,0 2458,9 2100,6 3230,6 5230,0 4117,8 3972,7 3371,7 C20orf1
99
chromosome 20 open reading frame
199
219054_at 2593,2 3849,2 3076,5 4444,3 5834,4 4759,9 5804,1 4441,0 C5orf23 chromosome 5 open
reading frame 23
212077_at 3319,6 5133,2 3297,6 4456,9 4280,9 4275,9 6101,2 5155,8 CALD1 caldesmon 1
207243_s_at 5198,5 4707,0 4113,2 4945,1 4306,3 4172,3 4708,2 5088,3 CALM1 calmodulin 1
(phosphorylase kinase, delta)
212097_at 4443,6 6981,2 5635,9 7757,9 4735,0 4961,3 5645,5 7191,3 CAV1 caveolin 1, caveolae
protein, 22kDa
221791_s_at 4122,6 4561,0 3738,5 5051,2 4231,6 4355,3 3573,5 3919,7 CCDC72 coiled-coil domain
containing 72
200663_at 5588,2 5639,0 11476,7 12707,8 10146,5 10446,4 6258,7 5694,5 CD63 CD63 molecule
200675_at 3548,1 4760,8 4836,4 8171,3 7158,0 7849,8 5241,6 4308,0 CD81 CD81 molecule
201029_s_at 4401,1 4816,0 6164,6 6330,6 7068,2 5731,6 5318,3 3958,6 CD99 CD99 molecule
207173_x_at 3073,6 2610,6 1263,7 1669,8 3723,3 3627,1 5144,7 3711,6 CDH11 cadherin 11, type 2,
OB-cadherin (osteoblast)
215388_s_at 3655,3 1930,5 2665,7 3914,4 5487,9 2054,7 3121,7 1110,3 CFH complement factor H
200021_at 4799,0 5803,0 8174,1 9885,7 6624,5 8151,1 6046,7 5805,6 CFL1 cofilin 1 (non-
muscle)
209395_at 6685,6 7031,2 8929,5 10038,9 16820,5 12231,3 8753,9 4750,0 CHI3L1 chitinase 3-like 1
(cartilage glycoprotein-39)
209396_s_at 6078,7 6398,0 10211,6 11145,6 17817,4 12801,9 8496,4 3781,0 CHI3L1 chitinase 3-like 1
(cartilage glycoprotein-39)
225664_at 3317,1 4528,6 4005,3 5508,9 6792,8 7119,9 6821,0 5414,4 COL12A
1 collagen, type XII,
alpha 1
1556499_s_at 7760,4 6765,4 14046,5 19603,4 13865,3 16985,0 9008,3 7644,0 COL1A1 collagen, type I,
alpha 1
202310_s_at 6451,1 6499,5 10484,6 14128,3 10948,3 15374,5 9624,9 8483,5 COL1A1 collagen, type I,
alpha 1
202403_s_at 7412,4 7452,9 15090,8 20575,3 18153,4 20256,0 10504,7 9701,2 COL1A2 collagen, type I,
alpha 2
202404_s_at 7091,1 7161,1 13040,5 16010,6 11899,2 14254,8 9586,1 7962,9 COL1A2 collagen, type I,
alpha 2
201852_x_at 7026,9 6296,9 12155,8 14010,9 14452,7 14309,8 9349,9 7605,9 COL3A1 collagen, type III,
alpha 1
211161_s_at 7202,7 6757,2 9073,7 10523,4 10753,2 10173,4 8886,0 6912,0 COL3A1 collagen, type III,
alpha 1
215076_s_at 9118,4 9263,0 16504,0 19491,7 19997,3 19698,1 12074,3 10507,3 COL3A1 collagen, type III,
alpha 1
221729_at 3218,8 2807,4 2513,5 2768,3 3193,3 2685,3 5510,0 4187,5 COL5A2 collagen, type V,
alpha 2

Tablas Suplementarias
101
213428_s_at 7083,5 7517,7 13391,6 16020,2 14063,8 15281,7 8950,2 7629,7 COL6A1 collagen, type VI,
alpha 1
212091_s_at 3006,5 1453,7 4103,8 4217,2 6371,3 5158,6 2017,8 1788,4 COL6A1 collagen, type VI,
alpha 1
209156_s_at 5190,9 5410,3 7042,5 8362,4 9856,4 10299,1 6487,7 5416,6 COL6A2 collagen, type VI,
alpha 2
201438_at 6292,1 6379,9 9990,0 13032,7 12551,1 13888,4 7931,6 7203,7 COL6A3 collagen, type VI,
alpha 3
1553538_s_at 7808,4 8238,0 11163,4 14451,1 13531,9 13987,5 10916,0 9603,9 COX1 cytochrome c
oxidase I
1553569_at 7633,9 8481,1 16058,5 18101,3 16172,2 13674,6 10434,4 9669,4 COX2 cytochrome c
oxidase II
1553570_x_at 7669,1 8410,7 18403,4 20393,9 18262,8 18097,7 10259,2 9477,4 COX2 cytochrome c
oxidase II
202698_x_at 3678,0 4265,2 4814,0 4870,1 5242,6 4457,4 3889,0 3486,8 COX4I1 cytochrome c
oxidase subunit IV isoform 1
206336_at 1239,6 3586,3 3744,5 6435,2 3586,9 3562,9 3329,0 3550,0 CXCL6
chemokine (C-X-C motif) ligand 6 (granulocyte
chemotactic protein 2)
201885_s_at 3200,9 3988,9 4422,2 5802,7 4955,8 4174,2 4826,4 4076,0 CYB5R3 cytochrome b5
reductase 3
202436_s_at 6314,1 7463,9 6819,5 10069,8 10671,5 8529,1 7367,5 6367,9 CYP1B1 cytochrome P450, family 1, subfamily
B, polypeptide 1
202437_s_at 5437,2 6951,1 6122,4 7461,9 7205,6 6045,9 5237,6 4420,1 CYP1B1 cytochrome P450, family 1, subfamily
B, polypeptide 1
202435_s_at 4222,8 5668,2 3765,2 5281,0 5951,4 4872,8 4571,5 3916,6 CYP1B1 cytochrome P450, family 1, subfamily
B, polypeptide 1
201893_x_at 6274,4 4924,5 6774,8 9245,7 10287,7 9885,7 7713,2 6694,2 DCN Decorin
211813_x_at 6144,1 2857,7 5489,0 6683,0 8392,5 8072,8 6335,9 5457,2 DCN Decorin
211896_s_at 6839,5 4060,2 5789,7 7033,0 8454,6 8356,6 7012,9 5924,8 DCN Decorin
201022_s_at 5185,6 7103,7 8345,1 10440,4 7157,3 8863,8 7711,7 7456,4 DSTN destrin (actin
depolymerizing factor)
201021_s_at 3981,0 4384,6 6127,2 7148,7 4733,0 6603,2 5442,6 5519,5 DSTN destrin (actin
depolymerizing factor)
201041_s_at 5066,8 3035,0 2200,8 2771,9 2719,3 1894,4 2868,9 2487,7 DUSP1 dual specificity phosphatase 1
204892_x_at 6806,4 6654,5 19802,3 26448,2 18924,9 21182,2 8789,8 8116,6 EEF1A1
eukaryotic translation
elongation factor 1 alpha 1
206559_x_at 6388,8 6720,5 20114,0 24945,3 20474,7 19537,5 9085,1 8436,5 EEF1A1
eukaryotic translation
elongation factor 1 alpha 1
213477_x_at 7367,5 7494,5 17128,0 24862,0 17593,6 21679,4 9249,5 7929,5 EEF1A1
eukaryotic translation
elongation factor 1 alpha 1
213614_x_at 7962,0 7464,1 17772,7 24952,7 20013,3 22150,5 9891,8 9322,6 EEF1A1
eukaryotic translation
elongation factor 1 alpha 1

Tablas Suplementarias
102
213583_x_at 6943,7 6642,1 16257,0 22839,7 16566,5 20185,3 8543,2 7501,5 EEF1A1
eukaryotic translation
elongation factor 1 alpha 1
200705_s_at 5661,1 4421,9 4087,9 6381,1 6657,6 6286,5 5035,7 5722,8 EEF1B2
eukaryotic translation
elongation factor 1 beta 2
214394_x_at 3772,0 4964,9 3242,5 4599,1 5394,9 5064,4 5186,1 4837,8 EEF1D
eukaryotic translation
elongation factor 1 delta (guanine
nucleotide exchange protein)
211927_x_at 5552,9 5424,0 7642,3 11184,2 10341,0 12204,3 6647,9 6254,3 EEF1G
eukaryotic translation
elongation factor 1 gamma
211345_x_at 5065,2 4658,2 4774,6 8177,1 8200,6 9485,7 5733,0 5473,8 EEF1G
eukaryotic translation
elongation factor 1 gamma
200689_x_at 4982,4 4710,1 4514,4 8263,2 8244,2 9120,1 5812,1 5368,8 EEF1G
eukaryotic translation
elongation factor 1 gamma
204102_s_at 4858,9 5539,3 7118,0 12269,3 8823,7 11829,6 6598,6 5966,1 EEF2 eukaryotic translation
elongation factor 2
200094_s_at 3785,2 4028,4 5196,9 5902,5 7361,6 9025,7 5387,5 5528,3 EEF2 eukaryotic translation
elongation factor 2
201842_s_at 7776,3 8666,5 10905,5 15308,7 15983,6 15923,1 11064,9 8984,7 EFEMP1
EGF-containing fibulin-like
extracellular matrix protein 1
211956_s_at 5898,9 5983,5 6201,0 9134,7 8676,5 9436,3 7191,9 6355,9 EIF1 eukaryotic
translation initiation factor 1
201231_s_at 7072,5 6178,1 8747,4 10796,1 8907,9 8905,5 8254,1 7343,3 ENO1 enolase 1, (alpha)
209392_at 4174,9 4615,3 3290,9 4546,5 5584,8 4499,9 4420,9 4295,1 ENPP2 ectonucleotide
pyrophosphatase/phosphodiesterase 2
396_f_at 207,9 126,7 10208,3 5031,8 3780,0 3578,1 158,8 144,4 EPOR erythropoietin
receptor
203980_at 5411,0 3398,7 6715,7 3056,7 4782,0 873,9 525,3 144,5 FABP4 fatty acid binding
protein 4, adipocyte
202766_s_at 3773,6 5119,9 4274,5 5832,1 5546,4 4576,8 5512,1 4176,8 FBN1 fibrillin 1
210495_x_at 5709,3 5985,1 13199,1 19708,3 14268,3 16507,7 6888,5 6634,9 FN1 fibronectin 1
211719_x_at 7135,4 7291,0 12701,6 20240,3 14992,7 17608,1 8326,8 7784,8 FN1 fibronectin 1
212464_s_at 7433,7 7029,4 14158,3 20426,6 14893,5 19502,6 7846,9 7617,1 FN1 fibronectin 1
216442_x_at 5930,1 5720,5 12016,9 20021,4 14237,6 15869,8 7023,8 6549,1 FN1 fibronectin 1
205021_s_at 167,8 33,1 1364,6 5703,4 1219,4 2397,5 66,4 44,0 FOXN3 forkhead box N3
208782_at 6346,7 6375,5 8714,2 10098,6 10374,6 9356,3 7807,4 6132,2 FSTL1 follistatin-like 1
200748_s_at 7590,7 8839,2 17257,3 23732,9 20123,1 21373,6 10347,1 9409,5 FTH1 ferritin, heavy polypeptide 1
211628_x_at 6751,8 6710,8 14901,3 21145,3 16668,9 17754,0 8919,2 7896,6 FTHP1 ferritin, heavy polypeptide
pseudogene 1
212788_x_at 7152,9 7052,8 17561,5 24053,8 20327,2 22047,1 9470,5 8477,1 FTL ferritin, light polypeptide

Tablas Suplementarias
103
213187_x_at 6083,8 6532,1 11051,5 14392,7 15482,7 15765,9 7765,6 6842,2 FTL ferritin, light polypeptide
212581_x_at 5258,7 5282,4 12919,1 17779,1 14446,7 13913,5 7293,9 6914,4 GAPDH glyceraldehyde-3-
phosphate dehydrogenase
213453_x_at 6294,6 6119,8 12517,7 17089,4 13385,9 14141,6 7953,4 7324,7 GAPDH glyceraldehyde-3-
phosphate dehydrogenase
217398_x_at 5358,8 5049,0 12578,4 17036,8 13858,3 13688,6 7217,0 6501,2 GAPDH glyceraldehyde-3-
phosphate dehydrogenase
204457_s_at 4391,0 3685,9 2289,2 3197,7 6077,9 3747,9 7330,4 4485,2 GAS1 growth arrest-
specific 1
1598_g_at 2017,3 4444,2 6628,8 6878,0 5997,7 6078,1 4327,0 3222,8 GAS6 growth arrest-
specific 6
231951_at 48,7 49,5 3558,8 5572,9 1092,0 629,7 30,7 18,6 GNAO1
guanine nucleotide binding protein (G
protein), alpha activating activity
polypeptide O
214548_x_at 4505,8 4032,7 5362,4 6698,1 6081,8 7324,6 5267,6 4681,8 GNAS GNAS complex locus
200780_x_at 4474,7 3932,6 4073,8 5332,0 6016,9 5640,6 5359,5 4839,5 GNAS GNAS complex locus
200981_x_at 4189,4 3393,2 3662,3 5355,4 5205,8 5513,4 5183,6 4249,0 GNAS GNAS complex locus
212273_x_at 4254,7 3836,9 3996,7 5315,9 5646,3 5692,6 5079,0 4232,6 GNAS GNAS complex locus
200651_at 5055,7 5056,7 5436,9 9085,0 8723,5 10098,4 6111,2 6158,0 GNB2L1
guanine nucleotide binding protein (G
protein), beta polypeptide 2-like 1
201348_at 4763,0 1724,6 3592,4 5687,4 6454,0 5943,8 3114,0 971,4 GPX3 glutathione
peroxidase 3 (plasma)
208825_x_at 7770,3 7523,6 9408,2 16768,2 15735,0 16246,5 9350,4 9235,6 hCG_16
001 similar to ribosomal
protein L23A
208834_x_at 7917,4 7635,2 9561,4 16625,4 15666,3 16585,8 9259,3 9211,7 hCG_16
001 similar to ribosomal
protein L23A
213084_x_at 7899,7 7741,1 10627,1 16503,3 15740,0 15793,5 9208,8 9223,0 hCG_16
001 similar to ribosomal
protein L23A
203034_s_at 7354,3 7536,4 11082,8 15802,9 16218,7 16496,2 9358,1 8662,2 hCG_21
078 hCG21078
200989_at 5375,6 5139,3 4154,4 5689,2 6659,0 6144,6 7067,2 6030,5 HIF1A
hypoxia-inducible factor 1, alpha
subunit (basic helix-loop-helix
transcription factor)
213932_x_at 3980,5 4510,5 4672,2 5153,9 6891,2 6212,1 5408,4 3869,3 HLA-A major
histocompatibility complex, class I, A
215313_x_at 3750,7 3864,5 4638,2 5381,4 7291,9 6816,1 4591,4 3366,0 HLA-A major
histocompatibility complex, class I, A
216526_x_at 2923,3 2748,5 3595,4 4423,4 5218,6 5453,5 4221,7 3277,0 HLA-B major
histocompatibility complex, class I, B
200016_x_at 5929,1 5658,1 3843,3 5607,6 5849,3 6388,6 6294,3 6895,9 HNRNP
A1
heterogeneous nuclear
ribonucleoprotein A1
213356_x_at 5051,6 4804,2 3337,5 5370,2 5220,1 6169,1 5330,8 6013,6 HNRNP
A1
heterogeneous nuclear
ribonucleoprotein A1

Tablas Suplementarias
104
1555653_at 3344,6 3081,2 5282,7 2719,6 6461,2 4967,5 3928,6 4229,6 HNRNP
A3
heterogeneous nuclear
ribonucleoprotein A3
224373_s_at 7975,4 8469,4 16459,5 23426,2 19111,2 18731,9 10476,7 9127,3 HNRNP
M
heterogeneous nuclear
ribonucleoprotein M
214328_s_at 5243,6 5825,2 4075,0 5574,6 4731,9 4468,9 5152,0 5424,7 HSP90A
A1
heat shock protein 90kDa alpha
(cytosolic), class A member 1
200599_s_at 5279,6 5184,1 2916,5 3474,9 4464,8 3233,1 4630,5 4525,5 HSP90B
1
heat shock protein 90kDa beta (Grp94),
member 1
211936_at 5371,2 4996,0 4817,9 6418,5 5109,5 4770,9 5043,2 4584,2 HSPA5
heat shock 70kDa protein 5 (glucose-regulated protein,
78kDa)
224187_x_at 5050,5 5682,6 5899,8 7150,2 5758,9 5260,7 3920,4 4252,9 HSPA8 heat shock 70kDa
protein 8
221891_x_at 5348,8 5710,8 5310,0 7136,3 5758,4 4779,5 4421,0 4396,1 HSPA8 heat shock 70kDa
protein 8
208687_x_at 4342,9 4095,1 4410,7 5619,1 4748,1 4307,9 3188,1 3540,8 HSPA8 heat shock 70kDa
protein 8
201185_at 4099,0 4467,0 7100,8 7235,5 8168,1 8687,5 5912,5 4422,2 HTRA1 HtrA serine peptidase 1
207783_x_at 6629,5 6209,6 16425,8 20307,2 19361,7 18066,3 9410,7 8371,5 HUWE1 HECT, UBA and WWE domain containing 1
224625_x_at 4425,4 4191,2 3426,6 5773,4 5057,7 5450,7 4015,2 3713,4 HYPK Huntingtin
interacting protein K
201315_x_at 5546,8 6032,0 10207,6 12696,9 13189,9 13313,7 7318,3 5727,9 IFITM2 interferon induced
transmembrane protein 2 (1-8D)
212203_x_at 4669,1 4878,8 7297,6 9875,0 10943,3 11268,8 6788,9 5203,7 IFITM3 interferon induced
transmembrane protein 3 (1-8U)
202409_at 2958,6 2502,8 4468,3 4675,9 6792,8 8488,3 6292,1 3460,0 IGF2 insulin-like growth
factor 2 (somatomedin A)
202718_at 3847,8 5692,3 9006,3 7322,1 9100,8 8987,1 5251,9 1957,8 IGFBP2 insulin-like growth
factor binding protein 2, 36kDa
210095_s_at 5882,8 8919,5 20221,0 24015,8 12571,0 12131,8 7567,5 6802,9 IGFBP3 insulin-like growth
factor binding protein 3
212143_s_at 2631,9 3449,6 10281,6 10304,9 5804,5 5670,3 2449,6 2390,9 IGFBP3 insulin-like growth
factor binding protein 3
201508_at 5479,1 6011,7 16507,7 19063,2 15745,2 16910,4 7714,5 6711,7 IGFBP4 insulin-like growth
factor binding protein 4
201162_at 2939,7 3667,0 4329,8 5626,7 4432,9 5284,0 3223,9 2553,0 IGFBP7 insulin-like growth
factor binding protein 7
224372_at 6713,2 7845,4 15926,1 23350,9 17561,7 18740,2 9509,3 8883,0 IQWD1 IQ motif and WD
repeats 1
211945_s_at 5016,5 5255,6 7407,7 8884,3 8169,1 7767,8 6483,3 5773,4 ITGB1
integrin, beta 1 (fibronectin
receptor, beta polypeptide, antigen CD29 includes MDF2,
MSK12)
217731_s_at 5618,1 5771,6 3091,9 3199,9 5114,3 2716,9 5958,8 4551,7 ITM2B integral membrane
protein 2B

Tablas Suplementarias
105
217732_s_at 4765,2 4423,5 5104,7 5687,0 7008,3 5310,8 5190,4 3771,3 ITM2B integral membrane
protein 2B
201553_s_at 5308,1 5909,0 4751,4 5824,5 6013,4 6132,7 6428,5 4837,4 LAMP1 lysosomal-associated membrane protein 1
200673_at 4523,4 4872,9 5026,3 6017,3 6185,7 5787,3 5301,5 4667,9 LAPTM4
A
lysosomal protein transmembrane 4
alpha
200650_s_at 6317,6 6018,7 7658,0 9494,9 7947,3 5798,5 7606,1 6826,6 LDHA lactate
dehydrogenase A
213564_x_at 4879,5 5472,6 4597,9 6141,2 5938,4 6576,3 5032,1 5334,8 LDHB lactate
dehydrogenase B
201105_at 5269,2 5444,7 7553,8 10010,3 6827,3 8065,9 6228,9 6578,5 LGALS1 lectin, galactoside-binding, soluble, 1
216342_x_at 4009,0 4727,6 4749,1 7156,3 6504,8 7425,3 5535,8 4782,4 LOC100128140
similar to ribosomal protein S4, X-linked
229563_s_at 7151,3 6963,5 10845,1 16821,2 14676,5 15910,6 8627,2 7842,8 LOC100128936
similar to ribosomal protein L10a
201049_s_at 6067,6 5966,8 9115,7 15952,2 14838,6 15736,4 8147,1 7265,1 LOC100130553
hypothetical protein
200823_x_at 4641,6 4322,8 2195,7 4004,9 5031,0 5146,3 4107,1 4312,6 LOC100131713
similar to ribosomal protein
1569522_at 47,7 52,5 6784,5 7039,6 3869,9 3318,9 117,4 72,5 LOC100132767
hypothetical protein LOC100132767
1553567_s_at 6042,9 5759,0 14953,1 19057,3 16057,0 15482,8 9022,1 7839,4 LOC100133315
similar to hCG1640299
201463_s_at 2942,2 5048,9 2644,2 2947,1 2531,3 2441,2 2596,8 3074,7 LOC100133665
similar to transaldolase
1558641_at 218,2 112,6 29342,6 30087,2 8335,4 11080,6 224,3 202,6 LOC202
051 hypothetical protein
LOC202051
234873_x_at 6040,3 5612,6 5337,3 6430,1 8043,4 6975,9 6980,7 7053,4 LOC388
474 similar to ribosomal
protein L7a
213801_x_at 6812,6 6662,1 9046,9 13608,3 12314,3 14527,9 8295,0 7674,2 LOC388
524 ribosomal protein SA
pseudogene
200869_at 7074,7 6167,4 3673,3 6893,6 8666,3 8189,8 7282,3 6829,1 LOC390
354 ribosomal protein L18a pseudogene
200099_s_at 7919,6 6830,1 9412,5 14883,2 14157,4 16050,6 8414,9 7951,8 LOC439
992
similar to v-fos transformation effector protein
1558688_at 916,7 369,7 9384,0 3227,6 4169,7 4208,6 1472,7 1099,1 LOC441
461
hypothetical gene supported by
BC030123
222229_x_at 3415,7 3299,0 5077,4 5028,7 5486,2 4993,7 3467,5 3931,9 LOC441
533
60S ribosomal protein L26 pseudogene
215963_x_at 5924,7 5215,9 6607,5 10200,9 11370,4 9862,0 7187,3 6989,1 LOC642
741 similar to ribosomal
protein L3
217753_s_at 5125,0 4425,7 3388,4 4449,0 5278,7 4764,0 5024,7 5079,2 LOC644
191 similar to hCG15685
200082_s_at 4764,7 4449,2 2424,0 4501,8 5061,3 4108,6 4910,5 5424,2 LOC644
315 similar to ribosomal
protein S7
200012_x_at 4881,8 4749,2 4980,3 7648,3 8569,4 8534,8 5891,7 5397,1 LOC653
737 hypothetical LOC653737
234512_x_at 4704,8 4249,6 2355,0 5459,6 5794,5 6135,2 4171,4 4051,6 LOC728
179
similar to LOC100037086
protein
201744_s_at 5897,9 3335,4 3129,7 3787,4 4808,0 3829,5 4477,3 3445,3 LUM Lumican
1558678_s_at 861,2 1119,0 141,4 276,2 3164,4 498,3 5016,1 3877,0 MALAT1
metastasis associated lung
adenocarcinoma transcript 1 (non-protein coding)
208646_at 4728,6 4248,5 2780,2 4955,7 5224,1 4831,8 4667,8 4267,6 MGC878
95 similar to ribosomal
protein S14

Tablas Suplementarias
106
205828_at 6939,0 686,9 1458,3 1254,8 614,6 374,9 909,6 1504,6 MMP3
matrix metallopeptidase 3
(stromelysin 1, progelatinase)
217982_s_at 4723,0 4232,1 2914,3 4016,8 4274,2 3849,7 5079,5 4458,5 MORF4L
1 mortality factor 4
like 1
201994_at 5059,2 4282,4 2073,4 3010,1 2800,4 2445,8 3712,4 3869,3 MORF4L
2 mortality factor 4
like 2
201318_s_at 3145,8 3585,2 3484,3 5114,1 3532,1 3608,9 3419,0 4231,5 MRCL3 myosin regulatory light chain MRCL3
226091_s_at 4495,9 5050,9 3828,5 4385,2 4819,8 4181,7 4968,6 5054,9 MRFAP1 Mof4 family
associated protein 1
212185_x_at 6074,4 5465,6 5575,9 9540,9 4635,1 6484,9 3518,4 5603,1 MT2A metallothionein 2A
212509_s_at 5598,5 5559,2 6030,7 6927,3 7278,2 5862,6 6833,7 5527,5 MXRA7 matrix-remodelling
associated 7
212082_s_at 5286,2 5707,2 5707,8 7892,2 5449,1 6276,0 6004,2 5685,8 MYL6
myosin, light chain 6, alkali, smooth
muscle and non-muscle
200735_x_at 5086,5 4987,9 3666,6 5691,1 7045,8 6770,2 5755,4 5779,5 NACA nascent polypeptide-associated complex
alpha subunit
208635_x_at 5082,3 4986,9 3742,0 5619,7 7011,4 6805,5 5718,0 5809,9 NACA nascent polypeptide-associated complex
alpha subunit
1553551_s_at 5362,9 5724,9 8343,6 8404,0 9868,4 10003,3 6926,9 6576,0 ND2 NADH
dehydrogenase, subunit 2 (complex I)
1553588_at 7669,1 7773,8 12116,6 14312,1 14945,7 13266,1 9052,0 7697,3 ND3 NADH
dehydrogenase, subunit 3 (complex I)
217963_s_at 5034,8 5760,5 6119,5 7303,5 6509,8 7528,4 5768,9 5919,3 NGFRAP
1
nerve growth factor receptor (TNFRSF16) associated protein 1
202237_at 6229,0 6560,9 5423,8 7254,0 9748,8 8079,5 7952,7 5879,9 NNMT nicotinamide N-
methyltransferase
202238_s_at 4530,8 3813,8 3651,9 4091,5 6053,3 4847,0 4869,0 3196,6 NNMT nicotinamide N-
methyltransferase
200063_s_at 5361,8 5355,4 5804,1 8586,8 7327,3 8182,6 6321,4 6047,7 NPM1
nucleophosmin (nucleolar
phosphoprotein B23, numatrin)
201468_s_at 1869,7 6274,5 3201,2 5017,0 3070,6 4417,0 3287,9 4669,0 NQO1 NAD(P)H
dehydrogenase, quinone 1
210519_s_at 1788,2 5202,8 3591,3 4997,0 3369,7 4730,6 2802,2 4359,8 NQO1 NAD(P)H
dehydrogenase, quinone 1
200077_s_at 5166,1 4904,7 6066,3 8166,0 6733,7 7642,0 5515,8 5552,3 OAZ1 ornithine
decarboxylase antizyme 1
215952_s_at 5388,5 4295,4 4269,2 4840,9 4542,2 5091,5 4652,4 4919,0 OAZ1 ornithine
decarboxylase antizyme 1
200654_at 5516,5 5275,4 7210,9 8826,2 7643,4 8777,9 6113,6 5354,9 P4HB
procollagen-proline, 2-oxoglutarate 4-
dioxygenase (proline 4-hydroxylase), beta
polypeptide
215157_x_at 4075,0 4037,6 4141,3 4961,9 5056,1 6171,1 5099,4 5184,1 PABPC1 poly(A) binding
protein, cytoplasmic 1
208113_x_at 3060,9 3479,1 2422,6 3843,1 4053,0 5187,1 4449,7 4528,3 PABPC3 poly(A) binding
protein, cytoplasmic 3

Tablas Suplementarias
107
224940_s_at 1729,4 6069,3 1633,6 2258,6 2389,1 1709,4 5143,1 2707,9 PAPPA
pregnancy-associated plasma
protein A, pappalysin 1
200006_at 4728,0 5336,4 2654,2 3731,4 3573,2 3076,4 4624,2 5215,7 PARK7
Parkinson disease (autosomal
recessive, early onset) 7
202465_at 3528,0 3212,3 2350,0 2768,8 5419,6 5860,4 4289,8 3297,6 PCOLCE procollagen C-endopeptidase
enhancer
203131_at 5170,9 4723,5 3078,7 4188,8 7745,2 5281,0 6680,0 5263,0 PDGFRA
platelet-derived growth factor
receptor, alpha polypeptide
200886_s_at 4926,0 4325,2 4728,6 5640,0 5031,6 3717,4 4698,9 4211,6 PGAM1 phosphoglycerate mutase 1 (brain)
200738_s_at 5678,2 3910,1 4853,4 6506,0 6657,9 3983,2 5567,7 4444,4 PGK1 phosphoglycerate
kinase 1
201251_at 3906,8 3656,4 4560,6 5436,7 3933,9 4320,4 3511,7 3789,1 PKM2 pyruvate kinase,
muscle
211730_s_at 3197,8 5805,5 4214,3 4829,7 3863,6 4084,6 3709,9 4458,3 POLR2L
polymerase (RNA) II (DNA directed) polypeptide L,
7.6kDa
209147_s_at 3640,9 3597,2 3591,7 7334,7 8177,2 7442,7 6189,0 4619,8 PPAP2A phosphatidic acid
phosphatase type 2A
210946_at 3984,2 4296,5 4063,2 6128,4 7685,1 7619,6 7078,2 4724,4 PPAP2A phosphatidic acid
phosphatase type 2A
212226_s_at 4065,4 5454,1 3557,7 6085,6 6741,0 5955,6 7163,9 5654,1 PPAP2B phosphatidic acid
phosphatase type 2B
212230_at 2405,1 5386,5 4139,0 5604,6 5636,7 5776,5 5848,4 4612,6 PPAP2B phosphatidic acid
phosphatase type 2B
209355_s_at 3188,3 4266,5 1971,4 4034,5 5044,9 4292,9 6170,6 4734,9 PPAP2B phosphatidic acid
phosphatase type 2B
201293_x_at 6700,2 6243,6 9078,0 11209,8 8837,7 9367,2 6128,5 6537,5 PPIA peptidylprolyl isomerase A
(cyclophilin A)
211378_x_at 6452,3 5976,3 8595,6 10589,0 8286,7 8881,2 6120,5 6545,8 PPIA peptidylprolyl isomerase A
(cyclophilin A)
211765_x_at 6701,9 6377,7 9092,5 11349,2 8729,8 9230,9 6358,2 6693,6 PPIA peptidylprolyl isomerase A
(cyclophilin A)
211978_x_at 6744,2 6324,1 8625,9 10943,7 8793,3 9136,9 6469,7 7124,0 PPIA peptidylprolyl isomerase A
(cyclophilin A)
212661_x_at 6941,7 6542,0 9152,5 11314,8 9290,3 9617,2 7026,1 7138,0 PPIA peptidylprolyl isomerase A
(cyclophilin A)
200967_at 4827,9 5328,5 3193,3 4792,1 4785,3 4446,5 5207,6 4671,7 PPIB peptidylprolyl isomerase B
(cyclophilin B)
200773_x_at 3159,5 2293,5 2626,6 4503,0 4966,7 5198,7 3482,6 3049,8 PTMA prothymosin, alpha
206157_at 2179,8 3738,0 5326,2 9836,2 7274,8 7657,0 6137,1 5927,9 PTX3 pentraxin-related
gene, rapidly induced by IL-1 beta
219140_s_at 4452,6 5466,5 6555,5 6548,5 7229,1 5996,2 4671,9 3794,0 RBP4 retinol binding
protein 4, plasma
200059_s_at 4604,8 3918,0 4515,1 5430,2 4231,3 5049,4 4444,0 4874,5 RHOA ras homolog gene family, member A
200725_x_at 4500,8 4246,2 4739,7 6567,9 7796,1 8229,7 5360,7 5041,0 RPL10 ribosomal protein
L10
200036_s_at 4756,9 4376,3 2960,2 4502,9 5870,2 5327,1 4943,3 4891,4 RPL10A ribosomal protein
L10a

