







Idiomas
Páginas
Jurídico


IMMUNOLOGIA Apunts 3r de Biotecnologia
2020 UNIVERSITAT AUTÒNOMA DE BARCELONA
Consell d’Estudiants de Biociències

Immunologia 2020
CEBIO 1
Índex Tema 0 – Presentació de l’assignatura ............................................................................... 13
Tema 1 – Introducció a la Immunologia............................................................................... 14
Què és la immunologia? .................................................................................................. 14
Elements bàsics del sistema immunitari. ......................................................................... 14
Immunogenicitat. .......................................................................................................... 14
El Funcionament del sistema immunitari. ........................................................................ 16
Immunitat innata o Immunitat adaptativa. ..................................................................... 16
Generalitat i Especificitat de la resposta immunitària. .................................................. 16
Velocitat de la Resposta immunitària. .......................................................................... 17
Memòria de la Resposta immunitària. .......................................................................... 18
Característiques de la resposta adaptativa. ................................................................. 18
Tema 2 – Immunitat Innata, Elements moleculars. .............................................................. 20
La Immunitat Innata ......................................................................................................... 20
Etapes de la Immunitat innata. ..................................................................................... 20
Inflamació. ................................................................................................................... 25
Tema 3 – El sistema del Complement ................................................................................. 26
Receptors de Patrons moleculars Solubles. .................................................................... 26
Nomenclatura del sistema del Complement..................................................................... 26
Vies d’activació del Complement. .................................................................................... 26
Via clàssica .................................................................................................................. 26
Via de les lectines ........................................................................................................ 27
Via alternativa. ............................................................................................................. 27
C5 convertasa: la convergència de les tres vies d’activació. ............................................ 27
Complex d’atac a la membrana. ................................................................................... 27
Funcions del sistema del complement. ............................................................................ 28
Inducció de la resposta inflamatòria. ............................................................................ 28
Opsonització i Fagocitosi de bacteris. .......................................................................... 28
Eliminació d’Immunocomplexos ................................................................................... 28
Mecanismes efectors del Complement ............................................................................ 28
Regulació del complement. ............................................................................................. 29
PRRs solubles. Proteïnes de fase aguda. ....................................................................... 29
Funció del complement. .................................................................................................. 30
Tema 4 - Cèl·lules de la immunitat innata, Part I. ................................................................ 31
Mastòcits. ........................................................................................................................ 31
Efectes biològics de la desgranulació. ......................................................................... 31
Basòfils. ........................................................................................................................... 32

Immunologia 2020
CEBIO 2
Eosinòfils. ........................................................................................................................ 32
Neutròfils. ........................................................................................................................ 33
Mecanismes de matar. ................................................................................................. 33
Macròfags. ...................................................................................................................... 34
Fagocitosi i destrucció del patogen. ................................................................................. 34
Esclat respiratori. ............................................................................................................. 34
Activació dels macròfags. ................................................................................................ 35
Funcions efectores dels macròfags activats ................................................................. 35
Tema 5 - Cèl·lules de la immunitat innata. Part II ................................................................ 36
Cèl·lules NK. ................................................................................................................... 36
Receptors activadors i inhibidors de les cèl·lules NK. .................................................. 36
Mecanisme d’acció de les cèl·lules NK. ....................................................................... 37
Citotoxicitat cel·lular depenent d’anticòs. ..................................................................... 38
Innate Lymphoid Cells. .................................................................................................... 38
Cèl·lules dendrítiques. ..................................................................................................... 38
Diferents TLRs de les cèl·lules dendrítiques. ............................................................... 39
De la resposta innata a l’adaptativa. ................................................................................ 39
Tema 6 – Estructura de les immunoglobulines. ................................................................... 40
BCR o Anticòs. ................................................................................................................ 40
Estructura de les immunoglobulines. ............................................................................... 40
Fragments d’Immunoglobulines sortints de la digestió amb proteases. ........................... 40
Regió o domini variable de les immunoglobulines. .......................................................... 41
Interacció antigen-anticòs. ........................................................................................... 41
Regió constant de la cadena pesada de les immunoglobulines. ...................................... 42
Subclasses de IgG i la seva flexibilitat. ............................................................................ 42
Flexibilitat i moviment de les immunoglobulines. ......................................................... 43
Isotips de les immunoglobulines: monòmers o multímers. ........................................... 43
Determinants que defineixen les diferències entre les Igs. ........................................... 43
Funció efectora de les immunoglobulines. ....................................................................... 43
Funció efectora de les immunoglobulines depenent de FcR. ....................................... 44
Tema 7 - Immunoglobulines. Reordenament gènic ............................................................. 45
Generació immunoglobulines .......................................................................................... 45
Gens de les immunoglobulines i la seva localització cromosòmica. ................................. 45
Organització dels gens de les cadenes pesades de les immunoglobulines. ................. 45
Organització dels gens de les cadenes lleugeres de les immunoglobulines. ................ 45
Reordenament gènic. ................................................................................................... 45
Tema 8 - Limfòcits B. .......................................................................................................... 48

Immunologia 2020
CEBIO 3
Desenvolupament limfòcits B. ......................................................................................... 48
Desenvolupament dels limfòcits B a la medul·la òssia. ................................................ 48
Etapes del desenvolupament dels Limfòcits B. ............................................................ 48
Ontogènesi i maduració a la medul·la òssia. ................................................................ 48
Subpoblacions de limfòcits B. .......................................................................................... 49
Receptor del limfòcit B. .................................................................................................... 49
Immunoglobulines dels BCRs. ..................................................................................... 49
Tema 9 - Receptor d’antigen del limfòcit T .......................................................................... 51
Immunoglobulina ............................................................................................................. 51
Estructura del Receptor d’antigen del limfòcit T. .............................................................. 51
Organització dels gens del Receptor d’antigen del limfòcit T. .......................................... 51
Mecanisme de reordenament dels gens del Receptor d’antigen del limfòcit T. ............ 51
Organització gens del Receptor d’antigen del limfòcit T γδ. ......................................... 52
Generació diversitat del Receptor d’antigen del limfòcit T. ........................................... 52
Epítops reconeguts per limfòcits T. .............................................................................. 52
El TCR està associat amb el complex senyalitzador CD3. ........................................... 53
Reconeixement d’antigen per part del TCR. ................................................................. 53
Tema 10 - Limfòcits T. ........................................................................................................ 54
Origen dels limfòcits T. .................................................................................................... 54
Estructura del timus. .................................................................................................... 54
Desenvolupament dels limfòcits T. .................................................................................. 54
Timòcits DN. ................................................................................................................ 55
Diferenciació a timòcits DP. ......................................................................................... 55
Diferenciació dels limfòcits T. ....................................................................................... 55
Desenvolupament limfòcits T reguladors...................................................................... 56
Maduració dels limfòcits T. ........................................................................................... 56
Limfòcits Tγδ. .................................................................................................................. 56
Limfòcits NKT. ................................................................................................................. 56
Altres poblacions de limfòcits T. ...................................................................................... 56
Tema 11 – Complex Principal d’Histocompatibilitat. ............................................................ 58
Presentació i reconeixement d’antigen. ........................................................................... 58
Molècules presentadores d’antigen. ................................................................................ 58
Estructura del Complex Principal d’Histocompatibilitat de classe I. .............................. 58
Estructura del Complex Principal d’Histocompatibilitat de classe II. ............................. 58
Interacció Complex Principal d’Histocompatibilitat – Pèptid. ........................................ 59
Lloc d’unió a l’antigen................................................................................................... 59
Molècules del Complex Principal d’Histocompatibilitat clàssiques i no clàssiques. ....... 59

Immunologia 2020
CEBIO 4
Organització dels gens de les molècules MHC humanes. ............................................ 59
Codominància del Complex Principal d’Histocompatibilitat. ............................................. 60
Genotip, Fenotip, Haplotip i Desequilibri de lligament del MHC. ...................................... 60
Polimorfisme del Complex Principal d’Histocompatibilitat. ............................................... 61
Residus polimòrfics a les molècules MHC de classe I i classe II. ................................. 61
Avantatge evolutiu dels polimorfismes del Complex Principal d’Histocompatibilitat. ..... 61
Molècules del Complex Principal d’Histocompatibilitat clàssiques i no clàssiques ........... 62
Expressió de molècules HLA clàssiques. ..................................................................... 62
Molècules no clàssiques del Complex Principal d’Histocompatibilitat. .......................... 62
Co-reconeixement de pèptid i Complex Principal d’Histocompatibilitat. ........................... 63
Efecte del Complex Principal d’Histocompatibilitat en el transplantament. ....................... 63
Efecte de les molècules HLA i la susceptibilitat a malalties. ............................................ 64
Altres aspectes de les molècules HLA. ............................................................................ 64
Temes 12 i 13 – Processament i presentació d’antigen. ..................................................... 65
Vies de presentació d’antigen. ......................................................................................... 65
Formes de presentació dels patògens. ............................................................................ 65
Processament i Presentació d’antigen per molècules del MHC de classe I. .................... 66
Processament i Presentació d’antigen per molècules del MHC de classe II. ................... 67
Vies alternatives de presentació d’antigen. ...................................................................... 68
Via de la Crospresentació. ........................................................................................... 68
Via de la Autofàgia. ...................................................................................................... 69
Via de les CD1. ............................................................................................................ 70
Interacció HLA-Pèptid. ..................................................................................................... 70
Pèptids presentats per molècules del MHC. ................................................................. 70
Tema 14 – Cèl·lules Presentadores d’Antigen. ................................................................... 72
Funcions de les cèl·lules Presentadores d’Antigen. ......................................................... 72
Tipus de Cèl·lules Presentadores d’antigen. ................................................................... 73
Cèl·lules Dendrítiques. ................................................................................................. 73
Els limfòcits B com a cèl·lules presentadores d’antigen. .............................................. 75
Cèl·lules presentadores d’antigen professionals. ............................................................. 76
El fenomen de la CrosPresentació. ................................................................................. 76
Tema 15 – Citocines i Quimiocines ..................................................................................... 77
Citocines ......................................................................................................................... 77
Funcions de les citocines. ............................................................................................ 77
Classes de Citocines.................................................................................................... 78
Quimiocines..................................................................................................................... 82
Tema 16 – Anatomia Funcional del sistema Immunitari ...................................................... 84

Immunologia 2020
CEBIO 5
Els Òrgans limfoides. ....................................................................................................... 84
Els òrgans limfoides primaris. .......................................................................................... 84
La medul·la òssia. ........................................................................................................ 84
El timus. ....................................................................................................................... 84
Òrgans limfoides secundaris. .......................................................................................... 85
Vasos limfàtics. ............................................................................................................ 85
Limfonodes. ................................................................................................................. 85
La Melsa. ..................................................................................................................... 87
Sistema immunitari associat a les mucoses. ................................................................ 87
Tema 17 – Recirculació Limfocitària ................................................................................... 89
Molècules d’Adhesió Cel·lular. ........................................................................................ 89
Circulació dels Limfòcits. ................................................................................................. 89
Molècules de la recirculació limfocitària. .......................................................................... 90
Les Selectines i les Sialomucines. ............................................................................... 90
Les Quimiocines. ......................................................................................................... 90
Les integrines. ............................................................................................................. 91
Les adhesines. ............................................................................................................. 91
Extravassació dels Neutròfils i dels Monòcits. ................................................................. 91
Recirculació als Òrgans Limfàtics Secundaris. ................................................................ 92
Diferències entre Limfòcits T efectors i limfòcits T naive. ................................................. 93
Extravasació dels limfòcits T efectors i memòria. ............................................................ 93
Extravasació a la pell ................................................................................................... 93
Paper de les molècules d’adhesió en la presentació d’antigen. ....................................... 93
Tema 18 – Resposta cel·lular. Activació de les cèl·lules T. ................................................. 95
Presentació d’Antigen. ..................................................................................................... 95
Etapes de la resposta cel·lular T. .................................................................................... 95
Activació del limfòcit T. ................................................................................................. 95
Interleucina 2. .............................................................................................................. 97
Expressió de Gens per l’activació. ............................................................................... 98
Presentació d’antigen i reconeixement. ........................................................................... 98
Tema 19 – Resposta cel·lular T. Mecanismes Efectors. .................................................... 100
Diferenciació a limfòcits T CD4+ efectors. ..................................................................... 100
Canvis epigenètics i citocines. ................................................................................... 100
Diferenciació de limfòcits TCD4 helper efectors. ........................................................ 101
Mecanisme efector dels limfòcits T helper. .................................................................... 103
Diferenciació de limfòcitsTCD8 efectors. ....................................................................... 103
Dendritic cell Licensing. ................................................................................................. 104

Immunologia 2020
CEBIO 6
Mecanisme efector de la citotoxicitat dels limfòcits TCD8. ............................................. 104
Limfòcits T citotòxics. .................................................................................................... 105
Limfòcits T de memòria. ............................................................................................. 105
Cèl·lules NKT. ............................................................................................................... 106
Tema 20 – Activació de la Resposta Humoral. .................................................................. 107
Subpoblacions de limfòcits B. ........................................................................................ 107
Activació dels limfòcits B. .............................................................................................. 107
1er senyal d’activació. ................................................................................................ 107
2n senyal d’activació. ................................................................................................. 108
Resposta dels limfòcits B a Antígens T-dependents i a Antígens T-independents. .... 109
Tema 21 – Mecanismes efectors de la resposta humoral. ................................................ 110
Arribada de l’antigen als fol·licles. ................................................................................. 110
Respostes humorals T-dependents. .............................................................................. 110
Cèl·lules T helper fol·liculars. ........................................................................................ 111
Centres germinatius. ..................................................................................................... 112
Cèl·lules Fol·liculars Dendrítiques. ................................................................................ 113
Limfòcits B al centre germinal. ....................................................................................... 113
Limfòcits B de memòria. ................................................................................................ 114
Tema 22 – Regulació de la resposta immunològica. ......................................................... 116
Tolerància Immunològica. .............................................................................................. 116
Mecanismes de la Tolerància Immunològica Perifèrica. ................................................ 116
Regulació per inducció d’anèrgia. .............................................................................. 117
Regulació per inducció de mort per apoptosi. ............................................................ 118
Regulació per cèl·lules T reguladores. ....................................................................... 119
Tolerància Perifèrica als Limfòcits B. ......................................................................... 120
Regulació de la resposta un cop s’ha iniciat. ................................................................. 120
Modulació Neuroendocrina de la resposta. ................................................................ 121
Tema 23 – Resposta a bacteris. ....................................................................................... 122
Microbiota. ..................................................................................................................... 122
Mecanismes d’acció de la microbiota sapròfita sobre el Sistema Immunitari. ............. 122
Funcions de la microbiota sapròfita. ........................................................................... 123
Entrada dels patògens. .................................................................................................. 123
Bacteris Extracel·lulars. ................................................................................................. 124
Resposta innata davant de Bacteris Extracel·lulars. .................................................. 125
Resposta adaptativa humoral davant de Bacteris Extracel·lulars. .............................. 126
Resposta adaptativa cel·lular davant de Bacteris Extracel·lulars. .............................. 126
Conseqüències patològiques de la resposta a Bacteris Extracel·lulars. ..................... 127

Immunologia 2020
CEBIO 7
Mecanismes d’evasió dels bacteris extracel·lulars. .................................................... 128
Resposta a Bacteris Intracel·lulars. ............................................................................... 128
Resposta innata a bacteris intracel·lulars. .................................................................. 129
Resposta adaptativa a bacteris intracel·lulars per limfòcits TCD4 i CD8. ................... 129
Resposta adaptativa a bacteris intracel·lulars per limfòcits Th17. .............................. 130
Conseqüències Patològiques de la resposta a bacteris intracel·lulars. .......................... 130
Mecanismes d’evasió dels bacteris intracel·lulars. ........................................................ 130
Evitar l’activitat microbicida com a mecanisme d’evasió dels bacteris intracel·lulars. . 131
Tema 24 – Resposta a Virus. ............................................................................................ 132
Els virus. ........................................................................................................................ 132
Rutes d’entrada i infecció de virus humans. .................................................................. 132
Tropisme cel·lular. ..................................................................................................... 133
Cicle vital dels virus. ...................................................................................................... 134
Detecció dels virus. ....................................................................................................... 134
Receptors de Tipus Toll. ............................................................................................ 135
Receptors de Tipus R. ............................................................................................... 135
Receptors NOD. ......................................................................................................... 136
Cèl·lules Dendrítiques Plasmacitoides. .......................................................................... 136
Resposta Innata. ........................................................................................................... 137
Interferons. ................................................................................................................. 137
Cèl·lules Dendrítiques Plasmacitoides. ...................................................................... 137
Fases de la infecció vírica i la resposta immunològica. .............................................. 138
Citotoxicitat per cèl·lules NK. ..................................................................................... 138
Macròfags i sistema del complement. ........................................................................ 139
Resposta adaptativa humoral. ....................................................................................... 140
Resposta adaptativa cel·lular. ....................................................................................... 140
Limfòcits TCD4 i Th1. .................................................................................................... 141
Resposta immunològica als virus. ................................................................................. 141
Mecanismes d’evasió. ................................................................................................... 141
Evitar la senyalització per mitjà dels PRR. ................................................................. 141
Inhibició de la presentació per molècules del MHC de classe I. ................................. 142
Variació antigènica. .................................................................................................... 143
Inhibició de la sinapsi immunològica. ......................................................................... 143
Altres mecanismes. .................................................................................................... 143
Conseqüències patològiques de la resposta a la infecció vírica..................................... 144
Tema 16 – Anatomia Funcional del sistema Immunitari .................................................... 145
Els Òrgans limfoides. ..................................................................................................... 145

Immunologia 2020
CEBIO 8
Els òrgans limfoides primaris. ........................................................................................ 145
La medul·la òssia. ...................................................................................................... 145
El timus. ..................................................................................................................... 146
Òrgans limfoides secundaris. ........................................................................................ 146
Vasos limfàtics. .......................................................................................................... 147
Limfonodes. ............................................................................................................... 147
La Melsa. ................................................................................................................... 148
Sistema immunitari associat a les mucoses. .............................................................. 149
Tema 17 – Recirculació Limfocitària ................................................................................. 151
Molècules d’Adhesió Cel·lular. ...................................................................................... 151
Circulació dels Limfòcits. ............................................................................................... 151
Molècules de la recirculació limfocitària. ........................................................................ 152
Les Selectines i les Sialomucines. ............................................................................. 152
Les Quimiocines. ....................................................................................................... 152
Les integrines. ........................................................................................................... 153
Les adhesines. ........................................................................................................... 153
Extravassació dels Neutròfils i dels Monòcits. ............................................................... 153
Recirculació als Òrgans Limfàtics Secundaris. .............................................................. 154
Diferències entre Limfòcits T efectors i limfòcits T naive. ............................................... 155
Extravasació dels limfòcits T efectors i memòria. .......................................................... 155
Extravasació a la pell ................................................................................................. 155
Paper de les molècules d’adhesió en la presentació d’antigen. ..................................... 155
Tema 18 – Resposta cel·lular. Activació de les cèl·lules T. ............................................... 157
Presentació d’Antigen. ................................................................................................... 157
Etapes de la resposta cel·lular T. .................................................................................. 157
Activació del limfòcit T. ............................................................................................... 157
Interleucina 2. ............................................................................................................ 159
Expressió de Gens per l’activació. ............................................................................. 160
Presentació d’antigen i reconeixement. ......................................................................... 160
Tema 19 – Resposta cel·lular T. Mecanismes Efectors. .................................................... 162
Diferenciació a limfòcits T CD4+ efectors. ..................................................................... 162
Canvis epigenètics i citocines. ................................................................................... 162
Diferenciació de limfòcits TCD4 helper efectors. ........................................................ 163
Mecanisme efector dels limfòcits T helper. .................................................................... 165
Diferenciació de limfòcitsTCD8 efectors. ....................................................................... 165
Dendritic cell Licensing. ................................................................................................. 166
Mecanisme efector de la citotoxicitat dels limfòcits TCD8. ............................................. 166

Immunologia 2020
CEBIO 9
Limfòcits T citotòxics. .................................................................................................... 167
Limfòcits T de memòria. ............................................................................................. 167
Cèl·lules NKT. ............................................................................................................... 168
Tema 20 – Activació de la Resposta Humoral. .................................................................. 169
Subpoblacions de limfòcits B. ........................................................................................ 169
Activació dels limfòcits B. .............................................................................................. 169
1er senyal d’activació. ................................................................................................ 169
2n senyal d’activació. ................................................................................................. 170
Resposta dels limfòcits B a Antígens T-dependents i a Antígens T-independents. .... 171
Tema 21 – Mecanismes efectors de la resposta humoral. ................................................ 172
Arribada de l’antigen als fol·licles. ................................................................................. 172
Respostes humorals T-dependents. .............................................................................. 172
Cèl·lules T helper fol·liculars. ........................................................................................ 173
Centres germinatius. ..................................................................................................... 174
Cèl·lules Fol·liculars Dendrítiques. ................................................................................ 175
Limfòcits B al centre germinal. ....................................................................................... 175
Limfòcits B de memòria. ................................................................................................ 176
Tema 22 – Regulació de la resposta immunològica. ......................................................... 178
Tolerància Immunològica. .............................................................................................. 178
Mecanismes de la Tolerància Immunològica Perifèrica. ................................................ 179
Regulació per inducció d’anèrgia. .............................................................................. 179
Regulació per inducció de mort per apoptosi. ............................................................ 180
Regulació per cèl·lules T reguladores. ....................................................................... 182
Tolerància Perifèrica als Limfòcits B. ......................................................................... 184
Regulació de la resposta un cop s’ha iniciat. ................................................................. 184
Modulació Neuroendocrina de la resposta. ................................................................ 184
Tema 23 – Resposta a bacteris. ....................................................................................... 186
Microbiota. ..................................................................................................................... 186
Mecanismes d’acció de la microbiota sapròfita sobre el Sistema Immunitari. ............. 186
Funcions de la microbiota sapròfita. ........................................................................... 187
Entrada dels patògens. .................................................................................................. 188
Bacteris Extracel·lulars. ................................................................................................. 189
Resposta innata davant de Bacteris Extracel·lulars. .................................................. 189
Resposta adaptativa humoral davant de Bacteris Extracel·lulars. .............................. 191
Resposta adaptativa cel·lular davant de Bacteris Extracel·lulars. .............................. 192
Conseqüències patològiques de la resposta a Bacteris Extracel·lulars. ..................... 193
Mecanismes d’evasió dels bacteris extracel·lulars. .................................................... 194

Immunologia 2020
CEBIO 10
Resposta a Bacteris Intracel·lulars. ............................................................................... 195
Resposta innata a bacteris intracel·lulars. .................................................................. 195
Resposta adaptativa a bacteris intracel·lulars per limfòcits TCD4 i CD8. ................... 196
Resposta adaptativa a bacteris intracel·lulars per limfòcits Th17. .............................. 196
Conseqüències Patològiques de la resposta a bacteris intracel·lulars. .......................... 197
Mecanismes d’evasió dels bacteris intracel·lulars. ........................................................ 197
Evitar l’activitat microbicida com a mecanisme d’evasió dels bacteris intracel·lulars. . 198
Tema 24 – Resposta a Virus. ............................................................................................ 199
Els virus. ........................................................................................................................ 199
Rutes d’entrada i infecció de virus humans. .................................................................. 199
Tropisme cel·lular. ..................................................................................................... 200
Cicle vital dels virus. ...................................................................................................... 201
Detecció dels virus. ....................................................................................................... 202
Receptors de Tipus Toll. ............................................................................................ 202
Receptors de Tipus R. ............................................................................................... 202
Receptors NOD. ......................................................................................................... 203
Cèl·lules Dendrítiques Plasmacitoides. .......................................................................... 203
Resposta Innata. ........................................................................................................... 204
Interferons. ................................................................................................................. 204
Cèl·lules Dendrítiques Plasmacitoides. ...................................................................... 205
Fases de la infecció vírica i la resposta immunològica. .............................................. 206
Citotoxicitat per cèl·lules NK. ..................................................................................... 206
Macròfags i sistema del complement. ........................................................................ 207
Resposta adaptativa humoral. ....................................................................................... 207
Resposta adaptativa cel·lular. ....................................................................................... 208
Limfòcits TCD4 i Th1. .................................................................................................... 209
Resposta immunològica als virus. ................................................................................. 209
Mecanismes d’evasió. ................................................................................................... 209
Evitar la senyalització per mitjà dels PRR. ................................................................. 210
Inhibició de la presentació per molècules del MHC de classe I. ................................. 210
Variació antigènica. .................................................................................................... 211
Inhibició de la sinapsi immunològica. ......................................................................... 212
Altres mecanismes. .................................................................................................... 212
Conseqüències patològiques de la resposta a la infecció vírica..................................... 212
Tema 25 – Resposta a paràsits i fongs ............................................................................. 214
Paràsits: Protozous i helmints........................................................................................ 214
Resposta a protozous. ................................................................................................... 214

Immunologia 2020
CEBIO 11
Tripanosoma cruzi. .................................................................................................... 214
Resposta innata a protozous. ..................................................................................... 214
Resposta adaptativa a protozous. .............................................................................. 215
Resposta a Plasmòdium. ........................................................................................... 215
Resposta a Leishmània. ............................................................................................. 217
Mecanismes d’evasió dels Protozous. ....................................................................... 218
Conseqüències patològiques. .................................................................................... 219
Resposta a helmints. ..................................................................................................... 219
Resposta innata a helmints. ....................................................................................... 220
Resposta adaptativa a helmints. ................................................................................ 221
Mecanismes d’evasió del sistema immunitari. ............................................................ 221
Conseqüències patològiques de la infecció per helmints. .......................................... 222
Resposta a Fongs. ........................................................................................................ 222
Candida albicans. ...................................................................................................... 222
Resposta innata a fongs. ........................................................................................... 222
Resposta adaptativa a fongs. ..................................................................................... 223
Mecanismes d’evasió dels fongs. ............................................................................... 224
Tema 26 – Reaccions d’Hipersensibilitat........................................................................... 225
Tipus d’hipersensibilitat. ................................................................................................ 225
Hipersensibilitat de tipus I .............................................................................................. 226
Atòpia. ....................................................................................................................... 227
Procés previ a la reacció d’hipersensibilitat de tipus I................................................. 227
Anticossos IgE. .......................................................................................................... 228
Reacció al·lèrgica. ..................................................................................................... 229
Conseqüències de la resposta. .................................................................................. 230
Al·lèrgia alimentària. .................................................................................................. 231
Tractament de les Hipersensibilitats de tipus 1. ......................................................... 232
Raons ambientals. ..................................................................................................... 232
Estil de vida. .............................................................................................................. 233
Hipersensibilitat de tipus II. ............................................................................................ 233
Anèmia hemolítica. .................................................................................................... 234
Eritroblastosi Fetal. .................................................................................................... 234
Hipersensibilitat de Tipus III. .......................................................................................... 234
Immunocomplexes. .................................................................................................... 234
Mecanisme patogènic. ............................................................................................... 235
Reacció d’arthus. ....................................................................................................... 235
Malaltia del sèrum. ..................................................................................................... 236

Immunologia 2020
CEBIO 12
Hipersensibilitat de tipus IV. .......................................................................................... 236
Sensibilització. ........................................................................................................... 236
Fase Efectora. ........................................................................................................... 236
Exemples. .................................................................................................................. 237
Taula resum. ................................................................................................................. 237
Tema 27 – Autoimmunitat ................................................................................................. 238
Autoimmunitat i malaltia autoimmunitària ...................................................................... 238
Trencament de la tolerància. ......................................................................................... 239
Factors genètics. ........................................................................................................ 239
Factors sistèmics o ambientals. ................................................................................. 241
Alliberació d’antígens segrestats. ............................................................................... 242
Mimetisme molecular o crosreactivitat. ...................................................................... 243
Expressió aberrant del MHC de classe II i de molècules coestimuladores. ................ 243
Classificació de les malalties autoimmunitàries. ............................................................ 244
Malaltia de Graves. .................................................................................................... 244
Miastènia gravis. ........................................................................................................ 245
Síndrome de Goodpasture. ........................................................................................ 245
Lupus eritematós sistèmic. ......................................................................................... 246
Artritis reumatoide. ..................................................................................................... 246
Diabetis tipus I. .......................................................................................................... 247
Esclerosi múltiple. ...................................................................................................... 248
Celiaquia. ................................................................................................................... 249
Tema 28 – Immunodeficiències ........................................................................................ 250
Classificació de les immunodeficiències. ....................................................................... 250
Immunodeficiències Primàries. ...................................................................................... 250
Símptomes de les Immunodeficiències Primàries. ..................................................... 251
Severitat de les Immunodeficiències Primàries. ......................................................... 251
Classificació de les Immunodeficiències Primàries. ................................................... 252
Immunodeficiències Secundàries o adquirides. ............................................................. 256

Immunologia 2020
CEBIO 13
Tema 0 – Presentació de l’assignatura Què és la immunologia? Què és aquesta paraula estranya que ens defineix una assignatura
del grau en Biotecnologia? I, encara més important, per què necessitem saber-ne?
La immunologia és la ciència que estudia els sistemes de defensa de l’organisme
davant d’agents estranys. Per aconseguir-ho, el nostre cos ha de protegir-nos no només de
l’entrada dels múltiples microbis que hi ha al nostre voltant, sinó que també ho ha de fer dels
seus atacs un cop ja han entrat. El sistema de resistència a les infeccions està constituït per
molècules i factors solubles produïts per cèl·lules que s’estructuren en els òrgans limfoides.
I doncs? Què hi tenen a veure les cèl·lules, les molècules i les reaccions que realitza el meu
cos quan detecta alguna cosa estranya amb els meus estudis en Biotecnologia?
Més del que ens pensem, ja que la immunologia posa en context els coneixements previs de
Microbiologia, Bioquímica i Fisiologia alhora que ajuda a integrar processos biològics que es
donen en organismes complets enlloc d’una de les seves parts.
Apart, la immunologia necessita molt de la Biotecnologia. La necessita per erradicar
infeccions mitjançant la creació de vacunes, la requereix per dissenyar teràpies efectives
contra el càncer o el rebuig als transplantaments i li és essencial per millorar la salut animal
(dins la qual hi trobem la salut humana).

Immunologia 2020
CEBIO 14
Tema 1 – Introducció a la Immunologia
Què és la immunologia? La Immunologia és la ciència que estudia els sistemes de defensa de l’organisme
davant agents estranys. Els agents estranys tenen vàries definicions. Una d’elles és la de
la teoria de la selecció clonal de Burnet:
“Les substàncies no-pròpies (o agents estranys) són totes aquelles
substàncies que NO entren en contacte amb els limfòcits durant el període
embrionari.”
Aquesta concepció s’ha demostrat clarament falsa ja que, per exemple, la mare li transmet
anticossos propis al nadó, i aquests no entren en contacte amb els limfòcits durant el període
embrionari.
Arran d’una manca de definició més exacta, definim tres agents estranys: Substàncies
exògenes, Substàncies al·logèniques i Substàncies alliberades. Les primeres són
patògens o els seus productes que poden causar dany. En les al·logèniques trobem els grups
sanguinis i el MHC. En les substàncies endògenes hi trobem components alliberats, cèl·lules
malmenades i cèl·lules transformades que canvien estructures moleculars.
Elements bàsics del sistema immunitari. El terme Immunitat és un derivat de la paraula llatina immunitas, la qual es referia a la
protecció de la persecució legal dels senadors romans durant la seva estada en ofici.
Històricament, la immunitat significa “protecció de la malaltia i de les malalties infeccioses.”
Les cèl·lules i molècules responsables de la immunitat constitueixen el sistema immunitari i
la seva resposta coordinada i col·lectiva a la presència d’agents estranys s’anomena
resposta immunitària.
La funció fisiològica del sistema immunitari és la defensa contra els microbis
infecciosos. Així i tot, fins i tot substàncies externes no infeccioses i productes de
cèl·lules malferides poden provocar una resposta immune. Encara més, els mecanismes
que normalment serveixen per protegir individus de la infecció i eliminen substàncies externes
també són capaços de provocar danys als teixits en algunes situacions. Conseqüentment,
una millor definició de la resposta immunitària és que és “una reacció a microbis i a molècules
reconegudes com estranyes, independentment de la conseqüència fisiològica o patològica
d’aquesta reacció.” Sota algunes condicions, fins i tot les pròpies cèl·lules poden provocar
respostes immunitàries (precisament anomenades resposta autoimmune).
Aquestes molècules que interaccionen amb els mecanismes de defensa del sistema
immunitari s’anomenen antígens, tot i que reben el nom d’immunògens si indueixen una
resposta del sistema immunitari.
Immunogenicitat.
Estructura i Naturalesa Química.
Els antígens poden ser de diferents tipus, cadascun amb una immunogenicitat ben diferent
segons els punts d’interacció i la qualitat de la mateixa amb el sistema immunitari.

Immunologia 2020
CEBIO 15
Mida i Estabilitat.
La immunogenicitat d’una molècula es troba relacionada no només amb els seus components
químics, que també, sinó que contempla la mida i l’estabilitat de la molècula antigènica.
D’aquesta manera, les molècules de major mida solen generar una immunogenicitat més
potent que no pas les molècules petites, que quasi no en causen. De manera inversa passa
amb l’estabilitat, ja que les molècules més estables són més difícils de degradar que les
molècules menys estables. Això en dificulta la presentació a les cèl·lules que han de
reconèixer l’antigen i provoca una immunogenicitat més dèbil.
Així i tot, la resposta del sistema immunitari als diferents antígens ve determinat per una sèrie
de factors que en determinen el funcionament. Entre aquests hi trobem:
• Les proteïnes són un tipus d'antigen que presenta diferents epítops, cadascun amb unaespecificitat diferent. Això atorga molta immunogenecitat a aquest tipus de molècules.
• En són una excepció les proteïnes homopolimèriques, ja que es redueix la quantitatd'interaccions diferents que es poden donar.
Proteïnes
• Els polisacàrids són uns antígens poc immunogenicistes, ja que tenen diferents epítops peròtots ells amb la mateixa especifitat.
• En són un excepció les glicoproteïnes, que presenten moltíssima immunogenicitat gràcies atenir diferents epítops.
Polisacàrids
• Un hapté és un antigen que no genera una resposta del sistema immunitari per si sol perquèsón tan petits que no poden arribar a interaccionar amb els elements del mateix per acabargenerant una resposta.
• Sovint s'associen a proteïnes transportadores. En aquest cas, el complex dóna lloc a unaresposta immunitària. Aquesta és tant contra l'hapté com contra la proteïna transportadora.
• Són haptens alguns metalls, antibiòtics, productes de síntesi, esteroides...
Haptens
• Degradació deficient
• Presentació deficient
• Baixa Immunogenicitat
Molècules Molt Estables
• Degradació eficient
• Presentació eficient
• Immunogenicitat Potent
Molècules poc
estables
• PM>6000Da
• Molta Immunigenicitat
• Toxina del Tètanus
Molècules grans
• 1000 < PM > 6000Da
• Immunogenicitat dèbil
• Insulina
Molècules Mitjanes
• PM < 1000Da
• No Immunogèniques
• Penicil·lina
Molècules Petites

Immunologia 2020
CEBIO 16
Aquests factors es veuen reflectits en els òrgans efectors del sistema immunitari, uns òrgans
limfoides dispersos connectats per vasos limfàtics. Aquests òrgans poden ser primaris,
com la medul·la òssia i el timus, o secundaris com els limfonodes, la melsa o el MALT.
El Funcionament del sistema immunitari. Dins de cadascun dels òrgans mencionats succeeixen molts esdeveniments per separat, però
molts d’ells responen a les necessitats del sistema immunitari, que quan entra en contacte
amb l’antigen fa servir components pre-formats i genèrics per intentar eliminar-lo i alhora
fabrica components nous que l’eliminen específicament.
Immunitat innata o Immunitat adaptativa.
Aquestes dues accions de resposta que al cap i a la fi es duen a terme entre tots els òrgans
del sistema immunitari responen a les respostes innata i adaptativa. La resposta innata
inicia, condiciona i augmenta la resposta adaptativa, que augmenta i modula la resposta
innata.
La resposta immunitària innata, la primera que interacciona i interactua amb l’antigen, és
una resposta genèrica, ràpida i fixa, capaç de donar-se en hores d’haver entrat l’antigen. Així
i tot, és limitada i no genera memòria, pel que només és eficaç en casos d’infeccions poc
intenses. La resposta immune innata està composta per barreres físiques, químiques i
fagòcits d’entre d’altres; fent-la present a plantes, invertebrats i vertebrats. És per això que
aquesta resposta immune es considera evolutivament anterior a la resposta immunitària
adaptativa, una resposta específica, lenta i eficient que genera memòria de la infecció
mitjançant limfòcits T i limfòcits B així com receptors d’antigen i anticossos.
Generalitat i Especificitat de la resposta immunitària.
Molècules del sistema immunitari.
Per tal que es puguin donar les respostes innata i adaptativa, però, calen els elements en què
es basa el sistema immunitari: les interaccions intermoleculars i intracel·lulars entre
lligands i receptors. D’aquests últims, els tres tipus més importants són els receptors de
Factors Genotípics
•Determinen si un individu respondrà a un agent estrany i com ho farà.
Edat
•Canvia la capacitat de resposta.
•El Sistema Immunitari dels nadons no està ben desenvolupat i el de les persones grans funciona menys bé
Factors Metabòlics i físics
•En formen part els canvis dels nivells hormonals o la connexió entre el sistema immunitari i el sistema nerviós
Les característiques anatòmiques
•Els punts d'infecció responen diferent al mateix estímul
Factors ambientals
•En formen part la nutrició, la microbiota, la contaminació...

Immunologia 2020
CEBIO 17
patrons moleculars (PRRs), unes estructures bioquímiques comunes en diferents
microorganismes, les immunoglobulines, que poden ser solubles o de membrana als
limfòcits B, i els Receptors específics d’antigen dels limfòcits T (TCR).
Resposta Adaptativa.
Especificitat.
La combinació de les activitats i interaccions de les molècules que hem mencionat permeten
generar els dos tipus de respostes immunitàries del que hem parlat: innata i adaptativa. La
adaptativa, malgrat ser més lenta, ofereix una característica molt important que la fa destacar
per sobre de la immunitat innata: l’especificitat, la qual es combina amb la diversitat.
L’especificitat de la resposta adaptativa garanteix que cada antigen indueix una resposta
específica que estimula les respostes més adients i eficients per eliminar-lo. Aquesta resposta
específica, però, es sol veure reflectida en la segona infecció de l’agent estrany.
Diversitat.
La mencionada diversitat de la resposta adaptativa permet al sistema immunitari respondre
a una gran diversitat d’antígens ja que el nombre total de limfòcits que es generen, i que es
denomina repertori de limfòcits T i B, permet reconèixer més de 109 antígens o determinants
antigènics diferents. Cal dir que tant els BCR (immunoglobulines de membrana als limfòcits
B) com els TCR (Receptors específics d’antigen dels limfòcits T) s’originen per mecanismes
moleculars de recombinació gènica complexos.
Velocitat de la Resposta immunitària.
Com ja hem dit, la resposta immunitària innata és el tipus de resposta ràpida i general mentre
que la resposta adaptativa és més lenta a canvi de ser molt més específica. Això succeeix
perquè la resposta immunitària innata es troba sempre present i a punt mentre que la
resposta immunitària adaptativa s’activa per exposició a l’agent estrany, el que
requereix de dies per poder ser més potent que la immunitat innata.
Per poder-hi haver aquestes diferències, cal que les dues respostes facin servir dos tipus de
cèl·lules diferents en el seu funcionament. És per això que la resposta innata és duta a terme
per fagòcits, granulòcits i cèl·lules NK mentre que la resposta adaptativa ho està per Limfòcits
B, cèl·lules plasmàtiques i limfòcits T. Un tipus especial de cèl·lula és la cèl·lula Dendrítica,
ja que forma part de la resposta innata com a fagòcit i de la resposta adaptativa com a cèl·lula
presentadora d’antígens.
Fases de la resposta adaptativa
Hem dit que la resposta adaptativa, la que duen a terme els limfòcits B, les cèl·lules
plasmàtiques i els limfòcits T, requereix de dies per actuar. Per ser el màxim de precisos que
la biologia ens permet hem de dir que la resposta immunitària adaptativa triga entre 4 i 7
dies per adquirir la màxima potència ja que requereix de la proliferació i diferenciació de
les seves cèl·lules. La resposta adaptativa consta de 5 fases:

Immunologia 2020
CEBIO 18
La diferència entre immunitat humoral i cel·lular (o mediada per cèl·lules) a la que es fa
referència en l’esquema es pot resumir en si la molècula efectora és soluble (resposta
humoral) o és una cèl·lula (cel·lular).
També hi ha una altre diferència: la funció que fa cadascuna. Mentre que la immunitat
humoral serveix per bloquejar les infeccions i eliminar els agents extracel·lulars, la
immunitat cel·lular pot tenir dues funcions diferents segons si la realitzen els Limfòcits T
Helper o els Limfòcits T citotòxics.
Els limfòcits T Helper tenen la funció d’activar els macròfags per eliminar els agent
fagocitats i els limfòcits T citotòxics maten les cèl·lules infectades i eliminen els reservoris
de la infecció.
Memòria de la Resposta immunitària.
Hem dit en l’esquema superior (i unes quantes pàgines enrere també) que la immunitat
adaptativa genera memòria, al contrari que la innata. La memòria és una característica del
sistema immunitari que li produeix canvis quan és exposat a un antigen. Aquests canvis
milloren la seva capacitat de respondre al mateix antigen quan hi torna a entrar en contacte.
Els principis de la vacunació es basen en aquesta característica de la resposta adaptativa.
Característiques de la resposta adaptativa.
Si fem un repàs del que s’ha dit fins ara veurem que les dues característiques que distingeixen
més clarament la immunitat innata i l’adaptativa són l’especificitat antigènica i la memòria.
Així i tot, hi ha altres característiques que diferencien encara més la immunitat innata de
l’específica. Aquestes són la diversitat, la clonalitat i l’autoregulació. La diversitat ja s’ha
explicat, no cal repetir-la, però farem una suau indagació sobre la clonalitat i l’autoregulació
de la resposta adaptativa.
Clonalitat de la Resposta Adaptativa.
La clonalitat és un concepte derivat de la paraula clon, de significat similar a còpia. És un
terme aplicable a la immunitat adaptativa perquè quan un limfòcit s’activa, aquest prolifera i
es multiplica, donant lloc a múltiples cèl·lules amb el mateix receptor d’antigen. Com que són
totes iguals, es considera que les cèl·lules que deriven del limfòcit activat són clons entre
elles.
Per explicar la immunitat humoral es va desenvolupar una adaptació de la hipòtesi de Burnet
amb els següents postulats:
Fase de Reconeixe-
ment
•Els limfòcits B i T naive interactuen amb la cèl·lula presentadora d'antigen.
•Comença la proliferació.
Fase d'activació
•Les cèl·lules es diferencien.
•Es generen cèl·lules productores d'anticossos i limfòcits T efectors.
Eliminació d'antigen
•Actuen les immunitats humoral i mediada per cèl·lules
•S'elimina l'antigen que ha provocat la resposta immunològica.
Contracció
•Apoptosi de la majoria de cèl·lules de la immunitat adaptativa.
Memòria
•Supervivència d'algunes de les cèl·lules generades.

Immunologia 2020
CEBIO 19
Autoregulació de la resposta adaptativa
L’autoregulació és una característica que fa que totes les respostes immunològiques
normals disminueixin d’intensitat amb transcurs del temps des de l’estimulació, fent que el
sistema immunitari retorni al seu estat basal i de repòs anomenat homeòstasi.
El sistema immunitari té una gran diversitat
d’anticossos preformats que actuen com a
receptors d’antigen dels limfòcits B
La interacció d’alta afinitat entre una molècula
estranya (antigen) i el receptor d’antigen
desencadena l’activació del limfòcit
Les cèl.lules efectores diferenciades originades a partir d’un limfòcit activat expressaran receptors
d’especificitat idèntica als de la cèl·lula original

Immunologia 2020
CEBIO 20
Tema 2 – Immunitat Innata, Elements moleculars.
La Immunitat Innata El terme Immunitat innata fa referència als mecanismes de defensa que estan sempre
presents i preparats per combatre qualsevol agent extern. És ràpida però, alhora, limitada
i fixa. El sistema immunitari innat està compost per molts tipus de cèl·lules i molècules que
eviten l’entrada i infecció dels microorganismes. Aquesta funció, la d’evitar l’entrada i infecció,
es fa de manera genèrica, és a dir, no específica. Això es deu, en part, a no presentar una
capacitat de memòria, pel que no protegeix davant d’infeccions posteriors.
Així i tot, la gran funció de la immunitat innata és la de facilitar i determinar el
desenvolupament de la immunitat adaptativa.
Etapes de la Immunitat innata.
La resposta immunitària innata té dues fases: Immediata, que es dona durant les primeres 4
hores des de la infecció, i la induïda, que dura des que han passat les primeres 4 hores fins
que n’han passat 96. Això és perquè la resposta adaptativa, l’altre tipus de resposta
immunitària, requereix de dies per generar mecanismes efectors i, per tant, és eficient quan
els mecanismes de la immunitat innata no poden eliminar la infecció.
La immunitat innata immediata reconeix l’antigen de manera inespecífica i l’intenta eliminar,
mentre que la immunitat innata induïda reconeix patrons microbians, inflama el teixit on es
troba l’agent estrany i mira d’eliminar-lo amb les cèl·lules efectores reclutades amb la
inflamació.
Immunitat innata immediata
De les dues fases de la immunitat innata, la immediata és aquella que succeeix en les
primeres 4 hores i proporciona una barrera per impedir la infecció o la seva disseminació.
Aquesta barrera s’aconsegueix mitjançant l’impediment del contacte entre l’agent infecciós
i les cèl·lules epitelials, evitant així la infecció per adhesió i colonització d’aquestes
superfícies.
Barreres físiques o mecàniques de la immunitat innata immediata.
Per evitar el contacte amb les cèl·lules epitelials, la immunitat innata immediata disposa d’una
sèrie de barreres físiques a totes les superfícies mucoses de l’epiteli nasofaringi, oral, del
tracte respiratori, intestinal, urogenital i de la pell. En tots els casos estan les tight junctions,
les unions entre cèl·lules, i a cada teixit hi ha una mesura “extra”. En el cas de la pell i l’intestí,
un mecanisme de defensa mecànica és el flux d’aire o d’un altre fluid de manera longitudinal,
en el cas dels pulmons és el moviment del moc pels cilis i en el cas dels ulls i del nas és la
presència de cilis nasals o de llàgrimes.
Barreres Químiques de la immunitat innata immediata.
Paral·lelament a les barreres físiques, el cos humà disposa, en el seu mecanisme d’immunitat
innata immediata, de substàncies microbicides o que inhibeixen el creixement microbià. Són
les barreres químiques. A la saliva i a les llàgrimes, per exemple, hi ha lisozim i altres
enzims bactericides. A l’estómac i al tracte intestinal superior hi trobem un pH baix
acompanyat d’enzims digestius com la pepsina, sals biliars i àcids grassos. Finalment, a les
cèl·lules epitelials del tracte respiratori hi trobem surfactants.
Apart d’aquests múltiples compostos químics, tant a la pell com a l’intestí com als pulmons hi
trobem una altra substància química que forma part d’aquesta barrera: els pèptids
antimicrobians. Els pèptids antimicrobians són pèptids petits, d’entre 12 i 50 aminoàcids,

Immunologia 2020
CEBIO 21
que s’uneixen a les membranes bacterianes mitjançant interaccions electrostàtiques tot
causant una distorsió de les mateixes. Això els permet entrar i inhibir el metabolisme bacterià.
Hi ha més de 800 pèptids antimicrobians descrits, tot i que majoritàriament es poden
classificar en 4 classes:
Barreres microbiològiques de la immunitat innata immediata.
Finalment, i com a mesura extra d’aquesta fortificació teòricament infranquejable que és la
immunitat innata immediata, ens trobem amb la flora bacteriana sapròfita, altrament
anomenada microbiota. Aquesta es troba associada a la majoria de superfícies epitelials i
està composta per bacteris comensals que competeixen per nutrients i per l’adherència a
aquestes superfícies. Els organismes formants de la microbiota produeixen substàncies
microbicides com, per exemple, l’àcid làctic o bactericines, les quals funcionen com a
pèptids antimicrobians.
Resum de l’efecte barrera de la immunitat innata immediata.
Recapitulant tota la informació que hem recollit fins ara, podem generar el següent mapa
conceptual que defineix tot el bloqueig que ha de superar un agent infecciós per entrar al cos
humà:
Localització de la barrera
Tipus de barrera Pell Intestí Pulmons Ulls i nas
Mecànica Cèl·lules epitelials unides per tight junctions
•Produïdes constitutivament en una forma inactiva.
•Distorsionen la membrana plasmàtica i inhibeixen el creixement bacterià.
•Es troben en les cèl·lules epitelials intestinals i en els neutròfils.
α-Defensines
•Produïdes només com a resposta a la presència de microorganismes.
•Actuen distorsionant la membrana plasmàtica i inhibint el creixement bacterià.
•Es poden trobar a les cèl·lules epitelials dels tractes respiratori i urogenital, de la pell i de la llengua.
β-Defensines
•Produïdes consitutivament.
•Són unes lectines de tipus C que s'uneixen al peptidoglicà de la paret bacteriana i en faciliten la destrucció.
•Es troben a les cèl·lules epitelials intestinals.
Reg3γ
•Produïdes constitutivament en una forma inactiva.
•Són pèptids catiònics amfipàtics que trenquen les membranes i que són tòxics per molts microorganismes.
•Es troben en cèl·lules epitelials intestinals i pulmonars en resposta a la infecció, en queratinòcits i en neutròfils i macròfags.
Catelicidines
Cèl·lules epitelials unides per tight junctions

Immunologia 2020
CEBIO 22
Flux longitudinal Moviment de
mucosa Llàgrimes i cilis
Química Àcids Grassos
Baix pH
Lisoenzim a les llàgrimes Enzims (pepsina)
Pèptids antibiòtics
Microbiològica Flora normal
Immunitat innata induïda.
Detecció de patrons moleculars.
PAMPs, MAMPs i DAMPs.
Si un microorganisme traspassa la barrera epitelial i es comença a replicar en els teixits de
l’hoste, és detectat immediatament per cèl·lules del sistema immunitari residents en aquests
teixits. Aquestes cèl·lules detecten els agents infecciosos mitjançant sistemes de
reconeixement genèrics i no adaptatius que activen molècules que ataquen el
microorganisme i faciliten la seva eliminació.
Per detectar-los fan servir PAMPs: Pathogen-Associated Molecular Patterns. Els PAMPs són
estructures moleculars conservades en diferents patògens que els són essencials per la
supervivència. Solen ser productes del metabolisme cel·lular o components microbians de
la paret cel·lular, com LPS, PGN o LTA. També poden ser elements microbians interns com
DNA o RNA o qualsevol estructura diferent a la de l’hoste. Altres patrons que també poden
detectar les cèl·lules de la immunitat innata són les MAMPs (Microbiota-Associated Molecular
Patterns) i les DAMPs (Danger-Associated Molecular Patterns).
Diferents classes de microorganismes expressen diferents PAMPs. Aquestes estructures són
els àcids nucleics exclusius del microbis, com l’ARN bicatenari que es troba en els virus
que s’estan replicant o les seqüències CpG de DNA no metilades que es troben en bacteris;
les característiques de les proteïnes que es troben en microbis, com la iniciació per N-
formilmetionina típica de les proteïnes bacterianes; i lípids i glúcids complexes que
sintetitzen els microorganismes però no les cèl·lules de mamífer, com el lipopolisacàrid
(LPS), l’àcid lipoteicoic i els oligosacàrids rics en manosa.
En realitat, només hi ha un nombre limitat de diferències fonamentals entre les molècules
microbianes i les que produeixen els organismes superiors. D’aquesta manera, el sistema
immunitari innat ha evolucionat per reconèixer tan sols una quantitat determinada de
molècules, la majoria exclusives dels microbis, mentre que el sistema immunitari adaptatiu
és capaç de reconèixer una sèrie molt més gran de substàncies estranyes, siguin o no dels
microorganismes.
Receptors de patrons moleculars o PRRs.
El sistema immunitari innat utilitza varis tipus de receptors cel·lulars, presents en
diferents localitzacions en les cèl·lules, i molècules solubles a la sang i a les secrecions
mucoses que reconeixen PAMPs i DAMPs. Aquests receptors cel·lulars de
microorganismes patògens i de molècules associades a la lesió s’anomenen Receptors pel
Reconeixement del Patró o PRRs i estan expressats per cèl·lules del sistema immunitari.
Les cèl·lules que els expressen són els fagòcits, les cèl·lules dendrítiques, les cèl·lules
epitelials externes i molts altres tipus de cèl·lules que ocupen els teixits i els òrgans.
Pèptids Antibiòtics
Flux Longitudinal
Flora Bacteriana

Immunologia 2020
CEBIO 23
Els PRRs s’expressen a la membrana plasmàtica o a les membranes endosòmiques de varis
tipus cel·lulars, així com en el citoplasma d’aquestes cèl·lules. Quan aquestes molècules de
reconeixement del patró s’uneixen a PAMPs o DAMPs, activen la transducció de senyals que
promouen les funcions antimicrobiana i pro-inflamatòria de les cèl·lules on s’expressen.
Hi ha 3 tipus de PRRs. Els PRRs de membrana serveixen per dur a terme la fagocitosi i la
senyalització al nucli. Són PRRs de membrana els receptor de tipus Toll (TLRs), els receptors
Scavenger i els receptors de lectina tipus-C. Els PRRs de citoplasma tenen com a única
funció fer una senyalització al nucli. Són PRRs de citoplasma els Nucleotide binding
oligomerization domain (NOD)-like receptors (NLRs) i els Retinoic acid-inducible gene I
(RIGI)-like receptors (RLRs). Finalment, els PRRs solubles són els encarregats de la
opsonització del microorganisme i de l’activació tant del sistema del complement com de les
proteïnes de la Fase Aguda, les quals comprenen la Lectina d’unió a la Manosa (MBL), la
Proteïna C Reactiva (RCP) i la Proteïna Amiloide del Sèrum (SAP).
Toll-Like Receptors o TLRs.
Els receptors de tipus Toll (TLR), una família de receptors pel reconeixement del patró
conservada al llarg de l’evolució que expressen molts tipus cel·lulars, reconeixen productes
d’una àmplia varietat de microorganismes. S’han descrit 11 TLRs diferents en humans. Els
TLRs són glicoproteïnes integrals de membrana que contenen repeticions riques en leucina
en les seves regions extracel·lulars (o intravesiculars), que participen en la unió al lligand, i
un domini TIR en les seves cues citoplasmàtiques que és essencial per la producció dels
senyals.
Els TLRs s’expressen a la membrana plasmàtica o de compartiments citoplasmàtics de
cèl·lules dendrítiques (DC), macròfags, neutròfils, cèl·lules de l’endoteli vascular, adipòcits,
miòcits, cèl·lules epitelials intestinals i d’altres tipus cel·lulars com els limfòcits.
Els TLRs 1, 2, 4, 5 i 6 s’expressen a la membrana plasmàtica, on reconeixen varis PAMPs
en l’ambient extracel·lular. Segons quines combinacions de TLRs es donin es reconeix un
o altre antigen. D’aquesta manera, la unió dels TLR 1 i 2 permeten reconèixer lipopèptids
bacterials, dos TLR2 permeten reconèixer els peptidoglicans bacterials, dos TLR4 permeten
identificar LPS, dos TLR5 permeten interaccionar amb la flagelina Bacterial i un TLR2 unit a
un TLR6 permet detectar lipopèptids bacterials.
Els TLRs 3, 7, 8 i 9, en canvi, s’expressen al reticle endoplasmàtic i a les membranes
endosòmiques de les cèl·lules, on detecten varis lligands d’àcids nucleics. Per ser més
precisos, els dímers de TLR3 reconeixen RNA de doble cadena, els de TLR7 reconeixen RNA
de cadena senzilla, els de TLR8 reconeixen RNA de cadena senzilla i els dímers de TLR9
reconeixen DNA amb motius CpG desmetilats, propi del DNA bacterià.
La unió dels PAMPs als TLRs indueix un canvi conformacional d’aquests que afavoreix
l’oligomerització de TLRs, acostant els dominis TIR i creant llocs d’unió per molècules
adaptadores. Les molècules adaptadores més importants són MyD88 (myeloid differentiation
primary response gene 88), TRIF (TIR-domain-containing adaptor-inducing IFN-β) i TRAM
(TRIF-related adaptor molecule). El resultat de la cascada de senyalització que provoca la
unió de PAMPs a TLRs és la producció de citocines pro-inflamatòries i d’interferons de tipus
1.
Els principals factors de transcripció activats per les vies de transmissió de senyals dels TLR
són el factor nuclear kB, la proteïna d’activació 1, el factor de resposta a l’interferó 3 (IRF3) i
IRF7. NK-kB i AP-1 estimulen l’expressió de gens que codifiquen per moltes de les molècules
que es necessiten per les respostes inflamatòries com les citocines inflamatòries, les

Immunologia 2020
CEBIO 24
quimosines i les molècules d’adhesió endotelials. IRF3 i IRF7 promouen la producció
d’interferons dels tipus 1, importants per les respostes immunitàries innates antivíriques.
Els receptors Scavenger i receptors de lectina tipus C
Els receptors scavenger detecten lipoproteïnes modificades com oxLDL, components
microbians i cèl·lules apoptòtiques. Aquests receptors s’expressen en monòcits, macròfags i
en cèl·lules dendrítiques.
Els receptors de lectines tipus C detecten carbohidrats, motius rics en manosa, fucosa i β-
glucà. Aquests receptors s’expressen en monòcits, macròfags i cèl·lules dendrítiques.
Rig-Like Receptors o RLRs.
Els receptors de tipus Rig són detectors citosòlics del ARN víric que responen induint la
producció d’interferons antivírics del tipus I. Els RLRs millor caracteritzats són RIG-I, MDA5 i
LGP2. Aquestes proteïnes contenen un domini C-terminal que uneix RNA víric. Alguns RLRs
tenen, a més, dos dominis CARD de reclutament i activació de caspasa-1. Quan un RLR
s’uneix amb el seu lligand, inicia la via de producció d’IFNs tipus I i d’altres citocines pro-
inflamatòries. Els RLRs s’expressen en una àmplia varietat de tipus cel·lulars, inclosos els
leucòcits derivats de la medul·la òssia i vàries cèl·lules tissulars.
Receptor tipus NOD o NLRs.
Els receptors de tipus NOD són una família de més de 23 proteïnes citosòliques diferents
algunes de les quals detecten la presència al citoplasma de PAMP i DAMP. Recluten altres
proteïnes per formar complexes transmissors de senyals que promouen la inflamació.
L’estructura d’aquests receptors consisteix en un Domini C terminal (Leucine-rich repeat) que
reconeix el lligand, un Domini NOD (Nucleotide-binding oligodimerization) i un Domini N-
terminal (CARD, PYD, BIR) efector.
Aquests receptors s’expressen en cèl·lules sistema immunitari (limfòcits, cèl·lules
dendrítiques i macròfags) però també a cèl·lules epitelials. Els seus lligands són proteoglicans
bacterians, DNA o RNA microbians i la seva unió dona lloc a l’activació del factor de
transcripció NF-kB, les MAPK o la caspasa-1.
La funció del receptor de tipus NOD ve definida pel seu domini N-Terminal, el qual pot ser un
domini CARD, que provoca l’activació de la caspasa i de l’inflamasoma, un domini Pyrina,
PYD, de transducció de senyal i d’activació de l’inflamasoma, o un domini inhibidor del
baculovirus, BIR, que regula la transcripció.
NOD1 i NOD2, membres de la subfamília NOD que contenen el domini CARD, s’expressen
al citoplasma de varis tipus cel·lulars com les cèl·lules epitelials mucoses o els fagòcits, i
responen a peptidoglicans de la paret bacteriana.
NOD1 reconeix substàncies derivades sobretot de bacteris gramnegatius, mentre que NOD2
reconeix una molècula diferent anomenada dipèptid muramil procedent de microorganismes
gramnegatius i grampositius.
Els inflamasomes, allò que activen els receptors de tipus NOD amb un domini N-terminal de
tipus CARD, són complexes multiproteics formats quan els NLRs amb dominis N-terminal
PYD o CARD reconeixen PAMPs o DAMPs. En aquestes casos, els dominis s'uneixen a ASC
(Apoptosis associated speck-like containing a CARD domain) per interaccions homotípiques
i ASC recluta la procaspasa-1, formant l’inflamasoma. La formació del complex permet
l’activació de la caspasa-1, que interactua directament amb NLRC-4 mitjançant el seu domini
CARD.

Immunologia 2020
CEBIO 25
La caspasa-1 té dues funcions: trenca els precursors de les citocines IL-1β i IL-18, que inicien
la inflamació, i indueix la lisi de la cèl·lula (piroptosi), exposant el patogen.
IL-1β i IL-18 es secreten en forma de precursor inactiu que cal processar per tal que la citocina
sigui activa. IL-1β és una citocina pirogènica que promou l’activació de respostes
inflamatòries en general. IL-18 és important per la producció d’IFNγ. Pro-IL-18 és produïda
constitutivament a macròfags però la producció de pro-IL-1β és regulada pel factor de
transcripció NF-kB. HMGB1, citocina secretada per macròfags i monòcits activats, és un
mediador d’inflamació
Inflamació.
El procés d’inflamació, mediat en part per l’inflamasoma, consisteix en diversos passos.
Primer, el dany tissular provoca l’entrada dels bacteris. Després, aquests bacteris activaran
els macròfags, que alliberaran citosines i quimiosines. Aquests dos components estimulen
un increment local del flux sanguini i de la permeabilitat capil·lar, el que deriva en rubor,
tumor, calor i dolor. Seguidament, els capil·lars permeables permetran un influx de fluid i de
cèl·lules inflamatòries que migraran al teixit, alliberant mediadors inflamatoris que provoquen
dolor. Finalment, els fagòcits migraran al lloc de la inflamació i aquests destruiran els
bacteris.
Destrucció del patogen.
Els patògens poden ser destruïts per macròfags o per neutròfils. Els neutròfils són cèl·lules
de vida curta, circulants i absents als teixits en condicions de salut. Respecte els macròfags,
en canvi, n’hi ha gran quantitat al teixit connectiu, a la submucosa del tracte gastrointestinal,
al pulmó, al fetge i a la melsa.
Quan una d’aquestes cèl·lules identifica un bacteri, l’enganxa a la membrana mitjançant unes
invaginacions anomenades pseudòpods. Aleshores el bacteri és ingerit, formant un
fagosoma. El fagosoma es fusiona aleshores amb el lisosoma, extremadament àcid. Els
enzims lisosomals digereixen el material capturat i els productes d’aquesta digestió són
expulsats de la cèl·lula.
Activació de les cèl·lules de la immunitat innata.
Les cèl·lules de la immunitat innata es poden activar mitjançant els receptors de
reconeixement de patrons, els receptors de pèptids formilats, els receptors del complement,
els receptors de citosines, els receptors de fragments d’Immunoglobulines o els receptors de
quimiocines. Aquesta activació de les cèl·lules de la immunitat innata acabarà provocant la
producció citocines i quimiocines i la producció de substàncies citotòxiques, microbicides,
de lípids bioactius, enzims...

Immunologia 2020
CEBIO 26
Tema 3 – El sistema del Complement
Receptors de Patrons moleculars Solubles. El sistema del complement és un sistema d’uns 30 enzims proteolítics inactius (zimògens)
molt abundants al plasma que s’activen per proteòlisi i que, en fer-ho, comencen una reacció
en cascada molt potent. El resultat final d’aquesta cascada pot ser l’opsonització de la
cèl·lula bacteriana, la quimiotaxi de cèl·lules del sistema immunitari, la lisi cel·lular de la
cèl·lula infectada o l’aglutinació d’agents infecciosos.
Els PRRs són proteïnes sintetitzades bàsicament per hepatòcits però també per monòcits,
macròfags tissulars i cèl·lules epitelials dels aparells digestiu i genitourinari. Per poder ser
funcionals, les proteïnes s’han d’unir per enllaços covalents a les superfícies cel·lulars o als
seus receptors (no són actives en suspensió a la sang). A més a més, les PRRs estan
sotmeses a una regulació severa mediada per proteïnes solubles i expressades a la pròpia
membrana de les cèl·lules normals de l’hoste i absents als microorganismes.
Nomenclatura del sistema del Complement. Per entendre el sistema del complement cal tenir en compte la seva nomenclatura. De forma
general, el primer component s’anomena C1, el segon C2 i així consecutivament. D’aquests
components, la forma soluble s’anomena CXa i la forma unida CXb. Aquesta segona norma
no es compleix per C2, on la forma soluble és C2b i la forma unida és C2a.
Altres elements del sistema del complement s’anomenen “Factor” seguit d’una lletra
majúscula, B, H, I… Quan s’escindeixen els fragments s’anomenen afegint una lletra
minúscula seguint el mateix mecanisme que pels components: FactorXa per la forma soluble
i FactorXb per la forma unida.
Vies d’activació del Complement. El sistema del complement es pot activar per 3 vies diferents. La via clàssica d’activació és
aquella que es dona a partir dels complexes antigen-anticòs. La via de la lectina és aquella
que es dona quan hi ha una unió de la lectina a les superfícies del patogen. Finalment, la via
alternativa és una que funciona directament sobre la superfície dels patògens. Totes tres
vies acaben portant a l’activació del complement sense cap diferència al seu resultat final,
que consisteix en l’opsonització de la cèl·lula bacteriana, la quimiotaxi de cèl·lules del
sistema immunitari, la lisi cel·lular de la cèl·lula infectada o l’aglutinació d’agents infecciosos.
Via clàssica
La via clàssica s’inicia amb la unió de dues immunoglobulines G (IgG) a dos antígens propers
en la superfície de l’agent estrany o amb la unió d’una IgM. Aquesta proximitat permet que
els dominis H del complex C1 interaccionin amb els dominis CH2 de la IgG o CH3 de la IgM.
Així i tot, és important saber que també són lligands de C1 la proteïna C reactiva i la proteïna
amiloide del sèrum.
Un cop hi ha hagut la unió C1-Immunoglobulina, la qual ha de ser per més d’un domini a
aquestes i quan estiguin unides a una superfície cel·lular, C1r2s2 talla C4 en C4b, que es
fixa sobre la paret bacteriana, i C4a, que queda lliure al medi extracel·lular.
C2 també es tallat per C1r2s2 en C2b, que s’allibera, i en C2a, que s’uneix a C4b. C4b2a és
la C3 convertasa de la via clàssica. La C3 convertasa talla aleshores C3 en C3a, que
s’allibera, i en C3b, que queda unit a la convertasa formant C4b2a3b o C5 convertasa.
Paral·lelament, C3b es pot unir a la superfície del patogen mitjançant un enllaç tioèster. Això
li permet funcionar com una opsonina. C4a i C3a són mediadors d’inflamació quan estan
solubles al medi.

Immunologia 2020
CEBIO 27
C3, un component clau del complement.
C3 és la proteïna central i més abundant del sistema del complement. Participa en les
cascades de les vies clàssica i alternativa. L’escissió proteolítica de la cadena α del C3 la
converteix en una forma metaestable en la que els enllaços tioèster interns s’exposen i es fan
sensibles a l’atac nucleòfil dels àtoms d’oxigen o de nitrogen. El resultat és la formació
d’enllaços covalents amb proteïnes o glúcids a les superfícies cel·lulars.
Via de les lectines
La via de les lectines comença amb la unió de la lectina d’unió a la manosa (MBL) a les
Manoses terminals de glicoproteïnes i glicolípids microbians o amb la unió de la Ficolina a
Oligosacàrids que tenen N-acetil glucosamina microbians. Els glicans de les proteïnes de
membrana dels llevats acaben normalment amb residus de manosa mentre que als vertebrats
ho fan amb àcid siàlic i per tant no permeten la unió de les proteïnes MBL i Ficolina.
A la via de les lectines hi participen MASP1 i MASP2, dues serino-proteases amb funció
homòloga a C1r2s2, que s’activen quan la MBL o la Ficolina estan unides a una superfície.
MASP-1 activa MASP-2, que hidrolitza C4 i C2 com a la via clàssica. Això permet la formació
del complex C4b2a, la C3 convertasa, i del complex C4b2a3b, la C5 convertasa, igual que
a la de la via clàssica. C3a, C3b i C4a es comporten com a la via clàssica en quant a
mediadors d’inflamació i opsonització de l’agent estrany.
Via alternativa.
A la via alternativa del sistema del complement no és necessària la unió d’immunoglobulines
ni tampoc de lectines. Aquesta via comença amb una escissió espontània de C3 o per C3b
generat per la via clàssica o de les lectines que s’uneix a la paret bacteriana. C3b unit a la
paret bacteriana uneix el Factor B. Aleshores el factor D trenca el Factor B i Bb queda unit a
C3b. El complex C3bBb és la C3 convertasa de la via alternativa. Si a aquest complex C3bBb
s’hi uneix un altre C3b es forma el complex C3bBb3b: la C5 convertasa de la via alternativa.
A la via alternativa també hi participa la properdina, que estabilitza el complex C3bBb3b i que
és alliberada per neutròfils en resposta a infecció. Al no haver-hi C4, el mediador de la
inflamació és només C3a. Així i tot, en tant que es continua produint C3b, aquest manté la
seva funció d’opsonització.
C5 convertasa: la convergència de les tres vies d’activació. Independentment de si es segueix la via clàssica o la via de les lectines s’acaba formant el
complex C4b2a3b, altrament anomenada C5 convertasa. Si es segueix la via alternativa, en
canvi, la C5 convertasa està formada pel complex C3bBb3b. Aquests complexes transformen
la C5 en C5a, un mediador de la inflamació, i en C5b, que s’uneix a la paret de l’agent
infecciós i inicia la fase final d’activació del complement.
Complex d’atac a la membrana.
Un cop C5b està a la membrana, uneix a C6, la qual s’uneix a C7. Això permet que s’incorpori
C8, la qual ancora el complex a la membrana. La presència de C8 a la membrana permet la
polimerització de les entre 12 i 16 unitats de C9. El resultat d’aquesta polimerització és la
formació del complex d’atac a la membrana o MAC (membrane attack complex) que genera
porus a la membrana. Aquest porus és d’uns 100A i es troba a la membrana de l’agent
infecciós, causant la mort del bacteri per trencament de l’equilibri osmòtic (alteració gradient
protons i penetració d’enzims com lisozim).

Immunologia 2020
CEBIO 28
Funcions del sistema del complement.
Inducció de la resposta inflamatòria.
C4a, C3a i C5a, les fraccions solubles dels components del sistema del complement, reben
el nom d’anafilotoxines. Aquestes indueixen la inflamació. Ho aconsegueixen mitjançant la
desgranulació que provoca la unió del receptor amb aquests elements, un receptor que
s’expressa a màstòcits, basòfils, eosinòfils, neutròfils i cèl·lules endotelials. La desgranulació
produeix una inflamció local arran de l’alliberament d’histamina.
A més a més, C5a és un factor quimiotàctic per neutròfils i monòcits com els macròfags i
C3a és un factor quimiotàctic per mastòcits i basòfils. Les anafilotxines C4a, C3a i C5a
provoquen la vasodilatació i un augment de la permeabilitat vascular, una expressió de
molècules d’adhesió a les cèl·lules endotelials, neutròfils i monòcits i un augment de la
capacitat bactericida de macròfags i granulòcits.
Opsonització i Fagocitosi de bacteris.
Tant C3b com els productes de la seva degradació i C4b actuen com Opsonines, molècules
que s’uneixen al patogen, etiquetant-lo i facilitant la seva detecció. En conseqüència, doncs,
estan facilitant la fagocitosi, la qual es veu augmentada 1000 vegades.
La fagocitosi és el procés d’internalització de microorganismes, cèl·lules mortes o debris
cel·lulars (<0.5µm de diàmetre) mediada per receptors de membrana. Es dona en 5 passos:
Eliminació d’Immunocomplexos
El sistema del complement també serveix per eliminar immunocomplexes, complexes de
molècules d’anticossos amb antigen unit. Com que cada molècula d’anticòs té un mínim de
dos llocs d’unió a l’antigen i molts antígens són multivalents, els immunocomplexes poden
variar molt en mida. Per eliminar-los cal que els eritròcits carregats amb immunocomplexos
(IC) arribin a la melsa, on són fagocitats pels macròfags residents a través de receptors de
proteïnes del Complement que s’uneixen als IC.
Mecanismes efectors del Complement El resultat final de l’activació del sistema del complement pot consistir en 4 accions diferents:
opsonització de la cèl·lula bacteriana, quimiotaxi de cèl·lules del sistema immunitari, lisi
cel·lular de la cèl·lula infectada o aglutinació d’agents infecciosos. Cadascuna d’aquestes
funcions és duta a terme per diferents mecanismes del complex. Per començar, C3b i C4b
són els encarregats de dur a terme l’opsonització i l’aglutinació d’agents infecciosos, aquesta
última mitjançant els immunocomplexes. La lisi cel·lular és duta a terme pel complex MAC
dels components C5, C6, C7, C8 i C9; i la quimiotaxi de les cèl·lules del sistema immune és
Unió del bacteri a invaginacions de la membrana anomenades Pseudòpodes.
Ingestió del bacteri per formar el fagosoma.
Fusió del fagosoma amb el lisosoma.
Digestió per part d’enzims lisosomals.
Expulsió dels productes de la digestió.

Immunologia 2020
CEBIO 29
realitzada mitjançant C5a, C3a i C4a, que provoquen la quimiotaxi en si i la inflamació del
teixit.
Regulació del complement. El complement és un sistema molt potent i que consumeix molts recursos. Per aquest motiu
és important que no estigui actiu si no és necessari. Per aconseguir regular-lo hi ha 4
mecanismes.
Un d’ells és impedir l’activació de C1 mitjançant C1INH, que impedeix que C1r2s2 s’uneixin
a C1q i adquireixin activitat proteolítica. Un altre és impedir la formació de C3 convertasa
mitjançant DAF i MCP, que desplacen C2a de C4b i impedeixen la formació de la C3
convertasa de la via clàssica o mitjançant DAF i CR1, que desplacen Bb de C3b i impedeixen
la formació de la C3 convertasa de la via alternativa. Una altre opció és inactivar C3b amb
MCP (i CR1), que actuen com a cofactors per l’escissió proteolítica de C3b, mediada pel factor
I, generant iC3b. L’última opció es inhibir la formació del MAC. Això es pot fer amb CD59
inhibint la unió de poli-C9, o amb la proteïna S inhibint la inserció de C5b-C7 a la membrana.
De la mateixa manera que el cos humà disposa de mètodes per regular i frenar el
complement, els patògens també disposen de mecanismes d’evasió del mateix. Aquests
mecanismes d’evasió actuen a tots els nivells mitjançant proteòlisi o unions irreversibles i
inhibitòries.
PRRs solubles. Proteïnes de fase aguda. Les proteïnes de fase aguda són proteïnes plasmàtiques la concentració de les quals
augmenta molt en un procés inflamatori i que pertanyen a diferents famílies de proteïnes. Els
seus tipus més destacats són la MBL, la RCP i la SAP, les quals són lligands de C1.
Impedir l’activació de C1.
• C1INH impedeix que C1r2s2 s’uneixin a C1q i adquireixin activitat proteolítica.
Impedir la formació de C3 convertasa.
• DAF i MCP desplacen C2a de C4b i impedeixen la formació de la C3 convertasa de la via clàssica.
• DAF i CR1 desplacen Bb de C3b i impedeixen la formació de la C3 convertasa de la via alternativa
Inactivar C3b
• MCP (i CR1) actuen com a cofactors per l’escissió proteolítica de C3b, mediada pel factor I, generant iC3b.
Inhibir la formació del MAC.
• CD59 inhibeix la unió de poli-C9
• La proteïna S inhibeix la inserció de C5b-C7 a la membrana.

Immunologia 2020
CEBIO 30
Els patògens indueixen secreció d’IL-6 per part dels macròfags. La IL-6 indueix la producció
de proteïnes de fase aguda per hepatòcits. Un cop produïdes, la proteïna C reactiva s’uneix
a la fosfoacetilcolina de les superfícies bacterianes, actuant com opsonina i també activant el
Complement. Paral·lelament, la MBL uneix residus de manosa a superfícies bacterianes,
actuant com opsonina i activant el Complement.
Funció del complement. El sistema del complement té com a objectiu final l’opsonització de la cèl·lula bacteriana, la
quimiotaxi de cèl·lules del sistema immunitari, la lisi cel·lular de la cèl·lula infectada o
l’aglutinació d’agents infecciosos. En tots aquests objectius un dels elements que hi
intervenen són les cèl·lules de la immunitat innata, les quals s’activen mitjançant la unió dels
receptors dels corresponents lligands, els quals poden ser citosines, quimiocines, fragments
d’immunoglobulina, pèptids formilats, patrons moleculars o elements del sistema del
complement.
Aquesta unió donarà lloc a l’activació de les cèl·lules de la immunitat innata, les quals
secretaran 5 tipus de citoquines. IL-1β activa l’endoteli vascular, els limfòcits, la destrucció
del teixit local i incrementa l’accés de totes les cèl·lules efectores. TNF-α activa l’endoteli
vascular i incrementa la permeabilitat vascular, el que porta a un increment en l’entrada d’IgG,
complement i cèl·lules als teixits i un drenatge incrementat als limfonodes. IL-6 provoca una
activació dels limfòcits i augmenta la producció d’anticossos. CXCL8 és un factor quimiotàctic
que recluta neutròfils, basòfils i cèl·lules T al lloc de la infecció. IL-12 activa les cèl·lules NK i
indueix la diferenciació de les cèl·lules CD4-T a cèl·lules TH1.
oCol·lectines
oUneixen residus de manosa, fucosa i altres CHO
oOpsonitzen i activen el sistema del Complement
MBL (Mannose Binding Lectin)
oPentraxines
oUneixen lipopolisacàrids de la paret cel·lular de bacteris i fongs, fosfocolina
oOpsonitzen i activen el sistema del Complement
RCP (Reactive C protein)
oPentraxines
oUneixen lipid A del lipopolisacàrid
oIndueixen enzims que degraden la matriu extracel·lular, fan quimiotaxi de leucòcits i activen el sistema del Complement
SAP (Serum Amyloid protein)

Immunologia 2020
CEBIO 31
Tema 4 - Cèl·lules de la immunitat innata, Part I. Les cèl·lules de la immunitat innata, com totes les cèl·lules del sistema immunitari, sorgeixen
d’una cèl·lula troncal hematopoiètica. Aquesta es transforma o bé en un precursor limfoide
comú o en un precursor mieloide comú. En cas que es transformi en aquesta última, es pot
transformar en un progenitor de granulòcits i macròfags per derivar en un precursor de
macròfags i cèl·lules dendrítiques.
Hi ha 7 cèl·lules de la immunitat innata diferents. Les cèl·lules dendrítiques presenten
antígens i realitzen fagocitosis. Els macròfags realitzen la fagocitosi alhora que provoquen
una inflamació, reparen el teixit i presenten antígens. Els neutròfils són els encarregats de
fagocitar i provocar una inflamació. Els basòfils, igual que els mastòcits en els que es poden
convertir, produeixen una inflamació. Els eosinòfils provoquen inflamacions i realitzen la
fagocitosi i les cèl·lules NK provoquen la lisi de les cèl·lules infectades o tumorals. D’aquesta
llista de cèl·lules és interessant destacar dues coses: els neutròfils, els basòfils, els mastòcits
i els eosinòfils són granulòcits o cèl·lules polimorfonuclears i les cèl·lules de la immunitat
innata no recirculen, són o van al teixit, fan la funció que els correspon i moren.
Mastòcits. Els mastòcits són cèl·lules derivades de la medul·la òssia presents a tots els teixits però
sobretot a la pell i a la mucosa epitelial dels tractes respiratori, digestiu i urogenital. Els
mastòcits contenen abundants grànuls citoplasmàtics plens de citosines, histamina i
altres mediadors.
Els mastòcits responen a les anafilotoxines C3a, C4a i C5a; a PAMPs reconeguts per TLRs,
NLRs i RLRs; i a l’entrecreuament de les Immunoglobulines E unides als seus receptors,
FcεR, expressats a la membrana.
Els mastòcits fan una resposta inflamatòria local alliberant el contingut dels grànuls
citoplasmàtics, procés anomenat desgranulació. En aquests grànuls hi tenen histamina,
heparina, serotonina, peroxidasa, proteases (triptasa, quimasa i carboxipeptidasa) i
hidrolases i TNF-α. També hi tenen mediadors lipídics d’inflamació com la prostaglandina, els
leucotriens, i el factor activador plaquetes. Els mastòcits, que tenen un paper clau en el
desenvolupament d’hipersensibilitat i al·lèrgies, són els encarregats de sintetitzar citocines
de novo (IL-4, IL-5 i IL13), quimiocines i factors de creixement.
Efectes biològics de la desgranulació.
La desgranulació dels mastòcits provoca diferents efectes en base als diferents compostos
que allibera. La Histamina, per començar, provoca la dilatació dels capil·lars sanguinis, un
augment de la permeabilitat vascular, una contracció de la musculatura llisa dels bronquis i
una estimulació de les terminacions nervioses.
Les proteases alliberades, en canvi, degraden proteïnes que es troben als teixits. Entre
aquestes proteases trobem la quimasa, que acabarà generant encara més citocines.
Finalment, els mastòcits alliberen Citosines, les quals també tenen diferents efectes: IL- 4
estimula la producció de IgE per part de les cèl·lules B, IL-5 activa els eosinòfils i IL-13
estimula la secreció de mucus per part de les cèl·lules epitelials. IL-1β activa l’endoteli
vascular, els limfòcits, la destrucció del teixit local i incrementa l’accés de totes les cèl·lules
efectores i TNF-α activa l’endoteli vascular i incrementa la permeabilitat vascular, el que porta
a un increment en l’entrada d’Immunoglobulina G, complement i cèl·lules als teixits i un
drenatge incrementat als limfonodes.

Immunologia 2020
CEBIO 32
Mediadors lipídics d’inflamació.
L’activació del mastòcit dona lloc a una ràpida síntesi de mediadors d’inflamació nous i a
l’alliberament de mediadors lipídics que tenen diversos efectes sobre els vasos sanguinis, el
múscul llis i els leucòcits. Els més importants d’aquests mediadors deriven de l’àcid
araquidònic, el qual es genera per la hidròlisi de fosfolípids de membrana mediada per la
PLA2. L’àcid araquidònic el metabolitzen després les vies de la ciclooxigenasa o la
lipooxigenasa per produir mediadors de les reaccions al·lèrgiques.
El principal mediador derivat de l’àcid araquidònic produït per la via de la ciclooxigenasa en
els mastòcits és la prostaglandina G2. La PGG2 alliberada s’uneix a receptors situats a
cèl·lules musculars llises i actua com a vasodilatador i broncoconstrictor. La PGG2 també
promou la quimiotaxi del neutròfil i la seva acumulació en zones inflamatòries.
Els principals mediadors derivats de l’àcid araquidònic que produeixen la via de la
lipooxigenasa són els leucotriens, especialment el LTC4 i els seus productes de degradació
LTD4 i LTE4. El LTC4 el produeixen els mastòcits mucosos i els basòfils, però no el produeixen
els mastòcits del teixit conjuntiu. Els leucotriens derivats del mastòcit s’uneixen a receptors
específics situats a les cèl·lules musculars llises, diferents dels receptors de la PGG2, i causen
una broncoconstricció prolongada. En el seu conjunt, LTC4, LTD4 i LTE4 constitueixen el que
una vegada es va dir Substància de Reacció lenta de l’Anafilaxi (SRS-A), i es creu que són
mediadors importants de la broncoconstricció asmàtica.
Mecanismes efectors.
Els mediadors dels mastòcits i dels basòfils són amines biògenes i enzims emmagatzemats
preformats en grànuls, així com citosines i mediadors lipídics, que en gran part es sintetitzen
un cop hi ha hagut la activació cel·lular. Les amines biògenes i els mediadors lipídics com
PAF, PGD2 i LTC4 indueixen la fuga vascular, la broncoconstricció i la hipermotilitat
intestinal, tots components de la resposta immediata. Les citosines IL-5, TNF-α i IFNs i els
mediadors lipídics PAF, PGD2 i LTC4 contribueixen a la inflamació, el que forma part de la
fase tardana. Els enzims contribueixen, probablement, a la remodelació tissular.
Basòfils. Els basòfils són un tipus de granulòcit circulant derivat de la medul·la òssia amb similituds
estructurals i funcionals amb els mastòcits, que contenen molts dels mateixos mediadors
inflamatoris que els mastòcits i que expressen un receptor d’alta afinitat pel Fc de la IgE. Els
basòfils que es recluten en els teixits on està l’antigen poden contribuir a les reaccions
d’hipersensibilitat immediata.
Els basòfils es poden arribar a considerar, doncs, els equivalents als mastòcits però a la
circulació sanguínia. Malgrat no tenir funció fagocítica, són uns elements importants en la
resposta inflamatòria, on realitzen una desgranulació en rebre estímul (FcεRI). Amb
aquesta desgranulació alliberen histamina, serotonina, proteases neutres, hidrolases àcides,
catepsina G, carboxipeptidasa i mediadors lipídics (PAF). Els basòfils, durant la seva
activació, sintetitzen citocines de novo (IL-4). També participen en el procés de les
al·lèrgies i de resposta a helmints. Els basòfils són importants per tal que altres cèl·lules
generin immunoglobulina E i per fer quimiotaxi a les plaquetes.
Eosinòfils. Els eosinòfils són granulòcits derivats de la medul·la òssia que abunden en els infiltrats
inflamatoris de la fase tardana de les reaccions d’hipersensibilitat immediata i que
contribueixen a molts dels processos patològics a les malalties al·lèrgiques. Els eosinòfils són
importants en la defensa contra els paràsits extracel·lulars, inclosos els helmints.

Immunologia 2020
CEBIO 33
Els Eosinòfils són cèl·lules tissulars, amb una presència predominant a les mucoses i, en
menor proporció, circulants. Responen a C3a, C5a i a la fracció Fc de les Immunoglobulines,
a PAMPs (TLR7), receptors de citocines (IL5 i IL4) i a quimiocines (CCR3). Durant una
resposta inflamatòria, els Eosinòfils pateixen un desgranulació una mica diferent a la de la
resta de granulòcits arran de la morfologia del seus grànuls.
Als Grànuls primaris, els eosinòfils hi tenen enzims (hidrolases lisosòmiques i
lisofosfolipasa) que degraden la membrana dels microorganismes, histamina i mediadors
lipídics d’inflamació com prostaglandines, leucotriens i PAF.
Els Grànuls específics, en canvi, contenen MBP (proteïna bàsica principal) i ECP (proteïna
eosinofílica catiònica, ribonucleasa): proteïnes tòxiques per helmints i bacteris i que activen
mastòcits. En aquests grànuls específics també s’hi emmagatzema EPO (peroxidasa
eosinofílica), composta pels àcids hipoclorós i hipobromós.
Els Eosinòfils també tenen la opció de secretar les citocines IL3, IL5, IL10 i GM-CSF, la
quimiocina CXCL-8 i l’eotaxina CCL11. Els eosinòfils també tenen una funció fagocítica, tot
i que no és gaire rellevant. Participen de la resposta a paràsits com els helmints arran de
la seva toxicitat cel·lular depenent d’Anticòs. Els eosinòfils tenen un paper important en la
fase tardana de les al·lèrgies.
Una altra de les característiques especials dels Eosinòfils és la seva capacitat d’atraure
Neutròfils.
Neutròfils. Els neutròfils són cèl·lules fagocítiques caracteritzades per un nucli globular segmentat i
grànuls citoplasmàtics plens d’enzims catabòlics. Els neutròfils, també anomenats leucòcits
PoliMorfoNucleats,, són el tipus més abundant de leucòcits circulants i són el principal tipus
de cèl·lula que media les respostes inflamatòries agudes a les infeccions bacterianes .
En resposta a infeccions, la medul·la òssia produeix una elevada quantitat de neutròfils
(leucocitosi) i són les primeres cèl·lules que entren o extravasen al teixit i arriben al focus
inflamatori per acció de la quimiocina CXCL8 o la IL8 produïda per macròfags o altres
leucòcits. Els neutròfils són de vida curta (duren entre 6 i 48h), són fagòcits i presenten
grànuls que contenen enzims lítics i substàncies bactericides.
Els seus Grànuls primaris són més densos i contenen mieloperoxidasa (MPO), lisozim,
diverses proteases (catepsina G, elastasa, esterasa) i α-defensines. Els Grànuls secundaris,
en canvi, contenen col·lagenasa, lactoferrina, lisozim i subunitats de la NADPH oxidasa. Els
seus Grànuls terciaris tan sols contenen metaloproteïnasa.
L’alliberació de tots els seus grànuls contribueix a produir espècies reactives d’oxigen
(ROS) i un esclat respiratori. Els neutròfils també s’encarreguen de la síntesi de citocines
(TNFα, IL1β, IL12, TGFβ) i de quimiocines (CXCL8 i altres).
Mecanismes de matar.
Els neutròfils disposen de tres grans mecanismes per matar els microorganismes:
l’alliberament de substàncies antimicrobianes com les catepsines, les defensines, la
lactoferrina, el MPO, la elastasa i el lisonzim; la producció d’espècies reactives d’oxigen
(ROS) i la internalització dels patògens. Per a aquest últim mecanisme els Neutròfils
disposen de NETs, Neutrophil Extracellular Traps. Aquestes consisteixen en DNA nuclear al
qual s’enganxen histones, proteïnes (com lactoferrina i catepsines) i enzims (MPO i elastasa)
dels grànuls i que després s’expulsen a l’exterior del neutròfil. Aquesta expulsió serveix per
immobilitzar els patògens, evitant així la disseminació i facilitant la fagocitosi. Aquesta trampa,

Immunologia 2020
CEBIO 34
que és molt efectiva arran de l’alta viscositat del DNA, és un últim recurs que fan servir els
neutròfils. La destrucció d’aquesta trampa la realitzen una sèrie de proteases associades.
Macròfags. Un macròfag és una cèl·lula fagocítica present en els teixits derivada de dels monòcits
sanguinis que desenvolupa funcions importants en les respostes immunitàries innata i
adaptativa. Poden assumir diferents formes en diferents teixits, com la micròglia del sistema
nerviós central, les cèl·lules Kupffer del fetge, els macròfags alveolars del pulmó, les cèl·lules
mesangials del ronyó, els macròfags peritoneals i els osteoclasts de l’os.
Els macròfags s’activen gràcies a productes microbians, com la endotoxina, i a citocines del
limfòcit T, com el IFN-γ. Quan ho fan, expressen MHC-II i molècules coestimuladores, el que
els converteix en Cèl·lules Presentadores d’Antigen (APCs) pels limfòcits T cooperadors.
Els macròfags activats fagociten i maten microorganismes mitjançant proteïnes
bactericides com el lisozim, enzims lítics com la col·lagenasa, l’elastasa, i la lipasa; i radicals
lliures oxidants. Paral·lelament, els macròfags secreten molècules pro-inflamatòries com
elements del sistema del complement, factors de coagulació, quimiocines i les citocines IL-
1β, IL-6, TNF-α, IL-12.
Els macròfags, a més, expressen receptors de membrana, des de FcγR, FcαR i, FcεR fins
els PRRs CD14, TLRs i scavenger tot passant pels receptors per C3b, C3bi i C5a i pel receptor
a la manosa o per la proteïna C reactiva.
Fagocitosi i destrucció del patogen. La fagocitosi és un procés d’ingestió de partícules grans en les vesícules que precisa
energia. Les vesícules fagocítiques es fusionen amb els lisosomes, on es destrueixen les
partícules ingerides i, d’aquesta forma, s’aïllen de la resta de la cèl·lula els mecanismes de
lisi, que podrien ferir el fagòcit.
Els neutròfils i els macròfags expressen receptors que reconeixen de forma específica els
microbis, i la unió dels microbis a aquests receptors és el primer pas en la fagocitosi. Una
vegada un microbi o una partícula s’uneixen a receptor del fagòcit, la membrana plasmàtica
a la regió dels receptors comença a redistribuir-se i estén una projecció en forma de copa al
voltant del microorganisme. Quan la copa membranaria que sobresurt s’estén més enllà del
diàmetre de la partícula, l’extrem de la copa es tanca sobre ella i es separa a l’interior per
formar una vesícula intracel·lular. Aquesta vesícula, anomenada fagosoma, es desprèn de
la membrana plasmàtica.
Els microbis poden ser ingerits per diferents receptors de membrana dels fagòcits. Alguns
s’uneixen directament a microbis i altres a microbis opsonitzats.
Els microbis s’interioritzen en fagosomes, que es fusionen amb els lisosomes per formar
fagolisosomes, on els microorganismes moren per l’acció d’espècies reactives de l’oxigen i
del nitrogen i per enzims proteolítics. Entre els enzims proteolítics trobem proteïnes
bactericides com el lisozim i enzims lítics com la col·lagenasa, l’elastasa i la lipasa.
Esclat respiratori. L’Esclat respiratori o esclat oxidatiu consisteix en l’alliberament ràpid d’espècies reactives
d’oxigen (anió superòxid i peròxid d’hidrogen) per part de cèl·lules fagocítiques del sistema
immunitari (neutròfils, macròfags, eosinòfils) en resposta a infecció per bacteris o fongs. Això
es genera quan els fagosomes es fusionen amb els grànuls primaris i secundaris.

Immunologia 2020
CEBIO 35
Aleshores Rac2 indueix l’assemblatge d’una NADPH oxidasa funcional en la membrana del
fagolisosoma. Aquesta NADPH oxidasa catalitza la transferència d’electrons del NADPH
citosòlic a l’oxigen intrafagosomal, generant anió superòxid (O2-). Aquest procés, unit a la
superòxid dismutasa que genera peròxid d’hidrogen, i a la mieloperoxidasa que converteix el
peròxid d’hidrogen en hipoclorit i en radicals hidroxil, incrementa molt la presència d’agents
oxidants. A més a més, es produeixen halògens que provoquen halogenació de la paret
cel·lular o la descarboxilació d'aminoàcids, donant lloc a aldèhids tòxics.
Les espècies reactives del nitrogen que es formen als macròfags es generen gràcies a
l’acció de la Sintetasa d’òxid nítric induïble (iNOS), que transforma una arginina i un oxigen
molecular en òxid nítric i en L-citrulina. L’òxid nítric pot derivar en les altres dues espècies
reactives de nitrogen: Peroxinitrit i diòxid de nitrogen.
Activació dels macròfags. Els macròfags es poden activar de dues maneres diferents. Quan reconeixen PAMPs o
citocines inflamatòries, es diu que els macròfags s’activen de manera clàssica o que s’activa
el macròfag M1, el que també passa per la unió dels lligands de IFN-γ. En aquests casos,
els macròfags activats realitzen les accions microbicides, com la fagocitosi i la mort de
moltes bactèries i fongs mitjançant les espècies reactives de l’oxigen, del nitrogen i enzims
lisosomals. També en aquest cas realitzen acció inflamatòria per mitjà de les quimiocines,
la IL-1, la IL-12, la IL-23, la IL6, la TNFα i la IL18.
Pel contrari, quan l’activació és per DAMPs, helmints, PGE2, glucocorticoides, IL-13 o IL-4 es
considera que s’han activat els macròfags M2 o que s’han activat per la via alternativa, el
que provoca efectes diametralment oposats que si s’activa de la manera clàssica. Per un
costat es secreta IL-10 i TGF-β, que tenen efecte anti-inflamatori. Per l’altre, la secreció de
poliamines de prolina i també de TGF-β serveixen per realitzar una fibrosi i reparar ferides.
Funcions efectores dels macròfags activats
Els macròfags activats tenen 4 funcions efectores, cadascuna amb les seves corresponents
molècules produïdes en els mateixos macròfags activats.
La primera de les funcions efectores és la destrucció de microorganismes, el que es duu a
terme amb l’oxidasa del fagòcit i la iNOS, els quals generen intermediaris reactius d’oxigen i
òxid nítric.
La segona funció efectora és la síntesi de proteïnes de la fase aguda i del Complement,
el que provoca una quimiotaxi i l’activació de l’endoteli per una estimulació de la immunitat
adaptativa, el que es duu a terme gràcies a les citocines TNF i IL-12.
Una altra de les funcions efectores és la que es realitza amb els factors de creixement de
fibroblasts, els factors angiògens i les metaloproteinases: la remodelació dels teixits.
Finalment, l’última funció efectora és la que es pot realitzar gràcies a l’augment de les
molècules del MHCi dels coestimuladors, és a dir, la potenciació de la presentació
d’antígens.

Immunologia 2020
CEBIO 36
Tema 5 - Cèl·lules de la immunitat innata. Part II Hi ha 7 cèl·lules de la immunitat innata diferents. Les cèl·lules dendrítiques presenten
antígens i realitzen fagocitosis. Els macròfags realitzen la fagocitosi alhora que provoquen
una inflamació, reparen el teixit i presenten antígens. Els neutròfils són els encarregats de
fagocitar i provocar una inflamació. Els basòfils, igual que els mastòcits en els que es poden
convertir, produeixen una inflamació. Els eosinòfils provoquen inflamacions i realitzen la
fagocitosi i les cèl·lules NK provoquen la lisi de les cèl·lules infectades o tumorals. D’aquesta
llista de cèl·lules és interessant destacar dues coses: els neutròfils, els basòfils, els mastòcits
i els eosinòfils són granulòcits o polimorfonuclears i les cèl·lules de la immunitat innata no
recirculen, són o van al teixit, fan la funció i moren.
Cèl·lules NK. Les cèl·lules NK o limfòcits citolítics naturals són un subgrup de limfòcits derivats de la
medul·la òssia que intervenen en les respostes immunitàries innates. Actuen contra cèl·lules
infectades per microbis mitjançant mecanismes lítics directes i la secreció de IFN-γ. Els
limfòcits NK no tenen un receptor específic d’antigen, com podria ser un TCR, ni CD3, ni
generen memòria. Les cèl·lules NK no necessiten activació, però són més eficients quan
aquesta es dona per part de cèl·lules dendrítiques i citocines. L’activació en aquests casos
està regulada per una combinació de receptors estimuladors i inhibidors que es troben a
la superfície cel·lular. Aquests receptors reconeixen molècules pròpies del MHC. Els
limfòcits NK representen entre el 5 i el 10 % dels limfòcits a la sang (CD56dim), la resta
(CD56bright) es troben als òrgans limfoides, abreviats sovint com a OLs.
Receptors activadors i inhibidors de les cèl·lules NK.
L’equilibri o desequilibri entre receptors activadors i inhibidors permet als limfòcits NK
distingir entre cèl·lules normals i cèl·lules alterades (ja perquè són tumorals o infectades).
Són receptors activadors i coestimuladors de les cèl·lules NK el CD94/NKG2C, NKG2D,
NKR-P1c, CD16, NKp30, NKp44 i NKp46 i KIRs. Alguns d’aquests receptors activadors són
receptors de lectines de tipus C. Pel contrari, els limfòcits NK només disposen de 3 receptors
inhibidors: CD94/NKG2A, ILT-2 i KIRs, que també n’és un activador.
Als receptors activadors de les cèl·lules NK la unió del lligand fosforila les tirosines del domini
ITAM, el que fa que es reclutin tirosina quinases que fosforilen proteïnes adaptadores i
continuen la cascada de senyalització. En el cas dels receptors inhibidors, en canvi, la unió
del lligand fosforila la tirosina del domini ITIM, el que provoca que es recluti tirosina fosfatasa
que inhibeix la cascada de senyalització.
Receptors activadors de les cèl·lules NK.
Els receptors activadors dels limfòcits NK reconeixen un grup heterogeni de lligands,
alguns dels quals poden expressar-se en cèl·lules normals i altres, sobretot, en cèl·lules que
hagin patit estrès, estiguin infectades per microbis o s’hagin transformat. Les característiques
moleculars dels lligands per molts d’aquests receptors no estan ben caracteritzades. Està
estudiat que també són lligands les proteïnes víriques i els proteoglicans de les cèl·lules
tumorals.
La majoria dels receptors activadors NK comparteixen la característica comuna d’una
estructura en les seves cues citoplasmàtiques, anomenada estructura tirosínica d’activació
del receptor immunitari (ITAM), que participa en la transmissió de senyals que promouen la
mort de la cèl·lula diana i la secreció de citocines. En alguns d’aquests receptors, una sola
cadena conté ITAM, així com la porció que s’uneix al lligand extracel·lular. En altres receptors,
les ITAM estan en cadenes polipeptídiques separades, com FcεRIγ, ζ i DAPI12, que no
s’uneixen al lligand, però s’associen de forma no covalent a la cadena que sí que ho fa.

