Contracción de Los Distintos Tipos Celulares
8
Contracción de la célula cardiaca, célula del musculo liso y célula del musculo estriado esquelético Las comunicaciones intercelulares son eventos biológicos esenciales en los organismos multicelulares, asociadas con el control del crecimiento y la diferenciación celular, la apoptosis, las respuestas adaptativas de células diferenciadas y la sincronización de funciones celulares. Las uniones intercelulares, conocidas como uniones gap, estructuralmente constituidas por conexinas, tienen una participación activa en estos procesos. A nivel cardiovascular, la comunicación célula a célula es indispensable, en condiciones normales, para la embriogénesis cardíaca, la transmisión del impulso eléctrico, la sincronización de la actividad contráctil cardíaca, la transmisión de señales reflejas vasculares, entre otras funciones biológicas, mientras que en condiciones patológicas, a causa de mutaciones genéticas heredadas o adquiridas, participan en el desarrollo de cardiopatías congénitas, arritmogénesis y remodelación eléctrica cardíaca, aterosclerosis e isquemia miocárdica, hipertensión arterial y remodelación miocárdica. La contracción muscular está regulada por variaciones en los niveles citosólicos de Ca++, lo que afectan las interacciones entre las cabezas de miosina y los filamentos de actina a través de las dos proteínas accesorias asociadas a la actina en el filamento fino: tropomiosina y troponina. En el músculo en reposo la concentración citosólica de Ca++ es de 10-7 M, la miosina no puede asociarse a la actina debido a que los sitios de unión para las cabezas de miosina en la G-actina, están bloqueados por la tropomiosina. Al aumentar las concentraciones citosólicas de Ca++ a 10-5 M, la subunidad TnC de la troponina une
-
Upload
sarah-matthews -
Category
Documents
-
view
5 -
download
0
description
fisiologia
Transcript of Contracción de Los Distintos Tipos Celulares

Contracción de la célula cardiaca, célula del musculo liso y célula del musculo
estriado esquelético
Las comunicaciones intercelulares son eventos biológicos esenciales en los organismos
multicelulares, asociadas con el control del crecimiento y la diferenciación celular, la
apoptosis, las respuestas adaptativas de células diferenciadas y la sincronización de
funciones celulares. Las uniones intercelulares, conocidas como uniones gap,
estructuralmente constituidas por conexinas, tienen una participación activa en estos
procesos. A nivel cardiovascular, la comunicación célula a célula es indispensable, en
condiciones normales, para la embriogénesis cardíaca, la transmisión del impulso eléctrico,
la sincronización de la actividad contráctil cardíaca, la transmisión de señales reflejas
vasculares, entre otras funciones biológicas, mientras que en condiciones patológicas, a
causa de mutaciones genéticas heredadas o adquiridas, participan en el desarrollo de
cardiopatías congénitas, arritmogénesis y remodelación eléctrica cardíaca, aterosclerosis e
isquemia miocárdica, hipertensión arterial y remodelación miocárdica. La contracción
muscular está regulada por variaciones en los niveles citosólicos de Ca++, lo que afectan
las interacciones entre las cabezas de miosina y los filamentos de actina a través de las
dos proteínas accesorias asociadas a la actina en el filamento fino: tropomiosina y
troponina. En el músculo en reposo la concentración citosólica de Ca++ es de 10-7 M, la
miosina no puede asociarse a la actina debido a que los sitios de unión para las cabezas
de miosina en la G-actina, están bloqueados por la tropomiosina. Al aumentar las
concentraciones citosólicas de Ca++ a 10-5 M, la subunidad TnC de la troponina une Ca+
+, produciéndose un cambio conformacional en la molécula de troponina y el
desplazamiento de la molécula de tropomiosina hacia la parte más profunda de la
hendidura de la hélice de la actina. Como resultado, los sitios en la G-actina, capaces de
interactuar con las cabezas de la miosina quedan libres. Las variaciones en las
concentraciones de Ca++, se producen en respuesta a los estímulos nerviosos que
inducen la contracción muscular y que actúan desencadenando la liberación de Ca++
desde el retículo sarcoplásmico hacia el citosol. La concentración del músculo cardiaco es
debido a la presencia de Ca++ en el citosol de las células. Una vez disparado el Ca++, las
GAP hacen que se contraigan todas las células comunicadas.
La fisiología de la contracción cardíaca es la siguiente: Cuando el sarcolema
(membrana plasmática) de una célula muscular cardíaca no estimulada está polarizado,
quiere decir que se encuentra negativamente cargado con respecto al exterior. Este estado
de no estimulación de la célula muscular, conocido como potencial de reposo, se crea con