Tablas Suplementarias
108
200010_at 5482,9 5249,0 4686,5 5750,2 7305,9 8079,0 6048,1 6005,0 RPL11 ribosomal protein
L11
200088_x_at 6794,0 6806,2 5696,3 9100,1 10175,2 10639,7 7708,5 7377,2 RPL12 ribosomal protein
L12
200809_x_at 5897,2 5678,0 5127,6 8334,4 9299,0 10570,1 6332,3 6408,6 RPL12 ribosomal protein
L12
214271_x_at 5436,8 5554,6 3959,7 8478,0 8489,3 10141,8 5972,7 6132,5 RPL12 ribosomal protein
L12
212734_x_at 5791,3 5541,3 5539,1 8075,5 9287,9 9592,0 6295,6 6211,3 RPL13 ribosomal protein
L13
208929_x_at 5256,7 5135,5 4925,5 7107,5 8694,0 7868,5 5836,4 5720,8 RPL13 ribosomal protein
L13
214351_x_at 6477,4 6001,4 3595,8 6734,7 8066,5 6057,4 6429,5 6442,2 RPL13 ribosomal protein
L13
212191_x_at 5112,2 4763,9 3906,2 5397,5 6992,1 6326,0 5425,3 5106,0 RPL13 ribosomal protein
L13
212933_x_at 4665,5 5141,6 5020,0 7548,8 7110,5 8070,1 5455,8 5379,6 RPL13 ribosomal protein
L13
200715_x_at 5222,1 5407,8 5189,7 8068,8 8169,7 10386,7 6008,1 5828,2 RPL13A ribosomal protein
L13a
200716_x_at 7042,3 6502,6 9505,8 15094,3 14591,0 16201,8 8361,1 7949,5 RPL13A ribosomal protein
L13a
210646_x_at 7715,6 7706,8 10959,9 16281,6 15448,7 16648,9 9365,9 8866,9 RPL13A ribosomal protein
L13a
211942_x_at 5740,5 5319,4 7949,8 10135,6 11500,5 12652,9 6116,6 5819,6 RPL13A ribosomal protein
L13a
212790_x_at 7385,0 7192,0 9546,7 14567,7 14249,5 15854,8 8814,1 8421,5 RPL13A ribosomal protein
L13a
213588_x_at 3503,2 3925,0 3344,9 5697,5 5432,8 5572,8 4443,8 4228,8 RPL14 ribosomal protein
L14
221475_s_at 6104,8 5647,1 3344,0 5821,8 6522,9 7226,5 5738,8 6047,3 RPL15 ribosomal protein
L15
221476_s_at 3292,7 3745,6 2657,2 4303,0 5149,3 5250,0 4178,8 4176,4 RPL15 ribosomal protein
L15
200038_s_at 7314,5 7101,1 6950,9 10920,7 11760,0 12249,3 7941,0 7882,2 RPL17 ribosomal protein
L17
212270_x_at 5704,2 5608,8 5012,4 9202,4 9259,4 9854,9 6133,6 6317,9 RPL17 ribosomal protein
L17
212537_x_at 5863,3 5640,7 5955,1 9622,4 10026,0 10471,7 6410,2 6554,8 RPL17 ribosomal protein
L17
200022_at 5801,0 5280,5 5447,1 8746,7 9257,2 10489,4 6102,9 5784,1 RPL18 ribosomal protein
L18
200029_at 5457,5 5949,5 6981,0 10355,2 8760,0 8493,7 6644,8 6552,0 RPL19 ribosomal protein
L19
208768_x_at 4467,9 4196,4 4280,3 5633,7 6186,0 6301,4 4722,6 4911,6 RPL22 ribosomal protein
L22
220960_x_at 4453,6 4247,8 3997,7 5365,4 6056,9 6250,1 4681,1 4861,6 RPL22 ribosomal protein
L22
221775_x_at 4347,1 4176,7 3616,4 5172,2 6003,9 6182,5 4918,9 4884,7 RPL22 ribosomal protein
L22
200888_s_at 4731,2 4029,0 6082,2 8632,3 8475,2 9262,4 4520,1 5200,7 RPL23 ribosomal protein
L23
203012_x_at 6025,8 5773,2 6932,6 12342,1 12315,9 12671,5 7174,3 6686,1 RPL23A ribosomal protein
L23a
200013_at 5776,8 5479,3 4227,8 5163,2 6262,3 4957,3 5218,8 6055,8 RPL24 ribosomal protein
L24
214143_x_at 5990,6 5993,6 4802,5 6505,8 8088,2 7212,8 6320,4 6389,7 RPL24 ribosomal protein
L24
200025_s_at 6562,8 6959,0 7623,0 9013,8 10657,6 10242,3 7542,5 6822,1 RPL27 ribosomal protein
L27
200003_s_at 8083,0 7521,3 8621,2 14179,0 13512,5 15910,2 8737,9 8362,9 RPL28 ribosomal protein
L28
213969_x_at 5296,0 5176,3 3301,6 5810,7 6933,3 7077,7 5488,8 5423,6 RPL29 ribosomal protein
L29
201217_x_at 6908,4 6562,0 7655,5 13307,7 13436,4 14871,5 8960,8 8566,4 RPL3 ribosomal protein L3
211073_x_at 6362,4 6026,1 6706,9 12098,9 12933,3 13328,8 7822,6 7942,5 RPL3 ribosomal protein L3

Tablas Suplementarias
109
212039_x_at 6269,5 6043,2 6547,6 12093,2 12946,6 14466,0 7800,1 7752,2 RPL3 ribosomal protein L3
211666_x_at 5032,8 3471,8 4781,5 6767,2 7653,4 9391,2 5112,8 5367,8 RPL3 ribosomal protein L3
200062_s_at 5817,9 6682,8 6664,6 8702,1 10031,5 10547,0 7334,1 7144,3 RPL30 ribosomal protein
L30
200963_x_at 7187,9 7457,0 8376,3 12499,3 13243,7 13539,0 8130,5 7966,4 RPL31 ribosomal protein
L31
200674_s_at 4938,1 5112,2 5333,8 8180,9 8761,2 8736,0 5793,5 5557,8 RPL32 ribosomal protein
L32
200026_at 5896,7 5490,5 7038,5 10392,4 10933,8 10825,3 6723,5 6200,0 RPL34 ribosomal protein
L34
200002_at 5152,8 5136,7 4023,0 6044,2 6907,0 6408,8 5499,1 5299,4 RPL35 ribosomal protein
L35
219762_s_at 3187,8 5070,5 3536,1 3536,1 4528,1 4670,2 4231,2 3967,1 RPL36 ribosomal protein
L36
201406_at 6551,1 6092,2 6172,2 9080,0 10205,4 10470,4 7107,0 6720,3 RPL36A ribosomal protein
L36a
200092_s_at 6109,0 6309,0 7560,4 10393,7 11673,0 11515,5 7507,8 6865,9 RPL37 ribosomal protein
L37
201429_s_at 7948,5 7933,4 11588,5 18614,3 18264,7 19750,0 9985,2 9038,0 RPL37A ribosomal protein
L37a
202029_x_at 4824,6 4998,7 4742,3 7041,8 7654,3 7976,7 5097,7 5141,7 RPL38 ribosomal protein
L38
208695_s_at 7635,5 8267,1 8481,0 13321,9 15478,2 12368,3 9632,1 7893,7 RPL39 ribosomal protein
L39
200089_s_at 6279,8 6175,9 4360,5 8486,7 8488,9 9345,2 7225,2 6325,5 RPL4 ribosomal protein L4
201154_x_at 4812,6 5119,3 3754,5 7616,0 7189,7 9423,4 5944,4 6122,7 RPL4 ribosomal protein L4
211710_x_at 4854,5 5205,1 4120,4 8826,4 8265,6 10879,6 6111,0 6226,0 RPL4 ribosomal protein L4
201492_s_at 6633,6 6608,6 14806,0 18203,8 16266,0 17137,2 8848,0 8284,3 RPL41 ribosomal protein
L41
213080_x_at 6835,6 7527,7 6082,5 9760,4 12692,5 12330,9 8859,2 7839,8 RPL5 ribosomal protein L5
200937_s_at 4761,3 4040,2 4483,2 6473,2 7159,0 8199,3 5041,3 5212,8 RPL5 ribosomal protein L5
200034_s_at 6746,8 6142,2 7900,5 12272,0 10559,3 11874,5 7103,7 6284,7 RPL6 ribosomal protein L6
200717_x_at 7834,8 7737,7 9642,4 14112,7 13640,8 14724,6 9293,2 8822,5 RPL7 ribosomal protein L7
212042_x_at 6730,9 6433,1 7796,5 11222,3 11392,8 12537,2 8039,8 7307,4 RPL7 ribosomal protein L7
217740_x_at 6976,0 6367,1 8487,4 11682,2 11677,7 13870,0 8146,7 8298,1 RPL7A ribosomal protein
L7a
224930_x_at 6125,3 5661,4 6988,6 10895,7 10328,9 12741,5 7039,6 6866,9 RPL7A ribosomal protein
L7a
200936_at 5875,1 5065,9 6694,8 8076,7 8120,7 8583,1 5815,9 6051,7 RPL8 ribosomal protein L8
200032_s_at 7136,3 6514,4 8798,9 12409,7 13105,0 13283,9 7929,1 7631,3 RPL9 ribosomal protein L9
201033_x_at 6207,5 6046,7 9871,8 14253,2 13900,3 14493,7 7863,7 7197,6 RPLP0 ribosomal protein,
large, P0
208856_x_at 6214,3 6067,8 9521,2 14496,9 14185,7 14613,5 7952,1 7624,7 RPLP0 ribosomal protein,
large, P0
211720_x_at 6298,9 5839,2 10435,9 14763,0 14787,6 15555,1 8142,4 7730,5 RPLP0 ribosomal protein,
large, P0
211972_x_at 5928,7 5406,8 9079,7 12929,1 13012,4 13364,6 7233,3 6793,9 RPLP0 ribosomal protein,
large, P0
214167_s_at 5675,1 5035,2 7005,3 11508,6 11078,0 12094,5 6496,4 6146,0 RPLP0 ribosomal protein,
large, P0
200763_s_at 7077,6 7109,6 10067,6 15462,6 13540,3 14698,2 8192,4 7624,8 RPLP1 ribosomal protein,
large, P1
200909_s_at 6461,0 6897,0 8119,6 11270,3 11682,8 12329,1 7399,3 6810,9 RPLP2 ribosomal protein,
large, P2
200095_x_at 7311,5 7036,0 8170,3 11937,8 11599,1 12240,6 8755,8 8265,5 RPS10 ribosomal protein
S10
200817_x_at 7547,1 7500,4 10450,8 15048,7 14494,6 14628,1 9026,5 8509,5 RPS10 ribosomal protein
S10

Tablas Suplementarias
110
211542_x_at 7587,8 7797,8 10182,2 14513,3 14000,0 14244,0 8882,0 8346,0 RPS10 ribosomal protein
S10
200031_s_at 7538,5 7095,7 8972,1 13495,9 13913,5 14725,2 8938,5 7967,4 RPS11 ribosomal protein
S11
213377_x_at 7173,4 7117,2 7951,9 11671,5 11597,0 13046,1 8232,3 7822,7 RPS12 ribosomal protein
S12
200018_at 7349,1 7606,1 7590,7 10176,4 11274,6 11447,2 8255,1 8244,0 RPS13 ribosomal protein
S13
208645_s_at 6814,1 6458,1 9311,4 14255,1 14037,4 13518,6 8178,6 7634,1 RPS14 ribosomal protein
S14
200819_s_at 5343,8 5747,0 5035,4 8309,3 8026,0 8765,4 5264,1 6021,5 RPS15 ribosomal protein
S15
200781_s_at 5472,2 5255,3 4853,3 9048,2 10755,0 9126,4 6528,7 6332,1 RPS15A ribosomal protein
S15a
226131_s_at 7840,1 7706,0 11049,9 16331,7 16069,1 16933,6 9816,6 8703,3 RPS16 ribosomal protein
S16
213890_x_at 5535,9 4553,9 5207,9 7644,4 8494,8 8617,0 6035,4 5947,1 RPS16 ribosomal protein
S16
201258_at 5646,3 4624,1 3777,8 6209,4 7778,9 7822,0 5719,2 5439,0 RPS16 ribosomal protein
S16
211487_x_at 5249,3 5259,9 7151,7 9660,8 9756,8 10379,5 5982,2 5348,6 RPS17 ribosomal protein
S17
201665_x_at 5400,6 4729,7 7062,1 9544,6 10345,2 10509,5 6400,5 6169,4 RPS17 ribosomal protein
S17
212578_x_at 4728,2 4934,8 4728,4 7347,8 7677,6 8403,2 5416,1 5123,5 RPS17 ribosomal protein
S17
202649_x_at 6706,8 6741,9 6104,4 8029,0 10406,8 10858,4 7440,1 6883,2 RPS19 ribosomal protein
S19
213414_s_at 7384,8 7248,6 8175,8 11183,6 13089,7 12982,3 8515,8 7735,7 RPS19 ribosomal protein
S19
203107_x_at 6529,5 6093,2 14304,8 20537,5 16650,9 18823,0 8381,7 7870,1 RPS2 ribosomal protein S2
212433_x_at 6916,3 6243,9 10640,7 17333,0 15121,3 17841,4 8382,9 7932,8 RPS2 ribosomal protein S2
217466_x_at 4362,3 3576,6 356,0 1576,2 3671,3 6874,7 4797,7 4781,4 RPS2 ribosomal protein S2
200949_x_at 5175,1 5026,3 5378,5 5890,9 7329,3 7420,0 5623,1 5633,1 RPS20 ribosomal protein
S20
214003_x_at 7090,4 7100,2 9098,0 10242,2 14214,6 11399,6 8735,1 9016,5 RPS20 ribosomal protein
S20
200834_s_at 6320,4 5866,0 3813,9 5713,0 7895,5 5471,0 6226,2 6486,7 RPS21 ribosomal protein
S21
200926_at 8648,6 8295,9 13230,6 18987,0 16390,0 19413,9 10101,4 9357,3 RPS23 ribosomal protein
S23
200061_s_at 5827,7 5914,9 9659,2 13075,3 11996,4 13104,0 6966,1 6660,2 RPS24 ribosomal protein
S24
200091_s_at 5376,5 5245,0 4511,5 6737,9 7730,6 7200,3 5712,8 5615,6 RPS25 ribosomal protein
S25
200741_s_at 7159,9 7165,4 10253,5 14202,8 15402,5 14871,5 8576,3 7551,4 RPS27 ribosomal protein
S27
211296_x_at 5915,0 5863,3 14209,3 18299,8 13207,6 14432,6 8004,3 6973,9 RPS27A ribosomal protein
S27a
208980_s_at 6132,5 3664,7 5557,1 6020,8 7089,4 7253,6 6146,7 5735,2 RPS27A ribosomal protein
S27a
200633_at 5937,9 6275,2 7536,9 10367,3 7548,1 8327,2 6861,3 6694,0 RPS27A ribosomal protein
S27a
200017_at 7251,6 7230,8 4579,5 7318,4 9140,6 9191,1 8135,0 7631,6 RPS27A ribosomal protein
S27a
218007_s_at 3448,7 5269,1 3368,5 4048,6 3768,0 3280,2 3536,2 3953,4 RPS27L ribosomal protein
S27-like
208904_s_at 4851,6 4569,0 6119,7 8945,2 8880,5 10005,0 5625,7 4960,6 RPS28 ribosomal protein
S28
201094_at 6486,8 5834,8 4934,2 5511,5 8167,0 6705,6 5896,2 6171,0 RPS29 ribosomal protein
S29
208692_at 6629,4 5446,8 6383,8 9558,0 10405,0 11245,0 6687,0 6724,9 RPS3 ribosomal protein S3
201257_x_at 7671,4 7306,5 10682,1 16601,7 15758,9 17046,4 9494,3 9073,0 RPS3A ribosomal protein
S3A

Tablas Suplementarias
111
212391_x_at 8201,1 7483,4 10932,9 17391,4 16750,0 17668,6 9577,7 9424,3 RPS3A ribosomal protein
S3A
200933_x_at 6959,5 6877,0 12294,2 18427,1 15783,9 16897,1 9160,5 8273,6 RPS4X ribosomal protein
S4, X-linked
213347_x_at 6248,9 6337,9 10691,9 16743,1 13127,7 14566,5 8277,6 7894,3 RPS4X ribosomal protein
S4, X-linked
200024_at 5001,2 4802,6 3649,4 4071,1 7035,8 4459,2 5386,8 4901,8 RPS5 ribosomal protein S5
201254_x_at 6962,0 6946,3 6557,9 10131,9 11152,0 12421,7 7517,5 7580,2 RPS6 ribosomal protein S6
209134_s_at 6399,3 5809,1 7913,5 10491,6 10540,4 12326,4 6533,3 6454,2 RPS6 ribosomal protein S6
200081_s_at 5336,1 4375,9 5563,8 7611,6 8209,2 9546,7 5118,2 5725,9 RPS6 ribosomal protein S6
213941_x_at 4935,2 4893,6 5623,1 7322,8 7065,6 6992,5 5856,4 5506,9 RPS7 ribosomal protein S7
200858_s_at 5571,5 4944,7 4934,3 7457,2 7910,9 9313,8 5328,0 5170,8 RPS8 ribosomal protein S8
217747_s_at 4910,0 4428,6 3202,8 4397,5 5335,6 5707,0 4350,8 4246,9 RPS9 ribosomal protein S9
214629_x_at 5025,1 5526,2 6637,7 9710,5 6881,7 7898,2 6182,3 5761,0 RTN4 reticulon 4
210968_s_at 4098,6 5698,2 5529,5 7505,8 5628,6 5999,6 5645,3 5393,4 RTN4 reticulon 4
211509_s_at 4261,0 5303,9 5946,5 7084,9 5251,9 5475,9 5650,9 5236,2 RTN4 reticulon 4
200872_at 5031,0 5648,8 4597,2 5184,6 3480,8 3449,0 4108,6 5399,0 S100A1
0 S100 calcium binding
protein A10
203186_s_at 2631,4 7907,7 4460,0 6283,3 3738,0 4688,1 6009,4 6719,7 S100A4 S100 calcium binding
protein A4
217728_at 6127,3 6781,9 8016,4 13404,3 10310,0 13074,8 8535,0 8438,5 S100A6 S100 calcium binding
protein A6
210592_s_at 3785,9 5017,1 3633,3 4006,0 4761,7 3129,2 4877,4 2881,8 SAT1 spermidine/spermin
e N1-acetyltransferase 1
203455_s_at 4662,1 4801,6 3410,0 3616,8 4172,5 2744,7 5166,3 3335,0 SAT1 spermidine/spermin
e N1-acetyltransferase 1
205475_at 5072,3 1812,8 2414,3 1978,6 2676,1 1224,1 3612,4 1072,2 SCRG1 scrapie responsive
protein 1
212190_at 7249,1 8356,3 18255,5 19940,6 9974,7 8553,8 6600,6 6074,6 SERPINE
2
serpin peptidase inhibitor, clade E
(nexin, plasminogen activator inhibitor type 1), member 2
200718_s_at 5373,5 6508,4 5247,5 6982,9 6180,1 5376,1 6086,3 6125,7 SKP1 S-phase kinase-
associated protein 1
200030_s_at 3749,0 4478,9 4187,7 5558,0 4924,9 5658,6 4570,3 4605,5 SLC25A3
solute carrier family 25 (mitochondrial carrier; phosphate carrier), member 3
212085_at 3524,4 3235,6 2646,2 4360,4 5166,1 5574,5 4323,6 3879,2 SLC25A6
solute carrier family 25 (mitochondrial carrier; adenine
nucleotide translocator),
member 6
212826_s_at 3359,8 4610,5 2448,3 3977,2 4764,4 4812,2 5133,4 4411,1 SLC25A6
solute carrier family 25 (mitochondrial carrier; adenine
nucleotide translocator),
member 6
222982_x_at 4720,8 5618,6 5222,0 6886,5 5722,3 7232,1 5957,2 6304,8 SLC38A2 solute carrier family
38, member 2
218041_x_at 4036,4 4100,4 3748,3 4982,5 4269,9 5312,2 4548,5 5190,4 SLC38A2 solute carrier family
38, member 2
220924_s_at 3876,0 3772,0 3633,2 4894,8 4253,2 5378,4 4375,0 4951,6 SLC38A2 solute carrier family
38, member 2
225155_at 3413,0 5485,2 5042,2 7400,6 9109,2 8885,9 6767,2 6617,0 SNHG5 small nucleolar RNA
host gene 5 (non-protein coding)

Tablas Suplementarias
112
200665_s_at 5279,2 4740,0 10026,8 12911,3 10637,0 11525,0 6390,4 5539,8 SPARC secreted protein,
acidic, cysteine-rich (osteonectin)
212667_at 3912,4 3775,7 3572,4 4761,8 5275,2 4440,3 5105,1 4221,5 SPARC secreted protein,
acidic, cysteine-rich (osteonectin)
201859_at 4799,8 4954,5 3628,0 5732,6 5391,1 4428,9 4225,3 4085,0 SRGN Serglycin
225987_at 2012,6 5707,9 4646,6 4857,3 5150,2 3855,5 5051,6 3715,3 STEAP4 STEAP family
member 4
201061_s_at 1935,9 3100,8 2479,9 4074,9 5128,5 3919,1 4711,2 3478,3 STOM Stomatin
227480_at 1517,1 3586,3 2476,4 3425,1 2729,7 5220,0 1723,3 2284,8 SUSD2 sushi domain containing 2
201506_at 6829,3 6669,9 11400,8 15293,7 15466,2 13771,5 9312,5 8077,4 TGFBI transforming growth factor, beta-induced,
68kDa
201110_s_at 2322,9 3884,1 2123,6 3349,4 3713,0 2821,9 6364,3 4268,7 THBS1 thrombospondin 1
201666_at 7594,2 7463,6 12276,7 14447,6 14762,0 12916,9 8704,9 6801,5 TIMP1 TIMP
metallopeptidase inhibitor 1
231579_s_at 3210,6 3807,4 4618,8 4265,3 5818,8 4922,5 5069,3 3418,1 TIMP2 TIMP
metallopeptidase inhibitor 2
217733_s_at 6720,2 8046,8 10702,5 13183,8 10669,1 10955,2 9285,2 8838,6 TMSB10 thymosin beta 10
216438_s_at 4128,9 4842,3 6042,9 8660,0 5620,3 8647,0 5687,6 6514,6 TMSB4X thymosin beta 4, X-
linked
201645_at 6946,8 4393,8 7049,9 5808,4 5862,2 4533,1 5413,9 3569,6 TNC tenascin C
224565_at 3226,3 5422,6 1317,9 1721,4 5982,3 2582,3 5698,8 4409,7 TncRNA trophoblast-derived
noncoding RNA
200822_x_at 4288,8 4047,7 4226,4 6007,4 5063,3 3916,4 4458,6 3768,4 TPI1 triosephosphate
isomerase 1
213011_s_at 4869,5 4831,1 3779,6 4615,8 5199,3 3377,7 5013,3 4645,4 TPI1 triosephosphate
isomerase 1
204083_s_at 2165,0 4365,4 3620,5 5875,4 3707,7 5711,0 4633,9 4936,0 TPM2 tropomyosin 2 (beta)
211943_x_at 6577,8 6560,2 15284,2 19143,0 16929,6 17734,4 8134,6 7081,5 TPT1 tumor protein, translationally-
controlled 1
212284_x_at 7452,5 7616,5 18886,9 25430,2 21531,7 21938,4 10019,3 8821,3 TPT1 tumor protein, translationally-
controlled 1
212869_x_at 7113,9 6927,0 17088,7 24147,8 20713,2 20515,4 9564,8 8553,3 TPT1 tumor protein, translationally-
controlled 1
214327_x_at 5558,8 5690,8 14369,3 20088,7 15738,0 17155,0 7238,4 6053,4 TPT1 tumor protein, translationally-
controlled 1
216520_s_at 6944,6 7217,7 14720,3 19519,7 18680,9 17790,2 9218,9 8365,8 TPT1 tumor protein, translationally-
controlled 1
212639_x_at 6575,4 4998,6 5460,1 6578,2 3084,0 4264,4 3871,5 5577,3 TUBA1B tubulin, alpha 1b
211058_x_at 5533,1 4240,4 4075,6 6780,1 2976,3 3517,0 3731,6 5069,5 TUBA1B tubulin, alpha 1b
211072_x_at 5052,0 4011,7 4563,6 6483,9 2819,4 3217,4 3640,1 4612,5 TUBA1B tubulin, alpha 1b
213646_x_at 5539,0 3954,3 3788,0 5664,0 2715,0 3163,1 3279,8 4383,5 TUBA1B tubulin, alpha 1b
201090_x_at 5103,6 3598,5 3228,1 4891,0 2256,3 2933,5 2966,8 4131,4 TUBA1B tubulin, alpha 1b
209251_x_at 4601,3 3508,0 3753,4 5207,3 2759,9 2812,7 2943,2 3818,5 TUBA1C tubulin, alpha 1c
211714_x_at 5251,5 2095,0 3236,2 3388,6 2292,6 2780,8 2184,0 2890,1 TUBB tubulin, beta
201010_s_at 1754,7 1805,8 1694,1 2530,5 5011,7 6132,1 3102,3 1899,7 TXNIP thioredoxin
interacting protein

Tablas Suplementarias
113
225413_at 3854,1 5295,6 2602,1 2846,5 3173,7 2128,1 3734,2 4005,9 USMG5
up-regulated during skeletal muscle
growth 5 homolog (mouse)
204620_s_at 3539,8 3170,7 2326,2 3741,0 5767,0 2802,2 3640,1 1602,4 VCAN Versican
221731_x_at 3390,4 3321,2 1797,5 3179,1 5560,0 2388,5 3858,4 1699,8 VCAN Versican
201426_s_at 6023,1 5940,8 13715,6 18176,0 15271,3 16389,8 8249,8 7489,1 VIM Vimentin
235751_s_at 1124,2 3726,2 4790,5 4691,8 7551,1 3817,9 3110,4 1653,8 VMO1 vitelline membrane
outer layer 1 homolog (chicken)
208628_s_at 4258,9 4723,8 4031,0 5088,2 3788,0 4428,9 4225,4 5145,1 YBX1 Y box binding
protein 1

Tablas Suplementarias
114
Tabla 5.- Tabla con las 242 funciones génicas correspondientes al primer subcultivo que se
encontraron sobreexpresadas utilizando el programa Cytoscape-BINGO®.
Refernecia Gene Ontology
Valor estadístico p
Valor estadístico p corregido
Funciones Génicas
3740 5,79E-72 4,61E-69 structural constituent of
ribosome
5830 1,85E-71 7,38E-69 cytosolic ribosome (sensu
Eukaryota)
5840 2,55E-70 6,76E-68 Ribosome
5198 1,40E-61 2,79E-59 structural molecule activity
44445 3,60E-59 5,73E-57 cytosolic part
30529 5,55E-57 7,37E-55 ribonucleoprotein complex
6416 1,97E-53 2,24E-51 protein biosynthesis
9059 1,44E-50 1,43E-48 macromolecule biosynthesis
16283 9,66E-44 7,69E-42 eukaryotic 48S initiation complex
5843 9,66E-44 7,69E-42 cytosolic small ribosomal subunit
(sensu Eukaryota)
44249 2,26E-39 1,64E-37 cellular biosynthesis
5829 2,14E-38 1,42E-36 Cytosol
16282 1,19E-36 7,26E-35 eukaryotic 43S preinitiation
complex
9058 4,19E-36 2,38E-34 Biosynthesis
43232 2,25E-35 1,12E-33 intracellular non-membrane-
bound organelle
43228 2,25E-35 1,12E-33 non-membrane-bound organelle
43234 5,26E-35 2,46E-33 protein complex
5737 7,91E-35 3,50E-33 Cytoplasm
15935 3,43E-34 1,44E-32 small ribosomal subunit
44444 6,06E-32 2,41E-30 cytoplasmic part
44260 1,05E-23 3,97E-22 cellular macromolecule
metabolism
44267 1,82E-23 6,57E-22 cellular protein metabolism
5842 8,45E-23 2,93E-21 cytosolic large ribosomal subunit
(sensu Eukaryota)
19538 4,76E-22 1,58E-20 protein metabolism
15934 7,57E-20 2,41E-18 large ribosomal subunit
3723 7,27E-18 2,22E-16 RNA binding
43170 1,88E-16 5,55E-15 macromolecule metabolism
44422 1,86E-15 5,10E-14 organelle part
44446 1,86E-15 5,10E-14 intracellular organelle part
44424 4,73E-10 1,26E-08 intracellular part

Tablas Suplementarias
115
19843 6,55E-08 1,68E-06 rRNA binding
5622 6,82E-08 1,70E-06 Intracelular
43229 6,31E-06 1,49E-04 intracellular organelle
43226 6,37E-06 1,49E-04 Organelle
8151 1,62E-05 3,68E-04 cellular physiological process
5578 1,12E-04 2,41E-03 extracellular matrix (sensu
Metazoa)
6455 1,12E-04 2,41E-03 translational elongation
44237 1,16E-04 2,43E-03 cellular metabolism
5202 2,96E-04 6,05E-03 Collagen
44238 1,64E-03 3,26E-02 primary metabolism
8152 3,50E-03 6,80E-02 Metabolism
31012 6,68E-03 1,27E-01 extracellular matrix
7582 1,31E-02 2,43E-01 physiological process
5583 1,91E-02 3,46E-01 fibrillar collagen
43037 2,25E-02 3,97E-01 Translation
5201 2,69E-02 4,66E-01 extracellular matrix structural
constituent
44421 4,67E-01 7,91E+00 extracellular region part
3676 9,41E-01 1,49E+01 nucleic acid binding
9987 9,69E-01 1,49E+01 cellular process
8043 9,89E-01 1,49E+01 ferritin complex
30049 9,89E-01 1,49E+01 muscle filament sliding
6123 9,89E-01 1,49E+01 mitochondrial electron transport,
cytochrome c to oxygen
5589 9,89E-01 1,49E+01 collagen type VI
5575 1,01E+00 1,49E+01 cellular_component
5576 2,19E+00 3,17E+01 extracellular región
42775 2,76E+00 3,92E+01 ATP synthesis coupled electron
transport (sensu Eukaryota)
47115 2,95E+00 4,05E+01 trans-1,2-dihydrobenzene-1,2-
diol dehydrogenase activity
5584 2,95E+00 4,05E+01 collagen type I
5746 3,12E+00 4,19E+01 mitochondrial electron transport
chain
6817 3,16E+00 4,19E+01 phosphate transport
48513 3,36E+00 4,38E+01 organ development
42773 3,93E+00 5,04E+01 ATP synthesis coupled electron
transport
44420 4,03E+00 5,10E+01 extracellular matrix part
8147 5,86E+00 7,28E+01 structural constituent of bone
5518 5,98E+00 7,32E+01 collagen binding

