BM-1.Estructura Primaria Del DNA
20
Tema 4: Estructura primària del DNA 0. Passos que es van fer per poder arribar a la seqüenciació Per tal de poder arribar a la seqüenciació, mètode amb el qual podem conèixer perfectament l’estructura primària en primer lloc es van tenir que fer una seria de passos importants en els quals destaquem: • Descobriments de Mendel (1866) Lleis de l’herència biològica, defineix en concepte de gen (era una definició teòrica, no feia referencia a cap molècula). Aquest concepte es l’antítesi de la biologia molecular • Teoria cromosòmica de l’herència: Es un punt molt important perquè va ser el primer moment que el concepte de gen adopta un significat físic ja que segons la teoria un gen es allò que podem trobar en els cromosomes. • Afirmació de que el material genètic es troba en el DNA • Creació dels mapes genètics • Creació dels mapes físics • Hibridació (desnaturalització +reasociació del DNA) • Anàlisis de les dimensions de l’estructura primària • Estudi de les endonucleases de restricció 1. Mapes genètics i mapes físics 1.1 Mapes genètics Definició: Es una representació de la situació dels gens en els cromosomes i de les distancies relatives que hi han entre elles. Aquets mapes es realitzen de forma indirecte ja que es generen gracies a l’observació fenotípica de la recombinació (producció de gàmetes recombinants durant la meiosi), d’aquesta manera es coneix que la freqüència de recombinació es inversament proporcional a la distancia relativa entre dos gens (dos gens que sempre van junts presenten una distancia molt petita) = ! !" !"#$%&’()(*+ ! !"!#$ !" !"#$"%!"%&# · 100 Hem de tenir en compte que 1% de unitat de mapa=1centiMorgan. Aquestes unitats son relatives ja que el mapa es relatiu. Els primers mapes genètics van ser realitzats per Morgan que va utilitzar els cromosomes de la Drosophila, tot i que després s’han realitzats mapes genètics de molt altres individus com poden ser els de Ecoli. Aquets mapes genètics també van permetre demostrar que els cromosomes bàsicament son com una línia en els quals els gens estan ordenats seqüencialment
-
Upload
ignasigrasscosta -
Category
Documents
-
view
27 -
download
21
description
Tema 1Professor: Ramon DabanAssignatura: Biologia Molecular
Transcript of BM-1.Estructura Primaria Del DNA
-
Tema 4: Estructura primria del DNA
0. Passos que es van fer per poder arribar a la seqenciaci Per tal de poder arribar a la seqenciaci, mtode amb el qual podem conixer perfectament lestructura primria en primer lloc es van tenir que fer una seria de passos importants en els quals destaquem:
Descobriments de Mendel (1866) Lleis de lherncia biolgica, defineix en concepte de gen (era una definici terica, no feia referencia a cap molcula). Aquest concepte es lanttesi de la biologia molecular
Teoria cromosmica de lherncia: Es un punt molt important perqu va ser el primer moment que el concepte de gen adopta un significat fsic ja que segons la teoria un gen es all que podem trobar en els cromosomes.
Afirmaci de que el material gentic es troba en el DNA Creaci dels mapes gentics Creaci dels mapes fsics Hibridaci (desnaturalitzaci +reasociaci del DNA) Anlisis de les dimensions de lestructura primria Estudi de les endonucleases de restricci
1. Mapes gentics i mapes fsics
1.1 Mapes gentics Definici: Es una representaci de la situaci dels gens en els cromosomes i de les distancies relatives que hi han entre elles. Aquets mapes es realitzen de forma indirecte ja que es generen gracies a lobservaci fenotpica de la recombinaci (producci de gmetes recombinants durant la meiosi), daquesta manera es coneix que la freqncia de recombinaci es inversament proporcional a la distancia relativa entre dos gens (dos gens que sempre van junts presenten una distancia molt petita) = ! !" !"#$%&'()(*+! !"!#$ !" !"#$"%!"% 100 Hem de tenir en compte que 1% de unitat de mapa=1centiMorgan. Aquestes unitats son relatives ja que el mapa es relatiu. Els primers mapes gentics van ser realitzats per Morgan que va utilitzar els cromosomes de la Drosophila, tot i que desprs shan realitzats mapes gentics de molt altres individus com poden ser els de E-coli. Aquets mapes gentics tamb van permetre demostrar que els cromosomes bsicament son com una lnia en els quals els gens estan ordenats seqencialment
JR.Daban BIOLOGIA MOLECULAR Figura 4.2
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.3
-------------------------------------------------------------------------------------------------
-
Aquets mapes no sn absoluts, sin que son relatius
1.2 Mapes fsics Definici: Son representacions reals de la collocaci dels gens en els cromosomes. Aquets mapes es realitzen de forma directa ja es poden fer utilitzan tres tcniques
Histolgiques: Es basen en tcniques dhibridaci in situ amb radioactivitat o FISH, es a dir sondes (cDNA) que son complementaries a un gen i que estan marcades amb radioactivitat o amb fluorescncia Ex: Marcaci de gens amb cDNA marcat radioactivament en el cromosoma gegants (politnics) de Drosophila.
Bioqumiques: Prpiament es diuen mapes de restricci ja que es basen en els diferents fragments que generen les endonucleases de restricci
Mitjanant seqenciaci: Es la manera en el qual obtenim un mapa 100% fiable.
