







Idiomas
Páginas
Jurídico


Estrategias de Manejo de la Roña Cladosporium cladosporioides (FRESEN) G.A. de
VRIES de la Gulupa Passiflora edulis f. edulis Sims.
Carlos Fernando Castillo Londoño
Universidad Nacional de Colombia
Facultad de Ciencias Agropecuarias
Maestría en Ciencias Agrarias
Palmira, 2014

Estrategias de Manejo de la Roña Cladosporium cladosporioides (FRESEN) G.A. de
VRIES de la Gulupa Passiflora edulis f. edulis Sims.
Carlos Fernando Castillo Londoño
Trabajo de investigación presentado como requisito parcial para optar al título de:
Magister en Ciencias Agrarias con énfasis en Fitopatología
Director:
Ph.D. Carlos German Muñoz Perea
Codirector:
Ph.D. Elizabeth Álvarez
Codirector:
Ph.D. Kriss Wichechkus
Universidad Nacional de Colombia
Facultad de Ciencias Agropecuarias
Maestría en Ciencias Agrarias
Palmira. 2014

A Dios por darme la inmensa
voluntad y capacidad de entrega para la
hacer las cosas con amor y dedicación. A mi
madre Carmen y a Geovanny por su gran
confianza, apoyo, consejos y su constante
exigencia para formar la persona que soy.

AGRADECIMIENTOS
A la Universidad Nacional de Colombia sede Palmira por las oportunidades y facilidades
brindadas en el transcurso de mi carrera profesional.
A la Universidad Jorge Tadeo Lozano y al Centro Internacional de Agricultura Tropical
(CIAT) por brindarme la oportunidad de trabajar en el proyecto.
A mi director Carlos German Muñoz, y codirectores Elizabeth Álvarez y Kriss
Wichechkus, por sus valiosos aportes y orientación en esta investigación.
A la Dra. Martha Cárdenas directora del Centro de Transformación Genética (Riverside),
por su valiosa colaboración en la revisión del documento.
A mis profesores, a quienes les debo gran parte de mis conocimientos, gracias por
prepararnos para un futuro competitivo no solo como los mejores profesionales sino
también como mejores personas.
Gracias a todas las personas que ayudaron directa e indirectamente en la realización de este
proyecto.

Contenido
2. LA GULUPA (Passiflora edulis f. edulis Sims) ................................................................ 1
2.1 ORIGEN Y DISTRIBUCIÓN ...................................................................................... 1
2.2 TAXONOMÍA DE Passiflora edulis f. edulis Sims. ................................................... 1
2.3 DESCRIPCIÓN ............................................................................................................ 2
2.4 IMPORTANCIA DEL CULTIVO ............................................................................... 3
2.5 PROPIEDADES DE LA GULUPA ............................................................................. 5
2.5.1 VALOR NUTRICIONAL DE LA GULUPA ........................................................... 5
2.5.2 CONDICIONES Y REQUERIMIENTOS AGRO-ECOLÓGICOS ......................... 5
2.6 GENÉTICA EN MATERIALES COMERCIALES DE GULUPA (Passiflora edulis f.
edulis SIMS) EN COLOMBIA .............................................................................................. 7
2.6.1 Variabilidad genética ................................................................................................. 7
2.6.2 Mejoramiento genético y variedades ......................................................................... 9
2.6.3 CARACTERIZACIÓN MOLECULAR EN PASIFLORAS .................................. 11
2.7 ENFERMEDADES EN GULUPA ................................................................................ 12
2.7.1 Antracnosis ocasionada por Glomerella cingulata (Stoneman) Spauld. & H.
Schrenk (Anamorfo Colletotrichum gloeosporioides (Penz.) Pen0z. & Sacc. in Penz). . 13
2.7.2 Características morfológicas................................................................................... 13
2.7.3 Epidemiología de la Antracnosis de la gulupa. ...................................................... 14
2.7.4 Factores que favorecen el desarrollo de Colletotrichum gloeosporioides. ............. 14
2.7.6. La Roña (Cladosporium cladosporioides) (Fresen.) G. A. de Vries ...................... 18
2.7.6.1. Características morfológicas................................................................................ 19
2.7.6.2 Epidemiología de la Roña de la gulupa. .............................................................. 19
2.7.6.3 Sintomatología ..................................................................................................... 20
2.8 MANEJO DE LA ENFERMEDAD .............................................................................. 20
2.8.1 Semillas (Material genético) .................................................................................... 21
2.8.2 Siembra .................................................................................................................... 21
2.8.3 Podas ........................................................................................................................ 22
2.8.4 Fertilización ............................................................................................................. 23
2.8.5 Manejo de arvenses ................................................................................................. 23
2.8.6 Cosecha .................................................................................................................... 24
2.8.7 Manejo químico ...................................................................................................... 24
2.9 MARCADORES MOLECULARES ............................................................................. 30
2.9.1. ITS (Región espaciadora interna transcrita) ........................................................... 31

2.9.3. Microsatélites RAMS ............................................................................................. 31
3. OBJETIVOS .................................................................................................................... 33
3.1 OBJETIVO GENERAL ............................................................................................ 33
3.2 OBJETIVOS ESPECÍFICOS .................................................................................... 33
4. ASPECTOS METODOLÓGICOS .................................................................................. 34
4.1 Recolección de material vegetal y aislamiento de cepas ........................................... 34
4.2.1 Reactivación de los aislamientos ............................................................................. 34
4.2.3 Aislamientos de cepas de Cladosporium spp .......................................................... 35
4.2.4 Desinfestación del material vegetal ......................................................................... 36
4.2.5 Siembra directa en medio de cultivo ....................................................................... 36
4.2.6 Purificación .............................................................................................................. 36
4.2.7 Obtención de cultivos monospóricos ....................................................................... 37
4.2.8 Pruebas de patogenicidad: Postulados de Koch ..................................................... 38
4.2.9 Preparación de los frutos y del material antes de la inoculación ............................. 38
4.2.10 Ajuste de la metodología de inoculación de frutos de gulupa con Cladosporium
spp ..................................................................................................................................... 39
4.2.11. Pruebas de patogenicidad ..................................................................................... 40
4.2.11 Evaluación de la enfermedad ................................................................................. 40
5.0. Pruebas moleculares ...................................................................................................... 41
5.1. Extracción de ADN ................................................................................................... 41
4.5.2. Amplificación de las regiones ITS del ADNr ........................................................ 42
4.5.3. Visualización y análisis de productos amplificados. .............................................. 43
4.5.4. Limpieza de producto PCR para secuenciación .................................................... 43
5.4.5 Análisis de secuencia ............................................................................................... 44
4.5.6. Amplificación aleatoria de microsatélites RAMS. ................................................. 44
4.6. Análisis estadístico ........................................................................................................ 46
4.6.1. Patogenicidad ......................................................................................................... 46
4.6.2. RAMS ..................................................................................................................... 47
4.6.3. Análisis de agrupamiento ....................................................................................... 48
4.6.4 Análisis de correspondencia múltiple ...................................................................... 49
4.6.5 Diversidad genética ................................................................................................. 49
4.6.6. Distancia genética ................................................................................................... 50
5.0. Evaluación in vitro de los extractos vegetales para el control de Cladosporium spp ... 51
5.1. Selección de aislamientos de Cladosporium spp. ...................................................... 51
5.2 Pruebas antagónicas .................................................................................................... 52

5.2 Preparación de inóculo de Cladosporium spp. ........................................................... 52
5.3. Preparación de extractos vegetales ............................................................................ 53
5.3.1. Extracto de Furcraea macrophylla (Fique) ............................................................ 53
5.3.2. Extracto de Swinglea glutinosa (Ecoswing®) ........................................................ 53
5.3.3 Extracto de Tagetes patula ...................................................................................... 53
5.3.4. Lixiviado de Compost de Raquis de Plátano (LRP) ............................................... 54
5.3.5. Stratego® (Ingrediente activo: Trifloxystrobin, Propiconazol) ............................. 54
5.3.6 Fungibiol .................................................................................................................. 54
5.3.7. Prueba de sensibilidad in vitro para biofungicidas. ................................................ 55
5.3.8 Evaluación del crecimiento del hongo y análisis de la información. ...................... 55
6.0. Evaluación in vivo de la capacidad de control de los extractos vegetales en frutos
desprendidos de gulupa. ....................................................................................................... 56
6. RESULTADOS Y DISCUSIÓN ..................................................................................... 58
6.1. Pruebas de patogenicidad (Postulados de Koch) y Estandarización de inoculación 58
6.2 .Caracterización patogénica ........................................................................................ 61
6.3. Secuenciación de las regiones ITS ............................................................................ 66
6.4. Caracterización molecular de Cladosporium cladosporioides ................................. 70
6.4.1. Análisis de Correspondencia Múltiple ................................................................... 75
6.4.2. Distancia e Identidad Genética ............................................................................... 76
6.4.3. Análisis de Varianza Molecular ............................................................................. 78
6.5. Evaluación de Biofungicidas ..................................................................................... 80
6.6. Pruebas antagónicas ................................................................................................... 87
6.8. Evaluación in vivo de la capacidad de control de los extractos vegetales en frutos
desprendidos de gulupa. ....................................................................................................... 91
7. CONCLUSIONES ............................................................................................................ 96
8. RECOMENDACIONES .................................................................................................. 97
BIBLIOGRAFIA .................................................................................................................. 98

INTRODUCCIÓN
Las frutas tropicales son una de las mejores opciones en la agricultura colombiana, por el
crecimiento de la demanda nacional e internacional, que en la actualidad es suplida por
importaciones (Cabra et al., 2002). Dentro de estas frutas tropicales, las Passifloras se
posicionan dadas sus características organolépticas que aumentan sus posibilidades de
mercado nacional e internacional.
En Colombia las Pasifloras se han venido consolidando dentro de las frutas de mayor
exportación. Para el 2005 ocuparon el cuarto lugar en exportación después del banano,
mango y uchuva, representando el 5,96% del total de frutas exportadas (Asohofrucol,
2007). De acuerdo con la FAO, las pasifloraceas, a pesar de estar ubicadas dentro de los
cultivos tropicales producidos en los más pequeños volúmenes, durante los últimos años
han presentado una gran expansión en el mercado (FAO, 2003). Este incremento se basa
principalmente en la comercialización del jugo, el cual, gracias a sus cualidades
organolépticas, se puede mezclar con otros jugos. Además de su valor comercial como
jugo, el fruto se puede utilizar en la elaboración de pulpas, dulces, néctares, jaleas y
mermeladas entre otros.
Principalmente la gulupa (Passiflora edulis f. edulis Sims.), es una planta nativa de Brasil y
se ha distribuido ampliamente en los países de los Andes. El fruto es muy apetecido para el
consumo en fresco debido a su sabor y aroma (Zárate, 2007).
Sin embargo, la Roña es una enfermedad limitante reduciendo entre un 20 y 40% de la
producción siendo favorecida por temperaturas altas prolongadas y alta humedad relativa
(Zárate, 2007). Además, la incidencia de la enfermedad se aumenta por diversos factores
como la selección inadecuada de zonas agroecológicas para el cultivo, distancias de
siembra, drenaje, podas, fertilización, deficiente recolección de frutos afectados e
inadecuado uso de fungicidas.
El uso indiscriminado de fungicidas para el control de la Roña, ha inducido resistencia del
patógeno a los productos en varios cultivos (Agrios, 2005), obligando a los agricultores a

utilizar nuevos fungicidas que aumentan los costos de producción. Además estos químicos
dejan residuos en el fruto disminuyendo la calidad del producto final, lo que reduce su
aceptación para exportar a otros países, convirtiéndose el conocimiento y manejo de esta
enfermedad de vital importancia para la producción nacional de la gulupa.
La gulupa se han constituido en una de las frutas exóticas más apetecidas en el mercado
internacional. Debido a que se han realizado escasas investigaciones en aspectos
fitopatológicos en Colombia, los agricultores han adoptado algunas prácticas realizadas en
diferentes cultivos. En la actualidad las enfermedades en el cultivo de gulupa se han
convertido en una limitante para la producción; para estos problemas no se han diseñado
estrategias de manejo. Por lo anterior y debido a que la gulupa es una fruta exótica, cuyo
destino principal es el mercado externo, se requiere diseñar una serie de estrategias que
integren distintos métodos de control en el ámbito del manejo integrado fitosanitario.
La fruticultura será, en los próximos años, el eje de reconversión del sector agroalimentario
colombiano, líder por su notable contribución a la creación de empleo e ingresos, a la
generación neta de divisas y a la modernización empresarial del sector agrícola, siempre
que supere los obstáculos de planeación de la producción con una solución integral de las
problemáticas fitosanitarias, transformación tecnológica y control de costos. Por lo tanto un
desarrollo frutícola en un país como Colombia debe estar sustentado tanto en un aporte
importante de innovación tecnológica, como en la identificación y control de los problemas
fitosanitarios y su comercialización.

1
2. LA GULUPA (Passiflora edulis f. edulis Sims)
2.1 ORIGEN Y DISTRIBUCIÓN
La gulupa, es originaria del Brasil y se encuentra ampliamente distribuida en los países de los
Andes (Ulmer y MacDougal, 2004).
La familia Pasifloraceae, está conformada aproximadamente por 630 especies, agrupadas en 18
géneros (Holm-Nielsen et a 1988.), dentro de los cuales el género Passiflora es el más
importante, ya que las especies ubicadas dentro de éste son apetecidas por sus propiedades
medicinales, como ornamentales y por las características deseables de sus frutos (Coppens et al.,
1997). El género Passiflora contiene alrededor de 530 especies (Cronquist, 1981) de las cuales
más de 350 se encuentran en las regiones tropicales de Sur América (Ulmer y MacDougal,
2004). La mayoría de estas especies han sido cultivadas durante varios años por coleccionistas de
ornamentales (Ulmer y MacDougal, 2004) pero únicamente la gulupa (Passiflora edulis f. edulis
Sims), el maracuyá (P. edulis f. flavicarpa), la badea (P.quadrangularis) y la granadilla (P.
ligulares) revisten importancia económica para la industria de jugos y para consumo en fresco en
Colombia (Guzzo et al., 2004).
2.2 TAXONOMÍA DE Passiflora edulis f. edulis Sims.
DIVISIÓN: Magnoliophyta
CLASE: Magnoliopsida
ORDEN: Violales
FAMÍLIA: Passifloraceae
GÉNERO: Passiflora
ESPECIE: P edulis f. edulis

2
2.3 DESCRIPCIÓN
El fruto es púrpura, redondo u ovalado con el pericarpio poco grueso, con 4 a 8 cm de diámetro,
con arilo pulposo de color naranja, de sabor ligeramente ácido y con buenas cualidades
organolépticas de sabor y aroma. Al finalizar la maduración, el pericarpio cambia su color verde
a púrpura, y la corteza se vuelve más resistente a la presión (CCI, 2006).
Morfológicamente : P edulis f. edulis Sims es una enredadera con hojas pequeñas, flores
coloridas y zarcillos de color verde. El tallo es subangular, hueco y estriado. Las frutas son de
tamaño pequeño de 4 a 8 cm. con una corteza ligeramente resistente de color morado. La pulpa
es amarilla oscura muy fragante (36% del peso de fruta) y menos ácida que la del maracuyá
(Garcia, 2006,). La gulupa presenta autopolinización (Ulmer y MacDougal, 2004), sin embargo
se ha reportado que cuando las flores de Passiflora edulis Sims tienen un estilo parcialmente
curvo la polinización cruzada es posible (CCI, 2006). Las altitudes mas apropiadas para su
desarrollo son de 1500 a 2000 msnm con temperaturas entre 15-20ºC con precipitaciones de
1500 a 2500 mm (CCI, 2006). Los suelos aptos son aquellos con textura arenosa y buen drenaje
(CCI, 2006). La planta presenta un crecimiento indeterminado e inicia su producción ocho meses
después de su establecimiento y con un manejo adecuado puede durar entre dos y tres años (CCI,
2006).
Según Ulmer y MacDougal (2004), entre las variedades más importantes de Passiflora edulis se
encuentran “Frederik”, “Perfecta” y “Nolfolk” las cuales son el producto de cruces entre gulupa
(P edulis f. edulis Sims) y maracuyá (P. edulis f. flavicarpa) y se cultivan principalmente en Sur
América (Ulmer y MacDougal 2004). La especie mas conocida en Europa es P edulis f. edulis
Sims, la cual es principalmente cultivada en Sur América y en menor escala en Sur Africa, India,
Malasia, Australia, Nueva Zelanda y Hawai.

3
2.4 IMPORTANCIA DEL CULTIVO
En Colombia las Pasifloras se han venido consolidando dentro de las frutas de mayor
exportación. Para el 2012 ocuparon el cuarto lugar en exportación después del banano, mango y
uchuva, representando el 5,96% del total de frutas exportadas (Asohofrucol, 2012). De acuerdo
con la FAO, las pasifloraceas, a pesar de estar ubicadas dentro de los cultivos tropicales
producidos en los más pequeños volúmenes, durante los últimos años han presentado una gran
expansión en el mercado (FAO, 2002).
Este incremento se basa principalmente en la comercialización del jugo, el cual, gracias a sus
cualidades organolépticas,se puede mezclar con otros jugos. Además de su valor comercial como
jugo, el fruto se puede utilizar en la elaboración de pulpas, dulces, néctares, jaleas y mermeladas
entre otros.
Actualmente no existen estadísticas específicas para la gulupa en cuanto a producción y área
sembrada, ya que esta es considerada como una variedad dentro del maracuyá y por lo tanto la
información no aparece desglosada para las dos especies. De acuerdo al reporte realizado por la
Corporación Colombiana Internacional para Junio de 2012, el volumen exportado de jugo de
maracuyá y gulupa ha crecido en un 94% respecto al mismo período en el 2011, con volumen de
exportación de jugo en el 2012 de 958 toneladas comparado con las 492 toneladas exportadas
entre enero y julio del año anterior. Adicionalmente se evidenció una penetración de este
producto en 21 países, siendo los principales países de destino Puerto Rico, Holanda, Alemania,
Reino Unido y Estados Unidos.
El principal país importador es Estados Unidos que para el 2006 incrementó las importaciones
con respecto al año anterior en un 72 % en valor y 56% en volumen de jugo. En Colombia el
área de cultivo de Gulupa registrada para exportación fue de 86,8 Ha y su producción se
encuentra distribuida en los municipios de Cundinamarca, Tolima, Boyacá y Quindío, La planta
produce anualmente 100 kg de fruta en promedio y dependiendo de la calidad del fruto, entre el
70% y 90% de dicha producción es destinada a la exportación, razón por la cual la fruta es poco
conocida a nivel nacional (CCI, 2006).

4
La gulupa se cataloga como cultivo promisorio exportable ya que presenta un gran potencial de
demanda internacional (FAO, 2002; Martínez, 2002; Asohofrucol, 2012). Generalmente las
estadísticas de la gulupa no se presentan individualmente pero se enmarcan dentro del grupo de
las pasifloras. De acuerdo a los cálculos de la Corporación Colombia Internacional (CCI, 2011),
las exportaciones de pasifloras en el año 2012 fueron de 1,7 millones de dólares FOB y para el
2006 el grupo se ubicó en el tercer renglón de frutas exportadas al mercado europeo después del
banano y la uchuva (MADR, 2006). Adicionalmente Tafur et al., (2010) en el Plan Frutícola
Nacional destacaron al grupo de las pasifloras por su dinámica en la exportación ya que presentó
una importante tendencia al aumento, pasando de exportar 1,2 millones de dólares FOB en el
2010 a 4,4 millones de dólares FOB en 2012 .
El destino de las exportaciones de pasifloras en 2006 fueron en su orden, Alemania, Holanda
(Países Bajos), Francia, Venezuela, Reino Unido, Costa Rica, España, Suiza, Canadá, Suecia,
Italia, Bélgica y Luxemburgo, Panamá, Portugal, Estados Unidos, Emiratos Árabes Unidos,
Brasil, Antillas Holandesas, Aruba y Guatemala. La participación en el área cultivada es poca
pero ha venido creciendo desde el año 1994. Con respecto al total nacional de área sembrada en
frutales, las pasifloras representaron el 15% en el 2007 con 17.028 hectáreas de las cuales 1.289
fueron destinadas al cultivo de la gulupa (CCI, 2008). Específicamente para el caso colombiano
es evidente el aumento en la producción y niveles de exportación que ha venido presentando el
cultivo de la gulupa.
Las estadísticas más recientes reportan que en el 2013 se produjeron 523 toneladas por un valor
de 1.936 dólares cuya cifra aumentó en el 2010 a 1.687 toneladas por un valor de 6.611 dólares.
Es decir que en tan solo dos años se triplicó la producción y el valor de las exportaciones
crecieron aproximadamente cinco veces más. Hasta Junio de 2012 se reportó una producción de
981 toneladas por valor de 4.802 dólares. Si se conserva dicha tendencia se esperaría que para
final del año 2012 la producción esté alrededor de 1.900 toneladas lo cual es superior al año
anterior (Agronet, 2012). Los seis principales destinos de exportación de la gulupa colombiana
son: Bélgica y Luxemburgo, Alemania, Holanda, Reino Unido, Francia y Suecia.

5
2.5 PROPIEDADES DE LA GULUPA
Principalmente de la gulupa son apetecidos los frutos, los cuales pueden ser consumidos como
fruta fresca o en productos procesados tales como pulpas, jugos, mermeladas etc. Como todas las
frutas de pasión, la Gulupa se utiliza como tranquilizante ya que funciona como sedante natural.
Baja la presión arterial y es una fuente rica de vitamina C. De igual forma la Gulupa ayuda a
conciliar el sueño, controla la tensión y mejora las funciones digestivas (Bridg, 2005).
2.5.1 VALOR NUTRICIONAL DE LA GULUPA
La Gulupa ayuda a proveer vitaminas esenciales que el cuerpo necesita como la vitamina A, B12
y C. Esta fruta es fuente de calcio, fibra, fósforo, hierro, proteínas y magnesio. También contiene
potasio y carbohidratos. Así mismo, en los frutos se pueden encontrar otras moléculas de
importancia, que se presentan en la Tabla 1.
2.5.2 CONDICIONES Y REQUERIMIENTOS AGRO-ECOLÓGICOS
En Colombia el cultivo de la gulupa se desarrolla en climas medios con temperaturas que varían
entre 15°C - 22°C y en un rango de altitud que va desde los 1.600 a los 2.400 msnm. Estos dos
factores son fundamentales en el desarrollo y productividad del cultivo ya que a mayor altitud el
tiempo entre siembra y cosecha se va haciendo más prolongado. Por ejemplo los cultivos
establecidos en el en altitudes cercanas a los 2000 msnm entran a cosecha 12 meses después de
la siembra, mientras que aquellos establecidos con altitudes alrededor de los 1.600 msnm inician
la cosecha siete meses después de establecido el cultivo (Ortíz et al., 2010).
Adicionalmente un elemento ambiental fundamental para la productividad del cultivo de la
gulupa es la radiación solar ya que está directamente relacionado con el desarrollo de los órganos
florales de modo que lugares o días nublados reducen el crecimiento y número de botones
florales, la floración y por ende la cosecha. Sin embargo es importante evitar el exceso de
radiación solar debido a que este puede causar daños irreversibles en el fruto denominados golpe
de sol (Nakasone y Paull, 1998; Lüdders, 2010). Precipitaciones alrededor de los 900 mm

6
anuales son ideales para el buen desarrollo de la planta ya que esta requiere una buena
disponibilidad de agua, especialmente en periodos críticos como brotación de yemas florales, cuajado
y llenado de fruto. Suelos fértiles francos a francos-arenosos son necesarios para facilitar el
desarrollo del sistema radical ya que éste es superficial y por ende requiere de un suelo liviano y
poco arcilloso (Ortíz et al., 2012).
Tabla 1. Contenido químico de la gulupa en 100 gramos (parte comestible)
Componente Concentración
Agua 88,9 g
Humedad 82.8 g.
Carbohidratos 11.0 g.
Cenizas 0.7 g.
Grasa 0.5 g.
Fibra cruda 0.4 g.
Proteína 1.5 g.
Calcio 9.0 mg.
Magnesio 23.9 mg.
Hierro 1.7 g.
Fósforo 21.0 g.
Vitamina A 1730 UI
Tiamina (B1) 0.1 g.
Riboflavina (B2) 0.17 mg.
Niacina 0.8 mg.
Acido ascórbico 20.0 mg.
Fuente: Secretaria Agricultura del Huila. Acuerdo de productividad cadena frutícola,
2012
Tabla 2. Condiciones y Requerimientos agro-ecológicos de la gulupa

7
Condiciones Agroecologicas
Altitud m.s.n.m <1200
Radiación H/día >4
Temperatura °C 20-30
Precipitación Mm 1000-2000
Humedad % 70-90%
Pendiente % <25%
Zona de Vida Bosque espinoso subtropical
Niveles de
nutrientes del
suelo
N Kg/ha 150
P2O5 Kg/ha 45
K2O Kg/ha 160
pH Kg/ha 5.5-
6.5
Profundidad cm. 50
Textura Clase Franca,
franco
arenosa,
franco
arcillosa
Distancia de siembra 4X3, 4X4
Distancia de siembra (Plantas/ Ha) 833,625
Vida útil 2 años
Cosecha Inicia de los 8 después de la siembra y
es continua durante minimo 14 meses :FUENTE: Fundación Sonrisas de colores (2004). Secretaria
Agricultura del Huila. Acuerdo de productividad cadena frutícola, 2012
2.6 GENÉTICA EN MATERIALES COMERCIALES DE GULUPA (Passiflora edulis f. edulis
SIMS) EN COLOMBIA
2.6.1 Variabilidad genética
La variabilidad intraespecífica en P. edulis f. edulis ha sido poco estudiada. Hasta el momento
únicamente existe un trabajo de variabilidad genética especifico para gulupa, realizado por
Trujillo et al., (2010), en el que se evaluaron 27 materiales procedentes de tres diferentes zonas
geográficas de Colombia empleando los marcadores moleculares denominados RAM (Random
Amplified Microsatellite). De acuerdo a los resultados obtenidos, la especie presenta una alta
diversidad genética. Sin embargo, este trabajo constituye un primer acercamiento y es necesario

8
desarrollar otros proyectos que permitan profundizar en el conocimiento de la especie para
entender y confirmar este primer resultado. Si bien la exploración a nivel intraespecífico es
escasa, la variabilidad interespecífica del género Passiflora ha sido relativamente bien estudiada
en los últimos años. Caracterizaciones morfoagronómicas, bioquímicas, fisicoquímicas y
moleculares han sido empleadas y enfocadas principalmente en la evaluación de los materiales
pertenecientes a los subgéneros Tacsonia y Passiflora los cuales reúnen las especies más
importantes económicamente.
Evaluaciones morfológicas fueron empleadas inicialmente con el fin de explorar la variabilidad
presente en siete especies del subgénero Tacsonia y Manicata, descriptores morfológicos de la
flor, hojas, tallo, brácteas y estípulas fueron útiles para detectar la variabilidad genética a nivel
interespecífico pero fueron poco discriminativos en el estudio de la variabilidad intraespecífica
(Villacis et al., 1998; Ocampo, 2007). Una mejor representación de la variación intra e
interespecífica fue obtenida empleando isoenzimas en las mismas especies evaluadas por
caracteres morfológicos y fue posible identificar cuáles de ellas pertenecían al mismo “pool”
primario y de esta forma abrir posibilidades para la transferencia interespecífica de genes entre
los materiales (Segura et al., 1998).
Posteriormente, la distancia genética de 12 especies representantes de los subgéneros Passiflora,
Decaloba y Tacsonia fue determinada mediante el análisis de fragmentos polimórficos del DNA
cloroplasmático empleando RFLP (Restriction Fragment Lenght Polymoprhism). En este trabajo
una alta variabilidad interespecífica fue nuevamente observada y se reportó adicionalmente la
evidencia de variabilidad intraespecífica en cuatro de las especies estudiadas (Sánchez et al.,
1999).
En estudios similares de variabilidad genética en Passiflora se emplearon diferentes técnicas de
evaluación tales como los marcadores moleculares AFLPs (Segura, 2002; Ocampo, 2004),
marcadores moleculares RAPDs (Random Amplification of Polymorphic DNA) (Fajardo et al,
1998; Aukar et al, 2002; Chrochermore et al, 2006), variación isoenzimática (Segura et al.,
1998) y caracteres agromorfológicos (Crochemore et al., 2003). En general los resultados
obtenidos han permitido confirmar la alta variabilidad interespecífica encontrada en el género