Tablas Suplementarias
116
42254 7,08E+00 8,53E+01 ribosome biogenesis and
assembly
42592 7,65E+00 9,08E+01 Homeostasis
7519 7,95E+00 9,30E+01 striated muscle development
46790 9,70E+00 1,12E+02 virion binding
16461 1,45E+01 1,60E+02 unconventional myosin
5747 1,45E+01 1,60E+02 respiratory chain complex I
(sensu Eukaryota)
45271 1,45E+01 1,60E+02 respiratory chain complex I
8182 1,49E+01 1,63E+02 translation elongation factor
activity
7028 1,52E+01 1,64E+02 cytoplasm organization and
biogénesis
51082 1,65E+01 1,73E+02 unfolded protein binding
8150 1,66E+01 1,73E+02 biological_process
31406 2,01E+01 2,03E+02 carboxylic acid binding
51238 2,01E+01 2,03E+02 sequestering of metal ion
42255 2,01E+01 2,03E+02 ribosome assembly
5730 2,27E+01 2,26E+02 Nucleolus
6096 2,34E+01 2,30E+02 Glycolysis
46916 2,66E+01 2,49E+02 transition metal ion homeostasis
5793 2,66E+01 2,49E+02 ER-Golgi intermediate
compartment
30162 2,66E+01 2,49E+02 regulation of proteolysis
5853 2,66E+01 2,49E+02 eukaryotic translation elongation
factor 1 complex
5507 3,40E+01 3,11E+02 copper ion binding
30934 3,40E+01 3,11E+02 anchoring collagen
7517 3,55E+01 3,21E+02 muscle development
16477 3,71E+01 3,24E+02 cell migration
44275 3,71E+01 3,24E+02 cellular carbohydrate catabolism
16052 3,71E+01 3,24E+02 carbohydrate catabolism
30005 3,86E+01 3,34E+02 di-, tri-valent inorganic cation
homeostasis
8201 4,03E+01 3,45E+02 heparin binding
15698 4,21E+01 3,49E+02 inorganic anion transport
5770 4,22E+01 3,49E+02 late endosome
44455 4,25E+01 3,49E+02 mitochondrial membrane part
6007 4,25E+01 3,49E+02 glucose catabolism
6875 5,09E+01 4,14E+02 metal ion homeostasis
5768 5,25E+01 4,22E+02 Endosome
45182 5,70E+01 4,53E+02 translation regulator activity

Tablas Suplementarias
117
Tabla 6.- Tabla con las 232 funciones génicas correspondientes al segundo subcultivo que se
encontraron sobreexpresadas utilizando el programa Cytoscape-BINGO®.
Refernecia Gene Ontology
Valor estadístico p
Valor estadístico p corregido
Funciones Génicas
3740 1,88E-72 1,59E-69 structural constituent of ribosome
5840 8,31E-71 3,52E-68 Ribosome
5830 1,09E-66 3,06E-64 cytosolic ribosome (sensu
Eukaryota)
5198 3,37E-62 7,13E-60 structural molecule activity
30529 1,85E-57 3,14E-55 ribonucleoprotein complex
44445 1,51E-53 2,13E-51 cytosolic part
6416 1,24E-52 1,49E-50 protein biosynthesis
9059 8,23E-50 8,70E-48 macromolecule biosynthesis
44249 8,71E-39 8,19E-37 cellular biosynthesis
5737 1,49E-37 1,26E-35 Cytoplasm
9058 1,35E-36 1,04E-34 Biosynthesis
16283 3,49E-36 2,27E-34 eukaryotic 48S initiation complex
5843 3,49E-36 2,27E-34 cytosolic small ribosomal subunit
(sensu Eukaryota)
43232 6,76E-36 3,81E-34 intracellular non-membrane-
bound organelle
43228 6,76E-36 3,81E-34 non-membrane-bound organelle
5829 1,25E-34 6,62E-33 cytosol
43234 1,14E-32 5,65E-31 protein complex
44444 1,48E-32 6,94E-31 cytoplasmic part
16282 1,78E-30 7,91E-29 eukaryotic 43S preinitiation
complex
15935 1,29E-28 5,46E-27 small ribosomal subunit
5842 3,24E-25 1,31E-23 cytosolic large ribosomal subunit
(sensu Eukaryota)
44260 1,07E-22 4,13E-21 cellular macromolecule
metabolism
44267 1,88E-22 6,91E-21 cellular protein metabolism
15934 9,16E-22 3,23E-20 large ribosomal subunit
19538 4,29E-21 1,45E-19 protein metabolism
43170 2,33E-16 7,57E-15 macromolecule metabolism
3723 3,17E-15 9,94E-14 RNA binding
44422 2,20E-14 6,43E-13 organelle part
44446 2,20E-14 6,43E-13 intracellular organelle part
44424 1,67E-12 4,72E-11 intracellular part
5622 3,52E-10 9,60E-09 intracellular
8151 3,46E-09 9,15E-08 cellular physiological process

Tablas Suplementarias
118
19843 5,99E-08 1,54E-06 rRNA binding
43229 8,85E-08 2,16E-06 intracellular organelle
43226 8,95E-08 2,16E-06 organelle
7582 5,05E-06 1,19E-04 physiological process
44237 5,49E-06 1,25E-04 cellular metabolism
8152 8,32E-05 1,85E-03 metabolism
44238 2,66E-04 5,77E-03 primary metabolism
6455 3,27E-03 6,92E-02 translational elongation
5575 5,75E-03 1,19E-01 cellular_component
5202 7,39E-03 1,49E-01 collagen
5201 2,45E-02 4,82E-01 extracellular matrix structural
constituent
9987 3,39E-02 6,51E-01 cellular process
43037 1,36E-01 2,56E+00 translation
5578 1,40E-01 2,58E+00 extracellular matrix (sensu
Metazoa)
6007 4,64E-01 8,35E+00 glucose catabolism
44464 6,89E-01 1,17E+01 cell part
5623 6,93E-01 1,17E+01 cell
44275 7,08E-01 1,17E+01 cellular carbohydrate catabolism
16052 7,08E-01 1,17E+01 carbohydrate catabolism
19320 9,55E-01 1,46E+01 hexose catabolism
8043 9,64E-01 1,46E+01 ferritin complex
30049 9,64E-01 1,46E+01 muscle filament sliding
6123 9,64E-01 1,46E+01 mitochondrial electron transport,
cytochrome c to oxygen
5589 9,64E-01 1,46E+01 collagen type VI
46365 1,03E+00 1,53E+01 monosaccharide catabolism
5583 1,07E+00 1,56E+01 fibrillar collagen
46164 1,11E+00 1,59E+01 alcohol catabolism
42592 1,36E+00 1,92E+01 homeostasis
8150 1,45E+00 2,01E+01 biological_process
6096 2,22E+00 3,03E+01 glycolysis
31012 2,44E+00 3,28E+01 extracellular matrix
42775 2,63E+00 3,48E+01 ATP synthesis coupled electron
transport (sensu Eukaryota)
47115 2,87E+00 3,59E+01 trans-1,2-dihydrobenzene-1,2-diol
dehydrogenase activity
5584 2,87E+00 3,59E+01 collagen type I
6091 2,89E+00 3,59E+01 generation of precursor metabolites and energy
51082 2,94E+00 3,59E+01 unfolded protein binding
6817 2,95E+00 3,59E+01 phosphate transport

Tablas Suplementarias
119
5746 2,97E+00 3,59E+01 mitochondrial electron transport
chain
42773 3,74E+00 4,46E+01 ATP synthesis coupled electron
transport
44421 3,81E+00 4,47E+01 extracellular region part
6006 4,15E+00 4,81E+01 glucose metabolism
44455 4,80E+00 5,48E+01 mitochondrial membrane part
40011 5,09E+00 5,59E+01 locomotion
51674 5,09E+00 5,59E+01 localization of cell
6928 5,09E+00 5,59E+01 cell motility
16477 5,41E+00 5,87E+01 cell migration
8147 5,71E+00 6,12E+01 structural constituent of bone
42254 6,62E+00 7,00E+01 ribosome biogenesis and assembly
6092 7,30E+00 7,63E+01 main pathways of carbohydrate
metabolism
7519 7,58E+00 7,82E+01 striated muscle development
48513 9,19E+00 9,36E+01 organ development
46790 9,46E+00 9,53E+01 virion binding
5576 1,10E+01 1,09E+02 extracellular region
6119 1,16E+01 1,14E+02 oxidative phosphorylation
5520 1,26E+01 1,22E+02 insulin-like growth factor binding
16461 1,41E+01 1,31E+02 unconventional myosin
5747 1,41E+01 1,31E+02 respiratory chain complex I (sensu
Eukaryota)
19642 1,41E+01 1,31E+02 anaerobic glycolysis
45271 1,41E+01 1,31E+02 respiratory chain complex I
7028 1,43E+01 1,31E+02 cytoplasm organization and
biogenesis
8182 1,44E+01 1,31E+02 translation elongation factor
activity
44420 1,87E+01 1,68E+02 extracellular matrix part
31406 1,96E+01 1,71E+02 carboxylic acid binding
51238 1,96E+01 1,71E+02 sequestering of metal ion
42255 1,96E+01 1,71E+02 ribosome assembly
19318 2,28E+01 1,97E+02 hexose metabolism
46916 2,56E+01 2,18E+02 transition metal ion homeostasis
30162 2,60E+01 2,18E+02 regulation of proteolysis
42632 2,60E+01 2,18E+02 cholesterol homeostasis
5996 2,73E+01 2,27E+02 monosaccharide metabolism
16491 3,25E+01 2,67E+02 oxidoreductase activity
30934 3,32E+01 2,67E+02 anchoring collagen
4459 3,32E+01 2,67E+02 L-lactate dehydrogenase activity
7517 3,37E+01 2,69E+02 muscle development

Tablas Suplementarias
120
30005 3,66E+01 2,89E+02 di-, tri-valent inorganic cation
homeostasis
8201 3,85E+01 3,02E+02 heparin binding
15698 3,96E+01 3,07E+02 inorganic anion transport
4457 4,12E+01 3,14E+02 lactate dehydrogenase activity
5770 4,12E+01 3,14E+02 late endosome
15980 4,49E+01 3,39E+02 energy derivation by oxidation of
organic compounds
6875 4,83E+01 3,62E+02 metal ion homeostasis
3674 5,00E+01 3,69E+02 molecular_function
5768 5,02E+01 3,69E+02 endosome
3676 5,29E+01 3,86E+02 nucleic acid binding
45182 5,40E+01 3,91E+02 translation regulator activity
30003 6,03E+01 4,32E+02 cation homeostasis
5743 6,47E+01 4,60E+02 mitochondrial inner membrane
30218 7,01E+01 4,94E+02 erythrocyte differentiation

Tablas Suplementarias
121
Tabla 7.- Tabla con las 227 funciones génicas correspondientes al tercer subcultivo que se
encontraron sobreexpresadas utilizando el programa Cytoscape-BINGO®.
Refernecia Gene Ontology
Valor estadístico p
Valor estadístico p corregido
Funciones Génicas
3740 1,47E-59 1,14E-56 structural constituent of
ribosome
5840 3,54E-58 1,37E-55 ribosome
5198 5,22E-52 1,35E-49 structural molecule activity
5830 7,51E-52 1,46E-49 cytosolic ribosome (sensu
Eukaryota)
30529 6,25E-47 9,70E-45 ribonucleoprotein complex
6416 1,80E-43 2,33E-41 protein biosynthesis
44445 3,53E-42 3,92E-40 cytosolic part
9059 4,66E-41 4,52E-39 macromolecule biosynthesis
16283 2,32E-32 1,80E-30 eukaryotic 48S initiation complex
5843 2,32E-32 1,80E-30 cytosolic small ribosomal subunit
(sensu Eukaryota)
44249 9,30E-32 6,56E-30 cellular biosynthesis
5737 2,28E-31 1,47E-29 cytoplasm
43232 8,01E-30 4,44E-28 intracellular non-membrane-
bound organelle
43228 8,01E-30 4,44E-28 non-membrane-bound organelle
9058 5,33E-29 2,76E-27 biosynthesis
16282 2,02E-27 9,79E-26 eukaryotic 43S preinitiation
complex
43234 3,16E-27 1,44E-25 protein complex
5829 3,54E-26 1,53E-24 cytosol
15935 5,80E-26 2,37E-24 small ribosomal subunit
44444 2,13E-25 8,25E-24 cytoplasmic part
44260 3,96E-16 1,46E-14 cellular macromolecule
metabolism
44267 7,44E-16 2,63E-14 cellular protein metabolism
19538 6,61E-15 2,23E-13 protein metabolism
5842 7,43E-15 2,40E-13 cytosolic large ribosomal subunit
(sensu Eukaryota)
15934 1,02E-13 3,18E-12 large ribosomal subunit
3723 3,47E-11 1,04E-09 RNA binding
44422 5,68E-10 1,57E-08 organelle part
44446 5,68E-10 1,57E-08 intracellular organelle part
43170 7,60E-10 2,03E-08 macromolecule metabolism
44424 1,88E-08 4,85E-07 intracellular part
5622 4,17E-07 1,04E-05 intracellular

Tablas Suplementarias
122
19843 4,00E-06 9,70E-05 rRNA binding
8151 3,16E-05 7,44E-04 cellular physiological process
7582 2,32E-04 5,31E-03 physiological process
43229 4,55E-04 9,89E-03 intracellular organelle
43226 4,59E-04 9,89E-03 organelle
5578 1,23E-03 2,57E-02 extracellular matrix (sensu
Metazoa)
6455 1,81E-03 3,71E-02 translational elongation
5202 4,11E-03 8,17E-02 collagen
5575 5,56E-03 1,08E-01 cellular_component
5576 1,18E-02 2,24E-01 extracellular region
31012 4,29E-02 7,93E-01 extracellular matrix
43037 5,64E-02 1,02E+00 translation
9987 7,31E-02 1,29E+00 cellular process
44421 9,42E-02 1,62E+00 extracellular region part
8150 1,14E-01 1,92E+00 biological_process
5201 1,34E-01 2,22E+00 extracellular matrix structural
constituent
44237 3,88E-01 6,20E+00 cellular metabolism
5520 3,91E-01 6,20E+00 insulin-like growth factor binding
8043 7,90E-01 1,14E+01 ferritin complex
30049 7,90E-01 1,14E+01 muscle filament sliding
6123 7,90E-01 1,14E+01 mitochondrial electron
transport, cytochrome c to oxygen
5589 7,90E-01 1,14E+01 collagen type VI
5583 7,96E-01 1,14E+01 fibrillar collagen
48513 9,66E-01 1,36E+01 organ development
6817 1,72E+00 2,38E+01 phosphate transport
42775 1,79E+00 2,44E+01 ATP synthesis coupled electron
transport (sensu Eukaryota)
5746 2,03E+00 2,71E+01 mitochondrial electron transport
chain
47115 2,35E+00 3,05E+01 trans-1,2-dihydrobenzene-1,2-
diol dehydrogenase activity
5584 2,35E+00 3,05E+01 collagen type I
42773 2,56E+00 3,26E+01 ATP synthesis coupled electron
transport
44238 3,15E+00 3,95E+01 primary metabolism
8152 3,70E+00 4,56E+01 metabolism
42254 3,91E+00 4,74E+01 ribosome biogenesis and
assembly
8147 4,68E+00 5,59E+01 structural constituent of bone
7519 5,21E+00 6,13E+01 striated muscle development

Tablas Suplementarias
123
40008 8,18E+00 9,47E+01 regulation of growth
7028 8,53E+00 9,73E+01 cytoplasm organization and
biogenesis
44420 1,05E+01 1,18E+02 extracellular matrix part
8182 1,08E+01 1,20E+02 translation elongation factor
activity
16461 1,16E+01 1,23E+02 unconventional myosin
5747 1,16E+01 1,23E+02 respiratory chain complex I
(sensu Eukaryota)
45271 1,16E+01 1,23E+02 respiratory chain complex I
19838 1,45E+01 1,52E+02 growth factor binding
31406 1,61E+01 1,64E+02 carboxylic acid binding
42255 1,61E+01 1,64E+02 ribosome assembly
42592 1,83E+01 1,85E+02 Homeostasis
46916 1,93E+01 1,92E+02 transition metal ion homeostasis
30162 2,13E+01 2,07E+02 regulation of proteolysis
42632 2,13E+01 2,07E+02 cholesterol homeostasis
7517 2,19E+01 2,09E+02 muscle development
7148 2,21E+01 2,09E+02 cellular morphogenesis
5507 2,26E+01 2,11E+02 copper ion binding
16477 2,28E+01 2,11E+02 cell migration
15698 2,42E+01 2,21E+02 inorganic anion transport
30934 2,73E+01 2,46E+02 anchoring collagen
44455 2,85E+01 2,54E+02 mitochondrial membrane part
1558 3,04E+01 2,68E+02 regulation of cell growth
44464 3,24E+01 2,81E+02 cell part
5623 3,26E+01 2,81E+02 Cell
45182 3,54E+01 3,02E+02 translation regulator activity
40011 4,92E+01 4,06E+02 Locomotion
51674 4,92E+01 4,06E+02 localization of cell
6928 4,92E+01 4,06E+02 cell motility
40007 5,53E+01 4,52E+02 Growth
6119 5,68E+01 4,59E+02 oxidative phosphorylation
30218 5,77E+01 4,62E+02 erythrocyte differentiation
6820 5,84E+01 4,62E+02 anion transport

Tablas Suplementarias
124
Tabla 8.- Tabla con las 312 funciones génicas correspondientes al cuarto subcultivo que se
encontraron sobreexpresadas utilizando el programa Cytoscape-BINGO®.
Refernecia Gene Ontology
Valor estadístico p
Valor estadístico p corregido
Funciones Génicas
5830 7,34E-77 7,69E-74 cytosolic ribosome (sensu
Eukaryota)
3740 2,99E-76 1,57E-73 structural constituent of
ribosome
5840 2,11E-74 7,38E-72 ribosome
44445 6,71E-64 1,76E-61 cytosolic part
5198 1,25E-63 2,61E-61 structural molecule activity
30529 1,79E-59 3,12E-57 ribonucleoprotein complex
6416 1,21E-55 1,82E-53 protein biosynthesis
9059 1,21E-53 1,59E-51 macromolecule biosynthesis
44249 7,23E-42 8,42E-40 cellular biosynthesis
9058 4,15E-41 3,59E-39 biosynthesis
16283 4,30E-41 3,59E-39 eukaryotic 48S initiation complex
5843 4,30E-41 3,59E-39 cytosolic small ribosomal subunit
(sensu Eukaryota)
5829 4,45E-41 3,59E-39 cytosol
5737 3,95E-38 2,95E-36 cytoplasm
43232 1,41E-36 9,23E-35 intracellular non-membrane-
bound organelle
43228 1,41E-36 9,23E-35 non-membrane-bound organelle
43234 3,93E-36 2,42E-34 protein complex
44444 8,42E-35 4,90E-33 cytoplasmic part
16282 5,09E-34 2,81E-32 eukaryotic 43S preinitiation
complex
15935 1,84E-31 9,62E-30 small ribosomal subunit
5842 1,42E-30 7,09E-29 cytosolic large ribosomal subunit
(sensu Eukaryota)
15934 4,02E-25 1,91E-23 large ribosomal subunit
44260 8,60E-19 3,92E-17 cellular macromolecule
metabolism
44267 1,10E-18 4,81E-17 cellular protein metabolism
19538 1,19E-17 4,97E-16 protein metabolism
3723 3,95E-15 1,59E-13 RNA binding
43170 4,28E-15 1,66E-13 macromolecule metabolism
44422 1,15E-13 4,15E-12 organelle part
44446 1,15E-13 4,15E-12 intracellular organelle part
44424 3,51E-09 1,23E-07 intracellular part
19843 3,06E-07 1,04E-05 rRNA binding

Tablas Suplementarias
125
5622 3,60E-07 1,18E-05 intracellular
8151 6,87E-06 2,18E-04 cellular physiological process
43229 1,36E-04 4,12E-03 intracellular organelle
43226 1,38E-04 4,12E-03 organelle
7582 1,67E-04 4,86E-03 physiological process
6096 2,66E-04 7,54E-03 glycolysis
6455 5,05E-04 1,39E-02 translational elongation
6007 1,17E-03 3,14E-02 glucose catabolism
5202 1,32E-03 3,47E-02 collagen
44237 1,53E-03 3,91E-02 cellular metabolism
5578 2,40E-03 5,98E-02 extracellular matrix (sensu
Metazoa)
19320 3,60E-03 8,78E-02 hexose catabolism
46365 4,04E-03 9,63E-02 monosaccharide catabolism
46164 4,53E-03 1,06E-01 alcohol catabolism
44275 5,98E-03 1,33E-01 cellular carbohydrate catabolism
16052 5,98E-03 1,33E-01 carbohydrate catabolism
5201 1,43E-02 3,12E-01 extracellular matrix structural
constituent
44238 1,88E-02 4,03E-01 primary metabolism
8152 2,99E-02 6,26E-01 metabolism
6006 3,53E-02 7,26E-01 glucose metabolism
5576 4,90E-02 9,88E-01 extracellular region
5520 6,20E-02 1,23E+00 insulin-like growth factor binding
6092 8,51E-02 1,65E+00 main pathways of carbohydrate
metabolism
31012 1,16E-01 2,20E+00 extracellular matrix
44421 1,21E-01 2,26E+00 extracellular region part
43037 1,81E-01 3,32E+00 translation
16477 4,24E-01 7,66E+00 cell migration
19318 5,01E-01 8,90E+00 hexose metabolism
6091 5,89E-01 1,03E+01 generation of precursor metabolites and energy
5996 6,64E-01 1,14E+01 monosaccharide metabolism
5575 8,99E-01 1,52E+01 cellular_component
9987 1,08E+00 1,80E+01 cellular process
15980 1,44E+00 2,26E+01 energy derivation by oxidation of
organic compounds
6817 1,46E+00 2,26E+01 phosphate transport
30934 1,50E+00 2,26E+01 anchoring collagen
8201 1,52E+00 2,26E+01 heparin binding
8043 1,53E+00 2,26E+01 ferritin complex

Tablas Suplementarias
126
30049 1,53E+00 2,26E+01 muscle filament sliding
6123 1,53E+00 2,26E+01 mitochondrial electron
transport, cytochrome c to oxygen
5589 1,53E+00 2,26E+01 collagen type VI
8182 1,69E+00 2,46E+01 translation elongation factor
activity
5583 2,12E+00 3,04E+01 fibrillar collagen
44420 3,70E+00 5,24E+01 extracellular matrix part
48513 4,16E+00 5,82E+01 organ development
47115 4,56E+00 6,05E+01 trans-1,2-dihydrobenzene-1,2-
diol dehydrogenase activity
8195 4,56E+00 6,05E+01 phosphatidate phosphatase
activity
8354 4,56E+00 6,05E+01 germ cell migration
5584 4,56E+00 6,05E+01 collagen type I
19838 5,88E+00 7,70E+01 growth factor binding
8150 6,05E+00 7,83E+01 biological_process
5539 6,16E+00 7,87E+01 glycosaminoglycan binding
42775 6,30E+00 7,95E+01 ATP synthesis coupled electron
transport (sensu Eukaryota)
40011 6,74E+00 8,22E+01 locomotion
51674 6,74E+00 8,22E+01 localization of cell
6928 6,74E+00 8,22E+01 cell motility
5746 7,10E+00 8,55E+01 mitochondrial electron transport
chain
30247 7,42E+00 8,84E+01 polysaccharide binding
40008 8,60E+00 1,01E+02 regulation of growth
42773 8,91E+00 1,03E+02 ATP synthesis coupled electron
transport
8147 9,04E+00 1,03E+02 structural constituent of bone
4128 9,04E+00 1,03E+02 cytochrome-b5 reductase
activity
44455 1,35E+01 1,53E+02 mitochondrial membrane part
1871 1,38E+01 1,54E+02 pattern binding
6066 1,42E+01 1,56E+02 alcohol metabolism
7148 1,46E+01 1,59E+02 cellular morphogenesis
7519 1,78E+01 1,92E+02 striated muscle development
44265 2,12E+01 2,26E+02 cellular macromolecule
catabolism

Tablas Suplementarias
127
42254 2,17E+01 2,26E+02 ribosome biogenesis and
assembly
16461 2,22E+01 2,26E+02 unconventional myosin
5747 2,22E+01 2,26E+02 respiratory chain complex I
(sensu Eukaryota)
19642 2,22E+01 2,26E+02 anaerobic glycolysis
45271 2,22E+01 2,26E+02 respiratory chain complex I
1558 2,49E+01 2,51E+02 regulation of cell growth
15698 2,90E+01 2,87E+02 inorganic anion transport
1501 2,90E+01 2,87E+02 skeletal development
45182 2,98E+01 2,92E+02 translation regulator activity
42592 3,04E+01 2,94E+02 homeostasis
31406 3,09E+01 2,94E+02 carboxylic acid binding
42255 3,09E+01 2,94E+02 ribosome assembly
6119 3,17E+01 2,99E+02 oxidative phosphorylation
6826 3,54E+01 3,31E+02 iron ion transport
5743 3,68E+01 3,41E+02 mitochondrial inner membrane
7409 3,91E+01 3,60E+02 axonogenesis
30162 4,08E+01 3,66E+02 regulation of proteolysis
5853 4,08E+01 3,66E+02 eukaryotic translation elongation
factor 1 complex
42632 4,08E+01 3,66E+02 cholesterol homeostasis
48812 4,48E+01 3,93E+02 neurite morphogenesis
48667 4,48E+01 3,93E+02 neuron morphogenesis during
differentiation
7028 4,50E+01 3,93E+02 cytoplasm organization and
biogenesis
46916 4,91E+01 4,21E+02 transition metal ion homeostasis
5793 4,91E+01 4,21E+02 ER-Golgi intermediate
compartment
9057 4,98E+01 4,24E+02 macromolecule catabolism
19866 5,06E+01 4,27E+02 organelle inner membrane
904 5,11E+01 4,28E+02 cellular morphogenesis during
differentiation
4459 5,21E+01 4,33E+02 L-lactate dehydrogenase activity
31175 5,44E+01 4,49E+02 neurite development
5730 5,75E+01 4,71E+02 nucleolus
8040 5,97E+01 4,85E+02 axon guidance
3674 6,10E+01 4,91E+02 molecular_function

Tablas Suplementarias
128
Tabla 9.- Tabla con las 319 funciones génicas correspondientes al quinto subcultivo que se
encontraron sobreexpresadas utilizando el programa Cytoscape-BINGO®.
Refernecia Gene Ontology
Valor estadístico p
Valor estadístico p corregido
Funciones Génicas
5830 1,68E-84 1,75E-81 cytosolic ribosome (sensu
Eukaryota)
3740 1,98E-80 1,03E-77 structural constituent of
ribosome
5840 1,70E-78 5,89E-76 ribosome
44445 1,80E-67 4,69E-65 cytosolic part
5198 4,11E-64 8,57E-62 structural molecule activity
30529 6,62E-63 1,15E-60 ribonucleoprotein complex
6416 3,27E-61 4,86E-59 protein biosynthesis
9059 4,99E-59 6,50E-57 macromolecule biosynthesis
16283 4,80E-49 5,00E-47 eukaryotic 48S initiation complex
5843 4,80E-49 5,00E-47 cytosolic small ribosomal subunit
(sensu Eukaryota)
44249 2,27E-46 2,15E-44 cellular biosynthesis
9058 2,01E-43 1,75E-41 biosynthesis
5829 5,26E-42 4,22E-40 cytosol
43234 6,89E-41 5,13E-39 protein complex
16282 3,78E-40 2,63E-38 eukaryotic 43S preinitiation
complex
5737 4,43E-37 2,88E-35 cytoplasm
15935 6,58E-37 4,03E-35 small ribosomal subunit
43232 7,82E-37 4,29E-35 intracellular non-membrane-
bound organelle
43228 7,82E-37 4,29E-35 non-membrane-bound organelle
44444 5,37E-34 2,80E-32 cytoplasmic part
5842 1,94E-30 9,64E-29 cytosolic large ribosomal subunit
(sensu Eukaryota)
15934 5,66E-25 2,68E-23 large ribosomal subunit
44260 1,86E-21 8,43E-20 cellular macromolecule
metabolism
44267 2,34E-21 1,01E-19 cellular protein metabolism
19538 3,36E-20 1,40E-18 protein metabolism
3723 1,67E-17 6,69E-16 RNA binding
43170 1,54E-15 5,95E-14 macromolecule metabolism
44422 2,84E-13 1,02E-11 organelle part
44446 2,84E-13 1,02E-11 intracellular organelle part
44424 5,91E-08 2,05E-06 intracellular part
19843 3,40E-07 1,14E-05 rRNA binding

Tablas Suplementarias
129
5622 1,26E-05 4,09E-04 intracellular
6455 1,89E-05 5,95E-04 translational elongation
7582 2,86E-05 8,77E-04 physiological process
8151 5,78E-05 1,72E-03 cellular physiological process
5578 5,13E-04 1,45E-02 extracellular matrix (sensu
Metazoa)
44237 5,14E-04 1,45E-02 cellular metabolism
43229 9,51E-04 2,57E-02 intracellular organelle
43226 9,61E-04 2,57E-02 organelle
5202 1,47E-03 3,82E-02 collagen
6096 4,66E-03 1,19E-01 glycolysis
43037 5,61E-03 1,39E-01 translation
44238 6,69E-03 1,62E-01 primary metabolism
5576 8,63E-03 2,04E-01 extracellular region
8152 1,17E-02 2,72E-01 metabolism
5575 1,46E-02 3,24E-01 cellular_component
44421 1,46E-02 3,24E-01 extracellular region part
5201 1,62E-02 3,51E-01 extracellular matrix structural
constituent
6007 1,69E-02 3,60E-01 glucose catabolism
31012 3,33E-02 6,95E-01 extracellular matrix
19320 4,52E-02 9,23E-01 hexose catabolism
46365 4,99E-02 1,00E+00 monosaccharide catabolism
46164 5,51E-02 1,08E+00 alcohol catabolism
44275 6,03E-02 1,14E+00 cellular carbohydrate catabolism
16052 6,03E-02 1,14E+00 carbohydrate catabolism
5520 6,67E-02 1,24E+00 insulin-like growth factor binding
8182 8,44E-02 1,54E+00 translation elongation factor
activity
6006 3,29E-01 5,91E+00 glucose metabolism
19883 5,37E-01 9,49E+00 antigen presentation, endogenous antigen
19885 6,24E-01 1,08E+01 antigen processing, endogenous
antigen via MHC class I
6092 7,06E-01 1,21E+01 main pathways of carbohydrate
metabolism
8150 8,32E-01 1,40E+01 biological_process
45182 9,69E-01 1,60E+01 translation regulator activity
5853 1,05E+00 1,71E+01 eukaryotic translation elongation
factor 1 complex
30934 1,56E+00 2,38E+01 anchoring collagen
8043 1,58E+00 2,38E+01 ferritin complex

Tablas Suplementarias
130
30049 1,58E+00 2,38E+01 muscle filament sliding
6123 1,58E+00 2,38E+01 mitochondrial electron
transport, cytochrome c to oxygen
5589 1,58E+00 2,38E+01 collagen type VI
6817 1,61E+00 2,39E+01 phosphate transport
48513 1,69E+00 2,48E+01 organ development
44455 1,80E+00 2,60E+01 mitochondrial membrane part
5583 2,21E+00 3,16E+01 fibrillar collagen
8135 2,87E+00 4,04E+01 translation factor activity, nucleic
acid binding
16477 3,24E+00 4,49E+01 cell migration
19318 3,28E+00 4,49E+01 hexose metabolism
44420 4,13E+00 5,58E+01 extracellular matrix part
5996 4,18E+00 5,58E+01 monosaccharide metabolism
47115 4,70E+00 5,97E+01 trans-1,2-dihydrobenzene-1,2-
diol dehydrogenase activity
8195 4,70E+00 5,97E+01 phosphatidate phosphatase
activity
8354 4,70E+00 5,97E+01 germ cell migration
5584 4,70E+00 5,97E+01 collagen type I
30106 4,85E+00 6,09E+01 MHC class I receptor activity
42612 5,18E+00 6,35E+01 MHC class I protein complex
42611 5,18E+00 6,35E+01 MHC protein complex
9987 5,39E+00 6,53E+01 cellular process
6091 5,97E+00 7,15E+01 generation of precursor metabolites and energy
19838 6,29E+00 7,45E+01 growth factor binding
42775 6,66E+00 7,80E+01 ATP synthesis coupled electron
transport (sensu Eukaryota)
30333 6,84E+00 7,88E+01 antigen processing
1501 6,88E+00 7,88E+01 skeletal development
5746 7,51E+00 8,30E+01 mitochondrial electron transport
chain
40011 7,57E+00 8,30E+01 locomotion
51674 7,57E+00 8,30E+01 localization of cell
6928 7,57E+00 8,30E+01 cell motility
3676 8,10E+00 8,76E+01 nucleic acid binding
15980 8,15E+00 8,76E+01 energy derivation by oxidation of
organic compounds
8147 9,31E+00 9,90E+01 structural constituent of bone