1.3 Importncia daquets mapes Per tal de poder-se plantejar la seqenciaci, i el projecte del genoma hum, ha sigut molt important tota la feina feta pels mapes gentics i els mapes fsics ja que sense aquets coneixements basics dels cromosomes i dels gens i per tant de lestructura primria hagus sigut impossible aquesta seqenciaci
2. Hibridaci del DNA Definici: La hibridaci del DNA ens permet tenir una idea de la complexitat que presenta aquest i conseqentment tamb la repetici, sent que la complexitat depn de la seqencia que presenta. Per tant ens dona una primera idea de com pot ser les seqencies dels diferents genomes i per tant una primera idea de lestructura primria. Com veurem tamb les condicions dhibridaci tamb amb permetran regular el gru de Actualment la hibridaci te moltes aplicacions i es una de les potes de lenginyeria gentica
2.1 Preparaci del DNA Definici: Per tal de poder fer estudis dhibridaci o de resociaci del DNA es necessari fer un tractaments previs:
JR.Daban BIOLOGIA MOLECULAR Figura 4.4
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.5
-------------------------------------------------------------------------------------------------
-
Purificaci del DNA Fragmentaci del DNA
Purificaci del DNA: Antigament es feien les purificacions mitjanant solvents orgnics, actualment exsteixen kits de purificaci que es basen en els segents passos: 1. Lisis cellular: Es fa mitjanant un pH alcal que es capa de provocar que les
cllules rebentin. Hem de tenir en compte que aquest pH tamb provca que el DNA es desnaturalitzi per aquest quan eliminem el medi alcal recuperar de forma rapida la seva estructura
2. Introducci del DNA a una columna de bescanvi inic: Aquesta columna conte rena que es capa dunir-se el DNA. Podrem diferenciar els segents passos: 2.1. Adsorci: Es basa en la introducci del material lisat. Com a conseqncia es
quedar unit els diferents tipus de DNA per no o faran ni les protenes ni el RNA
2.2. Neteja: Es basa en la eliminaci dels RNA o protenes residuals que puguin haver quedat en la columna. La neteja es fa amb una soluci salina de NaCl. Aquesta mateixa soluci es la que utilitzar per fer la eluci per tant es necessari que coneguem quina es la [NaCl] en la qual salta el DNA.
2.3. Eluci: Es basa en la obtenci del DNA. Per aix utilitzar una concentraci creixent de NaCl. Hem de tenir en compte que els diferents tipus de DNA eluiran a concentracions diferents de NaCl
Aquets mtode es fora ptim ja que per una banda com podem veure la electroforesis desprs de leluci aconseguim una banda prima i estreta i per tant aix vol dir que esta ben purificat, i per laltre banda el temps que es necessita per aconseguir el DNA es molt curt ja que amb 30m ja en tens prou Cal destacar tamb que aquet mtode de purificaci tamb es pot aplicar a gran escala amb sistemes que contenen mes de 90 columnes de rena, en els qual la elusi no es fa per gravetat sin per bomba de buit, i per tant la purificaci encara es ms rapida, i a ms a ms estan automatitzats. Daquesta manera en poques hores puc obtenir una gran quantitat de DNA i aquest fet es molt important per la seqenciaci massiva o a nivell clnic Fragmentaci del DNA: Aquesta fragmentaci es pot fer mitjanant dos tcniques:
Ultrasonicaci: Es basa en sotmetre la preparaci del DNA a ultrasons dalta freqncia que sn capaos de produir una pertorbaci mecnica en l DNA. Saconsegueixen fragments heterogenis de 100pb
Xeringa hipodrmica: Es basa en xuclar i deixar anar la preparaci del DNA unes quantes vegades ja que daquesta manera saconsegueixen unes forces hidrodinmiques capaces de fraccionar el DNA
Hem de tenir en comte que el DNA ja es fragmenta nomes a partir de la seva manipulaci.
JR.Daban BIOLOGIA MOLECULAR Figura 4.6
-------------------------------------------------------------------------------------------------
-
Temperatura optima dhibridaci: Es va poder determinar que aquesta temperatura era TM-25 Astringncia/severitat de la hibridaci: Hem de tenir en compte quines son les condicions en les quals hibridem ja que segons aquestes el grau de complementarietat dels hbrids formats pot ser un o un altre. Les variables que podem regular son aquelles que son capaces destabilitzar o desatabilitzar la doble hlix, es a dir, la temperatura i la concentraci salina (apantallament de carregues negatives). Daquesta manera podem diferenciar entre dos tipus de condicions diferents:
Condicions astringents: Son condicions en les quals la doble hlix esta desestabilitzada ja que les condicions sn:
o Temperatura elevada o Fora inica baixa
En aquestes condicions nomes les seqencies que presentin una complementarietat total seran les que es podran hibridar
Condicions relaxades: Son condicions en les quals la doble hlix esta estabilitzada ja que les condicions sn:
o Temperatura baixa o Fora inica alta
En aquestes condicions nomes les seqencies que no son totalment complementaries i per tant poden aparixer hbrids inespecfics que no presentin una complementarietat total.
2.2 Punts claus de la hibridaci Fases de la hibridaci:
Nucleaci: Es la fase lenta i es el pas mitjanant el qual shan de comenar a hibridar les bases complementaries. Es la fase que marca la velocitat de lhibridaci i la que podem observa. Per aquest mateix motiu es necessari que fragmentem el DNA ja que en el cas del DNA circular la nucleaci es molt ms fcil i per tant mes rapida i aix ens fa impossible poder observar la hidridaci
Propagaci o zipperng: Es la fase rapida i es pas mitjanant el qual es propaga la hibridaci com si fos una cremallera a partir de les bases que shan ajuntat en la nucleaci. Es una fase molt rapida i que no podem observar
Detecci de la hibridaci: Per tal de detectar el procs dhibridaci es poden utilitzar diferents tcniques
Efecte hipercromic (baixada de labsorvancia) Bromur detidi: La fluorescncia augmentar a mesura que obtinguem dsDNA Columna dhidroxiapatita: Es una columna que es capa dunir el dsDNA,
mentre que el ssDNA passa lliurament. Per tant si anem afegint solucions en el qual sest hibridant el DNA a diferents temps podrem veure com cada vegada sunir ms DNA a la columna i per tant obtindrem menys ssDNA. Es el sistema ms utilitzat.
Dependncia de la longitud: La hibridaci depn completament de la longitud del DNA de forma directament proporcional sent que a major longitud ms temps es necessitar per aconseguir la hibridaci complerta. Ex: Temps dhibridaci de diferents DNA. Tenim tres tipus de DNA
-
Vedella: La hibridaci es produeix de forma molt lenta i quasi mai es aconsegueix una hibridaci complerta (histresino es recuperen les condicions inicials)
Bacterifag M13: La hibridaci es produeix de forma rapida DNA clonat del nucleosoma de 146pb: La hibridaci es produeix de forma
instantnia i no es pot detectar
2.3 Cintica dhibridaci Raonament matemtic: Hem de tenir en compte dos definicions:
C0: Concentraci de DNA reasociat a t=0 C: Concentraci de DNA reasociat a t=t
Per tant hem de tenir en compte que ! !! es una manera dexpressar la quantitat de S si fssim una analogia amb una reacci normal ja que es una relaci entre el DNA que tenim respecte el DNA inicial. La concentraci de DNA es mesura en pb/L per tant es una manera de representar la longitud del DNA
Podem veure que els factors que son variables i que per tant podem controlar sn el t i la C0 tenint en compte que a ms t i ms C0 ms fracci saturada. Per tant dit dun altre manera i com ja hem dit abans a ms longitud ms temps es necessita. Representaci: Una representaci que es pot fer i ens pot ajudar a representar els resultats es:
Eix x: C0t Eix y: 1 ! !! (fracci reassociada)
En aquesta grfica poden definir un nou parametre que es C0t1/2 que es el valor de C0t on hem obtingut la mitat de la reasociacio. Es un valor que ens ajuda a comparar diferents DNA.
