
MICROPALEONTOLOGiA - aragosaurus.com · casos imp ide conocer las caracteristicas completas del...
21
MICROPALEONTOLOGiA Eustoquio Molina (Editor) " COLECCION TEXTOS DOCENTES
-
Upload
trinhquynh -
Category
Documents
-
view
217 -
download
0
Transcript of MICROPALEONTOLOGiA - aragosaurus.com · casos imp ide conocer las caracteristicas completas del...

MICROPALEONTOLOGiA Eustoquio Molina (Editor) "
COLECCION TEXTOS DOCENTES

Micropaleontologfa (E. Molina, ed., 2002)
19.1. Introducci6n
Capitulo 19
Algas calcareas
Jose Ignacio Canudo
365
«Algas» calcareas es un termino informal que agrupa a distintos grupos de protoctistas afines a las plantas y que tienen facilidad para calcificarse total 0
parcialmente. Este canicter hace que se encuentren f6siles con relativa frecuencia. Son un componente fundamental de los dep6sitos carbonatados del Mesozoico y del Cenozoico (fig. 19.1). Su interes es doble, por una parte como bioconstructores de arrecifes y por otra por ser uno de los componentes biocl<lsticos mas abundante en las roc as carbonatadas. En este capitulo se describen dos grupos de medios marinos: algas rojas y algas verdes, y otro de aguas continentales: las carofitas.
FIGURA 19.1. Laminas delgadas de asociaciones algales. Segun Abate et at. , en Flugel (1977) .

366 Algas calcareas (J.I. Canudo)
19.2. Aspectos generales sobre su biologfa
Las algas calcareas son protoctistas, bentonicas, y fotosinteticas (necesitan la luz del sol). Sus celulas eucariotas suelen ser verdes al contener clorofila a (las clorofitas), pero pueden estar pigmentadas de otros colores como el rojo, debido al alto contenido en ficoeritrina (las rodofitas) . El plan corporal basico es el de un tala sin vascularizar, que las diferencia de las plantas. Algunos de los caracteres usados en la clasificacion de las algas vivientes son los pigmentos (moleculas responsables de los colores) y estructuras presentes en las partes blandas, que en ocasiones no son identificables en el registro fosil. Los detalles de las estructuras vegetativas y organos reproductores son imprescindibles para la clasificacion de las algas actuales en ordenes, familias, generos y especies.
19.2.1 Calcificaci6n
La calcificacion es un proceso biologico fundamental y variable en estos organismos. En general, se realiza durante la fotosintesis y esta catalizada por la enzima rubisco. La mineralogia y el lugar don de se produce la calcificacion son determinantes en la conservacion de los caracteres morfologicos. La calcificacion puede producirse en diferentes carbonatos, pero nunca se mezclan en el mismo organismo. Las algas rojas precipitan generalmente calcita, pero varia la cantidad de magnesio presente en la estructura cristalina. Las algas verdes depositan aragonito. Los organos reproductores de las carofitas son de calcita. La mineralizacion se puede producir en la pared celular, en la parte externa de la celula 0 como deposito superficial del organismo.
La presencia de un esqueleto carbonatado no es en sf un criterio diagnostico. En la actualidad hay especies del mismo genero con y sin calcificacion. Ademas, la calcificacion puede ser parcial, produciendose exclusivamente en una de las partes del organismo. A pesar de estas dificultades, un solo fragmento calcificado suele ser suficiente para su identificacion, 10 que es especialmente util en el estudio micropaleontologico.
19.2.2 Forma del crecimiento y morfologfa externa
La forma y el tipo del crecimiento de las algas suele ser el resultado del habitat donde se desarrollan . Gran parte de la variabilidad en la forma externa de las algas calc areas se explica por la adquisicion de diferentes habitos: incrustante, erguido articulado, erguido foliar y tubular (fig. 19.2).
Las incrustaciones pueden tener la superficie lisa 0 irregular, y formar nodulos o ramas rfgidas. Su tamafio puede variar de pocos milfmetros a algunos centfmetros. Este tipo de habito se encuentra en varias familias de algas rojas. Las