9
Passiflora y para cada estudio particular se logró determinar la cercanía y afinidades entre las
diferentes especies evaluadas.
Un resultado importante de algunos de los trabajos mencionados fue la baja variabilidad
intraespecífica encontrada por Crochemore et al. (2003) y por Fajardo et al. (1998) entre las
accesiones de P. edulis f. flavicarpa evaluadas, los cuales fueron diferentes a los obtenidos por
Sánchez et al. (1999) y Ocampo et al. (2004) quienes reportaron una alta variabilidad
intraespecífica en P. edulis f. flavicarpa. En estos casos la diferencia en los resultados estuvo
relacionada con el número y la procedencia de individuos analizados.
Estudios de variabilidad específicos para P. edulis f. flavicarpa, en donde se incluyeron
materiales comerciales y silvestres provenientes de diferentes regiones permitieron en cierta
medida aclarar la controversia de los anteriores resultados, ya que fue posible demostrar que la
variabilidad genética de los materiales comerciales de maracuyá es escasa pero al incluir
materiales silvestres las diferencias a nivel genético aumentan y permiten discernir diferencias a
nivel intraespecífico (Bellon et al., 2007).
Otro resultado interesante que se desprende de estos estudios es el planteamiento de una
separación de las dos formas botánicas de P. edulis, teniendo en cuenta que en la evaluación de
22 descriptores agromorfológicos definidos por Chrochermore et al. (2003) las dos especies se
diferenciaron ampliamente y en los estudios con marcadores moleculares han presentaron
diferencias significativas en los patrones de bandeo (Aukar et al, 2002; Ocampo et al, 2004).
Adicionalmente Sánchez et a.l (1999) obtuvieron índices de similaridad más bajos entre P. edulis
f. edulis y P. edulis f. flavicarpa que entre Pasiflora. sp. india y Pasiflora. mollissima las cuales
son consideradas especies distintas (Sánchez et al., 1999).
2.6.2 Mejoramiento genético y variedades
En Colombia actualmente no se han registrado cultivares comerciales de gulupa, sin embargo
recientemente se inició un programa de mejoramiento genético que se encuentra en un primer

10
ciclo de selección (John Ocampo, comunicación personal 2011). En Brasil tampoco existen
variedades comerciales de gulupa, sin embargo individuos de esta especie han sido incluidos en
los programas de mejoramiento para la forma amarilla que es de mayor importancia económica
para Brasil (Molina et al., 2005).
Cultivares híbridos producto del cruzamiento de la F1 de un cruce entre los materiales Sul Brasil
ou Marilia (P. edulis f. flavicarpa) x Roxo australiano (P edulis f. edulis) y su reciproco frueron
cruzados cada uno a su vez con el hibrido denominado GA-2 que proviene del cruce del material
Sul Brasil ou Marilia (P. edulis f. flavicarpa) x redondao (P. edulis f. flavicarpa).
Paralelamente se han llevado a cabo experimentos para el desarrollo de injertos de plantas de
gulupa (copa) provenientes de Australia, sobre dos materiales de maracuyá (patrón), Marilia
selección cerrado y el hibrido EC-2-0 las cuales presentan resistencia a Fusarium spp. Desde el
punto de vista técnico la injertación gulpa maracuyá fue posible ya que no se presentaron
problemas de incompatibilidad y se obtuvieron altas tasas de unión de los injertos, sin embargo
aún falta evaluar la aceptación comercial de los frutos en el mercado (Carvalho et al., 2005).
Las variedades comerciales de gulupa conocidas hasta el momento han sido desarrolladas
durante los programas de mejoramiento llevados a cabo principalmente en Australia. Sin
embargo variedades ó materiales de gulupa con características diferentes no han sido obtenidas o
encontradas naturalmente. Todas las variedades comerciales corresponden principalmente a los
materiales desarrollados mediante cruzamientos entre P. edulis f. edulis x P. edulis f. flavicapa ó
P. edulis f. edulis x P. incarnata. El uso de injertos en las plantaciones comerciales de gulupa en
Australia es muy común, por esta razón los programas de fitomejoramiento están enfocados al
desarrollo de patrones ó portainjertos mejorados y paralelamente al desarrollo de injertos o copas
mejoradas con el objetivo de desarrollar posteriormente un único injerto comercial (APIA,
2008).
La incorporación de P. incarnata a los programas de fitomejoramiento de patrones ha sido de
gran utilidad en el desarrollo de variedades con resistencia a virus y tolerancia a frío. Numerosos
patrones, copas, variedades e híbridos han sido desarrollados desde 1998 con el programa de

11
fitomejoramiento en Australia, sin embargo no todos han tenido aceptación en el mercado y su
competitividad se ha visto comprometida debido al deterioro a nivel genético y al aumento
significativo de la presión de virus denominado Passionfruit Woodiness Virus (Anderson, 2009).
Los cultivares comerciales australianas más conocidas son Misty Gem, Sweetheart y variedades
rojas como Panama y Pandora (Apia, 2008).
Recientemente se ha trabajado en el desarrollo de dos cultivares denominados McGuffies Red y
Samba los cuales han presentado resultados satisfactorios en términos de contenido de azúcar y
de pulpa, apariencia y tiempo de vida después de cosecha. Sin embargo, algunas evaluaciones
adicionales son necesarias antes de liberarlas al mercado (Anderson, 2009).
2.6.3 CARACTERIZACIÓN MOLECULAR EN PASIFLORAS
Estudios recientes reportan una baja variabilidad genética en 70 materiales de gulupa
provenientes de cultivos comerciales ubicados en ocho departamentos de Colombia empleando
marcadores moleculares AFLPs y SSRs (Ortiz, 2010)
Los AFLPs han sido de gran utilidad para evaluar diferencias genéticas entre especies sin un
conocimiento previo del genoma, tales como Manihot esculenta Crantz (Elias et al., 2000), Vicia
sativa L (Potokina et al., 2002) y Valerianella locusta (Muminovic et al., 2004), entre otras.
Actualmente, el limitado conocimiento genético del género Passiflora ha permitido posicionar
los AFLPs como una de las técnicas más poderosas para revelar polimorfismos en el DNA de las
especies de este grupo (Segura et al., 2002). Una alta variabilidad interespecífica en los
subgéneros Passiflora y Tacsonia fue identificada a partir de fragmentos con alto poder
discriminativo (50% fueron polimórficos) obtenidos empleando AFLPs (Segura et al., 2002;
Ocampo et al., 2004).
Resultados similares fueron encontrados utilizando AFLPs en el estudio de la variabilidad
intraespecífica de materiales mejorados de maracuyá provenientes de diferentes estados del
Brasil (Devos et al., 2004).

12
Ortiz (2010) obtuvó 419 fragmentos con seis combinaciones de primers de AFLPs, sin embargo
solo el 7% fueron polimórficos en al menos cuatro de los 70 individuos evaluados. Este
porcentaje es significativamente bajo si se compara con el 93,89% de fragmentos polimórficos
obtenidos con solo dos combinaciones de primers evaluando individuos de maracuyá (Devos et
al., 2004).
Un resultado similar fue reportado para materiales comerciales de maracuyá en Brasil empleando
RAPDs (Viana et al., 2003; Faleiro et al., 2005). De manera contrastante, una amplia diversidad
intraespecífica fue encontrada en individuos de maracuyá provenientes de programas de
mejoramiento empleando AFLPs (Devos et al., 2004), en individuos silvestres y comerciales
evaluados por RAPDs (Bellon et al., 2007) y en materiales procedentes de diferentes países
utilizando AFLPs (Ocampo et al., 2004).
Un resultado particular fue la alta variabilidad genética reportada por Trujillo et al. (2009) para
materiales comerciales de gulupa en Colombia empleando RAM´s. Estos resultados son
contrastantes con el estudio realizado por Ortiz (2010) y en otros trabajos en donde se evaluaron
individuos cultivados comercialmente para otras especies tales como maracuyá (Viana et al.
2003).
2.7 ENFERMEDADES EN GULUPA
Las enfermedades constituyen una de las mayores limitantes en la producción de los cultivos,
afectando el rendimiento y la calidad de los frutos en la cosecha y poscosecha (Agrios, 2005). El
conocimiento de los agentes causales de las enfermedades en gulupa es limitado a nivel nacional
y se orienta a partir de los hallazgos en enfermedades de otras pasifloras cultivadas, tales como
maracuyá, granadilla y curuba (Riascos et al.2010). Dentro de las enfermedades de importancia
que se han encontrado relacionadas con la gulupa se destacan: la Antracnosis causada por
Glomerella cingulata (anamorfo: Colletotrichum gloeosporioides), Secadera ocasionada por
Fusarium oxysporum Schltdl., Roña del fruto ocasionada por Cladosporium cladoporioides
Frensen., y la Bacteriosis o Mancha aceitosa ocasionada por Xanthomonas axonopodis, las

13
cuales se pueden presentar juntas en un mismo cultivo (Riascos, 2010). La incidencia y severidad
de estas enfermedades pueden llegar a ser muy altas cuando existen condiciones ambientales
adecuadas que favorezcan el desarrollo de estos patógenos.
2.7.1 Antracnosis ocasionada por Glomerella cingulata (Stoneman) Spauld. & H. Schrenk
(Anamorfo Colletotrichum gloeosporioides (Penz.) Pen0z. & Sacc. in Penz).
Ubicación taxonómica de Glomerella cingulata (Stonem.) Spauld & Schrenk
REINO: Hongo
DIVISIÓN: Mycota
SUBDIVISIÓN: Eumycotina
CLASE: Ascomycetes
SUBCLASE: Euascomycetidae
SERIE: Pyrenomycetes
ORDEN: Diaporthales
FAMILIA: Diaporthaceae
GÉNERO: Glomerella
ESPECIE: cingulata
2.7.2 Características morfológicas.
Colletotrichum gloeosporioides se caracteriza por presentar colonias variables, de color blanco
grisáceo a gris oscuro, micelio aéreo plano y en penachos asociados con conidióforos, por el
envés presenta color desigual de blanco grisáceo a gris oscuro especialmente con la edad. Los
conidios se observan formadas en masas de color salmón, y al microscopio son cilíndricas, con
ápices obtusos y base truncada (Bailey & Jeger 1992).
El estado perfecto (sexual o teleomorfo) de Colletotrichum spp., Glomerella Spauld. & H.
Schrenk, forma peritecios ostiolados, obpiriformes hasta subglobosos, glabros o con vellosidades

14
rodeando el ostíolo, parcialmente o completamente inmersos en el tejido del hospedante, solos y
dispersos o agregados, algunas veces poco desarrollados; cuello ostiolar inconspicuo o corto,
frecuentemente más pálido que el resto del ascoma, presencia de perifísis y parafísis. La pared
del ascoma es pseudoparenquimatosa, con células externas de pared delgada y pigmentada;
células internas hialinas y planas. Ascas unitunicadas de pared delgada, ampliamente cilíndricas,
ligeramente clavadas o elipsoides y sésiles o con un pedicelo corto. Las ascosporas son
unicelulares, hialinas, elipsoides o subcilíndricas, rectas o curvas y de tamaño menor a 20 μm de
longitud (Hanlin, 1990).
2.7.3 Epidemiología de la Antracnosis de la gulupa.
La enfermedad es favorecida por temperaturas y humedad relativa altas, así como, por las lluvias
periódicas; otros factores que favorecen la enfermedad son: estado nutricional deficiente de las
plantas, suelos pesados, material de siembra susceptible, selección inadecuada de zonas
agroecológicas, distancias de siembra inapropiadas, mal drenaje, podas, recolección de frutos
afectados e inadecuado uso de fungicidas (Escobar y Sánchez, 1992).
Colletotrichum gloeosporioides, forma los conidios en masas mucilaginosas denominadas cirros,
razón por la cual necesita de la lluvia seguida de humedad relativa alta y una capa de agua que
persista sobre el tejido infectado para la diseminación del patógeno. Estas condiciones sumadas
a la capacidad del hongo de penetrar en ausencia de heridas (penetración directa), hace más
difícil su prevención y manejo. La diseminación del patógeno por el salpique de agua, viento,
insectos, animales o herramientas también contribuye a que el manejo se haga más complejo
(Escobar y Sánchez, 1992).
2.7.4 Factores que favorecen el desarrollo de Colletotrichum gloeosporioides. El proceso
infectivo que desarrolla el patógeno causante de la Antracnosis es favorecido por precipitaciones
altas, humedad ambiental y temperaturas altas; sin embargo, cada uno de esos componentes debe
estar presente en niveles óptimos en forma simultánea, ya que con uno solo de ellos que no esté
en el rango requerido por el patógeno se afecta negativamente el proceso de infección (Hartung

15
et al., 1981; Zárate, 1998; Zárate, 2002). Mora et al. (1996), encontraron que la Antracnosis es
favorecida bajo estrés por efecto de saturación de agua en el suelo.
En cuanto a la temperatura, la mayoría de los aislamientos tropicales de C. gloeosporioides se
desarrollan a temperatura de 25°C y humedad alta durante 10 h continuas en la hoja, condiciones
que favorecen la germinación de conidias del hongo. La diferenciación en la patogenicidad de la
cepa puede ser consecuencia de las interacciones del medio ambiente, secreción de enzimas del
hongo y el genotipo del hospedante (Dodd et al., 1992).
Respecto al efecto de la temperatura en el crecimiento radial de C. gloeosporioides, la
temperatura óptima para su desarrollo es 20°C, con un menor crecimiento a 27°C y un desarrollo
escaso a 5, 10 y 30°C (Hartung et al., 1981).
El apresorio es importante en la penetración porque es el sitio donde se acumulan las enzimas
celulasas, cutinasas y pectinasas, que degradan la pared celular de la planta y contribuyen en la
colonización del tejido inter e intracelularmente. La temperatura ideal para la formación del
apresorio es de 25°C, lo cual ocurre 12 h después de la incubación. La formación de hifas
infecciosas, la cual se mide en el laboratorio cuando la hifa penetra una membrana de celulosa,
se da en mayor proporción a temperatura de 25°C, y comienza a las 30 h. Las temperaturas
comprendidas entre 10 y 12.5°C reducen la formación de apresorios (Fitzell et al., 1984).
Las precipitaciones altas aumentan la incidencia y la severidad de la enfermedad (Escobar y
Sánchez, 1992). En cultivos de tomate de árbol estos resultados reafirman que la estación seca
es la época más favorable para la producción, puesto que las condiciones de sequía disminuyen
la incidencia y la severidad de la Antracnosis al desfavorecer la diseminación y germinación del
patógeno (Araúz, 1986; Girard et al., 1987).
La función de la precipitación como tal, es como agente diseminador de los conidios; es el
principal condicionante de la humedad ambiental, junto con la temperatura. Si bien la cantidad
de lluvia caída es importante, más importante es la intensidad con que llueve, debido a que la
permanencia del agua sobre el tejido (humedad foliar) influye sobre la germinación de las

16
esporas; se requieren por lo menos 4 h de humedad permanente sobre la superficie del
hospedante para que se desarrolle la enfermedad; además lluvias cortas de 2 mm son suficientes
para la dispersión de conidias de Colletotrichum spp. ((Escobar y Sánchez, 1992). Los conidios
con ayuda del mucílago hidrofílico en el que se forman son arrastradas desde las partes
vegetativas hasta los órganos florales y frutos (Estrada et al., 1993; Escobar y Sánchez, 1992).
Otro factor importante es la humedad relativa, la cual tiene gran influencia en el proceso
infectivo de C. gloeosporioides: humedad relativa mayor del 95% es de gran importancia para
que ocurra la germinación de las esporas, formación de apresorios y diseminación del hongo,
siendo necesario que se tenga una temperatura óptima (Fitzell et al., 1984).
En el laboratorio se han podido determinar las condiciones requeridas por C. gloeosporioides
para desarrollar la enfermedad; evaluando humedades relativas entre 39 y 100% a una
temperatura de 25°C, se ha encontrado que la mayor germinación ocurre entre 97 y 100%, siendo
del 50% a las 20 h después de la incubación. A una humedad relativa del 100% se forman
apresorios e hifas infecciosas, iniciándose después de 30 h de incubación (Estrada et al., 1993).
Rangos de humedad relativa del 82 al 86%, inducen reducción en el porcentaje de germinación.
La humedad relativa alta sobre la superficie de los tejidos del hospedante y temperaturas entre
20-30°C, constituyen un factor importante para la liberación de conidias de Colletotrichum spp.
(Estrada et al., 1993)
Hay aspectos que proporcionan condiciones climáticas favorables para el desarrollo del hongo,
los cuales no influyen sobre la patogenicidad de Colletotrichum spp. pero que pueden facilitar
condiciones ambientales óptimas para su desarrollo, tales como la arquitectura de la planta y
aspectos agronómicos del cultivo. En cultivos como mango y tomate de árbol se pueden
desarrollar copas voluminosas con excesivo follaje, lo que crea condiciones climáticas favorables
para el desarrollo del patógeno en esta área de la planta en la cual la humedad ambiental es alta,
hay poca penetración de la radiación solar y ventilación baja, creándose un microclima favorable
para el desarrollo de la enfermedad (Saldarriaga et al., 1997).

17
Los factores agronómicos que favorecen el desarrollo del patógeno son: distancias de siembra
inapropiadas, épocas y dosis de fertilización inadecuada, principalmente de nitrógeno; en
cultivos de mango y tomate de árbol cuando no se realizan podas, los árboles tienden a
desarrollar una copa cerrada, condición que favorece el crecimiento rápido del patógeno y la
presencia de inóculo (Saldarriaga et al., 1997; Arango, 1993).
En el cultivo de maracuyá se realizan tres podas: la primera de ellas, conocida como poda de
formación, se hace a uno o 1.5 m de altura para obtener una planta limpia y de porte bajo, la
segunda poda o poda de crecimiento, tiene como objetivo promocionar un excelente equilibrio y
simetría del arbusto, esta va acompañada del deschuponado. La última poda (tercera), conocida
como poda de producción, se aconseja llevarla a cabo después de cada cosecha, se cortan todas
las ramas que se encuentran secas y enfermas; y debe realizarse de tal forma que se permita
buena entrada de luz y aireación a la planta, manejando de esta manera problemas fitosanitarios
como la Antracnosis (Oliveros, 2000).
Los sistemas de riego también son importantes para el manejo fitosanitario del cultivo. Para
guanábano el mejor sistema de riego es por goteo, su uso contribuye a bajas en el ataque por
Glomerella cingulata, mientras que el riego por microaspersión lo aumentan (Oliveros, 2000).
El uso indiscriminado de fungicidas para el control de Antracnosis, ha inducido resistencia del
patógeno a los productos en varios cultivos (Agrios 2005) como pera, uva, manzano (Chung et
al., 2006), mango y aguacate (Sanders et al., 2000) afectados por C. gloeosporioides exhibiendo
diferentes grados de resistencia hacia los bencimidazoles y, en otros cultivos como el pasto Azul
(Poa annua) afectado por aislamientos de Colletotrichum cereale resistentes a fungicidas
inhibidores de la demetilación del esterol, DMIs (sterol demethylation inhibitor, por sus siglas en
inglés) como el propiconazol, myclobutanil, tebuconazol, y triadimefon (Wong y Midland,
2007), obligando a los agricultores a utilizar nuevos fungicidas que aumentan los costos de
producción. Además estos químicos dejan residuos en el fruto disminuyendo la calidad y
seguridad en el consumo del fruto y el producto final, lo que reduce su aceptación para exportar a
otros países (Consumer.es, 2003).

18
2.7.5 Sintomatología. Este hongo afecta a hojas, guías y frutos. En las hojas los síntomas
aparecen en los márgenes y se manifiesta como lesiones acuosas de forma circular de 5 mm de
diámetro, presentan un halo de color verde oscuro y en las guías se observan lesiones alargadas.
En los frutos las lesiones se presentan como depresiones o áreas hundidas con pudrición seca,
causando un arrugamiento precoz del área afectada, la pudrición llega a la parte interna y
finalmente el fruto cae. En las áreas necróticas se observan anillos concéntricos de puntos
negros, que son las fructificaciones del hongo. El control químico que para contrarrestar este
hongo se realiza empleando productos cúpricos autorizados por las normas orgánica. Se pueden
hacer aplicaciones de productos quelatados de rápida absorción a base de calcio, potasio, boro,
zinc (Consumer.es, 2003).
2.7.6. La Roña (Cladosporium cladosporioides) (Fresen.) G. A. de Vries
REINO: Hongo
DIVISIÓN: Mycota
CLASE: Ascomycota
SERIE: Dothideomycetes
ORDEN: Capnodiales
FAMILIA: Dividiellaceae
GÉNERO: Cladosporium
ESPECIE: cladosposporioides
En Colombia es escasa la información relacionada a la etiología de las enfermedades que
afectan al cultivo de gulupa (Passiflora edulis Sims.) a nivel comercial. Por lo anterior, se han
adoptado los conocimientos existentes para cultivos de maracuyá (Passiflora edulis f.
flavicarpa) y otras pasifloras. Si bien en la actualidad, el principal problema fitosanitario es
ocasionado por virus, también existen otras alteraciones causadas por hongos que pueden ser
limitantes para la producción y comercialización. En el caso de enfermedades fungosas que
afectan la parte aérea de las plantas de gulupa, existen varios síntomas en frutos comúnmente
llamados roña que pueden ocasionar pérdidas económicas considerables para los productores; sin

19
embargo, otras sintomatologías son observadas en diferentes partes de las plantas que podrían
contribuir a estas pérdidas (Riascos, 2011).
2.7.6.1. Características morfológicas. Colonias de crecimiento moderadamente rápido (25-35
mm, OA, 25 °C, 7 días), aterciopeladas volviéndose pulverulentas, de color verde oliva a verde
clara. Conidióforos de hasta 250 µm de longitud, ramificados especialmente hacia el ápice, con
nódulos a lo largo del conidióforo que coinciden con los locus formadores de conidios. Conidios
en cadenas acrópetas y ramificadas, pardos, generalmente verrugosos, unicelulares, elipsoidales
o subcilíndricos, de 5-11 x 4-5 µm, con cicatrices prominentes y más oscuras que el resto del
conidio, pueden nacer de células soporte de mayor tamaño con uno o más septos transversales,
de hasta 14 x 6 µm, que se separan del conidióforo con mucha facilidad (Yasmiro, 1991).
2.7.6.2 Epidemiología de la Roña de la gulupa. El crecimiento, esporulación y dispersión de
Cladosporium spp. es altamente sensible a los cambios en las condiciones climáticas. Se ha
demostrado que varios factores ambientales favorecen la esporulación, entre ellos altas
temperaturas, alta humedad relativa y lluvia antes de la diseminación de las esporas. La lluvia
diaria en conjunto con temperaturas medias diarias de más de 15 ºC, pueden provocar un
incremento en la humedad relativa (alrededor del 80%), contribuyendo significativamente al
aumento en la producción de esporas (Rondon, 1995).
En estudios realizados en nueces, los conidios de Cladosporium caryigenum germinaron a 23 ºC
bajo oscuridad y humedad relativa de 100% en un periodo de 3 días; el periodo de incubación
ocurrió entre los 7 y 9 días a una temperatura entre 10 a 35 ºC y su máximo desarrollo de
síntomas se produjo luego de las 48 horas de infección con agua libre en las hojas y una
temperatura entre 15 a 25 ºC (Rivero, 2000); a partir de 9 horas continuas de HR en 100% se
incrementó significativamente la expresión de los síntomas (Gottwald, 1985).
La liberación de esporas de Cladosporium carpophilum en durazno, ocurrió cuando la humedad
relativa fue inferior al 40% (Gottwald, 1983).

20
C. herbarum ataca los frutos del maracuyá en sus diferentes estados de desarrollo, en ellos se
presentan lesiones ulcerosas de color pardo y tamaño variable, distribuidas en forma aislada,
aunque en algunos casos pueden agruparse (Chacón y Serna, 1990); internamente el fruto no
sufre daño, limitándose la enfermedad a la parte externa de la cáscara (García, 2006). En las
hojas los síntomas se manifiestan como lesiones circulares de 3.0-5.0 mm rodeadas de un halo
amarillo cuando inicia la enfermedad, pero después toda la lesión se vuelve de color rojizo. En
los tallos, las lesiones son longitudinales y forman una ralladura de color marrón asemejándose a
una canoa (García, 2006).
2.7.6.3 Sintomatología. Esta enfermedad ataca el tallo principal, ramas, hojas y frutos; en tallos
y ramas se observa una roña color café claro que se localiza sobre tejidos semileñosos, en cuyo
centro se observan puntos negros que corresponden a las estructuras reproductivas del hongo; en
las hojas, la roña se presenta en el pecíolo y a lo largo de las
nervaduras; en los frutos, las lesiones son ligeramente hundidas, secas, de color café claro, de
forma redondeada que al avanzar pueden ir coalesciendo, siguiendo el movimiento del agua
sobre el fruto (Franco y Zuleta, 2006)
2.8 MANEJO DE LA ENFERMEDAD
Todas las labores en el cultivo deben estar dirigidas a disminuir la incidencia de los agentes
causantes de problemas fitosanitarios, eliminando o modificando las condiciones que favorecen
su desarrollo, aumentando la tolerancia de las plantas al ataque de enfermedades, reduciendo en
forma oportuna y preventiva el riesgo de su presencia en el cultivo, antes de que éstas se
conviertan en un foco de contaminación y no puedan ser controlables (Neira, 2006).
La mayoría de los problemas fitosanitarios se pueden manejar llevando a cabo las siguientes
prácticas y labores preventivas:

21
2.8.1 Semillas (Material genético)
La siembra de semilla de buena calidad es el punto de partida para una óptima cosecha de
gulupa. En Colombia hay graves problemas por la deficiente calidad del material vegetal; es
frecuente la presencia de virus, bacterias y hongos. Esto se debe a que la mayoría de la semilla
que se utiliza no es sometida a control de calidad fitosanitaria y generalmente la obtiene el
agricultor de sus propios cultivos, comprada a vecinos, viveristas o proveedores comerciales, sin
que medie ninguna garantía sobre su calidad.
Una buena semilla se selecciona a partir de frutos sanos provenientes de una plantación que no
haya presentado problemas fitosanitarios, que haya demostrado buen rendimiento en las mismas
condiciones agroclimáticas donde se va establecer el cultivo. Además, los frutos deben estar
maduros, tener un buen color, tamaño y peso, y estar libres de daños mecánicos.
La gulupa también se puede reproducir por medio de estacas de unos 15 cm con 2 o 3 entrenudos
o mediante injertos obtenidos de plantas con buenas características. Esta última alternativa se
conoce como propagación vegetativa o asexual y aún es poco utilizada. Es recomendable
adquirir el material de siembra en viveros registrado ante el ICA, que cumplan todos los
requisitos técnicos de calidad y manejo del material (Albert, 1991; ICA 2012)
2.8.2 Siembra
• La preparación del suelo debe evitar intervenciones drásticas que alteren la capa fértil y lo
protejan de la erosión causada por la acción del viento y el agua, es decir, la labranza debe ser
mínima.
• La siembra depende del sistema de tutorado que se vaya a emplear: emparrado o espaldera. El
primero –aunque facilita las labores agronómicas del cultivo y permite frutas de mejor calidad,
evitando el golpe de sol–, favorece el desarrollo de enfermedades debido al microclima
favorable bajo el emparrado. Desde el punto de vista fitosanitario, el sistema de espaldera ofrece

22
ventajas en condiciones de ola invernal, pues permite una mejor aireación y facilita las labores
de aplicación de plaguicidas (Jiménez, Carranza y Rodríguez, 2009).
• La gulupa necesita suelos fértiles (fertilización básica con elementos mayores debe
complementarse con adición de microelementos) y bien drenados. Si los son arcillosos y llueve
mucho, la retención de humedad puede favorecer los problemas fungosos; en suelo pesados, se
recomienda abrir hoyos de mayor tamaño para la siembra y adicionar materia orgánica bien
descompuesta.
• Como la mayoría de los suelos aptos para el cultivo de gulupa son ácidos, debe considerarse la
necesidad de aplicar enmiendas antes de la siembra, de acuerdo con los resultados del análisis de
suelos y bajo la orientación técnica de un profesional.
La información y prácticas agronómicas para el sistema de gulupa son extrapoladas de otros cultivos
de pasifloras
2.8.3 Podas
Dentro del manejo cultural de las enfermedades que ocasionan pudriciones en los frutales, se
destaca la poda. En el cultivo de granadilla, se realizan cuatro tipos de podas conocidas como de
formación, mantenimiento, inducción de floración y renovación. La poda de mantenimiento o
sanitaria consiste en la eliminación de las ramas que ya produjeron cosecha, como también las
improductivas, las enfermas y las secas, con esta práctica se evita el encamamiento, es decir, la
superposición exagerada de cantidad de ramas, que contribuye a la presencia de enfermedades y
a la producción de frutos de baja calidad (Castro, 2001). En el cultivo de maracuyá, también se
realiza la denominada poda de limpieza, en la cual se eliminan las ramas enfermas o dañadas con
el fin de destruir focos de infección, disminuir el peso de la planta sobre la estructura de soporte
(tutorado), mejorar la aireación, favorecer la captación de luz y facilitar la penetración de los
plaguicidas a todas las partes de la planta (García, 2002).