2.4 Complexitat del DNA Definici: Es una caracterstica intrnseca al DNA que es capa de modificar la k de velocitat de la reacci i la C0 i daquesta manera la velocitat de reasociaci Explicaci: Imaginem un conjunt de seqencies de la mateixa longitud i determinarem els diferents valors de k i de C0
Seqencia pol(A)-pol(T): En aquesta seqencia la velocitat de nucleaci la considerarem mxima ja que durant la nucleaci tots els xocs que es produeixin seran efectius i conseqentment tota la C0 ser efectiva
o k=kmax
!
v = k2 [ssDNA(+)][ssDNA()]
v = k2C 2 =dCdt
dCdC 2 =C0
C
k2 dt0
t
11C
1C0
= k2t 21C =
1C0
+ k2t xC0 C0C = 1+ k2C0t
invers CC0= 11+ k2C0t
=
fracci no reasociada CC0= 11+ k2C0t
fracci reasociada 1 CC0
-
o C0 efectiva = C0 Seqencia pol(AG)-pol(TC): En aquesta seqencia la velocitat de nucleaci
ser la mitat de la velocitat mxima ja que durant la nucleaci nomes els xocs en els quals la A semparelli amb la T i la G amb la C el xoc ser efectiu, conseqentment nomes la mitat de C0 ser efectiva
o k= !!"#! o C0 efectiva = !!!
Seqencia pol(ACG)-pol(TGC): En aquesta seqencia la velocitat de nucleaci ser una tercera part de la velocitat mxima ja que durant la nucleaci nomes els xocs en els quals la A semparelli amb la T i la G amb la C i C amb G el xoc ser efectiu, conseqentment nomes una tercera part de C0 ser efectiva
o k= !!"#! o C0 efectiva = !!!
Seqencia aleatria de longitud x amb cap base repetida de forma consecutiva i repetida n vegades: En aquesta seqencia la velocitat de nucleaci ser x vegades la velocitat inicial de la velocitat mxima ja que durant la nucleaci nomes els xocs en els quals siguin completament complementaris ser efectiu, conseqentment nomes una x part de C0 ser efectiva
o k= !!"#! o C0 efectiva = !!!
Per tant podem veure com a partir destudis simples dhibridaci puc conixer quina es la complexitat del DNA i per tant puc tenir una idea de quina es lestructura primria del DNA ja que aquesta complexitat depn de les propietats de la seqencia. Unitat: La manera que tenim de compara la complexitat de genoma es a partir de al quantitat de pb que shan de daparellar durant la nucleaci perqu es doni un xoc efectiu, es a dir, determinar el valor x. Aquest valor x el puc obtenir a partir de les dades cintiques C0t sent que mes gran sigui aquest valor ms complexitat tindrem Ex: Daquesta manera poden dir que
o Seqencia pol (A)-pol(T): Presenta un complexitat de 1 o DNA de MS2: Presenta un complexitat de 3500 o DNA de T4: Presenta un complexitat de 1,7105 o DNA de E.coli: Presenta un complexitat de 4,2106
2.5 Complexitat dels organismes complexos Si mirem la complexitat dels organismes complexos, podem veure que aquets presenten diferents tipus de complexitat ja que presenten C0t1/2 , en concret 3, es a dir
JR.Daban BIOLOGIA MOLECULAR Figura 4.7
-------------------------------------------------------------------------------------------------
-
que dalguna manera presenten 3 tipus diferents destructures primries del DNA: una DNA dhibridaci rapida, un DNA dhibridaci intermdia i un DNA dhibridaci lenta . Si fem la representaci podem veure com hi han 3 corbes solapades.
*Quantitat de pb: Per tal de poder-ho determinar primer hem de conixer la quantitat de DNA que presenta una cllula haploide (valor C) i per tant el que es fa es agafar cllules, contar quantes en tenim i quan DNA tenim mitjanant algun mtode com pot ser lespectroscpia. Posteriorment dividirem la quantitat de DNA en vers les cllules que tinguem i el resultat el anirem multiplicant pels diferents percentatges. **Freqncia de repetici: Ens indica el n de vegades que es repeteix una seqencia en concret. Sobt a partir de dividir la quantitat de pb entre la complexitat. A partir de la freqncia de repetici tamb podem determinar que els diferents tipus de DNA el podem classificar entre:
DNA repetitiu (500.000) DNA poc repetitiu (350) DNA no repetitiu (1)
Com ms repetida estigui una seqencia , menys complexitat vol dir que presenta i per tant tindr un C0t1/2 ms petit. Daquesta manera a partir de la complexitat i de la longitud del DNA a partir danalisis cintics puc conixer com de repetitiu es un DNA, i conseqentment conixer ms informaci de lestructura primria del DNA. Shan realitzats aquets experiments en molts altres especies i sha pogut observar que la quantitat de DNA no repetiu varia ja que pot anar des de un 100% com en el cas dels bacteris o llevats, fins a un valor inferior al 50% com pot ser les plantes. Si observem la taula podem veure com a mesura que lorganisme es mes complex apareix mes DNA repetitiu i per tant menys complex.
2.7 Paradoxa del valor C Definici: Es una contradicci entre la longitud del genoma haploide (valor C) i la complexitat del organisme. Malgrat que seria lgic pensa que contra ms complexitat ms quantitat de genoma aix no te perqu ser aix Demostraci: Es van determinar els diferents valors C de difernts organismes amb diferents complexitats i es va definir que la complexitat depenia de la quantitat de cllules diferents que presentava un organisme (a ms cllules ms complexitat) Es va veure que existien alques que presentaven una gran quantitat de DNA malgrat que eren organismes molt poc complexos. Per aquest motiu es va determinar la teoria del DNA selfish o egoista
JR.Daban BIOLOGIA MOLECULAR Figura 4.7
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.8
-------------------------------------------------------------------------------------------------
-
DNA selfish o DNA egoista: Es un DNA que no codifica per cap protena i que la seva nica voluntat es replicar-se
2.8 Protein-conding-Gens vs Genome size Si ens fixem en la relaci entre la quantitat de gens que codifica un DNA i la seva longitud podem veure que hi ha una relaci ja que contra mes grans ms gens. Aquesta relaci per no es lineal (presenta diversos pendents i es una escala logartmica) i aix es degut al fet de lexistncia del DNA repetitiu que fa que organismes amb genomes molt grans presentin molt del seu DNA ocupat per seqencies que no es codifiquen.