Micropaleontologfa (E. Molina, ed., 2002) 367
que tienen un habito erguido articulado, como Halimeda 0 Corallina, suelen ser segmentadas y ramificadas. Dada su fragilidad, se suelen encontrar desarticuladas en elementos individuales, cuya forma externa puede variar de segmentos cilfndricos a formas mas 0 men os aplanadas de po cos milfmetros de tamano. Estos segmentos desarticulados son suficientes para la clasificaci6n, pero en muchos casos imp ide conocer las caracteristicas completas del alga productora, particularmente si carecemos de analogos actuales. Los habitos articulados estan presentes en algas rojas, verdes y en carofitas. EI habito erguido foliar se da en algas como Udotea 0 Caulerpa. EI habito tubular comun en las carofitas 0 algunas codiaceas f6siles presenta tal os de aspecto «filamentoso», compuesto por numerosos filamentos empaquetados.
FIGURA 19.2. Tipos de habitos en algas calcareas: a) erguido articulado, b) erguido foliar, c) incrustante irregular, d) incrustante nodular, e) incrustante ramificado, f) tubular. Modificado de Wray (1977).
19.2.3 Anatomia interna
La forma, tamano y la estructura interna es fundamental en la identificaci6n taxon6mica de las algas. La anatomia interna basica de las algas calcareas es filamentosa. Los diferentes tipos de empaquetamiento de estos filamentos dan lugar a diferentes tipos de talos. Los 6rganos reproductores varian de tamano, forma y distribuci6n. Pueden estar incluidos en la estructura vegetativa esqueletica,

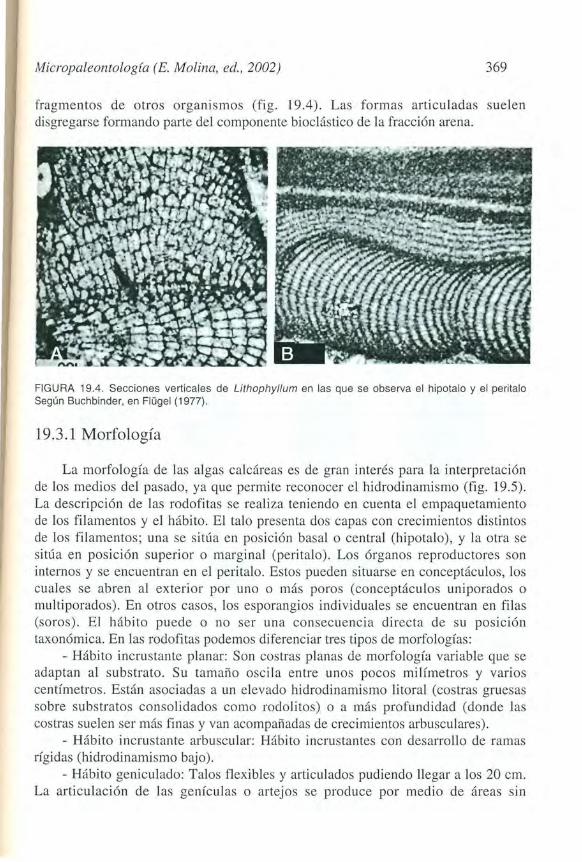
Micropaleontologia (E. Molina, ed., 2002) 369
fragmentos de otros organismos (fig. 19.4). Las formas articuladas suelen disgregarse forman do parte del componente biocl<istico de la fraccion arena.
FIGURA 19.4. Secciones verticales de Lithophyl/um en las que se observa el hipotalo y el peritalo Segun Buchbinder, en Flugel (1977).
19.3.1 Morfologfa
La morfologfa de las algas calcareas es de gran interes para la interpretacion de los medios del pasado, ya que permite reconocer el hidrodinamismo (fig. 19.5). La descripcion de las rodofitas se realiza teniendo en cuenta el empaquetamiento de los filamentos y el habito. El tala presenta dos capas con crecimientos distintos de los filamentos; una se situa en posicion basal 0 central (hipotalo), y la otra se situa en posicion superior 0 marginal (peritalo). Los organos reproductores son internos y se encuentran en el peritalo. Estos pueden situarse en conceptaculos, los cuales se abren al exterior por uno 0 mas poras (conceptaculos uniporados 0
multiporados). En otros casos, los esporangios individuales se encuentran en filas (soras). El habito puede 0 no ser una consecuencia directa de su posicion taxonomica. En las rodofitas podemos diferenciar tres tipos de morfologfas:
- Habito incrustante planar: Son costras planas de morfologfa variable que se adaptan al substrato. Su tamafio oscila entre unos pocos milfmetros y varios centfmetras. Estan asociadas a un elevado hidrodinamismo litoral (costras gruesas sobre substratos consolidados como rodolitos) 0 a mas profundidad (donde las costras suelen ser mas finas y van acompanadas de crecimientos arbusculares).
- Habito incrustante arbuscular: Habito incrustantes con desarrollo de ramas rfgidas (hidrodinamismo bajo).
- Habito geniculado: Talos flexibles y articulados pudiendo llegar a los 20 cm. La articulacion de las genfculas 0 artejos se produce por medio de areas sin