23
Los beneficios sanitarios de la poda han sido demostrados en varios cultivos de frutales. En un
experimento realizado en huertos de manzana orgánica en el Este de Hungría, se obtuvieron los
más bajos valores de severidad de la Roña del Manzano, ocasionada por Venturia inaequalis
Cke., tanto en hojas como en frutos en las parcelas donde se realizó remoción de hojas junto con
una poda en invierno, logrando reducir el uso de fungicidas a base de cobre (Holb, 2008). En
otro experimento llevado a cabo en un viñedo cerca a Montpellier (Francia), se encontró una
correlación positiva entre la incidencia del moho gris (Botrytis cinerea Pers.) y la densidad del
follaje en la zona de fructificación, debido a que una alta densidad del canopy está asociada con
un microclima más favorable para el desarrollo de la enfermedad (Valdés et al., 2008).
2.8.4 Fertilización
Se elabora a partir del análisis de suelo y bajo la orientación técnica de un ingeniero agrónomo.
El plan de fertilización debe tener en cuenta también el desarrollo del cultivo y las necesidades
de las plantas, especialmente en etapa de formación de frutos. La fertilización básica con
elementos mayores debe complementarse con adición de microelementos, específicamente boro
y magnesio para evitar deficiencias en el cultivo (Angulo, 2010).
2.8.5 Manejo de arvenses
Estas plantas compiten por nutrientes con las plantas de gulupa, dificultan las podas, la
fertilización. Pese a lo anterior, no se recomienda limpiar totalmente el cultivo de las arvenses,
debido a que estas plantas bien manejadas pueden ayudar a retener humedad y a frenar la
erosión, especialmente en terrenos de ladera. Los primeros meses, después de establecido el
cultivo, son los de mayor competencia por las arvenses y requieren control periódico; después,
esa competencia se reduce y el control depende de la invasión favorecida por los períodos de
lluvia (Tamayo, 2004)

24
2.8.6 Cosecha
La cosecha de los frutos de gulupa debe hacerse cuando estos presentan su epidermis con un
50% de color morado y un 50% de color verde; este es el indicador visual de la madurez
fisiológica y del mayor peso de los frutos. La recolección debe hacerse en horas de la mañana,
generalmente en forma semanal, pero dependiendo de las exigencias del mercado y de las
condiciones climáticas, se puede cosechar con más frecuencia; en épocas de lluvia se recomienda
hacerlo día de por medio. Los frutos se deben cosechar secos o si están húmedos se deben
cubrir con papel periódico para evitar su deterioro.
La preselección de frutos debe hacerse tan pronto son cosechados, separando frutos enfermos o
dañados por insectos. El empaque se hace en cajas de cartón o en canastillas de plástico, las
cuales deben haber sido desinfectadas para evitar la contaminación de los frutos. Se recomienda
colocar papel periódico entre las capas de fruta para evitar dañar la epidermis (Garcia, 2002).
2.8.7 Manejo químico
En cuanto al control químico de las enfermedades de las pasifloras se han realizado pocos
estudios. En Kenya, probaron varios fungicidas para el control de la mancha parda causada por
A. passiflorae, y encontraron que el ingrediente activo mancozeb (Manzate D y Dithane M-45)
era uno de los más efectivos para el control de esta enfermedad, en aplicaciones cada 2-3
semanas. Aunque los investigadores no midieron el efecto de la poda, mencionan que es muy
aconsejable no sólo porque mejora la penetración de las aspersiones sino también porque ayuda a
que el viento y el sol sequen más rápido el follaje, creando así un ambiente desfavorable para la
dispersión de la enfermedad (Ondieki et al, 1971).
En Australia evaluaron el efecto de la combinación de fungicidas del grupo de las estrobirulinas
(principalmente azoxystrobin) y un análogo funcional del ácido salicílico (acibenzolar) en el
control de la roña de la gulupa ocasionada por Cladosporium oxysporum Berk., los resultados
obtenidos mostraron un control efectivo de la enfermedad en campo por parte de las
estrobirulinas (Willingham et al., 2002)

25
Trifloxystrobin es un fungicida perteneciente al grupo de las estrobilurinas. Esta clase de
fungicidas tiene actividad contra un amplio rango de hongos filamentosos (Clough y Godfrey,
1998), el mecanismo de acción de este grupo de fungicidas es el bloqueo de la respiración.
Trifloxystrobin es un fungicida mesostémico, altamente activo, que brinda un control efectivo no
sólo en las plantas tratadas sino también en las plantas vecinas a ellas; tiene una alta afinidad con
la superficie vegetal, siendo absorbido y acumulado en las capas cerosas de la misma, quedando
protegido del lavado de las lluvias; desde aquí se redistribuye en la superficie de la planta por el
movimiento superficial en fase de vapor con posterior re-deposición; el producto penetra al
tejido de la planta, tiene actividad translaminar, existiendo poco o nada de transporte dentro del
sistema vascular (Moraga, 2003). Debido a esta última característica esta estrobilurina tiene un
modo de acción básicamente preventivo (Ocampo, 2010).
Los triazoles son una clase de fungicidas que afecta las células de los hongos a través de la
inhibición de la biosíntesis de los esteroles, y dentro de esta clase pertenece al grupo de los
inhibidores de la demetilación del C-14 (Uesugi, 1998). Tebuconazol, un miembro de este grupo,
es un fungicida sistémico y de contacto de largo efecto residual, el cual puede ser utilizado en
estrategias de control preventivo, curativo y erradicante (Moraga, 2003); triadimenol, al igual
que tebuconazol, hace parte del grupo de los triazoles, es un fungicida sistémico y de amplio
espectro, que es rápidamente absorbido por la planta y translocado dentro de ella. Tebuconazol
se ubica sobre los haces vasculares centrales (xilema y floema) mientras que triadimenol se
mueve más por la nervadura y los haces conductores de las hojas, esta característica le confiere
una mayor movilidad al primero (Ocampo, 2010).
Las Buenas Prácticas Agrícolas (BPA) se fundamentan en obtener productos que no afecten la
salud del consumidor, y que durante su proceso de producción no se deteriore el medio ambiente
y se asegure el bienestar laboral. El concepto de las BPA surgió de la iniciativa de los
distribuidores mayoristas de la Unión Europea (UE) en el año 1997, y se concretó a través de una
norma commercial de adopción voluntaria llamada EurepGap, la cual fue diseñada inicialmente
para los productores europeos (Marín, 2007); actualmente esta norma es conocida como
GlobalGap (GLOBALGAP, 2007). La aplicación de ésta requiere asegurar la trazabilidad de los

26
productos, diseñar y mantener un sistema de registros, cumplimiento de la regulación oficial,
diseñar un plan de Manejo Integrado de Plagas y Enfermedades (MIPE), uso correcto de
plaguicidas, entre otros (Villamil, 2008).
El manejo integrado de plagas (MIP) es una alternativa viable que la Unión Europea definió en
1991 como “la aplicación racional de una combinación de medidas biológicas, biotecnológicas,
químicas, de cultivo o de selección de vegetales de modo que la utilización de productos
fitosanitarios químicos se limite al mínimo necesario para mantener la población de la plaga en
niveles inferiors a los que producirían daños o pérdidas inaceptables desde un punto de vista
económico”. La regulación nacional, basada en los requerimientos internaciones para el uso de
plaguicidas, afirma que sólo se puede usar aquéllos registrados en el país de uso para el cultivo a
tratar y la plaga específica para la cual fueron registrados (Lugo, 2007); este es un serio
problema para el caso colombiano ya que los plaguicidas registrados para pasifloráceas son muy
escasos.
Teniendo en cuenta el manejo químico de la enfermedad, el riesgo de desarrollo de resistencia
por parte de los patógenos a estos ingredientes activos, el marco actual de la producción en un
ambiente limpio y la ausencia de resistencia o tolerancia genética de la gulupa a la Roña, se ha
promovido el desarrollo y evaluación de nuevas estrategias de manejo de las enfermedades para
el reemplazo de fungicidas en el manejo integrado, logrando hacer la producción
económicamente viable y sostenible
Castaño et al (2008), evaluaron la eficacia in vitro de los lixiviados de raquis de plátano y los
fungicidas Propiconazol y Clorotalonil sobre Paracercospora fijiensis, anamorfo de
Mycosphaerella fijiensis, El efecto de los lixiviados, 15 días después de la siembra tuvo una
mayor reducción en el número y tamaño de colonias del hongo con los lixiviados con el 90% de
agua evaporada con respecto a los demás tratamientos, mientras que la esporulación y
germinación de conidios con este tratamiento fueron reducidas de manera similar que con los
fungicidas.

27
Otro trabajo similar donde evaluaron extractos metanolicos y de diclorometano, sobre las fases
de reproducción de Mycosphaerella fijiensis, se encontró que un 30% de los extractos
metanolicos y un 76.2 % de los extractos de diclorometano evaluados, causaron deformidad en
los tubos germinativos de las ascosporas de M fijiensis. (Viveros, 2006)
Fayette (2007), utilizo 86 extractos etanólicos liofilizados a partir de raíces, hojas, tallos, flores,
frutos y rizomas. Se evaluó el efecto antimicrobiano de extractos etanólicos a una concentración
de 250 mg/L en dimetilsulfóxido sobre el crecimiento in vitro de Xanthomonas axonopodis
pv.vesicatoria, Phytophthora palmivora y Colletotrichum gloeosporioides.
Los extractos a base de caléndula JF/10/40A, JF/10/40B, JF/10/40C, JF/10/40D, JF/10/24A y
JF/10/24C mostraron actividad inhibitoria a una concentración de 250 mg/L sobre el crecimiento
de X. axonopodis pv. vesicatoria. La mínima concentración que induce actividad inhibitoria fue
de 50 mg/L para cada extracto. Estos resultados muestran que las especies vegetales de donde
provienen estos extractos son potenciales fuentes de compuestos antimicrobianos. Sin embargo,
ninguno de los extractos evaluados mostró actividad inhibitoria sobre el crecimiento micelial de
P. palmivora y C. gloeosporioides.
Mendoza (2006), evaluaron el efecto antifúngico de los extractos sobre el crecimiento in vitro de
Phytophthora palmivora Butl. y Colletotrichum gloeosporioides (Penz.) Penz. & Sacc., hongos
fitopatógenos aislados de la mazorca de cacao (Theobroma cacao L.) y de frutos de mango
(Mangifera indica L.), respectivamente y adicionalmente, se evaluaron 64 extractos más. Como
control positivo, se utilizó metalaxyl-M + mancozeb (2 %) para P. palmivora y mancozeb (2 %)
para C. gloeosporioides. Ninguno de los extractos vegetales mostró inhibición del crecimiento
micelial de los patógenos estudiados.
Rojas (2008) evaluó el efecto biofungicida in vitro donde reveló la disminución de la velocidad
de crecimiento del hongo, considerando el extracto de fique al 5 y 10% con el mejor efecto
inhibitorio, incluso en las concentraciones de 0.5 y 1%, que tuvieron mejor acción antifúngica
que los extractos de swinglea y el LRP en sus diferentes concentraciones. Entre el extracto de
swinglea en solución acuosa y el LRP al 5% no hubo diferencias significativas, mostrándose

28
estos sin un efecto marcado sobre la inhibición del crecimiento, así como, el LRP al 25, 50 y
100% que ocasionaron la menor inhibición en el desarrollo de los aislamientos.
Álvarez et al. (2000, 2002), utilizaron el extracto de hojas de swinglea (Swinglea glutinosa
(Blanco) Merr.), preparado en agua o etanol, para el control efectivo del Mildeo polvoso de la
rosa ocasionado por Sphaerotheca pannosa (Vallr.: Fr.) Lev. var. rosae Woronichin; también se
utilizo el ectracti de hojas swinglea para la Antracnosis ocasionada por Colletotrichum
lindemuthianum (Sacc. & Magnus) Lams.- Scrib., y el Añublo causado por Ascochyta spp. en
fríjol (Pastor-Corrales, 1991) y la Roya del cafeto ocasionada por Hemileia vastatrix (Ramírez y
Ñañez, 2002)
López y Salazar (2001), evaluaron diferentes concentraciones de extracto de fique (Furcraea
macrophylla Baker) para determinar el efecto de este sobre la germinación in vitro de
esporangios de Phytophthora infestans (Mont.) de Bary aislado de tomate de mesa, y la
inhibición en plantas de tomate infectadas con el patógeno; concluyendo que el extracto de fique
posee propiedades fungicidas aprovechables para el manejo de la enfermedad. El efecto
fungicidas sobre Colletotrichum gloeosporioides aislado de tomate de árbol también ha sido
registrado (Gómez, 2002).
El lixiviado de compost de raquis de plátano ha sido utilizado en el control de Moko del plátano
ocasionado por Ralstonia solanacearum (Smith 1896) Yabuuchi et al. raza 2 (CIAT, 2002) y
Mildeo polvoso de la rosa ocasionado por S. pannosa (Álvarez et al., 2001; Álvarez et al., 2002).
Con la utilización de ácidos fúlvicos producidos por la degradación de raquis de plátano
Dominico Hartón y aplicados foliarmente a diferentes dosis para el manejo de sigatokas
ocasionadas por Mycosphaerella spp. en la cv. Dominico Hartón, Escobar y Castaño-Zapata
(2005), determinaron una buena actividad con aplicaciones al 0.5%. Estos extractos también
mostraron ser efectivos al ser probados en el manejo de Sigatoka negra ocasionada por
Mycosphaerella fijiensis Morelet en banano (García y Apezteguia, 2001).

29
Se evaluó la actividad antifúngica de los aceites esenciales de Eucalyptus sp (Myrtaceae) y
cascara de naranja, Citrus sinensis (Rutaceae) a diferentes concentraciones frente a los modelos
biológicos: Trichoderma harzianum, Absidia sp y Fusarium oxysporum,. El aceite esencial de
eucalipto inhibió completamente el crecimiento de este hongo a 3000 ppm, mostrando mayor
potencial inhibitorio que el fungicida comercial Dithane empleado a 10000 ppm. (Murillo, 2009)
Estos biopreparados representan una alternativa de manejo ambientalmente amigable y segura,
que permite ampliar las fronteras de comercialización de una de las frutas colombianas más
promisorias en el mercado nacional e internacional (Alvarez, 2002).
En la década de 1990 se inició la búsqueda de métodos biológicos efectivos de protección contra
los hongos patógenos. Se han realizado estudios en condiciones de laboratorio, ensayos
semicontrolados y de campo hasta lograr la obtención de biopreparados de Trichoderma spp., los
cuales hoy en día ocupan un lugar importante dentro del manejo de los patógenos del suelo
(Stefanova 1997).
El potencial de especies de Trichoderma como agentes biocontroladores de enfermedades de
plantas fue reconocido en los años 1930 (Weindling 1934), principalmente en microorganismos
del suelo de los géneros Phytophthora, Rhizoctonia, Sclerotium, Pythium y Fusarium entre otros,
sin embargo se están realizando investigaciones para el control de muchos otros
microorganismos que causan problemas en cultivos de importancia económica y en años
siguientes el control de muchas enfermedades ha sido agregado a la lista (Howell 2003). Por
ejemplo, se ha demostrado el efecto antagónico de Trichoderma spp. sobre algunas especies de
Phytophthora en yuca, en pruebas in vitro, en invernadero (Alvarez et al. 2003) y campo
reduciendo significativamente la pudrición radical causada por este microorganismo (Álvarez et
al. 2008). Igualmente, se ha comprobado que la aspersión de Trichoderma harzianum (Rifai)
controla B. cinerea en cultivos de uva (Latorre et al. 1997).

30
2.9 MARCADORES MOLECULARES
La variabilidad genética de hongos fitopatógenos y material vegetal, puede ser evaluada
mediante caracteres morfológicos e isoenzimáticos o con marcadores moleculares, que son
secuencias de ADN que se encuentran en el genoma. Estos caracteres son llamados marcadores
debido a que pueden ser utilizados directamente para obtener información acerca de la genética
de caracteres de interés del organismo en estudio
Los marcadores moleculares tales como el polimorfismo en la longitud de los fragmentos de
restricción (RFLP), ADN polimórfico amplificado al azar (RAPD), microsatélites e isoenzimas
han sido desarrollados para estudiar la diversidad genética y domesticación, establecer mapas
moleculares, caracterizar variedades y ayudar en los programas de mejoramiento.
Los marcadores moleculares no son afectados por el ambiente y no varían con la edad de la
planta; pueden ser regiones de ADN codificantes (exones) o no codificantes (intrones). Se han
desarrollado varios tipos de marcadores moleculares basados en la técnica de PCR (Polymerase
change reaction, por sus siglas en inglés), la cual permite elaborar un número ilimitado de copias
de cualquier fragmento de ADN; a partir de una sola molécula de ADN, la PCR puede generar
100.000 millones de moléculas idénticas.
Los marcadores moleculares se han usado para mapeo genético, mejoramiento, estudios de
diversidad genética, flujo de genes y huellas genéticas. Los polimorfismos detectados mediante
RFLPs y AFLPs se deben a variaciones en sitios de corte por enzimas de restricción. Mientras
que los polimorfismos detectados mediante RAPDs y microsatélites se deben al número de
secuencias repetidas que se encuentran entre las regiones conservadas; esta variación en las
secuencias de ADN donde se alinean los cebadores y la diferencia en la longitud del ADN
amplificado (CIAT, 1999), permite identificar especies que son morfológicamente similares y
puede confirmar la identificación realizada con base en características morfológicas y
patogénicas (Iriarte, 2002).

31
2.9.1. ITS (Región espaciadora interna transcrita)
Debido a que las características morfológicas para la identificación de las especies del género
Cladosporium pueden llegar a ser altamente variables y por lo tanto inconsistentes,
Construcciones de cebadores especie específicos, como los ITS (Internal Transcribed Spacer por
sus siglas en inglés) del ADN ribosomal (ADNr) han sido propuestos como los más eficientes y
el sistema más confiable para la detección y diferenciación de las especies de Cladosporium
(Ureña–Padilla et al. 2002), por encima de los polimorfismos en la longitud de fragmentos de
restricción (RFLP), análisis de regiones ricas en A + T y ADN polimórfico amplificado al azar
(RAPD) (Saldarriaga 2005).
La región del espaciador interno transcrito ITS es no codificadora y variable, mientras que el gen
RNAr 5.8S es codificante y conservado, ambos son útiles para determinar las relaciones
filogenéticas de los hongos, debido a que las regiones ribosomales muestran un bajo
polimorfismo intraespecífico y una alta variabilidad interespecífica (Martínez - Culebras et al.
2000). La región 5.8 S - ITS ha sido empleada en diversos trabajos para identificar varias
especies, como en el reportado por Martínez- Culebras et al. (2000) en el que patrones generados
mediante la restricción de la región ITS1, ITS2 y 5.8S de 80
2.9.3. Microsatélites RAMS
Los marcadores moleculares tipo RAMS, son útiles para determinar la diversidad genética en
plantas y animales debido a que permiten seleccionar regiones al azar dentro de la molécula de
ADN, el número de polimorfismos detectables es teóricamente ilimitado y permiten analizar
tanto la información que se expresa como la que no lo hace (Morillo et al. 2005).
En general, los microsatélites son regiones de secuencias de ADN pequeñas y repetidas,
generalmente conformadas de dos a tres nucleótidos. Dado que la repetición por sí misma no
codifica para ningún producto, y debido a que las secuencias de ADN repetitivo pueden
recombinarse y expandirse más frecuentemente que otros tipos de secuencia, estas regiones son a
menudo altamente variables y muy útiles para medir similitudes entre especies o variedades
relacionadas. Este método es muy eficiente y proporciona tantos datos para estudios entre

32
especies, como microsatélites resulten del alto grado de mutaciones, sin embargo, la necesidad
de clonación, secuenciación y síntesis de los cebadores específicos de la especie, reduce su
universalidad. (Henríquez 2003)
La técnica RAMS combina los beneficios del análisis microsatélite con los del universalmente
análisis RAPD, Hantula et al. (1996) seleccionaron cuatro iniciadores (GT, ACA, CCA, CGA)
que fueron diseñados teniendo una longitud de 18 bases incluyendo un extremo 5´ degenerado, el
cual sirve de anclaje para asegurar la unión del cebador al inicio del microsatélite. La técnica se
ha aplicado para generar marcadores de ADN de los hongos Armillaria cepistipes, Gremmeniella
abietina, Heterobasidion annosum, Phytophthora cactorum, Phlebiopsis gigantea, Stereum
sanguinolentum, Colletotrichum spp., entre otros. La técnica es reproducible, y aplicable a todos
los hongos, incluyendo miembros de los Phycomycetes, Ascomycetes y Basidiomycetes; lo que
permite detectar polimorfismos de ADN interespecíficos e intraespecíficos (Henríquez 2003).

33
3. OBJETIVOS
3.1 OBJETIVO GENERAL
Identificar estrategias de manejo de la roña (Cladosporium spp) en gulupa (Passiflora edulis pv
edulis), con base en biofungicidas.
3.2 OBJETIVOS ESPECÍFICOS
Estandarizar pruebas de patogenicidad en laboratorio, y caracterizar patogénicamente
aislamientos de Cladosporium spp. asociados a la roña en cultivos de gulupa.
Caracterizar patogénica, y molecularmente los aislados de Cladosporium spp.
procedentes de varias regiones de Colombia.
Determinar mediante la técnica molecular RAMS, la variabilidad intraespecífica de
aislamientos de Cladosporium spp. causantes de la roña en gulupa.
Evaluar in vitro la actividad de diferentes biofungicidas contra aislamientos patogénicos
y molecularmente de Cladosporium spp.
Evaluar frutos de gulupa bajo condiciones en in vitro por su resistencia a Cladosporium
spp.

34
4. ASPECTOS METODOLÓGICOS
4.1 Recolección de material vegetal y aislamiento de cepas
En el marco del proyecto “Promoción de manejo integrado de plagas (enfocado en plagas
dipteras y roña) y desarrollo de paquetes de buenas prácticas agrícolas en los cultivos de
maracuya, grandilla y gulupa” se realizaron los ensayos de obtención y purificación de
aislamientos. Estos se llevaron a cabo en el laboratorio de Fitopatología del programa de
Agronomía de la Facultad de Ciencias Agropecuarias de la Universidad de Caldas a cargo del
Fitopatólogo Carlos Germán Delgado Méndez ( Tabla 3).
La caracterización patogénica y molecular de los aislamientos se realizó en el Laboratorio de
Patología de Yuca del Centro Internacional de Agricultura Tropical, ubicado en la ciudad de
Palmira, a una altura de 965 msnm, temperatura promedio 24ºC, humedad relativa de 76% y una
precipitación media anual de 965mm.
4.2.1 Reactivación de los aislamientos
Las cepas a reactivar se encontraban conservadas en papel filtro estéril y almacenados a -20 C y
-4C. Dichos asilamientos se llevaron a una cámara de flujo laminar en donde se tomaron
pequeños trozos de papel filtro que contenían el aislamiento a reactivar. Estos fueron sembrados
en cajas de petri que contenian agar (PDA) y se incubaron a 24° durante 6 días. Del crecimiento
allí obtenido se hicieron réplicas de cada aislamiento para la evaluación morfológica y para la
producción de inoculo para las pruebas de patogenicidad.

35
Tabla 3. Colección, codificación e identificación de aislamientos con síntomas de Roña.
Tabla 1
Ítem Código Departamento Municipio Vereda Hospedante Aislamiento
1 HUPEC-GU-1 Huila Pitalito El Cedro Gulupa Cladosporium
2 HUA-GU-1 Huila La
Argentina Gulupa Cladosporium
3 HUGA-GU-1 Huila Guadalupe Australia Gulupa Cladosporium
4 HUALG-GU-
1 Huila El Agrado
La
Guacana Gulupa Cladosporium
5 HUB-GU-1 Huila Belén Gulupa Cladosporium
6 VAYJ-GU-2 Valle Yotocó Jiguales Gulupa Cladosporium
7 BOJ-GU-1 Boyacá Jenesano Gulupa Cladosporium
8 BOB-GU-1 Boyacá Buenavista Gulupa Cladosporium
9 BOB-GU-2 Boyacá Buenavista Gulupa Cladosporium
10 BOBER-GU-1 Boyacá Buenavista El
Recuerdo Gulupa
Cladosporium y
Colletotrichum
11 CUCSL-GU-1 Cundinamarca Chioachi San Luis Gulupa Cladosporium
12 TOILM-GU-1 Tolima Ibagué La María Gulupa Cladosporium
13 CALT-GU-1 Caldas Manizales Tesorito Gulupa Cladosporium
14 *ANTR-GU-1 Antioquia Rionegro El Jardin Gulupa Cladosporium
15 *ANTR-GU-1 Antioquia Rionegro El
Poblado Gulupa Cladosporium
*Obtencion y purificación de aislamientos realizados en CIAT
4.2.3 Aislamientos de cepas de Cladosporium spp
De las muestras registradas y codificadas adecuadamente, se procedió a realizar el aislamiento de
Cladosporium spp, siguiendo el método directo para aislamiento de microorganismos
fitopatógenos (Castaño-Zapata, 1997) con algunas modificaciones; con el objetivo de conformar
un banco de aislamientos monospórico.

36
4.2.4 Desinfestación del material vegetal
LOS AISLAMIENTOS SE OBTUVIERON A PARTIR DE TEJIDO VEGETAL AFECTADO. LAS MUESTRAS SE COLECTARON EN CAMPOS EN LOS
CUALES LAS PLANTAS PRESENTABAN LESIONES DEPRIMIDAS Y CIRCULARES EN LOS FRUTOS. POSTERIOR A LA DESCRIPCIÓN DE LA
SINTOMATOLOGÍA OBSERVADA EN EL TEJIDO A ANALIZAR, SE CORTARON CINCO FRAGMENTOS DE TEJIDO VEGETAL DE APROXIMADAMENTE
0.25 CM2, LOS CUALES DEBÍAN ESTAR COMPUESTOS DE TEJIDO SANO Y TEJIDO ENFERMO; TENIENDO LOS TROZOS ASÍ CORTADOS, SE
PROCEDIÓ A DESINFESTARLOS DE LA SIGUIENTE FORMA: EN UN RECIPIENTE DE VIDRIO, MARCADO CON EL CÓDIGO RESPECTIVO Y TAPADO
CON GASA DOBLE Y SUJETADA CON BANDAS ELÁSTICAS SE LAVÓ EL MATERIAL VEGETAL; EL MONTAJE ASÍ REALIZADO SE COLOCARON BAJO EL
CHORRO DE AGUA CORRIENTE O DESTILADA PARA EFECTUAR EL LAVADO DE LOS TROCITOS POR 20 MINUTOS. PASADO EL TIEMPO DE
LAVADO, SE RETIRÓ EL AGUA Y SE PROCEDIÓ A HACER LA DESINFESTACIÓN EN CÁMARA DE FLUJO LAMINAR, CON LA ADICIÓN DE
HIPOCLORITO DE SODIO AL 2% DURANTE 2 MINUTOS Y EN AGITACIÓN CONTINUA; PASADOS LOS 2 MINUTOS, SE RETIRÓ EL HIPOCLORITO Y
SE ADICIONO ETANOL AL 70%, PERMITIENDO QUE ACTÚE POR 2 MINUTOS Y AGITANDO CONSTANTEMENTE. AL FINALIZAR LA
DESINFESTACIÓN, SE LAVARON 3 VECES CONSECUTIVAS CON AGUA DESTILADA ESTÉRIL Y CON PINZAS METÁLICAS ESTÉRILES SE COLOCARON
SOBRE TOALLAS DE PAPEL ESTÉRIL, PARA ELIMINAR EL EXCESO DE AGUA.
4.2.5 Siembra directa en medio de cultivo
Cuando el material vegetal estuvieron secos, se procede a tomar con pinzas metálicas estériles, 5
trocitos son colocados por separados y perpendiculares a la superficie seca de agar Papa-
Dextrosa (39 g agar PDA, 1000 mL agua destilada) en caja Petri de 10 cm. de diámetro; cada
análisis se efectuará por duplicado.
Las cajas así sembradas fueron incubadas invertidas a 28C / 3-5 días y 24 horas de luz.
4.2.6 Purificación
Finalizado el tiempo de incubación, con asa micológica esterilizada a la llama, se tomaron
fragmentos de los crecimientos presuntivos desarrollados de Cladosporium spp en cada uno de
los trocitos y se transfirieron a medio nuevo con ácido láctico (PDA+AL25%). Es importante la
transferencia de todos los crecimientos presuntivos, para determinar posteriormente diferencias
macroscópicas en las colonias desarrolladas procedentes de la misma muestra y obtener varios o
un único aislamiento.