Tablas Suplementarias
131
42773 9,42E+00 9,92E+01 ATP synthesis coupled electron
transport
5518 1,18E+01 1,22E+02 collagen binding
8201 1,35E+01 1,39E+02 heparin binding
7519 1,88E+01 1,92E+02 striated muscle development
1772 2,05E+01 2,08E+02 immunological synapse
19882 2,26E+01 2,21E+02 antigen presentation
16461 2,29E+01 2,21E+02 unconventional myosin
5747 2,29E+01 2,21E+02 respiratory chain complex I
(sensu Eukaryota)
19642 2,29E+01 2,21E+02 anaerobic glycolysis
45271 2,29E+01 2,21E+02 respiratory chain complex I
42254 2,33E+01 2,23E+02 ribosome biogenesis and
assembly
15698 3,15E+01 2,96E+02 inorganic anion transport
31406 3,18E+01 2,96E+02 carboxylic acid binding
42255 3,18E+01 2,96E+02 ribosome assembly
6119 3,38E+01 3,11E+02 oxidative phosphorylation
5743 3,96E+01 3,62E+02 mitochondrial inner membrane
5539 4,17E+01 3,77E+02 glycosaminoglycan binding
30162 4,20E+01 3,78E+02 regulation of proteolysis
40008 4,66E+01 4,15E+02 regulation of growth
30247 4,84E+01 4,24E+02 polysaccharide binding
7028 4,84E+01 4,24E+02 cytoplasm organization and
biogenesis
5793 5,11E+01 4,44E+02 ER-Golgi intermediate
compartment
44464 5,31E+01 4,51E+02 cell part
5623 5,34E+01 4,51E+02 cell
4459 5,36E+01 4,51E+02 L-lactate dehydrogenase activity
8191 5,36E+01 4,51E+02 metalloendopeptidase inhibitor
activity
19866 5,43E+01 4,53E+02 organelle inner membrane

Tablas Suplementarias
132
Tabla 10.- Tabla con las 300 funciones génicas correspondientes al sexto subcultivo que se
encontraron sobreexpresadas utilizando el programa Cytoscape-BINGO®.
Refernecia Gene Ontology
Valor estadístico p
Valor estadístico p corregido
Funciones Génicas
5830 3,29E-80 3,27E-77 cytosolic ribosome (sensu
Eukaryota)
3740 3,79E-79 1,88E-76 structural constituent of
ribosome
5840 2,87E-77 9,51E-75 ribosome
5198 6,04E-64 1,50E-61 structural molecule activity
44445 1,10E-62 2,17E-60 cytosolic part
30529 4,38E-62 7,25E-60 ribonucleoprotein complex
6416 8,50E-61 1,21E-58 protein biosynthesis
9059 1,85E-57 2,30E-55 macromolecule biosynthesis
44249 2,61E-45 2,88E-43 cellular biosynthesis
16283 4,31E-44 3,89E-42 eukaryotic 48S initiation complex
5843 4,31E-44 3,89E-42 cytosolic small ribosomal subunit
(sensu Eukaryota)
9058 1,70E-42 1,40E-40 biosynthesis
43234 2,00E-40 1,53E-38 protein complex
5737 1,51E-38 1,07E-36 cytoplasm
43232 5,94E-38 3,69E-36 intracellular non-membrane-
bound organelle
43228 5,94E-38 3,69E-36 non-membrane-bound organelle
5829 6,31E-38 3,69E-36 cytosol
16282 1,80E-36 9,90E-35 eukaryotic 43S preinitiation
complex
44444 7,02E-34 3,67E-32 cytoplasmic part
15935 1,03E-33 5,13E-32 small ribosomal subunit
5842 7,47E-31 3,53E-29 cytosolic large ribosomal subunit
(sensu Eukaryota)
15934 1,99E-25 8,98E-24 large ribosomal subunit
44260 4,31E-22 1,86E-20 cellular macromolecule
metabolism
44267 5,75E-22 2,38E-20 cellular protein metabolism
19538 6,77E-21 2,69E-19 protein metabolism
3723 2,03E-17 7,76E-16 RNA binding
43170 2,40E-15 8,84E-14 macromolecule metabolism
44422 7,53E-14 2,58E-12 organelle part
44446 7,53E-14 2,58E-12 intracellular organelle part
44424 7,05E-10 2,33E-08 intracellular part
5622 2,01E-07 6,43E-06 intracellular

Tablas Suplementarias
133
19843 2,47E-07 7,66E-06 rRNA binding
8151 4,92E-06 1,48E-04 cellular physiological process
6455 1,31E-05 3,84E-04 translational elongation
7582 2,35E-05 6,68E-04 physiological process
43229 6,23E-05 1,69E-03 intracellular organelle
43226 6,30E-05 1,69E-03 organelle
44237 6,00E-04 1,57E-02 cellular metabolism
5202 1,07E-03 2,74E-02 collagen
5520 1,79E-03 4,43E-02 insulin-like growth factor binding
43037 3,33E-03 8,06E-02 translation
44238 4,25E-03 1,01E-01 primary metabolism
8152 1,18E-02 2,72E-01 metabolism
5575 2,04E-02 4,60E-01 cellular_component
5578 4,62E-02 1,02E+00 extracellular matrix (sensu
Metazoa)
8182 6,76E-02 1,46E+00 translation elongation factor
activity
5201 1,07E-01 2,27E+00 extracellular matrix structural
constituent
5576 1,90E-01 3,92E+00 extracellular region
19883 4,32E-01 8,76E+00 antigen presentation, endogenous antigen
19838 4,99E-01 9,78E+00 growth factor binding
19885 5,02E-01 9,78E+00 antigen processing, endogenous
antigen via MHC class I
45182 7,05E-01 1,35E+01 translation regulator activity
5853 9,19E-01 1,72E+01 eukaryotic translation elongation
factor 1 complex
8150 9,61E-01 1,77E+01 biological_process
6817 1,21E+00 2,19E+01 phosphate transport
31012 1,24E+00 2,19E+01 extracellular matrix
9987 1,28E+00 2,23E+01 cellular process
30934 1,37E+00 2,27E+01 anchoring collagen
44455 1,40E+00 2,27E+01 mitochondrial membrane part
8043 1,44E+00 2,27E+01 ferritin complex
30049 1,44E+00 2,27E+01 muscle filament sliding
6123 1,44E+00 2,27E+01 mitochondrial electron
transport, cytochrome c to oxygen
5589 1,44E+00 2,27E+01 collagen type VI
5583 1,93E+00 3,00E+01 fibrillar collagen
44421 2,01E+00 3,07E+01 extracellular region part
8135 2,17E+00 3,27E+01 translation factor activity, nucleic
acid binding

Tablas Suplementarias
134
16477 2,45E+00 3,63E+01 cell migration
48513 2,91E+00 4,25E+01 organ development
30106 3,94E+00 5,66E+01 MHC class I receptor activity
47115 4,29E+00 5,77E+01 trans-1,2-dihydrobenzene-1,2-
diol dehydrogenase activity
8195 4,29E+00 5,77E+01 phosphatidate phosphatase
activity
8354 4,29E+00 5,77E+01 germ cell migration
5584 4,29E+00 5,77E+01 collagen type I
42612 4,36E+00 5,77E+01 MHC class I protein complex
42611 4,36E+00 5,77E+01 MHC protein complex
30333 5,57E+00 7,15E+01 antigen processing
6096 5,57E+00 7,15E+01 glycolysis
42775 5,62E+00 7,15E+01 ATP synthesis coupled electron
transport (sensu Eukaryota)
5743 5,90E+00 7,42E+01 mitochondrial inner membrane
5746 6,33E+00 7,86E+01 mitochondrial electron transport
chain
40008 7,19E+00 8,81E+01 regulation of growth
3676 7,33E+00 8,87E+01 nucleic acid binding
42773 7,95E+00 9,51E+01 ATP synthesis coupled electron
transport
8147 8,51E+00 1,01E+02 structural constituent of bone
19866 8,69E+00 1,01E+02 organelle inner membrane
5518 1,03E+01 1,19E+02 collagen binding
8201 1,10E+01 1,26E+02 heparin binding
6007 1,18E+01 1,33E+02 glucose catabolism
44420 1,37E+01 1,53E+02 extracellular matrix part
8143 1,41E+01 1,55E+02 poly(A) binding
44275 1,53E+01 1,65E+02 cellular carbohydrate catabolism
16052 1,53E+01 1,65E+02 carbohydrate catabolism
7519 1,59E+01 1,70E+02 striated muscle development
1772 1,74E+01 1,84E+02 immunological synapse
19882 1,86E+01 1,92E+02 antigen presentation
42254 1,86E+01 1,92E+02 ribosome biogenesis and
assembly
40011 2,08E+01 2,00E+02 locomotion
51674 2,08E+01 2,00E+02 localization of cell
6928 2,08E+01 2,00E+02 cell motility
19320 2,09E+01 2,00E+02 hexose catabolism
16461 2,09E+01 2,00E+02 unconventional myosin
5747 2,09E+01 2,00E+02 respiratory chain complex I
(sensu Eukaryota)

Tablas Suplementarias
135
19642 2,09E+01 2,00E+02 anaerobic glycolysis
45271 2,09E+01 2,00E+02 respiratory chain complex I
1558 2,14E+01 2,02E+02 regulation of cell growth
31966 2,19E+01 2,05E+02 mitochondrial membrane
46365 2,22E+01 2,06E+02 monosaccharide catabolism
46164 2,35E+01 2,16E+02 alcohol catabolism
15698 2,45E+01 2,21E+02 inorganic anion transport
1501 2,45E+01 2,21E+02 skeletal development
6119 2,78E+01 2,48E+02 oxidative phosphorylation
31406 2,91E+01 2,56E+02 carboxylic acid binding
42255 2,91E+01 2,56E+02 ribosome assembly
5539 3,43E+01 2,99E+02 glycosaminoglycan binding
44464 3,52E+01 3,03E+02 cell part
5623 3,54E+01 3,03E+02 cell
3729 3,64E+01 3,09E+02 mRNA binding
30162 3,85E+01 3,24E+02 regulation of proteolysis
7028 3,88E+01 3,24E+02 cytoplasm organization and
biogenesis
30247 3,99E+01 3,30E+02 polysaccharide binding
7148 4,09E+01 3,36E+02 cellular morphogenesis
5740 4,40E+01 3,58E+02 mitochondrial envelope
8283 4,49E+01 3,61E+02 cell proliferation
46916 4,50E+01 3,61E+02 transition metal ion homeostasis
31967 4,75E+01 3,78E+02 organelle envelope
4459 4,91E+01 3,87E+02 L-lactate dehydrogenase activity
5730 5,06E+01 3,96E+02 nucleolus
31975 5,11E+01 3,97E+02 envelope
4457 6,08E+01 4,65E+02 lactate dehydrogenase activity
5770 6,08E+01 4,65E+02 late endosome
40007 6,17E+01 4,68E+02 growth
6820 6,55E+01 4,85E+02 anion transport
5507 6,58E+01 4,85E+02 copper ion binding
6006 6,60E+01 4,85E+02 glucose metabolism
1871 6,60E+01 4,85E+02 pattern binding

Tablas Suplementarias
136
Tabla 11.- Tabla con las 289 funciones génicas correspondientes al séptimo subcultivo que se
encontraron sobreexpresadas utilizando el programa Cytoscape-BINGO®.
Refernecia Gene Ontology
Valor estadístico p
Valor estadístico p corregido
Funciones Génicas
5830 6,17E-78 5,96E-75 cytosolic ribosome (sensu
Eukaryota)
3740 1,46E-76 7,05E-74 structural constituent of
ribosome
5840 9,75E-75 3,14E-72 ribosome
5198 4,72E-66 1,14E-63 structural molecule activity
44445 4,57E-63 8,84E-61 cytosolic part
30529 5,44E-60 8,76E-58 ribonucleoprotein complex
6416 5,90E-59 8,14E-57 protein biosynthesis
9059 6,12E-57 7,40E-55 macromolecule biosynthesis
16283 5,02E-47 4,85E-45 eukaryotic 48S initiation complex
5843 5,02E-47 4,85E-45 cytosolic small ribosomal subunit
(sensu Eukaryota)
44249 6,70E-44 5,89E-42 cellular biosynthesis
9058 3,56E-41 2,87E-39 biosynthesis
16282 8,40E-39 6,25E-37 eukaryotic 43S preinitiation
complex
5829 2,64E-38 1,82E-36 cytosol
5737 7,04E-38 4,54E-36 cytoplasm
43234 1,59E-36 9,57E-35 protein complex
15935 8,01E-36 4,11E-34 small ribosomal subunit
43232 8,08E-36 4,11E-34 intracellular non-membrane-
bound organelle
43228 8,08E-36 4,11E-34 non-membrane-bound organelle
44444 5,40E-33 2,61E-31 cytoplasmic part
5842 4,16E-26 1,91E-24 cytosolic large ribosomal subunit
(sensu Eukaryota)
44260 8,67E-23 3,81E-21 cellular macromolecule
metabolism
44267 1,18E-22 4,96E-21 cellular protein metabolism
15934 6,02E-22 2,42E-20 large ribosomal subunit
19538 6,48E-21 2,50E-19 protein metabolism
43170 1,16E-16 4,31E-15 macromolecule metabolism
3723 6,76E-16 2,42E-14 RNA binding
44422 1,25E-13 4,17E-12 organelle part
44446 1,25E-13 4,17E-12 intracellular organelle part
44424 2,93E-09 9,43E-08 intracellular part
19843 2,21E-07 6,88E-06 rRNA binding

Tablas Suplementarias
137
5622 6,83E-07 2,06E-05 intracellular
8151 1,38E-06 4,04E-05 cellular physiological process
6455 1,16E-05 3,30E-04 translational elongation
7582 1,47E-05 4,07E-04 physiological process
5578 3,17E-05 8,50E-04 extracellular matrix (sensu
Metazoa)
5202 3,61E-05 9,42E-04 collagen
43229 2,26E-04 5,66E-03 intracellular organelle
43226 2,28E-04 5,66E-03 organelle
44237 8,23E-04 1,99E-02 cellular metabolism
5201 8,71E-04 2,05E-02 extracellular matrix structural
constituent
43037 2,78E-03 6,39E-02 translation
31012 3,02E-03 6,78E-02 extracellular matrix
44238 5,97E-03 1,31E-01 primary metabolism
5575 1,11E-02 2,38E-01 cellular_component
8152 1,50E-02 3,15E-01 metabolism
5576 1,77E-02 3,64E-01 extracellular region
5583 3,79E-02 7,63E-01 fibrillar collagen
6096 4,16E-02 8,21E-01 glycolysis
5520 4,95E-02 9,56E-01 insulin-like growth factor binding
44421 5,57E-02 1,05E+00 extracellular region part
8182 6,26E-02 1,16E+00 translation elongation factor
activity
6007 1,27E-01 2,31E+00 glucose catabolism
6817 1,33E-01 2,38E+00 phosphate transport
9987 1,98E-01 3,48E+00 cellular process
48513 2,33E-01 4,01E+00 organ development
19320 2,95E-01 4,99E+00 hexose catabolism
44275 3,05E-01 4,99E+00 cellular carbohydrate catabolism
16052 3,05E-01 4,99E+00 carbohydrate catabolism
46365 3,22E-01 5,18E+00 monosaccharide catabolism
46164 3,50E-01 5,55E+00 alcohol catabolism
44420 4,99E-01 7,78E+00 extracellular matrix part
45182 6,32E-01 9,68E+00 translation regulator activity
1501 8,32E-01 1,26E+01 skeletal development
5853 8,77E-01 1,30E+01 eukaryotic translation elongation
factor 1 complex
40011 1,07E+00 1,52E+01 locomotion
51674 1,07E+00 1,52E+01 localization of cell
6928 1,07E+00 1,52E+01 cell motility
30934 1,30E+00 1,82E+01 anchoring collagen
8150 1,36E+00 1,82E+01 biological_process

Tablas Suplementarias
138
8043 1,40E+00 1,82E+01 ferritin complex
30049 1,40E+00 1,82E+01 muscle filament sliding
6123 1,40E+00 1,82E+01 mitochondrial electron
transport, cytochrome c to oxygen
5589 1,40E+00 1,82E+01 collagen type VI
6006 1,64E+00 2,11E+01 glucose metabolism
8135 1,97E+00 2,51E+01 translation factor activity, nucleic
acid binding
16477 2,23E+00 2,80E+01 cell migration
6092 3,16E+00 3,91E+01 main pathways of carbohydrate
metabolism
8195 4,16E+00 4,96E+01 phosphatidate phosphatase
activity
8354 4,16E+00 4,96E+01 germ cell migration
5584 4,16E+00 4,96E+01 collagen type I
15698 4,59E+00 5,41E+01 inorganic anion transport
19838 4,77E+00 5,55E+01 growth factor binding
42775 5,29E+00 6,09E+01 ATP synthesis coupled electron
transport (sensu Eukaryota)
5746 5,97E+00 6,78E+01 mitochondrial electron transport
chain
42773 7,50E+00 8,42E+01 ATP synthesis coupled electron
transport
8147 8,25E+00 9,16E+01 structural constituent of bone
44464 9,83E+00 1,07E+02 cell part
5623 9,89E+00 1,07E+02 cell
8201 1,03E+01 1,10E+02 heparin binding
44455 1,10E+01 1,17E+02 mitochondrial membrane part
19318 1,18E+01 1,24E+02 hexose metabolism
5996 1,46E+01 1,51E+02 monosaccharide metabolism
6820 1,48E+01 1,51E+02 anion transport
3676 1,49E+01 1,51E+02 nucleic acid binding
7519 1,50E+01 1,51E+02 striated muscle development
42254 1,72E+01 1,71E+02 ribosome biogenesis and
assembly
16461 2,03E+01 1,94E+02 unconventional myosin
5747 2,03E+01 1,94E+02 respiratory chain complex I
(sensu Eukaryota)
19642 2,03E+01 1,94E+02 anaerobic glycolysis
45271 2,03E+01 1,94E+02 respiratory chain complex I
7275 2,10E+01 1,99E+02 development

Tablas Suplementarias
139
15980 2,59E+01 2,43E+02 energy derivation by oxidation of
organic compounds
42255 2,82E+01 2,62E+02 ribosome assembly
5539 3,21E+01 2,95E+02 glycosaminoglycan binding
40008 3,45E+01 3,15E+02 regulation of growth
7028 3,60E+01 3,25E+02 cytoplasm organization and
biogenesis
30162 3,73E+01 3,28E+02 regulation of proteolysis
42632 3,73E+01 3,28E+02 cholesterol homeostasis
30247 3,73E+01 3,28E+02 polysaccharide binding
5793 4,31E+01 3,75E+02 ER-Golgi intermediate
compartment
5730 4,74E+01 4,03E+02 nucleolus
4459 4,76E+01 4,03E+02 L-lactate dehydrogenase activity
8191 4,76E+01 4,03E+02 metalloendopeptidase inhibitor
activity
6091 5,26E+01 4,42E+02 generation of precursor metabolites and energy
4457 5,90E+01 4,87E+02 lactate dehydrogenase activity
5770 5,90E+01 4,87E+02 late endosome

Tablas Suplementarias
140
Tabla 12.- Tabla con las 252 funciones génicas correspondientes al octavo subcultivo que se
encontraron sobreexpresadas utilizando el programa Cytoscape-BINGO®.
Refernecia Gene Ontology
Valor estadístico p
Valor estadístico p corregido
Funciones Génicas
3740 4,23E-81 3,25E-78 structural constituent of
ribosome
5840 2,72E-79 1,05E-76 ribosome
5830 4,42E-76 1,13E-73 cytosolic ribosome (sensu
Eukaryota)
5198 3,81E-67 7,33E-65 structural molecule activity
30529 1,22E-64 1,88E-62 ribonucleoprotein complex
6416 4,27E-63 5,48E-61 protein biosynthesis
44445 1,33E-60 1,47E-58 cytosolic part
9059 6,98E-60 6,71E-58 macromolecule biosynthesis
44249 3,07E-47 2,63E-45 cellular biosynthesis
9058 1,45E-43 1,11E-41 biosynthesis
16283 1,94E-43 1,24E-41 eukaryotic 48S initiation complex
5843 1,94E-43 1,24E-41 cytosolic small ribosomal subunit
(sensu Eukaryota)
43232 1,86E-41 1,02E-39 intracellular non-membrane-
bound organelle
43228 1,86E-41 1,02E-39 non-membrane-bound organelle
5737 7,56E-41 3,88E-39 cytoplasm
43234 8,52E-38 4,10E-36 protein complex
5829 1,79E-36 8,13E-35 cytosol
16282 2,37E-36 1,01E-34 eukaryotic 43S preinitiation
complex
15935 7,04E-34 2,85E-32 small ribosomal subunit
44444 1,88E-33 7,26E-32 cytoplasmic part
5842 2,59E-27 9,51E-26 cytosolic large ribosomal subunit
(sensu Eukaryota)
44267 1,60E-25 5,59E-24 cellular protein metabolism
44260 5,89E-25 1,97E-23 cellular macromolecule
metabolism
15934 2,81E-23 9,01E-22 large ribosomal subunit
19538 3,24E-23 9,97E-22 protein metabolism
3723 2,08E-18 6,17E-17 RNA binding
43170 1,23E-16 3,51E-15 macromolecule metabolism
44422 3,96E-14 1,05E-12 organelle part
44446 3,96E-14 1,05E-12 intracellular organelle part
44424 2,50E-13 6,42E-12 intracellular part
5622 7,07E-11 1,76E-09 intracellular

Tablas Suplementarias
141
19843 7,80E-08 1,88E-06 rRNA binding
43229 1,02E-07 2,34E-06 intracellular organelle
43226 1,03E-07 2,34E-06 organelle
8151 1,40E-06 3,07E-05 cellular physiological process
6455 1,33E-04 2,84E-03 translational elongation
44237 3,44E-04 7,12E-03 cellular metabolism
5202 3,51E-04 7,12E-03 collagen
7582 6,39E-04 1,26E-02 physiological process
44238 1,79E-03 3,45E-02 primary metabolism
43037 3,94E-03 7,39E-02 translation
8152 4,39E-03 8,06E-02 metabolism
5578 3,85E-02 6,89E-01 extracellular matrix (sensu
Metazoa)
5575 2,14E-01 3,74E+00 cellular_component
5201 3,26E-01 5,58E+00 extracellular matrix structural
constituent
6817 4,35E-01 7,27E+00 phosphate transport
3676 4,69E-01 7,69E+00 nucleic acid binding
9987 5,00E-01 8,03E+00 cellular process
8182 8,02E-01 1,26E+01 translation elongation factor
activity
30934 8,43E-01 1,30E+01 anchoring collagen
31012 8,81E-01 1,33E+01 extracellular matrix
8043 1,04E+00 1,46E+01 ferritin complex
30049 1,04E+00 1,46E+01 muscle filament sliding
6123 1,04E+00 1,46E+01 mitochondrial electron
transport, cytochrome c to oxygen
5589 1,04E+00 1,46E+01 collagen type VI
5583 1,20E+00 1,64E+01 fibrillar collagen
48513 1,35E+00 1,83E+01 organ development
45182 1,69E+00 2,25E+01 translation regulator activity
42775 3,03E+00 3,96E+01 ATP synthesis coupled electron
transport (sensu Eukaryota)
5584 3,10E+00 3,97E+01 collagen type I
5746 3,42E+00 4,32E+01 mitochondrial electron transport
chain
42773 4,31E+00 5,35E+01 ATP synthesis coupled electron
transport
44420 4,76E+00 5,81E+01 extracellular matrix part
44464 5,14E+00 6,12E+01 cell part
5623 5,17E+00 6,12E+01 cell
8135 5,94E+00 6,93E+01 translation factor activity, nucleic
acid binding

Tablas Suplementarias
142
8147 6,15E+00 7,07E+01 structural constituent of bone
16477 6,59E+00 7,22E+01 cell migration
40011 6,66E+00 7,22E+01 locomotion
51674 6,66E+00 7,22E+01 localization of cell
6928 6,66E+00 7,22E+01 cell motility
42254 8,05E+00 8,61E+01 ribosome biogenesis and
assembly
7519 8,72E+00 9,19E+01 striated muscle development
15698 9,62E+00 1,00E+02 inorganic anion transport
8150 1,08E+01 1,11E+02 biological_process
16461 1,52E+01 1,48E+02 unconventional myosin
5747 1,52E+01 1,48E+02 respiratory chain complex I
(sensu Eukaryota)
19642 1,52E+01 1,48E+02 anaerobic glycolysis
45271 1,52E+01 1,48E+02 respiratory chain complex I
7028 1,73E+01 1,66E+02 cytoplasm organization and
biogenesis
42255 2,11E+01 2,01E+02 ribosome assembly
5730 2,53E+01 2,37E+02 nucleolus
6096 2,56E+01 2,38E+02 glycolysis
6820 2,70E+01 2,47E+02 anion transport
30162 2,79E+01 2,50E+02 regulation of proteolysis
5853 2,79E+01 2,50E+02 eukaryotic translation elongation
factor 1 complex
46916 2,85E+01 2,52E+02 transition metal ion homeostasis
3674 3,36E+01 2,94E+02 molecular_function
4459 3,57E+01 3,09E+02 L-lactate dehydrogenase activity
5507 3,71E+01 3,17E+02 copper ion binding
5576 3,95E+01 3,27E+02 extracellular region
42592 3,95E+01 3,27E+02 homeostasis
7517 3,95E+01 3,27E+02 muscle development
44421 4,10E+01 3,36E+02 extracellular region part
8201 4,40E+01 3,55E+02 heparin binding
4457 4,43E+01 3,55E+02 lactate dehydrogenase activity
44455 4,64E+01 3,65E+02 mitochondrial membrane part
6007 4,64E+01 3,65E+02 glucose catabolism
1501 4,75E+01 3,69E+02 skeletal development

Tablas Suplementarias
143
Tabla 13.- Correlación genes/secuencias expresadas (EST) con el número de subcultivo. Se
objetiva la existencia de 489 genes/secuencias expresadas (EST) que se correlacionan de
forma directa con el subcultivo celular (R> 0.8), y de 192 genes/secuencia expresadas (EST)
que se correlacionan de forma inversa con el subcultivo celular(R < -0.8).
Referen-cia
Affymetrix
1er
Subcultivo
2º Subcul
tivo
3 er
Subcultivo
4º Subcultivo
5º Subcultivo
6º Subcultivo
7º Subcultivo
8º Subcultivo
Pearson r
Símbolo Gen
Nombre del gen
241140_at 5,1 10,4 13,3 21,9 20,1 25,3 29,9 32,7 0,9833 LMO7 LIM domain 7
203637_s_at 200,5 182,2 218,2 249,8 289,1 338,5 356,0 356,3 0,9675 MID1
midline 1 (Opitz/BBB syndrome)
215501_s_at 110,0 111,1 134,6 125,7 161,8 174,2 193,0 222,7 0,9668 DUSP10
dual specificity phosphatase 10
241403_at 8,5 16,4 18,9 19,2 27,7 34,3 41,9 38,0 0,9665 CLK4 CDC-like kinase 4
209793_at 19,6 21,8 42,8 49,7 60,4 86,6 78,4 85,6 0,9642 GRIA1 glutamate receptor, ionotropic, AMPA 1
217371_s_at
12,2 33,0 31,5 42,1 56,5 58,9 94,0 88,4 0,9623 IL15 interleukin 15
213746_s_at 779,3 1061,3
1049,1
1224,3
1135,1
1422,9
1644,8
1784,5
0,9587 FLNA filamin A, alpha (actin binding protein 280)
229227_at 5,4 8,3 9,0 12,4 15,6 16,8 21,0 17,8 0,9550 FLJ4524
4
hypothetical locus FLJ45244
227074_at 79,8 188,3 229,4 251,8 303,0 299,2 402,8 372,1 0,9539 LOC100131564
hypothetical protein LOC100131564
201206_s_at 294,7 316,0 362,9 458,9 446,7 559,7 527,9 550,0 0,9523 RRBP1
ribosome binding protein 1 homolog
180kDa (dog)
236321_at 53,8 82,2 120,8 115,0 138,8 133,4 150,6 180,3 0,9520 --- ---
232297_at 123,8 177,2 137,4 175,1 223,9 241,4 261,6 295,5 0,9513 --- ---
240721_at 18,4 24,0 26,2 45,5 48,4 50,2 50,2 59,3 0,9511 --- ---
219468_s_at 40,6 41,5 46,9 92,1 95,2 104,3 146,3 128,3 0,9471
CUEDC1
CUE domain containing 1
203139_at 81,0 442,0 346,8 418,2 548,0 651,3 964,5 958,0 0,9455 DAPK1 death-associated protein kinase 1
227243_s_at
31,8 41,0 46,9 44,9 92,0 86,9 94,6 108,4 0,9447 EBF3 early B-cell factor 3
226847_at 116,8 195,6 227,0 203,7 269,4 279,5 310,7 311,5 0,9446 FST follistatin
231828_at 36,3 53,8 113,6 115,6 181,2 177,4 195,1 189,4 0,9445 --- ---
207233_s_at 131,0 130,4 133,4 132,9 169,8 174,1 191,6 208,7 0,9427 MITF
microphthalmia-associated
transcription factor
51192_at 59,3 82,0 77,0 91,2 96,1 95,4 104,1 108,8 0,9426 SSH3 slingshot homolog 3
(Drosophila)
223347_at 124,7 140,8 159,1 149,2 175,6 181,3 212,3 195,3 0,9421 MUM1 melanoma
associated antigen (mutated) 1 214604_at 8,5 5,7 22,2 27,8 44,2 41,3 52,0 46,8 0,9418 HOXD11 homeobox D11
233323_at 15,1 17,3 16,9 17,6 27,7 29,4 28,9 33,4 0,9413 --- ---
209890_at 99,2 127,9 113,6 162,1 194,6 169,4 208,4 258,4 0,9404 TSPAN5 tetraspanin 5
236494_x_at
42,0 39,4 36,7 60,0 84,2 90,7 109,0 100,4 0,9387 --- ---
230269_at 1,7 1,8 4,8 4,2 7,3 6,4 6,8 9,7 0,9384 --- ---
240690_at 23,0 9,0 29,0 37,9 36,3 54,5 55,6 65,1 0,9370 --- ---
229400_at 37,7 68,5 61,1 77,8 232,3 200,5 313,4 304,4 0,9366 HOXD10 homeobox D10
219523_s_at 33,6 11,6 55,5 57,4 60,5 82,6 117,3 143,6 0,9355 ODZ3
odz, odd Oz/ten-m homolog 3
(Drosophila)
1553129_at 26,0 28,5 46,3 31,5 49,3 55,1 70,2 82,0 0,9339 SVEP1
sushi, von Willebrand factor type A, EGF and
pentraxin domain containing 1