Immunologia 2020
CEBIO 37
La capacitat dels receptors activadors d’induir respostes funcionals en els limfòcits NK
augmenta amb les citocines. Les principals citocines del sistema immunitari innat que
estimulen la funció del limfòcits NK són la IL-12, la IL-15, la IL-18 i els interferons del tipus I.
Cadascuna de les citocines realça l’activitat citotòxica dels limfòcits NK i la quantitat de la
citocina IFN-γ que els limfòcits NK secreten. Aquesta citocina, la IFN-γ, té varis efectes
antimicrobians.
Són receptors activadors de les cèl·lules NK el CD16, els NCRs, el KIR2DS, el CD94 i el
NKG2C,E i el NKG2D.
Receptors Inhibidors de les cèl·lules NK.
La majoria dels receptors inhibidors, però no tots, en el sistema immunitari contenen
estructures ITIM orientades cap al citosol que poden reclutar fosfatases amb un domini SH2 i
així atenuar les senyals de moltes formes. Els receptors inhibidors desenvolupen funcions
clau en els limfòcits NK, els limfòcits T i els limfòcits B, així com en altres cèl·lules de la
immunitat innata.
En els limfòcits NK humans, els receptors inhibidors clau poden dividir-se àmpliament en 3
grups: KIR o receptors citolítics del tipus Ig, la família de proteïnes ILT, que es relacionen
estretament amb els KIR, i les lectines del tipus C, el principal dels quals és un heterodímer
que consisteix en la lectina del tipus C NKG2A i CD94.
Els KIR contenen dominis extracel·lulars d’Ig que poden reconèixer molècules de classe I
del HLA, i un subgrup d’aquests receptors conté estructures ITIM citosòliques. L’ILT-2, part
d’una família de receptors inhibidors més antiga evolutivament que els KIR, també té dominis
extracel·lulars de Ig que s’uneixen a la classe I de HLA i a estructures ITIM citosòliques.
El dímer CD94/NKG2A s’uneix a una molècula atípica de la classe I del MHC anomenada
HLA-E, i la cadena NKG2 d’aquest dímer conté estructures ITIM citosòliques.
Mecanisme d’acció de les cèl·lules NK.
Els receptors activadors dels limfòcits NK reconeixen lligands en les cèl·lules diana i
activen la proteïna tirosina quinasa (PTK). L’activitat d’aquesta proteïna és inhibida pels
receptors inhibidors, que reconeixen molècules de la classe I del MHC, i activen les proteïnes
tirosina fosfatasa (PTP). Els limfòcits NK no maten de forma eficient cèl·lules sanes que
expressin la classe I del MHC.
Si una infecció vírica o algun altre tipus d’estrès inhibeix l’expressió de la classe I MHC a les
cèl·lules infectades i indueix a l’expressió de lligands activadors addicionals, el receptor
inhibidor del limfòcit NK no s’uneix al seu lligand i les funcions del receptor activador sense
cap oposició desencadenen respostes de limfòcits NK, com la mort de cèl·lules diana i
la secreció de citocines.
Les cèl·lules estressades per la infecció o la transformació poden expressar majors quantitats
de lligands activadors, els quals s’uneixen als receptors activadors dels limfòcits NK i
indueixen a una major fosforilació de tirosines de les que poden compensar les fosfatases
associades al receptor inhibidor, el que provoca la mort de les cèl·lules estressades.
Per resumir els tres casos d’actuació, es podria dir que si MHC-I és reconegut per un
receptor inhibidor, aquest envia un senyal negatiu i la cèl·lula NK no mata a la cèl·lula
diana. En canvi, en absència o alteració de l’expressió de MHC I, manca el senyal negatiu i
s’activa la capacitat lítica de la cèl·lula NK, pel que allibera el contingut dels seus grànuls i
mata la cèl·lula diana.

Immunologia 2020
CEBIO 38
Les funcions efectores dels limfòcits NK són matar a les cèl·lules infectades i activar
els macròfags perquè destrueixin els microbis fagocitats. Els limfòcits NK tenen grànuls
que contenen proteïnes mediadores de la mort de les cèl·lules diana. Quan els limfòcits NK
s’activen, les exocitosis dels grànuls allibera aquestes proteïnes de manera adjacent a les
cèl·lules diana. Una proteïna del grànul del limfòcit NK, anomenada perforina, facilita
l’entrada d’altres proteïnes dels grànuls, anomenades granzimes, en el citoplasma de les
cèl·lules diana. Les granzimes són enzims que inicien una seqüència d’esdeveniments
transmissors de senyals que causen la mort de les cèl·lules diana per apoptosi.
Citotoxicitat cel·lular depenent d’anticòs.
Els limfòcits citolítics naturals i altres leucòcits s’uneixen a les cèl·lules cobertes
d’anticòs mitjançant receptors pel Fc i les destrueixen. Aquest procés rep el nom de
Citotoxicitat cel·lular depenent d’anticòs. Els limfòcits NK fan servir el seu receptor pel Fc,
el FcγRIIIA, per unir-se a cèl·lules cobertes d’anticòs. Arran de les característiques de
FcγRIIIA, la ADCC es produeix només quan la cèl·lula diana està coberta de molècules
d’anticòs i la IgG lliure en el plasma no activa els NK ni entra en competència eficaç contra
la IgG unida a la cèl·lula per la unió al FcγRIII.
Les cèl·lules NK, a més, són les principals productores d’IFNγ, que activa els macròfags que
secreten IL12, que indueix més producció d’IFNγ per cèl·lules NK. A més, IFNγ incideix en
la resposta adaptativa induint la diferenciació dels limfòcits T cap a efectors TH1, el que
indueix l’activació de limfòcits T citotòxics (CTLs). IFNα i IFNβ secretats per les cèl·lules
dendrítiques també estimulen les cèl·lules NK.
Innate Lymphoid Cells. Les Innate Lymphoid Cells són unes cèl·lules de la immunitat innata que detecten
mediadors d’inflamació i citocines secretades per cèl·lules epitelials i mieloides. Aquestes
cèl·lules responen de manera ràpida amb la producció de citocines i enzims citotòxics.
ILC1 és responsable de la resposta innata a virus, bacteris intracel·lulars i paràsits. ILC2 és
responsable de la resposta a paràsits extracel·lulars com els helmints. També són les
encarregades de la producció de factors solubles, citocines, que regulen l’activació d’altres
cèl·lules del sistema immunitari (com els macròfags o els granulòcits) de l’hoste que ajuden
a la seva expulsió i a la reparació del teixit. ICL3 provoquen la resposta innata a bacteris
extracel·lulars.
Cèl·lules dendrítiques. Les cèl·lules dendrítiques són cèl·lules derivades de la medul·la òssia a partir de progenitors
mieloides i limfoides que es troben en els teixits epitelials. La seva morfologia es caracteritza
per projeccions membranoses fines (dendros). Existeixen molts subgrups de cèl·lules
dendrítiques amb diverses funcions. Són poc abundants a la sang i es troben majoritàriament
als teixits. Reben diferents noms segons el teixit on es trobin, ja sigui Cèl·lules de
Langerhans de la pell, Cèl·lules dendrítiques interdigitants del timus i dels òrgans
limfoides o Cèl·lules dendrítiques dels òrgans limfoides perifèrics. Les cèl·lules
dendrítiques tenen una funció de pont entre immunitats innata i adaptativa.
Les cèl·lules dendrítiques activades (altrament dit madures) funcionen com a cèl·lules
presentadores d’antigen (APC) pels limfòcits T verges i són importants per l’inici de les
respostes immunitàries adaptatives a antígens proteínics. Les cèl·lules dendrítiques
immadures (o en repòs) són importants per la inducció de tolerància respecte antígens propis.

Immunologia 2020
CEBIO 39
Les cèl·lules dendrítiques presenten molts PRRs, ja siguin receptors pel fragment Fc de
la Ig, Receptors “scavengers”, receptors de proteïnes de xoc tèrmic, receptors per fragments
derivats de C3 o receptors de lectina tipus C (RCL).
Diferents TLRs de les cèl·lules dendrítiques.
Com que existeixen diferents cèl·lules dendrítiques i aquestes es troben en llocs
diferents, els Receptors de tipus Toll que han de produir són també diferents. En les
cèl·lules dendrítiques convencionals, per exemple, hi ha una elevada expressió de CD11c,
MHC-II i molècules coestimuladores per tal de poder fer una presentació d’antigen i activació
de cèl·lules T naive. El marcador de les Cèl·lules dendrítiques convencionals és el DC-SIGN
i DEC-205.
En les cèl·lules dendrítiques plasmacitoides, el que ens trobem és una producció elevada
d’interferons de tipus I per poder donar resposta a virus i una expressió baixa de CD11c,
molècules MHC-II i coestimuladores. Les Cèl·lules dendrítiques plasmacitoides tenen com a
marcador BDCA-2 i BDCA-4 en humans.
De la resposta innata a l’adaptativa. El procés de resposta immunològica comença amb una adhesió del patogen a l’epiteli, d’on
després realitzarà una infecció local superficial. Aquesta acabarà sent una infecció local del
teixit, atacada per part d’un drenatge limfàtic i activant la resposta immunitària adaptativa. En
el següent tema començarem a tractar la resposta adaptativa a través de les
immunoglobulines.

Immunologia 2020
CEBIO 40
Tema 6 – Estructura de les immunoglobulines.
BCR o Anticòs. Les immunoglobulines tenen dues funcions: reconèixer l’antigen i activar el sistema
immunitari. Les immunoglobulines es poden trobar en la seva forma unida a la membrana
del limfòcit B, cas que s’anomenen BCRs, o en la seva forma soluble, cas que s’anomenen
Anticossos.
El BCR està format per una immunoglobulina de membrana més un dímer Igα-Igβ.
Quan un limfòcit B naive veu un antigen, es produeix una cascada de senyals internes que fa
que la cèl·lula s’expandeixi clonalment i es diferenciï de cèl·lula B a cèl·lula plasmàtica, la
qual té la funció d’eliminar el patogen. Aquesta cèl·lula és més gran que la cèl·lula B, té molt
més reticle endoplasmàtic rugós, el qual cobreix quasi un 50% de la cèl·lula, ha perdut els
receptors de membrana (perquè no necessita buscar més antígens) i la seva funció
principal és produir anticossos. El temps que es triga en passar de limfòcit B a cèl·lula
plasmàtica és d’aproximadament 7 dies.
Les immunoglobulines són heterodímers en forma de Y compostos per 4 cadenes
unides per ponts disulfur i altres interaccions no covalents: 2 cadenes lleugeres (L) i
dues pesades (H). Les dues cadenes pesades són iguals entre elles, i les dues lleugeres
també. Poden ser de membrana, cas en què tindran una regió transmembrana que l’ancora
a la mateixa o poden ser solubles, cas en què no tindran aquesta regió.
Les immunoglobulines presenten una regió variable, que serà el lloc d’unió a l’antigen i
una regió constant. La regió variable d’una immunoglobulina està
formada per l’extrem d’una cadena pesada i d’una lleugera, mentre que la regió constant està
formada per la resta.
Estructura de les immunoglobulines. L’estructura de les immunoglobulines consisteix en dues cadenes de cada tipus unides entre
elles per ponts disulfur. La cadena lleugera és petita i pot ser de tipus κ o λ. La cadena pesada
és la més gran de les dues i conté una frontissa o xarnera i un colze, mentre que la cadena
lleugera només té un colze. A cada cadena hi trobem regions constants (1 a les cadenes
lleugeres i entre 3 i 4 a les pesades), una regió frontissa al mig de la zona constant que
entrellaça les cadenes pesades perquè l’estructura sigui més flexible i s’adapti a l’antigen i
una regió variable que serveix de zona de reconeixement d’antígens.
Les immunoglobulines tenen uns dominis globulars formats per uns 110aa que tenen un
plegament en làmina β formant un sandwitch estabilitzat per un enllaç disulfur. Cadascuna de
les làmines β està formada per cadenes β antiparal·leles unides per bucles sense estructura
terciària. Aquest plegament genera una estructura estable que es denomina plegament
d’immunoglobulina o domini de tipus Ig. Els dominis d’Ig es troben en moltes proteïnes del
sistema immunitari que formen part de la denominada superfamília de les immunoglobulines.
Les superfamílies d’immunoglobulines és un conjunt de molècules amb estructura molt similar
a les immunoglobulines tot i que no tenen perquè ser del sistema immunitari. Dins d’aquesta
família hi trobem les IgG, les TCR, les MCH classe I i II, el CD2, els receptors de Fc, els
receptors de cèl·lules T, receptors de poli-Ig i molècules d’adhesió cel·lular entre d’altres.
Fragments d’Immunoglobulines sortints de la digestió amb proteases. La proteòlisi d’immunoglobulines amb proteases específiques dóna fragments
funcionalment diferents. El tall amb Papaïna és per sobre del S-S de la regió xarnera i
separa la immunoglobulina en 3 parts: 2 fragments Fab que uneixen l’antigen i un Fc format

Immunologia 2020
CEBIO 41
per dominis constants C de la cadena pesada. El tall amb Pepsina és per sota del S-S i genera
un fragment F(ab’)2 bifuncional i fragments petits de la regió Fc.
S’ha vist que el Fragment Fab’ és el responsable del reconeixement de l’antigen, que el
fragment Fc és el responsable de l’activitat biològica de la immunoglobulina i que F(ab’)2 és
el que es genera per l’entrecreuament de receptors Fc.
Regió o domini variable de les immunoglobulines. La regió variable de les immunoglobulines és la que determina l’especificitat antigènica
de la mateixa. En aquesta, els loops que connecten les fulles β antiparal·leles i que es troben
exposades es consideren regions hipervariables. Això es deu a què la seva variabilitat
aminoacídica és molt més elevada que en qualsevol altre proteïna. Aquestes regions
hipervariables reben el nom de CDRs: Complementary Determining Region. Les fulles β
antiparal·leles que gairebé no varien reben el nom de FR i són les encarregades del
manteniment estructural del plegament. Aquests CDR seran les parts de la
immunoglobulina encarregades de la interacció amb l’antigen, sent una part essencial
del fragment Fab’. A les immunoglobulines hi ha 3 CDRs per cada cadena. Això significa que
hi ha 6 CDRs per immunoglobulina.
Interacció antigen-anticòs.
Determinants de la interacció Antigen-Anticòs.
Entre un anticòs i un antigen hi ha d’haver una interacció que permeti al primer unir-se
al segon. Això s’aconsegueix mitjançant Interaccions no covalents (reversibles) entre l’epítop
i el paratop. El paratop és el conjunt d’aminoàcids de la immunoglobulina que
interaccionen amb l’antigen, mentre que l’epítop és el conjunt d’aminoàcids de l’antigen
que interaccionen amb la immunoglobulina. La complementarietat del paratop i l'epítop
dependrà de la seva forma i de les possibilitats de formar múltiples interaccions no covalents.
Hi ha 4 possibles interaccions no covalents. Les forces hidrofòbiques són les més fluixes i
són fruit de la mala interacció entre els grups hidrofòbics i l’aigua, el que provoca que els
primers tinguin tendència a agregar-se per tal d’excloure molècules d’aigua. Aquesta atracció
també inclou forces de van der Waals, les quals consisteixen en fluctuacions en els núvols
d’electrons al voltant de molècules tot produint una polarització oposada d’àtoms veïns. Els
ponts d’hidrogen són similars a les forces de van der Waals amb la diferència que la
polarització ja està present i es soluciona mitjançant el compartir un àtom d’hidrogen entre
dos àtoms electronegatius. Les forces electroestàtiques són les interaccions no covalents
més fortes i es deuen a l’atracció entre càrregues oposades.
Afinitat i Avidesa de la interacció Antigen-Anticòs.
L’afinitat d’una Immunoglobulina per un antigen és la intensitat de la interacció no covalent
que s’estableix entre un únic epítop d’un Antigen i un sol paratop, recordem que és una
interacció reversible i que, per tant, hi ha una constant d’equilibri o afinitat que ve donada
per la fórmula de
𝐾𝑎 =[𝐴𝑔 − 𝐴𝑐]
[𝐴𝑔][𝐴𝑐]=
1
𝐾𝑑
On Kd és la concentració de lligand a la qual el 50% dels llocs d’unió d’Antigen de la
Immunoglobulina estan ocupats. En conseqüència, amb una Kd baixa és quan hi ha una major
afinitat. L’avidesa, en canvi, és la intensitat resultant de la suma de l’afinitat de les múltiples
interaccions entre un anticòs polivalent i un antigen.

Immunologia 2020
CEBIO 42
Epítops reconeguts per les immunoglobulines
Les immunoglobulines poden reconèixer 3 tipus d’epítops. Els epítops conformacionals són
aquells que poden ser reconeguts pels anticossos quan la proteïna antigènica es troba en la
seva forma plegada però que no es correspon amb una sèrie d’aminoàcids consecutius en
l’estructura primària de l’antigen. Els epítops lineals són el contrari que els epítops
conformacionals, ja que es corresponen a una sèrie d’aminoàcids consecutius en l’estructura
primària de l’antigen. Aquest epítop, el lineal, pot estar accessible a l’anticòs en la forma
plegada de l’antigen o pot ser que sigui necessària una desnaturalització per poder-lo
reconèixer. L’últim tipus és l’epítop resultant d’una proteòlisi limitada. Aquests només es
poden reconèixer per part de l’anticòs si abans ha actuat una proteïna proteolítica que ha
fragmentat d’alguna manera l’antigen i ara exposa al solvent una part abans amagada al
mateix.
Regió constant de la cadena pesada de les immunoglobulines. La regió constant de la cadena pesada de les immunoglobulines és el que abans hem definit
com a Fc dels fragments resultants d’una proteòlisi de les immunoglobulines. Aquest és el
fragment de la immunoglobulina que en determina l’activitat biològica o efectora. Hi ha
5 tipus de cadenes pesades: cadena γ, cadena µ, cadena δ, cadena α i cadena ε. És de
destacar la diferència entre el nombre i la localització dels ponts disulfur que uneixen les
cadenes. IgM i IgE manquen d’una regió xarnera, però cadascuna disposa d’un domini
addicional de cadena pesada. Els isotips també difereixen en la distribució dels grups de
carbohidrats amb unions en N.
Les diferents regions constants tenen una diferent cadena pesada, un diferent nombre de
dominis, un diferent patró de glicosilació, un diferent nombre i localització dels ponts disulfur
i una diferent xarnera. Així i tot, només hi ha dos grans tipus de cadena lleugera, κ (κ) o
lambda (λ).
Subclasses de IgG i la seva flexibilitat. En humans hi ha quatre subclasses de IgG: IgG1, IgG2, IgG3 i IgG4. Malgrat entre elles hi
ha una homologia molt gran respecte la regió constant Fc, tenen diferències importants
en quant a la regió xarnera. Per començar, IgG3 té una regió xarnera gran (de 62
aminoàcids) que li proporciona molta flexibilitat, pel que uneix més eficientment l’antigen quan
està a baixa concentració o molt dispers o quan és poc accessible. Pel contrari, IgG1 i IgG4
tenen una regió xarnera curta, el que els confereix menys flexibilitat. En conseqüència, si
haguéssim d’ordenar les subclacces humanes de IgG (en humans en són 4, però en ratolins
en són 5 perquè tenen dos tipus de IgG2) segons la seva flexibilitat en ordre descendent el
resultat seria el següent:
Tot i tenir un 90% d’homologia en la seqüència d’aminoàcids, cada subclasse té
característiques úniques en quant a la interacció amb l’antigen, la formació
d’immunocomplexes, l’activació de cèl·lules efectores, la vida mitja i el transport a través de
la placenta. El tipus d’Antigen determina la subclasse d’Immunoglobulina que
predominarà a la resposta. La deficiència d’una de les subclasses no és normalment letal
per un individu però sí que augmenta la susceptibilitat a patir infeccions per determinats
patògens.
lgG3 lgG1 lgG4 lgG2

Immunologia 2020
CEBIO 43
Flexibilitat i moviment de les immunoglobulines.
Les immunoglobulines es poden moure de dues maneres respecte la seva frontissa. La
primera manera és un moviment de plec segons el qual poden acostar més o menys la regió
variable a la regió constant. Aquest moviment depèn de la longitud de la xarnera. L’altre
moviment és el de rotació de la cadena variable, el qual es realitza a partir de la capacitat de
rotació dels enllaços covalents.
Isotips de les immunoglobulines: monòmers o multímers.
Les immunoglobulines són unes estructures en forma de Y que actuen, en teoria, de manera
solitària amb tan sols 2 llocs d’unió a l’antigen. Hi ha cops que s’uneixen dues o més
immunoglobulines per formar multímers i incrementar la quantitat de llocs d’unió a
l’antigen. Una de les immunoglobulines que forma multímers, més concretament dímers, és
la immunoglobulina A, que es pot presentar també com a monòmer. Formar un dímer permet
a aquest tipus d’immunoglobulina tenir 4 llocs d’unió a l’antigen. El cas més clar i present és
el de la IgM. En aquest cas, la unió és de 5 immunoglobulines, formant un multímer en forma
de pentàgon que disposa de 10 llocs d’unió a l’antigen. La unió d’immunoglobulines es fa per
mitjà de la cadena J.
Cada isotip d’immunoglobulina té associat un o varis tipus de receptors Fc, els quals
són responsables que puguin realitzar la seva funció.
Funció associada a l’isotip de les immunoglobulines
Cadascun dels isotips de les immunoglobulines té una funció associada. Per començar,
IgG és la més important i la que més funcions té. IgG és la immunoglobulina encarregada
d’opsonitzar els antígens per tal que els fagocitin els macròfags i els neutròfils, d’activar la via
clàssica del sistema del complement, de provocar la ADCC per part dels limfòcits NK, de
atorgar la immunitat neonatal i de fer inhibició per feedback de les cèl·lules B activades.
La resta d’immunoglobulines tenen menys tasques assignades, però no per això deixen de
ser importants. IgM és l’encarregada d’activar la via clàssica del complement i de ser el
receptor d’antigen dels limfòcits B naive, IgA té la funció de garantir la immunitat de la mucosa,
IgE s’encarrega de la desgranulació de les cèl·lules màstiques i IgD és també un receptor
d’antigen pels limfòcits B naive.
Determinants que defineixen les diferències entre les Igs.
Existeixen 3 tipus de determinants antigènics de les immunoglobulines. L’isotip, que ve
determinat per la regió constant de la cadena pesada, l’al·lotip, el qual està determinat per
petites diferències de seqüència (al·lels) en cada isotip (diferències entre individus) i l’idiotip,
el conjunt d’aminoàcids que formen la regió hipervariable de cada molècula
d’Immunoglobulina, i que formen el lloc d’unió a l’antigen.
Funció efectora de les immunoglobulines. Els anticossos contra microorganismes i les seves toxines neutralitzen aquests agents, els
opsonitzen per la fagocitosi, els sensibilitzen per la toxicitat mediada per anticossos i activen
el sistema del complement. Aquestes diverses funcions poden estar mediades per diferents
isotips d’anticossos.
Una possibilitat de funció efectora dels anticossos és mitjançant l’isotip de la IgM. En
aquest cas, les molècules d’IgM pentamèrica adopten la forma de grapa al unir-se a antígens
de la superfície bacteriana. Aleshores C1q s’associa a una molècula de igM unida i aquesta
unió a la immunoglobulina activa C1r, que talla i activa la serinoproteasa C1s.

Immunologia 2020
CEBIO 44
En el cas que la funció la dugui a terme la IgG, les molècules de IgG s’uneixen a antígens de
la superfície bacteriana, C1q s’associa amb al menys dues molècules properes de IgG i
aquesta unió a la immunoglobulina activa C1r, que talla i activa la serinoproteasa C1s.
En ambdós casos, el C1q del Sistema del Complement interacciona amb el domini CH3
de les IgMs i CH2 d’IgG1 i IgG3.
Si els anticossos han de neutralitzar toxines, el que fan és bloquejar la unió de la mateixa, la
qual podria provocar la seva endocitosi pels complexes del receptor i una posterior dissociació
de la toxina que provoqui l’alliberament de la cadena activa, la qual enverina les cèl·lules.
En el cas dels virus el procediment és similar, ja que l’anticòs bloqueja la unió del receptor
dels virus i també pot bloquejar l’esdeveniment de fusió.
Funció efectora de les immunoglobulines depenent de FcR.
La unió de l’anticòs a un agent estrany pot derivar, apart de en l’activació del sistema del
complement, en l’opsonització i fagocitosi de patògens per part dels fagòcits o en
l’eliminació de cèl·lules infectades o de paràsits per citotoxicitat cel·lular depenent
d’anticòs (ADCC). En el primer cas, la unió de l’anticòs al patogen permet que hi hagi una
interacció entre la immunoglobulina i un receptor FcγRI, el qual inicia una cascada de
senyalització intracel·lular en el fagòcit que la presenta. El resultat final d’aquesta cascada és
induir el fagòcit a fagocitar l’agent patogènic.
Per tal d’eliminar les cèl·lules infectades o els paràsits, els anticossos tenen dues vies.
En la primera, l’anticòs o anticossos s’uneixen als antígens de la superfície de la cèl·lula
infectada. Adherits a aquesta, la cèl·lula circula pel corrent sanguini o per algun altre medi
fins trobar-se amb un limfòcit NK. Aquest, en tenir una interacció dels anticossos amb FcγRIII
present a la membrana del NK, matarà la cèl·lula infectada i rodejada d’anticossos. La segona
via consisteix en envoltar d’anticossos el paràsit, provocant així una interacció dels primers
amb els receptors FcεRI dels eosinòfils, els quals mataran el paràsit.

Immunologia 2020
CEBIO 45
Tema 7 - Immunoglobulines. Reordenament gènic
Generació immunoglobulines Les immunoglobulines que es generen al cos humà es fan amb aproximadament 1013
especificitats antigèniques diferents. Entre aquestes diferents especificitats, n’hi que són
formes secretades (Ac) i n’hi ha que ho són de membrana (BCR). N’hi ha que tenen diferents
isotips i altres el que tenen és capacitat per adaptar-se a la resposta.
La generació d’immunoglobulines és un procés de recombinació somàtica entre
diferents segments gènics que codifiquen per les cadenes pesades i per les lleugeres.
Aquest procés de recombinació dona lloc a un reordenament únic per cada limfòcit B i té lloc
a la medul·la òssia durant el desenvolupament dels mateixos. Cada limfòcit B expressa un
BCR funcional d’especificitat antigènica única.
Gens de les immunoglobulines i la seva localització cromosòmica. Les cadenes pesades i lleugeres de les immunoglobulines estan codificades per
famílies de gens situades en diferents cromosomes. Les cadenes lleugeres tenen
codificades les seves varietats λ als cromosomes 22 i les varietats κ als cromosomes 4. La
cadena pesada de les immunoglobulines, en humans, està codificada al cromosoma 14.
Organització dels gens de les cadenes pesades de les immunoglobulines.
A l’ADN de la línia germinal, les famílies de gens contenen seqüències codificants o
segments gènics, separades per regions no codificants. Aquests segments gènics poden
ser de 5 tipus diferents segons si són de la regió variable, de la regió constant o simplement
són la seqüència Líder.
La seqüència líder ”L” es troba davant del segment gènic variable “V”, que es transcriurà
primer. Seguidament s’aniran transcrivint els segments gènics de diversitat “D” per acabar
transcrivint el segment gènic d’unió “J”. Al final es transcriurà la regió constant, composta
només pels segments gènics constants “C”.
Organització dels gens de les cadenes lleugeres de les immunoglobulines.
Les famílies de gens de les cadenes lleugeres no tenen un segment gènic de diversitat,
sinó que només hi ha seqüència líder, segment variable, segment d’unió i segment constant.
Això succeeix tant per les cadenes λ com per les κ.
Malgrat tenir el mateix nom, els locis de les immunoglobulines no tenen el mateix nombre
de segments gènics funcionals. Varien segons les cadenes que s’estan generant.
Reordenament gènic.
En el procés de generació de cadenes lleugeres i pesades, s’escull a “l’atzar” una
determinada combinació de segments V, (D) i J. La recombinació del segments gènics V,
D i J genera el domini variable de les cadenes H i la recombinació dels segments V i J el de
les cadenes L. (VDJ).
Diversitat de les Ig per la diversitat combinatòria.
Usant només els gens V, D i J funcionals podem determinar que hi ha 6,000
combinacions possibles d’immunoglobulines. (40 VH x 25 DH x 6JH). L’anàlisi dels
segments D de diferents immunoglobulines va demostrar que el segment D té tres codis de
lectura i que, per tant, es poden generar diferents seqüències d’aminoàcids i, en
conseqüència, es pot generar més diversitat d’anticossos. Per tant, multiplicant les possibles
combinacions dels gens V, D i J funcionals per les possibles pautes de lectura del segment D
obtenim 18,000 combinacions. Possibles.

Immunologia 2020
CEBIO 46
Respecte les cadenes senzilles hem de tenir en compte que hi ha 200 combinacions per la
cadena lleugera κ (40 Vκ x 5 Jκ) i que hi ha 120 combinacions de cadena lleugera λ (30 Vλ x
4 Jλ). En total, doncs, disposem de 320 cadenes lleugeres possibles diferents.
Si cadenes H i L s’aparellessin a l’atzar, tindríem 5,760,000 possibilitats de diversitat
COMBINATÒRIA (18,000 x 320). A la pràctica, però, algunes combinacions H+L són
inestables a part que alguns gens V i J s’usen més freqüentment que d’altres. Per això
s’estima que en total hi ha entre 1010 i 1013 combinacions possibles.
Mecanisme de reordenament del domini variable de les immunoglobulines.
La recombinació de segments gènics i la lligació no és perfecta, doncs hi ha flexibilitat
d’unió. La imprecisió i l’atzar durant el procés de trencament i reparació del DNA permet la
inserció o la pèrdua de nucleòtids al lloc d’unió entre segments. Per aquest motiu, dos limfòcits
B que escullin els mateixos segments V i J poden generar receptors amb regió variable
diferent, el que suposa una especificitat antigènica diferent.
Els gens d’algunes regions gèniques tenen el que s’anomena Recombination Signal
Sequence, un heptàmer palindròmic i un nonàmer ric en Adenina o Timina conservats i
separats per una seqüència intermèdia de 12 o 23 parells de bases no conservada. Aquesta
última rep el nom d’espaiador 12-RSS o 23-RSS. Només es produeix la recombinació entre
segments amb RSS de 12 i i RSS de 23. Mai 12-12 o 23-23, és la regla 12/23.
Per fer el recombinament del domini variable cal que passin una sèrie de coses. Primer, cal
que hi hagi un alineament entre els heptàmers i els nonàmers. Després cal que les RSS
alineades creïn el lloc d’unió del conjunt d’enzims anomenats recombinases. RAG-1 i
RAG-2 (recombination activating genes), les quals són exclusives dels limfòcits B i T,
reconeixen les RSS i tallen el DNA. Aquesta forma només es genera si es compleix la norma
12-23.
El reconeixement de les RSS per RAG1 i RAG2 acosta les dues seqüències RSS.
Seguidament tallen l’extrem 5’ de l’heptàmer, que es separa del segment gènic creant
bucles de DNA als seus extrems. Fet el tall, es dona l’inici de la reparació de la doble cadena
de DNA. En aquesta acció hi intervenen un conjunt d’enzims que s’anomenen complex
DSBR o double strand break repair enzyme. Aleshores, la unió dels enzims Ku70:80 als
extrems creats recluten la quinasa DNA-PK. La DNA-PK uneix Artemis al complex i el
fosforila. Artemis trenca els bucles de DNA creant seqüències palindròmiques o nucleòtids
P. Aleshores hi ha una unió de TdT o deoxinucleotidil transferasa terminal, que comença a
afegir nucleòtids a l’atzar o nucleòtids N. Si es produeix complementarietat entre les dues
cadenes del DNA, s’uneixen els extrems gràcies a la DNA IV lligasa del complex enzimàtic
DSBR.
Explicat d’una altra manera, les RSS són reconegudes per RAG1 i RAG2, que les
juxtaposen. Les recombinases fan talls simples en una de les cadenes del DNA a l’extrem
5’ de l’heptàmer. El grup OH que queda lliure a 3’ hidrolitza l’enllaç fosfodiester de la
cadena complementària de DNA, creant un bucle entre els nucleòtids dels dos extrems. El
complex Artemis:DNA-PK obre els bucles tallant a punts a l’atzar. Els nucleòtids que
s’obren s’incorporen a la cadena de DNA complementària generant seqüències
palindròmiques o Nucleòtids P. L’enzim deoxynucleotidyl transferasa terminal (TdT) afegeix
nucleòtids a l’atzar als extrems, són els nucleòtids N. Les bases complementàries
s’uneixen. Les exonucleases eliminen nucleòtids no aparellats. Finalment, el complex
enzimàtic DSBR acaba d’unir els segments. La DNA polimerasa omple els forats amb
nucleòtids seguint el motlle i la DNA lligasa IV uneix les cadenes.

Immunologia 2020
CEBIO 47
En conclusió, es crea una seqüència a l’atzar entre els segments V, D i J de la cadena pesada
i els segments V i J de les cadenes lleugeres.

Immunologia 2020
CEBIO 48
Tema 8 - Limfòcits B. Els limfòcits B són els únics tipus de cèl·lules que poden produir anticossos i, per tant,
són els mediadors de les respostes immunitàries humorals. Els limfòcits B o cèl·lules B es
desenvolupen a la medul·la òssia i se’n troben, sobre tot, en els fol·licles limfàtics dels teixits
limfàtics secundaris, a la medul·la òssia i a la circulació. En tots tres casos, els limfòcits B que
hi trobem són limfòcits B madurs. A la sang s’hi troben en poques quantitats.
Desenvolupament limfòcits B. El desenvolupament dels limfòcits B s’inicia a la medul·la òssia on, a partir d’una cèl·lula
mare progenitora, es formarà un altre progenitor. Aquest ho serà de la línia limfoide. A partir
del progenitor de la línia limfoide es formaran, entre d’altres cèl·lules, els limfòcits B. Aquests
maduraran a la medul·la.
El desenvolupament dels limfòcits té dues fases. Durant la primera, la fase independent
d’antigen, tindrem la maduració. Durant aquesta es donarà el reordenament dels gens de
les immunoglobulines i una selecció de les mateixes, el que ens permetrà tenir cèl·lules
immunocompetents en el reconeixement d’antígens externs.
Durant la segona fase del desenvolupament dels limfòcits B, la fase dependent d’antigen,
hi ha una activació i una diferenciació. L’activació és el procés en el qual els limfòcits B es
troben circulant i interaccionen amb l’antigen. La diferenciació, en canvi, és el procés
mitjançant el qual els limfòcits B passen a ser cèl·lules plasmàtiques secretores d’anticossos.
Desenvolupament dels limfòcits B a la medul·la òssia.
A mesura que els precursors dels limfòcits B es desenvolupen, van perdent l’expressió de
l’enzim TdT i es traslladen de la cara interna de l’os, cap el centre de la cavitat medul·lar. Allà
esperen als sinus portals fins que surten a la circulació sanguínia. S’estima que la producció
diària de limfòcits B arriba a 5*107.
El desenvolupament a la medul.la òssia depèn de la interacció amb l’estroma medul·lar
(cèl·lules reticulars): molècules d’adhesió, senyalització via c-Kit/SCF i IL7. En humans la IL7
no és estrictament necessària per al desenvolupament dels limfòcits B. En ratolins, en canvi,
sí.
Etapes del desenvolupament dels Limfòcits B.
En el procés de desenvolupament dels limfòcits B el primer que tenim és la proliferació dels
progenitors limfoides a la medul·la òssia. Això ens produirà cèl·lules Pro-B, les quals
només expressen el dímer Ig-α/Ig-β. A les mateixes cèl·lules hi comença a haver una
reordenació de la cadena pesada de les immunoglobulines. Abans d’iniciar la reordenació
de la cadena lleugera, la cèl·lula pro-B en rep un substitut que està format per dos polipèptids:
Vpre-B i λ5. Aquestes dues molècules serveixen per interaccionar amb una cèl·lula
estromal i saber si ha reordenat bé la cadena pesada. En cas que la reordenació s’hagi donat
correctament, no cal fer-la en l’altre cromosoma i es pot començar a reordenar la cadena
lleugera. La unió d’aquest substitut de la cadena lleugera converteix la nostra cèl·lula pro-B
en pre-B gran. La reordenació de la cadena lleugera serà el pas final en la transformació de
cèl·lula pre-B petita en limfòcit B immadur.
Ontogènesi i maduració a la medul·la òssia.
Als òrgans limfoides primaris no hi sol haver antígens exògens. Per tant, els antígens
presents durant el desenvolupament dels limfòcits B són antígens propis o
autoantígens. A l’estadi pre-B, si el pre-BCR uneix un lligand de l’estroma, les cèl·lules
segueixen el seu desenvolupament. És la selecció positiva dels limfòcits B.

Immunologia 2020
CEBIO 49
A l’estadi de limfòcits B immadurs passen un procés de selecció negativa. Els que
reconeixen antigen propi amb avidesa baixa entren en estat d’anèrgia. Els que reconeixen
antigen propi amb avidesa alta poden fer edició del BCR, procés pel que cal una re-expressió
de RAG, o bé poden entrar en apoptosi i ser eliminats.
L’edició del receptor mitjançant un canvi del segment V és el procés de reordenació del
BCR en una cèl·lula B que ja té una reordenació productiva de la cadena H i la L. Al voltant
del 50% del limfòcits B són inicialment autoreactius. L’edició del receptor és el mecanisme
predominant per a mantenir la tolerància i té lloc a nivell de medul·la òssia. Majoritàriament,
l’edició és de VL però també podria ser de VH.
Els limfòcits B que superen la selecció negativa surten de la medul·la òssia com a limfòcits B
immadurs i passen per dues etapes de transició (T1 i T2) abans de convertir-se en cèl·lules
B madures.
Subpoblacions de limfòcits B. Els limfòcits B es separen en B-1, un subgrup de limfòcits B que expressen un repertori reduït
de gens V amb escassa diversitat en la unió i que secreten anticossos IgM que s’uneixen a
antígens independents de T, i en B-2, un subgrup de limfòcits B que expressen un repertori
ampli de gens V.
Els limfòcits B-1 representen una població definida, sobretot en ratolins. En humans
presenten el receptor CD5, són abundants a la pleura i a la cavitat peritoneal. Els limfòcits
B-1 són cèl·lules productores de IgM a l’inici de la resposta immunitària.
Els limfòcits B-2 fol·liculars corresponen a la majoria de limfòcits B madurs, responen a
antígens proteics i depenen de la col·laboració limfòcits T. Els limfòcits B-2 marginals es
troben a la zona marginal de la melsa, responen a antígens provinents de la sang, són
independents de l’ajuda dels limfòcits T i responen ràpid secretant IgM.
Receptor del limfòcit B. El receptor dels limfòcits B és el BCR, el qual consisteix en una immunoglobulina de
membrana associada a les molècules accessòries Ig-α i Ig-β, responsables de la transducció
del senyal. Recordem que Ig-α és la que conté el domini ITAM que permet realitzar aquesta
transducció del senyal.
Immunoglobulines dels BCRs.
Les immunoglobulines que formen els BCR poden ser molt diferents entre els milions
de limfòcits B que existeixen. Aquesta diversitat d’immunoglobulines es pot fer independent
o dependent d’antigen. En la generació de diversitat independent d’antigen s’aprofita la
diversitat de segments gènics V, (D) i J a la línia germinal, la combinació de segments V–(D)–
J, la diversitat per la flexibilitat en la unió dels segments gènics amb l’addició de nucleòtids P
i nucleòtids N i l’associació combinatòria de cadenes pesades i lleugeres. En la generació de
diversitat dependent d’antigen hi pot haver un canvi de la forma de membrana a la forma
soluble de la immunoglobulina, una híper-mutació somàtica o un canvi d’isotip
d’immunoglobulina.
Immunoglobulina de membrana o secretada.
La regió constant de les immunoglobulines té exons opcionals addicionals. Cada
domini de la cadena pesada i la xarnera estan codificats en exons diferents. Al final dels exons
hi ha una seqüència codificant i un lloc de poliadenilació per si la immunoglobulina és
secretada i n’hi ha uns altres per si és de membrana. En cas que la immunoglobulina sigui
secretada, enlloc de regió transmembrana i citoplasmàtica tindrà uns 20 aminoàcids

Immunologia 2020
CEBIO 50
hidrofílics. En cas que sigui de membrana, disposa de 40 aminoàcids que inclouen una regió
transmembrana hidrofòbica i un segment citoplasmàtic molt curt hidrofílic.
Hipermutació somàtica.
La hipermutació somàtica és un dels mecanismes de generació de diversitat dependent
d’antigen que disposen les immunoglobulines. Té lloc als centres germinatius dels òrgans
limfoides secundaris, els canvis es donen sobretot a les regions CDR i té una freqüència
altíssima1 de mutació, de 1/1000 pb del gen V per divisió cel·lular. El procés d’hipermutació
somàtica, on hi intervenen l’enzim AID (citidina deaminasa induïda per activació) i els enzims
DSBR, es troba determinat per citocines i la interacció CD40-CD40L.
La hipermutació somàtica contribueix al procés anomenat maduració per afinitat, que
permetrà la generació d’immunoglobulines amb més afinitat per l’antigen que ha iniciat la
resposta.
Canvi d’isotip.
El procés de canvi d’isotip, un altre dels mecanismes de generació de diversitat entre
immunoglobulines que és dependent d’antigen, consisteix en la substitució de la regió
constant de la cadena pesada per una altra regió constant, de µ ó δ a γ, α ó ε. Mentre es
produeix la resposta immunitària, es manté constant l’especificitat antigènica d’una
immunoglobulina (tot i que en pot canviar l’afinitat). A mesura que la resposta progressa
es canvien les regions constants, que són les que determinen la funció efectora de la
immunoglobulina.
Davant de cada segment C, excepte en el cas del segment Cδ, hi ha una seqüència de DNA
repetitiu o regió switch o Sx. Estan formades per 150 repeticions en tàndem de DNA amb
una elevada freqüència de G/C [(GAGCT)n(GGGGGT)] on n té un valor entre 3 i 7. Això
provoca que tinguin moltes dianes per a l’enzim AID, les quals són els motius WRC (W = A
o T, R = purina, Y = pirimidina).
El canvi d’isotip ve determinat per citocines, les quals determinen quines regions switch
es recombinaran, i per CD40, que indueix l’expressió de l’enzim AID. En el procés de canvi
d’isotip hi intervenen l’enzim AID (citidina deaminasa induïda per activació) i els enzims
DSBR. AID converteix la citosina en uracil, la maquinària de reparació del DNA detecta
l’alteració i comença a reparar. Reparació vol dir la recombinació no homòloga amb la
regió Switch pròxima i l’eliminació del DNA entremig. Les citocines estimulen la
transcripció de les regions Switch i, per tant, s’obre la doble cadena de DNA. Això permet que
l’enzim AID s’uneixi a la cadena de DNA que no s’està transcrivint. A cada recombinació, les
regions constants d’entremig són eliminades del genoma. Les cèl·lules B que fan IgE mai
podran fer canvi d’isotip a IgM, IgD, IgG1-4 o IgA1.
Un limfòcit B sempre genera IgM o IgD de bones a primeres. Després, durant la resposta,
pot fer el canvi d’isotip. El canvi depèn de les citocines produïdes durant la resposta, el que
permet adequar la capacitat efectora dels anticossos a cada situació. Això ens permet
saber si la resposta a un antigen és primària o no.
1 Alta si es compara amb la de la mutació espontània que té lloc en altres gens, que és de 1/108pb

Immunologia 2020
CEBIO 51
Tema 9 - Receptor d’antigen del limfòcit T
Immunoglobulina Una immunoglobulina és un tipus de molècula glucoproteïnica produïda pels limfòcits B que
s’uneix als antígens, sovint amb un alt grau d’especificitat i afinitat. La unitat bàsica de les
immunoglobulines es compon de dues cadenes pesades idèntiques i de dues cadenes
lleugeres idèntiques. Les regions N terminal de les cadenes pesades i lleugeres formen les
zones d’unió a l’antigen, mentre que les regions C terminals constants de les cadenes
pesades interactuen amb altres molècules del sistema immunitari.
L’Anticòs és la forma soluble de la immunoglobulina i és la forma efectora. Els B cell
receptor és una immunoglobulina ancorada a la membrana que actua com a receptor
d'antigen dels limfòcits B.
Les immunoglobulines es generen per recombinació somàtica de segments gènics que
codifiquen per la regió constant i la regió variable, reconeixen antígens solubles, epítops tant
conformacionals com lineals i la composició d'aquestes regions canvia durant la resposta
immunitària, donant lloc a Immunoglobulines amb major afinitat per l'antigen.
Estructura del Receptor d’antigen del limfòcit T. El TCR és el receptor d’antigen de les cèl·lules T. Està format per dues cadenes (α i β o
γ i δ) unides per un pont disulfur. A la seva regió extracel·lular tenen dos dominis de tipus
Immunoglobulina: un domini variable (V) i un domini constant (C). Els TCR sempre es
troben ancorats a la membrana. En conseqüència, sempre tenen una regió transmembrana
i una cua citoplasmàtica curta. Hi ha al voltant de 30.000 molècules de TCR per cada
cèl·lula T. El lloc d’interacció amb l'Antigen està format per la combinació de les regions
variables de cada cadena. A la regió variable també es defineixen les regions determinants
de complementarietat CDR1, CDR2 i CDR3. El TCR té un sol lloc d’unió a l’antigen, és
monovalent. Els TCR tenen una distribució clonal, és a dir, presenten una sola especificitat
per limfòcit.
Organització dels gens del Receptor d’antigen del limfòcit T. L’organització dels segments gènics de la cadena α del TCR és similar a la cadena L de les
Immunoglobulines (VJC) i la de la β, a la cadena H (VDJ) i dóna lloc a un TCR per
recombinació somàtica. L'organització dels segments gènics constants és més senzilla que
a les Immunoglobulines. Cada segment constant codifica per una regió transmembrana i una
de citoplasmàtica i no hi ha isotips com a les Immunoglobulines. El reordenament té lloc
durant la diferenciació dels precursors al timus i usa els mateixos mecanismes que intervenen
en el reordenament de les Immunoglobulines, seqüències RSS i enzims (recombinases,
DSBR).
Mecanisme de reordenament dels gens del Receptor d’antigen del limfòcit T.
El reordenament dels gens del TCR comença per la cadena β. Primer reordena la D-J i
després V-DJ. Com que primer s’expressarà la cadena β, aquesta s’unirà a un substitut de la
cadena α al timus. Si el reordenament de la cadena β és funcional, hi haurà un reordenament
de la cadena α (V-J). En ambdues cadenes, α i β, es produeix el fenomen de l’exclusió
al·lèlica. Seguidament s’uneixen les dues cadenes reordenades i s’analitza que no sigui
autoreactiu. Com en les immunoglobulines, intervenen els mecanismes de combinatòria
d’associació de cadenes, reordenament de segments, unió flexible, addició de nucleòtids... El
que no es dona, i que si que ho fa en immunoglobulines, és la hipermutació somàtica. També
hi ha una altra diferència amb les immunoglobulines: el procés es fa al tim enlloc de a la
medul·la òssia.

Immunologia 2020
CEBIO 52
Hi ha un sol segment gènic Cα i, tot i que hi ha dos segments Cβ, són molt homòlegs i no hi
ha cap diferència funcional coneguda en les proteïnes generades. La possibilitat de produir
un reordenament D-J productiu és de 1/3 i també 1/3 de produir un reordenament V-DJ
productiu. Es pot intentar fer un segon reordenament.
La disposició d’RSS al TCR és diferent que a les immunoglobulines, doncs en segments V
les RSS són de 23 parells de bases, en segments J ho són de 12 i en segments D en són de
12 i 23.
Organització gens del Receptor d’antigen del limfòcit T γδ.
Les cadenes γ i δ són estructuralment semblants a les α i β, respectivament, i
l’organització dels gens que les codifiquen també. Els TCR γ i δ es van descriure després
que els que codifiquen per α i per β, però són filogenèticament més antics.
Els segments gènics que codifiquen per la cadena δ es troben dins el locus dels que
codifiquen per la cadena α. El reordenament de Vα i Jα deleciona Dδ, Jδ i Cδ i, per tant, les
cèl·lules amb TCRαβ no poden expressar un TCR γδ. Hi ha pocs segments V, però la cadena
δ pot incloure dos segments D que afegeixen diversitat igual que en el cas de la cadena β.
Generació diversitat del Receptor d’antigen del limfòcit T.
La diversitat del TCR és funció de la combinació de segments, la flexibilitat de la unió
dels segments (addició de nucleòtids P i N, entre 0-6 nucleòtids) i l'associació de cadenes
diferents (αβ o γδ). Es produeix exclusió al·lèlica a la cadena β però no és absoluta a la
cadena α. No es produeixen canvis al TCR durant la resposta ni a la regió variable ni a la
constant. No existeix una forma secretada, no hi ha canvi d’isotip, ni hipermutació somàtica.
Epítops reconeguts per limfòcits T.
Els limfòcits B i els limfòcits T reconeixen epítops diferents d’un mateix antigen proteic.
L’epítop reconegut pels limfòcits T, per començar, ha d’ésser un agent desnaturalitzat ja que
és un epítop lineal d’entre 8 i 30 aminoàcids. Sovint aquest epítop es troba intern a la
l’agent natiu, el qual sol ser una proteïna, un polisacàrid o un lípid. La unió de l’epítop al TCR
té una kd d’entre 10-5 i 10-7M, el que es considera una baixa afinitat. Això significa que té una
Unió i Desunió lentes. Una vegada unit, el pèptid pot mantenir-se així hores o alguns dies.
El TCR no interacciona amb l’antigen sol, sinó que ho fa amb el complex format per l’antigen
unit a la molècula del MHC ancorada a la membrana de la cèl·lula presentadora d’Antigen.
El MHC uneix una part de l’antigen processat per la cèl·lula presentadora, normalment
fragments de proteïnes (pèptids) resultants de la degradació proteica cel·lular. En una cèl·lula
presentadora d’antigen infectada, la major part de pèptids units a les molècules de MHC
provenen de proteïnes del patogen. La interacció del TCR amb el complex MHC-pèptid
activa el limfòcit (el TCR envia un senyal bioquímic al nucli de la cèl·lula).
La zona d’interacció amb l’antigen coincideix amb la zona N-terminal de les regions variables
on la flexibilitat d’unió entre segments genera més diversitat. Les regions CDR1, CDR2 i
CDR3 de la regió variable de cada cadena del TCR són les que interaccionen amb el complex
MHC-pèptid.
El reconeixement del complex MHC-pèptid el fan les regions CDR1, CDR2 i CDR3 de la
regió variable del TCR. En general, CDR1 reconeix els extrems del pèptid, CDR2 les hèlix
α del MHC i CDR3 la regió central del pèptid.

Immunologia 2020
CEBIO 53
El TCR interaccciona amb la superfície del complex pèptid-MHC. La hipermutació
somàtica podria canviar aminoàcids que interaccionen amb la molècula MHC i es podria
perdre el reconeixement, per això no es produeix.
El TCR està associat amb el complex senyalitzador CD3.
El complex CD3 és indispensable per a la transducció del senyal d’activació que s’inicia
amb el reconeixement d’Antigen. Pertany a la superfamília de les Immunoglobulines format
per 6 cadenes. Les cadenes γ, δ i ε tenen un domini extracel·lular i intracel·lular. La cadena
ζ , en dímer i de pocs aminoàcids extracel·lulars, és pràcticament intracel·lular.
El complex TCR/CD3 el formen una o dues molècules del TCR associada/es a les 6 cadenes
del CD3: una cadena δ, una γ, dues ε i el dímer de cadenes ζ. L’associació és de tipus no
covalent amb el TCR. A la regió transmembrana tenen aminoàcids polars amb càrrega
negativa. A la cua citoplasmàtica, el que tenen són motius ITAM (Immunoreceptor Tyrosine
based Activation Motifs) o motius d’activació d’immunoreceptors basats en tirosines.
Reconeixement d’antigen per part del TCR.
L’afinitat del TCR pel complex pèptid–MHC és baixa (Kd d’entre 10-5 i 10-7M) perquè les
cèl·lules amb TCR d’afinitat alta són eliminades per evitar la reactivitat a estructures
pròpies. Això requereix molècules accessòries que permetin una interacció estable del
limfòcit T amb la cèl·lula presentadora d’antigen. L’estabilitat de la interacció determina el
grau d’activació i les característiques de la resposta una vegada activats.
Molècules accessòries CD4 i CD8.
CD4 té 4 dominis extracel·lulars d’Immunoglobulina, una regió transmembrana i una cua
citoplasmàtica de 38 aminoàcids. CD4 interacciona amb dominis β2 o α2 no polimòrfics de la
cadena MHC de classe II. CD4 estabilitza la interacció del TCR amb el complex pèptid-
MHC de classe II.
CD8 és, predominantment, un heterodímer format per 1 cadena α i una β unides per un pont
disulfur, però també pot ser un homodímer CD8αα. CD8 conté un domini extracel·lular, una
regió transmembrana i una cua citoplasmàtica de 25 aminoàcids. CD8 interacciona amb el
domini α3 d’MHC-I i estabilitza la unió de TCR amb pèptid-MHC de classe I.

Immunologia 2020
CEBIO 54
Tema 10 - Limfòcits T. Els limfòcits T són els component clau de les respostes immunitàries cel·lulars del
sistema immunitari adaptatiu. Els limfòcits T maduren al tim, circulen per la sang, poblen
els teixits limfàtics secundaris i es recluten a les zones perifèriques d’exposició a l’antigen.
Expressen TCRs, que reconeixen fragments peptídics de proteïnes estranyes unides a
molècules pròpies del MHC. Com que totes les cèl·lules d’un individu hereten els mateixos
segments gènics, la població de limfòcits T només reconeix antígens presentats per
MHC propi.
Durant el desenvolupament, a cada limfòcit T es produeix un únic reordenament genètic per
recombinació somàtica. Això aconsegueix que, amb l’enorme diversitat de receptors TCR
possibles, no hi hagi limfòcits T que responguin a allò que és propi. En tant que els gens
reordenats donen lloc a dues cadenes que formen un receptor únic a cada cèl·lula, es
dona una distribució clonal que determina el tipus de limfòcit T.
Origen dels limfòcits T. Els precursors dels limfòcits T es generen a la medul·la òssia i s’eduquen, és a dir, es
diferencien, seleccionen i maduren al tim.
Estructura del timus.
El tim és l’òrgan limfoide primari on es produeix la maduració dels limfòcits T. El tim és
un òrgan situat al mediastí, a l’alçada del cor. És un òrgan que involuciona amb l’edat.
Durant els primers anys de vida, creix en mida i n’assoleix la màxima de greix, còrtex i
medul·la (components del tim) entre els 10 i 15 anys d’edat. A partir d’aleshores, el tim es va
fent cada cop més petit. El teixit adipós (greix) no disminueix tant, però el teixit funcional
casi desapareix. Això no significa que no estiguem generant limfòcits T al llarg de la nostra
vida, sinó que la màxima producció dels mateixos es dona durant l’adolescència.
El tim és un òrgan bilobulat que té una càpsula al voltant i s'invagina formant trabècules.
Aquestes trabècules formen petits lòbuls. En cadascuna de les porcions de teixit funcional
(lòbuls) podem distingir un còrtex al voltant, la medul·la al centre, i una zona corticomedul·lar
que delimita els dos espais. Dins de la medul·la hi ha unes estructures en forma de ceba que
s'anomenen corpuscles de Hassall.
Desenvolupament dels limfòcits T. Les cèl·lules progenitores de cèl·lules T es desenvolupen a la medul·la òssia i migren al Tim.
Allà, el timòcit reorganitza els seus gens de TCR. Al timus es dona una selecció positiva
i negativa de les cèl·lules T immadures que reconeixen el MHC. Aquelles que ho facin rebran
senyals per sobreviure. Aquelles que interaccionin fortament amb el propi antigen seran
eliminades del repertori. Les cèl·lules T naive migren als òrgans limfàtics perifèrics, on
tenen encontre amb antígens estranys i activen els òrgans on es troben. Un cop activats, hi
ha una migració de limfòcits T efectors als llocs d’infecció, on proliferen i acaben amb
aquesta.
El precursor limfoide arriba, des de la medul·la òssia via vasos sanguinis, a les vènules
postcapilars de la zona cortico-medul·lar. Dins el timus, expressa CCR9 i migra cap a la
zona subcapsular del còrtex per quimiotaxi positiva a un gradient de CCL25 produïda per
les cèl·lules epitelials tímiques corticals o cTEC. Com que no expressa TCR ni CD4 ni CD8
s'anomenen timòcits doble negatiu o DN.

Immunologia 2020
CEBIO 55
Timòcits DN.
Els Timòcits DN presenten, en la seva primera fase, receptors c-kit i CD44. C-kit interacciona
amb l’stem cell factor (SCF) i activa segons missatgers que permeten la supervivència,
proliferació i diferenciació dels timòcits. CD44, és un molècula d’adhesió i interacció tant
amb selectines com amb la matriu extracel·lular. Un cop ha entrat en la segona fase, el DN
comença a expressar CD25, una cadena α del receptor d’IL-2. En la tercera fase, els DN3
només expressen CD25, el qual perden en la 4a fase.
Diferenciació a timòcits DP.
El timòcit DN va madurant, el que implica un reordenament genètic del TCR (primer la
cadena β, i després la cadena α), i l’expressió del mateix a la superfície (sempre que estigui
acompanyat de CD3). A més, també expressa els dos coreceptors CD4 i CD8. Per això en
aquesta fase madurativa s'anomena timòcit doble positiu (DP), el qual migra cap al còrtex.
Com que expressen TCR i CD3, poden interaccionar amb els complexes pèptid/MHC-I o
pèptid/MHC-II que expressen les cèl·lules epitelials corticals (cTEC).
Diferenciació dels limfòcits T.
Diferenciació dels limfòcits T per selecció positiva.
Les cèl·lules epitelials corticals (cTEC) expressen molècules del MHC de classe I i de classe
II que presentaran pèptids propis. El timòcit DP començarà a interaccionar amb les cTECs i
amb les MHCI i MHCII que aquestes tenen. Aleshores es produirà la selecció positiva: se
seleccionen aquells timòcits que han rebut un senyal, que han reconegut alguna cosa. Si no
reconeix els MHC, no serveix de res. Per tant, no és tan important el pèptid sinó que
reconegui el MHC.
Si el timòcit DP no contacta amb suficient afinitat, aquest morirà per apoptosi, que és el que
li passarà a la majoria. Si la interacció té suficient afinitat, el limfòcit DP rebrà un senyal de
maduració i continuarà madurant. Aquesta maduració provoca que deixin d'expressar un
dels coreceptors i passin a ser single positive o SP.
Si el senyal l'ha enviat un MHCI deixaran d'expressar CD4. Si el senyal l'ha enviat un MHCII,
deixaran d'expressar CD8. Un cop són SP, comencen a expressar nivells alts de CCR7 i
migren cap a la medul·la tímica, on les cèl·lules epitelials medul·lars (mTEC) produeixen els
seus lligands, CCL19 i CCL21.
Diferenciació dels limfòcits T per selecció negativa.
Si el timòcit SP CD4 o CD8 no interacciona amb els complexos pèptid-MHC de les mTEC o
mDC passats 7 dies, morirà per apoptosi en un procés anomenat deleció clonal. Si
interacciona amb el complex amb una afinitat alta, també morirà per deleció clonal. Si
interacciona amb afinitat intermitja o baixa, el SP seguirà el procés de diferenciació. És a
dir, expressarà el receptor de S1P1 que el farà migrar per quimiotaxi positiva cap als vasos
sanguinis de la zona cortico-medul·lar. Finalment, sortirà cap a la circulació com a limfòcit
TCD4 o TCD8 madur i naive.
Selecció tímica.
Els precursors dels limfòcits T passen de la medul·la òssia al timus. Allà es genera un
repertori de limfòcits T amb TCRs que poden ser o bé capaços de reconèixer qualsevol
pèptid antigènic presentat per molècules MHC pròpies o bé incapaços de reconèixer antígens
propis amb suficient afinitat com per iniciar una resposta en condicions homeostàtiques.
La selecció per AutoImmune Regulator (AIRE) funciona mitjançant un factor de transcripció
expressat, predominantment, a les cèl·lules epitelials medul·lars del timus. AIRE regula la
transcripció de gens específics de teixit en base a la tolerància central cap a autoantígens de

Immunologia 2020
CEBIO 56
teixits perifèrics. Mutacions en aquest gen donen una malaltia autoimmunitària recessiva,
APECED (Autoimmune Polyendocrinopathy-Candidiasis-Ectodermal Dystrophy) que es
manifesta per una autoimmunitat generalitzada a molts òrgans.
L’afinitat de la interacció entre el TCR i el complex pèptid propi-MHC determina la selecció
tímica.
Educació tímica.
A un limfòcit T no l’hem ensenyat a lluitar contra els patògens, simplement a no atacar el que
sigui propi. Si, per aleatorietat, identifiqués un patogen, ho notaria com estrany, ja que no ho
hauria contactat abans. Arran d'això, el limfòcit T l'atacarà.
Desenvolupament limfòcits T reguladors.
Els timòcits SP CD4+ que reconeixen antígens propis amb una afinitat intermitja donen
lloc a limfòcits T reguladors. Aquests limfòcits fan una funció molt important en el control
de la resposta adaptativa. Una vegada activats, regulen la funció dels limfòcits T efectors.
Poden suprimir la seva proliferació i la seva funció efectora.
Maduració dels limfòcits T.
Les cèl·lules que entren al timus proliferen massivament, però només sobreviuen i surten del
timus entre un 2 i un 5% del total. Un 90% moren per negligència (no superen la selecció
positiva) i entre un 5 o un 8% són seleccionats negativament.
Limfòcits Tγδ. Els limfòcits Tγδ són uns limfòcits T amb uns TCRs de cadenes γδ unides per ponts disulfur.
Per tant, aquest TCR és un heterodímer i es troba associat a CD3. La majoria de limfòcits
Tγδ no expressa ni CD4 ni CD8, tot i que una petita proporció sí. Els limfòcits Tγδ reconeixen
estructures moleculars de cèl·lules estressades presentades per molècules del MHC no
clàssiques. Representen un percentatge de cèl·lules variable. En humans, són menys del
5% del total dels limfòcits T que es poden trobar a la perifèria. Són més abundants a les
mucoses i a la pell. En algunes espècies (com els remugants), són la població majoritària fins
i tot a la perifèria.
Tot i la major diversitat potencial del TCRγδ, a humans només s’expressen 6 segments Vγ i
8 de Vδ que siguin funcionals. A més, hi ha aparellaments de cadenes γ i δ molt dominants.
Limfòcits NKT. Els Limfòcits NKT són cèl·lules amb característiques de limfòcits T (TCR i CD3), de cèl·lules
NK (NK1.1) i d’altres. Són seleccionats positivament per reconeixement de lligands
endògens presentats per molècules del MHC no clàssiques com CD1d, expressada a timòcits
DP. Quan s'activen, produeixen ràpidament una gran quantitat de citosines de molts tipus
que activen les cèl·lules presentadores d'antigen i altres cèl·lules de la immunitat adaptativa
com les cèl·lules T i les cèl·lules B). En alguns casos poden incrementar la immunitat cel·lular
i en d'altres poden regular-la o suprimir-la.
Acaben donant lloc a NKTs CD4+, CD8+ o DN. Adquireixen marcadors de fenotip efector
i de memòria abans de sortir del timus. Els marcadors de tipus NK els adquireixen ja fora
del timus, als teixits perifèrics.
Altres poblacions de limfòcits T. Els Limfòcits Tαβ són limfòcits T de mucoses. Són limfòcits intraepitelials (IELs) amb un
TCR limitat i tenen una funció immunoreguladora i de vigilància a les mucoses. És poc
variable, ja que normalment veuen els mateixos antígens. Han de tenir tolerància a la

Immunologia 2020
CEBIO 57
microbiota amb la qual estan en contacte i capacitat de reacció enfront dels agents externs.
També poden ser Limfòcits de la làmina pròpia.
Els limfòcits MAIT, mucosa-associated invariant T cells, compten amb un TCR format per una
cadena TCRα invariant i una de β oligoclonal. Reconeixen metabòlits de la vitamina B
produïts per bacteris i presentats per una molècula del MHC no clàssica, MR1.

Immunologia 2020
CEBIO 58
Tema 11 – Complex Principal d’Histocompatibilitat.
Presentació i reconeixement d’antigen. Els limfòcits T només reconeixen antígens presentats per les molècules del complex
principal d’histocompatibilitat. Aquestes molècules carreguen pèptids que han estat
degradats prèviament per les cèl·lules presentadores d’antigen.
Molècules presentadores d’antigen. Les molècules del Complex Principal d’Histocompatibilitat (MHC) es troben codificades en
alguns gens concrets. Els productes proteics que es troben codificats per aquests gens reben
el nom de molècules MHC o molècules d’histocompatibilitat.
Depenent de l’espècie en la que es trobin, reben un nom diferent. Majoritàriament s’anomenen
amb un prefix depenent de la que provinguin. Per això, les humanes s’anomenen HLA, en
vaques BoLA, ELA en èquids, SLA en porcs, OLA en xais, CLA en cabres i DLA en gossos.
Cas excepcional és el dels ratolins, on reben el nom de H-2. La referència LA respon per
Leukocyte Antigen. Això és perquè les molècules del MHC són majoritàriament molècules
presentadores d’antigen.
Les molècules del MHC es separen en MHC de classe I, MHC de classe II i MHC de classe
III. Aquestes últimes, però, són estructuralment diferents i tenen una funció immunitària que
no està relacionada directament amb la presentació d’antigen.
Estructura del Complex Principal d’Histocompatibilitat de classe I.
El Complex Principal d’Histocompatibilitat de classe I està compost per una cadena α i
per una β-2-microglobulina.
La cadena α és una cadena pesada i glicosilada. En la seva regió extracel·lular hi trobem
els dominis variables α1 i α2 i el domini constant de tipus immunoglobulina α3. Els dominis
α1 i α2 estan formats per una làmina de cadenes β antiparal·leles i una hèlix α. α1 i α2 formen
el lloc d’unió d’antigen, ocupat per pèptids que interaccionen amb les molècules MHC per
mitjà d’enllaços no covalents. Els residus del “terra” i les “parets” d’aquest espai són molt
polimòrfics. Ancorada a la membrana, la cadena α conté una regió transmembrana i una
de citoplasmàtica.
La β-2-microglobulina és una cadena més petita, associada a la cadena α mitjançant
interaccions no covalents. Està formada per un domini de tipus immunoglobulina. La β-2-
microglobulina és idèntica en tots els individus de la mateixa espècie.
Estructura del Complex Principal d’Histocompatibilitat de classe II.
El Complex Principal d’Histocompatibilitat de classe II està compost per una dues
cadenes, una α i una β. Cadascuna d’aquestes cadenes té 2 dominis extracel·lulars
anomenats α1 i α2 i β1 i β2, un domini transmembrana i una cua citoplasmàtica. Les
cadenes α i β es troben associades mitjançant interaccions no covalents. El domini β1 es
plega mitjançant un pont disulfur, mentre que el domini α1 és més obert.
Els dominis α2 i β2 són de tipus immunoglobulina. α1 i β1 estan formats per 4 cadenes β
antiparal·leles i una hèlix α cadascun. El conjunt forma el lloc d’unió a l’antigen. α1 i β1 són
els dominis més polimòrfics. El domini transmembrana de cada cadena té una longitud
aproximada de 30 aminoàcids de naturalesa hidrofòbica. El domini citoplasmàtic, en canvi,
conté entre 10 i 16 aminoàcids i és de naturalesa hidrofílica.

Immunologia 2020
CEBIO 59
Interacció Complex Principal d’Histocompatibilitat – Pèptid.
Hi ha una diferència important entre les molècules MHC de classe I i de classe II. Les
molècules de MHC de classe I tenen els extrems del lloc d’unió a l’antigen tancats, el que
suposa una limitació en la llargada dels pèptids que s’hi poden unir. Aquesta limitació es
tradueix en què només es poden presentar de forma adient pèptids de l’ordre de 9
aminoàcids.
En canvi, les molècules de MHC de classe II tenen un lloc d’unió d’antigen amb els extrems
més oberts, el que permet que s’hi acomodin pèptids amb una llargada d’entre 10 i 20
aminoàcids. Veient l’estructura de la molècula amb tots els residus i totes les cadenes
laterals veiem que al lloc d’unió d’antigen hi queda un solc, que és on s’acomoda el pèptid.
Aquests llocs són polivalents i tenen una afinitat més aviat baixa.
Lloc d’unió a l’antigen.
Per tant, el lloc d’unió a l’antigen té una estructura on les cadenes β antiparal·leles formen la
base del lloc d’unió i les dues hèlixs α limiten l’espai del mateix.
Aquest complexes pèptid-MHC han d’interaccionar amb els receptors d’antigen dels
limfòcits T, els TCR. Per tant, hi haurà una sèrie de cadenes laterals de residus d’aminoàcids
del pèptid que interaccionaran amb el TCR i hi haurà unes altres cadenes laterals de residus
d’aminoàcids del pèptid que l’ancoraran a la molècula del Complex Principal
d’Histocompatibilitat. Aquests residus s’anomenen residus d’ancoratge i s’uneixen a les
butxaques del Complex Principal d’Histocompatibilitat.
Molècules del Complex Principal d’Histocompatibilitat clàssiques i no clàssiques.
Les molècules del MHC es separen en MHC de classe I, MHC de classe II i MHC de classe
III. Aquestes últimes, però, són estructuralment diferents i tenen una funció immunitària que
no està relacionada directament amb la presentació d’antigen. Així i tot, les molècules del
MHC de classe III es troben en el mateix locus.
D’entre les molècules del MHC de classe I i de classe II parlem de molècules clàssiques i
de molècules no clàssiques. Les molècules del MHC clàssiques són aquelles que actuen
com a presentadores d’antígens derivats del processament de proteïnes, és a dir, que
presenten pèptids. Les molècules de MHC de classe I són molècules molt polimòrfiques.
Les molècules del MHC no clàssiques tenen la mateixa estructura però la seva funció principal
no és la presentació d’antígens peptídics als limfòcits T. Està demostrat que les molècules
del MHC no clàssiques poden presentar altres tipus d’antigen o poden ser molècules que
participen en la presentació d’antigen sense ser elles qui ho facin directament. Les molècules
del MHC no clàssiques són molt menys polimòrfiques.
Organització dels gens de les molècules MHC humanes.
S’han descrit, en humans, 3 grans tipus de molècules del MHC de classe I clàssiques. Es
denominen afegint una lletra A, B o C al final de la seva nominació. Per tant, parlem de HLA-
A, HLA-B i HLA-C. Les molècules MHC de classe I no clàssiques en humans s’anomenen
HLA-E, HLA-F, HLA-G i MICA.
Les molècules MHC de classe II clàssiques s’agrupen sota els termes HLA-DP, HLA-DQ i
HLA-DR. Les molècules MHC de classe II no clàssiques, en canvi, s’agrupen sota els noms
DM i DOB.
Complex HLA

Immunologia 2020
CEBIO 60
Classe del MHC II I
Terme DP DQ DR B C A
Productes HLA-DP
αβ HLA-DQ
αβ HLA-DR
αβ HLA-B HLA-C HLA-A
Organització dels gens de les molècules MHC de classe I.
Les molècules de MHC de classe I clàssiques es troben codificades o bé pel gen HLA-A, pel
HLA-B o bé pel HLA-C. Cada gen codifica per una seqüència reguladora que permet que
la seva expressió es pugui induir o limitar. Hi ha una seqüència líder i després una sèrie
d’exons que codifiquen per cadascun dels dominis que formen la cadena α de la molècula
del MHC. Seguidament hi ha uns exons que codifiquen per la regió transmembrana i la cua
citoplasmàtica.
La cadena β-2-microglobulina està codificada en un altre cromosoma. El 15 en humans i el 2
en ratolins.
Organització dels gens de les molècules MHC de classe II.
Pel que fa a les molècules de MHC de classe II clàssiques, hi ha 3 gens: HLA-DP, HLA-DQ i
HLA-DR. En aquest cas trobem tant les seqüències que codifiquen per la cadena α com
les que codifiquen per la cadena β al mateix cromosoma, el 6. Cada gen codifica per una
seqüència reguladora que permet que la seva expressió es pugui induir o limitar. Hi ha una
seqüència líder i després una sèrie d’exons que codifiquen per cadascun dels dominis que
formen la cadena α de la molècula del MHC. Seguidament hi ha uns exons que codifiquen
per la regió transmembrana i la cua citoplasmàtica. En el cas de les cadenes β,
l’organització gènica és la mateixa.
Codominància del Complex Principal d’Histocompatibilitat. Els gens del Complex Principal d’Histocompatibilitat són codominants. Partint de la
premissa que la codominància és l’expressió simultània de gens homòlegs provinents
d’ambdues dotacions cromosòmiques, podem afirmar que, en el cas de l’MHC, una cèl·lula
expressarà dues molècules MHC de cada tipus. En aquest context, tipus de molècula MHC
fa referència a aquelles que es troben codificades en el cromosoma matern i en el patern, on
hi ha 2 al·lels de cada gen HLA-A, HLA-B, HLA-C...
Evidentment, aquesta codominància és un avantatge per la comunitat i per l’individu
perquè permet presentar una major diversitat d’antígens i així respondre a la seva presència.
Genotip, Fenotip, Haplotip i Desequilibri de lligament del MHC. El genotip d’un individu és la seva composició gènica, independentment de la seva expressió.
En el cas aplicat del HLA, el genotip ens informa de no només els al·lels, sinó que també ho
fa de la seva situació als cromosomes. Per posar un exemple, imaginem el següent genotip:
• A1, B8, Cw1, DR3, DQ2, DP1 / A2, B15, Cw2, DR4, DQ3, DP2.
L’haplotip d’un individu tan sols presenta una petita diferència respecte el genotip, ja que és
el conjunt de gens lligats entre si i heretats en bloc en un mateix cromosoma. Seguint
l’exemple anterior podríem definir dos haplotips malgrat haver-n’hi molts més de possibles:
• A1, B8, Cw1, DR3, DQ2, DP1.

Immunologia 2020
CEBIO 61
• A2, B15, Cw2, DR4, DQ3, DP2.
El fenotip, aplicat també al HLA, és el conjunt d’al·lels dels diferents loci de HLA que estan
expressats a les cèl·lules d’un individu. Seguint encara amb el mateix exemple i recordant
que el MHC té codominància, podríem definir el següent fenotip:
• A1,2; B8,15; Cw1,2; DR3,4; DQ2,3; DP1,2.
Finalment, el desequilibri de lligament és la tendència de certs al·lels a associar-se entre
sí, formant haplotips més freqüents dels que es produirien per associació aleatòria. Per
exemple. La freqüència de l’haplotip següent en la població caucàsica és molt més gran del
que s’esperaria per la combinació aleatòria dels 4 al·lels:
• A1, B8, DR3, DQ2.
Polimorfisme del Complex Principal d’Histocompatibilitat. Els gens de l’MCH són molt polimòrfics, doncs hi ha una gran variació genètica dins d’una
població. Dit d’una altra manera, hi ha diverses formes al·lèliques per un mateix gen.
La manera com es descriuen és que cada al·lel rep el nom de la molècula HLA seguida d’un
número. Per exemple, HLA-B27 seria l’al·lel 27 d’una molècula MHC de classe I HLA-B i seria
seguida per un altre número, que defineix quin dels 40 al·lels que codifiquen per HLA-B27 és
el de l’individu. Per tant, quan parlem del HLA-B27 de l’individu en qüestió estarem parlant
del HLA-B2705 o del HLA-B2736, per exemple.
Cada al·lel està distribuit a la població amb una freqüència característica que depèn del
grup ètnic. És el resultat de la pressió evolutiva de la selecció natural.
Aquest polimorfismes no són iguals per totes les cadenes o molècules MHC. A les molècules
MHC de classe II, la cadena β és més polimòrfica que la cadena α. De totes les molècules
MHC de classe II, HLA-DR és més polimòrfica que HLA-DP o que HLA-DQ.
En el cas de les molècules MHC de classe I, les que són clàssiques són molt polimòrfiques,
mentre que les no clàssiques ho són molt poc.
Residus polimòrfics a les molècules MHC de classe I i classe II.
Els polimorfismes es troben principalment al lloc d’unió a l’antigen, sigui als residus que
formen les hèlix α o els que formen les cadenes β antiparal·leles. Si seqüenciem les proteïnes
de les molècules del MHC, comprovarem que les principals diferències entre al·lels i individus
es troben en la zona d’unió a pèptid.
Avantatge evolutiu dels polimorfismes del Complex Principal d’Histocompatibilitat.
Aquest polimorfisme és un avantatge evolutiu, doncs la majoria dels individus som
heterozigots per a tots els loci i, per tant, disposem d’una bona diversitat de tipus de molècules
HLA que permeten presentar les proteïnes dels patògens.
A nivell poblacional, el polimorfisme suposa que hi hagi una enorme diversitat de
molècules HLA disponibles. És improbable que un patogen aconsegueixi modificar les
seves proteïnes fins al punt que no puguin ser presentades per les molècules de HLA de cap
individu. Tot això fa improbable que sorgeixi un microorganisme que elimini l’espècie,
tot i que pot passar en poblacions aïllades i molt consanguínies.
Per tant, les molècules MHC són poligèniques, tenim diferents gens que codifiquen per
molècules MHC de classe I i per molècules MHC de classe II, polimòrfiques i codominants,
cada cèl·lula expressa dues molècules HLA de cada tipus.

Immunologia 2020
CEBIO 62
Molècules del Complex Principal d’Histocompatibilitat clàssiques i no clàssiques
Expressió de molècules HLA clàssiques.
Però no totes les cèl·lules expressen el MHC amb la mateix grau d’expressió. Tot i que
les molècules MHC de classe I són d’expressió constitutiva a totes les cèl·lules nucleades de
tots els teixits, hi ha diferents nivells d’expressió entre ells. L’expressió de MHC-I augmenta
amb l’activitat de citocines com IFNγ, IFNα i IFNβ, que són les que actuen sobre les regions
reguladores dels gens que codifiquen per aquestes molècules MHC.
El cas de les molècules MHC de classe II la qüestió és diferent, doncs l’expressió està
restringida a les cèl·lules presentadores d’antigen professionals però es pot induir en
alguns tipus cel·lulars amb citocines perquè hi ha factors activadors com CIITA a la regió
promotora.
Molècules no clàssiques del Complex Principal d’Histocompatibilitat.
Molècules no clàssiques del Complex Principal d’Histocompatibilitat de classe I.
Les molècules no clàssiques es consideren molècules del MHC malgrat no realitzar les
mateixes funcions. Tenen una organització gènica i una estructura proteica molt similar als
nivells primari, terciari i quaternari amb les molècules MHC de classe I. Com aquestes, tenen
una cadena α associada a la β-2-microglobulina. Les molècules no clàssiques són, però, poc
polimòrfiques i tenen un patró d’expressió més restringit. N’hi ha varis tipus.
Les molècules HLA-E uneixen pèptids de la seqüència senyal de les molècules del MHC.
Amb un reconeixement per cèl·lules NK a través del receptor inhibidor NKG2A, les molècules
HLA-E tenen una expressió no restringida i no són polimòrfiques.
Les molècules HLA-G uneixen pèptids, com les HLA-E i les molècules HLA clàssiques. Les
HLA-G tenen una expressió restringida a trofoblasts fetals i a cèl·lules de l’epiteli tímic, el
que significa que pot intervenir en el procés de selecció tímica dels limfòcits T. Aquestes
molècules tenen polimorfisme limitat però hi ha diferents isoformes. Algunes d’aquestes són
de membrana i d’altres són solubles i generades per splicing alternatiu. Les molècules HLA-
G són reconegudes per receptors inhibidors com ILT2 i ILT4 de les cèl·lules NK i Tγδ. Arran
del seu patró d’expressió, aquestes són unes molècules molt importants per la tolerància
materno-filial.
Les molècules HLA-F són molècules MHC de localització intracel·lular i d’expressió
restringida a l’amígdala, la melsa i el tim. La veritat és que no es coneixen gaire les funcions
d’aquestes molècules del MHC.
Finalment, les molècules MICA i MICB són un tipus de molècules MHC que no uneixen
pèptids perquè tenen el lloc d’unió d’antigen tancat. Aquestes molècules tampoc s’associen
a la cadena β-2-microglobulina, el que les fa més diferents encara que la resta, però sí que
són polimòrfiques. Les MICA i les MICB tenen una expressió restringida a les cèl·lules
epitelials, als fibroblasts, els queratinòcits i a les cèl·lules endotelials. Són molècules que
s’expressen en situacions d’estrès i són reconegudes per les cèl·lules NK a través del receptor
activador NKG2D i per Tγδ.
També són molècules del MHC de classe I no clàssiques les molècules de CD1. Aquestes no
estan codificades al locus MHC. Se’n coneixen 5 gens en humans i codifiquen per les
molècules de CD1A, CD1B, CD1C, CD1D i CD1E; totes al cromosoma 1. Aquestes molècules
de CD1 no són polimòrfiques i tenen una distribució diferent entre elles. No s’expressen en
els mateixos tipus cel·lulars ni en les mateixes localitzacions cel·lulars. El més característic
és que presenten antígens de base lipídica que reconeixen els limfòcits NKT, limfòcits T

Immunologia 2020
CEBIO 63
que expressen un receptor d’antigen T i receptors de cèl·lules NK. El fet que presentin
antígens de base lipídica implica que han de tenir molts residus hidrofòbics en els seus
llocs d’unió a l’antigen.
Molècules no clàssiques del Complex Principal d’Histocompatibilitat de classe II.
Pel que fa a les molècules del MHC de classe II no clàssiques, són molècules no
polimòrfiques i d’expressió restringida com les MHC II clàssiques. Aquestes molècules es
localitzen exclusivament a la membrana de compartiments intracel·lulars. No s’expressen
a la membrana citoplasmàtica. Com en el cas de les Molècules no clàssiques del Complex
Principal d’Histocompatibilitat de classe I, tenim diferents tipus. En destaquen dos.
Les HLA-DM són molècules amb una seqüència i estructura terciària intermèdia entre classe
I i classe II i té un lloc d’unió d’antigen pràcticament tancat. La seva expressió és induïble
amb l’interferó γ. La seva funció és facilitar la càrrega de pèptids d’alta afinitat a les molècules
del MHC-II i estabilitzar les molècules del MHC-II inestables fins trobar un pèptid adient.
Les HLA-DO, al contrari que les HLA-DM, no s’indueixen amb IFNγ. La seva funció és
modular la de HLA-DM als limfòcits B, a les cèl·lules epitelials tímiques i a algunes
subpoblacions de les cèl·lules dendrítiques.
També es consideren molècules del MHC de classe II no clàssiques aquelles que estan
relacionades amb la via d’MHC de classe I com TAP, un transportador de pèptids, Tapasina,
una xaperona, i LMP2 i LMP7, unes subunitats de l’immunoproteasoma.
Co-reconeixement de pèptid i Complex Principal d’Histocompatibilitat. El TCR d’un limfòcit T només reconeix pèptids presentats per molècules del MHC
pròpies. Un limfòcit T específic per un pèptid X d’un individu no reconeixerà el pèptid X si és
presentat per una molècula del MHC d’un al·lel diferent i tampoc si és un pèptid diferent Y
presentat per la molècula del MHC pròpia. S’anomena restricció MHC perquè restringeix o
limita l’habilitat del limfòcit T de reconèixer l’antigen.
Efecte del Complex Principal d’Histocompatibilitat en el transplantament. En els casos de transplantament, els empelts s’accepten sense problemes entre
individus genèticament idèntics. El rebuig d’un empelt és una resposta dependent del
reconeixement dels limfòcits T, tot i que també hi intervenen anticossos i el sistema del
complement. La regió MHC, especialment els gens de les molècules de classe II i (en menor
grau) els de classe I, conté els gens més importants en la definició del rebuig.
Hi ha dues maneres perquè les molècules HLA provoquin rebuig en el transplantament
d’òrgans. Una opció és que els limfòcits T del receptor siguin activats per cèl·lules
presentadores d’antigen del donant de complexes pèptid-MHC. L’altre opció és que les
ACPs del receptor poden adquirir, processar i presentar proteïnes del donant i activar
els limfòcits T del receptor. El primer cas s’estaria donant un reconeixement directe i en el
segon, un d’indirecte.
El 1959, Joseph Murray va trasplantar amb èxit un ronyó entre germans bessons fraterns i va
funcionar durant 20 anys sense fer servir immunosupressors. Això és perquè la intensitat del
rebuig és proporcional a la disparitat genètica.
Un donant es considera compatible si es comparteixen els al·lels HLA-A, HLA-B i HLA-DRB1.
Un conjunt de 3 dels al·lels s’hereta de la mare i els altres 3 del pare. En conseqüència, per
tal que un donant es consideri compatible amb el seu receptor han de coincidir 6 al·lels. Es
requereix que un mínim de 5 dels 6 al·lels coincideixin per poder fer donació entre adults.

Immunologia 2020
CEBIO 64
Efecte de les molècules HLA i la susceptibilitat a malalties. Hi ha una llarga llista de malalties que s’associen a l’expressió de determinats al·lels HLA.
Aquests al·lels determinen la capacitat de resposta del sistema immunitari perquè
intervenen en la generació del repertori de limfòcits T en el procés de selecció tímica i
determina quins pèptids seran presentats eficientment.
És possible que haplotips que han estat útils en algun moment de l’evolució per respondre a
determinats microorganismes quedessin seleccionats però pot ser que siguin aquests
mateixos haplotips els que ara, en un ambient diferent, afavoreixin les respostes
autoimmunitàries.
Altres aspectes de les molècules HLA. Un altre aspecte de les molècules HLA és que sembla que la tria de parella en humans sembla
ser depenent del MHC. També pot ser que això passi en primats no humans.

Immunologia 2020
CEBIO 65
Temes 12 i 13 – Processament i presentació d’antigen. El processament d’un antigen és el conjunt de processos degradadors pels que els antígens
d’un patogen es redueixen a pèptids prou petits com perquè es puguin unir a les molècules
del sistema d’histocompatibilitat.
Les molècules del MHC clàssiques prenen els pèptids resultants de la degradació i els
presenten a la superfície de la cèl·lula presentadora d’antigen. Aquesta exposició de
fragments peptídics units a molècules del MHC permet la interacció amb els Receptors
d’antigen dels limfòcits T.
Per tal que es pugui donar el processament de l’antigen per part de la cèl·lula presentadora
d’antigen cal que aquesta sigui capaç de captar-lo. En cas que aquest antigen provingui d’un
element extern com un patogen, la integració es pot donar per fagocitosi o per endocitosi.
Si la molècula presentadora d’antigen és una MHC de classe II, podrà presentar l’antigen a
un limfòcit T amb el co-receptor CD4. Si el que passa és que la cèl·lula es troba infectada
per un virus, per exemple, l’exposició dels pèptids resultants de la degradació de proteïnes
citosòliques es farà per molècules del MHC de classe I. Per tant, podran presentar aquests
antígens a limfòcits T que porten el co-receptor CD8.
Vies de presentació d’antigen. Hi ha dues grans vies de presentació d’antigen. Una és la que ve donada per la
presentació d’antigen per part de molècules del MHC de classe I i l’altre, la que ve donada
per la presentació d’antigen per part de molècules del MHC de classe II. Això distingeix les
vies de presentació d’antigen.
Les molècules del MHC de classe I presentaran antígens endògens, propis de la cèl·lula,
o que es troben al citoplasma de la cèl·lula. Aquesta presentació, a més, la faran a limfòcits
TCD8, que expressen els co-receptors CD8. Les molècules de MHC de classe II
presentaran antígens exògens o que entrin a la cèl·lula per fagocitosi o endocitosi. Aquesta
presentació d’antigen, a més, la faran a limfòcits TCD4, que expressen els co-receptors CD4.
Això significa que hi ha tota una sèrie de mecanismes no compartits entre aquestes dues vies
de presentació d’antigen. Aquestes diferències causaran tipus de respostes immunitàries
diferents.
Formes de presentació dels patògens. La determinació de quina molècula del MHC presentarà l’antigen ve per part d’aquest,
doncs els patògens o els seus productes es poden trobar tant al citosol com en compartiments
cel·lulars.
Si és un antigen citosòlic, com els virus que entren al citoplasma de les cèl·lules, aquests
seran presentats per molècules del MHC de classe I, el que suposa l’activació de la resposta
dels limfòcits T amb co-receptor CD8. El resultat d’aquesta interacció entre la cèl·lula
presentadora d’antigen i els limfòcits TCD8 serà l’eliminació de la cèl·lula infectada arran
de l’alliberament de la maquinària citotòxica del limfòcit T.
D’altra banda, si és un antigen fagocitat o endocitat i entra a la cèl·lula en les seves
vesícules endocítiques, l’antigen serà processat i presentat per molècules del MHC de
classe II a limfòcits TCD4. Això suposarà la seva activació, la qual provoca una producció
de citocines. Les citocines activen la funció bactericida de la cèl·lula fagocítica o la cèl·lula
B i provoquen l’expansió de limfòcits TCD4.

Immunologia 2020
CEBIO 66
Processament i Presentació d’antigen per molècules del MHC de classe I. Els antígens que es presenten per molècules del MHC de classe I són propis de la cèl·lula o
que es troben al seu citoplasma. Són processats per un complex enzimàtic anomenat
proteasoma, el qual genera pèptids d’entre 9 i 16 aminoàcids que són transportats per la
proteïna transportadora TAP fins al reticle endoplasmàtic.
El proteasoma és un complex proteolític amb expressió constitutiva a totes les cèl·lules i
que degrada proteïnes citoplasmàtiques. Està format per dos anells centrals anomenats β
i 2 anells laterals α de 7 subunitats cadascun. Aquests anells laterals es troben ordenats
en forma de cilindre. Als extrems del proteasoma hi trobem subunitats reguladores.
Tres subunitats dels anells centrals β del proteasoma tenen activitat proteasa: β1, β2 i β5.
Són subunitats que trenquen enllaços peptídics depenent d’ATP. Les proteïnes
ubiquitinades entren al canal del proteasoma i en surten fragments peptídics d’entre 9 i 16
aminoàcids.
A les cèl·lules presentadores d’antigen, quan hi ha Interferó γ, s’expressa
l’immunoproteasoma. En aquest, les subunitats β1, 2 i 5 són substituïdes per subunitats β1i,
2i i 5i, les quals tenen una especificitat enzimàtica diferent. L’immunoproteasoma genera
pèptids amb aminoàcids C-terminals hidrofòbics o bàsics que són presentats més
eficientment per molècules del MHC de classe I.
Hi ha una altra forma del proteasoma expressat per les cèl·lules epitelials tímiques que
col·labora a generar un altre repertori de pèptids. Aquest proteasoma, anomenat
timoproteasoma, genera els pèptids diferents que la resta de cèl·lules arran de la seva
subunitat β5t, també diferent.
Els pèptids sortints del proteosoma arriben després a la proteïna TAP (Transporter
Associated with Antigen Processing). Aquesta és una proteïna que transporta els pèptids
generats pel proteasoma fins al reticle endoplasmàtic, on hi ha les molècules del MHC de
classe I. Aquesta proteïna TAP és un heterodímer de membrana format per dues subunitats:
TAP1 i TAP2. Es troben ancorades a la membrana del reticle endoplasmàtic.
El transport de pèptids a través de TAP és depenent d’ATP, però té més afinitat per
pèptids d’entre 9 i 16 aminoàcids amb un extrem C-terminal bàsic o hidrofòbic. Per tant, es
pot veure que TAP optimitza el transport de pèptids que s’uneixen de manera preferent
a les molècules del MHC de classe I.
Al reticle endoplasmàtic, on els pèptids sortints del proteosoma arriben després de ser
translocats per la proteïna TAP, actuen les aminopeptidases ERAP1 i ERP-2. Aquestes
aminopeptidases degraden l’extrem N-terminal del pèptid per generar-ne uns d’entre 8 i 9
aminoàcids, la mida òptima per interaccionar amb el lloc d’unió de pèptids de les molècules
del MHC de classe I.
Després d’aquest tall, els pèptids que tinguin els aminoàcids adients a les posicions
d’ancoratge es podran carregar a les molècules del MHC de classe I. Aquesta “càrrega”
farà que les molècules es tornin prou estables per ser transportades pel sistema de
vesícules de la cèl·lula fins a la seva membrana citoplasmàtica. Un cop allà, es presentaran
al limfòcit TCD8.
Aquestes molècules del MHC de classe I es sintetitzen al reticle endoplasmàtic. Primer es
genera la cadena α. Aquesta s’associa a una xaperona, la calnexina, per mantenir-se estable
fins que s’uneixi a la cadena β-2-microglobulina. En el moment en què hi ha aquesta unió
s’allibera la calnexina. Ja estaria formada, doncs, la molècula del MHC de classe I. El

Immunologia 2020
CEBIO 67
problema rau en què les molècules del MHC de classe I necessiten que hi hagi un pèptid
al lloc d’unió a l’antigen perquè siguin estables. Com que s’acaben de formar i no hi ha
cap pèptid associat, ràpidament es constitueix el complex de càrrega de pèptids.
El complex de càrrega de pèptids és una estructura formada per la calreticulina, la
tiolreductasa ERp57 i la tapasina. En aquest complex, ERp57 s’associa no covalentment a
la calreticulina per estabilitzar la seva interacció amb la molècula del MHC de classe I.
Tapasina, en canvi, connecta la proteïna transportadora TAP amb la resta del complex
al unir-se a ERp57 per mitjà d’un pont disulfur. La funció de la tapasina és seleccionar els
pèptids que s’uneixin amb més estabilitat a la molècula del MHC de classe I.
L’estabilització de la molècula de MHC de classe I per part del complex de càrrega de pèptid
permet que hi hagi una interacció al lloc d’unió d’antigen de la mateixa amb un pèptid. Això
la estabilitzarà, el que li permetrà sortir del reticle i ser expressada a la membrana plasmàtica.
Allà, l’antigen serà exposat pel reconeixement pels Receptors de les Cèl·lules T.
Processament i Presentació d’antigen per molècules del MHC de classe II. La via de presentació d’antigen per molècules de MHC de classe II és bastant diferent de la
de presentació d’antigen per part de molècules del MHC de classe I. Per començar, els
antígens que es presenten són exògens que s’han internalitzat i que entren a una sèrie de
compartiments intracel·lulars que s’anomenen endosomes. En aquests, les proteïnes són
degradades i arriben fins a uns compartiments anomenats compartiments MIIC, on hi ha
una gran expressió de molècules del MHC de classe II. Les molècules carregaran els pèptids
i migraran amb ells fins a la membrana citoplasmàtica, on s’exposaran al reconeixement dels
receptors dels limfòcits TCD4.
Els mecanismes d’entrada dels antígens exògens en una cèl·lula, en general, s’anomenen
endocitosi o fagocitosi. Si són partícules molt grans és quan reben el nom de fagocitosi o
macropinocitosi. Quan són més petits pot ser que siguin endocitosis mediades per
receptors. Aquests poden ser de lectines, del sistema del complement... El resultat d’aquesta
interacció és la invaginació de la membrana i la formació dels compartiments citoplasmàtics
anomenats endosomes.
El conjunt de compartiments intracel·lulars de pH cada vegada més baix, començant pels
endosomes primerencs, seguit dels endosomes tardans i els lisosomes per acabar sent
compartiments MIIC és el que es coneix com a via endocítica.
Els compartiments de la via endocítica contenen enzims lítics que actuen a diferents
pHs. Als lisosomes hi ha hidrolases dependents d’àcid com proteases, nucleases,
glicosidases, lipases, fosfolipases i fosfatases. Aquest conjunt d’enzims degraden proteïnes
tot generant pèptids d’entre 13 i 18 aminoàcids que són carregats a les molècules del MHC
de classe II.
Els enzims que hi ha en aquest compartiments no són els mateixos que hi ha en cèl·lules
epitelials tímiques corticals, cèl·lules epitelials tímiques medul·lars o cèl·lules dendrítiques.
Això vol dir que, també a partir d’aquesta via, es generen repertoris de pèptids diferents a
partir d’una mateixa proteïna depenent de la cèl·lula que l’estigui presentant.
Tant la cadena α com la cadena β de les molècules del MHC de classe II es generen al
reticle endoplasmàtic. En el mateix, les cadenes s’associen a una cadena invariant, la qual
està formada pels dominis de trimerització i endocític i una part central a la que anomenem
CLIP. Quan s’han format tres d’aquests complexes, el domini de trimerització forma el que

Immunologia 2020
CEBIO 68
s’anomenen Nonàmers, supercomplexos de 3 cadenes invariables, 3 cadenes α i 3 cadenes
β. Aquest supercomplex, a través dels dominis endocítics, passarà al golgi i d’allà als
compartiments de la via endocítica, ja siguin els endolisosomes o els lisosomes.
En aquests compartiments el pH és molt àcid i hi ha unes proteases, les catepsines. Les
mateixes degraden la cadena invariant, deixant el fragment CLIP ocupant el lloc d’unió al
pèptid de la molècula del MHC de classe II com porten fent tota l’estona. Això significa que
ni al reticle endoplasmàtic ni a l’aparell de golgi ni als compartiments lisosomals aquestes
molècules poden incorporar antigen perquè encara tenen el fragment CLIP unit al seu lloc
d’unió a l’antigen.
Els antígens que entren per endocitosi són degradats en els compartiments
endolisosomals i són convertits en pèptids.
El dímer de classe II amb el pèptid CLIP o amb pèptids de baixa afinitat és inestable i no es
transporta eficientment a la superfície cel·lular fins que uneix els pèptids òptims que
seran presentats a limfòcits TCD4. Això ho fa en els compartiments MIIC catalitzat per la
HLA-DM.
HLA-DM és una molècula del MHC de classe II no clàssica que es troba als
compartiments endocítics MIIC. Aquesta molècula té el solc d’unió a antigen pràcticament
tancat, pel que no en pot unir. La seva expressió és induïble amb l’interferó γ. La seva
funció és doble. Per un costat, actua com a xaperona, ja que estabilitza els dímers de les
molècules del MHC de classe II fins que troben un pèptid adient. Per l’altre banda, tenen
una funció catalitzadora de l’intercanvi de CLIP per pèptids d’alta afinitat en el lloc d’unió
d’antigen de les molècules del MHC de classe II.
HLA-DO és una altra molècula del MHC de classe II que es localitza als lisosomes o als
compartiments MIIC però que no és induïble per l’interferó γ. La seva funció és modular el
comportament de l’HLA-DM a les cèl·lules B, a les cèl·lules epitelials tímiques i a algunes
subpoblacions de cèl·lules dendrítiques.
Vies alternatives de presentació d’antigen. Els antígens exògens són internalitzats per les cèl·lules presentadores d’antigen i són
degradats als compartiments endocítics, el que servirà per fer una associació peptídica
amb les molècules del MHC de classe II. Això permetrà que es faci una presentació
d’antigen als limfòcits TCD4. Els patògens intracel·lulars o propis de les cèl·lules
presentadores d’antigen es degraden al citoplasma per associar-se a molècules MHC de
classe I, el que portarà a l’activació dels limfòcits TCD8.
El que passa és que aquestes dues vies de presentació d’antigen no expliquen com es duu a
terme la resposta immunològica quan un virus no infecta una cèl·lula presentadora d’antígens
professional, ni quan un tumor no es genera en cèl·lules presentadores d’antigen
professionals ni tampoc quan s’injecta una vacuna.
Via de la Crospresentació.
S’ha descrit que, si les cèl·lules presentadores d’antigen, per tal de poder presentar-lo als
limfòcits TCD8, l’han d’internalitzar. Això significa captar-lo de l’exterior i que passi a la via de
presentació per MHC-I.
Hi ha una via alternativa de presentació d’antigen que es va anomenar crospresentació.
Aquest procés permet que la presentació d’antígens exògens a limfòcits TCD8 per
molècules del MHC-I. En aquest procés es degraden els antígens en els compartiments
endocítics i després s’associen a molècules del MHC de classe I.