37
Bajo el estereoscopio se observó la esporulación típica del hongo sobre los trozos o en el
crecimiento desarrollado, con una microaguja estéril se toman cirros de los cuerpos fructíferos
desarrollados y se transfirieron a nuevo medio PDA+AL25%.
Las cajas Petri con las colonias presuntivas o conidias de Cladosporium spp así transferidas, se
incubaron a una temperatura de 28C / 24h luz por un tiempo de 3 a 5 días o hasta que se pueda
determinar claramente la presencia del hongo.
4.2.7 Obtención de cultivos monospóricos
De los cultivos purificados en los cuales se determino por características macroscópicas la
presencia de Cladosporium spp, inicialmente se hizo una observación del aislamiento utilizando
microscopio binocular compuesto y como colorante azul de lactofenol, con el objetivo de
confirmar mediante características morfológicas del hongo.
Cuando se confirmo la presencia de Cladosporium spp, se procedió a tomar con microaguja
estéril un cirro de esporas, el cual fue depositado en un tubo para microcentrífuga de 1.5 mL
(Eppendorf) al cual previamente se le ha adiciono 1.0 mL de agua destilada estéril; se
homogenizo en vórtex y se tomo 5 L de esta mezcla colocándola en portaobjetos para realizar
la observación al microscopio compuesto la presencia de esporas típicas del hongo y poder
establecer una concentración mínima de ellas.
De la mezcla realizada en los tubos para microcentrífuga, se tomó alícuotas con asa
bacteriológica de argolla las cuales se colocaron separadamente en la superficie seca de una
placa de agar agua 2% en caja Petri de 10 cm.; estas alícuotas se distribuyeron utilizando una asa
de vidrio sobre toda la superficie del agar, buscando distribuir de manera homogénea los
conidios del hongo permitiendo que estas queden separadas. Las cajas así inoculadas se
incubaron a una temperatura de 28C / 24h luz por un tiempo 18-24 horas.
Pasado el tiempo de incubación, se hizo la observación de las cajas sembradas utilizando para
ello estereoscopio. En las esporas observadas germinadas aisladamente, se tomó con microaguja

38
estéril el segmento de agar y se confirmó la presencia de sólo una espora al microscopio de luz;
esta espora se siembra en medio agar avena 4% (OA+S) modificado por adición con
estreptomicina (Agar 15 gr/L, avena molida 40 gr/L, estreptomicina 300 mg/L, 1000 mL agua
destilada), medio recomendado por Silveira et al (2004), por el buen crecimiento y esporulación
de C. cladosporioides spp.
Las cajas sembradas con 5 esporas cada una (para detectar posibles diferencias en crecimiento
y/o asegurar la germinación total de al menos una de ellas), se incubaron invertidas a 28C / 24h
luz por 3-5 días. De cada una las colonias crecidas, se tomo independientemente con una asa
micológica, se transfirieron a cajas petri de agar avena 4% y se incubaron bajo las mismas
condiciones, permitiendo un buen desarrollo de micelio y esporas.
4.2.8 Pruebas de patogenicidad: Postulados de Koch
Con el fin de determinar si los aislamientos obtenidos a partir de hojas y frutos de gulupa
afectados por roña, es el causante de este disturbio, se realizaron pruebas de patogenicidad in
vitro. Estas pruebas son rápidas, efectivas y facilitan la detección de un agente infeccioso gracias
a la rapidez con que se pueden observar sus efectos (Agrios, 2005).
4.2.9 Preparación de los frutos y del material antes de la inoculación
Se recibieron frutos provenientes de una misma finca del municipio de Santa Rosa de Cabal
(Risaralda), los cuales se dividieron en cinco grupos usando como criterio similitudes en tamaño
y grado de madurez. A cada grupo fue asignado un número de repetición. 1 fue el más maduro,
siguieron los grupos 2 y 3 y luego los grupos 4 y 5 que fueron los menos maduros. Los grupos 3
y 5 fueron de tamaño un poco superior a los de los grupos 2 y 4. El número del grupo se colocó
al otro lado del pedúnculo.
Los frutos fueron lavados con agua corriente y detergente, después fueron sumergidos en
hipoclorito al 2,5 % durante 2 minutos, luego en alcohol al 70% durante 5 minutos y finalmente

39
fueron lavados con abundante agua. Una vez realizado este procedimiento se dejaron secar a
temperatura ambiente.
4.2.10 Ajuste de la metodología de inoculación de frutos de gulupa con Cladosporium spp
Para la estandarización y ajuste de metodología de inoculación de frutos de gulupa se realizaron
tres métodos de inoculación.
El primer método consistía en inocular frutos de gulupa sanos, a cada uno de ellos se le hicieron
punciones en cinco áreas de aproximadamente 5mm de diámetro con una aguja de disección
estéril. En cada una de esas áreas se aplicaron alícuotas de 1µ, 5µ y 10µ de suspensión de
esporas 1x 10 6. Una vez realizado este procedimiento los frutos fueron depositados en cajas
plásticas con tapas (Estra REF 2-15-272, las cuales contenían 20 mL de agua destilada estéril y
una rejilla plástica sobre la cual se colóca el tejido inoculado evitando que haya contacto de la
superficie del fruto con el agua. Esta caja se cierra herméticamente para realizar una cámara
húmeda y favorecer el desarrollo de la infección.
Para la segunda metodología se procedió en inocular frutos de gulupa, con ayuda de un
sacabocado estéril se marcaron circulos de aproximadamente 6 mm de diámetro sobre las
colonias de Cladosporium spp reactivadas en agar Marthur. Con el mismo sacabocado estéril se
hicieron 2 orificios opuestos en la zona ecuatorial de las frutas. Cada uno de los orificios fue
marcado con la letra a y b, en cada uno se depositó un disco de agar con el hongo, cubriendo el
área afectada con cinta PARAFILM "M' Chicago. IL, 60631. Igualmente fueron colocados en
cajas plásticas con tapa con sus respectivos nombres.
Para la tercera inoculación a cada fruto se les realizo punciones y posteriormente se asperjaron
con una solución acuosa de esporas a una concentración de 1x106 esporas / mL de agua. Para
este método se utilizó un aspersor de agua de plástico (123RF 400 x 267 aprox). Igualmente se
colocaron los frutos en sus respectivas cajas. Cada ensayo piloto se inocularon 3 frutos por
aislamiento, y se consideró cada fruto una repetición.

40
4.2.11. Pruebas de patogenicidad
Con el fin de evaluar la patogenicidad de todos los aislamientos obtenidos de muestras de
gulupa con la enfermedad, y establecer diferencias entre ellos, se realizaron inoculaciones
artificiales con el método de aspersión de esporas. Previo a esto todos los frutos del grupo 5
fueron sometidos al proceso de desinfección.
Se inocularon seis frutos por aislamiento y se consideró cada fruto una repetición, posteriormente
fueron incubados en una caja plástica con tapa (Estra REF 2-15-272), la cual estaba
acondicionada con unas rejillas plásticas y una película de agua destilada en el fondo, para hacer
las veces de una cámara húmeda.
4.2.11 Evaluación de la enfermedad
Se realizaron evaluaciones al día 1, 2, 3, 6, 7, 9, y 12 después de la inoculación.
La incidencia y la severidad se determinó así:
1. Se contaron todos los frutos inoculados con cada aislamiento.
2. Para el cálculo de incidencia, se utilizó la siguiente fórmula:
Nº de frutos con síntomas de la enfermedad x 100
Nº total de frutos
3. La determinación de la severidad se hizo con base en el porcentaje de área afectada (figura 1).
La escala de severidad fue diseñada por Mora (2011).
Figura 1. Escala de severidad en frutos de gulupa

41
Fuente: Mora (2011)
5.0. Pruebas moleculares
5.1. Extracción de ADN La extracción se hizo de la siguiente forma, siguiendo el protocolo reportado por Álvarez et al.
(2004) con algunas modificaciones:
1. Se pesó 0.1 g de tejido macerado (equivalente a 100 μL) y se le adicionó 600 μL de buffer de
extracción más 1 μL de proteinasa K (10 mg/mL), posteriormente se mezcló en un vortex hasta
obtener una solución homogénea y se incubó a 65°C por 1 h.
2. Se adicionó 200 μL de acetato de amonio 7.5 M dejando a temperatura ambiente por 10 min.
3. Se centrifugó a 13.400 g por 20 min a temperatura ambiente, luego se transfirió el
sobrenadante a otro tubo y se le adicionó igual volumen de cloroformo: isoamil alcohol (24:1)
para luego mezclar por inversión durante 15 min.
4. Después se centrifugó a 13.400 g por 5 min a 4°C y se recuperó el sobrenadante.
5. Se repitió el lavado con cloroformo: isoamil alcohol 2 veces.
6. Se transfirió el sobrenadante a un tubo nuevo, se le adicionó igual volumen de isopropanol frío
y se llevó a -20°C durante toda la noche. Se centrifugó a 13.400 g por 10 min y se descartó el
sobrenadante.

42
7. Se lavó la pelotilla o pellet con 500 μL de etanol frío al 70%, golpeando suavemente para
desprender el pellet del tubo, luego se centrifugó a 13.400 g por 5 min a 4°C y se lavó 2 v eces
descartando siempre el sobrenadante.
8. Se secó por 1 h a 37°C en incubadora con los tubos invertidos sobre una toalla.
9. Finalmente se resuspendió en 50 μL de TE (Tris-EDTA) y se le adicionó 1 μL de RNAsa y se
incubó a 37°C durante 1 h. Luego se almacenaron las muestras a 4°C.
El ADN total extraído de cada aislamiento fue visualizado en geles de agarosa al 0.8%,
adicionado con Gel red 10000X (Biotium), 2 μL por cada 100 mL, mediante electroforesis a 100
voltios / 40 min., y visualizados bajo luz ultravioleta en un analizador de imágenes Eagle eye II
de Strategene, con el objetivo de verificar su integridad. El ADN fue cuantificado en un
espectrofotómetro (Nanodrop™ 2000C Technologies, Wilmington, DE) y luego se ajustó su
concentración de 5 ng/µL (solución de trabajo).
Para la obtención de micelio del hongo, cada aislamiento fue reactivado en medio PDA por 8
días, y a partir de estas colonias se tomaron dos discos de agar con micelio y se sembraron en
medio PDA líquido más ampicilina y estreptomicina sin agitación, bajo condiciones de luz y
temperatura ambiente (±24ºC; 10 horas de luz y 14 de oscuridad), al cabo de 10 días se cosechó
el micelio por filtración al vacío, se secó en cajas de petri estériles entreabiertas en un horno a
36ºC durante 3 días; el micelio seco se maceró en un mortero con nitrógeno líquido hasta obtener
un polvo muy fino, el cual se conservó en tubos para microcentrifugación Eppendorf®.
4.5.2. Amplificación de las regiones ITS del ADNr
Para la amplificación de las regiones ITS se utilizaron los cebadores universales ITS1/ITS4 e
ITS4/ITS5 ITS1: (5´-TCCGTAGGTGAACCTGCGG- ´3), ITS4:
(5´TCCTCCGCTTATTGATATGC-´3), ITS5: 5´GGAAGTAAAAGTCGTAACAAGG-´3)
(White et al., 1990). Las reacciones de PCR fueron calculadas para 25 μL; se utilizando las
siguientes condiciones de amplificación:

43
Las reacciones de PCR fueron calculadas para 30 μL; se utilizando las siguientes condiciones de
amplificación: 0.3 μL de cada primer a una concentración de 50 μm, 0.25μL de la Taq
polimerasa a 5U/ μL, 2.4 μL de los dinucleotidos (dATP, dCTP, dGTP y dTTP) a una
concentración de 2.5mM, 6 μL de buffer (5X) (10 mM Tris-HCl, pH 8.3; 50 mM KCl, 15 mM
MgCl2) y 1.2 μL del AND del hongo a una concentración de 5 ng/μL (Tabla ).
Tabla 2
Tabla 3. Condiciones de amplificación de ADN de Cladosporium spp., con los
cebadores ITS1 e ITS4.
Etapa Temperatura en grados centígrados
(ºC)
Tiempo en
segundos
1. Denaturación inicial 94 180
2. Denaturación por ciclo 94 60
3. Apareamiento 59 60
4. Extensión 72 120
5. Ciclos de amplificación 30 ciclos de 2 a 4
6. Extensión final 72 180
7. Finalización 4 Indefinido
4.5.3. Visualización y análisis de productos amplificados.
Para la visualización de los productos amplificados y su análisis, estos se separaron en gel de
agarosa (1.2% p/v, gel de 15 cm. de ancho por 10 cm. de largo), adicionado con GEL RED
10000X (Biotium); la electroforesis se realizó a 90 voltios/1h y fueron visualizados en analizador
de imágenes Eagle eye II de Strategene.
4.5.4. Limpieza de producto PCR para secuenciación
Se realizó la limpieza del producto de PCR de 15uL, adicionando el mismo volumen de PEG al
20% y 2.5M NaCl. Luego se mezcló por vortex y fue dejado a temperatura ambiente por 15

44
minutos. Se hizo una centrifugación a 13000 rpm por 15 minutos y se descartó el sobrenadante.
Posteriormente se agregó 100 uL de etanol al 70% y se centrifugó a 13000 rpm durante 2
minutos, del producto se descartó el sobrenadante con ayuda de una pipeta teniendo cuidado de
no retirar el pellet. Finalmente se resuspendió el pellet en 15 uL de agua y se revisó en un gel de
agarosa 1.2%.
5.4.5 Análisis de secuencia
Se hizo el análisis de secuencia de la región ITS, para la identificación de especies de
Cladosporium spp. Las amplificaciones obtenidas con los cebadores ITS 1, 4 y 5 fueron enviadas
a Iowa State University en el departamento de DNA sequencing facility en donde se llevó a cabo
la secuenciación automática de los fragmentos obtenidos.
Las secuencias fueron editadas y alineadas para obtener una secuencia consenso parcial de la región
ITS para cada una de las aislados. El análisis de la secuencia se realizó con los programas
ChromasPro© versión 1.49 beta y MEGA 4.0©. Posteriormente se realizó una búsqueda de
homología en el Banco de genes GenBank (www.ncbi.nlm.nih.gov) mediante la herramienta Blast.
4.5.6. Amplificación aleatoria de microsatélites RAMS.
Para determinar la variabilidad genética entre los aislamientos pertenecientes a la misma especie
de Cladosporium previamente determinada por la amplificación de la región ITS, se utilizó la
técnica de amplificación aleatoria de microsatélites, RAMS (Random Amplification of
Microsatellites, por su sigla en inglés) basada en la reacción en cadena de la polimerasa PCR
(Hantula et al., 1996).
En la reacción de PCR se utilizó: Buffer Taq (BIOLASE™ DNA Polymerase, BIOLINE)
10xNH4 (160mM (NH4)2SO4, 670 Tris-HCl pH 8.0, 0.1% Tween-20), a una concentración final
de 1X, 0.2 mM de cada uno de los dNTPs, 0.5mM de cada cebador, 1.5mM de MgCl2, 5ng/L
de ADN, agua HPLC esterilizada por filtración (MILLEX®-GV; Millipore Products Division,
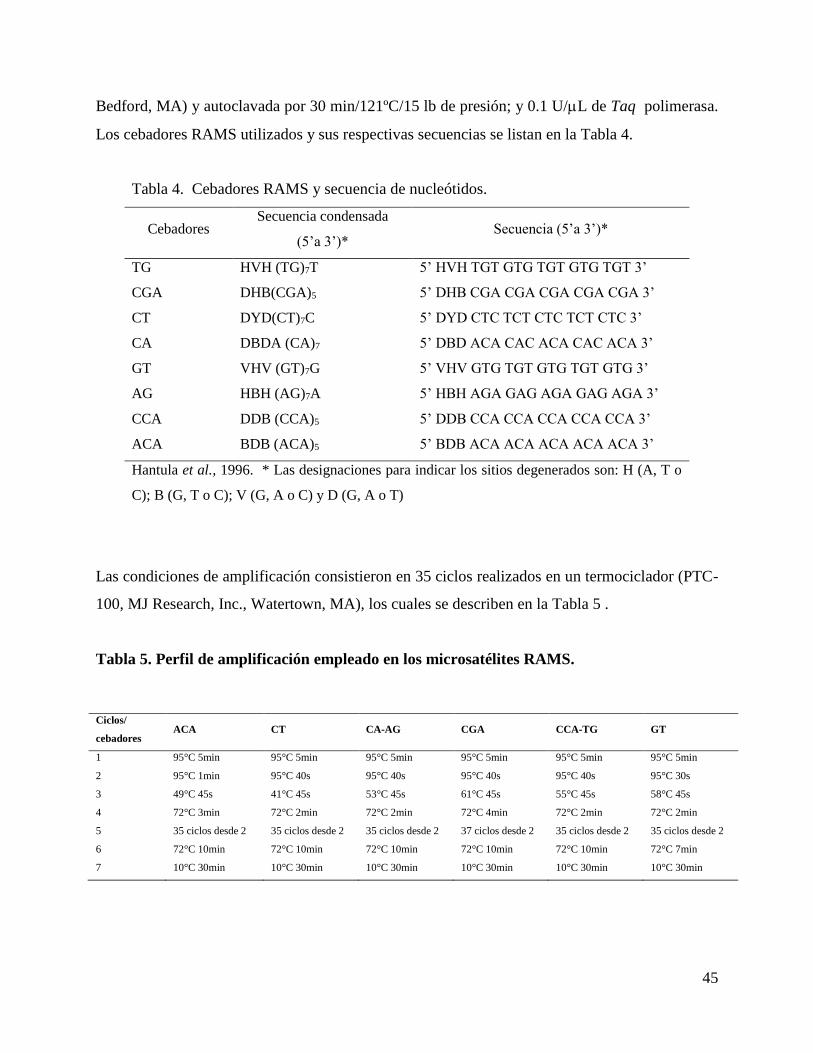
45
Bedford, MA) y autoclavada por 30 min/121ºC/15 lb de presión; y 0.1 U/L de Taq polimerasa.
Los cebadores RAMS utilizados y sus respectivas secuencias se listan en la Tabla 4.
Tabla 4. Cebadores RAMS y secuencia de nucleótidos.
Cebadores Secuencia condensada
(5’a 3’)* Secuencia (5’a 3’)*
TG HVH (TG)7T 5’ HVH TGT GTG TGT GTG TGT 3’
CGA DHB(CGA)5 5’ DHB CGA CGA CGA CGA CGA 3’
CT DYD(CT)7C 5’ DYD CTC TCT CTC TCT CTC 3’
CA DBDA (CA)7 5’ DBD ACA CAC ACA CAC ACA 3’
GT VHV (GT)7G 5’ VHV GTG TGT GTG TGT GTG 3’
AG HBH (AG)7A 5’ HBH AGA GAG AGA GAG AGA 3’
CCA DDB (CCA)5 5’ DDB CCA CCA CCA CCA CCA 3’
ACA BDB (ACA)5 5’ BDB ACA ACA ACA ACA ACA 3’
Hantula et al., 1996. * Las designaciones para indicar los sitios degenerados son: H (A, T o
C); B (G, T o C); V (G, A o C) y D (G, A o T)
Las condiciones de amplificación consistieron en 35 ciclos realizados en un termociclador (PTC-
100, MJ Research, Inc., Watertown, MA), los cuales se describen en la Tabla 5 .
Tabla 5. Perfil de amplificación empleado en los microsatélites RAMS.
Ciclos/
cebadores ACA CT CA-AG CGA CCA-TG GT
1 95°C 5min 95°C 5min 95°C 5min 95°C 5min 95°C 5min 95°C 5min
2 95°C 1min 95°C 40s 95°C 40s 95°C 40s 95°C 40s 95°C 30s
3 49°C 45s 41°C 45s 53°C 45s 61°C 45s 55°C 45s 58°C 45s
4 72°C 3min 72°C 2min 72°C 2min 72°C 4min 72°C 2min 72°C 2min
5 35 ciclos desde 2 35 ciclos desde 2 35 ciclos desde 2 37 ciclos desde 2 35 ciclos desde 2 35 ciclos desde 2
6 72°C 10min 72°C 10min 72°C 10min 72°C 10min 72°C 10min 72°C 7min
7 10°C 30min 10°C 30min 10°C 30min 10°C 30min 10°C 30min 10°C 30min

46
4.5.7. Electroforesis en agarosa
El producto amplificado de RAMS se separó por electroforesis en un gel de agarosa al 2%
teñido con GEL RED 10000x (Biotium) empleando buffer TBE 0.5X (Trisma base, ácido
bórico y ETDA), a 100 V. En cada gel se incluyó 5 μl de marcador de peso molecular de 1 Kb;
las bandas se visualizaron bajo luz ultra violeta y la imagen obtenida se capturó utilizando un
lector de geles Gel Doc 2000 (BioRad, USA Inc.).
4.6. Análisis estadístico
4.6.1. Patogenicidad
Para establecer los niveles de patogenicidad, se emplearon datos obtenidos a partir del área de la
lesión desarrollada sobre el fruto, y se analizaron por medio del programa Statistix versión 8.0
(Analytical Software, St. Paul, Minn) según un diseño completamente al azar para cada día de
evaluación y para el área bajo la curva del progreso de la enfermedad (ABCPE). Para todos los
días de evaluación se realizó un análisis de varianza (ANOVA) y una prueba de comparación
múltiple de Dunnett (POSTANOVA) con un nivel de significancia del 5% que permitió
determinar los aislamientos que presentaron diferencias significativas frente al control.
El ABCPE en porcentaje por día para cada repetición se calculó a partir de la siguiente ecuación
1:
1
1
2ABCPE
ii
n
i
ii ttxx
, Ecuación 1.
Donde:
xi = Severidad (porcentaje de área afectada) de la enfermedad a la i-ésima evaluación
ti = Tiempo (días) de la i-ésima evaluación
n = Número de evaluaciones.

47
Para comprobar que existen diferencias significativas entre las especies, y los niveles de
patogenicidad establecidos, se realizó una ANOVA con su respectiva prueba de separación de
medias LSD empleando los datos del ABCPE.
4.6.2. RAMS
Para el análisis de los productos, cada fragmento generado fue considerado como un carácter
independiente. Los fragmentos de ADN del mismo tamaño se asumieron como representantes
del mismo locus genético y se evaluaron como ausente o presente. Para cada banda individual
presente se asignó un valor de “1” y para ausencia un valor de “0”, generando así una matriz
binaria de datos para todos los aislamientos de Cladosporium spp.
Con la matriz generada, se obtuvo una matriz de similitud aplicando el coeficiente de similitud
de DICE (ecuación 2) a partir de la cual se construyó un dendrograma utilizando el método de
unión media aritmética no ponderada UPGMA (Unweighted pair group method with arithmetic
mean, por sus siglas en inglés), mediante el programa estadístico NTSYS-PC versión 2.02j
(Rohlf, 1998).
Ecuación 2: Sij = 2a , donde:
(2a + b + c)
Sij: Similitud entre el individuo i y el j.
a: Número de bandas presentes simultáneamente en los individuos i y j.
b: Número de bandas presentes en i, pero ausentes en j.
c: Número de bandas presentes en j, pero ausentes en i.
Con el fin de determinar la confiabilidad de cada nodo generado en el dendrograma, se utilizó el
método de repetición de muestras con reemplazamiento o bootstrap, mediante el programa
Winboot (Yap et al., 1996). Con el dendrograma obtenido se determinó el grado de similitud

48
entre y dentro de las especies de Cladosporium spp. Adicionalmente se establecieron las
frecuencias del hongo en cada departamento y los grados de patogenicidad de los aislamientos en
cada uno de los grupos observados en el dendrograma.
4.6.3. Análisis de agrupamiento
La matriz de datos fue utilizada para realizar un análisis de agrupamiento empleando el programa
estadístico NTSYS-PC versión 2.02 (Rohlf 1998), utilizando el coeficiente de Nei-Li (1978) o
DICE (1945, ecuación 2), para calcular el coeficiente de similitud también conocido como
similitud de DICE el cual omite la consideración de pares negativos (ausencia de bandas, 0-0) y
da doble peso a los pares positivos (presencia de bandas, 1-1), lo que lo hace muy útil en
términos de similaridad a nivel del DNA, en la que la ausencia compartida de una banda no es
necesariamente una indicación de similaridad entre los aislamientos.
)2(
2
cba
aSij
Ecuación 2.
Donde :
Sij = similaridad entre el individuo i y el j
a= Número de bandas presentes simultáneamente en los individuos i y j
b= Número de bandas presentes en i pero ausentes en j
c= Número de bandas presentes en j pero ausentes en i.
A partir de la matriz de similitud generada, se construyó un dendrograma usando el método de
unión media aritmética no ponderada UPGMA (del inglés unweighted pair group method with
arithmetic mean), posteriormente se empleó el programa Winboot (Yap et al. 1996) con el fin de
determinar la confiabilidad de cada nodo usando el método de repetición de muestras con
reemplazamiento ó bootstrap. Con el dendrograma obtenido se determinó el grado de similitud
en el que ambas especies se separan filogenéticamente. Igualmente, con el dendrograma obtenido
a partir de la matriz binaria de cada especie se determinó el coeficiente de similitud en el que se

49
establecieron las diferentes agrupaciones. Adicionalmente para cada grupo establecido se
determinaron las frecuencias de los municipios y niveles de patogenicidad que lo conforman.
4.6.4 Análisis de correspondencia múltiple
Se realizó un análisis de correspondencia múltiple mediante el programa estadístico SAS (SAS
Institute Inc. 2000) con el fin de facilitar la representación multidimensional de determinados
grupos hallados con características descriptivas de ellos en un plano. El análisis asocia todos los
aislamientos con las características con que fueron calificados, es decir, asocia todas las filas con
las columnas determinando el nivel de proximidad o asociación.
4.6.5 Diversidad genética
Empleando el paquete estadístico POPGEN versión 1.32 (Yeh et al 1999), se calculó la
heterocigocidad promedio (h) de acuerdo a la fórmula de Nei (1978) y el porcentaje de loci
polimórficos, para cada uno de los cebadores, las especies y la población general.
La heterocigosidad esperada para la población total y las dos especies se calculó mediante la
ecuación 3:
q
i
iXH1
21
Ecuación 3.
Donde:
Xi =Frecuencia en la población del alelo i
q = Número de alelos.
La diversidad total y la diversidad dentro de los municipios para la población total y las especies
evaluadas se calcularon con la ecuación 4.
STST HHH , Donde
)...(
)...(
21
2211
j
jj
STnnn
HnHnHnH
Ecuación 4.

50
Donde:
n = Número de individuos totales
HT = Total de la diversidad
HS = Diversidad dentro de cada municipio evaluado
HST = Media de la diversidad entre los municipios evaluados
El programa AMOVA-PREP versión 1.01 (Miller 1998) permitió generar matrices de distancia
con base en el coeficiente de DICE y crear archivos de distancia entre los grupos analizados para
automatizar los procesos de preparación de archivos que se usaron en el programa Análisis de
Varianza Molecular AMOVA versión 1.55 (Excoffier et al. 1992), el cual permitió realizar
comparaciones entre y dentro de los municipios evaluados, y calcular el coeficiente de
diferenciación genética FST para estimar la variabilidad observada entre los municipios
evaluados, estimándose con la ecuación 5
T
ST
STH
HF
Ecuación 5
Adicionalmente se hallo la diversidad de Nei (1978) para los niveles de patogenicidad
establecidos y se realizó un AMOVA con el fin de determinar diferencias significativas (P <
0.05) entre los mismos.
4.6.6. Distancia genética
La distancia genética e identidad genética entre municipios se calculó empleando el coeficiente
de distancia genética (Ecuación 6) y la identidad insesgada de Nei (1978) (Ecuación 7),
utilizando el paquete estadístico POPGEN, lo que generó un dendrograma (UPGMA) que
muestra las relaciones entre los municipios del estudio.
Ecuación 6. Identidad genética de Nei (1978) Donde:
JX y JY = y
nx y ny = Tamaños poblacionales en la población X e Y, respectivamente.

