







Idiomas
Páginas
Jurídico


MOLUSKUEN LISERI-GURUINEKO EPITELIOAREN BERRIZTAPEN ZELULARRA ETA BERE INPLIKAZIOA INGURUMEN-OSASUNAREN EBALUAKETAN
Jakintza-arloa: Natur Zientziak
Egilea: BEÑAT ZALDIBAR ARANBURU Urtea: 2006 Zuzendaria: JUAN ANTONIO MARIGÓMEZ ALLENDE Unibertsitatea: UPV/EHU ISBN: 978-84-8438-190-7

Hitzaurrea Dagoeneko ia bi urte igaro dira tesia aurkeztu nuenetik eta denbora tarte horretan argiak diren zenbait aldaketa nabarmentzen dira. Tesiaren zati teorikoan aipatzen den moduan bi ildo nagusi jorratu dira. Lehendabizi moluskuen eta nagusiki muskuiluen liseri-guruineko zelulen berriztapenaren azterketa burutzen da. Horretarako, gaur egun indarrean dauden zenbait teknika immunohistokimiko erabili dira. Ohikoak eta aski ezagunak ugaztunetan baina zenbait kasutan oraindik ere ornogabeetan guztiz hedatu gabe daudenak (antigorputz espezifikoen faltagatik edota ehunen ezagutza oraindik ere oso sakona ez delako). Bigarren ildoa, moluskuen liseriepitelioan gerta daitezkeen aldaketak ingurumen-osasunaren ebaluaketan erabiltzen diren biomarkatzaileetan nolako eragina duten ikertzea izan da. Biomarkatzaileak antolakuntza maila biologiko sinpleenetan (maila zelular, molekular zein tisularrean) gertatzen diren aldaketak dira eta antolakuntza maila biologiko konplexuagoetan (populazio edo ekosistema) sortaraz daitezkeen alterazioak aurresateko lagungarriak izan daitezke. Ahalik eta erantzun goiztiarragoen bilaketa dela eta, azkenengo urteotan biologia molekularreko tresnak gero eta gehiago nabarmenduz joan dira eta “omics” bezala ezagutzen diren jarduerak (genomics, proteomics, metabonomics…) gero eta pisu zein garrantzi gehiago hartuz joan dira. Egungo biologiaren tendentzia ikusita, zalantzarik gabe gaur berriz ere nire tesiarekin hasiko banintz, tresna molekularren erabilpenak pisu gehiago izango luke. Hau da, zelulen proliferazioa aztertzeko immunohistokimikak egiteaz gain, hau da, proteina jakin bat ehunean lokalizatzeaz gain, proteina hori kodetzen duen RNA mezularia lokalizatzen saiatuko nintzen in situ hibridazioak eginez. Eta, biomarkatzaileen atalari dagokionez, zelula- eta ehun-mailako biomarktzaileak erabiltzeaz gain, ziur aski, maila molekularreko biomarkatzaileren bat ere (gene ezagun baten adierazpen-maila, adibidez) erabiliko nuke. Dena den, ugaztunetan aski garatuak dauden teknologia hauek, ornogabeetan, aldiz, oraindik lehendabiziko urratsak betetzen ari dituzte. Aurretik aipatutako guztia egia bada ere, 2006. urtean Valery Forbes ikertzaileak argitaratutako artikulu batean, gaur egungo biomarkatzaileen erabilpena eta tendentzia (aipatutako “omics” teknologia) zalantzan jartzen ditu. Bertan, biomarkatzaileak, kutsatzaileak nolako mekanismo zelular eta molekularren bitartez eragiten duten ezagutzeko baliogarriak direla esaten du, baina bere izaera prediktiboaren gainean hodei ilun bat ezartzen du. Hori dela eta, ingurumen toxikologian lan egiten dugunoi, epe luzeko esperimentu konplexuak egitera bultzatzen gaitu, beraz, ez dirudi hurrengo urteotan lan falta izango dugunik. Aldaketa klimatikoa ahaztu gabe, noski, baina hori beste historia bat da…

ZOOLOGIA ETA ANIMALI ZELULEN BIOLOGIA SAILA
DEPARTAMENTO DE ZOOLOGÍA Y BIOLOGÍA CELULAR ANIMAL
MOLUSKUEN LISERI-GURUINEKO EPITELIOAREN BERRIZTAPEN
ZELULARRA ETA BERE INPLIKAZIOA INGURUMEN-OSASUNAREN EBALUAKETAN
Beñat Zaldibar AranburuBiologian Doktore Gradua lortzeko aurkeztutako memoria
Leioa 2006ko Otsaila


Ikerlan honek hurrengo laguntzak izan ditu:
Euskal Herriko Unibertsitatea (240110) proiektua (1999-2000) etaIkerkuntza-talde kontsolidatuei emandako laguntzak (2001. tik aurrera)
Eusko Jaurlaritza (PI-1999-23) proiektua (2001-2002) eta ETORTEK-BERRILUR proiektua (2004-2005)


ZOOLOGIA ETA ANIMALI ZELULEN BIOLOGIA SAILA
DEPARTAMENTO DE ZOOLOGÍA Y BIOLOGÍA CELULAR ANIMAL
MOLUSKUEN LISERI-GURUINEKO
EPITELIOAREN BERRIZTAPEN
ZELULARRA ETA BERE INPLIKAZIOA
INGURUMEN-OSASUNAREN
EBALUAKETAN
Beñat Zaldibar AranburuBiologian Doktore Gradua lortzeko aurkeztutako memoria
Leioa 2006ko Otsaila

I.- ZATI TEORIKOA.............................................................................................................1
1.- Moluskuen liseri-guruina....................................................................................5
1.1.- Bibalbioak (Mytilus eredua).................................................................5
1.2.- Gastropodo itsastarrak (Littorina eredua).........................................14
1.3.- Lehorreko gastropodoak (Arion eredua)...........................................171.4.- Liseri-guruinaren plastikotasuna.......................................................21
1.4.1.- Epitelioaren aldaketa morfologikoak..................................211.4.2.- Zelula-moten ordezkapena................................................221.4.3.- Liseri-guruineko epitelioko zelulen plastizitatea.................23
2.- Epitelio-ehunen berriztapena...........................................................................25
2.1.- Epitelioetako zelula amak.................................................................26
2.2.- Zelulen proliferazioa aztertzeko teknikak..........................................272.2.1.- Fluxu-zitometria..................................................................282.2.2.- Nukleotidoen analogoen erabileran oinarrituriko teknikak..........................................................................................28
3H-timidinaren bitartezko markaketa....................28Bromodeoxiuridina bitartezko markaketa..............29
2.2.3.- Antigeno nuklearren immunohistokimikan oinarrituriko teknikak..........................................................................................31
Ki67 antigenoaren immunohistokimika.................31PCNA antigenoaren immunohistokimika...............32Ziklinen menpeko kinasen immunohistokimika.....33Ziklinen immunohistokimika..................................34
2.2.4.- Geneen adierazpenaren azterketan oinarrituriko teknikak..........................................................................................34
2.3.- Zelula amen bitartez berriztaturiko epitelioak...................................372.3.1.- Ugaztunen epidermisa.......................................................372.3.2.- Ugaztunen hestea..............................................................402.3.3.- Ugatz-guruina.....................................................................422.3.4.- Krustazeoen hepatoarea....................................................45
2.4.- Desberdintzatutako zelulen bitartez berriztatzen diren epitelioak.....472.4.1.- Area....................................................................................472.4.2.- Listu-guruinak.....................................................................492.4.3.- Ugaztunen gibela...............................................................512.4.4.- Moluskuen liseri-guruina....................................................52

II.- AUZIAREN EGOERA, HIPOTESIA ETA HELBURUAK.............................................69
III.- EMAITZAK ETA EZTABAIDA.....................................................................................75
1.- Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean...............................................................................................................77
2.- Epitelio-zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean eta urdailean........................................................................................103
3.- Zelula epitelialen berriztapena muskuiluen liseri-guruin eta urdailean: sasoi, adin eta erregimen marealari lotutako aldamoldeak............................................119
4.- Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.................................................................139
5.- Liseri-zelulen tberriztapena kadmio eta kerosenozko nahasketa baten pean esperimentalki jarritako bareen liseri-guruineko epitelioan.................................163
6.- Zelula-moten ordezkapena bareen liseri-guruinean eta bere eragina biomarkatzaileen gainean: metalez kutsatutako eta kutsatu gabeko tokien artean lekuzaldatze-esperimentuak ...............................................................................187
IV.- ONDORIOAK ETA TESIA.........................................................................................217

Lan honetan, estuki erlazionaturiko bi ildo nagusi jorratu dira. Lehenengoanmolusku desberdinen liseri-epitelioko zelulen berriztapena nola burutzen den, eta zelula-mota desberdinen kopurua eta proportzioa nola eta zein faktoreen arabera aldatzen direnaztertu da. Bigarrenean, moluskuen liseri-epitelioaren osaketa zelularrean zenbait faktorekbultzatutako aldaketek, ingurumen-toxikologian erabiltzen diren parametroen gaineannorainoko eragina duten ikertu egin da. Bi ildo horiek, memoria honetan emaitza etaondorioak aurkeztu dituzteneko 6 kapituluetan behin eta berriro uztartuko dira, jorratutakogaiak honako hauek izanik:
1) Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean.
2) Epitelio-zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean eta urdailean.
3) Zelula epitelialen berriztapena muskuiluen liseri-guruin eta urdailean: sasoi,adin eta erregimen marealari lotutako aldamoldeak.
4) Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littoreamolusku itsastarraren liseri-guruinean.
5) Liseri-zelulen berriztapena kadmio eta kerosenozko nahasketa baten peanesperimentalki jarritako bareen liseri-guruineko epitelioan.
6) Zelula-moten ordezkapena bareen liseri-guruinean eta bere eragina biomarkatzaileen gainean: metalez kutsatutako eta kutsatu gabeko tokien arteanlekuzaldatze-esperimentuak.
Zati teorikoa, bere aldetik, bi atal nagusitan antolatuta dago. Lehenengo atalean,ikerlan honetan erabilitako material biologikoa aurkeztu egin da, Mytilus galloprovincialismuskuilu, Littorina littorea magurio eta Arion ater barearen liseri-guruinen biologia zelulareta tisularrari buruz dakizkigun oinarrizko kontzeptuak azalduz, hain zuzen ere. Halaber,beharrezkoa iruditu zaigunean, beste moluskuen espezieen ezaugarriak ere aintzat hartudira, moluskuen liseri-guruinaren azalpen integratu eta konparatua osatzeko asmoz. Orohar, liseri-guruinaren funtzioak eta bere osagai zelularren deskribapena eta funtzioakazaldu dira. Gainera, moluskuen liseri-epitelioa plastikotasun handikoa denez, zelula-moten osaketa ingurumen-faktoreen eraginaren arabera nola eta noraino alda daitekeenadierazi dugu. Halaber, zelula-moten artean nagusiak ohi diren liseri-zelulen lisosomenedukia eta egitura ingurumen-faktoreen arabera ere alda daitekeenez, eta aldaketa horiekingurumen-toxikologian biomarkatzaile gisa erabiltzen direnez, zelula-moten osaketarenaldaketek biomarkatzaileen gainean eduki dezaketen eraginaz ere aritu gara atal honenazken parrafoetan.
Zati Teorikoa
3

Bigarren atalean, epitelio-ehunetan zelulen berriztapena nola eman daitekeenazaltzen saiatu gara, oso laburki izan bada ere. Arestian, zelula amei buruzko zenbaitoinarrizko orokortasunak adierazi dira. Gero, zelulen proliferazioa detektatzeko gaur arteerabilitako metodo ezagunenak azaldu dira. Azkenik, epitelioen berriztapenerakomekanismoen barianteak azaldu dira, hoberen ezagutzen diren zenbait epitelioren kasuendeskribapena emanez. Barianteak, duten estrategia orokorraren arabera bi taldetansailkatu dira: (a) zelula amen bitartez berriztatzen diren epitelioak, eta (b) zelula amenpartehartze zuzena ez duten epitelioak.
Zati teorikoaren ostean, ikerlan honen hipotesia eta helburuak azaldu dira; emaitzeiburuzko 6 kapituluen ostean, tesia eta ondorioak. Bukatzeko, memoria honen azken atala,erabilitako metodo, prozedura eta protokolo nagusiak zehazten dituen Metodologi-Eranskina dugu.
Zati Teorikoa
4

1.- MOLUSKUEN LISERI-GURUINA
Moluskuen liseri-guruina heste ertaineko egitura da, eta elikagaiak xurgatu egitendireneko adarkatutako dibertikuluz osatuta dago. Nahiz eta molusku talde desberdinenartean aldakortasun handia egon, eskuarki organo handia da oso. Dibertikuluetanliseriketa intrazelularra gauzatzen da. Halaber, liseriketa estrazelularrean parte hartzenduten entzima batzuen sintesia ere bertan burutzen da. Oro har, elikagaien liseriketaz gainbestelako hainbat funtzio betetzen ditu, organismoaren eraenketa metabolikoa etahomeostasiaren arduradun nagusia izatea, besteak beste.
Liseri-guruinak, ikerketa esparru desberdinetan interes handia piztu du. Izan ere,animali talde desberdinen liseri-traktuaren eboluzio orokorra ulertu ahal izateko gako-organoa da. Alde batetik, liseriketa intrazelular (berezkoa izaki primitiboetan) etaestrazelularraren (berezkoa izaki eboluzionatuetan) arteko bidegurutze ebolutiboa da.Bestalde, liseri-traktu ertainari asoziaturiko guruin primitiboenetakoa dugu.
Ikerlan honetan zehar erabilitako hiru molusku espezietan liseri-guruinarenantolakuntza morfofuntzionalean hiru eredu desberdin bereiz ditzakegu, zehaztasungehiagoz banan-banan jarraian azalduko direnak.
1.1.- Bibalbioak (Mytilus eredua)
Mytilus galloprovincialis muskuilua bibalbio janari-iragazlea dugu. Iragazibeharreko partikulak, mukiaren bitartez bildu eta ezpai-palpoetarantz, eta ondorenahorantz, garraiatzen dira. Gerora, hestegorri laburra igaro ondoren, iragazitako partikulakkonplexua den urdailera heltzen dira (Salvini-Plawen, 1988). Behin urdailean, partikulakestilo kristalinoaren eraginaren ondorioz birrindu egiten dira, urdailean dauden entzimahidrolitikoek lehendabiziko liseriketa estrazelularra burutuz. Urdaila traktu ziliatudesberdinetan banatuta dago. Bertan, partikulak euren tamainaren arabera sailkatu etabanandu ondoren, liseri-guruineko dibertikuluetarantz bideratzen dira, liseri-konduktuensistematik. Liseri-konduktu primarioak urdailetik abiatzen dira, eta beraietatik adarkatzendiren kalibre txikiagoko liseri-konduktu sekundarioak liseri-guruineko dibertikuluetarairisten dira (Morton, 1983).
Liseri-guruina, anatomikoki bereizgarriak diren bi lobuluz osatuta dago,ezkerraldekoa eta eskubialdekoa, hain zuzen (Owen, 1966). Antolakuntza histologikoaridagokionez, hierarkizatutako antolakuntza lobularra ere antzeman da (1. Ird.; Basalo,
Moluskuen liseri-guruina
5

1998). Ehun konektibo dentsozko kapsula liseri-guruinaren bi lobuluak inguratzen ditu.Kapsula horrek loditze-gune nabarmen batzuk agertzen ditu, zeintzuetatik urdailaenkapsulatzen duen ehun konektibo dentsozko kapsularekin bat egin arte trenkadasendoak proiektatzen baitira. Horrela, dibertikuluen multzoak trenkada sendoen arteanmurgilduta agertu dira (lobulutxo primarioak). Lobulutxo primario bakoitza konduktuprimario baten inguruan antolatuta dago. Lobulutxo primarioen barruan ehun konektibozkoxaflek inguratutako lobulutxo sekundarioen taldekapenak bereizi dira. Horrela lobulutxosekundario bakoitzean 5-10 dibertikulu daude, konduktu sekundario berari lotuta daudenak(Basalo, 1998). Urdaila zein konduktu primarioen inguruan muskulu leuna dago; konduktusekundario eta dibertikuluetan, ordea, gero eta muskulu gutxiago agertzen da. Izatez,dibertikuluetan ez da muskulu leunaren zantzu esangarririk aurkitu (Basalo, 1998).
Gaur egun molusku bibalbioentzat onartuta dagoen liseri-guruinaren ereduaMortonek proposatu zuen 1983. urtean. Eredu horren arabera, liseri-guruina, urdailetikabiatutako konduktu (primario eta sekundario) adarkatuen bitartez konektaturik dauden
Zati Teorikoa
6
1. Irudia: Masson-Golder tindaketatrikromikoaz tindatutako muskuiluaren liseri-guruinaren ebakien irudiak. (A) Muskulu-ehun etaehun konektibozko trenkada bat ikus daiteke. Geziestuek muskulu-zelulak adierazten dituzte. Gezilodiek trenkadaren zatirik zabalena adieraztendute. Izarrek, ehun konektibotan joriak direntrenkadaren zatirik estuenak adierazten dituzte. (B)eta (C) irudiek lagin baten gune beretsuanmikrometro batzuen distantzian hartutako bi ebakinerakusten dituzte. Geziek trenkadak adieraztendituzte. Eskala-marra: 100 μm. (Basalo-tik (1998)hartua).

tubulu itsuz osaturik dago (2. Ird.). Dena den, ebakien gainean interpretazioak egiterakoaneredu tubular honek zenbait arazo sortu ditu. Tubuluak oso adarkatuta baldin baleude, osoluzeak ez balira behintzat, ukondo eta bihurguneen hainbat irudi behatuko liratekeebakietan, eta irudi hauek, berriz, urriak dira. Halaber, tubuluak oso adarkatuta ez baldinbaleude (Salvini-Plawen, 1988), konduktuak oso handiak izan beharko lirateke, etaebakietan agertzen diren konduktuen ebakidurak, berriz, urriak dira. Azkenik, konduktu etatubuluen arteko loturen bi dimentsiotako irudi eskematikoak plazaratu diren arren (Owen,1966; Salvini-Plawen, 1988; Beninger & LePennec, 1991), eskema horiek 3-dimentsiotako(3-D) antolakuntza espaziala ezin dute argiro azaldu. Gainera, liseri-dibertikuluetanematen den fluidoen sarrera- eta irteera-prozesuak eta liseriketa intrazelularrerakodibertikuluen epitelio-zeluletan ematen diren xurgapen-, jariapen- eta iraizpen-prozesukoordinatuak nekez azaldu daitezke eredu tubular honen arabera.
Aitzitik, 3-D berreraiketak eginez, muskuiluen liseri-guruinean unitatemorfofuntzionala liseri-albeoloa dela frogatu da (3. Ird.; Marigómez et al., 1995a; 1995b;Quincoces, 1995; Lekube, 1997). Liseri-albeoloak, konduktu sekundariotik irekitzen dira,
Moluskuen liseri-guruina
7
2. Irudia: Hematoxilina-eosinaz tindatutako muskuiluaren liseri-guruinaren ebakinen irudiak. (A) Liseri-guruinarenikuspegi orokorra, bertan (K) liseri-konduktuak, (G) liseri-albeoloen ganbarak eta (A) ehatz itxurako luzakinak bereizdaitezke. Ehun konektiboak liseri-egiturak inguratzen ditu. (B) Hatz itxurako luzakin baten soslaia xehetasun gehiagozikusten da. Bertan liseri-zelulak (LZ), eta zelula basofilikoak (ZB) bereiz daitezke.
Zati Teorikoa
8
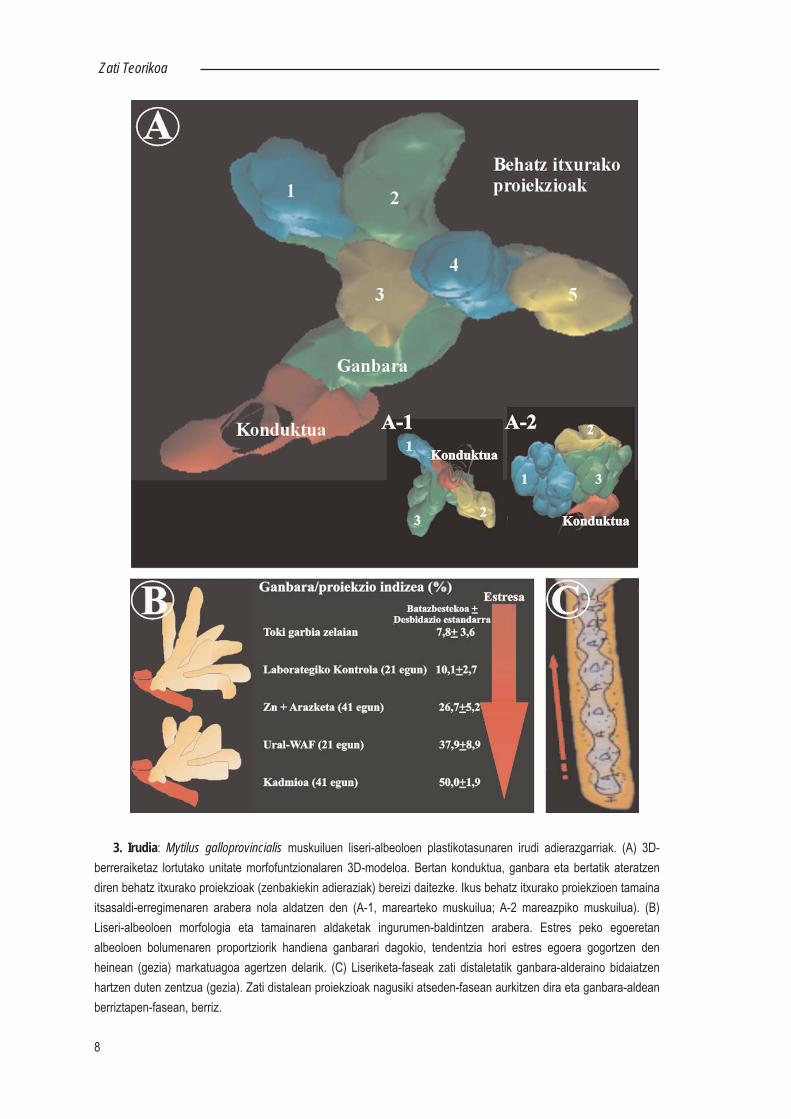
3. Irudia: Mytilus galloprovincialis muskuiluen liseri-albeoloen plastikotasunaren irudi adierazgarriak. (A) 3D-berreraiketaz lortutako unitate morfofuntzionalaren 3D-modeloa. Bertan konduktua, ganbara eta bertatik ateratzendiren behatz itxurako proiekzioak (zenbakiekin adieraziak) bereizi daitezke. Ikus behatz itxurako proiekzioen tamainaitsasaldi-erregimenaren arabera nola aldatzen den (A-1, marearteko muskuilua; A-2 mareazpiko muskuilua). (B)Liseri-albeoloen morfologia eta tamainaren aldaketak ingurumen-baldintzen arabera. Estres peko egoeretanalbeoloen bolumenaren proportziorik handiena ganbarari dagokio, tendentzia hori estres egoera gogortzen denheinean (gezia) markatuagoa agertzen delarik. (C) Liseriketa-faseak zati distaletatik ganbara-alderaino bidaiatzenhartzen duten zentzua (gezia). Zati distalean proiekzioak nagusiki atseden-fasean aurkitzen dira eta ganbara-aldeanberriztapen-fasean, berriz.

eta, Mortonen ereduko liseri-tubuluak ez bezala, estrukturalki konplexuak dira. Konduktutikgertuen dagoen albeoloaren zatia zabalduta agertzen da (ganbara) eta bertatik behatzitxurako luzakin tubularrak (4-10 proiekzio) proiektatzen dira. Zenbait kasutan, luzakinhoriek alde distalean banatuta egon daitezke, eta konduktu sekundario eta ganbararenartean estugunea (lepoa) ere ager daiteke. Ganbara zapalduta dago, eta bere dimentsioriktxikienean ez da behatz-itxurako luzakinen kalibrea baino handiagoa. Hori dela eta,tubuluen ukondo edo adarkatze-gune moduan interpretatu da hainbat lanetan, eta zenbaitkasutan ere gemazio bidez agertutako tubulu berrien sorgune gisa (Palmer, 1979). 3-Dberreraiketak emandako ikuspegi berri hori, kriohausturaz lortutako laginak ekorkuntzazkomikroskopio elektronikoan aztertuz egiaztatu egin da (Basalo, 1998) (4. Ird.).
Aipatutako albeoloen deskribapena muskuiluen kasuan indibiduo helduentzatbaliagarria da, baina muskuilu gazteetan (maskorraren luzera <1 cm) liseri-albeoloadesberdina da oso. Ganbarak oso zabalak dira, eta bertatik dozenaka behatz-itxurakoluzakin ateratzen dira. Dirudienez, muskuilu gazteen liseri-guruinean unitatemorfofuntzional horien kopurua txikia izango litzateke, eta zenbait kasutan konduktuprimario bakoitzetik unitate bakarra aterako litzateke (Lekube, 1997).
Moluskuen liseri-guruina
9
4. Irudia: Muskuiluen liseri-guruinaren eredu albeolotubularraren irudi eskematikoa (V. Asensioren kortesiaz)

Liseri-albeoloen epitelioan nagusiki 2 zelula-mota aurkitu dira, liseri-zelulak etazelula basofilikoak (Owen, 1972) (2. eta 5. Ird.).
Liseri-zelulak (5. eta 6. Ird.) azidofilikoak dira. Itxurari dagokionez prismatikoak dira,baina kubikoak ere izan daitezke. Erpinaldea, mikrobiloxkez hornituta dago. Mikrobiloxkenkopurua eta garapena oso aldakorra da, zelulen egoera metabolikoaren arabera
Zati Teorikoa
10
6. Irudia: Transmisiozko mikroskopio elektroniko bidez lortutako Mytilus galloprovincialis muskuiluaren (A) liseri-zelula (handipena x6000) eta (B) zelula basofilikoaren (handipena x9000) irudiak. Liseri-zelulan sistemaendolisosomiko garatua argiro beha daiteke. (N) nukleoa, (M) muskulua, (BL) xafla basala, (RB) hondakin-gorputza,(HL) heterolisosoma, (HP) heterofagosoma, (JC) lotura-konplexua. Zelula basofilikoa, ordea, elektrodentsoagoa daeta erretikulu endoplasmatiko pikortsua eta Golgi sistema oso garatua ditu. (N) nukleoa, (GC) Golgi-konplexua, (CT)ehun konektiboa (LF) lipofuszina. (Cajaraville-tik(1989) birmoldatua).
5. Irudia: Liseri-zelula eta zelula basofilikoaren ultrastrukturaren irudi eskematikoa. (A) Liseri-zelularen sistemaendo-lisosomiko garatuaren osagaiak bereiz daitezke. (B) Zelula basofikoaren soslaia piramidala da, eta erretikuendoplasmatiko pikortsua eta Golgi sistema oso garatuak ditu. (Owen-tik (1973) birmoldatua).

(Cajaraville et al., 1991). Zelula hauek, sistema endo-lisosomiko oso garatua dute,liseriketa intrazelularra beraien funtziorik nagusiena delako (Robledo & Cajaraville, 1996).Alboetako eta oinaldeko mintzek interdigitazio eta inbaginazioak erakusten dituzte.Endozitosi-besikula, heterofagosoma, heterolisosoma, eta hondakin-gorputz ugari daude.Endozitosi-besikulak mikrobiloxkekin erlazionaturik agertzen dira, zelularen erpinaldean(Owen, 1970; Pal, 1972; Henry et al., 1991). Heterofagosomak, besikula erregularrak etatamaina aldakorrekoak dira, eta, orokorrean, hauek ere erpinaldean deskribatu dira (Owen,1973). Heterolisosomak, zelularen erdialdean agertzen dira, heterofagosomak bainohandiagoak eta esferikoak dira, eta beraien barneko materiala oso heterogeneoa da.Azkenik, hondakin-gorputzak zelularen oinaldean agertzen dira. Hondakin-gorputz horiekargira askatzen dira, konduktu primario eta sekundarioen argietan ere aurkitu direlarik(Cajaraville, 1989).
Zelula-basofilikoak (5. eta 6. Ird), liseri-zelulak baino kopuru txikiagotan agertzendira liseri-hodietan. Itxura piramidala dute, eta erpinaldean mikrobiloxkak dituzte. Mintzak,oinaldean zein alboetan, inbaginazio txikiak ditu. Zelula hauen ezaugarri garrantzitsuenabasofilia da, duten erretikulu endoplasmatiko pikortsu ugaria dela eta (Henry, 1987).Horretaz gain, Golgi aparatua ere oso garatua dute. Nukleoa orokorrean zelularenoinaldean agertzen da. Autore batzuk, bigarren zelula basofiliko bat deskribatu dute,Cardium edule (Owen, 1970) edo Mytilus edulis (Thompson et al., 1974) moluskuetan.Bigarren zelula basofiliko hori, flagelatua omen da.
Bibalbioen liseri-epitelioaren morfologia aldakorra da, izan ere liseriketa prozesuaziklikoa eta dinamikoa denez, aurrera doan heinean liseri-albeoloen epitelioek liseriketa-fase desberdinei dagozkien itxura desberdinak erakusten dituzte (7. Ird., Langton, 1975;Robinson & Langton, 1980; Morton, 1983): (a) I mota edo atseden-fasea; (b) II mota edoxurgatze-fasea, (c) III mota edo desegite-fasea; eta (d) IV mota edo berregite-fasea.
Prozesuaren koordinazioa dela eta, eredu tubularrean oinarrituta moluskubibalbioen artean klasikoki bi motatako liseriketa-jarduera bereiztu dira: monofasikoa etabifasikoa. Liseriketa monofasikoan, liseri-guruinean tubulu-mota bakarra izango litzatekenagusi une bakoitzean, fotoperiodo edo eta itsasaldiaren arabera. Jarduera-mota horiDreissena polymorpha, Cardium edule, Ostrea edulis eta Scrobicularia plana espezieetandeskribatu da (Morton, 1969; 1970; 1971; Owen, 1972). Liseriketa monofasikoan liseriketa-ziklo bakoitzean, hau da, itsasaldi-ziklo bakoitzean, II motako tubuluek, I motako tubuluakordezkatzen dituzte, eta amaieran III motako tubuluak dira nagusi. Liseriketa bifasikoan,ordea, aldi berean bi tubulu-mota nagusituko lirateke. Prozesu hau Lasaea rubra, Cardiumedule eta Ostrea edulis espezieetan deskribatu da (McQuiston, 1969; Owen, 1972). Kasu
Moluskuen liseri-guruina
11

horretan, III motako tubuluen proportzioa liseriketa prozesuan zehar konstante mantentzenda, eta I eta II motako tubuluen artean txandakatzea suertatzen da itsasaldi-zikloan zehar(McQuiston, 1969). Edozein modutan, tubulu guztietara elikagaia aldi berean, neurriberean eta konposizio berarekin heltzen ez denez, liseri-guruinean 4 liseriketa-faseak(tubulu-motak) bereiz daitezke aldi berean, proportzio oso desberdinetan bada ere(McQuiston, 1969; Robinson & Langton, 1980; Robinson, 1983).
Lau tubulu-moten existentzian oinarrituz, liseriketa intrazelularra azaltzeko teoriabat plazaratu zen (Morton, 1983; Henry, 1987; Beninger & Le Pennec, 1991). Elikagaiakheldu bitartean, tubuluak atseden-fasean egongo ziren. Elikagaiak tubuluetarairisterakoan, liseri-zelulek elikagaiak xurgatuko lituzkete (xurgatze-fasea), eta liseriketabukatzearekin batera zelula asko deuseztatuko ziren (desegite-fasea). Azkenik, hurrengoliseriketa-zikloari ekin ahal izateko, epitelioa zelula berriez hornituko zen (berregite-fasea). Teoria horren arabera, liseriketa-zikloa eta epitelioaren berriztapen-zikloa koordinaturikegongo ziren (Morton, 1983), eta zelulen berriztapen masiboa ordu gutxitako epeansuertatu beharko zen. Hau da, liseri-epitelioan zelulen berriztapen-maila izugarri handiaizan beharko litzateke, eta liseri-tubuluetako epitelioa partzialki (liseriketa difasikoa) edoguztiz (liseriketa monofasikoa) berriztatua izango litzateke elikatze-ziklo bakoitzeko,etengabe. Aitzitik, epitelio-zelulen zatiketa ez da batere konspikuoa liseri-epitelioan(Marigómez et al., 1998a).
Zati Teorikoa
12
7. Irudia: Liseriketa prozesuan zehar muskuiluenliseri-epitelioak hartzen dituen morfologia desberdinak.(A) Atseden-fasea, (B) Xurgatze-fasea, (C) Desegite-fasea (D) Berregite-fasea. (Langton-tik (1975)birmoldatua).

Eredu albeolarra aintzat hartzean, tubulu-moten existentziaren eta eurentxandakatze-ereduaren esangura fisiologikoa oso bestelakoa dela frogatu da. Izan ere, M.galloprovincialis muskuiluaren liseri-guruinean egindako 3-D berreraiketek, liseriketa-faseak zati distaletatik ganbara-alderaino bidaiatzen duten eta albeolo bereko luzakinetanzehar modu sinkronikoan aurrera egiten duten uhinen moduan agertzen direla erakutsidute (3C Ird.; Marigómez et al., 1995a; 1995b). Hau da, luzakinen zati distalen epitelioanatseden-fasea nagusitzen den bitartean, hurrengo lakainetan xurgatze- eta desegite-fasetxandakatuak agertzen dira, ustezko berregite-fasea ganbara-alderantz nagusituz doalarik(Quincoces, 1995). Bestalde, entzima- eta lektina-histokimika, eta morfometria aplikatuz,liseri-zelulen ezaugarrietan oinarritutako liseriketa-faseen esangura fisiologikoa berraztertu
Moluskuen liseri-guruina
13
8. Irudia: ß-Glukuronidasa entzimaren (A) eta WGA lektinaren bidezko N-azetil glukosamina hondarren (B)detektapen histokimikoa liseri-guruineko epitelioan liseriketa-fase desberdinetan. Sailkapen “klasikoaren” arabera: A-1/B-1 atseden-fasea izango litzateke, A-2/B-2 xurgatze-fasea, A-3/B-3 desegite-fasea eta A-4/B-4 berregite-fasea.Jarduera entzimatiko eta lektinen emaitzen arabera, liseriketa-faseen sailkapena aldatu beharko litzateke. Atseden-fasea, xurgatze-fasea izan beharko litzateke (WGA marka bortitza epitelioaren erpinaldean (B-1)), xurgatze-fasea,liseritze-fasea (entzimaren jarduera nabarmena eta WGA markaketa lisosometan (A-2 eta B-2)), desegite-fasea,liseritze-fase berantiarra (oso lisosoma handiak (A-3)) eta berregite-fasea, atseden-fasea (lisosoma txiki eta urriak etaWGA markaketa difuso eta ahula epitelioan zehar barreiatuta (A-4 eta B-4)).
da (8. Ird., Marigómez, argitaratu gabe). Berregite-fase "klasikoa" delakoa seguruenikatseden-faseari legokioke, liseri-zeluletan lisosoma urriak izanik, ezaugarri histokimikoekhori adierazi baitute. Izan ere, marearteko muskuiluak uretatik kanpo nahiko denbora igaroondoren (hau da, elikatzerik ez dutenean), IV-motako tubuluen morfologia nagusitzen da(Cajaraville et al., 1991; 1992). Halaber, datu berriek mantentze-fase "klasikoa" xurgapen-faseari legokiokeela erakutsi dute, eta desegite-fase delakoan dauden albeoloakxurgapen-fase berantiarrari edo liseriketa intrazelularraren fase aurreratuari legozkieke(Marigómez, argitaratu gabe). Interpretazio berri honetan, epitelioaren berriztapena inolazere kontuan hartu ez dena azpimarratzekoa da.
1.2.- Gastropodo itsastarrak (Littorina eredua)
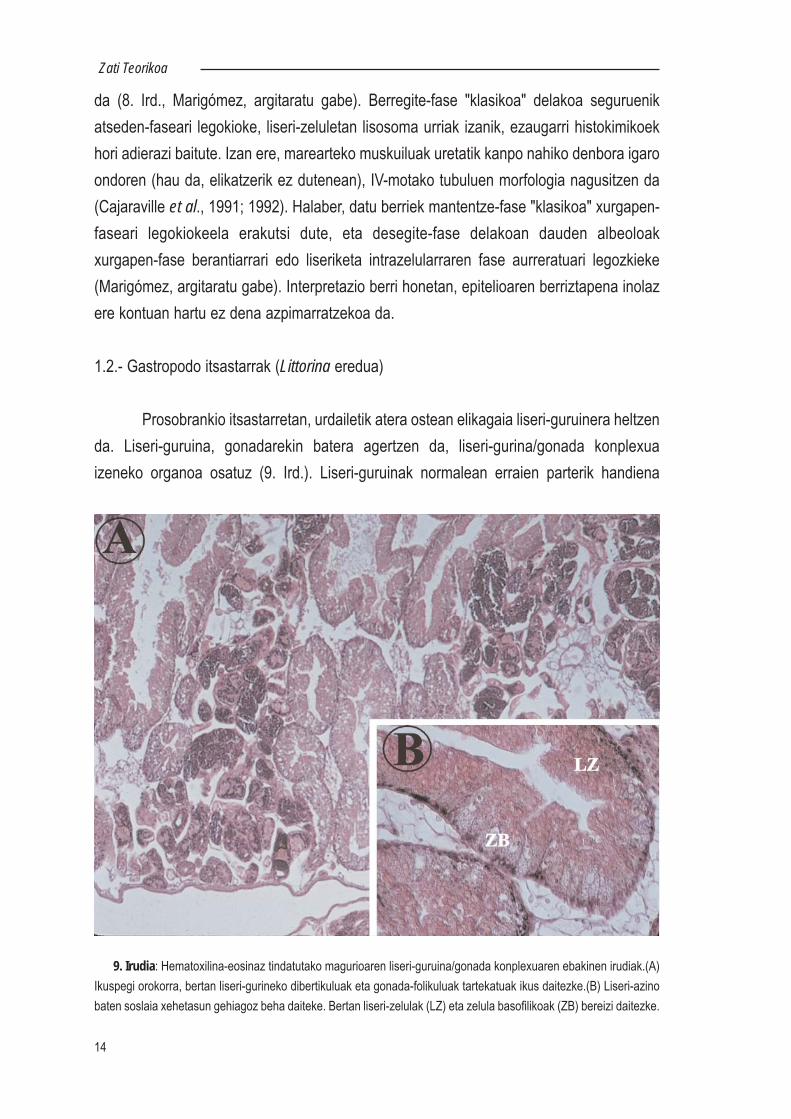
Prosobrankio itsastarretan, urdailetik atera ostean elikagaia liseri-guruinera heltzenda. Liseri-guruina, gonadarekin batera agertzen da, liseri-gurina/gonada konplexuaizeneko organoa osatuz (9. Ird.). Liseri-guruinak normalean erraien parterik handiena
Zati Teorikoa
14
9. Irudia: Hematoxilina-eosinaz tindatutako magurioaren liseri-guruina/gonada konplexuaren ebakinen irudiak.(A)Ikuspegi orokorra, bertan liseri-gurineko dibertikuluak eta gonada-folikuluak tartekatuak ikus daitezke.(B) Liseri-azinobaten soslaia xehetasun gehiagoz beha daiteke. Bertan liseri-zelulak (LZ) eta zelula basofilikoak (ZB) bereizi daitezke.

betetzen du; ugalketa garaian izan ezik, noiz gonadaren tamaina asko emendatzen baita.Hala ere, liseri-guruinean tamaina desberdineko 2 lobulu bereiz daitezke, orokorreanezkerreko lobulua handiagoa delarik. Urdaileko aurrealdera 3 konduktu labur etaadarkatugabez lotuta dago liseri-guruina. Liseri-guruina hainbat azino adarkatuez osatutadago (Fretter & Graham, 1962), eta bertan elikagaien xurgapena eta liseriketa zeinhondakinen andeatzea eta iraiztea burutzen dira (Merdsoy & Farley, 1973). Prosobrankioitsastarren liseri-guruinean oinarrizko bi zelula-mota daude, liseri-zelula eta zelulabasofilikoa, alegia (10. Ird.).
Liseri-zelula, (syn., guruin-zelula, xurgapen-zelula, zelula hepatiko, gibel-zelula,bakuolo-zelula, zelula jariatzaile) kolumnarra da, eta erpinaldean mikrobiloxka ugari etaproteinetan joriak diren pikor zitoplasmatikoak dauzka (Lufty & Demian, 1967; Mason1983; Voltzow, 1994). Oinaldean, zenbaitetan errefringentea izan daitekeen edukiheterogeneoa duten besikulak agertzen dira. Tamaina txikiko nukleoa, orokorreanerdialdean edo oinaldean kokatzen da. Zelula-mota honen zeregin nagusia liseriketaintrazelularra burutzea da (Voltzow, 1994).
Moluskuen liseri-guruina
15
10. Irudia: Liseri-zelula eta zelula basofilikoaren irudi eskematikoak. (A) Zelula basofikoaren soslai piramidala etaerretiku endoplasmatiko garatua ikus daitezke, beste organuluekin batera. (B) Liseri-zelula egitura kolumnarra daukaeta morfologia desberdinak har ditzake liseriketa fasearen arabera. (cl), zilioa; (cr), zilioaren gorputz basala; (mv),mikrobiloxkak; (ps), proteinazko jariapen-pikorrak; (cy), zitosola; (gr), pikor mineralizatuak; (rer), erretikuluendoplasmatiko pikortsua; (mi), mitokondrioa; (sv), iraizte-pikorra (gb), diktiosoma; (lin), tolesdura laterala; (nu),nukleoa; (nue), nukleoloa; (bin), tolesdura basala; (bl), xafla-basala; (m), muskulua; (hae), hemozianina; (lu), lumena;(dv), liseri-bakuoloa; (li), lipidoa, (pv), pinozitosi pikorra; (rb), hondakin-gorputza. (Mason-etik (1983) hartua).

Zelula basofilikoa, (syn., zelula jariatzailea, kripta-zelula, iraizte-zelula, hartzitze-zelula) liseri-zelula baino urriagoa da (Lufty & Demian, 1967; Mason 1983; Voltzow, 1994).Hematoxilina-eosinaz tindatzerakoan, zitoplasma liseri-zelularena baino askozbasofilikoagoa suertatzen da, izena hortik datorkiolarik. Itxura piramidala edo konokaraizanik, banaka zein 3-4 zeluletako multzoetan ager daiteke, askotan liseri-tubuluen ertzperiferikoei asoziatuta. Nukleolo oso nabarmena daukan nukleo handia ageri da oinaldean.Erretikulu endoplasmatiko pikortsua oso garatua dago, eta nukleoaren inguruan kokatzenda. Bestalde, mintz-pikor zitoplasmatikoen presentzia nabarmena da. Batez ereerpinaldean dauden pikor batzuk oso elektrodentsoak dira, eta seguruenik jariapen-proteinez osatuta daude. Oinalderagontz sakabanaturik dauden beste pikorretan, ordea,fosfato kaltzikoa dugu osagai nagusi (Mason, 1983; Marigómez et al., 1990). Zelulabasofilikoek, kaltzioaren garraioan eta metaketan parte hartzen dute, eta liseriketaestrazelularrerako entzimak jariatzen dituzte (Owen, 1956; Fretter & Graham, 1962;Merdsoy & Farley, 1973; Mason, 1983).
Magurioen liseri-guruinean, nahiz eta muskuiluetan deskribatzen diren "tubulu-mota" desberdinak ez agertu, liseri-zelulen pikorren kopuru eta morfologiaren araberaliseriketa-prozesuarekin erlazionatutako 3 edo 4 fase bereiztu dira (11. Ird., Sáez et al.,1990). Xurgapen-fasean pikor eosinofilikoak agertzen dira erpinaldeko ertzean, eta apurbat beherago purpura kolorea hartzen duten pikor ilunak. Liseriketa-fase goiztiarrean
Zati Teorikoa
16
11. Irudia: Liseri-zelulek liserike-fase desberdinetan erakusten dituzten morfologia desberdinen irudi eskematikoa.(A) Xurgatze-fasea. Bertan zelularen erpinaldean pikor eosinofilikoak agertzen dira, eta euren azpian pikor ilun etatxikiagoak agertzen dira. (B) Liseriketa-fase goiztiarra. Pikor eosinofilikoak erpinaldean ageri dira eta pikor ilunakoinalderantz zabaltzen dira. (C) Liseriketa-fase berantiarra. Erpinaldean pikor eosinofilikoek darraite eta oinaldeanpikor ilunekin batera lipofuszinen antzeko pikor horiak ageri dira. Pikor horiok fase honen karakteristikoak dira. (D)Iraizte-fasea. Erpinaldean urriak diren pikor eosinofilikoak zenbait kasutan bakuoloen barruan ageri dira. Lipofuszinenantzeko pikorrak oso nabarmenak dira (Sáez et al.-etik (1990) hartuta).

aipatutako pikor ilunagoak zelula osoan zehar barreiaturik agertzen dira. Liseriketa-faseberantiarrean, pikor ilunetaz gain lipofuszinen antzeko pikor horiak oinaldeannabarmentzen hasten dira (Sáez et al., 1990). Azkenik, iraizte-fasean pikor gutxiagoagertzen dira, eta bakuolo txikien zein lipofuszina-pikorren presentzia nabaria da. Prozesuhori, itsasaldien erregimenarekin estuki erlazionatuta dago. Izan ere, liseriketa batez ereitsasbeherako egoeretan gauzatzen da (Sáez et al., 1990).
1.3.- Lehorreko gastropodoak (Arion eredua)
Arion ater barearen liseri-guruina gorputzaren atal handia betetzen du, etaanatomikoki bereizgarriak diren bi guruin-zatitan bananduta dago: aurreko guruin-zatia,normalean txikiena, eta atzeko guruin-zatia. Bi guruin-zati horiek, papoarekin (aurrekoguruin-zatia) eta urdailarekin (atzeko guruin-zatia) bi konduktu bereizien bitartezkomunikatuta daude (Roach, 1968). Guruin-zati horiek, aurrekoa zein atzekoa, hainbatlobulutan daude bananduta (12. Ird., Runham & Hunter, 1970). Lobulu bakoitza,adarkatuak zein multzoka agertzen diren hainbat dibertikuluz osatuta dago. Liseri-dibertikuluak unitate adenomerikoz daude osaturik. Unitate adenomerikoan (13. Ird.), liseri-konduktu izeneko gune zentral zabala dago, eta bertatik azino-itxurako zabalguneakadarkatzen dira (liseri-azinoak).
Moluskuen liseri-guruina
17
12. Irudia: Arion ater barearen liseri-sistemaren irudi eskematikoa. Bertan elikagaien liseriketan parte hartzenduten organo desberdinak ageri dira eta liseri-guruinaren izaera lobularra bereiz daiteke. (Argaud & Bounoure-tik(1910) birmoldatua).
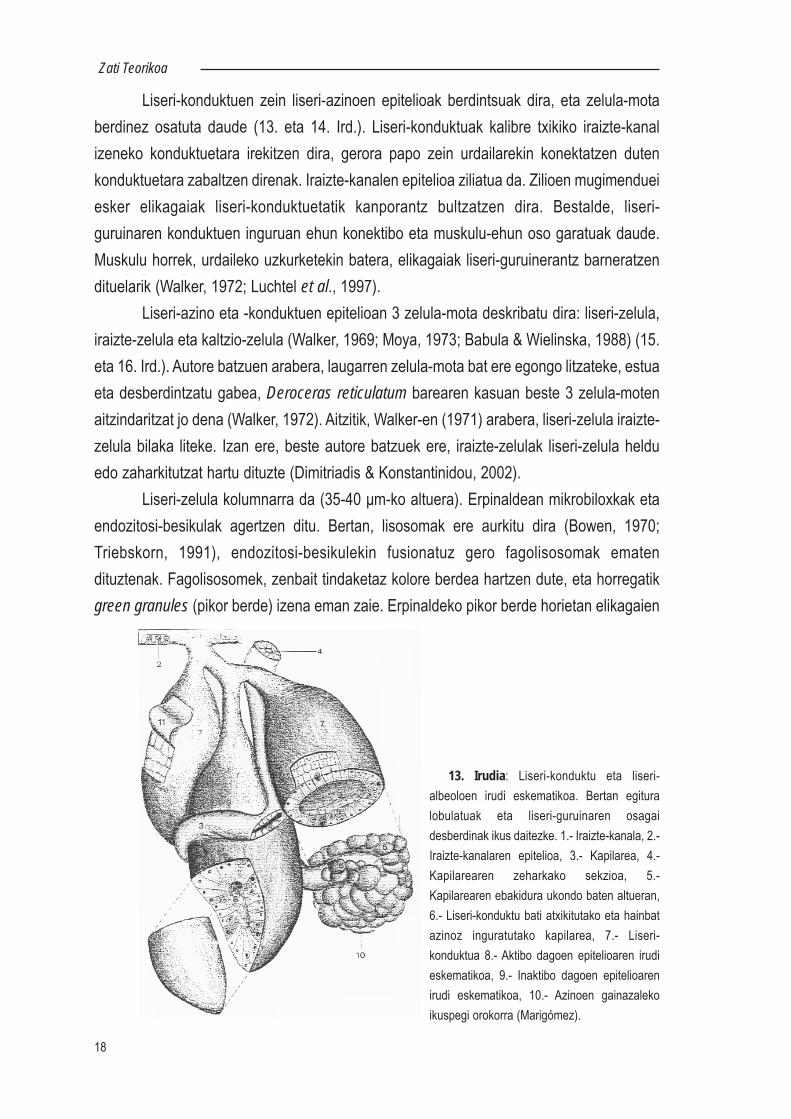
Liseri-konduktuen zein liseri-azinoen epitelioak berdintsuak dira, eta zelula-motaberdinez osatuta daude (13. eta 14. Ird.). Liseri-konduktuak kalibre txikiko iraizte-kanalizeneko konduktuetara irekitzen dira, gerora papo zein urdailarekin konektatzen dutenkonduktuetara zabaltzen direnak. Iraizte-kanalen epitelioa ziliatua da. Zilioen mugimendueiesker elikagaiak liseri-konduktuetatik kanporantz bultzatzen dira. Bestalde, liseri-guruinaren konduktuen inguruan ehun konektibo eta muskulu-ehun oso garatuak daude.Muskulu horrek, urdaileko uzkurketekin batera, elikagaiak liseri-guruinerantz barneratzendituelarik (Walker, 1972; Luchtel et al., 1997).
Liseri-azino eta -konduktuen epitelioan 3 zelula-mota deskribatu dira: liseri-zelula,iraizte-zelula eta kaltzio-zelula (Walker, 1969; Moya, 1973; Babula & Wielinska, 1988) (15.eta 16. Ird.). Autore batzuen arabera, laugarren zelula-mota bat ere egongo litzateke, estuaeta desberdintzatu gabea, Deroceras reticulatum barearen kasuan beste 3 zelula-motenaitzindaritzat jo dena (Walker, 1972). Aitzitik, Walker-en (1971) arabera, liseri-zelula iraizte-zelula bilaka liteke. Izan ere, beste autore batzuek ere, iraizte-zelulak liseri-zelula helduedo zaharkitutzat hartu dituzte (Dimitriadis & Konstantinidou, 2002).
Liseri-zelula kolumnarra da (35-40 μm-ko altuera). Erpinaldean mikrobiloxkak etaendozitosi-besikulak agertzen ditu. Bertan, lisosomak ere aurkitu dira (Bowen, 1970;Triebskorn, 1991), endozitosi-besikulekin fusionatuz gero fagolisosomak ematendituztenak. Fagolisosomek, zenbait tindaketaz kolore berdea hartzen dute, eta horregatikgreen granules (pikor berde) izena eman zaie. Erpinaldeko pikor berde horietan elikagaien
Zati Teorikoa
18
13. Irudia: Liseri-konduktu eta liseri-albeoloen irudi eskematikoa. Bertan egituralobulatuak eta liseri-guruinaren osagaidesberdinak ikus daitezke. 1.- Iraizte-kanala, 2.-Iraizte-kanalaren epitelioa, 3.- Kapilarea, 4.-Kapilarearen zeharkako sekzioa, 5.-Kapilarearen ebakidura ukondo baten altueran,6.- Liseri-konduktu bati atxikitutako eta hainbatazinoz inguratutako kapilarea, 7.- Liseri-konduktua 8.- Aktibo dagoen epitelioaren irudieskematikoa, 9.- Inaktibo dagoen epitelioarenirudi eskematikoa, 10.- Azinoen gainazalekoikuspegi orokorra (Marigómez).

eskurapena eta liseriketa burutzen dira. Oinalderagontz askoz handiagoak diren etaliseriketaren hondakinak diren lipofuszinez osatuta dauden yellow granules (pikor hori)agertzen dira (Walker, 1972). Nukleoa oinaldean kokaturik dago, eta bere inguruanerretikulu endoplasmatikoa, Golgi aparatua eta zenbait kasutan glukogenoa eta lipido-tantak ere agertzen dira (Sumner, 1969; Walker 1969; Moya, 1973). Ezaugarriutrastruktural berezia manosoma izeneko mintz-egitura dugu (Baumforth et al., 1998).Manosomek, sei hodiko errosetatan antolatuta daude, eta hornidura entzimatikoaespezifikoa (manitol oxidasa) dute (Moya & Rallo, 1975; Czarna et al., 1985; Baumforth etal., 1998). Dirudienez, manosomak gastropodo lurtarren liseri-zeluletan arruntak dira(David & Götze, 1963). Oinaldeko zitoplasman, ioi-garraioarekin erlazionatutako fosfatasaalkalino jarduera behatu da (Bowen & Davies, 1971). Liseri-zelulek liseriketa-prozesuanzehar itxura desberdina erakutsi dezakete, altueraz zein mikrobiloxken eta pikorzitoplasmatikoen kopurua aldatuz (Sumner, 1965).
Kaltzio-zelulak (syn., kripta-zelula; zelula jariatzailea) itxura piramidala dutenzelulak dira (Sumner, 1969; Walker 1969; Moya, 1973). Erpinaldean mikrobiloxkak dituzte,
Moluskuen liseri-guruina
19
14. Irudia: Hematoxilina-eosinaz tindatutako barearen liseri-guruinaren ebakien irudiak. (A) Liseri-guruinarenikuspegi orokorra, bertan liseri-azino (A) eta -konduktuak (K) ikus daitezke. (B) Liseri-azino baten soslaia detailegehiagoz beha daiteke. Bertan liseri-zelulak (LZ) , iraizte-zelulak (IZ) eta kaltzio-zelulak (KZ) bereizi daitezke.

eta horien azpian mitokondrioak, erretikulu endoplasmatiko leuna eta hainbat besikulapleomorfiko agertzen dira. Zelularen erdialdean proteinetan joriak diren besikulak paraturikdaude. Oinaldean, nabarmenki handia den nukleoa aurkitu da. Lipido-tantak xaflabasalaren gaineko zitoplasmaren eremuan ager daitezke. Zelulan zehar materialheterogeneoa duten pikor biribilak ageri dira tipikoki. Pikor horiek, geruza kontzentrikozparatutako matrizea dute, X-izpien mikroanalisia eta beste metodoen bitartez, behintzatHelix aspersa barraskilo eta Arion rufus barearen kasuetan frogatu denez, kaltzio,
Zati Teorikoa
20
16. Irudia: Transmisiozko mikroskopio elektroniko bidez lortutako bareen (A) liseri-zelula (handipena x 2500), (B)kaltzio-zelula (handipena x31400) eta (C) iraizte-zelularen (handipena x3500) irudiak. Liseri-zelulan sistema endo-lisosomiko garatua beha daiteke. Kaltzio-zelulan, pikorren geruza kontzentrikoak bereiz daitezke. Iraizte-zelularenerpinaldean dagoen bakuolo handian izaera desberdineko pikorrak ikus daitezke. (Luchtel et al-tik (1997)birmoldatua).
15. Irudia: Liseri-zelula, kaltzio-zelula eta iraizte-zelularen (A) eta (B) Liseri-zelularen egitura kolumnarra etasistema endo-lisosomiko garatua bereiz daiteke. (C) Kaltzio-zelularen soslaia piramidala da eta kaltzio pikorrak errezbereiz daitezke. (D) Iraizte-zelularen bakuolo handia da zelula-mota honen egiturarik bereizgarriena. (Walker-tik(1972) birmoldatua).

magnesio eta fosforoz (CaMgP2O7 moduan) osatuta dagoena (Howard et al., 1981; Mason& Simkiss, 1982; Janssen, 1985). Horregatik, pikor horiei kaltzio-esferula izena eman zaie,eta zelulei kaltzio-zelularena. Hala ere, beste zenbait elementu ere badaudela egiaztatuda, manganesoa, burdina, kobaltoa eta zinka, besteak beste (Simkiss, 1981; Mason &Simkiss, 1982, Recio et al., 1988). Kaltzio-zelularen funtzioari dagokionez, proposamenezberdinak plazaratu dira. Ikertzaile batzuen arabera, barne-medioaren pH-a orekatzekoindargetzaile gisa funtzionatuko luke kaltzioak (Krigjsman, 1928). Beste autore batzuenaburuz, kaltzio-zelulek metabolismo orokorrean eta muki-ekoizpenean ezinbestekoa denkaltzio kantitate handien biltegi gisa funtzionatuko lukete (Walker, 1971).
Iraizte-zelulek erpinaldean mikrobiloxka ugari dituzte, eta hauen azpian hainbatpikor eta mitokondrio, erretikulu endoplasmatikoa eta diktiosomak. Hala ere, zelula hauenezaugarririk deigarriena zelularen bolumen gehiena betetzera irits daitekeen bakuolohandia da (Sumner, 1969; Walker 1969; Moya, 1973). Bakuoloaren edukina heterogeneoada, eta batez ere lipofuszinez osatuta dago. Lehen aipatu den bezala, autore batzuenarabera, iraizte-zelulak liseri-zeluletatik eratorriak izan litezke (Walker, 1971), afera horioraindik ere eztabaidan dagoen arren (Dimitriadis & Konstantinidou, 2002).
1.4.- Liseri-guruinaren plastikotasuna
1.4.1.- Epitelioaren aldaketa morfologikoak
Liseri-guruinaren egitura ez da egonkor mantentzen; aitzitik, egoera desberdinenaurrean plastikotasun handia erakusten du. Hainbat ikerlanek deskribatutakoaren arabera,ingurugiroaren estres-iturrien pean, estrusio apokrino/holokrino eta aufogafia prozesuenemendioa behatu da liseri-epitelioan (Soto et al., 1996). Izan ere, kutsatzaile organiko zeinez-organikoen pean egondako moluskuetan eta baita bestelako estres orokorrekoegoeratan (adb., baraualdiaren ondorioz), liseri-epitelioan aldaketa morfologikoesanguratsuak behatu dira, epitelioaren altueraren murrizpena eta zelula-motenproportzioaren aldaketak, bestek beste (Lowe et al., 1981; Couch, 1984; Marigómez et al.,1986; 1993; 1996; 1998b; 2005; Recio et al., 1988; Vega et al., 1989; Cajaraville et al.,1991).
Liseri-epitelioaren batez besteko lodiera adierazteko MET (ingelesezko MeanEpithelial Thickness) parametroa erabili da. Dena den, MET parametroak ezin du bereosotasunean liseri-guruineko epitelioak pairatutako aldaketa estruktural guztiak azaldu.Liseri-dibertikuluaren tamaina adierazten duen MDR (ingelesezko Mean Diverticular
Moluskuen liseri-guruina
21

Radius) eta lumenaren kalibrearen adierazlea den MLR (ingelesezko Mean LuminalRadius) parametroek ere informazio baliotsua eman dezakete, eta baita MLR/MET etaMET/MDR parametro erlatiboek ere (Vega et al., 1989). Orokorrean, estresatutakomoluskuen MET eta MDR balioak baxuak eta MLR balioak altuak dira. Hala ere, aldaketamorfologikoak ez dira beti espero bezala. Zenbait ikerlanetan, MET jaistearekin batera,MDR mantendu edo igo daiteke (Cajaraville et al., 1991; Cajaraville et al., 1992). MDR,batez ere, estres-iturri iraunkorren aurrean jaisten da esangarriki. Bestalde, mareartekobibalbioen kasuan, liseriketa-jarduera fasikoek ere liseri-epitelioaren altueraren gaineragina dute. I eta II motetako tubuluen morfologia duen liseri-epitelioa IV motetakoenabaino garaiagoa da (7. Ird.). Gainera, estres-egoeretan, MLR goratzearekin batera, IVmotako tubuluen morfologia duen epitelioaren hedapena ere emenda daiteke (Cajaravilleet al., 1991). Azkenik, zenbait egoera patologikotan, kasik epiteliorik gabeko tubuluatrofikoei legokioken V motako epitelio-morfologia aurkitu da (Couch, 1984).
Halaber, muskuiluen liseri-guruinaren kasuan liseri-albeoloen morfologia etatamaina ere ingurumen-baldintzen arabera alda daitezke. Alde batetik, hatz itxurakoproiekzioen tamaina, itsasaldi-erregimenaren arabera desberdina izan daiteke. Bestalde,egoera normaletan hatz itxurako proiekzioak nabarmenagoak diren bitartean, estres pekoegoeretan albeoloen proportziorik handiena ganbarari dagokio, 3D-berreraiketetanoinarrituta frogatu den moduan (3. Ird.; Quincoces, 1995; Lekube 1997).
Oro har, ingurumen-baldintzen arabera suertatutako aldaketa morfologikoek,epitelioaren masa, hots zelulen masa edo kopurua, oso aldakorra dela agerian utzi dute.Gainera, aldaketak, itzulgarriak izanik, ordu gutxitan suertatzen dira gehienetan. Horriesker, epitelio-zelulen berriztapenaren garrantziaz jabetu gara, nahiz eta, gerora azaldukodenez, epitelio honen berriztapenari buruz ezer gutxi dakigun.
1.4.2.- Zelula-moten ordezkapena
Morfologian ez ezik, liseri-guruineko epitelioa osatzen duten zelula-motenpresentzia erlatiboan ere aldakortasun esanguratsua aurkitu da. Oro har, egoera fisiologikoeta patologiko jakin desberdinetan zelula basofilikoen presentzia erlatiboa areagotudaitekeela deskribatu da. Erantzun hori orokorra da molusku espezien artean, xenobiotikoorganiko eta metal pean egondako magurio zein bibalbioetan (Rasmussen et al., 1983;Lowe, 1988; Cajaraville et al., 1990; 1991; Marigómez et al., 1990; Soto et al., 1997, 2002),eta metal pean egondako bareetan (Marigómez et al., 1986, 1996, 1998b) deskribatu denbezala. Gainera, bestelako estres-egoeretan (adb., baraualdia eta bizkarroein presentzia)
Zati Teorikoa
22

ere erantzun-mota bera aurkitu da (Yoshino, 1976; Marigómez et al., 1992; 1993).Esaterako, Mytilus edulis muskuiluek baraualdi luzea jasan ostean, flagelodun etaflagelobako zelula basofilikoen kopuru erlatiboak gora egin zuela ondorioztatu zen(Thompson et al., 1974). Gainera, dirudienez, zelula-moten proportzioaren aldaketa behinbehinekoa dirudi. Adibidez, Littorina littorea magurioaren kasuan, petrolio-eratorrien peanegotearen ondorioz zelula basofilikoen proportzioak gora egin arren, erantzuna itzulgarriadela ikusi da (Widdows et al., 1984).
Testuinguru honetan, behatutako zelula-moten kopuru erlatiboen aldaketa liseri-zelulen kopuruaren jaitsiera edo zelula basofilikoen proliferazioaren bitartez burutzen oteden argitzeke dago. Litekeena da prozesu biak, liseri-zelulen narriadura eta zelulabasofilikoen proliferazioa, aldi berean gertatzea. Izan ere, zelula basofilikoen kopuru etadesegite-faseko liseri-epitelioaren hedapenaren artean korrelazio positibo esangarriaaurkitu da (Cajaraville et al., 1990). Beste autore batzuen arabera, ordea, zelulabasofilikoen proliferazioa estresak induzitutako liseri-zelulen galeraren aurretik gertatukolitzateke (Thompson et al., 1974). Zentzu horretan, Widdows et al.-ek (1984) diotenez,xenobiotikoek zelula basofilikoen kopuruaren igoera eragingo lukete, estres-egoerei aurreegiteko zelula hauek ekoitzitako entzimen eskari handiagoa bailego.
Estres kronikoa pairatu duten bareetan ere, kaltzio-zelulen kopuru erlatiboren balioaltuak ikusi dira. Esaterako, kupre-meatzaldean bizi diren bareen liseri-guruinekoepitelioan zelula-mota nagusia kaltzio-zelula dugu, liseriketa-jarduerei zein kaltzioarenmetabolismoari dagokienez liseri-guruina berezia izanik (Marigómez et al., 1998b).
1.4.3. Liseri-guruineko epitelioko zelulen plastizitatea
Lehenago aipatu den bezala, liseri-zelulak oso sistema endo-lisosomikoa garatuadauka, lisosomek zelularen bolumen gehientsua betetzen dutelarik (Marigómez & Baybay-Villacorta, 2003). Lisosomen gainean estres-iturri desberdinek eragina izan dezakete,kutsadura kimikoak, gazitasun-estresak, muturreko tenperaturek altua, baraualdiak etaugalketari asoziaturiko estres-egoerak, besteak beste (Lowe et al., 1981; Moore et al.,1987; Lowe, 1988; Cajaraville et al., 1991; 1995a; Marigómez et al., 1991, 1996; Regoli,1992; Krishnakumar et al., 1995; Domouhtsidou & Dimitriadis, 2001). Oro har, liseri-zelulenlisosometan gerta daitezkeen aldaketak hiru taldetan sailka daitezke: tamainazemendatzea, mintza desegonkortzea eta edukinen izaera aldatzea (Marigómez et al.,2005). Lisosomen erantzunak berehalakoak dira, eta zenbait kasutan tamainazemendatzea eta mintza desegonkortzea kitzikadura jaso eta ordu gutxitara dagoeneko
Moluskuen liseri-guruina
23

behatu dira (Lekube et al., 2000; Izagirre et al., 2005). Hori dela eta, lisosomen erantzunakestres-biomarkatzaile goiztiar gisa kontsideratu dira (Cajaraville et al., 2000; Marigómez &Baybay-Villacorta, 2003; Marigómez et al., 2005).
Dena dela, lisosomen erantzunak bai kutsatzaile-motaren arabera, bai esposizio-denboraren arabera eta baita esposatutako kontzentrazioaren arabera desberdinak izandaitezke. Metalen pean egondako muskuilu eta magurioen liseri-zelulen lisosomendentsitate bolumetrikoaren emendatzen den bitartean (Marigómez et al., 1990), kutsatzaileorganikoen pean egondakoen kasuan, kutsatzailearen izaera, esposizio-denbora etakontzentrazioaren arabera erantzun ezberdinak behatu dira. Esaterako, petrolioarenhidrokarburo eta azetonaren pean egondako muskuiluetan, hasiera batean lisosomaktxikiagoak diren arren (Cajaraville et al., 1995b; Cancio et al., 1998, Marigómez & Baybay-Villacorta, 2003), esposizio luzeagoak tamainaz emendatzea dakarkie (Cajaraville et al.,1995b). Aitzitik, di(2-etilhexil)ftalato gisako zenbait kutsatzaile organiko pean egoteak,hasiera-hasieratik lisosomak tamainaz emendatzea eragin dezake (Marigómez & Baybay-Villacorta, 2003).
Azkenik, zelula basofilikoei dagokienez, aipatutako kopuru erlatiboaren emendioagertatzeaz gain, zelulen morfologia eta ultrastrukturan ere aldaketak aurkitu dira(Marigómez et al., 1998b; Triebskorn & Kohler, 1996). Kadmio kontzentrazio subletalenpean egondako magurioetan, zelula basofilikoen hipertrofia eta basofiliaren galera behatudira, eraldatutako zelula basofiliko hauek liseri-zelula eta zelula basofilikoen artekomorfologia erakutsi dutelarik (Marigómez et al., 1990). Ingurumen kutsatuetanmantendutako Mizuhopecten yessoensis bibalbioaren liseri-epitelioan ere, tarteko itxurakozelula basofilikoak deskribatu dira (Syasina et al., 1997).
Zati Teorikoa
24

2.- EPITELIO-EHUNEN BERRIZTAPENA
Zelula eukariotikoak, jatorriz aske bizi ziren banakako izakiak izan arren eboluzioanzehar izaki zelulanitzen partaide espezializatuak bilakatuz joan dira. Hori dela eta, moduindependentean bizitzeko ezinbestekoak ziren ezaugarriak galduz joan ziren, izakizelulanitzak beren osotasunean bizirik mantentzeko derrigorrezko ezaugarri berriakeskuratuz joan ziren bitartean. Izaki zelulanitza osatzen duten zelulak, nahiz eta genomabera partekatu, elkarren artean arras desberdinak dira. Zelulek, aniztasun zabalekozereginak buru ditzaketen organoetan antolatutako hamaika ehun desberdin itxuratzekoelkarlanari ekiten diote. Ehunak eta organoak behar bezala ulertzeko, zelulen lan egitekomodua eta habitat desberdinetan nola bizi eta hiltzen diren ezagutzea beharrezkoa da(Alberts et al., 2002).
Epitelio-ehuna, animaliak osatzen dituzten lau oinarrizko ehun-motetako bat dugu.Oro har, bi era desberdinetan antolatuta dago, zelulen xafla jarrai moduan edo guruinmoduan. Epitelio-ehunek, oso funtzio desberdinak burutzen dituzte, babespena, zelulenarteko garraioa, xurgapena, jariapena eta iragazkortasuna, besteen artean. Epitelioensorrera eta mantentzearen inguruan oinarrizko hainbat galdera erantzuteke daude oraindikere. Zenbait epiteliok, hestea edo larruazala kasu, zelulen ordezkatze-tasa handia erakutsidute (Potten & Loeffler, 1990), beste zenbaitek, ordea, gibela edo area kasu, egoeranormalean ordezkatze-tasa geldoagoa dute, berriztapen azkarrak behar izatekotanmoldaera bereziak erakutsi dituzten arren (Finegood et al., 1995; Slack, 1995; Alison et al.,1997).
Hurrengo orrialdeetan azalduko diren barne-faktore dibertsoen eraginaz gain,farmakoek, gaixotasunek, ebakuntza kirurgikoek eta ingurumen-faktoreek ere epitelioenberriztapen-tasa eraendu dezakete. Euren artean, argia/iluntasun zikloa faktorenagusitakoa omen da, askotan zelulen proliferazioaren dinamikaren erritmoak eta zikloakpatroi zirkadiarren arabera gauzatzen baitira. Adibidez, hainbat zianobakterio etaprotistotan, DNAren sintesia gauez baino ez da ematen. Hori, babes-mekanismo moduaninterpretatu da, egunez erradiazio ultramoreak DNAren kopia akastunen ekoizpena bultzalezakeelako (Reddy et al., 2005). Animalien artean ere, antzeko portaerak behatu dira, baiarrainen epidermisean (Dekens et al., 2003) eta baita ugaztunen hezur-muinean(Abrahamsen et al., 1997) zein bestelako epitelio-ehunetan ere (Bjarnason & Jordan,2002; Barbeito et al., 2003; Brandi et al., 2004).
Epitelio-ehunen berriztapena
25