Immunologia 2020
CEBIO 69
Per tal que això sigui possible, els antígens extracel·lulars, enlloc de ser transportats als
lisosomes on es degradaran ràpidament i es carregaran a molècules del MHC-II, passen a la
via de presentació per molècules del MHC de classe I des dels endosomes primerencs. S’ha
descrit més d’una opció per aquest mecanisme.
Per començar, una de les opcions que tenen els pèptids per passar a la via de presentació
del MHC de classe I és aprofitar que els compartiments endocítics tenen o bé fuites o bé
transportadors de proteïnes cap al citoplasma, on segueixen la via de processament del
MHC de classe I.
L’altre opció és carregar els pèptids a les molècules del MHC de classe I als
compartiments endosomals o al fagosoma. Aquests adquireixen els components de
càrrega de pèptids a molècules del MHC-I del reticle endoplasmàtic. La senyalització via
receptors de tipus toll contribueix a la presència de TAP en aquests endosomes
primerencs.
Per tant, la crospresentació explica vàries coses. Per un costat, la crospresentació explica
com pot ser que molts virus no infectin les cèl·lules dendrítiques i tot i així s’activin les cèl·lules
TCD8 citotòxiques. Això és perquè la crospresentació permet a les cèl·lules dendrítiques
presentar antígens extracel·lulars, produïts en altres cèl·lules, i activar limfòcits TCD8
citotòxics.
Per altra banda, la crospresentació explica com pot ser que la resposta immunitària a
patògens fagocitats com Salmonella, Brucella i Leischmania incloguin la resposta dels
limfòcits TCD8 citotòxics. Això ho aconsegueix ja que transfereix antígens dels patògens
del fagosoma al citosol, el que permet induir respostes de classe I.
Finalment, la crospresentació explica com els immunògens de les vacunes, sent
extracel·lulars, donen lloc a respostes de cèl·lules TCD8 citotòxiques. Això és perquè els
antígens extracel·lulars es transfereixen al citosol per ser presentats per MHC de classe
I i induir així una resposta dels limfòcits TCD8.
El resultat de la crospresentació té conseqüències importants per la cèl·lula presentadora
d’antigen. Això és perquè el fet que una cèl·lula presenti un antigen al limfòcit TCD8 l’activa.
Aquesta activació porta a la cadena que desemboca en l’activitat citotòxica del mateix
limfòcit TCD8.
La mort de la cèl·lula presentadora d’antigen no suposa un problema per la resposta
immunològica, doncs una vegada s’han activat els limfòcits TCD8 ja no és necessària la
cèl·lula dendrítica i, a més, se’n generen de noves a la medul·la òssia.
Aquest mecanisme de la crospresentació no explica, però, que els limfòcits TCD4 participin
en la resposta a virus.
Via de la Autofàgia.
S’ha descrit una altra via alternativa de presentació d’antigen anomenada autofàgia que
permet la presentació d’antígens citosòlics per molècules del MHC de classe II, el que
provoca l’activació de limfòcits TCD4.
L’autofàgia és el mecanisme constitutiu de les cèl·lules que els permet obtenir nutrients en
situacions de carència alhora que eliminen orgànuls citoplasmàtics malmenats. L’autofàgia
facilita l’entrada de proteïnes citosòliques a la via endocítica i, per tant, facilita la presentació
d’antigen per molècules del MHC de classe II i l’activació de limfòcits TCD4.

Immunologia 2020
CEBIO 70
Existeixen 3 mecanismes d’autofàgia possibles. En l’autofàgia depenent de xaperones,
aquestes transmeten proteïnes als endosomes. En la microautofàgia, la membrana
lisosomal envolta components del citoplasma. Finalment, en la macroautofàgia els
components del citoplasma són envoltats per una doble membrana formant un auto-fagosoma
que es fusionarà amb els endosomes.
Via de les CD1.
Finalment, la presentació d’antigen per molècules del MHC de classe I no clàssiques es
duu a terme per part de la CD1 entre d’altres. Aquestes molècules de CD1 tenen, a la seva
cua citoplasmàtica, uns motius que permeten la unió de diferents adaptadors que fan que,
des del reticle citoplasmàtic on són generades, vagin a diferents compartiments endosomals
de manera que CD1a la trobem als endosomes primerencs, CD1c als endosomes intermedis
i CD1b i CD1d es trobin als endosomes tardans, fins i tot als lisososmes.
Això permet que les diferents molècules de CD1 puguin carregar antígens de base lipídica
de diferent estructura, els quals seran presentats a limfòcits Tγδ i a Limfòcits NKT.
Interacció HLA-Pèptid.
Pèptids presentats per molècules del MHC.
Les molècules del MHC de classe I, al tenir uns extrems de lloc d’unió a l’antigen tancats,
només poden carregar pèptids d’entre 8 i 11 aminoàcids. Aquests normalment tenen entre
2 i 3 posicions del pèptid que proporcionen la major part de l’energia d’unió a la molècula del
MHC, són els residus d’ancoratge. Els principals residus d’ancoratge es troben a l’extrem
N-terminal i a l’extrem C-terminal, sent els residus 2 i 9. Aquests residus solen ser els
determinants de quins pèptids s’uniran i de quins no. Els aminoàcids dels llocs d’unió de
pèptid de les molècules del MHC de classe I són els residus més conservats. El repertori
de pèptids que presenta cada molècula del MHC de classe I és restringit i es poden identificar
els motius d’ancoratge per cada molècules del MHC de classe I.
Les molècules del MHC de classe II presenten pèptids d’entre 9 i 22 aminoàcids. El pèptid
en qüestió sol sobresortir peles extrems del lloc d’unió d’antigen. La zona central del pèptid,
de 9 aminoàcids i que ocupa el lloc d’unió de pèptids s’anomena seqüència core, i és la que
interacciona amb les molècules de MHC de classe II. Normalment, els pèptids tenen 4
residus d’ancoratge, els quals poden canviar entre diferents al·lels. Així i tot, la interacció
de l’esquelet del pèptid amb la molècula del MHC contribueix a l’energia d’unió. El repertori
de pèptids presentats per cada molècula del MHC de classe II és més gran que el de les
molècules del MHC de classe I.
Les molècules del MHC de classe I poden presentar pèptids més llargs sempre que els
residus d’ancoratge estiguin fixats i mantinguin l’estabilitat de la interacció. En aquest cas, la
part central pot sobresortir del lloc d’unió d’antigen.
En les molècules del MHC de classe I, les diferències en la forma d’unió es tradueixen en
diferències conformacionals del complex pèptid-MHC-I i, per tant, també es tradueixen en
diferències en els residus exposats a la interacció amb el TCR.
El lloc d’unió a pèptids de les molècules del MHC de classe II té els extrems oberts, permetent
que els pèptids sobresurtin. El nucli central del pèptid es troba fixat al lloc d’unió de pèptid
mitjançant interaccions entre els residus d’ancoratge i les butxaques. Els extrems N i C
terminals del pèptid que sobresurten són fàcilment degradats per proteases, mentre que la
seqüència principal està protegida.

Immunologia 2020
CEBIO 71
La unió pèptid-MHC depèn dels aminoàcids del pèptid a les posicions d’ancoratge que
s’uneixen a les butxaques o pockets de la molècula del MHC. Aquests residus d’ancoratge
formen el motiu d’unió del pèptid a una determinada molècula del MHC. La
complementarietat estereoquímica, conformacional i bioquímica està determinada pels
aminoàcids de l’al·lel MHC i selecciona els pèptids que s’hi poden unir.
La manca de residus complementaris a les butxaques del MHC impedeix l’acoblament estable
del pèptid i la seva presentació. Si els motius d’unió del pèptid estan ocupats per
aminoàcids que no encaixen a les butxaques, el pèptid no s’unirà correctament.
Parlem de mimetisme molecular quan una mateixa molècula del MHC és capaç de presentar
pèptids derivats d’una proteïna vírica o pròpia. Això ho fa perquè es mantenen els residus
d’ancoratge independentment de la proteïna de la que provingui el pèptid.
També es pot unir un mateix pèptid a diferent molècules del MHC. Això passa per un canvi
conformacional del pèptid. Això passa, per exemple, amb el pèptid viral gp33, el qual pren
diferents conformacions per poder interaccionar amb dues molècules del MHC de classe I
diferents. En aquest cas, el TCR reconeix el complex pèptid-MHC. Aquest complex té una
conformació diferent depenent de l’al·lel i del pèptid. Per tant, cada complex format per un
al·lel MHC i el pèptid tenen una conformació única que serà reconeguda per un TCR i no un
altre.
Les molècules del MHC de classe I que uneixen el pèptid han d’interaccionar amb el
TCR. Això vol dir que hi ha uns aminoàcids del pèptid determinats que són els d’ancoratge a
les molècules del MHC de classe I i que, d’altra banda, hi haurà unes cadenes laterals dels
aminoàcids que interaccionaran amb els CDR dels TCR. Aquestes seran les cadenes
laterals dels aminoàcids 1, 4, 7 i 8.
Hi ha un tipus d’antigen anomenats súper antígens que no funcionen de la mateixa manera.
Els súper antígens són proteïnes que interaccionen amb la regió constant d’un domini Vβ
del TCR i una regió constant de la molècula del MHC de classe II que, en funció del súper
antigen, pot ser la cadena α o la cadena β. Cada súper antigen interacciona amb la regió
constant d’una regió Vβ concreta. Poden ser d’origen endogen o exogen, víric o bacterià.
La interacció indueix una activació policlonal i la proliferació dels limfòcits T
independentment de la seva especificitat antigènica.

Immunologia 2020
CEBIO 72
Tema 14 – Cèl·lules Presentadores d’Antigen.
Funcions de les cèl·lules Presentadores d’Antigen. Al cos humà hi ha uns 1011 limfòcits T, els quals es divideixen en unes 109 especificitats
antigèniques possibles. Per altra banda, al cos humà hi ha unes 1014 cèl·lules que cal
monitoritzar. Monitoritzar, en aquest context, significa analitzar si es troben infectades o si
són canceroses, per exemple. Tenint en compte que cada cèl·lula hauria de ser escanejada
per totes i cadascuna de les especificitats antigèniques, es necessitarien 1023 contactes
cèl·lula-cèl·lula per escanejar tot el cos per part dels limfòcits T.
Si cada contacte cèl·lula-cèl·lula requereix d’un mínim de 10 minuts, es necessitarien un
mínim de 1021 minuts per fer un anàlisi complet del cos. Aquesta immensitat de temps,
corresponent a anys, ni tan sols té en compte que les cèl·lules han de migrar d’un lloc a un
altre ni tampoc que hi haurà contactes redundants.
Així i tot, els bacteris que infecten l’ésser humà es divideixen cada 20 o 30 minuts i els virus
es multipliquen a majors velocitats.
Per tal que es pugui generar una resposta immunològica eficient existeixen les cèl·lules
presentadores d’antigen. Aquest tipus de cèl·lules, entre les quals es troben les cèl·lules
dendrítiques, recullen els antígens als teixits i els transporten als limfonodes. Allà
s’acumulen i visiten els limfòcits T. Així doncs, als limfonodes s’acumulen tant les cèl·lules
presentadores d’antigen com els limfòcits T.
Per tant, les Cèl·lules Presentadores d’Antigen (APCs) tenen 4 funcions principals que
desenvolupen per poder ajudar a combatre les infeccions.
La seva primera funció és capturar antígens i portar-los al lloc anatòmic adient. Les
cèl·lules dendrítiques, les APCs per excel·lència, transporten els antígens als limfonodes o
a la melsa, on circulen els limfòcits T naive. Aquests limfòcits T escanegen les cèl·lules
presentadores d’antigen buscant complexes pèptid-MHC pels quals tenen un TCR específic.
Els antígens, doncs, es concentren als òrgans limfoides secundaris, per on circulen els
limfòcits T naive per monitoritzar la presència d’antígens.
Una altre de les funcions de les cèl·lules presentadores d’antigen és el processament dels
antígens. Aquest processament consisteix en la degradació proteolítica de proteïnes en
forma de petits fragments que es poden unir a molècules del MHC que s’exposaran a la
membrana de la cèl·lula presentadora d’antigen per tal que els reconegui el limfòcit T
corresponent. Cal recordar que els limfòcits T només reconeixen antígens que han estat
processats i degradats.
La tercera funció que tenen les cèl·lules presentadores d’antigen és, precisament aquesta:
mostrar els antígens d’una manera que puguin ser reconeguts per limfòcits T. La
presentació d’antígens peptídics al TCR dels limfòcits T suposa l’expressió de molècules del
MHC, molècules coestimuladores i molècules d’adhesió.
Finalment, l’última funció que duen a terme les cèl·lules presentadores d’antigen és
proporcionar segons senyals per a l’activació dels limfòcits T. Els limfòcits T naive
reconeixen antígens presentats per cèl·lules presentadores d’antigen. El reconeixement del
pèptid en una molècula del MHC és el que es coneix com a primer senyal. Depenent de
si la cèl·lula presentadora d’antigen és no professional o no activada, la resposta que provoca
en un limfòcit T pot ser entrar en estat d’anèrgia o no donar resposta.

Immunologia 2020
CEBIO 73
Tipus de Cèl·lules Presentadores d’antigen. La majoria de cèl·lules nucleades del cos expressen molècules del MHC de classe I. En
conseqüència, la majoria de cèl·lules nucleades del cos poden presentar antígens. En
general, però, el terme de Cèl·lula Presentadora d’Antigen fa referència a les cèl·lules amb
capacitat per presentar antigen fent servir molècules del MHC de classe II a més de les
de classe I.
Dins de les cèl·lules presentadores d’antigen n’hi ha de dos tipus. Les cèl·lules
presentadores d’antigen no professionals són aquelles on no hi sol haver una expressió
de molècules del MHC de classe II però que se n’hi pot induir l’expressió. Són cèl·lules
presentadores d’antígens no professionals els fibroblasts, les cèl·lules endotelials vasculars,
les cèl·lules epitelials i les cèl·lules de la glia. Les cèl·lules presentadores d’antigen
professionals, en canvi, són aquelles que presenten molècules del MHC de classe II alhora
que presenten co-estimuladors, pel que poden activar limfòcits T naive. Són cèl·lules
presentadores d’antigen professionals les cèl·lules dendrítiques, els macròfags i els limfòcits
B. Aquests últims tenen veus en contra que se’ls consideri cèl·lules presentadores d’antigen,
però.
Cèl·lules Dendrítiques.
L’any 1973, Ralph Steinman i Zanvil Cohn van descriure l’existència d’un tipus de cèl·lules
que constituïen un 1% del conjunt cel·lular dels limfonodes. Eren les cèl·lules Dendrítiques.
En aquesta descripció, Steinman i Cohn van destacar que eren cèl·lules amb una
distribució tissular ubiqua i que eren poc abundants a la sang. Les cèl·lules dendrítiques
reben aquest nom per les elongacions del citoplasma que presenten, anomenades dendros.
S’han descrit dos grans tipus de cèl·lules dendrítiques: cèl·lules dendrítiques
convencionals, com les cèl·lules de Langerhans i les cèl·lules interdigitants, i cèl·lules
dendrítiques plasmacitoides.
Les cèl·lules dendrítiques convencionals expressen un repertori de TLR (toll like receptors)
diferent del de les cèl·lules dendrítiques plasmacitoides. Això implica que reaccionen a la
presència de patògens diferents. A més, la seva funció principal és presentar antígens i
activar els limfòcits T naive. Les cèl·lules dendrítiques plasmacitoides, en canvi, tenen
la funció de produir interferons de tipus I com a resposta a la infecció del cos per part de
virus. Les cèl·lules dendrítiques convencionals, a més, tenen una elevada expressió de
CD11c, de molècules del MHC de classe II i de molècules co-estimuladores. Elements dels
que les cèl·lules dendrítiques plasmacitoides tenen una baixa expressió.
Totes les cèl·lules dendrítiques es generen a la medul·la òssia a partir del precursor
mieloide comú. A partir d’aquí es diferenciaran les cèl·lules dendrítiques convencionals i les
plasmacitoides. Les cèl·lules dendrítiques migren als teixits on capturen l’antigen i el
transporten als limfonodes proximals, on el presentaran a les cèl·lules T naive. En canvi, els
macròfags romanen al teixit.
En el viatge de les cèl·lules dendrítiques cap als teixits, passen per un procés de
maduració. Aquest fa que les cèl·lules dendrítiques passin de ser capturadores d’antigen a
presentadores d’antigen.
El procés de maduració s’inicia als teixits perifèrics, on la interacció amb el patogen provoca
l’activació de la cèl·lula dendrítica arran de la presència de PAMPs. La senyalització per
Receptors de tipus Toll indueix CCR7 i incrementa el processament dels antígens derivats
del patogen. CCR7 dirigeix la migració de la cèl·lula dendrítica als teixits limfàtics alhora
que augmenta l’expressió de molècules co-estimuladores i de molècules del MHC. La cèl·lula

Immunologia 2020
CEBIO 74
dendrítica, ara ja madura, entra al limfonode i pot transmetre alguns dels antígens a les
cèl·lules dendrítiques residents. Les cèl·lules dendrítiques positives en el co-receptor B7
estimulen les cèl·lules T naive.
Cèl·lules Dendrítiques convencionals.
Les cèl·lules dendrítiques convencionals que es troben als teixits reben el nom de cèl·lules
dendrítiques convencionals immadures. Aquestes tenen dendrites abundants i moltes
vesícules endocítiques amb elevat contingut de molècules del MHC. A més, les cèl·lules
dendrítiques convencionals immadures tenen una expressió elevada de PRRs, TLRs i
s’activen per interacció amb PAMPs. Aquestes són les cèl·lules presentadores d’antigen
que millor activen els limfòcits T naive, fent que s’iniciï la immunitat adaptativa.
Aquestes cèl·lules dendrítiques també expressen molts receptors que els atorguen una
elevada capacitat endocítica. Aquests receptors ho són de Fc d’immunoglobulines, com
FcγRI, FcγRII, FcγRIII, FcεRI i FcεRII; de molècules del sistema del complement com CR3 i
CR4; i de lectines del tipus C, com DC-SIGN o BDCA. El resultat de la interacció dels
antígens amb aquests receptors deriva en la internalització. Les cèl·lules dendrítiques
convencionals immadures realitzen la fagocitosi de microorganismes opsonitzats i de cèl·lules
i cossos apoptòtics. Aquestes cèl·lules dendrítiques també poden realitzar la macropinocitosi.
A més, les cèl·lules dendrítiques immadures expressen diferents receptors de quimiocines,
el que suposarà que puguin dur a terme la seva funció, és a dir, que puguin capturar
l’antigen.
Aquestes cèl·lules dendrítiques presents als teixits detecten l’antigen. Això farà que el deixin
i migrin a través dels vasos limfàtics als òrgans limfoides. En aquest procés de migració, les
cèl·lules dendrítiques redueixen el nombre de dendros fins convertir-se en una cèl·lula
velada. Aquest canvi no és només morfològic, sinó que també implica un canvi en la
distribució de les molècules del MHC de classe II. Aquestes molècules abandonen els
lisosomes on es troben, pel que ja no es co-localitzen amb les proteïnes lisosomals. Les
cèl·lules dendrítiques convencionals activades expressen CCR7 a la membrana, un receptor
de quimiocines que els permet accedir als limfonodes per les vènules d’endoteli alt.
Finalment, quan les cèl·lules arriben als limfonodes, ja podem parlar de cèl·lules
dendrítiques convencionals madures. Aquestes cèl·lules dendrítiques tenen dendros molt
llargs per augmentar la superfície de la cèl·lula i la possibilitat d’entrar en contacte amb els
limfòcits T. Com que tenen una elevada expressió de CCR7, es situen a l’àrea del limfonode
més rica en cèl·lules T. Com és d’esperar, tenen menys capacitat fagocítica i més expressió
a la membrana de complexes MHC-pèptid i de molècules co-estimuladores com CD80 i CD86,
altrament anomenades B7-1 i B7-2 respectivament. Aquests mateixos elements que es troben
a la membrana ja són pràcticament indetectables a les vesícules del citoplasma.
La funció de les cèl·lules dendrítiques convencionals és activar els limfòcits TCD4 i els
limfòcits TCD8 naive. Per fer-ho, produeixen elevades quantitats de citocines, que
determinaran el tipus de resposta per part dels limfòcits T.
A mesura que va avançant la ciència i les capacitats d’anàlisi per citometria de flux al tenir
més anticossos, es van fent subdivisions de les cèl·lules dendrítiques convencionals en funció
dels receptors de membrana i dels PRRs que expressen. Les subpoblacions de cèl·lules
dendrítiques són diferents en funció de si s’estudia la sang i els teixits limfàtics o els teixits
cutanis.

Immunologia 2020
CEBIO 75
Cèl·lules Dendrítiques Plasmacitoides.
Aquestes cèl·lules dendrítiques reconeixen àcids nucleics com a PAMPs, principalment.
Aquest reconeixement dona lloc a la secreció d’interferons de tipus I, com IFN-α i IFN-β,
els quals tenen un paper fonamental en la resposta davant de virus. El reconeixement d’àcids
nucleics també deriva en la inducció d’una sèrie de gens induïbles per IFN de tipus I. Són
gens que codifiquen per citocines, quimiocines i gens relacionats amb la inducció de
l’apoptosi. La interacció amb PAMPs augmenta l’expressió de molècules del MHC de classe
II i de molècules co-estimuladores, però es dona a un nivell més baix que a les cèl·lules
dendrítiques convencionals.
Les cèl·lules dendrítiques plasmacitoides (pDC) no són bones presentadores d’antigen, en
contraposició a les Cèl·lules Dendrítiques Convencionals (cDCs). Això es deu a que són
menys eficients en la fagocitosi, el processament i la càrrega a molècules del MHC de
l’antigen i a que no activen limfòcits T naive. Així i tot, poden activar limfòcits TCD4 i limfòcits
TCD8 de memòria.
Les cèl·lules dendrítiques plasmacitoides interactuen amb les cèl·lules NK, amb les cèl·lules
dendrítiques convencionals, amb les cèl·lules T i amb les cèl·lules B per mitjà de la secreció
de citocines i quimiocines, així com per l’expressió de molècules co-estimuladores.
Les pDC suposen menys del 1% del total de cèl·lules mononuclears de la sang en humans.
Poblacions de Cèl·lules Dendrítiques.
Les poblacions de cèl·lules dendrítiques expressen uns conjunts de receptors Toll no
solapants. Les cèl·lules dendrítiques de la sang perifèrica en humans i els monòcits
expressen diferents conjunts de TLRs tant en les seves superfícies cel·lulars com en els seus
compartiments lisosomals.
Les cèl·lules dendrítiques convencionals expressen una varietat de receptors Toll de
superfície i poden reconèixer patògens bacterials, fúngics i vírics. Secreten les citocines
inflamatòries IL-12, el factor de necrosi del tumor i la IL-6. Tot i tenir un conjunt similar de
TLRs, els monòcits no expressen TLR3, la regulen a mesura que maduren a cèl·lules
dendrítiques.
Tant les cèl·lules dendrítiques plasmàtiques humanes com les de ratolí expressen TLR7 i
TLR9, responen a virus i secreten interferons de tipus I. En els òrgans limfàtics secundaris,
els precursors de cèl·lules dendrítiques derivats de la sang estimulen la presència de
cèl·lules dendrítiques.
Aquestes cèl·lules dendrítiques, quan detecten un antigen, participaran en la resposta
adaptativa i la orientaran. Això es deu a la capacitat de sintetitzar diferents citocines. Per tant,
es pot dir que les cèl·lules dendrítiques actuen com a pont entre la resposta innata i la
resposta adaptativa.
Els limfòcits B com a cèl·lules presentadores d’antigen.
El limfòcit B és una bona cèl·lula presentadora de l’antigen que el reconeix mitjançant el BCR.
El BCR reconeix l’antigen i l’internalitza per endocitosi mediada per receptor. L’afinitat
del BCR per l’antigen és tan alta que necessita concentracions d’antigen 1000 vegades
inferior a la que necessita un macròfag per a unir-lo i internalitzar-lo. L’antigen internalitzat
és processat a la via endocítica i, per tant, és presentat per molècules del MHC de classe
II. Els complexes MHC-pèptid seran reconeguts per limfòcits TCD4 amb un TCR amb afinitat
per aquest mateix antigen. El limfòcit T activat col·laborarà en l’activació complerta del limfòcit
B, que entrarà al cicle de divisió cel·lular.

Immunologia 2020
CEBIO 76
La unió de l’antigen al BCR activa la senyalització BCR-Igα/Igβ, desencadenant la cascada
d’activació de la cèl·lula B. Com a conseqüència, s’accelera el transport als endosomes, es
reorganitzen i s’acidifiquen els endosomes tardans i s’acumulen molècules del MHC de classe
II i HLA-DM no associat a HLA-DO als MIIC. Com que el complex BCR-antigen és molt
estable, no es degrada fins als lisosomes, potenciant la presentació d’antígens reconeguts
pel BCR.
L’activació de la cèl·lula B indueix l’expressió de CD40, el qual interacciona amb CD40L
a les cèl·lules T, augmentant encara més la funció co-estimuladora de les cèl·lules B.
Cèl·lules presentadores d’antigen professionals. Les cèl·lules epitelials del timus, les quals inclouen cTEC i mTEC, expressen molècules del
MHC de classe I i de classe II. Aquestes cèl·lules epitelials són presentadores d’antigen al
timus. No participen a la resposta adaptativa.
El fenomen de la CrosPresentació. De les vies alternatives de la presentació d’antigen, en destaca la crospresentació. Els
macròfags són la primera cèl·lula presentadora d’antigen en què es va veure la capacitat de
crospresentar. Així i tot, la rellevància d’aquesta opció in vivo encara no està clara fruit de la
baixa investigació d’aquest fet. Les cèl·lules B, malgrat ser molt bones en les seves funcions,
poden ajudar a les cèl·lules dendrítiques a crospresentar quan hi ha poc antigen disponible.
No pot realitzar la crospresentació ella mateixa, però. Finalment, les cèl·lules dendrítiques
són les més estudiades en quant a crospresentació fa referència. Són excepcionalment bones
en aquest procés.

Immunologia 2020
CEBIO 77
Tema 15 – Citocines i Quimiocines
Citocines Les citocines són polipèptids d’entre 8 i 80kDa que actuen com a missatgers entre cèl·lules
quan són secretades durant les fases efectores de la resposta immunitària innata i de la
resposta immunitària adaptativa. Les citocines actuen com a mediadors i reguladors de
l’hematopoesi, de la resposta immunitària, de la inflamació i de la reparació del teixit.
Les citocines, generalment, no existeixen com a molècules pre-formades actives i la seva
secreció requereix d’una síntesi proteica de novo, la qual és autolimitada i breu al tenir un
mRNA inestable. Moltes citocines tenen una vida mitjana molt curta. Arran d’això, només en
rebran l’efecte les cèl·lules més properes a la secreció. Les citocines estimulen programes
específics de proliferació i diferenciació de les cèl·lules diana. Aquestes cèl·lules diana solen
formar part del sistema immunitari, però altres tipus cel·lulars també es poden veure afectats
per les citocines. Les citocines són un element clau en el sistema immunitari, doncs regulen
la intensitat, la durada i el tipus de resposta immunològica que es dona.
Les citocines s’uneixen a receptors específics expressats a la membrana plasmàtica de les
cèl·lules, pel que només actuen sobre aquelles que els expressen. En conseqüència,
l’expressió dels receptors controla la capacitat de la citocina per actuar sobre la cèl·lula.
La interacció citocina-receptor genera senyals que canvien l’expressió gènica, ja que
augmenta l’expressió del receptor i la secreció d’altres citocines. Aquestes són actives a
concentracions picomolars perquè l’afinitat dels receptors és alta, d’una Kd d’entre 10-9 i
10-12M. Així i tot, la concentració de citocines a nivell local és molt alta, el que contribueix a
que l’efecte es doni principalment sobre les cèl·lules més properes malgrat pugui ser
necessari un contacte cèl·lula-cèl·lula.
Funcions de les citocines.
Les citocines són produïdes, principalment, per cèl·lules del sistema immunitari. Així i tot, hi
ha altres tipus cel·lulars que secreten citocines que modulen la resposta immunitària.
D’entre aquests tipus cel·lulars en destaquen les cèl·lules del fetge, del cervell o de les
glàndules endocrines.
Les citocines participen en l’hematopoesi tot regulant la proliferació i la diferenciació
cel·lulars. Participen en la immunitat innata al ser les responsables de la inducció de la
resposta inflamatòria. També participen a la immunitat adaptativa, on són presents en el
desenvolupament de les respostes humoral i cel·lular.
Les citocines poden tenir efectes autocrins, actuant sobre la mateixa cèl·lula que les han
produïdes, paracrins, actuant sobre una cèl·lula veïna, o poden tenir efectes endocrins si
entren a la circulació sanguínia i actuen lluny d’on s’han produït.
Un cas d’efecte autocrí és el que presenten les citocines interleuquines 2, les quals són
produïdes per les cèl·lules T per tal que s’uneixin als receptors d’IL2 i indueixin la producció
de més citocines. Un cas d’efecte paracrí és el que es dona amb la secreció de citocines per
part dels limfòcits T en el procés de col·laboració entre cèl·lules B i cèl·lules T. Finalment, un
cas de citocines amb efecte endocrí són les citocines inflamatòries com IL-1, TNF-α i IL6; les
quals arriben a l’hipotàlem i indueixen la resposta neurològica de l’augment de la temperatura.
Les citocines, en qualsevol d’aquests efectes, també actuen en cascades amplificadores. Una
d’elles pot consistir en un limfòcit T activat, el qual generarà la citocina IFN-γ, i activarà un
macròfag. Els macròfags activats generen una altra citocina, la IL-12, i aquesta s’encarrega
d’activar nous limfòcits T que acabaran produint més IFN-γ i altres citocines com IL-2.

Immunologia 2020
CEBIO 78
Una citocina pot tenir efectes diferents sobre diferents tipus cel·lulars i pot actuar
sinèrgicament o antagònicament amb d’altres citocines. Per exemple, IL-4 és una citocina que
presenta pleiotropisme. Pleiotropisme és la característica d’afectar diferent a diferents tipus
de cèl·lules. Això és doncs IL-4 pot inhibir els macròfags M1 i els limfòcits Th1 però també pot
activar els limfòcits Th2 i les cèl·lules B.
En un sentit bastant diferent al pleiotropisme hi trobem la redundància i el sinergisme, que
implica que diferents citocines tenen un efecte similar sobre un tipus cel·lular alhora que
l’efecte de la seva acció conjunta és superior al de la suma dels efectes de cadascuna per
separat.
De la mateixa manera que existeix el sinergisme, el qual es pot veure amb les citocines IL-4
i IL-5 induint el canvi d’isotip cap a IgE, existeix l’antagonisme. Aquest fenomen és aquell per
el qual una citocina evita que es dugui a terme l’efecte que ha provocat una altra. Un exemple
d’antagonisme que es dona en citocines el veiem amb IFN-γ, que bloqueja el canvi d’isotip a
IgE que provoca la IL-4.
Una citocina pot ser produïda per diferents tipus cel·lulars i afectar a molts tipus cel·lulars.
Això significa que una mateixa cèl·lula és exposada a moltes citocines alhora i que l’efecte
final és el resultat de la suma dels efectes sinèrgics i antagònics de totes elles.
Classes de Citocines.
S’han descrit 6 classes de citocines. Aquesta classificació és en funció dels receptors que
tenen.
La família de la interleuquina 1 inclou mediadors d’inflamació importants i en formen part la
IL-1α, la IL-1β, la IL-18, la IL-33 i la IL-1Ra. El seu receptor està compost per una regió
extracel·lular amb dominis de tipus immunoglobulina.
La gran família de les petites molècules de citocina a la que anomenem família hemopoietina
inclou una gran varietat de citocines i de funcions de les mateixes. En aquesta família de
citocines s’hi poden trobar factors de creixement, les Interleuquines 2, 3, 4, 5, 6, 7, 12, 13, 15,
21 i 23, la prolactina i alguns factors d’expressió. El seu receptor conté un motiu WSXWS a
part d’unes quantes cisteïnes conservades.
La família dels interferons és una família de citocines molt importants en les respostes anti-
víriques, però totes són importants moduladors de la resposta immunitària. En aquesta família
hi ha les IFN-α, β i γ, així com les interleuquines 10, 19, 20, 22 i 24. Els seus receptors són
similars als receptors de les hemopoetines sols que no tenen el motiu WSXWS.
Els membres de la família dels factors de necrosi del tumor, la qual inclou citocines com TNF-
α, TNF-β, CDL40, BAFF, APRIL o LTβ; tant poden ser solubles com de membrana. A més,
són citocines involucrades en el desenvolupament del sistema immunitari, en les funcions
efectores i en l’homeòstasi. Els seus receptors consisteixen en dominis extracel·lulars
conservats rics en Cisteïnes.
La família de la interleuquina 17 és la més recentment descoberta. Els seus membres, les IL-
17 A, B, C, D i F; promouen l’acumulació i l’activació de neutròfils alhora que són pro-
inflamatòries. Els receptors de la família de la interleuquina 17 està composta per 5 membres:
IL-17RA, IL-17RB, IL-17RC, IL-17RD i IL-17RE. Aquesta família de receptors poden formar
homodímers, heterodímers i, fins i tot, trímers.

Immunologia 2020
CEBIO 79
Finalment, les quimiocines són aquelles que tenen una funció de quimiotaxi. Inclou les IL-8,
les CCL2, 3, 19 i 21 i també inclou les RANTES. Els seus receptors consisteixen en 7 dominis
transmembrana.
La família de les Interleuquines 1.
IL-1α, IL-1β i IL-18 són interleuquines produïdes principalment per monòcits, macròfags i
cèl·lules dendrítiques com a precursors de 31kDa. IL-1α té activitat com a precursor. La IL-1β
requereix de l’acció de l’enzim caspasa 1 i és activada per l’inflamasoma, que la converteix
en la forma activa de 17 kDa. Els receptors IL-1R i IL-18R pertanyen a la superfamília d eles
immunoglobulines. IL-1R i IL-18R són receptors bicatenaris. Les formes solubles al sèrum,
monocatenàries i anomenades ILRa, actuen com a antagonistes.
La senyalització del receptor de la IL-1 i de la IL-18 és semblant a la senyalització del TLR-4.
Aquesta senyalització està mediada per l’adaptador Myd88, que recluta IRAKs i TRAF6.
IRAK4 fosforil·la TAK6, el que permet reclutar TAK1 i TAB1-TAB2. Aquestes molècules
aprofiten un complex de poliubiquitinació per formar cadenes d’ubiquitina que activen TAK-1,
el que desbloqueja l’NFκB del citosol i li permet translocar-se al nucli, on activarà la via de les
MAP quinases.
La unió de la citocina al receptor indueix la síntesi d’altres citocines proinflamatòries i també
indueix l’augment de l’expressió de molècules d’adhesió.
La família de les hemopoetines.
Les citocines de tipus I, altrament anomenades hemopoetines, tenen una estructura
tridimensional composta per 4 cadenes hèlix α i algunes cadenes β antiparal·leles. Hi ha
citocines com IL-2, IL-3 i IL-4, que tenen cadenes petites d’entre 8 i 10 residus. Altres
citocines, com IL-6 i IL-12, són d’entre 10 i 20 aminoàcids. Les hemopoetines tenen funcions
molt diverses. Entre aquestes, hi trobem la proliferació de limfòcits T per part de IL-2, la síntesi
de proteïnes del complement o diferenciació de cèl·lules B a cèl·lules plasmàtiques per part
de IL-6, la inducció de Th1 per IL-12 o de Th2 per IL-4 i la diferenciació de llinatges leucocitaris
per part de GM-CSF o G-CDF. Els receptors de les citocines de tipus I estan formats per dies
o més cadenes amb motius conservats consistents en la seqüència Triptòfan-Serina-X-
Triptòfan-Serina.
Els receptors de classe I solen estar formats per diverses subunitats on una uneix
específicament la citosina i l’altra o altres transmeten el senyal. Les subunitats específiques
solen ser de baixa afinitat, però la unió a la subunitat transductora de senyals forma receptors
d’alta afinitat. Les citocines amb receptors que tenen la mateixa subunitat de transducció de
senyals solen tenir els mateixos patrons de senyalització, el que resulta en una redundància
en l’efecte de la citosina. Hi sol haver competència per les subunitats transductores, produint
antagonisme entre citocines diferents.
Un exemple d’efectes de citocines de tipus I és la que es dona amb la interleuquina 2, la qual
té els seus receptors als limfòcits TCD4, TCD8, a les cèl·lules NK i a les cèl·lules B activades.
En estat de repòs, tots elles expressen la cadena γ del receptor unida a la cadena β, pel que
ja pot unir la citocina però amb una afinitat intermitja. Quan hi ha una activació, s’expressa la
cadena α del receptor de la IL-2. Aquesta cadena α, quan està sola, té una afinitat molt baixa.
Així i tot, l’associació de les tres cadenes, α, β i γ, forma un receptor amb una afinitat molt
alta. El fet de poder expressar receptors d’afinitats baixes, intermitja i altes permet a les
cèl·lules regular la seva resposta immunològica i la seva reacció a les citocines.

Immunologia 2020
CEBIO 80
La família de les citocines de tipus II.
La família de les citocines de tipus II va rebre aquest nom, el d’interferons, per la capacitat
d’interferir en el creixement i en la disseminació d’un virus. Se n’han descrit 3 grups. Les IFN
de tipus I inclou les α i les beta. Són produïdes en cèl·lules dendrítiques plasmacitoides en
resposta a la infecció per virus. Les IFN de tipus II també reben el nom de IFNγ i es produeixen
en leucòcits com els limfòcits T, les cèl·lules NK o les cèl·lules NKT en resposta a patògens
intracel·lulars i com a resposta citotòxica. Les IFN de tipus III reben el nom d’IFNlambda i són
produïdes per les cèl·lules epitelials de les mucoses intestinal i pulmonar com a resposta a
virus. La família de les citocines de tipus II també inclou la IL-10, una immunomoduladora, i
la IL-22, de defensa de les cèl·lules epitelials.
Senyalització intracel·lular de les citocines de tipus I i II.
Quan la citocina s’uneix al receptor específic de membrana, indueix la dimerització de les
cadenes que el formen. Aquesta dimerització recluta STATs (Signal Transducer and Activator
of Transcription), que s’uniran als residus de tirosina del dímer de receptors. Seguidament, la
proteïna JaK (Janus Kinase) fosforilarà aquests mateixos residus, provocant la dimerització i
activació de les STATs. Aquestes es translocaran aleshores al nucli, on s’uniran a seqüències
promotores per regular la transcripció i expressió gèniques. En funció del receptor, els efectes
canviaran.
Hi ha 4 quinases diferents que s’anomenen Jak 1, 2 i 3 i Tyk2 i hi ha 7 factors de transcripció
STATs diferents anomenats STAT 1, 2, 3, 4, 5a, 5b i 6. Cada cua citoplasmàtica del receptor
de citosina s’uneix a una quinasa diferent, permetent l’expressió de diferents factors de
transcripció.
En el cas del receptor de la IL-2, les diferents cadenes poden arribar a activar diferents factors
de transcripció, però domina STAT 5. En el cas de la IL-4, és STAT 6.
IL-12. El pont entre la immunitat innata i l’adaptativa.
De la IL-12 sempre s’ha dit que fa de pont entre la immunitat innata i l’adaptativa perquè és
produïda pels macròfags que, un cop detecten un senyal de perill, la produeixen. IL-12 activa
les cèl·lules presentadores d’antigen, determinant la seva secreció i el tipus de resposta
cel·lular de limfòcits T que es donarà. Això és perquè la IL-12 provoca que els limfòcits T
naive s’activi i es diferenciï a cèl·lula T efectora helper de tipus 1.
Les funcions biològiques de la IL-2.
La IL-2 és produïda sobretot per limfòcits TCD4 col·laboradors i limfòcits TCD8 en menor
quantitat. Actua tant sobre limfòcits TCD4 activats com limfòcits TCD8 citotòxics i cèl·lules
NK. Contribueix a la proliferació de limfòcits B i en la producció d’anticossos. Actua com a
factor de creixement i activació de limfòcits T, limfòcits B i de cèl·lules NK. La producció de
IL-2 És imprescindible per tal que un limfòcit B aconsegueixi proliferar, dividir-se i produir
citocines.
Funcions Biològiques de la IL4.
La citocina IL-4 és produïda, majoritàriament, per cèl·lules T, cèl·lules NKT i mastòcits.
Inhibeix l’activació de cèl·lules NK i de macròfags, indueix a la proliferació i diferenciació de
limfòcits B activats, promou el canvi d’isotip a IgE i a IgG1 i estimula el desenvolupament i
expansió de cèl·lules T helper de tipus 2.
La citocina IL-4 és una d’aquestes citocines que es veu involucrades en la interació amb altres
citocines. Per començar, el limfòcit TCD4 naive, en activar-se, prolifera i es diferència a
limfòcit T efector, Th. En funció de les citocines decretades es diferenciarà en un o un altre

Immunologia 2020
CEBIO 81
tipus cel·lular de Th. La IL-4 indueix la generació de limfòcits de tipus Th2 mentre que la IFNg
indueix Th1. Cadascuna d’aquestes citocines inhibeix l’efecte de l’altre.
Una cosa semblant passa amb l’interferó γ. IFNγ activa el limfòcit T-bet, que indueix la
transformació a Th1. Aquesta activació és anulada per IL-4. Similarment passa amb el factor
de transcripció GATA-3 als limfòcits Th2, el qual és activat per IL-4 i desactivat per IFNγ.
GATA-3 inhibeix el pas a T-bet.
Funcions biològiques de l’interferó γ.
D’altra banda, l’IFNγ també actua sobre macròfags activant-los i augmentant la seva
activitat anti-microbiana i antiviral, sobre els limfòcits B tot provocant-ne la diferenciació i el
canvi d’isotips, sobre els limfòcits TCD4 naive diferenciant-los a limfòcits T efectors Th1 i
sobre les cèl·lules presentaodres d’antigen incrementant l’expressió de molècules del MHC,
de molècules d’adhesió i de molècules co-estimuladores per tal que hi hagi un increment en
la presentació d’antigen.
La família dels factors de necrosi dels tumors.
Les citocines de la família del TNF tenen diverses funcions. D’entre elles en destaquen la de
regular el desenvolupament, la funció efectora i la d’homeòstasi de les cèl·lules del sistema
neuronal, esquelètic i immunitari.
TNFα i TNFbeta són citocines proinflamatòries produïdes principalment per macròfags en
resposta a la infecció o a les situacions d’estrès. El TNFbeta és produït per limfòcits T activats
i activa neutròfils, cèl·lules endotelials i osteoclasts. Aquesta activació suposa un augment de
l’expressió de molècules del MHC i de molècules d’adhesió.
Les citocines BAFF i APRIL es torben implicades en la proliferació i en l’activació de limfòcits
B i en la producció d’immunoglobulina A a la mucosa intestinal. BAFF i APRIL estan produïdes
per cèl·lules T, cèl·lules dendrítiques, macròfags i monòcits, però no ho estan per cèl·lules B.
Les cèl·lules B són les que expressen de forma quasi exclusiva els seus receptors: BCMA i
TACI.
Els receptors de la família del TNF, TNF-R1 i TNF-R2, són els receptors més estudiats. Tenen
una expressió ubiqua i a la seva cua citoplasmàtica hi ha un death domain. TNF-R1 pot
desencadenar senyals d’activació i de supervivència o pot fer-ho d’apoptosi i mort. El TNF
s’uneix al TNF-1R i n’indueix la trimerització. Aquesta provoca un canvi conformacional que
rpovoca la unió amb TRADD, una molècula amb Death Domains.
Si TRADD permet la unió de la quinasa RIP1 i TRAF2, s’ha iniciat la senyal d’apoptosi. En
aquest cas, el complex es lliura al citosol formant un nou complex amb la molècula FADD,
que uneix pro-caspasa 8 i la transforma en la seva forma activa. La caspasa 8 activa les
caspases 3 i 7, unes nucleases actives que trenquen el DNA.
Si TRADD permet la unió d’ubiquitines lligases que poliubiquitinen RIP1, s’ha iniciat la senyal
de supervivència. Això portarà a l’activació de TAK1-TAB1/TAB2, la qual activa NFκB i MAP-
quinases. NFκB és la responsable de la síntesi de cFLIP, que inhibeix la caspasa 8.
Família de la IL-17.
La família de citocines més recentment descrita inclou les interleucines 17A, 17B, 17C, 17D i
17F. Els receptors d’aquestes citocines es troben expressats en neutròfils, fibroblasts,
cèl·lules epitelials, queratinòcits i altres cèl·lules no limfoides.
Quan les citocines de la família IL-17 realitzen una senyalització via IL-17RA i IL-17RC, el
resultat és una activació de NFκB a partir del reclutament de ACT1, que s’uneix a TRAFs.

Immunologia 2020
CEBIO 82
Aquests, al seu torn, s’uneixen a TAK1-TAB1/TAB2, qui activa NFκB. NFκB, un cop entra al
nucli, provoca l’expressió de citocines. La senyalització via IL-17RA i IL-17RC també resulta
en l’activació de la cascada de les MAP quinases, que fosforilarà ERK1. ERK1 no està implicat
en la fosforil·lació de factors de transcripció, sinó que s’encarrega d’estabilitzar el mRNA.
Finalment, l’últim element que s’activa fruit de IL-17RA i IL-17RC són els factors de
transcripció C/EBPbeta i C/EBPsigma, que indueixen a la transcripció de IL-6.
Les citocines IL-17 tenen vàries funcions. Per començar, són les encarregades d’estimular
l’expressió de citocines pro-inflamatòries com IL-6, TNF-α, G-CSF, GM-CSF i IL1beta. Les IL-
17 també provoquen un augment de la prostaglandina E2 i d’òxid nítric en diferents tipus
cel·lulars, el que afavoreix l’esclat respiratori. Quan actuen les citocines IL-17 hi ha una
inducció de quimiocines, sobretot les CXC que afavoreixen la quimiotaxi de neutròfils. Són
quimiocines induïdes per IL-17 la CXCL1, 2, 5, 8 i 10.
Les IL-17 tenen importància regulatòria sobre la funció dels limfòcits, la qual realitzen tot
inhibint la polarització a Th1. Aquesta tasca és gràcies a la supressió de l’expressió de T-bet.
També tenen càrrecs regulatoris sobre la funció de cèl·lules no limfoides. IL-17, ja estigui sola
o combinada amb el factor activador B (BAFF), promou la supervivència, proliferació i
diferenciació dels limfòcits B i, per tant, la generació de cèl·lules plasmàtiques.
Quimiocines Les quimiocines o citocines quimiotàctiques són una superfamília de polipèptids petits d’entre
8 i 14 kDa molt glicosilats i amb la capacitat d’estimular la motilitat dels leucòcits i el seu
moviment dirigit mitjançant l’establiment d’un gradient. La capacitat d’estimular la motilitat dels
leucòcits rep el nom de quimiocinesi i la capacitat d’estimular el seu moviment dirigit rep el
nom de quimiotaxi.
Les quimiocines estan produïdes per una gran diversitat de cèl·lules limfoides i no limfoides
de diferents orígens. Les quimiocines s’emmagatzemen als grànuls citoplasmàtics, que
alliberen el seu contingut a l’espai intercel·lular. Les quimiocines són reconegudes per
receptors ancorats a la membrana i se n’han descrit més de 45 tipus. Les 45 quimiocines
tenen, entre elles, molta homologia de seqüència, de fins a un 95%.
Les quimiocines tenen una estructura comuna basada en 1 hèlix α i una làmina de cadenes
beta antiparal·leles. El plegament de la molècula està estabilitzat per dos ponts disulfur que
s’estableixen entre cisteïnes conservades.
Hi ha 4 tipus de quimiocines en funció del nombre d’aminoàcids n conservats que separen
els dos primers residus de cisteïna. Poden ser quimiocines C, CC, CXC o CX3C.
Les quimiocines també es poden separar en classes segons la seva funció, doncs poden ser
quimiocines amb funció inflamatòria, que promouen l’adhesió dels neutròfils a les cèl·lules
endotelials, l’extravassació dels neutròfils i la seva migració fins al lloc d’inflamació;
quimiocines amb funció homeostàtica, que s’expressen a teixits i òrgans en els que
aparentment no hi ha estímuls inflamatoris, i poden ser quimiocines amb una funció
angiogènica o angiostàtica.
Algunes quimiocines amb funció homeostàtica estan constitutivament expressades en alts
nivells als limfonodes, a la melsa, al fetge, als fol·licles limfoides, a les plaques de peyer, al
timus i a la medul·la òssia. La funció de les quimiocines amb funció homeostàtica és la
recirculació limfocitària, el desenvolupament de les cèl·lules T i B i el correcte
desenvolupament i compartimentalització dels teixits limfoides.

Immunologia 2020
CEBIO 83
Les quimiocines amb una funció angiogènica promouen la vascularització in vivo. A les
teràpies anti-tumorals s’utilitzen anticossos neutralitzants de quimiocines per reduir el
creixement i la capacitat de metàstasi dels tumors. Les quimiocines amb una funció
angiostàtica, del mateix grup que les angiogèniques, eviten el desenvolupament de xarxes
vasculars al voltant de determinats teixits.

Immunologia 2020
CEBIO 84
Tema 16 – Anatomia Funcional del sistema Immunitari
Els Òrgans limfoides. Els òrgans limfoides es separen en primaris, on s’originen i maduren les cèl·lules del sistema
immunitari, i els òrgans secundaris, on es concentren els limfòcits madurs, s’acumula
l’antigen i s’inicia la resposta immunitària. Són òrgans limfàtics primaris la medul·la òssia,
origen dels limfòcits B i T i encarregada de la maduració dels primers, i el timus, on es
produeixen limfòcits T i cèl·lules NKT. Són òrgans limfàtics secundaris els limfonodes, la
melsa i els MALT, els teixits limfàtics associats a mucoses. Aquests MALT es separen en
BALT (bronquials), GALT (intestí), NALT (nasofaringe) i SALT (pell). A més, tots aquests
òrgans es troben connectats entre si per vasos sanguinis i limfàtics.
Els òrgans limfoides primaris.
La medul·la òssia.
La medul·la òssia és on es generen els precursors hemopoètics que poblaran altres òrgans.
Els precursors es generen a la medul·la òssia per processos com l’eritropoesi2 i la
mielopoesi. També es generen els precursors dels limfòcits T i dels limfòcits B. Aquests
últims es desenvolupen a la mateixa medul·la, on obtindran diversitat i tolerància. Arran del
capteniment d’aquestes cèl·lules B que, quan es desenvolupen, es converteixen en cèl·lules
plasmàtiques, la medul·la òssia també és l’encarregada de la producció d’anticossos.
La medul·la es troba a tots els ossos llargs del cos. A l’interior de tots aquests hi ha, doncs,
capacitat hematopoètica. Així i tot, la capacitat es va perdent amb l’edat. A l’edat adulta
bona part de la medul·la es torna teixit adipós i això és el que n’hi fa perdre. La màxima
capacitat de producció és en el moment del naixement i, a mesura que passa el temps,
es va perdent. La pèrdua és inversament localitzada, doncs amb el pas dels anys la producció
i capacitat hemopoètica es va concentrant a les costelles, la pelvis i la columna vertebral.
El timus.
El timus és l’altre òrgan limfoide primari. Es troba situat al mediastí i consisteix en una
càpsula fibrosa al voltant de dos lòbuls dividits en lòbels separats per trabècules. En
cadascun d’aquests lòbels s’hi troba el còrtex, la part més perifèrica, la medul·la al centre i
la zona cortico-medul·lar entre les dues. Aquesta zona participa en el procés de
diferenciació tímica dels limfòcits T. En aquesta mateixa zona també es dona la selecció
positiva dels limfòcits T doble positius, mentre que a la medul·la es fa la selecció negativa
dels limfòcits single positive.
Al timus és on té lloc la diferenciació dels precursors en les diferents poblacions de
limfòcits T, ja siguin convencionals com els TCD4, els TCD8, els Tαβ o els Treg; o no
convencionals com els NKT o els Tγδ. En el timus també on té lloc el procés de selecció.
Això és gràcies que l’estructura anatòmica de Medul·la, còrtex i zona cortico-medul·lar,
juntament amb la composició cel·lular del tim, permeten aquesta diferenciació.
Igual que amb la medul·la òssia, el timus involuciona amb l’edat. Al néixer, el timus té un
pes d’uns 20 grams i augmenta durant uns 10 anys fins arribar a uns 30-35 grams. Mai
desapareix del tot, però a partir de la pubertat s’inicia la involució. Aquesta involució
consisteix en una disminució del volum i en què el parènquima3 del teixit és substituït per
teixit adipós, generant menys limfòcits T. Apart de les cèl·lules del teixit adipós, al tim també
es formen el que s’anomena Corpuscles de Hassall. En aquests hi ha una acumulació de
2 conjunt de processos de formació, maduració i alliberament al torrent sanguini dels eritròcits 3 Teixit essencial d’un òrgan

Immunologia 2020
CEBIO 85
cèl·lules epitelials tímiques, macròfags, cèl·lules dendrítiques i, de vegades, també limfòcits.
Malgrat no tenen una funció ben determinada, es sap que regulen la limfopoesi4.
Quan es parla de la substitució del parènquima s’està fent referència a la pèrdua de
l’estructura característica de l’òrgan. En conseqüència d’aquesta substitució per teixit
adipós hi ha una reducció de la xarxa de cèl·lules epitelials corticals, una regió medul·lar molt
més petita i difusa i es torna difícil distingir la regió cortico-medul·lar. Aquest envelliment del
timus, doncs, fa que aquest tingui una menor capacitat de generar limfòcits T madurs, el que
es tradueix en una acumulació poc diversa de limfòcits T de memòria a la perifèria.
Òrgans limfoides secundaris. El sistema immunitari és dispers i es troba escampat per totes les zones del cos. Però gracies
als vasos limfàtics, tots aquests elements que en formen part (Limfonodes, melsa i MALT) es
troben connectats entre si. El sistema limfàtic recull els antígens via vasos limfàtics i els
concentra als limfonodes. El conjunt de citocines, quimosines i molècules d’adhesió
permeten la circulació, el homing i la retenció dels limfòcits en els òrgans limfàtics
secundaris. La captació d’antigen i la recirculació limfàtica a través dels òrgans limfàtics
secundaris són essencials per a una resposta immune eficient.
Vasos limfàtics.
Els vasos limfàtics es troben als teixits i recullen el líquid intersticial: tot el que hi hagi al
teixit i no siguin cèl·lules. La diferència de pressió entre l’espai intersticial i l’interior dels
capil·lars limfàtics facilita l’entrada als capil·lars. Aquest mateix procediment és el que ha
provocat, prèviament, la sortida del líquid des dels capil·lars sanguinis fins al líquid intersticial.
Les vàlvules flaplike dels vasos limfàtics permeten l’entrada del líquid intersticial als
capil·lars limfàtics. La circulació limfàtica es produeix per compressió de les estructures
veïnes i per la contracció de la musculatura llisa associada als vasos. Les vàlvules internes
dirigeixen l’avançament de la limfa i n’impedeixen el reflux. El vas limfàtic arribarà com a vas
aferent a un altre limfonode o un vas limfàtic de major diàmetre.
La limfa recollida a un teixit determinat va passant el filtre dels limfonodes cap a vasos
limfàtics de cada cop més diàmetre fins arribar al ducte, el vas limfàtic de major calibre.
Aquest connecta amb la circulació sanguínia. Per tant, una vegada el líquid intersticial ha
passat per tots els limfonodes, torna al teixit del que ha sortit. Així i tot, gràcies a la tasca dels
limfonodes, aquest líquid ja no té antígens ni patògens. Hi ha dues grans àrees de
drenatge. Una d’elles és el circuit que desemboca en el ducte toràcic, que avoca el seu
conegut a la vena subclàvia esquerra, i l’altre és el ducte limfàtic dret, que aboca el contingut
a la vena subclàvia dreta.
Això és important perquè hi ha una gran diversitat de limfòcits T però n’hi ha poques quantitats
de cada especificitat i es troben escampats per la circulació. De la mateixa manera, per tal
que es puguin reconèixer els antígens cal un punt de trobada: els limfonodes. Aquest
genera un entorn que fa possible la interacció de l’antigen amb els limfòcits i l’activació del
sistema immunitari.
Limfonodes.
Els limfonodes o ganglis tenen una estructura molt concreta. Tenen una sèrie de vasos
limfàtics aferents que arriben al limfonode i ho fan a nivell de la càpsula, la qual té les
trabècules. Per sota d’aquesta càpsula hi ha el sinus subcapsular, un canaló on els vasos
aferents hi aboquen els seus continguts. Al limfonode, la part més perifèrica correspon al
4 Generació de limfòcits

Immunologia 2020
CEBIO 86
còrtex i és on hi trobem els limfòcits B. Anant cap a la part més interna hi ha el paracòrtex,
que és on es troben els limfòcits T. Finalment, al centre del limfonode, hi ha la medul·la. En
aquesta hi trobem les cèl·lules plasmàtiques, les que s’acaben de generar. La via de sortida
és un vas limfàtic eferent.
Als limfonodes es produeix l’entrada d’antigen a través dels vasos limfàtics aferents i es
filtra a través de l’estroma o bé és carregat a les cèl·lules dendrítiques madures per la mateixa
via. Un cop dins es dona el contacte de l’antigen amb les cèl·lules presentadores
d’antigen, que poden ser cèl·lules dendrítiques, macròfags o cèl·lules B. També es fa una
presentació d’antigen a les cèl·lules T i a les cèl·lules B per activar-les. Aproximadament,
cada dia passen 25*10^9 limfòcits pels limfonodes.
Hi ha 3 vies d’entrada de l’antigen al limfonode. En el cas dels immunocomplexes o les
partícules víriques, l’antigen és capturat i mostrat al còrtex per macròfags del sinus
subcapsular. En el cas dels antígens solubles petits o proteolitzats com les quimosines,
l’antigen accedeix al còrtex a través de gaps entre les cèl·lules que formen la paret del sinus
o a través de conductes de cèl·lules fibroblàstiques reticulars. Finalment, els antígens grans
o provinents de cèl·lules són transportats al còrtex dins les cèl·lules dendrítiques que l’han
capturat al teixit.
Cèl·lules Fibroblàstiques.
Les cèl·lules fibroblàstiques reticulars embolcallen fibres reticulars com col·lagen de tipus
III, elastina i laminina tot formant canals o conductes FRC que transporten antígens solubles.
Les cèl·lules fibroblàstiques reticulars formen l’estructura o esquelet dels teixits amb una
elevada cel·lularitat. Serveixen per crear vies per on circulen els limfòcits i les cèl·lules
dendrítiques, pel que faciliten la presentació d’antigen. Les cèl·lules Fibroblàtiques reticulars
expressen molècules com el MHC de classe I i molècules d’adhesió. També secreten
quimiocines com CCL19, CCL21 i CXCL13, que atrauen cèl·lules dendrítiques, limfòcits T i
limfòcits B.
Als limfonodes, el 90% dels limfòcits que hi entren són naive i ho fan a través de les cèl·lules
de les Vènules de l’endoteli alt. Aquestes són vènules5 especialitzades que es troben en
els llocs de migració del limfòcit des de la sang fins l’estroma d’un gangli limfàtic perifèric o
mucós. Les HEV (que reben el seu nom per les seves sigles en anglès) es caracteritzen pel
fet de tenir les cèl·lules endotelials prominents, cuboides i altes que expressen molècules
d’adhesió úniques implicades en la unió dels limfòcits T verges. El 10% dels limfòcits restants,
majoritàriament cèl·lules de memòria, entren a través dels vasos limfàtics aferents.
Els limfòcits T naive o verges tendeixen a concentrar-se als limfonodes. La vènula de l’endoteli
alt és la que permet el homing dels limfòcits T naive al limfonode. L’acumulació d’aquests es
sol donar al paracòrtex, que és on desemboquen les vènules d’endoteli alt.
Entrada i sortida de cèl·lules.
Els limfòcits surten dels vasos sanguinis a través de les HEV i es dirigeixen als limfonodes.
En aquests, els limfòcits B es dirigeixen al còrtex i els T al paracòrtex. Això passa perquè
els limfòcits T expressen receptors de quimiosines CCR7, que reconeixen CCL21 i CCL19.
Aquestes dues quimiocines s’expressen al paracòrtex. Els limfòcits B, en canvi, expressen
els receptors CXCR4 i CXCR5. Aquest permeten interaccionar amb les quimiocines CXCL12
i CXCL13.
5 Vena petita

Immunologia 2020
CEBIO 87
Els limfòcits B s’acumulen formant fol·licles. Si no hi ha resposta són primaris. Si hi ha
resposta són secundaris. En aquests últims es forma el centre germinal. En el centre
germinal hi ha una zona fosca, on trobem centroblasts dividint-se. Aquesta divisió implica
que el domini variable del BCR passa pel procés d’hipermutació somàtica. També hi ha una
zona clara, on s’està fent una selecció dels centròcits en funció de l’afinitat del seu BCR
mitjançant el contacte amb les cèl·lules fol·liculars dendrítiques. Els limfòcits B amb millor
afinitat poden fer un canvi d’isotip i desenvolupar-se en cèl·lula plasmàtica. També en
aquesta zona clara s’està donant la mort per apoptosi dels limfòcits B amb menys afinitat
per l’antigen. Existeix una corona o mantell. És on hi ha immunoglobulines D i cèl·lules B en
repòs, és a dir, que no són específiques per l’antigen al qual s’està responent al centre
germinal.
La Melsa.
La melsa és l’òrgan limfoide secundari més gran. No hi arriben els vasos limfàtics aferents
però està connectada directament amb la circulació sanguínia mitjançant l’artèria
esplènica. La melsa és un òrgan especialitzat en capturar i respondre a antígens que
entren per via sanguínia. Amb una estructura relativament semblant als limfonodes, es
separa en polpa roja i en polpa blanca.
La polpa roja és una xarxa de sinus amb glòbuls vermells, macròfags i algun limfòcits. La
polpa roja és la part encarregada d’emmagatzemar eritròcits i eliminar-ne els envellits. La
polpa blanca és la part de la melsa que envolta les artèries, en forma d'una beina de limfòcits,
principalment limfòcits T. La sigla PALS prové de l'anglès periarteriolar lymphatic sheath. En
aquesta polpa blanca també hi trobem fol·licles limfoides amb cèl·lules B. Hi ha cèl·lules
interdigitants, semblants a les FDCs dels limfonodes que actuen com a cèl·lules
presentadores d’antigen.
Sistema immunitari associat a les mucoses.
El sistema immunitari associat a les mucoses (MALT) és el més complex i abundant del
sistema immunitari. Amb una altíssima activitat immunitària, ha de desenvolupar immunitat
activa contra els patògens que travessen la superfície mucosa i, al mateix temps, controlar
que no hi hagi resposta davant els aliments o la microbiota. Amb el MALT s’estableix una
modulació fina de la resposta immune per tal d’induir una tolerància local (i sistèmica) a
antígens innocus alhora que es manté la vigilància respecte els microorganismes patogènics.
L’estímul antigènic de la microbiota participa en el desenvolupament i el manteniment del
MALT. El sistema immunitari associat a les mucoses es troba a les plaques de Peyer, a les
amígdales, als adenoides, a l’apèndix i a tot el teixit associat a la mucosa gàstrica, respiratòria
i reproductora apart de trobar-lo a la pell.
GALT.
El teixit limfoide associat a la mucosa intestinal es troba format per regions estructurades com
ho són les plaques de Peyer o els fol·licles aïllats. Aquest teixit limfoide fa servir limfòcits
intraepitelials i limfòcits dispersos per la làmina pròpia. El GALT té una irrigació sanguínia
amb la presència de vènules d’endoteli alt i de vasos limfàtics que drenen als limfonodes
mesentèrics. Els limfòcits intestinals es troben en teixits organitzats, on s’indueixen les
respostes immunes, i repartits per l’intestí, on poden realitzar altres funcions efectores.
El GALT està format per diversos tipus cel·lulars, com les cèl·lules de paneth, que generen
pèptids antimicrobians, les cèl·lules de goblet, que produeixen moc que forma una capa per
separar el lumen de les cèl·lules epitelials, les cèl·lules tuft, que produeixen interleuquina 25
i activen les cèl·lules ILC (que acaben estimulant les cèl·lules productores d’interleuquina 13)
i les cèl·lules M.

Immunologia 2020
CEBIO 88
Les plaques de Peyer de les que hem parlat són nòduls limfàtics situats a la submucosa de
l'intestí prim i que tenen una funció similar als ganglis limfàtics. Recobreixen una part
important de l’intestí prim i generalment es troben associats a cèl·lules M.
Les cèl·lules M són cèl·lules de l’epiteli de l’intestí prim amb una membrana apical molt
invaginada i una concavitat anomenada butxaca basolateral on s’acumulen els limfòcits B,
els limfòcits T i els macròfags. L’antigen entra a través d’aquestes cèl·lules, que es situen en
els llocs inductius. Un lloc inductiu és una regió curta de l’epiteli que s’emplaça sobre els
fol·licles limfoides.
Els limfòcits intraepitelials de la mucosa solen ser limfòcits TCD8. Aquests expressen una
integrina única, la α4β7. Es troba involucrada en el homing. En quant a la diversitat dels
limfòcits intraepitelials és destacable que hi ha més cèl·lules T γδ que a la circulació
sanguínia o als limfonodes. Sobre els limfòcits de la làmina pròpia, la gran majoria són
limfòcits T i limfòcits B activats, juntament amb basòfils, mastòcits i macròfags. La majoria de
les cèl·lules T de la làmina pròpia són TCD4. Els limfòcits B del sistema immunitari associat
a les mucoses produeixen immunoglobulines A i poden induir tolerància.
Aquesta immunoglobulina A dimèrica que es produeix es pot unir a un receptor polyIgA de la
membrana basolateral de les cèl·lules epitelials intestinals. Aquest receptor transporta la
immunoglobulina A a la membrana apical, la que està en contacte amb la llum de l’intestí.
Dins la cèl·lula es dona un tall del receptor polyIgA que provoca que un fragment es quedi
unit a la immunoglobulina quan aquesta es trobi al lumen de l’intestí per realitzar la seva
funció.
Les tonsil·les i les adenoides constitueixen les principals àrees inductores de resposta als
antígens intranasals i inhalats a humans, primats i ratolins. Formen una barrera de teixits
limfoides anomenada anell de waldeyer.
En l’epidermis, el teixit limfoide conté queratinòcits, encarregats de produir diverses citocines
proinflamatòries i expressar molècules del MHC de classe II, cèl·lules de langerhans, que
no deixen de ser cèl·lules dendrítiques però amb la diferència que resideixen a l’epidermis, i
limfòcits intraepidèrmics, que majoritàriament són limfòcits Tαβ i limfòcits Tγδ.

Immunologia 2020
CEBIO 89
Tema 17 – Recirculació Limfocitària Les cèl·lules de la immunitat innata no es mouen del teixit on van. Els leucòcits, en canvi, es
mouen pel sistema fent servir els vasos sanguinis i els vasos limfàtics. El moviment dels
leucòcits a través dels vasos sanguinis i limfàtics s’aconsegueix gràcies a les
Molècules d’adhesió cel·lular i a altres tipologies moleculars.
Molècules d’Adhesió Cel·lular. Les molècules d’adhesió cel·lular (CAMs) són molècules expressades a la membrana
cel·lular que permeten la interacció dels leucòcits amb altres leucòcits, amb cèl·lules dels
teixits o amb la matriu extracel·lular. Algunes molècules fan interaccions homofíliques. Altres,
heterofíliques.
Les Molècules d’Adhesió Cel·lular intervenen en els patrons de recirculació dels
leucòcits, en la seva migració als teixits i en la seva extravasació. També s’encarreguen
de l’estabilització del reconeixement d’antigen pel TCR, dels processos de maduració i
diferenciació i de la modulació de la resposta limfocitària.
En funció de les seves semblances estructurals i funcionals, les molècules d’adhesió cel·lular
es classifiquen en Selectines, Sialomucines, Integrines i en la Superfamília de les
immunoglobulines.
Circulació dels Limfòcits. Els limfòcits T naive van de la circulació sanguínia fins als limfonodes, on s’hi estan un
temps monitoritzant si hi ha antígens pels quals tenen el TCR específic. Si no s’activen, al
cap d’un temps surten cap a la circulació sanguínia a través dels vasos limfàtics
eferents, que desemboquen en els ductes. D’aquí, els limfòcits poden anar a la melsa, als
teixits... Els limfòcits estan contínuament en moviment. Aquest moviment continu és el
que es coneix com recirculació limfocitària.
Un limfòcit pot fer un circuit circulatori complet dues vegades al dia sempre i quan no
reconegui cap antigen pel qual tingui un TCR específic. El crcuit circulatori complert comença
a la sang, d’on es pot derivar als teixits o als òrgans limfàtics secundaris. D’aquests aniran a
parar a la limfa, d’on tornaran a la sang.
La cascada de processos que es dona en la interacció entre els leucòcits i la cèl·lula
endotelial és el que els permet realitzar una extravasació. Aquesta cascada consisteix en
un rodolament, una activació, un reforçament i una transmigració.
L’enganxament i rodolament o rebot és l’aproximació del leucòcit a l’endoteli alhora que
redueix la seva velocitat. En aquest procés on intervenen les selectines, els granulòcits i els
monòcits rodolen mentre que els limfòcits reboten.
L’activació és una etapa on la interacció entre les quimiosines i els receptors
d’aquestes determinaran quines cèl·lules reduiran encara més la seva velocitat i, en
conseqüència, seran les que podran fer l’extravassació. Les quimiocines, en aquesta etapa,
tenen un doble paper. Per un costat, les quimiocines fan una quimiotaxi al provocar la
retenció i concentració dels leucòcits sobre les cèl·lules endotelials on es troben. Per altra
banda, les quimiosines realitzen una activació de la funció integrina. La funció integrina
consisteix en la unió de la cèl·lula a la Matriu Extracel·lular o en la transmissió del senyal des
de la matriu fins dins la cèl·lula.

Immunologia 2020
CEBIO 90
El reforçament i aturada és un procés dut a terme per les integrines on els leucòcits aturen
la seva circulació i els canvia la morfologia per tal d’iniciar la transmigració a través de
l’endoteli. Aquest procés de canvi morfològic rep el nom de polarització.
La transmigració és l’últim pas de la cascada i consisteix en un extravasament per diapèdesi
dels leucòcits. Diapèdesi és el nom que rep la sortida dels leucòcits a través dels capil·lars
sanguinis i es duu a terme gràcies a les integrines, el citoesquelet i les col·lagenases.
Molècules de la recirculació limfocitària.
Les Selectines i les Sialomucines.
Una selectina és una qualsevol de les tres proteïnes lligands de glúcids estretament
relacionades que mitjança l'adhesió de leucòcits a les cèl·lules endotelials. Les selectines
reconeixen sialomucines.
Cada molècula selectina és una glicoproteïna transmembrana d'una sola cadena amb una
estructura modular similar, inclòs un domini lectina tipus C extracel·lular dependent de
calci. Aquest domini tipus C és el responsable de la interacció amb les sialomucines.
Les Selectines són la selectina L (CD62L), expressada constitutivament a limfòcits T i B
naive; la selectina P (CD62P), expressada per plaquetes i cèl·lules endotelials activades; i la
selectina E (CD62E), expressada per cèl·lules endotelials activades. La selectina L reconeix
les sialomucines GlyCAM-1, CD34 i Mad-CAM-1. Les selectines P i E reconeixen la
sialomucina PSGL-1.
Les sialomucines, també anomenades adresines vasculars, són proteïnes molt glicosilades
que permeten la unió a selectines. Són sialomucines la GlyCAM-1 i CD34, que s’expressen
de manera constitutiva a les vènules de l’endoteli alt dels limfonodes; Med-CAM-1, que
s’expressa a les cèl·lules endotelials dels vasos de les mucoses i condueix els limfòcits al
teixit limfoide del GALT; i PSGL-1, que s’expressa a neutròfils, monòcits i limfòcits T efectors
i és el principal responsable de la seva unió a l’endoteli activat.
Les selectines són les responsables de guiar els leucòcits cap a determinats teixits,
procés anomenat homing. Les selectines interaccionen amb els carbohidrats de les
sialomucines a través del domini lectina tipus C extracel·lular. Cada selectina uneix uns
carbohidrats determinats. La selectina L, expressada a limfòcits T i B naive, és responsable
de l’entrada dels limfòcits als Òrgans Limfàtics Secundaris a través de les vènules de l’endoteli
alt. La selectina P i la selectina E, expressades a l’endoteli vascular activat, faciliten el
reclutament de cèl·lules efectores al lloc d’inflamació.
En condicions homeostàtiques, els leucòcits viatgen a gran velocitat pels vasos
sanguinis i les cèl·lules endotelials no expressen selectines E i P. Així i tot, els leucòcits
sí expressen la selectina L i la sialommucina PSGL-1 en cas que estiguin activats. En el
moment d’inflamació, les cèl·lules endotelials comencen a expressar les selectines E i P, que
els permeten interaccionar amb la sialomucina PSGL-1, la qual indueix la extravassació dels
leucòcits.
Les Quimiocines.
Les Quimiocines són una gran família de citocines de baix pes molecular amb una estructura
homòloga que estimulen el moviment del leucòcit i regulen la migració dels leucòcits
des de la sang als teixits.
Les quimiocines interaccionen amb els glicosaminoglicans (GAG) carregats
negativament. Els GAGs solen estar units a proteïnes de la membrana cel·lular o de la matriu

Immunologia 2020
CEBIO 91
extracel·lular en estructures anomenades proteoglicans. Quan les quimiocines són
secretades, queden retingudes i concentrades a la superfície cel·lular i a la matriu
extracel·lular, pel que representen un suport sòlid per on les cèl·lules poden reptar i
migrar.
Les quimiocines, apart de determinar quines cèl·lules interaccionaran amb l’endoteli durant el
pas d’activació, activen les integrines tot canviant l’afinitat d’unió als seus lligands. Els
lligands de les integrines són molècules de la superfamília de les immunoglobulines.
Gràcies a l’acció de les quimiocines, les integrines adquireixen major afinitat pel seu lligand.
Les integrines.
Les integrines són proteïnes heterodimèriques formades per una cadena α i una
cadena β unides per enllaços no covalents. Són proteïnes de la superfície cel·lular
importants per a les interaccions entre el limfòcit T i les cèl·lules presentadores d'antigen i per
a la migració dels leucòcits des de la sang fins als teixits. Les integrines tenen un paper
crític en l’extravasació leucocitària.
Hi ha dues principals famílies de les integrines; els membres de cada família d’integrines
expressen una cadena β conservada (β1/CD29 i β2/CD18) associada a diferents cadenes
α. Les Integrines es classifiquen segons la cadena β que expressin. Entre les integrines més
importants es troben αLβ2 (LFA-1), expressada a limfòcits, monòcits, cèl·lules dendrítiques i
neutròfils; αMβ2, expressada a neutròfils, monòcits i macròfags; α4β7 (LPAM-1), expressada
a limfòcits que es dirigeixen a les mucoses; i α4β1 (VLA-4), que s’expressa en monòcits i
limfòcits.
Els leucòcits en repòs expressen integrines de baixa afinitat pels lligands. Als leucòcits
activats són d’alta afinitat. Hi ha diferents estímuls que poden regular l'afinitat pels lligands
de les integrines, entre els quals es troben les quimiocines. Els dominis citoplasmàtics de
les integrines s'uneixen al citoesquelet cel·lular, on es dona la senyalització intracel·lular
a través del receptor de la quimiocina.
Les adhesines.
Les adhesines són els lligands de les integrines i pertanyen a la superfamília de les
immunoglobulines, una gran família de proteïnes que contenen una estructura globular
anomenada domini de tipus Ig o plegament de Ig. Les adhesines també uneixen proteïnes
de la matriu extracel·lular com la fibronectina, el fibrinogen i la laminina.
Les adhesines tenen un baix nivell d’expressió a les cèl·lules de l’endoteli vascular en
repòs. A les zones inflamades, on es produeixen de forma local citocines proinflamatòries
com TNFα, hi ha una inducció de la seva producció.
Són exemples de les integrines les ICAM-1 i ICAM-2, que s’expressen a l’endoteli activat i en
repòs respectivament; VCAM-1, que s’expressa a l’endoteli activat; i PECAM, que s’expressa
als leucòcits activats i a les unions entre cèl·lules endotelials.
L’adhesina PECAM és una molècula crítica en el procés d’extravasació que s’expressa
a les unions intercel·lulars de les cèl·lules endotelials i al limfòcit. Facilitat per l’adhesina
PECAM, el procés d’extravasació requereix de la ruptura transitòria i reversible de les
proteïnes d’unió adherent així com d’una digestió de la membrana basal dels vasos
sanguinis. Aquesta última digestió la duen a terme metal·loproteases.
Extravassació dels Neutròfils i dels Monòcits. Els receptors de quimiocines expressats en el procés d’extravasació són diferents per
neutròfils i per monòcits. També s’observen diferències en l’expressió de les integrines i de

Immunologia 2020
CEBIO 92
les adhesines, el que determina la facilitat amb què aquests tipus cel·lulars extravasaran i
aniran al teixit a fer la seva funció efectora.
Els neutròfils interaccionen amb la selectina L per mitjà de la PSGL-1, el que permet que
hi hagi una interacció de la CXCR1 del neutròfil amb la CXCL18 de l’endoteli. Aquest
interacció suposarà un canvi de conformació de LFA-1 i de MAC-1, augmentant l’afinitat.
Quan seguidament hi hagi una interacció suficient amb ICAM-1, es donarà la transmigració
per mitjà de PECAM-1.
Els monòcits interaccionen amb la selectina L per mitjà de la PSGL-1, el que després els
permet que hi hagi una interacció de CCL2 o CX3CL1 del monòcit amb CCR2 o CX3CR1 de
l’endoteli, respectivament. Aquest interacció suposarà un canvi de conformació de VLA-4 o
de α4β1 i de LFA-1. Quan hi hagi una interacció suficient amb VCAM-1 i ICAM-2, es donarà
la transmigració per mitjà de PECAM-1.
Recirculació als Òrgans Limfàtics Secundaris. Al paracòrtex dels limfonodes hi ha les vènules d’endoteli alt. Aquestes expressen, de forma
constitutiva, les adresines vasculars CD34 i GlyCAM-1. Això permet el homing o l’entrada de
limfòcits T i B naive, que expressen la selectina L, al limfonode.
Les vènules d’endoteli alt són vènules post-capil·lars que tenen cèl·lules endotelials
amb forma cúbica permanentment activades. Tenen una expressió constitutiva de les
adresines CD34 i GlyCAM-1, que permeten el homing de limfòcits T i B naive per la seva
expressió de selectina L. Per això, les vènules de l’endoteli alt són responsables de la
recirculació dels limfòcits naive per tots els òrgans limfoides secundaris excepte la
melsa. S’estima que, al limfonode, s’extravasen 14,000 limfòcits per segon.
La majoria de limfòcits naive entren als òrgans limfàtics secundaris en busca d’antigen.
Si no el troben després d’un període de temps concret d’entre 12 i 18 hores per les cèl·lules
T i unes 24 hores pels limfòcits B, els limfòcits naive surten a través dels vasos limfàtics
eferents per dirigir-se a la circulació limfàtica.
Les cèl·lules T migren a la zona T del limfonode a una “velocitat” de 12µM per minut a
través d’un moviment que sembla aleatori. En realitat, la migració dels limfòcits T a la zona
T està dirigida per quimiocines. Es calcula que una única cèl·lula dendrítica interactua amb
aproximadament 5,000 cèl·lules T per hora a la zona T.
Les cèl·lules T i B naive recirculen a l’interior dels limfonodes per l’expressió dels
receptors de quimiocines CCR7 i CXCR5 i els seus lligands.
CCR7 és un receptor de les quimiocines CCL19 i CCL21. CCR7 permet la migració a
través de la zona T. CCL21 està produida per les vènules de l’endoteli alt i per cèl·lules
estromals dels teixits limfoides. CCL19 està produida per les cèl·lules estromals de l’àrea T i,
en menys quantitat, per les cèl·lules dendrítiques interdigitants.
CXCR5 és un receptor de CXCL13, una quimiocina produida per les cèl·lules fol·liculars
dendrítiques localitzades als fol·licles. La interacció CXCR5-CXCL13 determina
l’acumulació de les cèl·lules B per formar els fol·licles. CXCR5 s’indueix en les cèl·lules
T helper fol·liculars (Thf) i els permet migrar a la zona clara del centre germinal.
La migració de les cèl·lules T al paracòrtex es fa seguint els conductes que formen les
cèl·lules fibroblàstiques reticulars (FCR), on s’uneixen les quimiocines que permeten que
els limfòcits transitin com per carreteres on també circulen les cèl·lules dendrítiques. Això,

Immunologia 2020
CEBIO 93
doncs, facilita la interacció entre ambdues cèl·lules i, conseqüentment, facilita el
reconeixement d’antigen.
Una vegada han reconegut antigen, les cèl·lules efectores surten pel vas eferent a la
circulació limfàtica fins al ducte toràcic, on es volca la limfa a la circulació sanguínia a
través de la vena subclàvia.
Diferències entre Limfòcits T efectors i limfòcits T naive. Els limfòcits T activats redueixen l’expressió de molècules que els condueixen als
òrgans limfàtics i augmenten l’expressió de molècules que participen en la seva
migració als llocs perifèrics de la infecció i lesió tissular. Algunes de les molècules que
veuen reduïda la seva expressió són la selectina L i el receptor per quimiocines CCR7.
Algunes de les que veuen augmentada la seva expressió són les integrines LFA-1 i VLA-4,
els lligands per les selectines E i P i diversos receptors per quimiocines. L’activació del
limfòcit també augmenta l’expressió de CD44, un receptor per l’àcid hialurònic de la
matriu extracel·lular. La unió de CD44 al seu lligand ajuda a retenir els limfòcits T efectors
en els llocs d’infecció i lesió tissular.
Les cèl·lules T efectores també augmenten l’expressió de CD2 i LFA-1, unes molècules
d’adhesió; de CD45RO, que s’associa al TCR i a CD4 alhora que té una activitat fosfatasa
que baixa el llindar d’activació; i de VLA4, induint el homing a l’endoteli vascular inflamat. Les
cèl·lules T efectores, en canvi, redueixen l’expressió de la selectina L, que indueix al homing
al limfonode, i de CD45RA, que provoca el splicing diferencial del mRNA de CD45.
Extravasació dels limfòcits T efectors i memòria. Els limfòcits T efectors fan homing a les zones d’infecció i hi detecten l’endoteli inflamat
gràcies a les quimiocines que es generen al focus inflamatori. El perfil migratori de la
cèl·lula T efectora depèn del lloc on la cèl·lula dendrítica hagi capturat l’antigen que
presenta al limfòcit T naive. El tipus de receptor de quimiocines de les cèl·lules efectores,
juntament amb la integrina, determinen el homing de la cèl·lula T efectora.
Els receptors de quimiocines CCR9, que s’uneix a CCL25, i α4β7, que s’uneix a MadCAM-1,
indiquen homing a la mucosa. En canvi, els receptors de quimiocines CCR4, que s’uneix a
CCL17, CCR10, que s’uneix a CCL27, CLA, que s’uneix a la selectina E, i α4β1, que s’uneix
a VCAM-1, indiquen un homing a la pell.
Extravasació a la pell
Els limfòcits T efectors que s’extravasen a la pell són els que han estat activats als
limfonodes proximals, els quals reben la limfa procedent de la zona inflamada de la pell.
Durant el reconeixement d’antigen presentat per les cèl·lules dendrítiques activades in situ,
s’indueix l’expressió de les molècules necessàries per poder migrar a la pell. Les molècules
necessàries per poder migrar a la pell són CLA, CCR4 i /o CCR10 i α4β1.
Paper de les molècules d’adhesió en la presentació d’antigen. Les molècules d’adhesió són molècules de la superfície cel·lular la funció de les quals
és promoure les interaccions adhesives amb altres cèl·lules o amb la matriu
extracel·lular. Aquestes molècules són importants en les interaccions contínues dels
LFA-1 - ICAM-1 LFA-1 - ICAM-2 ICAM-3 - DCSIGN CD2 - CD58

Immunologia 2020
CEBIO 94
limfòcits T amb les cèl·lules dendrítiques als limfonodes. Aquestes interaccions són
transitòries i es donen entre molècules d’adhesió i els seus lligands. En destaquen 4:
Les integrines es troben en un estat de baixa afinitat a les cèl·lules en repòs. Les integrines
pateixen un canvi conformacional que augmenta l’afinitat per seu lligand quan hi ha
una senyalització a través del TCR. Per tant, s’estabilitza la interacció.

Immunologia 2020
CEBIO 95
Tema 18 – Resposta cel·lular. Activació de les cèl·lules T.
Presentació d’Antigen. Les cèl·lules dendrítiques que es troben als teixits de barrera capturen i processen l’antigen
per, seguidament, transportar-lo cap als limfonodes proximals a través dels vasos limfàtics.
Al paracòrtex dels limfonodes poden presentar l’antigen als limfòcits T naive que allà es
trobin. Aquesta presentació no és aleatòria. Requereix de moltes interaccions, les quals van
realitzant transitòriament els limfòcits T. En cas que hi hagi reconeixement, el limfòcit
s’activarà i serà un limfòcit T efector. Aleshores sortirà del limfonode cap al teixit, on realitzarà
la seva funció efectora.
Als limfonodes, els limfòcits T interaccionen contínuament amb les cèl·lules presentadores
d’antigen. Aquestes interaccions són transitòries entre les molècules d’adhesió i els seus
lligands. Aquestes interaccions poden ser de 4 tipus: LFA-1 – ICAM-1, LFA-1 – ICAM-2,
ICAM-3 – DC-SIGN i CD2 – CD58. LFA-1, ICAM-3 i CD2 es troben a la membrana de la
cèl·lula T, mentre que ICAM-1, ICAM-2, DC-SIGN i CD58 es troben a la membrana de les
cèl·lules presentadores d’antigen.
Etapes de la resposta cel·lular T. Una vegada s’ha produït l’acostament pot tenir lloc la primera etapa de la resposta cel·lular
T. Aquesta primera etapa consisteix en el reconeixement d’antigen per part dels limfòcits
T, que tan sols dependrà de la interacció TCR-complex MHC/pèptid. En aquest primer senyal
o etapa hi tenen un paper mot rellevant els coreceptors.
La segona etapa serà l’activació cel·lular. En l’activació, hi ha d’haver un senyal entre la
molècula coestimuladora del limfòcit T i el seu lligand. Totes aquestes interaccions resulten
en la transducció de senyals a l’interior de la cèl·lula, el que es coneix com a cascada
bioquímica de segons missatgers. Aquesta cascada consisteix en l’activació de la transcripció
de gens, la repressió de la transcripció d’altres gens, l’expressió de noves proteïnes de
membrana, la secreció de citocines activadores del creixement autocrí i paracrí com IL-2 i la
inducció de l’activitat mitòtica. Aquest últim fenomen de la cascada es tradueix en una
proliferació i expansió de les cèl·lules T.
La tercera etapa de l’activació cel·lular és la de l’activació de la funció efectora, la qual
requereix de senyals d’adhesió i, sobretot, de citocines. Finalment, la quarta etapa es
correspon amb la regulació de la resposta immunològica.
Activació del limfòcit T.
L’activació del limfòcit T és el resultat de la generació de 3 senyals. El primer senyal és
el derivat de la interacció entre el TCR i el complex pèptid/MHC. En la generació d’aquest
primer senyal també hi participa la senyalització dels coreceptors CD4 o CD8. El segon senyal
és el de la coestimulació mediada per la interacció de CD28 del limfòcit amb CD80 ó CD86
de la cèl·lula presentadora d’antigen. Enlloc d’això, el segon senyal es pot dura a terme per
senyals derivats de molècules de la immunitat innata. Finalment, el tercer senyal és el resultat
de la interacció de citocines amb el seu receptor. Aquest tercer senyal és principalment
responsable de la diferenciació de les cèl·lules T activades en cèl·lules efectores de diferents
tipus.
1r senyal d’activació: el reconeixement d’antigen pel complex TCR/MHCpèptid.
El TCR no senyalitza directament, ho fa a través del CD3. Les cadenes d’aquest tenen
motius ITAM en el seu costat citoplasmàtic. El complex TCR/CD3 el formen una o dues
molècules del TCR associada/es a les 6 cadenes del CD3: una cadena δ, una γ, dues ε i el
dímer de cadenes ζ. L’associació és de tipus no covalent amb el TCR. Les cadenes ζ, de

Immunologia 2020
CEBIO 96
pocs aminoàcids extracel·lulars, és pràcticament intracel·lular. Els motius ITAM
(Immunoreceptor Tyrosine based Activation Motifs) participen en la transmissió de senyals al
reclutar tirosina quinases un cop s’han fosforilat.
Quan el TCR interacciona amb el complex pèptid/MHC, la quinasa citoplasmàtica Fyn del
limfòcit fosforila la quinasa Lck. Aquesta, ja activada, fosforila els ITAMs de CD3γ, δ, ε i ζ.
Posteriorment, la tirosina quinasa ZAP-70 s’uneix als ITAM fosforilats de CD3ζ i Lck acaba
de fosforilar ZAP-70, que s’activa. La complerta activació necessita, per tant, la interacció del
coreceptor amb la molècula del MHC. La fosfatasa CD45 regula la fosforilació de Lck i Fyn
en estat de repòs però és apartada del TCR quan es produeix la interacció amb el complex
pèptid/MHC.
A la cèl·lula en repòs, la tirosina quinasa Lck, associada al coreceptor CD4 o CD8, es troba
separada del complex TCR/CD3 perquè està inclosa en els lípid rafts o balses lipídiques, com
moltes molècules d’adhesió. Aquestes són microdominis de membrana rics en esfingomielina,
glucoesfingolípids i colesterol, pel que són més rígides. El complex TCR/CD3, en canvi, es
troba fora dels lípid rafts. Quan el TCR interacciona amb el complex pèptid/MHC, migra als
lípid rafts i s’associa al coreceptor CD4 o CD8.
Quan ZAP-70 està activat, fosforila l’adaptador LAT. La fosforilació crea llocs lliures per a la
unió de SLP-76 i altres molècules, el que inicia una cascada de senyalització. SLP-76 és el
responsable dels canvis en el citoesquelet del limfòcit T. Per tant, és el que li canvia la forma
en activar-se. Per altra banda, A LAT fosforilat s’hi pot unir la fosfolipasa Cγ i el factor
intercanviador del nucleòtid guanina (GEF). La fosfolipasa Cγ (PLCγ) fosforilada catalitza la
hidròlisi d’un fosfolípid de la membrana plasmàtica anomenat PIP2que genera dos productes:
el 1,4,5-trifosfat d’inositol (IP3) i el diacilglicerol (DAG) unit a la membrana. IP3 i DAG actuen
en dues vies de transmissió de senyals dels limfòcits T.
IP3 es difon pel citosol fins al reticle endoplasmàtic per unir-se al seu receptor, un canal de
calci activat per lligand, i estimula la sortida d’ions calci del reticle endoplasmàtic cap al
citoplasma i l’entrada de calci del medi extracel·lular. El calci alliberat provoca un augment
ràpid del calci lliure citosòlic als pocs minuts de l’activació del limfòcit T. L’augment de Calci
citoplasmàtic permet unir a la calmodulina, que activa la calcineurina i aquesta activa NFAT.
El diacilglicerol (DAG), el segon producte de l’escissió del PIP2, és un lípid unit a la membrana
que atreu la proteïna quinasa C (PKC). Hi ha diferents formes de PKC que participen a la
generació de factors de transcripció actius. La combinació de l’augment del calci citosòlic lliure
i DAG activa certes isoformes de PKC associades a la membrana al induir un canvi
tridimensional que deixa accessible el lloc catalític de la quinasa als seus substrats. La PKC
activa el factor de transcripció NFκB.
D’altra banda, GEF estimula el canvi de GTP per GDP a la proteïna Ras, una petita proteïna
lligadora del nucli de guanina (o proteïna G petita). Arran d’aquest canvi, la proteïna Ras
pateix un canvi conformacional i pot aleshores reclutar la proteïna Raf, que serà l’encarregada
d’iniciar la via de pes MAP quinases tot activant la MAP quinasa Erk.
L’activació de la MAP quinasa Erk li permet entrar al nucli, on fosforila el factor de transcripció
Elk-1. Aquest estimula la transcripció del gen FOS. A través d’una altra cascada de
senyalització s’acaba fosforilant la proteïna quinasa Jun que, juntament amb FOS fosforilada,
provoca l’activació de moltíssims gens transcripcionals.
2n senyal d’activació: Molècules coestimuladores.
Les molècules coestimuladores proporcionen el segon senyal d’activació o coestimulació.
Són necessàries per a l’activació i diferenciació de limfòcits T naive a més del reconeixement

Immunologia 2020
CEBIO 97
d’antigen pel TCR. El coestimulador CD28 es troba expressat constitutivament als limfòcits T
naive mentre que ICOS s’expressa a limfòcits T activats. Els lligands de CD28 són B7-1 i B7-
2 i tenen una expressió baixa o negativa a les cèl·lules presentadores d’antigen en repòs però
s’indueixen amb PRRs i citocines de la resposta innata. Hi ha molècules coestimuladores com
CTLA-4 i PD-1 que s’expressen a limfòcits T activats i que participen en la regulació.
Els productes microbians i les citocines produïdes per les cèl·lules de la immunitat innata
indueixen l’expressió de molècules coestimuladores a les cèl·lules presentadores d’antigen.
En absència de coestímul, els limfòcits T que reconeixen l’antigen no hi responen, pel que
entren en estat d’anèrgia.
Quan es produeix la interacció de CD28 amb el seu lligand es genera tota una cascada de
senyalització. CD28 té, a la seva cua citoplasmàtica, un motiu de tirosina que permet la
interacció amb la proteïna reguladora de la fosfoinositol 3 fosfat quinasa, 2 motius rics en
prolina que permeten la unió de la tirosina quinasa ITK i de la quinasa LcK. Quan CD28
interactua amb els seu lligand (B7-1 i B7-2), fa servir GEF i activa Rac. Rac activarà ña via
de les MAP quinases. Això durà a l’activació de la quinasa Jnk, que es fosforila. Al estar
activada, Jnk fosforila Jun, que entrarà al nucli per unir-se a FOS i formar AP-1, que indueix
la transcripció de gens.
La cascada de les MAP quinases activada pel segon senyal d’activació comença per GEF,
segueix per Rac, fosforila MAPKKK, aquesta a MAPKK i aquesta a MAPK. Finalment hi ha
una translocació al nucli.
La suma del primer senyal d’activació, mitjançant Ras, dona lloc a FOS activat al nucli i el
segon senyal d’activació, mitjançant Rac, dona lloc a Jun activat al nucli. Tots dos formen AP-
1. Per tant, l’acció dels dos senyals d’activació fan, en comú, l’activació del factor AP-1.
NFAT, AP-1 i NFκB s’uneixen al promotor del gen de la IL2. Cal que s’hi uneixin tots perquè
comenci la transcripció del gen. Per tant, es pot considerar que el sistema d’expressió de la
IL2 està molt regulat.
Interleucina 2.
Les citocines desenvolupen funcions clau en les respostes immunitàries adaptatives. En elles,
les principals fonts de citocines són els limfòcits T. La citocina més important produïda pels
limfòcits T després de la seva activació és la Interleucina-2. La IL-2 és un factor de
creixement, supervivència i diferenciació pels limfòcits T. Desenvolupa una funció
important en la regulació de les respostes dels limfòcits T en virtut del seu paper crucial
en el manteniment dels limfòcits T reguladors. La Interleucina 2 actua a les mateixes
cèl·lules que la produeixen o a limfòcits T adjacents.
L’activació dels limfòcits T pels antígens i els coestimuladors estimula la transcripció del gen
de la IL-2 i la síntesi i secreció de la proteïna. La producció de IL-2 és ràpida i transitòria. Els
limfòcits T secreten la Interleucina a la interacció entre ells i les cèl·lules presentadores
d’antigen. Els receptors per les citocines dels limfòcits també tendeixen a localitzar la
interacció, pel que la citocina i el seu receptor arriben a concentracions locals prou altes com
per iniciar les respostes cel·lulars.
Els receptors funcionals per a la IL-2 s’expressen de forma transitòria en l’activació
dels limfòcits T verges i efectors. Els limfòcits T reguladors sempre expressen
receptors per la Interleucina. El receptor per la IL-2 (IL-2R) consta de tres proteïnes
associades de forma no covalent: IL-2Rα, IL-2/15Rβ i γc; alternativament anomenades CD25,
CD122 i CD132 respectivament.

Immunologia 2020
CEBIO 98
La IL-2 s’uneix a la cadena α sola amb una afinitat baixa i això no produeix cap senyal
citoplasmàtica ni cap resposta biològica detectable. IL-2/15Rβ contribueix a la unió de la IL-2
i connecta amb les vies de transducció de senyals. Malgrat la cadena γc no participa
directament la unió amb IL-2, es necessita la seva associació al complex del receptor per a
la unió amb alta afinitat a la IL-2 i per a l’activació completa de les vies de transducció del
senyal.
Els complexes IL-2/15Rβγc s’expressen en baixes quantitats en els limfòcits T en repòs i
s’uneixen a la IL-2 amb una Kd d’aproximadament 10-9M. Les cèl·lules que expressen IL-2Rα
i formen complexes IL-2Rαβγ poden unir-se a la IL-2 amb més força, amb una Kd al voltant
de 10-11M, i l’estimulació del creixement d’aquestes cèl·lules es produeix a concentracions
similars de IL-2.
La IL-2, produïda en resposta a l’estímul d’un antigen, és un estímul per la inducció de IL-
2Rα, el que proporciona un mecanisme pel qual les respostes dels limfòcits T s’amplifiquen a
si mateixes.
Expressió de Gens per l’activació.
A la gràfica següent es mostra la cinètica aproximada de l’expressió d’algunes molècules
després de l’activació dels limfòcits T per antígens i coestimuladors. Els exemples il·lustratius
comprenen un factor de transcripció (c-Fos), una citocina (IL-2) i proteïnes de la superfície
(CD69, IL-2Rα, lligand de CD40). Aquestes proteïnes es solen expressar en baixes quantitats
als limfòcits T naive i les indueixen senyals activadores. El CTLA-4 s’indueix al cap d’una
setmana de l’activació. Les cinètiques són estimacions i variaran amb la naturalesa de
l’antigen, la seva dosi i la seva persistència.
Presentació d’antigen i reconeixement. Perquè es produeixi l’activació hi ha d’haver un encaix entre un limfòcit T i la cèl·lula
presentadora d’antigen. Això requereix de moltes interaccions. Es formen interaccions molt
bones, ja que han de ser prou estables per tal que el limfòcit T desencadeni tots els
mecanismes que li pertoquen. . Aquesta interacció requereix d’una reestructuració de les
proteïnes de membrana a la que anomenem sinapsi immunològica.
La sinapsi immunològica és la superfície de contacte entre la cèl·lula presentadora d’antigen
i el limfòcit T. Quan es produeix la primera interacció entre LFA-1 i ICAM-1, que permet que
s’acostin la cèl·lula presentadora d’antigen i el limfòcit T, es pot donar la interacció entre el
TCR i el complex pèptid/MHC. Quan això passa, el TCR migra cap a l’interior de les balses
lipídiques, on hi havia gran quantitat molècules d’adhesió, i aquestes se separen de la regió
central i van cap al voltant del punt d’interacció. Això és perquè impedirien el bon contacte
entre el TCR i el complex pèptid/MHC al ser molt llargues. Enlloc d’això, es reorganitzen les
proteïnes de membrana per tal que els complexes quedin al centre i les molècules d’adhesió
es quedin fora, estabilitzant. En la sinapsi hi participen altres molècules.
Les molècules del limfòcit T que es mobilitzen ràpidament al centre de la sinapsi són el
complex del TCR, els coreceptors CD4 o CD8, receptors per coestimuladors, enzims quinases
i proteïnes adaptadores que s’associen a les cues citoplasmàtiques dels receptors
transmembranals. En aquesta posició de la sinapsi, anomenada c-SMAC, la distància entre
la membrana plasmàtica del limfòcit T i la de la cèl·lula presentadora d’antígens és d’uns
15nm. Les integrines es mantenen a la perifèria de la sinapsi, on funcionen estabilitzant la
unió del limfòcit T a la cèl·lula presentadora d’antigen, i formen la porció perifèrica del SMAC
anomenada p-SMAC. En aquesta part externa de la sinapsi, les dues membranes estan
separades uns 40nm. La part externa rep el nom de d-SMAC.

Immunologia 2020
CEBIO 99
El TCR i les molècules senyalitzadores (LAT) formen nanoagregats homotípics a limfòcits T
en repòs. Amb la interacció amb l’antigen el TCR i les molècules senyalitzadores s’agrupen
formant agregats de mida més gran, microagregats. Els microagregats es fusionen per formar
el complex supramolecular central o cSMAC a la zona de contacte entre cèl·lules T i cèl·lules
presentadores d’antigen.
La membrana plasmàtica és dinàmica i els receptors d’antigen i les molècules de senyalització
es desplacen en funció del metabolisme de la cèl·lula, de si el limfòcit està activat, si està en
repòs... La zona d’interacció entre el limfòcit T i la cèl·lula presentadora d’antigen varia al cap
del temps. Als 15 minuts el TCR es troba localitzat en un anell perifèric mentre que LFA-1 es
troba al centre. És el que s’anomena una sinapsi immunològica immadura. Als 30 minuts, en
canvi, el TCR és al centre i es troba envoltat per un anell perifèric de LFA-1. És el que
s’anomena una sinapsi immunològica madura.
La formació de la sinapsi immunològica permet el canvi de conformació de la LFA-1 a la forma
d’alta afinitat. Aquest canvi estabilitza la interacció cèl·lula T – Cèl·lula presentadora d’antigen.
Les principals interaccions implicades en la immobilització depenen de l’actina del
citoesquelet del TCR i de la integrina a la sinapsi immunològica, incloent-hi els factors que
contribueixen a la desactivació de LFA-1. El SLP-76 activat facilita l’acostament de kindlin-3 i
RIAM a la integrina. Rap/RapL/RIAM promouen l’associació de talin amb la cadena β del LFA-
1. El resultat de l’associació és el canvi conformacional cap a la forma d’alta afinitat de la LFA-
1.