51
Ecuación7.
5.0. Evaluación in vitro de los extractos vegetales para el control de Cladosporium spp
Debido a las consecuencias que lleva consigo el uso indiscriminado de agroquímicos en los
diferentes cultivos, se propuso una investigación donde se evaluó la capacidad inhibitoria de
tratamientos alternos de control de la roña . Se evaluó la capacidad antagónica de los extractos
de plantas como Tagetes patula, Swinglea glutinosa (Ecoswing®), Furcraea macrophylla y
lixiviado de raquis de plátano (LRP). Adicionalmente, se incluyó Stratego como testigo de la
prueba, el cual es utilizado por los productores de gulupa y de demostrada actividad inhibitoria
del agente causante de la roña en otros cultivos como maracuyá y granadilla.
La efectividad en el control se evaluó mediante la medición diaria del crecimiento circular del
hongo, hasta que el testigo absoluto cubrió totalmente la superficie del medio. Se comparó cada
tratamiento con el testigo absoluto, y a los diez días, cuando el hongo logró su esporulación total.
Adicionalmente se evaluó un biofungicida (Fungibiol) comercial utilizado comúnmente por los
productores para el manejo de varias enfermedades en arroz y pastos, con el objetivo de
determinar la sensibilidad o resistencia de aislamientos de Cladosporium spp altamente
patogénicos y provenientes de cultivos comerciales de gulupa y efectuar la comparación entre el
manejo convencional (manejo químico) y nuevas alternativas para el manejo de la enfermedad.
5.1. Selección de aislamientos de Cladosporium spp.
De acuerdo a los resultados de los grupos RAMS y de patogenicidad de las cepas sobre frutos de
gulupa, se seleccionaron los aislamientos, más patogénicos, efectuando con ellos las pruebas de
inhibición in vitro, enfrentando cada aislamiento con los productos antes descritos.

52
5.2 Pruebas antagónicas
Para la prueba de antagonismo, se evaluó la efectividad de Trichoderma spp. Para esto se
seleccionó dos cepas, T. harzianum de nombre comercial Agroguard y Trichoderma viride
(suministrado por Live Systems Technology – LST.
Las evaluaciones de antagonismo, se realizaron en medio PDA con ácido láctico (39 g PDA y 10
ml de ácido láctico 25% por litro de medio). Para tal efecto, se sembró un disco de medio de
cultivo fresco con micelio del patógeno, obtenido con sacabocado # 2 (5 mm de diámetro), el
cual se colocó en un extremo de una caja petri de 9 cm de diámetro y un disco de medio de
cultivo con crecimiento del antagonista en el otro extremo. Cada disco fue colocado a 2.25 cm
del borde de la caja, para asegurar el mismo espacio de crecimiento para cada uno de los hongos.
Los tratamientos testigos consistieron de un disco con micelio del patógeno creciendo sobre
medio PDA.
Las evaluaciones se realizaron día de por medio, iniciando dos días después de sembrado,
durante 10 días, registrando el radio de crecimiento de los patógenos hacia Trichoderma spp.,
para un total de cinco evaluaciones. Con los datos obtenidos al realizar la última evaluación, se
determinó el porcentaje de inhibición de crecimiento radial (ICR), mediante la fórmula de
Dennis y Webster (1971):
ICR (%) = (R1-R2) 100/R1
Donde R1 es la distancia más lejana recorrida por el patógeno y R2 es la distancia recorrida por
el patógeno hacia el antagonista.
5.2 Preparación de inóculo de Cladosporium spp.
Los seis aislamientos seleccionados y conservados en papel filtro se sembraron por triplicado en
cajas Petri con agar PDA y se incubaron invertidas a 28oC/15 días luz continua.

53
5.3. Preparación de extractos vegetales
Para evaluar los extractos de Swinglea glutinosa (Ecoswing®) al 2%, F. macrophylla al 10%,
Tagetes patula al 5% y el Lixiviado de Raquis de Plátano LRP al 10%, se prepararon medios de
cultivo PDA modificados con cada uno de los extractos.
5.3.1. Extracto de Furcraea macrophylla (Fique)
El extracto se obtuvo de hojas de plantas adultas que fueron cortadas desde la base y lavadas con
agua destilada estéril con la finalidad de reducir microorganismos contaminantes. El proceso de
extracción de jugo se realizó por corte y licuado; este licuado se filtró a través de 6 capas de
gasa, posteriormente a través de filtros de celulosa Whatman de 8.0 y 2.5 µm de tamaño de
poro y finalmente esterilizado mediante un filtro de jeringa de 25 mm con membrana de nilón
FisherBrand de 0.25 µm de tamaño de poro. El extracto estéril obtenido se valoró como el
100% y a partir de él se elaboraron los medios con las concentraciones a evaluar. Este extracto
concentrado fue utilizado para elaborar medios modificados y solidificados con PDA en
concentraciones de 0.5, 1, 5 y 10% (Gómez, 2002). Los medios así preparados fueron
almacenados en oscuridad por 24 h, para verificar la esterilidad de los mismos antes de su uso.
5.3.2. Extracto de Swinglea glutinosa (Ecoswing®)
Se evaluó el producto Ecoswing® el cual se recomienda en dosis de 2 cc/L, para aplicaciones en
campo. Para hacer la evaluación In Vitro se preparó 600 mL de medio, de los cuales 12 mL
correspondían al extracto puro que fue previamente esterilizado por filtrado, a través de un filtro de
jeringa de 25 mm con membrana nilón FisherBrand® de 0.2 μm.
5.3.3 Extracto de Tagetes patula
Se empleó el extracto de T. patula producido por SAFER, el cual viene preparado en una
concentración al 10%. Para la evaluación se prepararon 600 mL de medio del cual 300 mL
correspondieron al extracto de T. patula para completar una concentración de 5%. Este extracto fue

54
igualmente esterilizado por filtración a través de un filtro de jeringa de 25 mm con membrana nilón
FisherBrand® de 0.2 μm.
5.3.4. Lixiviado de Compost de Raquis de Plátano (LRP)
El lixiviado de Raquis de Plátano fue obtenido de la Finca la Guaira, Montenegro-Quindío y fue
valorado como el 100%. Para la evaluación se preparó 600mL de medio de los cuales 60 mL
correspondieron al lixiviado puro, previamente esterilizado en autoclave (121°C / 15 lb de presión /
15 min) para completar una concentración del 10% (Rojas 2008)
5.3.5. Stratego® (Ingrediente activo: Trifloxystrobin, Propiconazol)
Stratego 450 g/ litro de formulación. Categoría toxicológica III.
Como fungicida químico se utilizó stratego (Trifloxystrobin, Propiconazol ®), el cual fue evaluado
en dosis comercial de 1.0 mL/L de agua (450 g/L de ingrediente activo) y esterilizado a través de un
filtro de jeringa de 25 mm con membrana de nilón FisherBrand ® de 0.25 μm de tamaño de poro.
Este tratamiento constituyó el control (-), para hacer las comparaciones con los diferentes extractos
vegetales.
Adicionalmente al tratamiento con el fungicidad se agregó 100 μg.mL de SHAM, ya que la
oxidasa alternativa, que es funcional solo en mitocondrias aisladas de mutantes resistentes a
fungicidas y que es inhibida por el ácido salicilhidroxámico (Ziogas et al., 1997), distorsiona los
resultados de los tests de monitoreo de sensibilidad. Por cuanto la resistencia está controlada por
genes mitocondriales, su herencia se asume que es no mendeliana (Gisi et al., 2002).
5.3.6 Fungibiol
Su ingrediente activo lo constituyen extractos vegetales con propiedades fungicidas,
saponificados y homogenizados en una base oleosa orgánica, que mezcla fácilmente con el agua.

55
Actúa como endoelicitor y exoelicitor por los metabolitos presentes en los extractos. Alterando
el hábitat de algunos hongos. El cual comercialmente viene en formulación de concentrado
emulsionable al 45%, y el cual se evaluó en dosis comercial de 1.0 g/L de agua, y con la cual se
preparó el medio modificado con PDA, previa esterilización a través un filtro de jeringa de 25
mm con membrana de nilón FisherBrand de 0.25 µm de tamaño de poro.
5.3.7. Prueba de sensibilidad in vitro para biofungicidas.
Utilizando los medios modificados con cada uno de los biofungicidas, se procedió a colocar un
disco de 6.5 mm. de diámetro con crecimiento del hongo (seis aislamientos evaluados) en el
centro de la caja. El diseño constó de tres repeticiones por tratamiento, las cuales se dispusieron
de manera aleatoria en tres bloques.
En la prueba se incluyeron testigos conformados por PDA, el cual fue sembrado con discos de
6.5 mm. de diámetro con micelio del hongo de cada uno de los seis aislamientos y en un diseño
de bloques al azar con tres repeticiones. Las cajas Petri con los medios modificados sembradas
con los seis aislamientos, se sellaron e incubaron invertidas a 24ºC /10 días.
5.3.8 Evaluación del crecimiento del hongo y análisis de la información.
Durante los días 2, 4, 6, 8 y 10, se registró el valor en milímetros de los cuatro radios de
crecimiento de las colonias en cada uno de los medios, incluido los testigos; seleccionando el día
10 como el último día de evaluación, dado que a este tiempo en uno de los tratamientos la
colonia sembrada cubrió el área total de la caja Petri.
Estos datos fueron empleados para calcular el crecimiento por día, expresado como área y
velocidad (pendiente), durante el tiempo de evaluación (10 días), y con los cuales se realizó una

56
prueba de contraste múltiple de rangos de Duncan según los tratamientos con un nivel de
significancia del 5%, para determinar las medias que fueron significativamente diferentes.
6.0. Evaluación in vivo de la capacidad de control de los extractos vegetales en frutos
desprendidos de gulupa.
Para la prueba de inhibición en frutos de gulupa se utilizó un diseño completamente al azar, cada
unidad experimental consistió en una cámara humedad con seis frutos, con esta prueba de
inhibición del desarrollo de la roña en frutos desprendidos, se pretendió evaluar la protección que
puede ofrecer los extractos vegetales evaluados contra el desarrollo de los síntomas ocasionados
por Cladosporium cladosporioides, aislado de frutos de gulupa enfermos.
Para esta prueba se utilizaron los extractos vegetales Ecosiwing, extracto de fique 10% y
Fungibiol, seleccionados en las pruebas de inhibición del crecimiento micelial, debido a que
estos fueron los extractos vegetales que se presentaron mejores resultados en las pruebas in
Vitro. Adicionalmente se montó un control negativo que consistió en frutos sanos sumergidos en
agua destilada esteril.
Los frutos fueron lavados con agua corriente y detergente, después fueron sumergidos en
hipoclorito al 2,5 % durante 2 minutos, luego en alcohol al 70% durante 5 minutos y finalmente
fueron lavados con abundante agua. Una vez realizado este procedimiento se dejaron secar a
temperatura ambiente.
Una vez secos, los frutos de cada tratamiento seleccionado se sumergieron en una solución de
cada uno de los extractos vegetales por 10 min y se dejaron nuevamente al ambiente. Los
tratamientos se aplicaron en tres ocasiones: una semana antes, el mismo día y una semana
después de la inoculación del patógeno.
Los aislamientos evaluados fueron reactivados en PDA por 10 días a 27 ˚C en oscuridad,
posteriormente a cada fruto se les realizo pequeñas punciones y luego se asperjaron con una
solución acuosa de esporas a una concentración de 1x106 esporas / mL de agua. Para este método

57
se utilizó un aspersor de agua de plástico (123RF 400 x 267 aprox). Las condiciones óptimas
para el desarrollo de la enfermedad se proporcionaron colocando los frutos en cámaras húmedas
(HR > 90%) a temperatura de 22 ˚C.
Tres días después de la inoculación los frutos se observaron y en cada una de las lesiones
desarrolladas, por su forma circular, se tomaron cuatro radios con los que posteriormente se
calculó el área del círculo (Lesión) y el porcentaje de fruto afectado (severidad). Se realizaron
lecturas cada dos días hasta que alguna lesión cubriera completamente el fruto o hasta que más
de dos frutos por aislamiento se descompusieran. Todos los frutos fueron mantenidos a 22°C en
condiciones de laboratorio.

58
6. RESULTADOS Y DISCUSIÓN
6.1. Pruebas de patogenicidad (Postulados de Koch) y Estandarización de inoculación
Desde ya hace varios años, se ha venido presentando una enfermedad en los cultivos de gulupa,
denominada roña. Se han realizado gran número de investigaciones buscando el agente causal de
este disturbio. Las muestras procesadas para obtener microorganismos fueron tomadas a partir de
frutos de gulupa de diferentes estados de madurez.
Con base en estos antecedentes, en este ensayo se realizaron pruebas de patogenicidad en vitro
con microorganismos posiblemente patógenos asociados con gulupa afectados por roña. Se
realizaron aislamientos de frutos de gulupa que presentaban síntomas de la enfermedad. A partir
de estos tejidos se aislaron tres (3) hongos pertenecientes a los géneros, Cladosporium,
Colletotrichum y Fusarium usando medios de cultivos semiselectivos.
La patogenicidad de los microorganismos seleccionados fue determinada mediante la
inoculación de tejidos de frutos de gulupa bajo condiciones in vitro. Paralelamente se evaluó el
mejor método de inoculación en los diferentes estados de madurez del fruto.
El mejor método fue la inoculación con punciones y la aspersión de la solución acuosa de
esporas a una concentración de 1x106 esporas / mL de agua. De todos los frutos inoculados con
los diferentes hongos, solamente se pudo observar síntomas característicos y típicos de la
enfermedad con los aislamientos de Cladosporium spp (Figura 2).
De los frutos inoculados, a las 48 horas después no presentaron síntomas de la enfermedad; a las
72 horas se empezaron a mostrar pequeñas manchas con motas algodonosas sobre las heridas
que se le ocasionaron al fruto, esto concuerda con lo descrito por Delgado et al. (2012),
refiriéndose que la roña está asociada con las heridas causadas por insectos.

59
Los frutos con los síntomas más característicos de la enfermedad fueron del grupo 5. Estos frutos
presentaron síntomas de la roña, a excepción del testigo que permaneció completamente sano, a
los 10 días después de la inoculación se observó que en los frutos de los grupos (4 y 5) la
enfermedad ya había alcanzado un grado de severidad a comparación de los grupos 1, 2 y 3, los
cuales en el momento de la medición no tenían síntomas de la enfermedad, los frutos del grupo 5
se asemejaban más a los síntomas observados en campo, tales como lesiones de color marrón
oscuro produciendo un chancro con esporulación del hongo color oliváceo a veces marrón. Estos
resultados coinciden con lo descrito por Mora (2011), quien determino que después de la
inoculación en frutos de gulupa con Cladosporium ssp, primero se forma un chancro y
posteriormente la roña (Figura 3).
Santos (2008) explica que los frutos, en sus primeras etapas de desarrollo, aumentan
progresivamente su tasa respiratoria, ya que esta, está ampliamente relacionada con la actividad
bioquímica del mismo para lograr un crecimiento normal hasta llegar a su etapa de maduración,
en este momento los frutos se encuentran susceptibles a enfermedades por el esfuerzo que
deben realizar hasta llegar a la madurez fisiológica.
Rondón (2007) expresa que en la dinámica de la infección por tamaño del fruto de tomate, se
observa una mayor susceptibilidad de los frutos verde mediano, verde pequeño y verde grande,
debido muy posiblemente a sus características bioquímicas y quizás a la mayor duración de
estas etapas en el campo, como blanco de la infección.
Los frutos que fueron inoculados con Fusarium spp y Colletotrichum spp, presentaron una
pudrición húmeda color marrón observándose síntomas no típicos de la enfermedad en estudio.
El testigo al cual sólo se le ocasionaron heridas, se mantuvo sano durante todo el periodo de
evaluación, lo que quiere decir, que las lesiones presentes en los frutos, si fueron ocasionadas
por los géneros de hongos inoculados, descartando la posibilidad de que ya estos frutos
estuviesen inoculados por el patógeno.

60
Estos resultados coinciden con lo descrito por Delgado et al. (2012), quienes determinaron que a
los 6 dias el hongo Cladosporium spp., había completado el ciclo de la enfermedad
momificando completamente los frutos inmaduros de maracuyá (Passiflora edulis.), por tanto
que los resultados obtenidos en esta investigación permiten relacionar la roña con el daño
ocasionado por el hongo Cladosporium spp.
Figura 2. Estandarización de la metodología de inoculación en frutos de gulupa. a.
Aplicación de alícuotas de 1µ, 5µ y 10µ de suspensión de esporas 1x 10 6. b. Inoculación de
bloques de micelio del hongo en heridas realizadas en el fruto. c. Aspersión con una solución
acuosa de esporas a una concentración de 1x106 .d. Síntoma característico de la roña en gulupa
por el método de aspersión
a b
c d

61
6.2 .Caracterización patogénica
El análisis de la patogenicidad expresada por cada uno de los aislamientos evaluados, llevó a la
separación de tres grupos, identificados como: aislamientos de patogenicidad alta y, media, y un
grupo que se caracterizó por presentar patogenicidad muy alta.
Los frutos de gulupa presentaron diferencias en el progreso de la enfermedad con respecto a los
aislamientos, oscilando estas variaciones desde el 1 hasta el 90% de severidad (Tabla 6)
Los síntomas observados en los frutos de gulupa inoculados con Cladosporium spp. bajo
condiciones de laboratorio consistieron de lesiones generalmente ovaladas con bordes irregulares
de color oliváceo , que avanzaron en forma diferente entre los aislamientos lo que permitió
relacionar el área de lesión con el nivel de patogenicidad. (Los controles frutos inoculados
asperjados con agua destilada esteril) no presentaron ningún tipo de síntoma a través de los 21
días de duración del experimento.

62
Tabla 6. Niveles de patogenicidad determinada como ABCPE para los 15 aislamientos de
Cladosporium spp en Passiflora edulis fv edulis.
Tabla 3
NIVELES DE
PATOGENICIDAD AISLAMIENTO
ABCPE
MEDIA
GRUPOS ESTABLECIDOS POR LA
SEPARACIÓN DE MEDIA (LSD)
PMA HUPEC-GU-1 584.03 A
PMA HUA-GU-1 579.34 AB
PMA HUGA-GU-1 497.54 AB
PMA HUALG-GU-1 476.56 BC
PMA HUB-GU-1 470.65 BCD
PA VAYJ-GU-2 313.45 CDEF
PA BOJ-GU-1 250.19 CDEFG
PA BOB-GU-1 231.65 CDEFGH
PA BOB-GU-2 229.55 CDEFGHI
PA BOBER-GU-1 228.48 CDEFGHIJ
PA CUCSL-GU-1 222.89 CDEFGHIJ
PM TOILM-GU-1 191.89 DEFGHIJK
PM CALT-GU-1 163.84 DEFGHIJKL
PM ANTR-GU-1 141.45 DEFGHIJKLM
PM ANTR-GU-1 133.73 DEFGHIJKLMN

63
Figura 3. Frutos de gulupa 26 días después de inoculados con aislamientos de Cladosporium
spp.
HUPEC- GU-1 HUA_GU_1 HUGA_GU_1
HUALG_GU
HUB_GU-1 VAYJ-GU-
VAYJ-GU-2 BOJ-GU-1 BOB-GU-1

64
El análisis de varianza (Tabla 7) mostró diferencias significativas (P<0.001) entre los
aislamientos para el área de la lesión (cm2), durante todos los días de evaluación y para el área
bajo la curva del progreso de la enfermedad (ABCPE) (Tabla 7).
Tabla 7. Análisis de Varianza del área de la lesión (cm2) durante los 21 días de evaluación y el
ABCPE de los aislamientos de Cladosporium spp. aislados de gulupa.
Evaluación Fuente de
Variación
Valor
Fischer(F) P valorb
Nivel de
significancia
Día 5 Entre aislamientos 2.31 <0.001 ASc
Día 9 Entre aislamientos 3.20 <0.001 AS
Día 13 Entre aislamientos 4.97 <0.001 AS
Día 17 Entre aislamientos 4.65 <0.001 AS
Día 21 Entre aislamientos 4.84 <0.001 AS
Área bajo la curva Entre aislamientos 4.62 <0.001 AS
P valorb, probabilidad de encontrar valores mayores de F
ASc, altamente significativo
El análisis de varianza para los tres niveles de patogenicidad establecidos arrojó diferencias
significativas (F=59.5) a un nivel de confianza de 0.01%, y la separación de medias (LSD)
De los 15 aislamientos evaluados el 34.6% resultaron con un nivel de patogenicidad medio, el
43.4 % presentaron un nivel alto y el 22% presentaron un nivel de patogenicidad muy alto
(Figura 4).

65
Figura 4. Niveles de patogenicidad determinados para los aislamientos de Cladosporium spp. de
gulupa. A. nivel muy alto, B. nivel alto, C. nivel medio.
En la Figura 5 se muestra que los aislamientos de los municipios de Huila, Boyacá y Antioquia
son los más diversos en cuanto a niveles de patogenicidad, presentando niveles muy altos, altos e
intermedios. Valle, Caldas y Cundinamarca, concentraron la mayoría de los aislamientos con un
nivel intermedio de patogenicidad, y Tolima fue el único municipio con aislamientos sin nivel
intermedio.
0
5
10
15
20
25
30
35
40
45
0 5 9 13 17 21 Días
Po
rcen
taje
de
áre
a af
ecta
da
HUPEC-GU-1!
HUA-GU-1
HUGA-GU-1
HUALG-GU-1
VAYJ-GU-2
BOJ-GU-1
BOB-GU-1
BOB-GU-2
BOBER-GU-1
CUCSL-GU-1

66
Figura 5. Nivel de patogenicidad por municipio de 15 aislamientos de Cladosporium spp.
inoculados en frutos degulupa.
6.3. Secuenciación de las regiones ITS
Se seleccionaron 10 productos de amplificación de ITS y se secuenciaron con el fin de buscar
identidad con genes de patógenos de plantas y que han sido reportados previamente en la base de
datos del Gen- Bank (NCBI) (Tabla 8).
Se utilizaron aislamientos que tuvieron mayor diferencia macroscópica y que provenían de
regiones y hospedantes diferentes, ya que molecularmente no se había evidenciado una
diferencia específica para todos los aislamientos. Los aislamientos seleccionados se secuenciaron
con los cebadores ITS 1-4; e ITS 1-5.
Al utilizar el programa MEGA 5.0©, se confirmó la alta homología de las muestras secuenciadas
con respecto a Cladosporium cladosporioides. Se mencionan algunas secuencias obtenidas del
GenBank, reportadas por diferentes autores, para la relación de las aislados de Cladosporium
spp. Obtenidas por PCR (Tabla 9)
0
10
20
30
40
50
60
70
80
90
P
o
r
c
e
n
t
a
j
e
d
e
a
i
s
l
a
m
i
e
n
t
o
s
Departamentos
Muy alto
Alto
Intermedio
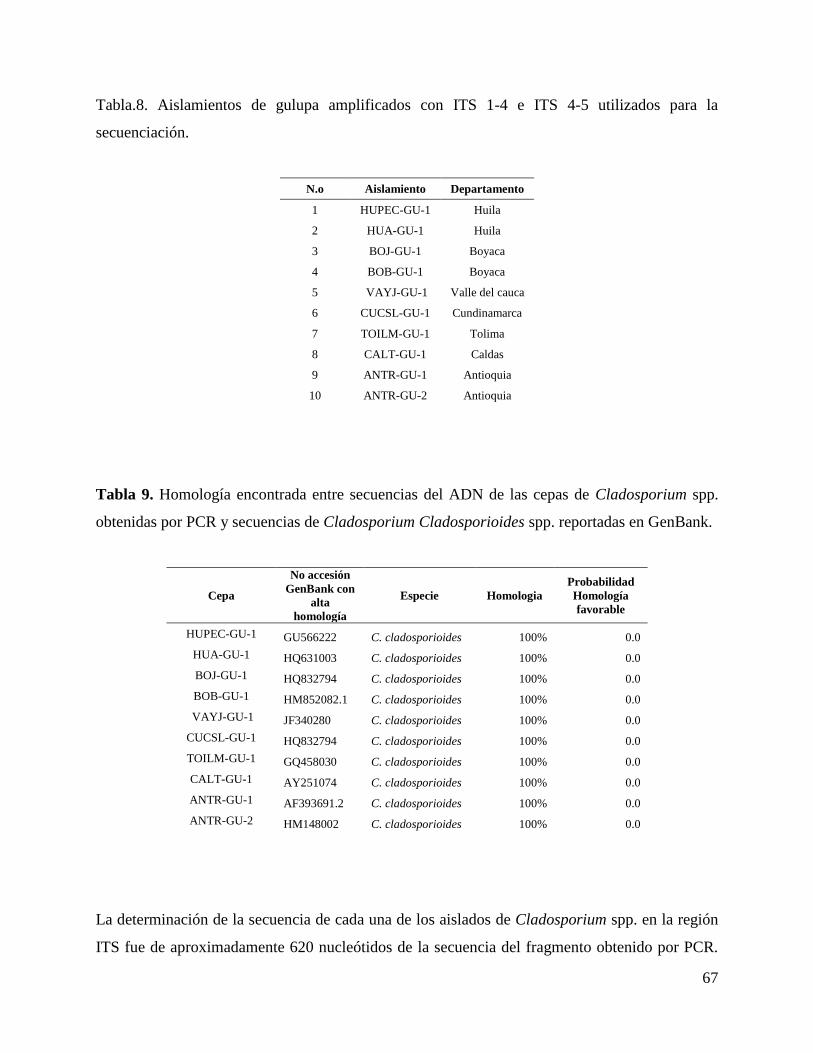
67
Tabla.8. Aislamientos de gulupa amplificados con ITS 1-4 e ITS 4-5 utilizados para la
secuenciación.
N.o Aislamiento Departamento
1 HUPEC-GU-1 Huila
2 HUA-GU-1 Huila
3 BOJ-GU-1 Boyaca
4 BOB-GU-1 Boyaca
5 VAYJ-GU-1 Valle del cauca
6 CUCSL-GU-1 Cundinamarca
7 TOILM-GU-1 Tolima
8 CALT-GU-1 Caldas
9 ANTR-GU-1 Antioquia
10 ANTR-GU-2 Antioquia
Tabla 9. Homología encontrada entre secuencias del ADN de las cepas de Cladosporium spp.
obtenidas por PCR y secuencias de Cladosporium Cladosporioides spp. reportadas en GenBank.
Cepa
No accesión
GenBank con
alta
homología
Especie Homologia
Probabilidad
Homología
favorable
HUPEC-GU-1 GU566222 C. cladosporioides 100% 0.0
HUA-GU-1 HQ631003 C. cladosporioides 100% 0.0
BOJ-GU-1 HQ832794 C. cladosporioides 100% 0.0
BOB-GU-1 HM852082.1 C. cladosporioides 100% 0.0
VAYJ-GU-1 JF340280 C. cladosporioides 100% 0.0
CUCSL-GU-1 HQ832794 C. cladosporioides 100% 0.0
TOILM-GU-1 GQ458030 C. cladosporioides 100% 0.0
CALT-GU-1 AY251074 C. cladosporioides 100% 0.0
ANTR-GU-1 AF393691.2 C. cladosporioides 100% 0.0
ANTR-GU-2 HM148002 C. cladosporioides 100% 0.0
La determinación de la secuencia de cada una de los aislados de Cladosporium spp. en la región
ITS fue de aproximadamente 620 nucleótidos de la secuencia del fragmento obtenido por PCR.

68
Mediante el análisis y comparación de las secuencias realizados con la herramienta Blast en
GenBank, se encontró que los aislados de Cladosporium spp analizadas, presentaron altos
niveles de homología con secuencias de la especie Cladosporium cladosporioides al 100%
reportadas en el Genbank.
Los aislamientos y las secuencias obtenidas del Gen-Bank de diferentes especies de
Cladosporium también se compararon con una secuencia de la especie Mycosphaerella
graminícola y Cercospora beticola con el fin de hacer evidente la similitud o la diferencia con
especies de Cladosporium, de esta manera se formó un clado completamente aparte y sin
ninguna relación con especies de Cladosporium (Figura 6).