Atal honetan, epitelio desberdinetan zelulen berriztapena zein mekanismorenbitartez suertatzen den aurkeztuko dugu. Lehendabizi, sarrera gisa, zelula amenkontzeptuaren inguruan orokortasun batzuk eta zelulen proliferazioa aztertzeko teknikaeskuragarriak azaldu dira laburki. Gerora, epitelioen berriztapenaren estrategiadesberdinak aurkeztu dira, bi estrategia bereiziz: (a) egoera normaletan zelula amenbitartez berriztatzen diren epitelioak; eta (b) nahiz eta bertan ere zelula amak egon,normalean desberdintzatutako zelula helduen bitartez berriztatzen diren epitelioak.
2.1. Epitelioetako zelula amak
Duela 40 urte inguru, ehun bereko bestelako zelula-motak emateko gai zirenzenbait zelula somatiko topatu ziren, zelula amak alegia. Zelula somatiko horiek,lehenengo aldiz saguen ehun hematopoietikoan deskribatu ziren (Till & McCullog, 1961).Azken urteotan, zelula amen presentzia bestelako ehun-mota desberdinetan eredemostratu da, epidermisean, muskuluan, gibelean eta burmuinean, besteak beste (Gage,2000; Slack, 2000). Epitelio gehienek, zelula amak dituzte (Slack, 2000). Zelula amakdefinitu nahian, modu funtzionalean egitera behartuta gaude; kasu gehienetan epiteliokogainontzeko zelulekiko inolako desberdintasun morfologikorik ez baitute. Zelula amakdeskribatzen dituzten ezaugarri funtzionalak honako hauek ditugu (Alberts et al., 2002):
1.- Guztiz desberdintzatu gabeko zelulak dira.2.- Mugarik gabe zati daitezke edo, behintzat, organismoak bizirik dirauen bitartean
zatitzeko gaitasuna manten dezakete.3.- Zatiketa zelularra burutu ostean, zelula kume bakoitzak bi aukera ditu; zelula
ama izaten jarraitu edo eta derberdintzapen-bidea bukaeraraino eraman.Epitelio bateko zelula-mota desberdin guztiak sortzeko gaitasuna
(multipotentzialitatea) zelula amak identifikatzeko sarritan erabili den ezaugarria da.Tamalez, gehienetan, gaitasun hori ehuna kaltetu denean baino ez da azaltzen. Hau da,egoera normalean zelula ama gehienek desberdintzatutako zelula-mota bakarraproduzitzen dute (unipotentzialitatea). Gibelaren kasuan, adibidez, hepatozitoenberriztapenaz hepatozitoak beraiek arduratzen dira (Michalopoulos & DeFrances, 1997),baina, edozein arrazoia dela medio, hepatozitoen zatiketa inhibituta baldin balego,hepatozitoen berriztapena konduktuetako zelulek burutuko lukete (Alison et al., 1997).
Zatiketa zelularraren zinetika ikertu ondoren, zelula amek orokorrean bikoizpen-tasa baxua dutela ondorioztatu da, espero ez bezala. Izatez, epitelio-ehunetan gehienzatitzen diren zelulak zelula anplifikatzaile iragankorrak edo TAC (Transit Amplifying Cell)
Zati Teorikoa
26

deitutakoak dira. TAC zelulak hainbat zatiketa-ziklo pairatu eta gero, desberdintzapen-prozesuan sar daitezke. Euren kopurua altua denean, ehunak desberdintzapenerakogaitasun handia duela ikusi da, zelula ama urriak izan arren (Hall & Watt, 1989).
Oro har, ehunetan zatitzen diren zelulak, zelula amak zein TAC zelulak, tokiberezietan daude kokaturik. Esaterako, hestearen kasuan zelula amak Lieberkühn kriptenzati distaletik gertu kokatuta dauden bitartean, TAC zelulak kriptaren altueraren bi herendistaletan paratuta daude, eta desberdintzatutako zelulak, aldiz, batez ere herenproximalean kokaturik agertu ohi dira (Potten & Loeffler, 1990).
2.2. Zelulen proliferazioa aztertzeko teknikak
Ziklo zelularraren kontzeptua, 1950. hamarkadaren inguruan plazaratu zenarratoien barrabiletan autorradiografiazko teknikak erabiliz 32P-aren inkorporazioa DNAnbehatu ondoren (Howard & Pelc, 1950). Ikerlan horren ondorioz, ziklo zelularra 4 fasetanbanandu zen G1-, S-, G2- eta M-faseak, alegia. Lau fase horietatik deigarriena, etaikuspuntu morfologiko batetik interesgarriena, M-fasea (mitosia) da. Bertan, nukleoa zatituegiten da, eta zelula batetik bi zelula kume sortzen dira. Mitosia gertatu eta gero, zelulakumeak, G1-fasean edo lehenengo hazkuntza-fasean sartzen dira. Bertan, zelula sortuberriak bere ingurua aztertzen du, eta hazteari ekiten dio. Tamaina nahikoa lortu eta gero,eta seinale egokia jasoz gero, S-fasean (sintesia) sartzen da. Bertan nukleoko DNAbikoiztu egiten da, eta, DNA zeharo erreplikatuta egon ondoren, zelula G2-fasean sartzenda. Bertan, zelulak DNA guztiz erreplikatu dela egiaztatzen du, berriro ere hurrengo M-fasean sartu aurretik. G1-fasean dagoen zelulak bere ziklo normala geldiarazi dezake, etaespezializatutako atsedenaldian sar daiteke (G0-fasea). Bertan, asteak, hilabeteak edourteak egon daiteke S-fasean berriro sartu arte, beharrezkoa den seinale espezifikoajasotzearen zain. (Alberts et al.,2002).
Ziklo zelularra eraentzen duen kontrol-sistema molekularra, eboluzioan zehar osokontserbatua da. Esaterako, zelula eukariotiko guztietan zatiketaren kontrolean antzekoproteina eraentzaileek dihardute (Hall & Levinson, 1990). Zelulen proliferazioa ehunetanikertzeko, proteina horien eta beren ekoizpenaren azterketaz baliatzen diren teknikadesberdinak eskuragarri daude. Euren artean erabilienak, fluxu-zitometria, nukleotidoenanalogoak erabileran oinarrituriko teknikak, immunohisto(zito)kimika eta geneenadierazpenaren azterketa molekularra (in situ hibridazioa, PCR...) dira. Moluskuetan zelulaproliferatzaileak detektatzeko teknikak oso gutxitan erabili dira (1. Taula). Izatez, nahiz etaPCNA edo Ki67 moduko proteinak eboluzioan zehar kontserbatuak izan (Prelich et al.,
Epitelio-ehunen berriztapena
27

1987; Endl & Gerdes, 2000), moluskuen ehunetan zelula proliferatzaileen identifikazioanukleotidoen analogoen erabileran oinarritu da batik bat.
2.2.1.- Fluxu-zitometria
Teknika honetan, zelulak seinale batez markatzen dira. Seinaleak, itu-proteina batespezifikoki ezagutzen duen antigorputzen bat zein azido nukleiko berezia ezagutzen duenzunda osagarriren bat izan daitezke. Zelulen esekidura batean zenbait zelula espezifikokihorrela markatu ondoren, laser-izpi batek zeharkatzen duen zulo batetik zelulen esekidurapasarazten da. Orduan, laser-izpiok zeluletan transmititu edo islatu egingo dira, zelulekseinalea daramatenentz arabera. Ondorioz, argia transmititu edo islatu den araberazelulak karga elektrostatiko desberdinez hornituko dira, eta eremu elektromagnetikobatean zehar pasarazi ondoren banandu daitezke (Alberts et al., 2002). Teknika honenbitartez, zelulen kopurua eta zelula bakoitzak duen DNA kopurua ezagutu daitezke. Izanere, aneuploidiarik ez egotekotan, G0- eta G1-fasean dauden zelulak diploideak izangodira, G2-fasean dauden zelulak tetraploideak eta S-fasean daudenek tarteko DNA-kopuruaizango dute. Beraz, zelulen esekidurak erabiliz eta modelo matematiko egokiak aplikatuz,zatiketa-zinetika ere iker daiteke (Baisch et al., 1982). Zenbait kasutan, histologiarakofixatutako eta parafinan inkluditutako zeluletan ere DNA kopurua kalkula daiteke, laginakdesparafinatu eta hidratatu ondoren (Hedley et al., 1983). Fluxu-zitometria, egoerapatologiko desberdinak aztertzeko erabili da, eta bere bidez lortutako proliferazio-indizeektumoreen garapenaren azterketarako ere balio prognostikoa dutela proposatu da (Joensuuet al., 1988). Fluxu-zitometriak, zelula proliferatzaileak detektatzeko bestelako teknikenaurrean daukan abantaila handienetakoa emaitzen objektibotasuna dugu. Gainera, zelula-kopuru handiekin lan egin daiteke, eta interesatzen zaizkigun zelulak isolatzeaahalbidetzen du. Bestalde, beharrezkoa den ekipamendua oso garestia da, eta laginasakabanatu eta desegituratu behar denez, zelula proliferatzaileen kokapena ehunean ezdago aztertzerik. Gainera, zelula proliferatzaileen azpipopulazioak ere galtzen dira.
2.2.2- Nukleotidoen analogoen erabileran oinarrituriko teknikak
3H timidinaren (3H-T) bitartezko markaketa
Zelulen zinetika ehunetan zuzenean aztertzeko, 1950. hamarkadan 3H timidinatritiatuaren (3H-T) bitarteko markaketa, teknika estandarrena bilakatu zen. Zelula
Zati Teorikoa
28

bideragarriek 3H-T DNAn eransten dute S-fasean daudenean (17. Ird.), eta ondorentimidina hori autorradiografia bitartez ikuskor bihur daiteke. Timidina erradioaktiboa,animaliari edo zelulen kultiboari in vivo ematen zaie (3H-T-ren pultsua), eta DNAsintetizatzen ari diren zelulek markatutako timidina beraien DNAn erantsiko dute. Halaber,biopsiaz lortutako materiala eta bestelako lagin-motak aztertzeko, fixapena burutu aurretikdenbora-tarte luzez timidinaz inkubatu behar dira. In vivo pultsuak burutzeko zailtasuna,batetik, zein material erradiaktiboa erabili beharra, bestetik, ezaugarri mugatzaileak izanik,azkeneko urteotan 3H-T gero eta gutxiago erabili da histopatologian. Edozein kasutan,zenbait ikerketa berezi egiteko edo eta ikerkuntz-zentro espezializatuetan teknika hau gauregun oraindik ere erabilia da (Hale et al., 2003).
Bromodeoxiuridinaren (BrdU) bitartezko markaketa
BrdU, timidinaren analogoa den molekula da, eta horregatik DNA sintetizatzen denfasean (S-fasea) nukleoan inkorporatzen da 3H-T moduan (17. Ird.). BrdU inkorporatuduten zelulak immunohisto(zito)kimika bitartez detekta daitezke, BrdUren aurkako
Epitelio-ehunen berriztapena
29
17. Irudia: Ziklo zelularraren fasearen arabera adierazten diren markatzaile molekular desberdinak

antigorputzak erabiliz, eta mikroskopioan aztertu (18. Ird.; Gratzner 1982; Alison, 1995).BrdU immunohisto(zito)kimika S-fasean dauden zelulak identifikatzeko oso erabilgarria da.Kasu gehienetan antigorputzarekiko eskuragarritasuna emendatzeko DNA desnaturalizatubeharra dago, azido klorhidrikoaz tratatuz edo liseriketa entzimatikoaren bidez (Gratzner etal., 1982; Roberts et al., 1985; Kikuyama et al., 1988). BrdU, injekzio bitartez emandakopultsu moduan edo etengabe ponpa peristaltikoak erabiliz eman daiteke. Gaur egun,teknika hau metodo estandarra bilakatu da, eta horren ondorioz ikerlan desberdinetan
Zati Teorikoa
30
18. Irudia: Zelula proliferatzaileak detektatzeko erabil daitezkeen teknika desberdinek arratoien hestegorrianeman dituzten irudiak. (A) Ki67 immunohistokimika; (B) BrdU immunohistokimika (C) PCNA immunohistokimika; (D)H2b, H3 eta H4 histonen mRNAren in situ hibridazioa. (Muskhelishvili et al.-etik (2003) birmoldatua).

lortutako emaitzak konparagarriak izatea lortu da. Hala ere, metodo honek beredesabantailak ere badauzka: (1) zenbait kasutan animalia injektatu beharra dago eta berazbaimen bereziak ezinbestekoak dira, (2) BrdU S-fasean eransteaz gain DNAkonpontzerakoan ere DNAri eransten zaio, zelula proliferatzaileen gainestimazioa sorlezakeena; eta (3) BrdU mutagenoa denez, epe luzeko ikerketa toxikologikoetan ez da osokomenigarria.
2.2.3- Antigeno nuklearren immunohistokimikan oinarrituriko teknikak
Ki67 antigenoaren immunohistokimika
Ki67 proteina nuklearra zelula proliferatzaileak detektatzeko oso antigeno erabiliadugu. Ki67 zelula-mota guztietan G1-, S- eta G2-faseetan adierazten den bitartean, G0-fasean ez da adierazten (17. Irudia; Gerdes et al., 1984; Gerlach et al., 1997). Hori delaeta, ehunaren hazkuntza-mailaren neurketetarako erabil daiteke. Hala ere, momentuz berefuntzioa zein den argitzeke dago (Endl & Gerdes, 2000). Minbiziaren ikerkuntzan zeinbestelako gaixotasunen diagnostikoan Ki67 antigenoaren immunohistokimika oso erabiliaizan da (18. Ird.; Scholzen & Gerdes, 2000). Hala ere, proteina honen adierazpenaren etaziklo zelularraren barruan duen kokapenaren inguruan eztabaida ugari dago, eta baitaproteinaren biziraupenaz ere (Littleton et al., 1991; Bruno & Darzynkiewicz, 1992;Goldblum & Appelman, 1995; Oka & Arai, 1996; Scholzen & Gerdes, 2000). Antigenohorren desabantailarik garrantzitsuena, antigenoak fixatzaileekiko daukan sentikortasunhandia da, kasu gehienetan izoztutako materiala erabiltzera bultzatzen duena, informaziomorfologikoaren galera ekarriz (Endl & Gerdes, 2000). Gainera, giza-antigenoaren aurkakoantigorputza erabili da gehienetan, hainbat espezieekiko erreakzionagarritasun gurutzatuabaxu samarra edo nulua izanik. Zorionez, arazo horri irtenbidea emateko asmoz,bakteriotan adierazitako gizakiaren Ki67 antigenoaren aurkako antigorputz berezia garatuda, MIB-5 izenaz ezagutua dena (Schluter et al., 1993; Gerlach et al., 1997). Izan ere, MIB-5 antigorputzak, karraskarien Ki67 antigenoarekiko erreakzionagarritasun gurutzatu osoaltua du, fixatzaileekiko horren sentikorra ez izanik (Gerlach et al., 1997).
Epitelio-ehunen berriztapena
31

PCNA (Proliferating Cell Nuclear Antigen) antigenoaren immunohistokimika
PCNA, δ eta ε DNA-polimerasen zein DNAren sintesirako beharrezkoak direnhainbat entzimen proteina laguntzailea dugu (Kurki et al., 1986; Bravo et al., 1987; Wood& Shivji, 1997). PCNA, Miyachi-ren taldeak 1978. urtean lehenengo aldiz deskribatu zuen(Miyachi et al., 1978). PCNAren adierazpena G1-fasearen bukaera aldera emendatzen da,eta S-fasean zehar lortzen du bere adierazpen mailarik altuena. Ondoren, G2- eta M-faseetan adierazpena berriro ere jaisten da (17. Ird.). Berezitasun hauek direla eta, ziklozelularraren fase desberdinak identifikatzea ahalbide dezake (Foley et al., 1993). PCNArenpisu molekularra 36 kDa-koa da, eta immunofluoreszentzia erabiliz bi azpipopulaziobereiztu dira. Lehenengoa, soilik DNAren erreplikapen-guneetan dago, eta DNA ekoiztenden bitartean bere kontzentrazioa igo egiten da. Bigarrena, berriz, nukleoplasmansakabanatuta dago, eta ziklo zelularrean zehar beti, gutxi gorabehera, maila beretsuanadierazten da (Bravo et al., 1987). Bi azpipopulazio horiek, fixatzaileekiko sentikortasundesberdina daukate. Fixatzaile aldehidikoz fixatutako ehunetan bi azpipopulazioakdetektatzen diren bitartean, fixatzaile alkoholikoak erabiltzen direnean soilik DNArenerreplikapen-guneetan dagoen PCNA detektatzen da (PCNA osoaren %20-30-a).PCNAren biziraupena 20 ordukoa da (Bravo & MacDonald., 1987). Nahiz eta PCNAzmarkatutako zelula proliferatzaileen indizeak bestelako teknikekin lortutako indizeekinkonparatzeko arazoak egon diren (Jain et al., 1991; Yu et al., 1991; Visakorpi, 1992; Sarliet al., 1995), PCNA immunohisto(zito)kimika oso erabilia izan da, bai oinarrizko ikerketa
Zati Teorikoa
32
19. Irudia: Mercenaria mercenaria bibalbioaren zelula proliferatzaileen markaketa PCNA immunohistokimikaerabiliz. Geziek marka positiboa adierazten dute eta gezi-buruek nukleo negatiboak. (A) Liseri-guruineko epiteliokozelula proliferatzaileak. B) Zelula proliferatzaileen marka bortitza zakatzen oinaldeko epitelioan. (C) PCNA-markaketapositiboa giltzurruneko granulometan murgildutako hemozitoetan. Eskala marra: 13 μm. (Hanselman et al.-etik (2000)birmoldatua).

egiteko eta baita pronostiko patologikoetan ere (18. Ird.; Faderl et al., 2002; Lillo et al.,2002). Hala ere, zelula proliferatzaileak detektatzeko PCNAk duen baliogarritasunakhainbat zalantza sortu ditu. Izan ere, PCNAren kopurua zelula-moten zein zelulen egoerafisiologiko edo patologikoen arabera esanguratsuki alda daiteke. Beraz, minbizi-zelulak etazelula arruntak bereiztu nahi izan direnean, ez da erreza izan zelulen egoera bakoitzarizegozkion PCNA-markaketaren balioak finkatzea (Morris & Mathews, 1989; Hall et al.,1990; Scholzen & Gerdes, 2000). Gainera, PCNAk DNAren konponketan ere parte hardezake, bere adierazpena bikoizten ari ez diren zeluletan ere beha daitekeelarik (Toschi &Bravo, 1988; Wood & Shivji, 1997).
PCNA, eboluzioan zehar nahiko kontserbatua den proteina dugu (Prelich et al.,1987). Garatutako antigorputz monoklonal komertzialek, batez ere PC10 izenaz ezagutzendenak, erreakzionagarritasun gurutzatu sendoa erakutsi dute (19. Ird.), ugaztun, intsektu,landare, molusku zein legamien PCNA antigenoekin (1 Taula; Waseem & Lane, 1990;González-Melendi et al., 1996; Marigómez et al., 1999; Hanselman et al., 2000).
Ziklinen menpeko kinasen immunohistokimika
Zelulen proliferazioa, ziklo zelularreko fase espezifikoetan proteina-kinasakonplexu desberdinek eraenduta dago. Konplexu horiek, azpiunitate katalitiko eta berarilotutako azpiunitate eraentzaileaz osaturik daude. Proteina-kinasa konplexuen azpiunitatekatalitikoak ziklinen menpeko kinasak (zmk) dira, eta ziklo zelularrean aurrera egiteareneraentzaile nagusiak dira. Kinasa horien jarduera, ziklo zelularrean zehar igo eta jaitsiegiten da, eta gorabehera horiek ziklo zelularraren gertaera nagusietan (DNArenerreplikapena, mitosia, zitokinesia) gertatzen diren proteina ezberdinen fosforilazioaneragina daukate (Morgan, 1997). Adibidez, zmk jakin baten jarduera mitosiaren hasieranigoz gero, kromosomen kondentsazioan edo mintz nuklearraren apurketan parte hartzenduten proteinen fosforilazioen emendioa bultzatuko litzateke. Hainbat zmk desberdindaude, aipagarrienak zmk-1, zmk-2, zmk-4 eta zmk-6 izanik. Ikerlan aplikatu zeinoinarrizko gehienek, lau zmk horiek izan dituzte aztergai (Palacios et al., 2005;Schmetsdorf et al., 2005).
Zmk aurkako antigorputzei dagokienez, zmk-1 delakoaren PSTAIRE deitutakogunea ezagutzen duten antigorputzak garatu dira. Gune hori kontserbatua da, etaugaztunetan gain beste espezie batzuetan antigorputz espezifikoetako elkargarritasungurutzatua frogatu da (Weinstein et al., 1994). Beste zmk aurkako antigorputzeidagokienez, ugaztunetaz kanpo apenas informaziorik dago.
Epitelio-ehunen berriztapena
33

Ziklinen immunohistokimika
Proteina hauek, aurrenez aipatutako ziklinen menpeko kinasekin erlazio zuzenadute. Izan ere, ziklinak zmk-konplexuaren azpiunitate eraentzaileak dira. Ziklinak, ziklozelular bakoitzean sintetizatu eta andeatu egiten dira. Kinasa jarduera, ziklinak zmkeiestuki lotzen zaizkienean baino ez da adierazten. Hainbat ziklina-mota desberdin daude,garrantzitsuenak A-, B-, D- eta E-ziklinak izanik (20. Ird.). Ziklo zelularrak aurrera jarraidezan, D-ziklina zmk4 eta zmk6-ri G1-fasean zehar lotu behar zaie (Sherr, 1996). E-ziklinak, zmk2 edo zmk4-ri lotuz, aktibatu egiten ditu, ziklo zelularra G1-fasetik S-faserapasaraziz. A-ziklina, bere aldetik, zmk2-ri S-fasean lotzen zaio, DNAren sintesiaahalbidetuz; eta B ziklina, bestetik, zmk1-ri lotzen zaio, mitosiari bide emanez (Nigg, 1995;Edgar & Lehener, 1996). Ziklinen adierazpena, immunohistokimika bitartez azter daiteke,minbiziarekin erlazionatutako kasuetan (Zhu et al., 2003) zein oinarrizko ikerketetan(Golias et al., 2004) jadanik egin den bezalaxe. Hala eta guztiz ere, ugaztunetatik kanpoziklinei buruzko ikerlan immunohistokimiko urriak burutu dira (1 Taula).
2.2.4.- Geneen adierazpenaren azterketan oinarrituriko teknikak
Aldez aurretik ikusi dugunez, ziklo zelularra eraentzen duten zenbait proteina uneedo fase jakinetan baino ez dira sintetizatzen zikloan zehar, bereziki histonen eta ziklinenkasuan alegia (17. Ird.). Proteina horien adierazpena, antigorputzak erabiliz iker daiteke.
Zati Teorikoa
34
20. Irudia: Ziklina desberdinen adierazpen-mailak ziklo zelularraren fase desberdinetan. Eskuarki ziklina-motabakoitzak bere adierazpen-piko maximoa ziklo zelularraren fase jakin batean lortzen du.

Izan ere, beren funtzioa bete bezain pronto antigenoak andeatu egiten direnez, ziklinenkasu, oso informazio baliagarria ematen dute ziklo zelularraren fasea identifikatu ahalizateko. Esaterako, A-ziklina S-fasearen bukaera-aldera eta G2-fasean zehar soilik topadezakegu, eta B-ziklina, bestalde, G2-fasearen bukaera-aldera eta M-fasean zehar bainoez (20. Ird.). Aldiz, kromatinaren proteina estrukturalak diren histonak oso proteinaegonkorrak dira, sintetizatu eta gero zelularen ziklo osoan zehar aurki ditzakegularik.Adierazpen iragankorra duten ziklinen kasuan zein adierazpen iraunkorra duten histonenkasuan, proteina horiek ekoizten dituzten RNA mezularien presentzia, proteinaksintetizatzen diren une jakinetan emendatzen dira, gerora maila basaletara itzuliz (17. Ird.).Hau da, histonak egonkorrak diren arren, beren geneak S-fasean zehar soiliktranskribatzen dira. Beraz, zelularen batean histonen RNA mezulariak topatzekotan,zalantzarik gabe zelula hori S-fasean dagoena badakigu. Halaber, ziklinen kasuan, ziklinadesberdinen RNA mezularien sintesia ziklina bakoitza topatu dugun baino aurrenekofasean zehar areagotuz doa. Adibidez, B-ziklinaren genearen transkribapena G2-fasean
Epitelio-ehunen berriztapena
35
Teknika Espeziea Organoa Erreferentzia 3H timidina Achatina fulica Bihotz-muskulua Martynova & Bystrova, 2002 Crenomytilus grayanus Liseri-traktua Leibson & Frolova, 1994 Aplysia californica Nerbio-sistema Hickmott & Carew, 1991 Margaritifera
margaritifera Zakatza Tomasovic & Mix, 1974
BrdU Planorbarius corneus Mikrogliako zelulen kultiboa
Peruzzi & Sonetti, 2004
Sepia officinalis Enbrioia Grimaldi et al., 2004 Melampus bidentatus Garroen nerbio-
sistema Bale et al., 2001
Pinctada fucata marte nsii Mantu paliala Awaji & Suzuki, 1998 Helix lucorum Protozerebroa Zakharov et al., 1998 Incilaria fruhstorferi Hemozeleko
zelulak Furuta et al., 1994
Mytilus galloprovincialis Zakatzak Martínez-Expósito et al., 1994 Ki67 Helix pomatia Listu-guruina Pirger et al., 2004 PCNA Mercenaria mercenaria Liseri-guruina,
zakatzak, hemozitoak
Hanselman et al., 2000
Mytilus galloprovincialis Liseri-guruina Marigómez et al., 1999 Zmk Aplysia californica Begia Sankrithi & Eskin, 1999 Bulla gouldiana Begia Krucher & Roberts, 1994 Ziklinak Pecten maximus Liseri-guruina Le Pennec & Le Pennec, 2002 Dreissena polymorpha Gonada arra Lamers et al., 1999 Patella vulgata Enbrioia Colas et al., 1993a Patella vulgata Oozitoak Colas et al., 1993b Spisula soldis sima Enbrioia Hunt et al., 1992 Patella vulgata Oozitoak Van Loon et al., 1991
1. Taula: Moluskuen ehunetan zelula proliferatzaileak detektatzeko erabili diren teknika desberdinen adibideak.

zehar areagotuz doa, M-fasearen hasiera-hasieran genearen adierazpenik altuena ematendelarik.
RNA mezulari horiek PCR kuantitatiboaren bidez kuantifika daitezke, DNA-hasleegokiak erabiliz. Era berean, ehunaren gainean molekula horiek lokalizatzeko in situhibridazioa erabili ahal da, RNA- zein DNA-zunda osagarriak erabiliz. DNA-hasle egokiakdiseinatzeko sekuentziaren ezagutza partziala ezinbestekoa da; DNA- eta RNA-zundakdiseinatzeko, bestalde, sekuentzia nukleotidikoaren ahalik eta zatirik handiena ezagutubeharra dago. Proteina homologoek espezien artean nolabaiteko kontserbazio-maila duteneta eboluzioan zehar egonkorrak izan daitezkeena jakina da. Haatik, hau ez da egia RNA-zunden edota DNA-hasleen kasuan, espezie desberdinen proteina beraren sekuentzianukleotidikoak oso desberdinak izan baitaitezke. Sekuentzia nukleotidikoek sekuentziaaminoazidikoak baino aldakortasun handiagoa erakusten dute espezien artean, kodegenetikoaren endekapenaren ondorioz (aminoazido askotarako 4 kodon ezberdinbaitaude), hain zuzen ere. Hau da, antigorputzak erabiliz proteina bera espeziedesberdinetan identifika daitekeen arren, RNA-zundek ez dute ezertarako baliogarriak izanbehar espezie desberdinetan proteina jakin baten RNA mezulariak identifikatzeko. Beraz,edozein kasutan, ikertu nahi den espeziearen sekuentzia nukleotidikoaren ezagutzaminimoa ezinbestekoa da.
Moluskuen kasuan, histonen hainbat mRNA sekuentzia geneen datu-baseetanargitaratu dira, 320 sekuentziatik gora daude eskuragarri (http://www.ncbi.nlm.nih.gov),hain zuzen ere. Taldez-talde, eta tesi honetan aztertuko diren espeziei erreparatuz, M.galloprovincialis-en histona guztien sekuentzia osoak argitaratu dira (Eirin-Lopez et al.,2002), eta beste mitilido askorenak ere (M. edulis, M. trossulus, M. californianus eta M.chilensis). A. ater eta L. littorea gastropodoen histonen sekuentziarik ez dugu ezagutzen,ordea, guztira pulmonatuen 35 sekuentzia desberdin eta orthogastropodoen 78 sekuentziadesberdin argitaratu diren arren. Oraintsu, muskuiluaren histonen sekuentziak 50 genedesberdineko microarray espezifiko batean erabili dira (Dondero & Viarengo, 2005).
Moluskuen ziklinen 18 sekuentzia desberdin ere argitaratuak izan dira. Hala ere, ezda Mytilus, Arion edo Littorina generoen sekuentziarik argitaratu. Argitaratutakosekuentzien artean Dreissenia polymorpha bibalbioaren A-, B-, C- eta D-ziklinen RNAmezularien sekuentzia osoak (Lamers et al., 1999) topa ditzakegu. Halaber, beste bibibalbioen sekuentziak identifikatu dira, hala nola Spissula solidissimaren A- eta B-ziklinaketa Crasssostrea virginicaren zenbait ziklinen antzekotasun handia duten sekuentzianukleotidiko laburrak edo EST-ak (ingeleraz, Expression Sequence Tags; CD646624 etaCD647240 A-ziklina; CD646450 eta 646472 B-ziklina, CD647301 C-ziklina; CD648974 D2-
Zati Teorikoa
36

ziklina eta CD649103 G1-ziklina; ikus NCBI web orrialdea). Crassostrearen sekuentziakgainera merkurio pean mantendutako animalien eta animali kontrolen transkriptomakkonparatuz aurkitu dira; proliferazio zelularra modulatuz ziklina horiek merkurio peanmantendutako ostretan gainadierazi edo azpiadierazi egin baitira (Peatman et al.,argitaratu gabe, ikus NCBI web orrialdea).
PCNA eta Ki67 sekuentziarik ez da klonatu orain arte. Hala ere, M.galloprovincialis-en 435 base paretako EST bat argitaratu da (AJ516167; Venier et al.,2003, ikus NCBI web orrialdea), zeinen sekuentzia aminoazidikoak Danio rerio etaIctalurus punctatus arrainen PCNAren sekuentziekiko homologia (homologia: 1e-29) altuabaitu.
2.3.- Zelula amen bitartez berriztaturiko epitelioak
2.3.1.- Ugaztunen epidermisa
Epidermisa, jatorri ektodermikoa duen larruazalaren oinarrizko epitelio-osagaia da.Zuzenean eta jarraian kanpo-medioarekin kontaktuan egotearen eragina jasotzen duenez,epitelioaren berriztapenak, hots zelulen proliferazioak, ikaragarrizko garrantzia du. Horidela eta, ugaztunen epidermisa organismo helduetan epitelio-ehunak nola berriztatzendiren ikertzeko ohiko eredu esperimentala izanez, bertako proliferazio zelularrarendinamika eta mekanismoak aski ezagunak dira.
Ugaztunen epidermisa, keratinozitoz osatutako epitelio geruzatua da.Keratinozitoek, epidermisari sendotasuna damaion keratina deitutako proteina ekoiztendute. Keratinozitoak epitelioaren oinaldetik erpinalderantz desberdintzatuz doaz, eta goraegiten duten heinean osaketaz eta itxuraz aldatzen dira. Xafla basalaren gainean ezarritadauden zelulak oinaldeko zelulak dira. Zelula hauek zatitzeko ahalmena dute, geruzagerminatiboa konfiguratuz. Euren gainean desmosometan joriak izateari eskerepidermisari trinkotasun mekanikoa eskaintzen dioten zelula handiagoak ditugu (Malpighizelulak). Desmosomen inguruan sortutako arantza itxurako egitura nabarmenakedukitzegatik, zelula hauek osatzen duten epitelioaren zatiari geruza arantzatsu izenaeman zaio, zenbait autorek Malpighi geruza ere deitu duten arren. Hurrengo mailan,geruza arantzatsuaren gainean paratuta, pikor zitoplasmatiko elektrodentsoz hornitutakozelulek, geruza pikortsu izenaz ezagututakoa osatzen dute. Geruza pikortsu horrek,metabolikoki aktiboa den epitelioaren barne-zati eta keratinaz betetako zelula hilezosatutako epitelioaren kanpo-zati arteko muga markatzen du. Geruza keratinotsua
Epitelio-ehunen berriztapena
37

izenekoa osatzen duten kanpo-zatiko zelulak etengabe askatzen dira. Gizakiaren kasuan,adibidez, geruza keratinotsua milaka aldiz berriztatu behar da bizitzan zehar. Geruzagerminatiboan zelula amez osatutako keratinozitoen azpipopulazio berezia dago,epidermiseko zelula, ile-folikulu eta izerdi-guruinetako zelulen aitzindari izango dena(Lavker & Sun, 2000). Zelula amok bi tokitan kokaturik daude nagusiki, folikuluen artekoepidermisean eta ile-folikuluaren konkorraren aldean (21. Ird.), azken horiek zatiketangogotsuago dihardutelarik (Ghazizadeh & Taichman, 2001).
Larruazaleko zelula amak normalean nahiko mantso zatitzen diren bitartean, zelulaametatik eratorritako TAC zelulak askoz bizkorrago zatitzen dira (Cotsarelis et al., 1990),zelula ama batetik sortu berria den TAC zelula bakoitzak desberdintzatzeari ekin bainolehen hiruzpabost zatiketa-ziklo pairazten dituelarik (Jones et al., 1995). TAC zeluletatiksortutako zelula kumeek erpinalderantz migratzeari ekiten diote, xafla basaletik urrunduzeta bidean zehar integrinen adierazpen-mailen jaitsiera jasoz. Integrinen beherakada
Zati Teorikoa
38
21. Irudia: Epidermiseko zelula amen kokapena eta beraien banaketa ugaztunetako larruazalean zehar. Babankokatzen diren zelula amek zatitu ondoren papila dermikora, gantz-guruinera zein epidermisaren oinaldera migratzendute, ondoren desberdintzapen-bidean sartuz. (Spradling et al.-etik (2001) birmoldatua).

horrek ziklo zelularra geldiaraztea eragiten du, azkenean desberdintzapena oso-osorikekarriz (Morasso & Tomic-Canic, 2005).
Epidermisaren zati bat kalteturik edo eta zeharo suntsiturik suertatzekotan, kalteainguruko zelula epidermikoek konpontzen dute, proliferazio-tasa areagotuz etasuntsitutako guneraino migratuz. Dena den, kalte-motaren arabera erreakzioa ezberdinaizan daiteke. Adibidez, izpi ultramoreen erradiazioaren aurrean, keratinozitoendesberdintzapena bultzatzen da, geruza keratinotsua trinkoago bihurtuz (Sesto et al.,2002). Haatik, epidermisa mekanikoki larri kaltetzekotan, keratinozitoen proliferazioa etamigrazioa gauzatzen dira (Tomic-Canic et al., 2004). Sortu berria den migrazioarenhelmuga-gunean, zelula ama berrien beharra dago. Horretarako, gune horren albokoertzetako zelula amek zatiketa simetrikoa pairatzen dute, eta zelula ama bakoitzetik bizelula ama kume ateratzen dira, zelula amen kopurua behar bezala emendatuz. Gainera,TAC zelulek eta guztiz desberdintzatu gabeko zelulek ere epitelioaren berreskurapeneanparte har dezakete (Li et al., 2004). Izan ere, epidermiseko xafla basalarekin kontaktuandarraiten zelulek, zelula amak izateko gaitasuna gorde dezakete, kontaktua galdu artedesberdintzapen-bidean sartzen ez baitira.
Desberdintzapenaren eraenketa, ß1-integrinak izeneko proteinen menpean dago.Dirudienez, ß1-integrinen adierazpen-maila baxua duten zelulek, altua dutenek bainozatitzeko ahalmen gehiago dute, zelulak isolatuz gero kolonia berriak sortzeko dutengaitasuna ikertzean ondorioztatu dena (Watt, 2000). Adibidez, TAC zelulek ß1-integrinenmaila baxua dute, zatitzeko gaitasun handia edukiz. Izatez, TAC zelulak zatiketen kopurumugatu batetik aurrera ezin dira berriro ere zatitu; eta, maiztasun handiz zatituz gero, xaflabasaletik askatu eta desberdintzapen prozesuan murgiltzen dira (Jensen et al., 1999). Halaere, zenbait kasutan, xafla basalarekin kontaktuan darraiten zelulak ere desberdintzatzenhas daitezke, kontaktu horien iraunkortasuna keratinozitoen patua muga dezakeen faktorebakarra ez baita. Izan ere, keratinozitoen patua hainbat aldagaien eragin pean legoke,esaterako: zelula amen zatiketa-tasa, zelula amen kumeak zelula ama bilakatzekoprobabilitatea, TAC zeluletan desberdintzatzen hasi aurretik zenbat zatiketa-ziklo emanbehar diren, xafla basaletik askatzeko keratinozitoek behar duten denbora, etadesberdintzapen-prozesua bere osotasunean erdiesteko behar den denbora, besteakbeste. Aldagai guzti horien atzetik, hainbat seinale eta informazio-molekulen saredesberdinetako partaideen mailan gertatutako aldaketak ditugu. Izatez, epitelioenhazkuntza-faktorea (EGF), fibroblastoen hazkuntza-faktorea (FGF), Wnt, Hedgehog,Notch, hezurren proteina morfogenetikoa/ß eraldatze-hazkuntza faktorea (BMP/TGF ß) eta
Epitelio-ehunen berriztapena
39

integrinen gisako faktore proteikoek epidermisaren hazkuntza eta desberdintzapeneanparte hartzen dutena badakigu (Oro & Scott, 1998; Dellambra et al., 2000; Watt, 2000).
2.3.2.- Ugaztunen hestea
Ugaztunen heste-epitelioan lau zelula-mota nagusi aurkitu dira, hala nola:enterozitoak, muki-zelulak (ertz itxiko mukozitoak) edo zelula kaliziformeak (ertz irekikomukozitoak), zelula endokrinoak eta Paneth zelulak. Epitelio horrek, endodermotikeratorritako epitelio pseudogeruzatu batean du bere jatorria. Hestearen garapenean zehar,jatorrizko epitelioa inbaginatu egiten da, eta inbaginazio eratu berrien zati distaleko zelulakzatiketan arituz, inbaginazioak hazi egiten dira. Azkenean, inbaginazio horiek, hestemeharrean Lieberkühn kriptak eta heste lodian kolon-guruinak bilakatzen dira (Totafurno etal., 1987).
Saguen kasuan, heste-epitelioaren berriztapena bizkor eta etengabe Lieberkühnkriptetan gertatzen da. Kripta horietako bakoitzean 250 zelula kokatzen dira batezbeste,
Zati Teorikoa
40
22. Irudia: Saguan timidina tritiatua injektatu ondoren, zelula amen kokapena eta geroko migrazio-bidea erakutsidituzten hestearen ebakin histologikoak. A) 40 minutu injektatu ondoren, B) egun bat injektatu ondoren, C) 2 eguninjektatu ondoren, D) 3 egun injektatu ondoren. Irudian segidan markatutako zelulak kriptaren erpinaldera migratuzdoazela ikus daiteke, baita seinalea diluituz doala ere. (Potten-etik (1998) birmoldatua).

horien artean zelula amak, zelula desberdintzatuak eta zelula migratzaile sortu berriakaurki ditzakegularik. Eskuarki, zelula amak kripten heren distalean banatuta daude (Pottenet al., 2003). TAC zelulak, zelula ametatik eratorriak eta 4-6 aldiz zatitzera irisdaitezkeenak, kripten bi heren distaletan lokalizatu dira. Heste-kriptak eta -guruinen arteanhestearen argirantz proiektatutako heste-ebaginazio hazkarak heste-biloxka izenazezagutu dira. Heste meharrean, biloxka bakoitzaren inguruan, beraz, hainbat Lieberkühnkripta daude paratuta, eta kripta horietako zelula amek bai biloxketako eta bai kriptetakoepitelioak osatzen dituzten lau zelula-motak sor ditzakete. Enterozitoak, muki-zelulak etazelula endokrinoak, heste-biloxken mutur distaleranzko migrazio-bidean dauden bitarteandesberdintzatu egiten dira. Migrazio hori, 2-5 egunez gauzatzen dena, oso era antolatuanburutzen da, zutabeka (22. Ird.; Potten, 1998), eta biloxken mutur distalera heltzerakoanzelulak hil egiten dira (Potten, 1998). Bestalde, kolonean zein urdailean, kriptetatik
Epitelio-ehunen berriztapena
41
23. Irudia: Heste epitelioko zelulen migrazio eta desberdintzapen-bide orokorrak adierazten dituen irudieskematikoa. (Sancho et al.-etik (2004) birmoldatua).

migratzen duten zelula gazteak hestearen argi-alderantz edo mukosa-alderantz abiatzendira, zelula amak guruinen zati proximalean kokaturik egonik (Yamada et al., 1992).
Heste meharreko Paneth zelulek, kripten mutur distalean paratuta daudenak,mikrobioen aurkako peptidoak eta hazkuntza-faktoreak jariatzen dituzte, eta heste-epitelioko beste zelula-motek ez bezala, kriptetako mutur distalerantz migratzen dute,bidean desberdintzatuz doazelarik (23. Ird.). Helmugara, hau da Lieberkühn kriptetara,iritsi ondoren biziraupen luze samarra dute (20 egun inguru), hil eta fagozitatuak izan bainolehen.
Azkenik, bestelako zelula amekin alderatuz, hesteko zelula amak apoptosiansartzeko gaitasun handiagoa dutela aipatu beharra dago. Halaber, desberdintzapenprozesua nahiko itzulgarria dirudi, zelula kume desberdintzatuak, zelula amenberezitasunak berriro ere berreskuratzeko gai baitira (Marshman et al., 2002).
2.3.3.- Ugatz-guruina
Ugaztun helduen ugatz-guruina, ehun konektibo adipotsuz inguratuta dagoen etaepitelio bakunez osatuta dagoen konduktu-sistema da, indibiduo helduen konduktuetandistalki albeoloak bereiz daitezkeelarik. Konduktuen artean, primarioak eta sekundarioakbereiztu dira, kalibre eta kokapenaren arabera. Konduktu zein aleboloetan, bi zelula-motadesberdin deskribatu dira, jariatzaileak eta mioepitelialak alegia (Kordon & Smith, 1998).Jatorriz epitelio-zelulak diren zelula mioepitelialek, muskulu-zelulen antzera uzkurkorrakizanik, esnea albeoloetatik konduktuetara eroatearen ardura dute, oxitozinarenpresentzian uzkurtuz. Ugatz-guruinak, jatorri ektodermikoa du. Bere garapenarenabiapuntuari dagokionez, ektodermoaren loditze-gune bat aldameneko mesenkimanmurgilduz zelula ametan joria den baba edo konkor moduko egitura sortzen da. Gerora,baba horretatik adarkamenduak desberdintzatuz doaz (Smalley & Ashworth, 2003). Ugatz-guruinaren tamainaren behin-behineko emendioa eta adarkatutako egituren hedapena,hormona-kinada eraginda (Topper & Freeman, 1980), indibiduoaren bizitzan zehar garaijakin batzutan ematen da, pubertarora heldutakoan eta haurdunaldian, alegia, bukaera-babetan zelula amen presentzia nabarmena izanik (Smalley & Ashworth, 2003).Konduktuen adarkatze eta murrizketan, zelula amen ekintza eta apoptosiaren bidezkoheriotza zelularra berebiziko garrantzia duten prozesuak dira (24. Ird.). Hori dela eta,ugatz-gurina, desberdintzapen mekanismoak ikertzeko, zelulen proliferazioa etaheriotzaren arteko lankidetza barne, askotan erabili den eredu esperimentala dugu (Albertset al., 2002).
Zati Teorikoa
42

Epitelio-zelulen proliferazioari dagokionez, guztiz garatutako ugatz-guruinekozelula-mota guztiak zelula ama amankomun batetik (zelula ama multipotentziala) eratorriakdirela behatu da saguen kasuan (Kordon & Smith, 1998). Alde batetik, zelula esne-jariatzaileei dagokionez, haurdunaldian zelula ama zatitzen hastea hormonek eragitendute, beren kopurua hamar-hogei biderrez igo araziz. Horrela, konduktuen bukaera-zatiakhazi egiten dira, adarkamendu berriak eta, azkenean, zelula jariatzaile desberdintzatuezosatutako albeolo dilatatuak eratzen dira (Smalley & Ashworth, 2003). Beste alde batetik,zelula mioepitelialak ere, konduktuetako epitelioan dauden zelula ama multipotentzialetikeratorriak dira (Liu et al., 2005); albeoloak tapizatzen paratu aurretik oso desberdintzapen
Epitelio-ehunen berriztapena
43
24. Irudia: (A) Ugatz-guruineko bukaera-konduktuaren alde distalean zelula amak simetrikoki (gezi estuak) zeinasimetrikoki (gezi lodiak) zatitzen dira. Zatiketa asimetriko baten ostean, zelula kumeak zelula mioepitelialak edokonduktu-zelulak emateko desberdintzatzen dira. Argia sortzeko, epitelioko erpinaldean dauden guztiz desberdintzatugabeko zelulak apoptosian sartzen dira. (B) Ugatz-guruineko zelula ama batzuk zatiketa simetrikoa (gezi estuak) etaasimetrikoak (gezi lodiak) pairatzen dituzte epitelioa berriztatzeko. Beste zelula amek, ordea, soilik zatiketaasimetrikoa pairatzen dute, beraien zelula kumeak albeoloen zelula mioepitelialak eta zelula jariatzaileak ematekodesberdintzatuz (Smalley & Ashworth-etik (2003) birmoldatua).

prozesu luzea pairatu behar duten arren. Zelula ama multipotentzialak, garapen etadesberdintzapen fase guztietan xafla basalari lotuta behatu dira (Chepko & Smith, 1997;Chepko & Dickson, 2003). Epitelioaren berriztapenean zelula amen partehartzeakxehetasun handiz deskribatu da (24. Ird.; Smalley & Ashworth, 2003). Zelula amek, oro har,ezaugarri ultrastruktural nabarmenik ez duten arren (arestian aipatu den bezala), ugatz-guruineko zelula ama multipotentzialak berezko ezaugarri ultrastrukturalak baditu (Chepko& Dickson, 2003).
Aitzitik, zenbait ikerlanek hiru zelula amen mota desberdin daudela iradoki dute.Ugatz-guruinaren transplantea jaso duten arratoiak ikertuz, zelula ama multipotentzialazgain beste bi zelula ama desberdin, konduktu sekundariokoak eta albeolokoak alegia,egon daitezkeela ondorioztatu da (Kamiya et al., 1999; Kim et al., 2000). Azken biok zelulaama multipotentziala aitzindari amankomuna dute, eta bai zelula jariatzaileak zein zelulamioepitelialak emango lituzketelarik, norberak bere eskualdean (Kordon & Smith, 1998).Horrela orden hierarkikoa jarraituz, konduktu primarioetako zelula ama beste zelula amabion aitzindaria litzateke.
Esan bezala, zelulen heriotza ere ugatz-guruineko epitelioaren garapenean etaberriztapenean oso fenomeno garrantzitsua da. Izatez, esnea ekoizteko beharraamaitutakoan, zelula jariatzaileak apoptosi bitartez hiltzen dira, albeolo gehienakerregresioz desagertuz. Orduan, makrofagoek, hildako zelulen hondakinak fagozitatzendituzte, ugatz-guruina berriro ere latentzia egoerara itzuliz. Apoptosia hainbat faktore direlamedio aktibatzen da, esne-jariapena blokeatzen den tokietan TGFß3 (ß3 eraldaketahazkuntza-faktorea; ingeleraz, Transforming Growth Factor ß3) metatuz, adibidez.
Zati Teorikoa
44
25. Irudia: Penaeus semisulcatus krustazeoaren hepatoarearen tubuluaren irudi eskematikoa. Bertan zelula-motadesberdinen kokapena ikus daiteke. (E), zelula amak; (R), zelula birxurgatzaileak; (F), zuntz-zelulak; (B), babaitxurako zelulak; (M), zelula txikiak. (Al-Mohanna & Nott-etik (1989) birmoldatua).
Ugatz-guruinaren garapena, obarioko hormona, hormona adenokortikoide etahormona pituitarioen arteko elkarrekintzek eraenduta dago (Topper & Freeman, 1980);hormona horiek hazkuntza-faktoreen gainean duten eragina dela medio (Roberts & Sporn1992). Hazkuntza-faktore horien artean, hazkuntza-faktore epiteliala (EGF), fibroblastoenhazkuntza-faktorea (FGF), α eta ß eraldaketa hazkuntza-faktoreak (TGFα eta TGFß),intsulina antzeko hazkuntza-faktorea (ILGF) eta faktore kolonia-kitzikatzailea (CSF1)ditugu (Fischer & Lakshmanan, 1990; Derynck, 1992; Roberts & Sporn, 1992; Pollard &Hennighausen, 1994).
2.3.4.- Krustazeoen hepatoarea
Hepatoarea, krustazeoen heste ertaineko egitura nagusia da, bolumenari zeinkonplexutasunari dagokionez. Bertan, liseriketa-prozesuaren azkeneko faseak betetzen
Epitelio-ehunen berriztapena
45
26. Irudia: Astacus astacus krustazeoaren hepatoarearen tubuluaren enbrioi-gunearen histologia.orokorra (a-c)eta irudi mitotikoen zehaztasunak (d-k).(HS), gune hemolinfatikoa, (A), anafasea, (E), E-zelula, (N), nukleoa, (Me),metafasea, (TL), tubuluen argia, (P), profasea, geziburuek PAS gune positiboak adierazten dute, geziek mikrobiloskakadierazten dute.(Vogt-etik (1994)-tik hartua.

dira, esaterako: 1) aurreko hestetik jasotako elikagaien azken liseriketa; 2) aurrekohestean beharrezkoak diren entzimen ekoizpena, 3) liseriketa produktuen xurgapena etametaketa eta 4) hondakinen askapena. Hepatoarearen egitura-ardatz nagusia, konduktuprimario batek osatzen du; konduktu hori konduktu sekundariotan eta horiek bukaera-mutur itsuko tubuluetan adarkatuz bukatzen delarik (Gibson & Barker, 1979). Krustazeoenespezie desberdinetan, hepatoarearen sorrera eta desberdintzapena une desberdinetangerta daiteke. Zenbaitetan, Penaeus setiferus espeziea kasu, hepatoarea larbaondokofase berantiarrean itxuratzen hasten da; Homarus americanus espeziearen moduko bestekasuetan, berriz, enbrioi-fasean (Biesiot, 1986; Lovett & Felder, 1989).
Dena den, oro har, hepatoarearen epitelioan lau zelula-mota nagusi bereizi dira,hala nola: E-, R-, F- eta B-zelulak (25. Ird.). Zenbait espezietan, bosgarren zelula-mota batgehiago ere identifikatu da, M-zelula izena eman zaiona. E-zelulak (E, embrionary cell),tubuluen mutur distalean kokaturik, zelula ama gisa jokatzen du, eta beste zelula-motaguztien aitzindaria da. R-zelula (R, resorptive cell) birxurgatzailea da, ugariena izanik, etabertan lipido eta glukogenoaren metaketa gauzatzen dira. F-zelulak (F, fiber cells),tubuluen eskualde distalean aurki ditzakegu batik bat. B-zelulak (B, blisterlike cells)hepatoareako epitelioan deskribatu diren zelula handienak dira, tubuluen bi herenproximaletan nagusiak izanik. Azkenik, M-zelulak (M, midget cells), liseri-epitelioarenoinaldean kokaturik lumeneraino iristen ez diren zelula gutxi batzuk baino ez dira (Icely &Nott, 1992).
Aurrenez esan bezala, E-zelulek zelula ama gisa dihardute (26. Ird. Vogt, 1994).Izan ere, E-zelulek desberdintzapen-maila baxua erakusten dute. Haatik, zenbait autorekdiotenaren arabera, zelula-mota guztiak ez datoz zuzenean E-zeluletatik. Zehazki, R- etaF- zelulak, E-zelulen aldamenean kokaturik daudenak, E-zeluletatik eratorritako bi leinuzelular ezberdintzat kontsideratu diren arren, B-zelulak antzerako ezaugarriultrastrukturalak dituzten F-zeluletatik garatuko lirateke. (Loizzi, 1971; Hopkin & Nott,1979). Beste autore batzuk, Penaeus monodon espeziean B-zelula gazteak F- eta R-zelulagazteetatik zalantzarik gabe bereizi daitezkeenaz jabetu ondoren, B-zelulek euren aldetikE-zeluletatik zuzenean sortzen den leinu zelularra osatuko luketeela proposatu dute (Icely& Nott, 1992).
Zati Teorikoa
46

2.4.- Desberdintzatutako zelulen bitartez berriztatzen diren epitelioak
2.4.1.- Area
Arean, bi epitelio nagusi daude, entzimak liseri-traktura jariatzen dituzten zelulaexokrinoz osatutakoa eta odolera hormonak jariatzen dituzten zelula endokrinozosatutakoa. Area exokrinoa, epitelio bakunez osatutako azinoetan antolatuta dago, nonzelula nagusiak (jariatzaileak), zelula zentroazinarrak eta konduktu-zelulak deskribatubaitira. Langerhans irletan mataza moduan korapilatutako epitelio endokrinoaridagokienez, 4 zelula-mota bereizi dira, α-zelulek glukagona, ß-zelulek intsulina, γ-zelulek
Epitelio-ehunen berriztapena
47
ZELULA-MOTA FUNTZIOA JATORRIA Ugaztunen epidermisa
Keratinozitoak
Keratinaren ekoizpena (babesa)
Epidermiseko zelula amak
Ugaztunen hestea Enterozitoak Muki-zelulak Zelula endokrinoak Paneth-zelulak
Hidrolasen jariapena/elikagaien xurgapena Mukiaren ekoizpena Hormonen jariapena Hazkuntza-faktore eta mikrobioen aurkako peptidoen jariapena
Kriptetako zelula amak.
Ugatz-guruina Zelula jariatzaileak Zelula mioepitelialak
Esnearen jariapena Uzkurketa/mugimendua
Ugatz-epitelioko zelula amak
Krustazeoen hepatoarea E zelulak R zelulak F zelulak B zelulak M zelulak
Zelula amak Lipido eta glukogenoaren metaketa Entzimen jariapena Elikagaien xurgapena/liseriketa Hondakinen metaketa
E zelulak
2. Taula: Zelula amen bitartez berriztatzen diren epitelioen zelula-motak, euren funtzioa eta jatorria.
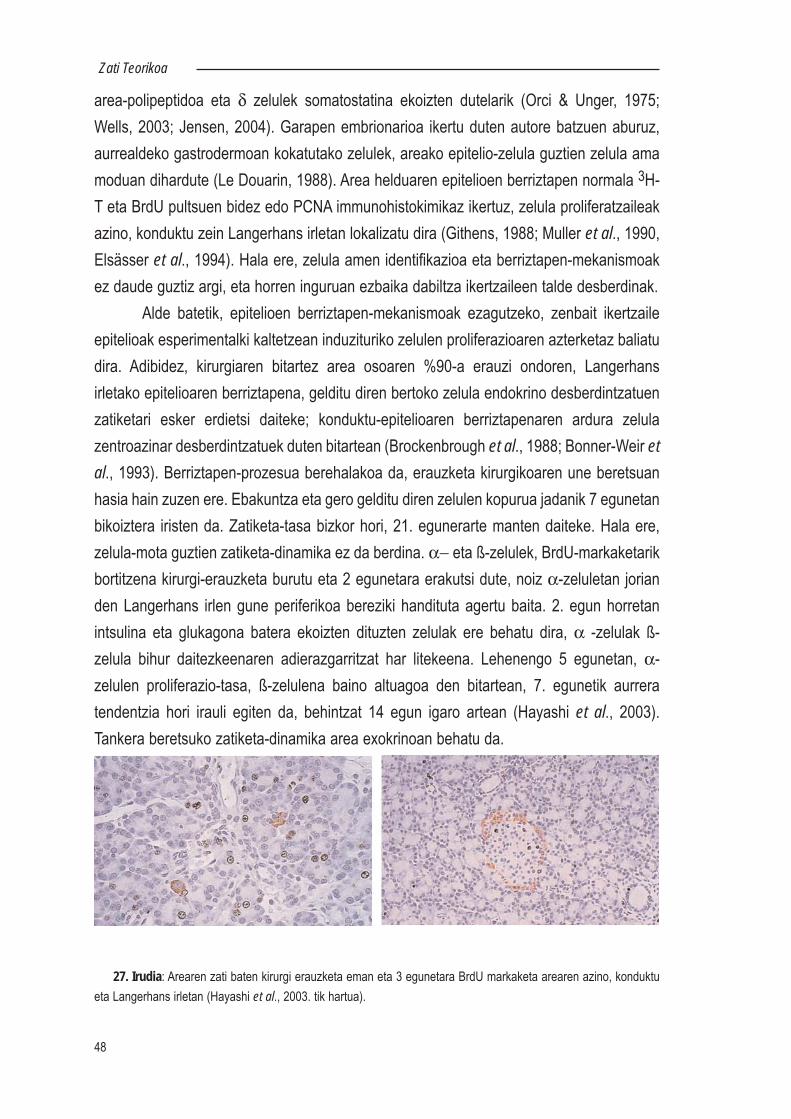
area-polipeptidoa eta δ zelulek somatostatina ekoizten dutelarik (Orci & Unger, 1975;Wells, 2003; Jensen, 2004). Garapen embrionarioa ikertu duten autore batzuen aburuz,aurrealdeko gastrodermoan kokatutako zelulek, areako epitelio-zelula guztien zelula amamoduan dihardute (Le Douarin, 1988). Area helduaren epitelioen berriztapen normala 3H-T eta BrdU pultsuen bidez edo PCNA immunohistokimikaz ikertuz, zelula proliferatzaileakazino, konduktu zein Langerhans irletan lokalizatu dira (Githens, 1988; Muller et al., 1990,Elsässer et al., 1994). Hala ere, zelula amen identifikazioa eta berriztapen-mekanismoakez daude guztiz argi, eta horren inguruan ezbaika dabiltza ikertzaileen talde desberdinak.
Alde batetik, epitelioen berriztapen-mekanismoak ezagutzeko, zenbait ikertzaileepitelioak esperimentalki kaltetzean induzituriko zelulen proliferazioaren azterketaz baliatudira. Adibidez, kirurgiaren bitartez area osoaren %90-a erauzi ondoren, Langerhansirletako epitelioaren berriztapena, gelditu diren bertoko zelula endokrino desberdintzatuenzatiketari esker erdietsi daiteke; konduktu-epitelioaren berriztapenaren ardura zelulazentroazinar desberdintzatuek duten bitartean (Brockenbrough et al., 1988; Bonner-Weir etal., 1993). Berriztapen-prozesua berehalakoa da, erauzketa kirurgikoaren une beretsuanhasia hain zuzen ere. Ebakuntza eta gero gelditu diren zelulen kopurua jadanik 7 egunetanbikoiztera iristen da. Zatiketa-tasa bizkor hori, 21. egunerarte manten daiteke. Hala ere,zelula-mota guztien zatiketa-dinamika ez da berdina. α− eta ß-zelulek, BrdU-markaketarikbortitzena kirurgi-erauzketa burutu eta 2 egunetara erakutsi dute, noiz α-zeluletan jorianden Langerhans irlen gune periferikoa bereziki handituta agertu baita. 2. egun horretanintsulina eta glukagona batera ekoizten dituzten zelulak ere behatu dira, α -zelulak ß-zelula bihur daitezkeenaren adierazgarritzat har litekeena. Lehenengo 5 egunetan, α-zelulen proliferazio-tasa, ß-zelulena baino altuagoa den bitartean, 7. egunetik aurreratendentzia hori irauli egiten da, behintzat 14 egun igaro artean (Hayashi et al., 2003).Tankera beretsuko zatiketa-dinamika area exokrinoan behatu da.
Zati Teorikoa
48
27. Irudia: Arearen zati baten kirurgi erauzketa eman eta 3 egunetara BrdU markaketa arearen azino, konduktueta Langerhans irletan (Hayashi et al., 2003. tik hartua).

Bestalde, zelula exokrinoak, endokrinoak emateko ere desberdintza daitezkeelaesperimentalki frogatu da, saguak espezifikoki ß-zelulak deuseztatzen dituenestreptozotozina izeneko farmakoaz tratatuz (Gu et al., 1997) zein area-konduktuakirurgikoki buxatuz (Bertelli & Bendayan, 1997). Esperimentu horietan amilasa eta intsulinabatera ekoitz ditzaketen zelulak deskribatu dira, zelula exokrinoak endokrino bihurdaitezkeenaren adierazgarritzat har litekeena. Zentzu berean, arratoi transgenikoengarapena aztertzean, zelula ametatik eratorritako zelula endokrino eta exokrinoenezaugarriak batera dituen zelula-mota bat aurkitu da (Gu et al., 1994). Halaber,konduktuetako zelula amak ß-zelulak emateko desberdintza daitezke, induzituriko kalteeierantzunez zein ezohiko eskari metabolikoen aurrean areagotu daitekeena (Bonner-Weir& Sharma, 2002).
Azkenik, arratoi transgenikoak eredu esperimental gisa erabiliz, areako zelulaendokrinoak zatiketa autologoz baino ez direla zatitzen ere proposatu da, partikularki ß-zelulen kasuan, egoera normalean zein arearen zati handia erauztekoan ß-zeluladesberdintzatuetatik soilik sor daitezkeenak (Dor et al., 2004).
Beraz, α− eta ß-zelulek jatorri berbera omen dutenez, eta zelula endokrino etaexokrinoen jatorria ere amankomuna omen denez, guztiok zelula ama beratik eratorriakizan daitezkeela pentsa genezake (Herrera 2000). Jatorrizko zelula ama horrek areakotranskribapen-faktorea edukiko lukeen arren, ez luke hormonen adierazpen-gaitasunikizango. Gainera, zelula ama amankomunetik zelula ama endokrino eta exokrinoa bereiztudaitezkeenik ezin da baztertu (Seaberg et al., 2004); ezta norberak gerora zelula amaespezifikoagoak eman litzakeenik ere. Izan ere, aitzindari endokrinoak, konduktutik gertuegon arren, ez lirateke konduktu-epitelioaren osagaiak izango (Gu et al., 2002).
2.4.2.- Listu-guruinak
Listu-guruinak, garatze-bidean dagoen mihi-epitelioaren loditze-gune batetiksortzen dira, zeinen ondoan, listu-guruinak izango diren baba itxurako formazio bat eratuz,kondentsazio mesenkimatikoa ematen baita (Melnick & Jaskoll, 2000). Listu-guruinhelduek itxura albeolotubularra dute, eta ehun konektiboak enkapsulatzen dituzten hainbatlobuluz osaturik daude. Eskualde albeolotubularra, azino jariatzaileen multzoek etakonduktuen sistema konplexu batek osatzen dute. Azinoen epitelioan zelula jariatzaileaketa zelula mioepitelialak daude; konduktu-epitelioetan, berriz, ez dago zelula jariatzaileriketa osmoeraenketa moduko funtzioak bertan betetzen dira (Actis et al., 2002).
Epitelio-ehunen berriztapena
49

Timidina tritiatuaz markatutako arratoi helduetan, konduktuen zelula pikortsuakkonduktu ertaineko oinaldeko zeluletatik eratorriak direla behatu da. Hasiera batean,konduktuen zelula pikortsu ildaxkatu bihurtzen dira (tarteko egoera), azkenean zelulapikortsu helduak bilakatu arte (Chai et al., 1993; Denny et al., 1993). 3H-T markaketa-indizeek erakutsi dutenaren arabera, ez dirudi konduktu ertainetako zelula amek bizkorzatitzen direnik. Haatik, jarduera proliferatzaile altuena zelula pikortsu ildaxkatuek erakutsidute. Hau da, zelula amak astiro zatitzen diren bitartean, zelula pikortsu ildaxkatuak, agianTAC zelulen analogotzat har genitzakeenak, bizkor zatitzen dira, gerora guztizdesberdintzatuak direnean zatiketa-tasa berriro ere motelduz doalarik (Chai et al., 1993).
Listu-guruinetan esperimentalki kalteren bat induzituz, azinoen kopuruan etatamainan murrizpena sortaraztekotan, (adb., apoptosia bultzatuz), konduktuen antzekoegitura tubularren agerpena suertatu da (Takahashi et al., 2002). Dirudienez, egituratubular horiek konduktu ertainetatik eratorriak izango lirateke, eta azino-zelulen zelulaaitzindarien ordezko sortuberriez osatuta egongo lirateke (Takahashi et al., 1998; 2002).Beste autore batzuen arabera, berriz, tubulu horiek zelula jariatzaileak galdu dituztenazinoak baino ez lirateke izango, zelulen kopuru egokia berreskuratuz gero berriro erefuntzionalak bilakatuko liratekeenak (Scott et al., 1999). Bistan denez, momentuz egituratubular horien jatorria eta esangura biologikoa ez daude batere argi.
Listu-guruinek, kirurgi-buxadura 5 egunez jasan ondoren, konduktuetako zelulapikortsu helduek proliferazio-jarduera handia erakutsi dute, 3H-T zein PCNA markaketakerabiliz frogatu dena (Takahashi et al., 2000). Aitzitik, listu-guruinen kirurgi-erauzketapartziala egin ondoren, azinoen berreskurapena konduktu nagusietako zelulen proliferazioeta desberdintzapenaren bitartez gauzatzen dela behatu da, ebakuntza eta gero geratzendiren azinoek nolabait ere parte hartzen dutelarik (Hanks & Chaudhry, 1971).
Zati Teorikoa
50
28. Irudia: Listu-guruineko konduktu nagusia buxatuta duen arratoian burututako PCNA immunohistokimika.Azino zein konduktuetako zelulek marka positiboa (geziak) erakutsi dute (Takahashi et al.-etik (2001) hartua).

Bestalde, arratoien listu-guruinetako epitelioan dimorfismo sexuala dagoelaaipagarria da. Arratoi ar gazteetan, konduktuetan agertzen diren zelula berri gehienakkonduktu-zelula helduen zatiketatik datoz; hau da, berezko zatiketa edo zatiketa autologoaizenekoa ematen da. Zelula berri desberdintzatuen %20 inguru baino ez dira konduktuertainetako zelula ametatik eratorriak, azinoetan dauden gainerako zelula guztiak zelulahelduen proliferazioz sortuak direlarik. Eme gazteetan, ordea, bai azinoetan zeinkonduktuetan dauden zelulen %60-a zelula ametatik eratorriak dira (Denny et al., 1997).Hala ere, emeak heltzen doazen heinean, azinoetako zelula berriak zeluladesberdintzatuen zatiketa autologoz sortzen dira, zelula amen parte hartzea adinarekinbatera gutxituz doalarik. Aitzitik, eme helduetan, konduktu-zelulak oraindik ere konduktuertainetako zelula ametatik datoz. Desberdintasun honen gakoa, hazkuntza-abiaduranegon daiteke. Izan ere, eme gazteen listu-guruinek oraindik ere hazten jarrai dezakete,batez ere azino-eskualdean; nolabaiteko izaera enbrionarioa mantentzera behartutadaudela erakutsi lezakeena (Denny et al., 1990).
2.4.3.- Ugaztunen gibela
Gibelak jatorri endodermikoa du, endodermoaren alde bentrala loditu egiten dagibelaren dibertikulua eratuz. Hepatoblastoek endodermotik migratzen dute baba modukoeskualde bereizia itxuratuz eta septum transversum izeneko gunean zehar proliferatzendute (Tosh & Strain, 2005). Gibeleko epitelioen berriztapena, ez da egoera normaletanzelula ama gutxi batzuei esker ematen, zelula helduen zatiketa autologoz baizik.Esaterako, gibelaren erauzketa kirurgikoa partzialaren ondoren, berriztapena gainontzekohepatozito zein behazun-epitelioko zelula helduen zatiketaz burutzen da (Fausto, 2001).Beraz, epitelio-zelula horiek oraindik ere guztiz desberdintzatu gabeak direla esangenezake. Are gehiago, hepatozitoak, esaterako, organismoaren biziraupen osoan zeharproliferatzeko gai dira. Kirurgi-erauzketa aplikatu eta hepatozitoetan DNAren sintesia hasibitarteko denbora-tartea hainbat faktoreen pean dago, argiztapen-erregimena eta elikatze-patroia, besteak beste (Yanagi & Potter, 1977). Aitzitik, farmako kartzinogenikoz trataturikoarratoien gibelean, epitelio-zelulen proliferazioa, zelula amatzat har litezkeen eta porta-guneko Hering kanaletan dauden itxura obaleko zelula txikien zatiketaz hasten da. Zelulahoriek bipotentzialak lirateke, hepatozitoak zein behazun-epitelioko zelulak emangobailituzkete (Theise et al., 1999). Antzeko itxura obaleko zelula txikiak barneko porta-gunean ere deskribatu dira, hepatozito zein areako zelulen aitzindaritzat hartu direnak(Tosh & Strain, 2005).
Epitelio-ehunen berriztapena
51

Hepatozitoen proliferazioa zerk pizten duen jakiteko hainbat ikerketa burutu dira.Esaterako, gibel-ehunaren zatiak edo eta hepatozito isolatuak gibela ez den ehun bateratransplantatu ostean transplante-hartzailearen gibelaren zati bat ebakuntza bidezerauztean, transplantatutako zelulek, gibela ez den ehun horretan dauden arren, DNAsintetizatzen hasten dira. Horrek, gibela erregeneratzen den bitartean odolean nolabaitekoseinale mitogenikoa agertzen dela iradoki du (Overturf et al., 1999). Izan ere, hepatozitoenhazkuntza-faktoreak (HGF; ingeleraz hepatocyte growth factor), berebiziko garrantziaomen dauka prozesu horretan. Are gehiago, in vitro mantendutako hepatozitoak, HGFrenpresentzian, eta bestelako zitokinen seinalerik bitarteko izan gabe, zatitzen hasten dira.Hala ere, in vivo, HGFz gain, epitelio-hazkuntza faktore (EGF; ingeleraz, epithelial growthfactor) moduko beste faktore batzuen plasma-kontzentrazioak ere igotzen dira kirurgi-erauzketaren ondorioz. Halaber, intsulina, interleukina 6 (IL6) eta norepinefrina molekulenbidezko seinalizazioa ere gibelaren erregenerazioa burutzeko ezinbestekoak dira, nahizeta hepatozitoentzat oinarrizko mitogenoak ez izan. Antza denez, nondik norako orokorrakezagutu diren arren, hepatozitoen proliferazioan diharduten seinale molekularrei buruzoraindik ere argilun dexente dago (Michalopoulos & DeFrances, 1997).
2.4.4.- Moluskuen liseri-guruina
Krustazeo eta intsektu (Vogt 1994; Loeb et al., 2003; Illa, 2005) zein ornodunetan(ikus aurreneko atalak) ez bezala, moluskuen epitelio-ehunen berriztapenaz oso ikerlangutxi burutu dira (Leibson & Frolova 1994). Konkretuki, moluskuen liseri-guruinekoepitelioaren berriztapenaz egindakoak bereziki urriagoak dira (Hanselmann et al., 2000;Marigómez et al., 1999), ehun horren egitura histologikoaz ikerketa ugari egin diren arren(Ikus 1. atala).
Zati Teorikoa
52
29. Irudia: Ostren liseri-epitelioko balizko zelula basofiliko baten anafasea (Yongue-tik (1926) hartua)

Urdaileko zelula amak identifikatu ez diren arren, zatiketa zelularren irudiakargitaratzeaz gain, proliferazio zelularra aztertzeko 3H-T bidezko markaketaz baliatu direnikerlanak burutu egin dira (Leibson & Frolova, 1994). Dirudienez, mitosia gertatutakoan,zatitzear dauden zelulek xafla basaletik urrundu behar dute, eta lumenaren alderamigratzen dute, ondoren zelula kumeek berriro ere epitelioaren oinalderaino itzuliz (Mix,1972). Antzeko prozesua Crenomytilus grayanus muskuiluaren heste-epitelioan deskribatuda, zelulen proliferazio-tasan sasoi-aldaketa nabarmenak aurkitu direlarik (Leibson &Frolova, 1994). Hala ere, zunda espezifikoak erabiliz edo ultrastruktura sakonki aztertuz,zelula amak identifikatzera zuzendutako lanik ez dugu ezagutu.
Bestalde, liseri-guruineko tubuluen epitelioari dagokionez, datu egokien gabeziabera dagoen arren, zelula amen presentzia proposatu da. Ostretan, liseri-epitelioko zelulenaitzindariak izan omen daitezkeenak, liseri-tubuluetako kriptetan aurkitu dira (29 Ird.;Yonge, 1926; Shaw & Battle, 1957; Mix & Sparks, 1971). Izatez, erradiazio ionizatzaileztrataturiko liseri-tubuluen epitelioan, kriptetako zelula horiek epitelio berriaren sorrerarenerantzule nagusiak dirudite (Mix & Sparks, 1971). Mytilus edulis muskuiluan eta Pectenmaximus beiran ere, desberdintzatu gabeko zelula ama bat dagoela proposatu da
Epitelio-ehunen berriztapena
53
30. Irudia: PCNA immunohistokimikamuskuiluaren liseri-guruinean etakrustazeoaren hepatoarean. Muskuiluenliseri-guruinean (a-b eta tarteko irudi txikia)positibotasun gehiena liseri-zelulei dagokie(geziak), nahiz eta zelula basofilikobatzuetan markaketa ere ageri den (geziburuak). Urdaileko zelulek markaketapositiboa erakutsi dute (c), eta baitahemozitoek (H) ere (irudi txikia). Kontrol-ebakinetan (d) ez da inolako seinalerikatzeman. Krustazeoen hepatoarearen E-zeluletan (e) markaketa bortitza behatu da.Zelulak desberdintzatuz doazen heineanmarkaketa arinago bihurtuz. E-zeluleneskualdean irudi mitotikoak (m) ikusdaitezke. Hepatoarearen kontrol-ebakinetan (f) ez da inolako markarikatzeman. Eskala-marra: 10 μm (a-c) eta 20μm (d-f) (Marigómez et al.-etik (1999)hartua).