Micropaleontolog£a (E. Molina, ed., 2002) 371
19.3.2 Solenoporaceae
Esta familia, exclusivamente fosil, se caracteriza por formas incrustantes que crecen en nodulos 0 columnas. Su tamano oscila entre milimetrico y centimetrico. Su forma mas comun son masas redondeadas y hemisfericas. Su estructura interna esta compuesta por filamentos calcificados verticales y divergentes. La seccion de estos filamentos es circular 0 poligonal. Las celulas son de tamano mayor que las coralinaceas actuales. La estructura vegetativa es simple y se han identificado conceptaculos externos, en la superficie del talo.
Las solenoporaceas son algas predominantemente paleozoicas y mesozoicas, existiendo algun taxon en el Cenozoico como Parachaete tes. Su maxima abundancia y diversidad ocurre en el Paleozoico. En el Mesozoico comienzan a declinar hasta desaparecer definitivamente en el Cenozoico.
Su distribucion paleoambiental es similar a las actuales algas coralinas, pero son menos ubicuas respecto a la profundidad y a la temperatura. EI registro sedimentologico indica que Solenoporaceae ocupo paleoambientes marinos abiertos de salinidad normal.
En el Paleozoico, se las encuentra asociadas con invertebrados marinos, formando una parte importante de los carbonatos de plataforma, incluyendo los biohermos y los depositos arrecifales. En el Mesozoico ocupan nichos ecologicos diferentes a los de los cOl·ales hexacoralarios, encontrandose en facies diferentes . Las solenoporaceas del Cenozoico estan adaptadas a un mayor numero de ambientes y coinciden con hexacoralarios y coralinaceas.
19.3.3 Gymnocodiaceae
Las algas que se asignan a esta familia extinguida tienen su estructura vegetativa compacta que se diferencia bien de otras algas rojas. Su habito es erguido y ramificado. Los talos son alargados, cilfndricos con una estructura interna formada por filamentos gruesos ramificados, analoga a la de algunas algas verdes. Sin embargo, en la periferia del talo se observan conceptaculos, por 10 que actualmente se situa la familia dentro de las rodofitas.
Su registro fosil es limitado y discontinuo. Unicamente hay dos generos claramente asignados a esta familia: Gymocodium del Permico y Permocalculus del Permico, Cretacico y Paleoceno. Ambos generos han sido descritos en distintos afloramientos de Asia, Europa y Norteamerica, 10 que indica una distribucion cosmopolita. Suelen estar asociados con codiaceas y dasycladaceas y presumiblemente ocupan el mismo ambiente costero.

372 Algas calcareas (1. 1. Canudo)
19.3.4 Peyssonneliaceae
Estas rodofitas tienen un tfpico habito incrustante. Muchas de las especies ac tuales forman costras aplanadas bien cementadas a un substrato consolidado, mientras que otras son petaloideas, cubren toda una superficie y pueden presentar rizoides basales que contribuyen a la fijacion del substrato. Las formas incrustantes suelen tener un espesor que suele variar entre 0,2 y 0,5 mm, extendiendose por una superficie que puede llegar hasta los 10 cm2
. Algunos generos actuales estan mineralizados en aragonito y otros carecen de mineralizacion. EI crecimiento y la estructura interna de Squamariaceae fosiles tiene bastante parecido con algunas coralinaceas incrustantes. Internamente, el talo esta formado por filamentos celulares fuertemente empaquetados, en los que se diferencia el hipotalo y el peritalo. Las celulas presentan una importante variacion en el tamano.
Su regi stro fosil es escaso. Los generos Peyssonnelia y Polystrata se han encontrado en var ias localidades del Cenozoico. Es necesario des tacar que en lamina delgada las peisoneliaceas se suelen encontrar recristalizadas en calcita, formando costras total mente translucidas a la luz transmitida del microscopio optico. Este es el caracter principal para su reconocimiento. Las actuales Squamariaceae suelen ocupar ambientes marinos tropicales y subtropicales. Tambien son abundantes en el Mediterraneo y se han descrito especies sin calcificar en medios subarticos. Se encuentran en areas de salinidad marina normal en depositos someros que van desde la mi sma Ifnea mareal hasta unos pocos metros de profundidad; en areas tropicales como las islas Hawai han l\egado a encontrar hasta los 90 m. Son tfpicas de fondos no consolidados en el Mediterraneo.
19.3.5 Corallinaceae y Sporolithaceae
Estas so n las dos familias de rodofitas mas diversas y abundantes en el registro fosil por 10 que se van a tratar con mas detalle. Son componentes comunes en las rocas carbonatadas marinas del Cretacico y del Cenozoico.
19.3.5.1 Morfologia
Las coralinaceas presentan distintos tipos de habitos (fig. 19.5). La estructura basica anatomica de las coralinaceas son celulas distintivamente empaquetadas con las paredes celulares bien ca lcificadas. La estructura vegetativa se diferencia en tres regiones: hipotalo, peritalo y epitalo. En las formas incrustantes los filamentos del hipotalo se orientan mas 0 menos paralelamente al substrato, formando la estructura vegetati va basal. E I hi potalo basal puede desarrollar multi pIes capas 0