Tablas Suplementarias
144
229190_at 26,4 27,7 36,4 33,2 41,5 50,5 72,7 68,2 0,9337 --- ---
210757_x_at 523,6 505,3 578,1 569,1 590,6 605,6 710,0 720,3 0,9325 DAB2
disabled homolog 2, mitogen-responsive
phosphoprotein (Drosophila)
237773_at 8,9 1,6 36,8 45,8 53,1 68,7 65,3 68,8 0,9319 --- ---
200906_s_at 79,7 40,4 122,4 168,8 203,0 201,3 200,9 262,2 0,9315 PALLD
palladin, cytoskeletal associated protein
208790_s_at 226,3 261,9 305,8 423,1 370,0 370,2 528,1 548,8 0,9306 PTRF
polymerase I and transcript release
factor
214098_at 17,4 16,7 18,5 20,5 22,1 22,9 24,7 31,5 0,9304 KIAA11
07 KIAA1107
227565_at 83,4 146,3 92,0 142,2 191,1 186,8 239,3 277,5 0,9302 --- ---
202719_s_at 78,7 78,7 106,8 127,3 142,1 156,9 183,5 152,5 0,9290 TES
testis derived transcript (3 LIM
domains)
224704_at 156,4 176,9 161,0 187,6 248,4 231,5 289,3 273,6 0,9289 TNRC6A trinucleotide repeat
containing 6A
237317_at 14,0 8,4 15,0 18,8 23,8 22,1 27,4 28,2 0,9269 --- ---
212390_at 199,1 331,6 331,2 338,2 393,5 369,8 441,0 505,1 0,9264 PDE4DI
P
phosphodiesterase 4D interacting
protein (myomegalin) ///
similar to phosphodiesterase
4D interacting protein
(myomegalin)
210305_at 62,1 88,2 103,4 108,8 124,9 127,0 180,4 243,0 0,9257 PDE4DI
P
phosphodiesterase 4D interacting
protein (myomegalin)
1553260_s_at 2,9 0,5 5,1 6,9 4,7 10,2 11,8 13,2 0,9254
ALS2CR11
amyotrophic lateral sclerosis 2 (juvenile) chromosome region,
candidate 11
204004_at 141,6 254,0 248,6 284,0 333,6 331,1 492,8 420,0 0,9246 PAWR PRKC, apoptosis,
WT1, regulator
229238_at 55,1 73,6 76,9 84,5 81,6 107,4 101,6 102,3 0,9242 LOC400
566
hypothetical gene supported by
AK128660
230137_at 71,3 420,5 367,7 396,7 666,1 611,1 847,6 763,1 0,9220 TMEM15
5 transmembrane
protein 155
236344_at 27,3 20,0 51,8 75,7 86,0 119,1 101,9 104,8 0,9219 PDE1C
phosphodiesterase
1C, calmodulin-dependent 70kDa
204497_at 132,1 183,6 180,9 210,4 199,8 215,5 267,1 247,4 0,9216 ADCY9 adenylate cyclase 9
1566825_at
15,2 8,3 20,2 18,0 22,7 25,3 36,5 39,5 0,9208 --- ---
243688_at 6,6 16,3 33,0 36,5 46,9 31,4 70,5 77,4 0,9204 LOC285
431 hypothetical protein
LOC285431
227846_at 179,3 198,7 224,6 416,1 330,5 402,9 533,5 478,7 0,9192 GPR176 G protein-coupled
receptor 176
224926_at 236,3 281,9 294,6 274,4 297,9 308,5 329,7 338,5 0,9188 EXOC4 exocyst complex
component 4
222457_s_at 375,6 247,0 344,6 431,5 752,4 677,1
1057,0
983,5 0,9188 LIMA1 LIM domain and actin binding 1

Tablas Suplementarias
145
210959_s_at 85,8 78,1 102,5 87,2 102,2 130,2 143,8 145,4 0,9169 SRD5A1
steroid-5-alpha-reductase, alpha
polypeptide 1 (3-oxo-5 alpha-steroid delta
4-dehydrogenase alpha 1)
218468_s_at 179,0 1094,3 884,0
1064,9
1117,9
2041,9
2346,4
3638,0
0,9167 GREM1
gremlin 1, cysteine knot superfamily,
homolog (Xenopus laevis)
204005_s_at 58,0 112,2 94,5 115,2 138,2 121,3 194,4 214,4 0,9155 PAWR
PRKC, apoptosis, WT1, regulator
236987_at 11,3 24,5 25,8 40,3 29,6 53,9 44,4 59,3 0,9152 --- ---
202305_s_at 302,8 557,1 600,7 802,2 657,9 782,5
1028,8
946,7 0,9152 FEZ2 fasciculation and elongation protein
zeta 2 (zygin II)
229518_at 82,6 167,3 124,9 147,1 181,3 365,7 386,5 539,6 0,9143 FAM46B family with sequence
similarity 46, member B 216859_x_
at 18,1 21,5 23,1 17,8 35,8 45,2 46,1 46,7 0,9121 --- ---
239409_at 19,6 14,5 18,2 35,4 30,0 39,1 49,2 43,7 0,9114 --- ---
240351_at 2,2 3,9 7,4 10,2 10,5 17,1 22,4 15,3 0,9104 --- ---
241726_at 26,2 18,2 43,3 44,2 62,1 52,5 65,9 65,0 0,9091 --- ---
233866_at 9,0 9,5 3,2 16,5 17,4 24,4 26,6 28,3 0,9089 KLHL5 kelch-like 5 (Drosophila)
204493_at 42,2 53,7 62,1 69,6 65,4 76,6 68,2 87,9 0,9088 BID BH3 interacting domain death
agonist
1558017_s_at
6,0 13,1 20,2 22,3 31,3 23,0 39,6 33,5 0,9080 --- ---
242789_at 50,3 16,5 66,2 71,3 90,6 134,2 143,0 126,4 0,9077 PDE1A phosphodiesterase
1A, calmodulin-dependent
203695_s_at 121,9 141,2 138,3 131,4 190,7 237,9 311,8 410,3 0,9072 DFNA5
deafness, autosomal dominant 5
225372_at 9,4 43,4 45,8 36,6 63,5 62,8 73,2 73,9 0,9062 C10orf5
4
chromosome 10 open reading frame
54
226971_at 9,9 7,8 13,8 20,0 19,8 19,3 22,0 22,4 0,9061 CCDC13
6 coiled-coil domain
containing 136
235019_at 137,7 270,9 395,1 535,6 539,9 489,7 1180,
8 1081,
2 0,9061 CPM carboxypeptidase M
228847_at 15,7 20,2 23,3 18,4 27,9 25,6 29,6 32,0 0,9057 EXOC3 exocyst complex
component 3
219552_at 55,7 59,9 52,0 62,5 87,1 92,4 149,7 151,7 0,9050 SVEP1
sushi, von Willebrand factor type A, EGF and
pentraxin domain containing 1
238192_at 8,7 13,9 11,6 11,4 19,8 16,8 22,1 27,3 0,9047 --- ---
228707_at 65,6 88,6 65,4 119,8 144,7 169,1 162,0 155,8 0,9038 CLDN23 claudin 23
1570048_at 3,2 4,4 10,1 14,4 11,2 13,8 16,3 15,6 0,9036 DPH4
DPH4, JJJ3 homolog (S. cerevisiae)
212686_at 36,7 61,6 56,5 61,9 64,3 72,4 104,2 93,2 0,9034 PPM1H protein phosphatase
1H (PP2C domain containing)
215576_at 13,8 10,0 14,1 17,1 20,0 23,2 21,0 22,8 0,9025 --- ---
226614_s_at 40,2 69,5 107,2 131,7 87,6 146,5 163,2 162,9 0,9024 C8orf13
chromosome 8 open reading frame 13
244055_at 8,5 9,7 5,2 15,3 17,0 31,8 30,7 29,6 0,9022 --- ---
203819_s_at 5,8 22,9 16,6 32,3 34,1 37,8 84,5 108,9 0,9020
IGF2BP3
insulin-like growth factor 2 mRNA
binding protein 3
1557685_at 9,3 20,4 10,6 23,0 36,2 32,9 37,9 38,0 0,9014 C4orf38
chromosome 4 open reading frame 38

Tablas Suplementarias
146
207980_s_at 546,3 673,4 840,9
1049,3
955,0 989,4 991,3 1197,
1 0,9007 CITED2
Cbp/p300-interacting transactivator, with
Glu/Asp-rich carboxy-terminal
domain, 2
236848_s_at 1,3 0,8 3,9 2,5 4,3 5,0 6,0 5,3 0,9007 TEX13A testis expressed 13A
215163_at 3,8 1,0 5,4 2,2 6,8 12,0 13,9 19,1 0,9001 --- ---
228462_at 10,7 16,7 16,8 17,0 40,3 32,5 72,5 64,9 0,8997 IRX2 iroquois homeobox 2
203157_s_at
91,2 107,4 175,0 135,0 159,6 212,4 219,1 207,3 0,8994 GLS glutaminase
233313_at 11,4 13,2 12,5 13,8 17,0 25,5 34,1 28,7 0,8993 HDAC7 Histone deacetylase
7
1564424_at
22,8 22,3 27,3 28,0 50,8 35,6 60,7 62,6 0,8992 --- ---
1556216_s_at 15,0 21,3 23,5 21,2 22,5 23,7 30,8 32,2 0,8992 --- ---
222433_at 1345,0 1190,8 1170,
3 1309,
9 1941,
5 1847,
0 2764,
8 2911,
7 0,8991 ENAH
enabled homolog (Drosophila)
219014_at 14,7 12,5 16,1 20,9 28,3 43,4 38,4 72,1 0,8991 PLAC8 placenta-specific 8
239842_x_at
36,3 24,5 44,2 64,0 78,8 56,6 97,8 95,0 0,8987 --- ---
203988_s_at 126,9 155,6 106,3 153,1 182,4 220,0 267,1 249,9 0,8984 FUT8
fucosyltransferase 8 (alpha (1,6)
fucosyltransferase)
1558345_a_at 18,6 28,5 18,9 22,1 33,1 40,4 41,8 45,4 0,8977
LOC439911
hypothetical gene supported by NM_194304
238769_at 20,1 18,1 25,7 32,4 57,2 50,9 48,9 55,0 0,8974 --- ---
213748_at 28,2 40,4 40,0 47,3 55,6 74,9 78,2 62,9 0,8974 TRIM66 tripartite motif-containing 66
233547_x_at 132,0 206,9 259,5 466,6 500,2 483,9 566,1 488,0 0,8971 PDE1A
phosphodiesterase 1A, calmodulin-
dependent
219718_at 88,4 85,6 97,5 117,9 139,9 126,8 134,0 135,0 0,8968 FGGY FGGY carbohydrate
kinase domain containing
1561956_at
8,4 7,6 8,9 13,6 12,4 13,0 20,6 17,6 0,8967 --- ---
233261_at 11,2 11,9 17,9 20,5 20,7 32,6 45,4 33,0 0,8967 EBF1 Early B-cell factor 1
239218_at 15,1 9,9 12,2 8,1 48,5 48,8 52,5 69,0 0,8953 PDE1C phosphodiesterase
1C, calmodulin-dependent 70kDa
223384_s_at 223,0 242,9 192,4 258,0 325,1 315,4 351,0 351,7 0,8948 TRIM4
tripartite motif-containing 4
1567272_at 1,1 1,2 6,0 6,3 9,1 7,5 8,2 9,4 0,8934 OR2K2
olfactory receptor, family 2, subfamily
K, member 2
1563497_at 22,1 21,5 21,3 25,8 42,4 41,4 44,6 41,4 0,8928 USP25
ubiquitin specific peptidase 25
217820_s_at 1154,9 850,0
1489,9
1125,8
1710,5
1621,1
2206,0
2263,8
0,8924 ENAH enabled homolog
(Drosophila)
224936_at 573,0 673,9 577,4 827,7 902,6 876,0 932,4 918,2 0,8921 EIF2S3
eukaryotic translation initiation factor 2, subunit 3
gamma, 52kDa
226435_at 37,7 49,3 46,4 52,7 72,5 113,0 80,9 130,4 0,8918 PAPLN papilin,
proteoglycan-like sulfated glycoprotein
205510_s_at 35,7 44,9 48,7 50,7 47,3 52,3 72,6 67,1 0,8914
FLJ10038
hypothetical protein FLJ10038
203820_s_at 8,0 63,7 56,1 85,2 78,2 68,0 117,3 153,0 0,8910
IGF2BP3
insulin-like growth factor 2 mRNA
binding protein 3
214594_x_at 71,1 89,5 76,0 80,1 132,5 106,6 141,0 164,8 0,8891 ATP8B1
ATPase, class I, type 8B, member 1

Tablas Suplementarias
147
37590_g_at
47,1 56,2 62,2 81,1 86,7 76,3 131,0 107,5 0,8890 --- ---
229773_at 15,9 14,7 18,9 16,8 34,6 25,2 48,4 56,1 0,8889 SNAP23 Synaptosomal-
associated protein, 23kDa
213313_at 244,2 504,4 371,1 451,6 586,2 669,7 981,1 781,6 0,8889 RABGA
P1 RAB GTPase
activating protein 1
238608_at 8,4 11,9 4,3 19,5 16,7 20,3 23,8 30,5 0,8888 --- ---
201462_at 354,1 424,5 346,9 452,4 432,7 456,3 547,3 592,6 0,8888 SCRN1 secernin 1
1558680_s_at 157,8 35,0 225,6 320,3 401,2 443,5 466,5 420,9 0,8886 PDE1A
phosphodiesterase 1A, calmodulin-
dependent
222463_s_at 151,6 173,2 226,2 219,2 294,6 284,2 286,6 274,3 0,8883 BACE1
beta-site APP-cleaving enzyme 1
1557906_at 3,2 5,5 3,7 11,2 9,5 14,0 13,4 12,6 0,8876 MUC12
mucin 12, cell surface associated
226684_at 145,8 127,2 145,6 151,6 188,1 182,2 257,1 228,5 0,8872 ATG2B ATG2 autophagy
related 2 homolog B (S. cerevisiae)
212117_at 424,9 385,0 393,7 547,5 559,9 604,5 626,1 592,3 0,8869 RHOQ ras homolog gene family, member Q
222434_at 97,1 72,9 114,5 99,5 191,3 138,6 251,1 291,3 0,8862 ENAH enabled homolog
(Drosophila)
238604_at 114,2 111,8 104,1 131,8 136,8 139,3 201,8 197,3 0,8861 --- ---
241936_x_at
22,7 11,0 8,4 24,7 45,7 42,1 90,7 88,9 0,8857 --- ---
218469_at 215,0 1135,1 764,9 952,6 958,0 1558,
2 2386,
1 3397,
6 0,8854 GREM1
gremlin 1, cysteine knot superfamily,
homolog (Xenopus laevis)
221900_at 50,5 32,1 58,5 66,0 70,8 142,0 164,7 132,6 0,8849 COL8A2 collagen, type VIII,
alpha 2
233003_at 15,7 29,1 17,9 40,4 58,2 47,3 49,9 61,3 0,8839 --- ---
233063_s_at
19,6 22,6 24,0 23,8 32,2 31,7 29,4 31,4 0,8833 --- ---
235406_x_at
19,2 28,5 26,9 31,2 32,3 42,9 41,0 37,1 0,8831 --- ---
201669_s_at 1359,6 1856,9
1607,8
2195,7
2331,3
2490,0
2593,6
2349,7
0,8829 MARCK
S
myristoylated alanine-rich protein kinase C substrate
222184_at 32,2 18,1 62,6 65,5 86,9 107,5 132,4 92,7 0,8828 --- ---
244859_at 14,8 19,3 21,3 51,9 38,8 39,0 54,8 56,1 0,8822 --- ---
1560209_at
1,6 1,7 3,5 3,0 1,8 8,9 10,6 11,3 0,8819 --- ---
242875_at 8,9 8,5 8,7 5,5 26,1 31,5 31,3 32,8 0,8817 --- ---
202197_at 87,3 86,2 124,6 117,8 120,1 128,0 141,2 134,0 0,8817 MTMR3 myotubularin related
protein 3
224806_at 332,6 301,2 303,3 447,3 406,6 438,2 492,6 479,7 0,8815 TRIM25 tripartite motif-containing 25
208396_s_at 247,4 406,8 427,4 522,7 944,2 809,9
1128,7
833,5 0,8813 PDE1A phosphodiesterase
1A, calmodulin-dependent
211370_s_at 87,0 126,7 104,6 130,4 174,0 183,0 172,2 171,9 0,8812 MAP2K5
mitogen-activated protein kinase
kinase 5
225964_at 53,5 46,1 53,9 64,3 61,7 77,9 94,8 79,2 0,8809 ZXDC ZXD family zinc
finger C
223638_at 73,3 103,4 110,4 94,2 138,7 114,5 142,6 155,6 0,8801 NBPF3 neuroblastoma
breakpoint family, member 3
238017_at 4,1 6,8 4,8 6,2 5,6 10,4 10,6 13,5 0,8799 RDHE2 epidermal retinal dehydrogenase 2
208690_s_at 384,0 675,5 438,5 676,5 789,5 934,6 787,3
1115,7
0,8798 PDLIM1 PDZ and LIM domain
1 (elfin)
1563414_at
3,5 1,0 5,8 3,1 15,3 12,0 36,1 34,8 0,8796 --- ---
200907_s_at 466,7 663,7 644,4 802,0 711,8 805,4
1446,7
1625,2
0,8791 PALLD palladin, cytoskeletal
associated protein

Tablas Suplementarias
148
210778_s_at 141,3 126,2 98,2 161,7 181,6 181,6 210,3 243,0 0,8788 MXD4
MAX dimerization protein 4
230472_at 5,6 6,4 8,6 6,0 25,5 40,8 44,0 33,7 0,8787 IRX1 iroquois homeobox 1
231963_at 9,6 32,3 35,6 58,4 48,0 69,0 49,7 97,3 0,8781 --- ---
238711_s_at 62,7 49,5 64,1 67,5 69,0 70,2 79,0 82,7 0,8780 ZNF148
zinc finger protein 148
201108_s_at 1425,1 944,4
1803,8
1850,3
2639,4
2609,3
3417,6
2642,9
0,8777 THBS1 thrombospondin 1
226056_at 53,8 67,6 67,6 85,0 76,4 83,4 80,3 90,5 0,8772 CDGAP Cdc42 GTPase-
activating protein
1553764_a_at 109,5 94,9 92,4 133,8 127,6 151,3 252,2 254,3 0,8770 JUB
jub, ajuba homolog (Xenopus laevis)
236335_at 3,5 8,6 1,8 11,6 13,9 13,8 15,6 17,3 0,8760 --- ---
215575_at 6,2 5,1 9,9 6,4 14,6 10,0 18,7 21,0 0,8755 PDE4DI
P
phosphodiesterase 4D interacting
protein (myomegalin) 228932_at 33,5 34,6 36,4 45,6 55,2 48,2 71,4 57,3 0,8751 --- ---
212136_at 456,7 399,0 483,4 444,2 545,5 556,2 726,4 897,8 0,8750 ATP2B4 ATPase, Ca++
transporting, plasma membrane 4
206965_at 4,4 3,9 9,7 7,2 10,8 13,5 21,0 14,5 0,8748 KLF12 Kruppel-like factor
12
239809_at 15,5 20,6 19,6 15,7 30,8 26,8 49,0 47,9 0,8745 --- ---
229819_at 19,3 32,6 26,6 44,5 45,9 72,3 56,3 58,8 0,8744 A1BG alpha-1-B
glycoprotein
236883_at 21,6 35,7 28,1 26,7 45,5 39,4 62,2 59,5 0,8743 --- ---
213948_x_at 11,3 2,4 37,2 40,9 62,9 97,1 107,4 68,9 0,8742 CADM3
cell adhesion molecule 3
1563719_a_at
7,5 4,7 8,0 5,9 8,1 12,2 16,0 17,3 0,8741 --- ---
222786_at 91,3 99,5 103,0 114,9 133,5 129,4 142,9 124,2 0,8739 CHST12 carbohydrate
(chondroitin 4) sulfotransferase 12
236042_at 30,1 31,1 33,6 25,5 37,6 50,7 58,2 56,8 0,8736 LOC100130219
similar to hCG2041987
233288_at 1,5 0,7 1,3 2,2 3,1 8,1 7,1 6,5 0,8736 ATR ataxia telangiectasia
and Rad3 related
1565951_s_at 23,5 21,0 23,1 19,9 29,7 35,8 53,0 64,8 0,8727 CHML
choroideremia-like (Rab escort protein
2)
242080_at 16,6 12,2 32,0 29,3 54,0 30,9 51,8 68,3 0,8726 --- ---
209460_at 71,0 83,7 85,6 80,5 86,0 86,6 110,4 117,3 0,8724 ABAT 4-aminobutyrate
aminotransferase
204693_at 166,8 252,5 245,7 308,3 264,0 465,5 376,9 419,4 0,8723 CDC42E
P1
CDC42 effector protein (Rho GTPase
binding) 1
212614_at 838,9 1243,0 901,2 1522,
0 1542,
2 1507,
3 1952,
5 1713,
1 0,8712 ARID5B
AT rich interactive domain 5B (MRF1-
like)
1559419_at 5,8 13,7 19,3 18,9 45,3 40,9 99,6 66,2 0,8704
CACNB2
calcium channel, voltage-dependent,
beta 2 subunit
221541_at 796,1 917,9 650,8 1086,
4 1149,
2 1441,
4 1351,
2 1366,
4 0,8703
CRISPLD2
cysteine-rich secretory protein
LCCL domain containing 2
217744_s_at 227,1 246,2 497,8 550,2 538,0 478,0 738,2 639,2 0,8702 PERP
PERP, TP53 apoptosis effector
1566696_at
7,3 7,5 8,9 8,3 7,9 12,1 16,2 20,4 0,8697 --- ---

Tablas Suplementarias
149
226311_at 585,4 1135,1 758,0 1047,
2 864,3
1465,4
1875,6
1882,1
0,8695 --- ---
209869_at 89,1 273,4 602,2 545,4 1383,
4 1222,
1 1406,
6 1082,
3 0,8693
ADRA2A
adrenergic, alpha-2A-, receptor
1558636_s_at 86,6 35,2 62,9 90,3 106,2 154,1 157,0 152,4 0,8689
ADAMTS5
ADAM metallopeptidase
with thrombospondin
type 1 motif, 5 (aggrecanase-2) 202790_at 25,1 27,0 42,7 40,9 36,2 77,7 61,6 110,8 0,8687 CLDN7 claudin 7
212057_at 220,0 150,3 144,1 219,5 249,5 290,8 308,7 365,2 0,8678 KIAA01
82 KIAA0182
201285_at 376,8 424,2 337,1 443,6 463,0 473,9 554,0 532,2 0,8676 MKRN1 makorin, ring finger
protein, 1
203071_at 17,2 48,7 21,4 65,4 74,6 59,0 90,3 87,1 0,8672 SEMA3
B
sema domain, immunoglobulin
domain (Ig), short basic domain,
secreted, (semaphorin) 3B
209286_at 116,4 211,9 149,0 188,6 236,6 198,4 298,3 328,7 0,8671 CDC42E
P3
CDC42 effector protein (Rho GTPase
binding) 3
217762_s_at 864,3 1181,3 733,6
1272,7
1371,2
1546,9
1781,8
1613,3
0,8666 RAB31 RAB31, member RAS
oncogene family
237218_at 16,5 14,3 10,4 26,9 39,1 34,8 44,1 37,4 0,8665 --- ---
241765_at 31,3 111,9 56,3 101,1 130,1 113,3 229,2 200,8 0,8655 CPM carboxypeptidase M
221555_x_at 70,8 77,8 64,6 88,2 91,3 105,5 163,6 137,5 0,8654
CDC14B ///
CDC14C
CDC14 cell division cycle 14 homolog B
(S. cerevisiae) /// CDC14 cell division cycle 14 homolog C
(S. cerevisiae)
1556461_at
0,4 0,8 2,0 3,4 1,1 3,2 6,3 5,8 0,8653 --- ---
206100_at 361,7 839,4 621,8 724,5 984,7 797,5 1741,5
1828,5
0,8648 CPM carboxypeptidase M
203485_at 19,0 36,2 45,3 37,3 36,8 60,4 55,7 58,6 0,8645 RTN1 reticulon 1
209602_s_at 1,5 0,4 2,8 1,8 4,8 11,3 6,9 10,4 0,8642 GATA3
GATA binding protein 3
233496_s_at
180,7 161,8 192,2 216,9 227,8 273,5 255,9 414,9 0,8641 CFL2 cofilin 2 (muscle)
219403_s_at
43,7 89,1 109,8 149,4 153,7 209,7 337,7 205,8 0,8641 HPSE heparanase
229590_at 62,2 110,8 103,6 138,4 132,4 157,8 134,5 157,0 0,8639 RPL13 ribosomal protein
L13
243455_at 15,5 23,3 28,4 12,3 33,8 34,8 46,5 53,4 0,8639 --- ---
220856_x_at
730,1 829,9 722,5 851,6 1066,0
915,0 1347,9
1215,4
0,8635 --- ---
242074_at 16,5 12,6 10,0 24,1 35,1 31,2 46,1 35,8 0,8628 --- ---
205691_at 2,0 5,7 18,1 33,3 24,0 23,0 44,3 34,7 0,8626 SYNGR
3 synaptogyrin 3
210847_x_at 1,6 6,1 8,4 7,9 9,3 12,9 35,2 27,6 0,8613
TNFRSF25
tumor necrosis factor receptor
superfamily, member 25
231766_s_at 719,6 567,7 682,4 983,6
1475,9
1736,8
1439,3
1459,9
0,8608 COL12A
1 collagen, type XII,
alpha 1
231822_at 74,7 55,7 47,3 66,8 80,9 105,8 124,9 126,7 0,8607 CTTNBP
2NL CTTNBP2 N-terminal
like
228310_at 144,8 157,9 262,0 180,3 336,5 324,8 756,2 608,3 0,8607 ENAH enabled homolog
(Drosophila)
233105_at 17,6 16,8 11,8 20,0 25,0 22,5 27,5 34,7 0,8606 --- ---

Tablas Suplementarias
150
200859_x_at 1444,4 1990,4
2108,1
3073,9
1999,3
3055,7
3218,9
3374,4
0,8604 FLNA filamin A, alpha (actin binding protein 280)
233824_at 14,8 13,3 17,4 15,2 34,8 29,3 28,8 35,4 0,8601 --- ---
233748_x_at 116,9 84,3 242,9 213,4 199,5 221,4 263,4 355,2 0,8600
PRKAG2
protein kinase, AMP-activated, gamma 2
non-catalytic subunit
211358_s_at 93,1 110,5 75,7 116,8 116,5 130,8 142,2 178,4 0,8599 CIZ1
CDKN1A interacting zinc finger protein 1
209740_s_at 52,8 61,4 63,1 83,4 87,4 86,3 103,5 83,4 0,8599 PNPLA4
patatin-like phospholipase
domain containing 4
1570078_a_at 1,8 2,5 5,8 2,3 2,9 9,1 11,5 11,8 0,8597 DOCK5
dedicator of cytokinesis 5
212748_at 81,5 70,7 57,1 81,6 95,5 114,4 117,1 120,5 0,8596 MKL1 megakaryoblastic
leukemia (translocation) 1
226066_at 93,5 135,0 114,9 194,6 211,9 218,0 185,2 223,6 0,8595 MITF microphthalmia-
associated transcription factor
241796_x_at
0,2 0,1 1,4 1,2 1,3 0,9 2,2 3,0 0,8594 --- ---
229843_at 12,6 3,3 23,5 31,7 24,3 53,5 35,7 50,6 0,8592 FAM82B Family with
sequence similarity 82, member B
200606_at 35,2 35,4 67,1 99,8 68,1 69,7 149,8 138,4 0,8592 DSP desmoplakin
212567_s_at 130,1 232,1 234,9 265,3 267,1 249,9 288,8 312,2 0,8590 MAP4
microtubule-associated protein 4
240655_at 13,8 20,1 12,7 32,1 43,0 28,3 53,9 46,0 0,8583 --- ---
241607_at 58,1 54,2 59,9 60,0 73,0 101,1 78,3 101,5 0,8577 LOC730
102 hypothetical protein
LOC730102
223000_s_at
18,3 17,4 18,7 10,5 27,7 34,8 42,4 42,3 0,8573 F11R F11 receptor
242894_at 9,9 14,3 12,3 5,8 25,0 27,8 27,0 35,8 0,8573 --- ---
215179_x_at 249,2 331,7 273,5 274,0 346,2 394,7 407,6 396,5 0,8572 PGF
Placental growth factor
229357_at 482,6 729,0 452,4 801,8 838,5 968,6 1830,
1 1478,
6 0,8568
ADAMTS5
ADAM metallopeptidase
with thrombospondin
type 1 motif, 5 (aggrecanase-2)
1562474_at
74,5 95,0 75,9 90,6 88,9 124,4 142,6 127,2 0,8565 --- ---
205805_s_at 44,7 80,3 98,6 100,8 128,3 125,9 111,1 127,0 0,8549 ROR1
receptor tyrosine kinase-like orphan
receptor 1
236671_at 50,1 60,2 61,1 62,2 74,4 82,5 75,9 72,6 0,8549 --- ---
204718_at 38,0 165,6 130,0 138,6 178,8 340,4 285,7 268,6 0,8542 EPHB6 EPH receptor B6
213142_x_at 19,6 35,1 26,8 35,5 44,4 35,9 56,0 48,4 0,8541
tcag7.1314
hypothetical protein LOC54103
243367_at 26,5 35,9 39,4 34,5 45,0 41,9 48,4 73,3 0,8539 --- ---
215823_x_at 2886,1 3416,2
2608,6
3280,6
3766,1
4404,9
4293,2
4250,9
0,8536 PABPC3 poly(A) binding
protein, cytoplasmic 3
225235_at 103,3 150,7 129,9 196,2 152,3 211,5 186,0 227,9 0,8533 TSPAN17
tetraspanin 17
236999_at 4,6 3,1 2,6 2,5 6,1 7,8 9,5 10,6 0,8531 --- ---
235373_at 8,5 7,7 6,4 17,7 19,9 19,4 25,7 19,4 0,8530 --- ---
231016_s_at
66,5 66,6 77,0 84,4 85,3 74,5 99,0 94,6 0,8529 --- ---
221039_s_at 243,7 211,0 219,0 285,7 252,1 283,0 351,4 435,9 0,8529 DDEF1
development and differentiation
enhancing factor 1

Tablas Suplementarias
151
211004_s_at 88,8 98,2 162,7 162,9 172,3 220,1 164,5 210,3 0,8528
ALDH3B1
aldehyde dehydrogenase 3
family, member B1
215813_s_at 285,0 282,8 566,5 398,4 623,6 449,7 745,0 786,6 0,8526 PTGS1
prostaglandin-endoperoxide
synthase 1 (prostaglandin G/H
synthase and cyclooxygenase)
235409_at 35,4 19,0 26,7 28,0 39,5 45,6 57,8 57,7 0,8523 MGA MAX gene associated
1561190_at 15,6 11,6 11,0 23,4 26,3 19,6 33,4 32,3 0,8522 CDKL3
cyclin-dependent kinase-like 3
225897_at 606,8 874,7 548,3 726,2 1051,
6 915,2
1419,6
1385,3
0,8520 MARCK
S
myristoylated alanine-rich protein kinase C substrate
36554_at 158,1 201,2 153,9 186,1 204,3 222,9 218,7 247,0 0,8517 ASMTL acetylserotonin O-methyltransferase-
like
225685_at 136,1 366,6 232,7 419,1 404,0 348,7 508,1 568,5 0,8509 --- ---
208727_s_at 130,8 182,2 162,8 207,1 326,4 221,2 574,1 795,5 0,8505 CDC42
cell division cycle 42 (GTP binding
protein, 25kDa)
239913_at 0,4 0,4 1,3 0,5 1,1 3,9 3,2 3,2 0,8500 SLC10A
4
solute carrier family 10 (sodium/bile acid
cotransporter family), member 4
227242_s_at 21,1 11,0 20,7 14,2 24,5 39,1 40,5 65,5 0,8500 EBF3 early B-cell factor 3
221893_s_at 43,6 74,7 56,0 74,6 71,7 84,1 84,1 90,0 0,8498 ADCK2
aarF domain containing kinase 2
238934_at 4,7 9,0 5,6 16,3 19,0 12,3 18,3 21,2 0,8496 --- ---
219489_s_at 397,2 510,9 539,2 557,4 515,0 574,1
1008,8
1125,3
0,8494 NXN nucleoredoxin
230258_at 153,5 101,7 169,9 173,2 282,1 223,2 381,7 297,2 0,8493 GLIS3 GLIS family zinc
finger 3
215717_s_at 8,6 7,0 2,1 24,5 12,8 21,1 55,1 72,1 0,8492 FBN2
fibrillin 2 (congenital contractural
arachnodactyly)
225807_at 23,1 32,6 44,0 39,8 40,4 35,5 59,6 70,3 0,8491 JUB jub, ajuba homolog
(Xenopus laevis)
203110_at 26,9 46,5 53,4 54,3 91,6 124,0 93,0 92,1 0,8486 PTK2B PTK2B protein
tyrosine kinase 2 beta
201109_s_at 1889,5 1611,8
1698,3
2222,5
3233,5
2677,1
4734,0
3644,0
0,8484 THBS1 thrombospondin 1
1560026_at
7,5 5,6 9,8 9,6 12,0 9,3 21,7 20,7 0,8483 --- ---
237452_at 25,9 21,8 29,6 53,6 63,4 41,6 73,3 64,6 0,8481 --- ---
241860_at 5,3 16,7 5,7 6,4 33,0 27,0 51,5 40,6 0,8479 --- ---
202986_at 11,5 14,0 14,1 11,0 20,4 37,5 54,1 38,4 0,8469 ARNT2 aryl-hydrocarbon receptor nuclear
translocator 2
236234_at 76,0 160,9 142,0 141,2 196,2 176,3 340,7 265,3 0,8468 PDE1A phosphodiesterase
1A, calmodulin-dependent
237868_x_at
34,1 27,5 29,2 26,4 51,2 42,0 57,1 66,8 0,8468 --- ---
219790_s_at 299,3 310,6 296,9 426,4 415,9 372,3 614,4 527,6 0,8467 NPR3
natriuretic peptide receptor C/guanylate
cyclase C (atrionatriuretic
peptide receptor C)
228752_at 33,9 33,8 47,2 31,4 69,6 55,9 77,9 70,8 0,8464 CXorf10 chromosome X open
reading frame 10