(Thompson et al., 1974; Henry et al., 1991). Beste autore batzuen aburuz, epitelioarenberriztatzearen arduraduna ziliatua, hau da nahiko desberdintzatua, den zelula basofilikoberezia izango litzateke. Horrelakorik Cardium edule bibalbio eta Maricrypta monoxylaprosobrankio itsastarren kasuan iradoki zen (Owen, 1970; Nelson & Morton, 1979). Datupartzial eta eskas horietan oinarrituta, zelula amak zelula basofiloak direna zenbaithamarkadetan ontzat hartu da oro har, datu esperimental berrien sustapenik gabe.
Aitzitik, azken urteotan, Mytilus galloprovincialis muskuiluan, liseri-albeoloetakoepitelioko 2 zelula-motek, liseri-zelula eta zelula basofilikoa hain zuzen, PCNAimmunohistokimikaren bitartez S-fasean sartzeko gai direla frogatu da (30. Ird.; Marigómezet al., 1999). Gainera, bi zelula-motok EGF hartzaileak erakusten dituzte (Canesi et al.,2000). Hori dela eta, behintzat espezie honetan, liseri-epitelioko bi zelula helduak zatitzekogai direla dirudi, ugaztunen hepatozito moduan zatiketa autologoaz baliatuz. Ekarpenhorrek, muskuiluen liseri-guruinean zelula amaren existentzia kolokan utzi du. Hala ere,erradiazio ionizatzailearen eraginaren ondorioz jasandako kalte bortitzen aurrean zelula
Zati Teorikoa
54
ZELULA-MOTA FUNTZIOA JATORRIA Ugaztunen area á-Zelulak ß-Zelulak ã-Zelulak ä-Zelulak
Glukagonaren ekoizpena Intsulinaren ekoizpena Are-polipeptidoaren ekoizpena Somatostatinaren ekoizpena
á-Zelulak/Zelula exokrinoak ß-Zelulak/Zelula exokrinoak ã-Zelulak/Zelula exokrinoak ä-Zelulak/Zelula exokrinoak
Ugaztunen listu -guruina Zelulak jariatzaileak (exokrinoak) Konduktuetako zelulak Zelula jariatzaileak Zelula mioepitelialak Zelula parenkimatikoak
Liseri-entzimen jariapena Oreka osmotikoaren eraenketa Entzimen jariapena Uzkurketa/mugimendua Egituraketa
Zelula exokrinoak Konduktuetako zelula amak Konduktuetako zelula amak/ Zelula jariatzaileak Konduktuetako zelula amak Zelula parenkimatikoak
Ugaztunen gibela Hepatozitoak
Funtzio anitzak
Hepatozitoak/Hering kanaleko zelulak
Moluskuen liseri -guruina Liseri-zelula Zelula basofilikoa
Elikagaien liseriketa Entzimen jariapena
Liseri-zelulak/Kriptetako zelula amak Zelula basofilikoak/Kriptetako zelula amak
3. Taula: Desberdintzatutako zelulen bitartez berriztatzen diren epitelioen zelula-motak, beraien funtzioa etajatorria.

amek liardukete. Azken finean, zelula basofiliko desberdintzatuak (jariatzaileak edoziliatuak) zelula amak direla onartzeko zailtasunak aurkitu ditugun bitartean (Marigómez etal., 1999), Mix eta Sparks (1971) ikertzaileek deskribatutako zelula basofilikoak, txikiak etabiribilak izanik, zelula amak ez direla esateko ez dugu oinarri sendorik.
Epitelio-ehunen berriztapena
55

Zati Teorikoa
56
BIBLIOGRAFIA
Abrahamsen JF, Smaaland R, Sothern RB, LaerumOD (1997) Circadian cell cycle variations oferythro- and myelopoiesis in humans. Eur. J.Haematol. 58:333-345.
Actis AB, Lampe PD, Eynard AR (2002) Cellularbasis and clinical implications of biologicalmarkers in salivary tissues: Their topologicaldistribution in murine submandibular gland. OralOncol. 38:441-449.
Alberts B, Jonson A, Lewis J, Raff M, Roberts K,Walter P (2002) Molecular biology of the cell. 4.edizioa. Taylor & Francis New York. 1463 orr.
Alison MR (1995) Assessing cellular proliferation:What's worth measuring? Human Exp. Technol.14:935-944.
Alison M, Golding M, Lalani EN, Nagy P,Thorgeirsson S, Sarraf C (1997) Wholesalehepatocytic differentiation in the rat fromductular oval cells, the progeny of biliary stemcells. J. Hepatol. 26:343-352.
Al-Mohanna SY, Nott JA (1989) Functional cytologyof the hepatopancreas of Penaeus semisulcatus(Crustacea: Decapoda) during the moult cycle.Mar. Biol. 101:535-544.
Argaud A, Bounoure L (1910) Contribution a l´etudeanatomique et histologique du tube digestifd´Arion rufus. S. Anat. Physiol. N. 46:146-174.
Awaji M, Suzuki T (1998) Monolayer formation andDNA synthesis of the outer epithelial cells frompearl oyster mantle in coculture withamebocytes. In Vitro Cell Dev. Biol. Anim.34:486-491.
Babula A, Wielinska L (1988) Ultrastructural studiesof the digestive system of the slug, Derocerasreticulatum (Müller) (Pulmonata). Bull. Soc. AmisSci. Lettres de Poznam. Ser. D. Sci. Biol. 26:73-78.
Baisch H, Beck HP, Christiensen IJ, Hartmann NR,Fried J, Dean PN, Gray JW, Jett JH, JohnstonDA, White RA, Nicolini C, Zeitz S, Watson JV(1982) A comparison of mathematical methodsfor the analysis of DNA histograms obtained byflow cytometry. Cell Tiss. Kin. 15:235-249.
Bale SD, Howard TA, Moffett SB (2001) Neuronaland non-neuronal responses to nerve crush in apulmonate snail, Melampus bidentatus. Invert.Neurosci. 4:105-117.
Barbeito CG, González NV, Badrán AF (2003) Sexand Age-Related temporal variations inintestinal-epithelium proliferation in the sucklingmouse. Chronobiol. Int. 20:97-107.
Basalo C (1998) Organización morfofuncional delos alvéolos de la glándula digestiva del mejillónMytilus galloprovincialis. Lizentziatura-Tesia.UPV/EHU. 134 orr.
Baumforth KR, Grewal N, Large AT, Jones CJ, PerryCJ, Connock MJ (1998) Zonal rotor purificationand characterization of "mannosomes": atubular membrane system in gastropod molluscdigestive gland. Anal. Biochem. 263:189-197.
Beninger PG, Le Pennec M (1991) Functionallanatomy of the scallops. 133-166 orr. Non:Scallops: biology, ecology and aquaculture.Shunway SE (ed). Elsevier, Amsterdam.
Bertelli E, Bendayan M (1997) Intermediateendocrine-acinar pancreatic cells in duct ligationconditions. Am. J. Physiol. C 273:1641-1649.
Biesiot PM (1986) Changes in midgut glandmorphology and digestive enzyme activitiesassociated with development in early stages ofthe American lobster, Homarus americanus.Woods Hole Oceanographic Istitution, Publ.WHOI-86-20, PB89 190276: Doktoradutza-Tesia, 222 orr.
Bjarnason GA, Jordan R (2002) Rhythms in humangastrointestinal mucosa and skin. Chronobiol.Int. 19:129-140.
Boghen A, Farley J (1974) Phasic activity in thedigestive gland cells of the intertidalprosobranch Littorina saxatilis (Olivi) and itsrelation to the tidal cycle. Proc. Malacol. Soc.London 41:41-56.
Bonner-Weir S, Baxter LA, Schuppin GT, Smith FE(1993) A second pathway for regeneration ofadult exocrine and endocrine pancreas. Apossible recapitulation of embryonicdevelopment. Diabetes 42:1715-1720.
Bonner-Weir S, Sharma A (2002) Pancreatic stemcells. J. Pathol. 197:519-526.

Bibliografia
57
Bowen ID (1970) The fine structural localization ofacis phosphatase in the gut epithelial cells of theslug, Arion ater (L.). Protoplasma 70:73-81.
Bowen ID, Davies P (1971) The fine structuraldistribution of acid phosphatase in the digestivegland of Arion hortensis (Fre.). Protoplasma73:73-81.
Brandi GM Calabrese C, Pantaleo MA, MorselliLabate A, Di Febo G, Hakim R, De Vivo A, DiMarco MC & Biasco G (2004) Circadianvariations of rectal cell proliferation in patientsaffected by advanced colorctal cancer. Can.Lett. 208:193-196.
Bravo R, Frank R, Blundell PA, Mcdonald-Bravo H(1987) Cyclin/PCNA is the auxiliar protein ofDNA polimerase-D. Nature 326:515-517.
Bravo R, Macdonald-Bravo H (1987) Existence oftwo cyclin proliferating cell nuclear antigenduring the cell cycle: Association with DNAreplication sites. J. Cell Biol. 105:1549-1554.
Brockenbrough JS, Weir GC, Bonner-Weir S (1988)Discordance of exocrine and endocrine growthafter 90% pancreatectomy in rats. Diabetes37:232-236.
Brouard M, Barrandon Y (2003) Controlling skinmorphogenesis: hope and despair. Curr. Opin.Biotechnol. 14:520-525.
Bruno S, Darzynkiewicz Z (1992) Cell cycledependent expression and stability of thenuclear protein detected by Ki-67 antibody inHL-60 cells. Cell Prol. 25:31-40.
Cajaraville MP (1989) Efectos histopatológicos ycitotóxicos de los hidrocarburos derivados delpetróleo, y su cuantificación en el mejillónMytilus galloprovincialis (Lmk). Doktoradutza-Tesia. 577 orr.
Cajaraville MP, Díez G, Marigómez JA, Angulo E(1990) Responses of basophilic cells of thedigestive gland of mussels to petroleumhydrocarbon exposure. Dis. Aquat. Org. 9:221-228.
Cajaraville MP, Díez G, Marigómez I, Angulo E(1991) Consequences of keeping Mytilus in thelaboratory as assessed by different cellularcondition indices. Helgol. Meeresunter. 45:445-464.
Cajaraville MP, Marigómez I, Díez G, Angulo E(1992) Comparative effects of the wateraccommodated fraction of three oils on mussels2.- Quantitative alterations in the structure of thedigestive tubules. Comp. Biochem. Physiol. C102:113-123.
Cajaraville MP, Robledo Y, Etxeberria M, MarigómezI (1995a) Cellular biomarkers as useful tools inthe biological monitoring of environmentalpollution: molluscan digestive lysosomes. 29-55orr. Non:. Cell biology in environmentaltoxicology. Cajaraville MP (ed.). University of theBasque Country Press Service. Bilbo.
Cajaraville MP, Abascal I, Etxeberria M, MarigómezI (1995b) Lysosomes as cellular markers ofenvironmental pollution: time- and dose-dependent responses of the digestive lysosomalsystem of mussels after petroleum hydrocarbonexposure. Environ. Toxicol. Water. Qual. 10:1-8.
Cajaraville MP, Bebianno MJ, Blasco J, Porte C,Sarasquete C, Viarengo A (2000) The use ofbiomarkers to assess the impact of pollution incoastal environments of the Iberian Peninsula: apractical approach. Sci. Tot. Environ. 247:295-311.
Cancio I, Orbea A, Völkl A, Fahimi D, Cajaraville MP(1998) Induction of peroxisomal oxidases inmussels: comparison of effects of lubricant oiland benzo(a)pyrene with two typical peroxisomeproliferators on peroxisome structure andfunction in Mytilus galloprovincialis. Toxicol.Appl. Pharmacol. 149:64-72.
Canesi L, Malatesta M, Battistelli S, Ciacci C, GalloG, Gazzanelli G (2000) Immunoelectronmicroscope analysis of epidermal growth factorreceptor (EGFR) in isolated Mytilusgalloprovincialis (Lam.) digestive gland cells:Evidence for ligand-induced changes in EGFRintracellular distribution. J. Exp. Zool. 286:690-698.
Chai Y, Klauser DK, Denny PA, Denny PC (1993)Proliferative and structural differences betweenmale and female mouse submandibular glands.Anat. Rec. 235:303-311.

Zati Teorikoa
58
Chepko G, Dickson RB (2003) Ultrastructure of theputative stem cell niche in rat mammaryepithelium. Tiss. Cell 35:83-93.
Chepko G, Smith GH (1997) Three division-competent, structurally-distinct cell populationscontribute to murine mammary epithelialrenewal. Tiss. Cell. 29:239-253.
Colas P, Launay C, van Loon AE, Guerrier P(1993a) Protein synthesis control cyclin stabilityin metaphase I-arrested oocytes of Patellavulgata. Exp. Cell Res. 208:518-521.
Colas P, Serras F, Van Loon AE (1993b)Microinjection of suc 1 transcripts delays the cellcycle clock in Patella vulgata embryos. Int. J.Dev. Biol. 37:598-594.
Cotsarelis G, Sun TT, Lavker RM (1990) Label-retaining cells reside in the bulge area ofpilosebaceous unit: implications for follicularstem cells, hair cycle, and skin carcinogenesis.Cell 29:1329-1337.
Couch J (1984) Atrophy of diverticular epithelium asan indicator of environmental irritants in theoyster Crassostrea virginica. Mar. Environ. Res.14:525-526.
Czarna Z, Krasowka E, Turska R (1985) The tubularstructures in the cells of the digestive gland ofArion rufus (Mollusca Gastropoda). Zool.Poloniae 32: 23-28.
David H, Götze J (1963) ElektronenmikroskopischeBefunde an der Mitteldarmdrüse vonSchnecken. Z. Mikrosk.-Anat. Forsch. (Leipzig).70:252-272.
Daughaday WH, Rotwein P (1989) Insulin-likegrowth factors I and II. Peptide, messengerribonucleic acid and gene structures, serum, andtissue concentrations. Endocr. Rev. 10:68-91.
Dellambra E, Golisano O, Bondanza S, Siviero E,Lacal P, Molinari M, D'Atri S, De Luca M (2000)Downregulation of 14-3-3sigma prevents clonalevolution and leads to immortalization of primaryhuman keratinocytes. J. Cell Biol. 29: 149:1117-1130.
Dekens MPS, Santoriello C, Vallone D, Grassi G,Withmore D, Foulkes NS (2003). Light regulatesthe cell cycle in zebrafish. Curr. Biol. 13:2051-2057.
Denny PC, Chai Y, Klauser DK, Denny PA (1990)Three-dimensional localization of DNA synthesisin secretory elements of adult female mousesubmandibular gland. Adv. Dent. Res. 4:34-44.
Denny PC, Chai Y, Klauser DK, Denny PA (1993)Parenchymal cell proliferation and mechanismsfor maintenance of granular duct and acinar cellpopulations in adult male mouse submandibulargland. Anat. Rec. 235:475-485.
Denny PC, Ball WD, Redman RS (1997) Salivaryglands: a paradigm for diversity of glanddevelopment. Crit. Rev. Oral Biol. Med. 8:51-75.
Derynck R (1992) The physiology of transforminggrowth factor-α. Adv. Canc. Res. 58:27-52.
Dimitriadis VK, Konstantinidou V (2002) Origin ofexcretory cells in the digestive gland of the landsnail Helix lucorum. Malacologia 44:145-151.
Domouhtsidou GP, Dimitriadis VK (2001) Lysosomaland lipid alterations in the digestive gland ofmussels, Mytilus galloprovincialis (L.) asbiomarkers of environmental stress. Environ.Pollut. 115:123-137.
Dondero F, Viarengo A (2005) Development of novelsensitive molecular markers for marineenvironment biomonitoring employing mussels.15th SETAC Europe Anual Meeting. Lille.
Dor Y, Brown J, Martinez OI, Melton DA (2004) Adultpancreatic β-cells are formed by self-duplicationrather than stem-cell differentiation. Nature429:41-46.
Edgar BA, Lehner CF (1996) Developmental controlof cell cycle regulators: a fly´s perpective.Science 274:1664-1672.
Eirin-López JM, González-Tizón AM, Martínez A,Méndez J (2002) Molecular and evolutionaryanalysis of mussel histone genes (Mytilus spp.):possible evidence of an "orphon origin" for H1histone genes. J. Mol. Evol. 55:272-83.
Elsasser HP, Biederbick A, Kern HF (1994) Growthof rat pancreatic acinar cells quantitated with amonoclonal antibody against the proliferatingcell nuclear antigen. Cell Tiss. Res. 276:603-609.
Endl E, Gerdes J (2000) The Ki-67 protein:fascinating forms and an unknown function. Exp.Cell Res. 257:231-237.

Bibliografia
59
Faderl S, Keating MJ, Do KA, Liang S.-Y, KantarjianH.M, O'Brien S, Garcia-Manero G, Manshouri T,Albitar M (2002) Expression profile of 11 proteinsand their prognostic significance in patients withchronic lymphocytic leukemia (CLL). Leukemia16:1045-1052.
Fausto N (2001) Liver regeneration: Fromlaboratory to clinic. Liver Trasn. 7:835-844.
Finegood DT, Scaglia L, Bonner-Weir S (1995)Dynamics of beta-cell mass in the growing ratpancreas. Estimation with a simplemathematical model. Diabetes 44:249-256.
Fisher DA, Lakshmanan J (1990) Metabolism andeffects of epidermal growth factor and relatedgrowth factors in mammals Endocr. Rev. 11:418-442.
Foley J, Ton T, Maronpot R, Butterworth R,Goldworthy TL (1993) Comparison ofproliferating cell nuclear antigen to tritiatedthymidine as a marker of proliferatinghepatocytes in rats. Environ. Health Prespec.101:199-205.
Fretter V, Graham A (1962) British prosobranchmolluscs. Ray Society. 755 orr.
Furuta E, Yamaguchi K, Shimozawa A (1994) Blood-cell producing site in the land slug Incilariafruhstorferi. Kaibogaku. Zasshi. 69:751-764.
Gage FH (2000) Mammalian neural stem cells.Science 287:1433-1438.
Gerdes J, Lemke H, wacker H, Schwab HJ, Stein H(1984) Cell cycle analysis of a cell proliferationasociated human nuclear antigen defined by themonoclonal antibody Ki-67. J. Immunol.133:1710-1715.
Gerlach C, Golding M, Larue L, Alison MR, GerdesJ (1997) Ki-67 immunoexpression is a robustmarker of proliferative cells in the rat. Lab.Invest. 77:697-698.
Ghazizadeh S, Taichman LB (2001) Multiple classesof stem cells in cuteneous epithelium; a lineageanalysis of adult mouse skin. EMBO J. 20:1215-1222.
Gibson R, Barker PL (1979) The decapodhepatopancreas. Oceangr. Mar. Biol. Ann. Rev.17:285-346.
Githens S (1988) The pancreatic duct cell:proliferative capabilities, specific characteristics,metaplasia, isolation, and culture. J. Pediatr.Gastroenterol. Nutr. 7:486-506.
Goldblum JR, Appelman HD (1995) Stromal tumorsof the duodenum: A histologic andimmunohistochemical study of 20 cases. Am. J.Sur. Pathol. 19:71-80.
Golias CH, Charalabopoulos K, Chralabopoulos K(2004) Cell proliferation and cell cycle control: amini review. Int. J. Clin. Pract. 58:1134-1141.
González-Melendi P, Testillano PS, Ahamadian P,Fadón B, Risueño MC (1996) New in situapproaches to study the induction of pollenembryogenesis in Capsicum anuum L. Eur. J.Cell Biol. 69:373-386.
Gratzner HG (1982) Monoclonal antibody to 5-bromo- and 5-iododeoxyuridine: A new reagentfor detection of DNA replication. Science218:474-475.
Grimaldi A, Tettamanti G, Rinaldi L, Brivio MF,Castellani D, de Eguileor M (2004) Muscledifferentiation in tentacles of Sepia officinalis(Mollusca) is regulated by muscle regulatoryfactors (MRF) related proteins. Dev. GrowthDiffer. 46:83-95.
Gu D, Lee MS, Krahl T, Sarvetnick N (1994)Transitional cells in the regenerating pancreas.Development 120:1873-1881.
Gu D, Arnush M, Sarvetnick N (1997)Endocrine/exocrine intermediate cells instreptozotocin-treated Ins-IFN-γ transgenicmice. Pancreas 15:246-250.
Gu G, Dubauskaite J, Melton DA (2002) Directevidence for the pancreatic lineage: NGN3+cells are islet progenitors and are distinct fromduct progenitors. Development 129:2447-2457.
Hale SA, Capuco AV, Erdman RA (2003) Milk yieldand mammary growth effects due to increasedmilking frequency during early lactation. J. DairySci. 86:2061-2071.
Hall PA, Levinson DA (1990) Review: assessment ofcell proliferation in histological material. J. Clin.Pathol. 43:184-192.
Hall PA, Levison DA, Woods AL, Yu CCW, KellockDB, Watkins JA, Barnes DM, Gillett CE,

Zati Teorikoa
60
Camplejohn R, Dover R, Waseem NH, Lane DP(1990) Proliferating cell nuclear antigen (PCNA)immunolocalization in paraffin sections: An indexof cell proliferation with evidence of deregulatedexpression in some neoplasms. J. Pathol.162:285-294.
Hall PA, Watt F (1989) Stem cells: the generationand maintenance of cellular diversity.Development 106:619-633.
Hanks CT, Chaudhry AP (1971) Regeneration of ratsubmandibular gland following partialextirpation. A light and electron microscopicstudy. Am. J. Anat. 130:195-208.
Hanselmann R, Smolowitz R, Gibson D (2000)Identification of proliferating cells in hard clams.Biol. Bull. 199:199-200.
Hayashi KY, Tamaki H, Handa K, Takahashi T,Kakita A, Yamashina S (2003) Differentiationand proliferation of endocrine cells in theregenerating rat pancreas after 90%pancreatectomy. Arch. Histol. Cytol. 66:163-174.
Hedley DW, Friedlander ML, Taylor IW, Rugg CA,Musgrove EA (1983) Method for analysis ofcellular DNA content of paraffin-embeddedpathological material using flow cytometry. J.Histochem. Cytochem. 31:1333-1335.
Henry M (1987) Glande digestive de la palourdeRuditapes decussatus, en métabolisme deroutine. I. La cellule digestive. II La cellulesécrétice. Vie Marine 6:7-24.
Henry M, Bocaud-Camou E, Lefort Y (1991)Functional micro-anatomy of digestive gland ofthe marine bivalves. Mar. Environ. Res. 17:286.
Herrera PL (2000) Adult insulin- and glucagon-producing cells differentiate from twoindependent cell lineages. Development127:2317-2322.
Hickmott PW, Carew TJ (1991) An autoradigraphicanalysis of neurogenesis in juvenile Aplysiacalifornica. J. Neurobiol. 22:313-326.
Hopkin SP, Nott JA (1979) Some observations onconcentrically structured, intracellular garnulesin the hepatopancreas of the shore crabCarcinus maenas (L.). J. Mar. Biol. Assoc. U.K.59:867-877.
Hotchin NA, Gandarillas A, Watt FM (1995)Regulation of cell surface beta 1 integrin levelsduring keratinocyte terminal differentiation J.Cell Biol. Mar. 128:1209-1219.
Howard A, Pelc SR (1950) P32 autoradiographs ofmouse testis; preliminary observations of thetiming of spermatogenic stages. Br. J. Radiol.23:634-641.
Howard B, Mitchell PC, Ritchie A, Simkiss K, TaylorM (1981) The composition of intracellulargranules from the metal-accumulating cells ofthe common garden snail (Helix aspersa).Biochem. J. 194:507-511.
Hunt T, Luca FC, Ruderman JV (1992) Therequirements for protein synthesis anddegradation, and the control of destruction ofcyclins A and B in the meiotic and mitotic cellcycles of the clam embryo. J. Cell Biol. 116:707-724.
Icely JD, Nott JA (1992) Digestion and absortion:digestive system and associated organs. 147-201 orr. Non: Microscopic Anatomy ofInvertebrates. Harrison FW & Kohn AJ (ed). 10;Decapod Crustacea: FW. Wiley-Liss. Inc. NewYork.
Illa I (2005) Un nuevo modelo para el estudio de ladiferenciación de las células madre (stem cells):el nido de regeneracion de Locusta.Doktoradutza-Tesia. Universidad de Navarra.164 orr.
Izagirre U, Ruiz P, Marigómez M (2005) Earlylysosomal membrane destabilisation onexposure to pollutants in mussel digestive cells.13rd Symposium PRIMO.
Jain S, Filipe MI, Hall PA, Waseem N, Lane DP,Levison DA (1991) Prognostic value ofproliferating cell nuclear antigen in gastriccarcinoma. J. Clin. Pathol. 44:655-659.
Janssen HH (1985) Some histological findings ofthe mid-gut gland of the common garden snail,Arion rufus (L.) (Syn. A. ater rufus (L.), A.emporicorum Férrusac), Gastropoda:Stylommatophora. Zool. Anz. 215:33-51.
Jensen UB, Lowell S, Watt FM (1999) The spatialrelationship between stem cells and theirprogeny in the basal layer of human epidermis:

Bibliografia
61
a new view based on whole-mount labelling andlineage analysis. Development 126:2409-2418.
Jensen J (2004) Gene regulatory factors inpancreatic development. Dev. Dyn. 229:176-200.
Joensuu H, Klemi PJ, Korkeila E (1988) Prognosticvalue of DNA ploidy and proliferative activity inHodkin's disease. Am. J. Clin. Pathol. 90:670-673.
Jones PH, Harper S, Watt FM (1995) Stem cellpatterning and fate in human epidermis. Cell80:83-93.
Kamiya K, Higgins PD, Tanner MA, Gould MN,Clifton KH (1999) Kinetics of mammaryclonogenic cells and rat mammary cancerinduction by X-rays or fission neutrons. J.Radiat. Res. 40 Suppl:128-137.
Kikuyama S, Kubora T, Watanabe M, Ishibiki K, AbeO (1988) Cell kinetic study of human carcinomasusing bromodeoxyuridine. Cell Tiss. Kin. 20:1-6.
Kim ND, Oberley TD, Yasukawa-Barnes J, CliftonKH (2000) Stem cell characteristics oftransplanted rat mammary clonogens. Exp. CellRes. 10: 260:146-159.
Klauser DK, Denny PA, Denny PC (1993)Proliferative and structural differences betweenmale and female mouse submandibular glands.Anat. Rec. 235:303-313.
Kordon EC, Smith GH (1998) An entire functionalmammary gland may comprise the progeny froma single cell. Development 125:1921-1930.
Krijgsman, B (1928) Arbeitsrhythmus derVerdauungsdüsen bei Helix pomatia. II.Sekretion, Resorption und Phagocytose. Z.Vergl. Physiol. 8:187-280.
Krishnakumar PK, Casillas E, Vanarasi U (1995)Effect of environmental contaminants on thehealth of Mytilus edulis from Puget Sound,Washington, U.S.A. II. Cytochemical detection ofsubcellular changes in digestive cells. Mar. Biol.124:251-259.
Krucher NA, Roberts MH (1994) Identification ofCDK- and cyclin-like proteins in the eye of Bullagouldiana. J. Neurobiol. 25:1200-1206.
Kurki P, Vanderlaan M, Dolbeare F (1986)Expression of proliferating cell nuclear antigen
(PCNA)/cyclin during the cell cycle. Exp. CellRes. 166:209-218.
Lamers AE, Heiney JP, Ram JL (1999) Cloning andsequence analysis of two cDNAs encodingcyclin A and cyclin B in the zebra musselDreissena polymorpha. Biochim. Biophys. Acta1448:519-24.
Langton RW (1975) Syncrony in the digestivediverticula of Mytilus edulis L. J. Mar. Biol. Ass.U.K. 55:221-229.
Lavker RM, Sun TT (2000) Epidermal stem cells:properties, markers, and location. Proc. Natl.Acad. Sci. U.S.A. 97:13473-13475.
Le Douarin NM (1988) On the origin of pancreaticendocrine cells. Cell 22:169-171.
Leibson NL, Frolova LT (1994) Winter-springessential reorganization of cell proliferation inthe digestive tract epithelia in the musselCrenomytilus grayanus. Mar. Biol. 118: 471-477.
Lekube X (1997) Mytilus galloprovincialismuskuiluaren liseri-guruinaren plastizitatea etadinamika zelularra: 3D-berreraiketa eta PCNAbidezko azterketa. Lizentziatura-tesia.UPV/EHU. 130 orr.
Lekube X, Cajaraville MP, Marigómez I (2000) Useof polyclonal antibodies for the detection ofchanges induced by cadmium in lysosomes ofaquatic organisms. Sci. Tot. Environ. 247:201-212.
Le Pennec G, Le Pennec M (2002) Molecularanalysis of the seasonal expression of genescoding for different functional markers of thedigestive gland of the bivalve mollusk Pectenmaximus (L.). Comp. Biochem. Physiol. B133:417-426.
Li A, Normand P, Redvers R, Kaur P (2004)Extensive tissue-regenerative capacity ofneonatal human keratinocytes stem cells andtheir progeny. J. Clin. Invest. 113:390-400.
Lillo C, Velasco A, Jimeno D, Cid E, Lara JM, AijónJ (2002) The glial design of a teleost optic nervehead supporting continuous growth. J.Histochem. Cytochem. 50:1289-1302.
Littleton RJ, Baker GM; Soombro IN, Adams RL,Whimster WF (1991) Kinetic aspects of Ki-67

Zati Teorikoa
62
antigen expression in a normal cell line. VirchowArch. B Path. Incl. Mol. Path. 60:15-19.
Liu S, Dontu G, Wicha MS (2005) Mammary stemcells, self-renewal pathways, andcarcinogenesis. Breast Canc. Res. 7:86-95.
Loeb MJ, Clark EA, Blacburn M, Hakim RS, ElsenK, Smagghe G (2003) Stem cells from midgutsof Lepidopteran larvae: clues to the regulation ofstem cells fate. Arch. Insect. Biochem. Physiol.53:186-198.
Loizzi RF (1971) Interpretation of crayfishhepatopancreatic function based on finestructural analysis of epithelial cell lines andmussel networks. Z. Zellforsch. Mikrosk. Anat.113:420-440.
van Loon AE, Colas P, Goedemans CE, Neant I,Dalbon P, Guerrier P (1991) The role of cyclinsin the maturation of Patella vulgata oocytes.EMBO J. 10:3343-3349.
Lovett DL, Felder DL (1989) Ontogeny of gutmorphology in the white shrimp Penaeussetiferus (Decapoda, Penaeidae). J. Morphol.201:53-68.
Lowe DM, Moore MN, Clarke KR (1981) Effects ofoil in the digestive cells of mussels: quantitativealterations in cellular and lysosomal structures.Aquat. Toxicol. 1:213-226.
Lowe DM (1988) Alterations in the cellular structureof Mytilus edulis resulting from exposure toenvironmental contaminants under field andexperimental conditions. Mar. Ecol. Prog. Ser.46:91-100.
Luchtel DL, Martin AW, Deyrup-Olsen I, Boer HH(1997) Gastropoda: Pulmonata. 459-718 orr.Non: Microscopic Anatomy of Invertebrates. FWHarrison FW & Kohn AJ (ed). 6B; Mollusca II.Wiley-Liss. Inc.
Lufty RG, Demian ES (1967) The histology of thealimentary system of Marisa cornuarietis(Mesogastropoda: Ampullariidae). Malacologia5:375-422.
Marigómez I, Angulo E, Moya J (1986) Coppertreatment of the digestive gland of the slug Arionater L. 2. Morphometrics and histophysilogy.Bull. Environ. Contam. Toxicol. 36:608-615.
Marigómez I (1989). Aportacion citohistológicas a laevaluación de efectos subletales de cadmio enel medio marino: estudios con el prosobranquioLittorina littorea. Doktoradutza-Tesia. UPV/EHU.430 orr.
Marigómez I, Vega MM, Cajaraville MP, Angulo E(1989) Quantitative responses of the digestivelysosomal system of winkles to sublethalconcentrations of cadmium. Cell Mol. Biol.35:555-562.
Marigómez I, Cajaraville MP, Angulo E (1990)Histopathology of the digestive gland-gonadcomplex of marine prosobranch Littorina littorea(L.) exposed to cadmium. Dis. Aquat. Organ.9:229-238.
Marigómez I, Soto M, Angulo E (1991) Responsesof winkles digestive cell and their lysosomalsystem to environmental salinity changes. CellMol. Biol. 37:29-39.
Marigómez I, Soto M, Angulo E (1992) Seasonalvariability in the quantitative structure of thedigestive tubules of Littorina littorea. Aquat.Living. Resour. 5:299-305.
Marigómez I, Soto M, Etxenberria M, Angulo E(1993) Effects of size, sex, reproduction andtrematode infestation on the quantitativestructure of digestive tubules in stressedwinkles. Zool. J. B. Anat. 123:319-336.
Marigómez I, Cajaraville MP, Quincoces I, SalisburyJR (1995a) Computer assisted 3-Dreconstruction techniques may provide newinsights into the pattern of histologicalorganisation of the bivalvian digestive gland.12th Inter. Malacol. Congr. Vigo.
Marigómez I, Cajaraville MP, Quincoces I, SalisburyJR (1995b) A novel model for the histologicalstructure of the molluscan digestive gland basedon computer aided 3D-reconstruction andscanning microscopy. VI Cong. SEBC. Congr.Lleida.
Marigómez I, Orbea A, Olabarrieta I, Etxeberria M,Cajaraville MP (1996) Structural changes in thedigestive lysosomal system of sentinel musselsas biomarkers of environmental stress inMussel-Watch programmes. Comp. Biochem.Physiol. C 113:291-297.

Bibliografia
63
Marigómez I, Cajaraville MP, Soto M, Lekube X(1998a) Cell-type replacement, a succesfulstrategy of molluscs to adapt to chronicexposure to pollutants. Cuad. Invest. Biol.18:431-435.
Marigómez I, Kortabitarte M, Dussart, GBJ (1998b)Tissue-level biomarkers in sentinel slugs ascost-effective tools to assess metal pollution insoils. Arch. Environ. Contam. Toxicol. 34:167-176.
Marigómez I, Lekube X, Cancio I (1999)Immunochemical localisation of proliferatingcells in mussel digestive gland tissue.Histochem. J. 31:781-788.
Marigómez I, Baybay-Villacorta L (2003) Pollutant-specific and general lysosomal responses indigestive cells of mussels exposed to modelorganic chemicals. Aquat. Toxicol. 64:235-257.
Marigómez I, Soto M, Cancio I, Orbea A, GarmendiaL, Cajaraville MP (2005) Cell and tissuebiomarkers in mussel, and histopathology inhake and anchovy from Bay of Biscay after thePrestige oil spill (Monitoring campaign 2003).Mar. Poll. Bull. (prentsan).
Marshman E, Booth C, Potten CS (2002) Theintestinal epithelial stem cell. BioEssays 24:91-98.
Martínez-Expósito MJ, Pasantes JJ, Méndez J(1994) Proliferation kinetics of mussel (Mytilusgalloprovincialis) gill cells. Mar. Biol. 120: 41-45.
Martynova MG, Bystrova OA (2002)Undifferentiated cells in the snail myocardiumare capable of DNA synthesis andmyodifferentiation. Biol. Bull. 203:104-111.
Mason AZ (1983) The uptake, accumulation andexcretion of metals by the marine prosobranchgastropod mollusc Littorina littorea (L).Doktoradutza-Tesia. University of Wales,Gwyneed (UK). 575 orr.
Mason AZ, Simkiss K (1982) Sites of mineraldeposition in mineral-accummulating cells. Exp.Cell Res. 139:383-391.
McQuiston RW (1969) Cyclic activity in the digestivediverticula of Lasae rubra (Montagu) (Bivalvia:Eulamellibranchia). Proc. Malacol. Soc. London38:483-492.
Melnick M, Jaskoll T (2000) Mouse submandibulargland morphogenesis: a paradigm for embryonicsignal processing. Crit. Rev. Oral Biol. Med.11:199-215.
Merdsoy B, Farley I (1973) Phasic activity in thedigestive gland of the marine prosobranchgastropod, Littorina littorea (L.). Proc. Malacol.Soc. London 40:473-482.
Michalopoulos GK, DeFrances MC (1997) Liverregeneration. Science 276:60-66.
Mix MC (1972) Chronic tissue degeneration in thePacific oyster, Crassostrea gigas, followingacute irradiation. Radiat. Res. 49:176-189.
Mix MC, Sparks AK (1971) Repair of digestivetubule tissue of the pacific oyster, Crassostreagigas, damaged by ionizing radiation. J.Invertebr. Pathol. 17:172-177.
Miyachi K, Fritzler MJ, Tan EM (1978) Autoantibodyto a nuclear antigen in proliferating cells. J.Immunol. 121:2228-2234.
Moore MN, Pipe RK, Farrar SV (1987) Induction oflysosomal lipid accumulation and fattydegeneration by polycyclic aromatichydrocarbons in molluscan digestive cells. Mar.Environ. Res. 17:230-233.
Morasso M, Tomic-Canic M (2005) Epidermal stemcells: the cradle of epidermal determination,differentiation and wound healing. Biol. Cell97:173-183.
Morgan DO (1997) Cyclin-dependent kinases:Engines, Clocks, and Microprocessors. Ann.Rev. Cell Dev. Biol. 13:261-291.
Morris GF, Mathews MB (1989) Regulation ofproliferating cell nuclear antigen during the cellcycle. J. Biol. Chem. 264:13856-13864.
Morton JE (1956) The tidal rhythm and action of thedigestive system of the lamellibranch Lasaearubra. J. Mar. Biol. Ass. U.K. 35:563-586.
Morton BS (1969) Studies on the biology ofDreissenas polymorpha Pal. II. Correlation of therhythms of adductor activity, feeding, digestionand excretion. Proc. Malacol. Soc. London38:401-414.
Morton BS (1970) The tidal rhythm and rhythm offeeding and digestion in Cardium edule. J. Mar.Biol. Assoc. U.K. 50:499-512.

Zati Teorikoa
64
Morton BS (1971) The diurnal rhythm and tidalrhythm of feeding and digestion in Ostrea edulis.Biol.J. Linn. Soc. 3:329-342.
Morton B (1979) The diurnal rhythm and the cycle offeeding and digestion in the slug Derocerascaruanae. J. Zool. 187:135-152.
Morton B (1983) Feeding and digestion in bivalvia.65-147 orr. Non: The Mollusca. 5. AcademicPress. Saleuddin ASM & Wilburg M (ed.) NewYork.
Moya J (1973) Estructura fina de algunos elementosglandulares y sus anejos en el aparato digestivodel limaco común Arion empiricorum, Ferussac(Gasterópodo pulmonado). Universidad deBilbao. Doktoradutza-Tesia.
Moya J, Rallo AM (1975) Intracisternalpolycylinders: a cytoplasmic structure in cells ofthe terrestrial slug Arion empiricorum Férussac(Pulmonata, Stylommatophora). Cell Tiss. Res.159:423-433.
Muller R, Laucke R, Trimper B, Cossel L (1990)Pancreatic cell proliferation in normal ratsstudied by in vivo autoradiography with 3H-thymidine. Virchows. Arch. B. Cell Pathol. Incl.Mol. Pathol. 59:133-136.
Muskhelishvili L, Latendresse JR, Kodell RL,Henderson EB (2003) Evaluation of cellproliferation in rat tissues with BrdU, PCNA, Ki-67(MIB-5) Immunohistochemistry and in situhybridization for histone mRNA. J. Histochem.Cytochem. 51:1681-1688.
Nelson L, Morton JE (1979) Cyclic activity andepithelial renewal in the digestive gland tubulesof the marine prosobranch Maoricryptamonoxyla (Lesson). J. Moll. Stud. 45: 262-283
Nigg EA (1995) Cyclin-dependent protein kinases:key regulators of the eukaryotic cell cycle.BioEssays 17:471-480.
Oka K, Arai T (1996) MIB1 growth fraction is notrelated to prognosis in cervical squamous cellcarcinoma treated with radiotherapy. Int. J. Gyn.Pathol. 15:23-27.
Orci L, Unger RH (1975) Functional subdivision ofislets of Langerhans and possible role of D cells.Lancet 20: 2:1243-1244.
Oro AE, Scott MP (1998) Splitting hairs: dissectingroles of signaling systems in epidermaldevelopment. Cell 25: 95:575-578.
Overturf K, Al-Dhalimy M, Finegold M, Grompe M(1999). The repopulation potential of hepatocytepopulation differing in size and prior mitoticexpansion. J. Am. Pathol. 155:2135-2143.
Owen G (1956) Observations on the stomach anddigestive diverticula of the Lamellibranchia II.The Nuculidae. Q. J. Microsc. Sci. 97:541-567.
Owen G (1966) Digestion. Non: Physiology of themolluscan. 53-96 orr. Wilbur KM & Yonge CM(ed.). 2. Academic Press. New York.
Owen G (1970) The fine structure of the digestivetubules of the marine bivalve Cardium edule.Phil. Trans. R. Soc. London Ser. 60:299-318.
Owen G (1972) Lysosomes, peroxisomes andbivalves. Sci. Prog. 60: 299-318.
Owen G (1973) The fine structure andhistochemistry of the digestive diverticula of theprotobranchiate bivalve Nucula sulcata. Proc. R.Soc. London B. 183:249-264.
Pal SG (1972) The fine structure of the digestiv etubules of Mya Arenaria L. II. Digestive cell.Proc. Malacol. Soc. London. 40:161-170.
Palacios J, Honrado E, Ososio A, Cazorla A, SarrióD, Barroso A, Rodríguez S, Cigudosa JC, DiezO, Alonso C, Lerma E, Dopazo J, Rivas C,Benítez J (2005) Phenotypic characterization ofBRCA1 and BRCA2 tumors based in a tissuemicroarray study with 37 immunohistochemicalmarkers. Breast Cancer Res. Treat. 90:5-14.
Palmer RW (1979) Histological and histochemicalstudy of digestion in the bivalve Arctica islandicaL. Biol. Bull. 156:115-129.
Peatman E, Kucuktas H, Li P, He C, Feng J, Wei X,Liu Z Differentially expressed oyster(Crassostrea virginica) genes after exposure tomercury. 2005. eko uztailan eskuratua.http://www.ncbi.nlm.nih.gov/entrez/viewer.fcgi?db=nucleotide&val=31900849”.
Peruzzi E, Sonetti D (2004) Microglia proliferationas a response to activation in the freshwatersnail Planorbarius corneus: A BrdU incorporationstudy. Acta Biol. Hung. 55:287-291.

Bibliografia
65
Platt AM (1971) Doktoradutza-Tesia. The Queen´sUniversity. Belfast.
Pirger Z, Elekes K, Kiss T (2004) Functionalmorphology of the salivary gland of the snail,Helix pomatia: a histochemical andimmunocytochemical study. Acta Biol. Hung.55:221-232.
Pollard JW, Hennighausen L (1994) Colonystimulating factor 1 is required for mammarygland development during pregnancy. Proc.Natl. Acad. Sci. U.S.A. 91:9312-9316.
Potten CS (1998) Stem cells in gastrointestinalepithelium: Numbers, characteristics and death.Philo. Trans. R. Soc. London Ser. B Biol. Sci.353:821-830.
Potten CS & Loeffler M (1990) Stem cells: attributes,cycles, spirals, pitfalls and uncertainties.Lessons for and from the crypt. Development110:1001-1020.
Potten CS, Booth C, Tudor GL, Booth D, Brady G,Hurley P, Ashton G, Clarke R, Sakakibara S,Okano H (2003) Identification of a putativeintestinal stem cell and early lineage marker;musashi-1. Differentiation 71:28-41.
Prelich G, Tan CK, Kostura M, Mathews MB, SØAG, Downey KM, Stillman B (1987) Functionalidentity of proliferating cell nuclear antigen and aDNA polymerase-δ auxiliar protein. Nature326:517-520.
Quincoces I (1995) Un nuevo modelo de la glánduladigestiva de Mytilus galliprovincialis (Bivalvia,Eulamellibranchia): reconstruccióntridimensional asistida por ordenador ymicroscopía electrónica de barrido.Lizentziatura-Tesia. UPV/EHU. 112 orr.
Rasmussen LPD, Hage E, Karlog O (1983) Lightand electron microscopic studies of the acuteand chronic toxic effects of N-nitrosocompounds on the marine mussel Mytilus edulis(L.). II. N-methyl-N-nitro-N-nitroso guanidine.Aquat. Toxicol. 3:301-311
Reddy AB, Wong GKY, O´Neill J, Maywood ES,Hastings MH (2005) Circadian clocks: Neuraland peripheral pacemakers that impact upon thecell division cycle. Mut. Res. 574:76-91.
Recio A, Marigómez JA, Angulo E, Moya J (1988)Zinc tratment of the digestive gland of the slugArion ater L. 1. Cellular distribution of zinc andcadmium. Bull. Environ. Contam. Toxicol.41:858-864.
Regoli F (1992) Lysosomal responses as a sensitivestress index in biomonitoring heavy metalpollution. Mar. Ecol. Prog. Ser. 84:63-69.
Roach DK (1968) Rhythmic muscular activity in thealimentary tract of Arion ater (L) (Gastropoda:Pulmonata). Comp. Biochem. Physiol. 24:865-878.
Roberts CG, De Fazio A, Tattersal MH (1985) Asimple rapid meted for the identification of cyclincells in freshly excised tumors. Cytobios 43:313-318.
Roberts AB, Sporn MB (1992) Differentialexpression of the TGF-β isoforms inembryogenesis suggests specific roles indeveloping and adult tissues. Mol. Reprod. Dev.32:91-98.
Robinson WE, Langton RW (1980) Digestion in asubtidal population of Mercenaria mercenaria(Bivalvia). Mar. Biol. 58:73-79.
Robinson WE (1983) Assessment of bivalveintracellular digestion based on directmeasurements. J. Moll. Stud. 49:1-8.
Robledo Y, Cajaraville MP (1996) Ultrastructural andcytochemical study of the Golgi vomplex ofmollusca (Mytilus galloprovincialis) digestivecells. Cell Tiss. Res. 284:449-458.
Rudolph KL, Chang S, Millard M, Schreiber-Agus N,DePinho RA (2000) Inhibition of experimentalliver cirrhosis in mice by telomerase genedelivery. Science 287:1253-1258.
Runham NW, Hunter PJ (1970) Terrestrial Slugs.Hutchinson University Library, London. 184 orr.
Sáez V, Marigómez I, Angulo E, Moya J (1990)Histomorphology and histochemistry of thedigestive gland of Littorina littorea (L.) in relationto experimental tidal conditions, food availability,and digestion. Zool. Jb. Anat. 120:185-196.
Salvini-Plawen LV (1988) The structure and functionof molluscan digestive systems. 301-379 orr.Non: The Molluscan. Form and function 11.

Zati Teorikoa
66
Trueman ER & Clarke MR (ed). AcademicPress, Orlando, Florida.
Sancho E, Batlle E, Clvers H (2004) Signalingpathways in intestinal development and cancer.Ann. Rev. Cell Dev. Biol. 20:695-723.
Sanskrithi N, Eskin A (1999) Effects of cyclin-dependent kinase inhibitors on transcription andocular circadian rhythm of Aplysia. J.Neurochem. 72:605-613.
Sarli G, Benazzi C, Preziosi R, Marcato PS (1995)Assessment of proliferative activity by anti-PCNA monoclonal antibodies in formalin-fixed,paraffin-embedded samples and correlation withmitotic index. Vet. Pathol. 32:93-96.
Seaberg RM, Smukler SR, Kieffer TJ, Enikolopov G,Asghar Z, Wheeler MB, Korbutt G, van der KooyD (2004) Clonal identification of multipotentprecursors from adult mouse pancreas thatgenerate neural and pancreatic lineages. Nat.Biotechnol. 22:1115-1124.
Schluter C, Duchowr M, Wohlenberg C, BeckerMHG, Key G, Flad HD, Gerdes J (1993) The cellproliferation-associated antigen of antibody Ki-67: A very large, ubiquitous nuclear protein withnumerous repeated elements, representing anew kind of cell cycle-maintaining proteins. J.Cell Biol. 123:513-522.
Schmetsdorf S, Gärtner U, Arendt T (2005)Expression of cell cycle-related proteins indeveloping and adult mouse hippocampus. Int.J. Dev. Neurosci. 23:101-112.
Scholzen T, Gerdes J (2000) The Ki-67 protein:From the known and the unknown. J. CellPhysiol. 182:311-322.
Scott J, Liu P, Smith PM (1999) Morphological andfunctional characteristics of acinar atrophy andrecovery in the duct-ligated parotid gland of therat. J. Dent. Res. 78:1711-1719.
Sesto A, Navarro M, Burslem F, Jorcano JL (2002)analysis of the ultraviolet B response in primaryhuman keratinocytes using oligonucleotidemicroarrays. Proc. Natl. Acad. Sci. U.S.A.99:2965-2970.
Sherr CJ (1996) Cancer cell cycles. Science274:1672-1677.
Simkiss K (1981) Cellular discrimination processesin metal accumulating cells. J. Exp. Biol. 94:317-327.
Slack JM (1995) Developmental biology of thepancreas. Development 121:1569-1580.
Slack JM (2000) Stem cells in epithelial tissues.Science 287:1431-3.
Smalley M, Ashworth A (2003) Stem cells and breastcancer: a field in transit. Nat. Rev. Canc. 3:832-844.
Sminia T (1981) Gastropods. Non: Invertebrateblood cells. 1. 191-232 orr. Ratcliffe NA &Rowley AF (ed). New York: Academic Press.
Smith GH, Gallahan D, Diella F, Jhappan C, MerlinoG, Callahan R (1995) Constitutive expression ofa truncated INT3 gene in mouse mammaryepithelium impairs differentiation and functionaldevelopment. Cell Growth. Differ. 6:563-577.
Soto M, Agirregoikoa MG, Pérez MA, Marigómez I(1990) A planimetric study of morphologicalvariability in the digestive diverticula of Littorinalittorea (Linnaeus) and Mytilus edulis (Linnaeus).J. Moll. Stud. 56:339-344.
Soto M, Cajaraville MP, Marigómez I (1996) Tissueand cell distribution of copper, zinc and cadmiumin the mussel, Mytilus galloprovincialis,determined by autometallography. Tiss. Cell28:557-568.
Soto M, Marigómez I (1997) Metal bioavailabilityassessment in "Mussel-Watch" programmes byautomated image analysis of BSD in digestivecell lysosomes. Mar. Ecol. Prog. Ser. 156:141-150.
Soto M, Zaldibar B, Cancio I, Taylor MG, Turner M,Morgan AJ, Marigómez I (2002) Subcellulardistribution of cadmium and its cellular ligands inmussel digestive gland cells as revealed bycombined autometallography and X-raymicroprobe analysis. Histochem. J. 34:273-280.
Spradling A, Drummond-Barbosa D, Kai T (2001)Stem cells find their niche. Nature 414:98-104.
Sumner AT (1965) The cytology and histochemistryof the digestive gland cells of Helix. Q. J.Microsc. Sci. 106:173-192.

Bibliografia
67
Sumner AT (1966) The fine structure of the digestivegland of Helix, Succinea and Testacella. J. Roy.Microsc. Soc. 85:181-192.
Sumner AT (1969) The distribution of some hydrliticenzymes in cells of the digestive gland of certainlamellibranchs and gastropods. J. Zool. 158:277-291.
Suzuki A, Nakauchi H, Taniguchi H (2004)Prospective isolation of multipotent pancreaticprogenitors using flow-cytometric cell sorting.Diabetes 53:2143-2552.
Syasina IG, Vaschenko MA, Zhandan PM (1997)Morphological alterations in the digestivediverticula of Mizuhopecten yessoensis(Bivalvia: Pectenidae) from polluted areas ofPeter the Great Bay, Sea of Japan. Mar. Environ.Res. 44:85-98
Takahashi S, Nakamura S, Suzuki R, Islam N,Domon T, Yamamoto T, Wakita M (2000).Apoptosis and mitosis of parenchyal cells in theduct-ligated rat submandibular gland. Tiss. Cell32:457-463.
Takahashi S, Nakamura S, Suzuki R, Shinzato K,Domon T, Yamamoto T, Wakita M (2001)Apoptosis and mitosis of myoepithelial cells inatrophic rat submandibular glands. J.Histochem. Cytochem. 49:1557-1563.
Takahashi S, Schoch E, Walker NI (1998) Origin ofacinar cell regeneration after atrophy of the ratparotid induced by duct obstruction. Int. J. Exp.Pathol. 79:293-301.
Takahashi S, Shinzato K, Nakamura S, Domon T,Yamamoto T, Wakita M (2002) The roles ofapoptosis and mitosis in atrophy of theraltsublingual gland. Tiss. Cell 34:297-304.
Theise ND, Saxena R, Portmann, BC, Thung SN,Yee H, Chiriboga L, Kumar A, Crawford JM(1999) The canals of Hering and hepatic stemcells in humans. Hepatology 30:1425-1433.
Thompsom RJ, Ratcliffe NA, Bayne BL (1974)Effects of starvation on structure and function inthe digestive gland of the mussel (Mytilusedulis). J. Mar. Biol. 54:699-712.
Till JE, McCulloch EA (1961) A direct measurementof the radiation sensitivity of normal mouse bonemarrow cells. Radiat. Res.14:213-222.
Tosh D, Strain A (2005) Liver stem cells-prospectsfor clinical use. J. Hepatol. 42:75-84.
Tomasovic SP, Mix MC (1974) Cell renewal in thegill of the freshwater mussel Margaritiferamargaritifera: an autoradiographic study usinghigh specific activity tritiated thymidine. J. CellSci. 14:561-569.
Tomic-Canic M, Agren MS, Alvarez OM (2004)Epidermal repair and the chronic wound. Non:The epidermis in wound healing. 25-57 orr.Rovee D & Maibach H (ed). CRC Press, U.S.A.
Topper YJ, Freeman CS (1980) Multiple hormoneinteractions in the developmental biology of themammary gland. Physiol. Rev. 60:1049-1106.
Toschi L, Bravo R (1988) Changes incyclin/proliferating cell nuclear antigendistribution during DNA repair synthesis. J. CellBiol. 107:1623-1628.
Totafurno J, Bjerkes M, Cheng H (1987) The cryptcycle. Crypt and villus production in the adultintestinal epithelium. Biophys. J. 52:279-294.
Triebskorn R (1991) The impact of molluscicides onenzyme activities in the hepatropancreas ofDeroceras reticulatum (Müller). Malacologia 33:255-272.
Triebskorn R, Kohler HR (1996) The impact ofheavy metals on the grey garden slug,Deroceras reticulatum (Muller): Metal storage,cellular effects and semi-quantitative evaluationof metal toxicity. Environ. Poll. 93:327-343.
Vega MM, Marigómez I, Angulo E (1989)Quantitative alterations in digestive cell structureof the marine gastropod Littorina littorea onexposure to cadmium. Mar. Biol. 103: 547-553.
Venier P, Pallavicini A, De Nardi B, Lanfranchi G(2003) Towards a catalogue of genestranscribed in multiple tissues of Mytilusgalloprovincialis. Gene 314:29-40.
Visakorpi T (1992) Proliferative activity determinedby DNA flow cytometry and proliferating cellnuclear antigen (PCNA) immunohistochemistryas a prognostic factor in prostatic carcinoma. J.Pathol. 168:7-13.
Vogt G (1994) Life-cycle and functional cytology ofthe hepatopancreatic cells of Astacus astacus.Crustacea, Decapoda. Zoomorphol. 114:83-101.

Zati Teorikoa
68
Voltzow J (1994) Gastropoda: Prosobranchia. Non:Microscopic Anatomy of Invertebrates. 5. 111-252 orr. Harrison FW & Kohn AJ (ed). Wiley-Liss. Inc.
Walker G (1969) Studies on digestion of the slugAgriolimax reticulatus (Müller) (Mollusca,Pulmonata, Limacidae). Doktoradutza-Tesia,University of Wales.
Walker G (1971) The cytology, histochemistry, andultrastructure of the cell types found in thedigestive gland of slug, Agriolimax reticulatus.Protoplasma 71: 91-109.
Walker G (1972) The digestive system of the slug,Agriolimax reticulatus (Müller): Experiments onphagocytosis and nutrient absortion. Proc.Malacol. Soc. 40:33-43.
Waseem NH, Lane DP (1990) Monoclonal antibodyanalysis of the proliferating cell nuclear antigen(PCNA). Structural conservation and thedetection of a nuclear form. J. Cell Sci. 96:121-129.
Watt FM (2000) Epidermal stem cells as targets forgene transfer. Hum. Gene Ther. 16:2261-2266.
Weinstein J, Jacobsen FW, Hsu-Chen J, Wu T,Baum LG (1994) A novel mammalian protein,p55CDC, present in dividing cells is associatedwith protein kinase activity and has homology tothe Saccharomyces cerevisiae cell division cycleproteins Cdc20 and Cdc4. Mol. Cell Biol.14:3350-3363.
Wells JM (2003) Genes expressed in the developingendocrine pancreas and their importance forstem cell and diabetes research. DiabetesMetab. Res. Rev. 19:191-201.
Widdows J, Moore MN, Lowe DM. 1984. Sublethalbiological effects and short-term recovery ofmussels (Mytilus edulis) and winkles (Littorinalittorea) following chronic exposure to petroleumhydrocarbon. Non: Long term effects of oil onmarine benthic communities in enclosures.Norwegian Institute for Water Research/NIVATechnical Report No. 0-8200.
Wood RD, Shivji MKK (1997) Which DNApolymerases are used for DNA-repair ineukaryotes? Carcinogenesis 18:605-610.
Yamada K, Yoshitake K, Sato M, Ahnen DJ (1992)Proliferating cell nuclear antigen expression innormal, preneoplastic, and neoplastic colonicepithelium of the rat. Gastroenterology 103:160-167.
Yanagi S, Potter VR (1977) Sequential changes inornithine decarboxylase thymidine kinase, andother enzyme activities in regenerating liver inrats on controlled feeding schedules. Life Sci.1:1509-1519.
Yonge CM (1926) The digestive diverticula inLamellibranchia. Trans. Roy. Soc. Edinbourgh54: 703-718.
Yoshino TP (1976) Histopathological effects of larvaldigenea on the digestive epithelia of the marineprosobranch Cerithidea californica: finestructural changes in the digestive gland. J.Invert. Pathol. 28:209-216.
Yu CCW, Hall PA, Fletcher CDM, Camplejohn RS,Waseem NH, Lane DP, Levison DA (1991)Haemangiopericytomas: The prognostic value ofimmunohistochemical staining with amonoclonal antibody to proliferating cell nuclearantigen (PCNA). Histopathol. 19:29-33.
Zakharov IS, Hayes NL, Ierusalimsky VN,Nowakowsky RS, Balaban PM (1998)Postembryonic neurogenesis in theprotocerebrum of the terrestrial snail, Helixlucorum L. J. Neurobiol. 35:271-276.
Zhu MH, Ni CR, Zhu Z, Li FM, Zhang SM (2003)Immunohistochemical demonstration of cyclinsA, B, D1, D3 and E in hepatocellular carcinomasusing tissue microarrays. Zhonghua. Bing. Li.Xue. Za. Zhi. 32:440-443.