69
Figura 6. Cladograma del análisis de las secuencias de nucleótidos de la region ITS del ADNr
para 10 aislamientos de Cladosporium spp. aislados de frutos de gulupa Passiflora edulis fv
edulis

70
Resultados similares registra Delgado (2012), donde el análisis de secuencias utilizando ITS en
muestra de maracuyá, gulupa y granadilla afectadas por la roña, permitió identificar altos niveles
de homología con Cladosporium cladosporioides. Por otro lado Riascos et al (2011), registraron
datos de análisis morfológicos por microscopía de barrido electrónico y molecular por ITS
mostrando correspondencia, al determinar por ambas vías a C. cladosporioides sensu lato como
el complejo que incluye la especie a la cual pertenecen los aislamientos de gulupa con síntomas
de roña que analizaron.
6.4. Caracterización molecular de Cladosporium cladosporioides
Los marcadores moleculares pueden usarse con varios propósitos como estimar la variabilidad
genética entre y dentro de poblaciones, muestras o accesiones de especies cultivadas o silvestres
relacionadas; para detectar la duplicación de los materiales en las colecciones; para evaluar la
delimitación de los centros de origen; para el estudio de las relaciones filogenéticas entre
especies y taxones superiores y para la determinación de mejores estrategias de conservación de
recursos genéticos y la formación de colecciones nucleares.
La técnica molecular utilizada en este estudio mostró un alto grado de polimorfismo y
sensibilidad para la discriminación de los aislamientos de la colección, lo que sugiere que los
marcadores moleculares RAMs son útiles para la evaluación de la diversidad genética de
colecciones, bancos de germoplasma y poblaciones naturales, además, la técnica ha sido
reportada para la caracterización molecular de especies vegetales como soya (Glycine max),
(Akkaya et al. 1992) y frutales como uchuva (Physalis peruviana) (Bonilla et al. 2008), mora
(Rubus sp) (Morillo et al. 2006), guayaba (Psidium sp) (Sanabria et al. 2006), y Heliconias (H.
latispatha) ( Muñoz et all 2009)
Sus ventajas incluyen bajo costo, reproducibilidad, utilización de un solo cebador, no requiere
información previa, alto polimorfismo y fácil implementación (Muñoz et al. 2009)

71
El análisis mediante el coeficiente de Nei-Li, a un nivel de Similaridad para los 40 aislamientos
fue de 0.75, diferenciado la población en 6 grupos (A,B,C,D,E,F), también se visualiza una
separación marcada entre las especies de gulupa, maracuyá y granadilla. Esto es entendido
debido a que cada una de las estas especies posee una composición genética diferente, lo que le
otorga una identidad propia (Figura 7).
El análisis de diversidad de la población muestra que existe una variabilidad alta del patógeno,
representada en cultivos de pasifloras que son asociadas a la enfermedad, cabe destacar que la
muestra GRLUK-MA-1, la cual se separó del resto de genotipos a un nivel de similaridad de
0.50, tal vez tenía patrones de bandas únicos, que la diferencian del resto de los grupos.
Resultados similares reporta Henríquez (2003) en el hallazgo de uso de marcadores moleculares
para clarificar la relación entre aislamientos Andinos, Mesoamericanos y aislamientos Afro-
Andinos de P. griseola. Encontraron que para 131 aislamientos de P griseola recolectados de
África y Latinoamérica. En contraste con los marcadores de virulencia, los marcadores
moleculares mostraron un alto nivel de diferenciación geográfica dentro de cada grupo de P.
griseola definido. Los aislamientos andinos de África fueron claramente separados de los
aislamientos andinos de Latinoamérica, así como los aislamientos Mesoamericanos.
Estos resultados son opuestos obtenidos por Mahuku et al (2002), donde registra que la
diversidad genética de P. griseola, es la especialización del hospedero, con la aparente
diferenciación“ geográfica jugando un rol secundario. Los resultados presentados en este estudio,
mostraron que los aislamientos Afro- Andinos no son un grupo separado sino aislamientos
típicos andinos que han co-evolucionado para colonizar el fríjol desde el acervo genético
Mesoamericano.
Como puede observarse el grupo A contiene algunos aislamientos de gulupa, mientras que el
grupo B la mayoría de los aislamientos son de gulupa y maracuyá.

72
Figura 7. Dendrograma de la estructura genética de los 40 aislamientos de C.cladosporioides .
basados en el Coeficiente de Similitud de Nei-Li y calculado de los datos combinados de los
cinco cebadores Microsatélites RAMs, con el método de clasificación UPGMA.
En el caso de Cladosporium fulvum, Peteira et all (2011) registra que la caracterización
molecular de 37 aislados de C. fulvum provenientes de plantas de tomate de diferentes
localidades. La caracterización incluyó la identificación de los aislamientos por la amplificación
de la región 28S del gen que codifica para el ARNr, la PCR con cebadores específicos dirigidos a
diferentes genes relacionados con la patogenicidad (Avr2, Avr4, Avr4E, Avr9, Ecp1, Ecp2,
Ecp4 y Ecp5) y los RAPDs. Todos los aislados fueron clasificados como C. fulvum.
Es importante anotar que este trabajo se orientó a caracterizar los aislamientos existentes en el
cepario de pasifloras de la Universidad de Caldas, por lo anterior, existieron caracterizaciones
con dos aislamientos como es el caso de granadilla y otras con más de 15 aislamientos, esto
implica analizar los resultados con cautela, debido al sesgo introducido por el muestreo.

73
Arenas (2007), registra la diferenciación molecular de la especie Colletotrichum sp. de la especie
C. gloeosporioides, mediante el empleo de electroforesis SSCP, de la región 5.8S interna
transcrita del ADN ribosomal empleando la metodología desarrollada por Kong et al. (2004),
concluyendo que los patrones de esta especie no identificada son parcialmente compartidos con
la especie C. gloeosporioides, pero no idénticos a ésta.
Estos resultados concuerdan con estudios realizados por Rojas et al. (2008) y Sanabria (2008),
donde observaron que a un coeficiente de similitud de 0.26 se produce el agrupamiento del
género Colletotrichum, procedente de guanábano. La separación de las especies identificadas se
produce a un coeficiente de similitud de 0.28, formándose seis subgrupos RAMS, así: dos
subgrupos de la especie Colletotrichum sp. (subgrupos A y B), tres subgrupos de la especie C.
gloeosporioides (subgrupos C, D y E) y un subgrupo de la especie C. acutatum (subgrupo F).
Para la evaluación de la diversidad genética de C. cladosporioides, se seleccionaron cinco de los
siete cebadores utilizados; el primer GT no amplificó y fue descartado.
Los valores de heterocigosidad encontrados mediante el programa TFPGA® muestran que los
promedios mas altos corresponden a los cebadores CT (0.48), CGA (0.48) y TG (0.48). El
cebador CT fue el que mayor aporte hizo a la variación encontrada con un valor de Fst de
0.3298, lo que significa que puede ser útil para lograr una mayor diferenciación entre los
especies de Cladosporium cladosporioides (Tabla 10 )
Posiblemente el polimorfismo encontrado con este cebador se deba a una mayor frecuencia entre
los sitios de hibridación y las cadenas de ADN.

74
Tabla.8. Valores de Heterocigosidad, Fst, y porcentaje de loci polimorficos para los
primers RAMs.
Cebador No. Loci
polimórficos
He
estimada
% Loci
polimórficos
(95%)
Fst SD
CA 16 0,46 100 0,1161 0,06
CCA 18 0,45 100 0,1086 0,05
CT 12 0,48 100 0,3298 0,03
CGA 12 0,48 100 0,1113 0,05
TG 18 0,48 100 0,1621 0,04
Población total 76 0,47 100 0.1644 0.04
La heterocigosidad promedio (He) para la población total fue de 0.47, lo que revela un gran
polimorfismo genético en los grupos analizados, lo cual puede estar asociado a la probable
naturaleza alogama de la especie, la cual tiende a favorecer la conservación del alto porcentaje de
heterocigotos.
El Fst cuantifica la consanguinidad de las subpoblaciones en relación a la población total de la
cual forman parte, se interpreta como la proporción de la variación total atribuible a la
diferenciación entre poblaciones (Caujapé, 2006). El valor de Fst obtenido mediante el programa
TFPGA® para los 40 aislamientos estudiados asumiendo equilibrio Hardy-Weinberg fue de
0.1644, con una desviación estándar de 0.05. Según Wright (1978), valores por encima de 0.15
muestran una alta diferenciación genética (Tabla 11), por lo que el resultado obtenido evidencia
una alta variación entre las especie C. cladosporioides analizada corroborando los resultados
hasta el momento obtenidos. El Fst encontrado en este estudio es un parámetro importante puesto
que ayuda a entender la dinámica espacio-temporal de las especie del patógeno.

75
Tabla 11. Clasificación de la diferenciación genética de acuerdo al Fst propuesta por Wright
(1978).
Los resultados obtenidos con la colección del cepario sugieren la necesidad de emprender
trabajos encaminados a recolectar nuevos aislamientos de diferentes zonas del pais que
representen de una manera más amplia la diversidad que puede existir en esta especie,
especialmente en los departamentos Colombianos de Antioquia, Caldas y Cundinamarca en
donde se colectaron pocas accesiones; y Quindio y Sumapaz , en donde no se han realizado
colectas de aislamientos.
6.4.1. Análisis de Correspondencia Múltiple
El Análisis de Correspondencia Múltiple separó las introducciones en tres grupos, con
coeficientes de similitud dentro de grupos y entre grupos de 54 y 37 % respectivamente. En la
figura 8, se puede observar la distribución espacial de los 40 aislamientos de Cladosporium
cladosporioides que fueron analizados mediante la técnica RAMs. Se observa que los individuos
pertenecientes a la especie de maracuyá se unen más a la especie de gulupa, mientras que la
especie de granadilla está más disperso, aunque esto no es concluyente ya que se evaluaron muy
pocas aislamientos de esta especie.

76
Figura8. Representación espacial de los 40 aislamientos mediante el análisis de correspondencia
múltiple.
6.4.2. Distancia e Identidad Genética
Los valores de distancia genética calculados con el índice de Nei (1978) (Tabla 12) fueron altos
comparados con otros estudios de diversidad genética en bancos de germoplasma (Bonilla et al.
2008; Sanabria et al. 2006; Morillo et al. 2005; Rueda et al. 2006), la mayor distancia genética
fue para los grupos A y B (0.93), y la más baja para los grupos C y B (0.17). Corroborando de
nuevo la cercanía de los aislamientos de gulupa y maracuyá.
Tanto los valores de distancia como de identidad genética, concuerdan con los resultados
presentados hasta el momento, indicando que los seis grupos constituyen una unidad discontinua.

77
Por otra parte el árbol de distancia muestra dos agrupaciones, donde los grupos A, y B que
incluyen individuos de gulupa y maracuyá se unen a una distancia del 36% en tanto que el
grupos C corresponde a granadilla se encuentran a 54%, lo cual sugiere una amplia diversidad
genética. (Figura 13).
El dendrograma para los tres grupos construido mediante el método UPGMA (Figura 13) y con
un bootstrapp de 1000 repeticiones mostró un alto porcentaje de loci que soportan la formación
de los nodos (Tabla 12) lo que indica una alta confiabilidad de los loci utilizados en el análisis de
diversidad genética de la colección y la efectividad de la técnica molecular RAMs para el estudio
realizado.
Tabla 12. Identidad (bajo la diagonal) y distancia genética (sobre la diagonal) de los tres grupos
de accesiones de passifloras.
========================================
A B C
========================================
A **** 0.9361 0.7873
B 0.0661 **** 0.8377
C 0.2392 0.1771 ****
========================================
Tabla 13. Información de los loci que soportan la construcción del dendrograma
de distancias de los tres grupos formados.
Nodo
Proporción de
réplicas similares
(1000 repeticiones)
No. de loci
que soportan
el nodo
% Total de
loci que
soportan el
nodo
1 99,60% 28 36,84%
2 100% 76 100,00%

78
Figura 13. Dendrograma de distancia genética calculado con el índice de Nei (1978) construido
con el método de UPGMA para los tres grupos formados.
6.4.3. Análisis de Varianza Molecular
El análisis de varianza molecular AMOVA presentado en la tabla 18, permite detectar que de la
variación genética total, el 7% está explicada por la variación entre grupos mientras que para el
dentro del grupo la variación fue del 93%. Esta alta variación podría indicar la presencia de
niveles de subdivisión o jerarquización mayores o más importantes a los considerados en este
estudio.
GRLUK-
MA-7
Gulupa
Maracuya
Granadilla

79
Tabla 14. Análisis de Varianza Molecular (AMOVA) para entre grupos y dentro de grupos de
los aislamientos analizados.
AMOVA
Fuente de
Variación GL SC CM %
Entre grupos 2 62,302 31,151 7%
Dentro de grupos 38 584,088 15,371 93%
Total 40 646,390 100%
De acuerdo a lo anterior se hace necesario la realización de estudios geográficos que permitirán
entender la dinámica espacial y temporal de los aislamientos la cual ayudaría a determinar
cuáles son las causas que explican apropiadamente la distribución de la variación genética.
Figura 12. Porcentajes de variación del análisis de varianza molecular
Among Pops7%
Within Pops93%
Percentages of Molecular Variance

80
En grafica 12 se observa el análisis de varianza molecular para los tres grupos muestra que la
variación existente en la colección obedece a diferencias propias de los aislamientos dentro de
cada uno de los grupos (93%), El 3% restante se debe al componente de varianza genética entre
grupos.
6.5. Evaluación de Biofungicidas
En el análisis de la actividad antifúngica de los biofungicidas contra las especie de Cladosporium
evaluada, se detectó el efecto inhibitorio sobre la velocidad de crecimiento del hongo con
diferentes niveles según el tratamiento.
Se determinó que los biofungicidas evaluados redujeron la tasa de crecimiento de los
aislamientos en diferentes proporciones, permitiendo en la mayoría de los casos el desarrollo
tardío y alterado del crecimiento del hongo (Tabla 15).
Figura 13. Efecto de los biofungicidas evaluados y sus dosis sobre el crecimiento de C.
cladosporioides, Testigo (+) PDA, testigo (-) procloraz.
0
0,5
1
1,5
2
2,5
3
3,5
FIQ
UE
0.5
%
FIQ
UE
1%
FIQ
UE
5%
FIQ
UE
10
%
LRP
10
0%
Eco
swin
g 2
%
Tage
tes
10
%
Fun
gib
iol
TEST
IGO
Stra
tego
Ve
loci
dad
de
cre
cim
ien
to (
pe
nd
ien
te)
Tratamientos biofungicidas

81
Tabla 15. Prueba de rango múltiple de Duncan para determinar la actividad de los
tratamientos biofungicidas sobre los seis aislamientos de Cladosporium
cladosporioides.
Grupo Media N Tratamiento
A 27.6111 18 Testigo(PDA)
AB 25.8194 18 LRP 100%
B 24.6528 18 Tagetes
C 8..2222 18 Ecosiwing
D 8.0972 18 Fungibiol
E 14.4444 18 Fique 0.5%
F 12.0417 18 Fique 1%
G 9.6667 18 Fique 5%
G 8.7500 18 Fique 10%
H 0.0000 18 Stratego
Diferencia mínima significativa = 1.920. Alfa = 0.05
Figura 14 . Comportamiento general de los aislamientos de Cladosporium cladosporioides,
frente a todos los biofungicidas. Aislamientos: a: HUPEC-GU-1; b: VAYJ-GU-2; c: BOB-GU-1
; d: CUCSL-GU-1; e: CLAT-GU-1; y f: ANTR-GU-1.
0
0,5
1
1,5
2
2,5
3
3,5
Ve
loci
dad
cre
cim
ien
to (
pe
nd
ien
te)
Aislamientos

82
Tabla 16. Prueba de rango múltiple de Duncan para determinar la inhibición de los seis
aislamientos de Cladosporium cladosporioides. con los tratamientos biofungicidas.
Grupo Media N Aislamiento
A 26.2222 36 HUPEC-GU-1 B 23.2569 36 VAYJ-GU-2 C 14.5764 36 BOB-GU-1 C 13.9236 36 CUCSL-GU-1 C 13.6736 36 CLAT-GU-1 C 13.3819 36 ANTR-GU-1
Diferencia mínima significativa = 1.357 Alfa = 0.05
El comportamiento de los aislamientos frente al efecto biofungicida fue diferente, ya que se
observó que algunos aislamientos fueron capaces de desarrollarse más rápidamente que otros,
evidenciando las diferencias de sensibilidad entre aislamientos frente a la misma sustancia, y
marcando también diferencias en la sensibilidad a los biofungicidas evaluados dentro de C.
cladosporioides.; particularmente, el aislamiento HUPEC-GU-1, mostró resistente al extracto de
fique en concentraciones de 0.5, 1 y 5%, concentraciones en las cuales el hongo pudo
desarrollarse adecuadamente (comparado con el testigo PDA), inhibiendo su crecimiento
únicamente en la concentración de 10%; sin embargo este aislamiento presentó mayor
sensibilidad al extracto de ecosiwing (menor inhibición que con extracto de fique 10%) cuando
los demás aislamientos evaluados exhibieron más tolerancia a este tratamiento.

83
Figura 22. Efecto inhibitorio del crecimiento de las colonias de Cladosporium cladosporioides
con los tratamientos biofungicidas. Aislamientos: a: HUPEC-GU-1; b: VAYJ-GU-2; c: BOB-
GU-1 ; d: CUCSL-GU-1; e: CLAT-GU-1; y f: ANTR-GU-1.
Los anteriores resultados, suponen la implementación de estrategias de manejo de acuerdo con
las condiciones específicas del sitio, en las cuales se conozcan la o las poblaciones de patógenos
05
10152025
Stra
tego
FIQ
UE
0.5
%
FIQ
UE
1%
FIQ
UE
5%
FIQ
UE
10
%
LRP
10
0%
Eco
swin
g 2
%
Tage
tes
10
%
Fun
gib
iol
TEST
IGO
Cre
cim
ien
to c
olo
nia
(m
ed
ia r
adio
10
dia
s)
Tratamiento
05
10152025
Stra
tego
FIQ
UE
0.5
%
FIQ
UE
1%
FIQ
UE
5%
FIQ
UE
10
%
LRP
10
0%
Ecosw
ing…
Tage
tes
10
%
Fun
gib
iol
TEST
IGO
Cre
cim
ien
to c
olo
nia
(m
ed
ia r
adio
10
dia
s)
Tratamiento
05
10152025
Stra
tego
FIQUE…
FIQ
UE
1%
FIQ
UE
5%
FIQ
UE
10
%
LRP
10
0%
Ecosw
ing…
Tagetes…
Fun
gib
iol
TEST
IGO
Cre
cim
ien
to c
olo
nia
(m
ed
ia r
adio
10
dia
s)
Tratamiento
05
10152025
Stra
tego
FIQ
UE
0.5
%
FIQ
UE
1%
FIQ
UE
5%
FIQ
UE
10
%
LRP
10
0%
Ecosw
ing…
Tage
tes
10
%
Fun
gib
iol
TEST
IGO
Cre
cim
ien
to c
olo
nia
(m
ed
ia r
adio
10
dia
s)
Tratamiento
05
10152025
Stra
tego
FIQ
UE
0.5
%
FIQ
UE
1%
FIQ
UE
5%
FIQ
UE
10
%
LRP
10
0%
Eco
swin
g 2
%
Tage
tes
10
%
Fun
gib
iol
TEST
IGO
Cre
cim
ien
to c
olo
nia
(m
ed
ia r
adio
10
dia
s)
Tratamiento
0102030
Stra
tego
FIQUE…
FIQ
UE
1%
FIQ
UE
5%
FIQUE…
LRP
10
0%
Ecosw
in…
Tagetes…
Fun
gib
iol
TEST
IGO
Cre
cim
ien
to c
olo
nia
(m
ed
ia r
adio
10
dia
s)
Tratamiento

84
involucradas en la enfermedad, y las respuestas que estas tienen hacia los biofungicidas y a otras
prácticas como la resistencia varietal, que finalmente optimicen el paquete tecnológico adoptado
para este sitio en particular (Sánchez et al., 2007).
En un análisis general, se observó en el control con stratego, la inhibición total del crecimiento
en todos los aislamientos, asi como, en los tratamientos biofungicidas no se observó la inhibición
de los mismos, pero sí la reducción de la velocidad a la cual se desarrollaron las colonias. El
mejor efecto biofungicida se presentó con las cuatro concentraciones del extracto de fique,
resaltando que las concentraciones de 5 y 10% presentaron la mayor inhibición de los
aislamientos y entre los cuales no hubo diferencia significativa.
Figura 23. Aislamiento ANTR-GU-1 tratado con las diferentes concentraciones de biofungicidas.
A: Testigo (PDA); B: LRP 100 $; C: Tagetes 2 %; D: Stratego; E: fique 10%; F:fungibiol.
Gómez (2002) concluyó en un estudio llevado a cabo con aislamientos de C. gloeosporioides en
tomate de árbol, y evaluando concentraciones desde 0.1 a 25% de extracto de fique, que las
concentraciones mayores al 5% inhiben el desarrollo micelial del hongo, y se afecta la
A C B
E D F

85
germinación de los conidios cuando la concentración del extracto es superior al 1%; comparando
el anterior registro, se observó una marcada reducción del crecimiento, pero no la inhibición del
hongo en las concentraciones evaluadas.
El efecto de diferentes concentraciones de extracto de fique no solo es registrado sobre
Colletotrichum gloeosporioides; también se ha registrado el efecto de este extracto sobre la
germinación in vitro de esporangios de Phytophthora infestans aislados de tomate de mesa, así
como, la inhibición de este patógeno en plantas de tomate infectadas (López y Salazar, 2001).
Igualmente Aceros y Pineda (1997), registran su uso como un buen control de C.
lindemuthianum, teleomorfo Glomerella lindemuthiana, agente causante de la Antracnosis en
fríjol en condiciones in vitro, pero no en campo.
En orden de efectividad, el biofungicidad fungibiol, presentó un efecto inhibitorio marcado
después del extracto de fique en todas sus concentraciones , encontrándose diferencias
significativas con los demás tratamientos; también se pudo determinar en los aislamientos el
desarrollo anormal del micelio (Pastor-Corrales, 1991).
Álvarez et al. (2000, 2002), registran los resultados del uso del extracto de hojas de swinglea,
preparado en agua o etanol, para el control del Mildeo polvoso de la rosa ocasionado por
Sphaerotheca pannosa var. rosae. Un efecto similar se ha encontrado en el control de la
Antracnosis ocasionada por Glomerella lindemuthiana y el Añublo ocasionado por Ascochyta
spp. en fríjol (Pastor-Corrales, 1991) y para la Roya del cafeto ocasionada por Hemileia vastatrix
(Ramírez y Ñañez, 2002).
Efecto contrario se presentó con el lixiviado de raquis de plátano y el extracto de tagetes ,
quienes ejercieron un efecto inhibitorio bajo del crecimiento del hongo en concentraciones de
100 y 10%, sin observarse diferencias significativas, el hongo creció de forma similar al testigo
en PDA, no existiendo entre dichos tratamientos diferencias significativas a un nivel de
significancia del 95%.

86
Aunque los resultados obtenidos del efecto del LRP sobre los aislamientos de C. cladosporioides
evaluados, no afirman la utilización de este como una sustancia promisoria para el manejo del
patógeno en gulupa, con este se ha observado efectividad para el control de Moko del plátano
ocasionado por Ralstonia solanacearum raza 2 (CIAT, 2002), Mildeo polvoso de la rosa
ocasionado por S. pannosa (Álvarez et al., 2000; 2002) y en el manejo de las Sigatokas del
plátano ocasionadas por Mycosphaerella spp. (Escobar y Castaño-Zapata, 2005; García y
Apezteguia, 2001).
Resultados similares reporta Arango (2010), donde la inhibición de LRP para las especies de
Colletotrichum en mora pueden ser explicados por el hecho de que el lixiviado de plátano está
compuesto por una mezcla de sustancias no húmicas (azúcar, aminoácidos, polisacáridos,
proteínas), y sustancias húmicas (ácidos húmicos, AH; ácidos fúlvicos, AF y Huminas que son la
fuente más importante del Carbono orgánico en ambientes terrestres y acuáticos) (Proyecto
Residuos Rosario 2001, citado por Arenas et al. 2007), que aunque podrían inducir resistencia en
las plantas, también pueden ser fuente de 75 sustancias que favorecen el desarrollo de algunos
microorganismos. Lo anterior coincide con lo reportado por Muñoz & Madriñan (2005), quiénes
concluyeron que la aplicación de los lixiviados incrementó la actividad microbiana en un suelo
De todos los biofungicidas evaluados, el extracto de fique en concentraciones de 5 y 10%, fue
uno de los más promisorios para el manejo de la roña en gulupa, resultados que también han sido
observados en otros frutales (Gómez, 2002) y en enfermedades foliares de leguminosas, flores y
hortalizas (López y Salazar, 2001), una alternativa de control importante ya que puede contribuir
a disminuir el desarrollo de resistencia por los patógenos hacia sustancias químicas y al impacto
negativo de estas últimas en el ambiente.
Arango, (2010) reporto resultados donde mostraron que en la evaluación in vitro de los extractos
vegetales, T. patula fue el tratamiento más eficiente en la inhibición de C. gloeosporioides, B.
cinerea y C. acutatum, resultado comparable con lo reportado por Arenas et al. (2005), en donde
se registró a T. patula como un excelente tratamiento, al reducir en un 84,7 % la población de R.
solanacearum en suelo, al ser comparado con otros extractos y productos químicos para el
control de esta bacteria.