Micropaleontologfa (E. M olina, ed., 2002) 373
puede ser una capa unica de celul as alargadas y paralelas al substrata . E l peritalo se desarro lla encim a de l hipotalo y es ta fo rmado pa r filamentos ori entados perpendicul armente al substrata . Su grosor es muy variable, en algunos generos esta practicamente ausente a es muy fino, pero en otras forma la mayor parte del organi smo. El epitalo es una fina superficie de celul as sin calcificar, que no suele conservarse en los f6s iles . En las fo rmas e rguid as, al ex isti r un a es tru ctura vegetativa cortical y medular, los 6rganos reproductores son carac terfsticos para distinguir las coralinaceas de las esporolitaceas. Se encuentran en e l interior del peri tala incluidos en conceptaculos abiertos al exterior par una 0 mas aberturas .
GENICULADAS INCRUSTANTES
E u
o N , .,.,
1
Habito
Erguido con ramificaciones y segmentos articulados
(geniculas)
n
Estructura interna
Fragmentas de geniculas separadas
par uniones sin mineralizar (n)
Conceptaculo
p: peritalo
h: hipotalo
Habito
I+--- 1- 10 cm ~
FIGURA 19. 5. Habito y estructura interna en Corallinaceae. Modificado de Wray (1977).
19.3.S.2 C lasif icaci6 n
Los fragmentos ais lados de Corallinacea son suficientes para identi ficar gene ros f6s iles. Sin embargo, a lgun os de los generos ac tu ales no pueden identi ficarse en el registro f6s i I al estar defi ni dos can caracteres de las partes blandas. Desde el trabajo de Woelker ing (1988) , la antigua subdi visi6n de la famili a Corallinaceae en Melobi soidae y Corallinoidae a partir del habito ya no se util iza. Este autor di sti ngui 6 siete subfamilias, sabre todo a partir de caracteres de los 6rganos reproductores, que a veces son diffc il es de reconocer en el registro f6s il. Aspectos interesantes sabre la evo luci6n de las esporolitaceas y coralinaceas

374 Algas calcareas (J.I. Canudo)
se pueden consultar en Aguirre et al. (2000). Desde el punto de vista paleontol6gico se pueden distinguir dos grandes famiIias:
1) Sporolithaceae: Familia con tal os incrustantes y 6rganos reproductores en forma de soros, considerados primitivos. EI unico genero actual es Sporolithon.
2) Corallinaceae: Incluye todas las otras formas tanto incrustantes como geniculadas. De cara al registro f6sil se puede destacar las siguientes subdivisiones:
Subfamilia Melobesioidae, con conceptaculos multiporados (Lithothamnion, Mesophyllum y Melobesia)
Subfamilias Mastophoroidae y Lithophylloidae, con conceptaculos uniporados (Neogoniolithon, Lithophyllum, Titanoderma entre otros).
Subfamili as Corallinoidae y Amphiroidae, ambas geniculadas, que se diferencian por el proceso de formaci6n de las geniculas. Los representantes de Corallinoidae tiene genfculas primarias mientras que Amphiroidae tiene genfculas secundarias, formadas por descalcificaci6n de filamentos en la zona de intergenfcula. Este caracter es diffcil de reconocer en el registro f6sil.
19.3.5.3 Distribuci6n paleobiogeogrMica
Las coralinaceas actuales son cosmopolitas, exclusivamente marinas . Su distribuci6n va desde el tr6pico al polo y se extiende por toda la plataforma. La temperatura es el principal factor de distribuci6n geogrMica de los taxones. Hay generos de aguas tropicales como Lithoporella y de agua frla como Lithothamniom. Los taxones de agua frfa tam bien se encuentran en latitudes medias pero a mayores profundidades que en latitudes mas altas. Los generos articulados viven en aguas tropicales y templadas, siendo abundantes en los medios someros de alta energfa con menos de 10 m de profundidad.
La luz es el principal factor que controla la distribuci6n en profundidad de las coralinaceas incrustantes. Cada genero vive a una profundidad determinada relacionada con sus necesidades de luz. Las coralinaceas viven en aguas de salinidad normal. Su distribuci6n tambien esta controlada por el substrato y la energia de las olas. La preferencia por el tipo de substrato varia con los generos; muchos taxones necesitan un substrato duro, pero tambien pueden vivir en blandos aprovechandose de fragmentos donde fijarse. Respecto a la energia, sus requerimientos son variables, desde formas que necesitan vivir en medios agitados, hasta los que se desarrollan en regfmenes de baja energfa. Las coralinaceas son uno de los mas importantes biocronstructores de arrecifes del Cenozoico. En algunos arrecifes actuales IIegan a formar el 40% del edificio arrecifal. Los cOt·ales forman la mayor parte del volumen, pero las coralinaceas son el principal agente responsable de cohesi6n del edificio coralino.
Los primeros representantes de CoraIIinaceae aparecen progresivamente en el Jurasico, existiendo al menos siete generos al final del Cretacico. A 10 largo del