AUZIAREN EGOERA
Moluskuen liseri-guruina eraenketa metabolikoaren organo nagusia da. Horrela,
sistema immunean eta barne medioaren (pH, Ca-kontzentrazioa…) eraenketa
homeostatikoan parte hartzen du, eta elikagaien liseriketa intrazelular eta estrazelularraren
zein hondakinen jariapenaren arduradun nagusia da. Dena den, eta zati teorikoan azaldu
den moduan, moluskuen liseri-guruinaren epitelioa zelula-mota urriz osaturik dago, 2 edo
3 zelula-mota. Liseri-guruina, gainera, estres peko egoera desberdinetan modu
neurgarrian erantzuteko gai da, liseri-epitelioaren lodieraren murrizpenarekin edota liseri-
zelulen lisosomen tamaina eta osagaien aldaketarekin besteak beste. Hala ere, liseri-
guruineko epitelioan gertatutako aldaketen artean garrantzitsuena beharbada zelula-
moten ordezkapena da. Horrela, zelula basofilikoen dentsitate erlatiboa liseri-guruinean
liseri-zelularenaren gainetik emendatu egiten da. Dena den, prozesu hori liseri-zelulen
galera, zelula basofilikoen proliferazioa edo bi prozesuen elkarrekintzak sortua den argitu
beharra dago. Liseri-guruinaren antolakuntza-eredua desberdina da molusku espezie
desberdinen artean eta osaketa zelularra ere desberdina da. Horretaz gain, modelo
desberdinek erakusten duten plastikotasuna ere desberdina da, hori dela eta nagusiki
muskuilu itsastarrean deskribatu diren aldaketak beste molusku-ereduetan orokortu
daitezkeen ere ezagutzea komenigarria litzateke.
Ugaztunen epitelio-ehun desberdinetan kalteren baten eraginez, epitelio-zelulen
galera esanguratsua gertatzekotan, epitelioaren berreskurapena bi modu desberdinetan
ematen da nagusiki; epitelio-ehunen zelula amen zatiketaren bidez, edo oraindik ere
epitelioan darraiten zelula helduen zatiketen bidez. Moluskuen liseri-guruinean, hala ere,
zelula amen presentzia oraindik ere argitzeko dago, eta beraz beharrezkoa zaigu epitelio
honen berriztapena nola ematen den ulertzea, eta baita ere moluskuen bizi-zikloan zehar
berriztapena nola eraentzen den ezagutzea.
Bestalde, moluskuak ingurugiro kutsaduraren organismo zentinela gisa aspaldi
erabiliak izan dira. Izan ere, beraien ehunetan kutsatzaileak metatzen dituzte, eta
kutsatzaileen aurrean modu neurgarrian erantzuteko gai dira. Erantzun neurgarri hauen
artean biomarkatzaileak ditugu. Biomarkatzaileak antolakuntza-maila xinpleetan ematen
diren aldaketen neurketak dira, beranduago eta antolakuntza-maila konplexuagoetan
gertatuko diren aldaketak aurresateko baliogarriak izango zaizkigunak. Hori dela eta,
organismo hauen gainean, eta batez ere beraien liseri-guruinean, hainbat neurketa
biologiko (biomarkatzaileak) zein kimiko (biometaketa) burutzen dira. Liseri-guruinean
gertatzen diren zelula-moten aldaketek, beraz, eragin zuzena eduki dezakete ingurugiro-
71
Hipotesi eta helburuak

kalitatearen jarraipen programetan egindako hainbat neurketetan. Hortaz, zelula-moten
ordezkapenak neurketa hauetan hainbat alditan behatutako emaitza eztabaidatsuetan
eragina izan ote dezaken argitu beharra dago.
HIPOTESIA
Moluskuen liseri-guruineko epitelioaren berriztapena zelula helduen zatiketa
autologoz burutzen da eta aldagai naturalen pean alda daiteke. Esaterako, liseri-epitelioan
ingurumen-estresak bultzatutako zelula-moten ordezkapena, zelula basofilikoen
proliferazio areagotuak eragiten du. Bederen, zelula-moten osaketaren aldaketa horrek,
kutsaduraren jarraipen-programetan erabilitako zenbait biomarkatzaileen gainean eragina
du.
HELBURUAK
Aipatutako hipotesia frogatzeko asmoz, ikerlan honetarako hurrengo helburu
orokor hauek proposatu ditugu:
1.- Moluskuen liseri-guruinaren epitelioaren berriztapena zelula amen edo zelula
helduen zatiketaz ematen den argitzea.
2.- Epitelioaren berriztapenaren dinamika ezagutzea eta beren gain zeintzu
faktorek eragiten duten zehaztea.
3.- Moluskuen liseri-guruinean gertatzen den zelula-moten ordezkapena, zelula
basofilikoen proliferazioak eraginda dagoen antzematea.
4.- Aldaketa horiek kutsaduraren jarraipenerako erabiltzen diren ohiko hainbat
biomarkatzaileen gainean nolako eragina duten zehaztea.
Ikerlanaren helburu orokor horiek segidan azalduko diren eta “Emaitzak eta
Eztabaida” atalean ekin diegun honako helburu espezifiko hauetan xehatu dira.
1.- Kutsatzaile metaliko zein organikoek, muskuiluen liseri-guruineko epitelioan
zelula-moten ordezkapena noiz, nola eta zein puntutaraino eragin dezaketen jakitea.
2.- Muskuiluen liseri-guruineko epitelioko zelula-moten ordezkapenak
autometalografiaz errebelatutako zilarrezko hauspeakin beltzen hedapena gisako
biomarkatzaileen gainean nolako eragina duen zehaztea.
72
Hipotesi eta helburuak

3.- Muskuiluen liseri-guruineko epitelioaren berriztapenak patroi zirkadianoa edo
zirkamareala jarraitzen ote duen zehaztea.
4.- Tamaina, sasoi eta itsasaldi-erregimena gisako faktore naturalek Mytilus
galloprovincialis (Lmk) muskuiluen liseri-albeolo eta urdaileko zelulen jarduera
proliferatzailean nolako eragina duten zehaztea
5.- Kadmioz kutsatutako magurioetan zelula-moten ordezkapena liseri-zelulen
kopuruaren murrizpen zuzenarekin edota zelula basofilikoen proliferazioarekin
erlazionatuta ote dagoen zehaztea.
6.- Balizko zelula-moten ordezkapen horrek metalen esposizio eta efektu
bilogikozko biomarkatzaileen gainean eraginik ote duen antzematea.
7.- Kutsatzaile organiko eta metalikoen nahasketa pean egondako bareen liseri-
guruinean zelula-moten ordezkapena liseri-zelulen galerarekin edo kaltzio-zelulen
proliferazioarekin erlazionatuta dagoen eta prozesu hori itzulgarria ote den zehaztea.
8.- Prozesu horrek konposatu kimiko horien biometaketan eta efektu eta esposizio-
biomarkatzaileetan nola eragiten duen finkatzea.
9.- Kronikoki kutsadura pean egondako bareen liseri-guruineko osaketa zelularra
zein den zehaztea eta toki garbira lekuzaldatu ondoren liseri-guruinaren osaketa zelularrari
zein biomarkatzaileen erantzunei dagokionez, kutsadura kroniko horrek sorterazitako
aldaketak norainoko eragina izan dezaketen esagutzea.
73
Hipotesi eta helburuak

74
Hipotesi eta helburuak

Zelula-moten ordezkapena kutsatzaileen peanjarritako muskuiluen liseri-guruinean
Laburpena: Mytilus galloprovincialis (Lmk) muskuiluen liseri-epitelioa bi zelula-motez osaturik dago,liseri-zelula eta zelula basofilikoa. Egoera normaletan liseri-zelulak zelula basofilikoak baino ugariagoakdira, baina kutsatzaileen eraginez zelula basofilikoen proportzio erlatiboaren emendioa deskribatu da.Alterazio horri, zelula-moten ordezkapena (CTR; ingeleraz, cell-type replacement) deritzo.Autometalografia (AMG), metalak zeluletan lokalizatzearen bidez metal kutsatzaileen ingurune-mailakebaluatzeko erabiltzen den teknika histokimiko sentikorra da, metalak zilarrezko hauspeakin beltz (BSD;ingeleraz, black silver deposits) gisa liseri-zelulen lisosometan agertzen direlarik. Ikerlan honen helburuakhiru dira. Lehenik, Cd moduko kutsatzaileek CTR eragin dezakeenentz jakitea. Bigarrenik, AMGzerrebelatutako Cd-ren kokapena zelula-espezifikoa den ultrastruktura-mailan egiaztatzea eta, horrelabalitz, AMGren bidez egindako estimazioen gain CTRk zer nolako eragina duen ezagutzea. Azkenik, CTRgerta dadin kutsatzaile desberdinen zein kontzentrazio beharrezkoak diren eta prozesua itzulgarria denfinkatzea. Helburu horiek lortzeko, muskuiluak kutsatzaile organiko eta ez-organikoen kontzentrazio peaneta denbora-tarte desberdinetan mantendu dira laborategiko bost esperimentu osagarritan. CTRkuantifikatzeko, liseri-guruinaren parafinazko ebakietan zelula basofilikoen dentsitate bolumetrikoa(VvBAS) estimatu da estereologiaren bidez. Era berean, BSDen dentsitate bolumetrikoa (VvBSD) irudi-
analisia erabiliz kalkulatu da. Horrela, Cd-k CTR eragiten duela demostratu dugu. Bestalde, BSD liseri-zelulen lisosomen barruan eta beren mintzaren inguruan lokalizatu dira espezifikoki, eta inoiz ez zelulabasofilikoetan. BSD, metal espezifikoak ez diren arren, S-ri asoziatutako Cd ioien inguruan eratuak direlaX izpien mikroanalisia eta mapaketen bidez erakutsi dugu. Lokalizazio zelula-espezifiko horren ondorioz,CTRk VvBSD balioen gainean eragina duela frogatu dugu, ingurumen-metalen bioeskuragarritasunaren
estimazioak egiterakoan arazoak ekar litzakeena. Bestetik, kutsatzaile organiko, ez-organiko zein beraiennahasketek CTR eragin dezaketela ikusi dugu, kutsatzaile desberdinen kontzentrazio eraginkorradesberdina den arren.
Gako-hitzak: zelula-moten ordezkapena, autometalografia, kadmioa, benzo(a)pirenoa, X izpienmikroanalisia, muskuilua, liseri-guruina.
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
77

Cell-type replacement in the digestive gland of mussels exposed topollutants
Abstract: The digestive gland epithelium of the mussel Mytilus galloprovincialis (Lmk) is comprised oftwo cell types, namely digestive and basophilic cells. Under normal circumstances digestive cellsoutnumber basophilic cells but under the influence of pollutants a relative increase in the proportion ofbasophilic cells has been described. This alteration has been termed cell-type replacement (CTR).Autometallography (AMG) is a sensitive histochemical technique to localize metals in cells as black silverdeposits (BSD) and has been used in the assessment of environmental metal pollution. BSD are localizedin the digestive cells lysosomes. The objectives of this work are three. First, to know whether pollutants suchas Cd were able to induce CTR. Second, to check wether Cd localization revealed by AMG at theultrastructural level were cell-specific and, if so, to determine wether CTR influenced the estimations madeby AMG. Finally, to determine the effective concentration of various pollutants to provoke CTR and to checkwether CTR is reversible. For these purposes mussels were exposed to different concentrations of organicand inorganic pollutants at various exposure-times in five complementary laboratory experiments. In orderto quantify CTR, volume density of basophilic cells (VvBAS) was estimated on hematoxylin-eosin stained
sections by means of stereology. Similarly, the volume density of BSD (VvBSD) was calculated with the aid
of an image analysis. Results revealed that Cd provoked CTR. On the other hand, while BSD werespecifically localized within digestive cells lysosomes, basophilic cells were devoid of BSD. X-raymicroprobe analisys and X-ray mapping demonstrated that although BSD are not metal specific they weremainly formed around the Cd associated to S atoms. Due to this specific localization, CTR influencesVvBSD values, and therefore when environmental bioavailable metal levels are estimated this must be
taken into account. On the other hand, organic and inorganic pollutants or their mixtures can provoke CTRalthough critical concentrations may change.
Key Words cell-type replacement, autometallography, cadmium, benzo(a)pyrene, X-ray microanalysis,mussel, digestive gland.
Emaitzak eta eztabaida
78

Sustitución de tipos celulares en la glándula digestiva de mejillonesexpuestos a contaminantes
Resumen: El epitelio de la glándula digestiva del mejillón Mytilus galloprovincialis (Lmk) estáconstituido por dos tipos celulares, denominados célula digestiva y célula basófila, respectivamente. Encircunstancias normales las células digestivas son más numerosas que las basófilas pero bajo la influenciade contaminantes se ha descrito un aumento en la proporción relativa de células basófílas. Ésta alteraciónha sido denominada sustitución de tipos celulares (CTR; del inglés, cell-type replacement). Laautometalografía (AMG) es una técnica histoquímica sensible que mediante la localización de metales enlas células ha sido utilizada en la evaluación de la contaminación metálica del medio ambiente detectandolos metales como depósitos negros de plata (BSD; del inglés, black silver deposits) en los lisosomas de lascélulas digestivas. El presente estudio tiene tres objetivos. Primero, saber si contaminantes como el Cd soncapaces de provocar CTR. Segundo, determinar si el Cd revelado mediante AMGa nivel ultrastructural selocaliza específicamente en un tipo celular, y si así fuera, conocer si CTR tiene alguna influencia en lasestimaciones hechas sobre los datos obtenidos mediante AMG. Finalmente, establecer lasconcentraciones necesarias por diferentes contaminantes para provocar CTR y determinar si CTR es unproceso reversible. Para cumplir estos objetivos se han diseñado cinco experimentos de laboratoriocomplementarios en los que se han expuesto mejillones a diferentes concentraciones durante diferentesperiodos de tiempo a contaminantes orgánicos e inorgánicos. Para cuantificar CTR se ha estimado ladensidad volumetrica de células basofílicas (VvBAS) en cortes de parafina mediante esterología. Del
mismo modo, se ha calculado la densidad volumétrica de BSD (VvBSD) mediante análisis de imagen. Los
resultados demuestran que el Cd provoca CTR. Por otro lado, mientras los BSD se localizan asociados ala membrana de los lisosomas de las células digestivas, las células basofílicas no muestran BSD. Medianteel empleo de microanálisis y mapeo de rayos X se ha demostrado que aunque los BSD no son específicospara cada tipo de metal, éstos se forman mayormente alrededor de los átomos de Cd asociados a S.Debido a esta localización específica, CTR influye sobre los valores de VvBSD y por lo tanto puede crear
problemas al estimar la biodisponibilidad metálica en el medio ambiente. Por otro lado, los contaminantesorgánicos, inorgánicos y la mezcla de éstos provocan CTR aunque las concentraciones efectivas sondiferentes en cada caso.
Palabras clave: sustitución de tipos celulares, autometalografía, cadmio, benzo(a)pireno, microanálisis derayos X, mejillón, glándula digestiva.
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
79

Emaitzak eta eztabaida
80

Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
81
SARRERA
Muskuiluen liseri-guruinaren epitelioa bi zelula-
motez osaturik dago: liseri-zelulak eta zelula
basofilikoak (Morton, 1983). Liseri-zelulek oso
sistema endo-lisosomiko garatua dute, eta batez
ere barne-liseriketaz arduratzen diren arren,
kutsatzaileen metaketan eta detoxifikazioan ere
parte har dezakete (Viarengo, 1989; Cajaraville et
al., 1995). Zelula basofilikoek duten funtzioa
momentuz argitzeke dago, baina liseriketa
estrazelularrean parte har dezakete, horretarako
beharrezkoak diren entzimak sortu eta jariatzen
dituztelarik. Izan ere, beraien erretikulu
endoplasmatiko pikortsua oso garatua dago, eta
horrek beren basofilia bereizgarria emango lieke
(Cajaraville et al., 1990a; Marigómez et al., 1998;
Dimitriadis et al,. 2004). Egoera normaletan liseri-
zelulak zelula basofilikoak baino ugariagoak dira;
baina inguruneko estres-iturri desberdinek,
kutsatzaileek sortutako estresak barne,
epitelioaren osaketan aldaketak sortarazi ditzakete,
zelula basofilikoen proportzio erlatiboa emendatuz
(Thompson et al.,1974; Rasmussen et al., 1983;
Cajaraville et al., 1990a; Marigómez et al., 1990a;
Syasina et al., 1997; Marigómez et al., 1998).
Estresak induzitutako alterazio hori, zelula-moten
ordezkapena (CTR; ingeleraz, cell-type
replacement) izenaz ezagutzen dena, liseri-zelulen
galerak, zelula basofilikoen proliferazioak edo bi
prozesu horien konbinaketak sortarazia izan
daitekeela proposatu da (Marigómez et al., 1990a).
Muskuiluen kasuan, N-nitroso konposatuek
(Rasmussen et al., 1983), eta petrolioen eta olio
lubrifikatzailearen uretan egokitutako frakzioak
(WAF; ingeleraz, water accomodated fraction;
Cajaraville et al., 1990a) CTR induzi dezakete.
Halaber, zelai-ikerketen arabera, metal-kutsadura
duten lekuetako moluskuen liseri-guruinean ere
CTR behatu da (Soto & Marigómez,1997a; Syasina
et al., 1997).
Zenbait metalek (Cd, Cu, Zn) eta konposatu
organikok (benzo(a)pirenoa -BaP-), zein beren
nahasketek CTR eragin dezaketen zehaztea dugu
ikerlan honen lehengo helburu. Gainera,
kutsatzaile bakoitzaren kontzentrazio eraginkorra
zein den ere finkatu nahi dugu. Bestalde, CTR
prozesuaren mekanismoak ezagutzeko nahian,
estres-iturria eten ondoren prozesua itzulgarria
denentz jakitea ere planteatu dugu. Azkenik, CTR
pairatu duten muskuiluek (adb., zelaian leku
kutsatuan jasotakoak) laborategian berriro metalen
pean jarriz gero, CTRri dagokion
erantzunkortasuna ere aztertu nahi izan dugu.
Ingurumen-toxikologian, metal kutsatzaileen
maila bioeskuragarrien (esposizioa) zein beren
efektu biologikoak neurtzeko biomarkatzaileak
deritzen neurketak aplikatu ohi dira (Cajaraville et
al., 2000). Esposizio-biomarkatzaileen artean,
zeluletan metatutako metalak kuantifikatzeko oso
sentikorra den autometalografia (AMG) izeneko
teknika histokimikoa dugu. Teknika horren
ezaugarri nagusia, duen izugarrizko sentikortasuna
da, seinale ikuskorra emateko hamar atomo
katalitikoren presentzia nahikoa izan daitekeelarik
(Danscher, 1984). Beraz, moluskuen ehunetan
metalak argi-mikroskopioan lokalizatzeko nahiko
erabilia izan da. AMG aplikatu ondoren, Cu, Zn, Hg

eta Cd moduko metalak zilarrezko hauspeakin beltz
(BSD; ingeleraz, black silver deposits) moduan
agertzen dira markaturik, batez ere liseri-zelulen
lisosometan (Hemelraad & Herwig, 1988; Herwig et
al., 1989; Soto et al., 1996a; 1996b; Soto &
Marigómez, 1997a; 1997b; Domouhtsidou &
Dimitriadis, 2000; Marigómez et al., 2002).
Tamalez, teknika hau oso sentikorra den arren, ez
da batere espezifikoa, metal ezberdinak banan-
banan ezin baitira identifikatu. Beraz, BSDen
presentzia ezin da metal jakin baten kokapenarekin
erlazionatu. Testuinguru honetan, Cd pean
mantendutako animalietan BSDak espezifikoki Cd
ioien inguruan sortzen direla frogatu nahi izan dugu,
elektroi-zunden mikroanalisi (EPMA; ingeleraz,
electron probe microanalysis) moduko teknika
mikroskopiko osagarriak AMGrekin batera erabiliz.
AMG ez bezala, EPMA oso teknika espezifikoa da,
zelulen osaketa elemental zehatza
karakterizatzeko erabilia izan dena (Marigómez et
al., 1990a; Morgan & Morgan, 1998; Dimitriadis &
Papadaki, 2004). Aldi berean, EPMAren emaitzak,
Ag eta Cd moduko metal-ioiak kolokalizatzeko oso
teknika egokia den X izpien mapaketaren bidez
(Morgan & Morgan, 1998) baieztatzea planteatu
dugu.
Bestalde, AMG argi-mikroskopioan aplikatzean
metalak identifikatu ezin baditugu ere,
errebelatutako BSD-hedapena metatutako metalen
mailaren arabera emenda daiteke, bere
kuantifikazioak metal bioeskuragarrien maila
adieraz dezakeelarik (Soto & Marigómez, 1997b).
Izan ere, liseri-lisosometako BSD-hedapena irudi-
analisi bitartez kuantifikatuz, BSDen dentsitate
bolumetrikoa (VvBSD) deritzon parametroa kalkula
daiteke. VvBSD metal-esposizioaren
biomarkatzaile egokia da, zenbait espezie
desberdinetan ikusi denez (Soto & Marigómez,
1997a, 1997b; Da Ros et al., 2000; Porte et al.,
2001).
Hala ere, biomarkatzaile horren aplikazioari
dagokionez, CTR fenomenoa aintzakotzat hartu
beharrekoa da, metala metatzen duten liseri-zelulak
zelula basofilikoz ordezkatuak baitira, metalak
metatzeko liseri-guruinak duen gaitasuna aldarazita
suertatu daitekeelarik (Soto & Marigómez, 1997b;
Marigómez et al., 2002). Izan ere, metalak metatuz
doazen heinean, VvBSD balioek ez dute zertan
gora egin behar; metala metatzen ez duten zelula
basofilikoak, liseri-zelulak baino ugariak bilaka
baitaitezke. Hots, atalase batetik aurrera metalen
pean jarritako muskuiluetan liseri-epitelioan ematen
den metaleen metaketa zelula-moten
proportzioaren arabera alda liteke (Marigómez et
al., 1990b; 1998). Hori dela eta, VvBSD moduko
biomarkatzaileen gainean CTRk zer nolako eragina
izan dezaken jakitea dugu azken helburu.
Sarrera honetan aipatutako helburuak
erdiesteko, bost esperimentu desberdin diseinatu
ditugu. Guztietan Mytilus galloprovincialis
muskuiluak laborategiko baldintzen pean
kutsatzaile desberdinez tratatu dira, denbora tarte
eta kontzentrazio desberdinetan. Lehenengo
esperimentuan, Plentziatik hartutako muskuiluak
CTR induzi lezaken Cd kontzentrazio pean
mantendu ziren 21 egunez, eta VvBAS eta VvBSDdenbora tarte desberdinetan kalkulatu ziren.
Halaber, Cd-aren lokalizazio ultrastrukturala eta
Emaitzak eta eztabaida
82

Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
83
BSDen azterketa elementala burutu ziren, Cd-aren
lokalizazioa zelula-espezifikoa dela ikertzeko
asmoz. Bigarrenean, Plentziako muskuiluak, CTR
induzitzen ez duten Cd kontzentrazioak aurkitu
nahian, Cd kontzentrazio baxuago pean mantendu
ziren, eta VvBAS zein VvBSD kalkulatu ziren.
Hirugarren esperimentuan, Cu, Cd eta Zn bidezko
tratamenduen arteko konparaketari ekin zitzaion,
hiru metalen CTR induzitzeko gaitasuna eta
VvBSD balioen gaineko eragina alderatuz. Halaber
esperimentu honetan, muskuiluak hiru metal
horiekin 7 egunez tratatu ondoren, CTR itzulgarria
den jakiteko asmoz, 14 eguneko arazketa-aldia
aplikatu zen. Laugarren esperimentuan, Plentziako
muskuiluak, Benzo(a)pireno (BaP) eta Cd + BaP
pean mantendu ziren 21 egunez, kutsatzaile-mota
desberdinen arteko elkarrekintzak CTRren gainean
daukan eragina aztertzeko asmoz. Azkenik,
bosgarren esperimentuan, aldez aurretik
estresatutako muskuiluak (Zn-ez kutsatuak,
Meñakozetik hartuak), laborategira ekarri eta Cd
kontzentrazio desberdinen pean mantendu ziren 41
egunez, esposizio esperimentalak CTR areago
dezakeenentz ikertzeko.
MATERIAL ETA METODOAK
Erreaktibo kimikoak
Erabilitako erreaktibo kimiko guztiak maila
analitikokoak eta Sigma-Aldrich (St. Louis,
Missouri, Amerikako Estatu Batuak), merkatal-
etxekoak dira besterik agertu ezean.
Prozedura esperimentala
Bestelakorik esan ezean esperimentuetan
erabilitako muskuilu (Mytilus galloprovincialis Lmk)
mareartekoak, Plentzian (Bizkaia) (46º 26´I, 2º
55´M) hartu eta berehala laborategira garraiatu
ziren. Bertan, muskuiluak, argi ultramore bidez
esterilizatutako eta karbono aktiboz zein beirazko
zuntzetan zehar iragazitako itsas-uretan mantendu
ziren 7 egunez, tenperatura egonkorrean (15-
16ºC). Muskuiluak baraurik mantendu ziren,
laborategiko egoerara ohitu egin zitezen, eta
urdailean zeuzkaten hondakinak hustu zitzaten,
kutsatzaile peko tratamenduak hasi aurretik
(moldarazte-tartea). Muskuiluak, esperimentuak
iraun bitartean, fotoperiodo natural pean mantendu
ziren, eta ornogabe itsastarrentzako jaki
komertzialez (Marine Invertebrate Diet, Carolina,
Burlington, North Carolina, Amerikako Estatu
Batuak) elikatu ziren. Etengabeko aireztapena
aplikatu zen, eta ura egunero aldatu zen.
Lehenengo esperimentua
1998.eko apirilean hartutako muskuiluak,
laborategiko baldintzei moldatu eta gero, 80 µg
Cd/l-ko kontzentrazioa (Cd kloruro moduan
gehitua) zuen itsas-uretan mantendu ziren 21
egunez. Aldi berean, Cd gehitu gabeko kontrol
serie bat ere burutu zen. Azterketa histologiko,
autometalografia eta mikoanalisirako laginak 1., 7.
eta 21. egunetan jaso ziren; absortzio atomikozko
espektrofotometriarako laginak, ordea, bakarrik 18.
egunean.

Bigarren esperimentua
1999.eko martxoan hartutako muskuiluak,
laborategiko baldintzei moldatu eta gero, 10 µg
Cd/l-ko kontzentrazioa (Cd kloruro moduan gehitua)
zuen itsas-uretan mantendu ziren 7 egunez. Aldi
berean, Cd gehitu gabeko kontrol serie bat ere
burutu zen. Azterketa histologikoetarako laginak 1.
eta 7. egunetan jaso ziren.
Hirugarren esperimentua
1999.eko martxoan hartutako muskuiluak
laborategiko baldintzei moldatu eta gero, 10 µg
Cd/l, 10 µg Cu/l zein 10 µg Zn/l-ko kontzentrazioa
(Cd eta Zn kloruro, Cu sulfato moduan gehituak
izanik) zuen itsas-uretan 7 egunez tratatu ziren
(esposizio-taldeak), azkenik 14 egunez ur garbitan
mantendu zirelarik (arazketa-taldeak). Aldi berean,
metalik gehitu gabeko kontrol serieak ere burutu
ziren. Azterketa histologikoak eta autometalografia
egiteko laginak 1. orduan, eta 1. eta 7. egunetan
hartu ziren, baita 14 eguneko arazketa-aldiaren
ostean ere (21. eguna). Absortzio atomikozko
espektrofotometriaren bidez metal-maila tisularrak
neurtzeko laginak 1. orduan, eta 1., 7. eta 21.
egunetan hartu ziren.
Laugarren esperimentua
1998.eko apirilean hartutako muskuiluak,
laborategiko baldintzei moldatu eta gero, 21 egunez
tratamendu desberdinak aplikatu zitzaien: (a) 500
µg Benzo(a)pireno/l-ko kontzentrazioa zuen itsas-
uretan (BaP talde esperimentala); (b) 80 µg Cd/l
(kloruro moduan) eta 500 µg BaP/l batera zituen
itsas-uretan (Cd + BaP talde esperimentala); eta (c)
uretan disolbagarria izateko, BaP-a dimetil
sulfoxidotan (DMSO) disolbatu zenez, 500 µg
DMSO/l-ko kontzentrazioa zuen itsas-uretan
mantendu ziren (DMSO kontrola). Aldi berean, Cd-
rik, BaP-rik zein DMSO-rik gehitu gabeko kontrol
serie bat ere burutu zen (Itsas-ur kontrola).
Azterketa histologikoak eta autometalografia
egiteko laginak esperimentuaren 1., 7. eta 21.
egunetan jaso ziren. Absortzio atomikozko
espektrofotometriaren bidez metal-maila tisularrak
neurtzeko laginak 18. egunean hartu ziren.
Bosgarren esperimentua
Meñakozen (43º 24´I, 2º 93´M) 1989.eko
martxoan hartutako muskuiluak laborategiko
baldintzei moldatu eta gero, 10 µg Cd/l eta 80 µg
Cd/l-ko kontzentrazioa (Cd kloruro moduan) zuen
itsas-uretan 41 egunez mantendu ziren. Aldi
berean, Cd-rik gehitu gabeko kontrol serie bat ere
burutu zen. Azterketa histologikoak eta
autometalografia egiteko laginak, 1., 7., 21., 27. eta
41, egunetan hartu ziren.
Analisi kimikoak
Disekzionatu eta gero, lehenengo, hirugarren
eta laugarren esperimentuko liseri-guruinak ur
distilatuan garbitu ziren, eta 120ºC-tara 48 orduz
labean berotu ziren, guztiz lehortu arte. Lehortutako
materiala almerizaren laguntzaz txikitu zen, eta
azido nitriko kontzentratuan liseritu zen. Ondoren,
azido nitriko kontzentratua plaka beroan lurrundu
ostean, liseritutako materiala ur distilatuan
diluitutako 0.1 M azido nitrikoan berresekitu zen.
Laginak iragazi ondoren, absortzio atomikozko
Emaitzak eta eztabaida
84

espektrofotometroan (PerkinElmer 2280, Wellesley,
Massachusetts, Amerikako Estatu Batuak) neurtu
ziren. Lehenengo eta laugarren esperimentuetan
Cd kontzentrazioak, µg Cd/g ehun pisu lehor
modura kalkulatu ziren. Hirugarren esperimentuan,
ordea, metal/maskor-pisua indizeak kalkulatu ziren
(Soto et al., 1995). Erabilitako estandarrak Merck
(Merck & Co., Inc, Whitehouse Station, New Jersey,
Amerikako Estatu Batuak) merkatal-etxeak
zertifikatutakoak izan ziren.
Gertakuntza histologikoa
Liseri-guruinaren zati txikiak (10 talde
esperimentaleko) Carnoy fixatzailean fixatu ziren
4ºC-tan ordu batez (Martoja & Martoja-Pierson,
1970). Laginak goranzko graduazioko etanoletan
deshidratatu ziren, metilo bentzoatoan aklaratu
ziren eta bentzenozko bi bainutan murgildu eta
gero, parafinan (60ºC) inkluditu ziren. Ebaki
histologikoak (7 µm) Leitz 1512 eta American
Optical mikrotomoetan lortu ziren. Liseri-guruinaren
beste zati txiki batzuk, %2.5 glutaraldehidoa zuen
kakodilato indargetzailean (0.1 M, pH = 7.2) fixatu
ziren 4ºC-tan 2 orduz. Gerora, indargetzailean
garbitu, goranzko graduazioko etanoletan
deshidratatu eta, azkenik, Epon 812 erretxinan
inkluditu ziren. Ebaki semifinak (1-3 µm) Leica
Ultracut (UCT) ultramikrotomoan lortu ziren.
Autometalografia (argi-mikroskopia)
Parafinazko ebakiak xilolezko bainuetan
desparafinatu ostean, beheranzko graduazioko
etanoletan hidratatu ziren. Bai parafinazko zein
erretxinazko ebaki semifinak labean (40ºC)
mantendu ziren guztiz lehortu arte.
Ondoren, laginak argazki-emultsio geruza fin
eta uniformez estali ziren, gela ilunean (Ilford
Nuclear Emulsion L4, Ilford, Mobberley,
Ingalaterra). Gerora, laginak ilunpean mantendu
ziren lehortu bitartean (30 minutu), eta
errebelatzaile komertziala erabiliz errebelatu ziren.
Parafinazko laginen kasuan, Ultrafin Tetenal SF
(1:5 diluzioa ur distilatuan; (Tetenal AG & Co,
Norderstedt, Alemania) errebelatzailea 15 minutuz
aplikatu zen. Erretxinazko laginen kasuan, ordea,
Kodak D-19 (Kodak, Paris, Frantzia) (%15 ur
distilatuan) errebelatzailea, 30-45 minutuz aplikatu
zen. Ondoren, errebelatzailearen erreakzioa %1
azido azetikozko disoluzioan minutu batez
murgilduz geldiarazi zen. Azkenik, Agefix (Agfa
Agefix, Agfa-Gevaert N.V., Mortsel, Belgika, 1:9
uretan) fixatzaileaz 10 minutuz fixatu ziren. Laginak
Kaiser gelatina glizerinatuan (Merck & Co., Inc,
Whitehouse Station, New Jersey, Amerikako Estatu
Batuak) montatu ziren. Erretxinan inkluditutako
ebakiak toluidina urdinez kontrastatu ziren. Metal-
ioien presentzia, zilarrezko hauspeakin beltzen
agerpenak adierazten du (Danscher, 1984; Soto et
al., 1998b).
Autometalografia (mikroskopio elektronikoa),
EPMA eta X izpien mapaketa
Beirazko portetan itsatsitako erretxinan
inkluditutako 3 µm-ko ebaki semifinetan
autometalografia aplikatu ondoren, Epon 812
erretxinan inkluditu ziren berriro ere ebaki ultrafinak
egiteko. Mikroskopioan behaketa ultrastruktural
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
85

morfologikoak egiteko, ebaki ultrafinak kuprezko
euskarrietan bildu ziren, eta uranilo azetatoz eta
berun zitratoz kontrastatu ziren.
Bestalde, mikroanalisia egiteko, ebaki semifinak
aldez aurretik karbono/formvar-ez estalitako
aluminiozko euskarrietan (200 mesh) bildu ziren,
eta berriro ere karbonoz estali ziren. Mikroanalisia,
energia dispertsibozko espektrometroa lotuta
zeukan JEOL 1210 STEM mikroskopioan burutu
zen (kontaketa-denbora = 100 segundu). Espektru-
analisia egiteko Link ISIS 1.04A softwarea erabili
zen (Morgan & Morgan, 1998). X izpien mapaketa
sistema berean burutu zen, Ag (Lα1 -2.984 keV-,
Lß1 -3.151 keV-), Cd (Lß1 -3.316 keV-) eta S (Kα -
2.308 keV-) elementuen seinaleak batera
lokalizatzeko.
BSDen kuantifikazioa argi-mikroskopioan
irudi-analisiaren bitartez
Liseri-zelulen lisosometan zeuden BSDen
hedapena irudi-analisia erabiliz kuantifikatu zen
(Soto & Marigómez, 1997b), emaitzak BSDen
dentsitate bolumetriko gisa adieraziz (VvBSD). Argi-
mikroskopiazko irudiak (x100 handipenean) B&W-
CCD TV kamera, HR-TV kolorezko pantaila, Leitz
mikroskopioa, PC ordenagailua, PIP 1024 (Matrox
Electronic Systems Ltd., Dorval, Kanada) sistema
eta Sevisan S.A. (Bilbo, Espainia) entrepresak
Emaitzak eta eztabaida
86
1. Irudia: Mytilus galloprovincialis muskuiluen liseri-epitelioan metalen lokalizazio autometalografikoa Cd pean 21 egunezmantendu eta gero. (A) Parafinan inkluditutako materiala. Eskala-marra: 10 µm. (B) Erretxinan inkluditutako liseri-guruinaren ebakisemifina, toluidina urdinez kontrastatua. Gezi-buruek liseri-zelulen lisosometan dauden BSDak erakutsi dituzte, zelula basofilikoek(izarrak), ordea, ez dute BSDrik erakusten. Eskala-marra: 10 µm. (C) Erretxinan inkluditutako eta kontrastatutako ebaki ultrafina.Zelula basofilikoen zitoplasman (BC) ez da BSDrik ikusten, bai ordea, heterolisosometan (gezi-buruak). (N = nukleoa; HL =heterolisosoma; L = lumena). Eskala-marra: 1 µm.

garatutako softwarea erabiliz eskuratu ziren.
Ondoren, eskuz eta softwarearen laguntzarekin
liseri-zelulen lisosometako BSD eta zitoplasma
bereizteko segmentazio-prozedura aplikatu zen.
Neurketak muskuilu bakoitzean 5 eremutan burutu
ziren, eta talde esperimental bakoitzean 10
muskuilu erabili ziren; tratamendu bakoitzeko
guztira 50 neurketa eginez. Kalkuluak, Lowe et al.-
ek (1981) proposatutako formulak erabiliz egin
ziren. Hau da, BSD partikulen tamaina ebakiaren
lodiera baino txikiagoa zela kontuan hartuz, Holmes
efektua zuzentzeko faktorea aplikatu zen.
Zelula basofilikoen analisi estereologikoak
Parafinan inkluditutako ebakiak (7 µm) Masson-
Goldner trikromikoz (Martoja & Martoja-Pierson,
1970) tindatu ziren, zelula basofilikoak argiro
bereiztu ahal izateko. Ondoren, zelula basofilikoen
dentsitate bolumetrikoa (VvBAS) estereologiaren
bidez kalkulatu zen. Horretarako, Nikon Optiphot
mikroskopioari erantsitako marrazketa-tutua erabili
zen (bukaerako handipena = x670). Multipurpose
test system M-168 Weibel gratikula erabiliz, zelula
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
87
2. Irudia: Muskuilu kontroletan (zutabe zuria) zein 80 µg Cd /lkontzentrazio pean mantendutako (zutabe grisa) muskuiluetankalkulatutako BSDen eta zelula basofilikoen dentsitatebolumetrikoak. (A) Irudi-analisi bitartez neurtutako BSDendentsitate bolumetrikoa. (B) Liseri-guruineko zelulabasofilikoen dentsitate bolumetrikoa. Segmentu bertikalekdesbidazio estandarra adierazten dute. Goialdeko asteriskoektaldeen arteko desberdintasun esangarriak adierazten dituzte(p<0.05).
3. Irudia: Zelula basofilikoen identifikazioa (asterisko zuriak) Mytilus galloprovincialis muskuiluaren liseri-albeoloetan.( A) Kontrola(B) 7 egunez 80 µgCd/l pean mantendutako muskuilua eta (C) 21 egunez 80µg/lCd pean mantendutako muskuilua. Eskala marra:25 µm (A eta B); 10 µm (C).

basofiliko eta liseri-zelulen gainean ezarritako
puntuak zenbatu ziren, VvBAS Delesse
printzipioaren arabera kalkulatuz (Weibel, 1979).
Neurketak elkarrengandik urruti (gutxienez 45 µm)
zeuden 2 ebakietan egin ziren, ebaki bakoitzean
sigi-saga eginez azarez aukeratutako 5 eremutan,
hain zuzen. Guztira, talde bakoitzean 100
zenbaketa egin ziren.
Estatistika
Emaitzak estatistikoki aztertzeko SPSS/PC+
(SPSS Inc., Microsoft Co.) pakete estatistikoa
erabili zen. Konfidantza-tarteak Student t testa
erabiliz kalkulatu ziren, eta taldeen arteko
desberdintasun estatistiko esangarriak p<0.05
mailan ezarri ziren (Sokal & Rohlf, 1979). Aldagai
esperimentalen esangarritasuna bariantzaren
analisia (ANOVA) erabiliz estimatu zen.
EMAITZAK
Lehenengo esperimentua
Analisi kimikoek muskuiluen liseri-guruinean
Cd-a metatu zela erakutsi zuten. Muskuilu kontrolen
Cd kontzentrazio-maila detekzio-atalasetik behera
mantendu zen bitartean, Cd pean egondako
muskuiluen kontzentrazio tisularra 1131.36 µg Cd/g
ehun-pisu lehor mailara heldu zen.
Mikroskopioan egindako behaketek erakutsi
zutenez, Cd pean egondako muskuiluetan BSDen
zelulabarneko banaketa ez zen uniformea izan.
BSD espezifikoki liseri-zeluletako lisosometan
lokalizatu ziren, zelula basofilikoak BSDrik gabe
agertu ziren bitartean (1. Ird.). Emaitza horiek, 21
egunez Cd pean egondako muskuiluetan,
erretxinan inkluditutako ebaki semifin zein
ultrafinetan konfirmatu ziren. Bertan, BSD
heterolisosometan eta, batez ere, lisosomen
mintzari lotuta, eta inolaz ere zelula basofilikoetan
lokalizatu ziren (1. Ird.). Horretaz gain, liseri-
albeoloen argirantz askatutako hondakinetan ere
BSD behatu ziren maiz, hondakin horiek,
seguruenik, jatorri lisosomikoa zutelarik.
Bestalde, VvBSD balioak Cd pean egondako
muskuiluetan kontroletan baino esangarriki
altuagoak suertatu ziren, batez ere 1. egunean.
Gerora, 7. eta 21. egunen artean VvBSD balioen
beherakada behatu zen, oraindik ere, Cd-z
tratatutako muskuiluen balioak kontrolenak baino
esangarriki altuagoak izanik (2A Ird.).
Halaber, VvBAS balioei dagokienez, muskuilu
kontroletan ez zen aldaketarik erregistratu
esperimentuak iraun bitartean, Cd pean
Emaitzak eta eztabaida
88
4. Irudia: Cd pean 21 egunez mantendutako muskuilu batenliseri-zelulen lisosometako autometalografiazko zilarrezkohauspeakin beltz baten EDS espektrua, EPMA egin eta gero.
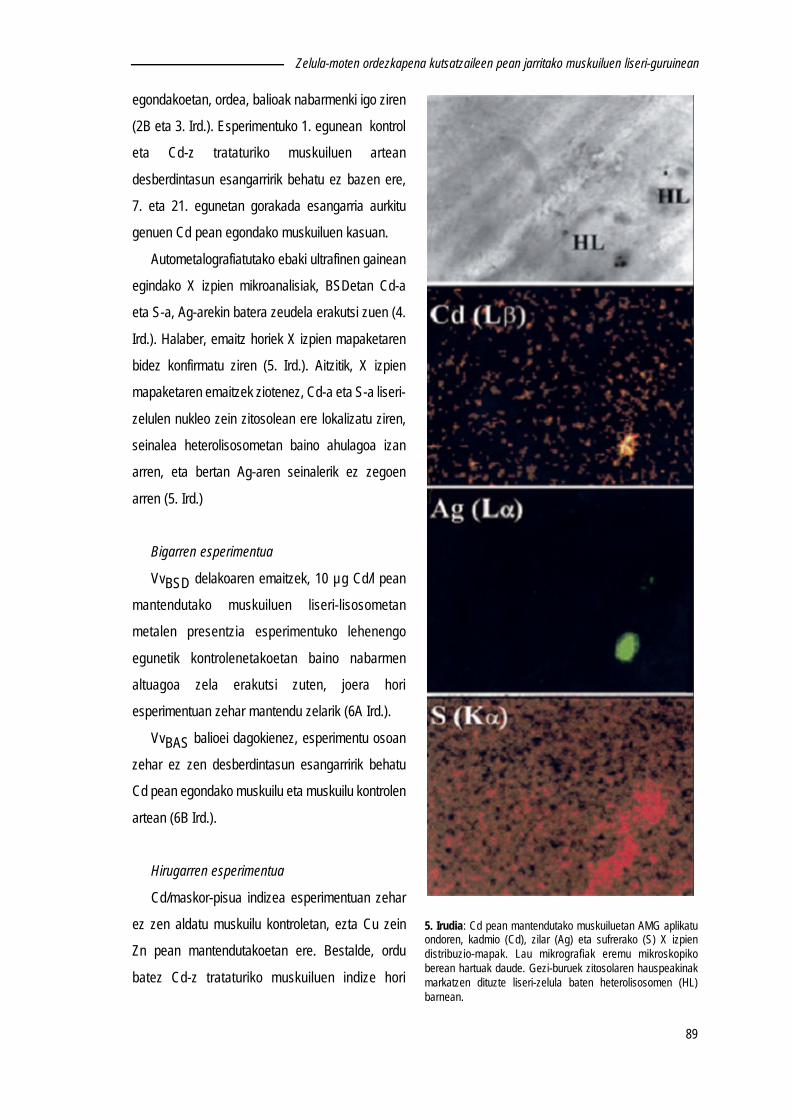
egondakoetan, ordea, balioak nabarmenki igo ziren
(2B eta 3. Ird.). Esperimentuko 1. egunean kontrol
eta Cd-z trataturiko muskuiluen artean
desberdintasun esangarririk behatu ez bazen ere,
7. eta 21. egunetan gorakada esangarria aurkitu
genuen Cd pean egondako muskuiluen kasuan.
Autometalografiatutako ebaki ultrafinen gainean
egindako X izpien mikroanalisiak, BSDetan Cd-a
eta S-a, Ag-arekin batera zeudela erakutsi zuen (4.
Ird.). Halaber, emaitz horiek X izpien mapaketaren
bidez konfirmatu ziren (5. Ird.). Aitzitik, X izpien
mapaketaren emaitzek ziotenez, Cd-a eta S-a liseri-
zelulen nukleo zein zitosolean ere lokalizatu ziren,
seinalea heterolisosometan baino ahulagoa izan
arren, eta bertan Ag-aren seinalerik ez zegoen
arren (5. Ird.)
Bigarren esperimentua
VvBSD delakoaren emaitzek, 10 µg Cd/l pean
mantendutako muskuiluen liseri-lisosometan
metalen presentzia esperimentuko lehenengo
egunetik kontrolenetakoetan baino nabarmen
altuagoa zela erakutsi zuten, joera hori
esperimentuan zehar mantendu zelarik (6A Ird.).
VvBAS balioei dagokienez, esperimentu osoan
zehar ez zen desberdintasun esangarririk behatu
Cd pean egondako muskuilu eta muskuilu kontrolen
artean (6B Ird.).
Hirugarren esperimentua
Cd/maskor-pisua indizea esperimentuan zehar
ez zen aldatu muskuilu kontroletan, ezta Cu zein
Zn pean mantendutakoetan ere. Bestalde, ordu
batez Cd-z trataturiko muskuiluen indize hori
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
89
5. Irudia: Cd pean mantendutako muskuiluetan AMG aplikatuondoren, kadmio (Cd), zilar (Ag) eta sufrerako (S) X izpiendistribuzio-mapak. Lau mikrografiak eremu mikroskopikoberean hartuak daude. Gezi-buruek zitosolaren hauspeakinakmarkatzen dituzte liseri-zelula baten heterolisosomen (HL)barnean.

kontrolen berdina izan arren, 1 eta 7 egun ondoren
kontrolarena baino esangarriki altuagoa suertatu
zen. Arazketa-aldiak, bere aldetik, ez zuen
Cd/maskor-pisua indizearen inolako beherakadarik
ekarri; haatik, balioak 7 eguneko tratamendu ostean
lortutakoak baino are altuagoak suertatu ziren (1.
Taula).
Cu/maskor-pisua indizea esperimentuan zehar
ez zen aldatu muskuilu kontroletan, ezta Cd zein
Zn pean mantendutakoetan ere. Bestalde, ordu
batez Cu-z trataturiko muskuiluen indize hori
kontrolen berdina izan arren, 1 eta 7 egun ondoren
kontrolarena baino esangarriki altuagoa suertatu
zen. Cu pean egondako muskuiluen hilkortasuna
handia gertatu zenez, 21. egunean ez genuen izan
analisi kimikoak egiteko behar bezain beste laginik,
eta, beraz, ezin dugu arazketa-aldiaren Cu/maskor-
pisua indizearekiko eraginari buruzko daturik eman
(1. Taula).
Zn/maskor-pisua indizea esperimentuan zehar
ez zen aldatu muskuilu kontroletan, ezta Cd zein
Cu pean mantendutakoetan ere. Bestalde, ordu
batez Zn-az trataturiko muskuiluen indize hori
kontrolen berdina izan arren, 1 eta 7 egun ondoren
kontrolarena baino esangarriki altuagoa suertatu
zen, aurreneko bi kasuen legez. Arazketa-aldiak,
bere aldetik, ez zuen Zn/maskor-pisua indizearen
inolako beherakadarik ekarri; haatik, balioak 7
eguneko tratamendu ostean lortutakoak baino are
altuagoak suertatu ziren (1. Taula).
Autometalografiaz errebelatutako BSD,
aurreneko esperimentuen emaitzekin bat, hiru
metal pean mantendutako muskuiluetan liseri-
zelulen lisosometan lokalizatu ziren, batez ere.
BSDen kuantifikazioa burutu ondoren, Cd, Cu
eta Zn pean egondako muskuiluetan lortutako
VvBSD balioak kontroletan baino altuagoak izan
ziren jadanik lehenengo ordutik, eta tendentzia hori
7 egunez mantendu zen (7A Ird.). Cd eta Cu pean
egondako muskuiluen VvBSD balio altuenak 7.
egunean erregistratu ziren; Zn-aren kasuan, ordea,
baliorik altuena 1. egunean gertatu zen. Izan ere,
balio altu hori esperimentu osoan zehar
erregistratutako baliorik altuena izan zen (7A Ird.).
Arazketa-aldiaren ostean VvBSD balioak
Emaitzak eta eztabaida
90
6. Irudia: Muskuilu kontrol (zutabe zuria) zein 10 µgCd/l peanmantendutako (zutabe grisa) muskuiluak. (A) Irudi-analisibitartez neurtutako BSDen dentsitate bolumetrikoa. (B) Liseri-guruineko zelula basofilikoen dentsitate bolumetrikoa.Segmentu bertikalek desbidazio estandarra adierazi dute.Goialdeko asteriskoek taldeen arteko desberdintasunesangarriak adierazi dute (p<0.05).

esangarriki jaitsi ziren hiru metalen kasuan, berriro
ere kontrolen mailara helduz (7A Ird.).
Liseri-epitelioan zelula basofilikoen proportzio
erlatiboa kuantifikatzeko erabilitako VvBAS balioak
ez ziren esperimentuan zehar aldatu muskuilu
kontroletan, ezta Cd eta Cu pean mantendutako
muskuiluetan ere (7B Ird.). Bestalde, 7 egunez Zn-
az trataturiko muskuiluen liseri-epitelioan VvBASbalioetan gorakada esangarria behatu zen, gerora,
arazketa-aldiaren ostean kontrolen mailara jaitsi
zena (7B Ird.).
Laugarren esperimentua
Analisi kimikoek, kadmioa BaP-z nahastuta
egon arren (Cd + BaP taldea), liseri-guruinean Cd-
a metatu zela erakutsi zuten. Muskuilu kontroletan
(DMSO-z tratatutakoak) Cd-aren kontzentrazio
balioak detekzio-atalasetik behera agertu ziren
bitartean, Cd + BaP pean mantendutakoen liseri-
guruinean 1079.25 µg Cd/g ehun-pisu lehor aurkitu
ziren.
Aurreneko esperimentuetan bezala, BSD
espezifikoki liseri-zeluletako lisosometan lokalizatu
ziren bitartean, zelula basofilikoetan ez zen seinale
esangarririk detektatu. Esperimentuko 1. egunetik
21. egunera, BaP eta Cd + BaP tratamenduetako
muskuiluetan neurtutako VvBSD balioak DMSO
kontroletan neurtutakoak baino esangarriki
altuagoak izan ziren, bereziki 1. egunean, gorakada
hori batik bat Cd + BaP tratamenduan nabarmena
gertatu zelarik (8A Ird.).
VvBAS balioei dagokienez, DMSO-z trataturiko
muskuilu kontroletan saiakuntza-denboran zehar
nolabaiteko igoera behatu zen. Hala ere, VvBAS
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
91
Lagina Cd indizea Cu indizea Zn indizea Kontrola Ordu 1 Egun 1 7 Egun Arazte-aldia
0.00126344 0.00214682 0.00100389 0.00608193
0.07669171 0.01559907 0.00763639 0.02290661
0.12930922 0.15481972 0.11437475 0.12991251
Cd-esposizioa Ordu 1 Egun 1 7 Egun Arazte-aldia
0.00313986 0.0098965* 0.0256267* 0.0370562*
0.01043518 0.01598671 0.00776714 0.02040212
0.1238165 0.18353205 0.12913258 0.12461644
Cu-esposizioa Ordu 1 Egun 1 7 Egun Arazte-aldia
0.00398981 0.0035511 0.00560253
###
0.0146126 0.0342118* 0.0681148*
###
0.1450487 0.19813616 0.15587894
###
Zn-esposizioa Ordu 1 Egun 1 7 Egun Arazte-aldia
0.00132747 0.00281531 0.00075713 0.0075807
0.0085813 0.01860038 0.00892704 0.02057063
0.13776012 0.26060344* 0.19286193* 0.32503914*
1. Taula: AAS bidez neurtutako liseri-guruineko Cd, Cu eta Zn/maskor-pisua indizea, metal pean ordu batez eta 1, 7 eta 21 egunezmantendutako muskuiluetan eta muskuilu kontroletan. Asteriskoek kontrolekiko desberdintasun esangarriak adierazten dituztep<0.05 mailan. ### muskuiluak hil zirenez ez zen gelditu animalia nahikorik arazketa esperimenturako.

balioen emendioa askoz nabarmenagoa izan zen
BaP eta Cd + BaP tratamenduetako muskuiluetan
(8B Ird.).
Bosgarren esperimentua
Esperimentuko 1. egunetik, VvBSD balioak 80
µg Cd/l kontzentrazio pean egondako muskuiluetan
kontroletan baino nabarmenki altuagoa izan ziren.
10 µg Cd/l kontzentrazio pean egondako
muskuiluetan, berriz, astebete behar izan zen
VvBSD balioak kontrolenetatik esangarriki
desberdinak izatera iristeko. Hala ere, Cd
kontzentrazio handiaz lau astez trataturiko
muskuiluetan, VvBSD balioen behin behineko
jaitsiera puntuala suertatu zen, 6 aste ostean berriro
ere VvBSD balio altuak neurtu zirelarik (9A Ird.).
VvBAS balioei dagokienez, muskuilu kontrolek
balio baxuak erakutsi zituzten esperimentuak iraun
bitartean. Bestalde, Cd kontzentrazioaren
menpekoa ez zen igoera esangarria behatu zen Cd
pean mantendutako muskuiluen VvBAS balioetan,
1. astetik aurrera behintzat (9B Ird.).
EZTABAIDA
Kadmioak CTR eragin dezake
Kutsatzaileek zein bestelako estres-iturriek
muskuiluen liseri-guruinean aldaketak sortarazten
dituztela ezaguna da; euren artean CTR izenekoa
(Thompson et al., 1974; Rasmussen et al., 1983;
Cajaraville et al., 1990a; Marigómez et al., 1990b;
Syasina et al., 1997; Marigómez et al., 1998).
Ikerlan honetan, Cd-ak muskuiluen liseri-guruinean
CTR prozesua bultza dezakeela baieztatu dugu.
Emaitzak eta eztabaida
92
7. Irudia: Muskuilu kontroletan zein 10 µg/lCd, Cu eta Zn pean jarritakoetankalkulatutako parametro estereologikoak. (A)Irudi-analisi bitartez neurtutako BSDendentsitate bolumetrikoa. (B) Liseri-guruinekozelula basofilikoen dentsitate bolumetrikoa.Segmentu bertikalek desbidazio estandarraadierazi dute. Goialdeko matrizetakoasteriskoek taldeen arteko desberdintasunesangarriak adierazi dituzte (p<0.05).

Laborategiko kontrol esperimentaletan VvBASbalioak 0.05-0.1 µm3/µm3 tartekoak diren
bitartean, Cd pean egondakoetan 0.15-0.22
µm3/µm3-koak izatera irits daitezke.
Hala ere, eraginkortasun horren atalase kritikoa,
desberdina da, muskuiluen jatorri eta baldintza
esperimentalen arabera. Esaterako, Plentziako
muskuiluetan, 80 µg Cd/l kontzentrazio pean 7
egunez egotea VvBAS-aren emendio esangarria
sortzeko beharrezkoa den bitartean, aldez aurretik
zelaian Zn-az kutsatutako Meñakozeko
muskuiluetan laborategian 10 µg Cd/l kontzentrazio
pean 7 egunez egotea nahikoa da VvBASemendatzeko. Aitzitik, 10 µg Cd/l kontzentrazio
pean 7 egunez egoteak VvBAS balioaren igoerarik
ez zien ekarri Plentziako muskuiluei.
Zelula-moten ordezkapen honen arrazoia
momentuz ezagutzen ez bada ere, autore batzuen
arabera, kutsaduraren aurkako babes-mekanismoa
izango litzateke. Alde batetik, zelula basofilikoen
emendioa, detoxifikazio-prozesuak eragindako
liseri-zelulen andeakuntzari aurre egiteko
erantzuna izango litzatekeela proposatu da, zelula
basofilikoak liseri-epitelioaren berriztapenaz
arduradunak direla suposatuz (Mix & Sparks, 1971;
Thompson et al., 1974). Bestalde, kutsatzaile
organikoen kasuan batik bat, zelula basofilikoek
aktiboki detoxifikazioan parte hartzen dutenez,
euren presentzia areagotzea, detoxifikazio-
prozesuaren ondorioa izan liteke (Widdows, 1984;
Cajaraville et al., 1990a; Triebskorn & Künast,
1990). Halaber, Widdows et al.-ek (1984),
kutsatzaileen aurrean zelula basofilikoen
kopuruaren gorakada, arazketa-aldiko eskakizun
energetiko gehigarriari erantzuteko beharrezkoak
diren entzimen jariapen areagotuarekin erlaziona
litekeela iradoki zuten. Azkenik, zelula basofilikoen
emendioa itxurazkoa baino ez dela, eta kutsatzaile
pean egoteak liseri-zelulen galera ekarri izanaren
ondorioz zelula basofilikoak nagusitzen direla ere
proposatu egin da (Marigómez et al., 1990b).
Izatez, hiru alternatiba horien artean VvBAS-en
emendioa zeinek azal dezaken argitzea tesi honen
helburuetako bat da, geroko kapituluetan jorratuko
den bezala.
Metalen metaketa zelula-espezifikoa
CTR eragiteak, liseri-epitelioko metaketa-
gaitasunean aldaketa garrantzitsuak sor ditzake.
Izan ere, zelula-mota desberdinek, kutsatzaileak
metatzeko gaitasun ezberdina eduki dezakete
(Marigómez et al., 2002). Moluskuetan, BSD liseri-
zelulen lisosometan agertu dira batez ere, eta
beraz, zelula basofilikoen proportzio erlatiboaren
igoerak, metalak metatzeko konpartimentua
murrizten duenez, metatzeko gaitasuna txikitzea
ekar lezake (Marigómez et al., 1998).
Ikerlan honetan, ere Cd pean egondako
muskuiluetan, BSD espezifikoki liseri-zeluletako
lisosometan agertu dira, argi-mikroskopian
oinarritutako beste moluskuen espezietan zenbait
ikerketek diotenarekin bat datorrena (Hemelraad &
Herwig, 1988; Holwerda, 1991; Giamberini et al.,
1996; Marigómez et al., 1996; 2002; Soto et al.,
1996a, 1996b, Soto & Marigómez, 1997b).
Lisosomen barruan metalak bahitzeak, metal horien
toxizitatea jaisten lagunduko luke, behin-behinekoz
behintzat. Gerora, metaletan joriak diren lisosomak
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
93

liseri-dibertikuluen argira askatu eta ondoren
gorotzekin batera kanporatuko lirateke (Ireland &
Marigómez, 1992; Viarengo & Nott, 1993;
Cajaraville et al., 1995; Langston et al., 1998;
Marigómez et al., 2002).
Hala ere, argi-mikroskopioaren bidez lortutako
emaitz horiek ez dira maiz mikroskopia
elektronikoaren bidez baieztatu, dakigunez. Izatez,
AMGren aplikazioak maila ultrastrukturalean urriak
dira (Domouhtsidou & Dimitriadis, 2000; Dimitriadis
& Papadaki, 2004).
Ikerlan honetan, mikroskopio elektronikoan
burututako AMGk, BSD espezifikoki liseri-zelulen
lisosometan lokalizatzen direla egiaztatu du.
Zehazki, BSD heterolisosometan, batez ere
mintzari lotuta, eta inolaz ere zelula basofilikoetan
lokalizatu dira. Emaitza horiekin ados, X izpien
mikroanalisia aplikatuz, Littorina littorea
magurioaren liseri-zeluletako lisosomek Cd-a
metatzeko konpartimentu nagusia osatzen dutela
frogatu zuten Marigómez et al.-ek (1991). Merkurio
dosi oso altuz tratatutako muskuiluetan, ordea,
liseri-zeluletako lisosometan egoteaz gain zelula
basofilikoetan ere aurkitu dira BSD (Domouhtsidou
& Dimitriadis, 2000). Beraz, banaketa intrazelularra
dosiaren menpekoa (metalaren izaeraren
menpekoa den bezala) izatea ezin da baztertu.
Izatez, Recio et al.-ek (1988), Zn pean egondako
bareetan, metala liseri- zein kaltzio- eta iraizte-
zeluletan esposizio-denbora eta dosiaren arabera
ager daitekeela erakutsi zuten, metodo histokimiko
konbentzionalak erabiliz, ditizona tindaketa, alegia.
Bestalde, AMGk ez du zuzenean Cd-aren
presentzia adierazi behar, teknika hau metal-
espezifikoa ez delako; eta gainera, BSDen eraketa
metal-ioien Ag-arekiko duten irisgarritasunaren
menpean dagoelako (Danscher, 1981). Alde
batetik, Cd-rik gabeko tokitan BSDak eratzea
posiblea da. Esaterako, Cd eta beste metal astunen
pean egoteak, ehunetako metal esentzialen (Fe-a,
Cu-a adibidez) berbanaketa sortarazi lezake
(Mason & Simkiss, 1983; Mason et al., 1984;
Mason & Jenkins, 1995), eta kasu horretan BSDek
lekuzaldatutako metal horien agerpena eta ez Cd-
aren presentzia erakutsiko lukete.
Emaitzak eta eztabaida
94
8. Irudia: DMSO-kontrol taldeko (zutabe zuria) zein BaP(zutabe gris argia) eta Cd+BaP (zutabe gris iluna) peanmantendutako muskuiluetan kalkulatutako parametroestereologikoak. (A) Irudi-analisi bitartez neurtutako BSDendentsitate bolumetrikoa. (B) Liseri-guruineko zelulabasofilikoen dentsitate bolumetrikoa. Segmentu bertikalekdesbidazio estandarra adierazi dute. Goialdeko asteriskoektaldeen arteko desberdintasun esangarriak adierazi dituzte(p<0.05).
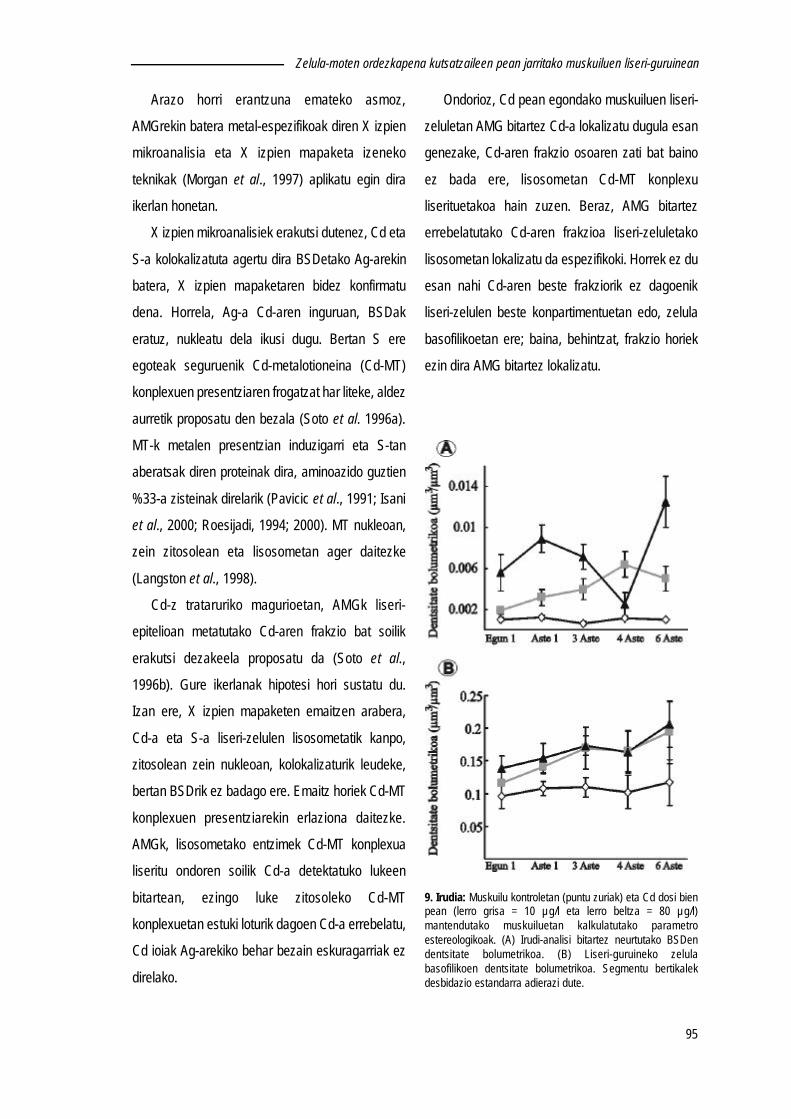
Arazo horri erantzuna emateko asmoz,
AMGrekin batera metal-espezifikoak diren X izpien
mikroanalisia eta X izpien mapaketa izeneko
teknikak (Morgan et al., 1997) aplikatu egin dira
ikerlan honetan.
X izpien mikroanalisiek erakutsi dutenez, Cd eta
S-a kolokalizatuta agertu dira BSDetako Ag-arekin
batera, X izpien mapaketaren bidez konfirmatu
dena. Horrela, Ag-a Cd-aren inguruan, BSDak
eratuz, nukleatu dela ikusi dugu. Bertan S ere
egoteak seguruenik Cd-metalotioneina (Cd-MT)
konplexuen presentziaren frogatzat har liteke, aldez
aurretik proposatu den bezala (Soto et al. 1996a).
MT-k metalen presentzian induzigarri eta S-tan
aberatsak diren proteinak dira, aminoazido guztien
%33-a zisteinak direlarik (Pavicic et al., 1991; Isani
et al., 2000; Roesijadi, 1994; 2000). MT nukleoan,
zein zitosolean eta lisosometan ager daitezke
(Langston et al., 1998).
Cd-z trataruriko magurioetan, AMGk liseri-
epitelioan metatutako Cd-aren frakzio bat soilik
erakutsi dezakeela proposatu da (Soto et al.,
1996b). Gure ikerlanak hipotesi hori sustatu du.
Izan ere, X izpien mapaketen emaitzen arabera,
Cd-a eta S-a liseri-zelulen lisosometatik kanpo,
zitosolean zein nukleoan, kolokalizaturik leudeke,
bertan BSDrik ez badago ere. Emaitz horiek Cd-MT
konplexuen presentziarekin erlaziona daitezke.
AMGk, lisosometako entzimek Cd-MT konplexua
liseritu ondoren soilik Cd-a detektatuko lukeen
bitartean, ezingo luke zitosoleko Cd-MT
konplexuetan estuki loturik dagoen Cd-a errebelatu,
Cd ioiak Ag-arekiko behar bezain eskuragarriak ez
direlako.
Ondorioz, Cd pean egondako muskuiluen liseri-
zeluletan AMG bitartez Cd-a lokalizatu dugula esan
genezake, Cd-aren frakzio osoaren zati bat baino
ez bada ere, lisosometan Cd-MT konplexu
liserituetakoa hain zuzen. Beraz, AMG bitartez
errebelatutako Cd-aren frakzioa liseri-zeluletako
lisosometan lokalizatu da espezifikoki. Horrek ez du
esan nahi Cd-aren beste frakziorik ez dagoenik
liseri-zelulen beste konpartimentuetan edo, zelula
basofilikoetan ere; baina, behintzat, frakzio horiek
ezin dira AMG bitartez lokalizatu.
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
95
9. Irudia: Muskuilu kontroletan (puntu zuriak) eta Cd dosi bienpean (lerro grisa = 10 µg/l eta lerro beltza = 80 µg/l)mantendutako muskuiluetan kalkulatutako parametroestereologikoak. (A) Irudi-analisi bitartez neurtutako BSDendentsitate bolumetrikoa. (B) Liseri-guruineko zelulabasofilikoen dentsitate bolumetrikoa. Segmentu bertikalekdesbidazio estandarra adierazi dute.

CTRk BSDen kuantifikazioaren gain duen
eragina
AMG bitartez errebelatutako lisosomen barneko
BSDak, irudi-analisiaren bitartez kuantifika
daitezke, horrela estimatutako VvBSD parametroak
ingurumeneko metal bioeskuragarrien mailak
adierazi litzakeelarik (Soto & Marigómez, 1997b).
BSD espezifikoki liseri-zeluletan eratzen direla
aintzat hartuz gero, metal kutsatzaileek beraiek
eragindako CTR moduko efektuek AMG bitartez
estimatutako metalen ingurumen-mailen gainean
eragina izan dezakete. Izan ere, Soto & Marigómez-
ek (1997b), arazo horretaz jabetuta, VvBAS eta
VvBSD batera aintzat hartzen zituen protokolo bat
proposatu zuten metalen esposizio-mailak
muskuiluen liseri-guruinean AMG aplikatuz
estimatzeko.
Ikerlan honetan, balizko eragin hori laborategiko
esperimentuetan oinarrituta aztertu dugu. Horrela,
Cd pean mantendutako muskuiluen kasuan, VvBASnabarmenki igotzerakoan, VvBSD balioek
beherantz egin edo, behintzat, ez zuten
tratamenduaren ondorioz gorantzago egin.
Esaterako, muskuiluak 80 µg/l-ko Cd
kontzentrazioaz tratatzean VvBSD balio altuenak 1.
egunean lortu ziren (1. esperimentua), eta denbora
aurrera joan ahala VvBSD balioak mantendu edo
jaitsi egin ziren, VvBAS balioen gorakadarekin
batera. Halaber, BaP eta Cd + BaP tratamenduen
ondorioz erantzun berbera ikusi dugu (4.
esperimentua), eta baita 10 µg/l-ko Zn
kontzentrazio pean 1 eta 7 egunez mantendutako
muskuiluen kasuan ere. Zn-aren kasuan, ordea,
CTR esperimentuko 7. egunean ageri zen. Bertan,
1. egunean Zn kontzentrazio tisularra eta VvBSDaltuak izan ziren bitartean, 7. egunean jaitsi egin
ziren, VvBAS gorakadarekin batera.
Gainera, muskuiluak CTR induzitzen ez duten
Cd eta Cu kontzentrazio baxuez tratatzerakoan
VvBSD balioen dinamikan ez da erregistratu
aldaketarik esperimentuan zehar (2. eta 3.
esperimentuak). Izan ere, 10 µg Cd/l
kontzentrazioz trataturiko bigarren esperimentuko
muskuiluetan, VvBSD esangarriki igo zen bitartean,
VvBAS ez zen aldatu.
Bestalde, Cd pean egon ostean 14 eguneko
arazketa-aldiak, Cd kontzentrazio tisularraren
igoera sortarazi zuen liseri-guruinean, aldi berean
VvBSD jaitsi zen arren. Itxuraz zentzu gutxiko
aldaketa horiek, liseri-epitelioaren zein ondoko
ehunen osaketa zelularraren aldaketekin erlaziona
litezke. Alde batetik, liseri-epitelioaz gain inguruko
ehun konektiboan eta hemozitoetan ere meta
daiteke kadmioa (Soto et al., 1996a), eta VvBSDbalioek liseri-lisosometan dauden BSDei soilik
dagozkie. Izan ere, VvBSD balioak kalkulatzeko,
beste zeluletan dauden BSD zein AMGk errebelatu
ez dituen metalak ez dira kontuan hartu, AAS
aplikatzean, berriz, metal frakzio guztiak batera
barneratzen dira. Gainera, liseri-lisosometan
metatutako Cd-a liseri-dibertikuluen argira
kanporatzen da, ondoren gorotzekin batera
eliminatua izateko (Marigómez et al., 2002).
Hondakin horiek BSDetan joriak dira, eta irudi-
analisi bitartez ez dira kuantifikatu; analisi
kimikoetan, ordea, hondakin horiek daramaten
metala ere neurtu den bitartean.
Emaitzak eta eztabaida
96

Ondorioz, metal-esposizioaren adierazleak
diren analisi kimiko zein AMG bidez lortutako
parametroen gain eragin esanguratsua du CTRk,
laborategiko esperimentuko metal pean egondako
muskuiluen kasuan behintzat, aldez aurretik zelai-
ikerketan oinarrituz proposatu zenarekin bat
datorrena (Soto & Marigómez, 1997b).
CTR eragin lezaketen kutsatzaileen maila
kritikoak
VvBSD zein beste biomarkatzaileen gainean
izan dezaken eragina kontuan hartuz, molusku
espezie desberdinetan CTR fenomenoa gerta dadin
beharrezkoak diren kutsatzaile desberdinen
kontzentrazio kritikoak ezagutzea komeni da.
Ikerlan honetan, 80 µg Cd/l, 10 µg Zn/l, 500 µg
DMSO/l eta 500 µg BaP/l-ko dosiek CTR eragin
dezaketela ondorioztatu dugu, beti ere denbora-
tarte desberdinetan kutsatzailearen arabera.
Halaber, 0.25 mg N-metil-N-nitro-N-nitroso-
guanidinaz 15 egunez zein fuel olioaren %6 WAFez
35 egunez tratatutako muskuiluetan eta 1.25 mg
Cd/l-z 17 egunez tratatutako magurioetan CTR
fenomenoa ere gertatu da (Rasmussen et al., 1983;
Cajaraville et al., 1990a; Marigómez et al., 1990b).
Behe-atalasearen bila, 3. esperimentuan
muskuiluak Cd, Cu eta Zn dosi baxuz tratatu ziren.
Bigarren esperimentua egin ondoren, 10 µg Cd/l-ko
kontzentrazioak 7 egunetan CTRrik eragin ez
zuena bagenekienez, kontzentrazio esperimental
hori aukeratu zen 3. esperimenturako. Bertan, 2.
esperimentuaren emaitzak konfirmatu ziren, eta
baita ere 10 µg Cu/l-ko kontzentrazioak ez zuela
CTRrik induzitzen. Zn-aren kasuan, ordea, 10 µg/l-
ko dosiak CTR jadanik 7. egunean eragin dezake.
Dosiaren menpeko erantzunak ere Cd pean
egondako magurioetan behatu dira. Bertan, 20
egunez 1.25 mg Cd/l kontzentrazioak liseri-epitelioa
nagusiki zelula basofilikoz osatuta egotea sortarazi
zuen bitartean, 500 µg Cd/l-ko kontzentrazioa ez
zen horren eraginkorra suertatu (Marigómez et al.,
1990b). Hain Cd kontzentrazio handiak magurioen
liseri-guruinean CTRrik ez eragiteak, espezien
arteko sentikortasuna oso desberdina izan
daitekeela utzi du agerian.
CTRren itzulgarritasuna
CTR fenomenoaren izaera eta beren
biomarkatzaileen gaineko eragina behar bezala
ulertzeko, prozesua itzulgarria denentz finkatzea
ezinbestekoa iruditu zitzaigun.
Arazketa-prozesuari dagokionez, Zn pean
mantendutako muskuiluetan, CTR prozesua
itzulgarria zela egiaztatu zen, behin estres-iturria
desagertutakoan, liseri-epitelioaren zelulen
proportzioa berriro ere bere hasierako balioetara
itzuli baitzen. CTR fenomenoaren itzulgarritasuna
ez da soilik muskuiluen kasuan ikusi; magurio eta
bareen kasuetan ere estres-egoera jasan eta gero,
zenbait egunez arazten utzitakoan, liseri-epitelioko
zelulen proportzioa berriro ere hasierako egoerara
itzul daitekeela frogatu da (4. eta 5. Kapituluak).
CTRaren gainean eragina izan dezaketen
faktoreak
CTR fenomenoa noiz eta nola ematen den
hobeto ulertzeko, bere gainean ingurumen-faktore
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
97

desberdinek nolako eragina duten ezagutzea
beharrezkoa zaigu. Faktore hauen artean,
muskuiluen adina, itsasaldi-erregimena, urteko
sasoia, kutsatzaileen arteko elkarrekintza eta aldez
aurretik kutsatzaile pean egon izana ditugu,
besteak beste.
Urteko sasoiari dagokionez, udan VvBASneguan baino altuagoa da. Sasoiari lotutako
aldaketa hori ziklo gonadalarekin erlazionatu da
(Méndez, 1993).
Kutsatzaileen arteko elkarrekintza ikertzeko
asmoz, laugarren esperimentuan Cd-arekin batera
muskuiluak BaP kutsatzaile organikoaren pean jarri
ziren. Analisi kimikoek diotenez, kutsatzaileen
nahasketa horrek ez zuen eraginik izan liseri-
guruinak Cd-a metatzeko duen gaitasunean.
Emaitza hori, beste ikerlan batzuekin bat dator. Izan
ere, Cd eta alkilbentzeno sulfonatua kutsatzaile
organikoaren artean ez da ere ez lehiaketa-
mekanismorik aurkitu (Da Ros et al., 1995). Hala
ere, BSDen emaitzek, BaP-k metalen mobilizazioa
eragin dezakela erakutsi digute. Hau da, BaP
taldeko muskuiluetan VvBSD balioak esangarriki
igo dira esperimentuko lehenengo egunetik.
Beharbada, BaP-aren eraginez, metal esentzialak
liseri-zelulen lisosometara mobilizatuak gerta
litezke.
Laugarren esperimentu honetan, Cd + BaP
nahasketak CTR eragiten du, baita BaPk eta
DMSOk (nahiz eta maila baxuagoan) ere. Cd + BaP
nahasketak emandako VvBAS balioak, Cd pean
mantendutako muskuiluen antzekoak izan dira,
beraz nahiz eta Cd-a eta BaP-a norberak bere
aldetik CTR eragin, bien nahasketak ez du erantzun
gehigarririk ekarri.
CTR fenomenoaren gainean eragina izan
dezakeen beste faktoreetako bat, aldez aurretik
estres-egoera batean egotea izan daiteke.
Bosgarren esperimentuan, Zn-ez kutsatuta zegoen
Meñakoz izeneko lekutik (Soto et al., 1995) hartu
ziren muskuiluak. 7 egunez laborategian ur garbitan
egon ondoren Plentziako muskuilu kontrolen
VvBSD eta VvBAS balio antzekoak erakutsi
zituzten. Izan ere, 3. esperimentuan CTR gutxienez
14 egunetan itzulgarria dela ikusi denez, 7
egunetan VvBAS balioak berreskuratzea ez zaigu
harrigarria gertatu. Gainera, kontrol gisa erabilitako
muskuiluetan, balioak beste esperimentuen
kontrolen maila baxuan mantendu ziren 5.
esperimentuak iraun bitartean. Cd pean jarritako
muskuiluetan, ordea, aurreneko esperimentuetan
ez bezala, 10 µg Cd/l-ko kontzentrazioak CTR
induzitu zuen. Muskuilu hauetan, aldez aurretik
estres-egoera batean egon izanak, beraien liseri-
guruineko VvBAS Plentziako muskuiluetan baino
errezago goratzea eragin zezakeen. Bestalde, bi
Cd kontzentrazioen desberdinen pean
mantendutako muskuiluetan VvBSD balioen
jeitsiera behatu zen, VvBAS balioen aldaketekin
sinkronian agertu ez ba ziren ere. Gainera, VvBSDbalioak Plentziako muskuiluetan (2. esperimentuko
datuak) baino denbora gehiago behar izan zuten
modu esangarrian igotzeko. Beraz, aldez-aurretik
estres-egoera batean egon izanak, dirudienez, CTR
erantzunaren nondik norakoak, zein metalen
detoxifikazio-mekanismoak eta -dinamika afekta
ditzake.
Emaitzak eta eztabaida
98