2.9 Aplicacions de la hibridaci Actualment la hibridaci te moltes aplicacions tant a nivell dinvestigaci, com a nivell de diagnstic clnic. Hi a ms a ms sha convertit en una de les potes de lenginyeria gentica
2.9.1 Separaci de la cadena de Watson(+) i la cadena de Crick(-) Objectiu: En aquest experiment sha clonat una seqencia de DNA de 146 pb a partir de la inserci daquesta en el bacterifag M13, i el que es vol obtenir es cadascuna de les dues cadenes daquesta seqencia per separat, es a dir, cadascuna en un tub. Per aquest motiu utilitzarem tcniques dhibridaci i tamb de precipitaci/purificaci del DNA. En aquest cas la precipitaci del DNA es far amb polietilenglicol (PEG) que es capa de discriminar entre dsDNA i ssDNA com tamb entre diferents mides de DNA Bacterifag M13: Es un virus molt utilitzat en biologia molecular gracies als diferents estats en que pot estar el seu DNA. Aquest bacterifag realitza la infecci a traves del pils del bacteri i presenta un DNA circular. Els diferents estats del seu DNA varien segons on es trobi el virus:
Fase extracellular: Es el estat es ssDNA Fase intracellular: El seu estat es dsDNA
Per tant segons si agafem els bacteris del medi extracellular o a partir dun lisat de bactries podrem obtenir o be ssDNA o dsDNA Separaci dels fragments: En aquest ens interessa tindre el ssDNA i per aquest motiu agafar els bacteris de la seva fase extracellular i el que tamb es necessari es el tros del DNA de 146pb en la seva forma dsDNA.
1. Afegir en un tub el vector que probe del bacterifag (ssDNA) i el fragment de 146pb (dsDNA).
2. 1er Desnaturalitzaci: Augmentem la temperatura fins a 100C i com a conseqncia el fragment de DNA de 146pb es separ en les seves dues cadenes
3. Renaturalitzaci: Baixem la temperatura i com a conseqncia es podr tornar a renaturalitzar el fragment de 146pb per una delles tamb ser capa dhibridar amb el fragment inserit en els ssDNA del bacterifag. Com a conseqncia obtindrem fragment de 146pb dsDNA, genoma del bacterifag amb el fragment inserit en formar dsDNA i fragmenten de 146pb ssDNA(+).
JR.Daban BIOLOGIA MOLECULAR Figura 4.8
-------------------------------------------------------------------------------------------------
-
4. Recuperaci del fragmenten de 146pb ssDNA(+): Afegirem PEG al 16,5% en el medi i centrifugarem. Com a resultat obtindrem un sobrenedant en el qual shi trobar el fragment de 146pb ssDNA(+)
5. Recuperaci del fragment de 146pb dsDNA: Resuspendre el sediment de la centrifugaci anterior amb una soluci que conte PEG al 9% i tornem a centrifugar. Com a resultat obtindrem un sobrenedant en el qual shi trobar el fragment de 146pb dsDNA. Aquest fragment podr ser utilitzat per altres cicles de purificaci
6. 2n Desnaturalitzaci: Generem un pH acid de 1,8 i com a conseqncia el , genoma del bacterifag amb el fragment inserit en formar dsDNA separar i per tant obtindrem el fragment de 146pb ssDNA(-)
7. Recuperaci del fragmenten de 146pb ssDNA(-): Afegirem PEG al 5% en el medi i centrifugarem. Com a resultat obtindrem un sobrenedant en el qual shi trobar el fragment de 146pb ssDNA(-)
3. Anlisi de les dimensions de lestructura primria del DNA El DNA presenta una dimensions molt grans ja que es el biopolimer mes gran que existeix. Per tal de poder estudiar les seves dimensions podem utilitzar diferents tcniques com poden ser la microscpia ptica i electrnica (es necessita una preparaci prvia), la electroforesis de camp pulsant i altres tcniques de suport com poden ser la citometria de flux (FACS)
3.1 Observaci del DNA amb micoscopia
3.1.1 Observaci del DNA amb microscpia electrnica. Tinci de Kleinschmidt Protocol: La preparaci del DNA per tal de poder-lo observar amb microscopi electrnic sanomena tinci de Kleinschmidt i existeix moltes variants. Aquesta preparaci es primordial perqu el DNA presenta una secci transversal molt petita (2nm) que fa que quasi be no es pugui observar amb TEM i al mateix temps presenta una secci axial (llargada) molt gran i aix fa que pugui forma grumolls de forma molt fcil i aix impossibilita la observaci. Per aix sutilitza una protena hidrofbica, anomenada citocrom C, que pot interaccionar amb el DNA si es troba en la seva forma desnaturalitzada. Aquest citocrom C permetr que la secci transversal del DNA augmenti i per tant sigui mes fcil observar el DNA. Els passos a segui sn:
1. Collocar sobre un parafilm una gota duna soluci que conte el DNA que volem observar i citocrom C, i una altre gota de formaldehid i a continuaci o taparem amb una placa de petri. El formaldehid es voltil i es un agent que es capa de
JR.Daban BIOLOGIA MOLECULAR Figura 4.10
-------------------------------------------------------------------------------------------------
-
desnaturalitzar el citocrom C. Per aquest motiu es formar una capa de citocrom C desnaturalitzat en la interfase aire-aigua, i aquest citocrom desnaturalitzat presentar una carrega parcial positiva que permetra interecciona r amb la carrega parcial negativa del DNA.
2. A continuaci ficarem just a sobre la gota que conte el DNA recobert de citocrom C la reixeta dobservaci i el DNA sunir a ella per capillaritat. Aquesta reixeta es de coure o de or i esta recoberta per un film de carboni.