Immunologia 2020
CEBIO 100
Tema 19 – Resposta cel·lular T. Mecanismes Efectors. Els limfòcits T, en reconèixer antigen presentat per la cèl·lula presentadora d’antigen,
s’activen. L’activació d’un limfòcit T el porta a produir IL-2, pas amb què comença la
proliferació i expansió clonal. A partir del procés d’expansió, els limfòcits es converteixen en
els diferents tipus que n’hi ha. Els limfòcits sortints de la diferenciació es separen en limfòcits
T efectors i limfòcits T de memòria.
El tercer senyal d’activació ve donat per citocines i molècules d’adhesió. Les citocines en són
l’element més rellevant, però, ja que determinen el tipus de limfòcit T efector.
Diferenciació a limfòcits T CD4+ efectors. El procés de diferenciació o polarització consisteix en 3 etapes. La primera etapa és la
inducció, que permet un expressió dels factors de transcripció específics de cada tipus efector
i que determinaran el patró de citocines que secretarà cada tipus de limfòcit T efector. La
segona etapa és la de compromís, on s’estabilitza el fenotip de limfòcit T efector perquè es
donen una sèrie de canvis epigenètics en les zones reguladores dels gens de les citocines
que porten a una producció estable d’algunes d’aquestes. Finalment, l’última etapa és la de
l’amplificació, en la qual les citocines secretades pels limfòcits T promouen la diferenciació de
més cèl·lules del mateix tipus efector i inhibeixen la diferenciació d’altres tipus efectors.
D’alguna manera, per tant, s’estabilitza el fenotip efector i s’inhibeix la diferenciació de la resta
de tipus de cèl·lules T efectores. En aquest cas parlem de cèl·lules T helper polaritzades. És
un fet que es veu, sobretot, en els limfòcits T helper perquè hi ha diferents tipus efectors. No
ho veiem en els limfòcits TCD8 que, malgrat parlar de limfòcits T efectors, no se n’han descrit
diferents tipus efectors. Així i tot, hi ha estudis que indiquen que hi ha diferents tipus de
limfòcits TCD8, com els reguladors.
Canvis epigenètics i citocines.
Els Canvis epigenètics, com modificacions de les histones o de la metilació del DNA,
indueixen canvis en l’estructura de la cromatina que deixen accessibles les zones reguladores
de davant del promotor dels gens de les diferents citocines als factors de transcripció de DNA.
Aquests canvis estabilitzen els programes d’expressió gènica durant la resposta T.
Per tant, en un limfòcit TCD4 que s’està convertint en un limfòcits T helper 2, el locus on es
troba la interleuquina 4 quedarà accessible als factors de transcripció. Quan es produeixi la
diferenciació, produirà aquesta interleuquina. Això li permetrà diferenciar-se i provocar la
diferenciació d’altres limfòcits en limfòcits T helper 2.
El que determina en quin tipus de limfòcit T efector es diferencia un limfòcit T són els factors
de transcripció de llinatge cel·lular, el quals venen determinats per les citocines. Per tant, es
pot afirmar que les citocines determinen en quin tipus de limfòcit T efector s’ha de diferenciar
una cèl·lula T.
Quan un limfòcit T reconeix antigen a través del TCR, comença tota la cascada de
senyalització associada. Aquesta cascada, juntament amb la que prové de la interacció de la
IL-4 amb el seu receptor (que produeix l’activació del factor de transcripció Stat-6), el limfòcit
expressa el factor de transcripció Gata3. El factor de transcripció Gata3 és el que fa possible
que el limfòcit T es converteixi en un que és productor de les interleuquines 4, 5 i 13. Per
tant, aquest limfòcit T secretarà les mencionades citocines. La interleuquina 4 actuarà de
forma autocrina, reforçant el fenotip Th2.
En canvi, si quan es dona la interacció TCR-MHC/pèptid hi ha interleuquina 12 al medi, aquest
s’unirà al seu receptor expressat a la membrana i activarà el factor de transcripció Stat-4. En

Immunologia 2020
CEBIO 101
conseqüència, hi haurà una expressió de T-Bet, un factor de transcripció que estimula la
transcripció del gen de l’interferó γ, provocant-ne l’expressió. Aquest interferó serà expulsat
al medi on de manera autocrina s’unirà al receptor del limfòcit T, activarà Stat-1 i reforçarà
l’expressió de T-Bet. T-Bet determina que el limfòcit sigui un T helper 1.
T-bet impedeix la diferenciació a Th2 inhibint Gata3. Gata3 impedeix la diferenciació a Th1
inhibint l’expressió de IL12Rβ2 i Stat4.
L’antigen i l’entorn determinen com són les cèl·lules presentadores d’antigen i quines citocines
secreten. El que determina quines citocines faran les cèl·lules T és l’antigen i on es trobi
aquest. Segons com estigui funcionant la immunitat innata, la cèl·lula dendrítica produirà
unes o altres citocines. En conseqüència, la cèl·lula presentadora d’antigen determinarà en
quin tipus de limfòcit es tronarà aquell que s’ha de diferenciar.
En presència de IL-12 i Interferó gamma, les cèl·lules T naive es diferenciaran en Limfòcits T
helper 1. En presència de IL-4, es diferenciaran en T helper 4. En cas que no hi hagi PAMPS
però sí que hi hagi IL-10 i TGFβ, els limfòcits T passaran a ser limfòcits T reguladors.
Per tant, es pot dir que la immunitat innata influeix en la adaptativa i que aquest és un dels
mètodes que té per fer-ho.
Diferenciació de limfòcits TCD4 helper efectors.
L’activació via TCR en funció de la presència de citocines produïdes determina el tipus de
limfòcit T efector. Els caracteritzem per l’expressió dels factors de transcripció. La
diferenciació d’un tipus efector inhibeix la diferenciació dels altres. Es dona el cicle de
potenciació del tipus efector diferenciat.
Les citocines actuen com a 3er senyal tot induint l’expressió de diferents factors de
transcripció (FoxP3, RORgT, T-bet, GATA-3) que determinen el desenvolupament a diferents
tipus de limfòcits TCD4 efectors. El tercer senyal fa possible la diferenciació dels limfòcits T
CD4+ activats a diferents tipus de limfòcits T helper o col·laboradors efectors.
La rellevància que, a partir d’un limfòcit T naive i d’una cèl·lula presentadora d’antigen hi hagi
una diferenciació en limfòcits T helper 1, T helper 2, T helper 17 o T reguladors induïts; és
que cadascun d’aquests tipus cel·lulars serà important per la resposta a diferents tipus
d’antigen.
Els Limfòcits T helper 1, per exemple, són molt importants en l’activació dels limfòcits TCD8
citotòxics i són molt rellevants en la resposta a patògens intracel·lulars. Els limfòcits T helper
2 seran rellevants en la resposta a patògens extracel·lulars i per ajudar a activar limfòcits B.
Els limfòcits T helper 17 són un nucli d’infecció i tenen molta importància en les respostes a
la pell i mucoses, així com als fongs. Els limfòcits T reguladors induïts destaquen per la seva
funció en el manteniment de la tolerància i la finalització de la resposta.
Diferenciació de limfòcits TH1.
Els limfòcits T helper 1 són importants en la resposta a patògens intracel·lulars. Quan els
limfòcits T es diferencien en presència de IL-12 i interferó gamma, es converteixen en limfòcits
T helper 1. Aquests destaquen per la producció tant de IL-12 com de INFgamma.
Són importants en la resposta a patògens intracel·lulars perquè una de les seves funcions
principals és induir l’activació dels macròfags. L’interferó gamma també influeix en el canvi
d’isotip a IgG.
Els limfòcits T helper 1 provoquen l’activació dels macròfags per acció de l’IFN-g i la interacció
CD40L-CD40. Aquesta interacció augmenta la producció de ROS i RNS, pel que s’obté més

Immunologia 2020
CEBIO 102
capacitat microbicida. A més, també secreta les citocines TNFα, IL12 i quimiocines.
Finalment, els limfòcits T helper 1 provoquen un augment de l’expressió de molècules del
MHC i de molècules coestimuladores (B7).
Els limfòcits Th1 també contribueixen a la bona diferenciació dels limfòcits TCD8 cap a
limfòcits TCD8 efectors arran de la seva producció de citocines. També secreten IL-3 i Factor
estimulador de colònies de monòcits i macròfags, pel que també indueixen la diferenciació de
macròfags i de diferenciació de les cèl·lules hematopoètiques a macròfags.
Amb la secreció de TNα, estimulen les cèl·lules endotelials i augmenten l’extravasació de
limfòcits T al lloc de la inflamació i infecció, facilitant el dur a terme la seva funció. També
produeixen la quimiocina CCL2, que atreu macròfags. D’alguna manera, doncs, els Th1
indueixen participació macròfags en resposta. Tenen un mecanisme on l’expressió de fas
provoca la mort de cèl·lules. Els Th1, amb expressió de lligand fas quan estan activats, poden
provocar la mort de la cèl·lula diana.
Diferenciació de limfòcits TH2.
Els limfòcits Th2, en presència de IL4 produïda per mastòcits o eosinòfils en resposta a
patògens extracel·lular com helmints, es diferencien. Es converteixen en limfòcits T helper
productors de IL-4, IL-5 i de IL-13. Tenen funció important en la producció de determinats
isotips d’immunoglobulines com IgE i en l’activació eosinòfils i de secrecions mucoses. Molt
rellevants en resposta a patògens extracel·lulars.
Les funcions dels limfòcits Th2 es duen a terme perquè la interleuquina 4 indueix el canvi
d’isotip d’immunoglobulina a IgE i, per tant, facilita la sensibilització de mastòcits. A més,
estimula la producció d’anticossos neutralitzants. També és possible gràcies a la producció
de IL-5, que activa eosinòfils, i a la producció de IL-13. Aquesta última interleuquina es
combina amb IL-4 per augmentar el peristaltisme intestinal i la secreció mucus així com per
afavorir l’activació de macròfags M2, que contribueixen a la reparació del teixits.
En conseqüència, es pot afirmar que la resposta dels limfòcits Th2 és una molt associada a
les mucoses gàstrica i respiratòria, on es produeixen les al·lèrgies.
Diferenciació de limfòcits TH17.
Els Th17 són aquells que es diferencien quan a l’entorn hi ha IL-6, IL-1 i TGFβ. Aquestes
citocines indueixen producció IL-21, que és qui estabilitza fenotip juntament amb IL-23.
Aquestes cèl·lules Th17 produeixen IL-17 i IL-22. IL-17 té un paper important en inflamació.
IL-22 té un paper important en barrera al estimular producció pèptids antimicrobians. Limfòcits
Th17 són rellevants en activació de neutròfils i de cèl·lules epitelials. Són molt importants a la
resposta a patògens extracel·lulars com bacteris i, sobretot, fongs.
Els limfòcits Th17 produeixen IL-17 i IL-22. IL-17 genera una resposta proinflamatòria potent
al induir l’expressió de citocines proinflamatòries com IL-1, IL-6 i TNFα; de quimiocines (CXC)
que indueixen la quimiotaxi de leucòcits, sobretot neutròfils, afavorint el manteniment de la
inflamació i infiltració de leucòcits al teixit; de factors creixement per a neutròfils com G-CSF
i al induir també un augment d’expressió d’ICAM-1 als teixits. La combinació de IL-17 i de IL-
22 indueix la secreció de pèptids antimicrobians per part de cèl·lules epitelials.
Un quart tipus de limfòcits T helper són els fol·liculars. Són essencials per la resposta dels
limfòcits B2 fol·liculars. Els limfòcits B2 fol·liculars són els més típics, els que es troben als
fol·licles limfoides al limfonode o a la melsa i que, a través de la col·laboració amb els limfòcits
T helper fol·liculars, desenvolupen la seva funció. Per tant, es pot dir que els Limfòcits T
helper fol·liculars contribueixen a l’activació dels limfòcits B, a la Hipermutació Somàtica i al
canvi d’isotip.

Immunologia 2020
CEBIO 103
Els limfòcits T helper fol·liculars, quan s’activen en presència d’IL-6 i IL-21, fan que s’expressi
BSL-6. Aquest potencia la producció de IL21. És essencial la coestimulació via interacció
ICOS-ICOSL per mantenir el fenotip. Els limfòcits T helper fol·liculars acompanyen limfòcits
T secundaris als fol·licles quan s’està produint la resposta.
Limfòcits Treg.
Els limfòcits T reguladors serveixen per regular l’activació i acció de les cèl·lules T efectores.
Es diferencien a partir precursors al timus, quan reben el nom de reguladors tímics. Els
limfòcits T reguladors sempre hi són arreu i mantenen l’estat de no resposta a la perifèria.
Quan es dona una resposta immunitària, es generen limfòcits T reguladors en presencia
d’antígens microbians, TGDFβ, IL-2, àcid retinoic, àcids grassos de cadena curta. Les
cèl·lules reguladores es caracteritzen per produir, algunes d’elles, FOXP3. Són cèl·lules
importants per finalitzar la resposta immunitària.
Les cèl·lules T reguladores expressen IL-10 i TGFβ per aturar la inflamació. També expressen
PD1 i CTLA4. La secreció d’aquests compostos permet als limfòcits T reguladors dur a terme
la seva funció d’aturar la resposta immunitària. Els Treg tenen una funció molt important en
la resposta a patògens. S’ha vist que poden inhibir la funció de limfòcits T efectors, inhibint la
resposta a tumors. Això incentiva el desenvolupament d’aquest.
Mecanisme efector dels limfòcits T helper. Els patrons de citocines secretat és el mecanisme efector dels limfòcits TH i determinen la
principal funció efectora de cada subtipus cel·lular. Th1, per exemple, també produeix IL-3 o
GM-CSF, que promouen la diferenciació dels precursors dels limfòcits T. El patró de secreció
de citocines i d’expressió de receptors de quimiocines proporciona funcions diferents a cada
tipus efectors de T helper durant la resposta immunològica.
Paral·lelament a la producció de citocines associades a tipus de limfòcit T helper, cadascun
expressa un diferent patró de receptors de quimiocines. Les diferències en patrons
d’expressió de receptors s’equival amb una diferència en els patrons de migració. Això
significa que el lloc on va cada tipus de limfòcit T efector és diferent perquè les quimiocines
es produeixen a llocs diferents. El conjunt del patró de citocines secretades més el patró de
receptors de quimiocines determina la capacitat funcional final de la cèl·lula T helper.
Diferenciació de limfòcitsTCD8 efectors. Malgrat no es coneixen diferents tipus de limfòcits TCD8 efectors, es parla de limfòcits TCD8
naive. Aquest limfòcit TCD8 naive no ha reconegut antigen. En el moment que ho fa, si es
donen les condicions adequades de suficient senyalització, intervenen el primer i segon
senyals d’activació amb col·laboració del correceptor i s’activa el limfòcit.
Aquesta activació li permet proliferar i es diferencia a limfòcit T efector. Hi ha diferents estadis
en aquest procés. Canvia expressió receptor IL-2 al aparèixer la cadena α, comença a
expressar poca IL-2, es desenvolupa la capacitat de proliferació i capacitat citotòxica dels
grànuls amb material citotòxic.
Quan es produeix una baixa inflamació, l’activació dels limfòcits T CD8 requereix la presència
de limfòcits T CD4 que responen al mateix antigen. Les cèl·lules T CD4 funcionen “activant”
les cèl·lules dendrítiques que presentaran l'antigen als limfòcits T CD8 i, interaccionant
directament amb limfòcits T CD8 mitjançant l'expressió de CD40 o la producció d’IL2. Si el
que es produeix és una alta inflamació, els limfòcits T CD8 es poden activar en absència de
cèl·lules T CD4 però no es genera una bona memòria T.

Immunologia 2020
CEBIO 104
Dendritic cell Licensing. La necessitat de TCD4 en l’activació ve donada per l’acció o efecte d’aquests sobre la cèl·lula
presentadora d’antigen. És el que s’anomena activació de les cel dendrítiques. Aquest
concepte defineix el procés mitjançant el qual les cèl·lules presentadores d’antigen, que solen
expressar un baix nivell de molècules coestimuladores, tenen un contacte amb els limfòcits
TCD4 mitjançant el reconeixement del complex pèptid/MHC i CD40/CD40L i això suposa un
augment en la presentació de molècules coestimuladores. Amb el cell licensing és possible
l’activació de CD8 i l’expressió de IL12 per activar i diferenciar CD8.
Si el limfòcit TCD4 i el TCD8 es troben alhora amb la cèl·lula dendrítica, les citocines
secretades per el TCD4 estimulen la diferenciació del T CD8 efector. Si no es troben alhora,
la interacció de la TCD4 amb la cèl·lula dendrítica li proporciona els senyals per a l’activació
amb l’augment d’expressió de molècules coestimuladores i de producció de citocines,
sobretot IL12, es diu dendrític cell licensing.
El dendrític cell licensing és important per l’activació dels limfòcits TCD8 naive. En cas que
estiguem parlant de limfòcits TCD8 de memòria, requereixen menys ajuda i la cèl·lula
dendrítica madura pot activar-les directament. Als limfòcits TCD8, el llindar d’activació és
més baix i necessita menys molècules estimuladores a la superfície de la cèl·lula dendrítica.
Les cèl·lules TCD8 de memòria produeixen IL-2 en rebre el 1er i 2on senyal, mentre que les
cèl·lules TCD8 naive necessiten les cèl·lules TH1 com a font de la mateixa citocina.
S’ha vist que senyalització en presència PAMPs fa augmentar presència molècules
coestimuladores CD80 i CD86 a cèl·lules dendrítiques. Això facilita activació limfòcits TCD8.
Mecanisme efector de la citotoxicitat dels limfòcits TCD8. Limfòcits TCD8 indueixen la mort cel·lular per apoptosi. Significa que la cèl·lula que presenta
l’antigen en complex amb el pèptid del MHC de classe I mor. En aquesta mort, el limfòcit
TCD8 indueix una pèrdua de potencial de membrana mitocondrial, una alliberació de citocrom
C, una condensació nuclear, una externalització de fosfatidilserina i una activació de les
caspases que provoquen la fragmentació del DNA. A nivell morfològic, hi ha un blebbing
nuclear i una alteració general de la morfologia cel·lular que genera cossos apoptòtics. Un
blebbing és una fragmentació i reorganització de la cromatina del nucli.
Això és així perquè els limfòcits TCD8 alliberen el contingut dels seus grànuls a zona contacte
entre les dues cèl·lules. Aquest grànuls lítics contenen perforina, que forma porus a la
membrana que permeten entrada d’aigua i sals provocant un trencant de l’equilibri osmòtic;
granzima, unes serina proteases que activen caspases del citoplasma per activar l’apoptosi;
i granulisina, que allibera Ca2+ intracel·lular i citocrom c.
La granzima activa la caspasa 8 que, directament o a través de l’alteració del potencial de
membrana a mitocondris que allibera el citocrom C i activa la caspasa 9, activa caspasa 3.
La caspasa 3 trenca el DNA doble cadena i provoca la mort cel·lular.
El procés d’inducció de la mort cel·lular és molt ràpid. Entre l’inici de la interacció i la inducció
de la mort cel·lular passen menys de 7 minuts. En aquests, el limfòcit TCD8 s’acosta a la
cèl·lula amb l’antigen, interacciona pel complex del TCR amb el complex pèptid/MHC,
desgranula sobre cèl·lula diana, es formen porus, la cèl·lula canvia de morfologia i s’indueix
la mort.
Hi ha altre mecanisme efector TCD8. Resultat interacció fas i lligand fas. Es produeix en Th1
activats. Limfòcits TCD8 expressen lligand fas. Moltes cèl·lules organismes expressen fas.
Interacció Fas-lligand fas té conseqüències. Interacció activa domini FADD associada a fas.

Immunologia 2020
CEBIO 105
FADD té dominis de mort que recluten caspasa 8 inactiva i indueixen activació. Caspasa 8
activa caspasa 3 que produeix trencament DNA i mort cel·lular.
L’alliberació del contingut dels grànuls es dona a la superfície de contacte entre el limfòcit
TCD8 i la cèl·lula diana. La cèl·lula TCD8 mata específicament les cèl·lules que expressen
l’antigen pel qual aquest limfòcit TCD8 és específic. És, doncs, un mecanisme molt diferent
del que tenen les cèl·lules de la immunitat innata, que alliberen el contingut dels grànuls a
l’exterior i afecten a totes les cèl·lules del voltant. El mètode que fan servir els limfòcits TCD8
és específic, per això requereix que l’alliberament sigui a la zona de contacte.
La citotoxicitat dels TCD8 depèn de la sinapsi immunològica. Fa que l’acció dels limfòcits es
produeixi només en la cèl·lula amb la que s’està interaccionant. Per tant, és específic.
Limfòcits T citotòxics. Les cèl·lules T citotòxiques maten selectivament les cèl·lules diana que expressen l’antigen
pel qual són específiques. L’alliberament de substàncies citotòxiques només es dona a la
zona de contacte entre el limfòcit i la cèl·lula diana.
Hi ha limfòcits T naive, que es diferencien. Una part ho fan a limfòcits T de memòria.
Responen de manera més ràpida i efectiva al mateix antigen quan aquest es presenta per
segona vegada a l’organisme. Els limfòcits T de memòria persisteixen en absència antigen.
Es mantenen durant temps llargs als teixits. Ho fan gràcies a IL-7 i IL-15. Hi ha diferents
limfòcits t memòria.
Limfòcits T de memòria.
Els limfòcits T de memòria s’activen més ràpid i de manera més eficient perquè tenen menys
requeriments d’activació. Necessiten menys senyals d’adhesió, no necessiten una alta
intensitat al 1r i al 2n senyal ni tampoc necessiten una alta concentració de molècules
coestimuladores. Aquests criteris s’apliquen tant als limfòcits TCD4 com als TCD8.
Hi ha diferents tipus de cèl·lules T de memòria. Aquestes difereixen per la seva localització.
Els limfòcits T de memòria centrals expressen el receptor CCR7, que provoca l’entrada als
òrgans limfàtics secundaris com els limfonodes on queden retinguts a limfonodes. També
expressen CD62L, que hi facilita la entrada. Els limfòcits T de memòria efectors no expressen
CCR7 i tenen un baix nivell d’expressió de CD62L. Un altre tipus de limfòcits T de memòria
que existeixen són els residents als teixits. Aquests limfòcits T de memòria no expressen
CCR7 ni CD62L, però expressen CD103 i CD11a.
Això fa que diferents limfòcits tinguin diferent capacitat de homing, és a dir, que es troben en
localitzacions concretes. Els limfòcits T de memòria centrals es troben en òrgans limfàtics
secundaris. Per contra, els limfòcits T de memòria efectors es troben en teixits no limfoides o
a la melsa. Finalment, els limfòcits T de memòria residents als teixits es troben en teixits amb
mucosa com poden ser els pulmons, el tracte urogenital o a la pell. Els limfòcits T efectors de
memòria actuen als teixits mentre que els de memòria centrals són un reservori per generar
altres limfòcits T de memòria si el sistema immunitari no dona la talla.
Les diferències de localització dels limfòcits T de memòria també es tradueix en una
diferenciació funcional.
Els limfòcits T amb receptor gammadelta també en tenen d’efectors. També en cèl·lules NKT.
Tenim cèl·lules T gamma delta amb capacitat citotòxica i col·laboradors. Patrons producció
citocines determinen participació en diferents processos de la resposta immunitària.

Immunologia 2020
CEBIO 106
Cèl·lules NKT. Hi ha 5 llinatges descrits de cèl·lules iNKT, que es generen a partir de les cèl·lules precursores
d'iNKT. Cada llinatge es troba diferenciat terminalment per l’expressió d'una selecció única
de citocines i factors de transcripció quan s’activen. NKT1: expressen alts nivells de Tbet,
secreten IFNg i algunes IL-4. NKT2: expressió continua de PLZF i secreten principalment IL-
4 i IL-13. NKT17: expressen RORgt i secreten IL-17. NKTFH: expressen Bcl6 i es
desenvolupen després de l'activació. No està clar si es desenvolupen a partir de precursors
NKT1. NKT10: es desconeixen els factors de transcripció que regulen NKT10 però es
defineixen com a secretores d’IL10, expressen NRP-1 i PD1.

Immunologia 2020
CEBIO 107
Tema 20 – Activació de la Resposta Humoral. La resposta immunitària humoral és la Resposta immunitària vehiculada per anticossos.
Subpoblacions de limfòcits B. Els limfòcits B es separen en diferents subpoblacions: limfòcits B1 i limfòcits B2. Els
limfòcits B1 són els més abundants a pleura i cavitat peritoneal i generen el fetge fetal durant
el desenvolupament embrionari. Es caracteritzen per tenir un repertori de BCRs reduït i solen
tenir i produir IgM.
Els limfòcits B2, en canvi, tenen un repertori ampli de BCRs i en distingim 2 grups: limfòcits
B2 fol·liculars i limfòcits B2 marginals. Els limfòcits B2 fol·liculars són els típics de la
resposta adaptativa, són la majoria de limfòcits B madurs, responen a antígens proteics i
depenen de la col·laboració amb els limfòcits T. Els limfòcits B2 de la zona marginal es
troben a la zona marginal de la melsa, responen a antígens provinents de la sang, són
independents de la col·laboració amb els limfòcits T i responen ràpid secretant IgM.
Activació dels limfòcits B. Per a una activació complerta dels limfòcits B també necessitem més d’un senyal, en aquest
cas en són 2. El primer senyal ve donat per la interacció entre l’antigen i el BCR. El segon
senyal d’activació són una sèrie d’interaccions que ajuden a l’activació completa dels limfòcits
B. Els segons senyals són diferents segons el tipus d’antigen i el tipus de limfòcit B.
1er senyal d’activació.
El primer senyal d’activació és el que ve donat per la interacció entre el BCR i l’antigen.
Aquesta interacció produeix tota una sèrie de canvis i moviment en aquest BCR. A les
cèl·lules B en repòs, el BCR es troba expressat a la membrana en forma de monòmers
associat a les cadenes Igα/Igβ, les quals tenen motius ITAM a les cues citoplasmàtiques.
Quan el BCR interacciona amb l’antigen a través d’algun dels seus braços, es produeix un
canvi conformacional al domini constant 4 de la cadena pesada. Això fa exposar la superfície
de contacte, permetent que es formin agregats de BCRs. En unir-se a l’antigen també es
produeix l’obertura dels dominis citoplasmàtics de Igα/Igβ. Aquesta conformació permet que
comenci la senyalització.
La unió d’antigen permet el crosslinking o entrecreuament del BCR. L’entrecreuament és la
formació d'un complex antigen-anticòs en què múltiples elements d'un mateix antigen, amb
més d'un determinant antigènic, s'uneixen als BCR corresponents, de manera que alguns
antígens queden units a més d'una molècula d'immunoglobulina.
Hi ha un tercer canvi quan hi ha una interacció amb l’antigen. Els oligòmers que es formen es
desplacen cap a les balses lipídiques o lipid rafts, on les cadenes Igα i Igβ poden interaccionar
amb tirosina quinasa (Lyn) i iniciar al senyalització a través dels seus motius ITAM.
Interacció BCR-antigen.
La fosforilació de les tirosines dels motius ITAM de la cua citoplasmàtica de les cadenes
Igα i Igβ per la quinasa Lyn permet que es recluti la quinasa Syk. Syk fosforila les quinases
BTK i BLNK. Btk contribueix a l’activació de BLNK, que crea els llocs d’unió per a la
fosfolipasa Cγ2 i per al GEF (el factor intercanviador de nucleòtids de guanina) anomenat
VAV. VAV indueix una remodelació del citoesquelet, pel que es canvia la morfologia de la
cèl·lula B. La fosfolipasa Cγ2 trenca el fosfoinositol-2-fosfat de la membrana citoplasmàtica
en diacilglicerol i en inositol-3-fosfat. L’activació de GEF també inicia la via de les MAP
quinases per part de les proteïnes G petites.

Immunologia 2020
CEBIO 108
L’inositol-3-fosfat fa augmentar la concentració de cations de calci al citoplasma. El calci
permet activar, a través de la calmodulina, la calcineurina. La calcineurina activa el factor de
transcripció NFAT, que es transloca al nucli. El diacilglicerol, a través de la fosfoquinasa C,
activa NFκB, que també es transloca al nucli. La fosfoquinasa C també contribueix a
l’activació del factor de transcripció Jul i la formació de Jun, que van al nucli.
Coreceptor activador del BCR.
De la mateixa manera que els limfòcits T tenen coreceptors que ajuden al TCR, els limfòcits
B tenen coreceptors que contribueixen a l’acció del BCR. En el cas dels limfòcits B hi ha
un coreceptor activador format per 3 proteïnes: CD19, CD21 i CD81. La contribució d’aquest
coreceptor és imprescindible perquè es produeixi l’activació completa del limfòcit B.
CD19 és una proteïna transmembrana el domini extracel·lular de la qual està format per
dominis immunoglobulina i té una cua citoplasmàtica amb un motiu ITAM que fa servir per
amplificar el senyal d’activació del BCR. CD21 és un receptor del fragment C3d de la proteïna
del sistema del complement. C3d és un fragment de C3. CD21 també rep el nom de CR2.
CD81 és una proteïna transmembrana associada a CD21 amb una funció desconeguda en el
marc de coreceptor activador del limfòcits B.
Interacció BCR-Coreceptor-Antigen.
Similar als limfòcits T, el coreceptor amplifica el senyal d’activació. CD21 interacciona amb
el fragment C3d, el qual està opsonitzant l’antigen que està reconeixent el BCR. El
coreceptor s’acosta aleshores al BCR, permetent una fosforilació dels motius ITAM de la
cua citoplasmàtica de CD19 per part de la quinasa Lyn. La fosforilació del motiu ITAM de
CD19 permet l’amplificació del senyal.
La fosforilació del motiu ITAM de la cua citoplasmàtica de CD19 crea el lloc d’unió per a la
quinasa Fosfoinositol-3-quinasa que amplifica la senyalització del BCR al inhibir
l’inhibidor del factor de transcripció NFAT, afavorint-ne l’activació.
Coreceptor inhibidor del BCR.
En condicions de repòs, el BCR està associat a CD22, un coreceptor inhibidor. CD22 és una
lectina que, a la cua citoplasmàtica, té un motiu ITIM. Aquest motiu, quan es troba fosforilat,
genera un lloc d’unió per una fosfatasa que manté desfosforilades les cues citoplasmàtiques
de Igα i Igβ. Aquest manteniment impedeix la transmissió de senyals d’activació a
l’interior de la cèl·lula. CD22, que uneix àcid siàlic, es troba associat o a prop del BCR,
mentre que el coreceptor activador només s’acosta al BCR quan aquest interacciona amb
l’antigen.
La senyalització del TCR i el BCR són semblants.
La transducció de senyal pel TCR i el BCR es basa en el mateix mecanisme general,
usant quinases i adaptadors específics que fan funcions similars i acabant en cascades de
senyalització comparables. Comparant element a element, es poden fer diverses analogies.
El BNLK del BCR és equiparable al SLP-76 del TCR tant com que la quinasa Syk es
correlaciona amb la quinasa ZAP70. A més, la fosfolipasa Cγ1 i la fosfolipasa Cγ2 realitzen la
mateixa funció, com les quinases equivalents Lck i Lyn.
2n senyal d’activació.
El 2n senyal d’activació és diferent segons quin sigui l’antigen i, en conseqüència, segons
quina sigui la subpoblació de limfòcits B que s’està activant. Distingim entre dos tipus de
segon senyal: segon senyal independent de limfòcits T i segon senyal dependent de la
interacció amb limfòcits T.

Immunologia 2020
CEBIO 109
2n senyal independent de limfòcits T.
Quan el segon senyal és independent de limfòcits T és perquè els limfòcits B no
requereixen de la col·laboració de limfòcits T helper per la seva activació i producció
d’anticossos. Els antígens que solen provocar aquest tipus de segon senyal són,
majoritàriament, components de la paret cel·lular, polisacàrids microbians o proteïnes
polimèriques. De segons senyals independents de limfòcits T també hi ha dos tipus: els
Antígens T independents de Tipus 1 (Antígens TI-1) i els Antígens T independents de Tipus
2 (Antígens TI-2).
Resposta de les cèl·lules B a Antígens T independents de Tipus 1.
El segon senyal en resposta a Antígens TI-1 són aquells on el propi antigen ja té activitat
coestimuladora perquè conté components reconeguts per PRRs. Els limfòcits B que reben
aquest segon senyal generen un canvi d’isotip molt poc eficient, pel que quasi tots els BCRs
produïts són IgM. Pràcticament no hi ha hipermutació somàtica.
Quan la concentració d’Antígens TI-1 és baixa, hi ha un limfòcit B que reconeix l’antigen via
BCR i, a més, rep el segon senyal provinent del PRR. Això provoca l’expansió clonal
d’aquest limfòcit B. S’ha vist que, quan els Antígens TI-1 amb activitat coestimuladora
pròpia es troben en una concentració molt elevada, poden donar lloc a l’activació policlonal
i no específica de la cèl·lula B. Això és perquè hi ha una gran quantitat d’aquests
components de l’antigen reconeguts per PRR que poden activar de manera poc eficient els
limfòcits B sense que hi hagi una interacció amb el BCR.
Resposta de les cèl·lules B a Antígens T independents de Tipus 2.
Els segons senyal en el cas de Antígens TI-2 són donat perquè aquests antígens contenen
epítops repetitius que poden estimular directament els limfòcits. És principalment el cas dels
limfòcits B1. Són carbohidrats, polisacàrids, elements repetitius... Els Antígens TI-2 només
poden donar lloc a l’activació de limfòcits B madurs, ja que cal una gran quantitat de
senyalització. En aquest cas, el primer senyal d’activació el dona la interacció de l’antigen
amb molts BCRs alhora, generant un crosslinking massiu. Com que molts BCRs alhora poden
reconèixer l’antigen, es dona un senyal suficient per produir una activació dels limfòcits B.
Sovint, aquest senyal repetitiu permet l’activació del limfòcits B i la producció
d’immunoglobulines M. No hi ha canvi d'isotip. Si en el context de la interacció hi ha citocines,
aquestes poden proporcionar un segon senyal que activi els limfòcits B per tal que es
diferenciïn a cèl·lules productores d’immunoglobulines i que, fins i tot, hi hagi un ineficient
canvi d’isotip.
Resposta dels limfòcits B a Antígens T-dependents i a Antígens T-independents.
Els tres grans tipus de limfòcits B són els limfòcits B2 fol·liculars, els Limfòcits B2 de la zona
marginal i els Limfòcits B1. Els limfòcits B1 i els limfòcits B2 de la zona marginal solen
respondre a antígens T independents. Els limfòcits B2 fol·liculars, en canvi, necessiten la
col·laboració dels limfòcits T per obtenir la màxima activació.
També canvia el que s’aconsegueix. Els limfòcits B2 de la zona marginal, activats
independentment de limfòcits T, produeixen IgM i cèl·lules productores d’immunoglobulines
però de vida curta, com els limfòcits B1. Els limfòcits B2, al necessitar la interacció amb els
limfòcits T, fan molt bé el canvi d’isotip, la hipermutació somàtica i cèl·lules plasmàtiques de
vida llarga.
Resposta dels limfòcits B a antígens T-dependents.
El primer senyal d’activació ve donat per la interacció entre l’antigen i el BCR. El resultat
d’aquesta interacció és un inducció de l’expressió de les molècules coestimuladores CD40,

Immunologia 2020
CEBIO 110
B7 i ICOSL, que serà molt important ja que dins la interacció T-B té un paper similar a la
interacció entre CD40 i el seu lligand. La interacció entre B7 al limfòcit B i CD28 al limfòcit T
és essencial per tal que es doni el primer senyal d’activació.
El primer senyal d’activació potencia l’expressió d’aquestes molècules coestimuladores. El
segon senyal ve donat per la interacció entre CD40 i el seu lligand o entre ICOSL i ICOS. A
més, la cèl·lula T col·laboradora produeix citocines que contribueixen en el segon
senyal i són importants en el canvi d’isotip. Les cèl·lules col·laboradores, amb la producció
de IL-4 i altres citocines, afavoriran la activació i la proliferació dels limfòcits B. El segon senyal
d’activació, doncs, és un conjunt de senyals donats per les molècules coestimuladores i les
citocines.
Col·laboració entre cèl·lules B i cèl·lules T.
La col·laboració entre els limfòcits T i B es pot donar perquè, en aquests casos concrets, els
limfòcits B actuen com a cèl·lules presentadores d’antigen reconegudes pel limfòcit T per
mitjà del TCR. L’antigen és el que el limfòcit B ha reconegut pel BCR. La interacció antigen-
BCR internalitza aquest complex per la via endocítica, generant pèptids que es poden
carregar a molècules del MHC-II i ser expressats a la membrana.
El limfòcit T i el limfòcit B reconeixen diferents epítops del mateix antigen. El resultat de
la interacció és l’augment d’expressió de CD40 a la membrna del limfòcit B per tal que
interaccioni amb el lligand de CD40 dels limfòcits T activats. Juntament amb les citocines,
CD40 reclutarà TRAF i iniciarà la cascada d’activació de NFκB i AP-1. Això proporciona més
senyal d’activació.
La interacció entre CD40 i el seu lligand proporciona el segon senyal d’activació. La interacció
entre B7 i CD28 senyalitza cap l’interior del limfòcit T i li proporciona el seu segon senyal
d’activació. El limfòcit B, fruit de la interacció, expressa receptors de quimiocines per ser més
sensible a l’expressió de les mateixes per part del limfòcit T.
Tema 21 – Mecanismes efectors de la resposta humoral. Hi ha un primer senyal, donat per la interacció antigen-BCR, i un segon senyal provinent de
la col·laboració entre els limfòcits T i B. Quan un limfòcit B reconeix un antigen pel seu BCR,
l’internalitza i el processa per presentar-ne pèptids derivats mitjançant les molècules del MHC
de classe II. Per tant, els limfòcits B poden actuar com a cèl·lules presentadores
d’antigen pels limfòcits TCD4. En aquesta interacció cèl·lula T – cèl·lula B, hi intervenen
CD40 al limfòcit B i CD40L al limfòcit T, proporcionant un senyal de coestimulació. A més,
aquest senyal augmenta l’expressió de receptors de citocines.
Arribada de l’antigen als fol·licles. L’antigen arriba a fol·licles dels òrgans limfàtics secundaris com els limfonodes. Els antígens
petits hi arriben mitjançant els vasos limfàtics aferents, que els recullen als teixits perifèrics.
Els antígens d’una mida mitjana són captats pels macròfags que es troben a la paret basal
del sinus subcapsular i des dels quals els poden transferir a l’interior dels fol·licles. Altres
antígens també poden ser transportats per les cèl·lules dendrítiques.
Respostes humorals T-dependents. Els limfòcits B que es troben al còrtex dels fol·licles limfàtics al expressar el receptor de
CXCL13, CXCR5. Poden interaccionar amb l’antigen. El resultat d’aquesta interacció és un
acostament a la frontera entre el paracòrtex i el còrtex. Això és perquè l’activació rebaixa
la presentació de CXCR5 i augmenta la de CCR7, decantant els pols d’atracció cap al
paracòrtex. El paracòrtex és la zona dels limfòcits T.

Immunologia 2020
CEBIO 111
Per altra banda, als limfonodes hi arriben limfòcits TCD4, que reconeixen antígens a la zona
del paracòrtex gràcies a les cèl·lules dendrítiques. Això els activa i, en conseqüència, són
retinguts al limfonode. L’activació pot donar lloc a la expressió de CXCR5 al limfòcit T. Si
es dona aquest fet, el limfòcit T s’acostarà al còrtex. A la zona propera entre el còrtex i el
paracòrtex es pot donar la interacció entre els limfòcits T i B. Aquesta interacció és la
primera part de la col·laboració B-T.
La interacció B-T comença amb el limfòcit B, que ha reconegut l’antigen a l’interior del
fol·licle i que l’ha processat, carregat i presentat al limfòcit T. Aquest limfòcit T és específic
per aquest mateix antigen que li presenta el limfòcit B. Per tant, tenim dos limfòcits específics
per diferents epítops del mateix antigen. Quan es dona aquesta interacció, el limfòcit B rep
el segon senyal d’activació arran de la interacció entre CD40 i CD40L. Per tant, el limfòcit
B pot convertir-se en una cèl·lula plasmàtica de vida curta, que produeixen
immunoglobulines als cordons medul·lars del limfonode i expulsen els anticossos per la
circulació, o poden migrar a l’interior dels fol·licles i començar la formació d’un centre
germinal.
Quan un limfòcit B naive reconeix antigen, li passen una sèrie d’alteracions i modificacions.
Entre aquestes trobem que comença a proliferar, augmenta l’expressió de les molècules
coestimuladores B7-1 i B7-2 (lligands de CD28 que li permeten proporcionar el senyal
coestimulador a les cèl·lules T), augmenta l’expressió de receptors de citocines i es produeix
un augment en l’expressió de CCR7 en detriment de la de CXCR5.
En ratolins, la interacció B-T als limfonodes dura uns 4 dies des de la entrada de l’antigen al
cos. Durant els dos primers dies, les cèl·lules T penetren cap a la frontera del còrtex amb el
paracòrtex mentre que les cèl·lules B fan el camí contrari. Al tercer dia ja ha començat la
proliferació de les cèl·lules B i aquestes es poden veure a la part més externa del limfonode.
El quart dia les cèl·lules B ja es poden veure formant centres germinals.
Cèl·lules T helper fol·liculars. La interacció entre els limfòcits T i els limfòcits B requereix de la molècula coestimuladora
ICOS. Aquesta molècula expressada al limfòcit T helper s’uneix a ICOSL, que es troba al
limfòcit B. Aquesta interacció indueix un augment de CD40 al limfòcit B i de CD40L al
limfòcit T. A més, la interacció ICOS-ICOSL regula la transcripció de la citocina
interleuquina 21. En conseqüència, la interacció fa que el limfòcit T helper es diferenciï al
limfòcit T efector T helper fol·licular.
La interacció ICOS-ICOSL augmenta la durada de la interacció entre els dos limfòcits, tenint
efectes en la maduració de l’afinitat, supervivència i diferenciació de les cèl·lules B. Dit d’altra
manera, té efecte sobre totes les reaccions del centre germinal.
Això és així perquè quan un limfòcit T reconeix antigen al paracòrtex, aquest s’activa i
comença a poder produir diferents citocines segons l’antigen, la cèl·lula dendrítica que li
hagi presentat, la quantitat d’antigen... Aquest fenotip, però, no és l’efector. El fenotip efector
queda fixat quan es produeix la interacció B-T i és el fenotip de limfòcit T helper fol·licular. Els
limfòcits T helper fol·liculars poden ser productors d’interferó γ, d’interleuquina 17,
interleuquina 4... I, malgrat de quin tipus siguin, tots produiran IL21. Aquesta citocina és
essencial en molt processos.
ICOS controla l’expressió de la IL21, que controla l’expressió de Bcl6. Bcl6, un factor de
transcripció característic dels limfòcits T helper fol·liculars, controla l’expressió del receptor
de quimiocines CXCR5. Dit d’altre manera, Bcl6 fa que els limfòcits T helper es mantinguin

Immunologia 2020
CEBIO 112
als fol·licles gràcies a l’expressió de CXCR5. En el fons, doncs, ICOS fixa el fenotip de
limfòcit T helper fol·licular i, per tant, aquests migren cap al fol·licle.
A la zona de contacte entre el paracòrtex (zona T) i el còrtex (zona B) el limfòcit B presenta
antigen al limfòcit T helper, del qual no s’ha fixat del tot el seu fenotip. Si aquest limfòcit T
pot reconèixer el mateix antigen que li presenta el limfòcit B, la parella de cèl·lules es
desplacen cap al centre dels fol·licles per desenvolupar la reacció dels centres germinals.
En aquest centre es pot donar lloc a cèl·lules B de memòria o cèl·lules plasmàtiques de
vida llarga.
Les cèl·lules productores d’immunoglobulines de vida llarga són diferents de les de vida
curta, produïdes per interaccions fora de la zona del còrtex. La diferència més important és
que les cèl·lules plasmàtiques de vida curta seran productores d’immunoglobulines del
mateix tipus que la cèl·lula B tenia com a BCR, mentre que les cèl·lules plasmàtiques de
vida llarga hauran passat pels centres germinals, on es pot donar un canvi d’isotip o una
hipermutació somàtica. En conseqüència, les immunoglobulines produïdes d’aquesta
manera seran més eficients contra l’antigen.
El que determina que un limfòcit B activat es diferenciï a cèl·lula plasmàtica o segueixi com a
limfòcit b fol·licular és el factor Bcl6. El limfòcit B activat expressa el receptor de la IL21.
Quan aquest receptor interacciona amb la citocina, el limfòcit B expressa Bcl6, que li manté
l’expressió de CXCR5. A més, es bloqueja l’expressió del factor de transcripció BLIMP-1, el
qual provocaria una transformació en una cèl·lula plasmàtica de vida curta. En conseqüència,
sortiria cap a la medul·la del limfonode per tal que els anticossos anessin cap a la circulació
sanguínia.
Centres germinatius. Generar un centre germinatiu o germinal és un procés complex. Per un costat, un cop es dona
l’activació del limfòcit T al paracòrtex arran de la presentació d’antigen per part de la
cèl·lula dendrítica, comença a migrar cap a la zona de contacte entre el còrtex i el
paracòrtex.
Per altra banda, el limfòcit B que ha trobat antigen també migra cap a la frontera entre la
zona B i la zona T. Allà es produeix la interacció i les parelles migren cap a l’interior del
fol·licle, on comencen a proliferar. Fins al dia 7 (depenent de la quantitat d’antigen i del lloc
d’on s’estigui parlant) no es veu clara l’estructura del centre germinatiu. Aquesta estructura
consisteix en una zona fosca composta principalment per cèl·lules B i una zona clara
formada per limfòcits B, limfòcits T helper fol·liculars i cèl·lules fol·liculars dendrítiques.
La zona fosca dels centres germinatius rep aquest nom perquè hi molta densitat de
limfòcits B que es troben proliferant, moment que s’anomenen centroblasts. Aquest gran
ritme de proliferació resultant d’haver rebut el coestímul dels limfòcits T permet que es doni
una hipermutació somàtica, petits canvis en les zones hipervariables del BCR que li poden
canviar l’afinitat per l’antigen.
A la zona clara s’hi troben els limfòcits B que han deixat de proliferar, anomenats centròcits,
limfòcits T helper fol·liculars i cèl·lules fol·liculars dendrítiques. A la zona clara es dona una
maduració de l’afinitat, procés on intervenen les cèl·lules fol·liculars dendrítiques, i el canvi
d’isotip. El canvi d’isotip depèn de les citocines que es produeixen al centre germinatius, les
quals alliberen els limfòcits T helper fol·liculars.
El moviment entre la zona fosca i la zona clara depèn del gradient de quimiocines. A la
zona fosca hi ha una concentració més alta de CXCL12, detectada pel receptor CXCR4, que

Immunologia 2020
CEBIO 113
no pas a la zona clara. Passa el contrari amb CXCL13. Això és perquè les cèl·lules fol·liculars
dendrítiques són les productores de CXCL13. Per tant, en funció del nivell d’expressió dels
receptors d’aquestes quimiocines, els limfòcits B es mourien de la zona fosca a la clara o a la
inversa.
Cèl·lules Fol·liculars Dendrítiques. Les cèl·lules fol·liculars dendrítiques reben aquest nom perquè tenen unes prolongacions
cel·lulars. No són d’origen hematopoètic. Sembla ser que deriven de cèl·lules l’estroma.
No són presentadores d’antigen ja que no tenen una gran capacitat d’internalització i
degradació del que capten a través dels receptors de la membrana. Tenen alts nivells de
receptors de la fracció constant de les immunoglobulines, com CD32 (receptor de la fracció
constant de les IgG i IgE), i alts nivells de receptors de proteïnes del sistema del
complement com CR1, CR2 i CR3.
Els receptors que aquestes cèl·lules presenten els permeten unir antígens opsonitzats i que
es troben formant immunocomplexes. Això inclou antígens opsonitzats per anticossos o per
proteïnes del sistema del complement. La interacció entre els immunocomplexes i els
receptors a la membrana són molt estables i es mantenen durant temps llargs perquè
aquestes cèl·lules fol·liculars dendrítiques no tenen un bon sistema d’internalització i
degradació. D’aquesta manera, els complexes ancorats a la membrana s’hi mantenen molt
més temps del que seria normal en una puta cèl·lula dendrítica normal i corrent. Aquestes
de seguida s’ho empassa per endocitosi o internalització.
Així i tot, aquests immunocomplexes queden retinguts a la membrana i formen acumulacions
que s’anomenen icosomes. Per tant, aquestes cèl·lules fol·liculars dendrítiques proporcionen
antigen al limfòcit B en un entorn on aquest és molt poc abundant ja que ens troben dins
dels centres germinals. Les cèl·lules fol·liculars dendrítiques mantenen l’antigen prou
temps per permetre l’activació dels collons. Puta Carme.
Les cèl·lules fol·liculars dendrítiques tenen la capacitat de produir CXCL13, el que atrau
els limfòcits B que expressen CXCR5 i també a les cèl·lules T helper fol·liculars que
expressen CXCR5. Les cèl·lules dendrítiques fol·liculars expressen molècules d’adhesió que
estabilitzen la interacció amb els limfòcits B que reconeixen els antígens que tenen
ancorats a la membrana a través dels immunocomplexes. També poden produir factors de
creixement i supervivència pels limfòcits B, com la interleuquina 6 i la citocina BAFF. BAFF
permet mantenir la supervivència dels limfòcits B en aquest entorn concret.
Limfòcits B al centre germinal. Les cèl·lules fol·liculars dendrítiques de la zona clara tenen els immunocomplexes ancorats
a la membrana. Els limfòcits B que han estat a la zona fosca han passat pel procés
d’hipermutació somàtica i poden haver canviat l’afinitat. Tan sols els que millor interaccionin
amb l’antigen seran els que podran reaccionar-hi i rebre tant el senyal de supervivència com
el segon senyal. Aquest segon ve per la interacció amb els limfòcits T helper fol·liculars, la
qual es dona amb CD40 i ICOS. Els limfòcits B de menys o gens afinitat per l’antigen per
culpa de la hipermutació somàtica, no tenen possibilitats d’interaccionar amb l’antigen i moren
per apoptosi al no rebre el senyal de supervivència.
Això vol dir que cada cop que es produeix una resposta per limfòcits B, els que queden al final
són els que tenen major afinitat per l’antigen. Per tant, els que responen inicialment a un
antigen són molt diferents dels que hi interaccionen quan aquest torna a entrar. A mesura que
es va donant la resposta a un determinat antigen, va augmentant la afinitat del BCR per
aquest. Això fa que la resposta sigui més eficient, més ràpida i millor.

Immunologia 2020
CEBIO 114
D’altra banda, a la zona clara del centre germinal hi ha la interacció entre el limfòcit B i el
limfòcit T helper fol·licular, que determina el canvi d’isotip. Amb la interacció amb el
limfòcit B, el T helper fol·licular s’activa i secreta citocines en funció de l’estímul inicial.
Aquest estímul determina si serà T helper fol·licular productora d’interferó, IL4, alguna altre
citocina... En funció de la citocina que es produeixi, el limfòcit B es diferenciarà a productor
d’un determinat isotip d’immunoglobulina. Si el T helper fol·licular produeix IL4 o IL13, el
limfòcit B es diferenciarà a un que pugui produir BCR de IgG1, IgG4 o IgE. Si, en canvi, és
IL12 o IL6, es diferenciarà a un limfòcit B de IgM. Si té IL10 i TGFβ, IgA. Si només té IL10,
IgG1 i IgG3. Si és IL21, serà IgG1, IgG3 i IgA1. Si té IL15, farà IgM, IgG1 i IgA. Gairebé cap
citocina fixa un isotip concret. El tipus de T helper fol·licular determinarà el canvi d’isotip que
es farà. El canvi d’isotip ve en funció de la inducció de la transcripció de les regions switch
que precedien els gens que codificaven per cadascuna de les regions constants.
Limfòcits B de memòria. Resultat de la interacció entre els limfòcits B i T a la zona clara, el limfòcit B es transformarà
en una cèl·lula plasmàtica o en una de memòria. Les plasmàtiques aniran als teixits
limfoides secundaris o aniran a emmagatzemar-se a la medul·la òssia per mantenir-s’hi durant
anys. IgA va a la mucosa intestinal. IgG van a la medul·la.
Les cèl·lules plasmàtiques de vida llarga es mantenen a la medul·la perquè hi troben el nínxol
que els permet sobreviure i mantenir la funció. Entre els factors que determinen que
s’emmagatzemin a la medul·la es troba la interacció entre CXCL12, que es produeix a la
medul·la, i CXCR4. La retenció també requereix de les cèl·lules de l’estroma de la medul·la,
que expressen molècules d’adhesió que ajuden a mantenir les cèl·lules allà. A més, també
expressen diferents factors de transcripció que mantenen aquest fenotip de cèl·lula
plasmàtica. Tant les cèl·lules de l’estroma com cèl·lules hematopoètiques produeixen
citocines que proporcionen els senyals de supervivència. Per tant, hi ha tota una sèrie de
factors que fan que les cèl·lules plasmàtiques de vida llarga es mantinguin vives durant anys
a la medul·la i que, quan és necessari, produeixin anticossos que es troben a la circulació
sanguínia i als diferents teixits.
Els limfòcits B seleccionats positivament es diferencien a cèl·lula de memòria o cèl·lula
plasmàtica. Ambdós tipus surten del centre germinatiu. Poden tornar a entrar-hi dins del
procés de proliferació i canvi d’isotip.
El resultat del procés d’interacció, diferenciació, activació, proliferació i generació de cèl·lules
plasmàtiques de vida llarga és la generació de cèl·lules B de memòria que van als teixits. El
concepte de memòria és el mateix que pels limfòcits T. Són limfòcits B amb un menor llindar
d’activació perquè presenten més molècules coestimuladores i necessiten menys senyal per
activar-se. A més, el menor llindar d’activació implica que es poden activar i desenvolupar els
seus mecanismes efectors de manera més ràpida. Les cèl·lules plasmàtiques productores
d’immunoglobulines de memòria ràpidament poden generar-ne en tornar a entrar en
contacte amb el mateix antigen que n’ha determinat l’activació i diferenciació. Les cèl·lules B
de memòria són més eficients. Ho són gràcies a tenir millor localització perquè es troben als
teixits i perquè tenen més precursors. Els precursors són limfòcits B amb afinitat per aquell
antigen arran de l’expressió clonal. En conseqüència, hi ha més limfòcits B amb la mateixa
especificitat antigènica. Aquests limfòcits B tenen major afinitat per l’antigen. Això es tradueix
en una major qualitat i una major quantitat. Aquest conjunt de característiques permet que,
quan aquests limfòcits B tornin a entrar en contacte amb l’antigen, la resposta sigui més ràpida
i molt més eficient.

Immunologia 2020
CEBIO 115
A la resposta primària s’aconsegueix uns nivells màxims de producció d’anticossos a partir
de 7 dies d’haver entrat en contacte amb l’antigen. Quan la resposta és secundària, és molt
més ràpida. La resposta secundària triga 3 dies en tenir inicia de resposta al tenir els limfòcits
de memòria. Entre 3 i 5 dies és el que es triga a obtenir el pic d’anticossos, la qual és entre
100 i 1000 vegades més gran que la primària. A la primera resposta predomina IgM mentre
que a la segona predominen altres isotips. Sobretot IgG si és resposta al plasma. A més, IgG
té més afinitat per l’antigen gràcies a la hipermutació somàtica.
Per tant, els limfòcits B que es converteixen en efectors poden produir immunoglobulines.
Aquests anticossos tenen funcions molt importants com la neutralització de microbis i toxines,
l’opsonització i fagocitosi dels microbis, ser un element clau en la citotoxicitat cel·lular
depenent d’anticòs i activar el sistema del complement. L’activació del sistema del
complement es tradueix en la lisi dels microorganismes, la inflamació i la fagocitosi dels
microorganismes opsonitzats amb fragments del sistema del complement. En si, el
mecanisme efector de les cèl·lules B és la producció d’anticossos.
Els anticossos són molt bons impedint la interacció de toxines amb els seus receptors a les
membranes de les cèl·lules. Aquest procés el realitzen interaccionant amb elles i impedint
l’adherència al receptor. Això és molt important en la resposta a virus, per exemple. Els
anticossos són importants en l’opsonització de bacteris, faciliten la seva internalització i
fagocitosi. Els anticossos també activen el complement i faciliten la destrucció d’alguns
bacteris.
Quan es produeix una resposta a un bacteri i aquest no està opsonitzat, té una supervivència
del 80%. Si ha estat opsonitzat, en canvi, la supervivència és del 1%. L’addició dels efectes
del sistema del complement redueix la població bacteriana al mínim en un temps de dues
hores. Tot això és perquè els anticossos faciliten la detecció per part de les cèl·lules
fagocítiques.
IgA i IgE tenen una funció molt important a les barreres de la pell i les mucoses associades
al tracte intestinal i respiratori. La IgA és la immunoglobulina secretada a les mucoses
associades al tracte intestinal i respiratori. S’hi troba en forma de dímer arran de la cadena J.
Per tal que la immunoglobulina passi al lumen de l’intestí, fa servir el receptor poliIg que
s’uneix al dímer i el transporta cap a la zona apical de les cèl·lules per transitosi. Abans de
sortir, es proteolitza el receptor poliIg i aquesta estructura de IgA dimèrica amb poliIg és el
que la fa tant estable al llum intestinal en condicions adverses. PoliIg és el que li dona
estabilitat i li permet fer la funció de neutralització.
Així dons, a l’intestí s’hi troba la IgA dimèrica, que opsonitza i facilita la detecció dels antígens
que allà es trobin. A l’interior del teixit tenim gran producció IgE, que pot sensibilitzar i activar
desgranulació mastòcits per atraure’ls i atraure altres cèl·lules i anticossos.

Immunologia 2020
CEBIO 116
Tema 22 – Regulació de la resposta immunològica. Les respostes immunitàries són resultat d’interaccions complexes entre cèl·lules i
mediadors diferents. Això fa necessitin estar molt regulades per tal de prevenir la resposta
a estructures pròpies o autoreactivitat. Si la regulació falla, es pot donar una resposta
excessiva que generi autoimmunitat, una resposta deficient que impedeixi la resposta a
tumors o una desviació de la resposta que doni lloc a reaccions d’al·lèrgia. Per altra banda,
la resposta immunitària s’ha de poder aturar quan ja s’ha eliminat l’antigen i el sistema
immunitari s’ha de mantenir en estat de no resposta enfront dels antígens propis. Així
i tot, el sistema immunitari ha d’aconseguir que hi hagi una gran estimulació quan interaccioni
amb un antigen forà i ha de retornar a l’estat de no resposta quan aquest desaparegui.
Tant important és aconseguir que no hi hagi
resposta a antígens propis com eliminar la
resposta quan ja no hi ha antígens. Si es
manté la resposta immunitària quan ja no
hi ha antígens forans es podria donar una
inflamació crònica, la qual comporta una
destrucció del teixit, un mal funcionament del
mateix i una alta possibilitat de desenvolupar
processos tumorals.
Tolerància Immunològica. La tolerància immunològica és la capacitat del sistema immunitari per decidir la no
resposta a un antigen determinat, sigui propi o no perillós. En principi, els al·lèrgens són
antígens no perillosos pels quals no hi ha una tolerància immunològica. Per tant, el sistema
immunitari ha d’aprendre a no respondre a allò que no sigui estrictament necessari. El
procés d’aprenentatge es fa en dues fases. La fase d’aprenentatge de la Tolerància Central
es genera als Òrgans Limfoides Primaris i serveix per generar un repertori de limfòcits T i B
poc o gens autoreactiu. Així es determina que hi hagi una poca capacitat de resposta al que
sigui propi. Així i tot, com que són poc reactius, hi ha una Tolerància Perifèrica que es genera
als Òrgans Limfoides Secundaris i que permet mantenir un estat de no resposta a estructures
pròpies.
Els mecanismes de manteniment de la tolerància central consisteixen en els processos
de selecció negativa dels limfòcits B autoreactius a la medul·la òssia i dels limfòcits T
autoreactius al timus. En canvi, la tolerància perifèrica es manté per altres vies i la seva
pèrdua es deu a gens inapropiats, a l’aparició de mutacions o a trobar-se en un entorn
inapropiat. La pèrdua de la Tolerància Perifèrica pot donar lloc a malalties
autoimmunitàries.
Mecanismes de la Tolerància Immunològica Perifèrica. La Tolerància Immunològica Perifèrica manté un repertori de cèl·lules que no genera
resposta a estructures pròpies quan es troben en condicions homeostàtiques. Per
aconseguir-ho, existeixen diferents mecanismes. Un mecanisme de la Tolerància Perifèrica
és mantenir un estat de no activació. En aquest és molt important la segregació de l’antigen,
segons el qual hi ha òrgans immunològicament privilegiats el parènquima d’aquests no
entra en contacte amb el sistema immunitari. En conseqüència, no s’hi pot donar una
resposta a antígens propis. L’estat de no activació també s’aconsegueix mantenint un estat
d’anèrgia, eliminant les cèl·lules autoreactives o fent un procés de supressió o regulació
mitjançada per limfòcits T reguladors i cèl·lules reguladores.

Immunologia 2020
CEBIO 117
Regulació per inducció d’anèrgia.
L’anèrgia és la manca de resposta a estímuls antigènics. Quan hi ha anèrgia, el resultat
de la interacció entre el limfòcit T i l’antigen presentat per una cèl·lula presentadora d’antigen
no es tradueix en una activació, proliferació i diferenciació efectora dels limfòcits. Enlloc
d’això, amb l’anèrgia s’indueix l’expressió d’una sèrie de gens que inhibeixen la
senyalització del TCR, inhibeixen l’expressió de citocines, degraden intermediaris de
la senyalització i bloquegen el cicle cel·lular. Els limfòcits T anèrgics, fruit dels gens
inductors de l’anèrgia, no secreten Interleuquina 2, la qual no pot actuar de manera autocrina
per permetre la seva proliferació. Els limfòcits B també poden entrar en un estat d’anèrgia.
En el seu cas, és el reconeixement de l’antigen per part del BCR el que no tindria efectes de
proliferació.
L’expressió dels gens inductors d’anèrgia es veu provocada per la presentació
d’antigen en absència de senyals coestimuladors activadors o d’elements de la
resposta innata. Quan això passa, s’indueix l’anèrgia clonal del limfòcit T autoreactiu. En
aquesta, el limfòcit té una baixa expressió dels complexes pèptid-MHC. Això provoca que,
quan una cèl·lula presentadora d’antigen interaccioni amb un limfòcit T anèrgic, tan
sols es donarà el primer senyal d’activació. Com que manca el segon senyal d’activació,
el primer senyal resultant de la interacció no serà prou intens com perquè es doni una
activació. Si hi ha un reconeixement d’antigen per part dels limfòcits T autoreactius però
no hi ha una senyalització mitjançant les molècules coestimuladores, el limfòcit es
converteix en un limfòcit T anèrgic. Això és perquè les molècules coestimuladores són les
que proporcionen el segon senyal d’activació. Un cop s’ha passat a un estat anèrgic, costa
molt que un limfòcit T respongui a l’antigen. Aquesta situació d’inducció d’anèrgia també
seria vàlida per l’activació del limfòcit B autoreactiu. Això és perquè el limfòcit B
autoreactiu no trobarà la col·laboració necessària en el limfòcit T activat. Per tant, l’anèrgia
és un sistema que funciona per evitar la resposta adaptativa.
Una altra manera d’induir anèrgia és la senyalització a través de molècules
coestimuladores inhibidores, com CTLA-4. Aquest inhibidor té els mateixos lligands que la
molècula coestimuladora activadora CD28. El lligand tant de CTLA-4 com de CD28 és B7,
altrament dit CD80/CD86. Els limfòcits T naive expressen CD28 de manera constitutiva i així
pot interaccionar amb B7 per transmetre un senyal de coestimulació activadora que
contribueix a l’activació del limfòcit. Quan s’expressa CTLA-4 es transmeten senyals
d’inhibició. Això és perquè CTLA-4 recluta fosfatases que defosforilen les cues
citoplasmàtiques dels receptors al limfòcit T. Així doncs, es pot dir que CTLA-4 indueix
anèrgia clonal.
PD-1 és una altra molècula coestimuladora inhibidora, però no competeix amb CD28 per
la unió amb CD80/CD86 al tenir el lligand propi PD-L1/PD-L2. El resultat de la interacció de
PD-1 i de CTLA-4 amb els seus lligands és la desfosforilació de les molècules
senyalitzadores. Això es tradueix en una aturada de la senyalització amb la conseqüent
disminució de la producció de citocines, de la proliferació i diferenciació de les cèl·lules
efectores i de la supervivència d’aquestes cèl·lules.
També es pot induir l’estat d’anèrgia als limfòcits B gràcies als receptors FcγRIIB per
tenir motius ITIM a la seva cua citoplasmàtica. Específicament parlant, el receptor és
CD32. Si es produeix un entrecreuament entre el BCR i la immunoglobulina que reconeix el
mateix antigen, la interacció de la immunoglobulina amb el receptor Fc activa les
fosfatases unides a la cua citoplasmàtica. Aquestes fosfatases defosforilen les cadenes
citoplasmàtiques de Igα i Igβ. En conseqüència, bloqueja la senyalització a través del BCR.

Immunologia 2020
CEBIO 118
Regulació per inducció de mort per apoptosi.
Una altra manera de regular la resposta del sistema immunitari és la inducció de la mort
per apoptosi. L’apoptosi és un procés fisiològic d’inducció de la mort cel·lular on intervenen
les caspases. L’apoptosi es caracteritza per canvis en la distribució dels lípids de la
membrana, una condensació i fragmentació nuclear, una fragmentació de l’ADN, una
vesiculació de la membrana i la formació de cossos apoptòtics. Les cèl·lules que entren en
apoptosi es fragmenten, però aquests fragments es mantenen envoltats de membrana.
És un procés molt diferent de quan una cèl·lula mor per necrosi. La necrosi és el procés pel
qual es perd la integritat de la membrana plasmàtica, el contingut cel·lular és degradat per
acció enzimàtica i el contingut citosòlic s’allibera a l’espai extracel·lular. En conseqüència, la
necrosi genera gran quantitat de DAMPs, Patrons Moleculars Associats a Perill. Per tant,
la necrosi genera una inflamació.
Des de fa poc s‘ha vist que quan els cossos apoptòtics són fagocitats, les cèl·lules que
ho han fet secreten citocines antiinflamatòries com IL-10 i TGF-beta. A més, les cèl·lules
que les han fagocitat disminueixen la secreció de citocines inflamatòries com TNF-α, IL-1β i
IL-12. És per això que es pot dir que el resultat de la mort per apoptosi és una mort silent,
que no genera activació i inflamació del sistema immunitari i que genera senyals que
inhibeixen la resposta del sistema immunitari.
La inducció de l’apoptosi es pot produir de dues maneres. La mort per omissió és la inducció
de l’apoptosi que es dona quan s’ha eliminat completament l’antigen o baixa tant la secreció
d’IL-2 que no n’hi ha a l’entorn que no la pot fer servir el limfòcit T. La mort per activació és
la inducció de l’apoptosi que es dona quan l’antigen és molt persistent i hi ha una estimulació
repetida del limfòcit T.
La mort per omissió es dona en cas que
hi hagi una absència d’estímuls de
supervivència, ja siguin factors de
creixement, antigen o interleuquina 2. En
la mort per omissió s’altera la permeabilitat
de la membrana dels mitocondris, el que
allibera el citocrom C al citoplasma. La unió
del citocrom C a la proteïna Apaf-1 activa la
caspasa 9. La caspasa 9, quan està activa,
activa la caspasa 3. La caspasa 3 trenca el
DNA de doble cadena i indueix l’apoptosi.
La inducció de l’apoptosi també es pot donar per
l’activació de Fas i del lligand de Fas. Fas
s’expressa a diferents tipus cel·lulars, com les
cèl·lules epitelials o els limfòcits. El lligand de Fas
només s’expressa als limfòcits T fruit d’una activació
repetida i l’acció de la IL-2. Per tant, quan hi ha una
activació continuada del limfòcit T, es comença a
expressar el lligand de Fas a la seva membrana. Es
pot produir la interacció entre Fas i el seu lligand estant
els dos a la membrana del limfòcit T. La interacció
entre Fas i FasL provoca que la proteïna FADD
s’uneixi al domini de mort cel·lular de la cua
citoplasmàtica de les proteïnes Fas. La interacció
activa la caspasa 8. La caspasa 8 activa la caspasa 3.
Alteració Membrana Mitocondris.
Alliberació Citocrom C al citoplasma.
Unió Citocrom C al Apaf-1.
Activació Caspasa 9.
Activació Caspasa 3.
Expressió de FasL.
Interacció Fas-FasL.
Unió FADD a Fas.
Activació Caspasa 8.
Activació Caspasa 3.

Immunologia 2020
CEBIO 119
Regulació per cèl·lules T reguladores.
Els timòcits que tenen una afinitat intermèdia pels antígens propis es diferencien en
cèl·lules T reguladores. Aquestes cèl·lules T reguladores es generen des del primer moment
del individu i contribueixen a mantenir l’estat de no resposta a la perifèria. Així i tot, es
generen cèl·lules T reguladores durant la resposta a la perifèria com a resultat de
l’activació. Quan hi ha una activació i una producció de TGF-β o àcid retinoic, els limfòcits T
que reconeixen antigen es poden diferenciar a cèl·lules T reguladores perifèriques o cèl·lules
T reguladores induïdes. Independentment de quin tipus siguin, la funció de les cèl·lules T
reguladores és aturar la resposta del sistema immunitari i impedir que les cèl·lules T
efectores continuïn activant-se.
S’han descrit vàries poblacions de cèl·lules T reguladores. Així i tot, la majoria de cèl·lules
T reguladores són limfòcits TCD4 que expressen CD25. Aquesta molècula és la cadena
α del receptor de la IL-2, aquella que permet generar IL2R d’alta afinitat. Les cèl·lules T
reguladores també expressen CTLA-4 de forma constitutiva, la molècula
coestimuladora inhibitòria GITR i el factor de transcripció Foxp3.
L’elevada expressió de CD25 significa que tenen un receptor d’alta afinitat per la IL-2.
En cas que aquesta citocina es trobi al medi, aquestes cèl·lules la podran captar a través de
la interacció amb aquest receptor d’alta afinitat. Per altra banda, l’expressió del factor de
transcripció Foxp3 té una funció molt important. Per una banda, Foxp3 s’uneix al promotor
dels gens de CD25, CTLA-4 i GITR alhora que indueix l’acetilació de les histones,
facilitant la transcripció dels gens que codifiquen per aquestes proteïnes. Per altra banda, la
unió de Foxp3 als promotors de la IL-2, IL22 o de l’interferó γ recluta enzims
desacetilases que en reprimeixen la transcripció al desacetilar les histones.
L’activitat de les cèl·lules T reguladores és dependent d’antigen. Dit d’una altra manera,
les cèl·lules T reguladores han de reconèixer antigen a través del seu TCR per manifestar la
seva funció reguladora. El fet que tinguin un TCR d’afinitat intermèdia implica que sol tenir
més afinitat que els TCR de les cèl·lules autoreactives. Per tant, les cèl·lules T reguladores
es poden activar a concentracions d’antigen molt baixes. Un cop s’han activat, les
cèl·lules T reguladores inhibeixen les cèl·lules T efectores, aquelles que estan responent o
respondrien.
Hi ha diferents maneres d’inhibir les cèl·lules T efectores. D’una banda, es pot inhibir la
transcripció de la IL-2 a la cèl·lula efectora. A més, ho faria competint per la IL-2 existent,
el que facilita la inducció de l’anèrgia a les cèl·lules T efectores. D’altra banda, les cèl·lules T
reguladores expressen molècules coestimuladores inhibidores com el CTLA-4, PD-1 i LAG-
3. Per tant, el competir per la interacció amb B7 impedeix que el limfòcit T rebi el segon
senyal i que es pugui activar. Quan CTLA-4 s’uneix a B7.1 o B7.2, augmenta la producció
d’enzims com IDO. IDO és un enzim que metabolitza el triptòfan, un aminoàcid essencial per
les funcions cel·lulars. Segrestant, eliminant i metabolitzant el triptòfan, les cèl·lules T
reguladores impedeixen o dificulten la activació i proliferació dels limfòcits T efectors.
A més, les cèl·lules reguladores poden produir citocines immunosupressores com IL-10, TGF-
β i la IL-35; que inhibeixen la inflamació i la resposta.
Les cèl·lules T reguladores fan la seva funció un cop han reconegut antigen, pel que la
supressió i regulació de la resposta immunitària que exerceixen les cèl·lules T reguladores és
específica d’antigen. Així i tot, el fet que puguin produir citocines immunosupressores fa
que, un cop activades, les cèl·lules T reguladores puguin suprimir limfòcits amb una
especificitat antigènica diferent només perquè es troben per la zona. Per tant, tot i que
l’activitat supressora és depenent d’antigen i això implica que regularan primer les cèl·lules

Immunologia 2020
CEBIO 120
efectores amb la mateixa especificitat, la seva funció efectora un cop activades ja no és
específica d’antigen. Poden inhibir la funció d’altres cèl·lules T efectores.
L’activitat reguladora a través de les citocines inhibidores com IL-10, TGF-β i IL-35 es pot
exercir directament sobre els limfòcits T inhibint la proliferació, inhibint la diferenciació de
limfòcits T naive a limfòcits T efectors, suprimint l’activitat de les cèl·lules T efectores quan ja
estan diferenciades, contribuint a la inducció de les cèl·lules T reguladores perifèriques o
mantenint les cèl·lules T reguladores d’origen tímic.
Així i tot, les cèl·lules T reguladores també poden fer el seu efecte sobre cèl·lules
presentadores d’antigen al convertir-les en cèl·lules tolerogèniques. Això es pot fer
inhibint l’expressió de molècules coestimuladores activadores a les cèl·lules presentadores
d’antigen. D’aquesta manera evitarà que proporcionin un segon senyal d’activació i així
anergitzen les potencials cèl·lules inhibidores. Un altre mètode descrit i que es fa servir per
immunoteràpia és que les cèl·lules T reguladores poden inhibir la maduració de les
cèl·lules dendrítiques. Això impedeix que es transformin en cèl·lules presentadores molt
potents, capaces d’activar els limfòcits T naive. A més, les cèl·lules T reguladores poden
incidir en les cèl·lules presentadores d’antigen tot induint l’activació de macròfags M2
per la secreció de les interleuquines 10, 4 i 13. Els macròfags de tipus M2 ajuden a acabar
la resposta i reparar el teixit malmenat.
Una altra manera per la qual les cèl·lules T reguladores poden regular la resposta dels
limfòcits T és intervenint en el seu metabolisme. Se sap que les cèl·lules T reguladores
expressen CD39 i CD73 a la membrana, ectoenzims que passen l’ATP a AMP i que
produeixen adenosina. Aquesta purina té activitat immunoreguladora i està molt ben descrita.
Per altra banda, les cèl·lules T reguladores expressen AMP cíclic intracel·lular que
transfereixen a les cèl·lules T efectores. L’acció de l’AMP cíclic és inhibir NFAT, pel que hi
ha una inhibició de la transcripció de la interleuquina 2. En absència de la IL-2, fàcilment
s’indueix l’apoptosi. Interferint en el metabolisme tant de les cèl·lules efectores com de
les cèl·lules presentadores d’antigen, les cèl·lules T reguladores poden evitar que hi
hagi una resposta dels limfòcits T.
Tolerància Perifèrica als Limfòcits B.
Els mecanismes bioquímics que s’apliquen sobre les cèl·lules T efectores també es
donen als limfòcits B per tal de mantenir-hi la tolerància perifèrica. Això es pot fer
mitjançant els receptors inhibidors CD22 i CD32 que impedeixen la senyalització, per la
inducció de l’apoptosi quan es reconeixen antígens propis sense la col·laboració dels limfòcits
T, quan es reconeixen antígens propis amb alta afinitat o quan es reconeixen antígens propis
repetidament, el que indueix l’anèrgia. Els mecanismes cel·lulars són semblants per
limfòcits T efectors i per a limfòcits B.
Regulació de la resposta un cop s’ha iniciat. Hi ha mecanismes de regulació que eviten la resposta del sistema immunitari i hi ha
mecanismes que controlen la resposta un cop iniciada. La regulació de la resposta un
cop ha estat iniciada tendeix a aturar-la o inhibir-la. El control de la resposta immunològica
iniciada pot ser:
Regulació per anticossos.
Regulació per antigen.
Regulació per limfòcits.
Modulació idiotípica de la
resposta.
Modulació neuroendocrina de la resposta.

Immunologia 2020
CEBIO 121
Modulació Neuroendocrina de la resposta.
Un exemple rellevant de la modulació neuroendocrina de la resposta és que l’estrès
pot provocar la supressió de la resposta immunològica. En situacions d’estrès, el sistema
immunitari funciona pitjor perquè aquest estrès redueix la capacitat de resposta a diferents
tipus de patògens. Això passa perquè existeix una interconnexió entre els sistemes
immunològic, endocrí i nerviós. La interconnexió no és casualitat, doncs la majoria dels
teixits limfoides estan innervats pel sistema nerviós simpàtic.
Per altra banda, el sistema nerviós controla la producció de diverses hormones da
manera directa o indirecta. Entre aquestes hormones estan la del creixement, la tiroxina,
l’adrenalina, els corticoesteroides... A més, els limfòcits expressen receptors per aquestes
hormones, neurotransmissors i neuropèptids. En conseqüència, la connexió entre els
sistemes immunitari, nerviós i endocrí és una manera de regular el funcionament del primer.
En això és important l’efecte dels glucocorticoides.
S’ha vist que els glucocorticoides modulen la transcripció dels gens de les citocines tot
suprimint els gens de les citocines pro-inflamatòries i regulant la producció de les
antiinflamatòries. Els glucocorticoides també modulen l’expressió de molècules
d’adhesió i la producció de quimiocines, pel que regulen el trànsit de cèl·lules del sistema
immunitari cap a les zones d’inflamació. En afegit, els glucocorticoides suprimeixen la
síntesi de mediadors d’inflamació com la prostaglandina i l’òxid nítric sintasa pel que,
d’alguna manera, els glucocorticoides rebaixen la inflamació. Els glucocorticoides
suprimeixen la maduració, diferenciació i proliferació de les cèl·lules del sistema
immunitari, incloses les de la immunitat innata, els limfòcits T i els limfòcits B. L’ús dels
glucocorticoides, quan es fan servir com a tractament per modular la resposta del sistema
immunitari, ha de ser limitada i controlada.
Per fer totes aquestes accions, els glucocorticoides fan servir el receptor de
glucocorticoides expressats a les cèl·lules del sistema immunitari. La interacció dels
glucocorticoides amb els seus receptors modula l’expressió gènica perquè intervé en
l’activació de factors com NFκB i en l’acció d’AP-1. Per tant, els glucocorticoides
impedeixen que el factors de transcripció NFκB i AP-1 actuïn al nucli, determinant la
transcripció de gens molt concrets.
Hi ha molts sistemes de regulació del sistema immunitari i tots han de funcionar
correctament. El mal funcionament de qualsevol d’aquests sistemes pot donar lloc a diferents
patologies o a l’absència de la resposta, doncs una mala regulació pot impedir que es
respongui a cèl·lules tumorals. Per tant, tots els mecanismes són molt importants i són
molt diversos.

Immunologia 2020
CEBIO 122
Tema 23 – Resposta a bacteris. Els microorganismes i els seus hostes estan en lluita constant per la supervivència.
Aquesta lluita tendeix a un equilibri inestable entre els dos, doncs l’objectiu del patogen és
infectar l’hoste i multiplicar-s’hi, mentre que l’objectiu de l’hoste és eliminar el patogen o reduir-
lo a un paràsit inofensiu. Per tal d’infectar l’hoste i multiplicar-s’hi dins, els bacteris evadeixen
els mecanismes de defensa de l’individu però sense immunodeprimir-lo prou com
perquè faciliti la infecció per altres patògens. De vegades, els mecanismes de defensa
aconsegueixen impedir l’entrada del patogen i fan predominar l’individu. Altres cops, els
patògens aconsegueixen infectar les cèl·lules i els teixits. Entre els mecanismes de defensa
de l’hoste contra bacteris també hi intervé la microbiota sapròfita, organismes comensals que
inhibeixen de manera competitiva la fixació de patògens a la cèl·lula hoste. En conseqüència,
té unes funcions molt importants en el manteniment de l’equilibri.
Microbiota. La microbiota associada a cada organisme és una comunitat ecològica de
microorganismes comensals simbiòtics que es troba a tots els organismes
multicel·lulars, des de plantes fins a animals. La microbiota inclou bacteris, arqueus, protists,
fongs i virus. Dels que més es coneix informació són els bacteris, però és molt important la
participació dels fongs i els virus. S'ha demostrat que la microbiota és crucial per a
l'homeòstasi metabòlica, hormonal i immunològica de l’hoste. Proporcionen vitamines i
nutrients. La microbiota participa en el manteniment i millora de la barrera epitelial dels teixits
de les mucoses, el desenvolupament del sistema immunitari i l’adquisició de factors que
contribueixen a la seva regulació. La microbiota té un paper molt important en el manteniment
de l’equilibri inestable entre individus i patògens.
Mecanismes d’acció de la microbiota sapròfita sobre el Sistema Immunitari.
La microbiota sapròfita actua de diverses maneres per proporcionar una protecció
directa contra els patògens. Entre aquestes maneres estan:
L’alteració de les condicions físiques i químiques del microambient s’aconsegueix consumint
oxigen i alterant el pH, el que provoca que els patògens tinguin menys capacitat de
proliferació. Les cèl·lules de Paneth es veuen induïdes, principalment, a produir RegIIIγ i AMP,
els quals tenen un paper molt important en la neutralització dels bacteris, en evitar que entrin
en contacte amb les cèl·lules que formen la barrera epitelial i en evitar que entrin al
parènquima del teixit.
La microbiota sapròfita també pot proporcionar una protecció indirecta al afavorir la
maduració del teixit limfoide associat a la mucosa, ajudar a estructurar les plaques de Peyer,
ajudar a mantenir la inflamació fisiològica per mitjà de Treg i la generació de Th17 i al induir
la resposta dels limfòcits B que donin lloc a la producció d’immunoglobulina A.
Competència directa per nutrients.
Alteració de les condicions físiques i
químiques del microambient local.
Producció de productes
bactericides.
Modulació de l’expressió de factors
de virulència.
Inducció de l’expressió de
mucines per part de les cèl·lules de
Goblet.
Inducció de la producció de pèptids antimicrobians per
les cèl·lules de Paneth.

Immunologia 2020
CEBIO 123
L’estructura del MALT restringeix les interaccions entre cèl·lules dendrítiques i els
limfòcits B o entre cèl·lules dendrítiques i cèl·lules T als ganglis mesentèrics, evitant la
resposta immunitària sistèmica. El MALT és el sistema immunitari associat a les mucoses.
La microbiota sapròfita és molt important per contribuir al desenvolupament del
sistema immunitari associat a les mucoses. Una de les coses que fa és activar els
macròfags intestinals, el que augmenta la forma pro-interleuquina-1β. La infecció per
patògens activa el pas de la forma inactiva a la forma activa de la IL-1β. En conseqüència, es
promou el reclutament dels neutròfils i l’erradicació dels patògens.
La microbiota és molt important en la promoció de la diferenciació i activació de les
cèl·lules Th17 i de les innate lymphoid cells. El conjunt de Th17 i les ILC controlen tant els
comensals com els patògens per mitjà de la secreció de citocines. Entre les citocines
secretades pels limfòcits Th17 i les ILCs es troben la IL-22. Per tant, en la separació entre el
món extern dels patògens i el mon intern dels hostes es troba la microbiota.
Funcions de la microbiota sapròfita.
Els elements ambientals, la contaminació, els pesticides, els factors genètics, els factors
epigenètics, l’estil de vida, els medicaments i els antibiòtics entre d’altres elements poden
alterar la microbiota. L’alteració de la microbiota provoca una alteració de tots els
mecanismes de control que exercien. En conseqüència, es veurà afavorida l’entrada dels
possibles elements patogèniques que estaven sent controlats per tots aquests mecanismes i
que puguin entrar al teixits causant una infecció. Si la infecció arriba als vasos sanguinis,
pot ser sistèmica. En el cas de les infeccions per bacteris, aquest és un pas clau. Quan es
produeix la disrupció de la barrera de la microbiota, a l’organisme entren tant patògens com
elements de la microbiota que poden tenir efectes patogènics.
És el cas de diferents especies bacterianes. Staphylococcus aureus colonitza les foses
nassals d’entre el 20 i el 50% dels individus. La presència d’aquest bacteri a les fosses no
té conseqüències patològiques, però si que les té quan és ingerit i passa a les vies
respiratòries. Diferents espècies de Clostrídium es troben normalment formant part de
la microbiota intestinal. Si entrés per una via diferent d’aquesta, pot generar el tètanus. Hi
ha molts altres microorganismes que poden causar meningitis o gonorrea depenent de la via
per on entrin. Una alteració de la microbiota suposa no només l’entrada de patògens
sinó també l’entrada d’elements de la mateixa microbiota, el que pot suposar un problema
si arriben al parènquima dels teixits.
Entrada dels patògens. Tant diversos són els bacteris com les seves vies d’entrada. Les vies d’entrada dels
bacteris poden ser les superfícies de les mucoses de les vies respiratòries, del tracte
gastrointestinal o el tracte reproductiu; les superfícies externes, les ferides o rascades o les
injeccions per la picada d’insectes. El mode de transmissió dels microorganismes serà
diferent per cadascun d’ells. La via d’entrada d’un microorganisme determina la resposta
immunitària que es faci servir.
Un cop han entrat, cal distingir entre els bacteris amb una vida extracel·lular d’aquells
que tenen una vida intracel·lular. Tenen una vida extracel·lular els bacteris que colonitzen
els espais intersticials, la sang, la limfa o les superfícies epitelials. Els bacteris intracel·lulars
entren a l’interior de les cèl·lules dels teixits. Dins les cèl·lules, els bacteris es poden
mantenir al citoplasma o a compartiments vesiculars. Els mecanismes del sistema
immunitari per eliminar un bacteri extracel·lular són diferents dels que farà servir per eliminar
bacteris intracel·lulars.

Immunologia 2020
CEBIO 124
Paral·lelament als bacteris extracel·lulars i intracel·lulars hi ha un grup de bacteris que
entren a l’interior de les cèl·lules per començar la seva replicació però que es troben a
l’espai extracel·lular. Dependents del moment de la vida es trobin aquests bacteris, seran
més eficients els mecanismes de resposta davant de bacteris extracel·lular o els de resposta
davant de bacteris intracel·lulars. Les infeccions per qualsevol tipus de bacteri consisteixen
en 4 etapes:
Les cèl·lules a les quals s’adhereixen els bacteris poden ser cèl·lules que fan de barrera
o poden ser les cèl·lules diana a les que entren. Són un exemple de cèl·lules que fan de
barrera les cèl·lules epitelials de la paret intestinal. La lesió de la cèl·lula es pot fer per diversos
mètodes, un dels quals són les toxines.
Bacteris Extracel·lulars. Els bacteris extracel·lulars proliferen fora de les cèl·lules de
l’hoste i causen malalties, problemes o simptomatologia per
dos mecanismes principals. Per un costat, els bacteris
extracel·lulars causen una gran inflamació. Aquesta deriva
en una important destrossa del teixit i en la formació de pus.
Aquest mecanisme de gran inflamació és el que fan servir els
bacteris Staphylococcus i Streptococcus entre d’altres. Són els
bacteris que trobem quan tenim mal de coll i causen una
inflamació.
L’altre mecanisme dels bacteris extracel·lulars consisteix
en la producció de toxines. Aquestes poden ser exotoxines
o endotoxines. Les exotoxines són toxines amb un efecte
directe sobre aquelles cèl·lules que tinguin un receptor per
elles. És el cas de la Diftèria toxinodiftèrica o el tètanus. Les
endotoxines són toxines amb un efecte immunopàtic.
L’endotoxina LPS activa els macròfags, els quals indueixen la
secreció de gran quantitat de citocines proinflamatòries com
TNF o IL-1, el que pot provocar una febre alta i un xoc sèptic.
Les conseqüències de la infecció per bacteris extracel·lulars són les causants de gran part de
la patologia. Davant bacteris extracel·lulars, el sistema immunitari té com a objectiu
eliminar el bacteri i neutralitzar els efectes de les toxines.
Adhesió dels bacteris a la cèl·lula de l’hoste.
Proliferació del Bacteri.
Invasió del teixit de l’hoste.
Lesió de la cèl·lula hoste.
Inflamació
oDestrossa del sistema immunitari.
oFormació de pus.
Toxines
oExotoxines.
oEndotoxines.

Immunologia 2020
CEBIO 125
Resposta innata davant de Bacteris Extracel·lulars.
Per eliminar els bacteris extracel·lulars, el principal mecanisme efector de la resposta
innata és la fagocitosi. La fagocitosi consisteix en internalització dels bacteris extracel·lulars
per tal que pugin ser eliminats per les cèl·lules fagocítiques. Les principals cèl·lules que
duran a terme la fagocitosi dels bacteris extracel·lulars són els neutròfils i els
macròfags, als qui detectaran mitjançant els receptors PRR. Els receptors PRR més
rellevants per aquesta funció són els receptors scavenger, els receptors de tipus toll i els
receptors de manosa. Tots aquests PRR poden detectar el bacteri. La fagocitosi s’afavoreix
amb el reconeixement de bacteris opsonitzats per immunoglobulines o proteïnes del
sistema del complement. Aquest reconeixement es fa a través de receptors de la fracció
constant de les immunoglobulines o receptors del sistema del complement. Aconseguir la
fagocitosi dels bacteris extracel·lulars és el mecanisme principal de resposta de la immunitat
innata als bacteris extracel·lulars. En la fagocitosi s’estimula la capacitat bactericida dels
fagòcits. La fagocitosi depèn molt dels receptors PRR. Hi ha molts snips que determinen
la capacitat o resposta dels PRR. Hi ha snips de TLR4 que afavoreixen la resposta a LTS
mentre que n’hi ha de TLR2 que afavoreixen la detecció del peptidoglicà. Per tant, els snips
fan que la identificació i fagocitosi dels bacteris extracel·lulars sigui més eficient.
Els elements del sistema del complement també contribueixen a la resposta innata
contra els bacteris extracel·lulars. Sobretot els elements de la via alternativa o de la via de
les lectines. La via alternativa contribueix perquè components com el LPS són capaços
d’iniciar el trencament espontani de C3. Per altra banda, la paret cel·lular de molts bacteris té
una alta concentració de carbohidrats i polisacàrids que es poden reconèixer per elements
que activen la via de les lectines. L’activació del complement genera opsonines i
anafilotoxines. Les opsonines faciliten la fagocitosi. Les anafilotoxines indueixen tant la
inflamació com el reclutament de leucòcits.
Els macròfags produeixen les citocines IL-12 i TNF-alpha. Les cèl·lules NK produeixen
Interferó gamma. El conjunt d’aquestes citocines proinflamatòries i les quimiocines que
atrauen als leucòcits al punt de la infecció seran molt importants per respondre als bacteris
extracel·lulars.
Es produeixen danys en el teixit. El teixit produirà DAMPS. Els DAMPS seran detectats per
PRR i contribuiran a la inflamació. La inflamació consisteix en l’atracció de cèl·lules i
productes del sistema immunitari per tal d’eliminar els bacteris.
La importància del sistema del complement en la resposta innata depèn parcialment de si són
gram positius o gramnegatius. Els bacteris gramnegatius són susceptibles a la lisi directa pel
complement. En el cas dels bacteris grampositius, el gran gruix de la paret bacteriana
impedeix la formació del MAC, ple que no solen ser susceptibles a la lisi pel complement. Així
i tot, els elements del sistema del complement actuen com a opsonines, el que facilita la seva
fagocitosi i eliminació.
Resumint la immunitat innata, l’entrada del bacteri extracel·lular provocarà l’activació del
complement. El sistema del complement produirà opsonines i anafilotoxines. Ambdues
activaran algunes cèl·lules de la immunitat innata, com els mastòcits perquè es desgranulin i
alliberin histamina. Les anafilotoxines també actuaran com a elements quimiotàctics
juntament amb la IL-8 i mediadors d’inflamació. Els elements quimiotàctics podran atraure
cèl·lules fagocítiques per tal que intentin fagocitar els bacteris i aconseguir la seva eliminació.
Les cèl·lules de la immunitat innata que s’activaran produiran citocines
proinflamatòries que augmentaran les molècules d’adhesió i la permeabilitat vascular.
Això activarà la fase aguda.

Immunologia 2020
CEBIO 126
Els bacteris procariotes extracel·lulars inicien les seves seqüències proteiques per metionines
formilades. Per tant, els bacteris generen pèptids formilats, els quals actuen com a agents
quimiotàctics que detecten les cèl·lules del sistema immunitari.
Resposta adaptativa humoral davant de Bacteris Extracel·lulars.
En el cas dels bacteris extracel·lulars, el principal mecanisme efector de la resposta
adaptativa és la resposta humoral. Això és així perquè els principals antígens bacterians
són polisacàrids. Aquest fet implica que generen una resposta a antígens T-independents, la
qual genera plasmoblasts de vida curta. Els plasmoblasts de vida curta produeixen anticossos
poc potents durant un temps limitat. La baixa potència dels anticossos es deu a no fer un
procés d’hipermutació somàtica, a generar poca memòria immunològica i a que les
cèl·lules plasmàtiques no tinguin una vida llarga. És per això que la majoria de resposta
a antígens T-independents són respostes de producció de IgM i algunes poques IgG.
També hi ha una resposta dels limfòcits B dependent de la col·laboració amb els
limfòcits T. En aquest cas, la resposta sol ser a antígens proteics, el que la fa més efectiva.
La resposta a antígens proteics provoca la resposta del centre germinatiu, el qual permet
produir anticossos d’alta afinitat i amb canvi d’isotip. L’isotip dels anticossos que surten
del centre germinal és el que millor s’adapta a la resposta. Generalment, els isotips són
IgG i IgA generats per cèl·lules plasmàtiques de vida llarga. Per tant, els anticossos tenen
una vida mitjana llarga i es genera una bona memòria.
La resposta humoral produeix anticossos. Els anticossos tenen tres mecanismes efectors
importantíssims. D’una banda, els anticossos han de neutralitzar els bacteris i les toxines
bacterianes. La neutralització consisteix en impedir que els bacteris i les toxines s’uneixin a
la cèl·lula de l’hoste. En la neutralització són molt importants la IgA i la IgG. La IgG depèn de
la superfície. Els anticossos ajuden en l’opsonització i fagocitosi dels bacteris. En
l’opsonització i fagocitosi és molt important la IgG al ser la millor per facilitar la fagocitosi a
través dels receptors Fcγ. Els anticossos faciliten l’activació de la via clàssica del
complement. Per tant, els anticossos facilitaran tots els mecanismes derivats de l’activació
del complement com la lisi directa via mac o la col·laboració en l’opsonització. Tots els
mecanismes han de tendir a impedir que la toxina o el bacteri entrin en contacte amb les
cèl·lules del teixit que fan de barrera.
Resposta adaptativa cel·lular davant de Bacteris Extracel·lulars.
La participació de la resposta adaptativa cel·lular és molt important en la resposta als
bacteris extracel·lulars. Si són bacteris extracel·lulars, seran reconeguts pels fagòcits a
través de la via endocítica. En conseqüència, els bacteris seran processats i presentats per
molècules del MHC de classe II a limfòcits Thelper CD4, el que en provocarà l’activació i
diferenciació. Els Limfòcits es diferenciaran en els seus diferents tipus efectors en funció del
Entrada del bacteri
extracel·lular.
Activació del sistema del
complement.
Producció d’opsonines i anafilotoxines.
Activació de cèl·lules de la
immunitat innata.
Alliberament d’histamina.
Quimiotaxi de les cèl·lules
fagocítiques.
Producció de citocines
proinflamatòries.
Augment de molècules
d’adhesió i de permeabilitat
vascular.

Immunologia 2020
CEBIO 127
patogen i la via d’entrada. Cada tipus de limfòcit T efector secreta unes citocines concretes i
té unes tasques específiques.
Si es produeix molta IL-17, els limfòcits es diferenciaran en limfòcits Th17. Si es produeix molt
de TNF, els limfòcits es diferenciaran en limfòcits Th1. Els limfòcits Th1 i Th17 atrauen i
activen macròfags i neutròfils. Tant els macròfags com els neutròfils tenen alhora una
capacitat fagocítica bactericida i una capacitat de produir citocines proinflamatòries.
Si es produeix molt INF-γ, s’activen els macròfags. L’activació dels macròfags fa augmentar
la fagocitosi i lisi bacterianes. Els macròfags provoquen el manteniment de la diferenciació en
limfòcits Th1, ela quals produeixen INF-γ.
En la resposta adaptativa cel·lular a bacteris extracel·lulars prenen molta rellevància les
citocines que realitzen accions en la col·laboració amb els limfòcits B, doncs el principal
element en la resposta adaptativa a bacteris és la resposta humoral. Els limfòcits TCD4 helper
fol·liculars contribuiran a l’activació dels limfòcits B en la producció del centre germinal i per a
totes les reaccions que hi tenen lloc. Les reaccions que es donen en un centre germinal són
la hipermutació somàtica i el canvi d’isotip. La col·laboració entre limfòcits T helper CD4 i
limfòcits B augmenta la producció d’anticossos.
Els ThCD4, produint IL4 i IL5, contribueixen en l’activació de les cèl·lules del sistema
immunitari com els eosinòfils. Aquests són importants en l’eliminació dels bacteris
extracel·lulars per la desgranulació i alliberament de factors microbicides. La IL-4 contribueix
a la activació i proliferació dels limfòcits B i la seva diferenciació a cèl·lules plasmàtiques
productores d’immunoglobulina.
La infecció per bacteris que colonitzen preferentment les mucoses, (i.e. S. pneumoniae o H.
pylori), sovint es tradueix en la generació de cèl·lules efectores Th17. L’apoptosi de cèl·lules
de la mucosa en un entorn inflamatori proporciona una combinació de PAMPs i DAMPs que
indueixen les cèl·lules dendrítiques a produir TGFβ i IL-6, el que promou la diferenciació a
TH17. En canvi, una cèl·lula que pateix apoptosi en absència d'infecció produeix només
DAMPs, que indueixen les cèl·lules dendrítiques a segregar només TGFβ i no de citocines
pro inflamatòries, pel que afavoreix la generació de cèl·lules T reguladores.
Les cèl·lules T helper 17 fan IL17 i provoquen la segregació de IL22 per part de les cèl·lules
de la mucosa. La combinació d’aquests dos elements fa que les cèl·lules epitelials produeixin
pèptids antimicrobians i factors quimiotàctics que serveixin per reclutar neutròfils. A més, les
cèl·lules dendrítiques activades secreten citocines proinflamatòries que diferencien cèl·lules
TCD4 a cèl·lules Th17 efectores.
Conseqüències patològiques de la resposta a Bacteris Extracel·lulars.
Sovint, la resposta a bacteris extracel·lulars té conseqüències patològiques. Es pot produir
una inflamació local molt aguda quan hi ha una bona combinació de resposta innata, limfòcits
Th17 i anticossos. Una resposta local molt aguda pot provocar danys als teixits o pot arribar
a tenir efectes sistèmics mitjançant la producció elevada de citocines. Si aquesta producció
és massiva, es pot arribar a produir un xoc sèptic.
Un xoc sèptic és l’activació de tots els endotelis. Per tant, un xoc sistèmic és la producció
massiva de citocines inflamatòries que es troben a tots els teixits i òrgans. Un xoc sèptic
provoca l’efecte exagerat de les citosines. Un xoc sèptic provoca hipotensió, col·lapse
circulatori sistèmic, coagulació intravascular disseminada i altres canvis metabòlics que
poden acabar amb la fallada multiorgànica.