Ondorioak
Ikerlan honetan lortutako emaitzek dioskutenez,
muskuiluen liseri-guruinean CTR eragiteko gai da
kadmioa. Bestalde, Cd-a ehunetan detektatzeko
erabili den AMG bitartez errebelatutako BSDak,
liseri-zelulen lisosometan espezifikoki lokalizatu
dira. Hori dela eta, CTRk VvBSD balioen gainean
eragina dauka, metala metatzeko
konpartimentuaren txikitzearen ondorioz. Bestalde,
CTR gertatzeko beharrezkoa den kutsatzaile
desberdinen kontzentrazio-atalaseak zehaztu dira,
kutsatzaile eta espeziearen arabera desberdinak
izanik. Halaber, estres-iturria etetean CTR
itzulgarria izan daitekeela atzeman dugula
azpimarratzekoa da. Azkenik, CTR prozesuaren
gainean zenbait faktorek eragina eduki dezaketela
ikusi dugu; kutsatzaileak nahasketetan agertzea eta
muskuiluak aldez aurretik estres-egoeran egon
izana, besteak beste.
BIBLIOGRAFIA
Cajaraville MP, Díez G, Marigómez I, Angulo E (1990a)
Responses of basophilic cells of the digestive gland of
mussels to petroleum hydrocarbon exposure. Dis. Aquat.
Org. 9:221-228
Cajaraville MP, Marigómez I, Angulo E (1990b) Short-term toxic
effects of 1-naphthol on the digestive gland-gonad complex
of the marine prosobranch Littorina littorea (L.): a light
microscopic study. Arch. Environ. Contam. Toxicol. 19:17-
24
Cajaraville MP, Robledo Y, Etxeberria M, Marigómez I (1995)
Cellular biomarkers as useful tools in the biological monito-
ring of pollution: molluscan digestive lysosomes. :29-55 orr.
Non: Cell Biology in Environmental Toxicology. Cajaraville
MP (ed.). University of the Basque Country Press Service.
Bilbo.
Cajaraville MP, Bebianno MJ, Blasco J, Porte C, Sarasquete C,
Viarengo A (2000) The use of biomarkers to assess the
impact of pollution in coastal environments of the Iberian
Peninsula: a practical approach. Sci. Total Environ.
247:295-311.
Danscher G (1981) Histochemical demonstration of heavy
metals. A revised version of the sulphide silver method sui-
table for both light and electronmicroscopy. Histochemistry
71:1-16.
Danscher G (1984) Autometallography. A new technique for
light and electron microscopic visualization of metals in bio-
logical tissues (gold, silver, metal sulphides and metal sele-
nides). Histochemistry 81:331-335.
Da Ros L., Nasci C, Campesan G, Sartorello P, Stocco G,
Menetto A (1995) Effects of linear alkylbenzene sulphona-
te (LAS) and cadmium in the digestive gland of mussel,
Mytilus sp. Mar. Environ. Res. 39:321-324.
Da Ros L, Nasci C, Marigómez I, Soto M (2000) Biomarkers
and trace metals in digestive gland of indigenous and
transplanted mussels, Mytilus galloprovincialis, in Venice
lagoon, Italy. Mar. Environ. Res. 50:417-423.
Dimitriadis VK, Domouhtsidou GP, Cajaraville MP (2004)
Cytochemical and histochemical aspects of the digestive
gland cells of the mussel Mytilus galloprovincialis (L.) in
relation to function. J. Mol. Histol. 35:501-509.
Dimitriadis VK, Papadaki M (2004) Field application of autome-
tallography and X-ray microanalysis using the digestive
gland of the common mussel. Ecotoxicol. Environ. Saf. 59:
31-37.
Domouhtsidou GP, Dimitriadis VK (2000) Ultrastructural locali-
zation of heavy metals (Hg, Ag, Pb, and Cu) in gills and
digestive gland of mussels, Mytilus galloprovincialis (L.).
Arch. Environ. Contam. Toxicol. 38:472-478.
Giamberini L, Auffret M, Pihan JC (1996) Hemocytes of the
freshwater mussel, Dreissena polymorpha Pallas: cytology,
cytochemistry and X-ray microanalysis. J. Moll. Stud. 62:
367-379.
Hemelraad J, Herwig HJ (1988) Cadmium kinetics in freshwa-
ter clams. IV. Histochemical localization of cadmium in
Anodonta cygnea and Anodonta anatina exposed to cad-
mium chloride. Arch. Environ. Contam. Toxicol. 17:337-
343.
Herwig HJ, Brands F, Kruitwagen E, Zandee DI (1989)
Bioaccumulation and histochemical localization of cad-
mium in Dreissena polymorpha exposed to cadmium chlo-
ride. Aquat. Toxicol. 15:269-286.
Holwerda DA (1991) Cadmium kinetics in freshwater clams. V.
Cadmium-copper interaction in metal accumulation by
Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
99

Anodonta cygnea and characterization of the metal-binding
protein. Arch. Environ. Contam. Toxicol. 21:432-437.
Ireland MP, Marigómez I (1992). The influence of dietary cal-
cium on the tissue distribution of Cu, Zn, Mg & P and his-
tological changes in the digestive gland cells of the snail
Achatina Fulica Bodwich. J. Moll. Stud. 58:157-168.
Isani G, Andreani G, Kindt M, Carpene E (2000)
Metallothioneins (MTs) in marine molluscs. Cell Mol. Biol.,
46: 311-330.
Langston WJ, Bebianno MJ, Burt GB (1998) Metal handling
strategies in molluscs. orr: 219-283 Non: Metal Metabolism
in aquatic envionments. Langston WJ & Bebianno MJ
(ed.).Chapman & Hall. London.
Lowe DM, Moore MN, Clarke KR (1981) Effects of oil on diges-
tive cells in mussels: quantitative alterations in cellular and
lysosomal structure. Aquat. Toxicol. 8:265-272.
Marigómez I, Cajaraville MP, Angulo E (1990a) Cellular distribu-
tion of cadmium in the common winkle Littorina littorea (L.)
determined by X-ray microprobe analysis and histoche-
mistry. Histochemistry 94: 191-199.
Marigómez I, Cajaraville MP, Angulo E (1990b) Histopathology
of the digestive gland-gonad complex of the marine proso-
branch Littorina littorea exposed to cadmium. Dis. Aquat.
Org. 9:229-238.
Marigomez JA, Soto M, Angulo E (1991) Responses of winkles
digestive cells and their lysosomal system to environmen-
tal salinity changes. Cell. Mol. Biol. 37:29-39.
Marigómez I, Soto M, Kortabitarte M (1996) Tissue-level bio-
markers of biological effect of mercury on sentinel slugs,
Arion ater. Arch. Environ. Contam. Toxicol. 31:54-62.
Marigómez I, Kortabitarte M, Dussart GBJ (1997) Tissue-level
biomarkers in sentinel slugs as cost-effective tools to
assess metal pollution in soils. Arch. Environ. Contam.
Toxicol. 34:167-176.
Marigómez I, Cajaraville, MP, Soto M, Lekube X (1998) Cell-
type replacement, a successful strategy of molluscs to
adapt to chronic exposure to pollutants. Cuad. Invest. Biol.
20:411-414.
Marigómez I, Soto M, Cajaraville MP, Angulo E, Giamberini L
(2002) Cellular and subcellular distribution of metals in
molluscs. Microsc. Res. Tech. 56:358-392.
Martoja R, Martoja-Pierson M (1970) Técnicas de Histología
Animal. Toray Masson. Barcelona. 350 orr.
Mason AZ, Jenkins KD (1995) Metal detoxification in aquatic
organisms. 479-608 orr. Non: Metal speciation and bioavai-
lability in aquatic systems Tessier A & Turner DR (ed.).
John Wiley and Sons. New York.
Mason AZ, Simkiss K (1983) Interactions between metals and
their distribution in tissues of Littorina littorea (L) collected
from clean and polluted sites. J. Mar. Biol. Ass. U.K. 63:
661-672.
Mason AZ, Simkiss K, Ryan KP (1984) The ultrastructural loca-
lization of metals in specimens of Littorina littorea collected
from clean and polluted sites. J. Mar. Biol. Ass. U.K.
64:699-720.
Méndez PA (1993) Composición celular y estructura tisular de
la glándula digestiva de mejillones en un programa de
seguimiento de la contaminación en el estuario de el Abra.
Lizentziatura-Tesia. UPV/EHU. 88 orr.
Mix MC, Sparks AK (1971) Repair of digestive tubule tissue of
the Pacific oyster, Crassostrea gigas, damaged by ionizing
radiation. J. Invert. Pathol. 17: 172-177.
Morgan AJ, Winters C, Stürzebaum S (1997) X-ray microanaly-
sis techniques. 245-276 orr. Non: Methods in molecular
Medicine Biology. 117: Electron Microscopy Methods and
Protocols Hajibagheri N (ed.). Humana Press Inc. Totowa
New Jersey.
Morgan JE, Morgan AJ (1998) The distribution and intracellular
compartmentation of metals in the endogeic earthworm
Aporrectodea caliginosa sampled from an unpolluted and a
metal-contaminated site. Environ. Pollut. 99:167-175.
Morton B (1983) Feeding and digestion in Bivalvia. 65-147orr.
Non: The Mollusca 5 Saleuddin ASM & Wilburg M (ed.).
Academic Press. New York.
Pavicic S, Balestreri E, Lenzi P, Raspor B, Branica MF(1991)
Isolation and partial characterization of cadmium induced
metallothionein-like proteins in Mytilus galloprovincialis.
Mar. Chem. 36:249-265.
Pearse AGE (1980) Histochemistry. Theoretical and applied.
Vol 1. preparative and optical technology. 4. edizioa.
Edinburgh: Churchill Livingstone. 1055 orr.
Porte C, Sole M, Borgi V, Martínez M, Chamorro J, Torreblanca
A, Ortiz-Zarragoitia M, Orbea A, Soto M, Cajaraville MP
(2001) Chemical, biochemical and cellular responses in
digestive gland of mussels Mytilus galloprovincialis from
the Spanish Mediterranean coast. Biomarkers 6:335-350.
Rainbow PS, Dallinger R (1993) Metal uptake, regulation and
excretion in freshwater invertebrates. 119-131 orr. Non:
Ecotoxicology of metals in invertebrates Dallinger R &
Rainbow PS (ed.). Lewis Publ. Boca Raton, Florida.
Emaitzak eta eztabaida
100

Zelula-moten ordezkapena kutsatzaileen pean jarritako muskuiluen liseri-guruinean
101
Rasmussen LPD, Hage E, Karlog O (1983) Light and electron
microscopic studies of the acute and chronic toxic effects
of N-nitroso compounds on the marine mussel Mytilus edu-
lis (L.). II. N-methyl-N-nitro-N-nitroso guanidine. Aquat.
Toxicol. 3:301-311.
Recio A, Marigomez JA, Angulo E, Moya J (1988) Zinc treat-
ment of the digestive gland of the slug Arion ater L. 1.
Cellular distribution of zinc and calcium. Bull. Environ.
Contam. Toxicol. 41:858-864.
Roesijadi G (1994) Metallothionein induction as a measure of
response to metal exposure in aquatic animals. Environ.
Health Persp. 102:91-95.
Roesijadi G (2000) Metal transfer as a mechanism for metallo-
thionein-mediated metal detoxification. Cell Mol. Biol. 46:
393-405.
Sokal RR, Rohlf FJ (1979). Biometría. Madrid: Blume. 832 orr.
Soto M, Kortabitarte M, Marigómez I (1995) Bioavailable heavy
metals in estuarine waters as assessed by metal/shell-
weight indices in sentinel mussels Mytilus galloprovincialis.
Mar. Ecol. Prog. Ser. 125:127-136.
Soto M, Cajaraville MP, Angulo E, Marigómez I (1996a)
Autometallographic localization of protein-bound copper
and zinc in the common winkle, Littorina littorea: a light
microscopical study. Histochem. J. 28:1-13.
Soto M, Cajaraville MP, Marigómez I (1996b) Tissue and cell
distribution of copper, zinc and cadmium in the mussel
Mytilus galloprovincialis determined by autometallography.
Tiss. Cell 28:557-568.
Soto M, Marigómez I (1997a) BSD extent, an index for metal
pollution screening based on the metal content within
digestive cell lysosomes of mussels as determined by
autometallography. Ecotox. Environ. Saf. 37:141-151.
Soto M, Marigómez I (1997b) Metal bioavailability assessment
in "Mussel-Watch" programmes by automated image
analysis of BSD in digestive cell lysosomes. Mar. Ecol.
Prog. Ser. 156:141-150.
Soto M, Quincoces I, Lekube X, Marigómez I. (1998a)
Autometallographed metal content in digestive cells of win-
kles: a cost-effective screening tool to monitor Cu and Zn
pollution. Aquat. Toxicol. 40:123-140.
Soto M, Quincoces I, Marigómez I (1998b)
Autometallographical procedure for the localization of
metal traces in molluscan tissues by light-microscopy. J.
Histotechnol. 21:123-127.
Syasina IG, Vaschenko MA, Zhandan PM (1997) Morphological
alterations in the digestive diverticula of Mizuhopecten
yessoensis (Bivalvia: Pectenidae) from polluted areas of
Peter the Great Bay, Sea of Japan. Mar. Environ. Res.
44:85-98.
Taylor, MG (1995). Mechanisms of metal immobilization and
transport in cells. 155-170 orr. Non: Cell Biology in
Environmental Toxicology. Cajaraville MP (ed.).University
of the Basque Country Press Service. Bilbo.
Thompson RJ, Ratcliffe NA, Bayne BL (1974) Effects of
starvation on structure and function in the digestive gland
of the mussel (Mytilus edulis, L.). J. Mar. Biol. Ass. U. K. 54:
699-712.
Triebskorn R, Künast C (1990) Ultrastructural changes in the
digestive system of Deroceras reticulatum (Mollusca:
Gastropoda) induced by lethal and sublethal
concentrations of the carbamate molluscicide cloethocarb.
Malacologia 32:89-106.
Viarengo A (1989) Heavy metals in marine invertebrates:
mechanisms of regulation and toxicity at the cellular level.
Rev. Aquat. Sci. 1:295-317.
Viarengo A, Nott JA (1993) Mechanisms of heavy metal cation
homeostasis in marine invertebrates. Comp. Biochem.
Physiol. C 103:355-372.
Viarengo A, Canesi L, Mazzucotelli A, Ponzano E (1993) Cu, Zn
and Cd content in different tissues of the Antarctic scallop
Adamussium colbecki, role of metallothionein in heavy
metal homeostasis and detoxification. Mar. Ecol. Prog. Ser.
95:163-168.
Weibel ER (1979) Stereological Methods. 1. Academic Press.
London 415 orr.
Widdows J, Moore MN, Lowe DM. 1984. Sublethal biological
effects and short-term recovery of mussels (Mytilus edulis)
and winkles (Littorina littorea) following chronic exposure to
petroleum hydrocarbon. Non: Long term effects of oil on
marine benthic communities in enclosures. Norwegian
Institute for Water Research/NIVA Technical Report No. 0-
82007.

Emaitzak eta eztabaida
102

Epitelio-zelulen proliferazioaren aldamoldezirkamareala muskuiluaren liseri-guruinean etaurdailean
Laburpena: Mytilus galloprovincialis (Lmk) muskuiluen liseri-guruineko eta urdaileko epitelio-zelulenberriztapena bromodeoxiuridinaren (BrdU) immunohistokimika bitartez ikertu da. Muskuiluak 4 mg BrdU/litsas-ur disoluzio pean etengabe mantendu ziren. Tratamenduari ekin eta 6 ordu ondoren, laginak 2 ordurohartu ziren 2 egunez. BrdU-markaketa argi-mikroskopioan zuzenean kontaketak eginez estimatu zen,balioak zelula BrdU-positiboen ‰ -tan emanez. BrdU erreakzio positiboa liseri-zelulen, zelula basofilikoen,eta konduktu zein urdaileko zelulen nukleoetan behatu zen, eta baita hemozitoetan ere. Liseri-dibertikuluetan, zelulen berriztapena eredu zirkamareala jarraituz sinkronizaturik agertu zen: BrdUmarkaketa, itsasbeheran igo eta itsasgoran jeisten zen. Liseri-zeluletan, irudi mitotikoa argiak behatu ziren.Lortutako emaitzek, zelula-mota desberdintzatuek proliferatzeari ekin diezaioketela egiaztatu ziguten, aldezaurretik egindako Proliferating Cell Nuclear Antigen (PCNA) immunohistokimikaren emaitzekin batzetorrena. Urdailean ere epitelio-zelulen berriztapena sinkronizatuta zegoela atzeman genuen.
Gako-hitzak: bromodeoxiuridina, berriztapen zelularra, mitosia, itsasaldia, fotoperiodoa, liseri-guruina,bibalbioak.
Zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean
103

Circatidal variation in epithelial cell proliferation in the mussel digestivegland and stomach
Abstract: Epithelial cell renewal in mussel (Mytilus galloprovincialis, Lmk) digestive gland and stomachwas investigated by bromodeoxyuridine (BrdU) immunohistochemistry. Mussels were exposed to 4 mgBrdU/l seawater continuously. Then, starting after 6 h treatment samples were collected every 2 h for 2 daysand BrdU labelling was estimated by direct counting at the light microscope as ‰ BrdU positive cells. BrdUpositive reaction was observed in the nuclei of digestive, basophilic, duct and stomach cells, as well as inhemocytes. Cell renewal in digestive diverticula is synchronised following a circatidal pattern: BrdU labellingincreased during low tide and decreased during high tide. Clearcut mitotic figures were identified indigestive cells, which confirmed that mature cell types proliferate, in agreement with the results fromimmunohistochemistry for proliferating cell nuclear antigen (PCNA) immunohistochemistry. Epithelial cellrenewal in the stomach also appeared to be synchronised.
Key Words: bromodeoxyuridine, cell renewal, mitosis, tide, photoperiod, digestive gland, bivalves.
Variación circamareal en la proliferación de células epiteliales en glánduladigestiva y estómago de mejillón
Resumen Se investigó la renovación de células epiteliales en glándula digestiva y estómago delmejillón (Mytilus galloprovincialis, Lmk) mediante inmunohistoquímica de bromodeoxiuridina (BrdU). Seexpusieron contínuamente mejillones a 4 mg BrdU/l agua de mar. Así,transcurridas 6 h de haberse iniciadoel tratamiento se recogieron muestras cada dos h durante 2 días y se estimó el marcaje de BrdU medianterecuento directo al microscopio fotónico en términos de ‰ de células BrdU positivas. Se observó reacciónBrdU positiva en los núcleos de las células digestivas, basófilas, de los conductos y del estómago, asícomo en hemocitos. La renovación celular en los divertículos digestivos estaba sincronizada de acuerdocon un patrón circamareal: el marcaje con BrdU aumentaba durante la marea baja y disminuía durante lamarea alta. Se identificaron cortes limpios de figuras mitóticas en células digestivas, lo que confirmó quelas células maduras son capaces de proliferar, de acuerdo con resultados anteriores deinmunohistoquímica de PCNA (Proliferating Cell Nuclear Antigen). También la renovación de células delepitelio del estómago parecía estar sincronizada.
Palabras clave: bromodeoxiuridina, renovación celular, mitosis, marea, fotoperiodo, glándula digestiva,bivalvos.
Emaitzak eta eztabaida
104

Zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean
105
SARRERA
Moluskuen liseri-guruin ehunaren oinarrizko
zelulen berriztapenari dagokionez kasik ikerlanik ez
dago, egitez. Muskuiluetan, liseri-guruina
azkenean urdailean zabaltzen diren konduktu
primarioei konduktu sekundarioen bitartez loturiko
unitate albeolotubularren multzotan antolatuta
dago. Unitate albeolotubular horien epitelioa bi
zelula-mota helduz osaturik dago, liseri-zelulak eta
zelula basofilikoak deitutakoak, hain zuzen
(Morton, 1983). Liseri-zelulek elikagaien liseriketa
intrazelularraren ardura dute, sistema endo-
lisosomiko erabat garatua daukatelarik. Zelula
basofilikoak, urriagoak, zelula jariatzaileak dira,
basofiliaren ezaugarria ematen dien erretikulu
endoplasmatiko pikortsu garatuaz hornituta
daudelarik (Dimitriadis et al., 2004). Epitelioa arras
dinamikoa da, eta elikagaien eskuragarritasunari
zein bestelako ingurumen-faktoreei erantzunez, bi
zelula-moten itxura, kopurua eta edukiak ordu gutxi
batzuetan bortizki alda daitezke (Morton 1983).
Izan ere, elikatze-ziklo bakoitzean, hots, ordu gutxi
batzuen buruan, liseri-epitelioa guztiz berriztatzen
dela proposatu da (Nelson & Morton 1979). Zelulen
berriztapen bizkor hori gertatu ala ez, zelula
basofilikoen proportzioa emendatzea dakarren
epitelioaren berrantolaketa, ingurumen-estresaren
iturri desberdinek bultzatzen dutena behin eta
berriz egiaztatu da (Thompson et al. 1974;
Rasmussen et al 1983; Cajaraville et al. 1990;
Marigómez et al. 1990; Syasina et al. 1997;
Marigómez et al. 1998a; 1. Kapitulua). Zelula-
moten ordezkapena (ingeleraz, Cell Type
Replacement; CTR) izenaz ezagutzen den
fenomeno hori, alde batetik, ingurumen-erasoa
jaso eta ordu gutxietara jada nahiko aurreratua
egon daiteke, eta, bestalde, zeharo itzulgarria da,
estresa eten eta egun gutxietara jada zelula-moten
proportzioa bere onera datorrelarik (Marigómez et
al., 1998b). Hastapenetako ikerlanetan, CTR zelula
basofilikoen proliferazioa areagotuak sor zezakeela
kontsideratu zen (Thompson et al. 1974;
Rasmussen et al 1983; Cajaraville et al. 1990;
Marigómez et al. 1990; 1998a). Haatik, zelula
basofilikoen proliferazioaren ideia guztiz baztertu
ezin bada ere, oraintsu eginiko esperimentuek
CTRren gorakadak zelula basofilikoen
proliferazioan baino gehiago liseri-zelulen galeran
datzala iradoki dute (Marigómez et al., 1998b;
Zaldibar et al., 2001). Bestalde, ingurumen-estresa
sortarazten duten iturriak desagertzekotan liseri-
zelulak proliferatzeko gai dira, laborategiko
baldintza kontrolatu pean buruturiko sendabidetze-
esperimentuetan erakutsi den moduan (Zaldibar et
al., 2001; 4. & 5. Kapituluak). Zoritxarrez, liseri-
guruinean zalantzabako irudi mitotikoak bilatzeak
ez du fruitu onik ekarri orain arte (Thompson et al.
1974; Rasmussen et al 1983; Cajaraville et al.
1990; Marigómez et al. 1990; Syasina et al. 1997;
Marigómez et al. 1998a; 1998b; Marigómez et al.
1999; Zaldibar et al. 2001).
Izan ere, mitosiak nekez aurkitu dira bibalbioen
liseri-guruinean. Ikerlan aurrendari batean, Yonge-
k (1926) anafasean omen zegoen zelula baten
marrazkia aurkeztu zuen. Autore horrek, zenbait
zelula basofilikok zelula ama (sic., young cell)
moduan joka dezaketela, eta, krustazeoen

hepatoarean bezalaxe (Icely & Nott 1992; Vogt
1994), zelula gaztez osatutako kriptetatik
(habietatik) zelulen berriztapena abia daitekeela
iradoki zuen. Are gehiago, erradiazio ionizatzaile
pean egondako ostretan liseri-guruineko epitelioa
ea zeharo desagerrarazi ondoren, epitelio-zelulen
birpopulatzea kripta-zelulen mitosien bidez
eratutako zelula-habien eraiketaz hasten da (Mix &
Sparks 1971). Hala ere, nahiz eta itxuraz
basofilikoak izan, kripta-zelulak txikiak dira, eta ezin
dira inolaz ere zelula basofiliko heldu (jariatzaile)
gisa kontsideratu, nahiz eta geroagoko artikuluetan,
bestelako argibide sustatzailerik gabe, zelula
basofilikoak bibalbioen liseri-guruineko epitelioaren
birsortzearen erantzuleak diren zelula gazteak
direla proposatu den (Fankboner 1971; Palmer,
1979; Morton 1983). Aitzitik, Proliferating Cell
Nuclear Antigen (PCNA) immunohistokimikak
oraintsu erakutsi duenez (Marigómez et al., 1999),
muskuiluetan bai liseri-zelulak eta baita zelula
basofilikoak ere desberdintzatzeko, S-fasean
sartzeko eta proliferatzeko gai dira, sagu helduaren
listu-guruinetan zatiketa autologoa pairatzen duten
zenbait zelula-mota helduek egiten duten bezalaxe
(Denny et al., 1993). Halaber, Mya arenaria-ren
liseri-tubuluetan ere, epitelio-zelula helduak PCNA -
positiboak dira (Hanselmann et al., 2000).
Ondorioz, zelula amek epitelioen berriztapenean
parte hartuko lukete baina soilik epitelioan kalte
masiboak ematekotan; bestela, baldintz
normaletan, epitelio-zelulen berriztapena liseri-
zelula eta zelula basofiliko helduen zatiketa
autologoa dela medio burutuko litzateke.
Ornodunen ehunetan, epitelio-zelulen berriztapen-
dinamika eta prozesu horretan inplikatutako zelula-
motek jasandako aldaketak, egoera fisiologiko eta
patologikoen menpean daudela modu sinesgarrian
erakutsi da (Karam & Leblond 1993; Santa Barbara
et al., 2003).
Muskuiluen liseri-guruineko epitelioaren
berriztapena sinkronizatuta egonez mitosiak gauez
burutuko balira, aurreneko ikerlanetan mitosirik
aurkitu ez izana azalduko litzateke (Marigómez et
al., 1998b; Zaldibar et al., 2001). Zelulen zatiketa-
tasan aldamolde zirkadiarrak algetatik hasita
ugaztunetaraino deskribatu izan dira (Makarov et al.
1995; Abrahamsen et al. 1998; Bjarnasson et al.
1999; García et al. 2001). Are gehiago, zenbait
eredu esperimentaletan behintzat, zelulen
proliferazioa eta ordezkatze-prozesuak batez ere
gauez gertatzen direna jakin badakigu (Scheving et
al., 1978; Abrahamsen et al., 1998; Bjarnasson et
al., 1999; García et al., 2001).
Muskuiluen liseri-guruineko zelulen
proliferazioak patroi zirkadiarra jarraitzen duen
antzemateko, zelula proliferatzaileen banaketa
bromodeoxiuridina (BrdU) immunohistokimika
aplikatuz ikertu da Mytilus galloprovincialis (Lmk.)
muskuiluan. BrdU timidinaren analogoa da, eta
ziklo zelularraren S-fasean zehar zelula
proliferatzaileen DNAri eransten zaio. Muskuiluak, 4
mg BrdU/l-ko kontzentrazioa zuen itsas-uretan
mantendu ziren 2 egunez. BrdUren agerpena
immunohistokimika bitartez detektatu zen, eta
zelula proliferatzaileen proportzio erlatiboa (BrdU-
markaketa) denbora-tarte desberdinetan
erregistratu zen, BrdU-markaketan aldaketa
zirkadiarrak identifikatu ahal izateko.
Emaitzak eta eztabaida
106

Zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean
107
MATERIAL ETA METODOAK
Erreaktibo kimikoak
Erabilitako erreaktibo kimiko guztiak maila
analitikokoak eta Sigma-Aldrich (St. Louis,
Missouri, Amerikako Estatu Batuak), merkatal-
etxekoak dira besterik agertu ezean.
Laginak eta prozedura esperimentala
Gutxi gorabehera 35-45 mm-ko luzerako
maskorra zuten mareazpiko M. galloprovincialis
(Lmk) muskuiluak Plentzian (Bizkaiko Golkoa
43°24'I, 2°56'M) hartu ziren, uztailan eta goizerdiko
itsasbeheran (10:12 a.m.). Laginketa mareazpiko
muskuiluetara mugatu zen, itsasaldien zikloa eta
fotoperiodoaren arteko elkarrekintza ekiditeko
asmoz. Muskuiluak (n=80) laborategira garraiatu
ziren eta 15 l-ko polietilenozko akuario batean
mantendu ziren 15°C-tan eta etengabeko
aireztapenaz. Itsas-ura argi ultramorea erabiliz
esterilizatu zen, eta karbono aktiboz zein beirazko
zuntzetan zehar iragazi egin zen; esperimentuak
iraun bitartean itsas-ura ordezkatu ez zelarik.
Muskuiluei, ornogabe itsastarrentzako jaki
komertziala (Marine Invertebrate Diet, Carolina,
Burlington, North Carolina, Amerikako Estatu
Batuak) egunero eman zitzaien, goizeko 12:00-tan.
Baldintza horietan, muskuiluak BrdU-tratamendu
pean 2 egunez mantendu ziren. Hasieran, itsas-
uretan 4 mg BrdU/l-ko pultsua aplikatu zitzaien, eta
gerora 2 mg/l-ko pultsuak 4 orduro aplikatu ziren,
maila zitotoxikoetara heltzeke BrdUren
eskuragarrita-suna etengabe ziurtatu ahal izateko.
Horrela, saiakuntza-denboran zehar BrdUren
galerarik gertatuko ez balitz ere, uretan metatutako
gehiegizko BrdUren kontzentrazioa 20 mg/l baino
baxuagoa izango litzateke. Ornogabe itsastarrekin
egindako pultsu-esperimentuetan BrdUren
kontzentrazioak 500 mg/l-ko bezain altuak erabili
dituzte beste ikertzaileek (Candia Carnevali et al.,
1997; Goergen et al., 2002). Hala ere, guk BrdU
kontzentrazio askoz baxuagoa aplikatzea erabaki
genuen honako arrazoi hauengatik: (1) muskuiluak
laborategian egoera egokian mantentzeko ur
bolumen handia ezinbestekoa zen, (2) 20-40 mg
BrdU/l baino kontzentrazio altuagoek, muskuiluen
ehunen zelulen berriztapena inhibi dezakete
(Martínez-Expósito et al., 1994); eta (3) aurreneko
ikerketek, 4 mg BrdU/l-ko kontzentrazioa liseri-
guruineko epitelioan BrdU-markaketa konspikuoa
erdiesteko nahikoa dela adierazi baitziguten.
Muskuiluak denbora-tarte desberdinetan sakrifikatu
ziren, BrdU-markaketaren fluktuazio-erritmoak
identifikatu ahal izateko.
Zelula BrdU-positiboen immunohistokimika
Liseri-guruinaren zati txikiak Carnoy fixatzailean
(%60 etanol absolutua, %30 kloroformoa eta %10
azido azetikoa) fixatu ziren ordu batez 4°C-tan.
Fixatu ondoren, laginak goranzko graduazioko
etanolezko bainu-serie batean deshidratatu ziren
eta parafinan inkluditu ziren 60°C-tan. Ebakiak (3-
4 µm-ko lodiera) American Optical mikrotomoan
ebaki ziren eta aldez aurretik (3-amino propil
trietoxisilanoz) silanizatutako portaobjektuetan jaso
ziren. Ondoren, laginak xilenoz desparafinatu ziren,
beheranzko graduazioko etanolezko bainu-serie
batean hidratatu eta fosfato indargetzaile salinoz

(PBS; ingeleraz, Phosphate-Buffered Saline)
garbitu ziren. Peroxidasa jarduera endogenoa
ekiditeko, ebakiak %3 zuen H2O2-an inkubatu ziren
5 minutuz, eta berriro PBSz garbitu ziren. DNA, 0.1
N HCl-ez desnaturalizatu zen ordu batez, 37°C-tan.
Ebakiak, Vectastain Elite ABC Kit-ak (Vector
Laboratories, Burlingame, California, Amerikako
Estatu Batuak) hornitutako suero blokeatzailean
ordu batez inkubatu ziren giro-tenperaturan eta
berriro ere PBSz garbitu ziren. Gero, ebakiak
BrdUren aurkako antigorputz espezifikoan (Roche,
Basilea, Suitza; %0.1 bovine serum albumine zuen
PBSan 1:10 diluitua) inkubatu ziren. Laginak PBSz
garbitu ziren, eta ondoren, Vectastain Elite ABC Kit-
aren bitartez, abidina-biotina entzima-konplexua
erabiliz antigorputza ikustarazi zen. Laburki,
ebakiak, PBSn diluitutako saguaren aurkako
zaldiaren IgG antigorputz sekundario biotinilatuan
inkubatu ziren, 30 minutuz, eta ondoren PBSn
garbitu ziren. Peroxidasa jarduera ikustarazteko
kromogenoaren disoluzioa (3-amino-9-
etilokarbazola, %0.015 H2O2 zuen sodio azetato
indargetzailean -pH 5.2- disolbatua) erabili zen.
Azkenik, ebakiak hematoxilinaz kontrastatu ziren
(10 s), dabiluretan garbitu eta Kaiser gelatina
glizerolatuan (Merck & Co., Inc, Whitehouse
Station, New Jersey, Amerikako Estatu Batuak)
muntatu ziren. Mikrografiak, Olympus BX50
(Olympus Tokio Japonia) mikroskopioari atxiki
zegoen Olympus PM20 argazki-kameraz erdietsi
ziren.
Zelula BrdU-positiboen kuantifikazioa
Zelula BrdU-positiboen kuantifikazioa, argi-
mikroskopioan kontateka zuzenaren bidez burutu
zen. Liseri-epitelioan zelula BrdU-positiboen
proportzioa antzemateko asmoz, 1000 zelula
zenbatu ziren lagin bakoitzean, beste ikerketetan
erabilitako lagin-tamainaren arabera (Okudela et
al., 1999). Aldez aurretik egindako behaketa
histologikoek, liseri-epitelioaren tubulu moduko
ebakidura bakoitzean gutxienez 20-30 zelula
daudela erakutsi zuten. Hori dela eta, eguneroko
lana errazteko asmoz, zelula BrdU-positiboen
zenbaketa aleatorioki hautatutako 40 tubulu
moduko ebakiduretan burutu zen (800-1200 zelula)
liseri-guruin bakoitzean. Halaber, eskuarki konduktu
sekundarioen ebakidura bakoitzean 70-90 zelulak
agertzen direnez gero, zelula BrdU-positiboen
zenbaketa 10 konduktu sekundarioen ebakiduretan
burutu zen (700-900 zelula). Horrek, laginaren
tamaina egoki lortzearen eta konduktu
sekundarioen zeharkako ebakidurak lortzearen
zailtasunaren arteko konpromisozko erabakia
suposatu zuen. Datuak adierazteko momentuan
‰--ra pasatu ziren. Urdailean, kontaketa trantsektu
bat eginez burutu zen, eta guztira 200 zelula
zenbatu ziren epitelioan barrena. Zelula-kopuru
baxuago hori, urdailean jarduera proliferatzailea
oso handia dela egiaztatu ondoren hautatu zen.
Datuak kasu honetan ere ‰-ko balioetan eman
dira, aurrekoekin konparagarriak izan ahal izateko.
Analisi estatistikoak, SPSS/PC+ programa
erabiliz burutu ziren (SPSS, Microsoft). Denboraren
eragina ikusi ahal izateko, faktore bateko ANOVA
burutu zen, eta konparaketa anitzak eginez
parekatutako batezbestekoen artean
desberdintasun esangarriak (p<0.05) finkatzeko
Emaitzak eta eztabaida
108

Duncan testa aplikatu zen. Azkenik, erregresio ez
linealaren analisiak Sigma Plot programaren
bitartez (SPSS, Microsoft) burutu ziren.
Mitosien bilaketa
Beste parafinazko ebaki batzuk (3 µm-koak)
aldez aurretik deskribatu bezala prozesatu ziren,
hematoxilina-eosinaz tindatu eta argi-
mikroskopioan behatu ziren, liseri-guruinaren
ehunean irudi mitotikoak identifikatzeko asmoz.
Mikrografiak, Olympus BX50 mikroskopioari atxiki
zegoen Olympus PM20 kameran lortu ziren.
EMAITZAK
Muskuiluen liseri-guruinan, zelula BrdU-
immunorreaktiboak urdail, konduktu sekundario eta
liseri-albeoloen epitelioetan lokalizatu egin ziren (1
Ird.). Liseri-epitelioan, liseri-zelulek zein zelula
basofilikoek positiboki markatutako nukleoa
erakutsi zuten, nahiz eta zelula erreakzionagarri
gehienak liseri-zelulak izan (1A Ird.). Konduktu
sekundarioetan, zelula BrdU-positiboak zati
ziliatuetan kokaturik agertu ziren gehien bat (1B
Ird.). Urdailean beste epitelioetan baino askoz
zelula immunopositibo gehiago behatu ziren (1C
Ird.). Azkenik, hemozitoen nukleoetan ere
markaketa bortitza aurkitu genuen. Hemozitoak,
sarritan epitelio-zelula BrdU-positiboen azpian
kokaturik agertu ziren, xafla basalaren (zentzu
histologikoan) azpian, hain zuzen ere (1B Ird.).
Gainera, zelulen soberakinak, nukleo BrdU-
positiboak barne, liseri-traktuaren argian askotan
behatu ziren, hondakin zelularrekin batera (1D Ird.).
BrdUren aurkako antigorputzarik gabe inkubatu
ziren kontrol-ebakiek ez zuten inolako markaketarik
erakutsi (1E Ird.).
Faktore bateko ANOVA analisien (1. Taula) zein
batezbestekoen arteko konparaketa anitza egiteko
Duncan testaren arabera, BrdU-markaketa
baloreak laginketa-denboren artean esangarriki
desberdinak izan ziren. Duncan testaren arabera
esangarriki desberdinak ziren zenbait azpitalde
homogeneo bereiztu ziren, 2. Ird.-an erakutsi den
bezalaxe. Liseri-epitelioan, zelulen ‰20-‰25
bitartean BrdUz markatuta agertu zen; konduktu
sekudarioetan, berriz, markaketa 8 aldiz baxuagoa
izan zen gutxi gorabehera (‰3), eta urdailean 6
biderrez altuagoa (‰150) (2 Ird.). Liseri-guruineko
zelulen BrdU-markaketak liseri-zelulen proliferazio-
tasaren aldaketak islatu zuen batik bat, zelula
basofiliko BrdU-positiboak oso noizean behin baino
behatu ez zirelako. Liseri-epitelioan, BrdU-
markaketaren jaitsiera nabarmenak 20:00-24:00
eta 08:00-10:00 tarteetan erregistratu ziren,
bitartean igoerak behatu zirelarik (2A Ird.).
Esperimentazio-egun bietan aldamolde erritmiko
berdintsua aurkitu genuen. Erregresio ez-linealaren
analisiaren ondoren, gutxi gorabehera 12 h-ko
maiztasuna zuen funtzio sigmoideo bati doitu
zitzaion, esangarriki. (3 Ird.). BrdU-markaketak
konduktu sekundarioetan eta urdaileko epitelioan
antzeko irudi erritmikoen patroia erakutsi zuen (2B,
C Ird.), baina erregresio ez-lineala eta korrelazio-
koefizientea ez ziren estatistikoki esangarriak izan.
Muskuiluen liseri-guruina ohi bezala behatu
ostean, irudi mitotikoak han-hemenka identifikatu
ziren liseri-zeluletan (1F Ird.), eta hemozito zein
urdaileko zeluletan ere.
Zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean
109

EZTABAIDA
PCNA erabiliz aldez aurretik egindako lanek,
liseri-epitelioa osatzen duten zelula-mota guztiek
proliferatzeko gaitasuna dutela erakutsi dute
(Marigómez et al., 1999). Liseri-zelulak zein zelula
basofilikoak desberdintzatzeko eta mitosian
sartzeko gai direla proposatu da. Dena den, bere
biziraupen luzeari esker, PCNA-immunoerreakzio-
nagarritasuna, zelula amen mitosien bidez
sortarazitako zelula desberdintzatuen nukleoetan
ere detekta daiteke (Casasco et al., 1995). Gure
ikerlan honetan, liseri-zeluletan irudi mitotikoak
agertu izanak eta BrdU-immunohistokimikaren
emaitzek, zelulen berriztapena liseri-zelula eta
zelula basofiliko helduen zatiketa autologoaren
bitartez erdiesten dela egiaztatu dute. Hala ere, oro
har, BrdU-markaketa PCNA-markaketa baino
ahulagoa da. Markatutako zelulen proportzioan
aurkitutako desberdintasunak, 3H-Timidina, PCNA
eta BrdU erabili direneko tasen araberakoak izan
daitezke (Elsässer et al., 1994; Bromley et al.,
1996; Moriki et al., 1996). Horrela, arratoiaren
areako zelula azinarretan, zelula PCNA-positiboak
3H-Timidina-positiboak baino 10 aldiz ugariagoak
dira, bi teknika histokimikoen bidez lortutako
emaitzen artean korrelazio esangarria dagoen
arren (Elsässer et al., 1994). Azalpenik egokiena,
seguru asko, PCNAren bizi-iraupena 20 ordukoa
izatean datza; S-fasearen iraupena baino luzeagoa
dena, hain zuzen. Hori dela eta, zelula PCNA-
positiboak ez dira soilik S-fasean daudenak baizik
eta S-fasean daudenak edo eta oraintsu S-fasea
gainditu berri dutenak ere. Ondorioz, PCNA
immunohistokimikak zelula proliferatzaileak
lokalizatzeko seinale bortitzagoa eskaini dezaken
arren; proteinaren biziraupen luzeagoak epitelioen
berriztapenaren dinamikak aztertzeko desabantaila
suposa diezaguke. BrdU-immunohistokimika, bere
aldetik, helburu hori betetzeko egokiagoa dela
dirudi, BrdUren biziraupena laburragoa denez,
pultsu-esperimentuetan aplikatzeko aukera eskaini
baitiezaguke. Izatez, proliferazioaren markatzaile
immunohistokimiko sendoena eta fidagarriena dugu
BrdU, S-faseak baino ez dituelako identifikatzen
(Bromley et al., 1996). BrdU-pultsu esperimentuak
hainbat animali eredutan aplikatu diren arren,
ornogabe itsastarren kasuan murritzak dira
(Harzsch et al., 1999; Awaji & Suzuki, 1995; Candia
Carnevali et al., 1997; Goergen et al., 2002).
Muskuilu itsastarretan, BrdU-pulsua nola aplikatu
arazo nagusietakoa izan genuen. Ikerlan honetan,
BrdU uretan disolbatu zen, eta beraz, elikatze-
iragazketa aurrera joan ahala BrdU zelulei iristen
zitzaien. Prozedura horrek, liseri-guruineko
ehunetan BrdU eranstea ahalbidetzen zuela
baieztatu zen, aldez aurretik zakatz-ehunetan
behatu izan zen bezalaxe (Martínez-Expósito et al.,
1994).
Immunohistokimika aplikatzekotan, pultsu batez
emandako BrdU, mitosiondoko zeluletan beren
bizitza osoan zehar detekta daiteke, nukleoan
seinalea esponentzialki diluituko luketen
proliferazio-prozesu suzesiboak ez gertatzekotan
behinik behin (Ward et al., 1991). Ikerlan honetako
baldintza esperimentalen arabera, BrdU-markaketa
esperimentuak aurrera egin ahala igoz joango zela
Emaitzak eta eztabaida
110

espero genuen. Aitzitik, BrdU-markaketa oso bizkor
jaitsi zen, 4 orduren buruan: ‰25-tik ‰2-ra unitate
albeolotubularretan eta ‰150-‰40-ra urdailean (2
Ird.). Nukleoetatik BrdU-markaketa desagertzea,
zelulen zatiketa suzesiboen ondorioa ote dugu, edo
markatutako zelulak jaio orduko hiltzearen ondorioa
ote da?.
Arratoiaren gibeleko epitelio-zeluletan egindako
Zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean
111
1. Irudia: BrdU-positiboak diren nukleoen identifikazioa (geziak) unitate albeolotubularretako liseri-zeluletan (A), konduktuetan (B),urdailean (C) eta urdailaren argian askatutako materialean (D). Kontrol-ebakiean ez da inolako zelula BrdU-positiborik behatu (E).Hematoxilina-eosinaz tintatutako kromosomak (gezia) mitosian dagoen liserri-zelula batean (F). A) unitate albeolotubularra; D)konduktuko epitelio-zelula; H) hemozitoa; S) urdaileko epitelio-zelula.DC) liseri-zelula; BC) zelula basofilikoa. Eskala-marrak: 25µm (A-E); 50 µm (F).

in vitro BrdU-pultsu esperimentuetan, BrdU-
immunomarkaketa hiru zatiketa zelular eta gero
desagertu dela demostratu da (Evarts et al., 1996).
Mihiko keratinozito murinoetan, BrdU-markaketaren
balio maximo eta minimoen artean gutxi
gorabehera 12 orduko aldea dago (Garcia et al.,
2001), baina indize mitotikoaren piko maximoa
BrdU-markaketaren piko maximoa baino 8 ordu
beranduago gertatzen da, BrdUren piko
minimoaren momentuan, hain zuzen. Horrela,
BrdU-markaketa zelulen zatiketaren ostean
diluituko balitz, horrek muskuiluen liseri-epitelioan
DNA bikoizketa eta zelulen zatiketa prozesu nahiko
azkarrak izatea suposatuko luke. Muskuiluetan
ziklo zelularraren iraupenari buruz ditugun datu
urriek, ordea, kontrakoa erakutsi dute. Esaterako,
zakatzetako epitelio-zeluletan ziklo zelularrak 30
ordu inguru irauten dituela frogatu da (Martínez
Expósito et al., 1994). Hala ere, epitelio-zelulek
behin eta berriro, zelula amen moduan,
proliferatuko balute, BrdU-markaketa ez zen
diluituta suertatuko; zatiketa berri bakoitzean gure
baldintza esperimentaletan etengabe eskuragarria
den BrdU gehiago behin eta berriro erantsiko
bailukete.
Bestalde, erabilitako BrdU kontzentrazioak
zitotoxikoak ez direla pentsatu arren (Martínez
Expósito et al., 1994), etengabeko BrdU-
esposizioak, zelula-mota bakoitzari dagokion modu
espezifikoan ziklo zelularraren progresioa alda
lezake (Diermeier et al., 2004). Batetik, BrdUk
apoptosia eragin dezake. Bestetik, BrdU eransteak
faboratutako DNA parekatze-akatsen ondorioz ziklo
zelularra eten daiteke (Yao et al., 1998). Are
gehiago, BrdUk, arearen kasuan esaterako,
garapen-programa espezifikoki eragin dezake,
proteinen sintesi- eta jariapen-patroietan aldaketa
deigarriak sortaraziz (Van Nest et al., 1983). Beraz,
gure baldintza esperimentaletan BrdU erantsi duten
liseri-guruineko epitelioko-zelulak alteratuta egonez
gero epiteliotik aska zitezkeen. Horrek, bertan
aurkitutako BrdU-markaketaren berehalako
beherakadak azalduko lituzke. Izan ere, gure
ikerlanean liseri-traktuaren argian zelula BrdU-
positiboak maiz behatu ditugu soberakinen artean
(1D Ird.).
Liseri-unitate albeolotubularreko epitelioan
BrdU-markaketaren kuantifikazioari buruzko
emaitzak liseri-zelulen proliferazio-tasaren
aldaketei dagozkie batez ere, BrdU-markatutako
zelula basofilikoak oso gutxitan behatu baitira. Gure
emaitzen arabera, epitelio-zelulen berriztapena
nabarmenagoa da urdailean unitate albeolotubula-
rretan eta konduktuetan baino. Urdail-epitelioan,
‰150 zelula ere egon daitezke aldi berean S-
fasean eta, dirudienez, epitelioa bere osotasunean
3 egunetan berrizta daiteke. Bestalde, liseri-zelulen
‰25-a eta bakarrik konduktuetako zelulen ‰3-a
sartzen dira S-fasean une batean. Beraz, unitate
Emaitzak eta eztabaida
112
1. Taula: Ikertutako hiru epitelioetan laginketa-denborak BrdU-markaketan duen eragina finkatzeko egindako faktore batekoANOVA-ren laburpena. P(F), Fisher ratioaren probabilitatea;a.g., askatasun-graduak. Estatistikoki esangarriak gertatu zirendesberdintasunak (p<0.05), asterisko batez adierazita daude.
F ratio a.g.(1) a.g. (2) p(F)
Unit. albeolotubularrak 5.060 13 61 <0.001*
Liseri-konduktuak 3.668 13 61 <0.001*
Urdaila 3.830 13 38 0.002*
(1) taldeen artean; (2) talde barruan
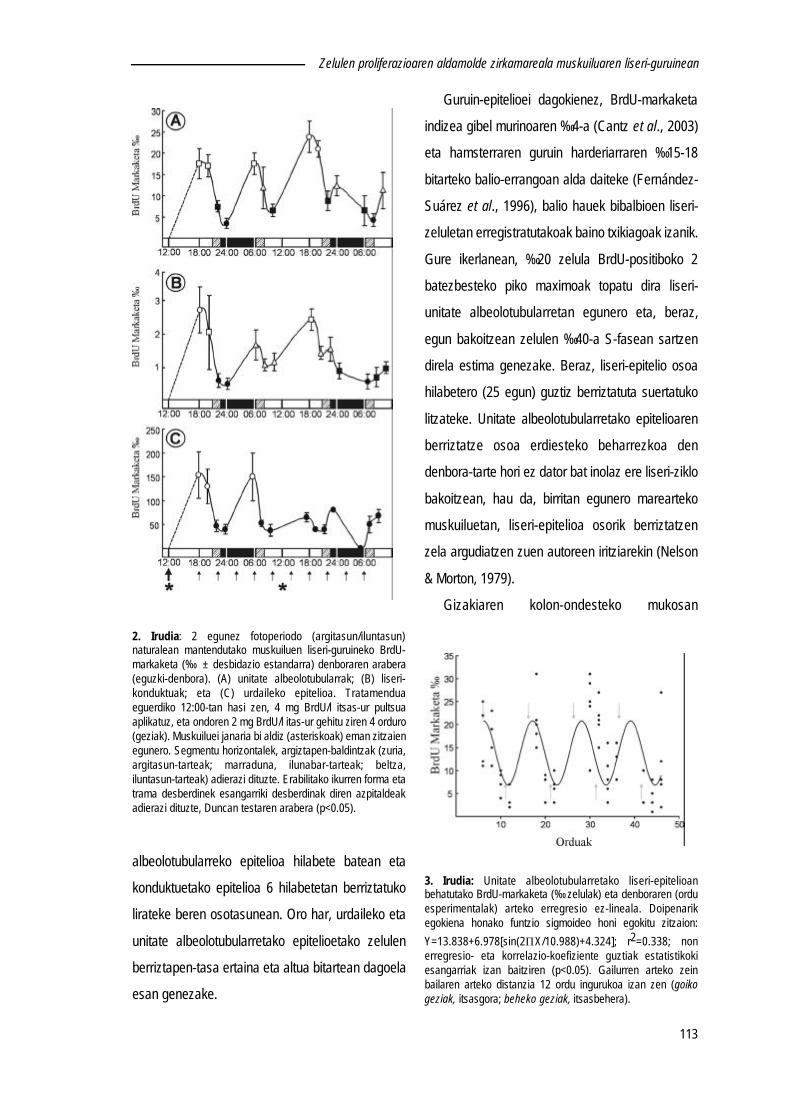
albeolotubularreko epitelioa hilabete batean eta
konduktuetako epitelioa 6 hilabetetan berriztatuko
lirateke beren osotasunean. Oro har, urdaileko eta
unitate albeolotubularretako epitelioetako zelulen
berriztapen-tasa ertaina eta altua bitartean dagoela
esan genezake.
Guruin-epitelioei dagokienez, BrdU-markaketa
indizea gibel murinoaren ‰4-a (Cantz et al., 2003)
eta hamsterraren guruin harderiarraren ‰15-18
bitarteko balio-errangoan alda daiteke (Fernández-
Suárez et al., 1996), balio hauek bibalbioen liseri-
zeluletan erregistratutakoak baino txikiagoak izanik.
Gure ikerlanean, ‰20 zelula BrdU-positiboko 2
batezbesteko piko maximoak topatu dira liseri-
unitate albeolotubularretan egunero eta, beraz,
egun bakoitzean zelulen ‰40-a S-fasean sartzen
direla estima genezake. Beraz, liseri-epitelio osoa
hilabetero (25 egun) guztiz berriztatuta suertatuko
litzateke. Unitate albeolotubularretako epitelioaren
berriztatze osoa erdiesteko beharrezkoa den
denbora-tarte hori ez dator bat inolaz ere liseri-ziklo
bakoitzean, hau da, birritan egunero marearteko
muskuiluetan, liseri-epitelioa osorik berriztatzen
zela argudiatzen zuen autoreen iritziarekin (Nelson
& Morton, 1979).
Gizakiaren kolon-ondesteko mukosan
Zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean
113
3. Irudia: Unitate albeolotubularretako liseri-epitelioanbehatutako BrdU-markaketa (‰ zelulak) eta denboraren (orduesperimentalak) arteko erregresio ez-lineala. Doipenarikegokiena honako funtzio sigmoideo honi egokitu zitzaion:
Y=13.838+6.978[sin(2ΠX/10.988)+4.324]; r2=0.338; nonerregresio- eta korrelazio-koefiziente guztiak estatistikokiesangarriak izan baitziren (p<0.05). Gailurren arteko zeinbailaren arteko distanzia 12 ordu ingurukoa izan zen (goikogeziak, itsasgora; beheko geziak, itsasbehera).
2. Irudia: 2 egunez fotoperiodo (argitasun/iluntasun)naturalean mantendutako muskuiluen liseri-guruineko BrdU-markaketa (‰ ± desbidazio estandarra) denboraren arabera(eguzki-denbora). (A) unitate albeolotubularrak; (B) liseri-konduktuak; eta (C) urdaileko epitelioa. Tratamenduaeguerdiko 12:00-tan hasi zen, 4 mg BrdU/l itsas-ur pultsuaaplikatuz, eta ondoren 2 mg BrdU/l itas-ur gehitu ziren 4 orduro(geziak). Muskuiluei janaria bi aldiz (asteriskoak) eman zitzaienegunero. Segmentu horizontalek, argiztapen-baldintzak (zuria,argitasun-tarteak; marraduna, ilunabar-tarteak; beltza,iluntasun-tarteak) adierazi dituzte. Erabilitako ikurren forma etatrama desberdinek esangarriki desberdinak diren azpitaldeakadierazi dituzte, Duncan testaren arabera (p<0.05).

batezbesteko BrdU-markaketa indizea ‰77-110
bitartekoa da (Potten et al., 1992), arratoiaren
epitelio gastrikoan ‰150 ingurukoa den bitartean
(Gama et al., 2000). BrdU-markaketa balio horiek
guruinak ez diren epitelio normaletan balio-atalase
maximoak direla dirudi (Bertalanffy 1962; Humme &
Potten 1980; Potten et al. 1992; Pawlinoski et al.,
1999; Gama et al. 2000), eta ikerlan honetan
urdailean aurkitutakoen antzekoak dira. Liseri-
unitate albeolotubularren kasuan aplikatutakoen
antzeko kalkuluetan oinarrituz, konduktuetako eta
urdaileko berriztapen osoa 6 hilabetetan (175 egun)
eta 3 egunetan, hurrenez hurren, burutuko
litzatekeela ondorioztatu da. Liseri-konduktuetako
epitelioaren BrdU-markaketa indizea baxua
kontsidera daiteke, ‰2 eta ‰3 bitarteko indizea
duen obarisektomizatutako arratoien endometrio-
aren moduko beste ehunen antzera (Pawlinoski et
al., 1999).
Gure emaitzen arabera, muskuiluen liseri-
unitate albeolotubularretako epitelio-zelulen
berriztapenak patroi fluktuagarria jarraitzen du.
Patroi horrek, gailur eta bailarak 6 ordutako
txandatan agertzen direneko erritmo zirkamareal
perfektoa irudikatzen du. Antzeko erritmo
zirkamarealak hainbat ornogabe itsastarren mota
askotako prozesu fisiologikoetan behatu dira
(Naylor 1996, Abello et al., 1997). Bi proliferazio-
ziklo gauzatzen dira egun batean zehar.
Proliferazio-eredua errepikatu egiten da, zelulak
itsasgorako orduetan S-fasean sartzen dira, geroko
orduetan, zatiketa zelular bizkorrak pairatzeko. Izan
ere, zelula BrdU-positiboak ia desagerturik daude 4
orduren buruan. Liseri-konduktuetako BrdU-
markaketa patroia ez da unitate albeolotubularretan
deskribatutakoaren oso desberdina, baina seinalea
hain baxua denez laginketa-denboren arteko
desberdintasunak BrdU-markaketan esangu-
ratasun biologiko eta esangarritasun estatistikorik
gabekoak dira. Urdailean ere epitelioaren
berriztapena sinkronizatuta dagoela dirudi.
Bibalboen liseri-traktuan epitelio-zelulek zatitu
baino lehen migratu egiten dutela deskribatu da
mitosiak epitelioaren erpinaldean gertatuz (Mix
1970; Leibson & Frolova 1994). Aitzitik, nukleo
BrdU-positiboak epìtelio-zelulen oinaldeko
herenean kokaturik daudela aurkitu dugu.
Gizakiaren aho-epitelioan (Bjarnasson et al.,
1999) eta bizkar-muinean (Abrahamsen et al.,
1998), zein arratoiaren liseri-traktuan ere (Scheving
et al., 1978), bereziki zelula ugari aurkitu dira S-
fasean ilunabarrean, gauan zehar beraien kopurua
nabarmenki jaitsiz. Xaguaren keratinozitoetan S-
fase gehienak gauez ematen dira (Garcia et al.,
2001). Otarrainaren zerebroan neurogenesiaren
mailarik altuena ilunabar aldean gertatzen da, gaua
jarduera fisiologiko maximoaren garaia izanik
(Goergen et al., 2002). Epitelio-zelulen proliferazio-
jardueraren aldaketa erritmikoen denbora-patroia
ehun- eta espezie-espezifikoa izan liteke
zalantzarik gabe, muskuiluen liseri-guruinean
elikatze eta liseriketa moduko jarduera zikliko zein
erritmo marealen menpekoa izanik (Nelson &
Morton 1979; Morton 1983). Ikerlan honetan,
marea-ziklo eta fotoperiodoaren arteko
interferentziak ekiditeko asmoz, bakarrik
mareazpiko muskuiluak aztertu dira. Hala ere,
Mytilusen habitata itsasbeherako mailatik metro
Emaitzak eta eztabaida
114

gutxi batzutara baino ez da iristen. Eskualde
horretan animaliek kinada fotikoak eta gorantz zein
beherantzko mareen pauta erritmikoak ere jasotzen
dituzte. Ondorioz, marea-erritmoa liseri-sistemaren
zelulen proliferazioaren prozesu erritmikoak
gobernatzeko Zeitgeber perfektua izan daiteke,
kasu honetan bezala muskuiluak mareazpikoak
badira ere.
Laburbilduz, liseri-zeluletan irudi mitotikoak
behatu izanak eta BrdUren emaitzek, zelulen
berriztapena liseri-guruineko zelula helduen
zatiketa autologoaren bitartez gauzatzen dela
egiaztatu dute. Urdaileko eta liseri-unitate
albeolotubularreko epitelioek, zelulen berriztapen-
tasa ertaina eta altua bitarteko dute. Epitelio-zelulen
berriztapena sinkronizatuta dago, 24 ordutan 2 ziklo
osoak betetzen dituen patroi zirkamareala jarraituz.
BIBLIOGRAFIA
Abello P, Warman CG, Naylor E (1997) Circatidal moulting
rhythm in the shore crab Carcinus maenas. J. Mar. Biol.
Assoc. U.K. 77:277-280.
Abrahamsen JF, Smaaland R, Sothern RB, Laerum OD (1998)
Circadian cell cycle variations of erythro- and myelopoiesis
in humans. Eur. J. Haematol. 58:333-345.
Awaji M, Suzuki T (1995) The pattern of cell proliferation during
pearl sac formation in the Pearl oyster. Fisheries Sci.
61:747-751.
Bertalanffy FD (1962) Cell renewal in the gastrointestinal tract
of man. Gastroenterology 11:472-475.
Bjarnasson GA, Jordan RCK, Sothern RB (1999) Circadian
variation in the expression of cell-cycle proteins in human
oral epithelium. Am. J. Pathol. 154:613-622.
Bromley M, Rew D, Becciolini A, Balzi M, Chadwick C, Hewitt
D, Li YQ, Potten CS (1996) A comparison of proliferation
markers (BrdUrd, Ki-67, PCNA) determined at each cell
position in the crypts of normal human colonic mucosa.
Eur. J. Histochem. 40:89-100.
Cajaraville MP, Díez G, Marigómez JA, Angulo E (1990)
Responses of basophilic cells of the digestive gland of
mussels to petroleum hydrocarbon exposure. Dis. Aquat.
Org. 9:221-228.
Candia Carnevali MD, Bonasoro F, Biale A (1997) Pattern of
bromodeoxyuridine incorporation in the advanced stages
of arm regeneration in the feather star Antedon
mediterranea. Cell Tiss. Res. 289:363-374.
Cantz T, Zuckerman DM, Burda DR, Dandri M, Goricke B,
Thalhammer S (2003) Quantitative gene expression
analysis reveals transition of fetal liver to mature
hepatocytes after transplantation in uPA/RAG-2 mice. Am.
J. Pathol. 162:37-45.
Casasco A, Casasco M, Icaro Cornaglia A, Mazzini G, De
Renzis R, Tateo S (1995) Detection of Bromo-
deoxyuridine- and Proliferating Cell Nuclear Antigen-
immunoreactivities in tooth germ. Connective Tiss. Res.
32:63-70.
Denny PC, Chai Y, Klauser DK, Denny PA (1993) Parenchymal
cell proliferation and mechanisms for maintenance of
granular duct and acinar cell populations in adult male
mouse submandibular gland. Anat. Rec. 235:475-485.
Diermeier S, Schmidt-Bruecken E, Kubbies M, Kuntz-Schughar
LA, Brockhoff G (2004) Exposure to continuous
bromodeoxyuridine (BrdU) differentially affects cell cycle
progression of human breast and bladder cancer cell lines.
Cell Prolif. 37:195-206.
Dimitriadis VK, Domouhtsidou GP, Cajaraville MP (2004)
Cytochemical and histochemical aspects of the digestive
gland cells of the mussel Mytilus galloprovincialis (L.) in
relation to function. J. Mol. Histol. 35:501-509.
Elsässer HP, Biederbick A, Kern HF (1994) Growth of rat
pancreatic acinar cells quantitated with a monoclonal
antibody against the proliferating cell nuclear antigen. Cell
Tiss. Res. 276:603-609.
Evarts RP, Hu Z, Omori N, Omori M, Marsden ER, Thorgeirsson
SS (1996) Precursor-product relationship between oval
cells and hepatocytes: comparison between tritiated
thymidine and bromodeoxyuridine as tracers.
Carcinogenesis 17:2143-2151.
Fankboner PV (1971) Intracellular digestion of symbiotic
zooxanthellae by host amoebocytes in giant clams
(Bivalvia: Tridacnidae), with a note on the nutritional role of
the hypertrophied siphonal epidermis. Biol. Bull. 141:222-
234
Zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean
115

Emaitzak eta eztabaida
116
Fernández-Suárez A, López JM, Carbajo S, Alvarez-Uria M,
Carbajo-Pérez E (1996) New insights into cytodynamics of
the hamster Harderian gland as provided by the
bromodeoxyuridine-labelling method. Histol. Histopathol.
11:351-355.
Gama P, Goldfeder EM, Bertacini de Moraes JC, Alvares EP
(2000) Cell proliferation and death in the gastric epithelium
of developing rats after glucocorticoid treatments. Anat.
Rec. 260:213-221.
García MN, Barbeito CG, Andrini LA, Badrán AF (2001)
Circadian rhythm of DNA synthesis and mitotic activity in
tongue keratinocytes. Cell Biol. Inter. 25:179-183.
Goergen EM, Bagay LA, Rehm K, Benton JL, Beltz BS (2002)
Circadian control of neurogenesis. J. Neurobiol. 53:90-95.
Hanselmann R, Smolowitz R, Gibson D (2000) Identification of
proliferating cells in hard clams. Biol. Bull. 199:199-200.
Harzsch S, Miller J, Benton J, and Beltz B, (1999) From embryo
to adult: Persistent neurogenesis and apoptotic cell death
shape the lobster deutocerebrum. J. Neurosci. 19:3472-
3485.
Humme WJ, Potten CS (1980) Changes in proliferative activity
as cells move along undulating basement membranes in
stratified squamous epithelium. British J. Dermatol.
103:499-504.
Icely JD, Nott JA (1992) Digestion absorption: digestive system
and associated organs. 147-201 orr. Non: Harrison FW, &
Humes AG (ed) Microscopic anatomy of invertebrates. 10:
Decapod Crustacea. Wiley-Liss, New York.
Karam SK, Leblond CP (1993) Dynamics of epithelial cells in
the corpus of the mouse stomach. I. Identification of
proliferative cell types and pinpointing of the stem cell.
Anat. Rec. 236:259-279.
Leibson NL, Frolova LT (1994) Winter-spring essential
reorganization of cell proliferation in the digestive tract
epithelia in the mussel Crenomytilus grayanus. Mar. Biol.
118:471-477.
Makarov VN, Schoschina EV, Luning K (1995) Diurnal and
circadian periodicity of mitosis and growth in marine
microalgae. 1. Juvenile sporophytes of laminariales
(phaeophyta). Eur. J. Phycol. 30:261-266.
Marigómez I, Cajaraville M P, Angulo E (1990) Histopathology
of the digestive gland-gonad complex of the marine
prosobranch Littorina littorea exposed to cadmium. Dis.
Aquat. Org. 9:229-238.
Marigómez I, Kortabitarte M, Dussart GBJ (1998a) Tissue-level
biomarkers in sentinel slugs as cost-effective tools to
assess metal pollution in soils. Arch. Environ. Contam.
Toxicol. 34:167-176.
Marigómez I, Cajaraville MP, Soto M, Lekube X (1998b) Cell-
type replacement, a successful strategy of molluscs to
adapt to chronic exposure to pollutants. Cuad. Invest. Biol.
18:431-435.
Marigómez I, Lekube X, Cancio I (1999) Immunochemical
localisation of proliferating cells in mussel digestive gland
tissue. Histochem. J. 31:781-788.
Martínez-Expósito MJ, Pasantes JJ, Méndez J (1994)
Proliferation kinetics of mussel (Mytilus galloprovincialis)
gill cells. Mar. Biol. 120:41-45.
Mix MC (1970) Cell renewal systems in the gut of the oyster,
Crassostrea gigas. Veliger 14:202-205.
Mix MC, Sparks AK (1971) Repair of digestive tubule tissue of
the Pacific oyster, Crassostrea gigas, damaged by ionizing
radiation. J. Invert. Pathol. 17:172-177.
Moriki T, Takahashi T, Kataoka H, Hiroi M, Yamane T, Hara H
(1996) Proliferation marker MIB-1 correlates well with
proliferative activity evaluated by BrdU in breast cancer: an
immunohistochemical study including correlation with
PCNA, p53, c-erbB-2 and estrogen receptor status. Pathol.
Internat. 46:953-961.
Morton B (1983) Feeding and digestion in Bivalvia. 65-147 orr.
Non: Saleuddin A S M. & Wilburg M (ed) The Mollusca. 5.
Academic Press, New York.
Naylor E (1996) Crab clockwork: the case for interactive
circatidal and circadian oscillators controlling rhythmic
locomotor activity of Carcinus maenas. Chronobiol.
Internat. 13:153-161.
Nelson L, Morton JE (1979) Cyclic activity and epithelial
renewal in the digestive gland tubules of the marine
prosobranch Maoricrypta monoxyla (Lesson). J. Moll. Stud.
45:262-283.
Okudela K, Ito T, Kameda Y, Nakamura N Kitamura H (1999)
Immunohistochemical analysis for cell proliferation-related
protein expression in small cell carcinoma of the
esophagus; a comparative study with small cell carcinoma
of the lung and squamous cell carcinoma of the
esophagus. Histol. Histopathol. 14:479-485.
Palmer RE (1979) A histological and histochemical study of
digestion in the bivalve Arctica islandica L. Biol. Bull.
156:115-129.