la presencia de proteínas y ácidos nucleicos con cargas negativas dentro de la célula
muscular. Un equilibrio entre iones de potasio positivo (K+) dentro de la célula e iones de
sodio positivo (Na+) fuera de la célula contribuye a la polarización. Durante un potencial de
acción, el equilibrio entre el K+ y el Na+ es alterado, de modo que la célula se despolariza.
La serie de acontecimientos que ocurre durante y seguido a un potencial de acción en las
fibras musculares contráctiles del corazón es similar al del músculo esquelético. A
continuación una descripción de estos acontecimientos fisiológicos de la contracción
muscular cardíaca:
-Una rápida despolarización tiene lugar cuando los canales de Na+ en el sarcolema se
abren y permiten la entrada de iones de Na+ hacia la célula muscular cardíaca; luego los
canales de Na+ se cierran rápidamente.
-Hay una fase de meseta durante la cual iones de calcio positivo Ca+ ingresan al citosol de
la célula muscular. Los Ca+ ingresan desde el retículo sarcoplasmático (retículo
endoplasmático) dentro de la célula, como así también desde fuera de la célula a través de
canales de Ca+ de apertura lenta en el sarcolema. Dentro de la célula, los iones de Ca+ se
adhieren a la troponina, la cual a su vez desencadena las uniones de puente que lleva al
deslizamiento de filamentos de actina cerca de los filamentos de miosina. Este
deslizamiento de filamentos provoca la contracción celular.
-Una repolarización tiene lugar cuando los canales de K+ se abren y el K+ se sale fuera de
la célula, al mismo tiempo que los canales de Ca+ se cierran. Estos acontecimientos
restaura la membrana a su polarización original, excepto que las posiciones de iones de
K+ y Na+ a cado lado del sarcolema se revierten.
-Luego sigue una fase refractaria durante la cual las concentraciones de iones K+ y Na+ se
restauran activamente a los lados correspondiente del sarcolema por medio de las bombas
de Na+/K+. La célula muscular no puede contraerse nuevamente hasta que los iones de
Na+ y K+ son restaurados a sus estados de potencial de reposo. La fase refractaria del
músculo cardíaco es dramaticamente más larga que en la del músculo esquelético. Esto
evita el tétano ocurra en el músculo cardíaco y asegura que cada contracción sea seguida
por el tiempo suficiente para permitir que las cavidades del corazón se llenen de sangre
antes de la próxima contracción.
Bases estructurales de la contracción del músculo liso
El aparato contráctil del músculo liso se contrae más lentamente que el del músculo
estriado, pero permite un acortamiento mayor de las fibras musculares lisas. El mecanismo
de contracción, en esta variedad de músculo, también se basa en el deslizamiento de los

filamentos finos sobre los filamentos gruesos. Los filamentos de actina de las fibras
musculares lisas son fáciles de detectar a nivel ultraestructural ; en cambio la visualización
de los filamentos gruesos requiere de condiciones de fijación especiales, que demuestran
que en el músculo liso por cada filamento grueso hay una proporción mucho mayor de
filamentos finos (1:14) que la que se observa en el músculo esquelético (1:6). En estas
células, la contracción es regulada también por alza en las concentraciones citosólicas de
Ca++. Sin embargo, la regulación de la contracción está asociada a miosina y no a actina.
Un alza en las concentraciones citosólicas de Ca++ induce la fosforilación de las cadenas
livianas de la miosina lo que: produce una modificación en la cola de la molécula que
permite la formación de filamentos gruesos (Fig 3) y genera un cambio conformacional en
la cabeza que permite su interacción con actina.
Los filamentos gruesos preparados in vitro, a partir de miosina de músculo liso, aparecen
polarizados en una sola dirección en un lado del filamento y en la dirección opuesta a lo
largo del otro lado (Fig 3). En esta configuración no existe una zona libre de puentes, como
la que se ve en el filamento grueso del músculo esquelético. Esta disposición tiene la
ventaja que actina y miosina pueden interactuar sin interrupción a lo largo de todo el
filamento grueso. Cuando la cabeza de la miosina se defosforila, los filamentos se
desensamblan y la miosina se disocia de la actina. La fosforilación es catalizada por una
enzima (quinasa de la cadena liviana de la miosina) cuya acción requiere de la presencia
del complejo Ca-calmodulina. El modelo aceptado de contracción de las fibras musculares
lisas (Fig 4) establece que manojos de filamentos finos de actina, asociados a filamentos
gruesos de miosina, se anclan por un extremo a cuerpos densos adheridos a la membrana
plasmática y por el otro a filamentos intermedios no contráctiles a través de cuerpos
densos citoplasmáticos. La a-actinina es uno de los componentes de los cuerpos densos.
El rol de los cuerpos densos es similar al de los discos Z de las miofibrillas del músculo
estriado. Los manojos contráctiles se orientarían oblicuos respecto del eje mayor de la
célula (Fig 5), lo que explicaría el acortamiento que experimentan las fibras musculares
lisas durante su contracción. En la superficie de las células musculares lisas existen
numerosas vesículas membranosas o cavéolas (Figs 7 y 8), vecinas a cisternas o túbulos
de retículo endoplásmico liso. Se cree que este sistema membranoso juega un papel en la
captura y liberación de calcio, similar al que desempeña el retículo sarcoplásmico en el
músculo estriado. Además de su actividad contráctil, las células musculares lisas tienen la
capacidad de sintetizar colágeno tipo III, elastina y proteoglicanos.
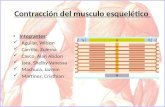
Contracción de la célula del musculo estriado esquelético