Immunologia 2020
CEBIO 128
D’altra banda, si la producció d’anticossos és exagerada, hi pot haver diverses
conseqüències:
• Els anticossos poden donar lloc a immunocomplexes de mida molt gran que
s’acumulen als glomèruls renals.
o Això produeix inflamació als glomèruls.
o Provoca un mal funcionament dels ronyons.
• Els anticossos poden tenir una reactivitat creuada amb proteïnes de la membrana de
la cèl·lula muscular i la miosina del cor.
o Genera problemes a nivell cardíac.
• Pot haver-hi una patologia autoimmunitària.
Mecanismes d’evasió dels bacteris extracel·lulars.
Hi ha quasi tants mecanismes d’evasió del sistema immunitari com espècies bacterianes que
poden infectar l’ésser humà. El primer pas d’una infecció és l’adhesió a una cèl·lula epitelial
o a una cèl·lula que s’infectarà. Molts bacteris tenen incrementada l’adhesió no específica
que els permet interaccionar amb les cèl·lules de barrera o hoste. Aquest increment en
l’adhesió augmenta la infecció.
També han desenvolupat mecanismes per combatre la fagocitosi. E. coli i Pseudomonas
aeruginosa secreten molècules que disminueixen la fagocitosi per neutròfils. Staphylococcus
aureus inhibeix la fagocitosi que es realitza a través dels receptors dels dominis Fc de les
immunoglobulines. Tenen una proteïna que expressen a la superfície bacteriana i s’hi uneix.
Per això són capaços de bloquejar la interacció d’aquests receptors amb les
immunoglobulines. Els bacteris també han desenvolupat modificacions a l’estructura
antigènica externa. És el que es coneix com Variació antigènica. Són modificacions que eviten
ser detectats per elements del sistema immunitari. E. coli, per exemple, presenta més de 106
antígens diferents que poden permetre evadir-se del sistema immunitari. Alguns bacteri
Gramnegatius presenten alteracions estructurals del LPS que el permeten passar
desapercebuts pels TLR4.
També hi ha bacteris que han desenvolupat mecanismes d’inhibició del sistema del
complement. El bacteri Salmonella typhimurium, per exemple, expressa un gen RcK que
inhibeix la formació del MAC, impedint la lisi mediada per complement. Però hi ha
mecanismes per evitar tots els passos del sistema del complement. Pot ser que la paret
cel·lular o la membrana externa amb càpsula o càlix impedeixi la unió dels components del
complement, pot ser que les molècules del bacteri dificultin la interacció de molècules del
sistema del complement amb els seus receptors, pot ser que el bacteri secreti inhibidors o
proteïnes que impedeixen inserció a membrana i la formació del MAC i pot ser que els bacteris
produeixin enzims que trenquin, inactivin o desacoblin els components del complement. Tots
aquests mecanismes diversos tendeixen a inhibir el sistema del complement sobre els
bacteris extracel·lulars.
Resposta a Bacteris Intracel·lulars. Pel que fa als bacteris intracel·lulars, s’entén que són inaccessibles per les molècules
efectores solubles un cop han entrat a la cèl·lula. Davant aquests patògens, els anticossos i
el sistema del complement són poc eficients. Així i tot, poden actuar en els moment previs a
la infecció cel·lular.
Els bacteris intracel·lulars es poden eliminar més fàcilment si són presentats a limfòcits T i
aquests són els que contribueixen a la seva eliminació. Els bacteris intracel·lulars viuen dins

Immunologia 2020
CEBIO 129
la cèl·lula i, en cas que mantinguin un estat latent, poden passar desapercebuts pel sistema
immunitari. El sistema només els detectarà quan causin la mort de la cèl·lula hoste.
Els bacteris intracel·lulars poden ser com Chlamydia, Rickettsia o Listeria i trobar-se al
citoplasma o poden ser com Mycobacteria, Salmonella, Legionella i Leishmania i trobar-se en
vesícules citoplasmàtiques. La possibilitat de resposta, detecció i escapada del sistema
immunitari també depèn d’aquesta localització intracel·lular.
Resposta innata a bacteris intracel·lulars.
En quant a la resposta innata, tan sols pot tenir efecte prèviament a l’entrada a la cèl·lula
hoste. S’aconsegueix per fagocitosi del patogen i per l’activació de l’esclat respiratori en els
macròfags infectats. Una vegada aquest bacteri ja és a l’interior de la cèl·lula, el mecanisme
principal és l’acció de les cèl·lules NK. En general, la resposta innata a bacteris intracel·lulars
és poc efectiva i és molt rellevant l’activitat de la resposta adaptativa.
Resposta innata a bacteris intracel·lulars per cèl·lules NK.
L’element principal quan el bacteri ja es troba dins la cèl·lula són les cèl·lules NK. Les cèl·lules
NK detecten la presència de la infecció i poden alliberar els seus grànuls, plens de perforines
i granzines que acabaran provocant la lisi de la cèl·lula infectada. Les cèl·lules NK tenen un
nivell baix de CD56 però que tenen molt actius els grànuls citoplasmàtics amb productes
microbicides.
D’altra banda, les cèl·lules NK també secreten citocines com l’interferó γ. L’IFNγ activa la
capacitat bactericida de macròfags. Els macròfags faran IL12, que activa les cèl·lules NK i les
fa més eficients. La IL12 i l’interferó γ fan que la resposta dels limfòcits TCD4 sigui del tipus
Th1. La resposta Th1 produeix IFNγ i manté el cicle d’activació, detecció i eliminació de
bacteris intracel·lulars. La contribució més importants és de les cèl·lules NK, que tenen un
nivell de CD56 baix i que actuen per producció d’enzims i citocines.
Les cèl·lules NK són imprescindibles en el primer control de la infecció per bacteris
intracel·lulars perquè contribueixen a l’eliminació dels bacteris en els primers dies de la
infecció. Sovint, aquests mecanismes no són suficients i els bacteris es queden dins dels
macròfags de mode latent. Això provoca infeccions cròniques que passen desapercebudes.
Resposta adaptativa a bacteris intracel·lulars per limfòcits TCD4 i CD8.
En el cas de la resposta a bacteris intracel·lulars, el principal mecanisme de la resposta
adaptativa és la resposta cel·lular mediada per limfòcits TCD4 i CD8. Un cop els bacteris han
entrat a la cèl·lula cal destruir la cèl·lula infectada per eliminar-los. En aquest procés
intervenen els limfòcits TCD4 helper 1 ja que secreten IFNgamma, el qual activa la capacitat
lítica dels macròfags i estimula la producció d’immunoglobulina G per part dels limfòcits B.
L’interferó promou aquest canvi d’isotip i, per tant contribueix a la producció d’aquests
anticossos. Les IgG són importants en facilitar l’eliminació dels bacteris intracel·lulars i en
activar el sistema del complement per augmentar l’opsonització dels bacteris intracel·lulars
mentre encara no hagin entrat a l’interior de la cèl·lula hoste.
D’altra banda, si els bacteris intracel·lulars s’escapen del fagosoma i entren a la cèl·lula hoste
al evitar la maquinària proteolítica del macròfag, es poden processar i presentar proteïnes del
citoplasma per part de molècules del MHC de classe I. Els limfòcits TCD8, amb capacitat
citotòxica, es poden activar i contribuir a l’eliminació dels bacteris intracel·lulars. Sense
l’actuació dels limfòcits TCD8, els bacteris intracel·lulars podrien donar lloc a infeccions
cròniques al mantenir-se fora de l’abast de molts dels mecanismes del sistema immunitari per
detectar i eliminar patògens, cosa que passa sovint.

Immunologia 2020
CEBIO 130
La col·laboració dels limfòcits TCD4 és important per tal d’aconseguir una bona activació i
funció dels limfòcits TCD8. Els limfòcits TCD4 ajuden a eliminar els patògens dels fagosomes,
mentre que els limfòcits TCD8 eliminen els patògens que han escapat al citoplasma. Per tant,
els dos tipus de limfòcits contribueixen en la resposta a bacteris intracel·lulars.
Resposta adaptativa a bacteris intracel·lulars per limfòcits Th17.
Els limfòcits T helper 17 sovint és prescindible. En el cas d’alguns patògens com
Micobacterium tuberculosis, es produeix l’activació dels macròfags via IL12. Aquesta activació
ja és suficient per activar una resposta de tipus Th1 i no fan falta els limfòcits del tipus Th17.
Davant altres patògens com Francisquella tularensis o Chlamydia muridarum, no s’indueix la
producció d’IL12 per part de les cèl·lules dendrítiques. En aquests casos és important la
col·laboració dels limfòcits Th17, que faran que les cèl·lules dendrítiques produeixin IL12. Així
s’afavoreix la diferenciació cap a limfòcits Th1, que produiran IFNγ i contribuiran a la seva
activitat bactericida.
Les cèl·lules Th17 també contribueixen al reclutament de neutròfils per la destrucció de
bacteris com Listeria monocytogenes i Salmonella typhimurium. D’altra manera, es podrien
escapar de l’acció dels macròfags.
Conseqüències Patològiques de la resposta a bacteris intracel·lulars. També hi pot haver conseqüències patològiques com a resultat de la resposta a bacteris
intracel·lulars. Es parla de l’aparició de reaccions d’hipersensibilitat i d’altres manifestacions
que depenen de factors genètics dels individus.
La resposta a bacteris intracel·lulars, en tant que es basa en la destrucció de cèl·lules
infectades, pot tenir conseqüències fisiològiques si es destrueixen aquelles cèl·lules amb una
funció en el teixit.
Moltes de les infeccions per bacteris intracel·lulars es cronifiquen i sovint generen
granulomes. S’han vist granulomes a les vies respiratòries en casos d’infecció per
Mycobacterium tuberculosis. Els granulomes són estructures força esfèriques que estan
formades per una capa interna de macròfags infectats i que al voltant tenen limfòcits TCD4
Th1 que intenten contribuir a l’activitat bactericida d’aquests macròfags. A la part més externa
hi ha tant limfòcits TCD4 com limfòcits TCD8.
L’objectiu de la formació de granulomes és intentar eliminar el bacteri intracel·lular i evitar la
seva disseminació ja que els bacteris queden atrapats al seu interior. Si el granuloma es
manté de manera estable, no hi ha conseqüències patològiques i la infecció queda
continguda. Si el granuloma es descomposa, la infecció es reactiva i es disseminen els
patògens fàcilment.
El granuloma és una conseqüència patològica perquè ocupa espai d’un òrgan i alhora és un
mecanisme per mantenir la disseminació.
Mecanismes d’evasió dels bacteris intracel·lulars. Els bacteris intracel·lulars han desenvolupat tota una sèrie de mecanismes d’evasió molt
diversos. Mycobacterium tuberculosis evita la fusió entre el fagosoma i el lisosoma. Això
impedeix que proteïnes del Mycobacterium arribin a la via endocítica, siguin processats,
carregats a molècules del MHC de classe II i es presentin al sistema immunitari.
Legionella monocytogenes escapa del lisosoma al citoplasma. Per tant, s’escapa dels
mecanismes intrínsecs de la via endocítica.

Immunologia 2020
CEBIO 131
Salmonella typhimurium expressa el gen Rck que confereix resistència a la lisi mediada per
complement, prevenint la formació del MAC.
Mycobacterium Leprae és capaç d’inhibir la formació d’espècies reactives de l’oxigen i del
nitrogen.
Evitar l’activitat microbicida com a mecanisme d’evasió dels bacteris intracel·lulars.
El conjunt d’espècies bacterianes intracel·lulars han desenvolupat mecanismes d’evasió de
l’activitat microbicida de la cèl·lula hoste. Aquests mecanismes inclouen des de la modificació
de la superfície de la cèl·lula hoste per resistir o trencar els pèptids antimicrobians, l’expressió
d’enzims com la catalasa que converteixen espècies reatives d’oxigen en components inocus
o fins i tot impedir la producció d’enzims que generen les espècies reactives d’oxigen i de
nitrogen.

Immunologia 2020
CEBIO 132
Tema 24 – Resposta a Virus.
Els virus. Els virus són paràsits intracel·lulars estrictes, pel que viuen a l’interior de les cèl·lules que
infecten i utilitzen la seva maquinària per a la síntesi de proteïnes. Quan es troben fora de la
cèl·lula, els virus són metabòlicament inerts. Usen la maquinària de síntesi de proteïnes
de la cèl·lula infectada per a la seva replicació, per tant depenen de la cèl·lula hoste per a la
seva replicació. Així doncs, les cèl·lules infectades expressen proteïnes el virus a la seva
membrana. Aquestes cèl·lules reben el nom d’hoste.
Els virus estan formats per material genètic envoltat per una càpsida proteica. Aquesta
estructura té un grau de complexitat variable, doncs difereixen en el tipus de material genètic
que poden tenir. Aquests àcids nucleics poden ser DNA o RNA, de cadena simple o de doble
cadena i poden ser lineals o circulars. A més, els virus difereixen en si disposen d’una envolta
lipídica al voltant de la càpsida proteica.
Tot això els hi dona una elevada complexitat afegida. Per altra banda, els virus tenen una
elevada freqüència de mutacions que dificulten la seva detecció i eliminació. Van canviant
més ràpid que l’adaptació del sistema immunitari. A més, els virus presenten una gran
diversitat de mecanismes d’evasió del sistema immunitari. Entre aquesta diversitat en
destaca la capacitat “d’amagar-se” a l’interior de les cèl·lules. Quan exerceixen aquesta
capacitat, es diu que els virus estan en la seva forma latent dins les cèl·lules. La forma
latent és invisible i reactiva la infecció en determinades condicions.
En general, els virus causen infeccions agudes que activen el sistema immunitari i, per
tant, l’hoste aconsegueix eliminar-los. Però hi ha certs tipus de virus que tenen la capacitat
de ser latents, pel que la infecció pot persistir de forma indefinida.
Rutes d’entrada i infecció de virus humans. Les infeccions de major incidència són les de les vies respiratòries i gastrointestinals.
L’entrada dels virus té lloc, sobretot, a través de les superfícies mucoses, lesions a la pell o
directament via sanguínia. Les superfícies mucoses susceptibles de ser la porta d’entrada de
virus són les mucoses respiratòria, intestinal, genital o ocular. L’entrada de virus per via
sanguina no es dona pas massa sovint. La via d’entrada, sobretot a traves de les superfícies
de les mucoses, determina les característiques del tipus de resposta.
Família Vírica Virus Malaltia
Herpesviridae
Herpes Simplex virus I Herpes labial
Virus Epstein - Barr Mononucleosi infecciosa
Limfoma de Burkitt
Varicella zòster virus Grans i pústules
Poxviridae Variola Virus Verola
Hepadnaviridae Hepatitis B virus Hepatitis
Papoviridae Papillomavirus Berruga
Orthomyxoviridae Influenza virus Malalties respiratòries
Togaviridae Rubivirus Rubèola
Paramyxoviridae
Mumps virus Paperes
Morbillivirus Xarampió
Respiratory Syncytal Virus Malalties Respiratòries
Rhabdoviridae Lyssavirus Ràbia
Retroviridae VIH SIDA
Coronaviridae SARS-CoV SARS

Immunologia 2020
CEBIO 133
Tropisme cel·lular.
Els virus presenten tropisme cel·lular. El tropisme cel·lular és el moviment d’orientació cap
a cèl·lules concretes. Dit d’una altra manera, els virus no infecten totes les cèl·lules del cos,
tan sols infecten aquelles cèl·lules que presenten el receptor que pot utilitzar el virus per entrar
a l’interior de la cèl·lula. En conseqüència, la infecció per virus tindrà una incidència o una
altre depenent del tipus de receptor que utilitzi per entrar a les cèl·lules.
El virus interacciona amb proteïnes de membrana de la cèl·lula hoste i hi introdueix el
seu material genètic. Els virus utilitzen receptors específics de cada tipus cel·lular i per això
cada espècie vírica infecta tipus cel·lulars concrets. Això no implica, però, que tan sols infectin
aquell tipus cel·lular. Els virus poden tenir tropisme cel·lular per diversos tipus de
cèl·lules. Els virus infecten una gran diversitat de poblacions cel·lulars.
Virus Receptor Cèl·lula
Poliovirus CD155
Adenovirus subfamília B2,
Virus xarampió, herpes virus 6
CD46 (Complement regulatory protein)
Rhinovirus CD54 (ICAM-1) Cèl·lules endotelials i epiteli
inflamat
Rhinovirus 2 humà Low-density lipoprotein
receptor (LDLR)
Flebovirus DC-SIGN
SARS coronavirus Liver-SIGN (L-SIGN)
Enterovirus 71 Scavenger receptor-B2 (SR-
B2)
Enterovirus 71 P-selectin glycoprotein
ligand-1 (PSGL-1)
HIV CD4 (CCR5 / CXCR4) TCD4+ i macròfags
Hepatitis C CD81 (SR-B1, claudin-1,
and occludin) Hepatòcits
Epstein-Barr CD21 (CR2, Complement
receptor) Limfòcits B
Ràbia Receptor acetilcolina Neurones
Les respostes immunitàries vindran determinades pel receptor que els virus facin servir, doncs
segons si infecten un tipus cel·lular molt específic o les cèl·lules de forma mes genèrica,
la resposta serà més o menys intensa. Un exemple de variabilitat en la infecció és el que
passa amb Flebovirus, el Coronavirus o el VIH, que infecten cèl·lules dendrítiques per mitjà
del DC-SIGN.
Receptors de lectines tipus C.
En el cas del DC-SIGN, un receptor de lectina de tipus C que reconeix carbohidrats, és usat
tant per Citomegalovirus (CMV), VIH i el virus del Dengue per tal d’infectar cèl·lules
dendrítiques. Gràcies al transport d’aquestes cèl·lules dendrítiques, els virus poden viatjar
dels teixits als òrgans limfoides secundaris, facilitant la disseminació del virus.
El receptor DC-SIGN es troba expressat a totes les cèl·lules dendrítiques que participen en
la resposta innata. En funció del tipus de cèl·lula, el receptor rep un nom diferent. Per les
cèl·lules dendrítiques c, el receptor és el DC-SIGN. En les cèl·lules dendrítiques
plasmacitoides, el receptor és el BCDA-2. Finalment, les cèl·lules de Langerhans tenen el
receptor anomenat langerina o CD207.

Immunologia 2020
CEBIO 134
La capacitat mòbil de les cèl·lules dendrítiques a través dels òrgans limfoides secundaris fa
que aquests receptors facilitin la disseminació dels virus. El cas més ben estudiat és del HIV,
que usa DC-SIGN per disseminar-se a través dels òrgans limfoides secundaris on pot infectar
els limfòcits TCD4.
Cicle vital dels virus. El cicle vital dels virus consta de 4 fases:
La maquinària de síntesi de proteïnes de la cèl·lula infectada pel virus és bàsicament
emprada per a la replicació del virus i no per a la producció de les proteïnes pròpies de la
cèl·lula. Això es deu a què el virus segresta tota la maquinària cel·lular. Aquestes proteïnes
víriques es poden expressar a la membrana i, per tant, la composició de proteïnes de
membrana en la cèl·lula infectada canviarà. Aquest és un element detectat per les cèl·lules
del sistema immunitari.
Detecció dels virus. Els principals mecanismes que la immunitat innata fa servir per detectar la presència de virus
en l’organisme són els receptors de tipus TOLL, els de tipus NOD i els de tipus RLR. Això és
perquè tots aquests receptors tenen els àcids nucleics en les seves diferents formes com a
principal lligand.
Entrada a la cèl·lula hoste per interacció amb
receptors específics de membrana.
Fusió amb la membrana de la
cèl·lula i alliberació dels àcids nucleics.
Ús de la maquinària
replicativa de l’hoste, per
sintetitzar les proteïnes
estructurals víriques.
Acoblament de genomes i proteïnes
estructurals formant les
partícules víriques que s’alliberen.

Immunologia 2020
CEBIO 135
Receptors de Tipus Toll.
Però no tota la defensa es basa en la detecció d’àcids nucleics. Alguns receptors de tipus
TOLL són capaços de detectar la presencia de la hemaglutinina o de l’envolta de determinats
virus. Aquesta detecció permet senyalitzar conforme s’ha detectat la presència d’un virus. Els
dos tipus principals de virus són el TLR2, TLR3, TLR4, TLR7, TLR8 i TLR9. Cada receptor de
tipus TOLL detecta coses diferents:
Les principals famílies víriques amb DNA de doble cadena són els Vesicular Stomatitis Virus
(VSV), el Lymphocytic Choriomeningitis Virus (LCMV), el Herpes Simplex Virus (HSV) i el
Virus Epstein-Barr (EBV).
Les principals famílies víriques amb RNA de cadena senzilla són els virus Influenza, VIH i
VSV.
El HSV és un dels virus amb el DNA de doble cadena amb les illes CpG desmetilades. Aquest
és un factor important ja que el TLR9 pot detectar el genoma del virus tant quan és alliberat
com quan es produeix la replicació.
D’aquests TLR, els més importants en la resposta a virus són el TLR7, TLR8 i TLR9 (més
importants que TLR3) perquè són els responsables de la detecció dels virus per part de les
cèl·lules dendrítiques plasmocitoides i per tant de l’elevada producció d’IFNs tipus I (els
interferons de tipus I són un dels mecanismes inicials en la resposta als virus).
Receptors de Tipus R.
D’entre els diferents receptors de tipus R en destaquen dos: RiG1 I MDA5. Cadascun detecta
diferents elements.
• RIG-1 detecta els virus
o Influenza.
o Hepatitis C.
o Vesicular Stomatitis.
o Encefalitis japonesa.
o Measles.
o Human Respiratory Syncytial.
• MDA-5 pot reconèixer
o Poly(I:C).
o Measles virus.
o Sendai.
o VSV: vesicular estomatitis virus.
TLR2 detecta MV hemagglutinin protein, HSV i
HCMV.
TLR3 detecta el DNA de doble
cadena.
TLR4 detecta MMTV envelope
protein i RSV.
TLR7 i TLR8 detecten RNA de cadena senzilla.
TLR9 detecta DNA de doble cadena amb les illes CpG
desmetilades.

Immunologia 2020
CEBIO 136
o RNA víric genòmic o generat durant la replicació.
El resultat de la interacció entre el receptor RLR i el seu lligand és sempre la inducció de la
producció d’interferó de tipus I.
Receptors NOD.
També hi ha tan sols dos tipus de receptors de tipus NOD que participin en la resposta a
virus. La interacció dels virus amb NOD2 acaba produint la senyalització amb la transcripció
de factors de transcripció IRF i, per tant, la producció d’interferons IFN-α i de IFN-β.
Quan la interacció és amb NLRP3, s’activa la formació del complex multi-proteic anomenat
inflamasoma. L’inflamasoma porta a l’activació de la caspasa 1, que farà de mitjancera en
l’activació proteolítica de la IL-1b emmagatzemada.
• NOD2 reconeix
o Influenza virus.
o Vesicular Stomatitis Virus.
o Human Respiratory Syncytial Virus
• NLRP3 reconeix els virus
o Influenza.
o Sendai.
o Adenovirus.
o Vaccinia virus (VACV).
Cèl·lules Dendrítiques Plasmacitoides. Les Cèl·lules Dendrítiques Plasmacitoides són importants en la resposta al virus per la seva
rapida i elevada producció d’interferons de tipus I.
Això és degut que aquestes cèl·lules dendrítiques plasmacitoides expressen de manera
constitutiva el factor de transcripció IRF-7, reclutat per MyD88 a través d’un complex
molecular que inclou TRAF6 i IRAK4. MyD88 és una molècula adaptadora que s’uneix a la
cua citoplasmàtica de TLR7, TLR8 i TLR9 quan aquests interaccionen amb el seu lligand. La
unió dels TLR al seu lligand i el reclutament de IRF-7 inicia una cascada de senyalització que
ràpidament indueix la transcripció en grans quantitats dels gens d’IFN i (interferó α i
β).
Aquestes cèl·lules dendrítiques plasmacitoides són diferents de les cèl·lules dendrítiques
convencionals, ja que la majoria de tipus cel·lulars requereixen de l’upregulació d’IRF-7 en
resposta a IFN tipus I per poder secretar IFN. És a dir, la unió de TLR7 i TLR9 als seus
lligands indueix la secreció de citocines proinflamatòries i quimiocines però poc IFN (si que
s’expressa però a molt baixa concentració).
Per tant, es necessita que es produeixi un augment de l’expressió de IRF-7 per a que es pugui
produir la resposta d’interferons tipus I per a secretar IFN i molt important per a la resposta
del virus (sobretot en les fases inicials). Aquesta necessitat de que hi hagi algun altre senyal
que indueixi la producció de IRF-7 abans de que passi res mes fa que les cèl·lules
dendrítiques plasmacitoides siguin molt eficients en aquesta funció.

Immunologia 2020
CEBIO 137
Resposta Innata.
Interferons.
La resposta innata principal als virus és la producció d’interferons, sobretot de tipus I però
també de tipus 2 (tipus ). Els interferons tenen un efecte indirecte sobre les cèl·lules
infectades ja que actuen sobre altres tipus cel·lulars.
Els interferons α, β i γ activen les cèl·lules NK i, per tant, les fan més eficients en matar
cèl·lules. A més, l’interferó γ activa els macròfags i les cèl·lules dendrítiques, pel que
augmenten la seva capacitat microbicida (macròfags) i l’expressió de molècules MCH de
classe I i de classe II, importants per a induir una bona resposta adaptativa als virus.
Així i tot, els interferons també tenen un efecte directe sobre les cèl·lules infectades i sobre
les cèl·lules del seu voltant. Aquests interferons, quan s’uneixen al receptor de les cèl·lules,
indueixen l’expressió de tota una sèrie de gens anomenats ISG (gens estimulats per
l’interferó).
Els gens ISG són essencials per a la generació de l’estat antivíric. És a dir, eliminen els virus
de les cèl·lules infectades i protegeixen les cèl·lules del seu entorn per a evitar que, si hi ha
una infecció, aquesta sigui potent i que acabi amb la destrucció de les cèl·lules. L’estat
antivíric consisteix en un estat d’inhibició de la síntesi de proteïnes i de la replicació del virus
arran de l’expressió dels gens estimulats per l’interferó.
La manera com es produeix aquest efecte directe és perquè la interacció dels interferons amb
el seu receptor desencadena la transcripció i expressió de tota una sèrie de proteïnes
(codificades pels gens estimulats per interferó). Destaquen quatre proteïnes.
La 2-5 oligoadenilat sintasa (OAS) és un enzim que activa la ribonucleasa (RNAsa L). És
essencial, ja que talla el mRNA víric i de la cèl·lula eucariota. Malgrat sembli una decisió
irresponsable, el sistema immunitari considera que, en aquest moment de la vida de la
cèl·lula, és més important poder tallar el mRNA víric. Per tant, impedeix la producció de
proteïnes del virus.
La Proteïna quinasa depenent de dsRNA (PKR) fosforila la proteïna elF2, inhibint l’inici de la
traducció a la síntesi proteica.
L’Adenosinadesaminasa (ADAR) edita l’RNA introduint-hi mutacions, el que genera RNA no
funcional.
Les Proteïnes Mx amb activitat GTPasa interaccionen amb les proteïnes de la càpsida del
virus interferint en el seu transport o alterant les seves propietats. Per tant, impedeixen que
les proteïnes de la càpsida del virus arribin a poder-se estructurar.
Per tant, l’efecte directe de l’interferó es pot resumir com que inhibeix la producció de
proteïnes i degrada el mRNA. Com no hi ha síntesi de proteïnes víriques, no es pot produir
la replicació del virus.
Cèl·lules Dendrítiques Plasmacitoides.
Els interferons de tipus I, a part d’induir l’estat antivíric, estan associats a la interleuquina 12
i indueixen la secreció d’interferons γ fruit de l’estimulació de les cèl·lules NK. Per tant, els
interferons també faciliten l’activació dels limfòcits TCD8 perquè promouen la diferenciació
cap a cèl·lules TCD4 Th1. Es pot dir, doncs, que promouen l’eliminació del virus.
Els IFN-I, juntament amb la interleuquina 6, promouen la diferenciació de limfòcits B a
cèl·lules plasmàtiques. El que afavoreix la producció d’anticossos específics contra el virus.

Immunologia 2020
CEBIO 138
Les cèl·lules plasmacitoides també produeixen quimosines com CCL3 i citocines
proinflamatòries que permeten reclutar leucòcits i per tant, cèl·lules que podran lluitar contra
la infecció vírica.
Les cèl·lules dendrítiques plasmacitoides no són bones presentadores d’antigen perquè són
molt poc eficients en l’activació dels limfòcits T naive de la resposta adaptativa en comparació
amb les cèl·lules dendrítiques convencionals. Aquesta menor eficiència es dona al nivell de
la fagocitosi, el processament i la càrrega dels pèptids a les molècules del MHC.
Els nivells d’expressió de molècules del MHC de classe II i de molècules coestimuladores en
aquestes cèl·lules plasmacitoides és molt més inferior al de les cèl·lules dendrítiques
convencionals. No s’ha pogut demostrar que puguin activar limfòcits T naïve però sí poden
activar i fer proliferar TH1 i TCD8+ de memòria. Per tant, participen sobretot en la resposta
primària als virus com a cèl·lules detectores del virus.
A les cèl·lules plasmacitoides activades per virus s’augmenta l’expressió de MHC-II i de
molècules coestimuladores (CD80, CD86, CD40) , pel que s’augmenta el seu potencial com
a cèl·lules presentadores d’antigen.
Fases de la infecció vírica i la resposta immunològica.
La magnitud d’una resposta a un virus des del contacte amb aquest fins que es resol la
infecció varia en components i intensitat segons el temps que ha passat des de la infecció.
Els primers elements que es produeixen són els interferons de tipus I, la Interleuquina 12 i
l’activació de les cèl·lules NK.
Els interferons de tipus I, juntament amb la interleuquina 12, provocarà l’activació dels
limfòcits Th1 i la conseqüent resposta adaptativa als virus. Per altra banda, les cèl·lules NK
controlen la infecció vírica durant els primers dies de la resposta. Així i tot, cal la participació
de la resposta adaptativa per poder erradicar completament la infecció.
Citotoxicitat per cèl·lules NK.
Les cèl·lules NK tenen un paper molt important en la resposta als virus. A través de l’equilibri
o la manca d’equilibri entre la senyalització dels seus receptors activadors i els inhibidors, es
determina si maten la cèl·lula diana.
A través d’aquests receptors, les cèl·lules NK reconeixen canvis en les cèl·lules infectades.
Aquests canvis poden ser:
• Modificacions en el patró de glicosilació de proteïnes de membrana de la cèl·lula
infectada.
• Alteracions en els nivells d’expressió de molècules del MHC de classe I a la cèl·lula
infectada.
• Canvis qualitatius en les molècules del MHC de classe I.
• Detecció de l’expressió de molècules induïdes per l’estrès o de lligands dels receptors
activadors induïts pel virus.
Es consideren canvis qualitatius en les molècules del MHC de classe I els canvis en els
pèptids carregats en el MHC-I. Arran d’aquests canvis en la unió, es pot impedir la interacció
de les molècules MHC-I amb els receptors inhibidors i provocar la lisi de les cèl·lules. Són
molècules induïdes per l’estrès les MICA, les MICB i les ILBP.
Les cèl·lules NK actuen alliberant el contingut dels seus grànuls, plens de perforina i
granzima, sobre la cèl·lula infectada. En rebre aquests enzims, la cèl·lula infectada veurà
induïda la seva apoptosi.

Immunologia 2020
CEBIO 139
Les cèl·lules NK presenten receptors activadors amb lligands diferents del MHC. El principal
és CD16, altrament dit FcγRIII. Són receptors d’elevada afinitat de la regió contant de les
immunoglobulines G, sobretot de IgG1 i IgG3 humanes. Aquest receptor es troba associat a
motius ITAM, el que els atorga la definició de receptors activadors. La interacció de la IgG
específica per una proteïna vírica amb el receptor FcγRIII provoca la desgranulació de les
cèl·lules NK sobre aquelles cèl·lules infectades que expressin les proteïnes víriques a la seva
membrana. En conseqüència, es pot considerar que FcγRIII és responsable de la citotoxicitat
cel·lular depenent d’anticòs.
Les cèl·lules NK també produeixen citocines, sobretot interferó γ. Les citocines secretades
tenen les funcions de:
• Promoure la diferenciació dels limfòcits TCD4 cap a Th1.
• Activar, fer créixer i diferenciar les cèl·lules T, B, NK i macròfags.
• Augmentar l’expressió de molècules MHC a les cèl·lules presentadores d’antigen.
• Determinar l’efecte anti-víric directe.
Les cèl·lules NK també sintetitzen TNF-α, el qual:
• Actua com a mediador paracrí i endocrí d’inflamació.
• Regula el creixement i la diferenciació de diferents tipus cel·lulars.
• Mostra citotoxicitat selectiva per a cèl·lules transformades.
Finalment, les cèl·lules NK sintetitzen factors de creixement com GM-CSF. Els factors de
creixement serveixen per:
• Assegurar la supervivència i creixement de les cèl·lules mares hematopoètiques.
• Diferenciar i activar granulòcits i monòcits.
• Induir al creixement de les cèl·lules endotelials, eritròcits, megacariòcits i limfòcits T.
Macròfags i sistema del complement.
Els macròfags també intervenen en la resposta innata als virus, però en aquest cas son molt
mes rellevants les cèl·lules NK. Els macròfags fan les funcions de:
• Destrucció de les partícules víriques opsonitzades.
• Fagocitosi de virus i de cèl·lules infectades per virus.
• Producció de molècules antivíriques.
• Presentació d’antígens vírics als limfòcits T.
Les molècules antivíriques són interferons de tipus I amb efecte antivíric o que produeixen
espècies reactives d’oxigen i nitrogen. Els principals tipus d’interferons són IFN-α, TNF-α i
NO.
En el cas del sistema del complement, la seva actuació no és massa rellevant de forma directe
ja que els virus són patògens intracel·lulars. Així i tot, el sistema del complement pot contribuir
a:
• Augmentar la neutralització de partícules víriques amb anticossos.
• Augmentar la fagocitosi de partícules víriques fent opsonines.
• Provocar la lisi directa de partícules víriques amb envolta lipídica.
• Provocar la lisi de cèl·lules infectades.
Per tal de provocar les lisis, el sistema del complement ho aconsegueix per mitjà de la
formació del complex d’atac a la membrana.

Immunologia 2020
CEBIO 140
Resposta adaptativa humoral. La resposta adaptativa humoral és poc eficient i normalment no elimina la infecció. És
important, però, durant l’etapa extracel·lular d’aquest. En aquest cas, l’objectiu principal és la
neutralització del virus, és a dir, impedir la infecció aprofitant anticossos. Per aconseguir-ho,
els anticossos poden actuar a 5 nivells.
Els anticossos impedeixen l’entrada del virus a la cèl·lula. Això es pot fer impedint la interacció
del virus amb la cèl·lula hoste unint-se a les proteïnes de la càpsida o de l’envolta vírica que
interaccionen amb el receptor a la membrana de la cèl·lula. Una altra opció és impedir la fusió
de l’envolta vírica amb la membrana de la cèl·lula bloquejant els epítops implicats. A les
mucoses, els principals anticossos antivírics són IgA.
Els anticossos també poden contribuir a activar la via clàssica del sistema del complements
i, per tant, provocar la lisi directa dels virus amb l’envolta lipídica o de les cèl·lules infectades.
També poden participar en la citotoxicitat mediada per anticossos, ja sigui a través de les
cèl·lules NK com per altres cèl·lules efectores. Un tipus de cèl·lules efectores que es poden
fer servir són els neutròfils, doncs també tenen reactors Fc. Així i tot, aquests receptors són
menys eficients.
Els anticossos faciliten la fagocitosi del virus opsonitzant-lo. També poden impedir la infecció
induint la secreció de citocines, les quals estimularan altres cèl·lules del sistema immunitari.
Resposta adaptativa cel·lular. La resposta adaptativa més important als virus és la resposta cel·lular. Aquesta és necessària
per inhibir la disseminació dels virus a les cèl·lules infectades i és essencial per eliminar el
virus una vegada s’ha produït la infecció.
El principal component de la resposta cel·lular als virus són els limfòcits TCD8, altrament
anomenats limfòcits T citotòxics. Aquests limfòcits acabaran eliminant aquelles cèl·lules
infectades. Per tal que hi hagi una bona resposta a limfòcits TCD8, és important que hi hagi
una bona activació dels limfòcits TCD4 i del tipus efector Th1. Aquestes cèl·lules Th1
contribueixen a l’activació dels limfòcits TCD8.
Els limfòcits TCD8 detecten la presència d’infeccions en cèl·lules perquè aquestes presenten
molècules del complex MHC de classe I carregades amb pèptids derivats del virus. Aquests
complexos MHC-pèptid són detectats pels limfòcits TCD8, que alliberaran la perforina i
gramzina que mataran les cèl·lules infectades. Els limfòcits citotòxics lisen, específicament,
la cèl·lula infectada tot induint-li l’apoptosi. Paral·lelament a la perforina i a la gramzina, els
limfòcits T citotòxics també poden fer servir les molècules Fas i FasL. Fas és una molècula
que es troba en totes les cèl·lules, mentre que FasL és el lligand de Fas i es troba en els
limfòcits TCD8.
L’activitat dels limfòcits T citotòxics contribueix a eliminar la possibilitat de la replicació vírica.
Els interferons de tipus I s’uneixen al receptor de la cèl·lula infectada i li indueixen un augment
de l’expressió de les molècules del MHC de classe I. Gràcies a això, es pot considerar que
els IFN-I faciliten la detecció i lisi per part dels limfòcits T citotòxics.
Si es tracta d’un virus que infecta cèl·lules dendrítiques, aquestes poden presentar els
antígens vírics units a les molècules MHC-I. En el cas de que sigui un virus que no presenta
tropisme cel·lular per les cèl·lules dendrítiques, aquestes poden internalitzar els antígens
vírics per fagocitosi dels virus o cèl·lules infectades, endocitosi o macropinocitosi,

Immunologia 2020
CEBIO 141
processar-los i presentar-los units a molècules MHC-I a limfòcits TCD8 per
crospresentació.
La mateixa cèl·lula presentarà antígens vírics internalitzats associats a MHC-II per a ser
presentats als limfòcits T CD4+, que tendiran a donar lloc a una resposta TH1,
imprescindible per a l’activació de limfòcits T CD8+.
Limfòcits TCD4 i Th1. Els limfòcits T helper 1 contribueixen a l’activació dels limfòcits TCD8 perquè són
imprescindibles per dur a terme l’activació de la cèl·lula presentadora d’antigen i per contribuir
a l’activació dels limfòcits TCD8 a través de la interacció entre CD40 i CD40L. CD40L és el
lligand de CD40 i s’expressa als limfòcits Th1. Els limfòcits Th1 també contribueixen a
l’activació de les cèl·lules TCD8 en tant que són productors de citocines.
Els limfòcits Th1 produeixen interleuquina 2, que contribueix a l’activació i proliferació dels
limfòcits TCD8 i a l’activació de les cèl·lules NK. També produeixen Interferó γ, que indueix
l’estat antivíric i activa tant les cèl·lules NK com els macròfags. Finalment, la tercera citocina
secretada pels limfòcits Th1 és el TNF-α, que recluta i activa macròfags.
Els limfòcits TCD4 helper 1 contribueixen de forma activa en què hi hagi una bona resposta
per part dels limfòcits TCD8. A més, la interacció entre Fas i FasL permet eliminar directament
les cèl·lules infectades. Fas és presentat per les cèl·lules infectades, mentre que FasL és
presentat pels limfòcits Th1.
Les cèl·lules T CD4+ són importants per a aconseguir una bona activació dels limfòcits B,
la qual dona lloc a una resposta humoral duradora i permet el canvi d’isotip. Per tant, la
producció de citocines per part de les cèl·lules T helper fol·liculars determinarà aquest canvi
d’isotip i la producció d’anticossos neutralitzats. També es produirà la maduració de
l’afinitat i la generació d’una bona de resposta i memòria a aquests virus.
El canvi d’isotip és molt important. Per exemple, l’interferó és important per a produir el canvi
d’isotip a IgG2A, que és una de les immunoglobulines que participa més activament en la
inducció de citotoxicitat cel·lular depenent d’anticòs en cèl·lules NK. Per tant, és un
mecanisme més per a la destrucció d’aquests virus.
Resposta immunològica als virus. La resposta als virus té com a objectiu impedir la infecció. Això es pot intentar aconseguir
mitjançant la producció d’IFN-I, que genera l’estat antiviral, i mitjançant la producció
d’anticossos neutralitzants, que impedeixen l’entrada dels virus a les cèl·lules.
Una vegada els virus han infectat les cèl·lules, els mecanismes de la immunitat innata i de la
adaptativa procuren l’eliminació de les cèl·lules infectades mitjançant les cèl·lules NK o els
limfòcits T CD8+ que les eliminen específicament.
Mecanismes d’evasió. Els virus, igual que els bacteris, han desenvolupat molts mecanismes per evitar l’acció del
sistema immunitari.
Evitar la senyalització per mitjà dels PRR.
Molts virus generen una sèrie de productes o proteïnes destinades a evitar la senyalització
mitjançant els PRR. Els passos que concretament bloquegen pot ser un o varis dels següents:
• La unió dels elements de la via de senyalització a l’adaptador MyD88.
• La interacció i producció de IRAK 2.

Immunologia 2020
CEBIO 142
• La unió dels adaptadors TRIF.
• La interacció dels TLR i els seus lligands.
Per tant, hi ha possibles interferències víriques de cadascuna de les vies de senyalització.
Inhibició de la presentació per molècules del MHC de classe I.
El principal mecanisme d’evasió de la immunitat adaptativa és inhibir la presentació per
molècules MHC-I de proteïnes víriques. Això es pot aconseguir per mitjà de:
• Inhibició de l’activitat del proteosoma per impedir la primera fase de la degradació de
proteïnes, com fan els virus Epstein Barr o els Citomegalovirus.
• Impedir el transport dels pèptids cap a l’interior del reticle endoplasmàtic per a ser
agregats a les molècules del MHC de classe I, com fan els Herpesvirus.
• Bloquejar la síntesi de les molècules del MHC de classe I en el reticle o retenir-les en
el mateix, com fan els adenovirus i els virus de la coriomeninigitis.
• Interferir en l’expressió i localització de les molècules del MHC.
• Facilitar la destrucció de les molècules del MHC de classe I en el reticle.
• Interferir en la presentació d’antigen MHC-I produint proteïnes similars a les MHC-I.
• Disminuir l’expressió de molècules coestimuladores.
Per tant, els virus codifiquen proteïnes que poden interferir amb gairebé cada etapa del
processament i presentació d’antigen.
Virus Proteïna Categoria Mecanisme
Herpes símplex
virus I ICP47
Bloqueig entrada
del pèptid al reticle
endoplasmàtic
Bloqueig d’unió del pèptid a
TAP
Citomegalovirus
Humà US6
Inhibeix activitat ATPasa de
TAP
Virus herpes
Boví UL49.5
Inhibeix transport del pèptid
TAP
Adenovirus E19
Retenció del MHC
de classe I al reticle
endoplasmàtic
Inhibició competitiva de la
tapasina
Citomegalovirus
humà US3 Bloqueig funció tapasina
Citomegalovirus
murí M152 Desconegut
Citomegalovirus
humà US2 Degradació dels
pèptids MHC de
classe I
Transporta molècules del MHC
de classe I acabades de
sintetitzar al citosol.
Herpes Virus γ
murí 68 mK3 Activitat E3-ubiquitina lligasa
Citomegalovirus
murí m4
Unió amb el MHC
de classe I a la
superfície cel·lular.
Interferència amb el
reconeixement per part dels
limfòcits Citotòxics

Immunologia 2020
CEBIO 143
Variació antigènica.
Un altre mecanisme d’evasió del sistema immunitari per part dels virus és la variació
antigènica. Això s’aconsegueix per canvis en el virus, la qual és molt gran i ràpida. Hi ha 2
tipus de variació antigènica.
La deriva antigènica es dona, sobretot, en virus de RNA. En aquests casos, les polimerases
víriques són poc fidels quan s’han de fer còpies del RNA i introdueixen mutacions al genoma
que deriven en proteïnes lleugerament diferents que no són reconegudes pels anticossos
generats durant la resposta primària. Són canvis de petita magnitud, però es tradueixen en
una resposta del sistema immunitari de menor intensitat.
El canvi antigènic no és el mateix que la deriva antigènica en tant que el canvi es produeix
per la combinació del genoma de virions humans amb els virions d’altres espècies que
infecten a la mateixa cèl·lula. Aquesta combinació dona lloc a noves espècies víriques amb
proteïnes molt diferents. Són canvis de gran magnitud, pel que passen desapercebudes pel
sistema immunitari. El cas més clar de virus que realitza un canvi antigènic és el virus
Influenza, causant de la grip i que realitza el canvi entre el virus humà i el virus de la grip.
Inhibició de la sinapsi immunològica.
Virus com el del VIH ha desenvolupat estratègies per a desorganitzar la formació de la sinapsi
immunològica de manera que no es pot produir una bona presentació d’antígens vírics als
limfòcits T. Per tant, no es produeix l’activació de la resposta cel·lular a aquest virus.
Altres mecanismes.
Un dels reservoris coneguts del virus del VIH son les cèl·lules fol·liculars dendrítiques.
Aquestes es troben als fol·licles limfoides i no tenen una maquinària de processament
d’antigen eficient ni molècules de MHC-II. Tenen un sistema de reciclatge de proteïnes de
membrana i d’immunocomplexes que estiguin associats en aquesta membrana, que fa que
el virus sigui internalitzat però que tornin a sortir a la membrana tal i com estaven, no hi ha
una degradació i, per tant, no hi ha una detecció d’aquestes cèl·lules per part del sistema
immunitari.
Una segona opció que tenen els virus per evitar el sistema immunitari és infectar cèl·lules no
permissives o semi-permissives de l’hoste per tal d’emmagatzemar-hi el seu material genètic
i passar desapercebuts al seu sistema immunitari. Els virus es mantenen de forma latent amb
una transcripció mínima o inexistent fins que, per alguna motiu, s’activa.
També hi ha altres virus que presenten mecanismes per a inhibir l’apoptosi, per endarrerir la
mort de la cèl·lula infectada tan com puguin per a poder generar mes virions infecciosos. Els
virus produeixen una sèrie de proteïnes que bloquegen la inducció de l’apoptosi en cadascuna
de les seves etapes, endarrerint la mort de la cèl·lula fins que els virions estan formats i són
infecciosos.
Finalment, hi ha virus, com l’Epstein Barr, que produeixen anàlegs d’algunes citocines com la
IL-10 i que s’uneixen als receptors substituint les citocines reals. En conseqüència,
interfereixen amb la senyalització de les citocines. Els virus contraresten l’activitat de les
citocines a la resposta produint molècules similars o homòlogues a aquestes citocines i els
seus receptors que s’uneixen i substitueixen les citocines reals i per tant interfereixen amb la
senyalització de les citocines de l’hoste.

Immunologia 2020
CEBIO 144
Conseqüències patològiques de la resposta a la infecció vírica. Malgrat la resposta immunitària als virus es dona per evitar que aquests acabin provocant
dany a l’organisme, hi ha cops que és el sistema immunitari qui provoca una conseqüència
negativa.
És el cas del dany tissular degut a la resposta dels limfòcits citotòxics, que poden donar lloca
l’hepatitis crònica quan hi ha una infecció per part de la coriomeninigitis. La destrucció de les
cèl·lules infectades per part d’aquest virus provoca una inflamació crònica, la qual dona lloc
a la hepatitis.
Per altra banda, està la infecció de les cèl·lules del sistema immunitari. Depenent de quines
cèl·lules del sistema immunitari es vegin afectades pel virus, el tipus d’immunodeficiència que
es pot donar és diferent. En el cas del virus del VIH, hi ha una mort selectiva dels limfòcits T,
el que genera una immunosupressió en els individus infectats. El virus Epstein – Barr infecta
els limfòcits B i genera una immunodepressió del sistema immunitari. Finalment, pot passar
que la infecció pels virus Epstein – Barr o per Virus Limfotròpic Humà de Tipus 1 (HTLV-1)
provoqui una transformació de les cèl·lules infectades.
També s’ha vist el lligam entre la resposta a determinats virus i el desenvolupament de
malalties autoimmunitàries. S’ha vist que els virus que no poden ser eliminats de manera
efectiva generen una activació crònica i una secreció d’IFN de tipus I de manera continuada.
S’ha vist que és important en el desenvolupament del lupus.
També, la resposta a antígens vírics pots donar lloc a la resposta a proteïnes pròpies. Per
exemple, la infecció del sistema nerviós central pel virus de Theiler’s, el virus de la hepatitis
murina, fa que la mielina sigui diana de la resposta T i B.
Hi pot haver el mimetisme molecular, que fa que hi hagi una similitud prou gran entre pèptids
derivats del virus amb pèptids derivats de proteïnes pròpies. Per tant, una conseqüència de
l’activació de la resposta a la infecció pel virus pot desencadenar una resposta a pèptids
derivats de la proteïna pròpia. Aquest és el cas de la cardiomiopatia induïda per resposta
prèvia a Coxsackie B virus. Finalment, hi ha certa relació entre una infecció per Coxsackie B
virus i el desenvolupament de la diabetis tipus 1.
Quan hi ha una infecció crònica o no ben eliminada de virus que produeixen gran quantitat
d’antígens i aquests no poden ser eficientment neutralitzats pels anticossos de l’hoste, es
formen immunocomplexes. Aquests immunocomplexes poden ser d’una mida gran i pot ser
que siguin difícils d’eliminar. Aquestes característiques provoca que es dipositin als ronyons
o als vasos sanguinis, el que causa una infecció crònica.
En alguns casos, els anticossos poc neutralitzants poden facilitar la internalització dels
complexes anticòs-virus i facilitar la infecció a traves de la unió a receptors Fc de macròfags.
Això s’anomena ADE, antibody dependent enhancement.

Immunologia 2020
CEBIO 145
Tema 16 – Anatomia Funcional del sistema Immunitari
Els Òrgans limfoides. Els òrgans limfoides es separen en primaris i secundaris. En els òrgans limfoides primaris
s’originen i maduren les cèl·lules del sistema immunitari. Són òrgans limfàtics primaris la
medul·la òssia i el timus. La medul·la és l’origen dels limfòcits B i T i s’encarrega de la
maduració dels primers. El timus és on es produeixen limfòcits T i cèl·lules NKT.
En els òrgans secundaris es concentren els limfòcits madurs, s’acumula l’antigen i s’inicia la
resposta immunitària. Són òrgans limfàtics secundaris els limfonodes, la melsa i els MALT,
els teixits limfàtics associats a mucoses. Aquests MALT es separen en BALT (bronquials),
GALT (intestí), NALT (nasofaringe) i SALT (pell). A més, tots aquests òrgans es troben
connectats entre si per vasos sanguinis i limfàtics.
Els òrgans limfoides primaris.
La medul·la òssia.
La medul·la òssia és on es generen els precursors hemopoètics que poblaran altres òrgans.
Els precursors es generen a la medul·la òssia per processos com l’eritropoesi6 i la
mielopoesi. També es generen els precursors dels limfòcits T i dels limfòcits B. Aquests
últims es desenvolupen a la mateixa medul·la, on obtindran diversitat i tolerància. Arran del
capteniment d’aquestes cèl·lules B que, quan es desenvolupen, es converteixen en cèl·lules
plasmàtiques, la medul·la òssia també és l’encarregada de la producció d’anticossos.
La medul·la es troba a tots els ossos llargs del
cos. A l’interior de tots aquests hi ha, doncs,
capacitat hematopoètica. Així i tot, la capacitat es
va perdent amb l’edat. A l’edat adulta bona part
de la medul·la es torna teixit adipós i això és el que
n’hi fa perdre. La màxima capacitat de
producció és en el moment del naixement i, a
mesura que passa el temps, es va perdent. La
pèrdua és inversament localitzada, doncs amb el pas dels anys la producció i capacitat
hemopoètica es va concentrant a les costelles, la pelvis i la columna vertebral.
6 Conjunt de processos de formació, maduració i alliberament al torrent sanguini dels eritròcits
Òrgans Limfoides
Òrgans Limfàtics Primaris
Medul·la Òssia
Timus
Òrgans Limfàtics Secundaris
Limfonodes Melsa MALT
Medul·la òssia
Precursors hemopoètics Precursors Limfòcits Producció d’anticossos
Melsa
Vèrtebres
Esternó
Costelles
Tíbia
Fèmurs
Hem
ato
poie
si re
lativa (
%) Medul·la
Òssia
Estructures
embriòniques
Fetge
Mesos Fetals Naixement Anys 70

Immunologia 2020
CEBIO 146
El timus.
El timus és l’altre òrgan limfoide
primari. Es troba situat al mediastí
i consisteix en una càpsula fibrosa
al voltant de dos lòbuls dividits en
lòbels separats per trabècules. En
cadascun d’aquests lòbels s’hi
troba el còrtex, la part més
perifèrica, la medul·la al centre i la
zona cortico-medul·lar entre les
dues. Aquesta zona participa en el
procés de diferenciació tímica
dels limfòcits T. En aquesta
mateixa zona també es dona la
selecció positiva dels limfòcits T
doble positius, mentre que a la
medul·la es fa la selecció negativa
dels limfòcits single positive.
Al timus és on té lloc la diferenciació dels precursors en les diferents poblacions de
limfòcits T, ja siguin convencionals com els TCD4, els TCD8, els Tαβ o els Treg; o no
convencionals com els NKT o els Tγδ. Al timus també té lloc el procés de selecció. Això
és gràcies a l’estructura anatòmica de la medul·la, del còrtex i de la zona cortico-medul·lar;
que permeten aquesta diferenciació juntament amb la composició cel·lular del tim.
Igual que amb la medul·la òssia, el timus involuciona amb l’edat. Al néixer, el timus té un
pes d’uns 20 grams i augmenta durant uns 10 anys fins arribar a uns 30-35 grams. Mai
desapareix del tot, però a partir de la pubertat s’inicia la involució. Aquesta consisteix en
una disminució del volum i en què el parènquima7 del tim és substituït per teixit adipós,
generant menys limfòcits T.
Apart de les cèl·lules del teixit adipós, al tim també es formen el que s’anomena Corpuscles
de Hassall. En aquests hi ha una acumulació de cèl·lules epitelials tímiques, macròfags,
cèl·lules dendrítiques i, de vegades, també limfòcits. Malgrat no tenen una funció ben
determinada, es sap que regulen la limfopoesi8.
Quan es parla de la substitució del parènquima, s’està fent referència a la pèrdua de
l’estructura característica de l’òrgan. En conseqüència d’aquesta substitució per teixit
adipós, hi ha una reducció de la xarxa de cèl·lules epitelials corticals, una regió medul·lar molt
més petita i difusa i es torna difícil distingir la regió cortico-medul·lar. Aquest envelliment del
timus, doncs, fa que aquest tingui una menor capacitat de generar limfòcits T madurs, el que
es tradueix en una acumulació poc diversa de limfòcits T de memòria a la perifèria.
Òrgans limfoides secundaris. El sistema immunitari és dispers i es troba escampat per totes les zones del cos però,
gracies als vasos limfàtics, tots els elements en que formen part (Limfonodes, melsa i MALT)
es troben connectats entre si. El sistema limfàtic recull els antígens via vasos limfàtics i
els concentra als limfonodes. El conjunt de citocines, quimosines i molècules d’adhesió
permeten la circulació, el homing i la retenció dels limfòcits en els òrgans limfàtics secundaris.
7 Teixit essencial d’un òrgan 8 Generació de limfòcits
Medul·la
Zona Còrtico-Medul·lar
Còrtex
Lòbel
Lòbul

Immunologia 2020
CEBIO 147
La captació d’antigen i la recirculació limfàtica a través dels òrgans limfàtics secundaris
són essencials per a una resposta immune eficient.
Vasos limfàtics.
Els vasos limfàtics es troben als teixits i recullen el líquid intersticial: tot el que hi hagi
al teixit i no siguin cèl·lules. La diferència de pressió entre l’espai intersticial i l’interior dels
capil·lars limfàtics facilita l’entrada als capil·lars. Aquest mateix procediment és el que ha
provocat, prèviament, la sortida del líquid des dels capil·lars sanguinis fins al líquid intersticial.
Les vàlvules flaplike dels vasos limfàtics permeten l’entrada del líquid intersticial als capil·lars
limfàtics. La circulació limfàtica es produeix per compressió de les estructures veïnes i
per la contracció de la musculatura llisa associada als vasos. Les vàlvules internes
dirigeixen l’avançament de la limfa i n’impedeixen el reflux. El vas limfàtic arribarà com a vas
aferent a un altre limfonode o un vas limfàtic de major diàmetre.
La limfa recollida a un teixit determinat va passant el filtre dels limfonodes cap a vasos
limfàtics de cada cop més diàmetre fins arribar al ducte, el vas limfàtic de major calibre.
Aquest connecta amb la circulació sanguínia. Per tant, una vegada el líquid intersticial ha
passat per tots els limfonodes, torna al teixit del que ha sortit. Així i tot, gràcies a la tasca dels
limfonodes, aquest líquid ja no té antígens ni patògens. Hi ha dues grans àrees de
drenatge. Una d’elles és el circuit que desemboca en el ducte toràcic, que avoca el seu
conegut a la vena subclàvia esquerra, i l’altre és el ducte limfàtic dret, que aboca el contingut
a la vena subclàvia dreta.
Això és important perquè hi ha una gran diversitat de limfòcits T però n’hi ha poques
quantitats de cada especificitat i es troben escampats per la circulació. De la mateixa
manera, per tal que es puguin reconèixer els antígens cal un punt de trobada: els limfonodes.
Aquest genera un entorn que fa possible la interacció de l’antigen amb els limfòcits i l’activació
del sistema immunitari.
Limfonodes.
Els limfonodes o ganglis tenen una estructura molt concreta. Tenen una sèrie de vasos
limfàtics aferents que arriben al limfonode i ho fan a nivell de la càpsula, la qual té les
trabècules. Per sota d’aquesta càpsula hi ha el sinus subcapsular, un canaló on els vasos
aferents hi aboquen els seus continguts. Al limfonode, la part més perifèrica correspon al
còrtex i és on hi trobem els limfòcits B. Anant cap a la part més interna hi ha el paracòrtex,
que és on es troben els limfòcits T. Finalment, al centre del limfonode, hi ha la medul·la. En
aquesta hi trobem les cèl·lules plasmàtiques, les que s’acaben de generar. La via de sortida
és un vas limfàtic eferent.
Als limfonodes es produeix l’entrada d’antigen a través dels vasos limfàtics aferents i es
filtra a través de l’estroma o bé és carregat a les cèl·lules dendrítiques madures per la mateixa
via. Un cop dins es dona el contacte de l’antigen amb les cèl·lules presentadores
d’antigen, que poden ser cèl·lules dendrítiques, macròfags o cèl·lules B. També es fa una
presentació d’antigen a les cèl·lules T i a les cèl·lules B per activar-les. Aproximadament,
cada dia passen 25*10^9 limfòcits pels limfonodes.
Hi ha 3 vies d’entrada de l’antigen al limfonode. En el cas dels immunocomplexes o les
partícules víriques, l’antigen és capturat i mostrat al còrtex per macròfags del sinus
subcapsular. En el cas dels antígens solubles petits o proteolitzats com les quimosines,
l’antigen accedeix al còrtex a través de gaps entre les cèl·lules que formen la paret del sinus
o a través de conductes de cèl·lules fibroblàstiques reticulars. Finalment, els antígens grans
o provinents de cèl·lules són transportats al còrtex dins les cèl·lules dendrítiques que l’han
capturat al teixit.

Immunologia 2020
CEBIO 148
Cèl·lules Fibroblàstiques.
Les cèl·lules fibroblàstiques reticulars embolcallen fibres reticulars com col·lagen de tipus
III, elastina i laminina tot formant canals o conductes FRC que transporten antígens solubles.
Les cèl·lules fibroblàstiques reticulars formen l’estructura o esquelet dels teixits amb una
elevada cel·lularitat. Serveixen per crear vies per on circulen els limfòcits i les cèl·lules
dendrítiques, pel que faciliten la presentació d’antigen. Les cèl·lules Fibroblàtiques reticulars
expressen molècules com el MHC de classe I i molècules d’adhesió. També secreten
quimiocines com CCL19, CCL21 i CXCL13, que atrauen cèl·lules dendrítiques, limfòcits T i
limfòcits B.
Als limfonodes, el 90% dels limfòcits que hi entren són naive i ho fan a través de les cèl·lules
de les Vènules de l’endoteli alt. Aquestes són vènules9 especialitzades que es troben en
els llocs de migració del limfòcit des de la sang fins l’estroma d’un gangli limfàtic perifèric o
mucós. Les HEV (que reben el seu nom per les seves sigles en anglès) es caracteritzen pel
fet de tenir les cèl·lules endotelials prominents, cuboides i altes que expressen molècules
d’adhesió úniques implicades en la unió dels limfòcits T verges. El 10% dels limfòcits restants,
majoritàriament cèl·lules de memòria, entren a través dels vasos limfàtics aferents.
Els limfòcits T naive o verges tendeixen a concentrar-se als limfonodes. La vènula de l’endoteli
alt és la que permet el homing dels limfòcits T naive al limfonode. L’acumulació d’aquests es
sol donar al paracòrtex, que és on desemboquen les vènules d’endoteli alt.
Entrada i sortida de cèl·lules.
Els limfòcits surten dels vasos sanguinis a través de les HEV i es dirigeixen als limfonodes.
En aquests, els limfòcits B es dirigeixen al còrtex i els T al paracòrtex. Això passa perquè
els limfòcits T expressen receptors de quimiosines CCR7, que reconeixen CCL21 i CCL19.
Aquestes dues quimiocines s’expressen al paracòrtex. Els limfòcits B, en canvi, expressen
els receptors CXCR4 i CXCR5. Aquest permeten interaccionar amb les quimiocines CXCL12
i CXCL13.
Els limfòcits B s’acumulen formant fol·licles. Si no hi ha resposta són primaris. Si hi ha
resposta són secundaris. En aquests últims es forma el centre germinal. En el centre
germinal hi ha una zona fosca, on trobem centroblasts dividint-se. Aquesta divisió implica
que el domini variable del BCR passa pel procés d’hipermutació somàtica. També hi ha una
zona clara, on s’està fent una selecció dels centròcits en funció de l’afinitat del seu BCR
mitjançant el contacte amb les cèl·lules fol·liculars dendrítiques. Els limfòcits B amb millor
afinitat poden fer un canvi d’isotip i desenvolupar-se en cèl·lula plasmàtica. També en
aquesta zona clara s’està donant la mort per apoptosi dels limfòcits B amb menys afinitat
per l’antigen. Existeix una corona o mantell. És on hi ha immunoglobulines D i cèl·lules B en
repòs, és a dir, que no són específiques per l’antigen al qual s’està responent al centre
germinal.
La Melsa.
La melsa és l’òrgan limfoide secundari més gran. No hi arriben els vasos limfàtics aferents
però està connectada directament amb la circulació sanguínia mitjançant l’artèria
esplènica. La melsa és un òrgan especialitzat en capturar i respondre a antígens que
entren per via sanguínia. Amb una estructura relativament semblant als limfonodes, es
separa en polpa roja i en polpa blanca.
La polpa roja és una xarxa de sinus amb glòbuls vermells, macròfags i algun limfòcits. La
polpa roja és la part encarregada d’emmagatzemar eritròcits i eliminar-ne els envellits. La
9 Vena petita

Immunologia 2020
CEBIO 149
polpa blanca és la part de la melsa que envolta les artèries, en forma d'una beina de limfòcits,
principalment limfòcits T. La sigla PALS prové de l'anglès periarteriolar lymphatic sheath. En
aquesta polpa blanca també hi trobem fol·licles limfoides amb cèl·lules B. Hi ha cèl·lules
interdigitants, semblants a les FDCs dels limfonodes que actuen com a cèl·lules
presentadores d’antigen.
Sistema immunitari associat a les mucoses.
El sistema immunitari associat a les mucoses (MALT) és el més complex i abundant del
sistema immunitari. Amb una altíssima activitat immunitària, ha de desenvolupar immunitat
activa contra els patògens que travessen la superfície mucosa i, al mateix temps, controlar
que no hi hagi resposta davant els aliments o la microbiota. Amb el MALT s’estableix una
modulació fina de la resposta immune per tal d’induir una tolerància local (i sistèmica) a
antígens innocus alhora que es manté la vigilància respecte els microorganismes patogènics.
L’estímul antigènic de la microbiota participa en el desenvolupament i el manteniment del
MALT. El sistema immunitari associat a les mucoses es troba a les plaques de Peyer, a les
amígdales, als adenoides, a l’apèndix i a tot el teixit associat a la mucosa gàstrica, respiratòria
i reproductora apart de trobar-lo a la pell.
GALT.
El teixit limfoide associat a la mucosa intestinal es troba format per regions estructurades com
ho són les plaques de Peyer o els fol·licles aïllats. Aquest teixit limfoide fa servir limfòcits
intraepitelials i limfòcits dispersos per la làmina pròpia. El GALT té una irrigació sanguínia
amb la presència de vènules d’endoteli alt i de vasos limfàtics que drenen als limfonodes
mesentèrics. Els limfòcits intestinals es troben en teixits organitzats, on s’indueixen les
respostes immunes, i repartits per l’intestí, on poden realitzar altres funcions efectores.
El GALT està format per diversos tipus cel·lulars, com les cèl·lules de paneth, que generen
pèptids antimicrobians, les cèl·lules de goblet, que produeixen moc que forma una capa per
separar el lumen de les cèl·lules epitelials, les cèl·lules tuft, que produeixen interleuquina 25
i activen les cèl·lules ILC (que acaben estimulant les cèl·lules productores d’interleuquina 13)
i les cèl·lules M.
Les plaques de Peyer de les que hem parlat són nòduls limfàtics situats a la submucosa de
l'intestí prim i que tenen una funció similar als ganglis limfàtics. Recobreixen una part
important de l’intestí prim i generalment es troben associats a cèl·lules M.
Les cèl·lules M són cèl·lules de l’epiteli de l’intestí prim amb una membrana apical molt
invaginada i una concavitat anomenada butxaca basolateral on s’acumulen els limfòcits B,
els limfòcits T i els macròfags. L’antigen entra a través d’aquestes cèl·lules, que es situen en
els llocs inductius. Un lloc inductiu és una regió curta de l’epiteli que s’emplaça sobre els
fol·licles limfoides.
Els limfòcits intraepitelials de la mucosa solen ser limfòcits TCD8. Aquests expressen una
integrina única, la α4β7. Es troba involucrada en el homing. En quant a la diversitat dels
limfòcits intraepitelials és destacable que hi ha més cèl·lules T γδ que a la circulació
sanguínia o als limfonodes. Sobre els limfòcits de la làmina pròpia, la gran majoria són
limfòcits T i limfòcits B activats, juntament amb basòfils, mastòcits i macròfags. La majoria de
les cèl·lules T de la làmina pròpia són TCD4. Els limfòcits B del sistema immunitari associat
a les mucoses produeixen immunoglobulines A i poden induir tolerància.
Aquesta immunoglobulina A dimèrica que es produeix es pot unir a un receptor polyIgA de la
membrana basolateral de les cèl·lules epitelials intestinals. Aquest receptor transporta la
immunoglobulina A a la membrana apical, la que està en contacte amb la llum de l’intestí.

Immunologia 2020
CEBIO 150
Dins la cèl·lula es dona un tall del receptor polyIgA que provoca que un fragment es quedi
unit a la immunoglobulina quan aquesta es trobi al lumen de l’intestí per realitzar la seva
funció.
Les tonsil·les i les adenoides constitueixen les principals àrees inductores de resposta als
antígens intranasals i inhalats a humans, primats i ratolins. Formen una barrera de teixits
limfoides anomenada anell de waldeyer.
En l’epidermis, el teixit limfoide conté queratinòcits, encarregats de produir diverses citocines
proinflamatòries i expressar molècules del MHC de classe II, cèl·lules de langerhans, que
no deixen de ser cèl·lules dendrítiques però amb la diferència que resideixen a l’epidermis, i
limfòcits intraepidèrmics, que majoritàriament són limfòcits Tαβ i limfòcits Tγδ.

Immunologia 2020
CEBIO 151
Tema 17 – Recirculació Limfocitària Les cèl·lules de la immunitat innata no es mouen del teixit on van. Els leucòcits, en canvi, es
mouen pel sistema fent servir els vasos sanguinis i els vasos limfàtics. El moviment dels
leucòcits a través dels vasos sanguinis i limfàtics s’aconsegueix gràcies a les
Molècules d’adhesió cel·lular i a altres tipologies moleculars.
Molècules d’Adhesió Cel·lular. Les molècules d’adhesió cel·lular (CAMs) són molècules expressades a la membrana
cel·lular que permeten la interacció dels leucòcits amb altres leucòcits, amb cèl·lules dels
teixits o amb la matriu extracel·lular. Algunes molècules fan interaccions homofíliques. Altres,
heterofíliques.
Les Molècules d’Adhesió Cel·lular intervenen en els patrons de recirculació dels
leucòcits, en la seva migració als teixits i en la seva extravasació. També s’encarreguen
de l’estabilització del reconeixement d’antigen pel TCR, dels processos de maduració i
diferenciació i de la modulació de la resposta limfocitària.
En funció de les seves semblances estructurals i funcionals, les molècules d’adhesió cel·lular
es classifiquen en Selectines, Sialomucines, Integrines i en la Superfamília de les
immunoglobulines.
Circulació dels Limfòcits. Els limfòcits T naive van de la circulació sanguínia fins als limfonodes, on s’hi estan un
temps monitoritzant si hi ha antígens pels quals tenen el TCR específic. Si no s’activen, al
cap d’un temps surten cap a la circulació sanguínia a través dels vasos limfàtics
eferents, que desemboquen en els ductes. D’aquí, els limfòcits poden anar a la melsa, als
teixits... Els limfòcits estan contínuament en moviment. Aquest moviment continu és el
que es coneix com recirculació limfocitària.
Un limfòcit pot fer un circuit circulatori complet dues vegades al dia sempre i quan no
reconegui cap antigen pel qual tingui un TCR específic. El crcuit circulatori complert comença
a la sang, d’on es pot derivar als teixits o als òrgans limfàtics secundaris. D’aquests aniran a
parar a la limfa, d’on tornaran a la sang.
La cascada de processos que es dona en la interacció entre els leucòcits i la cèl·lula
endotelial és el que els permet realitzar una extravasació. Aquesta cascada consisteix en
un rodolament, una activació, un reforçament i una transmigració.
L’enganxament i rodolament o rebot és l’aproximació del leucòcit a l’endoteli alhora que
redueix la seva velocitat. En aquest procés intervenen les selectines. Els granulòcits i els
monòcits rodolen mentre que els limfòcits reboten.
L’activació és una etapa on la interacció entre les quimiosines i els receptors
d’aquestes determinaran quines cèl·lules reduiran encara més la seva velocitat i, en
conseqüència, seran les que podran fer l’extravassació. Les quimiocines, en aquesta etapa,
tenen un doble paper. Per un costat, les quimiocines fan una quimiotaxi al provocar la
retenció i concentració dels leucòcits sobre les cèl·lules endotelials on es troben. Per altra
banda, les quimiosines realitzen una activació de la funció integrina. La funció integrina
consisteix en la unió de la cèl·lula a la Matriu Extracel·lular o en la transmissió del senyal des
de la matriu fins dins la cèl·lula.

Immunologia 2020
CEBIO 152
El reforçament i aturada és un procés dut a terme per les integrines on els leucòcits aturen
la seva circulació i els canvia la morfologia per tal d’iniciar la transmigració a través de
l’endoteli. Aquest procés de canvi morfològic rep el nom de polarització.
La transmigració és l’últim pas de la cascada i consisteix en un extravasament per diapèdesi
dels leucòcits. Diapèdesi és el nom que rep la sortida dels leucòcits a través dels capil·lars
sanguinis i es duu a terme gràcies a les integrines, el citoesquelet i les col·lagenases.
Molècules de la recirculació limfocitària.
Les Selectines i les Sialomucines.
Una selectina és qualsevol de les tres proteïnes lligands de glúcids estretament relacionades
que mitjança l'adhesió de leucòcits a les cèl·lules endotelials. Les selectines reconeixen
sialomucines.
Cada molècula selectina és una glicoproteïna transmembrana d'una sola cadena amb una
estructura modular similar, inclòs un domini lectina tipus C extracel·lular dependent de
calci. Aquest domini tipus C és el responsable de la interacció amb les sialomucines.
Les Selectines són la selectina L (CD62L), expressada constitutivament a limfòcits T i B
naive; la selectina P (CD62P), expressada per plaquetes i cèl·lules endotelials activades; i la
selectina E (CD62E), expressada per cèl·lules endotelials activades. La selectina L reconeix
les sialomucines GlyCAM-1, CD34 i Mad-CAM-1. Les selectines P i E reconeixen la
sialomucina PSGL-1.
Les sialomucines, també anomenades adresines vasculars, són proteïnes molt glicosilades
que permeten la unió a selectines. Són sialomucines la GlyCAM-1 i CD34, que s’expressen
de manera constitutiva a les vènules de l’endoteli alt dels limfonodes; Med-CAM-1, que
s’expressa a les cèl·lules endotelials dels vasos de les mucoses i condueix els limfòcits al
teixit limfoide del GALT; i PSGL-1, que s’expressa a neutròfils, monòcits i limfòcits T efectors
i és el principal responsable de la seva unió a l’endoteli activat.
Les selectines són les responsables de guiar els leucòcits cap a determinats teixits,
procés anomenat homing. Les selectines interaccionen amb els carbohidrats de les
sialomucines a través del domini lectina tipus C extracel·lular. Cada selectina uneix uns
carbohidrats determinats. La selectina L, expressada a limfòcits T i B naive, és responsable
de l’entrada dels limfòcits als Òrgans Limfàtics Secundaris a través de les vènules de l’endoteli
alt. La selectina P i la selectina E, expressades a l’endoteli vascular activat, faciliten el
reclutament de cèl·lules efectores al lloc d’inflamació.
En condicions homeostàtiques, els leucòcits viatgen a gran velocitat pels vasos
sanguinis i les cèl·lules endotelials no expressen selectines E i P. Així i tot, els leucòcits
sí expressen la selectina L i la sialommucina PSGL-1 en cas que estiguin activats. En el
moment d’inflamació, les cèl·lules endotelials comencen a expressar les selectines E i P, que
els permeten interaccionar amb la sialomucina PSGL-1, la qual indueix la extravassació dels
leucòcits.
Les Quimiocines.
Les Quimiocines són una gran família de citocines de baix pes molecular amb una estructura
homòloga que estimulen el moviment del leucòcit i regulen la migració dels leucòcits
des de la sang als teixits.
Les quimiocines interaccionen amb els glicosaminoglicans (GAG) carregats
negativament. Els GAGs solen estar units a proteïnes de la membrana cel·lular o de la matriu

Immunologia 2020
CEBIO 153
extracel·lular en estructures anomenades proteoglicans. Quan les quimiocines són
secretades, queden retingudes i concentrades a la superfície cel·lular i a la matriu
extracel·lular, pel que representen un suport sòlid per on les cèl·lules poden reptar i
migrar.
Les quimiocines, apart de determinar quines cèl·lules interaccionaran amb l’endoteli durant el
pas d’activació, activen les integrines tot canviant l’afinitat d’unió als seus lligands. Els
lligands de les integrines són molècules de la superfamília de les immunoglobulines.
Gràcies a l’acció de les quimiocines, les integrines adquireixen major afinitat pel seu lligand.
Les integrines.
Les integrines són proteïnes heterodimèriques formades per una cadena α i una
cadena β unides per enllaços no covalents. Són proteïnes de la superfície cel·lular
importants per a les interaccions entre el limfòcit T i les cèl·lules presentadores d'antigen i per
a la migració dels leucòcits des de la sang fins als teixits. Les integrines tenen un paper
crític en l’extravasació leucocitària.
Hi ha dues principals famílies de les integrines; els membres de cada família d’integrines
expressen una cadena β conservada (β1/CD29 i β2/CD18) associada a diferents cadenes
α. Les Integrines es classifiquen segons la cadena β que expressin. Entre les integrines més
importants es troben αLβ2 (LFA-1), expressada a limfòcits, monòcits, cèl·lules dendrítiques i
neutròfils; αMβ2, expressada a neutròfils, monòcits i macròfags; α4β7 (LPAM-1), expressada
a limfòcits que es dirigeixen a les mucoses; i α4β1 (VLA-4), que s’expressa en monòcits i
limfòcits.
Els leucòcits en repòs expressen integrines de baixa afinitat pels lligands. Als leucòcits
activats són d’alta afinitat. Hi ha diferents estímuls que poden regular l'afinitat pels lligands
de les integrines, entre els quals es troben les quimiocines. Els dominis citoplasmàtics de
les integrines s'uneixen al citoesquelet cel·lular, on es dona la senyalització intracel·lular
a través del receptor de la quimiocina.
Les adhesines.
Les adhesines són els lligands de les integrines i pertanyen a la superfamília de les
immunoglobulines, una gran família de proteïnes que contenen una estructura globular
anomenada domini de tipus Ig o plegament de Ig. Les adhesines també uneixen proteïnes
de la matriu extracel·lular com la fibronectina, el fibrinogen i la laminina.
Les adhesines tenen un baix nivell d’expressió a les cèl·lules de l’endoteli vascular en
repòs. A les zones inflamades, on es produeixen de forma local citocines proinflamatòries
com TNFα, hi ha una inducció de la seva producció.
Són exemples de les integrines les ICAM-1 i ICAM-2, que s’expressen a l’endoteli activat i en
repòs respectivament; VCAM-1, que s’expressa a l’endoteli activat; i PECAM, que s’expressa
als leucòcits activats i a les unions entre cèl·lules endotelials.
L’adhesina PECAM és una molècula crítica en el procés d’extravasació que s’expressa
a les unions intercel·lulars de les cèl·lules endotelials i al limfòcit. Facilitat per l’adhesina
PECAM, el procés d’extravasació requereix de la ruptura transitòria i reversible de les
proteïnes d’unió adherent així com d’una digestió de la membrana basal dels vasos
sanguinis. Aquesta última digestió la duen a terme metal·loproteases.
Extravassació dels Neutròfils i dels Monòcits. Els receptors de quimiocines expressats en el procés d’extravasació són diferents per
neutròfils i per monòcits. També s’observen diferències en l’expressió de les integrines i de

Immunologia 2020
CEBIO 154
les adhesines, el que determina la facilitat amb què aquests tipus cel·lulars extravasaran i
aniran al teixit a fer la seva funció efectora.
Els neutròfils interaccionen amb la selectina L per mitjà de la PSGL-1, el que permet que
hi hagi una interacció de la CXCR1 del neutròfil amb la CXCL18 de l’endoteli. Aquest
interacció suposarà un canvi de conformació de LFA-1 i de MAC-1, augmentant l’afinitat.
Quan seguidament hi hagi una interacció suficient amb ICAM-1, es donarà la transmigració
per mitjà de PECAM-1.
Els monòcits interaccionen amb la selectina L per mitjà de la PSGL-1, el que després els
permet que hi hagi una interacció de CCL2 o CX3CL1 del monòcit amb CCR2 o CX3CR1 de
l’endoteli, respectivament. Aquest interacció suposarà un canvi de conformació de VLA-4 o
de α4β1 i de LFA-1. Quan hi hagi una interacció suficient amb VCAM-1 i ICAM-2, es donarà
la transmigració per mitjà de PECAM-1.
Recirculació als Òrgans Limfàtics Secundaris. Al paracòrtex dels limfonodes hi ha les vènules d’endoteli alt. Aquestes expressen, de forma
constitutiva, les adresines vasculars CD34 i GlyCAM-1. Això permet el homing o l’entrada de
limfòcits T i B naive, que expressen la selectina L, al limfonode.
Les vènules d’endoteli alt són vènules post-capil·lars que tenen cèl·lules endotelials
amb forma cúbica permanentment activades. Tenen una expressió constitutiva de les
adresines CD34 i GlyCAM-1, que permeten el homing de limfòcits T i B naive per la seva
expressió de selectina L. Per això, les vènules de l’endoteli alt són responsables de la
recirculació dels limfòcits naive per tots els òrgans limfoides secundaris excepte la
melsa. S’estima que, al limfonode, s’extravasen 14,000 limfòcits per segon.
La majoria de limfòcits naive entren als òrgans limfàtics secundaris en busca d’antigen.
Si no el troben després d’un període de temps concret d’entre 12 i 18 hores per les cèl·lules
T i unes 24 hores pels limfòcits B, els limfòcits naive surten a través dels vasos limfàtics
eferents per dirigir-se a la circulació limfàtica.
Les cèl·lules T migren a la zona T del limfonode a una “velocitat” de 12µM per minut a
través d’un moviment que sembla aleatori. En realitat, la migració dels limfòcits T a la zona
T està dirigida per quimiocines. Es calcula que una única cèl·lula dendrítica interactua amb
aproximadament 5,000 cèl·lules T per hora a la zona T.
Les cèl·lules T i B naive recirculen a l’interior dels limfonodes per l’expressió dels
receptors de quimiocines CCR7 i CXCR5 i els seus lligands.
CCR7 és un receptor de les quimiocines CCL19 i CCL21. CCR7 permet la migració a
través de la zona T. CCL21 està produida per les vènules de l’endoteli alt i per cèl·lules
estromals dels teixits limfoides. CCL19 està produida per les cèl·lules estromals de l’àrea T i,
en menys quantitat, per les cèl·lules dendrítiques interdigitants.
CXCR5 és un receptor de CXCL13, una quimiocina produida per les cèl·lules fol·liculars
dendrítiques localitzades als fol·licles. La interacció CXCR5-CXCL13 determina
l’acumulació de les cèl·lules B per formar els fol·licles. CXCR5 s’indueix en les cèl·lules
T helper fol·liculars (Thf) i els permet migrar a la zona clara del centre germinal.
La migració de les cèl·lules T al paracòrtex es fa seguint els conductes que formen les
cèl·lules fibroblàstiques reticulars (FCR), on s’uneixen les quimiocines que permeten que
els limfòcits transitin com per carreteres on també circulen les cèl·lules dendrítiques. Això,

Immunologia 2020
CEBIO 155
doncs, facilita la interacció entre ambdues cèl·lules i, conseqüentment, facilita el
reconeixement d’antigen.
Una vegada han reconegut antigen, les cèl·lules efectores surten pel vas eferent a la
circulació limfàtica fins al ducte toràcic, on es volca la limfa a la circulació sanguínia a
través de la vena subclàvia.
Diferències entre Limfòcits T efectors i limfòcits T naive. Els limfòcits T activats redueixen l’expressió de molècules que els condueixen als
òrgans limfàtics i augmenten l’expressió de molècules que participen en la seva
migració als llocs perifèrics de la infecció i lesió tissular. Algunes de les molècules que
veuen reduïda la seva expressió són la selectina L i el receptor per quimiocines CCR7.
Algunes de les que veuen augmentada la seva expressió són les integrines LFA-1 i VLA-4,
els lligands per les selectines E i P i diversos receptors per quimiocines. L’activació del
limfòcit també augmenta l’expressió de CD44, un receptor per l’àcid hialurònic de la
matriu extracel·lular. La unió de CD44 al seu lligand ajuda a retenir els limfòcits T efectors
en els llocs d’infecció i lesió tissular.
Les cèl·lules T efectores també augmenten l’expressió de CD2 i LFA-1, unes molècules
d’adhesió; de CD45RO, que s’associa al TCR i a CD4 alhora que té una activitat fosfatasa
que baixa el llindar d’activació; i de VLA4, induint el homing a l’endoteli vascular inflamat. Les
cèl·lules T efectores, en canvi, redueixen l’expressió de la selectina L, que indueix al homing
al limfonode, i de CD45RA, que provoca el splicing diferencial del mRNA de CD45.
Extravasació dels limfòcits T efectors i memòria. Els limfòcits T efectors fan homing a les zones d’infecció i hi detecten l’endoteli inflamat
gràcies a les quimiocines que es generen al focus inflamatori. El perfil migratori de la
cèl·lula T efectora depèn del lloc on la cèl·lula dendrítica hagi capturat l’antigen que
presenta al limfòcit T naive. El tipus de receptor de quimiocines de les cèl·lules efectores,
juntament amb la integrina, determinen el homing de la cèl·lula T efectora.
Els receptors de quimiocines CCR9, que s’uneix a CCL25, i α4β7, que s’uneix a MadCAM-1,
indiquen homing a la mucosa. En canvi, els receptors de quimiocines CCR4, que s’uneix a
CCL17, CCR10, que s’uneix a CCL27, CLA, que s’uneix a la selectina E, i α4β1, que s’uneix
a VCAM-1, indiquen un homing a la pell.
Extravasació a la pell
Els limfòcits T efectors que s’extravasen a la pell són els que han estat activats als
limfonodes proximals, els quals reben la limfa procedent de la zona inflamada de la pell.
Durant el reconeixement d’antigen presentat per les cèl·lules dendrítiques activades in situ,
s’indueix l’expressió de les molècules necessàries per poder migrar a la pell. Les molècules
necessàries per poder migrar a la pell són CLA, CCR4 i /o CCR10 i α4β1.
Paper de les molècules d’adhesió en la presentació d’antigen. Les molècules d’adhesió són molècules de la superfície cel·lular la funció de les quals
és promoure les interaccions adhesives amb altres cèl·lules o amb la matriu
extracel·lular. Aquestes molècules són importants en les interaccions contínues dels
LFA-1 - ICAM-1 LFA-1 - ICAM-2 ICAM-3 - DCSIGN CD2 - CD58

Immunologia 2020
CEBIO 156
limfòcits T amb les cèl·lules dendrítiques als limfonodes. Aquestes interaccions són
transitòries i es donen entre molècules d’adhesió i els seus lligands. En destaquen 4:
Les integrines es troben en un estat de baixa afinitat a les cèl·lules en repòs. Les integrines
pateixen un canvi conformacional que augmenta l’afinitat per seu lligand quan hi ha
una senyalització a través del TCR. Per tant, s’estabilitza la interacció.

Immunologia 2020
CEBIO 157
Tema 18 – Resposta cel·lular. Activació de les cèl·lules T.
Presentació d’Antigen. Les cèl·lules dendrítiques que es troben als teixits de barrera capturen i processen l’antigen
per, seguidament, transportar-lo cap als limfonodes proximals a través dels vasos limfàtics.
Al paracòrtex dels limfonodes poden presentar l’antigen als limfòcits T naive que allà es
trobin. Aquesta presentació no és aleatòria. Requereix de moltes interaccions, les quals van
realitzant transitòriament els limfòcits T. En cas que hi hagi reconeixement, el limfòcit
s’activarà i serà un limfòcit T efector. Aleshores sortirà del limfonode cap al teixit, on realitzarà
la seva funció efectora.
Als limfonodes, els limfòcits T interaccionen contínuament amb les cèl·lules presentadores
d’antigen. Aquestes interaccions són transitòries entre les molècules d’adhesió i els seus
lligands. Aquestes interaccions poden ser de 4 tipus: LFA-1 – ICAM-1, LFA-1 – ICAM-2,
ICAM-3 – DC-SIGN i CD2 – CD58. LFA-1, ICAM-3 i CD2 es troben a la membrana de la
cèl·lula T, mentre que ICAM-1, ICAM-2, DC-SIGN i CD58 es troben a la membrana de les
cèl·lules presentadores d’antigen.
Etapes de la resposta cel·lular T. Una vegada s’ha produït l’acostament pot tenir lloc la primera etapa de la resposta cel·lular
T. Aquesta primera etapa consisteix en el reconeixement d’antigen per part dels limfòcits
T, que tan sols dependrà de la interacció TCR-complex MHC/pèptid. En aquest primer senyal
o etapa hi tenen un paper mot rellevant els coreceptors.
La segona etapa serà l’activació cel·lular. En l’activació, hi ha d’haver un senyal entre la
molècula coestimuladora del limfòcit T i el seu lligand. Totes aquestes interaccions resulten
en la transducció de senyals a l’interior de la cèl·lula, el que es coneix com a cascada
bioquímica de segons missatgers. Aquesta cascada consisteix en l’activació de la transcripció
de gens, la repressió de la transcripció d’altres gens, l’expressió de noves proteïnes de
membrana, la secreció de citocines activadores del creixement autocrí i paracrí com IL-2 i la
inducció de l’activitat mitòtica. Aquest últim fenomen de la cascada es tradueix en una
proliferació i expansió de les cèl·lules T.
La tercera etapa de l’activació cel·lular és la de l’activació de la funció efectora, la qual
requereix de senyals d’adhesió i, sobretot, de citocines. Finalment, la quarta etapa es
correspon amb la regulació de la resposta immunològica.
Activació del limfòcit T.
L’activació del limfòcit T és el resultat de la generació de 3 senyals. El primer senyal és
el derivat de la interacció entre el TCR i el complex pèptid/MHC. En la generació d’aquest
primer senyal també hi participa la senyalització dels coreceptors CD4 o CD8. El segon senyal
és el de la coestimulació mediada per la interacció de CD28 del limfòcit amb CD80 ó CD86
de la cèl·lula presentadora d’antigen. Enlloc d’això, el segon senyal es pot dura a terme per
senyals derivats de molècules de la immunitat innata. Finalment, el tercer senyal és el resultat
de la interacció de citocines amb el seu receptor. Aquest tercer senyal és principalment
responsable de la diferenciació de les cèl·lules T activades en cèl·lules efectores de diferents
tipus.
1r senyal d’activació: el reconeixement d’antigen pel complex TCR/MHCpèptid.
El TCR no senyalitza directament, ho fa a través del CD3. Les cadenes d’aquest tenen
motius ITAM en el seu costat citoplasmàtic. El complex TCR/CD3 el formen una o dues
molècules del TCR associada/es a les 6 cadenes del CD3: una cadena δ, una γ, dues ε i el
dímer de cadenes ζ. L’associació és de tipus no covalent amb el TCR. Les cadenes ζ, de

Immunologia 2020
CEBIO 158
pocs aminoàcids extracel·lulars, és pràcticament intracel·lular. Els motius ITAM
(Immunoreceptor Tyrosine based Activation Motifs) participen en la transmissió de senyals al
reclutar tirosina quinases un cop s’han fosforilat.
Quan el TCR interacciona amb el complex pèptid/MHC, la quinasa citoplasmàtica Fyn del
limfòcit fosforila la quinasa Lck. Aquesta, ja activada, fosforila els ITAMs de CD3γ, δ, ε i ζ.
Posteriorment, la tirosina quinasa ZAP-70 s’uneix als ITAM fosforilats de CD3ζ i Lck acaba
de fosforilar ZAP-70, que s’activa. La complerta activació necessita, per tant, la interacció del
coreceptor amb la molècula del MHC. La fosfatasa CD45 regula la fosforilació de Lck i Fyn
en estat de repòs però és apartada del TCR quan es produeix la interacció amb el complex
pèptid/MHC.
A la cèl·lula en repòs, la tirosina quinasa Lck, associada al coreceptor CD4 o CD8, es troba
separada del complex TCR/CD3 perquè està inclosa en els lípid rafts o balses lipídiques, com
moltes molècules d’adhesió. Aquestes són microdominis de membrana rics en esfingomielina,
glucoesfingolípids i colesterol, pel que són més rígides. El complex TCR/CD3, en canvi, es
troba fora dels lípid rafts. Quan el TCR interacciona amb el complex pèptid/MHC, migra als
lípid rafts i s’associa al coreceptor CD4 o CD8.
Quan ZAP-70 està activat, fosforila l’adaptador LAT. La fosforilació crea llocs lliures per a la
unió de SLP-76 i altres molècules, el que inicia una cascada de senyalització. SLP-76 és el
responsable dels canvis en el citoesquelet del limfòcit T. Per tant, és el que li canvia la forma
en activar-se. Per altra banda, A LAT fosforilat s’hi pot unir la fosfolipasa Cγ i el factor
intercanviador del nucleòtid guanina (GEF). La fosfolipasa Cγ (PLCγ) fosforilada catalitza la
hidròlisi d’un fosfolípid de la membrana plasmàtica anomenat PIP2que genera dos productes:
el 1,4,5-trifosfat d’inositol (IP3) i el diacilglicerol (DAG) unit a la membrana. IP3 i DAG actuen
en dues vies de transmissió de senyals dels limfòcits T.
IP3 es difon pel citosol fins al reticle endoplasmàtic per unir-se al seu receptor, un canal de
calci activat per lligand, i estimula la sortida d’ions calci del reticle endoplasmàtic cap al
citoplasma i l’entrada de calci del medi extracel·lular. El calci alliberat provoca un augment
ràpid del calci lliure citosòlic als pocs minuts de l’activació del limfòcit T. L’augment de Calci
citoplasmàtic permet unir a la calmodulina, que activa la calcineurina i aquesta activa NFAT.
El diacilglicerol (DAG), el segon producte de l’escissió del PIP2, és un lípid unit a la membrana
que atreu la proteïna quinasa C (PKC). Hi ha diferents formes de PKC que participen a la
generació de factors de transcripció actius. La combinació de l’augment del calci citosòlic lliure
i DAG activa certes isoformes de PKC associades a la membrana al induir un canvi
tridimensional que deixa accessible el lloc catalític de la quinasa als seus substrats. La PKC
activa el factor de transcripció NFκB.
D’altra banda, GEF estimula el canvi de GTP per GDP a la proteïna Ras, una petita proteïna
lligadora del nucli de guanina (o proteïna G petita). Arran d’aquest canvi, la proteïna Ras
pateix un canvi conformacional i pot aleshores reclutar la proteïna Raf, que serà l’encarregada
d’iniciar la via de pes MAP quinases tot activant la MAP quinasa Erk.
L’activació de la MAP quinasa Erk li permet entrar al nucli, on fosforila el factor de transcripció
Elk-1. Aquest estimula la transcripció del gen FOS. A través d’una altra cascada de
senyalització s’acaba fosforilant la proteïna quinasa Jun que, juntament amb FOS fosforilada,
provoca l’activació de moltíssims gens transcripcionals.
2n senyal d’activació: Molècules coestimuladores.
Les molècules coestimuladores proporcionen el segon senyal d’activació o coestimulació.
Són necessàries per a l’activació i diferenciació de limfòcits T naive a més del reconeixement

Immunologia 2020
CEBIO 159
d’antigen pel TCR. El coestimulador CD28 es troba expressat constitutivament als limfòcits T
naive mentre que ICOS s’expressa a limfòcits T activats. Els lligands de CD28 són B7-1 i B7-
2 i tenen una expressió baixa o negativa a les cèl·lules presentadores d’antigen en repòs però
s’indueixen amb PRRs i citocines de la resposta innata. Hi ha molècules coestimuladores com
CTLA-4 i PD-1 que s’expressen a limfòcits T activats i que participen en la regulació.
Els productes microbians i les citocines produïdes per les cèl·lules de la immunitat innata
indueixen l’expressió de molècules coestimuladores a les cèl·lules presentadores d’antigen.
En absència de coestímul, els limfòcits T que reconeixen l’antigen no hi responen, pel que
entren en estat d’anèrgia.
Quan es produeix la interacció de CD28 amb el seu lligand es genera tota una cascada de
senyalització. CD28 té, a la seva cua citoplasmàtica, un motiu de tirosina que permet la
interacció amb la proteïna reguladora de la fosfoinositol 3 fosfat quinasa, 2 motius rics en
prolina que permeten la unió de la tirosina quinasa ITK i de la quinasa LcK. Quan CD28
interactua amb els seu lligand (B7-1 i B7-2), fa servir GEF i activa Rac. Rac activarà ña via
de les MAP quinases. Això durà a l’activació de la quinasa Jnk, que es fosforila. Al estar
activada, Jnk fosforila Jun, que entrarà al nucli per unir-se a FOS i formar AP-1, que indueix
la transcripció de gens.
La cascada de les MAP quinases activada pel segon senyal d’activació comença per GEF,
segueix per Rac, fosforila MAPKKK, aquesta a MAPKK i aquesta a MAPK. Finalment hi ha
una translocació al nucli.
La suma del primer senyal d’activació, mitjançant Ras, dona lloc a FOS activat al nucli i el
segon senyal d’activació, mitjançant Rac, dona lloc a Jun activat al nucli. Tots dos formen AP-
1. Per tant, l’acció dels dos senyals d’activació fan, en comú, l’activació del factor AP-1.
NFAT, AP-1 i NFκB s’uneixen al promotor del gen de la IL2. Cal que s’hi uneixin tots perquè
comenci la transcripció del gen. Per tant, es pot considerar que el sistema d’expressió de la
IL2 està molt regulat.
Interleucina 2.
Les citocines desenvolupen funcions clau en les respostes immunitàries adaptatives. En elles,
les principals fonts de citocines són els limfòcits T. La citocina més important produïda pels
limfòcits T després de la seva activació és la Interleucina-2. La IL-2 és un factor de
creixement, supervivència i diferenciació pels limfòcits T. Desenvolupa una funció
important en la regulació de les respostes dels limfòcits T en virtut del seu paper crucial
en el manteniment dels limfòcits T reguladors. La Interleucina 2 actua a les mateixes
cèl·lules que la produeixen o a limfòcits T adjacents.
L’activació dels limfòcits T pels antígens i els coestimuladors estimula la transcripció
del gen de la IL-2 i la síntesi i secreció de la proteïna. La producció de IL-2 és ràpida i
transitòria. Els limfòcits T secreten la Interleucina a la interacció entre ells i les cèl·lules
presentadores d’antigen. Els receptors per les citocines dels limfòcits també tendeixen a
localitzar la interacció, pel que la citocina i el seu receptor arriben a concentracions locals
prou altes com per iniciar les respostes cel·lulars.
Els receptors funcionals per a la IL-2 s’expressen de forma transitòria en l’activació
dels limfòcits T verges i efectors. Els limfòcits T reguladors sempre expressen
receptors per la Interleucina. El receptor per la IL-2 (IL-2R) consta de tres proteïnes
associades de forma no covalent: IL-2Rα, IL-2/15Rβ i γc; alternativament anomenades CD25,
CD122 i CD132 respectivament.

Immunologia 2020
CEBIO 160
La IL-2 s’uneix a la cadena α sola amb una afinitat baixa i això no produeix cap senyal
citoplasmàtica ni cap resposta biològica detectable. IL-2/15Rβ contribueix a la unió de la
IL-2 i connecta amb les vies de transducció de senyals. Malgrat la cadena γc no participa
directament la unió amb IL-2, es necessita la seva associació al complex del receptor per a
la unió amb alta afinitat a la IL-2 i per a l’activació completa de les vies de transducció del
senyal.
Els complexes IL-2/15Rβγc s’expressen en baixes quantitats en els limfòcits T en repòs
i s’uneixen a la IL-2 amb una Kd d’aproximadament 10-9M. Les cèl·lules que expressen IL-2Rα
i formen complexes IL-2Rαβγ poden unir-se a la IL-2 amb més força, amb una Kd al voltant
de 10-11M, i l’estimulació del creixement d’aquestes cèl·lules es produeix a concentracions
similars de IL-2.
La IL-2, produïda en resposta a l’estímul d’un antigen, és un estímul per la inducció de IL-
2Rα, el que proporciona un mecanisme pel qual les respostes dels limfòcits T
s’amplifiquen a si mateixes.
Expressió de Gens per l’activació.
A la gràfica següent es mostra la cinètica aproximada de l’expressió d’algunes molècules
després de l’activació dels limfòcits T per antígens i coestimuladors. Els exemples il·lustratius
comprenen un factor de transcripció (c-Fos), una citocina (IL-2) i proteïnes de la superfície
(CD69, IL-2Rα, lligand de CD40). Aquestes proteïnes es solen expressar en baixes quantitats
als limfòcits T naive i les indueixen senyals activadores. El CTLA-4 s’indueix al cap d’una
setmana de l’activació. Les cinètiques són estimacions i variaran amb la naturalesa de
l’antigen, la seva dosi i la seva persistència.
Presentació d’antigen i reconeixement. Perquè es produeixi l’activació hi ha d’haver un encaix entre un limfòcit T i la cèl·lula
presentadora d’antigen. Això requereix de moltes interaccions i de molt bones, ja que han
de ser prou estables per tal que el limfòcit T desencadeni tots els mecanismes que li
pertoquen. Aquesta interacció requereix d’una reestructuració de les proteïnes de membrana
a la que anomenem sinapsi immunològica.
La sinapsi immunològica és la superfície de contacte entre la cèl·lula presentadora
d’antigen i el limfòcit T. Quan es produeix la primera interacció entre LFA-1 i ICAM-1, que
permet que s’acostin la cèl·lula presentadora d’antigen i el limfòcit T, es pot donar la interacció
entre el TCR i el complex pèptid/MHC. Quan això passa, el TCR migra cap a l’interior de
les balses lipídiques, on hi havia gran quantitat molècules d’adhesió, i aquestes se
separen de la regió central i van cap al voltant del punt d’interacció. Això és perquè
impedirien el bon contacte entre el TCR i el complex pèptid/MHC al ser molt llargues. Enlloc
d’això, es reorganitzen les proteïnes de membrana per tal que els complexes quedin al centre
i les molècules d’adhesió es quedin fora, estabilitzant. En la sinapsi hi participen altres
molècules.
Les molècules del limfòcit T que es mobilitzen ràpidament al centre de la sinapsi són el
complex del TCR, els coreceptors CD4 o CD8, receptors per coestimuladors, enzims quinases
i proteïnes adaptadores que s’associen a les cues citoplasmàtiques dels receptors
transmembranals. En aquesta posició de la sinapsi, anomenada c-SMAC, la distància entre
la membrana plasmàtica del limfòcit T i la de la cèl·lula presentadora d’antígens és d’uns
15nm. Les integrines es mantenen a la perifèria de la sinapsi, on funcionen estabilitzant la
unió del limfòcit T a la cèl·lula presentadora d’antigen, i formen la porció perifèrica del SMAC

Immunologia 2020
CEBIO 161
anomenada p-SMAC. En aquesta part externa de la sinapsi, les dues membranes estan
separades uns 40nm. La part externa rep el nom de d-SMAC.
El TCR i les molècules senyalitzadores (LAT)
formen nanoagregats homotípics a limfòcits
T en repòs. Amb la interacció amb l’antigen, el
TCR i les molècules senyalitzadores s’agrupen
formant agregats de mida més gran,
microagregats. Els microagregats es fusionen
per formar el complex supramolecular central o
cSMAC a la zona de contacte entre cèl·lules T i
cèl·lules presentadores d’antigen.
La membrana plasmàtica és dinàmica i els receptors d’antigen i les molècules de
senyalització es desplacen. Aquest desplaçament és en funció del metabolisme de la
cèl·lula, de si el limfòcit està activat, si està en repòs... La zona d’interacció entre el limfòcit
T i la cèl·lula presentadora d’antigen varia al cap del temps. Als 15 minuts, el TCR es
troba localitzat en un anell perifèric mentre que LFA-1 es troba al centre. És el que s’anomena
una sinapsi immunològica immadura. Als 30 minuts, en canvi, el TCR és al centre i es troba
envoltat per un anell perifèric de LFA-1. És el que s’anomena una sinapsi immunològica
madura.
La formació de la sinapsi immunològica permet el canvi de conformació de la LFA-1 a
la forma d’alta afinitat. Aquest canvi estabilitza la interacció cèl·lula T – Cèl·lula
presentadora d’antigen. Les principals interaccions implicades en la immobilització depenen
de l’actina del citoesquelet del TCR i de la integrina a la sinapsi immunològica, incloent-hi els
factors que contribueixen a la desactivació de LFA-1. El SLP-76 activat facilita l’acostament
de kindlin-3 i RIAM a la integrina. Rap/RapL/RIAM promouen l’associació de talin amb la
cadena β del LFA-1. El resultat de l’associació és el canvi conformacional cap a la forma d’alta
afinitat de la LFA-1.