Pawlinoski M, Melén-Mucha G, Mucha S (1999) The
involvement of the renin-angiotensin system in the
regulation of cell proliferation in the rat endometrium.
Cell.Mol. Life Sci. 55:506-510.
Potten CS, Kellett M, Roberts SA, Rew DA, Wilson GD (1992)
Measurement of in vivo proliferation in human colorectal
mucosa using bromodeoxyuridine. Gut 33:71-78.
Rasmussen LPD, Hage E, Karlog O (1983) Light and electron
microscopic studies on the acute and chronic toxic effects
of N-nitroso compounds on the marine mussel Mytilus
edulis (L.). II N-methyl-N-nitro-N-nitroso-guanidine. Aquat.
Toxicol. 3:301-311.
Santa Barbara P, van den Brink GR, Roberts DJ (2003)
Development and differentiation of the intestinal
epithelium. Cell Mol. Life Sci. 60:1322-1332.
Scheving LE, Burns RE, Pauly JE, Tsai TH, (1978) Circadian
variation in cell division of the mouse alimentary tract, bone
marrow and corneal epithelium. Anat. Rec. 191:479-486.
Syasina IG, Vaschenko MA, Zhandan PM (1997) Morphological
alterations in the digestive diverticula of Mizuhopecten
yessoensis (Bivalvia: Pectenidae) from polluted areas of
Peter the Great Bay, Sea of Japan. Mar. Environ. Res.
44:85-98.
Thompson RJ, Ratcliffe NA, Bayne BL (1974) Effects of
starvation on structure and function in the digestive gland
of the mussel (Mytilus edulis, L.). J. Mar. Biol. Ass. U. K.
54:699-712.
Van Nest G, Raman RK, Rutter WJ (1983) Effects of
dexamethasone and 5-bromodeoxyuridine on protein
synthesis and secretion during in vitro pancreatic
development. Dev. Biol. 98:295-303.
Vogt G (1994) Life-cycle and functional cytology of the
hepatopancreatic cells of Astacus astacus. Crustacea,
Decapoda. Zoomorphology 144:83-101.
Ward JM, Henneman JR, Osipova GY, Anisimov VN (1991)
Persistence of 5-bromo-2'-deoxyuridine in tissues of rats
after exposure in early life. Toxicology 70:345-352.
Yao XH, Sugihara H, Hattori T, Katsura K, Takamatsu T (1998)
Regulation of apoptotic cell death in the pyloric glands of
the canine stomach. Virchows Arch. 433:275-280.
Yonge CM (1926) The digestive diverticula in Lamellibranchia.
Trans. Roy. Soc. Edin. 54:703-718.
Zaldibar B, Iturralde J, Cancio I, Marigómez I (2001) Reversible
cell-type replacement in marine mollusc Littorina littorea
exposed to cadmium. SETAC Europe Ann. Conf., Vienna.
Zelulen proliferazioaren aldamolde zirkamareala muskuiluaren liseri-guruinean
117

Emaitzak eta eztabaida
118

Zelula epitelialen berriztapena muskuiluen liseri-guruin eta urdailean: sasoi, adin eta erregimenmarealari lotutako aldamoldeak
Laburpena: Tamaina, adin eta itsasaldi-erregimen desberdineko Mytilus galloprovincialis (Lmk)muskuiluen liseri-albeolo eta urdaileko epitelioko zelulen jarduera proliferatzailearen aldamolde naturalaikertu da sasoi desberdinetan. Muskuiluak, timidinaren analogoa den bromodeoxiuridinarekin (BrdU) 6orduz tratatu ondoren, 2 orduro hurrengo 36 ordutan BrdUren immunohistokimika burutu zen. Ondoren,zelula BrdU-positiboen proportzio erlatiboa BrdU-markaketa gisa (‰) kuantifikatu zen. Sasoiaridagokionez, desberdintasun esangarriak behatu ziren BrdU-markaketan. Udan, udazkenean eta neguanbaino askoz jarduera proliferatzaile altuagoa neurtu zen. Adin (tamaina) desberdineko animalien artean ezzen zelulen jarduera proliferatzaile desberdinarik behatu. Bestalde, marearteko eta mareazpiko muskuiluenliseri-guruineko epitelioen artean desberdintasunak aurkitu ziren, bai BrdU-markaketa mailetan, eta baitaaldamolde-patroian ere, non marearteko muskuiluetan BrdU-markaketa fotoperiodoak zein itsasaldi-erregimenak modulatua baitzegoen eta mareazpiko muskuiluetan, patroi zirkamareala jarraitu baitzuen.
Gako-hitzak: bromodeoxiuridina, immunohistokimika, liseri-zelula, zelula basofilikoa, berriztapenzelularra, erritmo zirkamareala, tamaina, adina, sasoia, marea, bibalbioak.
Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
119

Epithelial cell renewal in the digestive gland and stomach of mussels: season,age and tidal regime related variations
Abstract: The natural variability in cell proliferation activity in the epithelium of the digestive gland andstomach was investigated in mussels, Mytilus galloprovincialis (Lmk), of different age and tidal level atdifferent seasons. After treating mussels with the thymidine analogue bromodeoxyuridine (BrdU) for 6hours, BrdU immunohistochemistry was performed every 2 hours for the next 36. Then, the relativeproportion of BrdU positive cells was quantified as BrdU labelling (‰). Marked seasonal differences wererecorded in BrdU labelling, with much higher proliferating activity in summer than in autumn and winter. Cellproliferation seemed not to be significantly dissimilar between mussels of different age (size). In contrast,the digestive gland epithelium of mussels from intertidal and subtidal populations differed not only in thelevels but also in the pattern of variation of BrdU labelling, which in intertidal mussels appeared to bemodulated by photoperiod and tide, unlike in subtidal mussels, in which variations followed a circatidalpattern.
Key words: bromodeoxyuridine immunohistochemistry, digestive cell, basophilic cell, cell epithelialrenewal, circatidal rhythm, size, age, season, tide, bivalve.
Renovación de células digestivas en glándula digestiva y estómago demejillon: variación según la época, edad y régimen mareal
Resumen: Se ha investigado la variabilidad natural en la actividad proliferativa de las células de la glánduladigestiva y del estómago de mejillón, Mytilus galloprovincialis (Lmk), de diferente edad y régimen de mareaen diferentes épocas del año. Los mejillones fueron tratados con bromodeoxiuridina (BrdU), un análogo dela timidina, durante 6 horas y en las siguientes 36 horas se realizó la immunohistoquímica de BrdU cada 2horas. A continuación, se cuantificó la proporción relativa de células BrdU positivas (‰). Se observarondiferencias significativas en el marcaje de BrdU relacionadas con la época del año, siendo la proliferacióncelular mucho más marcada en verano que en otoño e invierno. La proliferación celular no resultó diferenteentre animales de diferentes edades (tamaño). Por el contrario, en el epitelio de la glándula digestiva demejillones intermareales y submareales no sólo resultó diferente el nivel, sino también el patrón devariación del marcaje de BrdU. En mejillones intermareales ese patrón parece modulado por el fotoperiodoy la marea, a diferencia de los submareales en los que las variaciones siguen un patrón circamareal.
Palabras clave: inmunohistoquímica de bromodeoxiuridina, célula digestiva, célula basofílica, renovacióncelular, ritmo circamareal, tamaño, edad, época, marea, bibalvio.
Emaitzak eta eztabaida
120

Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
121
SARRERA
Muskuiluen liseri-guruineko epitelioa
desberdintzatutako 2 zelula-motez osaturik dago,
liseri-zelula eta zelula basofilikoa, alegia. Liseri-
zelulek, oso sistema endo-lisosomiko garatua dute
eta elikagaien liseriketa intrazelularraz arduratzen
dira. Zelula basofilikoek, izaera basofiloa damaien
erretikulu endoplasmatiko pikortsu garatua dute,
eta liseri-zelulak baino urriagoak dira. Marigómez
et al.-ek, (1999), PCNA immunohistokimika erabiliz,
bi zelula-mota horiek proliferatzeko gai direla
frogatu zuten. Liseri-zeluletan irudi mitotikoak
aurkitu izanak eta BrdU immunohistokimikak,
zelulen berriztapena liseri-zelula eta zelula
basofiliko helduen zatiketa autologoaz burutzen
dela baieztatu dute (Zaldibar et al., 2004). Liseri-
dibertikuluen epitelio-zelulen berriztapena
sinkronizatuta dago patroi zirkamareala jarraituz,
BrdU-markaketaren aldaketek adierazi zutenez
(Zaldibar et al., 2004). Esperimentu horiek,
mareazpiko muskuiluekin soilik burutu ziren arren,
edonola ere itsasaldi-erregimena eraginkorra zela
proposatu zen. Dirudienez, liseri-guruineko
epitelioaren zelulen berriztapena, Izagirre-ren
(2000) arabera liseri-zelulen jarduera fisiologikoa
(adb, liseriketa intrazelularra) murrizten deneko
une jakinetara mugatuta dago. Beraz, jarduera
fisiologiko eta ziklo zelularraren arteko
elkarrekintza ezagutzea, liseri-epitelioaren zelulen
berriztapen ulertzeko oinarrizkoa izango litzateke.
Testuinguru horretan, liseri-guruinaren gainean
eragina duten aldagai naturalek (adina, sasoia,
itsasaldi-erregimena), liseri-guruineko zeluletan
aurkitutako jarduera proliferatzailearen eredu
zirkamareala, aldarazi edo modula dezaketen
finkatzea erronka erabakigarria da. Liseri
guruinaren forma eta funtzioa, liseritze-iraupena
eta intentsitatea eta beste prozesu fisiologikoak
barne, aldagai horien arabera aldatu egiten da
(Etxeberria et al., 1994; Cancio et al., 1999;
Fernández-Reiriz et al., 2001; Wong & Cheung,
2001; Le Pennec & Le Pennec, 2002; Pazos et al.,
2003). Are gehiago, tamaina (adin) desberdineko
muskuiluek, liseri-guruineko zelulen proliferazio-
eredu ezberdina dute. Izatez, SDS-PAGE
elektroforesiaren ondoko PCNAren
immunotransferentziak, muskuilu txikien (gazte
heldugabeak) liseri-guruineko zelulak muskuilu
handienetakoak (helduak) baino proliferazio
gaitasun handiagoa zutela erakutsi zuten
(Marigómez et al., 1999).
Hiru sasoi desberdinetan (negua, udara eta
udazkena) itsasarteko bi eskualde desberdinetan
(mareartekoa eta mareazpikoa) hartutako tamaina
desberdineko (txikiak-gazteak eta handiak-
helduak) Mytilus galloprovincialis (Lmk) muskuiluen
liseri-guruineko eta urdaileko zelula
proliferatzaileen banaketa, BrdU-
immunohistokimika bitartez ikertu zen.
MATERIAL ETA METODOAK
Erreaktibo kimikoak
Erabilitako erreaktibo kimiko guztiak maila
analitikokoak eta Sigma-Aldrich (St. Louis,
Missouri, Amerikako Estatu Batuak), merkatal-
etxekoak dira besterik agertu ezean.

Atariko esperimentua
Mareazpiko eta marearteko Mytilus
galloprovincialis muskuiluen liseri-guruineko eta
urdaileko zelulen proliferazioa konparatu ahal
izateko, BrdU-pultsua marearteko muskuiluak
pairatzen duten urperatze-aldira mugatu behar izan
zen (6 ordu). Aldez aurretik burututako
esperimentuak uretango BrdU-esposizio jarraia
aplikatu ondoren, BrdU-markaketa patroi
zirkamareala erakutsi zutenez, oraingo honetan
BrdU-pultsua egunaren unerik egokiena finkatzeko
zenbait aurretiko esperimentu diseinatu ziren.
Helburu hori betetzeko, 160 mareazpiko muskuilu
gazte (maskorraren luzera <15 mm) eta 160 heldu
(maskorraren luzera 35-45 mm) uztailean
itsasbeheran jaso ziren Plentzian (Bizkaiko Golkoa,
43º24´ I 2º56´ M). Muskuiluak laborategira garraiatu
ziren, eta polietilenozko 4 akuariotan (80 muskuilu
15 l itsas uretan) etengabeko aireztapenaz eta
15ºC-tara mantendu ziren. Itsas-ura argi ultramorea
erabiliz esterilizatu zen, eta karbono aktiboz zein
beirazko zuntzetan zehar iragazi egin zen. 12
orduro itsas-ura aldatu zen eta muskuiluei, uraren
aldaketarekin batera, janaria eman zitzaien (Marine
Invertebrate Diet, Carolina, Carolina Supply Co,
Burlington, North Carolina, Amerikako Estatu
Batuak). Bi esperimentu osagarri burutu ziren.
Lehenengo esperimentuan (Eguneko pultsuaren-
esperimentua), muskuilu gazte eta helduak,
bakoitza bere aldetik 4 BrdU mg/l zuen itsas-uretan
6 orduz jarri ziren, 14:00-tan hasita. Ondoren,
muskuiluak BrdUrik gabeko itsas-uretan murgildu
ziren, eta hurrengo 36 ordutan 2 orduro 5
espezimen sakrifikatu ziren. Bigarren
esperimentuan, (Gaueko pultsuaren esperimentua)
BrdU-pultsu berdina gauean zehar (24:00-tatik
06:00-ak arte) aplikatu zen eta muskuiluak
hurrengo 28 ordutan 2 orduro sakrifikatu ziren.
Esperimentu nagusia
Urteko hiru sasoi desberdinetan, negua
(martxoa), uda (uztaila) eta udazkena (urria),
muskuiluak (n = 320), Plentzian itsasbeheran hartu
ziren, laborategira garraiatu eta lehen aipatutako
baldintza berberetan mantendu ziren. Marearteko
eta mareazpiko populazioetan, adin desberdineko
(gazteak: <15 mm eta helduak: 35-45 mm
maskorraren luzera) muskuiluak hautatu ziren, eta
beraz, sasoi bakoitzean lau joku esperimental (80
muskuilu bakoitzean) ezarri ziren: marerazpiko
muskuilu helduak (LS), marearteko helduak (LI),
mareazpiko gazteak (SS), eta mareazpiko gazteak
(SI).
Talde esperimental guztiak, 4 BrdU mg/l zuen
itsas-uretan 6 orduz jarri ziren, 14:00-tan hasita,
marearteko muskuiluen urperatze-denboran zehar,
hain zuzen. Ondoren, muskuiluak BrdU gabeko
uretan murgildu ziren, eta denbora-tarte
desberdinetan talde bakoitzetik 5 espezimen
sakrifikatu ziren, BrdU-markaketa kuantifikatzeko.
LS eta SS taldeetan itsas-ura 12 orduro aldatzen
zen. LI eta SI taldeko muskuiluak marearteko
baldintzetan mantendu ziren, 6 orduz airean eta 6
orduz uretan murgildurik, euren jatorrizko lekuan
zegoen itsasaldi-erregimena jarraituz. Janaria
(Marine Invertebrate Diet) talde esperimental
guztietako muskuiluei batera 12 orduro ematen
zitzaien , marearteko muskuiluak uretan murgilduta
Emaitzak eta eztabaida
122

Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
123
zeuden momentuan.
BrdU immunohistokimika
Liseri-guruinaren zati txikiak Carnoy fixatzailean
(%60 etanol absolutua, %30 kloroformoa eta %10
azido azetikoa) ordu batez fixatu ziren 4°C-tan,
goranzko graduazioko etanolezko bainu-serie
batean deshidratatu ziren eta parafinan inkluditu
ziren, 60°C-tan. Ebakiak (3-4 µm-ko lodiera)
American Optical mikrotomo batean erdietsi ziren,
eta aldez aurretik (3-amino propil trietoxisilano)
silanizatutako portaobjektuetan jaso ziren.
Ondoren, ebakiak xilenoz desparafinatu,
etanolezko bainu-serie batean hidratatu, eta
PBSaz (fosfato indargetzaile salinoa; ingelesez,
Phosphate-Buffered saline) garbitu ziren.
Peroxidasa jarduera endogenoa ekiditeko, ebakiak
%3 zuen H2O2-an 5 minutuz inkubatu ziren eta
berriro PBSaz garbitu ziren. DNA, ordu batez
37°C-tan 0.1N HCl-z desnaturalizatu zen. Ebakiak,
borato indargetzailean (pH=8.5) neutralizatu eta
PBSaz garbitu ziren. Ondoren, ebakiak Vectastain
Elite ABC Kit-ak (Vector Laboratories, Burlingame,
Kalifornia, Amerikako Estatu Batuak) hornitutako
serum blokeatzaile normalean (NBS, ingelesez;
Normal Blocking Serum) ordu batez inkubatu ziren,
giro-tenperaturan, eta berriro PBSaz garbitu ziren.
Gero, ebakiak BrdUren aurkako antigorputz
espezifikoan (Roche, Basilea, Suitza; 1:10 %0.1
behi-albumina seruma zuen PBSan) inkubatu
ziren. Laginak PBSaz garbitu, eta ondoren,
Vectastain Elite ABC Kit-a erabiliz abidina-biotina-
entzima (ABC) metodoaren bidez
immunokonplexuak ikustarazi ziren. Laburki,
ebakiak, PBSan diluitutako sagu aurkako zaldiaren
IgG antigorputz sekundario 30 minutuz biotinilatuan
inkubatu ziren, eta ondoren PBSan biziki garbitu
ziren. Peroxidasa jardueraren ikustaraztea, sodio
azetato indargetzailean (pH = 5.2) 3-amino-9-
ethylkarbazola eta %0.015 H2O2 zituen
kromogeno-disoluzioa erabiliz erdietsi zen.
Azkenik, ebakiak hematoxilinaz kontrastatu (10
segundu), dabiluretan garbitu eta Kaiser gelatina
glizerinatuan (Merck & Co., Inc, Whitehouse
Station, New Jersey, Amerikako Estatu Batuak)
muntatu ziren. Mikrografiak, Olympus BX50
(Olympus, Tokio, Japonia) mikroskopioari atxiki
zegoen Olympus PM20 argazki-kameraz erdietsi
ziren.
Zelula BrdU-positiboen kuantifikazioa
Zelula BrdU-positiboen kuantifikazioa, argi-
mikroskopioan konkateka zuzenaren bidez burutu
zen. Liseri-epitelioan zelula BrdU-positiboen
proportzioa antzemateko asmoz, lagin bakoitzean
1000 zelula zenbatu ziren, beste ikerketetan
erabilitako lagin-tamainaren arabera (Okudela et
al., 1999; Zaldibar et al., 2004). Aldez aurretik
egindako behaketa histologikoek, liseri-epitelioaren
tubulu moduko ebakidura bakoitzean gutxienez 20-
30 zelula daudela erakutsi zuten. Hori dela eta,
eguneroko lana errazteko asmoz, zelula BrdU-
positiboen zenbaketa aleatorioki hautatutako 40
tubulu moduko ebakiduretan burutu zen (800-1200
zelula) liseri-guruin bakoitzean. Halaber, eskuarki
konduktu sekundarioen ebakidura bakoitzean 70-
90 zelulak agertzen direnez gero, zelula BrdU-
positiboen zenbaketa 10 konduktu sekundarioen
ebakiduretan burutu zen (700-900 zelula). Horrek,

laginaren tamaina egoki lortzearen eta konduktu
sekundarioen zeharkako ebakidurak lortzearen
zailtasunaren arteko konpromisozko erabakia
suposatu zuen. Datuak adierazteko momentuan
‰-ra pasatu ziren. Urdailean, kontaketa trantsektu
bat eginez burutu zen, eta guztira 200 zelula
zenbatu ziren epitelioan barrena. Zelula-kopuru
baxuago hori, urdailean jarduera proliferatzailea
oso handia dela egiaztatu ondoren hautatu zen.
Datuak kasu honetan ere ‰-ko balioetan eman
dira, aurrekoekin konparagarriak izan ahal izateko.
EMAITZAK
Liseri epitelioko liseri-zelula zein zelula
basofilikoek, BrdU-positibo ziren nukleoak erakutsi
zituzten, nahiz eta zelula BrdU-positibo gehienak
liseri-zelulak izan (1. Ird.). Urdaila, nukleo BrdU-
positibo gehien erakutsi zituen organoa izan zen (1.
Ird.). Tindaketaren kontrol-ebakietan ez zen inolako
positibotasunik behatu (1. Ird.).
Atariko esperimentuak
BrdU-markaketari dagokionez, liseri-guruin eta
urdaileko epitelioan patroi zirkamareal
konparagarria ezagutu zen, seinale/zarata erlazioa
urdailean txikiagoa izan arren. Beraz, liseri-
guruineko epitelioko BrdU-markaketa emaitzak
soilik aurkeztuko dira hemendik aurrera.
Eguneko pultsu-esperimentuan, muskuilu
helduetan liseri-guruineko ‰33 zelulek BrdU
markaketa aurkeztu zuten. Animali gazteetan,
ordea, markaketa baxuagoa izan zen (‰21) (2A eta
2C Ird.). Liseri-epitelioaren BrdU-markaketaren
kuantifikazioari dagozkion emaitzak, liseri-zelulen
proliferazio-tasan gertatutako aldaketei egokitu
zitzaizkien, zelula basofiliko BrdU-positiboak
noizean behin soilik behatu baitziren. Termino
kualitatiboetan, muskuilu gazte zein helduetan
aldamolde bera aurkitu zen. Aldamoldea itsasaldi-
erregimenak (IE) edo argia/iluntasuna zikloak (AIZ)
gobernatua dagoen zehazteko asmoz, uretan
murgiltzearen (itsasbeheratik itsasgoranzko bidea)
eta aire-esposizioaren (itsasgoratik itsasbeherantz)
uneetako batezbesteko BrdU-markaketa kalkulatu
zen (1. Taula, 3. Ird). Helburu horretarako, eta
esperimentua aurrera joan ahala behatutako BrdU-
markaketan beherakada kontuan hartuz, kalkuluak
egiteko BrdU-pultsu osteko lehenengo 12 ordutako
datuak baino ez ziren aintzat hartu. Batezbesteko
BrdU-markaketaren balio altuena, gaueko
murgiltze-aldian (marea altua) aurkitu zen muskuilu
gazte zein helduetan; muskuilu gazteetan, gainera,
eguneko murgilketa-aldian tarteko balioak
erregistratu zirelarik (3. Ird.). Bi bidetako ANOVA
analisiak muskuilu helduetan IE eta IE x AIZ
faktoreek batezbesteko BrdU-markaketan eragin
esangarria zutela adierazi zuten, muskuilu
gazteetan, ordea, IE x AIZ elkarrekintza faktorerik
esangarriena (p<0.05) suertatu zen (2. Taula).
Gaueko pultsu-esperimentuan, muskuilu
helduetan liseri-epitelioko zelulen ‰7-rarte BrdU-ko
markaketa izatera iritsi zen, muskuilu gazteetan
markaketa-pikoa altuagoa (‰12) izanik (2B eta 2D
Ird.). Termino kualitatiboetan muskuilu gazte zein
helduetan aldamolde bera aurkitu zen. Aldamoldea,
itsasaldi-erregimenak (IE) edo argia/iluntasuna
zikloak (AIZ) gobernatua den zehazteko asmoz,
Emaitzak eta eztabaida
124

urera murgiltzearen eta aire-esposizioaren
uneetako batezbesteko BrdU-markaketa kalkulatu
zen, egunez zein gauez (1. Taula, 3. Ird.). Helburu
horretarako, eta BrdU-pultsuaren ostean zein
hurrengo egun argiko unean BrdU-markaketan piko
nabarmenik behatu ez zelako kontuan hartuz,
kalkuluak egiteko BrdU-pultsua eman eta 24 orduko
osteko datuak baino ez ziren aintzat hartu.
Batezbesteko BrdU-markaketa maximoa gauez
behatu zen arren, bereziki muskulilu gazteetan, ez
zen talde esperimentalen artean desberdintasun
esangarririk topatu (3. Ird.). Honekin bat, bi bidetako
ANOVA analisiak, muskuilu gazteetan BrdU-
markaketaren gainean efektu esangarria soilik AIZ-
k zuela adierazi zuen. Muskuilu helduetan, bere
aldetik, IE, AIZ eta beraien elkarrekintzek ez zuten
efektu esangarririk eduki BrdU-markaketaren
gainean (2. Taula).
Esperimentu nagusiak
Urte-sasoi, adin eta itsasaldi-erregimenaren
efektua zelulen proliferazioaren gainean
Urteko hiru sasoi, eta bi itsasaldi-erregimen
desberdinetan hartutako bi adineko muskuiluen
zelulen proliferazioa modu integratu batean
konparatzeko hainbat parametro desberdin
kalkulatu ziren (3. Taula). Orotariko BrdU-
markaketa/egun indizea pultsu osteko lehenengo
12 orduetan behatutako piko maximoan
erregistratutako zelula BrdU-positiboen kopurua (A,
3. Taula), balio hau bikoiztuz 24 ordutako datuak
islatzeko asmoz (B, 3. Taula) eta zuzenketa-faktore
batez biderkatuz (C, 3. Taula) 3. Taulako balioak
lortzeko estimatu ziren. Zuzenketa-faktorea, (ZF)
aurreko esperimentuetan ez bezala (Zaldibar et al.,
2004), BrdU-gehipena jarraia izan beharrean 6
Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
125
1. Irudia: BrdU positiboak diren nukleoen identifikazioa (geziak) unitate albeolotubularretako liseri-zeluletan, konduktuetan etaurdailean (A-D); D) konduktuko epitelio-zelula), A) unitate albeolotubularra, H) hemozitoa, BC) zelula basofilikoa, DC) liseri-zelula,S) urdaileko epitelio-zelula. Ebaki kontrolean ez da inolako zelula BrdU positiborik behatzen (E). Eskala-marra: 50 µm

orduko pultsua erabili zela kontuan hartuz aplikatu
zen, konparazioak egiteko asmoz.
ZF= 1.21 = GJ/PO.
BrdU-markaketa/egun indizea = PO x 2 x ZF;
non, ZF: Zuzenketa-faktorea; GJ: BrdU-gehipen
jarraian lehenego 12 orduetan neurtutako zelula
BrdU positiboak; eta PO: 6 orduko BrdU pultsu
ostean llehenego 12 orduetan neurtutako zelula
BrdU positiboak, baitiran.
Azkenik, epitelioa bere osotasunean
berriztatzeko beharrezkoa den denbora, egunetan
(epitelio-berriztapen denbora; EBD) kalkulatu zen,
aldez aurretik deskribatutako kalkuluetan oinarrituz
(D, 3. Taulan).
Liseri-guruineko jarduera proliferatzailea,
nabarmenki aldatu zen sasoien artean, udan
altuena izanik, eta itsasaldi-mailen artean (3.
Taula). BrdU-markaketa/egun indizearen baliorik
altuenak (C, 3. Taulan) udan erregistratu ziren; noiz
liseri-guruineko epitelioaren EBD (D, 3. Taulan) adin
eta itsasaldi-erregimenaren arabera 2 eta 6 aste
bitartekoa izan baitzen. BrdU-markaketa/egun
indizearen baliorik baxuenak, eta beraz EBD
baliorik altuenak (6 hilabete inguru), udazkeneko
mareazpiko muskuiluetan aurkitu ziren. Neguan,
BrdU-markaketa/egun indizean ez zen
desberdintasun esangarririk behatu itsasaldi-
erregimenen artean, baina balioak apur bat
Emaitzak eta eztabaida
126
2. Irudia: 2 egunez fotoperiodo (argitasun/iluntasun) naturalean mantendutako mareazpiko muskuiluen liseri-guruineko BrdUmarkaketa (‰ ± desbidazio estandarra) denboraren arabera (eguzki-denbora). (A) Eguneko pultsuko muskuilu gazteak. (B)Gaueko pultsuko muskuilu gazteak. (C) Eguneko pultsuko muskuilu helduak (D) Gaueko pultsuko muskuilu helduak. Geziek BrdUpultsua eman zitzaien ordua adierazten dute. Marra horizontalek, argiztapen-baldintzak (zuria, argitasun-tarteak; marraduna,ilunabar-tarteak; beltza, iluntasun-tarteak),eta esperimentua egin zeneko egunetako itsasaldi baldintzak (zuri, airean; marraondulatuak, uretan adierazten dituzte).Erabilitako ikurren forma eta trama desberdinek esangarriki desberdinak diren azpitaldeakadierazi dituzte, Duncan testaren arabera (p<0.05).

handiagoak izan ziren muskuilu gazteetan
helduetan baino (3. Taula). Udazkeneko
marearteko eta mareazpiko muskuiluen artean
desberdintasun harrigarriak behatu ziren,
marearteko muskuiluetan zelulen proliferazioa
garrantzitsua zen bitartean, mareazpiko
muskuiluetan kasik ez baitzegoen. Udan bereziki,
BrdU-markaketa/egun indizea, muskuilu helduetan
gazteetan baino altuagoa izan zen (3. taula).
Epitelio-zelulen proliferazioaren aldamoldeak
Neguko eta udazkeneko mareazpiko
muskuiluetan, liseri-epitelioko zelulen proliferazio-
jarduera antzemanezina izan zen (3. Taula), beraz
epitelio-zelulen berriztapenean ez zen inolako
aldamolderik ageri. Aitzitik, desberdina izan arren,
udako muskuiluetan zein udazkeneko mareazpiko
muskuiluetan, zelulen proliferazioan aldamolde
garbiak antzeman ziren (2A, 2C eta 4. Ird.).
Marearteko eta mareazpiko muskuiluen artean
proliferazioaren aldamolde desberdinak identifikatu
zirela aipatzekoa da.
Mareazpiko muskuiluek, bai heldu zein gazteek,
BrdU-markaketaren aldamolde bera erakutsi zuten,
atariko esperimentuetan (Eguneko pultsu-
esperimentua 2A eta 2C Ird.) deskribatu zen
moduan. Hala ere, marearteko muskuiluen BrdU-
markaketaren aldamoldea arras desberdina izan
zen, eguneko piko bakarra ageri baitzen eta BrdU-
markaketa balioak ez baitziren aldatu itsasgoran
zehar, egunez ezta gauez ere (4. Ird.). Udan, liseri-
guruineko ‰17.25-a BrdU-positiboak izan ziren.
BrdU-markaketa, eguneko aire-esposizioan
emendatzen zen (10:00 eta 12:00-ak bitartean), eta
ia inolako aldaketarik gabe mantentzen zen aire-
esposizioko ilunabarreko ordua heldu arte (4A Ird.).
Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
127
EGUNEKO PULTSUA HELDUAK GAZTEAK
Airean Uretan Guztira AIREAN Uretan Guztira
Eguna 1±1.414 1±0.89 2 2.4±2.074 6.5±5.45 8.9
Gaua 0.44±0.73 16.15±16.49 16.59 2.44±2.92 11.25±+9.02 13.69
Guztira 1.44 17.15 18.59 4.84 17.75 22.59
GAUEKO PULTSUA HELDUAK GAZTEAK
Airean Uretan Guztira Airean Uretan Guztira
Eguna 3.75±2.5 2.8±0.83 6.55 1.33±1.53 1±1.41 2.33
Gaua 2.42±2.71 4.75±3.24 7.17 5.91±5.90 7.9±5.51 13.81
Guztira 6.17 7.55 13.72 7.24 8.9 16.14
1. Taula: Atariko esperimentuan ikertutako talde experimentaletako liseri-albeolotan egunez eta gabez zein airean edo uretanmurgilduta egotean, muskuiluetan kuantifikatutako batezbesteko BrdU-markaketa + desbidazio estandarra.

Udazkenean, BrdU-markaketa maximoa (‰18.5),
goizeko aire-esposizioari (gaua bukaera eta
eguerdi bitartean zabaltzen zena) egokitu zitzaiona
(08:00), eta berehala, berriz ere hurrengo
murgiketa-aldia heldu aurretik, maila basaletara
bueltatzen zen (4B Ird.).
EZTABAIDA
Mareazpiko eta marearteko muskuiluak
alderatzeko BrdU-pultsuaren aukeraketa
Eguneko pultsu-esperimentuan, BrdU-
markaketa gaueko murgilketa-aldian behatu zen
muskuilu gazte zein helduetan, eguneko
murgilketa-aldian tarteko balioak behatuz. Muskuilu
gazteetan itsasaldi erregimena efektu esangarria
Emaitzak eta eztabaida
128
3. Irudia: Atariko esperimentuan ikertutako taldeexperimentaletako liseri-albeolotan egunez eta gabez zeinairean edo uretan murgilduta egotean, muskuiluetankuantifikatutako batezbesteko BrdU-markaketa. (A) Egunekopultsuko muskuilu helduak; (B) Gaueko pultsuko muskuiluhelduak; (C) Eguneko pultsuko muskuilu gazteak; (D) Gauekopultsuko muskuilu gazteak; (E) BrdU-gehipen jarraikoesperimentuan (Zaldibar et al., 2004) lortutako batezbestekoBrdU-markaketa. Segmentu bertikalek errore estandarraadierazten dute. Goiko asteriskoek taldeen artekodesberdintasun esangarriak adierazi dituzte p<0.05 mailan.

Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
129
EGUNEKO PULTSUA
Askatasun-graduak F p(F)
Itsasaldi-erregimena (IE)
1 4.091 0.05*
Argia/iluntasuna zikloa (AIZ)
1 3.348 0.08
IE x AIZ 1 4.266 0.048*
Helduak
Barne-errorea 482.5 Itsasaldi-erregimena (IE)
1 8.943 0.06
Argia/iluntasuna zikloa (AIZ)
1 1.234 0.28
IE x AIZ 1 1.189 0.29
Gazteak
Barne-errorea 862.92
GAUEKO PULTSUA
Askatasun-graduak F p(F)
Itsasaldi-erregimena (IE)
1 0.412 0.53
Argia/iluntasuna zikloa (AIZ)
1 0.087 0.77
IE x AIZ 1 2.347 0.14
Helduak
Barne-errorea 190.48 Itsasaldi-erregimena (IE)
1 0.507 0.48
Argia/iluntasuna zikloa (AIZ)
1 7.268 0.01*
IE x AIZ 1 0.323 0.58
Gazteak
Barne-errorea 620.46
PULTSU JARRAIA
Askatasun-graduak F p(F)
Itsasaldi-erregimena (IE)
1 12.614 0.001*
Argia/iluntasuna zikloa (AIZ)
1 3.433 0.07
IE x AIZ 1 2.399 0.13
Helduak
Barne-errorea 2159 MAREARTEKO MUSKUILUAK
Askatasun-graduak F p(F)
Itsasaldi-erregimena (IE)
1 2.115 0.15
Argia/iluntasuna zikloa (AIZ)
1 1.75 0.19
IE x AIZ 1 0.002 0.96
Uda
Barne-errorea 1452.07 Itsasaldi-erregimena (IE)
1 3.362 0.07
Argia/iluntasuna zikloa (AIZ)
1 0.684 0.41
IE x AIZ 1 2.283 0.13
Udazkena
Barne-errorea 1750.11
2. Taula: Atariko esperimentuan, esperimentu nagusian eta argitaratutako BrdU-pultsu jarraia (Zaldibar et al., 2004) izan dutenmuskuiluetan itsasaldi-erregimenak, argia/iluntasuna zikloak zein beraien arteko elkarrekintzak duten eragina finkatzeko egindakobi bideetako ANOVA-ren laburpena. p(F), Fisher ratioaren probabilitatea. Estatistikoki esangarriak gertatu ziren desberdintasunak(p<0.05) asterisko batez adierazita daude.

eragin zuen BrdU-markaketan; muskuilu helduetan,
ordea, itsasaldi-erregimena eta argi/iluntasun
zikloaren interakzioa esangarria izan zen.
Aldamolde hori, BrdU-pultsu jarraia jaso zuten
mareazpiko muskuilu handietan behatutakoaren
antzekoa izan zen (Zaldibar et al., 2004). Seguru
asko, 6 ordutako pultsuaren ostean suertatutako
BrdUren eskuragarritasunaren beherakadak,
etengabeko BrdUren gehipenaren kasuan (Zaldibar
et al., 2004) ez bezala, BrdUren pultsua eman eta
12 ordutara proliferazio zelularrean aldamolde
zirkamareala zergaitik desagertuz joan zen
azalduko luke. Bereziki muskuilu helduen kasuan,
zeintzuetan, BrdU-markaketa bortitzagoa izanik
(piko maximo altuagoak), BrdU-ren hornidura
mugatua gazteetan baino kritikoagoa suertatuko
litzatekeen. Baldintza horietan, esperimentu
honetan zeharreko BrdU-maila eskuragarriak
muskuiluentzat baxuak izatea litekeena da (Potten
et al., 1992, Candia Carnevali et al., 1997); hala
ere, BrdU kontzentrazio ez kaltegarriak erabili
beharrak alde batetik (ikusi Zaldibar et al., 2004),
eta esperimentu nagusi hauetan ikertu nahi
genituen marearteko muskuiluak uretan BrdU pean
mantentzeko denbora mugatuak bestetik, diseinu
esperimentalaren nondik norakoak agindu
zizkiguten.
Emaitzak eta eztabaida
130
4. Irudia: 2 egunez fotoperiodo(argitasun/iluntasun) eta erregimen mareal(airean/uretan) naturalean mantendutakomuskuiluen liseri-guruineko BrdUmarkaketa (‰ ± desbidazio estandarra)denboraren arabera (eguzki-denbora). (A)udako marearteko muskuilu handiak, (B)udazkeneko marearteko muskuilu handiak.Geziek BrdU pultsua eman zitzaien orduaadierazten du. Marra horizontalek,argiztapen-baldintzak (zuria, argitasun-tarteak; marraduna, ilunabar-tarteak;beltza, iluntasun-tarteak), eta erregimenmareala (uhinak, uretan murgilduta)adierazten dituzte. Erabilitako ikurrenforma eta trama desberdinek esangarrikidesberdinak diren azpitaldeak adierazidituzte, Duncan testaren arabera (p<0.05).

Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
131
Gaueko pultsu-esperimentuan behatutako
zelulen proliferazioaren aldamoldea desberdina
izan zen. BrdU-markaketa, soilik gauez emendatu
zen modu esangarrian, elkarren segidako bi piko
maximo emanez. Egunez (argialdia) aldiz, soilik
maila basalak neurtu ziren, baita BrdU-pultsua
eman eta ondorengo laginketetan ere. Izan ere,
BrdU-markaketa baliorik baxuenak pultsua eman
osteko orduetan izatea deigarria da; zeren eta
eguneko pultsu-esperimentuan BrdU-markaketa
maximoak BrdU-pultsuaren osteko lehen
momentuetan, erregistratu ziren. Ugaztunetan,
liseri-traktuko epitelioaren proliferazio zelularra
gauez gertatzearen berri eman dute zenbait
ikerlanek (Scheving et al., 1978; Smaaland, 1996).
Molusku itsastarretan, prozesu zelular batzuk
nagusiki gabez eta beste batzuk nagusiki egunez
gertatzen direla deskribatu den arren (Levy et al.,
1994), muskuiluetan liseri-zelulen proliferazioak
aldamolde zirkamareala jarraitzen duela
demostratu da (Zaldibar et al., 2004). Ildo horretatik,
emaitzok itsasaldia zelulen proliferazio-zikloa
gobernatzen duen faktore nagusia dela adierazi
duten arren, fotoperiodoak, neurri batean
behintzat, zelulen proliferazio-prozesuak modula
ditzakeela dirudi. Dena den, fotoperiodoaren
eragina BrdU-hornidura mugatuak sortutako
artefaktuaren ondorio soila ere izan liteke. Eguneko
pultsu-esperimentuan, IE x AIZ elkarrekintza
esangarria suertatu zeneko animalia helduetan,
piko maximoak handiagoak izan ziren eta
lehendabiziko argialdian zehar erregistratu ziren,
BrdU-ren pultsuaren osteko lehen laginketetan.
Ikerlan honetako eta baldintza esperimental
berdinetan burututako BrdU-pultsu jarraian
lortutako emaitzak alderatzeko, egunez zein gauez
GAZTEAK HELDUAK A B C D A B C D
MAREARTEKOA 5.8 11.6 14.04 70 4 8 9.68 100 NEGUA
MAREAZPIKOA 6.25 12.5 15.12 66 4.25 8.5 10.28 95
MAREARTEKOA 10 20 24.2 40 17.25 34.5 41.74 24 UDA MAREAZPIKOA 21 42 50.82 20 33 66 79.86 12
MAREARTEKOA 10.66 21.32 25.79 38 18.5 37 44.77 22 UDAZKENA
MAREAZPIKOA 2.33 4.66 5.64 175 2.4 4.8 5.808 170
3. Taula: Esperimentu nagusiaren talde esperimentaletako muskuiluen liseri-albeolotan kuantifikatutako batezbesteko BrdU-markaketaren laburpena. (A) Lehendabiziko 12 ordutan eskuratutako BrdU-markaketaren piko maximoa. (B) 24 ordutan S-faseansartuko liratekeen zelulak egunero 2 piko ematen direla kontutan hartuta. (C) BrdU-pultsu jarraiako esperimentuaren datuak(Zaldibar et al., 2004) kontutan hartuta eta zuzenketa-faktorea sartu eta gero, egunero S-fasean sartuko liratekeen zelula kopuruteorikoa. (D) Liseri-epitelioa osoa berriztatzeko beharrezkoa liratekeen egun kopurua..

lortutako murgilketa- eta aire esposizio-aldietako,
eta batezbesteko BrdU-markaketa balioak kalkulatu
ziren Zaldibar et al., (2004) BrdU pultsu jarraiko
esperimentuan lortutako datuetatik abiatuz eta
ikerlan honetan aplikatutako kalkuluetan oinarrituz
(1. eta 2. Taulak, 3E Ird.). Eguneko pultsu-
esperimentuko gazteetan gertatzen den moduan,
batezbesteko BrdU-markaketan soilik itsasaldi-
erregimenak eragiten du modu esangarrian.
Ondorioz, atariko bi esperimentuen emaitzak
kontuan hartuz, esperimentu nagusia burutzeko
eguneko pultsu-esperimentuaren prozedura
jarraitzea erabaki zen bi arrazoi nagusi hauek direla
eta: (a) eguneko pultsu-esperimentuan BrdU-
markaketa handiagoa erregistratu zen, eta (b)
eguneko pultsu-esperimentuan aurkitutako
aldamoldeak, aldez aurretik BrdU-gehipen jarraia
aplikatu zeneko esperimentuan aurkituriko
aldamoldearen (Zaldibar et al., 2004) antz gehiago
du.
Sasoiaren eragina epitelioko zelulen
berriztapenean
Zelulen proliferazio-tasen baliorik altuenak udan
gertatu ziren. Muskuiluen liseri-guruinean urteko
sasoiaren arabera aldaketak behatu dira, entzimen
jardueran (Cancio & Cajaraville, 1999; Le Pennec &
Le Pennec, 2002), lipido-edukietan (Pazos et al.,
2003), parametro peroxisomikoetan (Cancio et al.,
1999) zein liseri-zelulen lisosomen egituran
(Etxeberria et al., 1994). Tenperatura epelagoak eta
eurei lotutako egokitze termikoak zein elikagaien
presentzia areagotua, liseri-epitelioko dinamikan
oso garrantzitsuak izan daitezke, liseri-epitelioaren
aktibazioa emanez. Beraz, udako liseri-epitelioko
zelulen proliferazio-tasa areagotua, alga- eta
fitoplankton-bloom-ek sortarazitako elikagaien
eskuragarritasun handiagoarekin erlazionatuta
egon daiteke, muskuiluen hazkuntza somatikoaren
emendioa ekar dezakeena. Are gehiago, udako
jarduera metabolikoaren igoerak zelulak kaltetzea
eta zelulen berriztapena handitzea sortu izana ere
posible da, izan ere, neguko muskuiluekin alderatuz
udakoen ehunetan DNA-harizpi matxuratu- eta
proteinen desnaturalizazio-tasa altuagoak ikusi
baitira (Hofmann & Somero, 1996; Shaw et al.,
2000). Beraz, liseri-guruinean zelulen proliferazio
areagotua, epitelioaren berriztapen eta egituren
mantenuarekin zein hazkuntza somatikoarekin
erlazionatuta egongo litzateke. Era berean,
udazkenean eta neguko azken hilabeteetan,
tenperaturaren eta elikagaien eskuragarritasunaren
jaitsierek, eta honekin batera jarduera metaboliko,
kalte zelular eta zelulen berriztapenaren jaitsierek
ikerlan honetan behatutako proliferazio-tasa
txikituak azalduko lituzkete. Halaber, Japoniako
itsasoko Cremomytilus grayanus espeziearen liseri-
traktuan ere, Leibson & Frolova-k (1994) zelulen
proliferazio-tasaren urteko sasoiarekin
erlazionatutako aldaketa esangarriak topatu
zituzten, zelulen proliferazioaren piko maximoak
maiatza-ekaina aldean eta piko minimoak otsaila-
martxoan topatu zituztelarik. Autore hauek, zelulen
proliferazioaren jaitsiera, neguko hilabeteetako
tenperatura baxuek eragindako jarduera metaboliko
eta elikadura-jarduera gutxituei egotzi zieten.
Emaitzak eta eztabaida
132

Adinarekin erlazionatutako aldaketak epitelio-
zelulen berriztapenean
Aldez aurretik PCNA immunohistokimika erabiliz
muskuilu gazteen liseri-guruinak helduena baino
proliferazio-jarduera altuagoa zuela erakutsi arren
(Marigómez et al., 1999), ikerlan honetan ez da
halakorik behatu. Izatez, muskuilu gazte eta
helduek antzeko proliferazio-tasak erakutsi
zituzten. Are gehiago, udan, BrdU-markaketa/egun
indizea muskuilu helduetan gazteetan baino
altuagoa izan zen. PCNA eta BrdUren
sentikortasunak desberdinak izatea (Sarli et al.,
1995; Muskhelishvili et al., 2003), emaitza
eztabaidatsu hauek azaldu lituzke. Are gehiago,
PCNAren kasuan ez bezala, BrdU-markaketa,
pultsu osteko BrdUren eskuragarritasunaren
araberakoa izango litzateke, eta hori gazte eta
helduen artean desberdina izan liteke,
muskuiluetan adinarekin alda daitezkeen elikadura-
jarduera, asimilazio-eraginkortasun eta beste
prozesu fisiologikoen menpean egonik (Thompson
et al., 1974; Bayne et al., 1976). Beraz, uretango
pultsu berak ez du ziurtatu behar muskuilu gazte
eta helduek BrdUren kopuru bera eta momentu
berean jaso behar dutenik. Haatik, termino
kualitatiboetan behintzat, muskuilu gazte eta
helduek aldamolde patroi bera erakutsi zuten, liseri-
epiteliora heldutako BrdUren kopurua faktore
mugatzaile nagusietakoa ez zela izan iradoki
zuena. Are gehiago, PCNAren bidez lortutako
emaitzak immunoblotetan oinarritu ziren, ikerlan
honetan lortutako BrdUren emaitzek
immunohistokimika oinarri duten bitartean. Ezin
daiteke baztertu, PCNA immunohistokimika aplikatu
ondoren, Marigomez et al.-ek, (1999) aurkitu
zituzten desberdintasunak, liseri-guruineko epitelio-
zelulen proliferazio-tasaren arteko
desberdintasunei baino gehiago, homogeneizatuan
zeuden beste zelula-mota batzuei egokitzea. BrdU
bidez lortutako emaitza, berriz, liseri-zelulen
jarduera proliferatzailearekin zuzenean
erlazionatuta egongo litzateke.
Itsasaldi-erregimenaren eragina epitelioko
zelulen berriztapenean
Marearteko muskuiluetan, aire-esposizio aldian
S-fase gehiago erregistratu ziren, baina uretan
murgildutakoan ez zen BrdU-markaketan
jeitsierarik behatu. Horrek, murgilketa zein aire-
esposizio aldietan, DNA erreplikapenak, aurrera
darraiela esan lezake, baina zatiketa zelularra
nagusia da animaliak airera esposatuak dauden
bitartean. Badirudi zelulen zatiketa ezingo zela inoiz
marea altuan gertatu jatearekin batera, DNAren
sintesia jatearekin batera eman litekeen bitartean.
Jatearen eta bestelako zenbait jarduera fisiologiko
garrantzitsuen arteko denboraren bereizketa
molusku itsastarren kasuan aski deskribaturik dago
(Susswein et al., 1983).
Bestalde, marearteko organismoek euren bide
metabolikoak egunero birritan aldatu beharrean
topa daitezke, hipoxiaren aldi luze zein laburrak
jasateren ondorioz. Hori, beherakada metabolikoen
bitartez lortzen da, bai energia lortzeko bide
anaerobikoak aktibatuz zein transkripzioaren
eraenketa edota proteinen fosforilazio itzulgarrien
bidezko entzima-jardueraren aldaketaz erdietsi
daitekeena (Greenway & Storey, 2000; Ton et al.,
Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
133

2003; David et al., 2005; Papandreou et al., 2005).
Oxigenoaren eskaera modu egokian ez
hornitzekotan, zelulen kopurua ere txikitu beharra
dago, beraz, hipoxiarekiko erantzuna, ziklo
zelularraren gelditze edo eta apoptosiaren indukzio
gisa deskriba daiteke (Ton et al., 2003; Papandreou
et al., 2005), ornodunetan hipoxiak induzitutako
faktorea (HIF-1) izeneko transkripzioaren
suspertzaile baten kontrolpean suertatzen dena
(Gracey et al., 2001, Ton et al., 2003; Papandreou
et al., 2005). Airean egoteak eta ondorengo
hipoxiak, edo uretan murgiltzearen ondorengo
beroxigenatzeak, DNAren erreplikapen eta prozesu
mitotikoen geldiaraztea sor izana aintzat har
genezake, baina, dirudienez hori ez da ikerlan
honetan behatu dena. Bi prozesuok, zelulei BrdU
berriaren eskuratzearen gutxitzea lekarkieke eta
baita uretan murgilduta egon bitartean S-faseari
ekin zioten zeluletako BrdU-markaketa metatzea
ekarri, eta hori ikerlan honetan ez da behatu. Ildo
beretik, eta HIF-1 tankerako proteina bat
Crassostrea virginica marearteko moluskuan
identifikatu den arren (NCBI, CD648099), hipoxia
larriaren ondorioz azpi eta gainadierazitako zenbait
gene Crassostrea gigas-en aztertu dituen ikerlan
berriago batek (David et al., 2005), ez du zelulen
proliferazio-prozesuan inolako eraenketaren berririk
eman, ezta areagotutako apoptosi-erantzunarena
ere. Edozein kasutan, ornodunetan gertatzen den
modu berean, moluskuen neuronetan ere, hipoxia-
baldintzek apoptosia piz dezaketela proposatu da
(Jonas et al., 2005; Papandreou et al., 2005).
Itsasbehera izan bitartean (6 ordu bakarrik) airean
egotea areagotutako apoptosi-erantzuna eragitea
ez da litekeena; baina, hori gertatzekotan, zelulen
berriztatze altuago hori, ikerlan honetan behatutako
marearteko indibiduoen liseri-guruineko zelulen
proliferazio gaitasun areagotuarekin orokorrean
erlaziona liteke. Edonola ere, marearteko
muskuiluak mareazpikoekin konparatu direnean
ehunetan areagotutako kalte-zelularra eta
proteinen desnaturaltzea behatu dira (Hofmann &
Somero, 1995).
Aerobiosi/anaerobiosi baldintzez gain, elikatze-
jarduera ere, muskuilu itsastarren fisiologia eragiten
duen faktore nagusienetarikoa da. Itsasaldi-
erregimenak, muskuiluen liseri-epitelioaren forma
eta funtzioan berebiziko eragina du, jarraiki
(mareazpiko muskuiluak) edo txandaka
(mareartekoak) jaten duten arabera liseri-epitelioak
modu desberdinean funtzionatzen baitu (Owen,
1972, Robinson et al. 1981; Izagirre, 2002). Hala
ere, itsasaldia eta fotoperiodoa estuki erlazionaturik
daude, bibalbioen elikatze-jarduerari dagokionez
behintzat. Animaliak, batez ere egunez elikatzen
dira, fotosintesia dela eta, algak ur-zutabean
nagusiki egunez agertzen baitira. Beraz, gaua
endekatutako epitelioa berrizta daitekeen unea
dugu. Mareateko muskuiluen kasuan, DNAren
bikoizpena edozein momentutan gerta daiteke,
baina mitosiak muskuiluak airera esposaturik
daudenean soilik suertatzen dira, jaten ez dauden
bitartean, hain zuzen. Are gehiago, liseri-guruineko
epitelioa dinamikoa da oso, eta berau osatzen
duten bi zelulen profila, kopurua zein edukia,
elikagai eta ingurumen baldintzen arabera ordu
gutxiren buruan bortizki alda daitezke (Thompson et
al., 1974; Soto & Marigómez, 1997; Syasina et al.,
Emaitzak eta eztabaida
134

1997). Ildo beretik, itsasbeheran elikatze-tasa eta
xurgapen-efizientzia emendatzen dira (Wong &
Cheung, 2001), eta liseri-entzima ugarien jarduerak
ur-azpian suspertzen dira (Fernandez-Reiriz et al.,
2001). Honela, elikatze-tasak eta entzima-jarduerak
doituz, xurgapena egonkor manten daiteke (Wong
& Cheung, 2001). Liseri-epitelioaren berriztapena
elikatze-ziklo bakoitzarekin batera burutzen dela
iradoki da, hau da, marearteko muskuiluetan
birritan egunero (Nelson & Morton, 1979). Horrela,
mareazpiko muskuiluetan liseri-zelulen
proliferazioak aldamolde zirkamareala darraien
bitartean (Zaldibar et al. 2004; ikerlan honetako
emaitzak), marearteko muskuiluetan, pairatu
beharreko baldintza mugatuagoak direla eta,
itsasaldiak eta fotoperiodoak liseri-zelulen
proliferazioa agindu lezakete, eta, ondorioz, BrdU-
markaketen emendioa egunean behin eta bortizki
gertatuko da.
Ondorioak
Talde esperimentalen arteko desberdintasun
nabarmenenak itsasaldi-erregimen eta urteko
sasoiaren aldaketekin antzeman dira. Azken finean,
ez dirudi adinak proliferazio-aldamoldean eragin
handiegirik duenik. Areagotutako jarduera
metabolikoa dela eta, uda hazkuntza somatiko
gehien edo zelulen berriztapen-tasa altuagoa duen
sasoia dela ondorioztatu dugu, muskuilu gazte zein
helduek antzeko proliferazio-aldamoldea erakutsiz.
Bestalde, itsasaldi-erregimen desberdina duten
muskuiluen artean desberdintasun handiak daude,
baina, oro har, liseri-epitelioan zelulen proliferazioa
liseriketa bukatutakoan gertatu behar da.
BIBLIOGRAFIA
Bayne BL, Widdows J, Thompson RJ (1976) Physiology I. 121-
206 orr. Non: Marine mussels, their ecology and
physiology. Bayne BL (ed). Cambridge University Press.
Cambridge.
Cancio I, Cajaraville MP (1999) Seasonal variation of xanthine
oxidoreductase activity in the digestive gland cells of the
mussel Mytilus galloprovincialis: A biochemical,
histochemical and immunochemical study. Biol. Cell 91:
605-615.
Cancio I, Ibabe A, Cajaraville MP (1999) Seasonal variation of
peroxisomal enzyme activities and peroxisomal structure in
mussels Mytilus galloprovincialis and its relationship with
lipid content. Comp. Biochem. Physiol. C 123:135-144.
Candia Carnevali MD, Bonasoro F, Biale A (1997) Pattern of
bromodeoxyuridine incorporation in the advanced stages
of arm regeneration in the feather star Antedon
mediterranea. Cell Tiss. Res. 289:363-374.
David E, Tanguy A, Pichavant K, Moraga D (2005) Response of
the Pacific oyster Crassostrea gigas to hypoxia exposure
under experimental conditions. FEBS J. 272:5635-5662.
Etxeberria M, Sastre I, Cajaraville MP, Marigómez I (1994)
Digestive lysosome enlargement induced by experimental
exposure to metals (Cu, Cd and Zn) in mussels collected
from a zinc polluted site. Arch. Environ. Toxicol. 27:338-
345.
Fernandez-Reiriz MJ, Labarta U, Navarro JM, Velasco A (2001)
Enzymatic digestive activity in Mytilus chilensis (Hupe
1854) in response to food regimes and past feeding history.
J. Comp. Physiol. 171:449-456.
Gracey AY, Troll JV, Somero GN (2001) Hypoxia-induced gene
expression profiling in the euryoxic fish Gillichthys
mirabilis. Proc. Nat. Acad. Sci. U.S.A. 98:1993-1998.
Greenway SC, Storey KB (2000) Seasonal change and
prolonged anoxia affect the kinetic properties of
phosphofructokinase and pyruvate kinase in oysters.
Comp. Biochem. Physiol. B 170:285-293.
Hofmann GE, Somero GN (1996) Interspecific variation in
thermal denaturation of proteins in the congeneric mussels
Mytilus trossulus and M. galloprovincialis: Evidence from
the heat-shock response and protein ubiquitination. Mar.
Biol. 126:65-75
Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
135

Izagirre U (2002) Itsasaldien, sasoien, elikadura-erregimenen
eragina liseri-zelulen lisosometan eta euren
kutsatzaileekiko erantzunetan. Lizentziatura-Tesia. 78 orr.
Jonas EA, Hickman JA, Hardwick JM, Kaczmarek LK (2005)
Exposure to hypoxia rapidly induces mitochondrial channel
activity within a living synapse. J. Biol. Chem. 280:4491-
4497.
Leibson, N.L & Frolova, L.T. 1994. Winter-spring essential
reorganization of cell proliferation in the digestive tract
epithelia in the mussel Crenomytilus grayanus. Mar. Biol.
118:471-477.
Le Pennec, G & Le Pennec, M. 2002. Molecular analysis of the
seasonal expression of genes coding for different
functional markers of digestive gland of the bibalve mollusk
Pecten maximus (L). Comp. Biochem. Physiol B 133:417-
426.
Levy M, Weller A, Susswein AJ (1994)Learned changes in the
rate of respiratory pumping in Aplysia fasciata in response
to increases and decreases in seawater concentration.
Behav. Neurosci. 108:161-170.
Marigómez I, Lekube X, Cancio I (1999) Immunochemical
localisation of proliferating cells in mussel digestive gland
tissue. Histochem. J. 31:781-788.
Muskhelishvili L, Latendresse JR, Kodell RL, Henderson EB
(2003) Evaluation of cell proliferation in rat tissues with
BrdU, PCNA, Ki-67(MIB-5) immunohistochemistry and in
situ hybridization for histone mRNA. J. Histochem.
Cytochem. 51:1681-1688.
Nelson L, Morton JE (1979) Cyclic activity and epithelial
renewal in the digestive gland tubules of the marine
prosobranch Maoricrypta monoxyla (Lesson). J. Moll. Stud.
45:262-283.
Okudela K, Ito T, Kameda Y, Nakamura N Kitamura H (1999)
Immunohistochemical analysis for cell proliferation-related
protein expression in small cell carcinoma of the
esophagus; a comparative study with small cell carcinoma
of the lung and squamous cell carcinoma of the
esophagus. Histol. Histopathol. 14:479-485.
Owen G (1972) Lysosomes, peroxisomes and bivalves. Sci.
Prog. Ser. Oxf. 60:229-318.
Papandreou I, Powel A, Lim AL, Denko N (2005) Cellular
reaction to hypoxia: sensing and responding to an adverse
environment. Mut. Res. 569: 87-100.
Pazos AJ, Sánchez JL, Román G, Pérez-Parallé ML, Abad M
(2003) Seasonal change in lipid classes and fatty acid
composition in the digestive gland of Pecten maximus.
Comp. Biochem. Physiol. B 134: 367-380.
Potten CS, Kellet M, Rew DA Roberts SA. (1992) Proliferation
in human gastrointestinal epithelium using
bromodeoxyuridine in vivo: data for different sites,
proximity to a tumour, and polyposis coli. Gut 33:524-529.
Robinson WE, Pennington MR, Langton RW (1981) Variability
of tubule type within the digestive glands of Mercenaria
mercenaria L., Ostrea edulis L., Mytilus edulis L. J. Exp.
Mar. Biol. Ecol. 54:265-276.
Sarli G, Benazzi C, Preziosi R, Marcato PS (1995) Assessment
of proliferative activity by anti-PCNA monoclonal antibodies
in formalin-fixed, paraffin-embedded samples and correla-
tion with mitotic index. Vet. Pathol. 32:93-96.
Scheving LE, Burns ER, Pauly JE, Tsai T (1978) Circadian
variation in cell division of the mouse alimentary tract, bone
marrow and corneal epithelium. Anat. Rec. 191:479-486.
Smaaland R. 1996. Circadian rhythm of cell division. Prog. Cell
Cycle Res. 2:241-266.
Shaw JP, Large AT, Livingstone DR, Doyotte A, Renger J,
Chipman JK, Peters LD (2000) Elevation of cytochrome
P450-immunopositive protein and DNA damage in mussels
(Mytilus edulis) transplanted to a contaminated site. Mar.
Environ. Res. 54:505-509.
Soto M, Marigómez I (1997) Metal bioavailability assessment in
“mussel-watch” programmes by automated image analysis
of autometallographical black silver deposits (BSD) in
digestive cell lysosomes. Mar. Ecol. Prog. Ser. 156:141-
150.
Susswein AJ, Gev S, Feldman E, Markovich S (1983) Activity
patterns and time budgeting of Aplysia fasciata under field
and laboratory conditions. Behav. Neural Biol. 39:203-220.
Syasina IG, Vaschenko MA, Zhandan PM (1997) Morphological
alterations in the digestive diverticula of Mizuhipecten
yessoensis (Bivalvia: Pectenidae) from polluted areas of
Peter the Great Bay, Sea of Japan. Mar. Environ. Res.
44:85-98.
Thompson RJ, Ratcliffe NA, Bayne BL (1974) Effects of
starvation on structure and function in the digestive gland
of the mussel (Mytilus edulis, L.). J. Mar. Biol. Assess. U.
K. 54:699-712.
Ton C, Stamatiou D, Liew CC (2003) Gene expression profile of
zebrafish exposed to hypoxia during development. Physiol.
Genom. 13:97-106.
Emaitzak eta eztabaida
136

Sasoi,tamaina eta erregimen marealarekin erlazionatutako berriztapen zelularra
137
Wong WH, Cheung SG (2001) Feeding rhythms of the green-
lipped mussel, Perna viridis (Linnaeus, 1758) (Bivalvia:
Mytilidae) during spring and neap tidal cycles. J. Exp. Mar.
Biol. Ecol. 257:13-36.
Zaldibar B, Cancio I, Marigómez I (2004) Circatidal variation in
epithelial cell proliferation in the mussel digestive gland
and stomach. Cell Tiss. Res. 318:395-402.

Emaitzak eta eztabaida
138

Zelula-moten ordezkapen itzulgarria kadmiopean jarritako Littorina littorea moluskuitsastarraren liseri-guruinean
Laburpena: Littorina littorea magurio itsastarren zelula basofilikoen proliferazioa eragiteko 1.25 mg Cd/lpean jarri ziren 21 egunez. Ondoren, magurioak itsas-ur garbitan 10 egunez arazten utzi ziren, zelula-moten ordezkapena itzulgarria zen antzemateko. Liseri-guruinak, Carnoy fixatzailean fixatu eta analisihistologikoak egiteko parafinan inkluditu ziren. Zelula basofilikoen (VvBAS) eta liseri-zelulen (VvDIG)
dentsitate bolumetrikoa kuantifikatzeko hematoxilina eosinaz tindatutako ebakien gainean analisiestereologikoak burutu ziren. Zelulen tamaina eta kopuru absolutua estimatu ondoren, behatutakoaldaketek liseri-zelulen galerari eta aldi bereko zelula basofilikoen hipertrofiari egotzi zitzaien, eta ez zelulabasofilikoen kopuruaren emendioari. Hamar egunetako arazketa-aldiaren ondoren zelula-moten osaketaeta tamaina kasik balio normaletara itzuli ziren. PCNA immunohistokimikak, Cd pean egondako etaarazketa taldeko magurioen liseri-epitelioan kontrolean baino liseri-zelula proliferatzaile gehiago zeudelaerakutsi zuen. Hau da, liseri-zelulen galera netoak liseri-zelulen proliferazioaren areagotzearekin batzetorrela iradoki du, zelula basofilikoen ekarpenak zerikusirik ez zuen bitartean. Beraz, Cd pean egoteak,Cd-aren detoxifikazioa burutzeko liseri-zelulen berriztapena areagotuko zuela dirudi. Autometalografia(AMG) bitartez errebelatzen diren zilarrezko hauspeakin beltzak (BSD; ingeleraz, black silver deposits),nagusiki Cd ioien inguruan eratuak, liseri-zelulen lisosometan agertu ziren, zelula basofilikoetan BSDenpresentziarik behatu ez zen bitartean. Arazketa-aldia burutu eta gero BSD ez ziren horren nabarmenakizan. Liseri-zelulen lisosometan gertatutako aldaketak kuantifikatzeko, kriostatoko ebakien gainean ß-glukuronidasa jardueraren erakusketa histokimikoa gauzatu zen. Gerora, eta irudi-analisi automatizatuarenlaguntzaz liseri-lisosometan gertatutako aldaketak neurtu ziren. Cd pean mantendutako magurioetanhandiagotutako lisosomak behatu ziren, arazketa-aldiaren hamar egunak eta gero berriro ere kontrolenbalioetara itzuliz. Liseri-zelulen proliferazioan ikusitako aldaketek, liseri-zelulen galerak eta zelulabasofilikoen hipertrofiak, ez zuten ikerlan honetan erabilitako esposiziozko (BSD) eta efektuzko (erantzunlisosomikoak) biomarkatzaileen gainean eragin nabarmenik eduki.
Gako-hitzak: zelula-moten ordezkapena, magurioa, kadmioa, PCNA, liseri-guruina, efektu- etaesposizio-biomarkatzaileak.
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
139

Reversible cell-type replacement in the digestive gland epithelium of themarine mollusc Littorina littorea exposed to cadmium
Abstract: Marine winkles Littorina littorea were exposed to 1.25 mg/l Cd for 21 days to provoke theproliferation of basophilic cells. Then, animals were depurated in clean seawater for 10 days to determinewhether CTR was reversible. Digestive glands were fixed in Carnoy and paraffin embedded for histologicalanalysis. The volume densities of basophilic cells (VvBAS) and digestive cells (VvDIG) were calculated by
stereology on haematoxylin-eosin stained sections. VvBAS raised and VvDIG decreased in Cd exposed
animals. After estimation of cell size and absolute cell numbers, these changes were attributed to digestivecell loss and concomitant basophilic cell hypertrophy but not to increased numbers of basophilic cells. Thecell type composition and cell size almost fully returned to normal values after 10 days depuration.Accordingly, PCNA immunohistochemistry demonstrated that proliferating digestive cells were moreabundant in winkles exposed to Cd and after 10 day depuration than in control specimens, suggesting thatnet digestive cell loss was accompanied by increased digestive cell proliferation and that CTR was notrelated to any contribution by basophilic cells. Thus, Cd-exposure seems to provoke an enhanced digestivecell turnover in order to cope with Cd detoxification. Autometallography revealed the presence of black silverdeposits (BSD), most likely formed around Cd ions, in digestive cell lysosomes whereas basophilic cellsappeared devoid of them. After depuration, BSD were less conspicuous. Changes occurring in digestive celllysosomes were quantified by automatized image analysis on cryostat sections stained to demonstrate ß-glucuronidase activity. Enlarged lysosomes were observed in Cd-exposed winkles while return to controllevels ocurred after 10 days depuration. Changes in digestive cell proliferation, digestive cell loss andbasophilic cell hypertrophy did not apparantly affect the exposure (BSD) and effect (lysosomal responses)biomarkers investigated herein.
Key Words cell-type replacement, winkle, cadmium, PCNA, digestive gland, effect and exposurebiomarkers.
Emaitzak eta eztabaida
140

Sustitución reversible de tipos celulares en el epitelio de la glánduladigestiva del molusco marino Littorina littorea expuesto a cadmio
Resumen Bígaros marinos Littorina littorea fueron expuestos a 1.25 mg/l de Cd durante 21 días conel fín provocar la proliferación de células basofílicas. Posteriormente, los animales se depuraron por unperíodo de 10 días en agua de mar limpia con el fin de observar si el recambio de tipos celulares erareversible. Las glándulas digestivas se fijaron en Carnoy fueron incluídas en parafina. La densidadvolumétrica de células basofílicas (VvBAS) y células digestivas (VvDIG) se calculó mediante estereología
en muestras teñidas con hematoxilina-eosina. En los bígaros expuestos a Cd se observó un aumento enlos valores VvBAS y un descenso en los valores VvDIG. Después de estimar el tamaño y el número
absoluto de células, éstos cambios se atribuyeron a la pérdida de células digestivas y a la hipertrofiaconcomitante de las células basofílicas, y no al aumento de éstas. Después del período de 10 días dedepuración, la composición de tipos celulares y el tamaño de las células volvieron a valores cercanos a losnormales. La inmunohistoquímica de PCNA demostró que la proliferación de células digestivas era mayoren los bígaros expuestos a Cd y después de 10 días de depuración que en los bígaros control, sugiriendoque la pérdida neta de células digestivas viene acompañada por el aumento en la proliferación de célulasdigestivas y no tiene relación alguna con la contribución de las células basofílicas. Por lo tanto, laexposición a Cd parece provocar una renovación aumentada de células digestivas. Los depósitos negrosde plata (BSD; del inglés, black silver deposits) mayormente formados alrededor de los iones de Cd yrevelados mediante autometalografía se localizaron en los lisosomas de las células digestivas, mientrasque no se detectaron en las células basofílicas. En el grupo de los bígaros depurados los depósitos fueronmenos llamativos. Se midieron los cambios ocurridos en los lisosomas de las células digestivas medianteanálisis de imagen sobre muestras de criostato, previamente teñidas para la demostración de la enzima ß-glucuronidasa. Se observó un mayor tamaño de lisosomas en los bígaros expuesto a Cd, que volvieron alos valores controles después de los 10 días de depuración. Los cambios en la proliferación de célulasdigestivas, la pérdida de células digestivas y la hipertrofia de células basofílicas no tuvieron efecto sobrelos biomarcadores de exposición (BSD) y efecto (respuestas lisosómicas) empleados en este trabajo.
Palabras clave: sustitución de tipos celulares, bígaro, cadmio, PCNA, glándula digestiva, biomarcadoresde efecto y exposición.
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
141

Emaitzak eta eztabaida
142

Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
143
SARRERA
Moluskuen liseri-guruina ingurumen-toxikologia
ikerketetarako itu-organoa da; izan ere
,kutsatzaileen metaketa eta detoxifikazioan parte
hartzen du (Marigómez et al., 2002). Orokorrean,
molusku gehienen liseri-guruina antzeko
antolaketa-eredua aurkezten du; hots, liseri-
dibertikulua epitelio bakun batez osaturik dago eta
bertan bi zelula-mota agertzen dira, liseri-zelula eta
zelula basofilikoa, alegia (Morton, 1983). Liseri-
zelulek, elikagaien liseriketa intrazelularrean parte
hartzen dute, eta oso sistema endo-lisosomiko
garatua dute; zelula basofilikoak, bestalde, zelula
jariatzaileak dira, eta gastropodotan pikor
mineralizatu zitoplasmatikoak izan ditzakete
(Mason, 1983). Baldintza fisiologiko normaletan,
liseri-zelulak zelula basofilikoak baino ugariagoak
dira, baina estres-egoera desbedinetan, zelula
basofilikoen proportzio erlatiboa emendatzen da,
fenomeno hau zelula-moten ordezkapena (CTR;
ingeleraz, cell type replacement) izenarekin
ezaguna delarik (Cajaraville et al., 1990a;
Marigómez et al., 1990). Zelula-moten
ordezkapena, kutsatzaileen eragin pean egondako
taxon desberdineko eta habitat desberdineko
moluskuetan ematen den fenomeno orokorra da
(Widdows et al., 1984; Lowe & Clarke, 1989;
Cajaraville et al., 1990a; 1990b; Marigómez et al.,
1990; 1996). Kutsatzaile organiko (Cajaraville et
al., 1990a) zein metaliko (Soto & Marigómez,
1997a; 1997b) desberdinen kontzentrazio
subletalen pean jarritako bibalbioek, nagusiki zelula
basofilikoz osatutako liseri-guruina aurkezten dute.
Modu berean, laborategian Cd pean jarritako
gastropodo itsastarrek, zelula basofiliko
hipertrofiatuz osatutako liseri-tubuluak aurkezten
dituzte (Marigómez et al., 1990). Zelula
basofilikoen kopuru erlatiboaren igoera esangarria
hidrokarburo aromatikoen pean jarritako
gastropodo itsastarretan ere behatu da (Cajaraville
et al., 1990b).
Zelula-moten ordezkapena, liseri-zelulen
kopuruaren murrizpenarekin zuzenean edo, zelula
basofilikoen proliferazioarekin erlazionatuta
dagoen erantzun beharra dago. Oraintsu, gure
taldeak muskuiluen liseri-guruineko liseri-zelulak
zein zelula jariatzaileak PCNA eta BrdUren
immunohistokimikarekiko erreaktiboak direla
erakutsi du (Marigómez et al., 1999, Zaldibar et al.,
2004). Beraz, zelula-mota bakoitzak ziklo zelularra
bere aldetik burutzen du . Bestalde, 1.25 mg Cd/l
itsas-ur kotzentrazioak, magurioen liseri-guruineko
zelula basofilikoen proportzio erlatiboen igoera
esangarria sorterazten duela deskribatu da
(Marigómez et al., 1990). Ekarpen horiek kontuan
hartuta, ikerlan honen helburua, zelula-moten
ordezkapena prozesu itzulgarria ote den, eta
zelula basofilikoen proliferazioa edo liseri-zelulen
galera dela medio ematen ote den argitzea da.
Gainera, prozesua itzulgarria izatekotan zelula-
moten ratio normalera itzultzea liseri-zelulen
proliferazioaren areagotze batek eragin ote duen
finkatu behar da. Helburu horiek betetzeko,
magurioak 1.25 mg Cd/l itsas-ur kontzentrazio
pean jarri ziren 3 astez, baldintza esperimental
horietan zelula-moten ordezkapena sortaraz
zitekeela bagenekielako (Marigómez et al., 1990),

eta, ondoren, 10 egunez itsas-ur garbitan eduki
ziren; izan ere, aldez aurretik arazketak zelula
basofilikoen kopuruan jaitsiera eragin dezakeela
ikusi da.
Zelula-moten ordezkapen-prozesuak hainbat
emaitza biokimiko eta bioenergetiko eztabaidatsu
azal ditzake, uretan eta muskuilu itsastarren ehun
bigunetan neurtutako metal-kontzentrazioak
erlazionatzerakoan behatutako emaitza
eztabaidakorrak, besteak beste. Marigómez et al.
(1996, 1998), Soto et al. (1998a) eta Soto &
Marigómez-ek (1997b) esposizio luze edo
kronikoen ondorioz liseri-epitelioetan metalen
metaketarik ematen ez zela aurkitu zuten; izan ere,
epitelio horietan ez da apenas liseri-zelularik
ikusten, eta euren lisosomak metaleen metaketa-
toki nagusia dira. Zelula-espezifiko diren hainbat
biomarkatzaile (parametro lisosomikoak,
peroxisomen proliferazioa...) erabiltzen direnean
lortutako emaitzak interpretatzerakoan, antzeko
ondorioak lortu daitezke. Kutsatzaileen eragin pean
jartzeak liseri-zeluletako lisosomen tamainaren
handipena eragiten du normalean (Cajaraville et al.,
1995a; Etxeberria et al., 1995; Marigómez et al.,
2005a), baina zelai ikerketetan denbora tarte
laburrez kutsatzaile organikoen pean jarritako
animalietan lisosomen tamainaren txikipena
deskribatu da baita (Marigómez & Baybay-
Villacorta, 2003). Metaketa-gaitasuna eta
biomarkatzaileen erantzun batzuk zelula-mota
espezifikoak direnez, zelula-moten osaketan
ematen diren aldaketak, biometatze eta
biomarkatzaileen ikerketetan, ustekabeko emaitza
gisa ager daitezkeela susma daiteke. Testuinguru
honetan, ikerketa honen bigarren helburua, zelula-
moten ordezkapenak, metal-esposizio eta efektu
biologikozko biomarkatzaile ezagunak diren AMGz
lortutako zilarrezko hauspeakin beltzen (BSD;
ingeleraz, black silver deposits) hedapenean eta
lisosomen tamaina aldaketan eraginik ote duen
ikustea da (Cajaraville et al., 1995a; Marigómez et
al., 2002).
MATERIAL ETA METODOAK
Erreaktibo kimikoak
Erabilitako erreaktibo kimiko guztiak maila
analitikokoak eta Sigma-Aldrich (St. Louis,
Missouri, Amerikako Estatu Batuak), merkatal-
etxekoak dira besterik agertu ezean.
Prozedura esperimentala
2.5-3.5 zm altuera kolumelarreko Littorina
littorea magurioak merkatu batean eskuratu ziren
2001. eko urtarrilean eta litro bateko akuarioetan
mantendu ziren moldatzen 17ºC-tan 7 egunez.
Girora moldatzeko denbora igaro ondoren,
magurioak 1.25 mg Cd/l-pean jarri ziren 21 egunez.
Kontzentrazio subletal horrek liseri-guruineko
epitelioko zelula basofilikoen proportzio
erlatiboaren igoera esangarria sortarazten duela
aldez aurretik egindako esperimentuetan
deskribatu da (Marigómez et al., 1990). Esposizio-
denbora bitartean, magurioak aldez aurretik
izoztutako Fucus vesiculosus algaz “ad libitum”
elikatu ziren. Janaria eta ura 2 egunero aldatu ziren.
Kadmio peko 21 egunak igaro ondoren, talde
esperimental bakoitzeko 10 magurio hartu ziren, eta
Emaitzak eta eztabaida
144

Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
145
liseri-guruina/gonada konplexua disekzionatu zen
gero deskribatuko den bezala. Gainontzeko
magurioak, itsas-ur garbitan mantendu ziren 10
egunez, arazteko asmoz, eta ondoren aurreko
magurioen modu berean prozesatu ziren.
Laginen prozesaketa
Liseri-guruina/gonada konplexuaren zati txikiak
Carnoy fixatzailean (%60 etanol absolutua, %30
kloroformoa eta %10 azido azetikoa) fixatu ziren
ordu batez 4°C-tan. Fixatu ondoren, laginak
goranzko graduazioko etanolezko bainu-serie
batean deshidratatu ziren eta parafinan inkluditu
ziren 60°C-tan. Ebakiak (7 µm-ko lodiera)
mikrotomoan ebaki ziren. Ehunaren beste zati txiki
bat, %10 sakarosa zuen 0.1 M fosfato
indargetzailean (pH 7.4) murgildu zen 10 minutuz
4ºC-tan. Ondoren, gehiegizko indargetzailea
iragazi-papera erabiliz lehortu, eta Cryo-M-Bed
(TAAB Laboratories Equipments Ltd., Aldermaston,
Ingalaterra) konposatuan inkluditu zen, amaieran
laginak nitrogeno likidotan izoztu ziren. Ehunaren
hirugarren zati bat, zati txikitan xehetu zen (<1
mm3) eta %2.5 glutaraldehido zuen kakodilato
indargetzailean (CB; 0.1 M; pH 7.2) fixatu zen 2
orduz 4ºC-tan. Ondoren, laginak %2 osmio
tetroxidoa eta %3 potasio ferrozianuroa zuen
CBan, 1:1 diluzioan, postfixatu ziren ordu batez
4ºC-tan. Laginak ur distilatuan garbitu ziren (3x10
minutu), goranzko graduazioko etanolezko bainu-
serie batean deshidratatu ziren, eta EPON 812
erretxinan inkluditu ziren. Ebaki semifinak Leica
Utracut ultramikrotomo baten bitartez eskuratu
ziren, eta toluidina urdinez tindatu ziren, argi
mikroskopioan behaketak egiteko.
Zelula basofilikoen tamainaren estimazio
morfometrikoa
Parafinan inkluditutako ebakiak (7 µm)
hematoxilina-eosinaz tindatu ziren liseri-zelulak
(eosinofiloak) eta zelula basofilikoak (ilunak)
bereizteko. Zelula basofilikoen tamainaren
estimazio zakar bat lortzeko Olympus BX50
mikroskopio (Olympus, Tokio, Japonia) (amaierako
handipena x540) bati atxiki zegoen irudi-analisi
baten bitartez (Visilog 5.4 softwarea, Noesis,
Crolles, Frantzia) tratamendu bakoitzeko
aleatorioki aukeratutako 20 zelula basofilikoen
soslaia eskuratu zen. Neurketak egiteko soilik
epitelioaren xafla basaletik (zentzu histologikoan)
lumenera argiro zabaltzen ziren soslai
triangeluarreko zelula basofilikoak aukeratu ziren.
Soslai bakoitzerako ebakiduraren azalera (As) eta
luzeera maximoa (Lm) neurtu zen eta ondoren
zelula basofilikoak konikoak zirela onartuz (Mason,
1983) beraien tamaina (SBAS) hurrengo oinarrizko
formula geometrikoen arabera kalkulatu zen. SBAS= (π x As2)/(3 x Lm), µm3-tan. Horrela As = (B x
Lm)/2; r = B/2; Vkonoa =(π/ 3) x r2 x Lm = (π/3) x
(As/Lm)2 x Lm, non B soslai triangeluarraren
basearen luzera den, eta r luzera maximoaren
ardatzarekiko elkartzuta den konoaren erradio
maximoa den.
Epitelioaren konposizio zelularraren analisi
estereologikoak
Hematoxilina-eosinaz tindatutako lagin

berberak erabiliz (7 µm), liseri-zelulen (VvDIG) eta
zelula basofilikoen (VvBAS) dentsitate
bolumetrikoak kuantifikatzeko prozedura
estereologikoa aplikatu zen. Horretarako
kontaketak lagin bakoitzean azarez aukeratutako
zelai batetan egin ziren kontaketak guztira talde
esperimental bakoitzeko 10 kontaketak burutuz.
Analisi estereologikoak burutzeko Nikon Optiphot
mikroskopio bati atxiki zegoen irudi-hodi bat erabili
zen (amaierako handipena x670). VvDIG eta
VvBAS kalkulatzeko Weibel gratikula
(multipurpouse test system M-168; Weibel, 1979)
erabili zen eta liseri-zelula zein zelula basofilikotan
eroritako puntuak kuantifikatu ziren eta kalkuluak
Delesse-ren printzipioen arabera gauzatu ziren
(Weibel, 1979).
Liseri-zelula eta zelula basofilikoen
bolumenaren estimazio absolutuak dentsitate
bolumetrikoen datuetan oinarrituz burutu ziren.
Liseri-zelulek eta zelula basofilikoek betetzen zuten
bolumena (VDIG = VvDIG x 17.8 x 106 µm3; VBAS= VvBAS x 17.8 x 106 µm3) arbitrarioki
aukeratutako liseri-guruinaren bolumen batekiko
kalkulatu zen (17.8 x 106 µm3 gutxi gorabehera).
Bolumen hori kalkulu estereologikoak egiteko
erabilitako Weibel gratikularen (356000 µm2)
arbitrarioki aukeratutako 50 µm-ko lodierako
ebakidura bati dagokio. Lodiera hori zelula
basofilikorik handiena guztiz barneratua egotea
ahalbidetzen du.
Ondoren, liseri-guruineko zati arbitrario horretan
liseri-zelula eta zelula basofilikoen kopurua
kalkulatzeko, liseri-zelulek tamaina konstantea
zutela onartu zen (talde esperimental
desberdinetan egindako behaketa mikroskopikoek
liseri-zelulen altueran aldaketa txikiak erakusten
dituzte, 30-33 µm, kalibrea konstante mantendu
zelarik, 6-8 µm), eta zelula basofilikoen tamainen
estimazio errealak aldez aurretik azaldu bezala egin
ziren.
PCNA immunohistokimika
Ebakiak (3-4 µm-ko lodiera) mikrotomo batean
lortu eta aldez aurretik (3-amino propil
trietoxisilanoz) silanizatutako portaobjektuetan jazo
ziren. Ondoren, laginak xilenoz desparafinatu ziren,
beheranzko graduazioko etanolezko bainu-serie
batean hidratatu eta fosfato indargetzaile salinoz
(PBS; ingeleraz, phosphate-buffered saline) garbitu
ziren. Peroxidasa jarduera endogenoa ekiditeko,
ebakiak %3 zuen H2O2an inkubatu ziren 5 minutuz,
eta berriro PBS-az garbitu ziren. Ebakiak,
Vectastain Elite ABC Kit-ak (Vector Laboratories,
Burlingame, Kalifornia, Amerikako Estatu Batuak)
hornitutako suero blokeatzailean 30 minutuz
inkubatu ziren giro-tenperaturan, eta berriro ere
PBS-an garbitu ziren. Gero, ebakiak PCNAren
aurkako antigorputz espezifikoan (PC10, 1:1500
PBSan) inkubatu ziren. Laginak PBS-az garbitu
ondoren, Vectastain Elite ABC Kit-aren bitartez eta
abidina-biotina entzima-konplexua erabiliz
antigorputza ikustarazi zen. Laburki, ebakiak, PBS-
an diluitutako saguaren aurkako zaldiaren IgG
antigorputz sekundario biotinilatuan inkubatu ziren,
30 minutuz, eta PBS-an garbitu. Peroxidasa
jarduera 3,3´-diaminobenzidina tetrahidrokloridoa
(DAB) eta %0.03 H2O2 zuen PBS indargetzailea
erabiliz ikustarazi zen. Azkenik, ebakiak
Emaitzak eta eztabaida
146

hematoxilinaz kontrastatu ziren (10 s), dabilen
uretan garbitu eta Kaiser gelatina glizerinatuan
(Merck & Co., Inc, Whitehouse Station, New Jersey,
Amerikako Estatu Batuak) montatu ziren.
Mikrografiak, Olympus BX50 mikroskopio batetara
atxiki zegoen Olympus PM20 argazki-kamera batez
erdietsi ziren.
Autometalografia
Metalak, Danscher-ek (1984) deskribatutako eta
Soto et al.-ek (1998b) modifikatutako prozedura
jarraituz determinatu ziren parafinan inkluditutako
laginetan. Ebakiak (7 µm), laburki xilenoz
desparafinatu ziren, beheranzko graduazioko
etanolezko bainu-serie batean hidratatu eta labean
(37ºC) utzi ziren guztiz lehortu arte. Laginak
ondoren, argazki-emultsio geruza fin eta uniforme
batez estali ziren gela ilunean (Ilford Nuclear
Emulsion L4, Ilford, Mobberley, Ingalaterra).
Laginak ilunpean mantendu ziren lehortu bitartean
(30 minutu) eta laginak Ultrafin Tetenal SF (1:5
diluzioa ur destilatuan) (Tetenal AG & Co,
Norderstedt, Alemania) errebelatzaile komertziala
erabiliz errebelatu ziren 15 minutuz. Ondoren,
errebelatzailearen erreakzioa %1 azido azetikoa
zuen diluzio batean geldiarazi zen minutu batez, eta
azkenik erreakzioa fixatzailean (Agfa Agefix, Agfa-
Gevaert N.V., Mortsel, Belgika, 1:9 uretan) fixatu
zen 10 minutuz. Hauspeakin beltz (BSD) moduan
agertutako metalak irudi-analisi baten bitartez
(Biological Measurement System software BMS-
Sevisan, Bilbo, Espainia.) kuantifikatu ziren.
Horrela, BSDen dentsitate bolumetrikoa (VvBSD)
kalkulatu zen: VvBSD = VBSD/VEH non VBSD
BSDen bolumena den eta VEH ehunaren
bolumena.
Lisosomen estereologia
ß-glukuronidasa entzimaren erreakzio
histokimikoa fixatu gabeko kriotomo ebakietan
burutu zen Moore-k (1976) deskribatu eta
Cajaraville et al.-ek, (1989) modifikatu legez.
Ebakiak (8 µm) -21ºC-ko kabinako tenperatura
zuen CM3000 kriotomoan (Leica Instruments
GmbH, Nussloch, Alemania) lortu ziren, ebakiak
giro-tenperaturan zeuden portaobjektuetan jazo
ziren eta -40ºC-tan mantendu ziren tindatu aurretik.
Ebakiak, 28 mg naphtol AS-BI glucuronide 1.2 ml
sodio bikarbonatoan (50 mM) disolbatutako eta 100
ml-arte %2.5 NaCl zuen 0.1 M sodio azetato
indargetzailean (pH 4.5) murgildu ziren. Disoluzio
honetan, %20 (W/V) polibinilo alkoholezko 10-21 g
disolbatu ziren egonkortzaile koloidal gisa. Ebakiak
40 minutuz 37ºC-tan inkubatu ziren etengabeko
irabiatzea zuen bainu batean. Ondoren, laginak
%2.5 NaCl-tan garbitu ziren 2 minutuz eta 37ºC-
tan,eta %2.5 NaCl eta 1 mg/ml Fast Garnet GBC
0.1 M fosfato indargetzailean (pH 7.4) tindatu ziren
10 minutuz giro tenperaturan. Ebakiak, Baker
formol kaltzikoan (%4 formaldehido, %1 CaCl2 eta
%2.5 NaCl) fixatu ziren 10 minutuz 4ºC-tan.
Laburki, uretan garbitu eta gero %0.1 zuen Fast
Green FCF disoluzioan kontrastatu zen 2 minutuz.
Azkenik, ebakiak Kaiser gelatina glizerinatuan
muntatu ziren.
Prozedura estereologikoa aplikatuz eta irudi-
analisia erabiliz liseri-zelulen lisosomen egitura
kuantifikatu zen (Cajaraville et al., 1995b;
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
147

Etxebarria et al., 1994). Sistema, aldez aurretik
Cajaraville et al.-ek, (1991) deskribatu dutenez
B&W-CDD bideo-kamera batez, Leitz Laborlux
mikroskopio batez eta Pentium ordenagailu batez
osaturik dago, softwarea BMS sistema izanik
(Sevisan, Bilbo, Espainia). x100 handipeneko
objektiboa erabili zen eta lisosomak eta liseri-
zelulen zitoplasma bereiziz irudi binarizatuak erabili
ziren. Lehenengo neurketak eskuz doitu ziren, lagin
desberdinen arteko tindaketa-intentsitate
desberdintasun txikiak ekiditeko asmoz. Irudi-
analisi sistemak hurrengo parametroak sortzen ditu:
lisosomen dentsitate bolumetrikoa (VvLYS =
VL/VC), dentsitate superfiziala (SvLYS = SL/VC),
azalera/bolumena ratioa (S/VLYS = SL/VL) eta
dentsitate numerikoa (NvLYS = NL/VC) non V =
bolumena, S = azalera, L = lisosomak eta C = liseri-
zelulen zitoplasma baitira. Formula
estereologikoak, batezbesteko diametroa
ebakiduraren lodiera baino txikiagoak duten
partikulentzako zuzenketa-faktorea dauka sartuta
(Lowe et al., 1981). Cajaraville et al.-en, (1995a)
arabera, 5 neurketa burutu ziren liseri-guruin
bakoitzeko.
Emaitzak eta eztabaida
148
1. Irudia: Magurio kontrol (A eta B) eta 1.25 mg Cd/l pean mantendutako magurioen (C eta D) irudi histologikoak, bai hematoxilina-eosinaz (A eta C) zein toluidina urdinez (B eta D) tindatutako ebakietan. Geziek zelula basofilikoak adierazten dituzte. L) liseri-tubuluen argia. Eskala-marrak: 50 µm (A, B eta D) eta 25 µm (C).
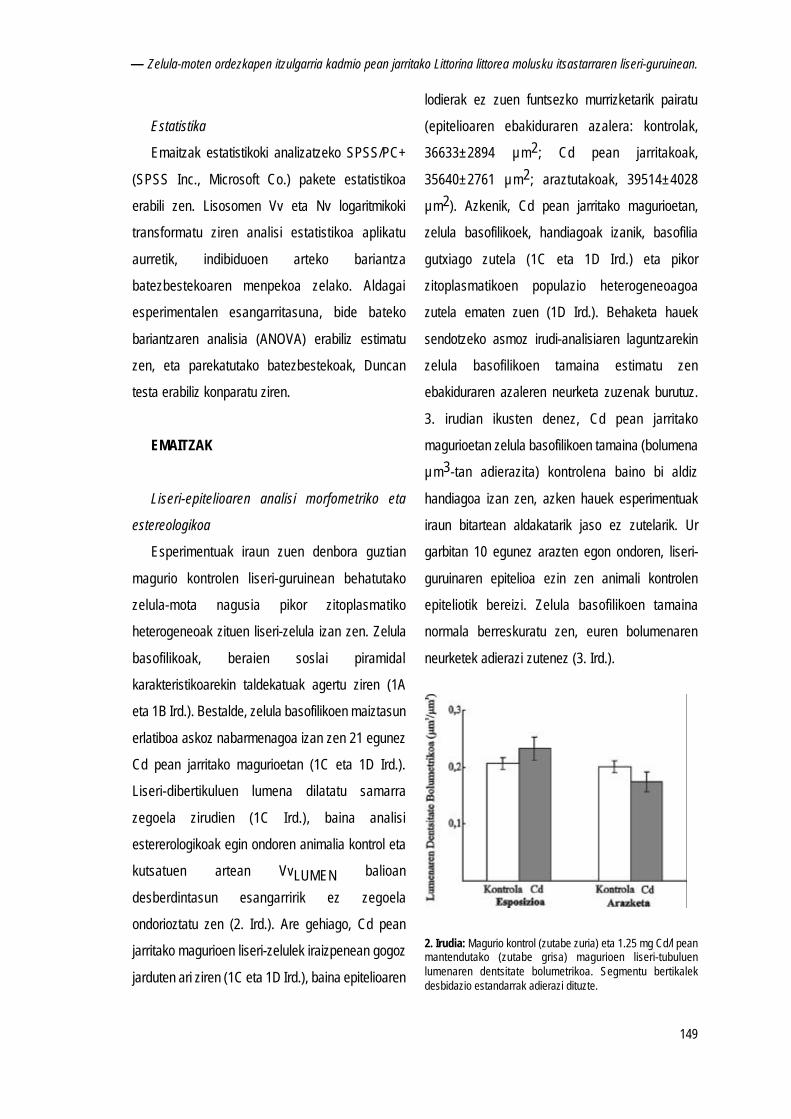
Estatistika
Emaitzak estatistikoki analizatzeko SPSS/PC+
(SPSS Inc., Microsoft Co.) pakete estatistikoa
erabili zen. Lisosomen Vv eta Nv logaritmikoki
transformatu ziren analisi estatistikoa aplikatu
aurretik, indibiduoen arteko bariantza
batezbestekoaren menpekoa zelako. Aldagai
esperimentalen esangarritasuna, bide bateko
bariantzaren analisia (ANOVA) erabiliz estimatu
zen, eta parekatutako batezbestekoak, Duncan
testa erabiliz konparatu ziren.
EMAITZAK
Liseri-epitelioaren analisi morfometriko eta
estereologikoa
Esperimentuak iraun zuen denbora guztian
magurio kontrolen liseri-guruinean behatutako
zelula-mota nagusia pikor zitoplasmatiko
heterogeneoak zituen liseri-zelula izan zen. Zelula
basofilikoak, beraien soslai piramidal
karakteristikoarekin taldekatuak agertu ziren (1A
eta 1B Ird.). Bestalde, zelula basofilikoen maiztasun
erlatiboa askoz nabarmenagoa izan zen 21 egunez
Cd pean jarritako magurioetan (1C eta 1D Ird.).
Liseri-dibertikuluen lumena dilatatu samarra
zegoela zirudien (1C Ird.), baina analisi
estererologikoak egin ondoren animalia kontrol eta
kutsatuen artean VvLUMEN balioan
desberdintasun esangarririk ez zegoela
ondorioztatu zen (2. Ird.). Are gehiago, Cd pean
jarritako magurioen liseri-zelulek iraizpenean gogoz
jarduten ari ziren (1C eta 1D Ird.), baina epitelioaren
lodierak ez zuen funtsezko murrizketarik pairatu
(epitelioaren ebakiduraren azalera: kontrolak,
36633±2894 µm2; Cd pean jarritakoak,
35640±2761 µm2; araztutakoak, 39514±4028
µm2). Azkenik, Cd pean jarritako magurioetan,
zelula basofilikoek, handiagoak izanik, basofilia
gutxiago zutela (1C eta 1D Ird.) eta pikor
zitoplasmatikoen populazio heterogeneoagoa
zutela ematen zuen (1D Ird.). Behaketa hauek
sendotzeko asmoz irudi-analisiaren laguntzarekin
zelula basofilikoen tamaina estimatu zen
ebakiduraren azaleren neurketa zuzenak burutuz.
3. irudian ikusten denez, Cd pean jarritako
magurioetan zelula basofilikoen tamaina (bolumena
µm3-tan adierazita) kontrolena baino bi aldiz
handiagoa izan zen, azken hauek esperimentuak
iraun bitartean aldakatarik jaso ez zutelarik. Ur
garbitan 10 egunez arazten egon ondoren, liseri-
guruinaren epitelioa ezin zen animali kontrolen
epiteliotik bereizi. Zelula basofilikoen tamaina
normala berreskuratu zen, euren bolumenaren
neurketek adierazi zutenez (3. Ird.).
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
149
2. Irudia: Magurio kontrol (zutabe zuria) eta 1.25 mg Cd/l peanmantendutako (zutabe grisa) magurioen liseri-tubuluenlumenaren dentsitate bolumetrikoa. Segmentu bertikalekdesbidazio estandarrak adierazi dituzte.

Liseri-guruinaren epitelioko zelula-moten
osaketan gertatu ziren aldaketak kuantifikatzeko,
VvBAS eta VvDIG kalkulatu ziren (4. Ird.). Bi
parametroak, osagarriak izanik, Cd pean jarritako
magurioetan eta magurio kontrol eta 10 egunez
arazten mantendutako magurioen artean
esangarriki desberdinak izan ziren (4A eta 4B Ird.).
VvBAS-en igoera, edota aldibereko VvDIG-en
jaitsiera, liseri-zelulen galerak edota zelula
basofilikoen ugaritzeak sortua den zehazteko
asmoz, liseri-zelula eta zelula basofilikoen bolumen
eta kopuru absolutuen estimazioa burutu zen,
planimetria eta estereologia batera aplikatuz.
Termino absolututan, Cd pean jarritako
magurioetan liseri-zelulen bolumena, berriro ere 10
egunetako arazketa-aldiaren ondoren jaitsi zen
kontrolen balioetara bueltatuz (4D Ird.). Bitartean,
zelula basofilikoen bolumena 10 egunez Cd pean
jarritako magurioetan igo zen, eta arazketaren egin
ondoren, berriro ere kontrolen balioetara bueltatu
zen (4D Ird.). Horrela, 17.8 x 106 µm3-ko liseri-
guruinaren zati bati dagokionez, kontroletan liseri-
zelulen bolumena 11.8 x 106 µm3 koa izan zen,
gutxi gorabehera. Cd pean jarri ondoren, liseri-
zelulen 1.6 x 106 µm3-ko bolumena galdu zen, eta
10 eguneko arazketa-aldia igaro ondoren, liseri-
zelulen bolumena berriro ere 12.7 x 106 µm3-ko
baliora bueltatu zen. Liseri-zelulen kopuruari
dagokionez, Cd pean jarritako magurioetan liseri-
guruinaren zati horretan 1850 liseri-zelula galdu
zitrela erakutsi zuten, magurio kontroletan liseri-
zelulen kopurua 13900 zeluletakoa izanik, gutxi
gorabehera. 10 egunez arazten utzi ondoren, liseri-
zelulen kopurua kontroletan baino apur bat
altuagoa izan zen, 14700 inguru. Zelula
basofilikoen bolumen absolutuari dagokionez, 17.8
x 106 µm3-ko zati arbitrarioan, magurio kontrolen
1.1 x 106 µm3-ko bolumena Cd pean jarritako
magurioetan 2.3 x 106 µm3-ra emendatu zen, eta
arazketaren ondoren berriro 1.2 x 106 µm3-ra itzuli
zen. Zelula basofilikoen kopurua, bere aldetik, 210-
315 tartean mantendu zen, eta talde esperimental
desberdinen artean esangarriki aldatu ez zelarik
(4E eta 4F Ird.).
PCNA immunohistokimika
Aurretik aipatutako zelula-kopuruen aldaketak
azaltzeko asmoz, zelulen proliferazio tasetan
balizko aldaketak aurkitzeko PCNA
immunohistokimika aplikatu zen. Behar bezalako
irudiak lortzeko, zailtasun handiak topatu ziren; izan
ere, lipofuszinak eta pikor zitoplasmatikoetan
seinale ez-espezifiko bortitza topatu zen; batez ere,
Cd pean jarritako magurioetan. Tindaketa-
prozedura desberdinekin, DAB, fosfatasa alkalinoa
eta tindagai fluoreszenteak barne, hainbat
Emaitzak eta eztabaida
150
3. Irudia: Magurio kontrol (zutabe zuria) eta 1.25 mgCd/l peanmantendutako (zutabe grisa) magurioen zelula basofilikoenbatezbesteko bolumena. Segmentu bertikalek desbidazioestandarra adierazi dituzte. Goiko asteriskoek taldeen artekodesberdintasun esangarriak adierazi dituzte p<0.05 mailan.

saiakuntza egin ondoren, nukleo PCNA-positiboak
bestelako egituretatik errez bereiztea lortu zen ,
baina aldez aurretik pentsatutako PCNAren analisi
kuantitatiboak egitea ezinezkoa suertatu zen. Dena
den, behaketa kualitatiboak, nahiz eta mugatuak
izan, interesgarriak suertatu ziren oso. Erabilitako
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
151
4. Irudia: Magurio kontrol (zutabe zuria) eta 1.25 mg Cd/l pean mantendutako (zutabe grisa) magurioen neurketa estereologikodesberdinak. (A) Zelula basofilikoen dentsitate bolumetrikoa; (B) Liseri-zelulen dentsitate bolumetrikoa; (C) Zelula basofilikoek
betetzen duten bolumena 17.8 x 106 µm3 bolumenarekiko; (D) Liseri-zelulek betetzen duten bolumena 17.8 x 106 µm3
bolumenarekiko; (E) Zelula basofilikoen kopurua 17.8 x 106 µm3 -tako bolumen batean: (F) Liseri-zelulen kopurua 17.8 x 106 µm3
-tako bolumen batean. Segmentu bertikalek desbidazio estandarrka adierazi dituzte. Goiko asteriskoek taldeen artekodesberdintasun esangarriak adierazi dituzte p<0.05 mailan.

arratoien PCNA aurkako PC10 antigorputz
komertziala, magurioen zelula proliferatzaileak
detektatzeko gai izan zen; izan ere, gonadan oso
erreakzio bortitza behatu zen (5A Ird.). Gonada
arran, espermatozito primarioetan, sekundarioetan
baino erreakzio bortitzagoa behatu zen, eta
espermatida helduek ez zuten PCNA-markaketarik
erakutsi (5A Ird.). PCNArik gabe inkubatutako
tindaketa kontroletan ez zen inolako seinalerik ikusi
(5B Ird.). Gonada emean, erreakzio positiboa
obozitoen nukleoan soilik behatu zen, nukleoloan
inolako seinalerik behatu ez zelarik (5C eta 5D Ird.).
Bestalde, magurio kontroletan liseri-zelulen nukleo
PCNA-positibo gutxi behatu ziren, eta lipofuszinen
erreaktibotasunagak sortutako zarata gutxi aurkitu
genuen (6A-C Ird.). Hala ere, Cd pean jarritako
magurioetan, lifofuszina eta lisosomen materialak
PCNA-markaketa bortitza eman zuten (6D-F Ird.),
magurio kontroletan eta araztuetan behatu ez zena
(6F-H Ird.).
Autometalografia
Autometalografiak, liseri-zelulen lisosometan
zilarrezko hauspeakin beltzen (BSD) presentzia
erakutsi zuen, zelula basofilikoetan BSDrik behatu
ez zen bitartean (7. Ird.). Magurio kontroletan
BSDen maila basalak behatu ziren, eta Cd pean
mantendutako magurioetan askoz erreakzio
bortitzagoak aurkitu ziren (7A-C Ird.). 10 eguneko
arazketa-aldia bukatu ondoren, tindaketa-patroia,
magurio kontroletan ikusitakoaren antzekoa izan
zen (7D Ird.). Behaketa horiek, VvBSD gisa
neurtutako BSDen hedapena neurtzerakoan
lortutako emaitza kuantitatiboekin bat egin zuten.
VvBSD balioak, Cd pean jarritako magurioetan
magurio kontroletan baino hiru aldiz handiagoak
izan ziren, eta are gehiago, araztutako magurioen
artean eta magurio kontrolen artean ez zen
desberdintasun esangarririk aurkitu (8. Ird).
Lisosomen egitura-aldaketak
Lisosomak, ß-glukuronidasa entzimaren
histokimika erabiliz ikustarazi ziren. Honela, arre-
Emaitzak eta eztabaida
152
5. Irudia: PCNA immunohistokimika burutu ondoren magurioen gonada ar eta emean behatutako immunomarkaketa. (A) PCNA-markaketa positiboa ikusten da espermatogonietan g), espermatozito primario p) eta sekundarioetan s) eta baita espermatidetanere t). Eskala-marra: 25 µm. (B) Antigorputz primariorik gabe inkubatutako lagin kontroletan (folikulu espermatikoa) ez da inolakomarkaketa positiborik behatzen. Eskala-marra: 25 µm. (C) eta (D) PCNAren lokalizazioa gonada emeetan, bai obozito helduetano), zein obozito goiztiarretan e). Nukleoek markaketa positibo bortitza erakusten duten bitartean (geziak), nukleoloetan ez damarkaketarik behatzen (gezi-buruak). Eskala marra: 50 µm.

purpura koloreko egitura esferiko gisa agertu ziren
liseri-zelulen zitoplasman. Lisosomak, Cd pean
jarritako magurioetan kontrol eta araztutako
taldeenetan baino apur bat handiagoak eta
ugariagoak ziren (9. Ird.). Lisosomen egituraren
analisi estereologikoak erakutsi zutenaren arabera,
nahiz eta S/VLYS eta NvLYS parametroetan
desberdintasun esangarririk ez aurkitu Cd-ak,
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
153
6. Irudia: PCNA immunohistokimika magurio kontrol eta 1.25 mg Cd/l pean mantendutako magurioetan. (A eta C) Maguriokontrolak, T) liseri-tubuloa, D) liseri-zelula, B) zelula basofilikoa, H) hemozitoa. Geziek liseri-zelula positiboak adierazten dituzte,eta gezi-buruek zelula basofiliko positiboak adierazten dituzte. Ikusi lipofuszinen erreaktibogarritasun urria magurio kontroletan (A).Eskala-marra: 50 µm (A) eta10 µm (C). (B) Antigorputz primariorik gabe inkubatutako laginetan ez da inolako markaketa positiborikbehatzen. Eskala-marra: 50 µm. (D-F) Cd pean 21 egunez mantendutako magurioetan lipofuszinak erreakzio bortitza erakustendute. T) liseri-tubuloa, D) liseri-zelula, B) zelula basofilikoa. Geziek liseri-zelula positiboak adierazten dituzte, eta gezi- buruek zelulabasofiliko positiboak adierazten dituzte. Eskala-marra: 50 µm (D) eta 10 µm (E eta F). (G-H) 10 egunez arazten utzitakomagurioetan lipofuszinen erreakzionagarritasuna ahulagoa da eta zelula PCNA positibo ugari (geziak) ikus daitezke. T) liseri-tubuloa. Eskala-marra: 50 µm (G) eta 10 µm (H).

VvLYS balioaren igoera (lisosomen handipena)
esangarria eragin zuen, arazketa burutu ondoren
balioak berriro ere kontrolen mailara itzuliz (10.
Ird.).
EZTABAIDA
Moluskuen liseri-guruinean kutsatzaileen eragin
pean behatutako bestelako erantzunen artean,
epitelioko zelula-moten ordezkapena (CTR)
bibalbio eta gastropodoetan fenomeno orokor gisa
deskribatu da (Widdows et al., 1984; Lowe &
Clarke, 1989; Cajaraville et al., 1990a; 1990b;
Marigómez et al., 1990; 1996; 2005a; 2005b).
Zenbait kasutan, bai zelula basofiliko normalek zein
hipertrofiatutako liseri-guruineko epitelioko parterik
nagusiena osatuz aurkitu dira, eta horregatik, CTR
prozesua zelula basofilikoen proliferazioa dela
medio burutzen dela proposatu da (Rasmussen et
al., 1983; Lowe & Clarke, 1989: Cajaraville et al.,
1990a; 1990b; Marigómez et al., 1990; Syasina et
al., 1997). Are gehiago, Yonge (1926) eta Mix &
Sparks-en (1971) lanetan oinarriturik, orokorrean,
liseri-guruinaren epitelioaren berriztapen-gaitasuna
zelula basofilikoei esleitu zaie (Thompson et al.,
1974; Nelson & Morton, 1979). Izatez, zelula
basofilikoen proliferazioa, babes-mekanismoa izan
daitekeela iradoki da, non zelula basofilikoak behar
bezalako baldintzak ematekotan liseri-zelula
helduetan desberdintzatu baitaitezken (Thompson
et al., 1974). Zelula basofilikoei eta liseri-zelulei
funtzio arras desberdinak egokitu zaizkiela kontuan
Emaitzak eta eztabaida
154
7. Irudia: Littorina littorea magurioaren liseri-epitelioan metalen lokalizazio autometalografikoa, bai 1.25 mg Cd/l pean 21 egunezmantendu eta gero, (A eta B); maguio kontroletan (C); zein 10 egunez arazten mantendutako magurioetan (D). Geziek liseri-zelulenlisosometan dauden zilarrezko hauspeakin beltzak adierazten dituzte, gezi-buruek liseri-epitelioko xafla basalean dagoen metalaadierazten dute. Eskala-marra: 25 µm.
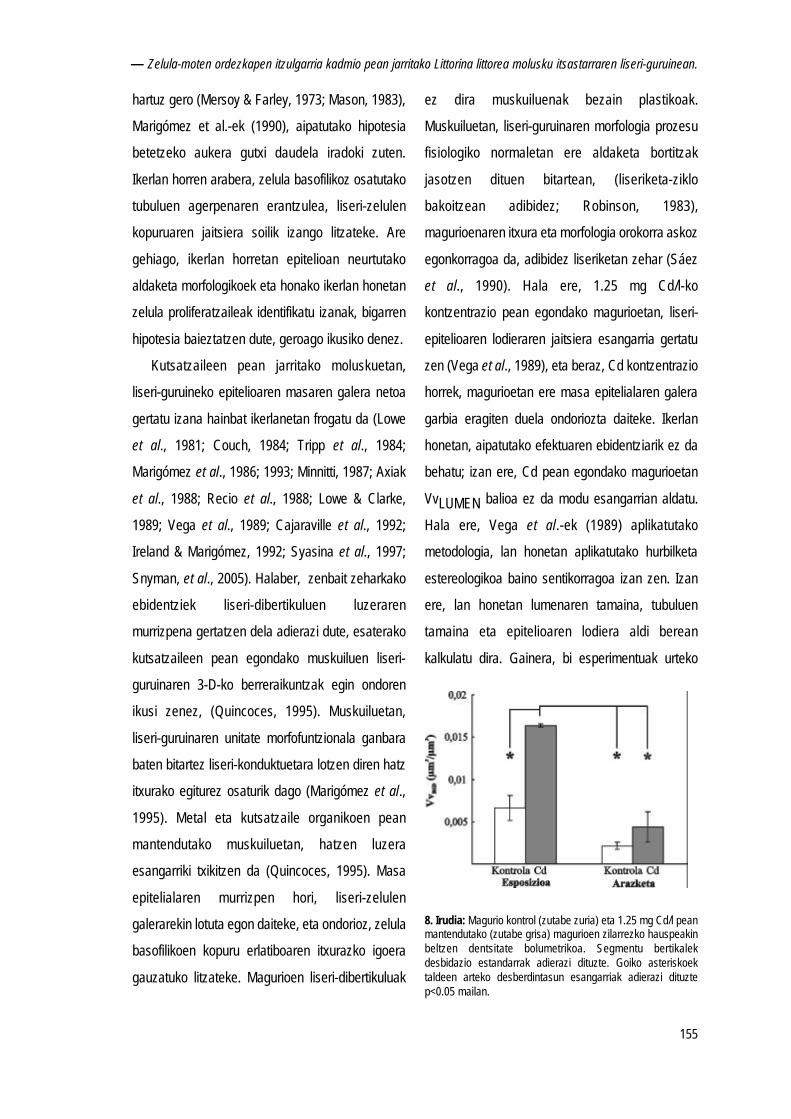
hartuz gero (Mersoy & Farley, 1973; Mason, 1983),
Marigómez et al.-ek (1990), aipatutako hipotesia
betetzeko aukera gutxi daudela iradoki zuten.
Ikerlan horren arabera, zelula basofilikoz osatutako
tubuluen agerpenaren erantzulea, liseri-zelulen
kopuruaren jaitsiera soilik izango litzateke. Are
gehiago, ikerlan horretan epitelioan neurtutako
aldaketa morfologikoek eta honako ikerlan honetan
zelula proliferatzaileak identifikatu izanak, bigarren
hipotesia baieztatzen dute, geroago ikusiko denez.
Kutsatzaileen pean jarritako moluskuetan,
liseri-guruineko epitelioaren masaren galera netoa
gertatu izana hainbat ikerlanetan frogatu da (Lowe
et al., 1981; Couch, 1984; Tripp et al., 1984;
Marigómez et al., 1986; 1993; Minnitti, 1987; Axiak
et al., 1988; Recio et al., 1988; Lowe & Clarke,
1989; Vega et al., 1989; Cajaraville et al., 1992;
Ireland & Marigómez, 1992; Syasina et al., 1997;
Snyman, et al., 2005). Halaber, zenbait zeharkako
ebidentziek liseri-dibertikuluen luzeraren
murrizpena gertatzen dela adierazi dute, esaterako
kutsatzaileen pean egondako muskuiluen liseri-
guruinaren 3-D-ko berreraikuntzak egin ondoren
ikusi zenez, (Quincoces, 1995). Muskuiluetan,
liseri-guruinaren unitate morfofuntzionala ganbara
baten bitartez liseri-konduktuetara lotzen diren hatz
itxurako egiturez osaturik dago (Marigómez et al.,
1995). Metal eta kutsatzaile organikoen pean
mantendutako muskuiluetan, hatzen luzera
esangarriki txikitzen da (Quincoces, 1995). Masa
epitelialaren murrizpen hori, liseri-zelulen
galerarekin lotuta egon daiteke, eta ondorioz, zelula
basofilikoen kopuru erlatiboaren itxurazko igoera
gauzatuko litzateke. Magurioen liseri-dibertikuluak
ez dira muskuiluenak bezain plastikoak.
Muskuiluetan, liseri-guruinaren morfologia prozesu
fisiologiko normaletan ere aldaketa bortitzak
jasotzen dituen bitartean, (liseriketa-ziklo
bakoitzean adibidez; Robinson, 1983),
magurioenaren itxura eta morfologia orokorra askoz
egonkorragoa da, adibidez liseriketan zehar (Sáez
et al., 1990). Hala ere, 1.25 mg Cd/l-ko
kontzentrazio pean egondako magurioetan, liseri-
epitelioaren lodieraren jaitsiera esangarria gertatu
zen (Vega et al., 1989), eta beraz, Cd kontzentrazio
horrek, magurioetan ere masa epitelialaren galera
garbia eragiten duela ondoriozta daiteke. Ikerlan
honetan, aipatutako efektuaren ebidentziarik ez da
behatu; izan ere, Cd pean egondako magurioetan
VvLUMEN balioa ez da modu esangarrian aldatu.
Hala ere, Vega et al.-ek (1989) aplikatutako
metodologia, lan honetan aplikatutako hurbilketa
estereologikoa baino sentikorragoa izan zen. Izan
ere, lan honetan lumenaren tamaina, tubuluen
tamaina eta epitelioaren lodiera aldi berean
kalkulatu dira. Gainera, bi esperimentuak urteko
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
155
8. Irudia: Magurio kontrol (zutabe zuria) eta 1.25 mg Cd/l peanmantendutako (zutabe grisa) magurioen zilarrezko hauspeakinbeltzen dentsitate bolumetrikoa. Segmentu bertikalekdesbidazio estandarrak adierazi dituzte. Goiko asteriskoektaldeen arteko desberdintasun esangarriak adierazi dituztep<0.05 mailan.

sasoi desberdinetan burutu ziren (errute-garaian,
abendua-urtarrila versus errute-ostean, otsaila-
martxoa). Erantzuna termino kualitatibotan
antzekoa izatea espero bada ere, erantzunaren
hedadura desberdina izan daiteke, abendua-
urtarrila garaian (Vega et al., 1989) otsaila-martxoa
garaian (ikerlan hau) baino erantzun sentikorragoak
behatuz. Modu berean, CTR erantzuna, nahiz eta
Cd kontzentrazio berdinak erabili, leunagoa
suertatu da ikerlan honetan, abendua-urtarrila
garaian burutu ziren esperimentuekin (Marigómez
et al., 1990) alderatuz. Are gehiago, Marigómez et
al.-en (1990) arabera, espero baino zelula
basofilikoen hipertrofia gutxiago behatu da oraingo
honetan .
Zelulen galeraren ebidentziarekin batera, orain
epitelioko zelulen berriztapen-mekanismoen
inguruan datu berriak daude eskuragarri.
Muskuiluen liseri-guruinean, bai liseri-zelulak zein
zelula basofilikoak PCNA zein BrdU
immunohistokimikak eginez erreakzionagarriak
direla frogatu da, eta beraz, bi zelula-motek bere
ziklo zelularra norberak bere aldetik betetzen dute
(Marigómez et al., 1999; Zaldibar et al., 2004). Hori
dela eta, kutsatzaileen eraginez desagertu diren
liseri-zelulak ordezkatzeko ez dirudi zelula
basofilikoak proliferatzen eta desberdintzatzen
direnik, liseri-zelulek bere aldetik bikoizteko
gaitasuna baitute. PCNA immunohistokimikaren
bitartez behatu denez, nahiz eta Cd pean
mantendutako magurioetan eta araztutako
magurioetan lipofuszinen erreakzionagarritasun
handiak emaitza kuantitatiboak lortzerik baimendu
ez, badirudi Cd-ak liseri-zelulen proliferazioaren
Emaitzak eta eztabaida
156
9. Irudia: Littorina littorea magurioaren liseri-epitelioan ß-glukuronidasa entzimaren jardueraren lokalizazioa histokimika bitartez,bai 1.25 mg Cd/l pean 21 egunez mantendu eta gero, (A eta B ); magurio kontroletan (C); zein 10 egunez arazten mantendutakomagurioetan (D). Geziek lisosomen barneko ß-glukuronidasa jarduera adierazten dute. Eskala-marra: 25 µm.

areagotzea eragiten duela. Areagotutako
proliferazio hori, 10 egunez arazten utzitako
magurioetan oraindik ere mantentzen da.
Proliferazio-tasak kalkulatzeko gai izan ez garenez,
arazketa ondoren liseri-guruinean gertatzen den
zelula-moten berreskurapena, liseri-zelulak gehiago
proliferatzen dutelako edo liseri-zelula gutxiago
galtzen direlako ondorioztatzea ezinezkoa zaigu.
Kadmio pean egondako magurioetan, liseri-
zelulen bolumen absolutua jaitsi zen 10 egunetako
arazketa-aldia eta gero, kontrolen balioetara itzuli
bazen ere. Zelula basofilikoen bolumen absolutua,
Cd peko tratamenduaren ondoren igo zen eta
arazketa-aldia eta gero berriro ere kontrolen
balioetara jaitsi zen. Liseri-zelulen bolumen
absolutuaren jaitsiera, liseri-zelulen galera netoa
dela medio eman zen bitartean (1850 liseri-zelula
galdu ziren liseri-guruinaren 17.8 x 106 µm3-ko zati
batean), zelula basofilikoen bolumenaren igoera
zelulen hipertrofiak sortua da, eta ez zelula
kopuruaren igoerak, sortua izan zen.
Ikerlan honetan lortutako emaitza guztiak batera
kontuan hartuta, CTR, zelula basofilikoen
proliferazioak sortua baino, gehiago, liseri-zelulen
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
157
10. Irudia: Magurio kontrol (zutabe zuria) eta Cd pean mantendutako (zutabe grisa) magurioen ß-glukuronidasa jarduerarenparametro estereologikoak (A) Dentsitate bolumetrikoa; (B) Dentsitate superfiziala; (C) Azalera/Bolumena ratioa; (D) Dentsitatenumerikoa. Segmentu bertikalek desbidazio estandarrak adierazi dituzte. Goiko asteriskoek taldeen arteko desberdintasunesangarriak adierazi dituzte p<0.05 mailan.

galera netoak sortua zela dirudi. Hala ere, liseri-
zelulen galera neto hori, liseri-zelulen proliferazio
areagotuarekin batera gertatu zen. Ornodunen
zeluletan, programatutako heriotza zelularra zein
zelulen proliferazioa Cd-ak eragiten duena jakina
da (Beyersmann & Hechtenberg, 1997; Habeebu et
al., 1998; Piechotta et al., 1999). Cd pean
egondako Helix pomatia barraskiloetan kontroletan
baino lau aldiz programatutako heriotza zelular
gehiago erregistratu da liseri-zeluletan
(Chavikovsky et al., 2004). Programatutako
heriotza zelular hori, elkar baztertzaileak ez diren
autofagia eta apoptosioren konbinaketak eraginda
dagoela dirudi (Bursch et al., 2000; Chavikovsky et
al., 2004). Izan ere, oraingo honetan Cd-ak liseri-
zelulen proliferazioa eragin dezakeela behatu da;
beraz, liseri-zelulen galera netoa zelulen heriotza
eta proliferazioaren arteko balantze gisa azal
daiteke. Beraz, magurioetan, Cd pean egoteak,
arazketari aurre egiteko liseri-zelulen berriztapena
areagotzen du.
Kadmio pean egondako magurioen zelula
basofilikoen hiperplasia, esposizio-denbora eta
kontzentrazioaren araberakoa da (Marigómez et al.,
1990). Zelula basofilikoen (kaltzio-zelulen)
hiperplasia zenbait metalen pean egondako
lehorreko gastropodoetan ere deskribatu da
(Marigómez et al. 1996; 1998; Chabicovsky et al.
2004). Hiperplasia hori fenomeno desberdinekin
erlazionatu da: (1) liseri-zelulen funtzioak (liseriketa
intrazelularra barne) kaltetuta egonez gero,
liseriketa estrazelularrerako entzimen areagotutako
ekoizpenarekin (Cajaraville et al., 1990b;
Marigómez et al., 1990); (2) zelula basofilikoetan
areagotutako detoxifikazio-jarduerekin (Cajaraville
et al., 1990a; Livingstone & Pipe, 1992; Triebskorn
& Köhler, 1996); eta (3) apoptosi bidez hildako
liseri-zelulen fagozitosiarekin (Chabicovsky et al.,
2004), hain zuzen ere. Bestalde, aipatutako
hiperplasia, liseri-zelulen galerak epitelioan toki
libre gehiago uzten duenez, zelula basofilikoen
tamaina eta soslai aldaketekin erlazionatuta egon
daiteke, ugaztunen urdaileko epitelioan deskribatu
den bezala (Clarke, 1970). Liseri-guruinaren
histofisiologiari dagokionez, duela gutxi egindako
behaketen arabera (argitaratu gabe), ez dirudi
areagotutako entzimen jariapena azalpen
sinesgarria denik. Halaber, kutsatzaile organikoen
pean, baina ez metalen pean, zelula basofilikoen
erretikulu endoplasmatiko pikortsuan areagotutako
jarduera detoxifikatzaileen berri eman da
(Livingstone & Pipe, 1992; Triebskorn & Köhler,
1996). Edonola ere, metalen detoxifikazioaren
kasuan partaide nagusiak liseri-zelulak direnez
(Vega et al., 1989), ikerlan honetako behaketak
azaltzeko pisu gutxiko aukera dirudi. Azkenik, zelula
basofilikoek hiltzen ari diren liseri-zelulen hondarren
fagozitosian parte hartzen dutela hiperplasia
azaltzeko aukera egokiena izan daiteke. Intsektuen
epidermisean (Locke, 1985), ugaztunen hepatozito
(Dini et al., 2002), Sertoli zelula (Nakanishi &
Shiratsuchi, 2004) eta bestelako epitelio-zeluletan
(Monks et al., 2005), eta gastropodoen liseri-
guruineko epitelio-zeluletan (Chavikovsky et al.,
2004), hiltzen diren epitelio-zelulak, alboko epitelio-
zelulek fagozitatzen dituztela deskribatu da.
Oraintsu, Cd-z elikatutako bareetan gorputz
apoptotikoak izan daitezkenak deskribatu dira (5.
Emaitzak eta eztabaida
158

Kapitulua). Edonola ere, azken hipotesi hori
baieztatzeko, teknika immunohistokimikoak erabiliz
zelula apoptotikoen identifikazio espezifikoak eta
mikroskopio elektroniko mailan egindako ikerketak
burutzea beharrezkoa da.
Ikerlan honen ekarpen garrantzitsuena, CTR
itzulgarria dela frogatzea izan da: 10 eguneko
arazketa-aldia eta gero, liseri-zelulak berriz ere
zelula basofilikoak baino ugariagoak suertatu dira.
Liseri-zelulen PCNA-markaketa Cd pean
mantendutako zein araztutako magurioetan
antzekoa izanik, CTRren itzulgarritasunak liseri-
zelulen galera moteltzearekin zerikusia behar du
izan, Cd-aren gabezian liseri-zelulen jarduera
metabolikoa berriz ere bere egoera normalera
itzultzea espero baita (Marigómez et al., 1986).
Beraz, liseri-zelulen berriztapena kutsatzaileen
gisako ingurumen-faktoreek modulatua dagoela
dirudienez, zelula-moten osaketa bereizgarria
ingurumen-baldintza berezi bati egotzi ahal zaio.
Ingurumen-erasoaren maila eta luzeraren
araberakoa izan daitekeen moldaera hori, liseri-
guruinaren fisiologiaren ulermenean eta
biomarkatzaileen aplikazioan eragina izan dezake,
itu-zeluletan ezartzen ez diren eta matrize
analitikoan (liseri-guruina) suertatzen diren
aldaketak aintzakotzat hartzen ez dituzten
prozedurak erabiltzen badira.
Metal-esposizioak sortarazitako CTRk, BSD-
hedapena bezalako esposizio-biomarkatzaile eta
lisosomen erantzunak bezalako efektu-
biomarkatzaileen gainean eragina izan dezakeela
frogatu da muskuiluetan (Soto et al., 2002; 1.
Kapitulua). Modu berean, metal-esposizioak
muskuilu eta magurioen liseri-guruinean
eragindako masa netoaren aldaketek, AAS bidez
estimatutako ehunetako metal-zaman eragina izan
dezakete (Soto et al., 1997a; 1997b). Dirudienez,
liseri-zelulen galerak eta berreskurapenak, ikerlan
honetan aplikatutako biomarkatzaileen (BSD-
hedapena, lisosomen parametro estereologikoak)
gainean eraginik ez dute. Magurioen liseri-
guruineko VvBAS balioak 0,2 µm3/µm3 baino
handiagoak izan arren, zelula basofilikoak
saihestuz; lisosomen barneko metal-kopuru eta
lisosomen handipenaren neurketak zuzenean liseri-
zelulen gainean egin daitezke.
Laburbilduz, ikerlan honen ondorioak honako
hauek dira: (1) "itxurazko" zelula-moten
ordezkapena liseri-zelulen galerak eta aldibereko
zelula basofilikoen hipertrofiak sortua dela; (2)
prozesua itzulgarria dela; (3) liseri-zelulen
proliferazioa Cd pean mantendutako magurioetan
areagotzen dela, eta tendentzia hori arazketa-
aldiaren ondoren mantentzen dela; eta (4) liseri-
zelulen galera eta berreskurapena ikerlan honetan
erabilitako maila zelular eta tisularreko
biomarkatzaileen gainean eraginik ez dirudi duenik.
BIBLIOGRAFIA
Axiak V, George JJ, Moore MN (1988) Petroleum hydrocarbons
in the marine bivalve Venus verrucosa: accumulation and
cellular responses. Mar. Biol. 97:225-230.
Beyersmann D, Hechtenberg S (1997) Cadmium, gene regula-
tion, and cellular signalling in mammalian cells. Toxicol.
Appl. Pharmacol. 144:247-261.
Bursch W, Ellinger A, Gerner CH, Fröhwein U, Schulte-
Hermann R (2000) Programmed cell death (PCD) apopto-
sis, autophagic PCD, or others?. Ann. N.Y. Acad. Sci.
926:1-12.
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
159

Cajaraville MP, Marigómez JA, Angulo E (1989) A stereological
survey of lysosomal structure alterations in Littorina littorea
exposed to 1-naphtol. Comp. Biochem. Physiol. C 93:231-
237.
Cajaraville MP, Díez G, Marigómez I, Angulo E (1990a)
Responses of basophilic cells of the digestive gland of
mussels to petroleum hydrocarbon exposure. Dis. Aquat.
Org. 9:221-228.
Cajaraville MP, Marigómez I, Angulo E (1990b) Short-term toxic
effects of 1-naphthol on the digestive gland-gonad complex
of the marine prosobranch Littorina littorea (L.): a light
microscopic study. Arch. Environ. Contam. Toxicol. 19:17-
24.
Cajaraville MP, Marifómez JA, Angulo E (1991) Automated
measurement of lysosomal structure alterations in oocytes
of mussels exposed to petroleum hydrocarbons. Arch.
Environ. Contam. Toxicol. 21:395-400.
Cajaraville MP, Marigómez I, Díez G, Angulo E (1992)
Comparative effects of the water accommodated fractions
(WAF) of three oils on mussels. 2.- Quantitative alterations
in the structure of the digestive tubules. Comp. Biochem.
Physiol. C 102:113-123.
Cajaraville MP, Abascal I, Etxeberria M, Marigómez I (1995a)
Lysosomes as cellular markers of environmental pollution:
time- and dose-dependent responses of the digestive lyso-
somal system of mussels after petroleum hydrocarbon
exposure. Environ. Toxicol. Water Qual. 10:1-8.
Cajaraville MP, Robledo Y, Etxeberria M, Marigómez I (1995b)
Cellular biomarkers as useful tools in the biological monito-
ring of environmental pollution: molluscan digestive lysoso-
mes. 29-45 orr. Non: Cell Biology in Environmental
Toxicology. Cajaraville MP (ed) University of the Basque
Country Press Service. Bilbo.
Chabicovsky M, Klepal W, Dallinger R (2004) Mechanisms of
cadmium toxicity in terrestrial pulmonates: Programmed
cell death and metallothionein overload. Environ. Toxicol.
Chem. 23:648-655.
Clarke RM (1970) A new method of measuring the rate of shed-
ding of epithelial cells from the intestinal villus of the rat.
Gut 11:1015-1019.
Couch JA (1984) Atrophy of diverticular epithelium as an indi-
cator of environmental irritants in the oyster, Crassostrea
virginica. Mar. Environ. Res. 14:525-526.
Danscher G (1984) Autometallography. A new technique for
light and electron microscopic visualization of metals in bio-
logical tissues (gold, silver, metal sulphides and metal sele-
nides). Histochemistry 81:331-335.
Dini L, Pagliara P, Carlà EC (2002) Phagocytosis of apoptotic
cells by liver: A morphological study. Microsc. Res. Tech.
57:530-540.
Etxeberria M, Sastre I, Cajaraville MP, Marigómez I (1994)
Digestive lysosome enlargement induced by experimental
exposure to metals (Cu, Cd and Zn) in mussels collected
from a zinc polluted site. Arch. Environ. Toxicol. 27:338-
345.
Etxeberria M, Cajaraville MP, Marigómez I (1995) Changes in
digestive cell lysosomal structure as biomarkers of environ-
mental stress in the Urdaibai estuary (Biscay Coast,
Iberian Peninsula). Mar. Poll. Bull. 30:599-603.
Habeebu SSM, Liu J, Klaassen CD (1998) Cadmium-induced
apoptosis in mouse liver. Toxicol. Appl. Pharmacol. 153:48-
58.
Ireland MP, Marigómez I (1992) The influence of dietary cal-
cium on the tissue distribution of Cu, Zn, Mn and P and his-
tological changes in the digestive cells in the snail Achatina
fulica Bowdich. J. Moll. Stud. 58:157-168.
Livingstone DR, Pipe RK (1992) Mussels and environmental
contaminants: molecular and cellular aspects. 425-464 orr.
Non: The Mussel Mytilus: ecology, Physiology, Genetics
and Culture. EM Gosling (ed). Elsevier Science Publs B.V.
Amsterdam.
Locke M (1985) A structural analysis of post embryonic deve-
lopment. 103-165 orr. Non: Parasites-Their world and ours.
Mattrick CF & Desser SS (ed). Elsevier Biomedical Press.
New York.
Lowe DM, Clarke KR (1989) Contaminant-induced changes in
the structure of the digestive epithelium of Mytilus edulis.
Aquat. Toxicol. 15:345-358.
Lowe DM, Moore MN, Clarke KR (1981) Effects of oil in the
digestive cells in mussels: quantitative alterations in cellu-
lar and lysosomal structure. Aquat. Toxicol. 1:213-226.
Marigómez I, Angulo E, Moya J (1986) Copper treatment of the
digestive gland of the slug Arion ater L. 2. Morphometrics
and histophysiology. Bull. Environ. Contam. Toxicol.
36:608-615.
Marigómez I, Cajaraville MP, Angulo E (1990) Histopathology of
the digestive gland-gonad complex of the marine proso-
branch Littorina littorea exposed to cadmium. Dis. Aquat.
Org. 9:229-238.
Emaitzak eta eztabaida
160

Marigómez I, Soto M, Etxeberria M, Angulo E (1993) Effects of
size, sex, reproduction, and trematode infestation on the
quantitative structure of digestive tubules in stressed win-
kles. Zool. Jb. Anat. 123:319-336.
Marigómez I, Cajaraville MP, Quincoces I, Salisbury JR (1995)
Computer assisted 3-d reconstruction techniques may pro-
vide new insides into the pattern of histological organisa-
tion of the bivalvian digestive gland. 12th Int. Malacol.
Congr. Vigo.
Marigómez I, Soto M, Kortabitarte M (1996) Tissue-level bio-
markers of biological effect of mercury on sentinel slugs,
Arion ater. Arch. Environ. Contam. Toxicol. 31:54-62.
Marigómez I, Cajaraville MP, Soto M, Lekube X (1998) Cell-
type replacement, a successful strategy of mollusks to
adapt to chronic exposure to pollutants. Cuad. Invest. Biol.
20:411-414.
Marigómez I, Lekube X, Cancio I (1999) Immunochemical loca-
lisation of proliferating cells in mussel digestive gland tis-
sue. Histochem. J. 31:781-788.
Marigómez I, Soto M, Cajaraville MP, Angulo E, Giamberini L
(2002) Cellular and subcellular distribution of metals in
molluscs. Microsc. Res. Tech. 56:358-392.
Marigómez I, Baybay-Villacorta L (2003) Pollutant-specific and
general lysosomal responses in digestive cells of mussels
exposed to model organic chemicals. Aquat. Toxicol.
64:235-257.
Marigómez I, Izagirre U, Lekube X (2005a) Lysosomal enlarge-
ment in the digestive cells of mussels exposed to cadmium,
bezo[a]pyrene and their combination. Comp. Biochem.
Physiol. C 141:188-193.
Marigómez I, Soto M, Cancio I, Orbea A, Garmendia L,
Cajaraville MP (2005b) Cell and tissue biomarkers in mus-
sel, anh histopathology in hake and anchovy from Bay of
Biscay after the Prestige oil spill (Monitoring Campaign
2003). Mar. Poll. Bull. (prentsan).
Mason AZ (1983) The uptake, accumulation and excretion of
metals by the marine prosobranch gastropod mollusc
Littorina littorea (L). Doktoradutza-Tesia. University of
Wales, Gwyneed (UK). 575 orr.
Merdsoy B, Farley I (1973) Phasic activity in the digestive gland
of the marine prosobranch gastropod, Littorina littorea (L.).
Proc. Malacol. Soc. London 40:473-482.
Minnitti F (1987) Effects of copper pollution on the hepatopan-
creas of Cyclope neritea L. (Mollusca:Gastropoda). Zool.
Anz. 219:141-146.
Mix MC, Sparks AK (1971) Repair of digestive tubule tissue of
the pacific oyster, Crassostrea gigas, damaged by ionizing
radiation. J. Invert. Pathol. 17:172-177.
Monks, J., Rosner, D., Geske, F.J., Lehman, L., Hanson, L.,
Neville, M.C., Fadok, V.A. (2005) Epithelial cells as pha-
gocytes: Apoptotic epithelial cells are engulfed by mam-
mary alveolar epithelial cells and repress inflammatory
mediator release. Cell Death Diff. 12:107-114.
Moore MN (1976) Cytochemical demonstration of latency of
lysosomal hydrolases in digestive cells of the common
mussel Mytilus edulis, and changes induced by thermal
stress. Cell Tiss. Res. 175:279-287.
Morton B (1983) Feeding and digestion in bivalvia. 65-147 orr.
Non: The Mollusca. ASM & Wilburg M. (ed). Academic
Press. Saleuddin. New York.
Nakanishi Y, Shiratsuchi A (2004) Phagocytic removal of apop-
totic spermatogenic cells by sertoli cells: Mechanisms and
consequences. Biol. Pharma. Bull. 27:13-16.
Nelson L, Morton JE (1979) Cyclic activity and epithelial rene-
wal in the digestive gland tubules of the marine proso-
branch Maoricrypta monoxyla (Lesson). J. Moll. Stud.
45:262-283.
Piechotta G, Lacorn M, Lang T, Kammann U, Simat T, Jenke
HS, Steinhart H (1999) Apoptosis in dab (Limanda liman-
da) as possible new biomarker for anthropogenic stress.
Ecotox. Environ. Saf. 42:50-56.
Quincoces I (1995) Un nuevo modelo de la glándula digestiva
de Mytilus galliprovincialis (Bivalvia, Eulamellibranchia):
reconstrucción tridimensional asistida por ordenador y
microscopía electrónica de barrido. Lizentziatura-Tesia.
UPV/EHU. 112 orr.
Rasmussen LPD, Hage E, Karlog O (1983) Light and electron
microscopic studies of the acute and chronic toxic effects
of N-nitroso compounds on the marine mussel, Mytilus
edulis (L) II. N-methyl-N-nitro-N-nitrosoguanidine Aquat.
Toxicol. 3:301-311.
Recio A, Marigómez JA, Angulo E, Moya J (1988) Zinc tratment
of the digestive gland of the slug Arion ater L. 1. Cellular
distribution of zind and cadmium. Bull. Environ. Contam.
Toxicol. 41:858-864.
Robinson WE (1983) Assessment of bivalve intracellular diges-
tion based on direct measuremets. J. Moll. Stud. 49:1-8.
Zelula-moten ordezkapen itzulgarria kadmio pean jarritako Littorina littorea molusku itsastarraren liseri-guruinean.
161

Sáez V, Marigómez I, Angulo E, Moya J (1990)
Histomorphology and histochemistry of the digestive gland
of Littorina littorea (L.) in relation to experimental tidal con-
ditions, food availability, and digestion. Zool. Jb. Anat.
120:185-196.
Snyman RG, Reinecke AJ, Reinecke SA (2005) Quantitative
changes in the digestive gland cells of the snail Helix
aspersa after exposure to fungicide copper oxychloride.
Ecotox. Environ. Saf. 60:47-52.
Soto M, Agirregoikoa MG, Pérez MA, Marigómez I (1990) A pla-
nimetric study of morphological variability in the digestive
diverticula of Littorina littorea (Linnaeus) and Mytilus edulis
Linnaeus. J. Moll. Stud. 56:339-344.
Soto M, Marigomez I (1997a) BSD extent, an index for metal
pollution screening based on the metal content within
digestive cell lysosomes of mussels as determined by
autometallography. Ecotox. Environ. Saf. 37:141-151.
Soto M, Marigómez I (1997b) Metal bioavailability assessment
in “mussel-watch” programmes by automated image analy-
sis of autometallographical black silver deposits (BSD) in
digestive cell lysosomes. Mar. Ecol. Prog. Ser. 156:141-
150.
Soto M, Ireland MP, Marigómez I (1997a) The contribution of
metal/shell weight index in target-tissues to metal body
burdens in sentinel marine mollusks. 1 Littorina littorea.
Sci. Tot. Environ. 198:135-147.
Soto M, Ireland MP, Marigómez I (1997b) The contribution of
metal/shell weight index in target-tissues to metal body
burdens in sentinel marine mollusks. 2 Mytilus galloprovin-
cialis. Sci. Tot. Environ. 198:149-160.
Soto M, Quincoces I, Lekube X, Marigomez I (1998a)
Autometallographed metal content in digestive cells of win-
kles: a cost-effective screening tool for monitoring Cu and
Zn pollution. Aquat. Toxicol. 40:123-140.
Soto M, Quincoces I, Marigómez I (1998b)
Autometallographical procedure for the localization of
metal traces in molluscan tissues by light microscopy. J.
Histotechnol. 21:123-127.
Soto M, Zaldibar B, Cancio I, Taylor MG, Turner M, Morgan AJ,
Marigómez I (2002) Subcellular distribution of cadmium
and its cellular ligands in mussel digestive gland cells as
revealed by combined autometallography and X-ray micro-
probe analysis. Histochem. J. 34:273-280.
Syasina IG, Vaschenko MA, Zhandan PM (1997) Morphological
alterations in the digestive diverticula of Mizuhopecten yes-
soensis (Bivalvia: Pectenidae) from polluted areas of Peter
the Great Bay, Sea of Japan. Mar. Environ. Res. 44:85-98.
Thompson RJ, Ratcliffe NA, Bayne BL (1974) Effects of starva-
tion on structure and function of the digestive gland of the
mussel (Mytilus edulis L) J. Mar. Biol. Assoc. U.K. 54:699-
712.
Triebskorn R, Köhler HR (1996) The impact of heavy metals on
the grey garden slug, Deroceras reticulatum (Muller): Metal
storage, cellular effects and semi-quantitative evaluation of
metal toxicity. Environ. Poll. 93:323-343.
Tripp MR, Fries CR, Craven MA, Grier CE (1984)
Histopathology of Mercenaria mercenaria as an indicator of
pollutant stress. Mar. Environ. Res. 14:521-524.
Vega MM, Marigómez I, Angulo E (1989) Quantitative altera-
tions in the structure of the digestive cell of Littorina littorea
on exposure to cadmium. Mar. Biol. 103:547-553.
Weibel ER (1979) Stereological Methods. 1. Academic Press.
London 415 orr.
Widdows J, Bakke T, Bayne BL, Donkin P, Livingstone DR,
Lowe DM, Moore MN, Evans SV, Moore SL (1984)
Responses of Mytilus edulis on exposure to the WAF of
North Sea oil. Mar. Biol. 67:15-31.
Yonge CM (1926) The digestive diverticula in Lamellibranchia.
Trans. Roy. Soc. Edin. 54:703-718.
Zaldibar B, Cancio I, Marigómez I (2004) Circatidal variation in
epithelial cell proliferation in the mussel digestive gland
and stomach. Cell Tiss. Res. 318:395-402.
162
Emaitzak eta eztabaida

Liseri-zelulen berriztapena kadmio etakerosenozko nahasketa baten peanesperimentalki jarritako bareen liseri-guruinekoepitelioan
Laburpena: Arion ater (L) bareak lurzoruen osasun-maila ebaluatzeko organismo-behale gisaproposatu dira. Kutsatzaile pean egondako bareen liseri-guruinean zelula-moten osaketan aldaketakbehatu dira, aldaketa horien artean esanguratsuena liseri-zelulen galera eta horrekin batera iraizte- etakaltzio-zelulen emendioa izanik. Lan honen helburua, liseri-zelulen galerak efektu- eta esposizio-biomarkatzaileen gainean eragina duen, eta zelulen galera hori laborategiko baldintzetan metal etakonposatu organikozko nahasketa baten pean jarritako bareetan itzulgarria ote den antzematea da.Horretarako, bareak kutsatu gabeko toki batetik (Delika, Euskal Herria) hartu ziren eta 10 µg Cd + %30kerosenotik lortutako urari egokitutako frakzioa zuen janariaz 27 egunez elikatu ziren. Gorputzeko metal-zama eta kerosenoaren mailak kalkulatu ziren erabilitako esposizioa eraginkorra zela baieztatzeko asmoz.Esposizio-biomarkatzaile gisa, autometalografia bitartez lortutako zilarrezko hauspeakin beltzen (ingeleraz,Black Silver Deposits; BSD) dentsitate bolumetrikoa eta azil-KoA oxidasa (AOX) entzimaren jarduera erabiliziren. Efektu-biomarkatzaile gisa, liseri-epitelioa osatzen duten 3 zelula-moten (liseri-, iraizte- eta kaltzio-zelulak) dentsitate bolumetrikoan neurtutako aldaketak erabili ziren. Zelula proliferatzaileakbromodeoxiuridinaren (BrdU) immunodetektapenaren bitartez identifikatu ziren. Ikerlan honen emaitzek,kutsatzaile pean jarritako bareetan AOX jarduera eta BSD-en denstitate bolumetrikoaren igoera eta liseri-zelulen kopuruaren jaitsiera gertatzen dela islatu zuten, dena den aldaketa horiek liseri-guruinekokutsatzaileen metaketa-gaitasunean eta erabilitako efektu- eta esposizio-biomarkatzaileen gaineaneraginik ez zuten izan. Gainera, liseri-epitelioko zelula-moten osaketan behatutako aldaketa eraldatu horiarazte-aldiko 7 egunen buruan itzulgarria zela behatu zen. BrdU-markaketak kutsatzaile pean egoteakliseri-zelulen proliferazio areagotua eragiten duela erakutsi zuen, proliferazio-jarduera areagotu hori arazte-aldian zehar ere mantentzen zelarik.
Gako-hitzak: liseri-guruina, bareak, zelula-moten ordezkapena, kadmioa, kerosenoa, efektu- etaesposizio-biomarkatzaileak..
Zelula-moten ordezkapena bareen liseri-guruinean: lanborategi-ikerketa
163