87
El efecto de F. macrophylla no solo ha sido comprobado con C. gloeosporioides; sino que
también ha sido reportado sobre la germinación In Vitro de esporangios de Phytophthora
infestans (Mont) De Bary, aislados de tomate, así como en la inhibición de este en plantas de
tomate infectadas (López & Salazar 2001). El efecto de este extracto fue igualmente evidente en
las pruebas in vitro con B. cinerea y con C. acutatum reportado por Arango (2010) en donde
hubo una inhibición intermedia, en comparación con los demás extractos evaluados, pero fue
más alta al compararlo con el testigo.
El extracto de fique al 5 y 10% presentó el mejor efecto sobre la reducción en la velocidad de
crecimiento de Cladospoirium cladosporioides . por lo cual puede considerarse su integración
como una práctica en el manejo de la enfermedad.
esto proporciona una alternativa en miras de minimizar el uso de agroquímicos perjudiciales, no
obstante, es importante hacer más investigaciones que proporcionen información acerca de los
componentes y el efecto que estos pueden tener sobre los cultivos, los consumidores y el medio
ambiente, para que puedan ser identificados como una alternativa potencial viable en el
reemplazo de los agroquímicos.
Los tratamientos evaluados, tanto biocontroladores como extractos vegetales, proporcionan una
alternativa para el manejo y control del patógeno, mostrando una alta especificidad, lo que puede
ser la base para incrementar y mejorar el manejo de las enfermedades.
6.6. Pruebas antagónicas
A partir del análisis de varianza para C. cladosporioides, se encontraron diferencias
significativas en la inhibición ejercida por las cepas de Trichoderma spp. evaluadas al
compararlas con un testigo (Tabla 17).
Trichoderma demostró un claro efecto antagónico contra los cinco aislamientos de
Cladosporium cladosporioides en los cultivos duales realizados previamente en in vitro,
aumentando su actividad antifúngica mediante la secreción del enzima hidrolítico (β-1,3-

88
glucanasa) (Ezziyyani 2004) y posiblemente otros, sobrecreciendo y reduciendo totalmente la
colonia del patógeno.
Tabla 17. Grupos homogéneos formados por comparación múltiple a partir del análisis de varianza,
del efecto de dos cepas de Trichoderma spp. sobre cinco aislamientos de Cladosporium
cladosporioides en cultivos duales.
Aislamientos Agroguard® T. viride Grupos
HUPEC-GU-1 20.14 22.1 A
VAYJ-GU-2 20.11 22 A
BOB-GU-1 20.15 22.2 A
CUCSL-GU-1 20.12 22.1 A
CLAT-GU-1 20.14 22 A
ANTR-GU-1 20.12 22 A
TESTIGO 2.5 2.5 B
La zona de inhibición producida por Trichoderma, frente al patógeno, aumenta a medida que
transcurre el tiempo, aumento que va acompañado de la destrucción del micelio fúngico
desarrollado hasta ese momento. Lo que se observa en primer lugar es una zona de
inhibición progresiva, en parte por la mayor velocidad de crecimiento de Trichoderma, Después
aparece un marcado efecto hiperparasítico, que se manifiesta por la inhibición del crecimiento
micelial no sólo por compartir el mismo sustrato sino también porque Trichoderma produce
antibióticos y enzimas: (β-1,3- glucanasa, quitinasa, proteasa y celulasa) degradadores de la
pared celular que juegan un importante papel en el micoparasitismo (Papavizas & Lumsden
1980, Herrera et al.. 1999, Sousa et al. 1998).
En la Fig. 23, se muestra el ensayo de antagonismo in vitro de Cladosporium cladosporioides
frente a Trichoderma en el medio PDA. Se puede observar que Trichoderma aumenta la
actividad antifúngica por encima del control sobrecreciendo y reduciendo la colonia del
patógeno. Es decir Trichoderma no sólo inhibe la expansión de C. cladosporioides creciendo en

89
su campo de acción, sino que a los cuatro días la colonia del patógeno C. cladosporioides se
quedó totalmente reducida en comparación con el
control.
Se encontró que las- cepas de Trichoderma permitieron un menor crecimiento del patógeno,
representando una inhibición de 42.77% y 37.77% respectivamente. Siendo significativamente
diferentes al testigo en el cual no se presentó inhibición. Los ensayos de antagonismo mostraron
inhibición en el crecimiento de los patógenos por parte de las dos cepas de Trichoderma spp
evaluadas. Entre los mecanismos que Trichoderma spp , se encuentra el micoparasitismo y la
competencia que ejerce sobre estos, lo cual pudo ser verificado en este ensayo.
Este resultado cobra importancia al tratarse de una especie diferente a la evaluada en este trabajo
y de la cual no se ha investigado mucho. No obstante, el porcentaje de inhibición fue mayor a
85%, lo cual permitió a Trichoderma spp crecer por todo el medio superando el 85% de
inhibición en la mayoría de casos, destacándose la cepa viride® por presentar los porcentajes de
inhibición más altos. El rápido crecimiento que presenta Trichoderma spp. le proporciona una
ventaja importante en la competencia por espacio y nutrientes, en comparación con los
patógenos, incluso antes que produzcan su arsenal de micotoxinas (Cook & Baker 1989, Deacon
& Berry 1992, citado por Barbosa et al. 2001).
Estos resultados son comparables con los obtenidos en otros trabajos en los que se busca la
selección de cepas de Trichoderma spp. con potencial de biocontrol.
Ensayos de Michel-Aceves et al. (2009) en la selección de las mejores cepas de Trichoderma
spp. para inhibir el crecimiento micelial de Fusarium spp. en el control de la “escoba de bruja”
en mango, fue considerando el mayor porcentaje de inhibición; que varió entre 42.0 - 62.9%.
Asimismo, Michel-Aceves et al. (2005), reportaron un porcentaje de inhibición de 77.8%, para
F. oxysporum. En un trabajo similar Arzate-Vega et al. (2006), evaluaron Trichoderma spp,
sobre Micosphaerella fijiensis (Morelet)., seleccionando 6 de 25 cepas que inhibieron al menos
el 45% del crecimiento micelial del patógeno. De igual manera, Sousa & Oliveira (1998)
reportan valores de 100% de inhibición sobre Colletotrichum gloeosporioides causante de la

90
antracnosis en el cultivo de maracuyá (Passiflora edulis ). Por otro lado, Guzmán et al. (2002)
reportaron inhibiciones entre 56 y 67% en el crecimiento de B. cinerea de lulo. De acuerdo con
los anteriores trabajos, los porcentajes de inhibición obtenidos con los patógenos evaluados en la
presente investigación, demuestran una alta viabilidad para seleccionar las cepas evaluadas como
potenciales biocontroladores.
Los resultados de este ensayo indican que en presencia de Trichoderma spp., C. cladosporioides,
presenta restricción en su crecimiento, debido a la competencia que se da al agotarse el espacio y
la cantidad de nutrientes disponibles, siendo más notorio en el caso de patógenos con
crecimiento lento. Este mecanismo que presenta Trichoderma spp. Podría llegar a ser muy útil al
emplearse en la protección de órganos como flores, hojas, tallos y frutos de plantas, ya que se
puede crear una fuerte competencia con los patógenos que las afectan (Barbosa et al. 2001).
En las distintas evaluaciones, se observó que Trichoderma spp. no sólo inhibió el crecimiento,
sino que además fue capaz de crecer sobre el patógeno, haciéndose visible crecimiento neto de
Trichoderma spp. en los últimos días de evaluación. Esta reacción es un indicio claro del
micoparasitismo que se pudo ejercer sobre los patógenos, lo cual se corroboró al realizar
observaciones en el microscopio, ya que se encontró degradación de las hifas del patógeno en la
zona de interacción con Trichoderma spp.
En estudios similares, Barbosa et al. (2001), registraron el efecto tóxico sobre aislamientos de
Cladosporium herbarum, el cual se vio reflejado en algunas alteraciones de la estructura hifal
cuando este fue puesto a crecer junto con algunas especies de Trichoderma spp. Como se ha
explicado, son varios los mecanismos mediante los cuales los antagonistas ejercen su acción, no
obstante, un análisis de los mecanismos de acción descritos anteriormente revela que en todos
ellos participan enzimas hidrolíticas de la pared celular (Rey 2000), siendo estas las que
determinan en gran parte el éxito inhibitorio.

91
Figura 23. Pruebas de cultivos duales. A: C. cladosporioides cepa (BOB-GU-1) contra
Trichoderma viridae ; B: C. cladosporioides cepa (BOB-GU-1)contra Agroguard.
6.8. Evaluación in vivo de la capacidad de control de los extractos vegetales en frutos
desprendidos de gulupa.
El objetivo de esta prueba fue determinar si los extractos lograban inhibir el desarrollo de los
síntomas en frutos desprendidos de gulupa, para esta prueba se seleccionaron los extractos
Ecosiwing y Fique 10% y Fungibiol, debido a que estos fueron los que presentaron los mejores
resultados en las pruebas de inhibición del crecimiento micelial. Se utilizó también el tratamiento
comercial de fungicida Stratego en su dosis recomendada.
Se determinaron y monitorearon los síntomas de la enfermedad en los frutos de gulupa durante
10 días. Debido a que en el día 10 la mayoría de los frutos presentaban infección del patógeno. A
partir del sexto día tras la inoculación. Se observaron los primeros síntomas de la enfermedad,
los cuales se evidenciaron por medio de un halo color verde olivo alrededor de la punción que se
colocó en cada fruto.
A B

92
Figura 24: Fruto de gulupa con los primeros síntomas de roña a los 6 días. Halo color verde con
micelio verde olivo.
Los resultados de esta prueba se determinaron realizando observaciones de síntomas y signos por
la enfermedad para esto se realizaron mediciones de severidad del patógeno, esto con el fin de
determinar cuál de los extractos causaba algún efecto en la manifestación de los síntomas de la
enfermedad.
Durante los 10 días de monitoreo de las cámaras húmedas se pudo observar que las cepa
HUPEC-GU-1, los extractos presentaron mayor efecto inhibitorio de los síntomas, se pudo ver
que la diferencia entre los tratamientos con los extractos de Ecosiwing y Fungibiol fue mayor
que para el extracto de fique en estas cepas.
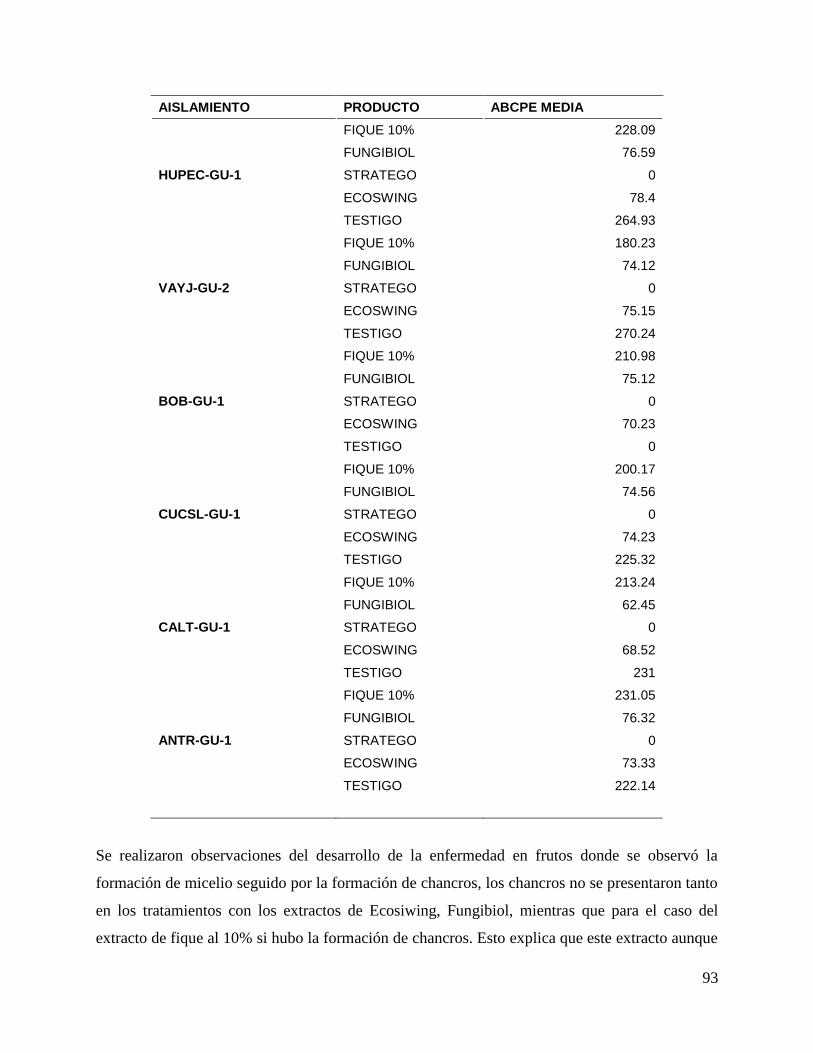
93
AISLAMIENTO PRODUCTO ABCPE MEDIA
HUPEC-GU-1
FIQUE 10% 228.09
FUNGIBIOL 76.59
STRATEGO 0
ECOSWING 78.4
TESTIGO 264.93
VAYJ-GU-2
FIQUE 10% 180.23
FUNGIBIOL 74.12
STRATEGO 0
ECOSWING 75.15
TESTIGO 270.24
BOB-GU-1
FIQUE 10% 210.98
FUNGIBIOL 75.12
STRATEGO 0
ECOSWING 70.23
TESTIGO 0
CUCSL-GU-1
FIQUE 10% 200.17
FUNGIBIOL 74.56
STRATEGO 0
ECOSWING 74.23
TESTIGO 225.32
CALT-GU-1
FIQUE 10% 213.24
FUNGIBIOL 62.45
STRATEGO 0
ECOSWING 68.52
TESTIGO 231
ANTR-GU-1
FIQUE 10% 231.05
FUNGIBIOL 76.32
STRATEGO 0
ECOSWING 73.33
TESTIGO 222.14
Se realizaron observaciones del desarrollo de la enfermedad en frutos donde se observó la
formación de micelio seguido por la formación de chancros, los chancros no se presentaron tanto
en los tratamientos con los extractos de Ecosiwing, Fungibiol, mientras que para el caso del
extracto de fique al 10% si hubo la formación de chancros. Esto explica que este extracto aunque
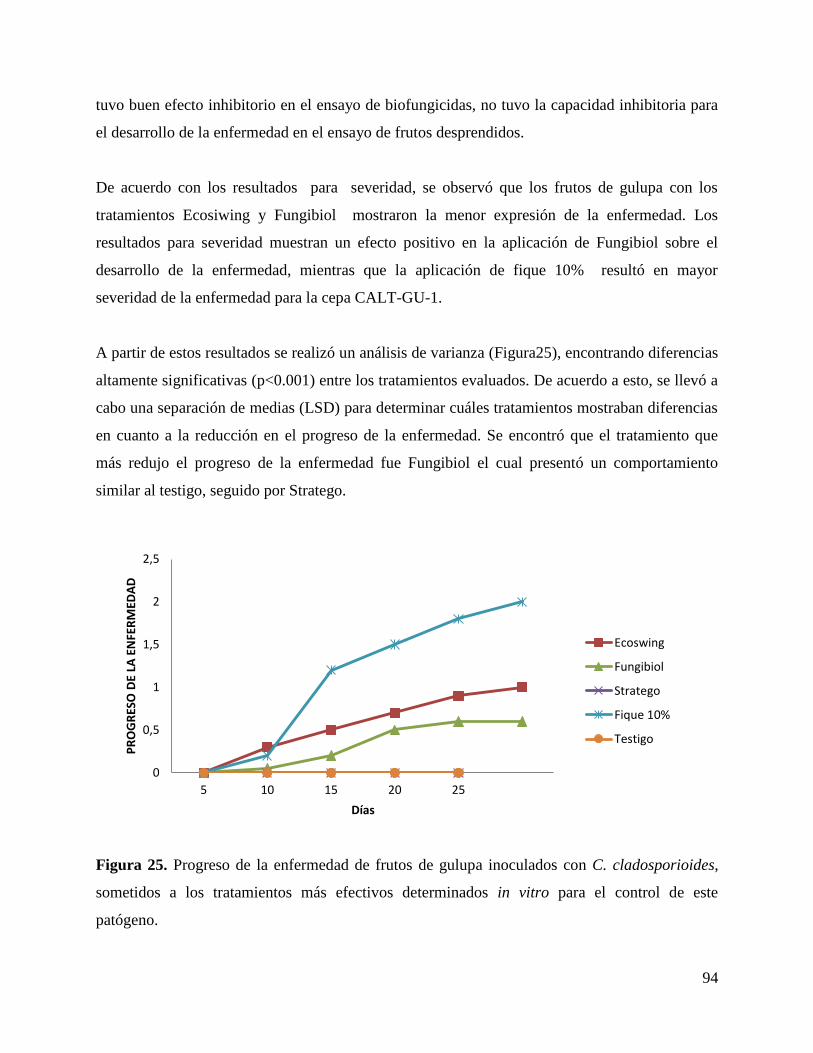
94
tuvo buen efecto inhibitorio en el ensayo de biofungicidas, no tuvo la capacidad inhibitoria para
el desarrollo de la enfermedad en el ensayo de frutos desprendidos.
De acuerdo con los resultados para severidad, se observó que los frutos de gulupa con los
tratamientos Ecosiwing y Fungibiol mostraron la menor expresión de la enfermedad. Los
resultados para severidad muestran un efecto positivo en la aplicación de Fungibiol sobre el
desarrollo de la enfermedad, mientras que la aplicación de fique 10% resultó en mayor
severidad de la enfermedad para la cepa CALT-GU-1.
A partir de estos resultados se realizó un análisis de varianza (Figura25), encontrando diferencias
altamente significativas (p<0.001) entre los tratamientos evaluados. De acuerdo a esto, se llevó a
cabo una separación de medias (LSD) para determinar cuáles tratamientos mostraban diferencias
en cuanto a la reducción en el progreso de la enfermedad. Se encontró que el tratamiento que
más redujo el progreso de la enfermedad fue Fungibiol el cual presentó un comportamiento
similar al testigo, seguido por Stratego.
Figura 25. Progreso de la enfermedad de frutos de gulupa inoculados con C. cladosporioides,
sometidos a los tratamientos más efectivos determinados in vitro para el control de este
patógeno.
0
0,5
1
1,5
2
2,5
5 10 15 20 25
PR
OG
RES
O D
E LA
EN
FER
MED
AD
Días
Ecoswing
Fungibiol
Stratego
Fique 10%
Testigo

95
Debido a la importancia que han adquirido los bioinsumos y la utilización de tecnologías
agrícolas que contribuyen al mejoramiento de la calidad de vida de los productores y la sanidad
ambiental y la introducción de tácticas de control más limpias y amigables con el medio permite
satisfacer al consumidor con un producto más sano, y de igual forma, reducir el número de
aplicaciones y costos de producción. Dentro de dichas estrategias se encuentran las plantas con
propiedades fungicidas y repelentes, que contienen compuestos promisorios (Downum,1993).
Las plantas producen compuestos con propiedades antimicrobianas que pueden ser empleadas
para controlar diferentes enfermedades en productos hortofrutícolas. La obtención de los
extractos vegetales y el estudio de sus compuestos activos propician su empleo contra diferentes
Fitopatógenos. En condiciones in vitro los extractos inhiben el crecimiento del patógeno, así
como la esporulación y germinación de esporas, de modo que ayudan a controlar las
enfermedades de frutos y hortalizas, in vivo, el efecto fungicida de los extractos vegetales varía
en función de la metodología de preparación (solvente, seco, fresco, tiempo de almacenamiento,
etc). Especie botánica, órgano de la planta (raíces, hojas, semillas, etc), sin embargo, la
combinación de los extractos vegetales con algún otro compuesto natural puede potenciar su
actividad fungicida. A la fecha son escasos los estudios básicos que incluyen el efecto de los
extractos vegetales en aspectos moleculares, bioquímicos y morfológicos del hospedero y del
patógeno (Hernández 2003).
Es importante implementar el paquete tecnológico recomendado para los agricultores por las
siguientes razones:
Prevención de impactos ambientales al emplear productos biológicos y químicos de
categorías toxicológicas III y IV (mediana y baja toxicidad), lo cual mejora las
posibilidades de mercado del producto.
Disminución del riesgo en la inversión
Mayor productividad
Mayores ingresos netos
Mejor calidad del producto, lo cual es una exigencia del mercado

96
7. CONCLUSIONES
La Roña de la gulupa en zonas productoras de este frutal en Colombia está asociada con la
especie Cladosporium Cladosporioides.
Los aislamientos de Cladosporium cladosporioides de gulupa presentan básicamente tres
niveles de patogenicidad sobre frutos: patogenicidad muy alta, patogenicidad alta y media.
Se encontró alta diversidad genética en la población de Cladosporium cladosporioides
procedente de gulupa, observándose una tendencia de los aislamientos a agruparse de acuerdo a
su especie.
En los ensayos in vitro, ecoswing y fungibiol fueron los extractos vegetales que mostraron mayor
inhibición (100%) del crecimiento de C. cladosporioides.
Las cepas Agroguard® (T. harzianum) y T. viride, mostraron una capacidad antagónica eficiente
con el patógeno , en las evaluaciones de cultivos duales, siendo la competencia por espacio-
nutrientes y el micoparasitismo los mecanismos a resaltar ejercidos por estas cepas.
Esta investigación, realizada directamente en el cultivo de gulupa, se propone como una
alternativa aplicada muy interesante ofreciendo soluciones prácticas a la fruticultura. Con este
modelo de trabajo se abren las puertas a investigaciones e interrogantes, con el ánimo final de
ofrecer al fruticultor alternativas viables en un manejo racional y ecológico del cultivo.
Los tratamientos evaluados, tanto biocontroladores como extractos vegetales, proporcionan una
alternativa para el manejo y control de los patógenos, mostrando una alta especificidad, hacia
algunos microorganismos, lo que puede ser la base para incrementar y mejorar el manejo de las
enfermedades.

97
8. RECOMENDACIONES
Colombia es una país con excelentes oportunidades comerciales en la producción de frutas
tropicales, la cual se ha visto limitada por prácticas de manejo de enfermedades inadecuadas,
esto ocasionado en muchos casos por el desconocimiento de los agentes causales; de acuerdo a lo
anterior, se recomienda la realización y/o ampliación de estudios tendientes a determinar la
composición y dinámica de las poblaciones de Cladosporium cladosporioides.
Para este ensayo los biofungicidas evaluados tuvieron un efecto significativo en la reducción de
la enfermedad aunque estadísticamente el comportamiento de fungibiol fue mejor que ecoswing.
Se recomienda la aplicación de estos dos productos para el control de la roña en campo.
Se recomienda hacer evaluaciones en plantas inoculadas por el método de aspersión tanto para el
patógeno, como para los tratamientos a evaluar (antagonistas y extractos vegetales) y de esta
manera determinar la efectividad de estos tratamientos para futuras aplicaciones en campo.
Partiendo del conocimiento actual sobre la etiología de la Roña en gulupa, es necesario realizar
estudios dirigidos a la selección de materiales con resistencia a la enfermedad. Igualmente, es
importante conocer la variabilidad del patógeno a través de estudios de la estructura genética de
las poblaciones.
El manejo de los insectos en campo, como los trips, que causan heridas al fruto desde estados
iniciales, sería de gran ayuda para evitar el desarrollo de la Roña.

98
BIBLIOGRAFIA
Aceros, P. y Pineda, E. 2001. Efecto de extractos vegetales sobre el crecimiento in vitro de
Colletotrichum lindemuthianum y sobre la severidad de la Antracnosis y los componentes de
rendimiento de fríjol Phaseolus vulgaris en dos regiones del departamento de Santander. Tesis
Ingeniería Agronómica. Bogotá, Colombia: Facultad de Agronomía, Universidad Nacional de
Colombia, Sede Bogotá, 64 pp.
Acuerdo de Frutas Huila 2006, Secretaria de Agricultura del departamento del Huila.
Agrios, G. 2005. Plant pathology. Quinta edición. Elsevier academic press. 922p.
AGRONET, Red de Información y Comunicación Estratégica del Sector Agropecuario.
2012. En: http://www.agronet.gov.co/agronetweb/; consulta: Enero 2011.
Akkaya, M. S, Bhagwat, A. A, Gregan, P. B. 1992. Lenght polymorphisms of simple Sequence
Repeat DNA in Soybean. Genetics 132: 1131-1139.
Albert, L. 1991. Primer Simposio Internacional de Pasifloras. Memorias. Universidad Nacional
de Colombia, Palmira. pp. 73-79.
Álvarez, E., C. X. Grajales, J. Villegas & J. B. Loke. 2001. Control del Mildeo Polvoso
(Sphaeroteca pannosa var. rosae) en Rosa (Rosa sp.) usando un lixiviado de Compost del Raquis
de Plátano (Musa AAB). Asocolflores 41-47.
Álvarez, E., J. B. Loke & C. Echeverri. 2002. Validación de un biofungicida y fertilizantes
foliares como alternativa de control del Mildeo Polvoso en cultivos de rosa. Asocolflores 31-40.
Anderson, J. 2009. Developing new comercial varieties, In: Annual Industry Report 08-09
Angulo Carmona, R. 2010. “Gulupa, la fruta de la pasión”. En El Agro, núm. 11, pp.12-13.

99
APIA, Association Australian Passionfruit Industry 2008. Annual levy payers meeting
minutes from 080822. Horticulture Australia Limited (HAL). 11 p.
Arango, Jimenez, L. 2010. Control de Botrytis cinerea y Colletotrichum spp. mediante el uso de
antagonistas y extractos vegetales. Tesis Ingeniería Agronómica. Universidad del Valle. Cali,
(Valle del Cauca), Colombia: Facultad de Ciencias.
Arango, Jorge. 1993. Evaluación de diferentes distancias de siembra y su incidencia en la
Antracnosis del tomate de árbol (Cyphomandra betacea (Cav.) Sendt.). Tesis Facultad de
Ciencias Agropecuarias. Medellín : Universidad Nacional de Colombia, 126 pp.
Arauz, Felipe. 1986. Enfermedades de postcosecha del mango y su combate. Programa de
Comunicación Agrícola. San José (Costa Rica): Escuela de Fitotecnia, Universidad de Costa
Rica, 20 p.
Arenas, Adriana. 2007. Caracterización molecular y patogénica de aislamientos de
Colletotrichum spp., asociados a la Antracnosis de mora en el Valle del Cauca. Trabajo de Tesis.
Cali, Colombia: Universidad del Valle, Facultad de Ciencias, Programa Académico de Biología.
p. 89.
Arzate-Vega, J., A. C. Michel-Aceves, V. M. Domíngez-Márquez y O. A. Santos-Eméstica.
2006. Antagonismo de Trichoderma spp., sobre Mycosphaerella fijiensis Morelet, agente causal
de la sigatoka negra del plátano (Musa sp. in vitro e invernadero. Revista Mexicana de
Fitopatología 24(2):98-104.
Asohofrucol, 2005.Asociación Hortofrutícola de Colombia. Exportaciones por producto
frutas:http://frutasyhortalizas.com.co/portal/includej/formatos/Exportaciones.pdf.consulta:Octubr
e 2012.

100
Asohofrucol, 2007. Asociación Hortofrutícola de Colombia. Exportaciones por producto frutas
en:htpp//frutahortalizas.com.co/portal/includej/formatos/Exportaciones.pdf.consulta 03 de
Octubre 2011
Aukar, A., E. Lemos y J. Oliveira. 2002. Genetic variations among passion fruit species using
RAPD markers. Rev. Bras Fruiticultura. 24: 738-740.
Bailey J. A. & M. J. Jeger. 1992. Colletotrichum: Biology, pathology and control. UK:
Redwood Press Ltd, Melksham. 388p.
Barbosa, M. A., K. G. Rehn, M. Menezes & R. L. Mariano. 2001. Antagonism of
Trichoderma species on Cladosporium herbarum and their enzymatic characterization. Brazilian
Journal of Microbiology. 32(2).
Bellon, G., F, Faleiro. K. Junqueira, N. Vilela, E. Carvalho, M. Braga y C. Teixeira. 2007.
Variabilidade genética de acessos silvestres e comerciais de Passiflora edulis Sims com base em
marcadores RAPD. Rev. Bras. Frutic., Jaboticabal – SP. 29:124-127.
Bellon, G., F, Faleiro. K. Junqueira, N. Vilela, E. Carvalho, M. Braga y C. Teixeira. 2007.
Variabilidade genética de acessos silvestres e comerciais de Passiflora edulis Sims com base em
marcadores RAPD. Rev. Bras. Frutic., Jaboticabal – SP. 29:124-127.
Bonilla M. L., Espinosa K., Posso A. M., Vasquez H., Muñoz J. E. 2008. Caracterización
molecular de 43 accesiones de uchuva de seis departamentos de Colombia. Acta Agronómica
57(2) 109-115.
Bridg, Hannia. 2005. Micropropagation and determination of the in vitro stability Passiflora
edulis. Trabajo de Tesis.: Universidad de Humboldt, Facultad de Ciencias Agrícolas,
Departamento de Ciencias de las Frutas,120 pp.

101
Cabra, Jorge et al. 2002. Material para siembra y selección de variedades en un programa de
frutales tropicales En: Taller para Colombia y el Mundo: Optimización de la Cadena Productiva.
Corporación BIOTEC y CIAT, p. 37- 47.
Carvalho, M., O. Kiyoshi, N. Junqueira, J. Peixoto y G. Fagundes. 2005. Producao de mudas
de tres genotipos de maracujazeiro-roxo pelo método de enxertia, en: IV reuniao técnica de
pesquisas em maracujazeiro. Brasil, EMPRAPA. 229 p.
Castaño, J. 2008. Enfermedades importantes de las pasifloráceas en Colombia. En: Memorias
Seminario Nacional sobre Pasifloráceas. Sociedad Colombiana de Ciencias Hortícolas. 25p.
Castaño-Zapata, Jairo. 1997. Manual para el diagnóstico de hongos, bacterias, virus y
nemátodos fitopatógenos. Manizales: Universidad de Caldas, Facultad de Ciencias
Agropecuarias,. 210 pp.
Castro, L. 2001. Guía Básica para el establecimiento y mantenimiento del cultivo de la
granadilla (P. ligularis). ASOHOFRUCOL Fondo Nacional de Fomento Hortofrutícola. pp. 40-
46.
Caujape, J. 2006. Brújula para botánicos desorientados en la genética de poblaciones. EXEGEN
Ediciones. Las Palmas de Gran Canaria. España. 132 p
Chacón, C. y J. Serna. 1990. El cultivo del Maracuyá. Federación Nacional de Cafeteros de
Colombia, Cali. pp. 3-30.
Chung, Wen-Hsin et al. 2006. Fungicide sensitivity and phylogenetic relationship of
anthracnose fungi isolated from various fruit crops in Japan. En: Plant Dis. Vol. 90 ; p. 506-512.
Clough, J. M. y C. Godfrey. 1998. The strobilurin fungicides. En: Fungicidal Activity.
Chemical and Biological Approaches to Plant Protection. Wiley, England. pp. 109-148.