Tablas Suplementarias
152
202847_at 315,2 282,5 345,9 481,8 425,3 467,1 556,1 466,3 0,8464 PCK2 phosphoenolpyruvat
e carboxykinase 2 (mitochondrial)
226140_s_at 120,7 131,4 119,9 134,4 162,8 148,7 185,5 161,8 0,8460 OTUD1
OTU domain containing 1
232898_at 27,6 18,5 30,0 23,4 62,2 65,2 65,0 59,5 0,8454 DAB2
disabled homolog 2, mitogen-responsive
phosphoprotein (Drosophila)
237157_at 32,3 40,4 17,2 44,3 63,4 45,3 94,7 105,4 0,8449 --- ---
212249_at 83,1 46,7 55,9 119,1 233,0 186,3 238,5 199,2 0,8447 PIK3R1 phosphoinositide-3-kinase, regulatory subunit 1 (alpha)
212209_at 151,1 121,1 164,3 151,8 260,1 276,9 233,2 263,4 0,8447 MED13L mediator complex
subunit 13-like
1567860_at
0,8 0,3 1,4 2,1 1,3 1,5 2,8 2,8 0,8446 --- ---
227485_at 30,5 30,1 35,6 51,1 42,6 51,4 51,1 48,5 0,8441 DDX26B DEAD/H (Asp-Glu-Ala-Asp/His) box polypeptide 26B
222133_s_at 42,0 38,7 48,0 76,6 73,1 61,2 107,7 85,9 0,8439
PHF20L1
PHD finger protein 20-like 1
226482_s_at 10,1 26,5 81,9 62,5 85,5 81,0 89,6 89,8 0,8437
RP11-544M22.
4 /// hCG_20
857
KAT protein /// hypothetical protein
LOC100134860
237724_at 0,8 1,3 2,9 3,0 2,6 4,8 4,5 11,1 0,8433 --- ---
242446_at 3,5 1,3 9,0 5,7 9,8 11,2 8,8 13,0 0,8430 C6orf16
3 chromosome 6 open
reading frame 163
1555989_at
43,6 43,9 74,6 62,0 56,7 82,0 86,1 82,6 0,8429 --- ---
213247_at 480,1 681,2 599,5 691,7 1066,
9 1055,
3 1555,
5 1056,
8 0,8425 SVEP1
sushi, von Willebrand factor type A, EGF and
pentraxin domain containing 1
242593_at 13,9 15,3 11,8 14,2 18,0 19,3 18,1 21,3 0,8422 --- ---
238000_at 11,5 21,4 21,2 11,1 23,1 28,2 44,3 40,3 0,8419 --- ---
1556341_s_at 50,7 82,3 64,4 80,0 105,2 93,0 89,0 124,4 0,8417 MAPK12
mitogen-activated protein kinase 12
1556886_a_at
3,6 14,6 22,2 22,4 36,2 30,4 31,9 29,7 0,8413 LAYN Layilin
214415_at 9,4 21,7 13,7 11,0 38,4 37,7 59,5 42,5 0,8413 PLGLB2
/// PLGLB1
plasminogen-like B2 /// plasminogen-like
B1
211177_s_at 62,4 93,1 79,0 96,3 84,6 138,7 110,9 139,5 0,8412 TXNRD2
thioredoxin reductase 2
206377_at 68,1 43,5 164,8 220,3 184,8 183,5 205,3 244,7 0,8409 FOXF2 forkhead box F2
244286_at 3,8 9,8 3,4 11,3 11,6 10,4 18,7 16,4 0,8409 --- ---
238970_at 9,8 6,0 3,3 3,8 24,4 21,3 58,2 49,0 0,8407 --- ---
234981_x_at 463,7 594,1 621,3 625,4 717,9 633,8 721,3 703,2 0,8405 CMBL
carboxymethylenebutenolidase homolog
(Pseudomonas)
211006_s_at 34,7 115,3 118,2 128,5 99,9 125,7 196,1 300,8 0,8399 KCNB1
potassium voltage-gated channel, Shab-
related subfamily, member 1

Tablas Suplementarias
153
215634_at 4,6 14,9 15,0 16,8 17,5 23,3 21,6 48,5 0,8399 --- ---
203886_s_at
86,6 129,2 279,3 316,2 417,1 509,0 415,6 369,5 0,8396 FBLN2 fibulin 2
242973_at 25,2 39,4 39,5 48,1 80,2 83,1 87,7 64,5 0,8388 CACNA
1C
calcium channel, voltage-dependent, L
type, alpha 1C subunit
202402_s_at 277,5 237,7 301,5 338,1 317,7 329,1 423,9 367,1 0,8388 CARS
cysteinyl-tRNA synthetase
225566_at 92,4 73,6 123,5 149,5 100,9 171,8 198,9 172,1 0,8381 NRP2 neuropilin 2
204568_at 119,4 164,7 115,4 150,1 203,1 178,3 215,2 209,6 0,8380 KIAA08
31 KIAA0831
223746_at 6,9 8,6 13,7 12,2 9,7 15,2 19,0 16,1 0,8377 STK4 serine/threonine
kinase 4
227762_at 368,6 238,4 279,4 318,2 592,2 493,9 636,5 612,7 0,8376 --- ---
235901_at 31,9 79,9 38,3 65,0 85,5 82,4 122,1 107,1 0,8375 --- ---
1557452_at
13,0 10,6 19,6 5,5 23,4 38,3 39,2 37,3 0,8375 --- ---
216215_s_at 679,0 685,0 964,5 908,5 814,2 985,0 947,4
1161,8
0,8373 RBM9 RNA binding motif
protein 9
208685_x_at 344,6 311,9 301,8 344,3 404,1 425,3 487,5 423,2 0,8372 BRD2
bromodomain containing 2
202016_at 289,9 614,4 959,4 939,7 1271,
1 1114,
4 3627,
1 2615,
6 0,8366 MEST
mesoderm specific transcript homolog
(mouse)
220241_at 46,3 35,2 51,6 55,4 61,4 62,5 59,5 63,3 0,8366 TMCO3 transmembrane and coiled-coil domains
3
224904_at 168,7 222,8 185,0 197,4 299,7 253,6 308,1 287,5 0,8365 PDPR
pyruvate dehydrogenase
phosphatase regulatory subunit
229741_at 73,4 153,4 89,0 110,3 141,4 162,3 194,5 196,0 0,8363 MGC326
0 Hypothetical LOC78993
225790_at 112,4 95,0 112,9 127,6 142,8 155,5 131,9 152,4 0,8363 MSRB3 methionine sulfoxide
reductase B3
239129_at 5,6 4,9 1,3 7,5 17,0 13,4 20,2 16,0 0,8356 --- ---
227929_at 20,2 101,8 65,9 102,9 136,6 100,2 123,9 168,7 0,8352 --- ---
242859_at 21,9 16,8 19,5 11,8 28,9 40,7 68,9 57,8 0,8349 --- ---
216080_s_at 171,9 252,1 304,2 441,8 278,5 376,8 413,1 480,0 0,8347 FADS3
fatty acid desaturase 3
217892_s_at 1096,6 1597,3
1377,1
1619,2
1783,6
1507,1
2519,4
2255,7
0,8346 LIMA1 LIM domain and actin binding 1
238281_at 9,1 12,9 17,8 17,1 25,8 13,0 37,3 47,4 0,8346 --- ---
232974_at 12,1 23,4 22,2 24,8 32,0 25,1 32,1 30,8 0,8343 --- ---
209409_at 241,3 378,4 285,1 387,5 408,4 469,9 479,7 430,3 0,8342 GRB10 growth factor
receptor-bound protein 10
204066_s_at 136,3 140,5 143,7 161,9 167,5 173,4 186,4 161,4 0,8341 CENTG2 centaurin, gamma 2
239153_at 11,1 39,2 16,4 20,2 33,3 37,6 72,7 77,0 0,8341 FLJ4174
7
hypothetical gene supported by
AK123741
235959_at 15,9 15,1 18,5 19,0 19,1 33,6 28,1 27,8 0,8341 --- ---
233480_at 51,6 75,2 49,2 70,1 87,4 70,9 99,6 109,4 0,8340 TMEM43 Transmembrane protein 43
239336_at 109,1 240,7 178,7 210,3 229,1 217,5 503,3 509,7 0,8339 THBS1 Thrombospondin 1
225228_at 105,4 124,7 123,3 161,2 197,9 232,6 175,4 198,6 0,8338 TMEM77 transmembrane protein 77

Tablas Suplementarias
154
221869_at 41,0 47,6 40,0 43,6 48,3 56,3 75,8 64,8 0,8336 ZNF512
B zinc finger protein
512B
220132_s_at 7,5 5,6 9,1 12,7 12,5 21,9 21,4 15,5 0,8330 CLEC2D
C-type lectin domain family 2, member D
224659_at 123,3 297,8 202,6 246,6 280,0 294,9 322,1 386,5 0,8328 SEPN1 selenoprotein N, 1
236617_at 5,4 8,0 6,8 3,7 19,3 12,9 35,9 29,6 0,8325 --- ---
227361_at 53,2 52,8 22,3 51,9 90,9 89,3 90,6 143,8 0,8325 HS3ST3
B1
heparan sulfate (glucosamine) 3-O-
sulfotransferase 3B1
203896_s_at 3,0 14,1 6,8 14,2 6,7 17,8 23,3 32,4 0,8324 PLCB4
phospholipase C, beta 4
233241_at 8,8 3,4 12,9 10,2 23,1 25,1 32,4 20,9 0,8323 C20orf1
9
chromosome 20 open reading frame
19
200779_at 2536,8 2447,1 2556,
8 3135,
8 3307,
1 3276,
3 3892,
9 3196,
1 0,8323 ATF4
activating transcription factor 4
(tax-responsive enhancer element
B67)
205934_at 13,3 23,2 31,0 41,8 40,0 55,1 51,0 40,0 0,8318 PLCL1 phospholipase C-like
1
240869_at 26,9 64,7 114,5 77,2 92,3 95,2 123,2 213,7 0,8318 --- ---
224833_at 378,8 305,6 204,4 363,4 607,9 581,0 659,7 641,3 0,8311 ETS1
v-ets erythroblastosis
virus E26 oncogene homolog 1 (avian)
240485_at 5,1 6,3 15,7 20,6 15,6 21,8 19,7 20,2 0,8310 --- ---
202998_s_at 1612,8 1490,3
2208,9
2826,5
2951,1
2693,9
3502,4
2745,1
0,8305 LOXL2
/// ENTPD4
lysyl oxidase-like 2 /// ectonucleoside
triphosphate diphosphohydrolase
4
224882_at 20,3 42,4 61,6 52,8 72,0 88,0 98,5 67,5 0,8304 ACSS1 acyl-CoA synthetase
short-chain family member 1
228111_s_at 9,6 7,6 12,1 19,3 22,8 26,3 20,8 21,2 0,8303 DNAH1
dynein, axonemal, heavy chain 1
1558922_at
18,9 10,5 17,7 12,5 21,2 26,7 35,6 32,5 0,8303 --- ---
243636_s_at
10,0 10,7 13,7 24,9 31,7 22,0 33,9 26,8 0,8302 --- ---
239234_at 11,9 13,5 7,3 24,8 19,1 24,9 33,6 26,8 0,8300 --- ---
240908_at 17,3 22,7 15,7 16,1 30,5 30,5 48,1 38,2 0,8299 --- ---
213375_s_at 27,3 31,6 33,7 41,2 50,4 47,2 68,2 47,0 0,8294
N4BP2L1
NEDD4 binding protein 2-like 1
1556212_x_at
11,1 12,2 11,7 15,2 16,9 12,0 20,0 21,8 0,8293 --- ---
212741_at 897,1 1357,8 969,1 1623,
4 1582,
8 2080,
5 1684,
2 1873,
7 0,8292 MAOA
monoamine oxidase A
230808_at 16,7 13,4 20,8 15,8 25,2 20,1 26,7 28,1 0,8288 FNTA farnesyltransferase,
CAAX box, alpha
207373_at 5,8 5,3 21,5 33,5 29,8 39,4 31,1 32,7 0,8286 HOXD10 homeobox D10
222336_at 11,9 19,0 15,3 21,9 34,4 27,5 26,6 31,2 0,8283 C4orf34 chromosome 4 open
reading frame 34
1563721_at
0,2 0,4 3,0 1,4 2,2 2,9 2,5 4,1 0,8276 --- ---
224318_s_at 230,5 314,4 275,6 401,4 462,1 401,9 439,7 417,4 0,8274
KIAA1310
KIAA1310
1565863_at 11,8 13,9 27,1 14,5 20,1 23,7 31,3 45,4 0,8274 --- ---

Tablas Suplementarias
155
210300_at 21,9 19,2 26,2 39,4 30,1 69,8 63,1 51,3 0,8272 REM1 RAS (RAD and GEM)-like GTP-
binding 1
213590_at 45,8 54,8 62,0 88,3 87,7 103,0 99,6 82,1 0,8271
SLC16A5 ///
LOC100133772
solute carrier family 16, member 5
(monocarboxylic acid transporter 6) ///
similar to MCT
207953_at 22,8 24,5 31,0 31,3 41,0 29,3 38,7 40,7 0,8270 RP5-
886K2.1 neuronal thread
protein AD7c-NTP
232420_x_at 61,6 103,0 64,7 74,3 112,7 106,6 177,7 153,5 0,8270
LOC286260
hypothetical protein LOC286260
244490_at 13,4 4,6 4,1 10,4 24,7 34,9 52,4 33,4 0,8261 --- ---
205609_at 811,0 1054,9 663,0 1023,
1 1536,
5 1552,
8 1887,
6 1476,
8 0,8257 ANGPT1 angiopoietin 1
237750_at 3,1 2,1 3,7 9,5 12,0 7,8 12,8 10,3 0,8257 XPNPEP
3
X-prolyl aminopeptidase
(aminopeptidase P) 3, putative
1558702_at
12,8 15,1 13,9 19,0 21,5 15,3 42,4 44,5 0,8255 TEX10 Testis expressed 10
242245_at 14,4 17,5 22,1 24,5 31,2 18,6 40,2 37,1 0,8254 --- ---
222909_s_at 26,0 28,4 38,5 40,5 44,2 30,8 51,4 65,1 0,8253 BAG4
BCL2-associated athanogene 4
220786_s_at 21,8 13,5 11,0 21,9 19,0 62,3 49,6 98,8 0,8249
SLC38A4
solute carrier family 38, member 4
222334_at 2,9 3,4 6,5 4,9 8,3 6,1 11,6 22,2 0,8248 --- ---
235508_at 40,8 48,9 41,6 38,6 67,0 51,3 75,3 79,7 0,8246 PML promyelocytic
leukemia
212580_at 27,1 17,5 24,1 18,8 33,7 40,1 37,7 43,7 0,8245 ERAP1 Endoplasmic
reticulum aminopeptidase 1
1558028_x_at
62,9 37,9 39,8 61,1 78,3 72,2 88,0 132,4 0,8245 --- ---
203184_at 496,0 622,9 617,8 839,3 476,4 1036,
0 2543,
2 2723,
8 0,8240 FBN2
fibrillin 2 (congenital contractural
arachnodactyly)
239561_at 20,0 20,6 13,0 31,5 53,8 37,5 56,2 47,1 0,8238 --- ---
230915_at 3,0 2,1 4,1 4,5 4,7 6,8 8,4 5,5 0,8237 DHRS7
C
dehydrogenase/reductase (SDR family)
member 7C
215095_at 24,9 29,3 24,8 33,5 36,4 40,7 45,9 35,7 0,8234 ESD Esterase
D/formylglutathione hydrolase
225806_at 125,4 202,1 101,8 158,0 204,3 199,8 403,4 395,7 0,8233 JUB jub, ajuba homolog
(Xenopus laevis)
202720_at 432,7 938,3 744,7 753,5 872,9 858,5 1326,
9 1225,
3 0,8230 TES
testis derived transcript (3 LIM
domains)
217253_at 49,6 41,4 30,2 32,0 64,9 64,7 88,8 95,3 0,8226 SH3BP2 SH3-domain binding
protein 2
201398_s_at 2752,6 3346,4
2699,5
3528,8
3797,3
3814,3
4453,9
3811,3
0,8226
TRAM1 ///
LOC100130862
translocation associated
membrane protein 1 /// PRO1292
208335_s_at 3,3 3,2 5,4 4,6 31,4 22,1 57,8 33,2 0,8226 DARC
Duffy blood group, chemokine receptor
203766_s_at 16,8 48,1 8,8 50,7 84,9 94,0 78,6 85,0 0,8225 LMOD1
leiomodin 1 (smooth muscle)

Tablas Suplementarias
156
212573_at 149,4 255,2 317,7 351,8 257,9 356,7 351,1 403,2 0,8225 ENDOD
1 endonuclease
domain containing 1
AFFX-HUMRGE/M10098_5
_at
47,1 126,3 48,3 211,6 265,0 188,3 422,6 277,0 0,8225 --- ---
208614_s_at 159,0 302,3 301,6 333,6 328,6 280,5 427,0 419,0 0,8225 FLNB
filamin B, beta (actin binding protein 278)
236692_at 34,4 19,4 23,6 23,3 42,3 50,6 51,0 54,4 0,8223 LOC729
839 hypothetical LOC729839
205883_at 360,0 419,6 298,5 361,9 697,8 640,4 902,7 687,9 0,8218 ZBTB16 zinc finger and BTB domain containing
16
214757_at 24,3 14,6 28,8 43,4 33,4 43,7 42,3 43,7 0,8216 PMS2L2 postmeiotic segregation
increased 2-like 2
208022_s_at 52,8 52,2 54,0 42,8 79,4 70,0 148,1 131,1 0,8215
CDC14B ///
CDC14C
CDC14 cell division cycle 14 homolog B
(S. cerevisiae)
236450_at 3,1 2,3 3,9 6,6 6,2 4,7 11,7 8,7 0,8210 --- ---
AFFX-HUMISGF3A/M97935_MB_at
220,9 208,3 284,8 291,8 263,4 307,8 278,7 336,5 0,8210 STAT1
signal transducer and activator of transcription 1,
91kDa
228465_at 27,1 33,4 47,2 64,4 75,4 45,0 76,5 76,7 0,8206 --- ---
1558390_at 5,3 1,1 3,2 5,4 4,3 10,0 8,9 11,0 0,8205 ZNF599
zinc finger protein 599
225220_at 745,6 1003,9 642,6 982,7 1305,
7 1436,
2 1352,
6 1305,
7 0,8204 SNHG8
small nucleolar RNA host gene (non-
protein coding) 8
209439_s_at 57,1 63,7 44,0 75,4 87,7 112,9 84,6 106,5 0,8200 PHKA2
phosphorylase kinase, alpha 2 (liver)
234994_at 159,7 90,9 83,2 127,6 179,0 168,9 287,5 347,0 0,8196 TMEM20
0A transmembrane
protein 200A
233914_s_at 27,4 33,6 34,4 31,5 35,1 49,5 95,9 75,6 0,8196 SBF2 SET binding factor 2
235482_at 31,0 63,5 40,6 54,8 72,3 85,8 107,4 76,9 0,8195 LOC400
960
hypothetical gene supported by
BC040598
201064_s_at 963,0 1311,1
1019,8
1378,8
1171,5
1628,1
1502,0
1637,9
0,8194 PABPC4 poly(A) binding
protein, cytoplasmic 4 (inducible form)
226978_at 45,7 72,4 74,1 110,6 89,5 113,4 106,7 102,4 0,8191 PPARA peroxisome
proliferator-activated receptor alpha
223253_at 863,6 974,0 914,5 999,6 1289,
7 1101,
9 2043,
7 1642,
4 0,8191 EPDR1
ependymin related protein 1 (zebrafish)
205608_s_at 544,4 342,4 334,0 651,2
1200,1
1199,3
1196,6
1034,5
0,8188 ANGPT1 angiopoietin 1
231235_at 4,8 7,0 9,4 0,9 25,9 28,1 33,8 24,5 0,8185 NKTR natural killer-tumor
recognition sequence
224818_at 140,5 164,5 241,6 181,8 214,2 243,4 310,7 250,5 0,8184 SORT1 sortilin 1

Tablas Suplementarias
157
65472_at 33,1 30,6 18,5 32,1 42,1 41,4 56,8 54,4 0,8183 LOC388
969 hypothetical LOC388969
231860_at 9,9 10,5 13,1 14,3 15,9 14,5 13,3 16,6 0,8181 BRWD1 bromodomain and WD repeat domain
containing 1
239432_at 39,0 17,6 31,9 24,6 52,6 49,5 61,8 65,9 0,8180 FLJ3130
6 hypothetical protein
FLJ31306
201926_s_at 148,1 186,5 129,9 128,4 240,8 206,6 279,8 293,5 0,8177 CD55
CD55 molecule, decay accelerating
factor for complement (Cromer
blood group)
1559739_at 9,0 11,5 10,0 9,0 17,7 16,7 15,5 17,5 0,8176 CHPT1
Choline phosphotransferase
1
AFFX-HUMRGE/M10098_M
_at
52,4 135,6 84,1 163,6 238,7 151,9 413,4 284,3 0,8176 --- ---
205992_s_at 39,8 182,3 117,0 143,2 186,2 137,5 253,2 283,0 0,8175 IL15 interleukin 15
207876_s_at 38,6 80,7 103,9 126,2 103,2 79,7 149,8 177,2 0,8172 FLNC
filamin C, gamma (actin binding protein 280)
241336_at 14,5 16,0 21,4 29,1 41,0 49,0 37,8 35,1 0,8172 --- ---
207776_s_at 15,8 18,3 46,7 49,0 121,4 104,5 107,9 82,4 0,8171
CACNB2
calcium channel, voltage-dependent,
beta 2 subunit
221071_at 11,7 13,6 17,4 17,0 24,2 32,9 30,9 22,8 0,8167 --- ---
210224_at 113,2 124,0 133,7 139,4 124,0 133,7 159,2 149,4 0,8167 MR1
major histocompatibility complex, class I-
related
233614_at 15,5 12,7 21,0 10,5 31,4 20,4 45,0 44,3 0,8167 --- ---
212208_at 120,9 119,7 153,5 136,1 227,4 183,7 237,1 196,3 0,8164 MED13L mediator complex
subunit 13-like
216173_at 6,9 9,4 11,3 2,2 16,2 14,6 21,9 24,8 0,8163 --- ---
225551_at 61,0 67,3 65,1 77,4 74,6 66,9 87,0 108,2 0,8159 C1orf71 chromosome 1 open
reading frame 71
1556606_at
11,7 64,3 47,0 20,6 99,8 91,4 86,9 119,8 0,8159 NAV2 neuron navigator 2
241949_at 1,0 0,8 3,9 4,4 4,8 6,3 3,6 6,4 0,8156 ACOT6 acyl-CoA
thioesterase 6
226876_at 204,7 149,3 175,7 208,7 221,4 228,0 319,9 272,6 0,8152 FAM101
B
family with sequence similarity 101,
member B
1553117_a_at 103,9 64,7 92,5 102,7 104,9 116,3 125,3 167,0 0,8152 STK38
serine/threonine kinase 38
222834_s_at 163,9 219,8 192,0 179,7 222,9 260,1 226,2 294,0 0,8150 GNG12
guanine nucleotide binding protein (G protein), gamma 12
238553_at 9,2 7,5 5,9 8,3 11,3 11,9 12,8 12,9 0,8150
CTGLF1 ///
BMS1P5 ///
CTGLF6 ///
CTGLF7
centaurin, gamma-like family, member 1 /// BMS1 pseudogene
5 /// centaurin, gamma-like family,
member 6 /// centaurin, gamma-
like family, member 7

Tablas Suplementarias
158
216115_at 29,4 18,2 28,4 33,4 32,8 35,3 43,5 38,5 0,8144 NF1
Neurofibromin 1 (neurofibromatosis, von Recklinghausen
disease, Watson disease)
219252_s_at 50,3 69,7 36,0 82,3 86,4 107,3 113,7 95,3 0,8143 GEMIN8
gem (nuclear organelle)
associated protein 8
1566446_at 12,1 7,3 11,8 13,6 20,7 17,2 16,3 30,6 0,8143 --- ---
205128_x_at 270,9 309,0 480,2 388,4 543,6 325,1 768,6 867,8 0,8141 PTGS1
prostaglandin-endoperoxide
synthase 1 (prostaglandin G/H
synthase and cyclooxygenase)
228456_s_at 122,1 157,0 131,7 196,6 203,6 214,1 200,5 196,6 0,8136
LOC149832
hypothetical protein LOC149832
206140_at 7,8 2,2 4,4 6,9 24,6 34,2 23,8 25,5 0,8136 LHX2 LIM homeobox 2
242797_x_at
11,2 8,5 10,1 22,5 26,2 20,5 19,2 30,6 0,8136 --- ---
1554447_at 30,0 33,7 20,0 35,5 32,3 42,5 52,6 51,2 0,8133
LOC554203
hypothetical LOC554203
212012_at 867,1 1523,5 1482,
6 2048,
0 1447,
5 2019,
1 2493,
8 2047,
1 0,8129 PXDN
peroxidasin homolog (Drosophila)
243031_at 14,4 10,0 9,2 22,1 14,9 19,8 40,9 34,7 0,8127 --- ---
213353_at 73,3 64,3 62,8 99,1 89,5 104,8 109,3 97,8 0,8126 ABCA5 ATP-binding
cassette, sub-family A (ABC1), member 5
209006_s_at 178,7 125,5 80,5 79,3 258,3 232,2 394,9 431,1 0,8125 C1orf63
chromosome 1 open reading frame 63
202085_at 222,0 239,9 161,7 186,2 301,0 295,3 493,9 728,9 0,8120 TJP2 tight junction protein 2 (zona occludens 2)
225688_s_at 159,5 271,1 122,5 173,4 461,0 334,2 574,0 483,7 0,8120 PHLDB2
pleckstrin homology-like domain, family
B, member 2 221010_s_at 4,6 4,7 20,6 15,6 17,1 27,0 20,7 22,6 0,8118 SIRT5
sirtuin (silent mating type information
regulation 2 homolog) 5 (S.
cerevisiae) 228326_at 32,9 46,6 34,2 47,0 46,4 53,2 57,4 51,6 0,8118 WDR27
WD repeat domain 27
239034_at 25,2 28,2 16,0 30,1 36,9 31,8 43,5 43,8 0,8113 CXorf24 chromosome X open
reading frame 24
204036_at 1275,1 1943,0 1376,
9 2169,
7 2248,
4 2096,
9 2402,
0 2329,
4 0,8112 LPAR1
lysophosphatidic acid receptor 1
235077_at 36,1 53,7 36,3 32,5 83,2 49,9 146,0 184,2 0,8108 MEG3 maternally
expressed 3
204460_s_at 169,4 162,4 186,9 230,0 195,4 238,1 202,5 260,7 0,8107 RAD1
RAD1 homolog (S. pombe)
230782_at 16,2 19,6 24,9 26,4 29,8 34,3 32,8 26,8 0,8106 SORD sorbitol
dehydrogenase
214752_x_at 675,7 985,0
1297,0
1287,7
1076,1
1496,5
1365,2
1435,6
0,8106 FLNA filamin A, alpha (actin binding protein 280)
37170_at 13,4 14,9 14,7 16,2 18,9 14,3 20,8 21,2 0,8104 BMP2K BMP2 inducible
kinase
238563_at 27,9 26,0 32,9 39,1 32,1 31,1 51,0 65,4 0,8104 --- ---

Tablas Suplementarias
159
243659_at 59,9 11,3 51,2 21,5 133,0 97,5 160,3 145,1 0,8102 --- ---
226736_at 397,8 235,9 393,4 409,4 718,1 582,6 662,2 637,5 0,8099 CHURC
1 churchill domain
containing 1
235706_at 145,5 755,9 896,9 811,8 722,8 873,1 1156,
3 1115,
2 0,8093 CPM carboxypeptidase M
231262_at 23,9 35,6 59,6 63,3 44,9 80,9 82,3 66,8 0,8090 --- ---
205705_at 16,9 16,3 15,1 22,4 28,6 23,5 22,7 29,5 0,8087 ANKRD
26 ankyrin repeat
domain 26
232722_at 8,5 6,4 12,3 20,0 20,8 18,0 18,7 19,5 0,8086
RNASET2 ///
LOC100131869
ribonuclease T2 /// hypothetical protein
LOC100131869
209288_s_at 148,9 292,4 183,3 275,4 312,0 259,1 347,0 527,1 0,8085
CDC42EP3
CDC42 effector protein (Rho GTPase
binding) 3
233518_at 35,4 23,2 10,6 24,2 58,8 48,9 59,0 83,9 0,8082 --- ---
1557814_a_at
41,7 50,1 61,4 92,2 113,2 130,2 119,0 91,2 0,8081 --- ---
239847_at 10,6 16,1 13,7 10,6 18,9 24,2 34,8 24,5 0,8080 --- ---
204818_at 6,1 9,2 37,6 17,3 15,8 25,6 57,4 60,4 0,8079 HSD17B
2
hydroxysteroid (17-beta) dehydrogenase
2
215435_at 12,3 15,3 12,6 16,4 14,2 19,4 47,2 42,4 0,8078 --- ---
219935_at 777,5 960,8 565,1 833,1 1076,
3 1019,
9 2308,
5 2164,
4 0,8071
ADAMTS5
ADAM metallopeptidase
with thrombospondin
type 1 motif, 5 (aggrecanase-2)
206986_at 8,7 11,5 8,4 14,2 11,5 16,0 15,8 14,7 0,8067 FGF18 fibroblast growth
factor 18
1560288_ at 1,7 1,5 4,1 1,9 13,3 10,6 11,6 9,7 0,8065 --- ---
213422_s_at 1694,9 2251,3
2441,8
2422,0
2712,1
3307,6
3199,0
2647,4
0,8065 MXRA8 matrix-remodelling
associated 8
242722_at 124,6 130,3 145,2 289,3 217,3 291,8 333,2 803,7 0,8062 LMO7 LIM domain 7
239476_at 31,2 42,2 22,1 24,0 51,3 46,1 85,7 82,9 0,8061 --- ---
221590_s_at 38,4 50,7 77,6 62,5 91,1 87,1 87,5 80,1 0,8057 --- ---
202323_s_at 155,6 137,3 114,8 183,2 198,9 159,6 224,3 253,4 0,8055 ACBD3
acyl-Coenzyme A binding domain
containing 3
204028_s_at
306,3 511,7 330,3 475,4 587,8 658,1 1221,7
813,8 0,8054 RABGAP1
RAB GTPase activating protein 1
242608_x_at 135,9 165,5 189,9 142,2 203,3 202,0 198,8 218,8 0,8053
C14orf44
Chromosome 14 open reading frame
44
232796_at 11,7 12,2 10,6 17,6 22,8 45,9 36,4 29,6 0,8047 --- ---
242829_x_at 20,2 26,4 33,0 10,1 32,4 50,4 70,1 58,4 0,8047 FBXL3
F-box and leucine-rich repeat protein 3
236484_at 20,2 17,9 16,2 26,1 23,6 41,4 30,3 35,9 0,8047 --- ---
224922_at 202,6 225,8 168,3 181,8 215,7 285,5 289,9 323,9 0,8046 CSNK2
A2
casein kinase 2, alpha prime polypeptide
217795_s_at 712,7 813,3 617,1 743,5 805,8 862,8 916,8 990,7 0,8043 TMEM43
transmembrane protein 43
222312_s_at 18,2 31,0 44,1 24,9 37,9 42,5 48,2 49,1 0,8036 --- ---
1557905_s_at 379,3 300,5 442,6 533,8 499,3 582,4 497,7 559,6 0,8035 CD44
CD44 molecule (Indian blood group)