3. Deixarem sacar la mostra i farem una tinci positiva amb acetat duranil i una posterior platinaci que permetr tenir un ombrejat bidireccional ja que la mostra anir rodant. La mostra resultat tindr una secci transversal de 20nm
El DNA que nomes presenta platinaci tamb es pot observar tot i que presenta ms dificultats Ex: DNA de Bacterifag T2 sotms a xoc osmtic
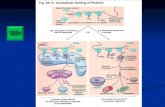
3.1.2 Observaci del DNA amb microscpia ptica. Teoria Uninmica Protocol: El DNA es una molcula massa petita com per poder-la veure amb microscpia ptica, per en canvi si que podem veure una autoradiografia. Els passos a seguir per fer aquesta autoradiografia sn:
1. Creixement de les cllules en un medi que presenta timidina tritiada (H3) com a nica font de timidina. Com a resultat el DNA que es crea estar marcat radioactivament
2. Allament del cromosoma i extensi daquest en una placa de vidre 3. Sobre aquesta placa de vidre si colloca una emulsi fotogrfica (presenta els
mateixos components que un carret fotogrfic) 4. Al cap de 5 mesos dexposici (el triti es poc radioactiu) en aquelles zones on hi
ha DNA i per tant ho ha radioactivitat hi haur precipitaci de plata. 5. Revelat.
Ex: Cromosoma politnic de Drosophila Teoria Uninmica: Es una teoria que es va realitzar gracies a lobservaci dels cromosomes i que diu que cada cromosoma conte una sola molecula de DNA i que presenta una longitud molt gran. La longitud del cromosoma lgicament varia segons lespecia ja que el dhumans es troba en lordre de cm, per el danfbis pot arriba ra
JR.Daban BIOLOGIA MOLECULAR Figura 4.12
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.12
-------------------------------------------------------------------------------------------------
-
medi fins i tot 1m i existeix el cas duna planta, la pavis japonica, que el DNA te una longitud de 3m. Per tant els cromosomes fets de cromosomes son el biopolimers mes llarg que existeix Amb experiments com el anterior tamb es va trobar empricament que 1m de DNA li corresponen 3000pb. Es un resultat fiable ja que = 13000 = 1 10!3000 = 0,33 0,34 3.2 Separaci dels diferents cromosomes. FACS Definici: El FACS (separador de cllules activat per fluorescncia) es una tcnica que utilitza la citometria de flux per poder serara diferents cllules, per tamb presenta la capacitat de separa els cromosomes segons la seva grandria. Procediment: En primer lloc es necessari aconseguir els cromosomes i per aix bloquejar les cllules en colchemid que es un agent qumic capa de sincronitzar-me les cllules en metafase. A continuaci aplicar una tinci que sigui capa de tenyir el DNA com pot ser el Hoetsh o la Cromomicin i per acabar far un xoc osmtic per tal de rebentar les cllules. Els cromosomes lliures els far passar pel FACS, aquest aparell es capa de crear un rajol tant prim que fa que els cromosomes es colloquin un darrera el altre. Aquest rajol passa per una zona en la qual hi ha un lser que permet veure la fluorescncia que presenta cada cromosoma i depenen de quina quantitat de fluorescncia (depn de la mida del cromosoma) que detecti carregar la gota duna manera o dun altre. Aquestes gotes passaran per un camp elctric i segons la seva carrega aniran a parar a un tub o a un altre i daquesta manera aconseguirem separar els diferents cromosomes.
3.3 Electroforesis de camp polsant Definici: Es una electroforesis en el qual el camp elctric no es constant, es a dir, que va variant la direcci al llarg del temps. Aquesta tcnica amb permet separar molcules de DNA fins a un mxim de 2000kb Aquesta tcnica va estar desenvolupada per Cantor i actualment ja esta patentada i te moltes utilitats, com per exemple detectar la presencia de bacteris en el hospital a partir de conixer la mida del DNA daquest.
3.1.1 Problemes de la electroforesis normal Problema: Els problemes de la electroforesis normal es que no ens permet separar molcules de DNA molt grans. Causa: El gels utilitzats sn el de poliacrilamida i el dagarosa (agents polimeritzadors) Aquets gels permeten separar correctament diferents rancs de mides de DNA segons quina sigui la seva concentraci, sent que com ms petita es la concentraci ms gran ser el DNA que podrem separar. Existeixen dos factors que son els que amb limiten la mxima longitud que puc separar
La no polimeritzaci del gel: Per tal de que polimeritzi el gel es necessita una concentraci mnima del agent polimeritzador:
o Poliacrilamida: 3,5%. Permet serapar un mxim de 1 o 2 kb
JR.Daban BIOLOGIA MOLECULAR Figura 4.11
-------------------------------------------------------------------------------------------------
-
o Agarosa: 0,3. Permet separar un mxim de 5-60kb. Presenta el porus ms gran que la poliacrilamida.
La resoluci: Existeix una dependncia entre la distancia migrada (Rf) i la quantitat de pb (logMr) i aix fa que es pugin separar els diferents molcules de DNA per la mida. Existeix per un mxim de 50kb en la qual la dependncia es pert i independentment de la quantitat de pb la migraci es la mateixa. Per tant a partir de 50pb ja no tenim resoluci ja que les molcules de DNA quasibe no es poden moure pel gel.
3.1.2 Tipus delectroforesis de camp polsant Existeixen diferents electroforesis de camp polsant segons quin sigui el sentit en el qual va canviant el camp electric. Destacarem:
Field inversion: El camp elctric canvia 180. Per tant primer va cap aball i desprs cap amunt. Hem de tenir en compte que el temps en el qual es migra cap avall ha de ser superior al temps en el qual es migra cap amunt ja que sin no ens mourem de lloc.
Ortogonal field: El camp elctric canvia 90. Per tant primer va cap a lesquerra i desprs cap a la dreta. No hem de tenir en compte la relaci entre els dos temps.