376 Algas calcareas (J.l. Canudo)
que una parte significativa del barro carbonatado de tamafio fino en los sedimentos proviene precisamente de su destruccion. Las algas verdes son abundantes en sedimentos detriticos de medios relativamente someros 0 en lagunas costeras, siendo raras a mayor profundidad. Las que tienen un mayor interes en Micropaleontologia son las halimedaceas, udoteaceas y las dasicladaceas.
19.4.1 Halimedaceas y udoteaceas
Las halimedaceas y las udoteaceas suelen ser erguidas y estan sujetas al fondo por rizoides . Su morfologia extern a varia desde talos compactos, como en Halimeda, a formas delicadamente ramificadas y poco calcificadas, como en Udotea 0 Caulerpa. Su estructura interna se caracteriza por una region central (medula) formada por filamentos gruesos entrelazados de los que parten filamentos finos ramificados. Estos ultimos forman una capa cortical, a menudo mejor calcificada. La forma y las ramificaciones de estos filamentos son caracteres diagnosticos para su clasificacion taxonomica.
Las generos fosiles de conocidos las halimedaceas y udoteaceas solo tienen habito erguido y se clasifican par el tipo de crecimiento (si es segmentado a no) , las variaciones internas en el tamafio, la forma y la distribucion de los filamentos medulares y corticales. Tambien es importante la forma y desarrollo de los organos reproductores. Se han encontrado codiaceas en todo el Fanerozoico, can excepci6n del Cambrico Inferior. Son un componente minoritario, aunque bastante comun de las asociaciones marinas someras de aguas tropicales, sabre todo en lagunas costeras y en el frente del arrecife. Se diversifican en el Carbonffero Superior, Permico, Cretacico Superior y Cenozoico. La mayorfa de las halimedaceas y udoteaceas actuales son tropicales, habiendo pocas especies subtropicales. Suelen colonizar substratos arenosos y arcillosos, donde sus rizomas pueden enraizar. En cuanto a la luz, suelen vivir a poca profundidad, especialmente en medios tropicales.
19.4.2 Dasicladaceas
Son algas erguidas, segmentadas y ramificadas de varios centfmetros de altura, formados par un unico citoplasma gigante (megacelula). Se anclan al substrata por medio de rizoides. De manera caracterfstica, el tala tiene una simetrfa radial segun un eje central alargado, a partir del cual se desarrollan ramificaciones laterales. Los organos reproductores se encuentran en ramificaciones a partir del eje central.
En la clasificacion de las dasicladaceas se utiliza la forma del tala, sus dimensiones, segmentacion del tala 0 esqueleto, arranque de las ramificaciones, forma de los extremos de las ramificaciones, forma del sistema central y forma de

Micropaleontologfa (E. Molina, ed., 2002) 377
los 6rganos reproductivos. Las dasicladaceas tienen una distribuci6n similar a las halimedaceas y udoteaceas en la actualidad. Son un componente minoritario respecto al resto de la fauna y flora marina. El gran numero de generos f6siles de las dasicladaceas respecto a las halimedaceas y udoteaceas parece indicar que en el pasado eran un grupo mas diversificado y abundante.
CHLOROPHYTA CODIACEAE DASYCLADACEAE
FIGURA 19. 7. Distribuci6n de algunos generos sign ificat ivos de Chlorophyta. Modificado de Wray (1978).
Las dasicladaceas suelen V1VIr en los primeros metros de profundidad; excepcional mente se han ci tado a mayor profundidad. Muchas dasicladaceas colonizan directamente el fondo arenoso, pero tambien se fijan a objetos como por