Immunologia 2020
CEBIO 162
Tema 19 – Resposta cel·lular T. Mecanismes Efectors. Els limfòcits T, en reconèixer antigen presentat per la cèl·lula presentadora d’antigen,
s’activen. L’activació d’un limfòcit T el porta a produir IL-2, pas amb què comença la
proliferació i expansió clonal. A partir del procés d’expansió, els limfòcits es converteixen
en els diferents tipus que n’hi ha. Els limfòcits sortints de la diferenciació es separen en
limfòcits T efectors i limfòcits T de memòria.
El tercer senyal d’activació ve donat per citocines i molècules d’adhesió. Les citocines
en són l’element més rellevant, però, ja que determinen el tipus de limfòcit T efector.
Diferenciació a limfòcits T CD4+ efectors. El procés de diferenciació o polarització consisteix en 3 etapes: inducció, compromís
i amplificació. La inducció permet una expressió dels factors de transcripció específics de
cada tipus efector. Els factors determinaran el patró de citocines que secretarà cada tipus de
limfòcit T efector. En l’etapa compromís s’estabilitza el fenotip de limfòcit T efector perquè es
donen una sèrie de canvis epigenètics en les zones reguladores dels gens de les citocines.
La regulació porta a una producció estable d’algunes citocines. Finalment, en l’amplificació
les citocines secretades pels limfòcits T promouen la diferenciació de més cèl·lules del mateix
tipus efector i inhibeixen la diferenciació d’altres tipus efectors.
D’alguna manera, per tant, s’estabilitza el fenotip efector i s’inhibeix la diferenciació de
la resta de tipus de cèl·lules T efectores. En aquest cas parlem de cèl·lules T helper
polaritzades. És un fet que es veu, sobretot, en limfòcits T helper perquè hi ha diferents tipus
efectors. No ho veiem en els limfòcits TCD8 que, malgrat parlar de limfòcits T efectors, no se
n’han descrit diferents tipus efectors. Així i tot, hi ha estudis que indiquen que hi ha diferents
tipus de limfòcits TCD8, com els reguladors.
Canvis epigenètics i citocines.
Els canvis epigenètics, com modificacions de les histones o de la metilació del DNA, indueixen
canvis en l’estructura de la cromatina. Això deixa accessibles les zones reguladores de
davant del promotor dels gens de les diferents citocines als factors de transcripció de
DNA. Aquests canvis estabilitzen els programes d’expressió gènica durant la resposta T.
Per tant, en un limfòcit TCD4 que s’està convertint en un limfòcits T helper 2, el locus on es
troba la interleuquina 4 quedarà accessible als factors de transcripció. Quan es produeixi la
diferenciació, produirà aquesta interleuquina. Això li permetrà diferenciar-se i provocar la
diferenciació d’altres limfòcits en limfòcits T helper 2.
El que determina en quin tipus de limfòcit T efector es diferencia un limfòcit T són els
factors de transcripció de llinatge cel·lular, el quals venen determinats per les citocines.
Per tant, es pot afirmar que les citocines determinen en quin tipus de limfòcit T efector s’ha
de diferenciar una cèl·lula T.
Quan un limfòcit T reconeix antigen a través del TCR, comença tota la cascada de
senyalització associada. Aquesta cascada, juntament amb la que prové de la interacció de la
IL-4 amb el seu receptor (que produeix l’activació del factor de transcripció Stat-6), fa que el
limfòcit expressi el factor de transcripció Gata3. El factor de transcripció Gata3 és el que fa
possible que el limfòcit T es converteixi en un que és productor de les interleuquines 4, 5 i 13.
Per tant, aquest limfòcit T secretarà les mencionades citocines. La interleuquina 4 actuarà de
forma autocrina, reforçant el fenotip Th2.
En canvi, si quan es dona la interacció TCR-MHC/pèptid hi ha interleuquina 12 al medi, aquest
s’unirà al seu receptor expressat a la membrana i activarà el factor de transcripció Stat-4. En

Immunologia 2020
CEBIO 163
conseqüència, hi haurà una expressió de T-Bet, un factor de transcripció que estimula la
transcripció del gen de l’interferó γ, provocant-ne l’expressió. Aquest interferó serà expulsat
al medi on de manera autocrina s’unirà al receptor del limfòcit T, activarà Stat-1 i reforçarà
l’expressió de T-Bet. T-Bet determina que el limfòcit sigui un T helper 1.
T-bet impedeix la diferenciació a Th2 inhibint Gata3. Gata3 impedeix la diferenciació a Th1
inhibint l’expressió de IL-12Rβ2 i Stat4.
L’antigen i l’entorn determinen com són les cèl·lules presentadores d’antigen i quines
citocines secreten. El que determina quines citocines faran les cèl·lules T és l’antigen i on
es trobi aquest. Segons com estigui funcionant la immunitat innata, la cèl·lula dendrítica
produirà unes o altres citocines. En conseqüència, la cèl·lula presentadora d’antigen
determinarà en quin tipus de limfòcit es tronarà aquell que s’ha de diferenciar.
En presència de IL-12 i Interferó gamma, les cèl·lules T naive es diferenciaran en Limfòcits T
helper 1. En presència de IL-4, es diferenciaran en T helper 2. En cas que no hi hagi PAMPS
però sí que hi hagi IL-10 i TGFβ, els limfòcits T passaran a ser limfòcits T reguladors.
Per tant, es pot dir que la immunitat innata influeix en la adaptativa i que aquest és un dels
mètodes que té per fer-ho.
Diferenciació de limfòcits TCD4 helper efectors.
L’activació via TCR en funció de la presència de citocines produïdes determina el tipus
de limfòcit T efector. Els caracteritzem per l’expressió dels factors de transcripció. La
diferenciació d’un tipus efector inhibeix la diferenciació dels altres. Es dona el cicle de
potenciació del tipus efector diferenciat.
Les citocines actuen com a 3er senyal tot induint l’expressió de diferents factors de
transcripció (FoxP3, RORgT, T-bet, GATA-3). Els factors determinen el desenvolupament a
diferents tipus de limfòcits TCD4 efectors. El tercer senyal fa possible la diferenciació dels
limfòcits T CD4 activats a diferents tipus de limfòcits T helper o col·laboradors efectors.
La rellevància que, a partir d’un limfòcit T naive i d’una cèl·lula presentadora d’antigen hi hagi
una diferenciació en limfòcits T helper 1, T helper 2, T helper 17 o T reguladors induïts; és
que cada tipus cel·lular de limfòcit T serà important per la resposta a diferents tipus
d’antigen.
Els Limfòcits T helper 1, per exemple, són molt importants en l’activació dels limfòcits TCD8
citotòxics i són molt rellevants en la resposta a patògens intracel·lulars. Els limfòcits T helper
2 seran rellevants en la resposta a patògens extracel·lulars i per ajudar a activar limfòcits B.
Els limfòcits T helper 17 són un nucli d’infecció i tenen molta importància en les respostes a
la pell i mucoses, així com als fongs. Els limfòcits T reguladors induïts destaquen per la seva
funció en el manteniment de la tolerància i la finalització de la resposta.
Diferenciació de limfòcits TH1.
Els limfòcits T helper 1 són importants en la resposta a patògens intracel·lulars. Quan els
limfòcits T es diferencien en presència de IL-12 i interferó γ, es converteixen en
limfòcits T helper 1. Aquests destaquen per la producció tant de IL-12 com de INFγ.
Els limfòcits T helper 1 són importants en la resposta a patògens intracel·lulars perquè
una de les seves funcions principals és induir l’activació dels macròfags. L’interferó
gamma també influeix en el canvi d’isotip a IgG.
Els limfòcits T helper 1 provoquen l’activació dels macròfags per acció de l’IFN-γ i la
interacció CD40L-CD40. Aquesta interacció augmenta la producció de ROS i RNS, pel que

Immunologia 2020
CEBIO 164
s’obté més capacitat microbicida. A més, els macròfags secreten les citocines TNF i IL12 i
quimiocines. Finalment, els limfòcits T helper 1 provoquen un augment de l’expressió de
molècules del MHC i de molècules coestimuladores (B7).
Els limfòcits Th1 també contribueixen a la bona diferenciació dels limfòcits TCD8 cap
a limfòcits TCD8 efectors arran de la seva producció de citocines. També secreten IL-3 i
Factor estimulador de colònies de monòcits i macròfags, pel que també indueixen la
diferenciació de macròfags i de diferenciació de les cèl·lules hematopoètiques a macròfags.
Amb la secreció de TNF-α, s’estimulen les cèl·lules endotelials i s’augmenta l’extravasació de
limfòcits T al lloc de la inflamació i infecció, facilitant el dur a terme la seva funció. També
produeixen la quimiocina CCL2, que atreu macròfags. D’alguna manera, doncs, els Th1
indueixen participació macròfags en la resposta. Tenen un mecanisme on l’expressió de fas
provoca la mort de cèl·lules. Els Th1, amb expressió de lligand fas quan estan activats, poden
provocar la mort de la cèl·lula diana.
Diferenciació de limfòcits TH2.
Els limfòcits Th2, en presència de IL4 produïda per mastòcits o eosinòfils en resposta a
patògens extracel·lulars com helmints, es diferencien. Es converteixen en limfòcits T
helper productors de IL-4, IL-5 i de IL-13. Tenen funció important en la producció de
determinats isotips d’immunoglobulines com IgE i en l’activació eosinòfils i de secrecions
mucoses. Molt rellevants en resposta a patògens extracel·lulars.
Les funcions dels limfòcits Th2 es duen a terme perquè la interleuquina 4 indueix el canvi
d’isotip d’immunoglobulina a IgE i, per tant, facilita la sensibilització de mastòcits. A
més, estimula la producció d’anticossos neutralitzants. També és possible gràcies a la
producció de IL-5, que activa eosinòfils, i a la producció de IL-13. Aquesta última interleuquina
es combina amb IL-4 per augmentar el peristaltisme intestinal i la secreció mucus així com
per afavorir l’activació de macròfags M2, que contribueixen a la reparació del teixits.
En conseqüència, es pot afirmar que la resposta dels limfòcits Th2 és una molt associada
a les mucoses gàstrica i respiratòria, on es produeixen les al·lèrgies.
Diferenciació de limfòcits TH17.
Els Th17 són aquells que es diferencien quan a l’entorn hi ha IL-6, IL-1 i TGFβ. Aquestes
citocines indueixen producció IL-21, que és qui estabilitza fenotip juntament amb IL-23.
Aquestes cèl·lules Th17 produeixen IL-17 i IL-22. IL-17 té un paper important en inflamació.
IL-22 té un paper important en barrera al estimular producció pèptids antimicrobians. Limfòcits
Th17 són rellevants en activació de neutròfils i de cèl·lules epitelials. Són molt importants a la
resposta a patògens extracel·lulars com bacteris i, sobretot, fongs.
Els limfòcits Th17 produeixen IL-17 i IL-22. IL-17 genera una resposta proinflamatòria
potent al induir l’expressió de citocines proinflamatòries com IL-1, IL-6 i TNFα; de quimiocines
(CXC) que indueixen la quimiotaxi de leucòcits, sobretot neutròfils, afavorint el manteniment
de la inflamació i infiltració de leucòcits al teixit; de factors creixement per a neutròfils com G-
CSF i al induir també un augment d’expressió d’ICAM-1 als teixits. La combinació de IL-17
i de IL-22 indueix la secreció de pèptids antimicrobians per part de cèl·lules epitelials.
Un quart tipus de limfòcits T helper són els fol·liculars. Són essencials per la resposta
dels limfòcits B2 fol·liculars. Els limfòcits B2 fol·liculars són els més típics, els que es troben
als fol·licles limfoides al limfonode o a la melsa i que, a través de la col·laboració amb els
limfòcits T helper fol·liculars, desenvolupen la seva funció. Per tant, es pot dir que els
Limfòcits T helper fol·liculars contribueixen a l’activació dels limfòcits B, a la
Hipermutació Somàtica i al canvi d’isotip.

Immunologia 2020
CEBIO 165
Els limfòcits T helper fol·liculars, quan s’activen en presència d’IL-6 i IL-21, fan que s’expressi
BSL-6. Aquest potencia la producció de IL21. La coestimulació via interacció ICOS-ICOSL és
essencial per mantenir el fenotip. Els limfòcits T helper fol·liculars acompanyen limfòcits T
secundaris als fol·licles quan s’està produint la resposta.
Limfòcits Treg.
Els limfòcits T reguladors serveixen per regular l’activació i acció de les cèl·lules T
efectores. Es diferencien a partir precursors al timus, quan reben el nom de reguladors tímics.
Els limfòcits T reguladors sempre hi són arreu i mantenen l’estat de no resposta a la perifèria.
Quan es dona una resposta immunitària, es generen limfòcits T reguladors en presencia
d’antígens microbians, TGFβ, IL-2, àcid retinoic i àcids grassos de cadena curta. Les cèl·lules
reguladores es caracteritzen per produir, algunes d’elles, FoxP3. Són cèl·lules importants per
finalitzar la resposta immunitària.
Les cèl·lules T reguladores expressen IL-10 i TGFβ per aturar la inflamació. També
expressen PD1 i CTLA4. La secreció d’aquests compostos permet als limfòcits T reguladors
dur a terme la seva funció d’aturar la resposta immunitària. Els Treg tenen una funció molt
important en la resposta a patògens. S’ha vist que poden inhibir la funció de limfòcits T
efectors, inhibint la resposta a tumors. Això incentiva el desenvolupament d’aquest.
Mecanisme efector dels limfòcits T helper. Els patrons de citocines secretat és el mecanisme efector dels limfòcits TH i determinen
la principal funció efectora de cada subtipus cel·lular. Th1, per exemple, també produeix
IL-3 o GM-CSF, que promouen la diferenciació dels precursors dels limfòcits T. El patró de
secreció de citocines i d’expressió de receptors de quimiocines proporciona funcions diferents
a cada tipus efectors de T helper durant la resposta immunològica.
Paral·lelament a la producció de citocines associades a tipus de limfòcit T helper,
cadascun expressa un diferent patró de receptors de quimiocines. Les diferències en
patrons d’expressió de receptors s’equival amb una diferència en els patrons de migració.
Això significa que el lloc on va cada tipus de limfòcit T efector és diferent perquè les
quimiocines es produeixen a llocs diferents. El conjunt del patró de citocines secretades
més el patró de receptors de quimiocines determina la capacitat funcional final de la
cèl·lula T helper.
Diferenciació de limfòcitsTCD8 efectors. Malgrat no es coneixen diferents tipus de limfòcits TCD8 efectors, es parla de limfòcits
TCD8 naive. Aquest limfòcit TCD8 naive no ha reconegut antigen. En el moment que ho fa,
si es donen les condicions adequades de suficient senyalització, intervenen el primer i segon
senyals d’activació amb col·laboració del coreceptor i s’activa el limfòcit.
L’activació li permet proliferar i diferenciar-se a limfòcit T efector. Hi ha diferents estadis
en aquest procés. Canvia expressió receptor IL-2 al aparèixer la cadena α, comença a
expressar poca IL-2, es desenvolupa la capacitat de proliferació i capacitat citotòxica dels
grànuls amb material citotòxic.
Quan es produeix una baixa inflamació, l’activació dels limfòcits TCD8 requereix la presència
de limfòcits TCD4 que responguin al mateix antigen. Les cèl·lules TCD4 funcionen
“activant” les cèl·lules dendrítiques que presentaran l'antigen als limfòcits TCD8 i
interaccionant directament amb limfòcits TCD8 mitjançant l'expressió de CD40 o la producció
d’IL2. Si es produeix una alta inflamació, els limfòcits TCD8 es poden activar en
absència de cèl·lules TCD4 però no es genera una bona memòria T.

Immunologia 2020
CEBIO 166
Dendritic cell Licensing. La necessitat de TCD4 en l’activació ve donada per l’acció o efecte d’aquests sobre la
cèl·lula presentadora d’antigen. És el que s’anomena activació de les cèl·lules
dendrítiques. Aquest concepte defineix el procés mitjançant el qual les cèl·lules
presentadores d’antigen, que solen expressar un baix nivell de molècules coestimuladores,
tenen un contacte amb els limfòcits TCD4 mitjançant el reconeixement del complex
pèptid/MHC i CD40/CD40L i això suposa un augment en la presentació de molècules
coestimuladores. Amb el cell licensing és possible l’activació de CD8 i l’expressió de IL-12
per activar i diferenciar CD8.
Si el limfòcit TCD4 i el TCD8 es troben alhora amb la cèl·lula dendrítica, les citocines
secretades per el TCD4 estimulen la diferenciació del TCD8 efector. Si no es troben
alhora, la interacció de la TCD4 amb la cèl·lula dendrítica li proporciona els senyals per a
l’activació amb l’augment d’expressió de molècules coestimuladores i de producció de
citocines, sobretot IL-12, es diu dendrític cell licensing.
El dendrític cell licensing és important per l’activació dels limfòcits TCD8 naive. En cas
que estiguem parlant de limfòcits TCD8 de memòria, requereixen menys ajuda i la cèl·lula
dendrítica madura pot activar-les directament. Als limfòcits TCD8, el llindar d’activació és més
baix i necessita menys molècules estimuladores a la superfície de la cèl·lula dendrítica. Les
cèl·lules TCD8 de memòria produeixen IL-2 en rebre el 1er i 2on senyal, mentre que les
cèl·lules TCD8 naive necessiten les cèl·lules TH1 com a font de la mateixa citocina.
S’ha vist que senyalització en presència PAMPs fa augmentar presència molècules
coestimuladores CD80 i CD86 a cèl·lules dendrítiques. Això facilita activació limfòcits
TCD8.
Mecanisme efector de la citotoxicitat dels limfòcits TCD8. Els limfòcits TCD8 indueixen la mort cel·lular per apoptosi. Significa que la cèl·lula que
presenta l’antigen en complex amb el pèptid del MHC de classe I mor. En aquesta mort, el
limfòcit TCD8 indueix:
• Una pèrdua de potencial de membrana mitocondrial.
• Una alliberació de citocrom C.
• Una condensació nuclear.
• Una externalització de fosfatidilserina.
• Una activació de les caspases que provoquen la fragmentació del DNA.
A nivell morfològic, hi ha un blebbing nuclear i una alteració general de la morfologia
cel·lular que genera cossos apoptòtics. Un blebbing és una fragmentació i reorganització
de la cromatina del nucli.
Això és així perquè els limfòcits TCD8 alliberen el contingut dels seus grànuls a zona
contacte entre les dues cèl·lules. Aquest grànuls lítics contenen perforina, granzima i
granulisina. La perforina forma porus a la membrana que permeten entrada d’aigua i sals
provocant un trencant de l’equilibri osmòtic. Les granzimes són serina proteases que activen
caspases del citoplasma per activar l’apoptosi. La granulisina que allibera Ca2+ intracel·lular i
citocrom c.
La granzima activa la caspasa 8 que, directament o a través de l’alteració del potencial de
membrana a mitocondris que allibera el citocrom C i activa la caspasa 9, activa caspasa 3.
La caspasa 3 trenca el DNA doble cadena i provoca la mort cel·lular.

Immunologia 2020
CEBIO 167
El procés d’inducció de la mort cel·lular és molt ràpid. Entre l’inici de la interacció i la
inducció de la mort cel·lular passen menys de 7 minuts. En aquests 7 minuts, el limfòcit TCD8:
1) S’acosta a la cèl·lula amb l’antigen.
2) Interacciona pel complex del TCR amb el complex pèptid/MHC.
3) Desgranula sobre cèl·lula diana.
4) Provoca la formació de porus.
5) Fa canviar la morfologia de la cèl·lula.
6) Indueix la mort.
Hi ha altre mecanisme efector TCD8. Resultat interacció fas i lligand fas. Es produeix en
Th1 activats. Limfòcits TCD8 expressen lligand fas. Moltes cèl·lules organismes expressen
fas. Interacció Fas-lligand fas té conseqüències. Interacció activa domini FADD associada a
fas. FADD té dominis de mort que recluten caspasa 8 inactiva i indueixen activació. Caspasa
8 activa caspasa 3 que produeix trencament DNA i mort cel·lular.
L’alliberació del contingut dels grànuls es dona a la superfície de contacte entre el
limfòcit TCD8 i la cèl·lula diana. La cèl·lula TCD8 mata específicament les cèl·lules que
expressen l’antigen pel qual aquest limfòcit TCD8 és específic. És, doncs, un mecanisme molt
diferent del que tenen les cèl·lules de la immunitat innata, que alliberen el contingut dels
grànuls a l’exterior i afecten a totes les cèl·lules del voltant. El mètode que fan servir els
limfòcits TCD8 és específic, per això requereix que l’alliberament sigui a la zona de contacte.
La citotoxicitat dels TCD8 depèn de la sinapsi immunològica. Fa que l’acció dels limfòcits
es produeixi només en la cèl·lula amb la que s’està interaccionant. Per tant, és específic.
Limfòcits T citotòxics. Les cèl·lules T citotòxiques maten selectivament les cèl·lules diana que expressen
l’antigen pel qual són específiques. L’alliberament de substàncies citotòxiques només es
dona a la zona de contacte entre el limfòcit i la cèl·lula diana.
Hi ha limfòcits T naive, que es diferencien. Una part ho fan a limfòcits T de memòria.
Responen de manera més ràpida i efectiva al mateix antigen quan aquest es presenta per
segona vegada a l’organisme. Els limfòcits T de memòria persisteixen en absència antigen.
Es mantenen durant temps llargs als teixits. Ho fan gràcies a IL-7 i IL-15. Hi ha diferents
limfòcits t memòria.
Limfòcits T de memòria.
Els limfòcits T de memòria s’activen més ràpid i de manera més eficient perquè tenen
menys requeriments d’activació. Necessiten menys senyals d’adhesió, no necessiten una
alta intensitat al 1r i al 2n senyal ni tampoc necessiten una alta concentració de molècules
coestimuladores. Aquests criteris s’apliquen tant als limfòcits TCD4 com als TCD8.
Hi ha diferents tipus de cèl·lules T de memòria que difereixen per la seva localització.
Els limfòcits T de memòria centrals expressen el receptor CCR7, que provoca l’entrada als
òrgans limfàtics secundaris com els limfonodes on queden retinguts a limfonodes. També
expressen CD62L, que hi facilita la entrada. Els limfòcits T de memòria efectors no expressen
CCR7 i tenen un baix nivell d’expressió de CD62L. Un altre tipus de limfòcits T de memòria
que existeixen són els residents als teixits. Aquests limfòcits T de memòria no expressen
CCR7 ni CD62L, però expressen CD103 i CD11a.
Això fa que diferents limfòcits tinguin diferent capacitat de homing, és a dir, que es
troben en localitzacions concretes. Els limfòcits T de memòria centrals es troben en òrgans
limfàtics secundaris. Per contra, els limfòcits T de memòria efectors es troben en teixits no

Immunologia 2020
CEBIO 168
limfoides o a la melsa. Finalment, els limfòcits T de memòria residents als teixits es troben en
teixits amb mucosa com poden ser els pulmons, el tracte urogenital o a la pell. Els limfòcits T
efectors de memòria actuen als teixits mentre que els de memòria centrals són un reservori
per generar altres limfòcits T de memòria si el sistema immunitari no dona la talla.
Les diferències de localització dels limfòcits T de memòria també es tradueix en una
diferenciació funcional.
Els limfòcits T amb receptor γδ també en tenen d’efectors. També en cèl·lules NKT.
Tenim cèl·lules T γδ amb capacitat citotòxica i col·laboradors. Patrons producció citocines
determinen participació en diferents processos de la resposta immunitària.
Cèl·lules NKT. Hi ha 5 llinatges descrits de cèl·lules iNKT, que es generen a partir de les cèl·lules
precursores d'iNKT. Cada llinatge es troba diferenciat terminalment per l’expressió d'una
selecció única de citocines i factors de transcripció quan s’activen.
• NKT1 expressen alts nivells de Tbet, secreten IFNγ i algunes IL-4.
• NKT2 tenen expressió continua de PLZF i secreten principalment IL-4 i IL-13.
• NKT17 expressen RORgt i secreten IL-17.
• NKTFH expressen Bcl6 i es desenvolupen després de l'activació. No està clar si es
desenvolupen a partir de precursors NKT1.
• NKT10 es desconeixen els factors de transcripció que regulen NKT10 però es
defineixen com a secretores d’IL10, expressen NRP-1 i PD1.

Immunologia 2020
CEBIO 169
Tema 20 – Activació de la Resposta Humoral. La resposta immunitària humoral és la Resposta immunitària vehiculada per anticossos.
Subpoblacions de limfòcits B. Els limfòcits B es separen en diferents subpoblacions: limfòcits B1 i limfòcits B2. Els
limfòcits B1 són els més abundants a pleura i cavitat peritoneal i generen el fetge fetal durant
el desenvolupament embrionari. Es caracteritzen per tenir un repertori de BCRs reduït i solen
tenir i produir IgM.
Els limfòcits B2, en canvi, tenen un repertori ampli de BCRs i en distingim 2 grups: limfòcits
B2 fol·liculars i limfòcits B2 marginals. Els limfòcits B2 fol·liculars són els típics de la
resposta adaptativa, són la majoria de limfòcits B madurs, responen a antígens proteics i
depenen de la col·laboració amb els limfòcits T. Els limfòcits B2 de la zona marginal es
troben a la zona marginal de la melsa, responen a antígens provinents de la sang, són
independents de la col·laboració amb els limfòcits T i responen ràpid secretant IgM.
Activació dels limfòcits B. Per a una activació complerta dels limfòcits B també necessitem més d’un senyal, en aquest
cas en són 2. El primer senyal ve donat per la interacció entre l’antigen i el BCR. El segon
senyal d’activació són una sèrie d’interaccions que ajuden a l’activació completa dels limfòcits
B. Els segons senyals són diferents segons el tipus d’antigen i el tipus de limfòcit B.
1er senyal d’activació.
El primer senyal d’activació és el que ve donat per la interacció entre el BCR i l’antigen.
Aquesta interacció produeix tota una sèrie de canvis i moviment en aquest BCR. A les
cèl·lules B en repòs, el BCR es troba expressat a la membrana en forma de monòmers
associat a les cadenes Igα/Igβ, les quals tenen motius ITAM a les cues citoplasmàtiques.
Quan el BCR interacciona amb l’antigen a través d’algun dels seus braços, es produeix un
canvi conformacional al domini constant 4 de la cadena pesada. Això fa exposar la superfície
de contacte, permetent que es formin agregats de BCRs. En unir-se a l’antigen també es
produeix l’obertura dels dominis citoplasmàtics de Igα/Igβ. Aquesta conformació permet que
comenci la senyalització.
La unió d’antigen permet el crosslinking o entrecreuament del BCR. L’entrecreuament és la
formació d'un complex antigen-anticòs en què múltiples elements d'un mateix antigen, amb
més d'un determinant antigènic, s'uneixen als BCR corresponents, de manera que alguns
antígens queden units a més d'una molècula d'immunoglobulina.
Hi ha un tercer canvi quan hi ha una interacció amb l’antigen. Els oligòmers que es formen es
desplacen cap a les balses lipídiques o lipid rafts, on les cadenes Igα i Igβ poden interaccionar
amb tirosina quinasa (Lyn) i iniciar al senyalització a través dels seus motius ITAM.
Interacció BCR-antigen.
La fosforilació de les tirosines dels motius ITAM de la cua citoplasmàtica de les cadenes
Igα i Igβ per la quinasa Lyn permet que es recluti la quinasa Syk. Syk fosforila les quinases
BTK i BLNK. Btk contribueix a l’activació de BLNK, que crea els llocs d’unió per a la
fosfolipasa Cγ2 i per al GEF (el factor intercanviador de nucleòtids de guanina) anomenat
VAV. VAV indueix una remodelació del citoesquelet, pel que es canvia la morfologia de la
cèl·lula B. La fosfolipasa Cγ2 trenca el fosfoinositol-2-fosfat de la membrana citoplasmàtica
en diacilglicerol i en inositol-3-fosfat. L’activació de GEF també inicia la via de les MAP
quinases per part de les proteïnes G petites.

Immunologia 2020
CEBIO 170
L’inositol-3-fosfat fa augmentar la concentració de cations de calci al citoplasma. El calci
permet activar, a través de la calmodulina, la calcineurina. La calcineurina activa el factor de
transcripció NFAT, que es transloca al nucli. El diacilglicerol, a través de la fosfoquinasa C,
activa NFκB, que també es transloca al nucli. La fosfoquinasa C també contribueix a
l’activació del factor de transcripció Jul i la formació de Jun, que van al nucli.
Coreceptor activador del BCR.
De la mateixa manera que els limfòcits T tenen coreceptors que ajuden al TCR, els limfòcits
B tenen coreceptors que contribueixen a l’acció del BCR. En el cas dels limfòcits B hi ha
un coreceptor activador format per 3 proteïnes: CD19, CD21 i CD81. La contribució d’aquest
coreceptor és imprescindible perquè es produeixi l’activació completa del limfòcit B.
CD19 és una proteïna transmembrana el domini extracel·lular de la qual està format per
dominis immunoglobulina i té una cua citoplasmàtica amb un motiu ITAM que fa servir per
amplificar el senyal d’activació del BCR. CD21 és un receptor del fragment C3d de la proteïna
del sistema del complement. C3d és un fragment de C3. CD21 també rep el nom de CR2.
CD81 és una proteïna transmembrana associada a CD21 amb una funció desconeguda en el
marc de coreceptor activador del limfòcits B.
Interacció BCR-Coreceptor-Antigen.
Similar als limfòcits T, el coreceptor amplifica el senyal d’activació. CD21 interacciona amb
el fragment C3d, el qual està opsonitzant l’antigen que està reconeixent el BCR. El
coreceptor s’acosta aleshores al BCR, permetent una fosforilació dels motius ITAM de la
cua citoplasmàtica de CD19 per part de la quinasa Lyn. La fosforilació del motiu ITAM de
CD19 permet l’amplificació del senyal.
La fosforilació del motiu ITAM de la cua citoplasmàtica de CD19 crea el lloc d’unió per a la
quinasa Fosfoinositol-3-quinasa que amplifica la senyalització del BCR al inhibir
l’inhibidor del factor de transcripció NFAT, afavorint-ne l’activació.
Coreceptor inhibidor del BCR.
En condicions de repòs, el BCR està associat a CD22, un coreceptor inhibidor. CD22 és una
lectina que, a la cua citoplasmàtica, té un motiu ITIM. Aquest motiu, quan es troba fosforilat,
genera un lloc d’unió per una fosfatasa que manté desfosforilades les cues citoplasmàtiques
de Igα i Igβ. Aquest manteniment impedeix la transmissió de senyals d’activació a
l’interior de la cèl·lula. CD22, que uneix àcid siàlic, es troba associat o a prop del BCR,
mentre que el coreceptor activador només s’acosta al BCR quan aquest interacciona amb
l’antigen.
La senyalització del TCR i el BCR són semblants.
La transducció de senyal pel TCR i el BCR es basa en el mateix mecanisme general,
usant quinases i adaptadors específics que fan funcions similars i acabant en cascades de
senyalització comparables. Comparant element a element, es poden fer diverses analogies.
El BNLK del BCR és equiparable al SLP-76 del TCR tant com que la quinasa Syk es
correlaciona amb la quinasa ZAP70. A més, la fosfolipasa Cγ1 i la fosfolipasa Cγ2 realitzen la
mateixa funció, com les quinases equivalents Lck i Lyn.
2n senyal d’activació.
El 2n senyal d’activació és diferent segons quin sigui l’antigen i, en conseqüència, segons
quina sigui la subpoblació de limfòcits B que s’està activant. Distingim entre dos tipus de
segon senyal: segon senyal independent de limfòcits T i segon senyal dependent de la
interacció amb limfòcits T.

Immunologia 2020
CEBIO 171
2n senyal independent de limfòcits T.
Quan el segon senyal és independent de limfòcits T és perquè els limfòcits B no
requereixen de la col·laboració de limfòcits T helper per la seva activació i producció
d’anticossos. Els antígens que solen provocar aquest tipus de segon senyal són,
majoritàriament, components de la paret cel·lular, polisacàrids microbians o proteïnes
polimèriques. De segons senyals independents de limfòcits T també hi ha dos tipus: els
Antígens T independents de Tipus 1 (Antígens TI-1) i els Antígens T independents de Tipus
2 (Antígens TI-2).
Resposta de les cèl·lules B a Antígens T independents de Tipus 1.
El segon senyal en resposta a Antígens TI-1 són aquells on el propi antigen ja té activitat
coestimuladora perquè conté components reconeguts per PRRs. Els limfòcits B que reben
aquest segon senyal generen un canvi d’isotip molt poc eficient, pel que quasi tots els BCRs
produïts són IgM. Pràcticament no hi ha hipermutació somàtica.
Quan la concentració d’Antígens TI-1 és baixa, hi ha un limfòcit B que reconeix l’antigen via
BCR i, a més, rep el segon senyal provinent del PRR. Això provoca l’expansió clonal
d’aquest limfòcit B. S’ha vist que, quan els Antígens TI-1 amb activitat coestimuladora
pròpia es troben en una concentració molt elevada, poden donar lloc a l’activació policlonal
i no específica de la cèl·lula B. Això és perquè hi ha una gran quantitat d’aquests
components de l’antigen reconeguts per PRR que poden activar de manera poc eficient els
limfòcits B sense que hi hagi una interacció amb el BCR.
Resposta de les cèl·lules B a Antígens T independents de Tipus 2.
Els segons senyal en el cas de Antígens TI-2 són donat perquè aquests antígens contenen
epítops repetitius que poden estimular directament els limfòcits. És principalment el cas dels
limfòcits B1. Són carbohidrats, polisacàrids, elements repetitius... Els Antígens TI-2 només
poden donar lloc a l’activació de limfòcits B madurs, ja que cal una gran quantitat de
senyalització. En aquest cas, el primer senyal d’activació el dona la interacció de l’antigen
amb molts BCRs alhora, generant un crosslinking massiu. Com que molts BCRs alhora poden
reconèixer l’antigen, es dona un senyal suficient per produir una activació dels limfòcits B.
Sovint, aquest senyal repetitiu permet l’activació del limfòcits B i la producció
d’immunoglobulines M. No hi ha canvi d'isotip. Si en el context de la interacció hi ha citocines,
aquestes poden proporcionar un segon senyal que activi els limfòcits B per tal que es
diferenciïn a cèl·lules productores d’immunoglobulines i que, fins i tot, hi hagi un ineficient
canvi d’isotip.
Resposta dels limfòcits B a Antígens T-dependents i a Antígens T-independents.
Els tres grans tipus de limfòcits B són els limfòcits B2 fol·liculars, els Limfòcits B2 de la zona
marginal i els Limfòcits B1. Els limfòcits B1 i els limfòcits B2 de la zona marginal solen
respondre a antígens T independents. Els limfòcits B2 fol·liculars, en canvi, necessiten la
col·laboració dels limfòcits T per obtenir la màxima activació.
També canvia el que s’aconsegueix. Els limfòcits B2 de la zona marginal, activats
independentment de limfòcits T, produeixen IgM i cèl·lules productores d’immunoglobulines
però de vida curta, com els limfòcits B1. Els limfòcits B2, al necessitar la interacció amb els
limfòcits T, fan molt bé el canvi d’isotip, la hipermutació somàtica i cèl·lules plasmàtiques de
vida llarga.
Resposta dels limfòcits B a antígens T-dependents.
El primer senyal d’activació ve donat per la interacció entre l’antigen i el BCR. El resultat
d’aquesta interacció és un inducció de l’expressió de les molècules coestimuladores CD40,

Immunologia 2020
CEBIO 172
B7 i ICOSL, que serà molt important ja que dins la interacció T-B té un paper similar a la
interacció entre CD40 i el seu lligand. La interacció entre B7 al limfòcit B i CD28 al limfòcit T
és essencial per tal que es doni el primer senyal d’activació.
El primer senyal d’activació potencia l’expressió d’aquestes molècules coestimuladores. El
segon senyal ve donat per la interacció entre CD40 i el seu lligand o entre ICOSL i ICOS. A
més, la cèl·lula T col·laboradora produeix citocines que contribueixen en el segon
senyal i són importants en el canvi d’isotip. Les cèl·lules col·laboradores, amb la producció
de IL-4 i altres citocines, afavoriran la activació i la proliferació dels limfòcits B. El segon senyal
d’activació, doncs, és un conjunt de senyals donats per les molècules coestimuladores i les
citocines.
Col·laboració entre cèl·lules B i cèl·lules T.
La col·laboració entre els limfòcits T i B es pot donar perquè, en aquests casos concrets, els
limfòcits B actuen com a cèl·lules presentadores d’antigen reconegudes pel limfòcit T per
mitjà del TCR. L’antigen és el que el limfòcit B ha reconegut pel BCR. La interacció antigen-
BCR internalitza aquest complex per la via endocítica, generant pèptids que es poden
carregar a molècules del MHC-II i ser expressats a la membrana.
El limfòcit T i el limfòcit B reconeixen diferents epítops del mateix antigen. El resultat de
la interacció és l’augment d’expressió de CD40 a la membrna del limfòcit B per tal que
interaccioni amb el lligand de CD40 dels limfòcits T activats. Juntament amb les citocines,
CD40 reclutarà TRAF i iniciarà la cascada d’activació de NFκB i AP-1. Això proporciona més
senyal d’activació.
La interacció entre CD40 i el seu lligand proporciona el segon senyal d’activació. La interacció
entre B7 i CD28 senyalitza cap l’interior del limfòcit T i li proporciona el seu segon senyal
d’activació. El limfòcit B, fruit de la interacció, expressa receptors de quimiocines per ser més
sensible a l’expressió de les mateixes per part del limfòcit T.
Tema 21 – Mecanismes efectors de la resposta humoral. Hi ha un primer senyal, donat per la interacció antigen-BCR, i un segon senyal provinent de
la col·laboració entre els limfòcits T i B. Quan un limfòcit B reconeix un antigen pel seu BCR,
l’internalitza i el processa per presentar-ne pèptids derivats mitjançant les molècules del MHC
de classe II. Per tant, els limfòcits B poden actuar com a cèl·lules presentadores
d’antigen pels limfòcits TCD4. En aquesta interacció cèl·lula T – cèl·lula B, hi intervenen
CD40 al limfòcit B i CD40L al limfòcit T, proporcionant un senyal de coestimulació. A més,
aquest senyal augmenta l’expressió de receptors de citocines.
Arribada de l’antigen als fol·licles. L’antigen arriba a fol·licles dels òrgans limfàtics secundaris com els limfonodes. Els antígens
petits hi arriben mitjançant els vasos limfàtics aferents, que els recullen als teixits perifèrics.
Els antígens d’una mida mitjana són captats pels macròfags que es troben a la paret basal
del sinus subcapsular i des dels quals els poden transferir a l’interior dels fol·licles. Altres
antígens també poden ser transportats per les cèl·lules dendrítiques.
Respostes humorals T-dependents. Els limfòcits B que es troben al còrtex dels fol·licles limfàtics al expressar el receptor de
CXCL13, CXCR5. Poden interaccionar amb l’antigen. El resultat d’aquesta interacció és un
acostament a la frontera entre el paracòrtex i el còrtex. Això és perquè l’activació rebaixa
la presentació de CXCR5 i augmenta la de CCR7, decantant els pols d’atracció cap al
paracòrtex. El paracòrtex és la zona dels limfòcits T.

Immunologia 2020
CEBIO 173
Per altra banda, als limfonodes hi arriben limfòcits TCD4, que reconeixen antígens a la zona
del paracòrtex gràcies a les cèl·lules dendrítiques. Això els activa i, en conseqüència, són
retinguts al limfonode. L’activació pot donar lloc a la expressió de CXCR5 al limfòcit T. Si
es dona aquest fet, el limfòcit T s’acostarà al còrtex. A la zona propera entre el còrtex i el
paracòrtex es pot donar la interacció entre els limfòcits T i B. Aquesta interacció és la
primera part de la col·laboració B-T.
La interacció B-T comença amb el limfòcit B, que ha reconegut l’antigen a l’interior del
fol·licle i que l’ha processat, carregat i presentat al limfòcit T. Aquest limfòcit T és específic
per aquest mateix antigen que li presenta el limfòcit B. Per tant, tenim dos limfòcits específics
per diferents epítops del mateix antigen. Quan es dona aquesta interacció, el limfòcit B rep
el segon senyal d’activació arran de la interacció entre CD40 i CD40L. Per tant, el limfòcit
B pot convertir-se en una cèl·lula plasmàtica de vida curta, que produeixen
immunoglobulines als cordons medul·lars del limfonode i expulsen els anticossos per la
circulació, o poden migrar a l’interior dels fol·licles i començar la formació d’un centre
germinal.
Quan un limfòcit B naive reconeix antigen, li passen una sèrie d’alteracions i modificacions.
Entre aquestes trobem que comença a proliferar, augmenta l’expressió de les molècules
coestimuladores B7-1 i B7-2 (lligands de CD28 que li permeten proporcionar el senyal
coestimulador a les cèl·lules T), augmenta l’expressió de receptors de citocines i es produeix
un augment en l’expressió de CCR7 en detriment de la de CXCR5.
En ratolins, la interacció B-T als limfonodes dura uns 4 dies des de la entrada de l’antigen al
cos. Durant els dos primers dies, les cèl·lules T penetren cap a la frontera del còrtex amb el
paracòrtex mentre que les cèl·lules B fan el camí contrari. Al tercer dia ja ha començat la
proliferació de les cèl·lules B i aquestes es poden veure a la part més externa del limfonode.
El quart dia les cèl·lules B ja es poden veure formant centres germinals.
Cèl·lules T helper fol·liculars. La interacció entre els limfòcits T i els limfòcits B requereix de la molècula coestimuladora
ICOS. Aquesta molècula expressada al limfòcit T helper s’uneix a ICOSL, que es troba al
limfòcit B. Aquesta interacció indueix un augment de CD40 al limfòcit B i de CD40L al
limfòcit T. A més, la interacció ICOS-ICOSL regula la transcripció de la citocina
interleuquina 21. En conseqüència, la interacció fa que el limfòcit T helper es diferenciï al
limfòcit T efector T helper fol·licular.
La interacció ICOS-ICOSL augmenta la durada de la interacció entre els dos limfòcits, tenint
efectes en la maduració de l’afinitat, supervivència i diferenciació de les cèl·lules B. Dit d’altra
manera, té efecte sobre totes les reaccions del centre germinal.
Això és així perquè quan un limfòcit T reconeix antigen al paracòrtex, aquest s’activa i
comença a poder produir diferents citocines segons l’antigen, la cèl·lula dendrítica que li
hagi presentat, la quantitat d’antigen... Aquest fenotip, però, no és l’efector. El fenotip efector
queda fixat quan es produeix la interacció B-T i és el fenotip de limfòcit T helper fol·licular. Els
limfòcits T helper fol·liculars poden ser productors d’interferó γ, d’interleuquina 17,
interleuquina 4... I, malgrat de quin tipus siguin, tots produiran IL21. Aquesta citocina és
essencial en molt processos.
ICOS controla l’expressió de la IL21, que controla l’expressió de Bcl6. Bcl6, un factor de
transcripció característic dels limfòcits T helper fol·liculars, controla l’expressió del receptor
de quimiocines CXCR5. Dit d’altre manera, Bcl6 fa que els limfòcits T helper es mantinguin

Immunologia 2020
CEBIO 174
als fol·licles gràcies a l’expressió de CXCR5. En el fons, doncs, ICOS fixa el fenotip de
limfòcit T helper fol·licular i, per tant, aquests migren cap al fol·licle.
A la zona de contacte entre el paracòrtex (zona T) i el còrtex (zona B) el limfòcit B presenta
antigen al limfòcit T helper, del qual no s’ha fixat del tot el seu fenotip. Si aquest limfòcit T
pot reconèixer el mateix antigen que li presenta el limfòcit B, la parella de cèl·lules es
desplacen cap al centre dels fol·licles per desenvolupar la reacció dels centres germinals.
En aquest centre es pot donar lloc a cèl·lules B de memòria o cèl·lules plasmàtiques de
vida llarga.
Les cèl·lules productores d’immunoglobulines de vida llarga són diferents de les de vida
curta, produïdes per interaccions fora de la zona del còrtex. La diferència més important és
que les cèl·lules plasmàtiques de vida curta seran productores d’immunoglobulines del
mateix tipus que la cèl·lula B tenia com a BCR, mentre que les cèl·lules plasmàtiques de
vida llarga hauran passat pels centres germinals, on es pot donar un canvi d’isotip o una
hipermutació somàtica. En conseqüència, les immunoglobulines produïdes d’aquesta
manera seran més eficients contra l’antigen.
El que determina que un limfòcit B activat es diferenciï a cèl·lula plasmàtica o segueixi com a
limfòcit b fol·licular és el factor Bcl6. El limfòcit B activat expressa el receptor de la IL21.
Quan aquest receptor interacciona amb la citocina, el limfòcit B expressa Bcl6, que li manté
l’expressió de CXCR5. A més, es bloqueja l’expressió del factor de transcripció BLIMP-1, el
qual provocaria una transformació en una cèl·lula plasmàtica de vida curta. En conseqüència,
sortiria cap a la medul·la del limfonode per tal que els anticossos anessin cap a la circulació
sanguínia.
Centres germinatius. Generar un centre germinatiu o germinal és un procés complex. Per un costat, un cop es dona
l’activació del limfòcit T al paracòrtex arran de la presentació d’antigen per part de la
cèl·lula dendrítica, comença a migrar cap a la zona de contacte entre el còrtex i el
paracòrtex.
Per altra banda, el limfòcit B que ha trobat antigen també migra cap a la frontera entre la
zona B i la zona T. Allà es produeix la interacció i les parelles migren cap a l’interior del
fol·licle, on comencen a proliferar. Fins al dia 7 (depenent de la quantitat d’antigen i del lloc
d’on s’estigui parlant) no es veu clara l’estructura del centre germinatiu. Aquesta estructura
consisteix en una zona fosca composta principalment per cèl·lules B i una zona clara
formada per limfòcits B, limfòcits T helper fol·liculars i cèl·lules fol·liculars dendrítiques.
La zona fosca dels centres germinatius rep aquest nom perquè hi molta densitat de
limfòcits B que es troben proliferant, moment que s’anomenen centroblasts. Aquest gran
ritme de proliferació resultant d’haver rebut el coestímul dels limfòcits T permet que es doni
una hipermutació somàtica, petits canvis en les zones hipervariables del BCR que li poden
canviar l’afinitat per l’antigen.
A la zona clara s’hi troben els limfòcits B que han deixat de proliferar, anomenats centròcits,
limfòcits T helper fol·liculars i cèl·lules fol·liculars dendrítiques. A la zona clara es dona una
maduració de l’afinitat, procés on intervenen les cèl·lules fol·liculars dendrítiques, i el canvi
d’isotip. El canvi d’isotip depèn de les citocines que es produeixen al centre germinatius, les
quals alliberen els limfòcits T helper fol·liculars.
El moviment entre la zona fosca i la zona clara depèn del gradient de quimiocines. A la
zona fosca hi ha una concentració més alta de CXCL12, detectada pel receptor CXCR4, que

Immunologia 2020
CEBIO 175
no pas a la zona clara. Passa el contrari amb CXCL13. Això és perquè les cèl·lules fol·liculars
dendrítiques són les productores de CXCL13. Per tant, en funció del nivell d’expressió dels
receptors d’aquestes quimiocines, els limfòcits B es mourien de la zona fosca a la clara o a la
inversa.
Cèl·lules Fol·liculars Dendrítiques. Les cèl·lules fol·liculars dendrítiques reben aquest nom perquè tenen unes prolongacions
cel·lulars. No són d’origen hematopoètic. Sembla ser que deriven de cèl·lules l’estroma.
No són presentadores d’antigen ja que no tenen una gran capacitat d’internalització i
degradació del que capten a través dels receptors de la membrana. Tenen alts nivells de
receptors de la fracció constant de les immunoglobulines, com CD32 (receptor de la fracció
constant de les IgG i IgE), i alts nivells de receptors de proteïnes del sistema del
complement com CR1, CR2 i CR3.
Els receptors que aquestes cèl·lules presenten els permeten unir antígens opsonitzats i que
es troben formant immunocomplexes. Això inclou antígens opsonitzats per anticossos o per
proteïnes del sistema del complement. La interacció entre els immunocomplexes i els
receptors a la membrana són molt estables i es mantenen durant temps llargs perquè
aquestes cèl·lules fol·liculars dendrítiques no tenen un bon sistema d’internalització i
degradació. D’aquesta manera, els complexes ancorats a la membrana s’hi mantenen molt
més temps del que seria normal en una puta cèl·lula dendrítica normal i corrent. Aquestes
de seguida s’ho empassa per endocitosi o internalització.
Així i tot, aquests immunocomplexes queden retinguts a la membrana i formen acumulacions
que s’anomenen icosomes. Per tant, aquestes cèl·lules fol·liculars dendrítiques proporcionen
antigen al limfòcit B en un entorn on aquest és molt poc abundant ja que ens troben dins
dels centres germinals. Les cèl·lules fol·liculars dendrítiques mantenen l’antigen prou
temps per permetre l’activació dels collons. Puta Carme.
Les cèl·lules fol·liculars dendrítiques tenen la capacitat de produir CXCL13, el que atrau
els limfòcits B que expressen CXCR5 i també a les cèl·lules T helper fol·liculars que
expressen CXCR5. Les cèl·lules dendrítiques fol·liculars expressen molècules d’adhesió que
estabilitzen la interacció amb els limfòcits B que reconeixen els antígens que tenen
ancorats a la membrana a través dels immunocomplexes. També poden produir factors de
creixement i supervivència pels limfòcits B, com la interleuquina 6 i la citocina BAFF. BAFF
permet mantenir la supervivència dels limfòcits B en aquest entorn concret.
Limfòcits B al centre germinal. Les cèl·lules fol·liculars dendrítiques de la zona clara tenen els immunocomplexes ancorats
a la membrana. Els limfòcits B que han estat a la zona fosca han passat pel procés
d’hipermutació somàtica i poden haver canviat l’afinitat. Tan sols els que millor interaccionin
amb l’antigen seran els que podran reaccionar-hi i rebre tant el senyal de supervivència com
el segon senyal. Aquest segon ve per la interacció amb els limfòcits T helper fol·liculars, la
qual es dona amb CD40 i ICOS. Els limfòcits B de menys o gens afinitat per l’antigen per
culpa de la hipermutació somàtica, no tenen possibilitats d’interaccionar amb l’antigen i moren
per apoptosi al no rebre el senyal de supervivència.
Això vol dir que cada cop que es produeix una resposta per limfòcits B, els que queden al final
són els que tenen major afinitat per l’antigen. Per tant, els que responen inicialment a un
antigen són molt diferents dels que hi interaccionen quan aquest torna a entrar. A mesura que
es va donant la resposta a un determinat antigen, va augmentant la afinitat del BCR per
aquest. Això fa que la resposta sigui més eficient, més ràpida i millor.

Immunologia 2020
CEBIO 176
D’altra banda, a la zona clara del centre germinal hi ha la interacció entre el limfòcit B i el
limfòcit T helper fol·licular, que determina el canvi d’isotip. Amb la interacció amb el
limfòcit B, el T helper fol·licular s’activa i secreta citocines en funció de l’estímul inicial.
Aquest estímul determina si serà T helper fol·licular productora d’interferó, IL4, alguna altre
citocina... En funció de la citocina que es produeixi, el limfòcit B es diferenciarà a productor
d’un determinat isotip d’immunoglobulina. Si el T helper fol·licular produeix IL4 o IL13, el
limfòcit B es diferenciarà a un que pugui produir BCR de IgG1, IgG4 o IgE. Si, en canvi, és
IL12 o IL6, es diferenciarà a un limfòcit B de IgM. Si té IL10 i TGFβ, IgA. Si només té IL10,
IgG1 i IgG3. Si és IL21, serà IgG1, IgG3 i IgA1. Si té IL15, farà IgM, IgG1 i IgA. Gairebé cap
citocina fixa un isotip concret. El tipus de T helper fol·licular determinarà el canvi d’isotip que
es farà. El canvi d’isotip ve en funció de la inducció de la transcripció de les regions switch
que precedien els gens que codificaven per cadascuna de les regions constants.
Limfòcits B de memòria. Resultat de la interacció entre els limfòcits B i T a la zona clara, el limfòcit B es transformarà
en una cèl·lula plasmàtica o en una de memòria. Les plasmàtiques aniran als teixits
limfoides secundaris o aniran a emmagatzemar-se a la medul·la òssia per mantenir-s’hi durant
anys. IgA va a la mucosa intestinal. IgG van a la medul·la.
Les cèl·lules plasmàtiques de vida llarga es mantenen a la medul·la perquè hi troben el nínxol
que els permet sobreviure i mantenir la funció. Entre els factors que determinen que
s’emmagatzemin a la medul·la es troba la interacció entre CXCL12, que es produeix a la
medul·la, i CXCR4. La retenció també requereix de les cèl·lules de l’estroma de la medul·la,
que expressen molècules d’adhesió que ajuden a mantenir les cèl·lules allà. A més, també
expressen diferents factors de transcripció que mantenen aquest fenotip de cèl·lula
plasmàtica. Tant les cèl·lules de l’estroma com cèl·lules hematopoètiques produeixen
citocines que proporcionen els senyals de supervivència. Per tant, hi ha tota una sèrie de
factors que fan que les cèl·lules plasmàtiques de vida llarga es mantinguin vives durant anys
a la medul·la i que, quan és necessari, produeixin anticossos que es troben a la circulació
sanguínia i als diferents teixits.
Els limfòcits B seleccionats positivament es diferencien a cèl·lula de memòria o cèl·lula
plasmàtica. Ambdós tipus surten del centre germinatiu. Poden tornar a entrar-hi dins del
procés de proliferació i canvi d’isotip.
El resultat del procés d’interacció, diferenciació, activació, proliferació i generació de cèl·lules
plasmàtiques de vida llarga és la generació de cèl·lules B de memòria que van als teixits. El
concepte de memòria és el mateix que pels limfòcits T. Són limfòcits B amb un menor llindar
d’activació perquè presenten més molècules coestimuladores i necessiten menys senyal per
activar-se. A més, el menor llindar d’activació implica que es poden activar i desenvolupar els
seus mecanismes efectors de manera més ràpida. Les cèl·lules plasmàtiques productores
d’immunoglobulines de memòria ràpidament poden generar-ne en tornar a entrar en
contacte amb el mateix antigen que n’ha determinat l’activació i diferenciació. Les cèl·lules B
de memòria són més eficients. Ho són gràcies a tenir millor localització perquè es troben als
teixits i perquè tenen més precursors. Els precursors són limfòcits B amb afinitat per aquell
antigen arran de l’expressió clonal. En conseqüència, hi ha més limfòcits B amb la mateixa
especificitat antigènica. Aquests limfòcits B tenen major afinitat per l’antigen. Això es tradueix
en una major qualitat i una major quantitat. Aquest conjunt de característiques permet que,
quan aquests limfòcits B tornin a entrar en contacte amb l’antigen, la resposta sigui més ràpida
i molt més eficient.

Immunologia 2020
CEBIO 177
A la resposta primària s’aconsegueix uns nivells màxims de producció d’anticossos a partir
de 7 dies d’haver entrat en contacte amb l’antigen. Quan la resposta és secundària, és molt
més ràpida. La resposta secundària triga 3 dies en tenir inicia de resposta al tenir els limfòcits
de memòria. Entre 3 i 5 dies és el que es triga a obtenir el pic d’anticossos, la qual és entre
100 i 1000 vegades més gran que la primària. A la primera resposta predomina IgM mentre
que a la segona predominen altres isotips. Sobretot IgG si és resposta al plasma. A més, IgG
té més afinitat per l’antigen gràcies a la hipermutació somàtica.
Per tant, els limfòcits B que es converteixen en efectors poden produir immunoglobulines.
Aquests anticossos tenen funcions molt importants com la neutralització de microbis i toxines,
l’opsonització i fagocitosi dels microbis, ser un element clau en la citotoxicitat cel·lular
depenent d’anticòs i activar el sistema del complement. L’activació del sistema del
complement es tradueix en la lisi dels microorganismes, la inflamació i la fagocitosi dels
microorganismes opsonitzats amb fragments del sistema del complement. En si, el
mecanisme efector de les cèl·lules B és la producció d’anticossos.
Els anticossos són molt bons impedint la interacció de toxines amb els seus receptors a les
membranes de les cèl·lules. Aquest procés el realitzen interaccionant amb elles i impedint
l’adherència al receptor. Això és molt important en la resposta a virus, per exemple. Els
anticossos són importants en l’opsonització de bacteris, faciliten la seva internalització i
fagocitosi. Els anticossos també activen el complement i faciliten la destrucció d’alguns
bacteris.
Quan es produeix una resposta a un bacteri i aquest no està opsonitzat, té una supervivència
del 80%. Si ha estat opsonitzat, en canvi, la supervivència és del 1%. L’addició dels efectes
del sistema del complement redueix la població bacteriana al mínim en un temps de dues
hores. Tot això és perquè els anticossos faciliten la detecció per part de les cèl·lules
fagocítiques.
IgA i IgE tenen una funció molt important a les barreres de la pell i les mucoses associades
al tracte intestinal i respiratori. La IgA és la immunoglobulina secretada a les mucoses
associades al tracte intestinal i respiratori. S’hi troba en forma de dímer arran de la cadena J.
Per tal que la immunoglobulina passi al lumen de l’intestí, fa servir el receptor poliIg que
s’uneix al dímer i el transporta cap a la zona apical de les cèl·lules per transitosi. Abans de
sortir, es proteolitza el receptor poliIg i aquesta estructura de IgA dimèrica amb poliIg és el
que la fa tant estable al llum intestinal en condicions adverses. PoliIg és el que li dona
estabilitat i li permet fer la funció de neutralització.
Així dons, a l’intestí s’hi troba la IgA dimèrica, que opsonitza i facilita la detecció dels antígens
que allà es trobin. A l’interior del teixit tenim gran producció IgE, que pot sensibilitzar i activar
desgranulació mastòcits per atraure’ls i atraure altres cèl·lules i anticossos.

Immunologia 2020
CEBIO 178
Tema 22 – Regulació de la resposta immunològica. Les respostes immunitàries són resultat d’interaccions complexes entre cèl·lules i
mediadors diferents. Això fa necessitin estar molt regulades per tal de prevenir la resposta
a estructures pròpies o autoreactivitat. Si la regulació falla, es pot donar una resposta
excessiva que generi autoimmunitat, una resposta deficient que impedeixi la resposta a
tumors o una desviació de la resposta que doni lloc a reaccions d’al·lèrgia. Per altra banda,
la resposta immunitària s’ha de poder aturar quan ja s’ha eliminat l’antigen i el sistema
immunitari s’ha de mantenir en estat de no resposta enfront dels antígens propis. Així
i tot, el sistema immunitari ha d’aconseguir que hi hagi una gran estimulació quan interaccioni
amb un antigen forà i ha de retornar a l’estat de no resposta quan aquest desaparegui.
Tant important és aconseguir que no hi
hagi resposta a antígens propis com
eliminar la resposta quan ja no hi ha
antígens. Si es manté la resposta
immunitària quan ja no hi ha antígens
forans es podria donar una inflamació
crònica, la qual comporta una destrucció
del teixit, un mal funcionament del mateix
i una alta possibilitat de desenvolupar
processos tumorals.
Tolerància Immunològica. La tolerància immunològica és la capacitat del sistema immunitari per decidir la no
resposta a un antigen determinat, sigui propi o no perillós. En principi, els al·lèrgens són
antígens no perillosos pels quals no hi ha una tolerància immunològica. Per tant, el sistema
immunitari ha d’aprendre a no respondre a allò que no sigui estrictament necessari. El
procés d’aprenentatge es fa en dues fases. La fase d’aprenentatge de la Tolerància Central
es genera als Òrgans Limfoides Primaris i serveix per generar un repertori de limfòcits T i B
poc o gens autoreactiu. Així es determina que hi hagi una poca capacitat de resposta al que
sigui propi. Així i tot, com que són poc reactius, hi ha una Tolerància Perifèrica que es genera
als Òrgans Limfoides Secundaris i que permet mantenir un estat de no resposta a estructures
pròpies.
Els mecanismes de manteniment de la tolerància central consisteixen en els processos
de selecció negativa dels limfòcits B autoreactius a la medul·la òssia i dels limfòcits T
autoreactius al timus. En canvi, la tolerància perifèrica es manté per altres vies i la seva
pèrdua es deu a gens inapropiats, a l’aparició de mutacions o a trobar-se en un entorn
inapropiat. La pèrdua de la Tolerància Perifèrica pot donar lloc a malalties
autoimmunitàries.
Resposta
im
munitària
Antígens propis - Antígens Foranis - Retorn a la normalitat
Aprenentatge de no-resposta a antígens propis
Tolerància Central
Òrgans Limfoides Primaris
Selecció negativa
Tolerància perifèrica
Òrgans Limfoides Secundaris
Manteniment tolerància central

Immunologia 2020
CEBIO 179
Mecanismes de la Tolerància Immunològica Perifèrica. La Tolerància Immunològica Perifèrica manté un
repertori de cèl·lules que no genera resposta a
estructures pròpies quan es troben en condicions
homeostàtiques. Per aconseguir-ho, existeixen diferents
mecanismes. Un mecanisme de la Tolerància Perifèrica és
mantenir un estat de no activació. En aquest és molt
important la segregació de l’antigen, segons el qual hi ha
òrgans immunològicament privilegiats el parènquima
d’aquests no entra en contacte amb el sistema
immunitari. En conseqüència, no s’hi pot donar una
resposta a antígens propis. L’estat de no activació també
s’aconsegueix mantenint un estat d’anèrgia, eliminant les
cèl·lules autoreactives o fent un procés de supressió o
regulació mitjançada per limfòcits T reguladors i cèl·lules
reguladores.
Regulació per inducció d’anèrgia.
L’anèrgia és la manca de resposta a estímuls antigènics.
Quan hi ha anèrgia, el resultat de la interacció entre el limfòcit
T i l’antigen presentat per una cèl·lula presentadora d’antigen
no es tradueix en una activació, proliferació i diferenciació
efectora dels limfòcits. Enlloc d’això, amb l’anèrgia s’indueix
l’expressió d’una sèrie de gens que inhibeixen la
senyalització del TCR, inhibeixen l’expressió de
citocines, degraden intermediaris de la senyalització i
bloquegen el cicle cel·lular. Els limfòcits T anèrgics, fruit
dels gens inductors de l’anèrgia, no secreten Interleuquina 2,
la qual no pot actuar de manera autocrina per permetre la
seva proliferació. Els limfòcits B també poden entrar en un
estat d’anèrgia. En el seu cas, és el reconeixement de
l’antigen per part del BCR el que no tindria efectes de
proliferació.
L’expressió dels gens inductors d’anèrgia
es veu provocada per la presentació
d’antigen en absència de senyals
coestimuladors activadors o d’elements
de la resposta innata. Quan això passa,
s’indueix l’anèrgia clonal del limfòcit T
autoreactiu. En aquesta, el limfòcit té una
baixa expressió dels complexes pèptid-MHC.
Això provoca que, quan una cèl·lula
presentadora d’antigen interaccioni amb
un limfòcit T anèrgic, tan sols es donarà el
primer senyal d’activació. Com que manca
el segon senyal d’activació, el primer senyal
resultant de la interacció no serà prou intens
com perquè es doni una activació.
Manteniment estat de no-activació
Segregació d’antigen
Estat d’anèrgia
Eliminació cèl·lules autoreactives
Regulació per Treg
Limfòcits T anèrgics
Inhibició senyalització TCR
Inhibició Expressió citocines
Degradació intermediaris senyalització
Bloqueig cicle cel·lular
No hi ha activació
Només 1r senyal activació
Interacció APC – Limfòcit T anèrgic
Baixa expressió dels complexes pèptid-MHC
Anèrgia clonal del limfòcit T autoreactiu
Expressió dels gens inductors d’anèrgia
Presentació d’antigen en absència de senyals coestimuladors activadors

Immunologia 2020
CEBIO 180
Si hi ha un reconeixement d’antigen per part dels limfòcits T autoreactius però no hi ha
una senyalització mitjançant les molècules coestimuladores, el limfòcit es converteix
en un limfòcit T anèrgic. Això és perquè les molècules coestimuladores són les que
proporcionen el segon senyal d’activació. Un cop s’ha passat a un estat anèrgic, costa molt
que un limfòcit T respongui a l’antigen. Aquesta situació d’inducció d’anèrgia també seria
vàlida per l’activació del limfòcit B autoreactiu. Això és perquè el limfòcit B autoreactiu no
trobarà la col·laboració necessària en el limfòcit T activat. Per tant, l’anèrgia és un sistema
que funciona per evitar la resposta adaptativa.
Una altra manera d’induir anèrgia és la
senyalització a través de molècules
coestimuladores inhibidores, com CTLA-4.
Aquest inhibidor té els mateixos lligands que
la molècula coestimuladora activadora CD28.
El lligand tant de CTLA-4 com de CD28 és
B7, altrament dit CD80/CD86. Els limfòcits T
naive expressen CD28 de manera
constitutiva i així pot interaccionar amb B7 per
transmetre un senyal de coestimulació
activadora que contribueix a l’activació del
limfòcit. Quan s’expressa CTLA-4 es
transmeten senyals d’inhibició. Això és
perquè CTLA-4 recluta fosfatases que
defosforilen les cues citoplasmàtiques dels
receptors al limfòcit T. Així doncs, es pot dir
que CTLA-4 indueix anèrgia clonal.
PD-1 és una altra molècula coestimuladora inhibidora, però no competeix amb CD28 per
la unió amb CD80/CD86 al tenir el lligand propi PD-L1/PD-L2. El resultat de la interacció de
PD-1 i de CTLA-4 amb els seus lligands és la desfosforilació de les molècules
senyalitzadores. Això es tradueix en una aturada de la senyalització amb la conseqüent
disminució de la producció de citocines, de la proliferació i diferenciació de les cèl·lules
efectores i de la supervivència d’aquestes cèl·lules.
També es pot induir l’estat d’anèrgia als limfòcits B gràcies als receptors FcγRIIB per
tenir motius ITIM a la seva cua citoplasmàtica. Específicament, el receptor és CD32. Si es
produeix un entrecreuament entre el BCR i la immunoglobulina que reconeix el mateix
antigen, la interacció de la immunoglobulina amb el receptor Fc activa les fosfatases
unides a la cua citoplasmàtica. Aquestes fosfatases defosforilen les cadenes
citoplasmàtiques de Igα i Igβ. En conseqüència, bloqueja la senyalització a través del BCR.
Regulació per inducció de mort per apoptosi.
Una altra manera de regular la resposta del sistema
immunitari és la inducció de la mort per apoptosi.
L’apoptosi és un procés fisiològic d’inducció de la mort
cel·lular on intervenen les caspases. L’apoptosi es
caracteritza per canvis en la distribució dels lípids de la
membrana, una condensació i fragmentació nuclear, una
fragmentació de l’ADN, una vesiculació de la membrana i la
formació de cossos apoptòtics. Les cèl·lules que entren en
apoptosi es fragmenten, però aquests fragments es
mantenen envoltats de membrana.
CTLA-4 s’uneix a B7
CTLA-4 recluta fosfatases
Desfosforilació cues citoplasmàtiques de receptors al limfòcit T
Inhibició CD28
Inducció d’anèrgia clonal
ApoptosiCanvis en distribució lípids
membrana
Condensació i fragmentació nuclear
Fragmentació DNA
Vesiculació membrana
Formació cossos apoptòtics

Immunologia 2020
CEBIO 181
És un procés molt diferent de quan una cèl·lula mor
per necrosi. La necrosi és el procés pel qual es
perd la integritat de la membrana plasmàtica, el
contingut cel·lular és degradat per acció enzimàtica
i el contingut citosòlic s’allibera a l’espai
extracel·lular. En conseqüència, la necrosi genera
gran quantitat de DAMPs, Patrons Moleculars
Associats a Perill. Per tant, la necrosi genera una
inflamació.
Des de fa poc s‘ha vist que quan els cossos
apoptòtics són fagocitats, les cèl·lules que
ho han fet secreten citocines
antiinflamatòries com IL-10 i TGF-β. A més,
les cèl·lules que les han fagocitat disminueixen
la secreció de citocines inflamatòries com TNF-
α, IL-1β i IL-12. És per això que es pot dir que
el resultat de la mort per apoptosi és una
mort silent, que no genera activació i
inflamació del sistema immunitari i que
genera senyals que inhibeixen la resposta
del sistema immunitari.
La inducció de l’apoptosi es pot produir de dues maneres. La mort per omissió és la inducció
de l’apoptosi que es dona quan s’ha eliminat completament l’antigen o baixa tant la secreció
d’IL-2 que no n’hi ha a l’entorn que no la pot fer servir el limfòcit T. La mort per activació és
la inducció de l’apoptosi que es dona quan l’antigen és molt persistent i hi ha una estimulació
repetida del limfòcit T.
La mort per omissió es dona en cas que hi hagi una
absència d’estímuls de supervivència, ja siguin
factors de creixement, antigen o interleuquina 2.
En la mort per omissió s’altera la permeabilitat de la
membrana dels mitocondris, el que allibera el citocrom
C al citoplasma. La unió del citocrom C a la proteïna
Apaf-1 activa la caspasa 9. La caspasa 9, quan està
activa, activa la caspasa 3. La caspasa 3 trenca el DNA
de doble cadena i indueix l’apoptosi.
La inducció de l’apoptosi també es pot donar per
l’activació de Fas i del lligand de Fas. Fas s’expressa a
diferents tipus cel·lulars, com les cèl·lules epitelials o els
limfòcits. El lligand de Fas només s’expressa als limfòcits T
fruit d’una activació repetida i l’acció de la IL-2. Per tant, quan
hi ha una activació continuada del limfòcit T, es comença a
expressar el lligand de Fas a la seva membrana. Es pot
produir la interacció entre Fas i el seu lligand estant els dos
a la membrana del limfòcit T. La interacció entre Fas i FasL
provoca que la proteïna FADD s’uneixi al domini de mort
cel·lular de la cua citoplasmàtica de les proteïnes Fas. La
interacció activa la caspasa 8. La caspasa 8 activa la
caspasa 3.
Alteració Membrana Mitocondris
Alliberació Citocrom C al citoplasma
Unió Citocrom C al Apaf-1
Activació Caspasa 9
Activació Caspasa 3
Expressió de FasL
Interacció Fas-FasL
Unió FADD a Fas
Activació Caspasa 8
Activació Caspasa 3
Necrosi
Pèrdua integritat membrana
Contingut cel·lular degradat per acció enzimàtica
Contingut citosòlic alliberat a espai extracel·lular
Generació DAMPs
Generació Inflamació
Fagocitosi cossos apoptòtics
Secreció citocines antiinflamatòries
IL-10 TGF-β
Disminució secreció citocines inflamatòries
TNF-α IL-1β IL-12

Immunologia 2020
CEBIO 182
Regulació per cèl·lules T reguladores.
Els timòcits que tenen una afinitat intermèdia pels antígens propis es diferencien en
cèl·lules T reguladores. Aquestes cèl·lules T reguladores es generen des del primer moment
del individu i contribueixen a mantenir l’estat de no resposta a la perifèria. Així i tot, es
generen cèl·lules T reguladores durant la resposta a la perifèria com a resultat de
l’activació. Quan hi ha una activació i una producció de TGF-β o àcid retinoic, els limfòcits T
que reconeixen antigen es poden diferenciar a cèl·lules T reguladores perifèriques o cèl·lules
T reguladores induïdes. Independentment de quin tipus siguin, la funció de les cèl·lules T
reguladores és aturar la resposta del sistema immunitari i impedir que les cèl·lules T
efectores continuïn activant-se.
S’han descrit vàries poblacions de cèl·lules T
reguladores. Així i tot, la majoria de cèl·lules T
reguladores són limfòcits TCD4 que expressen
CD25. Aquesta molècula és la cadena α del
receptor de la IL-2, aquella que permet generar
IL2R d’alta afinitat. Les cèl·lules T reguladores
també expressen CTLA-4 de forma constitutiva,
la molècula coestimuladora inhibitòria GITR i el
factor de transcripció Foxp3.
L’elevada expressió de CD25 significa que tenen un receptor d’alta afinitat per la IL-2.
En cas que aquesta citocina es trobi al medi, aquestes cèl·lules la podran captar a través de
la interacció amb aquest receptor d’alta afinitat.
Per altra banda, l’expressió del factor de
transcripció Foxp3 té una funció molt important.
Per una banda, Foxp3 s’uneix al promotor
dels gens de CD25, CTLA-4 i GITR alhora que
indueix l’acetilació de les histones, facilitant
la transcripció dels gens que codifiquen per
aquestes proteïnes. Per altra banda, la unió de
Foxp3 als promotors de la IL-2, IL-22 o de
l’interferó γ recluta enzims desacetilases que
en reprimeixen la transcripció al desacetilar
les histones.
L’activitat de les cèl·lules T reguladores és
dependent d’antigen. Dit d’una altra manera,
les cèl·lules T reguladores han de reconèixer
antigen a través del seu TCR per manifestar la
seva funció reguladora. El fet que tinguin un
TCR d’afinitat intermèdia implica que sol tenir
més afinitat que els TCR de les cèl·lules
autoreactives. Per tant, les cèl·lules T
reguladores es poden activar a
concentracions d’antigen molt baixes. Un
cop s’han activat, les cèl·lules T reguladores
inhibeixen les cèl·lules T efectores, aquelles que
estan responent o respondrien.
Cèl·lules T reguladores
Expressió CD25
Expressió CTLA-4
Expressió GITR
Factor transcripció Foxp3
Foxp3
Facilitar transcripció
CD25 CTLA-4 GITR
Repressió transcripció
IL-2 IL-22 INF-γ
Baixa concentració antigen
Reconeixement antigen per cèl·lula Treg
Inhibició cèl·lules T efectores

Immunologia 2020
CEBIO 183
Hi ha diferents maneres d’inhibir les cèl·lules T
efectores. D’una banda, es pot inhibir la
transcripció de la IL-2 a la cèl·lula efectora. A
més, ho faria competint per la IL-2 existent, el que
facilita la inducció de l’anèrgia a les cèl·lules T
efectores. D’altra banda, les cèl·lules T reguladores
expressen molècules coestimuladores inhibidores
com el CTLA-4, PD-1 i LAG-3. Per tant, el competir
per la interacció amb B7 impedeix que el limfòcit
T rebi el segon senyal i que es pugui activar.
Quan CTLA-4 s’uneix a B7.1 o B7.2, augmenta la
producció d’enzims com IDO. IDO és un enzim que
metabolitza el triptòfan, un aminoàcid essencial per
les funcions cel·lulars. Segrestant, eliminant i
metabolitzant el triptòfan, les cèl·lules T
reguladores impedeixen o dificulten la activació
i proliferació dels limfòcits T efectors. A més, les
cèl·lules reguladores poden produir citocines
immunosupressores com IL-10, TGF-β i la IL-35;
que inhibeixen la inflamació i la resposta.
Les cèl·lules T reguladores fan la seva funció un cop han reconegut antigen, pel que la
supressió i regulació de la resposta immunitària que exerceixen les cèl·lules T reguladores és
específica d’antigen. Així i tot, el fet que puguin produir citocines immunosupressores fa
que, un cop activades, les cèl·lules T reguladores puguin suprimir limfòcits amb una
especificitat antigènica diferent només perquè es troben per la zona. Per tant, tot i que
l’activitat supressora és depenent d’antigen i això implica que regularan primer les cèl·lules
efectores amb la mateixa especificitat, la seva funció efectora un cop activades ja no és
específica d’antigen. Poden inhibir la funció d’altres cèl·lules T efectores.
L’activitat reguladora a través de les citocines inhibidores com IL-10, TGF-β i IL-35 es pot
exercir directament sobre els limfòcits T inhibint la proliferació, inhibint la diferenciació de
limfòcits T naive a limfòcits T efectors, suprimint l’activitat de les cèl·lules T efectores quan ja
estan diferenciades, contribuint a la inducció de les cèl·lules T reguladores perifèriques o
mantenint les cèl·lules T reguladores d’origen tímic.
Així i tot, les cèl·lules T reguladores també poden fer el seu efecte sobre cèl·lules
presentadores d’antigen al convertir-les en cèl·lules tolerogèniques. Això es pot fer
inhibint l’expressió de molècules coestimuladores activadores a les cèl·lules presentadores
d’antigen. D’aquesta manera evitarà que proporcionin un segon senyal d’activació i així
anergitzen les potencials cèl·lules inhibidores. Un altre mètode descrit i que es fa servir per
immunoteràpia és que les cèl·lules T reguladores poden inhibir la maduració de les
cèl·lules dendrítiques. Això impedeix que es transformin en cèl·lules presentadores molt
potents, capaces d’activar els limfòcits T naive. A més, les cèl·lules T reguladores poden
incidir en les cèl·lules presentadores d’antigen tot induint l’activació de macròfags M2
per la secreció de les interleuquines 10, 4 i 13. Els macròfags de tipus M2 ajuden a acabar
la resposta i reparar el teixit malmenat.
Una altra manera per la qual les cèl·lules T reguladores poden regular la resposta dels
limfòcits T és intervenint en el seu metabolisme. Se sap que les cèl·lules T reguladores
expressen CD39 i CD73 a la membrana, ectoenzims que passen l’ATP a AMP i que
produeixen adenosina. Aquesta purina té activitat immunoreguladora i està molt ben descrita.
Inhibir cèl·lules T efectores
Inhibir transcripció IL-2
Competir per interacció amb B7
oCTLA-4
oPD-1
oLAG-3
Treure triptòfan
Citocines immunosupressores
oIL-10
oTGF-β
oIL-35

Immunologia 2020
CEBIO 184
Per altra banda, les cèl·lules T reguladores expressen AMP cíclic intracel·lular que
transfereixen a les cèl·lules T efectores. L’acció de l’AMP cíclic és inhibir NFAT, pel que hi
ha una inhibició de la transcripció de la interleuquina 2. En absència de la IL-2, fàcilment
s’indueix l’apoptosi. Interferint en el metabolisme tant de les cèl·lules efectores com de
les cèl·lules presentadores d’antigen, les cèl·lules T reguladores poden evitar que hi
hagi una resposta dels limfòcits T.
Tolerància Perifèrica als Limfòcits B.
Els mecanismes bioquímics que s’apliquen sobre les cèl·lules T efectores també es
donen als limfòcits B per tal de mantenir-hi la tolerància perifèrica. Això es pot fer
mitjançant els receptors inhibidors CD22 i CD32 que impedeixen la senyalització, per la
inducció de l’apoptosi quan es reconeixen antígens propis sense la col·laboració dels limfòcits
T, quan es reconeixen antígens propis amb alta afinitat o quan es reconeixen antígens propis
repetidament, el que indueix l’anèrgia. Els mecanismes cel·lulars són semblants per
limfòcits T efectors i per a limfòcits B.
Regulació de la resposta un cop s’ha iniciat. Hi ha mecanismes de regulació que eviten la resposta del sistema immunitari i hi ha
mecanismes que controlen la resposta un cop iniciada. La regulació de la resposta un
cop ha estat iniciada tendeix a aturar-la o inhibir-la. El control de la resposta immunològica
iniciada pot ser:
Modulació Neuroendocrina de la resposta.
Un exemple rellevant de la modulació neuroendocrina de la resposta és que l’estrès
pot provocar la supressió de la resposta immunològica. En situacions d’estrès, el sistema
immunitari funciona pitjor perquè aquest estrès redueix la capacitat de resposta a diferents
tipus de patògens. Això passa perquè existeix una interconnexió entre els sistemes
immunològic, endocrí i nerviós. La interconnexió no és casualitat, doncs la majoria dels
teixits limfoides estan innervats pel sistema nerviós simpàtic.
Per altra banda, el sistema nerviós controla la producció de diverses hormones da
manera directa o indirecta. Entre aquestes hormones estan la del creixement, la tiroxina,
l’adrenalina, els corticoesteroides... A més, els limfòcits expressen receptors per aquestes
hormones, neurotransmissors i neuropèptids. En conseqüència, la connexió entre els
sistemes immunitari, nerviós i endocrí és una manera de regular el funcionament del primer.
En això és important l’efecte dels glucocorticoides.
S’ha vist que els glucocorticoides modulen la transcripció dels gens de les citocines tot
suprimint els gens de les citocines pro-inflamatòries i regulant la producció de les
antiinflamatòries. Els glucocorticoides també modulen l’expressió de molècules
d’adhesió i la producció de quimiocines, pel que regulen el trànsit de cèl·lules del sistema
immunitari cap a les zones d’inflamació. En afegit, els glucocorticoides suprimeixen la
síntesi de mediadors d’inflamació com la prostaglandina i l’òxid nítric sintasa pel que,
d’alguna manera, els glucocorticoides rebaixen la inflamació. Els glucocorticoides
suprimeixen la maduració, diferenciació i proliferació de les cèl·lules del sistema
immunitari, incloses les de la immunitat innata, els limfòcits T i els limfòcits B.
Regulació per anticossos.
Regulació per antigen.
Regulació per limfòcits.
Modulació idiotípica de la
resposta.
Modulació neuroendocrina de la resposta.

Immunologia 2020
CEBIO 185
L’ús dels glucocorticoides, quan es fan servir com a tractament per modular la resposta del
sistema immunitari, ha de ser limitada i controlada.
Per fer totes aquestes accions, els glucocorticoides fan servir el receptor de
glucocorticoides expressats a les cèl·lules del sistema immunitari. La interacció dels
glucocorticoides amb els seus receptors modula l’expressió gènica perquè intervé en
l’activació de factors com NFκB i en l’acció d’AP-1. Per tant, els glucocorticoides
impedeixen que el factors de transcripció NFκB i AP-1 actuïn al nucli, determinant la
transcripció de gens molt concrets.
Hi ha molts sistemes de regulació del sistema immunitari i tots han de funcionar
correctament. El mal funcionament de qualsevol d’aquests sistemes pot donar lloc a diferents
patologies o a l’absència de la resposta, doncs una mala regulació pot impedir que es
respongui a cèl·lules tumorals. Per tant, tots els mecanismes són molt importants i són
molt diversos.
Glucocorticoides
Modulen transcripció gens citocines
Supressió gens citocines pro-inflamatòries
Regulació producció citocines
antiinflamatòries
Modulen expressió molècules adhesió
Modulen producció citocines
Supressió síntesi
mediadors inflamació
Supressió maduració,
diferenciació i proliferació cèl·lules S.I.

Immunologia 2020
CEBIO 186
Tema 23 – Resposta a bacteris. Els microorganismes i els seus hostes estan en lluita constant per la supervivència.
Aquesta lluita tendeix a un equilibri inestable entre els dos, doncs l’objectiu del patogen és
infectar l’hoste i multiplicar-s’hi, mentre que l’objectiu de l’hoste és eliminar el patogen o reduir-
lo a un paràsit inofensiu.
Per tal d’infectar l’hoste i multiplicar-s’hi dins, els bacteris evadeixen els mecanismes de
defensa de l’individu però sense immunodeprimir-lo prou com perquè faciliti la infecció
per altres patògens. De vegades, els mecanismes de defensa aconsegueixen impedir
l’entrada del patogen i fan predominar l’individu. Altres cops, els patògens aconsegueixen
infectar les cèl·lules i els teixits.
La microbiota sapròfita també intervé en els mecanismes de defensa de l’hoste contra
bacteris. La microbiota sapròfita són organismes comensals que inhibeixen de manera
competitiva la fixació de patògens a la cèl·lula hoste. En conseqüència, té unes funcions molt
importants en el manteniment de l’equilibri.
Microbiota. La microbiota associada a cada organisme és una comunitat ecològica de
microorganismes comensals simbiòtics que es troba a tots els organismes
multicel·lulars, des de plantes fins a animals. La microbiota inclou bacteris, arqueus, protists,
fongs i virus. Dels que més es coneix informació són els bacteris, però és molt important la
participació dels fongs i els virus.
S'ha demostrat que la microbiota és crucial per
a l'homeòstasi metabòlica, hormonal i
immunològica de l’hoste. Proporciona vitamines
i nutrients. La microbiota participa en el
manteniment i millora de la barrera epitelial dels
teixits de les mucoses, el desenvolupament del
sistema immunitari i l’adquisició de factors que
contribueixen a la seva regulació. La microbiota té
un paper molt important en el manteniment de
l’equilibri inestable entre individus i patògens.
Mecanismes d’acció de la microbiota sapròfita sobre el Sistema Immunitari.
La microbiota sapròfita actua de diverses maneres per proporcionar una protecció
directa contra els patògens. Entre aquestes maneres estan:
Competència directa per nutrients
Alteració de les condicions físiques i
químiques del microambient local
Producció de productes
bactericides
Modulació de l’expressió de factors
de virulència
Inducció de l’expressió de
mucines per part de les cèl·lules de Goblet
Inducció de la producció de pèptids antimicrobians per les
cèl·lules de Paneth
Microbiota
Proporcionar vitamines i nutrients
Manteniment i millora barrera epitelial
Desenvolupament sistema immunitari
Manteniment equilibri patògens vs individus

Immunologia 2020
CEBIO 187
L’alteració de les condicions físiques i químiques del microambient s’aconsegueix
consumint oxigen i alterant el pH, el que provoca que els patògens tinguin menys capacitat
de proliferació.
Les cèl·lules de Paneth es veuen
induïdes a produir RegIIIγ i AMP, els quals
tenen un paper molt important en la
neutralització dels bacteris, en evitar que
entrin en contacte amb les cèl·lules que
formen la barrera epitelial i en evitar que
entrin al parènquima del teixit.
La microbiota sapròfita també pot
proporcionar una protecció indirecta al
afavorir la maduració del teixit limfoide
associat a la mucosa, ajudar a estructurar
les plaques de Peyer, ajudar a mantenir la
inflamació fisiològica per mitjà de Treg i la
generació de Th17 i al induir la resposta dels
limfòcits B que donin lloc a la producció
d’immunoglobulina A.
L’estructura del MALT restringeix les interaccions entre cèl·lules dendrítiques i
limfòcits B o entre cèl·lules dendrítiques i cèl·lules T als ganglis mesentèrics, evitant la
resposta immunitària sistèmica. El MALT és el sistema immunitari associat a les mucoses.
La microbiota sapròfita és molt important
per contribuir al desenvolupament del
sistema immunitari associat a les
mucoses. Una de les coses que fa és activar
els macròfags intestinals, el que augmenta la
forma pro-interleuquina-1β. La infecció per
patògens activa el pas de la forma inactiva a
la forma activa de la IL-1β. En conseqüència,
es promou el reclutament dels neutròfils i
l’erradicació dels patògens.
La microbiota és molt important en la promoció de la diferenciació i activació de les
cèl·lules Th17 i de les innate lymphoid cells. El conjunt de Th17 i les ILC controlen tant els
comensals com els patògens per mitjà de la secreció de citocines. Entre les citocines
secretades pels limfòcits Th17 i les ILCs es troben la IL-22. Per tant, en la separació entre el
món extern dels patògens i el mon intern dels hostes es troba la microbiota.
Funcions de la microbiota sapròfita.
L’alteració de la microbiota provoca una alteració de tots els mecanismes de control
que exerceix. Els elements ambientals, la contaminació, els pesticides, els factors genètics,
els factors epigenètics, l’estil de vida, els medicaments i els antibiòtics entre d’altres elements
poden alterar la microbiota. En conseqüència, es veurà afavorida l’entrada dels possibles
elements patogènics que estiguessin sent controlats per tots aquests mecanismes i que
puguin entrar al teixits causant una infecció.
Si una infecció arriba als vasos sanguinis, pot ser sistèmica. En el cas de les infeccions
per bacteris, aquest és un pas clau. Quan es produeix la disrupció de la barrera de la
RegIIIγ + AMP
Neutralitzar bacteris
Evitar contacte amb cèl·lules barrera epitelial
Evitar entrada en parènquima teixit
Activació macròfags intestinals
Augment pro-interleuquina-1β
Activació forma activa IL-1β
Reclutament Neutròfils
Erradicació patògens
Protecció indirecta
Maduració del teixit limfoide associat a la mucosa
Estructuració de plaques de Peyer
Manteniment de la inflamació fisiològica
Inducció de resposta dels limfòcits B

Immunologia 2020
CEBIO 188
microbiota, a l’organisme entren tant patògens com elements de la microbiota que poden tenir
efectes patogènics. És el cas de diferents especies bacterianes.
Staphylococcus aureus colonitza les foses nassals d’entre el 20 i el 50% dels individus.
La presència d’aquest bacteri a les fosses no té conseqüències patològiques, però si que les
té quan és ingerit i passa a les vies respiratòries. Diferents espècies de Clostrídium es
troben normalment formant part de la microbiota intestinal. Si entrés per una via diferent
d’aquesta, pot generar el tètanus. Hi ha molts altres microorganismes que poden causar
meningitis o gonorrea depenent de la via per on entrin.
Una alteració de la microbiota suposa no només l’entrada de patògens sinó també
l’entrada d’elements de la mateixa microbiota, el que pot suposar un problema si arriben
al parènquima dels teixits.
Entrada dels patògens. Tan diversos són els bacteris com les seves vies d’entrada. Les vies d’entrada dels
bacteris poden ser les superfícies de les mucoses de les vies respiratòries, del tracte
gastrointestinal o el tracte reproductiu; les superfícies externes, les ferides o rascades o les
injeccions per la picada d’insectes. El mode de transmissió dels microorganismes serà
diferent per cadascun d’ells. La via d’entrada d’un microorganisme determina la resposta
immunitària que es faci servir.
Un cop han entrat, cal distingir entre els bacteris amb una vida extracel·lular d’aquells
que tenen una vida intracel·lular. Tenen una vida extracel·lular els bacteris que colonitzen
els espais intersticials, la sang, la limfa o les superfícies epitelials.
Els bacteris intracel·lulars entren a l’interior de les cèl·lules dels teixits. Dins les cèl·lules, els
bacteris es poden mantenir al citoplasma o a compartiments vesiculars. Els mecanismes del
sistema immunitari per eliminar un bacteri extracel·lular són diferents dels que farà
servir per eliminar bacteris intracel·lulars.
Paral·lelament als bacteris extracel·lulars i intracel·lulars hi ha un grup de bacteris que
entren a l’interior de les cèl·lules per començar la seva replicació però que es troben a
l’espai extracel·lular. Dependents del moment de la vida es trobin aquests bacteris, seran
més eficients els mecanismes de resposta davant de bacteris extracel·lulars o els de resposta
davant de bacteris intracel·lulars. Les infeccions per qualsevol tipus de bacteri consisteixen
en 4 etapes:
Adhesió dels bacteris a la cèl·lula de l’hoste.
Proliferació del Bacteri.
Invasió del teixit de l’hoste.
Lesió de la cèl·lula hoste.
Vies d’entrada de bacteris
Superfícies de mucoses
Vies respiratòries
Tracte gastrointestinal
Tracte reproductiu
Superfícies externes
Ferides Rascades
Injeccions
Picades d’insecte

Immunologia 2020
CEBIO 189
Les cèl·lules a les quals s’adhereixen els bacteris poden ser cèl·lules que fan de barrera
o poden ser les cèl·lules diana a les que entren. Són un exemple de cèl·lules que fan de
barrera les cèl·lules epitelials de la paret intestinal. La lesió de la cèl·lula es pot fer per diversos
mètodes, un dels quals són les toxines.
Bacteris Extracel·lulars. Els bacteris extracel·lulars proliferen fora de les cèl·lules de
l’hoste i causen malalties, problemes o simptomatologia per
dos mecanismes principals. Per un costat, els bacteris
extracel·lulars causen una gran inflamació. Aquesta deriva
en una important destrossa del teixit i en la formació de pus.
Aquest mecanisme de gran inflamació és el que fan servir els
bacteris Staphylococcus i Streptococcus entre d’altres. Són
els bacteris que trobem quan tenim mal de coll i causen una
inflamació.
L’altre mecanisme dels bacteris extracel·lulars consisteix
en la producció de toxines. Aquestes poden ser exotoxines
o endotoxines. Les exotoxines són toxines amb un efecte
directe sobre aquelles cèl·lules que tinguin un receptor per
elles. És el cas de la Diftèria toxinodiftèrica o el tètanus. Les
endotoxines són toxines amb un efecte immunopàtic.
L’endotoxina LPS activa els macròfags, els quals indueixen la
secreció de gran quantitat de citocines proinflamatòries com
TNF o IL-1, el que pot provocar una febre alta i un xoc sèptic.
Les conseqüències de la infecció per bacteris extracel·lulars són les causants de gran part de
la patologia. Davant bacteris extracel·lulars, el sistema immunitari té com a objectiu
eliminar el bacteri i neutralitzar els efectes de les toxines.
Resposta innata davant de Bacteris Extracel·lulars.
Per eliminar els bacteris extracel·lulars, el principal mecanisme efector de la resposta
innata és la fagocitosi. La fagocitosi consisteix en internalització dels bacteris extracel·lulars
per tal que pugin ser eliminats per les cèl·lules fagocítiques.
Les principals cèl·lules que duran a terme la fagocitosi
dels bacteris extracel·lulars són els neutròfils i els
macròfags, que els detectaran mitjançant els receptors PRR.
Els receptors PRR més rellevants per aquesta funció són els
receptors scavenger, els receptors de tipus toll i els receptors
de manosa. Tots aquests PRR poden detectar el bacteri.
La fagocitosi s’afavoreix amb el reconeixement de bacteris opsonitzats per
immunoglobulines o proteïnes del sistema del complement. Aquest reconeixement es fa
a través de receptors de la fracció constant de les immunoglobulines o receptors del sistema
del complement. En la fagocitosi s’estimula la capacitat bactericida dels fagòcits. La fagocitosi
depèn molt dels receptors PRR.
Hi ha molts snips que determinen la capacitat o resposta dels PRR. Hi ha snips de TLR4
que afavoreixen la resposta a LTS mentre que n’hi ha de TLR2 que afavoreixen la detecció
del peptidoglicà. Per tant, els snips fan que la identificació i fagocitosi dels bacteris
extracel·lulars sigui més eficient.
Toxines
Exotoxines
Endotoxines
Inflamació
Destrossa del sistema immunitari
Formació de pus
Receptors PRR
Scavenger
Tipus Toll
Manosa

Immunologia 2020
CEBIO 190
Els elements del sistema del complement també contribueixen a la resposta innata
contra els bacteris extracel·lulars. Sobretot els elements de la via alternativa o de la via de
les lectines. La via alternativa contribueix perquè components com el LPS són capaços
d’iniciar el trencament espontani de C3. Per altra banda, la paret cel·lular de molts bacteris té
una alta concentració de carbohidrats i polisacàrids que es poden reconèixer per elements
que activen la via de les lectines. L’activació del complement genera opsonines i
anafilotoxines. Les opsonines faciliten la fagocitosi. Les anafilotoxines indueixen tant la
inflamació com el reclutament de leucòcits.
Els macròfags produeixen les citocines IL-12 i TNF-α. Les cèl·lules NK produeixen
Interferó γ. El conjunt d’aquestes citocines proinflamatòries i les quimiocines que atrauen als
leucòcits al punt de la infecció seran molt importants per respondre als bacteris extracel·lulars.
Es produeixen danys en el teixit. El teixit produirà DAMPs. Els DAMPs seran detectats per
PRR i contribuiran a la inflamació. La inflamació consisteix en l’atracció de cèl·lules i
productes del sistema immunitari per tal d’eliminar els bacteris.
La importància del sistema del complement en la resposta innata depèn parcialment de
si són gram positius o gramnegatius. Els bacteris gramnegatius són susceptibles a la lisi
directa pel complement. En el cas dels bacteris grampositius, el gran gruix de la paret
bacteriana impedeix la formació del MAC, pel que no solen ser susceptibles a la lisi pel
complement. Així i tot, els elements del sistema del complement actuen com a opsonines, el
que facilita la seva fagocitosi i eliminació.
Resumint la immunitat innata, l’entrada del bacteri extracel·lular provocarà l’activació del
complement. El sistema del complement produirà opsonines i anafilotoxines. Ambdues
activaran algunes cèl·lules de la immunitat innata, com els mastòcits perquè es desgranulin i
alliberin histamina. Les anafilotoxines també actuaran com a elements quimiotàctics
juntament amb la IL-8 i mediadors d’inflamació. Els elements quimiotàctics podran atraure
cèl·lules fagocítiques per tal que intentin fagocitar els bacteris i aconseguir la seva eliminació.
Les cèl·lules de la immunitat innata que s’activaran produiran citocines
proinflamatòries que augmentaran les molècules d’adhesió i la permeabilitat vascular.
Això activarà la fase aguda.
Els bacteris procariotes extracel·lulars inicien les seves seqüències proteiques per
metionines formilades. Per tant, els bacteris generen pèptids formilats, els quals actuen com
a agents quimiotàctics que detecten les cèl·lules del sistema immunitari.
Entrada del bacteri
extracel·lular
Activació del sistema del complement
Producció d’opsonines i anafilotoxines
Activació de cèl·lules de la
immunitat innata
Alliberament d’histamina
Quimiotaxi de les cèl·lules
fagocítiques
Producció de citocines
proinflamatòries
Augment de molècules
d’adhesió i de permeabilitat
vascular

Immunologia 2020
CEBIO 191
Resposta adaptativa humoral davant de Bacteris Extracel·lulars.
En el cas dels bacteris extracel·lulars, el
principal mecanisme efector de la
resposta adaptativa és la resposta
humoral. Això és així perquè els principals
antígens bacterians són polisacàrids. Aquest
fet implica que generen una resposta a
antígens T-independents, la qual genera
plasmoblasts de vida curta. Els plasmoblasts
de vida curta produeixen anticossos poc
potents durant un temps limitat.
La baixa potència dels anticossos es deu a no fer un procés d’hipermutació somàtica,
a generar poca memòria immunològica i a que les cèl·lules plasmàtiques no tinguin
una vida llarga. És per això que la majoria de resposta a antígens T-independents són
respostes de producció de IgM i algunes poques IgG.
També hi ha una resposta dels limfòcits
B dependent de la col·laboració amb els
limfòcits T. En aquest cas, la resposta sol
ser a antígens proteics, el que la fa més
efectiva. La resposta a antígens proteics
provoca la resposta del centre germinatiu, el
qual permet produir anticossos d’alta afinitat
i amb canvi d’isotip. L’isotip dels
anticossos que surten del centre
germinal és el que millor s’adapta a la
resposta. Generalment, els isotips són IgG
i IgA generats per cèl·lules plasmàtiques de
vida llarga. Per tant, els anticossos tenen
una vida mitjana llarga i es genera una bona
memòria.
La resposta humoral produeix anticossos. Els anticossos
tenen tres mecanismes efectors importantíssims. D’una
banda, els anticossos han de neutralitzar els bacteris i les
toxines bacterianes. La neutralització consisteix en impedir
que els bacteris i les toxines s’uneixin a la cèl·lula de l’hoste.
En la neutralització són molt importants la IgA i la IgG. La IgG
depèn de la superfície.
Els anticossos ajuden en l’opsonització i fagocitosi dels
bacteris. En l’opsonització i fagocitosi és molt important la
IgG al ser la millor per facilitar la fagocitosi a través dels
receptors Fcγ.
Els anticossos faciliten l’activació de la via clàssica del
complement. Per tant, els anticossos facilitaran tots els
mecanismes derivats de l’activació del complement com la lisi
directa via MAC o la col·laboració en l’opsonització.
Tots els mecanismes han de tendir a impedir que la toxina o
el bacteri entrin en contacte amb les cèl·lules del teixit que fan de barrera.
Polisacàrids com antígens
Resposta a antígens T-independents
Plasmoblasts de vida curta
Anticossos poc potents durant temps limitat
Antígens proteics
Resposta del centre germinatiu
Anticossos d'alta afinitat i canvi d'isotip
Anticossos de vida mitjana
Memòria
Neutralització bacteris i toxines
Impedir unió a cèl·lula hoste
IgA + IgG
Opsonització i fagocitosi bacteris
A través de receptors Fcγ
IgG
Facilitar via clàssica complement
Facilitar lisi directa
Col·laborar en opsonitzar

Immunologia 2020
CEBIO 192
Resposta adaptativa cel·lular davant de Bacteris Extracel·lulars.
La participació de la resposta
adaptativa cel·lular és molt important
en la resposta als bacteris
extracel·lulars. Si són bacteris
extracel·lulars, seran reconeguts pels
fagòcits a través de la via endocítica. En
conseqüència, els bacteris seran
processats i presentats per molècules del
MHC de classe II a limfòcits Thelper CD4,
el que en provocarà l’activació i
diferenciació. Els limfòcits es diferenciaran
en els seus diferents tipus efectors en
funció del patogen i la via d’entrada. Cada
tipus de limfòcit T efector secreta unes
citocines concretes i té unes tasques
específiques.
Si els limfòcits es diferenciaran en limfòcits Th17, es produeix molta IL-17. Si els limfòcits es
diferenciaran en limfòcits Th1, es produeix molt de TNF. Els limfòcits Th1 i Th17 atrauen i
activen macròfags i neutròfils. Tant els macròfags com els neutròfils tenen alhora una
capacitat fagocítica bactericida i una capacitat de produir citocines proinflamatòries.
Si es produeix molt INF-γ, s’activen els
macròfags. L’activació dels macròfags fa
augmentar la fagocitosi i lisi bacterianes. Els
macròfags provoquen el manteniment de la
diferenciació en limfòcits Th1, els quals
produeixen INF-γ.
En la resposta adaptativa cel·lular a bacteris extracel·lulars prenen molta rellevància les
citocines que realitzen accions en la col·laboració amb els limfòcits B, doncs el principal
element en la resposta adaptativa a bacteris és la resposta humoral.
Els limfòcits TCD4 helper fol·liculars contribuiran a l’activació dels limfòcits B en la
producció del centre germinal i per a totes les reaccions que hi tenen lloc. Les reaccions
que es donen en un centre germinal són la hipermutació somàtica i el canvi d’isotip. La
col·laboració entre limfòcits T helper CD4 i limfòcits B augmenta la producció d’anticossos.
Bacteri extracel·lular
Reconeixement per fagòcits
Processament Bacteris
Presentació a limfòcits T helper per MHC II
Diferenciació en limfòcits T efectors
Secreció CitocinesActivació cel·lules
fagocítiques
IL-17 TNF
Th17
Th1
Macròfags
Neutròfils
Diferenciació limfòcits
Producció INF-γ
Activació macròfags
Diferenciació Th1
Producció INF-γ

Immunologia 2020
CEBIO 193
Els ThCD4, produint IL-4 i IL-5, contribueixen en l’activació de les cèl·lules del sistema
immunitari com els eosinòfils. Aquests són importants en l’eliminació dels bacteris
extracel·lulars per la desgranulació i alliberament de factors microbicides. La IL-4 contribueix
a la activació i proliferació dels limfòcits B i la seva diferenciació a cèl·lules plasmàtiques
productores d’immunoglobulina.
La infecció per bacteris que colonitzen
preferentment les mucoses, com S.
pneumoniae o H. pylori, sovint es tradueix en
la generació de cèl·lules efectores Th17.
L’apoptosi de cèl·lules de la mucosa en
un entorn inflamatori proporciona una
combinació de PAMP i DAMP, el que
indueix a les cèl·lules dendrítiques a produir
TGFβ i IL-6, el que promou la diferenciació a
TH17.
En canvi, una cèl·lula que pateix apoptosi en absència d'infecció produeix només
DAMP, que indueix les cèl·lules dendrítiques a segregar només TGFβ i no citocines pro
inflamatòries, pel que afavoreix la generació de cèl·lules T reguladores.
Les cèl·lules T helper 17 fan IL-17 i provoquen la segregació de IL-22 per part de les
cèl·lules de la mucosa. La combinació d’aquests dos elements fa que les cèl·lules epitelials
produeixin pèptids antimicrobians i factors quimiotàctics que serveixin per reclutar neutròfils.
A més, les cèl·lules dendrítiques activades secreten citocines proinflamatòries que diferencien
cèl·lules TCD4 a cèl·lules Th17 efectores.
Conseqüències patològiques de la resposta a Bacteris Extracel·lulars.
Sovint, la resposta a bacteris extracel·lulars té conseqüències patològiques. Es pot
produir una inflamació local molt aguda quan hi ha una bona combinació de:
Una resposta local molt aguda pot provocar danys als teixits o pot arribar a tenir efectes
sistèmics mitjançant la producció elevada de citocines. Si aquesta producció és massiva, es
pot arribar a produir un xoc sèptic.
ThCD4
IL-4
Activació i proliferació limfòcits
B
Diferenciació limfòcits B a
cèl·lules plasmàtiques
Activació Eosinòfils
Desgranulació
Alliberament factors microbicides
IL-5
Entorn inflamatori
Apoptosi cèl·lules de la mucosa
Producció PAMP i DAMP
Producció TGFβ i IL-6
Diferenciació a Th17
Resposta innata
Limfòcits Th17 Anticossos

Immunologia 2020
CEBIO 194
Un xoc sèptic és l’activació de tots els endotelis. Per tant, un xoc sistèmic és la producció
massiva de citocines inflamatòries que es troben a tots els teixits i òrgans. Un xoc sèptic
provoca:
• L’efecte exagerat de les citosines.
• Hipotensió.
• Col·lapse circulatori sistèmic.
• Coagulació intravascular disseminada.
• Altres canvis metabòlics que poden acabar en una fallada multiorgànica.
D’altra banda, si la producció d’anticossos és exagerada, hi pot haver diverses
conseqüències:
• Els anticossos poden donar lloc a immunocomplexes de mida molt gran que
s’acumulen als glomèruls renals.
• Els anticossos poden tenir una reactivitat creuada amb proteïnes de la membrana de
la cèl·lula muscular i la miosina del cor.
• Pot haver-hi una patologia autoimmunitària.
L’acumulació d’immunocomplexes de mida molt gran als glomèruls renals hi produeix una
inflamació i un mal funcionament dels ronyons. La reactivitat creuada dels anticossos amb
proteïnes de la membrana de la cèl·lula muscular i de la miosina del cor genera problemes a
nivell cardíac.
Mecanismes d’evasió dels bacteris extracel·lulars.
Hi ha quasi tants mecanismes d’evasió del sistema immunitari com espècies
bacterianes que poden infectar l’ésser humà. El primer pas d’una infecció és l’adhesió a
una cèl·lula epitelial o a una cèl·lula que s’infectarà. Molts bacteris tenen incrementada
l’adhesió no específica que els permet interaccionar amb les cèl·lules de barrera o hoste.
Aquest increment en l’adhesió augmenta la infecció.
També han desenvolupat mecanismes per combatre la fagocitosi. E. coli i Pseudomonas
aeruginosa secreten molècules que disminueixen la fagocitosi per neutròfils. Staphylococcus
aureus inhibeix la fagocitosi que es realitza a través dels receptors dels dominis Fc de les
immunoglobulines. Tenen una proteïna que expressen a la superfície bacteriana i s’hi uneix.
Per això són capaços de bloquejar la interacció d’aquests receptors amb les
immunoglobulines.
Els bacteris també han desenvolupat modificacions a l’estructura antigènica externa.
És el que es coneix com Variació antigènica. Són modificacions que eviten ser detectats per
elements del sistema immunitari. E. coli, per exemple, presenta més de 106 antígens diferents
que poden permetre evadir-se del sistema immunitari. Alguns bacteris gramnegatius
presenten alteracions estructurals del LPS que el permeten passar desapercebuts pels TLR4.
També hi ha bacteris que han desenvolupat mecanismes d’inhibició del sistema del
complement. El bacteri Salmonella typhimurium, per exemple, expressa un gen RcK que
inhibeix la formació del MAC, impedint la lisi mediada per complement. Però hi ha
mecanismes per evitar tots els passos del sistema del complement. Pot ser que la paret
cel·lular o la membrana externa amb càpsula o càlix impedeixi la unió dels components del
complement. Pot ser que les molècules del bacteri dificultin la interacció de molècules del
sistema del complement amb els seus receptors. Pot ser que el bacteri secreti inhibidors o
proteïnes que impedeixen inserció a membrana i la formació del MAC. Pot ser que els bacteris
produeixin enzims que trenquin, inactivin o desacoblin els components del complement.