3.1.3 Criteri de separaci Per entendreu imaginem un field inversion en el qual el DNA anir cap avall 10s i i 5s cap amunt i aix es repetir n vegades. En aquest cas aconseguim que el DNA recorri 1/3 part del que recorreria en una electroforesis normal: = 10 515 = 13 Hem de tenir en compte per que nomes les molcules petits de DNA seran les que realment es moguin 1/3 del recorregut normal ja que les gran recorreran encara menys distancia. Aquest fet es degut a que quan canviem el camp la molcula de DNA es te que reorientar i aquesta reorientaci ser mes rapida en DNA petit que no pas en DNA grans. Per tant podem dir que la reorientaci es el criteri de separaci. En un principi la reorientaci va ser una teoria, per aquesta es va confirmar experimentalment gracies a Carlos Bustamante. Aquest cientfic va preparar un gel electrofortic dagarosa que contenia bromur detidi i va utilitzar el sistema ortogonal field. A continuaci va collocar aquest gel sota un microscopi de fluorescncia dalta
JR.Daban BIOLOGIA MOLECULAR Figura 4.13
-------------------------------------------------------------------------------------------------
-
resoluci i va observar que passava a partir del moment en que es canviava el sentit del camp. Hem de tenir en compte que no podem collocar molta concentraci de DNA perqu sin no es veurem pegotes. El que va observar es que a mesura que passava el temps el camp elctric provocava que apareguessin una seria de pertorbacions en la nova direcci del camp elctric i una daquestes es la que es feina mes gran i es la que permetia genera la fora suficient per poder reorientar tota la molcula. Lgicament com mes gran sigui la molcula, mes gran es necessita que sigui el vector fora i per tant ms gran a de ser la pertorbaci, conseqentment es necessita ms temps
3.1.4 Fragilitat del DNA Com hem vist som capaos de separar DNA de longitud molt gran, per com ja hem dit altres vegades la simple manipulaci del DNA fa que aquest es trenqui en troos petits, per tant hem de desenvolupar una tcnica per tal devitar la degradaci del DNA abans daplicar la electroforesis. Per aconseguir-ho es fa servir un motlle de plstic que presenta uns forats en forma de cubs.
1. El primer pas es tancar una de les dues bandes amb un parafilm. 2. A continuaci es colloca en el interior de cada cub 50% de volum de cllules
que contenen el DNA que volem analitzar i 50% de volum dagarosa calenta (50) i deixem refredar
3. Eliminem el parafilm a afegim el cub en un eppendorf 4. Afegim tamp de lisi i protenasa K (proteasa molt estable que en presencia de
SDS encara te activitat) en el eppendorf. Hem de tenir present que com que lagarosa presenta porus tant el tamp com la protenasa podran fer la seva funci de forma normal.
5. Recuperarem el nostra cub amb una esptula i el collocarem en els forats que presentar el nostre gel. Entre el cub i el gel no hi pot haver-hi cap forat i per aix afegirem agarosa fosa
4. Endonucleases de restricci Definici: Les endonucleases de restricci amb permeten obtenir els fragment de DNA que jo vulgui ja que presenten un alt grau despecificitat, es a dir, les puc utilitzar com unes tisores que tallen per all on jo vull. Aix fa que sigui la tercera pota de la enginyeria gentica
JR.Daban BIOLOGIA MOLECULAR Figura 4.14
-------------------------------------------------------------------------------------------------
JR.D
abanBIO
LOG
IA MO
LECU
LAR
Figura 4.16
-------------------------------------------------------------------------------------------------
-
4.1 Importncia de la metilaci Els enzims de restricci formen part dun doble sistema que engloba dos activitats
Restricci: Permet el tall Metilasa o metiltransferasa: Permet metilar les bases nitrogenades del DNA.
Aquestes metilacions es produeixen en el C6 citosina i adenina (eucariotes) i el C5 dadenines (procariotes) de les bases nitrogenades.
La funci fisiolgica dels enzims de restricci es talla el DNA exogen de la cllula i per tant configura un mecanisme de defensa. Mitjanant la metilaci podem reconixer si el DNA es propi o no i evitar lautodegradaci, daquesta manera podem diferenciar tres tipus de DNA:
DNA amb les dos cadenes metilades: Es reconeix com a propi i per tant no se li podr aplicar lactivitat de restricci
DNA amb una cadena metilada: Es reconeix com a DNA propi que acaba de ser replicat i per saplicar lactivitat metilasa per metila la cadena filla.
DNA sense cap cadena metilada: Es reconeix com a DNA exogen i per tant se li podr aplicar lactivitat de restricci
Per tant aquesta metilaci ens dona ja una grau superior desepecificitat. Aix es va descobrir en un experiment que va donar el punt de sortida a les endonucleases es va infectar dos soques de E.coli amb el mateix bacterifag
Soca K12: Van aparixer moltes calves Soca B: Van aparixer molt poques calves
Posteriorment es va infectar un altre cop les dos soques amb els virus que shavien generat en la soca B12 i es veure que
Soca K12: Van aparixer poques calves Soca B: Van aparixer moltes calves
La interpretaci va ser la segent: Cada soca presenta una endonucleasa diferent, i que per tant reconeix una seqencia del DNA diferent en el cas de la primera infecci els bacteris que es van utilitzar presentaven una metilaci en la seqencia del DNA que era reconeguda per lendonucleasa de K12 i per aquest motiu aquesta no li podia produir un tall, en canvi la de B si. En la segona infecci, els virus que havien sortir de la soca B tenien que presentar si o si una metilaci en aquella seqencia que reconeixia lendonucleasa de B ja que els seus progenitors lhavien de presentar si havien pogut infectar.
4.2 Tipus de endonucleases de restricci Existeixen dos tipus de endonucleases de restricci i cadascuna delles presenta caracterstiques diferents:
Grup I i III o Es una protena oligomerica que presenta les dues subunitats en una
mateixa protena
Metilaci de les bases nitrogenades
http://www.worthpublishers.com/lehningerTema 15. Biocatlisi
Metilaci de les bases nitrogenades
http://www.worthpublishers.com/lehningerTema 15. Biocatlisi
-
o El lloc te tall esta molt allunyat de la seqencia de reconeixement del DNA (unes 1000pb)
o Necessita ATP: Lenergia que aporti el seu trencament sutilitzar per provocar un desplaament en el DNA que apropi la sequancia de reconeixement i el lloc de tall
Grup II: Son les utilitzades com eines de la biologia molecular degut a les seves caracterstiques
o No es una protena oligomerica i per tant les dos activitats es trobaren en protenes diferents. Una activitat nucelasa sempre implica una activitat metilasa el que pasa es que moltes vegades aquesta no es coneix ja que no te inters i per tant no sinvestiga
o El lloc de tall i el lloc de reconeixement del DNA es el mateix o No necessiten ATP per necessiten Mg
4.3 Seqencia de reconeixement i tipus de tall Seqencia de reconeixement: Es la seqencia del DNA que es reconeguda per lendonucleasa de restricci. En el cas de les endonucleases de tipus II totes aquestes seqencies sn palndroms es a dir que presenten un eix de simetria rotacional binari ja que els enzims de restricci tamb presenten uns dmers amb simetria rotacional binaria . Cada endonucleasa tindr una seqencia de reconeixement especifica. Aix permet tenir no solsmanet una especificitat pel S sin tamb una especificitat per una zona del S Cada endonucleasa presenta una seqencia de reconeixement i actualment hi ha mes de 100 seqencies diferents Tipus de tall: Les endonucleases de tipus II poden produir dos tipus de tall i generear aix dos tipus dextrems
Extrems cohesius: El tall es produeix en diferents llocs en les dues cadenes, daquesta manera es generen uns extrems monocadena
Extrem romo: El tall es produeix en el mateix lloc en les dues cadenes , daquesta manera es generen uns extrems sense monocadena.