378 Algas calcareas (I.!. Canudo)
ejemplo conchas de moluscos. Son tipicas de medios de poca energia, como lagunas costeras situ ados por debajo del nivel de las olas, prefiriendo salinidades normales. Las dasicladaceas actuales viven en medios que varian desde tropicales a tempI ados, estando ausentes en aguas frias.
Su registro fosil es am plio y diverso. Se conocen desde el Cambrico Inferior hasta la actualidad . Dentro del Paleozoico, son especialmente abundantes en el Carbonifero y en el Permico. La gran cantidad de especies descritas en el Mesozoico indica que es el periodo de mayor diversificaci6n de las dasicladaceas, 10 que ha permitido proponer biozonaciones locales para el Mesozoico en facies someras del Tetis.
19.5. Charophyta
En los sedimentos continentales lacustres suelen encontrarse 6rganos reproductores (oogonios) de carofitas . Este grupo de algas se habia incluido tradicionalmente en Chlorophyta. Sin embargo, su particular morfologia, sobre todo en 10 relacionado con su reproduccion, ha permitido reconsiderarla como una division diferente situada en la linea evolutiva de las plantas vasculares. Teniendo en cuenta que comparten un antepasado comun con las clorofitas , puede considerarse como una Ifnea evolutiva de «algas» verdes que ha colonizado eficazmente los medios acuaticos continentales.
Los organos reproductores femeninos (oogonios) suelen estar calcificados y suelen tener un tamano que oscila entre 100 {Lm y 2 mm. Tambien pueden encontrarse restos vegetati vos, pero tienen menor valor sistematico. La sistematica de las carofitas fosiles se realiza en funci6n de los oogonios . Las carofitas tienen una gran importancia en la dataci6n de los sedimentos continentales, siendo en muchos casos el unico microfosil que se puede utilizar para datar estas facies .
19.5.l. Estructura vegetati va de las carofitas actuales
La estructura vegetativa de las carofitas tiene un tamano que oscila entre algunos centfmetros y un metro. Se puede dividir en dos partes: el rizoide y el talo. EI tala esta sujeto al substrato por el rizoide. Puede tener engrosamientos Ilamados bulbos amflicos que son diagn6sticos en la sistematica. El talo es ramificado y erguido. Su estructura se organiza en nudos (formado por celulas nod ales) y entrenudos (formados por celulas internodales). Esta disposici6n permite tener gran flexibilidad a la estructura vegetativa. Los nudos son cortos y pluricelulares con celulas globulares dispuestas en un disco central rodeado por celulas perifericas. Los entrenudos estan formados por una gran celula conica alat"gada (fig.19.8).

Micropaleontologfa (E. Molina, ed., 2002) 379
Los verti ci los 0 f iloides parten de los nudos. Su numero depende de los taxones y suel e oscilar entre se is y diez. Estos filoides es tan compuestos a su vez por nudos y entrenudos distribuidos a una longitud determinada. En estos nudos se desarrolla un verticilo de bracteas uni celul ares.
n
\
\ A
- I ' B , '
b
FIGURA 19.8. Estructura vegetativa y reproductora de una carofita actual. A) esquema general del talo. b: bulbos amflicos , r: rizoide, n: nudos, i: talo, w. verticilos . B) Detalle de un fil oide y la inserci6n de los organos reproductores. a: anteriodi6foro, br: bracteas, 0: oogonio, co: cor6nula, ic: celula internodal, ct: cel ulas corticantes. Modificado de Martfn -Closas (2000).
19.5.2. Estructura reproductora de las carofitas actuales
Las carofitas actuales utili zan presentan tanto reproducci6n sex ual , como asex ual. La segunda es menos habitual, aunque puede darse de manera comun en algunas especies. Hay especies con ambos sexos situ ados en el mismo tala (monoicas) y las que tienen los sexos separados en talos di stintos (dioicas).
EI gametangio femenino se ll ama oosporangio (oogon ios). E I oosporangio en las espec ies actu ales esta fo rmado por un a oospora envuelta por una membran a organica resistente, la cual esta rodeada por celul as espirales. Estas distintivas celulas se enroll an dextrorsamente (sentido horario) desde la base hasta el ap ice. Cerca 0 en la base del oosporangio se encuentra la placa basal. Es ta estructura esta com puesta por una 0 tres celulas esteril es, hermana~ de la oospora y formadas como resultado de las divisiones que preceden a la genes is de l gameto femenino. En el fina l di sta l del oosporangio se encuentra la cor6nula. E I poro apica l s610