Tablas Suplementarias
160
235045_at 48,6 56,1 47,6 52,9 49,2 65,1 65,4 68,3 0,8034 RBM7 RNA binding motif
protein 7
215780_s_at 216,5 259,9 166,4 308,6 238,5 280,6 405,1 435,0 0,8033
SET /// hCG_1644608
SET nuclear oncogene /// SET
translocation (myeloid leukemia-
associated) pseudogene
225913_at 307,7 674,4 559,4 544,7 716,7 625,9 983,5 827,3 0,8033 SGK269 NKF3 kinase family
member
225241_at 635,2 658,3 735,7 1220,
0 1386,
9 1582,
2 2180,
2 1250,
5 0,8033 CCDC80
coiled-coil domain containing 80
216493_s_at 2,1 4,4 4,9 12,9 4,8 17,1 15,6 13,3 0,8033
IGF2BP3 ///
LOC645468
insulin-like growth factor 2 mRNA
binding protein 3
228647_at 44,8 61,9 74,0 86,8 79,3 86,5 105,3 80,5 0,8027 LOC100049716
hypothetical protein LOC100049716
222576_s_at 49,7 36,7 50,1 54,5 53,3 64,2 71,5 60,3 0,8027 EIF2C1
eukaryotic translation initiation
factor 2C, 1
222380_s_at 38,3 62,9 55,1 70,5 115,0 56,3 139,0 126,2 0,8023 PDCD6
Aryl-hydrocarbon receptor repressor
204063_s_at 48,3 50,7 54,3 49,6 61,0 73,0 66,4 62,5 0,8023 ULK2
unc-51-like kinase 2 (C. elegans)
243158_at 22,2 35,1 36,8 35,8 39,7 33,5 48,8 44,4 0,8022 --- ---
229622_at 202,1 238,4 281,6 242,3 445,5 278,1 779,5 613,7 0,8021 FAM132
B
family with sequence similarity 132,
member B
1566798_at 1,9 1,1 3,0 3,1 2,5 10,3 6,5 8,0 0,8021
SLC35E1
solute carrier family 35, member E1
219654_at 148,2 133,0 114,2 144,1 174,0 168,3 303,3 448,4 0,8018 PTPLA
protein tyrosine phosphatase-like (proline instead of catalytic arginine),
member A
226995_at 118,5 151,1 129,1 128,9 230,3 213,1 186,0 226,2 0,8018 C21orf8
6
Chromosome 21 open reading frame
86
205871_at 7,3 13,4 5,2 12,3 14,3 16,1 35,8 25,6 0,8016
PLGLB2 ///
PLGLB1 ///
PLGLA ///
LOC100134363
/// LOC100134366
plasminogen-like B2 /// plasminogen-like B1 /// plasminogen-
like A /// hypothetical protein
LOC100134363 /// hypothetical protein
LOC100134366
202975_s_at 732,6 380,1 357,5 649,3
1066,4
889,9 1301,
6 1162,
0 0,8016
RHOBTB3
Rho-related BTB domain containing 3
211986_at 1518,2 1536,4 1919,
4 1770,
9 1701,
4 1662,
0 2488,
1 2664,
3 0,8015 AHNAK
AHNAK nucleoprotein

Tablas Suplementarias
161
212200_at 143,7 155,6 164,7 178,4 168,3 169,9 178,9 173,1 0,8015 ANKLE2 ankyrin repeat and
LEM domain containing 2
239227_at 26,6 25,8 24,8 29,8 28,4 31,6 56,6 48,2 0,8013 --- ---
1552735_at 19,4 9,3 18,5 16,0 63,2 37,6 99,5 65,1 0,8012
PCDHGA4
protocadherin gamma subfamily A,
4
235660_at 30,8 31,1 18,9 39,7 34,7 44,7 51,1 47,8 0,8009 --- ---
235490_at 4,0 2,6 2,7 5,6 5,2 10,5 13,6 8,2 0,8007 TMEM10
7 transmembrane
protein 107
1552514_at 4,8 5,1 7,2 4,9 7,1 8,0 9,8 7,7 0,8007
WBP2NL
WBP2 N-terminal like
201349_at 266,9 405,1 359,1 434,5 503,0 420,8 448,0 506,0 0,8003 SLC9A3
R1
solute carrier family 9 (sodium/hydrogen exchanger), member
3 regulator 1
1552774_a_at 9,4 20,0 8,6 12,0 33,1 20,8 39,6 35,2 0,8003
SLC25A27
solute carrier family 25, member 27
242671_at 38,6 40,6 11,2 7,5 65,1 62,7 134,2 136,4 0,8001 --- ---
210117_at 64,3 74,4 57,2 38,2 26,6 39,8 37,6 34,3 -0,8005 SPAG1 sperm associated
antigen 1
221620_s_at
269,9 227,3 182,8 196,0 173,8 161,6 149,4 185,8 -0,8007 APOO apolipoprotein O
235471_at 106,7 99,1 128,9 85,2 64,2 82,7 66,3 58,1 -0,8008 C10orf7
2
Chromosome 10 open reading frame
72
228740_at 35,6 30,6 30,3 22,3 25,3 30,5 23,1 13,9 -0,8010 --- ---
202886_s_at 179,7 109,4 145,3 119,6 101,0 86,8 74,8 99,5 -0,8010
PPP2R1B
protein phosphatase 2 (formerly 2A),
regulatory subunit A, beta isoform
235197_s_at 85,3 103,1 68,9 83,1 59,5 38,7 64,1 45,4 -0,8013 OSTM1
osteopetrosis associated
transmembrane protein 1
203980_at 5411,0 3398,7 6715,
7 3056,
7 4782,
0 873,9 525,3 144,5 -0,8013 FABP4
fatty acid binding protein 4, adipocyte
219295_s_at 2017,4 1263,1 950,4 930,7 942,9 493,2 828,7 717,3 -0,8015
PCOLCE2
procollagen C-endopeptidase
enhancer 2
209276_s_at 2414,8 1918,4
1594,4
2190,4
1954,6
1694,0
1348,5
1184,7
-0,8017 GLRX glutaredoxin
(thioltransferase)
204784_s_at 95,1 74,5 84,9 65,3 73,3 55,9 62,3 64,4 -0,8023 MLF1
myeloid leukemia factor 1
225599_s_at 341,5 278,1 251,6 295,2 271,7 258,4 237,9 230,9 -0,8024
LOC286144
hypothetical LOC286144
214085_x_at 1334,0 1632,7 831,1
1075,9
519,8 525,9 482,6 698,0 -0,8027 GLIPR1 GLI pathogenesis-related 1 (glioma)
212813_at 572,0 599,6 681,5 622,4 418,2 356,6 437,9 344,9 -0,8033 JAM3 /// LOC100133502
junctional adhesion molecule 3 ///
hypothetical protein LOC100133502

Tablas Suplementarias
162
219155_at 442,8 356,9 187,6 232,7 133,3 171,4 199,6 144,9 -0,8037 PITPNC
1
phosphatidylinositol transfer protein, cytoplasmic 1
209242_at 49,4 22,4 13,8 17,9 15,0 13,6 4,1 8,7 -0,8041 PEG3 paternally expressed
3
203880_at 543,1 348,4 232,7 254,3 230,7 188,4 177,4 210,8 -0,8041 COX17
COX17 cytochrome c oxidase assembly
homolog (S. cerevisiae)
219686_at 61,6 42,3 45,3 55,9 30,3 12,7 34,3 16,1 -0,8041 STK32B serine/threonine
kinase 32B
209849_s_at 183,1 154,1 124,8 154,7 119,4 121,8 120,2 119,0 -0,8054 RAD51C
RAD51 homolog C (S. cerevisiae)
205913_at 359,0 153,0 323,0 285,8 267,5 86,0 26,8 9,4 -0,8060 PLIN perilipin
226712_at 274,9 254,3 250,1 206,7 206,8 243,1 191,5 202,4 -0,8064 SSR1 signal sequence receptor, alpha
218508_at 143,0 127,0 161,3 123,3 134,2 108,4 106,3 86,6 -0,8071 DCP1A DCP1 decapping
enzyme homolog A (S. cerevisiae)
230925_at 228,9 248,9 181,5 141,1 244,4 132,1 111,5 70,6 -0,8071 APBB1I
P
amyloid beta (A4) precursor protein-
binding,
235333_at 118,2 87,6 83,2 34,6 39,6 46,2 45,5 44,2 -0,8077 B4GALT
6
UDP-Gal:betaGlcNAc beta 1,4-
galactosyltransferase, polypeptide 6
225784_s_at 73,8 73,8 43,8 71,5 54,5 30,0 24,0 37,6 -0,8077
KIAA1166
KIAA1166
206697_s_at 639,9 169,9 306,1 73,3 153,6 56,4 30,5 5,4 -0,8077 HP haptoglobin
220266_s_at 50,8 38,0 38,7 42,0 35,3 39,2 30,7 31,6 -0,8081 KLF4
Kruppel-like factor 4 (gut)
225502_at 52,9 31,2 24,6 16,1 13,8 21,6 9,6 15,2 -0,8081 DOCK8 dedicator of
cytokinesis 8
204222_s_at 793,0 547,4 699,0 749,5 364,1 405,5 265,1 380,5 -0,8091 GLIPR1
GLI pathogenesis-related 1 (glioma)
203572_s_at 278,2 315,6 325,7 290,2 254,7 244,1 220,8 234,2 -0,8093 TAF6
TAF6 RNA polymerase II, TATA box binding protein
(TBP)-associated factor, 80kDa
202283_at 1114,1 785,2 311,5 393,4 528,3 463,6 279,5 167,1 -0,8094 SERPIN
F1
serpin peptidase inhibitor, clade F
(alpha-2 antiplasmin, pigment epithelium
derived factor), member 1
200699_at 2355,0 2497,5 2183,
0 2425,
0 1964,
4 2175,
9 1928,
5 1916,
5 -0,8106 KDELR2
KDEL (Lys-Asp-Glu-Leu) endoplasmic reticulum protein
retention receptor 2

Tablas Suplementarias
163
207551_s_at 209,7 204,7 184,0 189,1 141,4 161,7 140,1 166,3 -0,8118 MSL3L1
male-specific lethal 3-like 1 (Drosophila)
33307_at 151,0 115,9 94,3 105,6 84,8 63,1 73,3 88,5 -0,8120 CTA-
126B4.3 CGI-96 protein
214875_x_at 714,4 538,3 433,5 530,1 482,3 494,1 268,9 364,3 -0,8121 APLP2
amyloid beta (A4) precursor-like
protein 2
220495_s_at 930,1 1062,2 933,7 818,8 721,6 622,9 719,0 743,0 -0,8123
TXNDC15
thioredoxin domain containing 15
218922_s_at 11,0 7,5 5,9 3,4 5,4 4,3 2,4 4,3 -0,8127 LASS4
LAG1 homolog, ceramide synthase 4
244276_at 115,6 68,8 128,2 32,8 75,9 25,4 15,2 11,1 -0,8132 KLB klotho beta
235092_at 40,8 34,4 19,5 28,1 24,6 20,2 16,6 20,4 -0,8137 --- ---
206941_x_at 43,8 35,6 16,1 19,8 17,9 27,2 5,2 8,9 -0,8138
SEMA3E
sema domain
213285_at 181,2 119,0 94,8 91,4 92,5 81,4 94,1 63,0 -0,8138 TMEM30
B transmembrane
protein 30B
203679_at 259,3 300,5 244,7 254,4 224,0 195,4 209,1 214,7 -0,8140 TMED1
transmembrane emp24 protein
transport domain containing 1
222531_s_at 366,9 292,6 345,8 315,8 295,5 308,7 265,3 254,4 -0,8143
C14orf108
chromosome 14 open reading frame
108
209302_at 673,8 494,8 374,6 371,6 386,8 362,6 363,1 306,4 -0,8144 POLR2H polymerase (RNA) II
(DNA directed) polypeptide H
202966_at 24,5 11,6 19,9 16,6 13,9 7,1 5,2 8,3 -0,8145 CAPN6 calpain 6
227474_at 242,9 281,3 177,9 224,0 156,4 139,6 161,4 148,9 -0,8149 LOC654
433 Hypothetical LOC654433
231775_at 51,8 48,2 36,7 51,0 34,3 30,3 28,5 32,5 -0,8165 TNFRSF
10A
tumor necrosis factor receptor
superfamily, member 10a
238821_at 32,3 28,3 23,5 24,0 25,5 24,2 16,6 21,8 -0,8171 CSTF2 Cleavage stimulation
factor, 3' pre-RNA, subunit 2, 64kDa
203662_s_at 16,9 16,3 15,6 13,7 11,0 13,2 13,5 4,8 -0,8171 TMOD1 tropomodulin 1
200795_at 42,2 24,3 31,9 5,3 16,8 14,7 4,9 7,9 -0,8182 SPARC
L1 SPARC-like 1 (mast9,
hevin)
220647_s_at 360,4 299,4 275,4 342,1 239,5 278,4 193,6 215,0 -0,8188
CHCHD8
coiled-coil-helix-coiled-coil-helix
domain containing 8
212953_x_at 2146,4 2179,0
2739,5
1833,8
1999,6
1806,6
1112,0
964,3 -0,8190 CALR calreticulin
203800_s_at 667,3 594,2 548,6 377,2 493,6 367,7 421,5 411,4 -0,8196 MRPS14
mitochondrial ribosomal protein
S14
211564_s_at 266,5 301,7 251,7 263,4 197,6 229,2 187,2 209,4 -0,8203 PDLIM4
PDZ and LIM domain 4

Tablas Suplementarias
164
205282_at 137,5 175,7 87,8 88,2 73,4 56,7 45,4 70,5 -0,8207 LRP8
low density lipoprotein receptor-
related protein 8, apolipoprotein e
receptor
203382_s_at 481,9 331,6 195,0 137,1 256,2 235,2 110,7 68,3 -0,8217 APOE apolipoprotein E
209173_at 68,0 90,8 48,4 23,3 5,6 4,8 10,6 17,5 -0,8221 AGR2 anterior gradient
homolog 2 (Xenopus laevis)
204813_at 110,3 96,9 113,0 86,7 96,6 97,3 60,8 39,7 -0,8248 MAPK10 mitogen-activated protein kinase 10
217736_s_at 1018,9 757,2 613,9 605,3 529,7 525,1 523,7 536,2 -0,8253
EIF2AK1
eukaryotic translation initiation factor 2-alpha kinase
1
207316_at 119,1 77,5 80,5 108,2 49,0 38,5 37,0 44,8 -0,8262 HAS1 hyaluronan synthase
1
223340_at 209,0 203,6 126,0 107,6 107,6 104,1 94,8 108,9 -0,8262 SPG3A spastic paraplegia
3A (autosomal dominant)
220264_s_at 23,6 31,8 31,4 22,8 22,8 7,6 7,4 11,0 -0,8270 GPR107
G protein-coupled receptor 107
227889_at 50,1 40,4 39,1 51,6 39,5 30,6 28,9 23,4 -0,8277 LPCAT2 lysophosphatidylcholine acyltransferase
2
215311_at 10,5 7,1 7,3 3,6 3,6 2,0 1,3 4,4 -0,8279 NTRK3 neurotrophic
tyrosine kinase, receptor, type 3
201282_at 226,2 176,6 200,1 152,2 164,1 185,4 137,5 115,8 -0,8281 OGDH
oxoglutarate (alpha-ketoglutarate)
dehydrogenase (lipoamide)
229352_at 56,7 89,9 65,7 57,5 37,0 33,1 24,1 30,6 -0,8291 NOX5 /// SPESP1
NADPH oxidase, EF-hand calcium
binding domain 5 /// sperm equatorial
segment protein 1
209625_at 115,1 99,7 98,4 83,5 85,5 64,1 73,8 82,7 -0,8297 PIGH
phosphatidylinositol glycan anchor
biosynthesis, class H
221901_at 329,2 359,0 480,9 332,7 259,6 168,4 148,9 121,3 -0,8300 LL22NC
03-75B3.6
KIAA1644 protein
1562957_at 13,5 12,4 11,6 5,1 4,3 2,4 7,3 3,1 -0,8319 --- ---
201175_at 965,2 1053,4 894,2 741,6 703,4 577,5 720,4 687,7 -0,8330 TXNDC1
4 thioredoxin domain
containing 14
228635_at 10,2 5,6 3,8 4,3 3,3 2,2 0,8 2,9 -0,8340 PCDH10 protocadherin 10
205110_s_at 55,3 30,3 29,7 45,9 24,8 17,5 11,7 14,1 -0,8341 FGF13
fibroblast growth factor 13

Tablas Suplementarias
165
206932_at 98,7 40,9 51,8 63,2 35,0 24,7 11,9 21,2 -0,8343 CH25H cholesterol 25-
hydroxylase
202345_s_at 2720,9 1131,1
1611,0
1281,6
1283,4
526,4 257,2 670,9 -0,8350
FABP5 ///
FABP5L7 ///
FABP5L2
fatty acid binding protein 5 (psoriasis-
associated)
236398_s_at
8,5 7,8 4,9 5,0 3,6 5,1 4,6 3,3 -0,8362 --- ---
235573_at 27,5 30,5 23,1 28,6 24,0 21,2 21,9 17,3 -0,8363 --- ---
224824_at 350,9 244,4 242,1 232,6 147,8 170,0 151,3 183,5 -0,8364 FAM36A family with sequence
similarity 36, member A
1555740_a_at 10,0 9,5 3,1 3,7 2,8 1,4 1,2 2,5 -0,8365 MRAP
melanocortin 2 receptor accessory
protein
209030_s_at 89,1 72,1 85,6 63,9 42,4 33,4 53,9 42,1 -0,8370 CADM1
cell adhesion molecule 1
208892_s_at 597,9 614,5 262,2 279,2 371,4 169,3 197,6 186,7 -0,8370 DUSP6
dual specificity phosphatase 6
202348_s_at 494,6 412,1 315,8 324,3 260,4 247,4 263,2 285,9 -0,8374 TOR1A
torsin family 1, member A (torsin A)
227804_at 93,5 80,7 66,7 88,4 64,2 71,7 44,2 48,3 -0,8386 TLCD1 TLC domain containing 1
210501_x_at 1412,3 1469,7
1394,2
1454,1
1437,8
1260,2
1180,1
1159,1
-0,8390 EIF3K eukaryotic
translation initiation factor 3, subunit K
222317_at 50,5 22,4 37,2 17,6 28,8 7,8 2,5 8,4 -0,8394 --- ---
209550_at 998,0 519,9 364,4 318,0 188,4 178,4 192,9 160,9 -0,8405 NDN necdin homolog
(mouse)
200617_at 1010,8 808,0 684,0 772,7 746,8 614,6 620,5 629,1 -0,8409 KIAA01
52 KIAA0152
205347_s_at 116,5 93,5 58,6 47,3 25,6 39,5 24,9 41,6 -0,8419
TMSL8 ///
MGC39900
thymosin-like 8 /// thymosin beta15b
210792_x_at 271,1 159,8 142,6 141,1 130,1 67,2 101,2 91,6 -0,8419 SIVA1
SIVA1, apoptosis-inducing factor
209552_at 25,9 22,8 20,0 10,0 12,3 12,9 13,0 10,6 -0,8422 PAX8 paired box 8
224783_at 154,8 130,2 163,8 131,6 91,5 101,4 92,4 93,7 -0,8424 FAM100
B
family with sequence similarity 100,
member B
213757_at 974,1 397,2 441,4 559,8 294,2 305,0 140,2 129,7 -0,8442 --- ---
214920_at 55,0 49,9 30,3 11,5 29,0 18,2 17,2 11,7 -0,8446 THSD7A thrombospondin,
type I, domain containing 7A
1564475_s_at 34,6 22,7 23,2 19,1 17,2 14,1 18,7 14,1 -0,8456
LOC728723
hypothetical protein LOC728723
1554277_s_at 6,7 6,1 1,8 2,0 1,3 2,0 1,1 0,6 -0,8472 FANCM
Fanconi anemia, complementation
group M
237413_at 15,1 16,4 13,7 4,7 5,7 4,0 8,1 1,6 -0,8477 --- ---
225441_x_at 699,6 659,2 493,1 512,0 448,4 483,6 304,7 447,8 -0,8480 LSMD1
LSM domain containing 1

Tablas Suplementarias
166
230179_at 35,2 33,0 27,1 19,6 17,1 16,0 17,6 19,9 -0,8482 LOC285
812 hypothetical protein
LOC285812
204473_s_at 64,7 59,6 55,5 54,9 61,1 44,8 46,0 46,6 -0,8489 ZNF592
zinc finger protein 592
229059_at 113,4 79,5 106,0 78,8 49,5 75,1 53,3 42,2 -0,8489 C9orf10
9 chromosome 9 open
reading frame 109
201077_s_at 1492,7 1364,2
1198,2
1119,9
1017,4
988,5 954,4 1103,
2 -0,8490 NHP2L1
NHP2 non-histone chromosome protein
2-like 1 (S. cerevisiae)
205677_s_at 250,6 216,5 206,2 138,2 117,2 105,9 100,5 143,0 -0,8495 DLEU1
deleted in lymphocytic leukemia, 1
213706_at 242,7 117,0 191,3 63,9 153,9 12,8 10,6 1,5 -0,8505 GPD1 glycerol-3-phosphate
dehydrogenase 1 (soluble)
206243_at 1499,0 1198,6 892,2 701,3 1027,
6 500,6 751,8 478,1 -0,8508 TIMP4
TIMP metallopeptidase
inhibitor 4
210416_s_at 155,8 146,2 146,8 132,4 102,0 134,8 112,0 100,6 -0,8508 CHEK2
CHK2 checkpoint homolog (S. pombe)
227434_at 11,6 16,4 10,4 8,4 5,8 3,7 5,2 4,9 -0,8508 WBSCR
17
Williams-Beuren syndrome
chromosome region 17
218866_s_at 181,0 153,6 180,6 149,8 142,3 146,0 83,1 107,5 -0,8508 POLR3K
polymerase (RNA) III (DNA directed)
polypeptide K, 12.3 kDa
213830_at 53,1 31,5 17,7 12,8 18,8 16,5 5,0 6,7 -0,8510 TRD@ T cell receptor delta
locus
200845_s_at 2504,6 2276,0
1970,3
2305,3
2060,5
1882,2
1696,7
1859,5
-0,8511 PRDX6 peroxiredoxin 6
201281_at 580,5 464,8 453,9 540,5 374,4 411,7 317,1 338,8 -0,8513 ADRM1 adhesion regulating
molecule 1
244741_s_at 324,8 320,4 145,2 143,1 132,2 148,0 112,0 89,8 -0,8524
MGC9913
hypothetical protein MGC9913
212680_x_at 465,8 472,4 483,4 500,8 360,5 328,6 276,5 322,9 -0,8539
PPP1R14B
protein phosphatase 1, regulatory
(inhibitor) subunit 14B
217979_at 161,7 92,3 143,5 78,5 43,5 23,7 27,1 44,6 -0,8545 TSPAN1
3 tetraspanin 13
222532_at 273,4 245,4 193,9 194,2 140,8 171,0 145,8 165,5 -0,8556 SRPRB signal recognition particle receptor, B
subunit
200825_s_at 726,7 557,3 724,9 582,5 600,0 513,1 463,5 369,6 -0,8562 HYOU1
hypoxia up-regulated 1
213115_at 136,9 108,4 111,8 82,3 85,5 85,9 87,1 77,4 -0,8567 ATG4A ATG4 autophagy
related 4 homolog A (S. cerevisiae)
236236_at 24,8 26,2 20,8 9,8 9,9 14,0 8,3 9,4 -0,8574 --- ---
225128_at 525,0 415,0 463,5 451,6 333,1 369,1 296,2 340,5 -0,8582 KDELC2 KDEL (Lys-Asp-Glu-
Leu) containing 2
224179_s_at 15,8 18,4 11,2 13,8 3,9 10,0 6,0 2,3 -0,8592 MIOX
myo-inositol oxigenase

Tablas Suplementarias
167
210982_s_at 32,3 11,4 17,2 13,1 5,7 7,5 3,9 1,5 -0,8600
HLA-DRA
major histocompatibility
complex, class II, DR alpha
208547_at 9,3 12,1 9,6 12,6 6,0 4,2 1,7 1,2 -0,8600 HIST1H
2BB histone cluster 1,
H2bb
203903_s_at 99,0 127,8 57,9 66,2 42,0 41,2 42,1 26,4 -0,8602 HEPH Hephaestin
204865_at 24,0 10,9 12,7 18,1 8,7 7,8 4,5 0,9 -0,8602 CA3 carbonic anhydrase III, muscle specific
204249_s_at 92,4 117,6 57,7 68,9 36,1 36,6 27,4 34,9 -0,8603 LMO2
LIM domain only 2 (rhombotin-like 1)
223644_s_at 106,2 49,5 61,8 48,8 33,0 32,7 23,3 25,0 -0,8603 CRYGS crystallin, gamma S
201740_at 719,3 719,0 625,4 676,2 624,0 648,1 600,8 590,1 -0,8618 NDUFS3
NADH dehydrogenase
(ubiquinone) Fe-S protein 3, 30kDa
(NADH-coenzyme Q reductase)
223092_at 2842,4 2238,3 1780,
6 1486,
8 735,3 900,3
1034,1
1097,2
-0,8620 ANKH ankylosis,
progressive homolog (mouse)
217999_s_at 178,2 112,7 84,8 111,6 73,8 58,4 59,4 58,3 -0,8629 PHLDA1
pleckstrin homology-like domain, family
A, member 1
213524_s_at 1079,6 537,9
1076,3
723,9 561,3 404,6 179,8 83,7 -0,8632 G0S2 G0/G1switch 2
207154_at 44,9 24,0 37,2 19,9 14,3 12,0 10,1 11,8 -0,8639 DIO3 deiodinase,
iodothyronine, type III
225593_at 350,0 348,0 407,5 353,2 256,2 263,5 223,3 205,7 -0,8649 LSM10 LSM10, U7 small
nuclear RNA associated
228782_at 77,8 44,0 34,6 17,5 12,0 32,0 1,0 2,2 -0,8657 SCGB3
A2 secretoglobin, family
3A, member 2
237054_at 20,8 25,1 26,1 21,7 14,8 16,6 10,8 9,9 -0,8669 ENPP5
ectonucleotide pyrophosphatase/phosphodiesterase 5 (putative function)
207396_s_at 385,8 338,5 368,8 327,0 238,4 309,0 243,7 238,7 -0,8679 ALG3
asparagine-linked glycosylation 3
homolog (S. cerevisiae, alpha-1,3-mannosyltransferase
)
205794_s_at 124,1 130,3 82,8 104,5 105,6 88,1 49,4 45,8 -0,8699 NOVA1
neuro-oncological ventral antigen 1
225425_s_at 279,0 320,0 252,3 287,3 220,8 238,5 164,7 172,3 -0,8703 MRPL41
mitochondrial ribosomal protein
L41
204015_s_at 552,7 610,4 584,9 435,6 249,5 258,3 227,1 297,7 -0,8706 DUSP4
dual specificity phosphatase 4
1559399_s_at 74,5 71,3 65,5 42,5 43,6 30,9 32,7 42,7 -0,8708
ZCCHC10
zinc finger, CCHC domain containing
10
228224_at 1073,4 868,7 1146,
9 776,3 622,5 449,4 665,9 247,1 -0,8718 PRELP
proline/arginine-rich end leucine-rich repeat protein

Tablas Suplementarias
168
219508_at 46,0 45,7 36,3 37,5 26,0 12,4 17,3 24,3 -0,8722 GCNT3 glucosaminyl (N-
acetyl) transferase 3, mucin type
213894_at 46,8 40,9 30,5 12,7 17,2 10,3 12,5 13,9 -0,8723 THSD7A thrombospondin,
type I, domain containing 7A
226632_at 246,6 233,0 240,3 263,8 190,6 182,1 152,1 152,0 -0,8727 CYGB Cytoglobin
218008_at 539,3 447,4 367,4 325,5 350,0 342,6 296,7 297,6 -0,8751 C7orf42 chromosome 7 open
reading frame 42
205404_at 1326,9 765,5 432,2 449,7 391,4 171,9 248,1 131,3 -0,8759 HSD11B
1
hydroxysteroid (11-beta) dehydrogenase
1
227742_at 453,7 312,7 183,0 161,3 72,0 35,9 88,3 72,0 -0,8764 CLIC6 chloride intracellular
channel 6
204595_s_at 3601,6 2758,6
1326,1
1410,3
1017,4
414,3 1095,
0 411,8 -0,8785 STC1 stanniocalcin 1
226382_at 31,2 23,0 24,5 25,6 15,1 20,8 15,8 7,1 -0,8797 LOC283
070 hypothetical protein
LOC283070
1553602_at 77,0 89,7 83,6 54,7 73,4 12,9 23,9 8,7 -0,8799 MUCL1 mucin-like 1
200078_s_at 691,2 704,3 622,7 588,0 417,9 505,4 496,4 442,1 -0,8806
ATP6V0B
ATPase, H+ transporting,
lysosomal 21kDa, V0 subunit b
202771_at 695,4 649,0 568,3 606,5 569,8 519,2 467,6 536,2 -0,8826 FAM38A family with sequence
similarity 38, member A
217830_s_at 163,8 151,4 134,5 147,5 127,2 119,8 131,2 115,9 -0,8839 NSFL1C
NSFL1 (p97) cofactor (p47)
203029_s_at 38,7 43,8 38,0 43,2 13,4 12,8 2,5 7,6 -0,8845 PTPRN2
protein tyrosine phosphatase,
receptor type, N polypeptide 2
200096_s_at 3034,6 3049,2
2846,2
2900,9
2664,8
2273,7
2621,4
2311,9
-0,8867 ATP6V0
E1
ATPase, H+ transporting,
lysosomal 9kDa, V0 subunit e1
209859_at 64,5 37,0 49,5 34,3 34,7 13,1 21,3 15,8 -0,8875 TRIM9 tripartite motif-
containing 9
218516_s_at 379,9 299,1 349,1 333,8 266,4 231,4 231,4 231,2 -0,8880 IMPAD1
inositol monophosphatase
domain containing 1
203680_at 295,4 175,9 164,7 128,1 141,6 95,5 76,7 88,0 -0,8881 PRKAR
2B
protein kinase, cAMP-dependent, regulatory, type II,
beta
201147_s_at 292,3 201,1 255,8 250,2 164,8 164,5 80,4 103,6 -0,8889 TIMP3
TIMP metallopeptidase
inhibitor 3 (Sorsby fundus dystrophy,
pseudoinflammatory)
209301_at 323,4 196,5 157,8 115,8 68,7 36,0 44,2 65,6 -0,8893 CA2 carbonic anhydrase
II