Quantes vegades podem trobar una seqencia de reconeixement: La formula per calcular-ho es la segent: = ()4! On q es n de nucletids de la seqencia de reconeixement. Daquesta manera en el cas de EcoR1 on la seqencia de reconeixement es 5GAATTC3 = 2 10!4! = 4,9 10! Com a mxim un enzim de restricci es capa de reconixer 8 nucleotids, per tant el nombre de llocs de tall es molt gran i aix dificulta el estudi i el treball amb lestructura primria per axi mateix necessitem tcniques de suport com poden ser la tecica bloting . Per aquest motiu actualment sest treballant en el desenvolupament
Nucleases de restricci tipus II
Tema 15. Biocatlisi
-
denzims de restricci que sigui capaos de reconixer seqencies de fins a 12 nucletids
4.4 Tcniques de bloting o de transferncia a membrana Utilitat: Com ja hem dit els estudis de lestructura primria amb el DNA utilitzant nomes els enzims de restricci es molt difcil ja que es produeixen molts fragments i per tant si realitzem per exemple una electroforesis el resultat seria una llapisada formada per un conjunt de bandes solapades i no definides. Aquesta tcnica amb permet seleccionar quin fragment del DNA vull tenir informaci ja que amb permet marcar just la banda que minteressa. Passos:
1. Electroforesis: Fem una electroforesis de DNA en el qual un carril presentar marcadors de pes molecular (DNA tallat amb enzims ja caracteritzats) i els altres carrils amb solucions de DNA que ha estat sotms a lacci de nucleases.
2. Desnaturalitzaci: En el cas de que el gel no sigui desnaturalitzat es important que el desnaturalitzem ja que ens interesa tenir el ssDNA per la posterior hibridaci
3. Transferncia a membrana: En primer lloc eliminarem un dels vidres de la electroforesis i retallarem les butxaques i tot aquell gel que no presenta bandes. Sobre el gel i collocarem un paper de filte. Es necessari que aquest paper no presenti cap bombolla daire i per aix passarem la ma per sobre vigilant no fer una pressi excessiva. Posteriorment separarem el vidre que encara est unit al gel suaument per amb decisi i daquesta manera ens quedar el gel allat amb una de les seves costats tapat amb paper de filtre. Es dipositar aquest gel pel costat que presenta el paper de filtre sobre un vidre que presenta dos papers de filtre humitejats amb tamp de recorregut. Tambe hi ha la possibilitat de no utilitzars els papers de filtre indicats anteriorment i en el seu lloc diposita el gel en un recipient que conte el tamp de recorregut Per laltre canto on el gel esta exposat i collocarem la nostre membrana (pot ser de diferents materials com Nylon o nitrocellulosa), lhumitejarem i passarem una bareta de vidre per sobre per evitar la formaci de bombolles. Per acabar afegirem dos papers de secant i una placa de vidre. Damunt de tot aquest complex es dipositaran un pes. El tamp de recorregut anir pujant per capillaritat al paper secant i aix amb permetr que les bandes es transfereixin a la membrana.
4. Obtenci de la membrana i fixaci de les bandes: Desmuntem tot el complex format anteriorment i aconseguim una membrana que conte les nostres bandes, igual que la electroforesis, per en aquest cas les bandes/DNA est molt ms accessible i permet que es pugui hibridar ms fcilment. Hem de tenir present que el DNA pot separar-se de la membrana i per aix lhem de fixar. Aquesta fixaci es pot fer mitjanant dos mtodes:
a. Incubaci a una temperatura adequada b. Irradiaci ultraviolada: En el cas de fer servir NylonPermet la formaci
denllaos covalents entre la banda 5. Hibridaci: A continuaci es colloca la membrana al interior duna bosa de
plstic que conte, sondes marcades que son capaces dhibridar nomes amb la regi del DNA dinteres. Actualment cal destacar que existeixen cilindres en
-
els quals es fica la membrana i que permeten que el procediment sigui ms eficient.
6. Autofotografia: Trec la membrana de lloc on sha produt la incubaci i faig una autoradiografia ja que es sondes no estaran marcades indefinidament i per tant la fotografia amb permetr analitzar el gel tranquillament
4.5 Reconeixement de la seqencia especifica Reconeixement: Lnica part del DNA que es capa de trencar la monotonia del DNA son les bases nitrogenades ja que com veurem aquestes sn capaces de formar diferents ponts dH. Ja que cada parella de bases presenta un patr de ponts dH diferent que varia en la quantitat, en la presencia de donadors o acceptors i en la distancia entre tots aquets, a ms a ms tamb tot aix tamb depenen si ens troben en el sol major o en el solc menor
A-T o Solc major:
T: Acceptor A: Un donador i un acceptor
o Solc menor T: Acceptor A: Acceptor
C-G o Solc major
C: Donador G: Dos acceptor
o Solc menor C: Acceptor G: Un donador i un acceptor
Aquets diferents patrons son els que pot reconixer el enzim. Cal destacar que tamb hi han protenes que son capaces de reconixer de forma inespecfica el DNA Disciplina: Totes aquestes interaccions sn estudiades per la disciplina de la biloga molecular interacci especifica DNA-proteina que entre daltres coses permet conixer aspectes claus de la regulaci...