380 Algas calcareas (i.!. Canudo)
existe en las formas f6siles y se encuentra en el apice del girogonito, por debajo de la posici6n ocupada por la cor6nula.
Los oosporangios de las carofitas suelen calcificarse formando los denominados girogonitos; se diferencian facilmente en el registro f6sil por su forma elfptica y el enrollamiento de las celulas espirales . Los oogonios actuales presentan una marcada homomorffa, a pesar de pertenecer a diferentes familias . Los girogonitos f6siles presentan una mayor diversidad morfol6gica. Tienen diferentes sentidos de enrollamiento y presentan estructuras complejas ausentes en girogonitos actuales. EI girogonito puede estar desnudo 0 rodeado por una cubierta suplementaria llamada utrfculo, que es una especie de capsula que rodea el girogonito de algunas carofitas. En algunas c1avatoraceas, el utrfculo calcifica, por 10 que sus oogonios f6siles no muestran el girogonito espiral desnudo y tienen una morfologfa irregular.
EI anteriodi6foro (anteridio) es el gametangio masculino. Su morfologfa es variable en las especies actuales. Por ejemplo en Chara es una estructura globular formada poria uni6n de placas llamadas escudos . Los gametos masculinos 0
anterozoides son celulas flageladas que se forman en su interior. La ausencia de calcificaci6n en el anteriodi6foro dificulta su presencia en el registro f6sil.
19.5.3. Distribuci6n actual de las carofitas
Las carofitas son organismos tfpicamente lacustres y palustres. Algunas especies han colonizado algunos medios transicionales, como son deltas , albuferas, sistemas salobres de litoral 0 marinos de baja salinidad (Mar Baltico). La distribuci6n de las carofitas actuales se encuentra limitada por tres factores: las sales disueltas, la batimetrfa del lago y el grado de eutrofizaci6n de las aguas, a los que se podrfa anadir un cuarto factor relacionado con el tipo de sexualidad. a) Sales disueltas: Las especies actuales de carofitas se distribuyen segun el tipo y concentraci6n de sales disueltas (cloruros y sulfatos, principalmente) . Asf, por ejemplo, las especies de Chara son mas abundantes en aguas alcalinas. Por el contrario, Lamprothamnium papulosum es exclusiva de aguas saladas con variaciones en la concentraci6n salina, como pueden ser las lagunas esteparias. b) Batimetrfa dellago: Las carofitas son organismos pioneros en la colonizaci6n de medios recien inundados 0 effmeros. Su presencia es un buen indicador de una transgresi6n lacustre. Hay taxones adaptados a una profundidad determinada, como Nitellopsis obtusa que vive restringida a las zonas profundas de los lagos 0 de lagunas de inundaci6n permanentes. c) Grado de eutrofizaci6n: Las carofitas tienen su maximo desarrollo en lagos oligotr6ficos, siendo esc as as raras 0 ausentes en los medios eutrofizados. d) Factor sexual: Las especies monoicas suelen ser cosmopolitas; sin embargo, las dioicas suelen tener una distribuci6n mas restringida, presentando provincialismo. Las especies monoicas pueden reproducirse sexualmente des de el momento que