Estructura.- Cada fibra muscular está envuelta por una membrana celular eléctricamente
excitable, denominada “sarcolema”. El citoplasma recibe el nombre de “sarcoplasma”, y
contiene haces de filamentos fuertemente empaquetados. Estos filamentos se denominan
miofibrillas, compuestas por unas 20.000 unidades repetitivas llamadas “sarcómeros”.
Existe una forma especializada de retículo sarcoplásmico (RS) que envuelve a cada
miofibrilla. El RS está formado por túbulos membranosos longitudinales, los cuales se
disponen paralelos a las miofibrillas, y que finalizan en canales aplastados conocidos como
cisternas terminales. El RS contiene gran cantidad de Ca+2. Por otra parte, el sarcolema
de la fibra muscular presenta invaginaciones que forman el sistema de túbulos
transversales o túbulos-t, que se disponen perpendicularmente a las miofibrillas. Al final de
cada sarcómero hay una estructura de membranas denominada “triada”, formada por un
túbulo-t y por una estructura base. La contracción muscular esquelética se lleva a cabo por
un estímulo nervioso, que genera un potencial de acción que se extiende por el sarcolema
y a lo largo de la red de túbulos-t. La señal atraviesa la unión de la triada e induce la
liberación de Ca+2 del retículo sarcoplásmico al sarcoplasma, el cual interacciona con los
sarcómeros de la fibra muscular e induce a la contracción.
Estructura de la Fibra Muscular
CONTRACCIÓN MUSCULAR.- Para contraerse, el músculo necesita de energía en forma
de ATP. El ciclo de interacciones celulares es el siguiente:
Adhesión.- La cabeza de la miosina tiene 2 lugares de unión: uno para el ATP y otro para
la actina. Las cabezas de miosina se unen al ATP y lo descomponen en ADP+P (proceso
que requiere Mg+2). El complejo se une a la actina.
Rotación
El ADP y el P, son liberados por la miosina a la vez que las cabezas de ésta cambian de
conformación molecular: Se doblan formando un ángulo diferente y arrastrando los
filamentos delgados de actina. Esta tracción es ejercida en sentido opuesto en los dos
extremos del filamento de miosina. La región de superposición de los filamentos aumenta,
de manera que se produce más acortamiento del sarcómero.
Separación
La actina y la miosina continúan unidas formando un complejo estable hasta que una
nueva molécula de ATP se une a las cabezas de miosina. Durante la contracción muscular,
este ciclo puede repetirse entre 30 a 100 veces en un segundo. Otra manera de explicarlo
y representarlo sería mediante la siguiente forma:
Contracción Muscular

· Al inicio del ciclo, la cabeza de la miosina, que carece de un nucleótido unido, se
encuentra estrechamente unida al filamento de actina (estado I).
· La unión de ATP a la cabeza de la miosina, reduce la afinidad de la cabeza de la miosina
por la actina (estado II).
· La hidrólisis parcial del ATP (durante la cual ADP y Pi permanecen unidos a la miosina),
activa la cabeza de la miosina, la que experimenta un cambio conformacional y se
desplaza respecto del filamento fino (estado III).
· La miosina activada contacta a una molécula de actina y se une a ella produciéndose la
liberación de Pi (estado IV).
· Una vez unida a actina, la cabeza de la miosina experimenta un nuevo cambio
conformacional que se traduce en un desplazamiento del filamento fino y en la liberación
de ADP (estado V). De esta manera, cada cabeza de miosina se desplaza hacia el extremo
(+) del filamento fino adyacente. Mientras la concentración de Ca++ sea alta y exista ATP
disponible, los ciclos de formación de puentes actina-miosina continúan y el sarcómero
continúa contrayéndose.