4.6 Estructura de la EcoR1 i intereccions DNA-protena Estructura: Aquesta estructura es va poder resoldr a partir dun cristall que presentava la protena i un oligo (1986) que contenia la seqencia de reconeixement. Igual que qualsevol altre nucleasa aquesta protena presenta tamb una simetria rotacional binaria i per aquest motiu esta formada per dues subunitats iguals anomenades i . Aquesta protena te la capacitat de rodejar el DNA. En la imatge de lesquerra nomes es veu una subunitat
JR.Daban BIOLOGIA MOLECULAR Figura 4.20
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.20
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.26
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.26
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.26
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.26
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.26
-------------------------------------------------------------------------------------------------
-
Interaccions amb el DNA: Gracies al oligo tamb vem poder conixer quines intereccions es feien amb el DNA. Cadascuna de les subunitats realitza 6 ponts dH mitjanant 3aa i aquets sn
Arg145: Es donador de dos ponts dH amb N7 de dues A consecutives (la mateixa que la Glu de laltre subunitat)
Glu144: Es acceptor de dos ponts dH amb N6 de dues A consecutives (la mateixa que la Arg de laltre subunitat)
Arg200: Es donador de dos ponts dH amb N7 o O6 de una G del extrem de la seqencia de reconeixement.
4.7 Estructura de la metiltransferasa Estructura: La seva estructur en aquest cas es va resoldre a partir dun co-cristatll que presentava Metiltransferasa Hhal i una inhibidor que era capa de formar un enlla covalent amb el centre actiu. En aquest cas la protena no rodeja tot el DNA sin que nomes interacciona per un dels seus costat. El punt caracterstic es que gracies a la seva estructura i a les seves interaccions es capa de fer sortir la base que sha de metilar del interior del DNA i daquesta manera lapropa al centre actiu
4.8 Aplicacions de les endonucleases de restricci Les endonucleases de restricci ens han obert un gran ventall de possibilitats, entre les que hem de destacar la importncia en la creaci de DNA recombinant (enginyeria gentica, els ja esmentats mapes de restricci (mapes fsics) i tambe aplicacions clniques diverses basades en el polimorfismes dels fragments de restricci
JR.Daban BIOLOGIA MOLECULAR Figura 4.21
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.21
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.21
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.23
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.23
-------------------------------------------------------------------------------------------------
-
4.8.1 DNA recombinant Definici: Es basa en la inserci dun fragment de DNA a un vector amb diferents objectius:
Clonaci Expressi ....
Passos: 1. Obtenci del fragment dinteres: Es talla el gen que es vol amplificar o
expressar mitjanant enzims de restricci que siguin capaos de generar extrems cohesius, i al mateix temps tamb es fa un tall en el vector mitjanant la mateixa endonucleasa de restricci ja que daquesta manera els 4 extrems que obtindrem seran complementaris 2 a 2
2. Creaci del DNA recombinant: Es prepara una soluci que conte el fragment dinters, el vector i una lligasa. La lligasa permetr que suneixin els extrems i daquesta manera que sinsereixi el fragment del DNA en el vector
3. Introducci del vector: El vector normalment pot ser un plasmid i per aquest motiu moltes vegades sinternalitza per transformaci
Hem de tenir en compte que aquest nomes es un mecanisme que es pot seguir per que hi han moltes varietats
4.8.2 Obtenci de mapes de restricci Definici: Son mapes que es generen a partir de les diferents zones de tall que pot tenir un DNA Passos:
1. Marcatge dalguns dels extrems del DNA amb radioactivitat, fluorescncia... 2. Digesti: Afegim a la soluci del DNA una endonucleasa o varies per tal de que
es cren diversos fragment del DNA amb diverses longituds 3. Electroforesis: Es colloca tota la soluci dels DNA en un gel electrofortic que
en un dels carrils presentar un marcador de pes molecular (DNA tallat amb enzims ja caracteritzats) i en els altres carrils el DNA del qual farem el mapa
4. Observaci: En el DNA correran tots els DNA per nomes aquells que presentin en el seu extrem un marcatge seran els que podrem observar, daquesta manera podrem conixer els diferents fragments que es formen a les endonucleases, quina longitud tenen aquets fragments i conseqentment genera un mapa en el qual identificarem les diferents zones de tall
Comparaci mapa de restricci (fsic) amb mapa gentic: Charles Cantor va generar el mapa fsic de la soca K12 de E. Coli i posteriorment va voler identificar si hi havien moltes diferencies entre el mapa fsic i el mapa gentic. Per aquest motiu va fer diferents mapes de restricci amb diferents endonucleases de restricci que reconeixen una gran quantitat de bases i daquesta manera obtenem fragments
JR.Daban BIOLOGIA MOLECULAR Figura 4.24
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.24
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.24
-------------------------------------------------------------------------------------------------
JR.Daban BIOLOGIA MOLECULAR Figura 4.24
-------------------------------------------------------------------------------------------------
-
bastant grans entre les quals cal destacar la NOT que reconeix 8 nucletids. A continuaci es separaven els diferents fragments mitjanant una electroforesis de camp polsant ja que aquets eren molt grans i finalment shibridaven aquets gels amb diferents sondes que identificaven els diferents gens que shavien trobat en el mapa gentic. El resultat va permetre conixer quins gens contenia cada fragment de restricci i daquesta manera es va poder determinar que a grosso modo, no hi havia moltes diferencies entre el mapa gentic i el mapa de restricci tot i que si que existien unes petites desviacions.
4.8.3 Polimorfisme de la longitud dels fragments de restricci (RFLP) Definici: Depenent de la seqencia una endonucleasa una mateixa endonucleasa de restricci es capa de generar diferents fragment de DNA, amb diferents longituds entre dos DNA que son molt semblants per que presenten una variaci en els llocs de tall de lendonucleasa. Aplicacions clniques: Aquesta tcnica amb pot servir perqu si tenim molt ben caracteritzat la base gentica de lherncia o de una malaltia aquesta tcnica amb pot servir per obtenir els resultats de forma rapida i fiable. Si ens imaginem que tenim dos allels en els quals un produeix la malaltia i laltre no i sabem que aquets dos tenen una seqencia molt igual per que difereix en la seqencia de reconeixement dalguna endonucleasa quan fes us daquesta endonucleasa els fragments que es generaran per cada alles seran diferents i conseqentment podr identificar quin allel es el sa i quin es el patolgic. Tamb ens permet identificar individus de forma fiable i establir arbres geneologics ja que entre els progenitors els fragments de restricci que es crearan seran molt semblants per no dir idntics. Hem de tenir en compte que per fer totes aquestes tcniques tamb haurem de realitzar el bloting FALTA SINGLE STRAND CONFORMATION POLIMORFISM