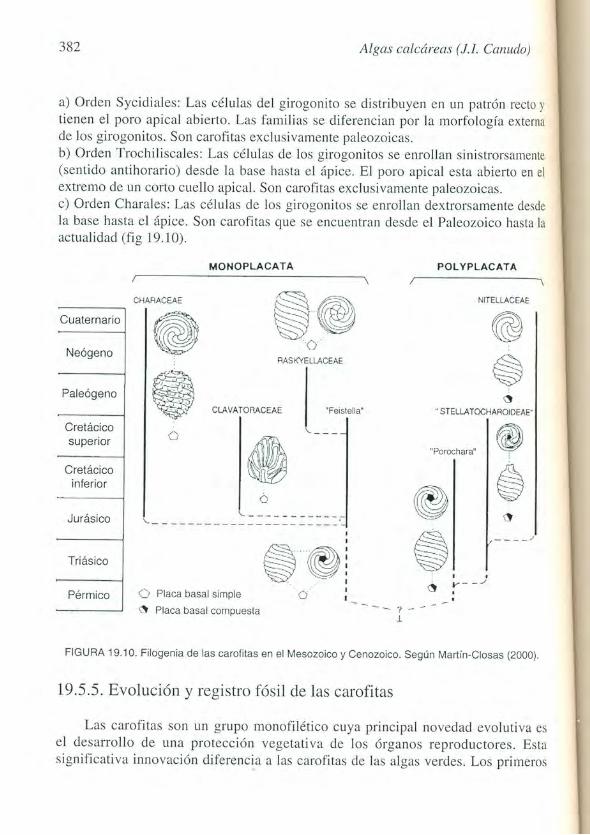

Micropaleontologfa (E. Molina, ed., 2002) 383
f6siles del Silurico poseen la tipica organizacion de nudos con verticilos y entrenudos; sus girogonitos estan calcificados y tienen ceIulas rectas 0 enrolladas. El desarrollo ontogenetico de las formas actuales parece indicar que el caracter derivado es la presencia de celulas enrolladas. En la base del Devonico, se produce una importante diversificacion; los girogonitos desarrollan estructuras novedosas que luego no van a vol verse a repetir en el resto de la historia evolutiva del grupo.
La primera extincion importante en las carofitas se produjo en el Ifmite Devonico/Carbonifero. Desaparecieron los grupos con girogonitos de celulas rectas, asi como algunos sinistrorsos. A partir del Carbonifero solo se encuentran girogonitos enrollados, y comienzan a diversificarse los girogonitos dextrorsos. La segunda importante extincion coincide con la crisis global del Ifmite Permico/Triasico, desapareciendo los girogonitos sinistrorsos. Al comienzo del Mesozoico se produce la desaparicion de las estructuras de los girogonitos desarrolladas por las carofitas durante el Paleozoico, sobreviviendo unicamente las carofitas con girogonitos de cinco celulas. Todas las carofitas post-paleozoic as se desarrollaron a partir de este grupo, por 10 que todas presentan un girogonito con cinco celulas espirales y el apice plano con el poro abierto.
Los girogonitos del Triasico y del Jurasico Inferior tienen poca variabilidad morfologica. A partir del Jurasico Superior se produce una nueva diversificacion, con la aparicion de las primeras caraceas y clavatoraceas. Las clavatoraceas tienen una espectacular radiacion global en el Cretacico Inferior, tanto en abundancia como en diversidad. Esta diversificacion coincide con el desarrollo de las facies continentales y de transicion del Jurasico Superior-Cretacico Inferior (facies «Purbeck» y «Weald») en Europa.
La diversidad de las clavatoraceas disminuye a partir del Aptiense. Hubo dos factores que afectaron a este grupo: la diversificacion de las angiospermas acuaticas, con las que entraban en directa competencia, y la sustitucion de los ambientes lacustres continentales (facies Weald) por los ambientes marinos de plataforma (facies «Urgon») y los ambientes deltaicos eutroficos (Formaciones Escucha y Utrillas en la Cordillera Iberica) . EI registro fosil de las carofftas es escaso durante el Cenomaniense y Santoniense. En este intervalo temporal hay escaso registro continental porque coincide con el maximo eustatico mesozoico. En el Campaniense y el Maastrichtiense se produce la radiacion de Characeae, y la definitiva extincion de Clavatoraceae.
Durante el Cenozoico, los representantes de Characeae dominan las asociaciones de carofitas . Se pueden diferenciar cuatro eta pas principales en la evolucion de esta familia. Durante el Paleoceno las especies se van relevando sucesivamente. La mayor diversificacion del grupo se produce en el Eoceno y se traduce en girogonitos fuertemente calcificados y ornamentados. En el transito Eoceno-Oligoceno se produce una importante extincion. La ultima etapa comienza en el Oligoceno y llega hasta la actualidad. Durante este periodo el numero de

384 Algas calcareas (1.1. Canudo)
espec les ha ido di sminuyendo progres ivamente y se ha ido incrementa ndo la homomorffa de los g irogonitos, aspecto que se observa en las asociaciones actua les.
19.5.6. Bioestratigraffa con carofitas
Su rel a tiva nipida evo lu c io n, s us morfo log fas bi en diferenciadas y su abu ndanc ia, hace de este g rupo un a excele nte herram ienta para la bioestratigraffa de sedimentos continenta les en a lgunos inte rva los temporal es. Las biozonaciones mas inte resantes coinciden con los momentos de maxima diversificacion , como en e l Cretacico Inferior, e l Cretacico Superior y e l Paleogeno. La biozonacion con carofitas de l Cretacico Infe rior es la de mayor importancia para la bioestratigrafIa de medios continentales en la Penfnsula Ibe ri ca. Las c1avatoraceas son abundantes en las facies «Purbeck» y «Weald» de la Cordillera Iberica, 10 que ha permitido realizar una detail ada biozonacion . Las biozonaciones del Cretacico Superior y Terciario se han definido a partir de las asoc iaciones de las fam ilias Raskye llaceae y Characeae. Las ultim as especies de Clavatoraceae tienen interes en la biozonacion del fina l del C retacico.
Bibliograffa
AGUIRRE, J., RlDING, R. Y BRAGA, J. C. 2000. Divers ity of cora lline red algae: or ig inati o n and ex tinct ion patterns from the Ea rl y Cretaceous to the Pleistocene. Palaeobiology, 26, 65 1-667.
BARATTOLO, F., DE CASTRO, P. Y PARENTE, M. eds. 1993. Studies on Fossil Benthic algae . Mucch i. 1-420.
FLUGEL, E. ed., 1977. Fossil Algae. Recent results and developments. Springer-Verlag, 1-375 .
MARTIN-CLOSAS, C. 2000. Els caroj/ts del luritssic superior i el Cretaci inferior de la Penlnsula Iberica. Institut d'estud is catalans, 1-304.
RIDING, R. ed . 1991. Calcareous algae and Stromatolites. Springer-Verlag, 1-571. TAYLOR, T . N. y TAYLOR, E . L. 1993. The biology and evolut ion of fossil plants.
Prentice Ha ll , 1-982. WOELKERLING , W . J. 1988. Th e Coralline Red Algae: An analysis of the genera and
subfamilies of Nongeniculate Corall inaceae. British Museum (Natural Hi story), Oxford Un ive rs ity Press, 1-268.
WRAY, J. L. 1977. Calcareous algae. Elsev ier Scientifi c Publish ing Company, 1- 185. WRA Y, J. L. 1978. Ca lcareous Algae. En: Introduct ion to Marin e Micropaleontology
(B.U. Haq y A. Boersma, eds.), Elsevier, 171-1 87.


