Tablas Suplementarias
169
52837_at 546,0 545,0 409,9 313,4 300,6 255,8 329,6 238,7 -0,8899 LL22NC
03-75B3.6
KIAA1644 protein
201525_at 2596,7 2439,4 964,7 799,6 1014,7
852,6 237,2 254,4 -0,8932 APOD apolipoprotein D
224184_s_at 421,6 456,5 383,6 287,9 233,4 197,9 290,2 76,0 -0,8936 BOC
Boc homolog (mouse)
221156_x_at 424,4 356,9 338,9 304,2 190,4 163,5 255,0 157,7 -0,8943 CCPG1
cell cycle progression 1
231688_at 73,9 42,3 34,0 54,4 36,9 17,2 6,1 3,0 -0,8975 --- ---
225402_at 187,9 160,9 174,9 116,2 111,0 128,7 114,7 74,3 -0,8977 TP53RK TP53 regulating
kinase
204401_at 25,2 27,0 18,9 11,8 7,7 13,7 3,4 6,2 -0,9018 KCNN4
potassium intermediate/small
conductance calcium-activated
channel, subfamily N, member 4
201150_s_at 673,1 622,0 517,5 567,6 531,8 508,2 369,6 431,2 -0,9037 TIMP3
TIMP metallopeptidase
inhibitor 3 (Sorsby fundus dystrophy,
pseudoinflammatory)
220980_s_at 625,5 574,2 546,7 548,7 496,1 528,2 517,5 455,3 -0,9041 ADPGK
ADP-dependent glucokinase
218736_s_at 110,1 63,4 81,8 68,6 62,7 50,1 43,5 14,3 -0,9054 PALMD palmdelphin
211991_s_at 57,9 43,5 33,4 21,6 11,6 9,4 12,2 12,3 -0,9064
HLA-DPA1
major histocompatibility
complex, class II, DP alpha 1
202920_at 101,6 133,2 79,7 94,3 48,2 40,8 33,7 25,1 -0,9073 ANK2 ankyrin 2, neuronal
223312_at 228,8 186,1 208,8 177,8 137,4 151,1 113,7 133,9 -0,9074 C2orf7 chromosome 2 open
reading frame 7
218290_at 126,9 137,2 120,0 121,7 100,3 94,0 97,9 94,1 -0,9083 PLEKHJ
1
pleckstrin homology domain containing, family J member 1
1552316_a_at 4,9 5,2 3,1 2,2 3,4 1,9 1,1 1,1 -0,9111 GIMAP1
GTPase, IMAP family member 1
230438_at 319,7 278,0 263,5 246,5 239,1 215,9 241,0 206,6 -0,9130 TBX15 T-box 15
218447_at 474,6 415,9 296,6 329,2 258,2 238,2 205,7 234,5 -0,9140 C16orf6
1
chromosome 16 open reading frame
61
236313_at 94,6 115,5 88,5 80,6 53,0 45,5 49,8 40,4 -0,9165 CDKN2
B
cyclin-dependent kinase inhibitor 2B
(p15, inhibits CDK4)
204596_s_at 3289,3 1573,2
2010,6
1337,3
990,5 413,5 524,8 213,7 -0,9174 STC1 stanniocalcin 1
226932_at 167,5 172,8 137,9 136,6 99,3 125,6 102,7 59,6 -0,9186 --- ---
224376_s_at 1146,5 1158,3
1116,1
986,1 1036,
9 1008,
2 801,5 819,5 -0,9197
C20orf24
chromosome 20 open reading frame
24

Tablas Suplementarias
170
202655_at 1411,0 1281,3 911,0 985,6 818,0 737,2 758,3 685,1 -0,9198 ARMET arginine-rich,
mutated in early stage tumors
205932_s_at 95,9 102,2 106,2 66,7 74,5 61,7 47,5 41,7 -0,9204 MSX1 msh homeobox 1
225511_at 9,8 10,1 10,4 5,1 2,6 2,5 1,9 1,3 -0,9241 GPRC5
B
G protein-coupled receptor, family C, group 5, member B
206707_x_at 178,2 167,6 147,1 112,0 100,7 60,9 94,6 72,7 -0,9254 C6orf32
chromosome 6 open reading frame 32
229963_at 102,2 121,5 84,4 72,5 28,5 28,6 17,6 20,8 -0,9306 BEX5 BEX family member
5
203136_at 1823,6 1665,5 1759,
8 1614,
4 1625,
1 1346,
2 1363,
9 1316,
6 -0,9318
RABAC1
Rab acceptor 1 (prenylated)
222657_s_at 174,5 175,8 156,6 133,4 133,8 98,6 121,7 98,4 -0,9337 UBE2W
ubiquitin-conjugating enzyme
E2W (putative)
219440_at 57,7 50,7 58,3 48,1 33,7 28,5 15,6 21,5 -0,9350 RAI2 retinoic acid induced
2
218454_at 115,2 95,8 76,4 82,4 59,0 67,2 59,6 35,2 -0,9395 FLJ2266
2 hypothetical protein
FLJ22662
202634_at 429,4 343,5 346,4 338,7 336,9 278,9 264,4 238,5 -0,9402 POLR2K
polymerase (RNA) II (DNA directed) polypeptide K,
7.0kDa
205348_s_at 112,0 105,5 98,8 100,3 69,1 50,3 51,1 48,3 -0,9500
DYNC1I1
dynein, cytoplasmic 1, intermediate chain
1
225914_s_at 124,0 112,2 113,8 106,6 89,3 97,0 70,6 62,7 -0,9513 CAB39L
calcium binding protein 39-like
205548_s_at 392,0 322,2 344,6 330,3 281,4 231,3 205,5 195,6 -0,9600 BTG3
BTG family, member 3
228614_at 323,9 273,4 276,1 231,9 240,4 205,6 202,7 182,3 -0,9624 LOC205
251 LOC205251
209003_at 381,7 342,5 305,4 292,9 269,0 252,5 238,4 227,6 -0,9769 SLC25A
11
solute carrier family 25 (mitochondrial
carrier; oxoglutarate carrier), member 11

INTRODUCCIÓN
OBJETIVOS
MATERIALES Y MÉTODOS
RESULTADOS
DISCUSIÓN
CONCLUSIONES
TABLAS SUPLEMENTARIAS
REFERENCIAS BIBLIOGRÁFICAS

Referencias Bibliográficas
172
Alaminos M, Sanchez-Quevedo MC, Muñoz-Avila JI, Serrano D, Medialdea S,
Carreras I, Campos A. Construction of a complete rabbit cornea substitute using a
fibrin-agarosa scaffold. Invest. Ophthalmol. Vis. Sci. 2006; 47,3311-3317.
Alaminos M, Garzón I, Sánchez-Quevedo MC, Moreu G, González-Andrades M,
Fernández-Montoya A, Campos A. Time-course study of histological and genetic
patterns of differentiation in human engineered oral mucosa. J Tissue Eng Regen Med.
2007; 1(5): 350–359.
Alaminos M, Sanchez-Quevedo MC, Muñoz-Avila JI, García JM, Crespo PV,
González-Andrades M, Campos A. Evaluation of the viability of cultured corneal
endothelial cells by quantitative electron probe X-ray microanalysis. J Cell Physiol.
2007b; 211(3): 692–698.
Allen JC. Sodium and potassium content and viability of mouse mammary gland tissue
and acini. J Dairy Sci. 1988; 71(3): 633–642.
Alvarez A, Casanova C, García Y. Tratamiento quirúrgico de la osteoartritis de rodilla.
Rev. Cubana Ortop Traumatol. 2004; 18(1):53-59.
Amato PA, Loizzi RF. The identification and localization of actin and actin-like filaments
in lactating guinea pig mammary gland alveolar cells. Cell Motil. 1981; 1(3): 329–347.
Ashkenazi A, Dixit VM. Death receptors: signaling and modulation. 1998. Science
281:1305–1308.
Beattie GM, Butler C, Hayek A. Morphology and function of cultured human fetal
pancreatic cells transplanted into athymic mice: a longitudinal study. Cell Transplant.
1994; 3(5): 421–425.
Bentley G, Greer RB. Homotransplantation of isolated epiphyseal and articular
cartilage chondrocytes into joint surfaces of rabbits. Nature. 1971; 230:385-388.
Bhuyan BK, Loughman BE, Fraser TJ, Day KJ. Comparison of different methods of
determining cell viability after exposure to cytotoxic compounds. Exp Cell Res. 1976;
97(2): 275–280.

Referencias Bibliográficas
173
Brink CB, Pretorius A, van Niekerk BP, Oliver DW, Venter DP. Studies on cellular
resilience and adaptation following acute and repetitive exposure to ozone in cultured
human epithelial (HeLa) cells. Redox Rep. 2008; 13(2):87-100.
Brittberg M, Lindhal A, Nilsson A, Ohlsson C, Isaksson O, Peterson L. Treatment of
deep cartilage defects in the knee with autologous chondrocyte transplantation. N Engl
J Med. 1994; 331: 889-895.
Buckwalter JA. Integration of science in orthopaedic practice: Implications for solving
the problem of articular cartilage repair. J Bone Joint Surgery Am. 2003; 85: 1-7.
Buckwalter JA, Martin J. Degenerative joint desease. Anatomy and physiology of
sinovial joints. Clinical Symposia. 1995. Ciba, vol 47, nº 2.
Buckwalter JA, Amendola A, Clark CR. Cartílago articular y menisco: biología,
biomecánica y cicatrización en Insall y Scott, Cirugía de la rodilla (307-316).
2007.Madrid Elsevier
Buja LM, Hagler HK, Parsons D, Chien K, Reynolds RC, Willerson JT. Alterations of
ultrastructure and elemental composition in cultured neonatal rat cardiac myocytes
after metabolic inhibition with iodoacetic acid. Lab Invest. 1985; 53(4): 397–411.
Campos A, López-Escámez J.A, Cañizarez FJ, Crespo PV. Electrón probe X-ray
microanálisis of Ca and K distrubutions in the otholithinc membrana, Micro and
Microscopica Acta. 1992; 23:349-350.
Campos A, López-Escámez J.A, Crespo PV, Cañizarez FJ, Baeyens J.M. Gentamicin
ototoxicity in otoconia: quantitative electrón probe x-ray microanlysis, Acta Otolaryngol.
1994; 114:18-23.
Campos A. Cuerpo, histología y medicina. De la descripción microscópica a la
ingeniería tisular. Discurso de ingreso en la Real Academia Nacional de Medicina.
2004.
Canosa R. Desbridamiento artroscópico y artroplastia por abrasión. Posibilidades
actuales. Rev Ortop Traumatol 2002; 5:451-457.

Referencias Bibliográficas
174
Carini, R. Autelli, G. Bellomo, M.U. Dianzani, and E. Albano. Sodium-mediated cell
swelling is associated with irreversible demage in isolated hepatocytes exposed to
hypoxia or mitochondrial toxin. Biochim Biophys Res commun. 1995; 206:180-185.
Carini, G. Bellomo, M.G. de Cesaris, and E. Albano. Glycine protect against hepatocyte
killing by kcn or hypoxia by preventing intracellular na- overload in the rat. Hepatology.
1997; 26:107-112.
Carini R, M.G. de Cesaris, R.Splendore, M. Bagnati, G. Bellomo, and E. Alabano.
Alterations of na-homeostasis in hepatocyte reoxygenation injury. Biochim biophys
acta. 2000; 1500:297-305.
Cebamanos J. Procedimientos actuales en la reparación de las lesiones del cartílago
articular. Monografía en internet: Instituto Veterinario de Ortopedia y traumatología.
1998.
Chang HI, Yang X. Proteases for Cell Suicide: Functions and Regulation of Caspases.
2000. Microbiology and molecular biology reviews, Dec. 2000, p. 821–846.
Choy JC, Granville DJ, Hunt DW, McManus BM. Endothelial cell apoptosis:
biochemical characteristics and potential implications for atherosclerosis. J Mol Cell
Cardiol. 2001; Sep;33(9):1673-90.
Coco-Martin JM, Oberink JW, van der Velden-de Groot TA, Beuvery EC. Viability
measurements of hybridoma cells in suspension cultures. Cytotechnology. 1992; 8(1):
57–64.
Colossi PA, Harvey NL, Shearwin-Whyatt LM, Kumar S. Conversion of procaspasa-3 to
an autoactivation caspasa by fusion to te casapasa-2 prodomain. 1998. J. Biol. Chem.
273: 26566-26570.
Couceiro J, Carpintero P, Silva MT. Fisiopatología del cartílago articular. En: Cáceres
E, Fernández A. Manual SECOT de cirugía ortopédica y traumatología. Madrid: Ed.
Médica Panamericana 2003. 128-141.
Crespo PV, López-Escámez JA, Cañizares FJ, Campos A. X-ray microanalytical
determination of P, S, and K concentrations in the gelatinuos membrana of utricle. Acta
Otolaryngol Stockh. 1993; 113:176-180, 1993.

Referencias Bibliográficas
175
Chesterman PJ, Smith AU. Homotransplantation of articular cartilage and insolated
chondrocytes. An experimental studying rabbits. J Bone Joint Surg 1968; 50B:184-197.
Costa da AO, de Assis MC, Marques Ede A, Plotkowski MC. Comparative analysis of
three methods to assess viability of mammalian cells in culture. Biocell. 1999; 23(1):
65–72.
Debbage P. Distinctive structural and cytoskeletal properties of the long-surviving
neurons in cell cultures of embryonic spinal cord. Neuroscience. 1985; 14(1): 207–224.
De Diego E, Villanueva A, Roca A, Martín B, Meana A, Gómez S y Gómez J. Estudio
experimental sobre la viabilidad del injerto libre de epitelio urinario autólogo cultivado in
vitro. Actas Urol Esp. 2004; 28: 714-731.
Deveraux QL, Roy N, Stennicke HR, Van Arsdale T, Zhou Q, Srinivasula SM, et al.
IAPs block apoptotic events induced by caspasa-8 and cytochrome c by direct
inhibition of distinct caspasas. EMBOj. 1998. 17:2215-2223.
Deveraux QL, Takahashi R, Salvessen GS, Redd JC. X-linked IAP is a direct inhibitor
of cell death proteases. Nature 1997. 388: 300-304.
Dove A. Cell-based therapies go olive. Nature Biotech. 2002; 20: 339-343.
Edidin M. A rapid, quantitative fluorescence assay for cell damage by cytotoxic
antibodies. J Immunol. 1970; 104(5): 1303–1306.
Elves M, W. A study transplantation antigens on chondrocytes from articular cartilage.
J Bone Joint Surg. 1974; 56A:178-185.
Emilson LB, Dougherty KA, Cockett AT, Urry RL. Simultaneous determination of
human sperm morphology and viability: simple office technique. Urology. 1978; 11(5):
488–491.
Fernández-Segura E, Cañizares FJ, Cubero MA, Campos A, Warley A. A procedure to
prepare cultured cells in suspention for electrón probe X-ray microanlysis: application
to scanning and transmission electron microscopy, Journal of Microscopy. 1999;
196:19-25.

Referencias Bibliográficas
176
Fernández-Segura E, Cañizares FJ, Cubero MA, Warley A, Campos A. Changes in
elemental content during apoptotic cell death studied by electrón probe X- ray
microalysis. Experimental cell Research. 1999; 253:454-462.
Finn G. Histología, Editorial Médica Panamericana, Tercera Edición, Madrid 2002; 263-
267.
Geschickter CF. The Application of Dyes in the Cancer Problem. Biotech Histochem.
1930; 5(2): 49–64.
Glowaki J, Yates KE, McLean R, Mizuno S. In Vitro engineering of cartilage: effects of
serum substitutes, TGF-B, and IL-1 alfa. Orthod Craniofacial Res. 2005; 8;200-208.
Goldstein AD, Roming DE, Newbury CE, Lyman P, Echlin PC, Fioror DC, Joy and E.
Lifshin. Scanning electron microscopy and X-ray microanalysis. Plenum Press, New
York. 1992.
Green WT. Basic science and patfology orthopaedic and related research. 1977; 124:
237-250.
Grogan SP, Barbero A, Diaz-Romero J, Cleton-Jansen AM, Soeder S, Whiteside R,
Hogendoorn PC, Farhadi J, Aigner T, Martin I, Mainil-Varlet P. Identification of markers
to characterize and sort human articular chondrocytes with enhanced in vitro
chondrogenic capacity. Arthritis Rheum. 2007 Feb;56(2):586-95.
Hamilton VT, Habbersett MC, Herman CJ. Flow microfluorometric analysis of cellular
DNA: Critical comparison of mithramycin and propidium iodide. J Histochem Cytochem.
1980; 28(10): 1125–1128.
Hall TA. Capabilities and limitations of probe methods for the microanalysis of chemical
elements in biology: a brief introduction. Ultramicroscopy 1988; 24: 181-184.
Handogy L, Sökösd L, Szabó Zs. Repair of cartilage defects. Technical aspects. Rev
Chir Orthop Reparatrice Appar Mot. 1999; Dec;85(8):846-57.
Hannallah D, Peterson B, Lieberman J, et al.: Gene therapy in orthopaedic surgery.
Instr Course Lect. 2003; 52: 753-768.

Referencias Bibliográficas
177
Hermann C, Assmus B, Urbich C, Zeiher AM, Dimmeler S. Insulin mediated stimulation
of protein Kinase Akt: a potent survival signalling cascade for endothelial cells.
Arterioescler. Thromb.. Vasc. Biol. 2000. 20:402-409.
Hoskins JM, Meynell GG, Sanders FK. A comparison of methods for estimating the
viable count of a suspension of tumour cells. Exp Cell Res. 1956; 11(2): 297–305.
Hu J, Xu YJ, Zhang ZX, Tian F. Effect of cigarette smoke extract on proliferation of rat
pulmonary artery smooth muscle cells and the relevant roles of protein kinase C. Chin
Med J (Engl). 2007; Sep 5;120(17):1523-8.
Hueber AO, Zornig M, Lyon T, Suda T, Nagata S, Evan GI. Requirement for the CD95
receptor-ligand pathway in c-Myc-induced apoptosis. 1997. Science 278:1305–1309.
Ikegami E, Fukazagua F, Kambe M, Watanabe M, Kamisago M, Hajikano M, Katsube
Y, Owaba S. 2007; 71:(11):1815-1820.
Irmle M, Thome M, Hahne M, Scheneider P, Hofmann K, Steier V, et al. Inhibition of
death reception signal by cellular flip. Nature. 1997. 388: 190-195.
Jato M, Ruano RA, Cerdá MT, Mejuto MT. Agencia de avalación de tecnoloxias
sanitarias de Galicia, Junta de Galicia, Conselieria de Sanidade, secretaria xeral
subdirección xeral de aseguramento y planificación. 2005; 01, 1-68.
Jiang CC, Chiang H, Liao CJ, Lin YJ, Kuo TF, Shieh CS, Huang YY, Tuan RS. Repair
of porcine articular cartilage defect with a biphasic osteochondral composite. J
Orthopaedics Res. 2007; Oct;25(10):1277-90.
Kaigler, D. Mooney, D. Tissue engineering´s impact on dentistry. J Dental Education.
2001; 65, 5: 456-462.
Koseki T, Inohara N, Chen S, Nuñez G. ARC an inhibitor of apoptosis expressed in
skeletal muscle and heart that interacts selectively with caspasas. Proc. Natl. Acad.
Sci. 1998. 95: 5156-5160.
Krishan A. Rapid flow cytofluorometric analysis of mammalian cell cycle by propidium
iodide staining. J Cell Biol. 1975; 66(1): 188–193.

Referencias Bibliográficas
178
Kuettner KE, Schelerbach R, Hascall VC. Articular cartilage biochemistry. New York:
Raven press. 1987.
Lesson T, Lesson R, Paparo A. Texto y Atlas de Histología, Editorial Interamericana
McGraw- Hill. 1989; 159-167.
Li P, Nijhawan D, Budihardjo I, Srinivasula SM, Ahmad M, Alnemri ES, WangX. Cytochrome c and dATP-dependent formation of Apaf-1/caspase-9 complex initiates
an apoptotic protease cascade. 1997. Cell 91:479–489.
Llames SG, Del Rio M, Larcher F, García E, García M, Escamez MJ, Jorcano JL,
Holguín P, Meana A. Human plasma as a dermal scaffold for the generation of a
completely autologous bioengineered skin. Transplantation. 2004; Feb 15;77(3):350-5.
Llusá, M. Merí, A. Ruano, D. Generalidades en Llusá, M Manual y atlas fotográfico de
anatomía del aparato locomotor (15). Buenos Aires; Madrid: Ed. Panamericana. 2007.
López-Escámez JA, Cañizarez FJ, Crespo PV, Campos A. Electrón probe
microanálisis of otholitic membrana. A methodological and quantitative study. Scanning
micrcoscopy. 1992a; 6:765-772.
López-Escámez JA, Crespo PV, Cañizares FJ, Campos A. Dependence between
saccule and utricle in Ca and K concentrations determined by electrón probe X-ray
microanalysis. Micron and microscopica Acta. 1992b; 23:367-368.
López-Escámez JA, Crespo PV, Cañizares FJ, Campos A. Standards for quantification
of element in the otholithic membrana by electrón probe x-ray microanalysis: calibration
curves and electron beam sensivity. Journal of Microscopy. 1993; 171:215-222.
López-Escámez JA, Campos A. Standards for X-ray microanalysis of calcified
structures. Scanning Microscopy. 1994; 8:171-185.
Mannick JB, Hausladen A, Liu L, Hess DT, Zeng M, Miao QX, et al. Fas induced
caspasa denitrosylation. 1999. Science 284: 651-654.
Maroudas A. Physicochemical properties of articular cartilage. In: Adult articular
cartilage, Freeman MAR. ed. Pitman Medical. Kent, England: 1979; 215-290.

Referencias Bibliográficas
179
Martín Hernández C. Estudio mecánico, histológico e histomorfométrico del
regenerado del cartílago a partir de injerto de periostio invertido. Tesis doctoral.
Barcelona: UAB; 2002.
Messner K. Reparación de las lesiones del cartílago articular. Insall & Scott. Cirugía
de la Rodilla. Madrid: Marbán; 2004; 327-340.
Minas T, Nehrer S. Current concepts in the treatment of articular cartilage defects.
Orthopedics. 1997; Jun;20(6):525-38.
Montalvo A. Evaluación genética y microanalítica de las células madre de la gelatina
de Wharton para su utilización en Ingeniería Tisular. Tesis Doctoral. Universidad de
Granada 2008a.
Montalvo A, Garzón I, González-Andrades M, Lobo M, Fernández-Montoya A,
Alaminos M, Campos A. Evaluación de la viabilidad de los cultivos de condrocitos
mediante microanálisis por energía dispersiva de rayos X. Boletín de la Asociación
Española de Bancos de Tejidos (AEBT). 2008b; 9:3-7.
Mow VC, Colmes MH, Lai WM. Fluid Transport and mechanical properties of articular
cartilage: A review. J Biomech. 1984; 102:73-84.
Muir H. Proteoglycans as organizers of the extracellular matrix. Biochem Soc Trans.
1983; 11:613-622.
Nakamura N, Miyama T, Engebretesen L, Yoshikawa H, Shino K. Cell-Based therapy
in articular lesions of the knee. Arthroscopy: The journal of arthroscopic and related
surgery. 2009; 25 (5): 531-552.
Nicholson DW, Ali A, Thornberry NA, Vaillancort JP, Ding CK, Gallant M, Gareau Y,
Griffin PR, Labelle M, Lazebnik YA, Munday NA, Raju SM, Smulson ME, Yamin TT, Yu
V, Miller DK. Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. 1995. Nature 376:37–43.
Norman Scott W. Cartílago articular y menisco: biología biomecánica y cicatrización.
Insall y Scott Cirugía de la rodilla (4ª Ed. Vol 2). Madrid Elsevier. 2007; 307-316.
O’Driscoll SW: Current Concept Review: The healing and regeneration of articular
cartilage. J Bone Joint Surg (AM). 1998; 80: 1795-1812.

Referencias Bibliográficas
180
Oliveira JM, Rodrigues MT, Silva SS, Malafaya PB, Gomes ME, Viegas CA, Dias IR,
Azevedo JT, Mano JF, Reis RL. Novel hydroxyapatite/chitosan bilayered scaffold for
osteochondral tissue-engineering applications: Scaffold design and its performance
when seeded with goat bone marrow stromal cells. Biomaterials. 2006
Dec;27(36):6123-37.
Park SS, Ward MJ. Tissue-Engineered cartilage for implantation and grafting. Facial
Plast Surg. 1995; 110:248-278.
Peterson L. Experiencia con trasplante autólogo de condrocitos. Insall & Scott.
Cirugía de la Rodilla. Madrid: Marbán. 2004; 341-56.
Park C, Moon DO, Ryu CH, Choi BT, Lee WH, Kim GY, Choi YH. Beta-sitosterol
sensitizes MDA-MB-231 cells to TRAIL-induced apoptosis. Acta Pharmacol Sin. 2008;
Mar;29(3):341-8.
Patterson MK Jr. In measurement of growth and viability of cells in culture. 1ª ed. New
York: IH Pastarf, Academic Press. 1979; pp. 150–155.
Perales S, Alejandre MJ. 2007. Apoptosis en células de músculo liso arteriales.
Granada. Universidad Granada.
Phillips HJ, Terryberry JE. Counting actively metabolizing tissue cultured cells. Exp Cell
Res. 1957; 13(2): 341–347.
Phillips HJ. In dye exclusion tests for cell viability. 2ª ed. New York: Academic Press.
1973; 406–411.
Posadas I, Vellecco V, Santos P, Prieto-Lloret J, Ceña V. Acetaminophen potentiates
staurosporine-induced death in a human neuroblastoma cell line. Br J Pharmacol.
2007; Mar;150(5):577-85.
Pineda, K; Merentes, E; Starosta, R. Caracterización Morfológica y Bioquímica de
Condrocitos in Vitro. Revista del Instituto Nacional de Higiene Rafael Rangel. 2004.
Poole AR. Proteoglycans in health and desease: structure and functions. Biochem J.
1986; 236:1-14.

Referencias Bibliográficas
181
Rinaldi M, Tricarico M, Bonmassar E, Parrella P, Barrera G, Fazio VM. Effect of 4-
hydroxynonenal, a product of lipid peroxidation on NK susceptibility of human K562
target cells. Anticancer Res. 1998; Sep-Oct;18(5A):3591-5.
Robert H. Techniques for repair of chondral and osteochondral lesions in the knee.
Principles of rehabilitation Ann Readapt Med Phys. 2002; Feb;45(2):62-8.
Rodriguez-Morata A, Garzon I, Alaminos M, Garcia-Honduvilla N, Sanchez-Quevedo
MC, Bujan J, Campos A. Cell viability and prostacyclin release in cultured human
umbilical vein endothelial cells. Ann Vasc Surg. 2008; 22(3): 440–448.
Roomans G.M, Von Euler A. X-ray microanalysis in cell biology and cell pathology. cell
biol int. 1996; 20(2): 103-109.
Rosales R. Aislamiento y control de calidad microanalítico para ingeniería tisular de los
condrocitos del tejido cartilaginoso hialino y fibroso. Tesis Doctoral. Universidad de
Granada, 2008.
Rotonda J, Nicholson DW, Fazil KM, Gallant M, Gareau Y, Labelle M, et al. The three
dimensial structureof apopain CPP32, a keymediator of apoptosis. 1996. Nature struct.
Biol. 1996. 3: 619-625.
Roy N, Deveraux QL, Takahashi R, Salvessen GS, Redd JC. The c-IAP-1 and c-IAP-2
proteins are direct inhibitors of specific caspasas. EMBOj. 1997. 16:6914-6925.
Sanchez-Quevedo MC, Alaminos M, Capitan LM, Moreu G, Garzon A, Crespo PV,
Campos A. Histological and histochemical evaluation of human oral mucosa constructs
developed by tissue engineering. Cellullar and Mollecular Biology. 2007; 22:631-640.
Saris DB, Vanlauwe J, Victor J, Haspl M, Bohnsack M. Characterized chondrocyte
implantation results in better structural repair when treating symptomatic cartilage
defects of the knee in a randomized controlled trial versus microfracture. Am J Sport
Med. 2008; Feb;36 (2):235-46.
Sellars RS, Peluso D, Morris EA. The effect of recombinant human bone
morphogenetic protein-2 (rhBMP-2) on the healing of full-thickness defects of articular
cartilage. J Bone Joint Surg Am. 1997; 79 1452-1463.

Referencias Bibliográficas
182
Sgaglione NA, Miniaci A, Gillogly SD, Carter TR. Update on advanced surgical
techniques in the treatment of traumatic focal articular cartilage lesions in the knee.
Arthroscopy. 2002; Feb;18(2 Suppl 1):9-32.
Shi, Y. (2002). Mol. Cell 9, 459–470.
Slee EA, Harte MT, Kluck RM, Wolf BB, Casiano CA, Newmeyer DD. Ordering the
cytochrome c-initiated caspasa cascade: hierarchical activation of caspases -2, -3, -6, -
7, -8, and -10 in a caspase-9-dependent manner. 1999. J. Cell. Biol. 144: 281-292.
So CL, Kaluarachchi K, Tam PP, Cheah KSE. Impact of mutations of cartilage matrix
genes on matrix structure, gene activity and chondrogenesis. Osteoarthritis and
cartilage, 2001; 9 (suppl. A): S160-S173.
Sopena JJ, Carillo JM, Rubio M, Redondo JI, Serra I, Soler C. Estructura y función
del cartílago articular. 2004. Argos. 52: 24-26.
Srinivasula SM, Ahmad M, Fernandes-Alnemri T, Alnemri ES. Autoactivation of
procaspasa-9 by Apaf-1-mediated oligomerization. 1998. Mol. Cell. 1: 949-957.
Stanton H, Rogerson FM, East CJ, Golub SB, Lawlor KE, Meeker CT, Little CB, Last K,
Farmer PJ, Campbell IK, Fourie AM, Fosang AJ. ADAMTS5 is the major
aggrecanasein mouse cartilage in vivo and in vitro. Nature. 2005 Mar 31;434
(7033):648-52.
Steinert AF, Ghivizzani SC, Rethwilm A, Tuan RS, Evans CH, Nöth U. Major biological
obstacles for persistent cell-based regeneration of articular cartilage. Arthritis Res Ther.
2007; 9(3):213.
Stennicke HR, Jurgensmeier JM, Shin H, Deveraux Q, Wolf BB, Yang X, Zhou Q,
Ellerby HM, Ellerby LM, Bredesen D, Green DR, Reed JC, Froelich CJ, Salvesen GS.
Pro-caspase-3 is a majorphysiologic target of caspase-8. 1998. J. Biol. Chem.
273:27084–27090.
Thornberry NA, Bull HG, Calaycay JR, Chapman KT, Howard AD, Kostura MJ, Miller
DK, Molineaux SM, Weidner JR, Aunins J, et al. A novel heterodimeric cysteine
protease is required for interleukin-1 beta processing in monocytes. 1992. Nature
356:768–774.

Referencias Bibliográficas
183
Thornberry NA, Lazabnick Y. Caspases, enemies within. 1998. Science. 281: 1312-
1316.
Tuli R, Li WJ, Tuan RS. Current state of cartilage tissue engineering. Arthritis Res
Ther. 2003; 5(5):235-8.
Vanthanouvong V, Hogman M, Roomans GM. In vitro and in situ experimental model
for X-ray microanalysis of intestinal epithelium. Microscopy Res Tech. 2003; 62:211-
217.
Wada M, Watanabe A, Yamashita T, Isobe T, Moriya H. Evaluation of articular cartilage
with 3D-SPGR MRI after autologous chondrocyte implantation. J Orthop Sci. 2003; 8
(4) 514-517.
Wakitani S, Kimura T, Hirooka A, Ochi T, Yoneda M, Owaki H, Ono K, Yasui N. Repair
of rabbit articular surfaces with allografts of chondrocytes embedded in collagen gels.
Nippon Seikeigeka Gakkai Zasshi. 1989; May 63 (5): 529-38.
Warley A, Fernández-Segura E, López-Escámez JA, Campos A. Changes in elemental
concentrations in K562 target cells after conjugation with human lymphocytes studied
by X-ray microanalysis. Cell Biology International. 1994; 18:915-916.
Warley A. X-ray microanalysis for biologists. Practical methods in electron microscopy.
Glauert AM. (ed) Portland Press, London. 1997.
Wiesel JM, Gamiel H, Vlodavsky I, Gay I, Ben-Bassat H. Cell attachment, growth
characteristics and surface morphology of human upper-respiratory tract epithelium
cultured on extracellular matrix. Eur J Clin Invest. 1983; 13(1): 57–63.
Wilson KP, Black JA, Kim EE, Griffith JP, Navia MA, et al. Structure and mechanism og
interleukin-1 beta converting enzyme. 1994. Nature 370: 270-275.
Zierold K. Effects of cadmium on electrolyte ions in cultured rat hepatocytes studied by
X-ray microanalysis of cryosections. Toxicol Appl Pharmacol. 1997; 144: 70–76.

FACULTAD DE MEDICINA
FACULTAD DE MEDICINA
UNIVERSIDAD DE GRANADA
UNIVERSIDAD DE GRANADA
TESISDOCTORAL
IDENTIFICACIÓN DE PATRONES DE VIABILIDAD Y EXPRESIÓN GÉNICA EN CONDROCITOS ARTICULARES HUMANOS PARA SU UTILIZACIÓN EN INGENIERÍA TISULAR
Álvaro Morales Villaescusa GRANADA 2010
TESIS DOCTORAL
Álvaro Morales Villaescusa