Immunologia 2020
CEBIO 195
Tots aquests mecanismes diversos tendeixen a inhibir el sistema del complement sobre els
bacteris extracel·lulars.
Resposta a Bacteris Intracel·lulars. Els bacteris intracel·lulars són inaccessibles per les molècules efectores solubles un
cop han entrat a la cèl·lula. Davant aquests patògens, els anticossos i el sistema del
complement són poc eficients. Així i tot, poden actuar en els moment previs a la infecció
cel·lular.
Els bacteris intracel·lulars es poden eliminar més fàcilment si són presentats a
limfòcits T i aquests són els que contribueixen a la seva eliminació. Els bacteris
intracel·lulars viuen dins la cèl·lula i, en cas que mantinguin un estat latent, poden passar
desapercebuts pel sistema immunitari. El sistema només els detectarà quan causin la mort
de la cèl·lula hoste.
Els bacteris intracel·lulars poden ser com Chlamydia, Rickettsia o Listeria i trobar-se al
citoplasma o poden ser com Mycobacteria, Salmonella, Legionella i Leishmania i trobar-se en
vesícules citoplasmàtiques. La possibilitat de resposta, detecció i escapada del sistema
immunitari també depèn d’aquesta localització intracel·lular.
Resposta innata a bacteris intracel·lulars.
La resposta innata tan sols pot tenir efecte prèviament a l’entrada a la cèl·lula hoste.
S’aconsegueix per fagocitosi del patogen i per l’activació de l’esclat respiratori en els
macròfags infectats. Una vegada aquest bacteri ja és a l’interior de la cèl·lula, el mecanisme
principal és l’acció de les cèl·lules NK. En general, la resposta innata a bacteris
intracel·lulars és poc efectiva i és molt rellevant l’activitat de la resposta adaptativa.
Resposta innata a bacteris intracel·lulars per cèl·lules NK.
L’element principal quan el bacteri ja es
troba dins la cèl·lula són les cèl·lules NK.
Les cèl·lules NK detecten la presència de la
infecció i poden alliberar els seus grànuls,
plens de perforines i granzines, que
acabaran provocant la lisi de la cèl·lula
infectada. Les cèl·lules NK tenen un nivell
baix de CD56 però que tenen molt actius els
grànuls citoplasmàtics amb productes
microbicides.
D’altra banda, les cèl·lules NK també
secreten citocines com l’interferó γ.
L’IFNγ activa la capacitat bactericida de
macròfags. Els macròfags faran IL-12, que
activa les cèl·lules NK i les fa més eficients.
La IL-12 i l’interferó γ fan que la resposta
dels limfòcits TCD4 sigui del tipus Th1. La
resposta Th1 produeix IFNγ i manté el cicle
d’activació, detecció i eliminació de bacteris
intracel·lulars. La contribució més
important és de les cèl·lules NK, que
tenen un nivell de CD56 baix i que actuen
per producció d’enzims i citocines.
Detecció de presència d’infecció
Alliberació dels grànuls
Lisi de la cèl·lula infectada
NK secreten INF-γ
INF-γ activa macròfags
Macròfags secreten IL-12
IL-12 activa cèl·lules NK i Th1
Limfòcits Th1 secreten INF-γ

Immunologia 2020
CEBIO 196
Les cèl·lules NK són imprescindibles en el primer control de la infecció per bacteris
intracel·lulars perquè les cèl·lules Natural Killer contribueixen a l’eliminació dels bacteris
en els primers dies de la infecció. Sovint, aquests mecanismes no són suficients i els
bacteris es queden dins dels macròfags de mode latent. Això provoca infeccions cròniques
que passen desapercebudes.
Resposta adaptativa a bacteris intracel·lulars per limfòcits TCD4 i CD8.
El principal mecanisme de la resposta
immunitària adaptativa a bacteris
intracel·lulars és la resposta cel·lular mediada
per limfòcits TCD4 i TCD8. Un cop els bacteris
han entrat a la cèl·lula, cal destruir la cèl·lula
infectada per eliminar-los. En aquest procés
intervenen els limfòcits TCD4 helper 1, ja que
secreten IFN-γ, el qual activa la capacitat lítica dels
macròfags i estimula la producció
d’immunoglobulina G per part dels limfòcits B.
L’interferó promou aquest canvi d’isotip i, per tant contribueix a la producció d’aquests
anticossos. Les IgG són importants en facilitar l’eliminació dels bacteris intracel·lulars
i en activar el sistema del complement. Activar el sistema del complement permet
augmentar l’opsonització dels bacteris intracel·lulars mentre encara no hagin entrat a l’interior
de la cèl·lula hoste.
D’altra banda, si els bacteris intracel·lulars
s’escapen del fagosoma i entren al
citoplasma de la cèl·lula hoste al evitar la
maquinària proteolítica del macròfag, els
bacteris intracel·lulars es poden
processar i presentar proteïnes del
citoplasma per part de molècules del
MHC de classe I. Els limfòcits TCD8, amb
capacitat citotòxica, es poden activar i
contribuir a l’eliminació dels bacteris
intracel·lulars. Sense l’actuació dels
limfòcits TCD8, els bacteris intracel·lulars
podrien donar lloc a infeccions cròniques al
mantenir-se fora de l’abast de molts dels
mecanismes del sistema immunitari per
detectar i eliminar patògens, cosa que
passa sovint.
La col·laboració dels limfòcits TCD4 és important per tal d’aconseguir una bona
activació i funció dels limfòcits TCD8. Els limfòcits TCD4 ajuden a eliminar els patògens
dels fagosomes, mentre que els limfòcits TCD8 eliminen els patògens que han escapat al
citoplasma. Per tant, els dos tipus de limfòcits contribueixen en la resposta a bacteris
intracel·lulars.
Resposta adaptativa a bacteris intracel·lulars per limfòcits Th17.
En el cas d’alguns patògens com Micobacterium tuberculosis, es produeix l’activació dels
macròfags i generen IL-12. Aquesta activació ja és suficient per activar una resposta de tipus
Th1 i no fa falta la participació dels limfòcits del tipus Th17. Això passa sovint.
Limfòcits Th1 secreten INF-γ
Activació capacitat lítica macròfags
Estimulació producció IgG
Bacteri escapa del fagosoma
Processat
Presentació per MHC-I
Reconeixement per TCD8
Activitat citotòxica

Immunologia 2020
CEBIO 197
Davant altres patògens com Francisquella tularensis o Chlamydia muridarum, no s’indueix la
producció d’IL-12 per part dels macròfags. En aquests casos és important la col·laboració
dels limfòcits Th17, que faran que les cèl·lules dendrítiques produeixin IL-12. Així s’afavoreix
la diferenciació cap a limfòcits Th1, que produiran IFNγ i contribuiran a la seva activitat
bactericida.
Les cèl·lules Th17 també contribueixen al reclutament de neutròfils per la destrucció
de bacteris com Listeria monocytogenes i Salmonella typhimurium. D’altra manera, es
podrien escapar de l’acció dels macròfags.
Conseqüències Patològiques de la resposta a bacteris intracel·lulars. També hi pot haver conseqüències patològiques com a resultat de la resposta a
bacteris intracel·lulars. Es parla de l’aparició de reaccions d’hipersensibilitat i d’altres
manifestacions que depenen de factors genètics dels individus.
La resposta a bacteris intracel·lulars es basa en la destrucció de cèl·lules infectades i
pot tenir conseqüències fisiològiques si es destrueixen aquelles cèl·lules amb una funció en
el teixit.
Moltes de les infeccions per bacteris
intracel·lulars es cronifiquen i sovint
generen granulomes. S’han vist
granulomes a les vies respiratòries en casos
d’infecció per Mycobacterium tuberculosis.
Els granulomes són estructures força
esfèriques que estan formades per una capa
interna de macròfags infectats i que al voltant
tenen limfòcits TCD4 Th1 que intenten
contribuir a l’activitat bactericida d’aquests
macròfags. A la part més externa hi ha tant
limfòcits TCD4 com limfòcits TCD8.
L’objectiu de la formació de granulomes és intentar eliminar el bacteri intracel·lular i
evitar la seva disseminació, ja que els bacteris queden atrapats al seu interior. Si el
granuloma es manté de manera estable, no hi ha conseqüències patològiques i la infecció
queda continguda. Si el granuloma es descomposa, la infecció es reactiva i es disseminen
els patògens fàcilment.
El granuloma és una conseqüència patològica perquè ocupa espai d’un òrgan i alhora
és un mecanisme per mantenir la disseminació.
Mecanismes d’evasió dels bacteris intracel·lulars. Els bacteris intracel·lulars han desenvolupat tota una sèrie de mecanismes d’evasió
molt diversos. Mycobacterium tuberculosis evita la fusió entre el fagosoma i el lisosoma.
Això impedeix que proteïnes del Mycobacterium arribin a la via endocítica, siguin processats,
carregats a molècules del MHC de classe II i es presentin al sistema immunitari.
Legionella monocytogenes escapa del lisosoma al citoplasma. Per tant, s’escapa dels
mecanismes intrínsecs de la via endocítica.
Salmonella typhimurium expressa el gen Rck, que confereix resistència a la lisi mediada per
complement, prevenint la formació del MAC.
Macròfags infectats
Limfòcits TCD4 helper 1
TCD4 + TCD8

Immunologia 2020
CEBIO 198
Mycobacterium Leprae és capaç d’inhibir la formació d’espècies reactives de l’oxigen i del
nitrogen.
Evitar l’activitat microbicida com a mecanisme d’evasió dels bacteris intracel·lulars.
El conjunt d’espècies bacterianes intracel·lulars han desenvolupat mecanismes
d’evasió de l’activitat microbicida de la cèl·lula hoste. Aquests mecanismes inclouen:
• Modificar la superfície de la cèl·lula hoste per resistir o trencar els pèptids
antimicrobians.
• Expressar enzims com la catalasa, que converteixen espècies reactives d’oxigen en
components innocus.
• Impedir la producció d’enzims que generen les espècies reactives d’oxigen i de
nitrogen.

Immunologia 2020
CEBIO 199
Tema 24 – Resposta a Virus.
Els virus. Els virus són paràsits intracel·lulars estrictes, pel que viuen a l’interior de les cèl·lules que
infecten i utilitzen la seva maquinària per a la síntesi de proteïnes. Quan es troben fora de la
cèl·lula, els virus són metabòlicament inerts. Usen la maquinària de síntesi de proteïnes
de la cèl·lula infectada per a la seva replicació, pel que depenen de la cèl·lula hoste per a la
seva replicació. Així doncs, les cèl·lules infectades expressen proteïnes del virus a la
seva membrana. Aquestes cèl·lules reben el nom d’hoste.
Els virus estan formats per material genètic envoltat per una càpsida proteica. Aquesta
estructura té un grau de complexitat variable, doncs difereixen en el tipus de material genètic
que poden tenir. Aquests àcids nucleics poden ser DNA o RNA, de cadena simple o de doble
cadena i poden ser lineals o circulars. A més, els virus difereixen en si disposen d’una envolta
lipídica al voltant de la càpsida proteica.
Tot això els hi dona una elevada complexitat afegida. Per altra banda, els virus tenen una
elevada freqüència de mutacions que dificulten la seva detecció i eliminació. Van canviant
més ràpid que l’adaptació del sistema immunitari. A més, els virus presenten una gran
diversitat de mecanismes d’evasió del sistema immunitari. Entre aquesta diversitat en
destaca la capacitat “d’amagar-se” a l’interior de les cèl·lules. Quan exerceixen aquesta
capacitat, es diu que els virus estan en la seva forma latent dins les cèl·lules. La forma
latent és invisible i reactiva la infecció en determinades condicions.
En general, els virus causen infeccions agudes que activen el sistema immunitari i, per
tant, l’hoste aconsegueix eliminar-los. Però hi ha certs tipus de virus que tenen la capacitat
de ser latents, pel que la infecció pot persistir de forma indefinida.
Rutes d’entrada i infecció de virus humans. Les infeccions de major incidència són les de les vies respiratòries i gastrointestinals.
L’entrada dels virus té lloc, sobretot, a través de les superfícies mucoses, lesions a la pell o
directament via sanguínia. Les superfícies mucoses susceptibles de ser la porta d’entrada de
virus són les mucoses respiratòria, intestinal, genital o ocular. L’entrada de virus per via
sanguina no es dona pas massa sovint. La via d’entrada, sobretot a traves de les superfícies
de les mucoses, determina les característiques del tipus de resposta.
Família Vírica Virus Malaltia
Herpesviridae
Herpes Simplex virus I Herpes labial
Virus Epstein - Barr Mononucleosi infecciosa
Limfoma de Burkitt
Varicella zòster virus Grans i pústules
Poxviridae Variola Virus Verola
Hepadnaviridae Hepatitis B virus Hepatitis
Papoviridae Papillomavirus Berruga
Orthomyxoviridae Influenza virus Malalties respiratòries
Togaviridae Rubivirus Rubèola
Paramyxoviridae
Mumps virus Paperes
Morbillivirus Xarampió
Respiratory Syncytal Virus Malalties Respiratòries
Rhabdoviridae Lyssavirus Ràbia
Retroviridae VIH SIDA
Coronaviridae SARS-CoV SARS
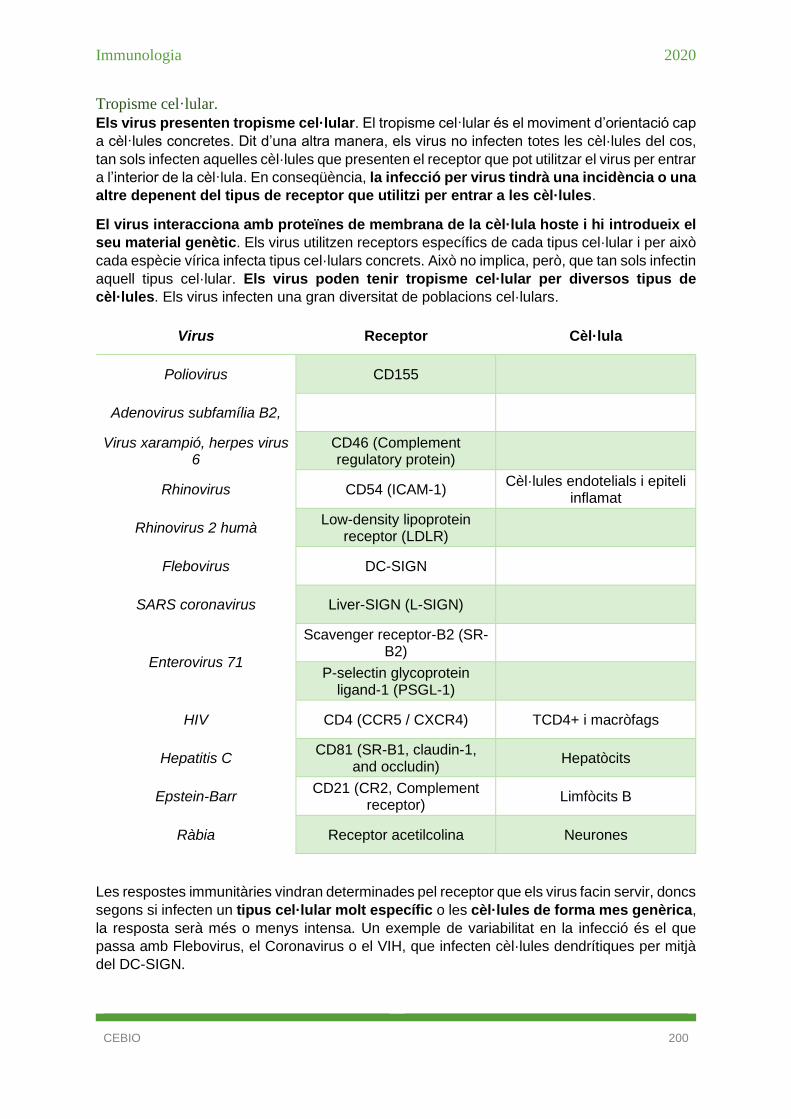
Immunologia 2020
CEBIO 200
Tropisme cel·lular.
Els virus presenten tropisme cel·lular. El tropisme cel·lular és el moviment d’orientació cap
a cèl·lules concretes. Dit d’una altra manera, els virus no infecten totes les cèl·lules del cos,
tan sols infecten aquelles cèl·lules que presenten el receptor que pot utilitzar el virus per entrar
a l’interior de la cèl·lula. En conseqüència, la infecció per virus tindrà una incidència o una
altre depenent del tipus de receptor que utilitzi per entrar a les cèl·lules.
El virus interacciona amb proteïnes de membrana de la cèl·lula hoste i hi introdueix el
seu material genètic. Els virus utilitzen receptors específics de cada tipus cel·lular i per això
cada espècie vírica infecta tipus cel·lulars concrets. Això no implica, però, que tan sols infectin
aquell tipus cel·lular. Els virus poden tenir tropisme cel·lular per diversos tipus de
cèl·lules. Els virus infecten una gran diversitat de poblacions cel·lulars.
Virus Receptor Cèl·lula
Poliovirus CD155
Adenovirus subfamília B2,
Virus xarampió, herpes virus 6
CD46 (Complement regulatory protein)
Rhinovirus CD54 (ICAM-1) Cèl·lules endotelials i epiteli
inflamat
Rhinovirus 2 humà Low-density lipoprotein
receptor (LDLR)
Flebovirus DC-SIGN
SARS coronavirus Liver-SIGN (L-SIGN)
Enterovirus 71
Scavenger receptor-B2 (SR-B2)
P-selectin glycoprotein ligand-1 (PSGL-1)
HIV CD4 (CCR5 / CXCR4) TCD4+ i macròfags
Hepatitis C CD81 (SR-B1, claudin-1,
and occludin) Hepatòcits
Epstein-Barr CD21 (CR2, Complement
receptor) Limfòcits B
Ràbia Receptor acetilcolina Neurones
Les respostes immunitàries vindran determinades pel receptor que els virus facin servir, doncs
segons si infecten un tipus cel·lular molt específic o les cèl·lules de forma mes genèrica,
la resposta serà més o menys intensa. Un exemple de variabilitat en la infecció és el que
passa amb Flebovirus, el Coronavirus o el VIH, que infecten cèl·lules dendrítiques per mitjà
del DC-SIGN.

Immunologia 2020
CEBIO 201
Receptors de lectines tipus C.
El DC-SIGN, un receptor de lectina de tipus C que reconeix carbohidrats, és usat tant per
Citomegalovirus (CMV), VIH i el virus del Dengue per tal d’infectar cèl·lules
dendrítiques. Gràcies al transport d’aquestes cèl·lules dendrítiques, els virus poden viatjar
dels teixits als òrgans limfoides secundaris, facilitant la disseminació del virus.
El receptor DC-SIGN es troba expressat a
totes les cèl·lules dendrítiques que
participen en la resposta innata. En funció
del tipus de cèl·lula, el receptor rep un nom
diferent. Per les cèl·lules dendrítiques c, el
receptor és el DC-SIGN. En les cèl·lules
dendrítiques plasmacitoides, el receptor és el
BCDA-2. Finalment, les cèl·lules de
Langerhans tenen el receptor anomenat
langerina o CD207.
La capacitat mòbil de les cèl·lules dendrítiques a través dels òrgans limfoides
secundaris fa que aquests receptors facilitin la disseminació dels virus. El cas més ben
estudiat és del HIV, que usa DC-SIGN per disseminar-se a través dels òrgans limfoides
secundaris, on pot infectar els limfòcits TCD4.
Cicle vital dels virus. El cicle vital dels virus consta de 4 fases:
La maquinària de síntesi de proteïnes de la cèl·lula infectada pel virus és bàsicament
emprada per a la replicació del virus i no per a la producció de les proteïnes pròpies de la
cèl·lula. Això es deu a què el virus segresta tota la maquinària cel·lular. Aquestes proteïnes
víriques es poden expressar a la membrana i, per tant, la composició de proteïnes de
membrana en la cèl·lula infectada canviarà. Aquest és un element detectat per les cèl·lules
del sistema immunitari.
Entrada a la cèl·lula hoste per interacció amb
receptors específics de membrana.
Fusió amb la membrana de la
cèl·lula i alliberació dels àcids nucleics.
Ús de la maquinària
replicativa de l’hoste, per
sintetitzar les proteïnes
estructurals víriques.
Acoblament de genomes i proteïnes
estructurals formant les
partícules víriques que s’alliberen.
DC-SIGN
DC-SIGN
CD207BCDA-
2

Immunologia 2020
CEBIO 202
Detecció dels virus. Els principals mecanismes que la immunitat innata fa servir per detectar la presència
de virus en l’organisme són els receptors de tipus TOLL, els de tipus NOD i els de tipus
RLR. Això és perquè tots aquests receptors tenen els àcids nucleics en les seves diferents
formes com a principal lligand.
Receptors de Tipus Toll.
Però no tota la defensa es basa en la detecció d’àcids nucleics. Alguns receptors de tipus
TOLL són capaços de detectar la presencia de la hemaglutinina o de l’envolta de determinats
virus. Aquesta detecció permet senyalitzar conforme s’ha detectat la presència d’un virus. Els
detectors dels principals tipus de virus són el TLR2, TLR3, TLR4, TLR7, TLR8 i TLR9. Cada
receptor de tipus TOLL detecta coses diferents:
Les principals famílies víriques amb DNA de doble cadena són els Vesicular Stomatitis Virus
(VSV), el Lymphocytic Choriomeningitis Virus (LCMV), el Herpes Simplex Virus (HSV) i el
Virus Epstein-Barr (EBV).
Les principals famílies víriques amb RNA de cadena senzilla són els virus Influenza, VIH i
VSV.
El HSV és un dels virus amb el DNA de doble cadena amb les illes CpG desmetilades. Aquest
és un factor important ja que el TLR9 pot detectar el genoma del virus tant quan és
alliberat com quan es produeix la replicació.
Els TLR més importants en la resposta a virus són el TLR7, TLR8 i TLR9 (més importants
que TLR3) perquè són els responsables de la detecció dels virus per part de les cèl·lules
dendrítiques plasmocitoides i, per tant, de l’elevada producció d’IFN tipus I (els
interferons de tipus I són un dels mecanismes inicials en la resposta als virus).
Receptors de Tipus R.
D’entre els diferents receptors de tipus R en destaquen dos: RiG1 I MDA5. Cadascun detecta
diferents elements.
• RIG-1 detecta els virus
o Influenza.
o Hepatitis C.
o Vesicular Stomatitis.
o Encefalitis japonesa.
o Measles.
o Human Respiratory Syncytial.
TLR2 detecta MV hemagglutinin protein, HSV i
HCMV.
TLR3 detecta el DNA de doble
cadena.
TLR4 detecta MMTV envelope
protein i RSV.
TLR7 i TLR8 detecten RNA de cadena senzilla.
TLR9 detecta DNA de doble cadena amb les illes CpG
desmetilades.

Immunologia 2020
CEBIO 203
• MDA-5 pot reconèixer
o Poly(I:C).
o Measles virus.
o Sendai virus.
o VSV: vesicular estomatitis virus.
o RNA víric genòmic o generat durant la replicació.
El resultat de la interacció entre el receptor RLR i el seu lligand és sempre la inducció
de la producció d’interferó de tipus I.
Receptors NOD.
Hi ha tan sols dos tipus de receptors de
tipus NOD que participin en la resposta a
virus. La interacció dels virus amb NOD2
acaba produint la senyalització amb la
transcripció de factors de transcripció IRF i,
per tant, la producció d’interferons IFN-α i de
IFN-β.
Quan la interacció és amb NLRP3, s’activa
la formació del complex multi-proteic
anomenat inflamasoma. L’inflamasoma
porta a l’activació de la caspasa 1, que farà
de mitjancera en l’activació proteolítica de la
IL-1β emmagatzemada.
• NOD2 reconeix
o Influenza virus.
o Vesicular Stomatitis Virus.
o Human Respiratory Syncytial Virus.
• NLRP3 reconeix els virus
o Influenza.
o Sendai.
o Adenovirus.
o Vaccinia virus (VACV).
Cèl·lules Dendrítiques Plasmacitoides. Les Cèl·lules Dendrítiques Plasmacitoides són importants en la resposta al virus per la
seva rapida i elevada producció d’interferons de tipus I.
Això és degut que aquestes cèl·lules dendrítiques
plasmacitoides expressen de manera constitutiva
el factor de transcripció IRF-7, reclutat per MyD88 a
través d’un complex molecular que inclou TRAF6 i
IRAK4. MyD88 és una molècula adaptadora que
s’uneix a la cua citoplasmàtica de TLR7, TLR8 i TLR9
quan aquests interaccionen amb el seu lligand. La
unió dels TLR al seu lligand i el reclutament de IRF-7
inicia una cascada de senyalització que ràpidament
indueix la transcripció en grans quantitats dels
gens d’IFN i (interferó α i β).
Interacció Virus – NOD2
Factors Transcripció IRF
IFN-α i IFN-β
Interacció Virus – NLRP3
Inflamasoma
Caspasa 1
IL-1β
MyD88 s'uneix a TLRs
Reclutament IRF-7
Cascada senyalització
Transcripció IFNα i IFNβ

Immunologia 2020
CEBIO 204
Aquestes cèl·lules dendrítiques plasmacitoides són diferents de les cèl·lules
dendrítiques convencionals, ja que la majoria de tipus cel·lulars requereixen de
l’upregulació d’IRF-7 en resposta a IFN tipus I per poder secretar IFN. És a dir, la unió de
TLR7 i TLR9 als seus lligands indueix la secreció de citocines proinflamatòries i quimiocines
però poc IFN (si que s’expressa però a molt baixa concentració).
Per tant, es necessita que es produeixi un augment de l’expressió de IRF-7 per a que es
pugui produir la resposta d’interferons tipus I per a secretar IFN i molt important per
a la resposta del virus (sobretot en les fases inicials). Aquesta necessitat de que hi hagi algun
altre senyal que indueixi la producció de IRF-7 abans de que passi res mes fa que les cèl·lules
dendrítiques plasmacitoides siguin molt eficients en aquesta funció.
Resposta Innata.
Interferons.
La resposta innata principal als virus és la producció d’interferons, sobretot de tipus I
però també de tipus 2 (tipus ). Els interferons tenen un efecte indirecte sobre les cèl·lules
infectades ja que actuen sobre altres tipus cel·lulars.
Els interferons α, β i γ activen les cèl·lules NK i, per tant, les fan més eficients en matar
cèl·lules. A més, l’interferó γ activa els macròfags i les cèl·lules dendrítiques, pel que
augmenten la seva capacitat microbicida (macròfags) i l’expressió de molècules MCH de
classe I i de classe II, importants per a induir una bona resposta adaptativa als virus.
Així i tot, els interferons també tenen un efecte directe sobre les cèl·lules infectades i
sobre les cèl·lules del seu voltant. Aquests interferons, quan s’uneixen al receptor de les
cèl·lules, indueixen l’expressió de tota una sèrie de gens anomenats ISG (gens estimulats per
l’interferó).
Els gens ISG són essencials per a la generació de l’estat antivíric. És a dir, eliminen els
virus de les cèl·lules infectades i protegeixen les cèl·lules del seu entorn per a evitar que, si
hi ha una infecció, aquesta sigui potent i que acabi amb la destrucció de les cèl·lules. L’estat
antivíric consisteix en un estat d’inhibició de la síntesi de proteïnes i de la replicació
del virus arran de l’expressió dels gens estimulats per l’interferó.
Interferons
α
βActivació
NK
γ
Activació macròfags
Activació DC

Immunologia 2020
CEBIO 205
La manera com es produeix aquest efecte directe és perquè la interacció dels interferons amb
el seu receptor desencadena la transcripció i expressió de tota una sèrie de proteïnes
(codificades pels gens estimulats per interferó). Destaquen quatre proteïnes:
• 2-5 oligoadenilat sintasa
• Proteïna quinasa depenent de dsRNA
• Adenosinadesaminasa
• Proteïnes Mx amb activitat GTPasa
La 2-5 oligoadenilat sintasa (OAS) és un enzim que activa la ribonucleasa (RNAsa L).
És una proteïna essencial en aconseguir un estat antivíric, ja que talla el mRNA víric i el de
la cèl·lula eucariota. Malgrat sembli una decisió irresponsable, el sistema immunitari
considera que, en aquest moment de la vida de la cèl·lula, és més important poder tallar el
mRNA víric. Per tant, impedeix la producció de proteïnes del virus.
La Proteïna quinasa depenent de dsRNA (PKR) fosforila la proteïna elF2, inhibint l’inici
de la traducció a la síntesi proteica.
L’Adenosinadesaminasa (ADAR) edita l’RNA introduint-hi mutacions, el que genera
RNA no funcional.
Les Proteïnes Mx amb activitat GTPasa interaccionen amb les proteïnes de la càpsida
del virus, interferint en el seu transport o alterant les seves propietats. Per tant, impedeixen
que les proteïnes de la càpsida del virus arribin a poder-se estructurar.
Per tant, l’efecte directe de l’interferó es pot resumir com que inhibeix la producció de
proteïnes i degrada el mRNA. Com no hi ha síntesi de proteïnes víriques, no es pot produir
la replicació del virus.
Cèl·lules Dendrítiques Plasmacitoides.
A part d’induir l’estat antivíric, els interferons de tipus I estan associats a la interleuquina
12 i indueixen la secreció d’interferons γ fruit de l’estimulació de les cèl·lules NK. Per
tant, els interferons també faciliten l’activació dels limfòcits TCD8 perquè promouen la
diferenciació cap a cèl·lules TCD4 Th1. Es pot dir, doncs, que promouen l’eliminació del virus.
Els IFN-I, juntament amb la interleuquina 6, promouen la diferenciació de limfòcits B a
cèl·lules plasmàtiques. El que afavoreix la producció d’anticossos específics contra el virus.
Les cèl·lules plasmacitoides també produeixen quimosines com CCL3 i citocines
proinflamatòries que permeten reclutar leucòcits i per tant, cèl·lules que podran lluitar
contra la infecció vírica.
Les cèl·lules dendrítiques plasmacitoides no són bones presentadores d’antigen
perquè són molt poc eficients en l’activació dels limfòcits T naive de la resposta adaptativa en
comparació amb les cèl·lules dendrítiques convencionals. Aquesta menor eficiència es dona
al nivell de la fagocitosi, el processament i la càrrega dels pèptids a les molècules del MHC.
Els nivells d’expressió de molècules del MHC de classe II i de molècules coestimuladores en
aquestes cèl·lules plasmacitoides és molt més inferior al de les cèl·lules dendrítiques
convencionals. No s’ha pogut demostrar que puguin activar limfòcits T naïve però sí poden
activar i fer proliferar TH1 i TCD8+ de memòria. Per tant, les cèl·lules dendrítiques
participen, sobretot, en la resposta primària als virus com a cèl·lules detectores del
virus.

Immunologia 2020
CEBIO 206
A les cèl·lules plasmacitoides activades per virus s’augmenta l’expressió de MHC-II i de
molècules coestimuladores (CD80, CD86, CD40) , pel que s’augmenta el seu potencial com
a cèl·lules presentadores d’antigen.
Fases de la infecció vírica i la resposta immunològica.
La magnitud d’una resposta a un virus des del contacte amb aquest fins que es resol
la infecció varia en components i intensitat segons el temps que ha passat des de la
infecció. Els primers elements que es produeixen són els interferons de tipus I, la
Interleuquina 12 i l’activació de les cèl·lules NK.
Els interferons de tipus I, juntament amb la interleuquina 12, provocaran l’activació dels
limfòcits Th1 i la conseqüent resposta adaptativa als virus. Per altra banda, les cèl·lules NK
controlen la infecció vírica durant els primers dies de la resposta. Així i tot, cal la participació
de la resposta adaptativa per poder erradicar completament la infecció.
Citotoxicitat per cèl·lules NK.
Les cèl·lules NK tenen un paper molt important en la resposta als virus. A través de
l’equilibri o la manca d’equilibri entre la senyalització dels seus receptors activadors i els
inhibidors, es determina si maten la cèl·lula diana.
A través d’aquests receptors, les cèl·lules NK reconeixen canvis en les cèl·lules
infectades. Aquests canvis poden ser:
• Modificacions en el patró de glicosilació de proteïnes de membrana de la cèl·lula
infectada.
• Alteracions en els nivells d’expressió de molècules del MHC de classe I a la cèl·lula
infectada.
• Canvis qualitatius en les molècules del MHC de classe I.
• Detecció de l’expressió de molècules induïdes per l’estrès o de lligands dels receptors
activadors induïts pel virus.
Es consideren canvis qualitatius en les molècules del MHC de classe I els canvis en els
pèptids carregats en el MHC-I. Arran d’aquests canvis en la unió, es pot impedir la interacció
de les molècules MHC-I amb els receptors inhibidors i provocar la lisi de les cèl·lules. Són
molècules induïdes per l’estrès les MICA, les MICB i les ILBP.
Les cèl·lules NK actuen alliberant el contingut dels seus grànuls, plens de perforina i
granzima, sobre la cèl·lula infectada. En rebre aquests enzims, la cèl·lula infectada veurà
induïda la seva apoptosi.
Les cèl·lules NK presenten receptors activadors amb lligands diferents del MHC. El
principal és CD16, altrament dit FcγRIII. Són receptors d’elevada afinitat de la regió contant
de les immunoglobulines G, sobretot de IgG1 i IgG3 humanes. Aquest receptor es troba
associat a motius ITAM, el que els atorga la definició de receptors activadors.
La interacció de la IgG específica per una proteïna vírica amb el receptor FcγRIII provoca la
desgranulació de les cèl·lules NK sobre aquelles cèl·lules infectades que expressin les
proteïnes víriques a la seva membrana. En conseqüència, es pot considerar que FcγRIII és
responsable de la citotoxicitat cel·lular depenent d’anticòs.
Les cèl·lules NK també produeixen citocines, sobretot interferó γ. Les citocines
secretades tenen les funcions de:
• Promoure la diferenciació dels limfòcits TCD4 cap a Th1.

Immunologia 2020
CEBIO 207
• Activar, fer créixer i diferenciar les cèl·lules T, B, NK i macròfags.
• Augmentar l’expressió de molècules MHC a les cèl·lules presentadores d’antigen.
• Determinar l’efecte anti-víric directe.
Les cèl·lules NK també sintetitzen TNF-α, el qual:
Finalment, les cèl·lules NK sintetitzen factors de creixement com GM-CSF. Els factors de
creixement serveixen per:
Macròfags i sistema del complement.
Els macròfags també intervenen en la resposta innata als virus, però en aquest cas son
molt mes rellevants les cèl·lules NK. Els macròfags fan les funcions de:
Les molècules antivíriques són interferons de tipus I amb efecte antivíric o que
produeixen espècies reactives d’oxigen i nitrogen. Els principals tipus d’interferons són:
L’actuació del sistema del complement no és massa rellevant de forma directe ja que
els virus són patògens intracel·lulars. Així i tot, el sistema del complement pot contribuir a:
Per tal de provocar les lisis, el sistema del complement ho aconsegueix per mitjà de la
formació del complex d’atac a la membrana.
Resposta adaptativa humoral. La resposta adaptativa humoral és poc eficient i normalment no elimina la infecció. És
important, però, durant l’etapa extracel·lular d’aquesta. En aquest cas, l’objectiu principal és
la neutralització del virus, és a dir, impedir la infecció aprofitant anticossos. Per aconseguir-
ho, els anticossos poden actuar a 5 nivells:
Impedir l’entrada del virus a la
cèl·lula
Activar la via clàssica del sistema del complement
Participar en la citotoxicitat
mediada per anticossos
Facilitar la fagocitosi del
virus
Induir la secreció de citocines
Augmentar la neutralització de
partícules víriques amb anticossos.
Augmentar la fagocitosi de
partícules víriques fent opsonines.
Provocar la lisi directa de partícules víriques amb envolta
lipídica.
Provocar la lisi de cèl·lules infectades.
IFN-α TNF-α NO
Destrucció de les partícules víriques
opsonitzades
Fagocitosi de virus i de cèl·lules
infectades per virus
Producció de molècules
antivíriques
Presentació d’antígens vírics
als limfòcits T
Actua com a mediador paracrí i
endocrí d’inflamació.
Regula el creixement i la diferenciació de
diferents tipus cel·lulars.
Mostra citotoxicitat selectiva per a
cèl·lules transformades
Assegurar la supervivència i creixement
de les cèl·lules mares hematopoètiques
Diferenciar i activar granulòcits i monòcits
Induir al creixement de les cèl·lules endotelials,
eritròcits, megacariòcits i limfòcits T

Immunologia 2020
CEBIO 208
Els anticossos impedeixen l’entrada del virus a la cèl·lula. Això es pot fer impedint la
interacció del virus amb la cèl·lula hoste unint-se a les proteïnes de la càpsida o de l’envolta
vírica que interaccionen amb el receptor a la membrana de la cèl·lula. Una altra opció és
impedir la fusió de l’envolta vírica amb la membrana de la cèl·lula bloquejant els epítops
implicats. A les mucoses, els principals anticossos antivírics són IgA.
Els anticossos també poden contribuir a activar la via clàssica del sistema del
complement i, per tant, provocar la lisi directa dels virus amb l’envolta lipídica o de les
cèl·lules infectades.
També poden participar en la citotoxicitat mediada per anticossos, ja sigui a través de
les cèl·lules NK com per altres cèl·lules efectores. Un tipus de cèl·lules efectores que es
poden fer servir són els neutròfils, doncs també tenen reactors Fc. Així i tot, aquests receptors
són menys eficients.
Els anticossos faciliten la fagocitosi del virus opsonitzant-lo.
També poden impedir la infecció induint la secreció de citocines, les quals estimularan
altres cèl·lules del sistema immunitari.
Resposta adaptativa cel·lular. La resposta adaptativa més important als
virus és la resposta cel·lular. Aquesta és
necessària per inhibir la disseminació dels
virus a les cèl·lules infectades i és essencial
per eliminar el virus una vegada s’ha produït
la infecció.
El principal component de la resposta cel·lular als virus són els limfòcits TCD8,
altrament anomenats limfòcits T citotòxics. Aquests limfòcits acabaran eliminant aquelles
cèl·lules infectades. Per tal que hi hagi una bona resposta a limfòcits TCD8, és important que
hi hagi una bona activació dels limfòcits TCD4 i del tipus efector Th1. Aquestes cèl·lules Th1
contribueixen a l’activació dels limfòcits TCD8.
Els limfòcits TCD8 detecten la presència d’infeccions en cèl·lules perquè aquestes
presenten molècules del complex MHC de classe I carregades amb pèptids derivats del
virus. Aquests complexos MHC-pèptid són detectats pels limfòcits TCD8, que alliberaran la
perforina i gramzina i mataran les cèl·lules infectades. Els limfòcits citotòxics lisen,
específicament, la cèl·lula infectada tot induint-li l’apoptosi.
Paral·lelament a la perforina i a la gramzina, els limfòcits T citotòxics també poden fer
servir les molècules Fas i FasL. Fas és una molècula que es troba en totes les cèl·lules,
mentre que FasL és el lligand de Fas i es troba en els limfòcits TCD8.
L’activitat dels limfòcits T citotòxics contribueix a eliminar la possibilitat de la
replicació vírica.
Els interferons de tipus I s’uneixen al receptor de la cèl·lula infectada i li indueixen un
augment de l’expressió de les molècules del MHC de classe I. Gràcies a això, es pot
considerar que els IFN-I faciliten la detecció i lisi per part dels limfòcits T citotòxics.
Resposta cel·lular
Inhibir disseminació virus
Eliminar virus
Interferons tipus I
Inducció augment
expressió MHC-I
Reconeixement per TCD8
Alliberació Perforina i Gramzina
Lisi cèl·lula infectada

Immunologia 2020
CEBIO 209
Si es tracta d’un virus que infecta cèl·lules dendrítiques, aquestes poden presentar els
antígens vírics units a les molècules MHC-I. En el cas de que sigui un virus que no
presenta tropisme cel·lular per les cèl·lules dendrítiques, aquestes poden internalitzar els
antígens vírics per fagocitosi dels virus o cèl·lules infectades, endocitosi o macropinocitosi;
processar-los i presentar-los units a molècules MHC-I a limfòcits TCD8 per
crospresentació.
La mateixa cèl·lula presentarà antígens vírics internalitzats associats a MHC-II per a ser
presentats als limfòcits TCD4, que tendiran a donar lloc a una resposta Th1, imprescindible
per a l’activació de limfòcits T CD8.
Limfòcits TCD4 i Th1. Els limfòcits T helper 1 contribueixen a l’activació dels limfòcits TCD8 perquè són
imprescindibles per dur a terme l’activació de la cèl·lula presentadora d’antigen i per contribuir
a l’activació dels limfòcits TCD8 a través de la interacció entre CD40 i CD40L. CD40L és el
lligand de CD40 i s’expressa als limfòcits Th1. Els limfòcits Th1 també contribueixen a
l’activació de les cèl·lules TCD8 en tant que són productors de citocines.
Els limfòcits Th1 produeixen interleuquina 2, que contribueix a l’activació i proliferació dels
limfòcits TCD8 i a l’activació de les cèl·lules NK. També produeixen Interferó γ, que indueix
l’estat antivíric i activa tant les cèl·lules NK com els macròfags. Finalment, la tercera citocina
secretada pels limfòcits Th1 és el TNF-α, que recluta i activa macròfags.
Els limfòcits TCD4 helper 1 contribueixen de forma activa en què hi hagi una bona
resposta per part dels limfòcits TCD8. A més, la interacció entre Fas i FasL permet eliminar
directament les cèl·lules infectades. Fas és presentat per les cèl·lules infectades, mentre que
FasL és presentat pels limfòcits Th1.
Les cèl·lules T CD4+ són importants per a aconseguir una bona activació dels limfòcits B,
la qual dona lloc a una resposta humoral duradora i permet el canvi d’isotip. Per tant, la
producció de citocines per part de les cèl·lules T helper fol·liculars determinarà aquest canvi
d’isotip i la producció d’anticossos neutralitzats. També es produirà la maduració de
l’afinitat i la generació d’una bona de resposta i memòria a aquests virus.
El canvi d’isotip és molt important. Per exemple, l’interferó és important per a produir el canvi
d’isotip a IgG2A, que és una de les immunoglobulines que participa més activament en la
inducció de citotoxicitat cel·lular depenent d’anticòs en cèl·lules NK. Per tant, és un
mecanisme més per a la destrucció d’aquests virus.
Resposta immunològica als virus. La resposta als virus té com a objectiu impedir la infecció. Això es pot intentar aconseguir
mitjançant la producció d’IFN-I, que genera l’estat antiviral, i mitjançant la producció
d’anticossos neutralitzants, que impedeixen l’entrada dels virus a les cèl·lules.
Una vegada els virus han infectat les cèl·lules, els mecanismes de la immunitat innata
i de la adaptativa procuren l’eliminació de les cèl·lules infectades. Això ho fan mitjançant
les cèl·lules NK o els limfòcits TCD8, que les eliminen específicament.
Mecanismes d’evasió. Els virus, igual que els bacteris, han desenvolupat molts mecanismes per evitar l’acció
del sistema immunitari.

Immunologia 2020
CEBIO 210
Evitar la senyalització per mitjà dels PRR.
Molts virus generen una sèrie de productes o proteïnes destinades a evitar la
senyalització mitjançant els PRR. Els passos que concretament bloquegen pot ser un o
varis dels següents:
• La unió dels elements de la via de senyalització a l’adaptador MyD88.
• La interacció i producció de IRAK 2.
• La unió dels adaptadors TRIF.
• La interacció dels TLR i els seus lligands.
Per tant, hi ha possibles interferències víriques de cadascuna de les vies de
senyalització.
Inhibició de la presentació per molècules del MHC de classe I.
El principal mecanisme d’evasió de la immunitat adaptativa és inhibir la presentació
per molècules MHC-I de proteïnes víriques. Això es pot aconseguir per mitjà de:
Inhibició de l’activitat del proteosoma
•Impedir la primera fase de la degradació de proteïnes
•Virus Epstein Barr o els Citomegalovirus.
Impedir el transport dels pèptids cap a l’interior del reticle endoplasmàtic per a ser agregats a les molècules del MHC de classe I
•Herpesvirus.
Bloquejar la síntesi de les molècules del MHC de classe I al reticle o retenir-les en el mateix
•Adenovirus i virus de la coriomeninigitis.
Interferir en l’expressió i localització de les molècules del MHC.
Facilitar la destrucció de les molècules del MHC de classe I en el reticle.
Interferir en la presentació d’antigen MHC-I produint proteïnes similars a les MHC-I.
Disminuir l’expressió de molècules coestimuladores.
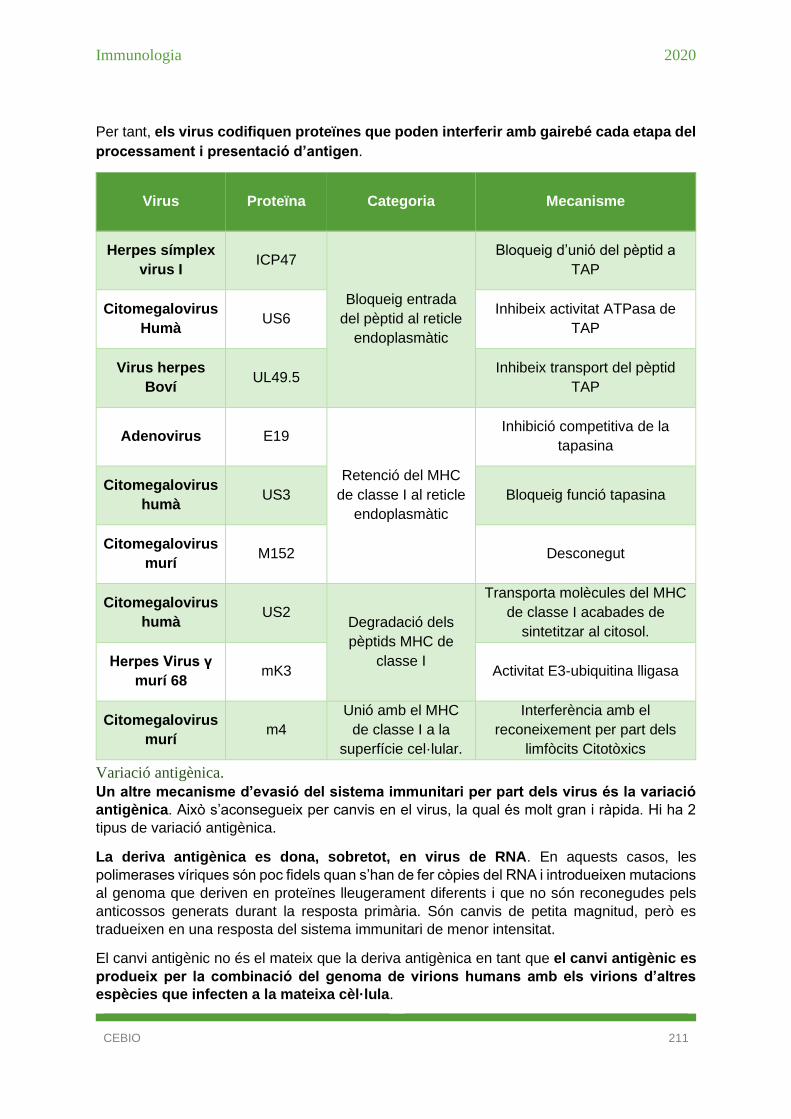
Immunologia 2020
CEBIO 211
Per tant, els virus codifiquen proteïnes que poden interferir amb gairebé cada etapa del
processament i presentació d’antigen.
Virus Proteïna Categoria Mecanisme
Herpes símplex
virus I ICP47
Bloqueig entrada
del pèptid al reticle
endoplasmàtic
Bloqueig d’unió del pèptid a
TAP
Citomegalovirus
Humà US6
Inhibeix activitat ATPasa de
TAP
Virus herpes
Boví UL49.5
Inhibeix transport del pèptid
TAP
Adenovirus E19
Retenció del MHC
de classe I al reticle
endoplasmàtic
Inhibició competitiva de la
tapasina
Citomegalovirus
humà US3 Bloqueig funció tapasina
Citomegalovirus
murí M152 Desconegut
Citomegalovirus
humà US2
Degradació dels
pèptids MHC de
classe I
Transporta molècules del MHC
de classe I acabades de
sintetitzar al citosol.
Herpes Virus γ
murí 68 mK3 Activitat E3-ubiquitina lligasa
Citomegalovirus
murí m4
Unió amb el MHC
de classe I a la
superfície cel·lular.
Interferència amb el
reconeixement per part dels
limfòcits Citotòxics
Variació antigènica.
Un altre mecanisme d’evasió del sistema immunitari per part dels virus és la variació
antigènica. Això s’aconsegueix per canvis en el virus, la qual és molt gran i ràpida. Hi ha 2
tipus de variació antigènica.
La deriva antigènica es dona, sobretot, en virus de RNA. En aquests casos, les
polimerases víriques són poc fidels quan s’han de fer còpies del RNA i introdueixen mutacions
al genoma que deriven en proteïnes lleugerament diferents i que no són reconegudes pels
anticossos generats durant la resposta primària. Són canvis de petita magnitud, però es
tradueixen en una resposta del sistema immunitari de menor intensitat.
El canvi antigènic no és el mateix que la deriva antigènica en tant que el canvi antigènic es
produeix per la combinació del genoma de virions humans amb els virions d’altres
espècies que infecten a la mateixa cèl·lula.

Immunologia 2020
CEBIO 212
Aquesta combinació dona lloc a noves espècies víriques amb proteïnes molt diferents.
Són canvis de gran magnitud, pel que passen desapercebudes pel sistema immunitari. El cas
més clar de virus que realitza un canvi antigènic és el virus Influenza, causant de la grip i que
realitza el canvi entre el virus humà i el virus de la grip.
Inhibició de la sinapsi immunològica.
Alguns virus com el del VIH han desenvolupat estratègies per a desorganitzar la
formació de la sinapsi immunològica de manera que no es pot produir una bona
presentació d’antígens vírics als limfòcits T. Per tant, no es produeix l’activació de la resposta
cel·lular a aquest virus.
Altres mecanismes.
Un dels reservoris coneguts del virus del VIH son les cèl·lules fol·liculars dendrítiques.
Aquestes es troben als fol·licles limfoides i no tenen una maquinària de processament
d’antigen eficient ni molècules de MHC-II. Tenen un sistema de reciclatge de proteïnes de
membrana i d’immunocomplexes que estiguin associats en aquesta membrana, que fa que
el virus sigui internalitzat però que tornin a sortir a la membrana tal i com estaven, no hi ha
una degradació i, per tant, no hi ha una detecció d’aquestes cèl·lules per part del sistema
immunitari.
Una segona opció que tenen els virus per evitar el sistema immunitari és infectar
cèl·lules no permissives o semi-permissives de l’hoste per tal d’emmagatzemar-hi el seu
material genètic i passar desapercebuts al seu sistema immunitari. Els virus es mantenen de
forma latent amb una transcripció mínima o inexistent fins que, per alguna motiu, s’activa.
També hi ha altres virus que presenten mecanismes per a inhibir l’apoptosi, per
endarrerir la mort de la cèl·lula infectada tan com puguin per a poder generar mes virions
infecciosos. Els virus produeixen una sèrie de proteïnes que bloquegen la inducció de
l’apoptosi en cadascuna de les seves etapes, endarrerint la mort de la cèl·lula fins que els
virions estan formats i són infecciosos.
Finalment, hi ha virus com l’Epstein Barr que produeixen anàlegs d’algunes citocines
com la IL-10 i que s’uneixen als receptors substituint les citocines reals. En
conseqüència, interfereixen amb la senyalització de les citocines. Els virus contraresten
l’activitat de les citocines a la resposta produint molècules similars o homòlogues a aquestes
citocines i els seus receptors que s’uneixen i substitueixen les citocines reals i per tant
interfereixen amb la senyalització de les citocines de l’hoste.
Conseqüències patològiques de la resposta a la infecció vírica. Malgrat la resposta immunitària als virus es dona per evitar que aquests acabin provocant
dany a l’organisme, hi ha cops que és el sistema immunitari qui provoca una
conseqüència negativa.
És el cas del dany tissular degut a la resposta dels limfòcits citotòxics, que poden donar
lloc a l’hepatitis crònica quan hi ha una infecció per part de la coriomeninigitis. La destrucció
de les cèl·lules infectades per part d’aquest virus provoca una inflamació crònica, la qual dona
lloc a la hepatitis.
Per altra banda, està la infecció de les cèl·lules del sistema immunitari. Depenent de quines
cèl·lules del sistema immunitari es vegin afectades pel virus, el tipus
d’immunodeficiència que es pot donar és diferent. En el cas del virus del VIH, hi ha una
mort selectiva dels limfòcits T, el que genera una immunosupressió en els individus infectats.

Immunologia 2020
CEBIO 213
El virus Epstein – Barr infecta els limfòcits B i genera una immunodepressió del sistema
immunitari. Finalment, pot passar que la infecció pels virus Epstein – Barr o per Virus
Limfotròpic Humà de Tipus 1 (HTLV-1) provoqui una transformació de les cèl·lules infectades.
També s’ha vist el lligam entre la resposta a determinats virus i el desenvolupament de
malalties autoimmunitàries. S’ha vist que els virus que no poden ser eliminats de manera
efectiva generen una activació crònica i una secreció d’IFN de tipus I de manera continuada.
S’ha vist que és important en el desenvolupament del lupus.
També, la resposta a antígens vírics pots donar lloc a la resposta a proteïnes pròpies.
Per exemple, la infecció del sistema nerviós central pel virus de Theiler’s, el virus de la
hepatitis murina, fa que la mielina sigui diana de la resposta T i B.
Hi pot haver el mimetisme molecular, que fa que hi hagi una similitud prou gran entre
pèptids derivats del virus amb pèptids derivats de proteïnes pròpies. Per tant, una
conseqüència de l’activació de la resposta a la infecció pel virus pot desencadenar una
resposta a pèptids derivats de la proteïna pròpia. Aquest és el cas de la cardiomiopatia
induïda per resposta prèvia a Coxsackie B virus. Finalment, hi ha certa relació entre una
infecció per Coxsackie B virus i el desenvolupament de la diabetis tipus 1.
Quan hi ha una infecció crònica o no ben eliminada de virus que produeixen gran quantitat
d’antígens i aquests no poden ser eficientment neutralitzats pels anticossos de l’hoste, es
formen immunocomplexes. Aquests immunocomplexes poden ser d’una mida gran i pot ser
que siguin difícils d’eliminar. Aquestes característiques provoca que es dipositin als ronyons
o als vasos sanguinis, el que causa una infecció crònica.
En alguns casos, els anticossos poc neutralitzants poden facilitar la internalització dels
complexes anticòs-virus i facilitar la infecció a traves de la unió a receptors Fc de
macròfags. Això s’anomena ADE, antibody dependent enhancement.

Immunologia 2020
CEBIO 214
Tema 25 – Resposta a paràsits i fongs
Paràsits: Protozous i helmints. Un paràsit és un organisme que viu temporalment o permanent damunt o dins d'un
altre organisme d'una espècie diferent, del qual obté l'aliment necessari per a la seva
subsistència sense aportar-li cap benefici a canvi i, sovint, perjudicant-lo. El terme paràsit
inclou els protozous, els helmints i els cucs. Els primers són organismes unicel·lulars i els
altres dos són pluricel·lulars.
Els protozous, en tant que són unicel·lulars i petits, tenen una vida bàsicament
intracel·lular. És també dins de les cèl·lules que es reprodueixen. Així i tot, poden ser
extracel·lulars en algunes fases del cicle extracel·lular.
Els helmints, al ser pluricel·lulars i bastant grans en comparació amb els protozous, no
poden ser internalitzats per les cèl·lules i viuen a la zona extracel·lular.
Aquesta característica determina els mecanismes del sistema immunitari que
s’activaran i contribuiran a eliminar-los.
Les infeccions per paràsits són molt freqüents en la població mundial. S’estima que el
30% de la població mundial en pateix infeccions. Un exemple de les malalties que aquests
organismes poden causar és la malària, que afecta a més de 100 milions de persones i que
en mata un milió anualment. En conseqüència, els paràsits suposen la major causa de
mortalitat entre les diferents malalties infeccioses existents, sobretot en països que es triben
en vies de desenvolupament.
Resposta a protozous. Generalment, els protozous fan infeccions cròniques difícils d’eliminar completament,
el que facilita la transmissió entre individus. Aquests protozous es transmeten utilitzant
animals com a vectors de transmissió, generalment insectes.
Una altra característica que determina la resposta del sistema immunitari davant d’aquests
patògens és que els protozous tenen un cicle vital complex amb canvis de morfologia,
composició antigènica i tropisme cel·lular. Per tant, es fa molt més difícil que el sistema
immunitari sigui eficient en la seva eliminació.
Els protozous són molt més complexes que els bacteris i que els virus, pel que han
desenvolupat molts i diversos mecanismes d’evasió.
Tripanosoma cruzi.
Tripanozoma cruzi és un protozou que presenta una elevada complexitat en les
interaccions amb la cèl·lula de l’organisme hoste. En aquesta interacció, que determina
la resposta del sistema immunitari, hi participen una gran quantitat de molècules
Resposta innata a protozous.
La resposta innata a protozous, en tant que la majoria d’aquests viuen i es repliquen dins
de la cèl·lules, està formada per elements semblants als que s’activen quan hi ha una
infecció per bacteris intracel·lulars. Així i tot, els protozous presenten una gran complexitat
i això provoca que la resposta innata sigui poc eficient. Gràcies a aquesta complexitat, els
protozous evadeixen la resposta innata molt més fàcilment que els bacteris intracel·lulars,
entren a la cèl·lula hoste i hi viuen i s’hi repliquen.

Immunologia 2020
CEBIO 215
Si bé que els protozous evadeixen la resposta innata molt més fàcilment que els bacteris
intracel·lulars, l’organisme no es rendeix a la primera de canvi i no es queda de braços
plegats.
Els mecanismes que fa servir l’organisme per defensar-se del protozou consisteixen
en la fagocitosi per macròfags i en l’activació del sistema del complement. El sistema
del complement dona lloc a la lisi directa dels protozous.
Els macròfags detecten els protozous per mitjà dels receptors del sistema del
complement o els receptors PRR, tot i que hi ha protozous que sobreviuen a l’interior dels
macròfags. Les molècules que els macròfags reconeixen són:
• El glicofosfatidil inositol del tripanosoma, leishmania i plasmòdium per mitjà del TLR2
i el TLR4.
• El lipofosfoglicà de leishmania per mitjà del TLR2.
• Els motius CpG del DNA de Tripanosoma cruzi per mitjà de TLR9.
El glicofosfatidil inositol és l’element del protozou que l’ajuda a ancorar les proteïnes a la
membrana del protozou.
Com també passa en el cas dels bacteris intracel·lulars, les cèl·lules NK continuen tenint
un paper molt important en la defensa de l’organisme davant els protozous. En alguns
casos s’ha vist que poden detectar directament antígens o elements del protozou expressats
a la membrana de les cèl·lules infectades o dels paràsits en la seva fase extracel·lular. El
reconeixement directe suposa la desgranulació de la cèl·lula NK i l’inici de la citotoxicitat
granular. De tota manera, és un fet no del tot comprovat.
Es sap segur, però, que la producció de INF-γ i de TNF-α per part de les cèl·lules NK pot
ajudar a activar els macròfags per fagocitar el protozou i a polaritzar la resposta
adaptativa cap a una resposta de tipus Th1 per afavorir l’eliminació de les cèl·lules
infectades.
Resposta adaptativa a protozous.
Es pot afirmar que, de forma genèrica, la resposta adaptativa a protozous és semblant a
la resposta a bacteris intracel·lulars. Està dominada, generalment, per una resposta
cel·lular de limfòcits TCD4 de tipus TH1 que contribueixen a l’activació dels limfòcits TCD8,
amb capacitat citotòxica. La seva activitat citotòxica els farà eficients en l’eliminació de les
cèl·lules infectades. Però hi ha truc, doncs la predominança de la resposta cel·lular depèn
molt del cicle vital de cada protozou i de cada etapa del cicle.
La resposta cel·lular és important en les fases cròniques, aquelles on el protozou es troba
a l’interior de les cèl·lules de l’hoste, ja siguin macròfags o eritròcits. D’aquesta forma
s’aconsegueix eliminar les cèl·lules infectades i, per tant, són eficients en eliminar la infecció.
Altrament, la resposta més rellevant en aquells casos on el protozou es trobi en la seva
fase extracel·lular serà la humoral mediada per anticossos.
Resposta a Plasmòdium.
Els protozous del gènere plasmòdium són els causants de la malària. Els principals
mecanismes de la immunitat que s’activen ho fan arran del cicle vital d’aquest protozou. Els
paràsits del gènere plasmòdium són injectats a l’organisme en forma d’esporozoïts mitjançant
la picada del mosquit anopheles. A través d’aquesta picada, el protozou entra a la circulació
sanguínia i arriba al fetge. Allà infecta els hepatòcits en 30 minuts, una alta velocitat.

Immunologia 2020
CEBIO 216
Al fetge, dins dels hepatòcits, l’esporozoït es transforma en merozoït i es multiplica de
manera molt ràpida. Això destrueix la l’hepatòcit i permet que el protozou surti a la circulació
sanguínia, on infecta els eritròcits. En aquesta etapa s’inicia de la simptomatologia i la
patologia clínica de la malària. Als eritròcits, es replica i trenca la cèl·lula. Això allibera
merozoïts que infectaran altres eritròcits. Per tant, aquest trencament dels eritròcits causa
anèmia, cansament i la resta de símptomes que es manifesten en la fase
simptomatològica inicial.
Alguns merozoïts alliberats a la circulació sanguínia es diferencien a gametòcits masculins o
femenins, que seran ingerits per un mosquit en el moment de la picada. A l’intestí del
mosquit, els gametòcits es diferencien a gàmetes masculins o femenins i es fusionen
per donar un zigot. Aquests zigots, a la glàndula salival del mosquit es transformen en
esporozoïts. Per tant, recomença el cicle infectiu.
Fase Pre-eritrocítica.
Des del moment que entra a l’individu fins que arriba a la circulació sanguínia, es
considera que el Plasmodium es troba en fase pre-eritrocítica. Durant aquesta fase hi ha
anticossos específics contra proteïnes de membrana de l’esporozoït. Tot i amb això, el temps
que passa entre la picada i l’entrada als hepatòcits és molt curta (uns 30 minuts). Per
consegüent, la resposta humoral per anticossos és poc eficient. Un cop els esporozoïts entren
als hepatòcits, la resposta essencial serà la que provingui dels limfòcits TCD8, que secreten
IFN-γ i provoquen la lisi dels hepatòcits infectats.
Per tal que hi hagi una bona activació de limfòcits citotòxics (els limfòcits TCD8), cal que
també hi hagi una resposta dels limfòcits TCD4 del tipus TH1. Aquests limfòcits TH1
produeixen IFN- que inhibeix el creixement del paràsit dins la cèl·lula infectada i
indueix la secreció d’IL-6 per part dels hepatòcits. La IL-6 indueix la síntesi de proteïna C
reactiva i de proteïnes del sistema del complement, les quals s’uneixen a l’esporozoït i
contribueix a la seva destrucció.
Infecció.
Fetge.
Merozoit.
Sang.
Eritròcits.
GametòcitsMosquit
Gàmetes.
Zigots.
Saliva.
Esporozoït.

Immunologia 2020
CEBIO 217
Encara més, aquest IFN- juntament amb la IL6, activa els macròfags del fetge. Les
cèl·lules de Kuppfer activades, que és el nom que reben aquests macròfags del fetge,
activaran la seva activitat microbicida tot produint espècies reactives de nitrogen (NO), les
quals ajuden a contribuir a la destrucció de les cèl·lules infectades.
Una altra funció de l’IFN- és poder induir a l’expressió de molècules MHC-II per part
dels hepatòcits i, per tant, conferir-los la capacitat de presentar antígens derivats dels
esporozoïts als limfòcits TCD4. Aquests limfòcits, en reconèixer l’antigen, podran contribuir a
l’activació dels limfòcits TCD8.
En conseqüència, es pot afirmar que hi ha tota una sèrie de mecanismes que poden intentar
eliminar els hepatòcits infectats i dificultar la transformació dels esporozoïts en merozoïts, els
quals sortirien cap a la circulació sanguínia. En quant ho intentessin, la producció d’IFN- γ
per part de les cèl·lules NK també contribueix a reforçar aquesta activació del sistema
immunitari per eliminar els hepatòcits infectats i evitar la proliferació.
Fase Eritrocítica.
En el moment que els esporozoïts es converteixen en merozoïts, trenquen l’hepatòcit i
surten a la circulació sanguínia. És el que es coneix com la fase Eritrocítica dels
Plasmodium. Durant aquesta fase, és essencial la resposta cel·lular mediada per limfòcits T
helper. En destaquen:
• Els limfòcits T helper 1
o Que activen macròfags
• Els limfòcits T helper fol·liculars
o Que col·laboren en el canvi d’isotip de les immunoglobulines.
L’activació dels macròfags per part dels limfòcits Th1 permet fagocitar i destruir
paràsits, així com cèl·lules parasitades. D’aquestes cèl·lules, en destaquen els eritròcits.
Durant la fase Eritrocítica, apareixen anticossos específics contra:
• Antígens dels merozoïts com MSP-2 i MSP-3.
• La proteïna PfEMP.
• Molècules tòxiques com GPI.
• Les formes sexuals de plasmòdium.
La proteïna PfEMP (Plasmodium falciparum erythrocyte membrane protein) és una proteïna
de membrana dels eritròcits que prové de plasmòdium, ja que és responsable de
l’adherència i aglutinació dels eritròcits. Aquest fenomen és el que causa les formes severes
de la malària. Per tant, els anticossos dirigits contra la proteïna PfEMP poden arribar a evitar
l’aglutinació alhora que en faciliten la destrucció.
Es pot dir que, durant la fase Eritrocítica, la resposta cel·lular proporciona i facilita la producció
d’anticossos contra els antígens dels merozoïts o dels eritròcits infectats. Això serà essencial
per combatre la infecció.
Resposta a Leishmània.
La Leishmània és un altre protozou que presenta un cicle cel·lular complex que consta de 3
estadis. En l’estadi de promastigot metacíclic, el paràsit es troba lliure a la circulació. Després
hi ha l’estadi majoritari, el qual viu a l’interior de les cèl·lules i hi prolifera. Aquestes cèl·lules
són, majoritàriament, macròfags. A partir d’aquí, l’estadi és d’amastigot. Quan es troba en
aquesta forma, el paràsit entra al vector, que transmet el protozou i li permet reiniciar el cicle
d’infecció.

Immunologia 2020
CEBIO 218
A l’estadi de promastigot metacíclic, la Leishmània és susceptible a la lisi pel sistema del
complement, ja que es permet l’estructuració del sistema d’atac a la membrana (MAC). A
partir de l’estadi d’amastigot, en el qual es troba a l’interior dels fagolisosomes de la cèl·lula
hoste (macròfags o altres), el paràsit evita l’acció dels anticossos. Per tant, el que combat la
infecció és la resposta TH1 que produirà IFN-γ, principalment. Aquesta resposta activa les
capacitats microbicides de macròfags i de cèl·lules dendrítiques, que podran presentar
antígens derivats del protozou.
En el cas de leishmània, els limfòcits TCD8 no tenen un paper important en la resposta a la
infecció primària, però sí que són molt importants en la resistència a la reinfecció. Un cop
s’han activat els limfòcits TCD8 i es disposa de limfòcits TCD8 de memòria, aquests seran
molt eficients en combatre la pròxima infecció.
La Leishmània s’ha estudiat en models de ratolí i s’ha vist que el model de ratolí BALB/c és
molt susceptible a la infecció per Leishmània perquè, en resposta a un epítop de L. Major,
genera una resposta TH2 amb una elevada producció d’ IL4 però no genera IFN-γ.
Mecanismes d’evasió dels Protozous.
Els protozous són resistents a la resposta innata i han desenvolupat molts mecanismes
d’evasió a la resposta adaptativa, pel que aconsegueixen evitar la resposta del sistema
immunitari. És per això que la majoria sobreviuen a la resposta primària i donen lloc a
infeccions cròniques que permeten la transmissió entre individus.
Per la majoria de protozous, la cronicitat es caracteritza per un període de latència en el que
la replicació del paràsit és mínima i la infecció és críptica i passa desapercebuda pel sistema
immunitari. (p.e. Toxoplasma, Leishmania).
A més, la cronicitat també és resultat de l’activació de mecanismes immunosupressors
per part del protozou per evitar la seva eliminació i suprimir la immunopatologia associada a
la resposta immunitària.
Evasió de la resposta innata.
La resposta innata intenta fagocitar els protozous i activar l’esclat respiratori per eliminar-los.
Davant d’això, l’evolució ha dotat els protozous de diferents mecanismes.
Per un costat tenim l’exemple de Tripanosoma cruzii, el qual és fagocitat en forma d’amastigot
però s’escapa al citoplasma produint una proteïna homòloga a C9, que és del sistema del
complement i que forma els porus a la membrana del fagosoma. Aquests porus permeten al
protozou arribar al citoplasma i evitar l’acció de l’esclat respiratori que es dona al fagosoma.
Per altra banda, ens trobem l’exemple de Leishmania. En aquests casos, el lipofosfoglicà
s’adhereix a la membrana interna del fagosoma i impedeix la fusió amb el lisosoma,
bloquejant l’activació de l’esclat respiratori.
També existeixen mecanismes de supervivència davant el sistema de complement que
eliminaria els protozous en les seves fases extracel·lulars.
Per un costat tornem a tenir l’exemple de Tripanosoma cruzii, el qual és susceptible al sistema
del complement activat per la via alternativa, en la seva fase d’epimastigot. Així i tot, aquesta
forma del protozou expressa una glicoproteïna homòloga a DAF anomenada gp160, que
inhibeix la formació de la C3 convertasa. D’aquesta manera, Tripanosoma impedeix l’activació
del sistema del complement.
En el cas de Leishmania, la susceptibilitat a la lisi per part del sistema del complement es
dona en la seva fase dins del vector com a promastigot. Per altra banda, les formes

Immunologia 2020
CEBIO 219
metacícliques d’aquest paràsit han desenvolupat dos mecanismes per evadir-lo. En un cas,
allarguen els fosfoglicans de la paret per impedir la inserció del MAC, evitant així el contacte
entre els components del sistema del complement. En l’altre cas, el paràsit presenta la
proteïna gp63, la qual trenca C3b i impedeix la formació del MAC. El que això no evita és
l’opsonització, ja que la forma resultant del trencament de C3b (iC3b) és reconeguda pels
receptors Mac-1 i CR1. En conseqüència, la forma metacíclica és sensible a l’acció del
sistema del complement fruit de la capacitat d’opsonització gràcies a iC3b.
Evasió de la resposta adaptativa.
El mecanisme d’evasió més important que presenten els protozous és la variació antigènica.
En altres paraules, els protozous introdueixen canvis en els antígens més immunogènics i
redueixen l’eficiència de la resposta mediada pels limfòcits T i B de memòria. Un antigen
immunogènic és aquell que és diana pel sistema immunitari.
En el cas de Tripanosoma, l’antigen més immunogènic és la Surface glycoprotein (VSG), que
recobreix el protozou. És la principal diana de la resposta humoral, pel que aquest protozou
introdueix canvis en aquesta proteïna VSG i provoca que, en una segona infecció, els
anticossos generats en resposta a ella no siguin ja eficients.
En el cas de Giardia lamblia, la família de proteïnes zinc-finger (VPS) són la diana principal
de la resposta T i B. La introducció de canvis en aquestes proteïnes dificulta que, en una
segona parasitèmia, els limfòcits responguin de forma eficient.
Finalment, el cas de Plasmodium falciparum consisteix en canviar molt fàcilment la proteïna
1 de la membrana de l’eritròcit (PfEMP-1), diana de la resposta cel·lular.
També forma part d’aquest conjunt de mecanismes d’evasió de la resposta adaptativa la
desviació cap a una resposta TH2. Una producció molt ràpida d’IL-4 inhibeix l’expressió de
la cadena beta del receptor de la IL-12 i, per tant, indueix una diferenciació de les TCD4 cap
a Th2, molt poc eficient per resoldre la infecció. És poc eficient ja que no facilita la destrucció
de les cèl·lules infectades com ho fan els limfòcits TH1.
La comprensió del conjunt de mecanismes que s’han mostrat expliquen com és possible la
cronificació de les infeccions per part d’aquests patògens.
Conseqüències patològiques.
Aquesta resposta del sistema immunitari pot tenir conseqüències patològiques. En la infecció
per Plasmodium, per exemple, s’ha vist que hi ha possibilitat de tenir una híper-resposta
de Th1 que derivi en anèmies, malàries cerebrals o una simptomatologia variada.
L’anèmia és fruit d’una destrucció exagerada dels eritròcits, estiguin o no infectats. La malària
cerebral es dona quan es produeix una adherència dels eritròcits en l’endoteli vascular i,
conseqüentment, una destrucció dels endotelis. Finalment, la simptomatologia variada que
pot sorgir fruit d’una resposta excessiva a Plasmodium per part dels limfòcits Th1 inclou mal
de cap, nàusees, hipotensió i diarrea. Cal anar amb molta cura amb aquestes malalties, doncs
poden donar lloc a complicacions importants en els individus amb aquesta infecció.
Cal anar amb cura amb els paràsits, doncs una activació de la resposta per part de Th1 i de
macròfags és important en la protecció i eliminació de la infecció però pot ser exagerada i
derivar en unes conseqüències patològiques molt greus.
Resposta a helmints. La resposta als helmints, els cucs, és totalment diferent de la resposta a protozous. Els
helmints són patògens pluricel·lulars i a les diferents etapes del cicle vital poden afectar

Immunologia 2020
CEBIO 220
diferents teixits dins de l’hoste parasitat. Aquesta pluricel·lularitat és un element molt important
a tenir en compte, ja que provoca que tinguin una certa mida que obliga que els helmints
hagin de ser paràsits extracel·lulars.
Resposta innata a helmints.
La resposta del sistema immunitari a helmints serà més semblant a la resposta a bacteris
extracel·lulars però amb la dificultat afegida que els helmints no poden ser fagocitats arran
de la seva mida. Per tant, la resposta innata a helmints està dirigida sobretot a expulsar-los
de l’organisme. Mentre s’intenta, les cèl·lules de la immunitat innata aprofiten per alliberar
substàncies microbicides per intentar destruir-los a l’espai extracel·lular.
Alguns helmints activen la via alternativa del sistema del complement, però molts han
desenvolupat mecanismes de resistència a la lisi mediada per aquest.
La necessitat d’aquesta activitat microbicida extracel·lular fa que la participació de les
cèl·lules innates limfoides de tipus II (ILC2) sigui molt important en aquesta resposta innata,
així com la desgranulació i l’alliberament de substancies microbicides per part d’eosinòfils i
mastòcits o macròfags i neutròfils, alliberant el contingut dels grànuls i destruint els
paràsits. Els eosinòfils i els mastòcits, recordem, es troben a les mucoses i alliberen el seu
contingut arran de la interacció de les immunoglobulines amb el receptor Fce. Els macròfags
i els neutròfils, per la seva banda, alliberen els seus grànuls fruit de la interacció amb els
receptors Fcg.
Entre aquests mecanismes, a part de l’alliberament dels enzims i substancies microbicides
dels grànuls, serà molt important la citotoxicitat cel·lular mediada per anticossos (ADCC).
D’aquesta manera, en els macròfags serà important l’acció de les IgG o les IgA donat que
aquestes cèl·lules tenen receptors Fc per aquestes immunoglobulines. Aquesta interacció
entre les immunoglobulines i els antígens dels helmints serà important per activar aquesta
citotoxicitat cel·lular.
En el cas dels neutròfils, aquesta citotoxicitat cel·lular també serà mediada per anticossos,
sobretot IgG. En el cas de eosinòfils i mastòcits, en canvi, serà mediada sobretot per IgE
unit al receptor Fc que tenen els eosinòfils activats o els mastòcits en estat de repòs. Per tant,
aquesta interacció via els anticossos produirà la desgranulació de les cèl·lules de la immunitat
innata i contribuiran molt a la destrucció del cuc.
Resumint, es pot dir que la citotoxicitat cel·lular mediada per anticossos consisteix en la
unió de les cèl·lules efectores als helmints opsonitzats per les immunoglobulines a
través dels receptors Fc de les cèl·lules fagocítiques. Aquesta unió allibera les molècules
citotòxiques contingudes als grànuls.
Per altra banda, les cèl·lules epitelials de les superfícies de barrera responen als
helmints secretant citocines anomenades alarmines. Les principals cèl·lules epitelials que
realitzen aquesta acció són les del tracte intestinal. Algunes de les alarmines alliberades són
IL-25, IL-33 i TSLP. Aquestes alarmines activen:
• Cèl·lules innates limfoides de tipus II.
• Mastòcits.
• Basòfils.
• Neutròfils.
• Limfòcits Tγδ.
• Cèl·lules dendrítiques.

Immunologia 2020
CEBIO 221
A més, aquestes citocines promouen la diferenciació dels precursors hematopoètics a
cèl·lules de la immunitat innata, pel que potencien la seva acció. Les cèl·lules diferenciades
més destacables són els basòfils, que contribueixen a la producció d’IL-4, i els mastòcits, que
contribueixen a la producció d’alarmines. En afegit, les neurones responen a productes
derivats d’helmints tot secretant Neural Peptide NeuroMedin U (NMU), la qual activa les
Cèl·lules innates limfoides de tipus II.
Aquestes cèl·lules innates, anomenades també ILC2, són molt importants en la
producció d’IL-13 i en la inducció de la secreció d’IL-25 per part de les cèl·lules tuft de
l’epiteli quan es troben activades. IL-25 i IL-33 promouen la secreció d'IL-13 per part de
les ILC2. La IL-13 augmenta la secreció de mucus per part de les cèl·lules de globet de
l’epiteli intestinal, facilitant l’expulsió dels helmints.
A part de la IL-13, els basòfils també produeixen una elevada quantitat de IL-4. Així doncs, el
resultat de tota aquesta cadena d’activació és la inducció d’una resposta adaptativa del tipus
Th2.
Resposta adaptativa a helmints.
La inducció d’una resposta adaptativa de tipus TH2 és important perquè els helmints no poden
ser fagocitats, sinó que han de ser expulsats de l’organisme. Per aconseguir-ho, les cèl·lules
epitelials produeixen factors (TSLP) que inhibeixen la secreció d'IL-12 per part de les cèl·lules
dendrítiques. Les cèl·lules dendrítiques que presenten antígens a cèl·lules TCD4 indueixen
la diferenciació d’aquestes a TH2 o a THF.
La diferenciació a Th2 o a T helper fol·licular farà que es produeixi una elevada quantitat
de IL-5, IL-4 i IL-13. La IL-5 activa els eosinòfils i n’indueix la desgranulació. IL-4 i IL-13
activen els macròfags immadurs per la via alternativa, el que els fa eficients en la reparació
del teixit afectat. Els macròfags immadurs reben el nom de macròfags M2 i la seva funció és
important perquè el pas dels helmints per un teixit hi causa una gran destrucció que
haurà de ser reparada. Aquesta destrossa és fruit de la gran mida dels helmints, la qual és
fruit de la seva pluricel·lularitat.
A més, aquestes cèl·lules TH2 o TH fol·liculars productores de IL-4 seran molt importants en
l’activació dels limfòcits B. La IL-4 promou el canvi d’isotip cap a IgE i a IgG1, pel que aquests
limfòcits B es converteixen en cèl·lules plasmàtiques productores en IgE i de IgG1.
Les immunoglobulines IgE s’uneixen als receptors de Fc dels mastòcits, eosinòfils activats i
dels basòfils. Això les faran molt importants en la destrucció dels helmints a través de la
toxicitat cel·lular depenent d’anticòs per part de les cèl·lules.
Les immunoglobulines IgG1 contribueixen a la citotoxicitat cel·lular a través dels neutròfils i
macròfags.
En conseqüència, aquest serà el mecanisme més important en la destrucció d’aquests
helmints.
Mecanismes d’evasió del sistema immunitari.
L’evolució, com no podria ser d’una altra manera, també ha dotat als helmints de mecanismes
d’evasió de la resposta del sistema immunitari. El seu principal tipus de mecanisme és la
resistència a la lisi mediada pel sistema del complement, el qual frenen per mecanismes
semblants als del protozous. Consisteix en intentar evitar l’estructuració del MAC.

Immunologia 2020
CEBIO 222
Una altre estratègia és “disfressar-se”, adsorbir proteïnes de l’hoste a la seva superfície.
D’aquesta manera, l’helmint intenta passar desapercebut i mimetitzar-se amb el proteoma de
l’hoste.
Finalment, els helmints també poden evadir el sistema immunitari al presentar un tegument
resistent del qual es poden desprendre. Això els permet evitar els mecanismes microbicides
de neutròfils i de macròfags.
Conseqüències patològiques de la infecció per helmints.
S’ha vist que una resposta adaptativa als helmints basada en una resposta de tipus TH2 és
capaç de provocar una producció d’una gran quantitat d’anticossos i donar lloc a
reaccions de hipersensibilitat.
Resposta a Fongs. Els fongs són microorganismes eucariotes que poden créixer, principalment, de dues formes.
Els llevats creixen com a microorganismes unicel·lulars que es reprodueixen per gemmació
asexual. Les floridures creixen en forma de filaments llargs anomenats hifes.
Els fongs són ubics, colonitzen totes les superfícies de barrera. En condicions normals no
suposen cap problema, però la interrupció de les barreres físiques, químiques o microbianes
degut a diferents problemes causen la infecció. Per tant, les infeccions fúngiques són
principalment oportunistes i aprofiten estats d’immunosupressió de l’hoste per proliferar.
Entre aquestes situacions d’immunosupressió es troben la SIDA, els trasplantaments, la
quimioteràpia, les intervencions mèdiques invasives i la malnutrició.
El tractament d’una infecció per fongs és complicada, ja que hi ha pocs anti-fúngics
comercials, la seva producció és difícil i tenen efectes secundaris. Són tractaments de llarga
durada i normalment hi ha l’aparició de resistències.
Candida albicans.
Les infeccions per part del fong Candida albicans s’estableixen en la seva fase de llevat,
durant la qual interaccionen amb la cèl·lula epitelial. Aquest contacte estimula el canvi de
llevat a hifa i la filamentació afavoreix l’adhesió a la cèl·lula epitelial i la internalització activa
o mediada per endocitosi.
Resposta innata a fongs.
La resposta innata a fongs és fonamental. Es produeix per una combinació de les respostes
contra bacteris intracel·lulars i extracel·lulars. A més, en el cas de la resposta a fongs hi
juguen un paper essencial els àcids grassos saturats que tenim a la pell al tenir una important
activitat fungicida. És per això que una neteja massa continuada s’associa amb una
proliferació dels fongs a la pell.
En al resposta a fongs també és molt important l’activació del sistema del complement,
principalment la que es dona per la via de les lectines. Els fongs tenen una elevada quantitat
de carbohidrats a la membrana, pel que la via de les lectines té un rol molt important. Així i
tot, també s’han vist casos on s’ha donat l’activació de la via alternativa del complement.
En el cas de llevats, destaca la intervenció de neutròfils i macròfags per provocar una
fagocitosi o mort intracel·lular del llevat mitjançant la secreció de substàncies fungicides com
les espècies reactives de l’oxigen.
Els fongs, un cop entren al cos, són detectats per PRR i per receptors de lectina tipus C fruit
de la presència de diversos carbohidrats en la membrana. Així i tot, també són detectats per

Immunologia 2020
CEBIO 223
TLR2. El reconeixement dels fongs per qualsevol d’aquests mecanismes indueix la seva
fagocitosi.
Reconeixement de fongs per PRR.
Els fongs tenen tota una sèrie de PAMP, majoritàriament de carbohidrats. En destaquen els
β-glucans, polímers de glucosa, la quitina, un polímer de N-acetilglucosamina, i els manans,
cadenes de centenars de molècules de manosa unides a proteïnes del patogen a través
d’enllaços N- ó O-.
Els PAMPs fúngics són detectats via receptors de lectina tipus C, anomenada dectina o
receptor de manosa, que suposen la senyalització cap a l’interior de la cèl·lula i l’activació de
la producció de diverses citocines. En alguns casos, també hi ha els TLR. N’és un exemple
TLR4, que detecta manans, o TLR2, que detecta fosfolipomanans. Detecten, doncs,
molècules complexes amb base de carbohidrats. TLR3, TLR7 i TRL9 detecten RNA o DNA
desmetilat, propi d’algunes fases d’aquests fongs. En alguns casos també hi ha receptors de
tipus NOD.
Resposta adaptativa a fongs.
La resposta adaptativa a fongs es dona, principalment, per part de la resposta cel·lular.
Aquesta es troba mediada per limfòcits TCD4 Helper 1, que produeixen IFN-γ i faciliten així
l’activació dels macròfags per tal que desenvolupin la seva activitat microbicida. També s’ha
vist una gran importància dels limfòcits Th17, els quals produeixen IL-17 i indueixen l’activitat
microbicida dels neutròfils. S’ha vist que la contribució dels neutròfils en la resposta als fongs
és essencial per una bona resolució de la infecció. Així i tot, no està ben estudiat el paper dels
limfòcits TCD8 en aquesta resposta.
Les cèl·lules dendrítiques, en resposta als fongs, secreten IL-6, IL-23 i IL-12 i indueixen la
proliferació i diferenciació de TCD4 a TH1. A continuació, TH1 via secreció d'IFN-γ activa
els macròfags. Els limfòcits TH17 secreten IL-17 i IL22. IL-17 indueix el reclutament de
neutròfils i IL-22 indueix la producció de β-defensines per part de cèl·lules epitelials, limitant
el creixement i proliferació del fong.
El resultat de la detecció d’aquests PAMPs fúngics a través de receptors de lectina de tipus
C o de TLRs acaben amb l’activació de limfòcits TH1 i de limfòcits TH17. Quin dels dos
predominarà es determina en funció del patró de citocines. En el cas del TLR2, pot donar la
producció de cèl·lules T reguladores que aconsegueixen que la infecció sigui persistent
però que no doni lloc a immunopatologia.
L’activació dels TLR4, juntament amb Dectin-1 i el Receptor de manosa (MR) a les cèl·lules
dendrítiques indueix una resposta per part dels limfòcits Th17. En canvi, la senyalització per
part dels TLR2 indueix una resposta amb cèl·lules T reguladores que fa que la infecció sigui
persistent però sense cap immunopatologia associada.
Resposta adaptativa humoral a fongs.
La resposta humoral no és la més important en la resposta a fongs, tot i que hi participa
contribuint en la opsonització dels fongs o la citotoxicitat cel·lular depenent d’anticòs amb
diferents isotips d’immunoglobulina, que són més o menys rellevants depenent del tipus de
fong. No es coneix que hi hagi una contribució de IgD en la resposta a fongs, com tampoc es
coneix massa si funciona la IgE. Els anticossos, però, són molt útils per diagnosticar la
infecció per fongs, pel que tenen una transcendència a la clínica.

Immunologia 2020
CEBIO 224
La resposta adaptativa humoral a fongs va dirigida contra antígens de la paret cel·lular.
Aquests poden ser β-glucans com els de A. Fumigatus i C. Albicans, Agglutinin-like sequence
3 com en C. Albicans o Glucuronoxylomannan com el de C. Neoformans.
Mecanismes d’evasió dels fongs.
D’entre els diferents mecanismes que la selecció natural i les mutacions han regalat als fongs,
en destaquen 5.
Per un costat, els fongs poden augmentar la mida per l’increment de la càpsula de la càpsula
de polisacàrids, el que impedeix i dificulta una fagocitosi efectiva. Això ho poden fer tant els
llevats tot formant les Giant cells com formant hifes.
Altrament, els fongs han aconseguit interaccionar amb els elements del sistema del
complement per així inhibir la formació de C3b.
En un altre ordre de coses, han aconseguit mecanismes que eviten la fagocitosi i mecanismes
que inhibeixen la funció i formació del fagolisosoma. En conseqüència d’aquests últims
mecanismes, eviten l’acció dels enzims que podrien dur a terme al seva destrucció en els
lisosomes. Un exemple n’és C. albicans, que altera el trànsit al citoplasma de la cèl·lula al
bloquejar l’associació entre el nucli i el reticle endoplasmàtic.
Finalment, l’últim mètode que tenen els fongs per evitar el sistema immunitari és la variació
dels polisacàrids de membrana, pel que passen desapercebuts més fàcilment en una segona
infecció.