102
CONSUMER.ES. Preocupación entre los investigadores por la elevada presencia de residuos
tóxicos.Colombia consultado ene., 2013.
http//www.consumaseguridad.com/web/es/sociedad_y_consumo/2003/01/29/4914.php
Coppens, G., S. Segura, J.E. Hodson y G. Góngora. 1997. Les fruits de la Passion. In:
Charrier, A., M. Jacquot, S. Hamon y D. Nicolas, L’amélioration Des Plantes Tropicales.
CIRADORSTOM Coedition, Montpellier. Pp 291-312.
Corporación Colombia Internacional (CCI). 2008. En: http://www.cci.org.co.; Consulta:
Octubre de 2012.
Corporación Colombiana Internacional (CCI). 2006. Inteligencia de mercados; maracuya.
Perfil del producto No. 19. ISSN 0124-1338 en http://cci.org.co/cci/cer_x/scripts/index.php;
Consulta Octubre 2012.
Crochemore, M., H. Correa y L. Esteves. 2006. Genetic Diversity in Passion Fruit (Passiflora
spp.) Evaluated by RAPD Markers. Brazilian Archives of Biology and Technology. 4: 521-527.
Crochemore, M., H. Molinari, N. Colauto. 2003. Caracterização agromorfológica do
maracujazeiro (Passiflora spp.). Rev. Bras. Frutic., Jaboticabal – SP. (25), 5-10.
Cronquist, A. 1981. An integrated system of classificacion of flowering plants. Columbia
University Press, New York.
Delgado, Mendez, C. 2012. Caracterización morfológica y molecular del agente causante de la roña
de las pasifloras. . Tesis de grado. Universidad de Caldas. Caldas (Manizales). Trabajo de tesis
presentado como requisito parcial para optar el título de: Magíster en Fitopatología.132p
Dennis, C. & J. Webster. 1971. Antagonistic properties of speciesgroups of Trichoderma III.
Hyphal interaction. Trans. Br. Mycol. Soc. 57:363-369.

103
Devos, R., C. Ruggiero, E. De Macedo, G. Garcia, M. Mantovani, E. Alves y T. Wickerrev.
2004. Diversidad genética em maracujazeiro-amarelo utilizando marcadores moleculares fAFLP.
Rev. Bras. Frutic. 26: 494-498.
Dodd, J.C.; Estrada A. & Jeger M.J. 1992. Epidemiology of Colletotrichum gloeosporioides
in the Tropics. En: Colletotrichum: Biology, pathology and control. Bayley J.A. & Jeger M.J.
(Eds). Wallingord : CAB International, p. 308-325.
Downum R, Kem. 1993. Phytochemical potential of tropical p l a n t s. I n Phytochemical
potential of tropical plants. Downum, K; Romeo, J; Stafford, H. Eds. New York, Plenum Press. p
287.
Elias, M., O. Panauda and T. Roberta. 2000. Assessment of genetic variability in a traditional
cassava (Manihot esculenta Crantz) farming system, using AFLP markers. Heredity. 85: 219–
230.
Escobar T. W. & Sánchez, L.A. 1992. Fruticultura colombiana, guanábano. Manual de
Asistencia Técnica No 57. Bogotá : ICA. PRODUMEDIOS,. 100 pp.
Escobar, J. H. & Castaño Zapata, Jairo 2005. Fulvic acid applications for the management of
diseases caused by Mycosphaerella spp. In: Infomusa, Francia. Vol. 14, No. 2; p. 15-17.
Escobar, J. H. y Castaño-Zapata, Jairo. 2005. Fulvic acid applications for the management of
diseases caused by Mycosphaerella spp. In: Infomusa, Francia. Vol. 14, No. 2 ; p. 15-17.
Estrada, A. B.; Dodd, J. C. & Jeddries, P. 1993. Effects of environment of the in-vitro growth
and development of Colletotrichum gloeosporioides isolates from The Philippines. En: Acta
Horticulturae. Vol. 341, mango IV, p. 360-369.

104
Excoffier, L., P. Smouse & J. Quatro. 1992. Analysis of molecular variance inferred from
metric distance among DNA haplotypes: application to human mitochondrial DNA restriction
data, Genetics 131:479–491.
Ezziyyani, M., C. Pérez, A. Sid ahmed, M. E. Requena & M. E. Candela. 2004. Trichoderma
harzianum como biofungicida para el biocontrol de Phytophtora capsici en plantas de pimiento
(Capsicum annuum L.). Anales de Biología 26: 35-45.
Fajardo, D., F. Angel, M. Grum, J. Tohme, M. Lobo. W. Roca and I. Sanchez. 1998.
Genetic variation analysis of the genus Passiflora L. using RAPD markers. Euphytica. 101: 341-
347.
Faleiro, G., N. Junqueira, M. Braga, G. Bellon, J. Peixoto. 2005. Diversidade genética de
variedades comerciais de maracujazeiro-azedo com base em marcadores RAPD. In: reunião
técnica de pesquisas em maracujazeiro. Planaltina, DF: Embrapa Cerrados.105-109 pp.
FAO, 2002. Commodities And Trade Division Food And Agriculture Organization Of The
United Nations Rome-Italia. En: http://www.fao.org.; consulta octubre 22 de 2012.
FAO, 2003. Commodities And Trade Division Food And Agriculture Organization Of The
United Nations Rome-Italia. En: http://www.fao.org.; consulta octubre 22 de 2010.
Fayette, J. 2007. Efecto in vitro del efecto microbiano de extractos vegetales sobre el
crecimiento de una bacteria y dos hongos fitopatógenos. Trabajo de grado. Universidad de Costa
Rica. p 122.
Fitzell, R.D.; Peak, C.M. & Darnell, R.E. 1984. A model for estimating infection levels of
anthracnose disease of mango. In: Ann. appl. Biol. Vol. 104; p. 451-458.
Franco, F. y J. Zuleta. 2006. Guía de campo para manejo de plagas y enfermedades en
granadilla. UNISARC, Santa Rosa de Cabal. pp. 2-4.

105
Fundación Sonrisas de colores. 2004. Secretaria Agricultura del Huila. Acuerdo de
productividad cadena frutícola, 2006 fungicides. Acta Horticulturae (ISSH) 21:85-89.
García, Eveling & Apezteguia 2001, Hermelinda. Estudio del lixiviado de compost y su efecto
sobre el control de Sigatoka negra (Mycosphaerella fijiensis Morelet) y el crecimiento del cultivo
del banano (Musa AAA). Tesis Ingeniero Agrónomo. Guácimo, Costa Rica : Universidad Earth,.
82 pp
García, Eveling & Apezteguia., Hermelinda. 2001. Estudio del lixiviado de compost y su
efecto sobre el control de Sigatoka negra (Mycosphaerella fijiensis Morelet) y el crecimiento del
cultivo del banano (Musa AAA). Tesis Ingeniero Agrónomo. Guácimo, Costa Rica :
Universidad Earth, 82 pp.
García, M. 2002. Guía técnica Cultivo de maracuyá Amarillo. Centro Nacional de Tecnología
Agropecuaria y Forestal. Ciudad Arce, La Libertad, El Salvador. 31p.
GenBank (www.ncbi.nlm.nih.gov)
Girard, E. & Lobo, M. 1987. El cultivo del tomate de árbol (Cyphomandra betacea (Cav.)
Sendt.). Manual de Asistencia Técnica No. 32. Bogotá : Instituto Colombiano Agropecuario
(ICA), 42 pp.
Gisi, U.; K. M. Chin; G. Knapova; R. Küng-Färber; U. Mohr; S. Parisi; H. Sierotski; U.
Stenfield 2000. Recent Developments in Elucidating Modes of Resistance to Phenylamides,
DMI and Strobilurin Fungicides, Crop Protect. 19:863-872,.
Gómez, Hurtado, J. E. 2002. Evaluación del extracto de fique en el desarrollo in vitro de
Colletotrichum gloeosporioides (Penz.) Penz. & Sacc. y Sclerotinia sclerotiorum (Lib.) de Bary.
En: IV Seminario Nacional. Frutales de Clima Frío Moderado. Medellín; p. 183-188.

106
Gottwald, T. R. 1983. Factors affecting spore liberation by Cladosporium carpophilum.
Phytopathology 73:1500-1505.
Gottwald, T. R. 1985. Influence of temperature, leaf wetness period, leaf age, and spore
concentration on infection of pecan leaves by conidia of Cladosporium caryigenum.
Phytopathology 75:190-194.
Guzzo, F., S, Ceoldo. F, Andreetta. F y Levi, Marisa.2004. In vitro culture from mature seeds
of passifora species. Scientia Agricola, v 61, n.1 108-113.
Hanlin, R. 1990. Illustrated genera of Ascomycetes. St. Paul, Minnesota : The American
Phytopathological Society, APS Press,. p. 40-41.
Hantula, J., M. Dusabenyagasani., R.C. Hamelin. 1996. Random Amplified Microsatellites
(RAMS) - a novel method for characterizing genetic variation within fungi. Eur. J. For. Path.
26:59-166.
Hartung, J. S.; Burton, C. L., & Ramsdell, D. C. 1981. Epidemiological studies of blueberry
anthracnose disease caused by Colletotrichum gloeosporioides. In: Phytopathology. Vol. 71; p.
449-453.
Henríquez, M. A. 2003. Análisis de diversidad genética de patógenos con énfasis en
Phaeoisariopsis griseola agente causal de la mancha angular. Facultad de Ciencias
Agropecuarias 1 (1): 30-37.
Holb, I. 2008. Timing of first and final sprays against apple scab combined with leaf removal
and pruning in organic apple production. Crop Protection 27:814-822.
Holm - Nielsen, L.B., P. Jorgensen, J.E. Lawesson.1988. Passifloraceae. In: Harling & L.
Andersson (eds.), Flora del Ecuador 31: 124pp.

107
Howell, C. R. 2003. Mechanisms employed by Trichoderma species in the biological control of
plant diseases: the history and evolution of current concepts. Plant Disease 87(01):4-10.
Instituto Colombiano Agropecuario (ICA). www.ica.gov.co. Consultada Enero, 2012.
Iriarte, G. A. 200.2. Identificación de marcadores asociados a QTLs para características
agronómicas en una retrocruza avanzada entre una accesión silvestre y una variedad andina
mejorada (ICA Cerinza) de fríjol común (Phaseolus vulgaris L.). Tesis de Grado Ingeniería
Agronómica. Ibagué, Colombia : Universidad del Tolima,. 160 pp.
Jiménez, Y.; Carranza, C. y Moreno, M. 2009. “Manejo integrado del cultivo de gulupa
(Passiflora edulis fv edulis Sims.)”. En: Miranda D. et al. (Eds.). Cultivo, poscosecha y
comercialización de las pasifloráceas en Colombia: maracuyá, granadilla, gulupa y curuba.
Colombia: Sociedad Colombiana de Ciencias Hortícolas.
Kong, P., C.X. Hong., P.W. Tooley., K. Ivors., M. Garbelotto & P.A. Richardson. 2004.
Rapid identification of Phytophthora ramorum using PCR-SSCP analysis of ribosomal DNA
ITS-1. Letters in Applied Microbiology 38:433-439.
Latorre, B. A., E. Agosin, R. San Martin & G. S. Vásquez. 1997. "Effectiveness of conidia of
Trichoderma harzianum produced by liquid fermentation against Botrytis bunch rot of table
grapes in Chile". Crop Protection 16:209-214.
López, M.A & Salazar, M., Lucía 2001. Evaluación del jugo de fique (Furcraea macrophylla
Baker), como biorregulador del hongo patógeno Phytophthora infestans (Mont.) de Bary del
tomate de mesa (Lycopersicum esculentum Mill). Tesis de Grado Ecología. Popayán : Facultad
de Ciencias Naturales, Fundación Universitaria de Popayán, 91 pp.
López, Miguel A y Salazar, Martha L. 2001. Evaluación del jugo de fique (Furcraea
macrophylla Baker), como biorregulador del hongo patógeno Phytophthora infestans (Mont.) de

108
Bary del tomate de mesa (Lycopersicum esculentum Mill). Tesis de Grado Ecología. Popayán :
Facultad de Ciencias Naturales, Fundación Universitaria de Popayán,. 91 pp.
Lugo, O. 2007. Parámetros para implementación de Buenas Prácticas Agrícolas (BPA) en
gulupa (Passiflora edulis Sims.) con énfasis en EUREPGAP. Trabajo de Grado. Facultad de
Agronomía. Universidad Nacional de Colombia, Bogotá. 20p.
Mahuku, G. S., Hsiang, T., and Yang, L. 1998. Genetic diversity of P. griseola isolates from
turfgrass. Mycol. Res. 102:559-567.
Marín, E. 2007. Avances en la implementación de las Buenas Prácticas Agrícolas (BPA) en
frutas de exportación en los departamentos de Boyacá y Cundinamarca (Colombia). En:
Memorias Segundo Congreso Colombiano de Horticultura: Colombia Hortícola: Retos y
Oportunidades. Sociedad Colombiana de Ciencias Hortícolas. pp. 77-84.
Martínez, A. 2002. Características y estructura de los frutales de exportación en Colombia. En:
Ministerio de agricultura y desarrollo rural observatorio de competitividad agrocadenas.
http://www.agronet.gov.co/agronetweb/; consulta: Enero 2013.
Martínez-Culebras, P. V., Barrio, E., Garcia, M. D. & A. Querol. 2000. Identification of
Colletotrichum species responsible for anthracnose of strawberry based on the internal
transcribed spacers of the ribosomal region. FEMS Microbiology letters 189: 97-101.
Mendoza, cb. y moreno, M. 2006. Evaluación del efecto de extractos vegetales sobre el
crecimiento in vitro de Phytophthora palmivora y Colletotrichum gloeosporioides.. [Proyecto de
Graduación Lic. Ing. Agr.]. Guácimo (CR): Universidad EARTH, 44 p.
MicheL-Aceves A.C., M.A. Otero-Sánchez, L. Y. Solano-Pascacio. 2009. Biocontrol in vitro
con Trichoderma spp. de Fusarium subglutinans (Wollenweb. y Reinking) Nelson, Toussoun y
Marasas y F. oxysporum Schlecht., agentes causales de la “Escoba de bruja” del mango
(Mangifera indica L.). Revista mexicana de Fitopatología. 27(1):18-26.

109
Miller, M.P. 1998. A program for the preparation of AMOVA input files from dominat-marker
raw data. AMOVA-PREP Versión 1.01.
Ministerio de Agricultura y Desarrollo Rural 2006. Apuesta exportadora agropecuaria 2006-
2010. En: Ministerio de Comercio Industria y Turismo, República de Colombia,
http://www.minicomercio.gov.co/eContent/documentos/Competitividad/InsumosApuestal.pdf;
consulta: Enero 2013.
Molina, L., M. Soares. L. Bernacci y R. Silva. 2005. Capitulo 3: Melhoramento genético do
maracujá: pasado e futuro. En: Maracujá germolasma e melhoramento genético. Brasil,
EMBRAPA. 677 p.
Mora, Jorge. 1996. Progreso de la antracnosis (Colletotrichum gloeosporioides) en el
guanábano (Anona muricata), bajo dos sistemas de manejo de la enfermedad en la zona atlántica
de Costa Rica. En: memorias X Congreso Nacional Agronómico y de Recursos Naturales Vol.
2. p. 86.
Mora. Hernández. R. 2012. Análisis epidemiológico de roña en gulupa (Passiflora edulis fv
edulis Sims.) en la región del Sumapaz, Colombia. Tesis de grado. Universidad Nacional de
Colombia. Sede Bogotá. Trabajo de tesis presentado como requisito parcial para optar el título
de: Magíster en Ciencias Agrarias con énfasis en Fitopatología.
Moraga, M. 2003. Efectividad de fungicidas sobre la incidencia y severidad de Botrytis cinérea
Pers. en vid cv. Sauvignon Blanc. Universidad de Talca. Tesis pregrado. Escuela de Agronomía.
Morillo, A. C., Morillo, Y., Muños, J.E., Vasquez, H.D & Zamorano. 2005. Caracterización
molecular con microsatélites aleatorios RAMS de la colección de mora, Rubus spp, de la
Universidad Nacional de Colombia sede Palmira. Acta Agronómica (Colombia) 54(2):15-24.

110
Morillo, Y., Morillo, A. C., Muños, J. E., Vasquez, H. D., Zamorano, A. 2005.
Caracterización molecular con microsatélites aleatorios RAMs de la colección de mora, Rubus
spp, de la Universidad Nacional de Colombia sede Palmira. Acta Agronómica 54 (2).
Muminovic, J., A. Melchinger, T. Lubberstedt. 2004. Genetic diversity in cornsalad
(Valerianella locusta) and related species as determined by AFLP markers. Plant Breed. 123:
460–466.
Muñoz, J. E., Morillo, A. C., Morillo, Y. 2008. Microsatélites amplificados al azar (RAMS) en
estudios de diversidad genética vegetal. Acta Agronómica 57 (4).
Muñoz, R. E. & R. Madriñan. 2005. Efecto de lixiviados del raquis de plátano sobre la
actividad y biomasa microbiana en floración y cosecha del tomate. Acta Agronómica. 54(1).
Murillo, W. 2009. Evaluación preliminar de la actividad fungicida de los aceites esenciales de
eucalipto (Eucalyptus tereticornis, Myrtaceae) y cáscara de naranja (Citrus sinensis, Rutaceae)
sobre algunos hongos filamentosos. Tesis de grado. Universidad Nacional de Colombia, sede
Medellin.
Nakasone, H y R. Paull. 1998. Tropical fruits. CAB International, Wallingford, UK. pp. 270-
291
Nei, M. 1978. Estimation of average heterozygosity and genetic distance from a small number of
individuals. Genetics 89: 583-590.
Neira, Y. 2006. El cultivo de la gulupa Passiflora edulis Sims. Trabajo de Especialización.
Universidad Nacional de Colombia. Facultad de Agronomía. Escuela de Posgrados. Bogotá. 49p
Ocampo, J. 2007. Capitulo 4: Chloroplast and mitochondrial DNA variation in the genus
Passiflora L. (Passifloraceae) as revealed by PCR-RFLP. En: Study of the diversity of genus
Passiflora L. (Passifloraceae) and its distribution in Colombia. Trabajo de grado para optar al

111
título de Doctor en Ciencias Agronómicas. Ecole Nationale Supérieure Agronomique De
Montpellier. pp 148-192.
Ocampo, J., G. D'Eeckenbrugge, C. Olano y R. Schnell. 2004. AFLPs analysis for the study
of genetic relationships among cultivated Passiflora species of the subgenera Passiflora and
Tacsonia Proceedings of the Interamerican Society for Tropical Horticulture. 48: 72-76
Oliveros C., C. M. 2000. Recopilación bibliográfica de seis especies de frutales tropicales: lulo,
mora, uchuva, pitaya, guanábano y tomate de árbol. Cali : CIAT, 245 pp.
Ondieki J. J. y J. M. Kori. 1971. Control of brown spot disease of passion fruit in Kenya using
Ortiz D. Carolina 2010. Estudio de la variabilidad genética en materiales comerciales de
gulupa (Passiflora edulis f. edulis Sims) en Colombia. Tesis para optar al título de Magíster en
Ciencias Agrarias Área de énfasis en Genética y Fitomejoramiento.
Ortíz, D., F. Cadena, M. Delgado y T. Mosquera. 2010. Capítulo 3: Descripción de los
sistemas productivos de gulupa. En: Cartilla avances del grupo de investigación en gulupa.
Facultad de Agronomía, Universidad Nacional de Colombia sede Bogotá. P 123
Papavizas, G. C., J. A. Lewis & T. H. abd-elmoity. 1982. Evaluation of new biotypes of
Trichoderma harzianum for tolerance to Benomyl and enhanced biocontrol capabilities.
Phytopathology 72: 126-132. Passionfruit. pp 6-7.
.
Pastor-Corrales, M. A. 1991. Management of bean diseases with plant extracts. In: Annual
Report, Bean Program. Centro Internacional de Agricultura Tropical (CIAT). Cali, Colombia.. p.
73-83.
Peteira B, Perez S, Fraga Y. 1999. A modified DNA extraction minipreparation protocol for
RAPDs analysis of genetic variability in C. fulvum. Rev. Prot Veg.;(2):121-124.

112
Potokina, E., F. Blattner, T. Alexandrova, K. Bachmann. 2002. AFLP diversity in the
common vetch (Vicia sativa L.) on the world scale. Theor Appl Genet 105:58–67.
Ramírez T., A. M. & Ñañez, M. A. 2002. Evaluación del extracto vegetal de Swinglea
glutinosa BL como biorregulador del hongo Hemileia vastatrix Berk & Br., patógeno que ataca
el cultivo del café. Tesis de Pregrado Ecología. Popayán : Fundación Universitaria de Popayán.
Riascos, D., E. Ortíz, R. Mora, C. García, L. Hoyos y C. De La Rotta. 2010. Capitulo 2:
Enfermedades de la gulupa. En: Cartilla avances del grupo de investigación en gulupa. Facultad
de Agronomía, Universidad Nacional de Colombia. Bogotá.
Rivero, M. 2000. Reconocimiento y diagnóstico de enfermedades de plantas: Guías para las
prácticas de laboratorio de la asignatura de fitopatología general. Pp. 21-31.
Rohlf, F. J. 1998. NTSYSpc: Numerical Taxonomy and Multivariate Analysis System version
2.0. Exeter Software, New Cork.
Rojas, E. A. 2008. Identificación de estrategias de manejo de la antracnosis (Glomerella cingulata
(Stonem.) Spauld & Schrenk. del Guanábano (Annona muricata L.). Tesis de maestría, Manizales-
Colombia, Universidad de Caldas, Facultad de Ciencias Agropecuarias.
.
Rondón, A.; Arnal, E.; Soto, E. y Solórzano, R. 1995. El manejo integrado de enfermedades
en algunos frutales de ciclo corto. FONAIAP 12: 45-49.
Saldarriaga, Cardona. Alegría. 1997. Enfermedades del cultivo del tomate de árbol en
Antioquia: Guía de reconocimiento y control. En: Boletín Técnico, Corporación Colombiana de
Investigación Agropecuaria Corpoica ; C.l. La Selva, Rionegro (Antioquia). p. 8-10.

113
Sanabria, H., García, M., Díaz, H., Muñoz, J.E. 2006. Caracterización molecular con
marcadores RAMs de árboles nativos de Psidium guajava (guayaba) en el Valle del Cauca. Acta
Agronómica 55 (1): 23-30.
Sánchez, Danilo et al. 2007. Agricultura específica por sitio en frutales. Tecnología desarrollada
en las fincas para cada finca. Corporación BIOTEC, Heig-vd, CIAT, SAG, Cenicaña, Asocaña,
Acción Social, Colciencias, Ministerio de Agricultura y Desarrollo Rural, 48 pp.
Sánchez, I., F. Angel. M. Grum., M. Duque, M. Lobo, J. Tohme and W. Roca. 1999.
Variability of chloroplast DNA in the genus Passiflora L. Euphytica 106: 15-26.
Sanders, G. M. y Korsten, L.1999. Survey of fungicides resistance of Colletotrichum
gloeosporioides from different avocado production areas. In: South African Avocado Growers’
Association Yearbook. Vol. 22; p. 35-38.
Santos, F. M. 2008. Seleção de maracujazeiro (Passiflora edulis) para resistência à
cladosporiose (Cladosporium herbarum). Universidade Federal de Viçosa. Programa de Pós-
graduação em Genética e Melhoramento. 104 p.
SAS Institute Inc. 2000. SAS OnlineDoc , Version 8, Cary, NC.
Segura, S., A. Bohorquez. P. Ollitrault y J. Tohme. 2002. An AFLP diversity study of the
genus Passiflora focusing on subgenus Tacsonia. Genetic Resources and Crop Evolution. 1: 1-
13.
Segura, S., G. Coppens d’Eeckenbrugge and P. Ollitrault. 1998. Isozyme variation in five
species of Passiflora subgenus Tacsonia and Passiflora manicata XLIV Annual Meeting of the
Interamer. Soc. Trop. Barquisimeto, Venezuela. September 28- October 2.

114
Sousa, J.R., & N.T. Oliveira. 1998. In vitro antagonistic potential of Trichoderma spp. against
Colletotrichum gloesporiodes agent of antracnose in the passion fruit (Passiflora). Boletín
Micológico 13:103-110.
STATISTIX version 8.0. Analytical Software, St. Paul, Minn.
Stefanova, N.M. & R.I. Sandoval. 1997. Efectividad de biopreparados de Trichoderma spp., en
el control de fitopatógenos de suelo. Boletín Técnico CIV INISAV. 2:10-22.
Tafur, R., P. Perfetti, D. Ruiz, J. Morales. 2006. Plan frutícola Nacional. 43 p. Talca (Chile). 42p.
Tamayo, P. 2004. Evaluación de desinfestantes de herramientas para el control de la secadera de
la granadilla. Ascolfi Informa 30:12-14.
Trujillo, N., C. Márquez, J. Moreno, W. Terán y I. Schuler. 2010. Caracterización molecular
de materiales cultivados de gulupa (Passiflora edulis f. edulis) Universitas Scientiarum. 14: 135-
140.
Uesugui, Y. 1998. Fungicide classes: chemistry, uses and mode of action. En: Fungicidal
Activity. Chemical and Biological Approaches to Plant Protection. Wiley, England. pp. 29-33.
Ulmer, T. y J. MacDougal. 2004. Passiflora, passionflowers of the world. Timber press,
Portland-Cambridge. 407pp.
.
Valdés, H., M. Fermaud, J. Roudet, A. Calonnec y C. Gary. 2008. Grey mould incidence is
reduced on grapevines with lower vegetative and reproductive growth. Crop Protection 27:1174-
1186
Viana, A., T. Santana, M. Pereira y M. De Souza, J. Martinez y A. Amaral. 2003.
Diversidade genética entre genótipos comerciais de maracujazeiroamarelo (Passiflora edulis f.
flavicarpa) e entre espécies de passifloras nativas determinada por marcadores RAPD. Rev.
Bras. Frutic. Jaboticabal – SP. 25: 489-493.

115
Villacis, L., J. Vega, M. Grum y G. Coppens d’Eeckenbrugge. 1998. Morphological
characterization of Andean passifloras (Passiflora spp.) from Ecuador. Plan Gen. Res. Newsl.
115: 51-55
Villamil, F. 2008. Las buenas prácticas agrícolas en pasifloráceas. En: Memorias Seminario
Nacional sobre Pasifloráceas. Sociedad Colombiana de Ciencias Hortícolas. 25p.
Viveros, J. & J. Castano-Zapata. 2006. Evaluación in vitro de extractos vegetales sobre
Mycosphaerella fijiensis Morelet. Agronomía 14(1):37-50.
Weindling, R. 1934. Studies on a lethal principle effective in the parasitic action of Trichoderma
lignorum on Rhizoctonia solani and other soil fungi. Phytopathology 24:1153-1179.
Willingham, S., K. Pegg, P. Langdon, A. Cooke, D. Peasley y R. McLennan. 2002.
Combinations of strobilurin fungicides and acibenzolar (Bion) to reduce scab on passionfruit
caused by Cladosporium oxysporum. Australasian Plant Pathology 31:333-336.
Wong, Francis y Midland, Sharon. 2007. Sensitivity distributions of California populations of
Colletotrichum cereale to the DMI fungicides propiconazole, myclobutanil, tebuconazole, and
triadimefon. En: Plant Dis. Vol. 91; p.1547-1555.
Wright, S, 1978. Evolution and the genetics of populations, variability within and among natural
populations, Vol 4. University of Chicago Press. Chicago.
Yamashiro, T. 1991. Principais doencas fúngicas e bacterianas no maracujazeiro, encontradas
no Brasil. In: Sao José, A., Ferreira, F., Vaz, R. (Eds.) A cultura do Maracuja no Brasil, FUNEP,
Jaboticabal, pp. 169 – 174.
Yap I.V & R.J. Nelson. 1996. WinBoot: A program for performing bootstrap analysis of binary
data to determine the confidence limits of UPGMA-based dendrograms.

116
Yeh, F.C., R.C. Yang., T. Boyle. 1999. Microsoft Window-based Freeware for Population
Genetic Analysis (POPGENE), Ver. 1.32.
Zárate, Rubén D. 2007. Estudios Etiológicos de las principales enfermedades del Valle del
Cauca. Palmira: Facultad de Ciencias Agropecuarias, 56 pp.
Ziogas, B. N.; C. Brian; B. C. Baldwin; J. E. Young 1997. Alternative Respiration: A
Biochemical Mechanism of Resistance to Azoxystrobin (ICIA 5504) in Septoria tritic, Pestic.
Sci. 50:28-44.
Top Related