Immunologia 2020
CEBIO 225
Tema 26 – Reaccions d’Hipersensibilitat El sistema immunitari ha de tenir l’habilitat de posar-se en marxa quan sigui necessari,
fer la seva funció i tenir mecanismes de control que aturin la resposta i tornin a la
situació d’homeòstasi. Així i tot, aquesta precisió pot fallar i provocar l’aparició de patologies
relacionades amb un mal funcionament del sistema immunitari. Les patologies que s’associen
a un mal funcionament són:
L’autoimmunitat és una resposta excessiva a estructures pròpies. Si es dona, el sistema
immunitari no ha aprés correctament a no respondre a allò que és propi. La immunodeficiència
consisteix en una resposta deficient a antígens exògens. La hipersensibilitat és una resposta
excessiva a molècules innòcues.
Les reaccions d’hipersensibilitat donen lloc a patologies utilitzant els mateixos
mecanismes que intervenen a una resposta immune normal. El problema es troba en què
la resposta s’està donant a molècules innòcues de forma excessiva o innecessària. Les
reaccions que es donen en casos d’hipersensibilitat estan intervingudes per elements
humorals o cel·lulars de la immunitat adaptativa. Els elements humorals principals de la
hipersensibilitat són els anticossos.
Tipus d’hipersensibilitat. Les reaccions d’hipersensibilitat es divideixen en 4 tipus, tot i que en alguns documents
podem trobar-ne fins i tot 5. Les reaccions hipersensibles es classifiquen en funció de
l’element del sistema immunitari que estigui implicat en el seu desenvolupament.
Poden ser anticossos, immunocomplexes o limfòcits T que donen lloc a diferents resultats.
Els mecanismes dels tipus de reacció d’hipersensibilitat es solen solapar en una mateixa
patologia.
No totes les hipersensibilitats es detecten amb la mateixa rapidesa. Hi ha que es donen de
resposta immediata, com les de tipus I, II i III. També hi ha respostes que són sistemes
d’hipersensibilitat tardana, com la resposta cel·lular de tipus IV. La resposta de tipus IV que
es deguda a limfòcits T i això és el que la fa més lenta.
Tipus d’Hipersensibilitat
Element del S. I. implicat
Resultat Temps de resposta
I IgE Al·lèrgia clàssica Immediata
II IgG citotòxica – IgG
activadora
Reconeixement de receptors, citotoxicitat
Immediata
III IgG -
Immunocomplexes Dipòsit
d’immunocomplexes Immediata
IV Limfòcits T Resposta cel·lular T Tardana
Autoimmunitat Immunodeficiència Hipersensibilitat

Immunologia 2020
CEBIO 226
Hipersensibilitat de tipus I Pràcticament, parlar d’hipersensibilitat de tipus I és parlar d’al·lèrgia. Aquesta és una
paraula molt bonica que prové del grec allos ergos, reactivitat alterada. La resposta de tipus
I es basa en una resposta exagerada i no protectora enfront d’un element innocu,
habitualment intervinguda per immunoglobulina. L’element innocu rep el nom d’al·lergen.
Es tracta d’una resposta a antígens generalment solubles i innocus. Per la població en general
no suposen un problema i no se sol respondre a ells. La hipersensibilitat de tipus I és
deguda a la producció exagerada d’anticossos tipus IgE, que causen una activació de
mastòcits. Desencadena en una rinitis al·lèrgica, asma o al·lèrgies alimentàries. La
hipersensibilitat de tipus I és el fenomen immunopatològic més freqüent i afecta a més d’un
20% de la població.
El terme al·lergen es refereix a antígens sovint ambientals i generalment innocus
l’exposició repetida als quals genera, en individus amb predisposició, una resposta
al·lèrgica. En general, els al·lèrgens:
• Tenen un baix pes molecular que els permet transmetre’s per l’aire.
• Són molt solubles i això fa que es difonguin bé pels teixits.
• Són molt estables, pel que es poden mantenir durant temps llargs i suposen un
contacte continu amb el sistema immunitari.
• Actuen com haptens en alguns casos, afegint-se a proteïnes de l’hoste i generant una
resposta que d’altre forma no generarien.
• Solen tenir activitat enzimàtica, la qual acostuma a ser proteolítica i els facilita el pas
a través de les mucoses.
Aquesta activitat enzimàtica els permet entrar en els teixits i generar la resposta al·lèrgica.
L’activitat enzimàtica es pot trobar en els al·lèrgens dels àcars, fongs, plantes i paràsits.
Un dels principals al·lèrgens que es coneix és els dels àcars. Molts dels que es coneixen
són proteases (Der p 1, 3, 6, 9). Aquests enzims es troben a les femtes dels àcars, pel que
l’al·lèrgia tampoc és contra els àcars en si.
Els al·lèrgens proteases dels àcars trenquen les unions estretes i penetren la barrera
de cèl·lules epitelials, interactuant amb les cèl·lules dendrítiques que activen la resposta del
sistema immunitari i generen la resposta dels mastòcits i la seva desgranulació.
També són proteases els al·lèrgens del pol·len i de la saliva de gat. La gent que és
al·lèrgica als gats, majoritàriament no és al·lèrgica a l’animal en si ni al pèl; sinó a la saliva
dels gats perquè s’hi troben aquestes proteases. Els gats es llepen el pèl per netejar-se, pel
que la saliva queda enganxada a aquest i és el que provoca la reacció majoritària d’al·lèrgia
als gats. Les proteases al·lergèniques també són injectades per picades d’insectes que
contenen larves de paràsits que contenen proteases.
Tanmateix, no tot són proteases, però si una bona part. Per exemple, components
alimentaris o de verins d’abelles o antibiòtics indueixen una reacció d’hipersensibilitat
de tipus I.

Immunologia 2020
CEBIO 227
Atòpia.
Generalment, els al·lèrgens no estimulen respostes innates potents, de forma que no
donen lloc a respostes de tipus Th1 o Th17. Sinó que una exposició repetida dels limfòcits T
sense una bona resposta innata promou una resposta de TH2.
Aquest fet no hauria de ser un inconvenient, però en individus atòpics fa que es generi una
resposta excessiva de producció d’IgE en resposta als al·lèrgens. Són individus atòpics
aquells que tenen una predisposició genètica a induir una resposta de tipus Th2. La
producció de IgE en individus atòpics és de l’ordre de 10 vegades superior a la resposta a
al·lèrgens en individus no atòpics.
La producció excessiva d’IgE és la que dona lloc a una reacció d’hipersensibilitat de
tipus I. En individus no atòpics, la resposta als mateixos al·lèrgens (antígens) indueix la
producció de baixos nivells d’IgG, que no dona lloc a cap mena de patologia.
Susceptibilitat.
Hi ha una predisposició genètica a donar lloc a reaccions d’hipersensibilitat de tipus I.
Un dels gens més implicats en l’atòpia és el que codifica per la IL-4. S’ha vist que hi ha snips
dels promotors de la IL-4 que fan que es produeixen quantitats exagerades d’aquesta
interleuquina. Sabem que aquesta IL-4 promou el canvi d’isotip cap a IgE. Per tant, una
producció excessiva de la interleuquina en resposta a un antigen que dona lloc a una resposta
desmesurada de producció d’IgE. La producció desmesurada de IgE davant un antigen
que hauria de ser innocu és una reacció d’hipersensibilitat de tipus I.
També s’han descrit reaccions d’hipersensibilitat de tipus I en individus amb variants
estructurals de la cadena α del receptor de la IL-4. La cadena α del receptor de la IL-4 fa
que s’assenyali amb una intensitat diferent la resposta a la IL-4.
En la taula també veiem canvis estructurals del receptor Fc d’alta afinitat de la IgE i d’altres
que no estan tan relacionats amb la IL-4 o la IgE, però el resultat de l’acció d’aquests gens
també afavoreix que hi hagi aquesta hipersensibilitat de tipus I.
Gen Tipus de polimorfisme Afectació en
hipersensibilitat
IL-4 Variació del promotor Variació en expressió
Cadena α IL-4R Variació estructural Increment en senyalització
Cadena β receptor IgE Variació estructural Canvis en conseqüències
unió IgE - antigen
MHC de classe II Variants estructurals Millor presentació pèptids
d’al·lèrgens
Receptor α de les cèl·lules T
Marcadors microsatèl·lits Increment reconeixement pèptids d’al·lèrgens per
cèl·lules T
TIM Variants de promotor i
estructurals Regulació de l’equilibri
Th1/th2
Procés previ a la reacció d’hipersensibilitat de tipus I.
La resposta d’hipersensibilitat és immediata però requereix que hi hagi un procés previ.
Hi ha una primera exposició a l’al·lergen en l’individu que segurament causa una mínima
reacció en l’individu que no es detecta. Tanmateix, en individus de predisposició genètica, tal
com hem descrit, suposa una sensibilització.

Immunologia 2020
CEBIO 228
Això provoca que, en una segona exposició a aquest mateix antigen, una mínima exposició
al al·lergen desencadena una resposta molt ràpida que pot ser molt severa. Aleshores
apareix tota la simptomatologia clínica associada a la hipersensibilitat de tipus I en qüestió de
minuts o segons. Algunes es donen unes hores més tard, sobretot si cal una digestió parcial
de l’al·lergen. Així i tot, la simptomatologia que es dona depèn de quin sigui el tipus d’al·lergen
o la via d’entrada.
La principal simptomatologia d’una reacció d’hipersensibilitat de tipus I és la següent llista:
L’anafilaxi és la resposta més severa i exagerada.
Anticossos IgE.
Durant la primera exposició a l’al·lergen, en l’individu es dona una resposta
immunitària majoritàriament de tipus Th2. Per tant, es tracta de limfòcits T helper
productors d’IL-4 que indueixen un canvi d’isotip cap a IgE. Aquesta col·laboració es basa en
una producció molt important i exagerada d’IgE en aquests individus atòpics. La
immunoglobulina E s’uneix als receptors Fc𝛆 dels mastòcits que l’expressen de forma basal.
La particularitat d’aquesta interacció és que la IgE pot persistir durant molts mesos unida als
receptors Fc de la membrana dels mastòcits. Aquesta duració porta a una sensibilització dels
últims.
La IgE té una vida mitjana molt curta a la circulació sanguínia. Presenta una gran afinitat pels
receptors Fcε dels mastòcits, basòfils activats i eosinòfils entre altres. Fruit de la
sensibilització, tots els receptors Fcε queden ocupats per una IgE que s’hi queda unida
durant dies o mesos.
Davant d’una segona exposició a l’al·lergen, aquest és reconegut per la IgE específica
unida als receptors Fcε dels mastòcits. Això indueix a una desgranulació massiva
d’aquestes cèl·lules. La segona resposta és ràpida, de segons o de minuts. Sovint es tracta
d’una resposta local, però té una alta potència en la localització concreta de la desgranulació.
Això és perquè hi ha molts mastòcits sensibilitzats.
Receptors Fcε.
En la resposta inicial, les cèl·lules més rellevants són els mastòcits perquè expressen
un receptor Fcε d’alta afinitat de forma permanent. El mateix tipus de receptor també és
expressat per part de basòfils i eosinòfils, però aquests només ho fan quan estan activats.
Això suposa que només participin en fases posteriors i no en a resposta inicial.
El receptor Fcε de mastòcits, basòfils i eosinòfils no és el mateix que el receptor Fcε
dels limfòcits B, les cèl·lules dendrítiques i els eosinòfils en repòs. Aquestes cèl·lules
presenten un receptor Fcε, certament, però té una afinitat molt més baixa. Estem parlant de
2 ordres de magnitud de diferència en l’afinitat.
Urticària / Angioedema
ÈczemaVòmits /
Diarrea / Poc creixement
Tos / Congestió
Dificultats respiratòries
Hipotensió Anafilaxi

Immunologia 2020
CEBIO 229
El receptor de baixa afinitat rep el nom de FcεRII i consta d’una sola cadena amb un domini
soluble, un domini transmembranal i un domini citoplasmàtic. En canvi, el receptor d’alta
afinitat rep el nom de FcεRI i està format per una cadena α, una cadena β i dues cadenes
γ.
La cadena α consta de dos dominis Ig extracel·lulars, un domini transmembranal i un domini
intracel·lular. Aquesta cadena α és la que s’uneix al Fc de la IgE. La cadena β consta de
4 dominis transmembranals i dos dominis intracel·lulars. Les cadenes γ consten d’un domini
transmembranal i un domini citoplasmàtic cadascuna. Les dues cadenes γ es troben unides
per ponts disulfur en els seus extrems N-terminals extracel·lulars.
Les cadenes β i γ transdueixen els senyals. Això és perquè la quinasa Lynn fosforila els
motius ITAM de les cadenes β i γ. El resultat d’aquesta fosforilació indueix el reclutament de
la quinasa Syk, que s’uneix als ITAM de la cadena γ i inicia la cascada de senyalització.
Fruit d’aquestes característiques, és normal que la resposta immunitària provingui,
principalment, de mastòcits. Mastòcits que, a més, es troben al teixit associat a les mucoses
com les superfícies de barrera.
Reacció al·lèrgica.
La interacció que es dona entre el l’al·lergen i la IgE associada al seu receptor Fcε de
la membrana dels mastòcits té diverses conseqüències.
En la reacció al·lèrgica aguda, es dona una activació dels mastòcits i una exocitosi dels
grànuls. Això significa que els macròfags es desgranulen i aboquen a l’exterior histamina,
serotonina i proteases de forma immediata. Aquesta immediatesa s’entén com una qüestió
de segons. En el cas de la histamina, que té una activació per secreció exocrina, actua en:
• La contracció del múscul llis bronquial.
• La contracció del múscul llis intestinal.
• La relaxació de la musculatura vascular.
• L’augment de la permeabilitat vascular.
• L’estimulació de les terminacions nervioses.
Per tant, no és d’estranyar que en una reacció al·lèrgica aguda es doni una broncoconstricció
que causa dificultats respiratòries, un augment del peristaltisme, un augment del calibre dels
vasos sanguinis que derivi en una hipotensió, un edema o dolor.
En la reacció al·lèrgica crònica es té és un efecte dels eosinòfils. Això és perquè en
aquests casos es produeix el metabolisme accelerat de l’àcid araquidònic amb la producció
d’una resposta més tardana. Els eosinòfils atrets i activats per l’acció de les citocines IL-3, IL-
5 i GM-CSF entre d’altres es desgranularan. Recordem que aquestes citocines IL-3, IL-5 i
GM-SCF afavoreixen la diferenciació a partir dels precursors. La desgranulació farà
abocar a l’exterior la proteïna bàsica principal, mediadors d’inflamació i altres molècules que
contribuiran a la simptomatologia associada a les reaccions al·lèrgiques.
És en aquesta segona fase on els basòfils, que hauran arribat al lloc de la reacció al·lèrgica
amb la producció d’IL-4, mantindran la resposta Th2 i, per tant, amb la diferenciació de
limfòcits B a cèl·lules plasmàtiques productores d’IgE.
En les imatges de microscòpia òptica, es veu que en l’estat inicial hi ha una desgranulació
dels mastòcits, el que causa una congestió de la musculatura. També es veu que, en una
reacció més tardana, hi ha una concentració d’eosinòfils que contribuiran a mantenir, junt amb
els basòfils, la reacció d’hipersensibilitat.

Immunologia 2020
CEBIO 230
Quan es dona un entrecreuament entre dos o més receptors FcεR, es dona una reacció
immediata al cap d’uns segons, una reacció immediata al cap d’uns minuts i una reacció
tardana al cap d’unes hores.
La reacció immediata que es dona en uns segons és fruit de la producció de fosfolipasa
A2, d’adenil ciclasa i de fosfolipasa C; els quals provoquen la formació de microtúbuls.
Aquesta formació permet la posterior exocitosi de grànuls i l’alliberament d’histamina,
serotonina i proteases. La histamina, a part de tot el que hem comentat que provoca, també
activa la secreció exocrina del cos.
La reacció immediata que es dona al cap d’uns minuts és només fruit de la producció
de fosfolipasa A2, la qual produeix àcid araquidònic. Aquest àcid s’aprofita posteriorment
per produir eicosanoides, els quals deriven en leucotriens i prostaglandines. Aquests
compostos provoquen una broncoconstricció, una hipotensió i la producció de secrecions
mucoses per part de les cèl·lules epitelials.
Finalment, la resposta tardana, la que es dona al cap dunes hores, és fruit de l’adenil
ciclasa i la fosfolipasa C, les quals acaben generant la proteïna quinasa C. Aquesta activa
la MAPK i la transcripció gènica que porta associada. Això porta a la secreció de citocines i
quimiocines com la IL-4, la IL-5, la IL-6, la IL-13, la GM-SCF, la TNF-α i la CCL3. Les citocines
secretades amplifiquen la resposta, creant el que s’anomena un Allergy loop.
Conseqüències de la resposta.
Els efectes clínics de la reacció al·lèrgica depenen del teixit on es produeix la
desgranulació dels mastòcits. Aquests són abundants a la mucosa dels tractes respiratori,
gastrointestinal i a la pell; teixits on es donen la majoria de les reaccions al·lèrgiques. És per
això que la introducció subcutània en diverses dosis genera només una resposta local. És el
que es mesura en les proves d’al·lèrgies.
Una de les possibles conseqüències de la resposta al·lèrgica és el que s’anomena
“resposta anafilàctica generalitzada”. Per tal que es doni aquest fenomen, l’antigen entra
al torrent sanguini i es generalitza la reacció, que afecta a gairebé tot el cos. Dit d’una altra
manera, la reacció acaba sent sistèmica. Això provoca una pèrdua de líquid des dels vasos
sanguinis fins als teixits, el que causa inflamació i un xoc anafilàctic. Les reaccions que
acaben en xoc anafilàctic poden ser fatals en qüestió de minuts. Les al·lèrgies que
representen la majoria de casos que acaben en anafilaxi són les que van dirigides a les
picades d’abella i a la penicil·lina.
A la següent taula es poden veure diferents hipersensibilitats de tipus I, els seus al·lèrgens
causants i les respostes que es produeixen. Cal tenir en compte que la resposta a un al·lergen
sol estar altament relacionada amb la via d’entrada d’aquest.
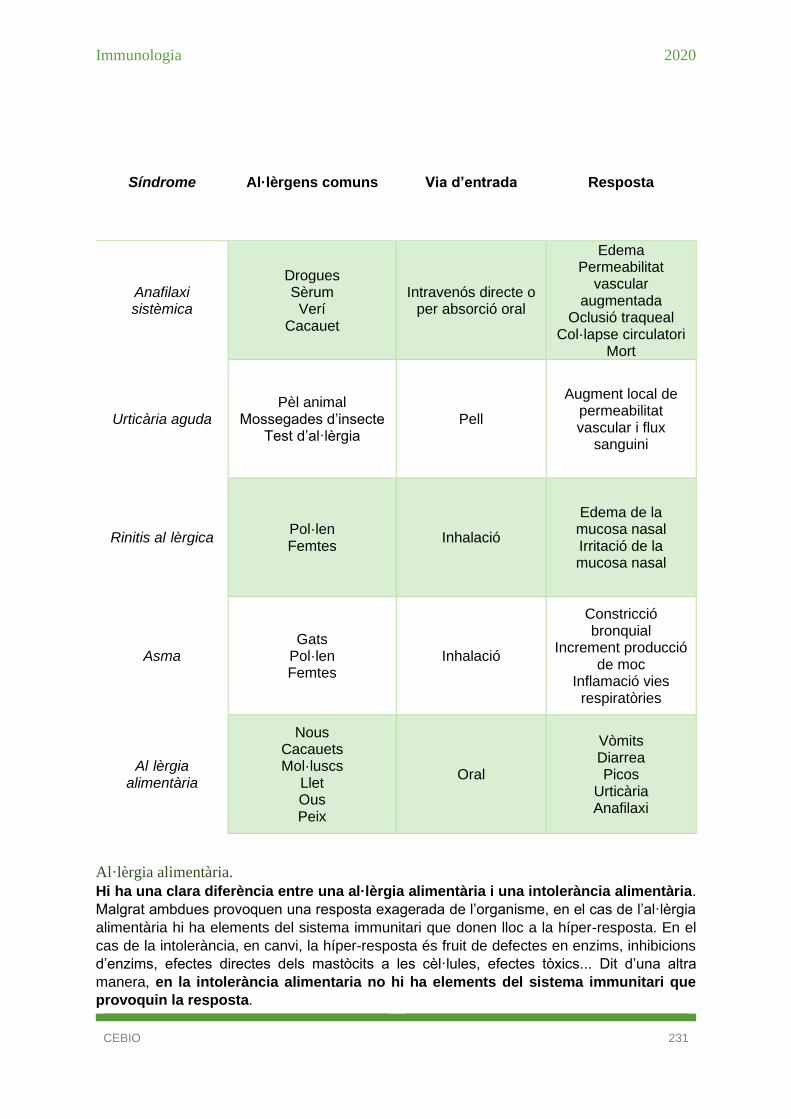
Immunologia 2020
CEBIO 231
Síndrome Al·lèrgens comuns Via d’entrada Resposta
Anafilaxi sistèmica
Drogues Sèrum Verí
Cacauet
Intravenós directe o per absorció oral
Edema Permeabilitat
vascular augmentada
Oclusió traqueal Col·lapse circulatori
Mort
Urticària aguda Pèl animal
Mossegades d’insecte Test d’al·lèrgia
Pell
Augment local de permeabilitat vascular i flux
sanguini
Rinitis al·lèrgica Pol·len Femtes
Inhalació
Edema de la mucosa nasal Irritació de la mucosa nasal
Asma Gats
Pol·len Femtes
Inhalació
Constricció bronquial
Increment producció de moc
Inflamació vies respiratòries
Al·lèrgia alimentària
Nous Cacauets Mol·luscs
Llet Ous Peix
Oral
Vòmits Diarrea Picos
Urticària Anafilaxi
Al·lèrgia alimentària.
Hi ha una clara diferència entre una al·lèrgia alimentària i una intolerància alimentària.
Malgrat ambdues provoquen una resposta exagerada de l’organisme, en el cas de l’al·lèrgia
alimentària hi ha elements del sistema immunitari que donen lloc a la híper-resposta. En el
cas de la intolerància, en canvi, la híper-resposta és fruit de defectes en enzims, inhibicions
d’enzims, efectes directes dels mastòcits a les cèl·lules, efectes tòxics... Dit d’una altra
manera, en la intolerància alimentaria no hi ha elements del sistema immunitari que
provoquin la resposta.

Immunologia 2020
CEBIO 232
Tractament de les Hipersensibilitats de tipus 1.
Per tractar les hipersensibilitats cal tenir clar l’objectiu del tractament. Si el causant
principal de la majoria de la simptomatologia és la histamina, es poden intentar eliminar els
símptomes. Els últims anys s’han usat molt els medicaments antihistamínics. Aquests són
molt efectius, ja que no creuen la barrera hematoencefàlica i això evita que tinguin efecte
sobre el sistema nerviós central. Si hi entrés, hi hauria un gran problema perquè la histamina
és un neurotransmissor important al sistema nerviós central per tenir l’estat d’alerta de
l’individu. Gràcies a no creuar la barrera, els medicaments es poden prendre de forma
habitual. A més, tenen una vida llarga que facilita la reducció de la simptomatologia.
El problema de tenir un bon tractament, però, és que ràpidament oblidem de curar la causa.
És el que passa amb les teràpies actuals, que tenen com a finalitat bloquejar els efectes de
la histamina i els símptomes associats enlloc d’intentar eliminar la recció d’hipersensibilitat.
Dessensibilització.
És per això mateix que hem exposat que s’estan començant a utilitzar diferents tractaments
que intenten evitar que hi hagi una reacció d’hipersensibilitat o tractaments que desviïn la
resposta del sistema immunitari. Per evitar una reacció d’hipersensibilitat s’està intentant
portar a terme una dessensibilització.
La dessensibilització consisteix en aconseguir que hi hagi una reducció de la resposta
IgE davant d’un al·lergen. Això es pot aconseguir per mitjà d’anticossos monoclonals
específics per IgE o amb injeccions repetides de quantitats molt petites de l’al·lergen al llarg
d’uns quants mesos. Amb aquestes injeccions s’aconsegueix que, enlloc d’IgE, es generin
IgG específiques contra l’al·lergen. Principalment, són IgG4. La gràcia d’aquest procediment
és que les IgG4 reaccionin amb l’antigen abans que aquest es pugui unir amb la IgE. El
resultat, doncs, és bloquejar la reacció de la IgE.
IgG4 és el subtipus d’IgG que menys activa el sistema del complement. És un detall
important, doncs una important producció d’IgG podria facilitar-ne l’activació. A més, aquesta
IgG4 també té una baixa afinitat pels receptors Fcγ, pel que té més dificultat per unir-se als
receptors Fc i produir la reacció de citotoxicitat cel·lular depenent d’anticossos.
Per tant, amb el tractament de dessensibilització s’intenta que no es produeixi IgE o
que es produeixin més IgG i que aquestes s’uneixin abans que les IgE a l’antigen.
També s’intenta que la resposta del sistema immunitari no sigui tant de Th2 i sigui més de
Treg, els quals permeten tenir una major tolerància a l’antigen.
Raons ambientals.
Les raons per les quals hi ha incidència de les hipersensibilitats de tipus I són molt
diverses. Una de les hipòtesis que hi ha és que la contaminació l’augmenti al transportar
al·lèrgens. Una altra de les hipòtesis, i que és molt plausible que sigui certa, és que la higiene
hi tingui alguna cosa a veure. La hipòtesi de la higiene afirma que, arran de l’actual estil de
vida, hi ha una menor exposició a agents infecciosos durant la infantesa. També afirma que
aquesta baixa exposició a agents infecciosos és la responsable de l’aparició de reaccions
d’hipersensibilitat en individus majors de 30 anys.
En el que es basa aquesta hipòtesi és que en l’entorn intrauterí es manté una resposta de
tipus Th2, coincident amb la que tenen els nounats. També té en consideració que les
exposicions a diferents agents infecciosos al llarg de la vida fan desenvolupar la resposta de
tipus Th1. En conseqüència, afirmen els qui la defensen, hi ha un equilibri entre la capacitat
de desenvolupar una resposta tipus Th1 o Th2 ja al voltant dels dos anys.

Immunologia 2020
CEBIO 233
Amb l’actual estil de vida, hi ha menys interaccions amb agents infecciosos durant els primers
anys de vida degut a una major higiene i a unes condicions d’habitabilitat que afavoreixen a
la menor exposició d’al·lèrgens. Tot això fa que el desenvolupament d’una capacitat de
resposta Th1 es retardi fins al punt que, als dos anys, sigui molt menor que la resposta Th2.
Això és el que fa possible que, en entrar en contacte amb un agent innocu, es desenvolupi
una resposta immune de tipus Th2 i es desencadeni una reacció al·lèrgica.
Aquesta hipòtesi permetria entendre, de forma genèrica, la causa de l’increment de les
reaccions d'hipersensibilitat de tipus I que està havent-hi a nivell mundial.
De fet, com que els nens i els adults passen més temps a l’interior, els al·lèrgens de la pols o
dels animals domèstics augmenten el risc de sensibilització al·lèrgica en proporció al grau
d’exposició. La majoria dels nens i adolescents amb asma estan sensibilitzats a al·lèrgens
d'interior.
Com que les cases d’avui dia tenen un bon aïllament, moquetes i calefacció central;
s’augmenta bastant la presència d’àcars de la pols. A l’exterior, en canvi, els al·lèrgens varien
segons l’estació i el clima.
Estil de vida.
Al desenvolupament de la hipersensibilitat de tipus I també hi contribueixen diferents elements
de l’estil de vida, sobretot relacionats amb l’alimentació. Per un costat, la disminució del
consum de fruites i hortalisses fresques provoca una disminució dels nivells d'antioxidants.
La presència de conservants en els aliments poden afectar la microbiota intestinal, donant
lloc a la sensibilització al·lèrgica en comptes de desenvolupar una tolerància als aliments. El
sistema immunitari està seriosament compromès en cas de mala nutrició. Paradoxalment, la
gran millora en la nutrició en els últims cinquanta anys podria haver donat lloc a què el sistema
immunitari d'alguns individus "sobre reaccioni" a substàncies en principi innòcues com els
al·lèrgens.
Per altra banda, certs àcids grassos poden canviar el sistema immune, augmentant-ne la
susceptibilitat al·lèrgica. Com que una higiene més continuada elimina àcids grassos de la
pell i els canvia, una higiene molt contínua podria augmentar la susceptibilitat al·lèrgica.
Hipersensibilitat de tipus II. Les reaccions d’hipersensibilitat de tipus II són degudes a una producció exagerada
d’immunoglobulines G i, en una menor quantitat, de IgM i proteïnes del sistema de
complement. Aquestes reaccions es donen en resposta a proteïnes de la matriu extracel·lular
o a proteïnes ancorades a la membrana i haptens. Les proteïnes de la matriu extracel·lular
són, en teoria, innòcues, mentre que les proteïnes ancorades a la membrana són endògenes.
El mecanisme que provoca les reaccions d’hipersensibilitat de tipus II són els
anticossos que, en unir-se a alguns receptors, els bloquegen o els activen. Un altre
mecanisme pel qual actua és la lisi mediada pel complement o la citotoxicitat cel·lular
depenent d’anticòs. És un exemple d’hipersensibilitat de tipus II l’anèmia hemolítica.
En les respostes d’hipersensibilitat de tipus II es produeixen IgG en quantitats
importants, a part que són específiques per antígens de les cèl·lules o per receptors de les
cèl·lules expressats a la membrana. Això, doncs, provoca:
• Una opsonització de les cèl·lules i la seva fagocitosi.
• Una citotoxicitat cel·lular depenent d’anticòs.
• Una activació o inhibició aberrant de les funcions cel·lulars.

Immunologia 2020
CEBIO 234
La unió dels anticossos als receptors de la membrana alteren el seu funcionament, ja
sigui activant la senyalització i les conseqüències funcionals que això té o bé impedint la
senyalització.
Anèmia hemolítica.
L’anèmia hemolítica és l’anèmia resultant de la lisi dels eritròcits. Es produeix en el cas
de que hi hagi una transfusió i no s’hagi tingut en compte ni el grup sanguini del donant ni el
del receptor.
Els grups sanguinis (A, B o 0) estan definits arrel de la presència o absència dels
antígens sanguinis A i B, que són carbohidrats. Segons la classificació, un individu amb
grup sanguini A genera anticossos contra epítops semblants a l’antigen B expressats per
microorganismes. En la situació contrària, un individu amb grup sanguini B produirà
anticossos contra epítops semblants a l’antigen A.
És per això que, quan hi ha una transfusió, els anticossos del receptor reconeixen
l’antigen sanguini expressat als eritròcits del donant i en provocaran la lisi a través de
l’activació del sistema del complement. El sistema es fixarà a la membrana dels eritròcits
i activarà la via clàssica, lisant els eritròcits i provocant l’anèmia.
Eritroblastosi Fetal.
L’eritroblastosi fetal també és un cas d’hipersensibilitat de tipus II. Es basa en
l’expressió de l’antigen RhD (Rhesus D antigen). Hi ha individus que no l’expressen, pel que
les mares RhD- que tenen un fetus RhD+ poden tenir problemes durant el part. En aquest, hi
pot haver un transvasament dels eritròcits RhD+ del fetus a la sang de la mare. Això
provocaria que ella generés una resposta a l’antigen RhD per mitjà d’anticossos i cèl·lules de
memòria contra l’antigen Rh.
Això, per si sol, no provocarà cap efecte negatiu contra el fetus o contra la mare. Tanmateix,
si es dona un segon embaràs en el qual el fetus també sigui RhD+, les immunoglobulines de
la mare especifiques per l’antigen RhD poden travessar la placenta al ser IgG. El resultat del
traspàs significa un atac als eritròcits del fetus i els lisaran. És el que es coneix com
eritroblastosi fetal.
La forma d’evitar que això passi és administrant a la mare, a les 28 setmanes d’embaràs i
durant les primeres 72h des del part, immunoglobulines específiques per l’antigen Rh. Gràcies
a això, s’impedirà l’acció dels anticossos al neutralitzar-ne la interacció.
Hipersensibilitat de Tipus III. Les reaccions d’hipersensibilitat de tipus III són causades per un antigen soluble que
causa l’acumulació d’immunocomplexes. Aquests són complexes formats per l’antigen i
la immunoglobulina, la qual sol ser IgG però també se n’han trobat amb IgM. Aquests
immunocomplexes provoquen l’activació del sistema del complement, la fagocitosi i el
reclutament de neutròfils. Cadascuna d’aquestes accions té unes conseqüències en el teixit
on es donen, pel que és una reacció local. Un exemple d’hipersensibilitat de tipus III és la
reacció arthus o la malaltia de la son.
Immunocomplexes.
El mecanisme o detonant de les reaccions d’hipersensibilitat de tipus III és la generació
d’anticossos contra un antigen soluble amb característiques especials. En general,
l’antigen és de mida petita i els immunocomplexes que es formen a partir d’ell, també. A més,
també s’han donat casos on l’antigen és molt abundant o persistent.

Immunologia 2020
CEBIO 235
En cas que l’antigen sigui molt abundant o persistent, es pot dificultar l’eliminació per
part del sistema immunitari fruit de la formació d’immunocomplexes que els neutròfils
i els macròfags no eliminen eficientment. La no eliminació els immunocomplexes pot tenir
dues causes:
• Que els immunocomplexes siguin massa petits.
• Que es produeixin massa immunocomplexes.
En cas que els immunocomplexes siguin massa petits, hi ha una mala opsonització per part
del sistema del complement que resulta en una mala unió i una difícil eliminació. La producció
d’un excés d’immunocomplexes pot ser fruit d’una quantitat abundant i persistent d’antigen,
el que genera un excés d’immunocomplexes que supera la capacitat d’eliminació del sistema
immunitari. L’excés d’immunocomplexes que supera la capacitat d’eliminació pot ser
degut a les altes dosis d’antigen o la producció contínua d’antigen.
Les altes dosis d’antigen poden ser resultat de diferents causes, com una infecció vírica
crònica o massiva com la de l’hepatitis B, una infecció bacteriana com la d’Endocartis
bacteriana, una inhalació continuada d’agents externs com passa amb les alveolitis
al·lèrgiques de grangers i forners o un tractament amb molècules antigèniques a altes dosis
com passa amb la malaltia del sèrum.
Els immunocomplexes que es produeixen circulen i es dipositen als teixits diana, que
poden ser les parets dels vasos sanguinis, els glomèruls renals o les articulacions. La
disposició dels immunocomplexes donen lloc a les reaccions d’hipersensibilitat.
Mecanisme patogènic.
Com que els immunocomplexes es dipositen, indueixen l’activació del sistema del
complement. Això deriva en la desgranulació dels mastòcits i, conseqüentment, en la lesió
tissular. Tanmateix, l’efecte patogènic de la reacció depèn del lloc d’entrada de l’antigen.
En cas que es tracti d’un antigen circulant, es produeixen lesions vasculars com les
vasculitis sistèmiques, lesions articulars com l’artritis i lesions renals com glomerulonefritis. Si
es tracta d’un antigen inhalat, les lesions que es produeixen són pulmonars com les alveolitis.
Finalment, quan l’antigen és subcutani, es produeixen lesions perivasculars a la zona
d’inoculació.
El procés pel qual en un individu immunitzat l’antigen interacciona localment amb IgG pre-
formades, es formen els immunocomplexes que activen el sistema del complement,
s’alliberen els mediadors d’inflamació que provoquen la desgranulació dels mastòcits i es
dona la inflamació local pot trigar entre 1 i 2 hores. Els mediadors alliberats són C5a, C3a
i C4a. C5a és el mediador que indueix a la desgranulació dels mastòcits. Amb la inflamació
local sorgida fruit dels mediadors, augmenta l’edema, l’arribada de noves proteïnes, de
fagòcits i pot provocar l’oclusió vascular.
Reacció d’arthus.
La reacció d’arthus és un exemple de reacció d’hipersensibilitat de tipus III. És una
reacció que s’observa quan han de fer una injecció intradèrmica o subcutània d’un antigen a
un individu immunitzat. És el cas d’individus que pateixen diabetis de tipus I, als quals se’ls
ha de fer una injecció subcutània d’insulina.
En aquests individus es produeixen anticossos específics contra la insulina, el que
donarà lloc a la formació d’immunocomplexes i d’intermediaris del sistema del
complement. Aquests intermediaris provoquen:

Immunologia 2020
CEBIO 236
• Una inducció de la desgranulació dels mastòcits.
• Una quimiotaxi sobre els neutròfils.
• Una estimulació de l’alliberació d’enzims lítics per part dels neutròfils que fagociten
immunocomplexes recoberts amb C3b.
En conseqüència, la simptomatologia de la reacció d’arthus consisteix en un eritema a nivell
local, un dipòsit d’immunocomplexes als capil·lars sanguinis i problemes sanguinis com
vasculitis, formació de microtrombes, oclusió vascular i necrosi tissular.
Malaltia del sèrum.
Aquest exemple d’hipersensibilitat de tipus III és ja poc freqüent, però està causada per la
injecció de grans dosis d’antigen proteic a la sang. Aquesta malaltia és possible de ser
observada si, per exemple, s’injecta l’antídot d’un verí d’una serp. En aquest cas, s’injecta
una gran quantitat de proteïnes de l’antídot, pel que es generen molts immunocomplexes i
aquests es dipositen a les parets dels vasos sanguinis de tot el cos però amb especial èmfasi
en els ronyons i les articulacions. En conseqüència, es donaria la reacció d’hipersensibilitat.
Als 7-10 dies de l’administració, l’individu pateix calfreds, febre, erupció cutània, urticària,
artritis i, sovint, glomerulonefritis.
Hipersensibilitat de tipus IV. La hipersensibilitat de tipus IV és l’única que és més tardana, d’entre 24 i 72 hores després
de l’exposició a l’al·lergen, motiu pel qual se l’anomena hipersensibilitat retardada. La
resposta hipersensible de tipus IV apareix independentment de la resposta humoral i
es dona tant en la primera com en la segona exposició a un al·lergen.
La independència a la resposta humoral es dona fruit de la dependència d’aquesta
hipersensibilitat dels limfòcits T específics per antígens solubles sols o associats a cèl·lules.
I és que en la resposta d’hipersensibilitat de tipus IV intervenen macròfags, eosinòfils i
limfòcits TCD4 o limfòcits TCD8. Dels limfòcits TCD4 en destaquen els limfòcits Th1.
Sensibilització.
En aquestes reaccions, la fase de sensibilització es produeix per la generació d’una
resposta TCD8 Th1 enfront de l’antigen, el que dona lloc a limfòcits TH1 de memòria. En
aquests casos, l’antigen també és innocu.
Fase Efectora.
Els limfòcits de memòria generats durant la sensibilització s’activen durant una segona
fase que s’anomena fase efectora. Quan estan activats, els limfòcits de memòria secreten:
• INF-γ, TNF-α i TNF-β.
• IL-2.
• Quimiocines.
• IL-3 i GM-SCF.
INF-γ, TNF-α i TNF-β causen inflamació i destrucció del teixit a nivell local. IL-2, en tant que
es produeix en gran quantitats, contribueix a la proliferació de limfòcits T i l’activació de
limfòcits TCD8 citotòxics. Les quimiocines recluten més macròfags i aquests, activats,
augmenten la inflamació. IL-3 i GM-SCF fan diferenciar més monòcits i macròfags a partir
dels precursors.
La combinació de totes aquestes secrecions provoca que es generi una resposta
exagerada a l’antigen que no hauria de generar cap mena de resposta. Si aquesta resposta
és continuada i es cronifica la inflamació, es pot donar lloc a la formació de granulomes.

Immunologia 2020
CEBIO 237
Exemples.
La hipersensibilitat retardada pura es troba en els efectes tardans de diversos verins
d’insectes i la resposta a la tuberculina entre d’altres. La tuberculina és una prova per
veure si un individu ha estat en contacte amb Mycobacterium tuberculosis. En aquests tipus
d’hipersensibilitat retardada, els antígens són pèptids de les proteïnes que els causen. El
temps que passa fins que es dona una reacció d’hipersensibilitat és d’entre 48 i 72 hores. Si
el teixit afectat és la pell, la infiltració és subepidèrmica.
Un cas particular és la malaltia celíaca, una enteropatia per culpa de la intolerància al gluten.
La celiaquia és una malaltia autoimmunitària on la hipersensibilitat de tipus IV sembla
tenir un paper central per explicar l’atròfia de vellositats malgrat haver-hi altres
mecanismes implicats.
La hipersensibilitat per contacte és un efecte haptènic de diverses substàncies que
modifiquen proteïnes pròpies de l’epidermis i generen aquest tipus de resposta
immunitària. És el tipus de resposta que es dona contra el làtex, el dinitrocorobenzè, el níquel
i el crom d’entre d’altres materials. El temps que passa fins que es dona la reacció
d’hipersensibilitat en aquests casos és d’entre 24 i 72 hores i la infiltració de l’al·lergen és
intraepidèrmica.
Taula resum. En aquesta taula podem veure resumides totes les característiques dels 4 tipus de reaccions
d’hipersensibilitat que hem comentat. Podem veure quin és l’element del sistema immunitari
causant de les reaccions, contra quin tipus d’antigen van, els mecanismes efectors del
sistema immunitari que donen lloc a aquesta resposta i alguns exemples.
Tipus I Tipus II
Reactiu immunitari
IgE IgG
Tipus d’antigen Antigen soluble Antigen de matriu o
membrana Receptor de superfície
Mecanisme efector
Activació mastòcits
Sistema del complement, Fagòcits, NK
Anticossos que alteren la senyalització
Exemple Al·lèrgia, Rinitis, Asma, anafilaxi
Al·lèrgies a drogues i a medicaments
Urticària crònica
Tipus III Tipus IV
Reactiu immunitari
IgG Th1 Th2 CTL
Tipus d’antigen Antigen soluble Antigen soluble
Antigen soluble Antigen cel·lular
Mecanisme efector
Sistema del complement,
Fagòcits
Activació Macròfags
Producció IgE, Activació
Eosinòfils, Activació
Mastocitosi
Citotoxicitat
Exemple Malaltia del
sèrum, Malaltia d’arthus
Dermatitis de contacte,
reacció de tuberculina
Asma crònic, al·lèrgia
crònica, rinitis Rebuig graft (?)

Immunologia 2020
CEBIO 238
Tema 27 – Autoimmunitat El segon grup de patologies lligades a un mal funcionament del sistema immunitari
correspon a les malalties autoimmunitàries. En una malaltia autoimmunitària es produeix
una resposta excessiva contra estructures i proteïnes pròpies. La resposta a les estructures
pròpies és fruit d’un reconeixement d’aquestes com a estranyes, el que dona lloc a una
resposta exagerada.
L’autoimmunitat es defineix com la capacitat de les cèl·lules del sistema immunitari de
reconèixer estructures pròpies. Aquesta propietat és inherent en el nostre sistema
immunitari, ja que la generació de limfòcits B i limfòcits T és per un mecanisme aleatori.
Aquesta generació aleatòria que es dona a la medul·la i al timus provoca la producció de
cèl·lules autoreactives. De forma que, els limfòcits han de tenir cert grau d’autoreactivitat.
Al timus es proporciona un sistema de selecció on els limfòcits T amb un TCR d’alta
afinitat per una estructura pròpia són eliminats. Només els limfòcits T amb una baixa
afinitat per estructures pròpies abandonen el timus per dirigir-se a la circulació i als teixits.
Els limfòcits B i T són capaços de reconèixer estructures pròpies però en unes
condicions anormals. A més, hi ha tota una sèrie de mecanismes de regulació que
mantenen l’estat de no reacció a estructures pròpies. Per si fos poc, els limfòcits necessiten
senyals de supervivència. Els senyals de supervivència venen donats per citocines, factors
de creixement i petites interaccions de baixa afinitat amb l’antigen. Mai en condicions normals
aquests senyals són suficients per activar l’autoimmunitat.
Autoimmunitat i malaltia autoimmunitària Hi ha 3 conceptes que hem de tenir clars: autoreactivitat, resposta autoimmunitària i malaltia
autoimmunitària.
L’autoreactivitat és la capacitat normal del sistema immunitari de reconèixer antígens
propis. L’autoreactivitat és un element necessari per mantenir la supervivència del pool de
limfòcits T i B.
La resposta autoimmunitària és la resposta dirigida per limfòcits T autoreactius i
limfòcits B productors d’autoanticossos. La resposta autoimmunitària és freqüent en la
població. De fet, el 25% de les dones de més de 50 anys tenen anticossos específics contra
hormones tiroidals. Sovint, la resposta autoimmunitària provoca lesions tissulars, però tan
sols una petita proporció de qui pateix respostes autoimmunitàries arriba a manifestar
simptomatologia clínica. La baixa presentació de simptomatologia clínica és fruit dels
mecanismes de regulació del sistema immunitari, que eviten que una resposta
autoimmunitària creixi en excés.
Una malaltia autoimmunitària és la simptomatologia clínica resultant directe d’una
resposta autoimmunitària. Entre un 4 i un 7% de la població mundial presenta malalties
autoimmunitàries. Les malalties autoimmunitàries són cròniques i amb exacerbacions, perquè
l’antigen és present de manera contínua. Les exacerbacions són períodes amb major i menor
intensitat resultants d’un millor o pitjor control per part del sistema immunitari.
De forma general, les malalties autoimmunitàries no afecten al sistema immunitari. Això
implica que tenir una malaltia autoimmunitària determina canvis en el sistema immunitari però
no altera la capacitat de resposta a patògens externs.

Immunologia 2020
CEBIO 239
Trencament de la tolerància. Les malalties autoimmunitàries necessiten d’un trencament de la tolerància per existir.
La tolerància del sistema immunitari pot ser central o perifèrica. La tolerància central es dona
en els òrgans limfoides primaris i la tolerància perifèrica es dona en els òrgans limfoides
secundaris.
Els mecanismes de la tolerància central que es donen al timus i a la medul·la òssia
determinen el repertori de limfòcits T i de limfòcits B autoreactius que surten a perifèria.
Els limfòcits autoreactius que han passat els mecanisme de la tolerància central tenen baixa
afinitat per l’antigen. Aquesta baixa afinitat determina que, en condicions normals, no hi hauria
d’haver una activació.
Així i tot, existeixen una sèrie de factors que poden suposar el trencament de la
tolerància perifèrica. Un trencament de la tolerància perifèrica equival al desenvolupament
d’una resposta autoimmunitària. Els factors que poden suposar el trencament de la tolerància
perifèrica són un mal funcionament de la tolerància central o la presència de factors externs.
Són factors externs que poden trencar la tolerància perifèrica els patògens, les toxines, els
fàrmacs, les hormones i altres elements forans.
Com a mínim, el 50% dels factors que trenquen la tolerància són de caràcter genètic.
Dels caràcters genètics que trenquen la tolerància en destaca HLA. Els Antígens Leucòcits
Humans (HLA) són molècules del MHC expressades en la superfície de les cèl·lules
humanes.
Si el 50% dels factors que trenquen la tolerància són genètics, un màxim del 50% des
factors que trenquen la tolerància tenen una naturalesa no genètica. Aquest fet ha estat
comprovat. En estudis sobre la diabetis en bessons univitel·lins, el fet que un dels dos tingui
diabetis de tipus 1 es correspon amb que l’altre tingui menys d’un 50% de possibilitats de no
desenvolupar-la. És convenient recordar que els bessons univitel·lins tenen la mateixa
dotació genètica.
També s’ha vist que aquells individus de poblacions amb baixa incidència de diabetis tipus 1
que van a viure en zones geogràfiques d’incidència molt alta desenvolupen la malaltia amb
una freqüència superior a la normal per la gent del seu país d’origen. Una zona geogràfica
amb una incidència molt alta de diabetis és Finlàndia. Així doncs, és correcte afirmar que hi
ha factors que contribueixen al trencament de la tolerància i al desenvolupament de les
malalties autoimmunitàries.
Molts factors fan prevaldre la immunitat sobre la regulació, afavorint el
desenvolupament de malalties autoimmunitàries.
El trencament de la tolerància ve determinat en un 50% per factors genètics i en un 50%
per factors ambientals. Els factors genètics inclouen el gen HLA entre d’altres. Els factors
ambientals inclouen les infeccions per enterovirus, la lactància, la dieta, la microbiota, la
higiene i la vitamina D entre d’altres.
Factors genètics.
Dels factors genètics que provoquen el trencament de la tolerància en destaquen amb
gran pes les Molècules del Complex principal d’Histocompatibilitat (MHC). Cal tenir en
compte les MHC perquè l’expressió de determinats al·lels HLA confereixen una susceptibilitat
molt elevada a patir malalties autoimmunitàries.
El cas més paradigmàtic és el de l’HLA-B27, una molècula MHC de classe I. La HLA-B27
confereix una alta susceptibilitat a patir espondilitis anquilosant. La susceptibilitat a

Immunologia 2020
CEBIO 240
l’espondilitis per part de la molècula HLA-B27 es deu a la seva cadena α. La cadena α de la
molècula HLA-B27 té tendència a plegar-se malament i expressar-se com homodímer a la
membrana. Aquesta modificació de l’estructura afavoreix la presentació d’autoantígens.
Un altre exemple on les molècules HLA són les responsables de la tendència a
desenvolupar patologies és el cas de la diabetis del tipus 1. Se sap que el MHC confereix
el 50% de susceptibilitat a patir la diabetis. La resta de la susceptibilitat depèn d’uns 20 o 30
gens que s’hi han associat. Així i tot, els gens associats no són responsables últims del
desenvolupament de la patologia.
Aquest fet indica que la majoria de malalties autoimmunitàries són poligèniques. Dit
d’una altra manera, hi ha múltiples loci gènics implicats en el desenvolupament de la diabetis
tipus I.
Possibles raons.
Determinats al·lels de HLA no poden presentar bé alguns pèptids de proteïnes pròpies.
La mala presentació de proteïnes pròpies es tradueix en una mala selecció tímica. Si hi ha
una mala selecció tímica, els limfòcits T que circulen i resideixen als teixits poden tenir una
afinitat massa alta per un antigen propi. Com que la tolerància perifèrica no fa un mecanisme
de selecció negativa, els limfòcits T amb afinitats massa altes per antígens propis es poden
activar. L’activació de limfòcits T amb afinitat contra antígens propis porta a la resposta contra
els propis teixits. Això explicaria perquè alguns al·lels de HLA poden provocar l’aparició
de malalties autoimmunitàries.
Hi ha altres gens que controlen la selecció negativa dels limfòcits T al timus i a la
medul·la. Una alteració en els gens reguladors de la selecció negativa porta a l’alliberament
de limfòcits T amb una afinitat massa alta pels antígens propis. Això explicaria perquè hi pot
haver una predisposició genètica no corresponent als gens HLA.
Altres gens.
Hi ha tota una sèrie de gens que contribueixen al trencament de la tolerància. El
trencament de la tolerància pot ser un mecanisme poligènic o monogènic. Quan el trencament
de la tolerància és per un mecanisme monogènic, els gens que deriven en el trencament
de la tolerància sovint estan relacionats amb la regulació del sistema immunitari. Alguns
dels gens afectats són AIRE, CTLA4, FOXP3, FAS i C1q.
El gen AIRE és un gen que permet l’expressió de proteïnes característiques d’altres
teixits a les cèl·lules epitelials medul·lars tímiques. L’expressió transitòria d’aquestes
proteïnes en el timus permet l’eliminació dels timòcits que tenen una alta afinitat per antígens
propis. Defectes en el gen AIRE són responsables de causar APECED. APECED és una
poliendocrinopatia autoimmune. En els individus amb APECED es donen afectacions a molts
òrgans diferents del cos.
CTLA4 que és un mecanisme de manteniment de la tolerància perifèrica per la seva
funció com a molècula coestimuladora inhibidora. Defectes en l’expressió de CTLA4
s’associen amb una pèrdua de tolerància perifèrica. S’han observat defectes en CTLA4 en
pacients de la malaltia de Graves i de diabetis de tipus 1.
FoxP3 és un factor de transcripció expressat pels limfòcits TCD4 reguladors i necessari
pel seu desenvolupament. La deficiència o alteració de FoxP3 i de la seva funció en cèl·lules
T reguladores s’ha associat a la patologia IPEX. Aquesta és una malaltia autoimmunitària
gairebé generalitzada.

Immunologia 2020
CEBIO 241
Fas o CD95 és un receptor mortal que s’expressa en la superfície dels limfòcits T i inicia
una cascada de senyals que condueixen a la mort apoptòtica de la cèl·lula. La via de
mort per fas s’activa en la unió de Fas al lligand de Fas. FasL s’expressa en els limfòcits T
activats. S’han vist mutacions o defectes en el gen Fas en malalties com ALPS. ALPS és una
síndrome limfoproliferativa autoimmunitària. La ALPS és fruit d’un mal funcionament del
mecanisme d’apoptosi de les cèl·lules autoreactives.
C1 és una proteïna del sistema del complement que inicia la via clàssica en unir-se a
les porcions Fc de les IgG o les IgM. S’han observat deficiències en gen C1q en malalties
com el Lupus Eritematós Sistèmic. Aquest tipus de lupus es caracteritza per una deficient
eliminació d’immunocomplexes i cèl·lules apoptòtiques.
De forma general, les síndromes causades per deficiències en els gens AIRE, CTLA4,
FOXP3, FAS i C1q causen autoimmunitat neonatal. Així i tot, és un clar exemple de com
un sol gen defectuós és responsable d’una patologia corresponent.
Hi ha gens amb efectes més subtils que afegeixen susceptibilitat a l’autoimmunitat. On
la majoria d’aquests tenen un paper important en la resposta immune. La patologia de la
diabetis pot tenir una major susceptibilitat en funció dels gens CTLA4, IL-2R, CLEC16A o
PTPN22.
La PTPN22 és una tirosina fosfatasa que intervé en la regulació de la via de
senyalització del TCR. S’ha vist que confereix susceptibilitat en les malalties del lupus, artritis
reumatoide i la diabetis de tipus 1.
El gen que codifica per la proteïna coestimuladora inhibidora PD-1 té susceptibilitat
amb el lupus eritematós sistèmic.
CTLA4 es troba associat tant a la diabetis tipus 1 com a malalties autoimmunitàries de
la tiroide.
Finalment, la presència de polimorfismes en el promotor de la R-TSH determina
l’eficàcia de la generació de la tolerància central. Defectes en aquest gen poden portar a
una mala selecció negativa al timus. Una mala selecció al timus per culpa de la R-TSH genera
un repertori de limfòcits T autoreactius contra la insulina.
Són malalties autoimmunitàries el lupus eritematós sistèmic, l’artritis reumatoide, les
malalties autoimmunitàries de la tiroide i la diabetis tipus 1.
Factors sistèmics o ambientals.
Les malalties autoimmunitàries necessiten d’un trencament de la tolerància per existir.
El trencament de la tolerància pot donar-se per factors genètics o per factors no genètics.
Dins dels factors no genètics que poden causar un trencament de la tolerància hi ha els factors
ambientals i els factors somàtics.
El trencament de la tolerància per factors somàtics és fruit d’un repertori clonotípic de
limfòcits T. Cada limfòcit T fa un reordenament genètic del TCR que li és específic. El
repertori de limfòcits T és determinant per la resposta immunitària. En conseqüència, la
susceptibilitat a generar una patologia autoimmunitària dependrà del reordenament genètic
del TCR.
En bessons monozigots, un màxim del 50% del repertori dels limfòcits T és idèntic. En
bessons dizigots, un màxim del 13%. En germans, només un 6%. Quant més repertori de
limfòcits T comparteixin, més possibilitats de contraure les mateixes malalties.

Immunologia 2020
CEBIO 242
En el trencament de la tolerància també hi intervenen una sèrie de factors ambientals.
Els principals factors ambientals que determinen el trencament de la tolerància són:
• L’alimentació
• Les infeccions
• Les toxines
• La latitud i la temperatura
• Els factors hormonals
• Els fàrmacs
Un exemple de l’efecte dels factors ambientals sobre la immunitat és el de l’alimentació per
mantenir la microbiota. La microbiota és important en el desenvolupament de sistema
immunitari de cada individu. La microbiota es pot veure alterada per, entre d’altres:
• Contacte amb toxines
• Temperatura
• Factors hormonals
• Alimentació
En conseqüència, es pot afirmar que els factors que alteren la microbiota modifiquen la
susceptibilitat a patir malalties autoimmunitàries.
La vitamina D s’obté gràcies a la llum del sol. La llum solar és capaç de transformar els
precursors en la forma activa de la vitamina D. El procés d’obtenció de la forma activa de
la vitamina D es veu alterat en funció de la temperatura i la latitud. La vitamina D influeix
al sistema immunitari als nivells de:
• Maduració de les cèl·lules dendrítiques
• Diferenciació dels macròfags
• Producció de citocines
• Disminució de la resposta Th1
• Augment de la resposta Th2
• Disminució de la producció d’immunoglobulines
Així doncs, la vitamina D incideix en molts elements del sistema immunitari. Un descontrol
de la vitamina D pot acabar afavorint el desenvolupament de malalties
autoimmunitàries.
Alliberació d’antígens segrestats.
L’alliberació d’antígens segrestats és un altre mecanisme que pot afavorir el
trencament de la tolerància perifèrica. Hi ha antígens que només es troben en un teixit i
que no es troben al timus. Aquests teixits són llocs immunològicament privilegiats. Són òrgans
immunològicament privilegiats:
• Cervell
• Ull
• Testis
• Úter (fetus)
No es fa una bona selecció tímica dels limfòcits T específics contra els llocs
immunològicament privilegiats. L’exposició dels autoantígens dels teixits privilegiats als
limfòcits T pot activar la resposta. L’exposició d’autoantígens privilegiats als limfòcits T no es
sol donar en condicions normals.

Immunologia 2020
CEBIO 243
Es pot donar un contacte dels limfòcits T amb els autoantígens privilegiats quan hi ha
un trauma del globus ocular, per exemple. El trauma provoca l’entrada de limfòcits als
limfonodes proximals. En els limfonodes proximals del globus ocular es permet el contacte
entre antígens de la còrnia i cèl·lules del sistema immunitari. El contacte entre limfòcits i
autoantígens privilegiats pot iniciar una resposta específica.
També es pot donar un contacte dels limfòcits T amb els autoantígens privilegiats quan
hi ha un trencament de la barrera hemato-encefàlica. El trencament de la barrera hemato-
encefàlica permet l’entrada dels limfòcits al sistema nerviós central. Al sistema nerviós central,
els limfòcits entren en contacte amb la proteïna bàsica de la mielina. La mielina forma una
beina que envolta els exons de les neurones del sistema nerviós central. La resposta a la
proteïna bàsica de la mielina desencadena en l’esclerosi múltiple.
Un contacte dels limfòcits T amb els autoantígens privilegiats es dona amb proteïnes
de l’esperma després de vasectomia o amb proteïnes del múscul cardíac després
d’infart de miocardi. En ambdós casos, proteïnes que estaven separades del sistema
immunitari entren en contacte amb els limfòcits a causa d’una lesió traumàtica.
Mimetisme molecular o crosreactivitat.
S’ha relacionat la pèrdua de la tolerància perifèrica amb el mecanisme del mimetisme
molecular, altrament anomenat crosreactivitat. El mimetisme molecular es dona quan els
antígens d’un organisme patogen són similars als antígens de proteïnes pròpies. Quan hi ha
un mimetisme molecular, els limfòcits T poden tenir un TCR que reconegui els epítops dels
patogen i que aquests epítops coincideixin amb els pèptids derivats de proteïnes pròpies. En
cas que es doni un mimetisme molecular, els limfòcits T activats en resposta al patogen poden
atacar proteïnes pròpies. L’atac dels limfòcits a les proteïnes pròpies acaba generant
autoimmunitat.
S’ha aconseguit identificar el mecanisme del mimetisme molecular en el virus del
papil·loma humà, el virus de la ràbia i en el receptor de la insulina. A la taula es pot veure
que hi ha una similitud prou important a nivell d’aminoàcid entre diferents virus i molècules
pròpies. El mimetisme molecular també es dona amb els citomegalovirus, que copien la
molècula HLA-DR humana.
Per tant, l’inici d’una resposta de limfòcits T contra epítops de proteïnes derivades de
patògens pot desencadenar una resposta autoimmunitària.
Expressió aberrant del MHC de classe II i de molècules coestimuladores.
Totes les cèl·lules nucleades del cos expressen molècules MHC de classe I. Les
molècules MHC de classe I poden activar limfòcits TCD8. Tanmateix, les la majoria de les
cèl·lules nucleades no tenen una expressió prou elevada de molècules coestimuladores. Per
aconseguir activar els limfòcits TCD8 en condicions no idònies cal que abans s’activin els
limfòcits TCD4.
Una expressió inapropiada de molècules del MHC de classe II i de molècules coestimuladores
en cèl·lules nucleades permetria el trencament de la tolerància. El trencament de la tolerància
seria per la presentació d’antígens propis de les cèl·lules nucleades als limfòcits TCD8.
Les cèl·lules β pancreàtiques produeixen insulina. En els individus amb diabetis tipus 1,
les cèl·lules β expressen nivells anormalment alts de MHC I i MHC II. Poden, per tant,
activar resposta T contra pèptids generats per aquestes cèl·lules.
La maquinaria de processament de les cèl·lules epitelials tímiques medul·lars no es la
mateixa que la de les cèl·lules β pancreàtiques. Les cèl·lules β pancreàtiques poden

Immunologia 2020
CEBIO 244
generar pèptids que siguin lleugerament diferent als generats en el timus. La presentació de
pèptids diferents implica que no hi hagi tolerància immunològica.
En condicions normals, les cèl·lules pancreàtiques no tenen capacitat per presentar
antigen als limfòcits T CD4 o CD8. En determinades condicions en què hi hagi una
expressió inapropiada de molècules MHC de classe II, podran convertir-se en cèl·lules
presentadores d’antigen. Els antígens presentats seran d’uns pèptids antigènics diferents i
pels quals la tolerància no estava ben resolta.
Una expressió inapropiada de molècules MHC-II i/o de molècules coestimuladores a cèl·lules
que no les expressen constitutivament permet que es generin i presentin pèptids diferents
pels quals no hi ha tolerància immunològica. Els pèptids pels quals no hi ha tolerància
immunològica s’anomenen neoantígens.
Classificació de les malalties autoimmunitàries. Les malalties autoimmunitàries es podem classificar en òrgan – específiques o en
sistèmiques. Algunes malalties però, són difícils d’ubicar en aquesta classificació.
Les malalties autoimmunitàries òrgan – específiques són aquelles on la resposta
autoreactiva està restringida a un òrgan determinat. Les malalties òrgan-específiques es
coneixen com malalties autoimmunitàries.
Les malalties autoimmunitàries sistèmiques són aquelles on la resposta autoreactiva
afecta a la majoria dels teixits de l’organisme . Es coneixen com malalties
autoimmunitàries sistèmiques. Entenem com a malalties autoimmunitàries sistèmiques
l’artritis reumatoide o el lupus eritematós sistèmic, entre d’altres. L’artritis reumatoide afecta
a les articulacions de tot el cos.
Les malalties autoimmunitàries també es diferencien en funció del mecanisme efector
i la clínica a la que donen lloc. Aquest fet depèn sobretot del tipus d’autoantigen diana.
L’autoantigen diana pot ser:
• Un receptor de membrana
• Un component ubic de membrana
• Proteïnes expressades a la membrana de les cèl·lules
• Factors de transcripció
• Reguladors de traducció
En els receptors de membrana, el mecanisme efector serà mediat per anticossos. Si és un
component ubic de la membrana, una acumulació de complexos serà el detonant de la
simptomatologia. Si l’autoantigen són proteïnes expressades a la membrana de les cèl·lules
(no receptors), factors de transcripció i reguladors de traducció, el mecanisme efector serà
degut a una resposta de limfòcits T.
Malaltia de Graves.
La malaltia de Graves és una malaltia òrgan-especifica que afecta la glàndula tiroide.
En la malaltia de Graves es generen anticossos específics pel receptor de la Hormona
Estimulant de la Tiroide (TSH-R). El TSH-R està expressat a les cèl·lules epitelials de la
glàndula tiroidal. Les cèl·lules de la glàndula tiroidal són secretores de les hormones tiroidals
que regulen el metabolisme de tot el cos.
La simptomatologia clínica relacionada amb la malaltia de Graves és molt variada i
afecta molts elements diferents. La malaltia de Graves s’associa amb:

Immunologia 2020
CEBIO 245
• Insomni
• Depressió
• Pèrdua de pes
• Taquicàrdia
• Suor
• Exoftàlmia (ulls molt sobresortits)
Totes les simptomatologies de la malaltia de Graves són degudes a un mal
funcionament de la TSH. El mal funcionament de la TSH es deu a la generació d’anticossos
específics contra el seu receptor. Els anticossos contra la TSH-R s’hi uneixen de forma
permanent. La unió dels anticossos al TSH-R provoca la contínua producció d’hormones
tiroidals.
Tot i el feedback negatiu cap a la pituïtària, no s’atura la producció d’hormones
tiroidals. La producció contínua d’hormones tiroidals provoca un funcionament excessiu de
la glàndula o hipertiroïdisme. L’hipertiroïdisme suposa l’abocament continu de les hormones
tiroidals al torrent sanguini.
La malaltia de Graves és una híper-resposta causada per la interacció d’una immunoglobulina
específica pel receptor de la TSH. Aquesta immunoglobulina altera el funcionament de TSH-
R. Es pot dir que la malaltia de Graves és deguda a un mecanisme semblant a la
hipersensibilitat de tipus II.
La malaltia de Graves està associada a determinats al·lels del HLA, especialment a
HLA-DR3. Sembla ser que HLA DQB1confereix protecció enfront la malaltia de Graves.
Graves és més freqüent en dones (ratio 7:1) i es dona quan tenen entre 30 i 40 anys. A partir
d’aquesta edat s’ha relacionat amb els canvis hormonals que tenen lloc.
Miastènia gravis.
La miastènia gravis és una malaltia òrgan – específica en la qual es generen anticossos
contra el receptor de l’acetilcolina. El receptor de l’acetilcolina s’expressa a les unions
neuromusculars de les cèl·lules del múscul esquelètic. Amb la miastènia Gravis es generen
anticossos que s’uneixen al receptor de l’acetilcolina i que bloquegen la transmissió de
l’impuls nerviós. Els malalts de miastènia gravis presenten una debilitat progressiva dels
músculs esquelètics. La miastènia gravis segueix un mecanisme semblant a la malaltia de
Graves, tot i que es dona en cèl·lules diferents i amb un resultat diferent. En la miastènia
gravis, al contrari que amb la malaltia de Graves, es bloqueja la interacció lligand – receptor.
Síndrome de Goodpasture.
La síndrome de Goodpasture és una malaltia òrgan – específica. En la síndrome es generen
anticossos específics per proteïnes expressades a la membrana. Es generen
autoanticossos d’elevada afinitat contra la fibra α3 del col·lagen tipus IV (α3(IV), domini NC1).
La fibra α3 s’expressa a la membrana basal de glomèruls renals i alvèols pulmonars.
La interacció dels anticossos amb les fibres α3 del col·lagen crea un possible lloc
d’interacció de la proteïna C1q del complement. La fibra de col·lagen és pròpia de
glomèruls renals i d’alvèols pulmonars. La fibra α3 s’expressa constitutivament. El patró
d’expressió de la fibra de col·lagen genera una activació continuada del sistema complement.
El sistema del complement actua sobre els glomèruls renals i els alvèols pulmonars.
L’actuació del sistema del complement provoca una lesió cel·lular directa. La lesió
cel·lular directe de la síndrome de goodpasture es deu a l’estructuració del MAC a la
membrana de les cèl·lules. El MAC provoca la mort directa.

Immunologia 2020
CEBIO 246
Alhora que hi ha una generació del MAC per part del sistema del complement, es genera una
inflamació. La inflamació dels de glomèruls renals i els alvèols pulmonars ve causada
per l’acumulació d’anafilotoxines. Les anafilotoxines indueixen la quimiotaxi de cèl·lules
fagocítiques. La quimiotaxi de cèl·lules fagocítiques és una acció continua. El resultat de la
inflamació als glomèruls renals és una lesió glomerular progressiva. El resultat d’una
inflamació als alvèols pulmonars és una hemorràgia pulmonar.
La generació d’anticossos contra les fibres del col·lagen provoca una activació
continua del sistema del complement. Hi ha una associació amb la molècula HLA-DR15 i
factors ambientals. Entre dels factors ambientals relacionats amb la generació d’anticossos
es troben:
• La infecció per virus respiratoris
• La infecció pel virus del dengue
• La inhalació de solvents hidrocarbonats
Lupus eritematós sistèmic.
El Lupus Eritematós Sistèmic (LES) consisteix en una malaltia autoimmunitària sistèmica. En
el lupus eritematós sistèmic es dona una inflamació sistèmica crònica. La malaltia
autoimmunitària del lupus provoca que es generin autoanticossos contra fosfolípids, antígens
nuclears i factors de coagulació. Són antígens nuclears:
• El DNA de doble cadena
• Les histones
• La combinació de DNA i histones
• Les ribonucleoproteïnes
Els anticossos que es generen en els casos de lupus eritematós sistèmic es dipositen
en forma d’immunocomplexes. El dipòsit d’anticossos suposa l’acumulació in situ
d’eritròcits, plaquetes i leucòcits. La majoria d’anticossos que es produeixen en un lupus
eritematós sistèmic estan dirigits contra fosfolípids i antígens nuclears. Sembla ser que la
producció d’anticossos es deu a una eritrosi massiva.
El dipòsit d’immunocomplexes que es dona en un lupus eritematós es tradueix en una
destrucció de les cèl·lules. La destrucció de cèl·lules en un lupus genera una
simptomatologia molt diversa. Entre els diversos símptomes es troben:
• Febre
• Debilitat general
• Artritis
• Erupcions cutànies
• Pleurèsia i disfunció renal
• Afectacions cardíaques i del sistema nerviós central
• Afectacions hematològiques com leucopènia, limfopènia o anèmia.
S’ha vist certa associació de patir Lupus eritematós sistèmic amb la presència HLA-
DR3 o DR2 I DQB1. Els LES és una malaltia més freqüent en dones (ratio 10:1).
Artritis reumatoide.
L’artritis reumatoide és una malaltia autoimmunitària sistèmica causada pel dipòsit
d’immunocomplexes. L’artritis es caracteritza per una inflamació crònica de la membrana
sinovial de les articulacions. La membrana sinovial de les articulacions és aquella que es troba

Immunologia 2020
CEBIO 247
entre el cartílag d’un os i el següent. La membrana sinovial manté el líquid sinovial de dos
ossos que formen l’articulació.
En l’artritis reumatoide es formen anticossos específics per la fracció Fc de les
immunoglobulines IgG. La fracció Fc de les IgG s’ha anomenat factor reumatoide. Els
anticossos que es formen en una artritis reumatoide són tant IgM com IgG. El conjunt
d’anticossos generats en l’artritis formen immunocomplexos que es dipositen a les
articulacions. La disposició dels anticossos provoca l’activació del sistema del complement i
la subsegüent inflamació in situ. La inflamació in situ atreu neutròfils i macròfags.
L’ambient inflamatori que es dona en l’artritis reumatoide permet la proliferació de
fibroblasts, macròfags i mastòcits que formen el pannus. El pannus és el conjunt de
cèl·lules que creixen a l’espai interarticular.
En conseqüència, es pot afirmar que l’artritis reumatoide provoca l’engruiximent excessiu
de la superfície de contacte entre dos ossos i el dolor a l’espai interarticular. En la
resposta de l’artritis hi col·laboren limfòcits Th1 i Th17. Els limfòcits Th17 ajuden a atraure
neutròfils i a mantenir la inflamació.
S’ha associat l’artritis reumatoide a l’expressió d’HLA-DR1 i HLA-DR4. L’artritis és més
freqüent en dones i entre els 20 i els 40 anys. L’artritis causa rigidesa matinal a les
articulacions i inflor del teixit tou al voltant d’un mínim de 3 articulacions.
Diabetis tipus I.
La diabetis tipus I és un dels exemples on la malaltia autoimmunitària és resultat de
l’acció directe de limfòcits T. En la diabetis, l’acció dels limfòcits T consisteix en la
destrucció de les cèl·lules β del pàncrees. Les cèl·lules β del pàncrees es troben situades als
illots de Langerhans. La destrucció de cèl·lules β la realitzen limfòcits TCD8 citotòxics
específics per proteïnes de les cèl·lules β.
Les cèl·lules β són productores d’insulina. Fruit de l’atac de les cèl·lules TCD8 ho ha una
destrucció de cèl·lules β i una producció insuficient d’insulina. La producció insuficient
d’insulina comporta un augment de glucosa a la sang. La diabetis de tipus I es relaciona amb:
• Hiperglucèmia
• Gana
• Augment de set
• Poliúria
• Glucosúria
S’ha associat la diabetis de tipus I a HLA-DR3 o DR4 i a HLA-DQ8.
L’inici de la resposta immunitària de la diabetis de tipus I és l’atac de les cèl·lules
productores d’insulina. Resultat de l’atac a les cèl·lules productores d’insulina es genera
una infiltració de limfòcits TCD8 i TCD4 als illots de Langerhans. La infiltració de les cèl·lules
TCD4 i TCD8 provoca la destrucció de les cèl·lules α i les cèl·lules 𝛿. Les cèl·lules α són
cèl·lules productores de glucagó. Les cèl·lules 𝛿 són cèl·lules productores de somatostatina
Diversificació d’epítops o Epitope Spreading.
La diabetis de tipus I és una resposta T dirigida inicialment contra la insulina. En individus que
pateixen la diabetis de tipus I hi ha anticossos contra aquestes proteïnes i també contra:
• Transportador de zinc (ZNT8)
• IA2

Immunologia 2020
CEBIO 248
• IGPR o descarboxil·lasa de l’àcid glutàmic
• Cromogranina A
Tot i que la resposta inicial va dirigida contra la insulina, després es produeix el
fenomen de l’epitope spreading. El fenomen de l’epitope spreading consisteix en una
diversificació d’epítops. La diversificació d’epítops provoca la diversificació d’autoantigens
diana de la resposta autoimmunitària. La diversificació d’autoantígens explicaria la destrucció
de totes les cèl·lules de l’illot de Langerhans.
S’ha suggerit que en pacients de diabetis tipus I es produeix mimetisme molecular amb
antígens vírics. Els virus que podrien tenir el mimetisme molecular són:
• Virus parotiditis
• Rubeòla
• CMV
• Coxsakie B4 (P2-C i GAD)
En individus amb diabetis de tipus I s’ha pogut detectar una infecció prèvia per
coxsakie virus B4. És difícil establir la relació perquè els virus infecten unes cèl·lules i en
quant s’acaba la infecció desapareixen els signes que aquesta hi ha sigut.
En individus amb diabetis tipus I, la resposta immunitària al virus comença molt abans
que es detecti simptomatologia clínica. Aquest fet és una de les coses que dificulta la seva
detecció.
Si el problema inicial que ha causat el desenvolupament de la resposta autoimmunitària
ha estat la infecció per un virus, els signes que hi ha hagut aquesta infecció vírica són
difícilment detectables. És difícil establir la causalitat de la malaltia autoimmunitària.
És una resposta humoral secundària però hi ha anticossos anti-GAD.
Esclerosi múltiple.
L’esclerosi múltiple és deguda a limfòcits T. En condicions fisiològiques, el sistema
immunitari fa la vigilància del sistema nerviós central. La vigilància del sistema nerviós central
es fa per una monitorització des del plexe coroïdeu i el líquid cefalorraquidi. Des del plexe
coroïdeu venen proteïnes del sistema immunitari al líquid cefalorraquidi, però no entren al
parènquima del sistema nerviós central, ja que es manté intacta la barrera hemato-encefàlica
(BBB).
Hi ha cops que es perd la integritat de la barrera hemato-encefàlica. La pèrdua de la barrera
suposa l’entrada de limfòcits TCD8 i limfòcits TCD4 efectors al sistema nerviós central.
Els limfòcits que entren al sistema nerviós intervenen en la destrucció de les beines de mielina
de les neurones. La destrucció es dona per l’especificitat dels limfòcits T contra la proteïna
bàsica de la mielina o altres proteïnes que formen beina de mielina. La beina de mielina
recobreix els axons neuronals.
L’entrada al sistema nerviós central suposa l’activació de limfòcits T autoreactius. Els
limfòcits Th1 i Th17 trenquen la transmissió de l’impuls neuronal entre neurones. La
destrucció de la beina de mielina de les neurones provoca l’acumulació de restes cel·lulars i
citocines pro-inflamatòries. Els components restants de la destrucció de la beina de mielina
promouen la inflamació local.

Immunologia 2020
CEBIO 249
La inflamació local provoca l’activació dels macròfags que es troben en el sistema nerviós
central i de la micròglia. L’activació de macròfags i de micròglia genera lesions en la
matèria blanca. Les lesions determinen l’agudització de la patologia.
La simptomatologia associada a l’esclerosi múltiple és progressiva. L’esclerosi múltiple
comença amb un adormiment lleu de les extremitats però pot arribar a una paràlisi i
pèrdua de visió. L’esclerosi múltiple és la causa més freqüent de discapacitat neurològica
en els països occidentals. S’ha detectat una associació als haplotips d’HLA de classe II
DRw15 i DRw6.
Celiaquia.
La celiaquia és una malaltia de base immunològica. En la celiaquia es genera una resposta
dels limfòcits T contra proteïnes del gluten. El gluten és una combinació de gliadina i
glutenina. Es pot trobar gluten en blat i altres cereals. La resposta al gluten provoca la
destrucció de les cèl·lules epitelials intestinals.
La destrucció de les cèl·lules epitelials no té res a veure amb l’autoimmunitat. Els
limfòcits T específics per pèptids derivats de proteïnes que formen el gluten no reconeixen
els pèptids presentats per cèl·lules epitelials intestinals. Succeeix és que les proteïnes del
gluten són molt riques en glutamina i prolina.
La prolina fa que les proteïnes del gluten siguin molt resistents a la degradació. Una
degradació proteica difícil ajuda a la generació de pèptids presentats per molècules MHC. La
glutamina és transformada per un enzim tissular a àcid glutàmic, un aminoàcid amb càrrega
neta negativa. L’enzim tissular és la transglutaminasa. Els pèptids derivats de la transformació
de les proteïnes del gluten amb l’acció de la transglutaminasa disposen de motius d’ancoratge
idonis per la càrrega de molècules HLA-DQ2 o HLA-DQ8.
L’actuació de la transglutaminasa genera una resposta específica als pèptids derivats
del gluten. Per transformar els pèptids del gluten, la transglutaminasa hi ha de formar un
complex. El complex gluten-transglutaminasa és reconegut per anticossos específics. El fet
de reconèixer el complex fa que s’anomeni malaltia autoimmunitària.
La resposta inicial és contra el gluten però es produeix una resposta contra l’enzim
tissular transglutaminasa. La resposta a l’enzim provoca la destrucció de les cèl·lules
epitelials intestinals l’expressen.

Immunologia 2020
CEBIO 250
Tema 28 – Immunodeficiències Les immunodeficiències són el conjunt de patologies produïdes per la manca o
funcionament deficient d’un o més components del sistema immunitari. Es comença a
sospitar que un individu té una immunodeficiència quan presenta signes clínics d’infeccions
cròniques recurrents. Aquestes infeccions són diferents en funció del component del sistema
immunitari que sigui deficient o que funcioni malament.
També es sospita d’una immunodeficiència quan es dona una elevada incidència de
càncer, el que sobretot passa en immunodeficiències dels limfòcits T causades per virus
oncogènics com l’Epstein Barr. Aquests virus substitueixen el parènquima dels teixits i fan
que hi hagi una deficiència de la resposta cel·lular.
Classificació de les immunodeficiències. Les immunodeficiències es classifiquen segons el seu origen. Les immunodeficiències
primàries són aquelles que tenen una base genètica. Arran d’això, solen aparèixer aviat
en el desenvolupament, com en la lactància o la infància. Les immunodeficiències primàries
són molt diferents en funció de quin component del sistema immunitari es vegi afectat per
aquestes alteracions genètiques.
Les immunodeficiències secundàries o adquirides són les més freqüents però les
menys severes. Apareixen a raó d’una malnutrició, càncers o els seus tractaments, fàrmacs
immunosupressors o per infeccions de cèl·lules del sistema immunitari. Aquest últim cas és
el que es dona amb el VIH.
Les immunodeficiències poden afectar elements o components tant de la immunitat
innata, com el sistema del complement, la fagocitosi o les cèl·lules NK; com de la immunitat
adaptativa. Això també determina, de forma molt important, l’afectació de les
immunodeficiències.
Immunodeficiències Primàries. Les immunodeficiències primàries, malgrat ser les menys habituals, són les més
severes. Tenen una afectació molt important pels individus que les pateixen. Les
immunodeficiències primàries són degudes a mutacions heretades, d’un o més gens, que
intervenen en el desenvolupament de les cèl·lules del sistema immunitari i/o en la seva funció
o en la regulació de la resposta.
Les immunodeficiències primàries donen lloc a alteracions quantitatives o qualitatives
d’un o més components del sistema immunitari. Aquests components, a més, poden ser
de la immunitat innata o de la immunitat adaptativa. S’han descrit unes 250
immunodeficiències primàries però es coneix el gen defectuós, deficient o alterat en menys
de la meitat d’aquestes immunodeficiències primàries. Això dificulta el seu tractament.
Les immunodeficiències, a diferència de l’autoimmunitat, tenen major incidència en
homes perquè molts dels defectes genètics que les causen es troben en gens autosòmics
recessius lligats al cromosoma X. En conseqüència, es manifesten de manera més clara en
els homes.
Les immunodeficiències primàries apareixen aviat i es diagnostiquen durant la
lactància i la infància. En aquests casos, la manifestació clínica principal de les
immunodeficiències primàries és una major incidència de càncer en cas que la
immunodeficiències afectin a la proliferació dels limfòcits T o B, una major presència de
malalties autoimmunitàries i una recurrència en la infecció.

Immunologia 2020
CEBIO 251
Símptomes de les Immunodeficiències Primàries.
Les infeccions recurrents són greus, repetides i sostingudes causades per
microorganismes que, per individus immunocompetents, no són patogènics. Per tant,
es solen detectar infeccions per bacteris encapsulats biogènics que provoquen la generació
de pus. Es sol associar l’aparició d’aquestes infeccions recurrents per bacteris encapsulats
biogènics a defectes en la producció d’anticossos, el sistema del complement i la fagocitosi.
Si es produeix una infecció cutània persistent per fongs o hi ha infeccions víriques
recurrents, és bastant plausible una immunodeficiència que afecti la resposta cel·lular,
és a dir, els limfòcits T.
En funció del tipus cel·lular, procés o funció afectat per la immunodeficiència primària es
distingeixen diferents tipus. Les immunodeficiències primàries humorals comprenen el
50% de les immunodeficiències primàries i van associades a infeccions bacterianes
cròniques.
Un 10% de les immunodeficiències són immunodeficiències en la resposta T o la
resposta cel·lular. Sovint acaben donant infeccions per organismes sapròfits com càndida o
per virus.
Un 20% de les immunodeficiències són el resultat de la combinació d’una
immunodeficiència en la resposta humoral i una deficiència en la resposta cel·lular. En
aquest cas, es parla de les immunodeficiències severes combinades. Són el que s’anomenen
“nens bombolla”: persones la supervivència de les quals està condicionada a un aïllament
perquè tenen una afectació total de la resposta adaptativa humoral i cel·lular.
Un 2% de les immunodeficiències són degudes a deficiències en elements del sistema
del complement. Es caracteritzen per infeccions recurrents per bacteris encapsulats que
acaben sent incontrolades i donen lloc a xocs sèptics.
El 18% de les immunodeficiències són degudes a problemes amb la fagocitosi i donen
lloc a infeccions bacterianes com les que causen els estafilococs.
Severitat de les Immunodeficiències Primàries.
La severitat de les immunodeficiències primàries depèn de la funció de l’element del
sistema immunitari que estigui afectat. No té les mateixes conseqüències que hi hagi una
deficiència en els mecanismes de regulació del sistema immunitari (que dona sovint malalties
autoimmunitàries), que la deficiència sigui de la resposta cel·lular o de la resposta humoral, o
de la fagocitosi o d’ambdues. En aquest últim cas estaríem parlant que la immunodeficiència
afecta a més d’un llinatge cel·lular.
La severitat també es diferent en funció del punt del desenvolupament o funció de les cèl·lules
del sistema immunitari en el qual actuï el gen alterat. Quant més primerenc sigui el paper
del gen alterat en el desenvolupament de les cèl·lules del sistema immunitari, més gran
serà l’afectació. No és el mateix que hi hagi una alteració a la JAK quinasa 3 o la cadena γ
comuna del receptor de citocines com la IL2, IL4 i la IL7; que una afectació o alteració en el
CD40 o el CD40L. Una alteració en el gen γ C dels receptors de les citocines afectarà tot el
llinatge B i no es donaran limfòcits B madurs. Una alteració o afectació de CD40 o CD40L
afectarà una part de la funció dels limfòcits B.
Quan abans en el desenvolupament de les cèl·lules del sistema immunitari tingui funció
el gen alterat, més gran serà la severitat de la immunodeficiència.

Immunologia 2020
CEBIO 252
Classificació de les Immunodeficiències Primàries.
Les immunodeficiències primàries també es poden classificar en funció de l’element
del sistema immunitari que està alterat. Les immunodeficiències s’agrupen en:
• Aquelles que afecten als fagòcits.
• Aquelles que afecten al sistema del complement.
• Les que afecten l’activació o funció dels limfòcits T
• Les que afecten la producció d’anticossos.
• Les immunodeficiències severes combinades.
Aquestes últimes, les immunodeficiències severes combinades, són aquelles que
afecten més d’un llinatge cel·lular i, conseqüentment, tenen una afectació molt més gran
en l’individu que les pateix.
Immunodeficiències Primàries per un dèficit en la funció fagocítica.
Les immunodeficiències que afecten la funció fagocítica es manifestem amb infeccions
recurrents de la pell i les vies respiratòries per bacteris. Destaquen les infeccions per
estafilococ aureus i pels fongs aspergillus i càndida.
Aquest tipus d’immunodeficiència pot resultar en una afectació intrínseca de la
fagocitosi i es diria que la causa és una deficiència en els mecanismes bactericides com la
generació d’espècies reactives d’oxigen i nitrogen. Això passa en individus amb mutacions
en les subunitats de la NADPH oxidasa. Per tant, tenen afectacions en la generació de l’esclat
respiratori, el mecanisme microbicida que tenen les cèl·lules fagocítiques.
També afecten la fagocitosi les mutacions en l’enzim de la glucosa-6-fosfat
deshidrogenasa. Això altera la producció d’energia a les cèl·lules fagocítiques. També hi ha
afectacions dels mecanismes d’estructuració dels microtúbuls. Passa quan hi ha defectes en
la polimerització de lectina, que impedeix el procés d’englobament o fagocitosi. És la causa
de la síndrome del leucòcit mandrós.
La síndrome de Chediak – Higashi és una on hi ha defectes en el gen list, impedint la
fusió entre el fagosoma i el lisosoma. Això impedeix que s’organitzi i s’estructuri l’esclat
respiratori. No s’aporten al fagosoma tots els components dels grànuls que permeten la
generació de l’esclat respiratori, pel que es veu afectat el procés de fagocitosi i destrucció
dels microorganismes.
Es pot afectar la fagocitosi amb una alteració o deficiència en els mecanismes
d’arribada de les cèl·lules fagocítiques al punt on es produeix l’entrada dels patògens.
Això afecta molt als neutròfils, que són els que arriben primer. S’han descrit problemes en la
funció fagocítica dels neutròfils com a resultat de l’expressió de molècules d’adhesió o els
seus lligands, el que afecta el moviment d’aquests leucòcits. S’han descrit deficiències en
CD18, que és la cadena β2 de les integrines com LFA-1. Aquest és el mecanisme alterat que
s’ha vist en la deficiència d’adhesió de leucòcits de tipus I. També s’ha descrit la deficiència
en els lligands de les selectines A i P. Són el cialil-X-louis carbohidrats. Aquesta deficiència
és la que es detecta en individus amb deficiència d’adhesió leucocitària de tipus II.
Immunodeficiències Primàries per un dèficit en els factors del sistema del complement.
S’han descrit moltes deficiències en diferents elements del sistema del complement.
En funció de l’element alterat i el tipus d’alteració hi ha unes conseqüències també diferents.
Defectes en els components C1, C2 i C4 causen malalties arran de l’acumulació
d’immunocomplexos, com passa amb el lupus.

Immunologia 2020
CEBIO 253
Alteracions en la lectina d’unió a la manosa provoquen alteracions o defectes en
l’activació de la via de les lectines. Això es sol traduir en infeccions recurrents per part de
bacteris extracel·lulars. Es detecta més en la infància i després passen desapercebudes.
Alteracions en el factor D de la via alternativa causen infeccions recurrents piògenes.
Són les que causen pus. Aquestes infeccions recurrents quan hi ha alteracions en el factor D
solen ser per part de bacteris com Neissèria. Les alteracions o deficiències en els
components del MAC també es tradueixen en infeccions recurrents per Neissèria.
Tenir alteracions de l’element C3 del complement suposa tenir infeccions recurrents
piògenes com per Neissèria i una acumulació d’immunocomplexes.
Immunodeficiències Primàries per altres dèficits.
S’han descrit dèficits en la senyalització d’infeccions per la via dels TLR. Donen lloc a
infeccions piògenes recurrents i a infeccions víriques greus. S’han descrit mutacions en
l’adaptador MyD88, molècula senyalitzadora important en la senyalització mitjançant molts
TLR de membrana plasmàtica. També s’han vist mutacions en IRAK4, l’element que s’associa
a MyD88. La mutació de MyD88 o IRAK4 pot provocar una deficiència total, parcial o
funcional però s’associa a infeccions bacterianes greus. Sobretot per pneumococs.
S’han descrit mutacions a TLR3 i a TBK1, element important en la senyalització pel TLR3.
Les mutacions en TLR3 o TBK1 s’associen a una menor producció dels interferons de
tipus I, pel que aquests tipus de defectes causen dificultats en la resposta a virus i les
infeccions per aquests organismes són recurrents.
La mutació NEMO és una mutació en l’inhibidor de l’activació de NFκB. La mutació
manté NFκB al nucli, impedint la seva translocació. S’ha vist que això dona lloc a infeccions
recurrents per micobacterium tuberculosis.
S’han vist dèficits en la via de senyalització de l’interferó γ i de la interleuquina 12.
Sobretot, les mutacions afecten la cadena β del receptor de la IL-12 i també de l’interferó γ.
Donen lloc a infeccions recurrents per patògens intracel·lulars, principalment per micobacteri.
La IL-12 és rellevant en la diferenciació de limfòcits T helper 1, que són importants en la
resposta a patògens intracel·lulars.
En funció de quin sigui l’element del sistema immunitari deficient, hi haurà una o altra
patologia.
Immunodeficiències Primàries Humorals per dèficit d’immunoglobulines.
Les immunodeficiències primàries que afecten la resposta humoral es detecten per
dèficits en la producció d’immunoglobulines. Hi ha tota una sèrie d’immunodeficiències
relacionades amb la resposta humoral. La més freqüent és la immunodeficiència que provoca
un dèficit selectiu d’immunoglobulina A. És molt freqüent però és molt lleu, no és gaire severa.
S’associa a mutacions en el gen TACI: el receptor de les citocines BAFF i APRIL. BAFF està
feta per les cèl·lules dendrítiques i col·labora en la diferenciació dels limfòcits B a cèl·lules
plasmàtiques secretores de IgA.
En els individus amb immunodeficiència de IgA, malgrat haver-hi limfòcits B que han fet
el canvi d’isotip i l’expressen a la membrana, no es produeix la diferenciació a cèl·lules
plasmàtiques productores de IgA. Per tant, la immunodeficiència s’associa a infeccions
respiratòries i intestinals. Són aquelles que afecten a les mucoses, doncs és on IgA té la
funció de neutralització.

Immunologia 2020
CEBIO 254
S’han descrit dèficits selectius de subclasses de IgG. La més habitual és la
immunodeficiència de IgG2. Es sol associar a una deficiència de IgA. També s’han descrit,
però, dèficits de IgG1 i de IgG3. Es detecten per infeccions bacterianes recurrents malgrat no
tenen una gran afectació clínica. A més, tant en el cas del dèficit selectiu de IgA, de IgG com
en el cas de la immunodeficiència variable comuna es poden tractar amb administracions
d’immunoglobulines de manera periòdica.
Quan es produeix una immunodeficiència de més d’un isotip d’immunoglobulines,
sempre i quan un d’ells sigui IgG, es parla d’immunodeficiència variable comuna.
Aquests individus tenen limfòcits B madurs però no tenen cèl·lules plasmàtiques. S’ha
associat a defectes intrínsecs dels limfòcits B, com mutacions a CD19 i alteracions o
deficiències entre la col·laboració entre limfòcits B i T. Recordar que aquestes interaccions
són molt importants perquè es produeixi la fase final de la diferenciació dels limfòcits B a
cèl·lules plasmàtiques, així com el canvi d’isotip. S’ha vist que hi ha mutacions de ICOS als
limfòcits T.
Les immunodeficiències variables comunes es detecten perquè hi ha nivells baixos
d’immunoglobulines al sèrum, pel que els individus són incapaços de fer una resposta
humoral potent contra antígens específics. La immunodeficiència variable comuna es
diagnostica més tard que les immunodeficiències d’IgA o IgG. Es detecten perquè es
produeixen infeccions bacterianes recurrents, sobretot als aparells respiratori i digestiu.
També s'associen a malalties autoimmunitàries i quan hi ha presència de limfomes. Això és
perquè amb la progressió tumoral dels limfòcits B s’afecta a la producció d’immunoglobulines.
També és una immunodeficiència primària humoral la Agammaglobulinèmia lligada al
cromosoma X, altrament dita Síndrome de Bruton. Agammaglobulinèmia significa “absència
d’immunoglobulines.” Això és perquè en aquests individus es produeix una alteració de
funcionament del gen BTK. Aquest gen és un element important en la transmissió del senyal
del BCR. Amb aquest gen mutat, no hi ha una correcta senyalització mitjançant el BCR i els
limfòcits B moren per apoptosi durant els primers estadis de maduració. De fet, el seu
desenvolupament s’atura a l’estadi pre-B, aquell on es fa el primer “test” del BCR. Durant el
mateix test, el BCR està format per cadena pesada i s’associa a una cadena lleugera
substitutòria. A partir d’aquest punt, s’atura el desenvolupament del limfòcit B i, segons quina
sigui l’alteració en el gen BTK, ja no es produeix cap limfòcit B o es produeixen nivells molt
baixos de limfòcits B madurs. La conseqüència és que hi hagi una absència o uns nivells molt
baixos d’immunoglobulines al torrent sanguini i una deficiència en la generació dels centres
germinals i una conseqüent absència de cèl·lules plasmàtiques. En aquest últim cas hi podria
arribar a haver una funció normal dels limfòcits B.
Aquestes immunodeficiències es detecten en nens que, a partir dels pocs mesos de vida, fan
infeccions piògenes de repetició. Sovint, aquests nens són molt sensibles al es infeccions
per enterovirus. Igual com les altres immunodeficiències, responen bé a un subministrament
periòdic d’immunoglobulines.
La síndrome d’hiper-IgM es tradueix en valors normals de limfòcits B, però el problema és
que no es fa el canvi d’isotip. Aquesta característica es deu a mutacions en diversos gens,
com CD40L. CD40L és important per la col·laboració T-B. Si es té la mutació en CD40, es
considera que es té una síndrome d’hiper-IgM de tipus 1. La síndrome també pot venir donada
per una mutació en CD40, cas en què es té una síndrome d’hiper-IgM de tipus 3. Finalment,
la síndrome d’hiper-IgM de tipus 2 és aquella que es veu provocada per una mutació en el
gen AID, la D-aminasa induïda per activació. Tots tres elements són essencials per a la
col·laboració T-B i per al canvi d’isotip.

Immunologia 2020
CEBIO 255
La immunodeficiència de tipus 1 està lligada al cromosoma X i es detecta quan hi ha nivells
molt elevats de IgM al sèrum. No només són nivells molt elevats, sinó que és l’únic tipus
d’immunoglobulina que es genera perquè no es dona un canvi d’isotip. També pot passar que
el canvi d’isotip es faci en molt poc grau, cas en què IgG i IgA són pràcticament indetectables.
En aquest cas, CD40 i CD40L també tenen una resposta cel·lular deficient.
La síndrome d’hiper-IgM sol aparèixer en nens majoritàriament barons que fan infeccions
piògenes de repetició. Sovint la infecció pot ser una pneumònia per Pneumocystis carinii.
També sovint, aquests nens tenen autoanticossos que compliquen la malaltia. Aquesta
producció exagerada de IgM i aquest mal control de la producció dona lloc a dipòsits
d’immunocomplexes.
Si es mira la imatge d’un limfonode d’un individu amb la síndrome d’hiper-IgM, es veu que
no hi ha formació de fol·licles limfoides de limfòcits B al còrtex i tampoc hi ha l’estructuració
dels centres germinals que es veu en els individus sense la immunodeficiència.
Les persones immunodeficients responen bé a la infusió intravenosa d’immunoglobulines o
al transplantament de moll d’os.
Immunodeficiències Primàries Cel·lulars combinades.
Hi ha immunodeficiències Primàries de la resposta cel·lular. Encara que siguin defectes
selectius de la resposta cel·lular (no hi ha limfòcits T a la sang), la col·laboració dels limfòcits
T i B i altres cèl·lules del sistema immunitari fa que aquestes immunodeficiències primàries
cel·lulars afectin tot el sistema immunitari. Per tant, es sol parlar d’immunodeficiència
combinada.
Un exemple d’immunodeficiència de resposta cel·lular és la síndrome de Di George. És una
Aplàsia Tímica Congènita. Es produeix un desordre degut a la deleció del braç llarg del
cromosoma 22, que afecta tota una sèrie de gens entre els que es troba T-box 1. Genera
defectes d’embriogènesi que afecten en el desenvolupament del timus, de la paratiroides i
dels vasos sanguinis. Per tant, aquests individus tenen un funcionament deficient de les
paratiroides, tenen problemes cardíacs degut a les deficiències i defectes dels grans vasos
sanguinis i tenen infeccions recurrents per micobacteris, virus i fongs. A més, tenen una
alteració facial característica amb les parpelles caigudes i una forma especial de boca. El
model animals de la síndrome és el ratolí node, atímic. No té pel i té tota una sèrie
d’afectacions.
Els problemes en el desenvolupament del timus fa que el desenvolupament de l’epiteli tímic
sigui deficient, així com també ho serà la maduració dels limfòcits T, el procés de selecció
tímica i la diferenciació dels limfòcits T. L’alteració de les glàndules paratiroides fa que hi hagi
una alteració en la homeòstasi del calci. El metabolisme del calci, entre altres coses, és molt
important en la senyalització de diferents vies del sistema immunitari com la del TCR.
En Conseqüència, els individus amb aquesta malaltia tenen aquesta sèrie d’afectacions. Té
de “bo” que, amb l’edat, pot disminuir. Millora en tant que el desenvolupament del timus perd
importància en el cos, pel que els efectes de la immunodeficiència es suavitzen una mica.
També són causa d’immunodeficiències primàries de la resposta cel·lular les alteracions o
mutacions en els gens que codifiquen per les proteïnes TAP1 i TAP2, així com el gen CIITA,
que és el que regula la transcripció dels gens que codifiquen per les molècules del complex
del MHC de classe II. Per tant, aquestes mutacions suposen dèficits en molècules de MHC
de classe I per TAP i de classe II per CIITA. Afecta tant als limfòcits TCD4 i TCD8, però també
afecta al conjunt de la resposta immunitària.

Immunologia 2020
CEBIO 256
S’han descrit dèficits en molècules o proteïnes com ZAP-70, que és essencial en la via de
senyalització del TCR, i en components del complex CD3. Aquestes mutacions afecten als
limfòcits T i també al conjunt de la resposta immunitària.
El cas més sever són les immunodeficiències severes combinades ja que afecten al
desenvolupament dels limfòcits T o dels limfòcits B. Sovint també tenen afectacions en el
desenvolupament de les cèl·lules NK. Per tant, són combinacions de defectes T, B o NK.
Aquests individus són nens que en els primers mesos de vida tenen infeccions de repetició
víriques, bacterianes, fúngiques i per protozous. Per tant, poden tenir infeccions diverses i
recurrents per diversos tipus de patògens. També es pot donar, si en el part hi ha un pas de
cèl·lules maternes als nadons, una malaltia de l’empelt contra l’hoste. Això és perquè tenen
un funcionament deficient de tot el sistema immunitari.
S’han descrit immunodeficiències severes combinades resultat de mutacions en gens com la
cadena γ comuna del receptor de la interleuquina 2 però que, a més, afecta els receptors de
totes les citocines que utilitzen aquesta cadena: IL2, IL4, IL7 i IL9. El resultat de la mutació és
una deficiència total de limfòcits T i de cèl·lules NK tot i que la funció B es manté en humans.
També s’han vist mutacions en la JAK quinasa 3, que és important en la senyalització
mitjançant el receptor de la IL2, IL4, IL7 i IL9. Per tant, té la mateixa afectació.
Les deficiències en el receptor de la IL7 afecten a la resposta T però no afecta a la resposta
NK.
També s’han descrit mutacions en els gens RAG1, RAG2 i Artemis; relacionats amb el procés
de recombinació somàtica per la generació de BCR i TCR. Per tant, dona lloc a deficiències
en la resposta T i la resposta B.
Deficiències en ADA alteren les respostes T, B i NK. Pot ser totalment deficient segons quina
sigui la mutació o alteració d’aquests gens. Són immunodeficiències molt severes i que limiten
molt la vida dels individus que la tenen.
Immunodeficiències Secundàries o adquirides. Les immunodeficiències secundàries són aquelles que no estan codificades en els gens, sinó
que són adquirides. Són de causa molt diversa. Poden sorgir d’una mala nutrició proteïno-
calòrica, que inhibeix la maduració dels limfòcits i genera una resposta adaptativa deficient.
Poden ser degudes a afectacions tumorals de la medul·la òssia, que redueix la formació de
precursors o que, si es produeix una infiltració paraneoplàstica d’aquesta medul·la, pot limitar
l’espai pel desenvolupament dels leucòcits i causar una immunodeficiència. També pot ser
deguda a l’administració de fàrmacs o resultat d’una irradiació que elimini determinats tipus
cel·lulars.
Així i tot, les més corrents són els que són resultat d’una infecció per paràsits, bacteris o virus
que causen immunodeficiències temporals o definitives en els individus que les pateixen. El
cas més típic és el del virus del VIH, que causa la SIDA. En aquest cas, és un virus que infecta
cèl·lules del sistema immunitari i que destrueix aquesta població, produint una
immunodeficiència. S’ha vist que la infecció pel virus del xarampió causa immunosupressió
temporal i que es recuperen les funcions al cap d’un temps.
Les immunodeficiències posteriors a infeccions són les més habituals i de les que es parla
més sovint. També s’han descrit immunodeficiències adquirides resultants de patologies
autoimmunitàries.