Digestive cell turnover in the digestive gland epithelium of slugsexperimentally exposed to a mixture of cadmium and kerosene
Abstract: Slugs, Arion ater (L), have been proposed as sentinel organisms to assess soil environmentalhealth. In slugs under the influence of pollutants changes in the cell-type composition of the digestive-glandhave been observed, among these changes, the most remarkable one is the loss of digestive cells and therelated increase of excretory and calcium cells.The aim of the present work was to determine whetherdigestive cell loss affects biomarkers of exposure and effect and whether is reversible in the digestive glandof slugs exposed to a mixture or metal and organic pollutants under laboratory conditions. For thesepurposes, slugs collected from a non-polluted site (Delika, Basque Country) were dosed with 10 µg Cd/gr+ 30% of the water accommodated fraction of kerosene in the food for 27 days. Metal body burdens andkerosene tissue levels were measured. The volume density of black silver deposits (VvBSD) revealed by
autometallography, and the activity of the enzyme acyl-CoA oxidase (AOX) were used as exposurebiomarkers. As effect biomarkes, changes in the volume density of the three cell types that constitute thedigestive gland epithelium in slugs (digestive, excretory and calcium cells) were calculated. Proliferatingcells were identified by means of bromodeoxyuridine (BrdU) immunohistochemistry. The results of thepresent work revealed that the mixture of pollutants provoked an increase in VvBSD and AOX activity as
well as a decrease in the number of digestive cells. Anyway, these changes had no effect in the capacity ofthe digestive gland to accumulate or in the effect and exposure biomarkers employed. Moreover, thechange in cell-type composition measured in the digestive epithelium was reversible after 7 days ofdetoxification. BrdU-labelling showed that exposure to pollutants provoked an enhanced digestive cellproliferation, that perdured during detoxification
Key words: digestive gland, slugs, cell-type replacement, cadmium, kerosene, effect and exposurebiomarkers.
Emaitzak eta eztabaida
164

Renovación de células digestivas en el epitelio de la glándula digestiva delimacos experimentalmente expuestos a una mezcla de cadmio y keroseno
Resumen: Los limacos, Arion ater (L) han sido propuestos como organismos centinela en la evaluaciónde la salud del suelo. Se han observado cambios en la composición de tipos celulares de la glánduladigestiva de limacos bajo la influencia de contaminantes. De entre todos estos cambios el más reseñablees la pérdida de células digestivas y el aumento de células excretores y de calcio que le acompaña. Elobjetivo del presente trabajo es determinar si la pérdida de células digestivas afecta a los biomarcadoresde exposición y efecto, y si dicha pérdida es reversible en limacos expuestos a una mezcla decontaminantes metálicos y orgánicos en condiciones de laboratorio. Con ese fín, se recogieron limacos deun lugar no contaminado (Delika, País Vasco) y se trataron con 10 µg Cd/gr + 30% de la fracciónacomodada al agua de keroseno en la dieta durante 27 días. Se midió la concentración de Cd y dekeroseno en la glándula digestiva. La densidad volumétrica de BSD (VvBSD), (BSD, del inglés; Black Silver
Deposit) revelados mediante autometalografía y la actividad del enzima acil-CoA oxidasa fueron losbiomarcadores de exposición estudiados. Como biomarcadores de efecto se calcularon los cambios en ladensidad volumétrica de los tres tipos celulares que contituyen el epitelio de la glándula digestiva delimacos (células digestivas, excretoras y de calcio). Las células proliferativas se identificaron medianteimmunohistoquímica de bromodeoxiuridina (BrdU). Los resultados obtenidos demostraron que la mezclade contaminantes provocan un aumento en VvBSD y en la actividad AOX así como la pérdida de células
digestivas en limacos, aunque los cambios observados en el epitelio digestivo no afectaron a la capacidadde acumulación de contaminantes ni a los biomarcadores de efecto y exposición empleados en el presentetrabajo. Además, el cambio en la composición de tipos celulares del epitelio digestivo fué reversibledespués de 7 días de depuración. Por otro lado, el marcaje de BrdU demostró que la exposición acontaminantes induce la proliferación de las células digestivas y que dicho aumento en la tasa deproliferación se mantiene durante el período de detoxificación.
Palabras clave: glándula digestiva, limacos, recambio de tipo celular, cadmio, keroseno,biomarcadores de efecto y exposición.
Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
165

Emaitzak eta eztabaida
166

Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
167
SARRERA
Moluskuetan, liseri-guruinaren epitelioko zelula-
moten osaketa larriki aldatuta suerta daiteke
kutsatzaile-esposizio subletalen ondorioz
(Rasmussen et al., 1983, Widdows et al., 1984;
Lowe & Clarke, 1989; Cajaraville et al., 1990a;
1990b; Marigómez et al., 1990; 1996; 1998a;
1998b; Soto et al., 2002). Efektu hori, zelula-moten
ordezkapena (CTR; ingeleraz, cell-type
replacement) izenaz ezagutu da, eta orokorki
zelula basofilikoen dentsitate bolumetriko edo
kopuru erlatibo gisa neurtu da (Rasmussen et al.,
1983, Widdows et al., 1984; Lowe & Clarke, 1989;
Cajaraville et al., 1990a; 1990b; Marigómez et al.,
1990; 1996; 1998a; 1998b; 1. Kapitulua). CTR,
kutsadura oean gerta daitekeen zelula basofilikoen
(kaltzio-zelulen) proliferazioaren ondorioa dela
iradoki da (Thompson et al., 1974; Rasmussen et
al., 1983, Widdows et al., 1984; Lowe & Clarke,
1989; Cajaraville et al., 1990a; 1990b; Marigómez
et al., 1990; 1996). Haatik, Cd pean mantendutako
magurioetan behatutako zelula basofilikoen
itxurazko areagotzea nagusiki liseri-zelulen galera
(DCL; ingeleraz, Digestive Cell Loss) eta aldi
bereko zelula basofilikoen hipertrofiaren (BDH;
ingeleraz, Basophilic Cell Hypertrophy) ondorioa
dela frogatu da berriki (4. Kapitulua). DCL, liseri-
zelulen proliferazio eta heriotzaren arteko
desorekaren ondorioa dela dirudi, nahiz eta liseri-
zelulen kopuruaren galera netoa suertatu, Cd peko
esposizioak liseri-zelulen proliferazioaren
emendioa eragiten baitu (4. Kapitulua).
Muskuilu zein magurioetan, liseri-guruinaren
epitelioa bi zelula-mota nagusiz osaturik dago,
liseri-zelula eta zelula basofilikoa, alegia; hortaz,
"itxurazko" CTR, epitelioko zelula-kopuruaren
galera netoa ematen denez, DCLren ondorio
zuzena dela onar genezake (1. eta 4. Kapituluak).
Lehorreko gastropodoen liseri-guruineko epitelioa,
ordea, morfologikoki desberdintzatutako hiru
zelula-motez osaturik dago: liseri-zelulak
(ugarienak), iraizte-zelulak eta kaltzio-zelulak
(basofilikoak) (Sumner, 1965). Hala ere, iraizte-
zelulak, bizi-zikloaren azken urratsetan dauden
liseri-zelulak, eta ez hirugarren zelula-mota
desberdintzatua, izango liratekeela proposatu da,
(Porcel et al., 1996; Dimitriadis & Konstantinidou,
2002). Molusku itsastarretan bezala, kaltzio-zelulen
(zelula basofilikoen) hipertrofia eta kopuru erlatibo
emendatuaren berri eman da metalen pean
mantendutako lehorreko bareetan (Marigómez et
al. 1996; 1998a). Hala ere, iraizte-zelulen
proportzio erlatiboa ere emenda daiteke
(Marigómez et al., 1998a) eta, beraz, bareen liseri-
guruinean hiru zelula-motetan gertatzen diren
aldaketak "itxurazko" CTR gainean nola eragiten
duten argitu beharra dago. Are gehiago, Cd eta Zn
pean mantendutako magurio eta muskuiluetan
behintzat, "itxurazko" CTR egun gutxiren buruan
itzulgarria denez (1. eta 4. kapituluak), moluskuen
liseri-guruineko epitelio-zelulen berriztatzearen
ikuspegi orokorra eskuratzeko asmoz, erantzun
hori zelula-moten osaketa konplexuagoa duten

Emaitzak eta eztabaida
168
bareen liseri-guruinean ere itzulgarria ote den
argitzea komenigarria iruditu zitzaigun.
Beste alde batetik, muskuiluen kasuan metal-
esposizioak eragindako CTRk autometalografia
(AMG) bidez errebelatutako zilarreezko haupeakin
beltzen (BSD; ingeleraz, Black Silver Deposits)
BSD-hedapenaren moduko esposizio-
biomarkatzaileen gainean eragina duena badakigu
(Soto et al., 2002; 1. Kapitulua). Hala ere, Cd pean
jarritako magurioetan ez dirudi DCLk BSD-
hedapenaren gainean eragina duenik, lisosomen
barneko BSDen neurketa mikroskopikoak liseri-
zeluletan modu errazean egin baitaitezke, zelula
basofilikoak taldekaturik agertu ohi direlako.
Bareetan, metal desberdinen arabera emaitza
desberdinak erdietsi dira. Dietan 1000 µg Hg/g dosi
pean 3 aste baino gehiagoz tratatutako bareetan,
liseri-guruineko epitelioan suertatutako zelula-
moten osaketaren aldaketak VvBSD (BSDen
dentsitate bolumetrikoa) esangarriki eragin zuen,
liseri-epitelioa nagusiki BSDrik gabeko iraizte-
zelulez osaturik baitzegoen (Marigómez et al.,
1996). Merkurio pean mantendutako bareetan, BSD
liseri-zeluletan bereziki ugariak, iraizte-zeluletan
esangarriki urriagoak eta kaltzio-zeluletan bakanak
zirela aipatu beharra dago. Aitzitik, Zn pean
mantendutako bareetan, zelula-moten osaketak ez
zuen BSD-hedapenean eraginik izan, AMG bidez
errebelatutako BSDak hiru zelula-motetan
lokalizatu baitziren (Marigómez et al., 1998a). Cd
pean jarritako bareetan, aurretiko behaketek
BSDak iraizte-zeluletan ere bazeudela erakutsi
zuten, eta, beraz, lotugai-multzo eskuragarrien
arabera metal desberdinak zelula-mota
desberdinetan bahitu daitezkeela kontuan hartuz
(Marigómez et al., 2002), Cd-aren presentzian
zelula-moten ordezkapenean behatutako aldaketek
BSD-hedapenaren moduko metal-esposizioaren
biomarkatzaileen gainean eragina duten ikertzea
planifikatu genuen.
Azkenik, metal indibidualen, metalen
nahasketen, zein kutsatzaile organikoen pean
jarritako bareen liseri-gurineko epitelioan ere
"itxurazko" CTR deskribatu da (Müller, 1994;
Marigómez et al., 1996; 1998a; 6. Kapitulua), baina
metal eta konposatu organikoen nahasketek
eragindako efektuei buruz ez dago datu
eskuragarririk, guk dakigunez. Moluskuen liseri-
guruinean, zelula-moten osaketari dagokionez
metal eta hidrokarburoen nahasketen eta
kutsatzaile indibidualen eragina desberdina denez
(1. Kapitulua), ikerlan honetan Cd eta kerosenozko
(beren urari egokitutako frakzioa, hain zuzen; WAF:
ingeleraz, Water Accommodated Fraction)
nahasketa kutsatzaile eredu gisa aukeratu zen.
Horrela, ikerlan honetako helburuak hurrengoak
dira: (1) metal (Cd) eta kutsatzaile organikozko
(keroseno-WAF) nahasketa pean mantendutako
bareen liseri-guruinean DCL dagoen den finkatzea;
(2) magurioetan bezala, kutsatzaileen pean
jarritako bareen liseri-guruinean, DCL prozesu
itzulgarria den frogatzea; eta (3) zelula-moten
osaketan gertatutako aldaketek BSD-hedapena
moduko esposizio-biomarkatzaileen gainean
eragina duten argitzea. Helburu horiekin, Delika
(Euskal Herria) inguruko toki garbi batetik Arion ater
bareak hartu ziren. Kadmio eta keroseno-WAF
nahasketaren dosi subletal pean 27 egunez jarri

Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
169
ziren, eta ondoren laborategiko egoera garbitan
mantendu ziren 7 egunez. Neurketa osagarri gisa,
elikatze-jarduera eta hazkuntza, egunero neurtu
ziren. Bareen liseri-guruineko Cd eta kerosenoaren
kontzentrazioak, absortzio atomikozko
espektrofotometria (AAS) eta gas-
kromatografia/masa-espektrometria (GC/MS)
bidez neurtu ziren, hurrenez hurren. Halaber, AMG
bidez errebelatutako BSDen dentsitate
bolumetrikoa (VvBSD) eta azil-KoA oxidasa (AOX)
entzimaren jarduera bareen liseri-guruinean,
metalen eta konposatu organikoen esposizio-
biomarkatzaile gisa aplikatu ziren, hurrenez hurren.
Bareen liseri-guruinaren epitelioa osatzen duten
hiru zelula-moten dentsitate bolumetrikoa (VvDIG,
VvEXC, VvCAL) hematoxilina-eosinaz tindatutako
parafinazko ebakien gainean estereologia erabiliz
kalkulatu zen. Gainera, liseri-guruinen bolumen-
unitate arbitrario bateko zelula-kopuru absolutua
ere estimatu zen. Azkenik, bromodeoxiuridina
(BrdU) immunohistoki-mika erabiliz zelula
proliferatzaileak identifikatu ziren.
MATERIAL ETA METODOAK
Erreaktibo kimikoak
Erabilitako erreaktibo kimiko guztiak maila
analitikokoak eta Sigma-Aldrich (St. Louis,
Missouri, Amerikako Estatu Batuak), merkatal-
etxekoak dira besterik agertu ezean.
Diseinu esperimentala eta laginen prozesaketa
Arion ater (Linnaeus) lehorreko bareak Iberiar
penintsularen iparraldeko Delikako (42º 58’I, 3º
1’M) gune landatar garbian ekainaren bukaeran
(ar-fasea) hartu ziren. Animaliak laborategira
garraiatu ziren, eta 15 aleko taldeetan mantendu
ziren, aireztapen egokia baimentzeko saretxo
batez estalitako 3 l-ko kutxetan. Kutxak, egunero
dabilur ugariz garbitu ziren, esperimentuak iraun
bitartean.
Elikagairik gabeko moldatze-denbora (7 egun)
igaro ondoren, bareak egunero azenario, sagar,
uraza eta kalabaza zati proportzionalak zituen eta
%1.5 agar uretan zuen "dieta naturalez" elikatu
ziren. Hogei bare, Cd (10 Cd µg/g) eta %30
keroseno-WAF nahasketaz tratatu ziren. WAFa,
900 ml ur distilatu eta 100 ml keroseno gau osoan
20ºC-tan etengabe irabiatzen nahastuz ekoiztu
zen. Kadmioa, agar-disoluzioa prestatzeko erabili
zen uretan kloruro gatz moduan disolbatu zen,
agar-disoluzioa %30 WAF eta %70 ur distilatuan
prestatu zelarik. Horrela janariaren Cd eta
kerosenoaren kontzentrazioak 395.93 µg/g ehun-
pisu lehor eta 415.36 µg/g ehun-pisu lehor izan
ziren, hurrenez hurren. Aldi berean, esposatu
gabeko kontrol seriea ere mantendu zen. 27 egun
ondoren, Cd + keroseno nahasketaz tratatutako
bareak 7 egunez "dieta naturalez" elikatu ziren,
arazte-prozesua ikertzeko asmoz. Tratamenduaren
erreplika-serie bat gehitu genuen, bestearen
baldintza berberetan mantendu zena.
Janaria egunero aldatu zen. Elikatze-jarduera
determinatzeko, emandako janaria eta utzitako
janaria egunero pisatu ziren. Bareak ere egunero
eta taldeka pisatu ziren.
Esperimentuan zehar analisi kimiko eta
biologikoak egiteko hainbat animalia beharrezkoak

zirenez, erabilitakoak erreplika-serieko barez
ordezkatzen ziren animalien dentsitatea konstantea
mantentzeko asmoz. Izan ere, lehorreko
gastropodoen fisiologia populazio-dentsitatearen
arabera esangarriki aldatzen dela deskribatu da
(Runham & Hunter, 1970).
Analisi kimikoak
Disekzionatutako liseri-guruinak (tratamenduko
18. egunean, eta 7 egunetako arazte-aldia eta
gero), ur distilatuan garbitu ziren, eta 120ºC-tara 48
orduz labean berotu ziren, guztiz lehortu arte.
Lehortutako materiala almerizaren laguntzaz txikitu
zen, eta azido nitriko kontzentratuan liseritu zen.
Ondoren, plaka beroan azido nitriko kontzentratua
lurrundu ostean, liseritutako materiala ur distilatuan
diluitutako (0.1 M) azido nitrikoan berresekitu zen.
Laginak absortzio atomikozko espektrofotometroan
(PerkinElmer 2280, Wellesley, Massachusetts,
Amerikako Estatu Batuak) neurtu ziren.
Kadmioaren kontzentrazioak, µg Cd/g ehun pisu
lehor modura kalkulatu ziren. Erabilitako
estandarrak Merck merkatal-etxeak (Merck & Co.,
Inc, Whitehouse Station, New Jersey, Amerikako
Estatu Batuak) zertifikatutakoak izan ziren.
Kerosenoa neurtzeko, disekzionatutako liseri-
guruinak (tratamenduko 18. egunean, eta 7
eguneko arazte-aldia eta gero), ur distilatuan
garbitu ostean izoztu egin ziren. Izoztutako
materiala, liofilizatu, txikitu eta pisatu zen. Ondoren,
azetonan murgilduta ultrasoinuen bidezko
erauzketari ekin zitzaion. Iragazi ondoren laginak
FID detektagailua zuen HP 6890 gas-
kromatografo/masa-espektrometroan (Hewlett-
Packard, Palo Alto, California, Amerikako Estatu
Batuak) neurtu ziren.
Analisi biologikoetarako esperimentuko 0., 3.,
18., eta 27. egunetan eta arazte-aldiko 7. egunean
5 bare disekzionatu ziren taldeko. Liseri-guruinaren
zati txikiak Carnoy fixatzailean (%60 etanol
absolutua, %30 kloroformoa eta %10 azido
azetikoa) fixatu ziren ordu batez 4°C-tan. Fixatu
ondoren, laginak goranzko graduazioko etanolezko
bainu-serie batean deshidratatu ziren, eta parafinan
inkluditu ziren 60°C-tan. 7 µm-ko lodiera zuten
ebakiak mikrotomoan lortu ziren, eta hematoxilina-
eosinaz tindatu ziren behaketa morfologikoak eta
zelula-moten osaketaren neurketak burutu ahal
izateko.
Autometalografia
Parafinazko ebakiak xilolezko bainuetan
desparafinatu ostean, beheranzko graduazioko
etanoletan hidratatu ziren. Parafinazko ebakiak
labean (40ºC) mantendu ziren guztiz lehortu arte.
Laginak argazki-emultsio geruza mehe eta
uniformez estali ziren, gela ilunean (Ilford Nuclear
Emulsion L4, Ilford, Mobberley, Ingalaterra).
Gerora, laginak ilunpean mantendu ziren lehortu
bitartean (30 minutu), eta errebelatzaile komertziala
erabiliz Ultrafin Tetenal SF (1:5 diluzioa ur
distilatuan) (Tetenal AG & Co, Norderstedt,
Alemania) errebelatu ziren 15 minutuz. Ondoren,
errebelatzailearen erreakzioa %1 azido azetikozko
disoluzioan minutu batez murgilduz geldiarazi zen.
Azkenik, fixatzaileaz (Agfa Agefix, Agfa-Gevaert
N.V., Mortsel, Belgika, 1:9 uretan) 10 minutuz fixatu
ziren. Laginak, Kaiser gelatina glizerinatuan
Emaitzak eta eztabaida
170

muntatu ziren. Metal-ioien presentzia, zilarrezko
hauspeakin beltzen agerpenak adierazten du
(Danscher, 1984; Soto et al., 1998).
BSDen kuantifikazioa argi-mikroskopioan irudi-
analisiaren bitartez
Liseri-zelulen lisosometan zeuden BSDen
azalera irudi-analisia erabiliz kuantifikatu zen (Soto
& Marigómez, 1997), emaitzak BSDen dentsitate
bolumetriko gisa adieraziz (VvBSD). Argi-
mikroskopiazko irudiak (x100 handipenean) B&W-
CCD TV kamera, HR-TV kolorezko pantaila, Leitz
mikroskopioa, PC ordenagailua, PIP 1024 (Matrox
Electronic Systems Ltd., Dorval, Kanada) sistema
eta Sevisan (Bilbo, Espainia) entrepresak
garatutako softwarea erabiliz eskuratu ziren.
Ondoren, eskuz eta softwarearen laguntzarekin
liseri-zelulen lisosometako BSD eta zitoplasma
bereizteko segmentazio-prozedura aplikatu zen.
Neurketak bare bakoitzean 5 eremutan burutu
ziren, eta talde esperimental bakoitzean 5 bare
erabili ziren; tratamendu bakoitzeko guztira 25
neurketa eginez. Kalkuluak, Lowe et al.-ek (1981)
proposatutako formulak erabiliz egin ziren. Hau da,
BSD partikulen tamaina ebakiaren lodiera baino
txikiagoa zela kontuan hartuz, Holmes efektua
zuzentzeko faktorea aplikatu zen.
Epitelioaren konposizio zelularraren analisi
estereologikoak
Hematoxilina-eosinaz tindatutako lagin
berberak erabiliz (7 µm), liseri-zelulen (VvDIG),
kaltzio-zelulen (VvCAL) eta iraizte-zelulen (VvEXC)
dentsitate bolumetrikoak kuantifikatzeko prozedura
estereologikoa aplikatu zen. Horretarako lagin
bakoitzean azarez aukeratutako zelai batean egin
ziren kontaketak, guztira talde esperimental
bakoitzeko 5 kontaketa burutuz. Analisi
estereologikoak burutzeko Nikon Optiphot
mikroskopioari atxiki zegoen irudi-hodia erabili zen
(orotariko handipena: x670). VvDIG, VvCAL eta
VvEXC kalkulatzeko Weibel gratikula
(multipurpouse test system M-168; Weibel, 1979)
erabili zen, eta liseri-zelula, iraizte-zelula zein zelula
basofilikotan eroritako puntuak zenbatu ziren,
dentsitate bolumetrikoak Delesse printzipioaren
arabera kalkulatzeko (Weibel, 1979).
Liseri-zelula, kaltzio-zelula eta iraizte-zelulen
estimazio absolutuak dentsitate bolumetrikoen
datuetan oinarrituz burutu ziren. Liseri-, kaltzio- eta
iraizte-zelulek betetzen zuten bolumenak
(VDIG=VvDIG x 17.8 x 106 µm3; VCAL=VvCAL x
17.8 x 106 µm3; VEXC=VvEXC x 17.8 x 106 µm3)
arbitrarioki aukeratutako liseri-guruinaren bolumen
batekiko kalkulatu ziren (17.8 x 106 µm3 gutxi
gorabehera): bolumen hori kalkulu estereologikoak
egiteko erabilitako Weibel gratikularen (356000
µm2) arbitrarioki aukeratutako 50 µm-tako
lodierako ebakidurari dagokio. Lodiera horrek,
kaltzio-zelula handiena guztiz barneratua egotea
ahalbidetzen du.
Ondoren, liseri-guruineko zati arbitrario horretan
liseri-, kaltzio- eta iraizte-zelulen kopurua
kalkulatzeko, zelula-mota guztiek tamaina
konstantea zutela onartu zen. Horrek iraizte- eta
kaltzio-zelulen tamainaren azpiestimazioa ekarriko
zuen, kutsatzaile pean egondako bareetan
behintzat.
Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
171

AOX peroxisomikoaren jarduera
AOX jarduera espektrofotometrikoki
determinatu zen, 502 nm-ko uhin-luzeera erabiliz,
Small et al.-ek (1985) ezarritako metodologia
jarraituz. Bareen liseri-guruinak 4 ml/g ehun pH 7.6
zuen TVBE indargetzailean (1 mM sodio
bikarbonatoa, 1 mM EDTA, %0.1 etanola eta %0.01
Triton X-100) teflonezko homegenizatzailea erabiliz
izotzezko bainuan homogenizatu ziren. Lagin
homogenizatuak 500 g-tara zentrifugatu ziren 15
minutuz 4ºC-tan eta, ondoren, AOX jarduera
neurtzeko, gainjalkina TVBE indargetzailean 10
aldiz diluitu zen. Saiakerak, 30 µM palmitoil-CoA
oxidasa substratu gisa erabiliz, peroxidasa
endogenoak katalizatutako diklorofluoreszeina
diazetatoaren (Molecular probes, Eugene, Oregon,
Amerikako Estatu Batuak) H2O2 menpeko
oxidazioa du oinarri. Orotariko proteina-
kontzentrazioa Lowry et al.-ek (1951) garatutako
metodoan oinarritutako DC protein assay (BioRad,
Hercules, California, Amerikako Estatu Batuak)
erabiliz kalkulatu zen, estandar gisa γ-globulina
erabiliz.
BrdU immunohistokimika
Liseri-guruineko zelula proliferatzaileak BrdU
immunohistokimika bidez lokalizatzeko, 27.
esposizio-eguneko eta arazte-aldi osteko bareei
3.5 mg BrdU injektatu zitzaien oinean zehar.
Carnoy-z fixatutako eta parafinan inkluditutako
ebakiak (3-4 µm-ko lodiera) American Optical
mikrotomoan lortu ziren, eta aldez aurretik (3-amino
propil trietoxisilanoz) silanizatutako
portaobjektuetan jaso ziren. Ondoren, laginak
xilenoz desparafinatu ziren, beheranzko
graduazioko etanolezko bainu-seriean hidratatu eta
fosfato indargetzaile salinoz (PBS; ingeleraz,
Phosphate-Buffered Saline) garbitu ziren. DNA, 0.1
N HCl-ez desnaturalizatu zen ordu batez, 37°C-tan.
Ebakiak, %5 bovine serum albumine (BSA) zuen
suero blokeatzailean ordu batez inkubatu ziren giro-
tenperaturan eta berriro ere PBS-z garbitu ziren.
Gero, ebakiak BrdUren aurkako antigorputz
espezifikoan (Roche, Basilea, Suitza); %0.1 BSA
zuen PBSan 1:10 diluitua) inkubatu ziren. Laginak
PBS-z garbitu ziren, eta ondoren, abidina-biotina
entzima-konplexua erabiliz antigorputza ikustarazi
zen. Laburki, ebakiak, PBS-an diluitutako saguaren
aurkako ahuntzaren IgG antigorputz sekundario
biotinilatuan inkubatu ziren, 30 minutuz, eta
ondoren PBSn garbitu ziren. Fosfatasa alkalinoa
jarduera ikustarazteko kromogenoaren disoluzioa
(naftol fast-red zuen PBS indargetzailean
disolbatua) erabili zen. Azkenik, ebakiak Mayer
hematoxilinaz kontrastatu (1 minutu), dabiluretan
garbitu eta Kaiser gelatina glizerinatuan (Merck &
Co., Inc, Whitehouse Station, New Jersey,
Amerikako Estatu Batuak) muntatu ziren.
Mikrografiak, Olympus BX50 (Olympus, Tokio,
Japonia) mikroskopioari atxiki zegoen Olympus
PM20 argazki-kameraz erdietsi ziren.
EMAITZAK
Bare kontrolek elikatze-jarduera konstantea
erakutsi zuten esperimentuak iraun bitartean (1B
Ird.). Maneiu esperimentalak eta laborategiko
baldintzek jaitsiera txikia eragin zuten bareen
Emaitzak eta eztabaida
172
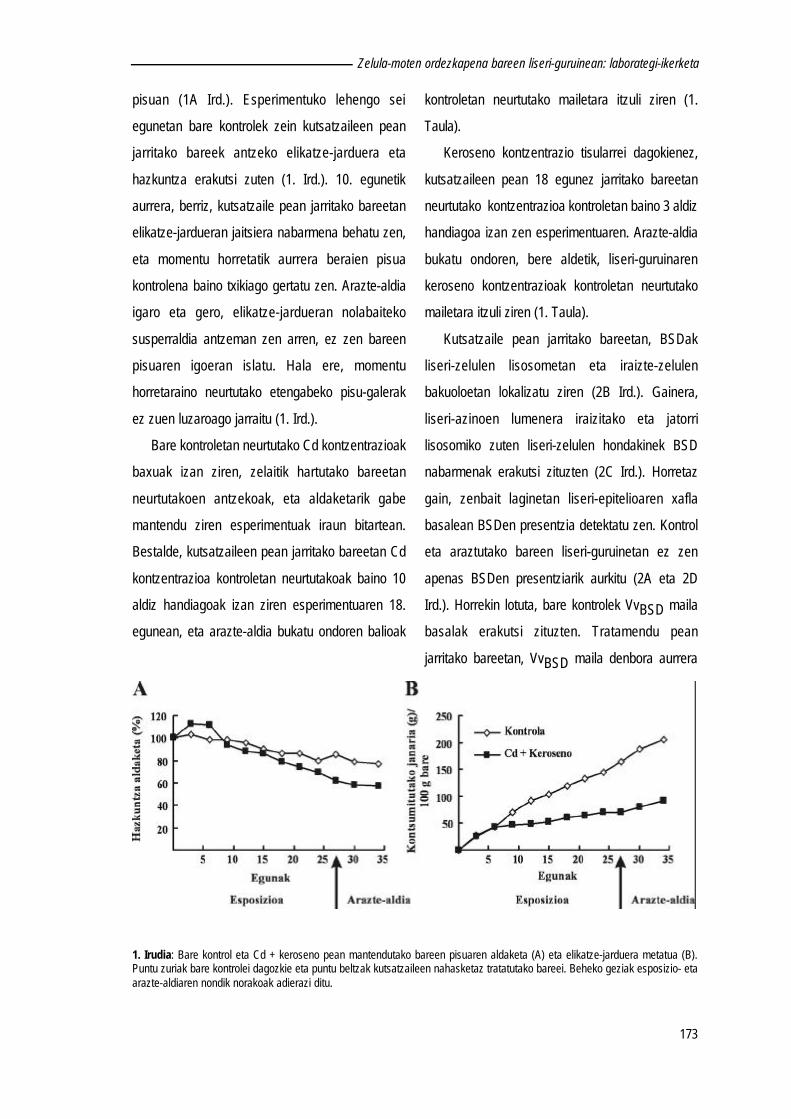
pisuan (1A Ird.). Esperimentuko lehengo sei
egunetan bare kontrolek zein kutsatzaileen pean
jarritako bareek antzeko elikatze-jarduera eta
hazkuntza erakutsi zuten (1. Ird.). 10. egunetik
aurrera, berriz, kutsatzaile pean jarritako bareetan
elikatze-jardueran jaitsiera nabarmena behatu zen,
eta momentu horretatik aurrera beraien pisua
kontrolena baino txikiago gertatu zen. Arazte-aldia
igaro eta gero, elikatze-jardueran nolabaiteko
susperraldia antzeman zen arren, ez zen bareen
pisuaren igoeran islatu. Hala ere, momentu
horretaraino neurtutako etengabeko pisu-galerak
ez zuen luzaroago jarraitu (1. Ird.).
Bare kontroletan neurtutako Cd kontzentrazioak
baxuak izan ziren, zelaitik hartutako bareetan
neurtutakoen antzekoak, eta aldaketarik gabe
mantendu ziren esperimentuak iraun bitartean.
Bestalde, kutsatzaileen pean jarritako bareetan Cd
kontzentrazioa kontroletan neurtutakoak baino 10
aldiz handiagoak izan ziren esperimentuaren 18.
egunean, eta arazte-aldia bukatu ondoren balioak
kontroletan neurtutako mailetara itzuli ziren (1.
Taula).
Keroseno kontzentrazio tisularrei dagokienez,
kutsatzaileen pean 18 egunez jarritako bareetan
neurtutako kontzentrazioa kontroletan baino 3 aldiz
handiagoa izan zen esperimentuaren. Arazte-aldia
bukatu ondoren, bere aldetik, liseri-guruinaren
keroseno kontzentrazioak kontroletan neurtutako
mailetara itzuli ziren (1. Taula).
Kutsatzaile pean jarritako bareetan, BSDak
liseri-zelulen lisosometan eta iraizte-zelulen
bakuoloetan lokalizatu ziren (2B Ird.). Gainera,
liseri-azinoen lumenera iraizitako eta jatorri
lisosomiko zuten liseri-zelulen hondakinek BSD
nabarmenak erakutsi zituzten (2C Ird.). Horretaz
gain, zenbait laginetan liseri-epitelioaren xafla
basalean BSDen presentzia detektatu zen. Kontrol
eta araztutako bareen liseri-guruinetan ez zen
apenas BSDen presentziarik aurkitu (2A eta 2D
Ird.). Horrekin lotuta, bare kontrolek VvBSD maila
basalak erakutsi zituzten. Tratamendu pean
jarritako bareetan, VvBSD maila denbora aurrera
Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
173
1. Irudia: Bare kontrol eta Cd + keroseno pean mantendutako bareen pisuaren aldaketa (A) eta elikatze-jarduera metatua (B).Puntu zuriak bare kontrolei dagozkie eta puntu beltzak kutsatzaileen nahasketaz tratatutako bareei. Beheko geziak esposizio- etaarazte-aldiaren nondik norakoak adierazi ditu.

joan heinean handitu zen (3. Ird.), 18 eta 27.
egunetan bereziki agerian gelditu zena. Araztearen
7 egunak igaro eta gero, VvBSD balioak kontrolen
mailetara itzuli ziren (3. Ird.).
Emaitzak eta eztabaida
174
Cd Kontzentrazioa 18. Eguna Arazte-aldia
Kontrola 102.84±78.16 94.36±36.6
Cd + keroseno 904.672±548.847 168.256±25.6
Keroseno Kontzentrazioa 18. Eguna Arazte-aldia
Kontrola 136.64±36.89 114.24±41.25
Cd + keroseno 415.32±170.14 163.25±52.13
1. Taula: Bare kontrol eta Cd + keroseno pean jarritako bareen liseri-guruinetan neurtutako Cd eta kerosenoaren adierazgarriakdiren hidrokarburoen gehiketaren kontzentrazioak (µg/g ehun-pisu lehor) esperimentuko 18. egunean eta arazte-aldia bukatuondoren.
2. Irudia: Metalen lokalizazio autometalografikoa Arion ater barearen liseri-epitelioan: bare kontroletan (A), Cd + keroseno pean 18egun egondako bareetan (B), Cd + keroseno pean 27 egun egondako bareetan (C eta irudi txikia), eta 7 eguneko arazte-aldia jasozuten bareetan (D). L, liseri-epitelioaren lumena; gezi-buruak, liseri-zelulen lisosometan dauden zilarrezko hauspeakin beltzak,geziak, liseri-epitelioko lumenean dagoen metala. Eskala-marra: 25 µm.

Peroxisomen proliferazioaren adierazle gisa
neurtutako azil-KoA oxidasa (AOX) jarduera Cd +
keroseno nahasketa pean induzitu zen 3. egunean
(4. Ird.). Gerora, AOX jarduera esangarriki jaitsi zen,
kontrol zein esposatutako bareetan, eta ez zen
aurkitu taldeen arteko desberdintasun esangarririk
(4. Ird.).
Bare kontrolen liseri-guruina nagusiki liseri-
zelulez osaturik agertu zen, eta iraizte- zein kaltzio-
zelula urriak taldekaturik agertu ziren (5A Ird.). 18
(5B Ird.) eta 27 (5C Ird.) egunez Cd + keroseno
nahasketaz tratatu ondoren, iraizte-zelulen kopurua
esposizio-denborarekin emendatu zela bistakoa
zen. Modu berean, kaltzio-zelulak ere
nabarmenagoak suertatu ziren, eta gainera itxuraz
hipetrofiaturik ageri ziren,(5B Ird. eta 5C irudi txikia).
Kutsatzaileen peko esposizioaren ondorio gisa,
odol-hodiak erlatiboki ugaritu ziren, eta euren
paretetako Leydig zelulak puztuta agertu ziren.
Zazpi eguneko arazte-aldiaren ondorioz, liseri-
guruinaren epitelioak kontrolen itxura berreskuratu
zuen (5D Ird.), liseri-zelula ugari eta iraizte-zelula
urriez eratuta egonik; kaltzio-zelulek oraindik ere
nolabaiteko hipertrofia erakusten jarraitu zuten
arren.
Aurrekoarekin bat, VvCAL, VvEXC eta VvDIGparametroek, Cd + keroseno nahasketa pean
jartzeak eta ondorengo arazte-aldiak bareen liseri-
guruinaren zelula-moten osaketan aldaketa
esangarriak eragin zituztela adierazi zuten (6. Ird.).
Bare kontroletan VvDC balioek ez zuten aldaketarik
erakutsi, eta modu iraunkorrean 0.8 baliotik gora
mantendu ziren. Kutsatzaileen pean jarritako
bareetan, berriz, parametro horretan jaitsiera
esangarria aurkitu zen esperimentuko 3. egunetik
aurrera, hortik aurrera antzeko balioak (inoiz ez 0.7
baliotik behera) lortu zirelarik. Araztearen ondorioz,
VvDC balioen igoera esangarria behatu zen, bare
kontrolen antzeko balioak eskuratuz (6A Ird.).
Liseri-zelulen batezbesteko tamaina (8 µm-ko
diametro eta 50 µm altuera) kontuan hartuz, gutxi
gorabehera liseri-guruineko 17.8 x 106 µm3 zati
batean (kalkulu estereologikoak burutzeko erabili
den Weibel gratikularen kaltzio-zelula handiena
barneratzea ahalbidetzen duen 50 µm-ko lodiera
arbitrarioaren ebakidurari dagokiona), liseri-zelulen
kopurua 4275±487 (3. eguna) eta 4797±111 (27.
eguna) bitartekoa izan zen bare kontroletan; eta
kutsatzaile pean jarritako bareetan, ordea,
3736±647 baliora jaitsi zen (21. eguna). Zazpi
eguneko arazte-aldiaren ondoren, liseri-zelulen
kopurua 5344 baliora igo zen (6B Ird.). VvEXC eta
iraizte-zelulen kopuruan aldaketa deigarriagoak
antzeman ziren (6C eta 6D Ird.). VvEXC 3. egunetik
aurrera hiru aldiz handiagoa izan zen kutsatzaile
pean jarritako bareetan kontroletan baino, arazte-
Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
175
3. Irudia: .Bare kontrol (zutabe zuria) eta Cd + keroseno peanmantendutako (zutabe grisa) bareen lisosometan neurtutakozilarrezko hauspeakin beltzen dentsitate bolumetrikoa(VvBSD). Segmentu bertikalek desbidazio estandarrak adierazi
dituzte. Goiko asteriskoek taldeen arteko desberdintasunesangarriak adierazi dituzte (p<0.05). Kutsatzaile peanmantendutako bareen arazte-aldian animalia bakarrazegoenez, ez dago desbidazio estandarrik.

aldiaren ondoren balioak berriro ere bare kontrolen
mailetara itzuli zirelarik (6C Ird.). Horrela, iraizte-
zelulen kopurua 243-tik 748-ra emendatu zen,
arazte-aldiaren ondoren 224-ra bueltatu zen
bitartean. Bestalde, VvCAL balioei dagokienez, ez
zen desberdintasun esangarririk behatu (6E eta 6F
Ird.). Orotara, epitelio-zelulen kopurua ez zen
desberdina izan ez esposizio ezta arazte-aldiaren
ondorioz ere (6G Ird.).
BrdU immunohistokimika
BrdU-positiboak ziren bareen liseri-guruineko
zelulen nukleoek kolore gorria erakutsi zuten (7.
Ird.). Liseri-epitelioan batez ere liseri-zelulak
positiboki markaturik aurkitu ziren, eta nahiz eta
iraizte-zelulen lipofuszinen materialak
erreakzionagarritasuna erakutsi, errez bereiz
zitekeen nukleo positibo eta sasipositibotasunaren
artean (7G Ird.). Urdailaren epitelioan ere nukleo
BrdU-positiboak behatu ziren (7E Ird.). Cd +
keroseno nahasketa pean jarritako bareetan, eta
baita arazte-prozesua pairatu zutenetan ere, talde
kontroletan baino nukleo BrdU-positibo gehiago
aurkitu ziren. (7B-D eta F Ird.). BrdU aurkako
antigorputzik gabe inkubatutako tindaketaren ebaki
kontroletan ez zen inolako positibotasunik topatu.
EZTABAIDA
Ikerlan honetako emaitzek, Cd + keroseno
nahasketaz tratatzeak bareen liseri-guruinean
zelula-moten osaketan aldaketak sortarazten
dituela erakutsi dute, aldaketa horiek liseri-zelulen
proportzioa gutxiagotzea eta iraizte-zelulena
handiagotzearekin zerikusia izanik. Gainera,
erantzuna 7 egunetan guztiz itzulgarria dela frogatu
egin da, ikerlan honetako baldintza
esperimentaletan behintzat.
Cd + keroseno pean jarritako bareek, metala eta
kerosenoa metatu zituzten euren ehunetan, liseri-
guruinaren AAS eta GC/MS bitaeteko analisiek
erakutsi zutenez. Lehorreko gastropodoen liseri-
guruineko kutsatzaileen kontzentrazioa
ingurumeneko esposizio-mailekin esangarriki
erlazionatuta dagoena jakina da (Popham &
D'Auria, 1980; Ireland, 1981; Marigómez et al.,
1998a; Beeby & Richmond, 2002; Hamers et al.,
2004). Beraz, esperimentalki trataturiko bareen
liseri-guruinean aurkitutako Cd eta keroseno
kontzentrazioen gorakada esangarriek, aplikatutako
kutsatzailearen esposizio-modua eraginkorra izan
zela egiaztatu zuten. Antzeko esposizio-modua
beste metal toxikoen kasuan aplikatu da aldez
aurretik (Marigómez et al., 1986a; Recio et al.,
1988a; 1988b; Marigómez et al., 1996), baina
kerosenoaren kasuan (WAFa lortzea eta dieta
Emaitzak eta eztabaida
176
4. Irudia: Bare kontrol (zutabe zuria) eta Cd +keroseno peanmantendutako (zutabe grisa) bareen liseri-guruineanneurtutako AOX jarduera. Segmentu bertikalek desbidazioestandarrak adierazi dituzte. Goiko asteriskoak taldeen artekodesberdintasun esangarria adierazi ditu (p<0.05). Kutsatzailepean mantendutako bareen arazte-aldian animalia bakarrazegoenez, ez dago desbidazio estandarrik.

naturalean gehitzea) zeharo hurbilketa berria izan
zen.
Esposizio-biomarkatzaileek ere, kimika
analitikoaren datuak baieztatu zituzten. Cd +
keroseno nahasketaz tratatutako bareen liseri-
zeluletako lisosometan VvBSD esangarriki igo
izana, metal-esposizioaren adierazle moduan har
dezakegu. Era berean, Hg pean egoteak, liseri-
zeluletako lisosometan BSDen presentzia
areagotua dakar (Marigómez et al., 1996). Metalen
metaketa, moluskuen liseri-guruinean oso erantzun
azkarra dena jakina da; izan ere, Cd pean
mantendutako muskuiluetan egun bakar batean
VvBSD balioak esangarriki goratu ziren (Soto et al.,
2002; 1. Kapitulua). Ikerlan honetan, VvBSD
parametroan behatutako aldaketak, denbora eta
dosiaren menpekoak dira. Hortaz, kerosenoaren
presentziak bareen liseri-guruinean Cd-aren
metaketa oztopatu ez zuela ondoriozta daiteke.
Halaber, Cd + bezo(a)pireno (BaP) pean jarritako
muskuiluetan, kutsatzaile organikoaren presentziak
ez zuen Cd-aren metaketa oztopatu liseri-
guruinean (Marigómez et al., 2005; 1. Kapitulua).
Bestalde, keroseno eta Cd nahasketa pean
jarritako bareetan, azil-KoA oxidasa (AOX) jarduera
bare kontroletan baino altuagoa izan zen
esperimentuko 3. egunean, bareak kutsatzaile
organikoen pean (kasu honetan kerosenoa) egon
izanaren seinale (Cancio & Cajaraville, 2000). AOX
jarduera estabulazio denborarekin jaitsi zen,
Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
177
5. Irudia: Arion ater barearen liseri-guruinaren ebakiak, hematoxilina-eosinaz tindatuak: bare kontrolak (A); Cd + keroseno pean18 egunez mantendutako bareak (B); Cd + keroseno pean 27 egunez mantendutako bareak (C eta irudi txikia); eta 7 egunekoarazte-aldia jaso duten bareak (D). Asteriskoak, iraizte-zelulak; geziak, kaltzio-zelulak; gezi-buruak, askatzen ari diren kaltzio-zelulak; L, liseri-azinoen lumena. Eskala-marra: 50 µm (A,-D); 25 µm (irudi txikia).
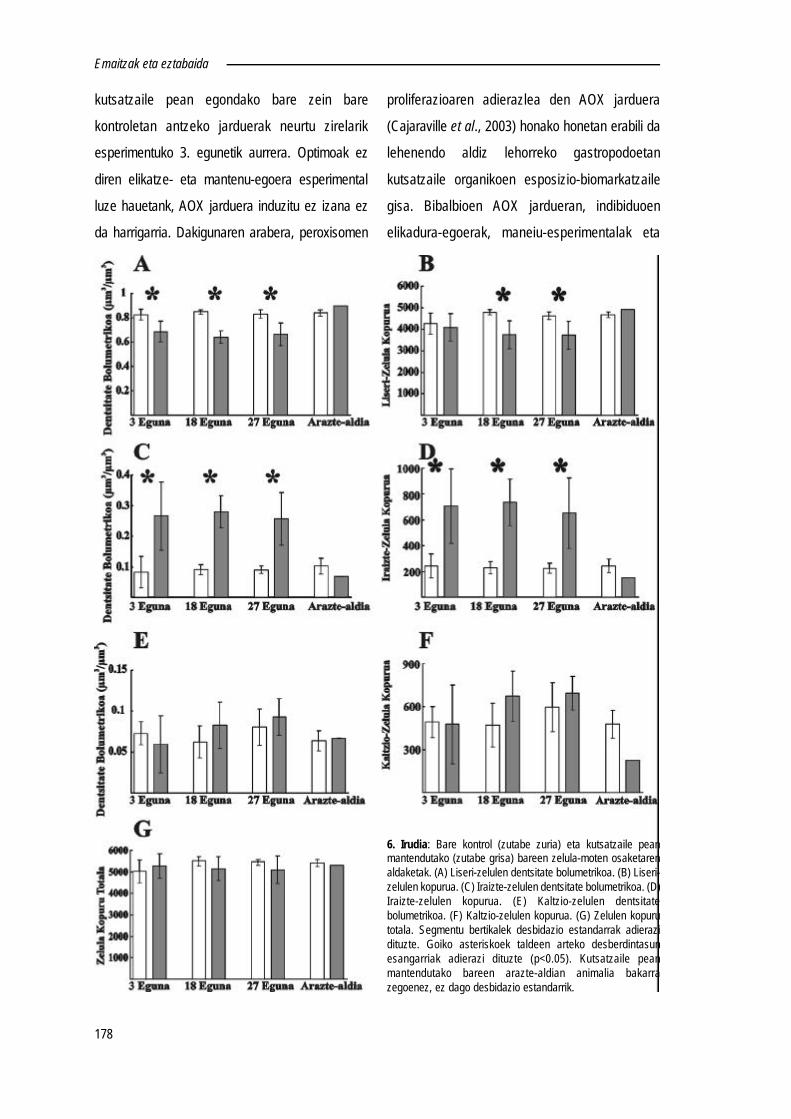
Emaitzak eta eztabaida
178
kutsatzaile pean egondako bare zein bare
kontroletan antzeko jarduerak neurtu zirelarik
esperimentuko 3. egunetik aurrera. Optimoak ez
diren elikatze- eta mantenu-egoera esperimental
luze hauetank, AOX jarduera induzitu ez izana ez
da harrigarria. Dakigunaren arabera, peroxisomen
proliferazioaren adierazlea den AOX jarduera
(Cajaraville et al., 2003) honako honetan erabili da
lehenendo aldiz lehorreko gastropodoetan
kutsatzaile organikoen esposizio-biomarkatzaile
gisa. Bibalbioen AOX jardueran, indibiduoen
elikadura-egoerak, maneiu-esperimentalak eta
6. Irudia: Bare kontrol (zutabe zuria) eta kutsatzaile peanmantendutako (zutabe grisa) bareen zelula-moten osaketarenaldaketak. (A) Liseri-zelulen dentsitate bolumetrikoa. (B) Liseri-zelulen kopurua. (C) Iraizte-zelulen dentsitate bolumetrikoa. (D)Iraizte-zelulen kopurua. (E) Kaltzio-zelulen dentsitatebolumetrikoa. (F) Kaltzio-zelulen kopurua. (G) Zelulen kopurutotala. Segmentu bertikalek desbidazio estandarrak adierazidituzte. Goiko asteriskoek taldeen arteko desberdintasunesangarriak adierazi dituzte (p<0.05). Kutsatzaile peanmantendutako bareen arazte-aldian animalia bakarrazegoenez, ez dago desbidazio estandarrik.

ugalketa-zikloak eragina izan dezakete (Cancio et
al., 1999; Cancio & Cajaraville, 2000; Orbea et al.,
2002). Bareetan, ugal-garapenaren aldaketak aste
gutxiren buruan gerta daitezke (Marigómez et al.,
1986; Zubiaga, 1986) eta beraz, uztailan zehar,
feminizazioaren aurretik, non gameto arrak ia guztiz
garatuak dauden, AOX jardueran aldaketak
gertatzea ezin daiteke baztertu. Ikerlan honetan
gertatu den bezala, Cd eta BaP nahasketaren pean
mantendutako muskuiluetan, AOX jarduera soilik
esposizio-egun baten ondoren erantzuteko gai izan
zen, momentu horretatik aurrera, AOX jarduerak
baxuak izan ziren, muskuilu tratatui eta muskuilu
kontrolen artean berdintsuak izan zirelarik (Orbea et
Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
179
7. Irudia: Arion ater barearen liseri-azinoen epitelioan eta urdaileko-epitelioan egindako BrdU immunohistokimika: Bare kontrola(A); Cd + keroseno pean 27 egunez egondako barea (B); 7 eguneko arazte-aldia jaso duten bareak (C-G). Gezi-buruak, nukleoBrdU-positiboak; zirkuluak, lipofuszinek sortutako marka sasipositiboak. Eskala-marra: 50 µm (A-E); 25 µm (F & G).

al., 2002). Era berean, BaP pean jarritako bareetan,
kutsatzaile kimikoen esposizio-biomarkatzaile gisa
erabilitako benzo(a)pireno hidroxilasa (BPH)
jarduera, soilik esperimentuko 3. egunean induzitu
zen, baina ez esposizio-denbora luzeagoetan
(Hamers et al., 2004). Halaber, BaP pean jarritako
muskuiluetan, esposizio osteko egun gutxitara
bakarrik induzitzen da BHP jarduera (Okay et al.,
2000). Beraz, nahiz eta esposizioaren 3. egunetik
aurrera erregistratutako AOX jarduera baxua izan,
AOXren indukzio azkarrak, ikerlan honetan
erabilitako kerosenoaren esposizio-modua
eraginkorra izan zela argiro erakutsi du.
Elikatze-jarduera eta hazkuntza, neurri osagarri
gisa erregistratu ziren esperimentuan zehar. Bare
zein barraskiloetan, kutsatzaile pean egoteak
elikatze-jardueran eta hazkuntzan beherakada
esangarriak sor ditzake (Russell et al., 1981;
Marigómez et al., 1986b; Laskowski & Hopkin,
1996; Swaileh & Ezzughayyar, 2000; 5. Kapitulua).
Hortaz, metalez (Cu, Zn eta Cd) kutsatutako
tokietatik hartutako bareek, kutsatugabeko tokietan
hartutako bareekin alderatuz, gorputz osoaren eta
liseri-guruinaren pisu lehorrean %50-eko
murrizketa erakutsi zuten (Marigómez et al.,
1998a). Efektu horri dagokionez, eragite-maila
metalez-metal alda daitekeela esan baharra dago.
Helix aspersa barraskiloan behatutakoaren
arabera, Cd-a, Zn-a eta Pb-a baino toxikoagoa da
elikatze-jarduera, hazkuntza eta heriotza-tasari
dagokienez (Laskowski & Hopkin, 1996).
Kutsatzaile organikoek ere, elikatze-jarduera zein
hazkuntzaren gainean eragina dute, laborategian
BaP pean jarritako bareek erakutsi zutenez
(Hamers et al., 2004). BaP pean jarritako bareetan,
elikatze-jarduera modu azkarrean txikiagotu zen,
baina gorputzaren pisuan aldaketak soilik
hirugarren egunetik aurrera gertatu ziren (Hamers
et al., 2004). Modu berean, ikerlan honetan Cd +
keroseno pean jarritako bareetan, elikatze-jarduera
aste batetik aurrera txikiagotu zen, hazkuntzaren
murrizpena bakarrik 3. astetik aurrera ikusgarria
izan zen bitartean.
Metal pean jarritako lehorreko gastropodoen
liseri-guruinaren xafla basalak loditu egiten dira,
liseri-zelulak lehendabizi bakuolizatuak ageri eta
beranduago urriagoak bilakatzen dira, kaltzio-
zelulak hipertrofiatu egiten dira, kaltzio-zelulak eta
iraizte-zelulak erlatiboki ugariagoak bilakatzen dira,
eta odol-zelulak eta ehun konektiboko zelula
migratzaileak liseri-azinoen inguruan metatzen dira
(Marigómez et al., 1986a; 1986b; 1996; 1998a;
Recio et al., 1988a; 1988b; Ireland & Marigómez;
1992, Chabikovsky et al., 2004; Snyman et al.,
2005; 6. Kapitulua). Oro har, Cd + keroseno pean
jarritako bareetan irudi beretsua ikusi da,
erantzuna, beraz, orokorra dela dirudi, eta ez metal-
kutsaduraren espezifikoa. Horrela, emaitza horiek,
behatutako aldaketak kutsatzaile espezifiko batek
sorterazitako erantzunak baino gehiago, estres-
egoera orokorraren aurrean gauzatutako
erantzunak direla konfirmatu dute, aldez aurretik
Marigómez et al.-ek, (1996) proposatu bezala. Hala
ere, kutsatzaile organikoek beraien kabuz efektu
berdinak, modu kualitatiboan behintzat, sortarazten
ote dituzten zehazteko ikerlan gehiagoren beharra
dago.
Emaitzak eta eztabaida
180

Efektu horien artean, deigarrienak liseri-
guruinaren epitelioan behatutako zelula-moten
osaketari dagozkionak dira. Aldez aurretik
laborategian egindako ikerlanek agerian utzi
dutenez, moluskuen liseri-guruinean zelula-moten
osaketan aldaketa esangarriak 3 eta 21 egun
bitartean, espeziearen, kutsatzaile, dosi eta
esposizio-moduaren arabera eta baita ingurumen-
egoeraren arabera, gerta daitezke (Marigómez et
al., 1996; 1 eta 4. Kapituluak). Ikerlan honetan
erabilitako esposizio-modu bera erabiliz, Cu eta Hg-
z tratatutako bareetan, liseri-epitelioa osatzen duten
zelula-moten proportzio erlatiboan aldaketak
behatu dira, iraizpen-jarduera eta estrusio apokrino
areagotuen ondorioz (Marigómez et al., 1986b;
1996). 1000 µg Hg/g janari dosiaz 27-30 egunez
elikatutako bareetan, liseri-zelulen kopurua izugarri
jaitsi zen, liseri-epitelioa funtsean kaltzio-zelulez eta
iraizte-zelulez osaturik geratuz (Marigómez et al.,
1996). Cd kontzentrazio subletalen pean
mantendutako magurioetan antzeko prozesua
deskribatu da (Marigómez et al., 1990). Ikerlan
honetan, VvDIG eta VvEXC modu esangarrian
aldatu ziren, Cd + keroseno esposizio zein 7
eguneko arazte-aldiaren eraginez. VvCAL balioak,
ordea, ez ziren esangarriki aldatu. Horrela, Cd +
kerosenoz tratatutako bareetan, VvDIGesperimentuko hirugarren egunetik aurrera jaitsi
zen, eta 7 eguneko arazte-aldiaren ondoren berriro
ere bare kontroletan neurtutako balioetara itzuli
zen. VvEXC balioen aldaketak nabarmenagoak
izan ziren, 3. tratamendu-egunetik aurrera
tratatutako bareetan, kontroletan baino hiru aldiz
handiagoak izatera iritsiz baina, VvDIG-ren kasuan
bezalaxe, arazte-aldiaren ondoren kontrol-mailetara
itzuli ziren. VvCAL balioei dagokienez, ez dirudi
tratamenduak kaltzio-zelulen proportzioaren gain
eragin esangarria izan zuenik. Aitzitik, Cu, Hg eta
metaleen nahasketan (Cu, Cd, Zn) peko
esposizioek kaltzio-zelulen dentsitate erlatiboaren
emendioa eragiten dutela deskribatu da
(Marigómez et al., 1986b; 1996; 1998). Ikerlan
honetan, VvCAL modu esangarrian aldatu ez zen
arren, itxurazko kaltzio-zelulen presentzia
erlatiboaren igoera susma zitekeen, mikroskopioan
egindako behaketa zuzenak kontuan hartuz,
behintzat. Horrekin bat, Cd + kerosenoz tratatutako
VvCAL balioen igoerako joera ageri zen,
estatistikoki esangarria izan ez arren. Edozein
kasutan, kaltzio-zelulen proportzioan behatutako
igoerak iraizte-zeluletan behatutakoen aurrean
garrantzi gutxikoak izango ziren, beste
moluskuetan ez bezala (Cajaraville et al., 1990a;
Marigómez et al., 1990; 1998b; 2005; Soto et al.,
2002; 1 eta 4. Kapituluak).
Liseri-zelulen batezbesteko tamaina kontuan
hartuta, beraien kopurua (DCN) 17.8 x 106 µm3-ko
liseri-guruineko ehun-zati arbitrario batetarako
(bolumen unitate arbitrarioa) kalkulatu zen. Bare
kontroletan, DCN ez zen esangarriki aldatu
esperimentuak iraun bitartean, batezbesteko balioa
unitate arbitrario horretan 4500 zelula baino apur
bat altuagoa izanik. Cd + keroseno tratamenduaren
eraginez, 750 (%17) liseri-zelula inguru galdu ziren,
7 eguneko arazte-aldiaren ondorioz unitate
arbitrario bakoitzeko 1500 liseri-zelula berri agertuz.
Era berean, Cd pean jarritako magurioetan 1850
(%13) liseri-zelula galdu ziren, 7 eguneko arazte-
Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
181

aldiaren ondorioz 2600 liseri-zelula berri sortu
zirelarik 17.8 x 106 µm3-ko liseri-guruineko unitate
arbitrarioko (4. Kapitulua). Cd pean jarritako
magurioetan, VvBAS eta zelula basofilikoen
bolumen absolutua handitu zen, baina ez zen
aldaketarik behatu zelula basofilikoen kopuruan (4.
Kapitulua), ikerlan honetan Cd + keroseno pean
jarritako bareetan behatutako emaitzekin bat
datorrena. Bestalde, Cd + keroseno pean 27
egunez mantendutako bareetan 500 (%300) iraizte-
zelula berri agertu ziren bolumen unitate
arbitrarioko, 7 eguneko arazte-aldiaren ondorioz
galdu zirenak, hain zuzen. Aipagarria da, muskuilu
eta magurioetan gertatu ez bezala, epitelio-zelulen
kopuru totala kutsatzaileen zein arazte-aldiaren
eraginez ez zela aldatu. Iraizte-zelula, zelula-mota
desberdina baino bizi-zikloaren amaieran dagoen
liseri-zelula dela proposatu da (Porcel et al., 1996;
Dimitriadis & Konstantinidou, 2002). Liseri- eta
iraizte-zelulen proportzio erlatiboek kontrako
zentzuan mugitzen dira baraualdian eta
kutsatzaileen esposizio pean (Porcel et al. 1996;
Marigómez et al. 1998; Dimitriadis & Konstantinidou
2002; 6. Kapitulua). Testuinguru horretan, ikerlan
honen emaitzek, iraizte-zelulak liseri-zelula “zahar”
edo “erabiliak” direnaren hipotesia sendotzen dute,
liseri-zelulen eta iraizte-zelulen kopuru absolutuek,
tratamendu eta arazte-aldiaren eraginez kontrako
zentzua jarraitzen baitute. Metalek, bareen liseri-
guruineko zelulen zahartzapen goiztiarra eragin
dezaketela aurretik ere proposatu da (Marigómez et
al., 1986b), Cu pean (dietan zein uretango
esposizioa) jarritako bareak aztertuz, hain zuzen
ere. Orokorki, lehorreko gastropodoen liseri-
guruinean ematen diren iraizte-jarduera eta estrusio
apokrino areagotuek, alterazio lisosomikoek eta
lipofuszinen metaketek, kutsatzaileen eragin
orokorren adierazgarritzat har ditzakegu
(Marigómez et al., 1996; 1998; Chandran et al.,
2005). Kuprez trataturiko bareetan, jarduera
fisiologikoaren fase desberdinak histokimikoki
determinatu ondoren, xurgapena, liseriketa eta
jariapena tratamenduko lehenengo egunetan
bizkortu ziren, fase bakoitzak gutxiago irauten
zuelarik; 300 eta 1000 µg Hg/g janari tratatutako
bareetan 15. egunetik aurrera, berriz, epitelioa
atrofiko edo, behintzat, ez horren aktibo bilakatu
zen (Marigómez et al., 1986b). Hiperaktibitate
horrek, zergaitik Cd + keroseno tratamenduaren 3.
egunetik aurrera liseri-zelula gehiago iraizte-zelula
bilakatzen joatea azalduko luke. Ondorioz, nahiz
eta magurioetan kutsatzaileek liseri-zelulen galera
netoa sortarazi, konplexuagoa den bareen liseri-
guruinean badirudi liseri-zelulak, galdu beharrean,
iraizte-zelulak bilakatzen direla. Hortaz, bareen
liseri-guruinean zelula-moten osaketan aldaketak
nabarmenago irudikatzeko, iraizte-zelula/liseri-
zelula ratioa (ECR; ingeleraz, Excretory to Digestive
Cell Ratio) parametro sentikorrago gisa proposatu
dugu, soilik liseri-zelula eta zelula basofilikoz
osatutako magurioen liseri-guruinean
proposatutako DCL parametroaren baliokidetzat
har genezakeena (4. Kapitulua).
Azkenik, zelula proliferatzaileak BrdU
immunohistokimika bitartez identifikatu ziren liseri-
guruineko epitelioan. Hainbat saiakera burutu
ondoren, fosfatasa alkalinoa (AlP; Chabicovsky et
al., 2003) aukeratu zen BrdU/anti-BrdU konplexuak
Emaitzak eta eztabaida
182

ikustarazteko, aldez aurretik magurio eta
muskuiluetan erabilitako diamino benzidinak
bareen liseri-guruinean pikor zitoplasmatiko eta
lipofuszina ugariekin erreakzionatzen baitzuen
(Zaldibar et al., 2004; 3 eta 4. Kapituluak). Hala ere,
iraizte-zelulen bakuoloen lipofuszinen zenbait
substantzia AlPrekin erreakzionagarriak badira.
Edonola ere, lipofuszina sasipositibook, liseri-
guruin eta urdaileko epitelioan behatutako nukleo
BrdU-positiboetatik zailtasun handirik gabe bereiztu
genituen. Muskuilu eta magurioetan gertatu zen
moduan, bareen kasuan ere, urdaileko epitelioan
liseri-guruineko epitelioan baino BrdU-markaketa
nabariagoa topatu dugu (Zaldibar et al., 2004; 4.
Kapitulua). Liseri-guruineko epitelioan, BrdU-
markaketa batez ere liseri-zeluletan behatu zen,
kaltzio-zelula urrietan markaketa ahula behatu zen
bitartean. Iraizte-zelulen nukleoan ez zen BrdU-
markaketa positiborik aurkitu. Hala ere,
bakuoloetako lipofuszinen markaketek nukleo
BrdU-positiboak itzal zitzaketeenez, iraizte-zelulak
proliferatzeko gai direnentz egiaztatzeko, behaketa
zehatzagoak egitea ezinbestekoa izango litzateke.
Edonola ere, Cd + keroseno tratamenduak eta
ondorengo arazte-aldiak, bareen liseri-zelulen
proliferazio-intentsitatearen areagotzea eragin dute,
ikusi dugunez. Izan ere, tratatutako zein araztutatko
bareetan nukleo BrdU-positibo gehiago daude talde
kontroleko bareetan baino. Era berean,
magurioetan, Cd-ak liseri-zelulen proliferazioa
areagotu zuen, eta proliferazio-tasa altu hori 7
eguneko arazte-aldiaren ondoren ere antzeko
mailan mantendu zen (4. Kapitulua).
Kutsatzaileek zelulen proliferazioa eragin
dezaketena jakina da (Beyersmann & Hechtenberg,
1997; Habeebu et al., 1998; Piechotta et al., 1999;
Chabicovsky et al., 2004). Magurioetan, Cd pean
egoteak, arazte-prozesuei aurre egin ahal izateko,
areagotutako liseri-zelulen berriztapena dakar,
zelulen heriotza eta proliferazioaren arteko balantze
gisa interpreta daitekeena (4. Kapitulua). Bareetan,
areagotutako liseri-zelulen proliferazioa iraizte-
zelula faseranzko progresio areagotuarekin batera
gertatu zela dirudi. Zentzu horretan, Cu-meatze
abandonatu batean hainbat belaunaldiz (kutsadura
kronikoa) bizi izan diren bareen liseri-guruinean,
iraizte-zelulen kopuru erlatibo altuaren berri eman
da; non liseri-zelulek pikor zitoplasmatikoen
iraizpenean bortizki baitziharduten eta oso
bakuolizaturik ageri baitziren (Marigómez et al.,
1998a).
Aldez aurretik metal pean jarritako magurio eta
muskuiluetan deskribatu den moduan (1 eta 4.
Kapituluak), Cd + keroseno nahasturaz trataturiko
bareetan suertatutako zelula-moten osaketaren
aldaketak ere, 7 eguneko arazte-aldiaren ostean
itzulgarriak izan ziren. Liseri-zelulen proliferazioa
tratatu eta araztutako bareen artean desberdina ez
dela dirudienez, behatutako leheneratzea liseri-
zeluletatik iraizte-zeluletaranzko progresioa
moteltzean datzala pentsa genezake, eta hori
epitelio-jarduera kutzatsailea ez egotean moteldu
izanarekin bat letorke (Marigómez et al., 1986b;
1998a; Chandran et al., 2005; 4. Kapitulua). Beraz,
ingurumen-baldintz jakin bakoitzari zelula-moten
osaketa bereizgarri bat eslei lekioke.
Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
183

Liseri-zelulen kopuru erlatiboaren murrizpen eta
kaltzio-zelulen, eta batez ere, iraizte-zelulen kopuru
erlatiboaren emendioaren berri, metalez
kutsatutako ingurunetan bizitako lehorreko bareen
liseri-guruinean ere eman da (Marigómez et al.
1998a). Ikerlan honek, zelula-moten osaketaren
aldaketok itzulgarriak direla erakutsi du,
laborategiko baldintzetan behintzat. Dena den,
ingurumen naturaletan animaliak beren bizitza
osoan zehar metal pean kronikoki egon daitezke,
eta beren erantzunak deskribatutakoen antzekoak
liratekeenentz argitu beharrean gaude oraindik ere
(Wang & Rainbow, 2005), hurrengo kapituluan
jorratuko denez.
Azkenik, zelula-moten osaketan metal-
esposizioak eragindako aldaketek, BSD-hedapena
gisako esposizio-biomarkatzaileen gainean eragina
izan ditzaketeenaren berri eman da muskuiluen
kasuan (Soto et al., 2002; 1. Kapitulua). Aitzitik, ez
dirudi Cd + keroseno esposizio eta ondoko arazte-
aldian zehar bareetan suertatutako aldaketek, BSD-
hedapena parametroaren gainean eragina izan
zutenik, VvBSD aldaketak dosi eta denboraren
menpekoak baitziren. Arrazoia, magurioetan
gertatzen den moduan, bareen liseri-guruinaren
epitelioko ezaugarri berezietan datza, non
lisosomen barneko metalen neurketa
mikroskopikoak liseri-zelulen gainean zuzenean
egin baitaitezken (4. Kapitulua), liseri-zelula
kopurua basala izanik ere.
BIBLIOGRAFIA
Beeby A, Richmond L (2002) Evaluation of Helix aspersa as a
sentinel for mapping metal pollution. Ecol. Indt. 1:261-270.
Beyersmann D, Hechtenberg S (1997) Cadmium, gene regula-
tion, and cellular signalling in mammalian cells. Toxicol.
Appl. Pharmacol. 144:247-261.
Cajaraville MP, Díez G, Marigómez I, Angulo E (1990a)
Responses of basophilic cells of the digestive gland of
mussels to petroleum hydrocarbon exposure. Dis. Aquat.
Org. 9:221-228.
Cajaraville MP, Marigómez I, Angulo E (1990b) Short-term toxic
effects of 1-naphthol on the digestive gland-gonad complex
of the marine prosobranch Littorina littorea (L.): a light
microscopic study. Arch. Environ. Contam. Toxicol. 19:17-
24.
Cajaraville MP, Cancio I, Ibabe I, Orbea A (2003) Peroxisome
proliferation as a biomarker in environmental pollution
assessment. Micr. Res. Tech. 61:191-202.
Cancio I, Ibabe A, Cajaraville MP (1999) Seasonal variation of
peroxisomal enzyme activities and peroxisomal structure in
mussels Mytilus galloprovincialis and its relationship with
the lipid content. Comp. Biochem. Physiol. C 123:135-144.
Cancio I, Cajaraville MP (2000) Cell biology of peroxisomes
and their characteristics in aquatic organisms. Int. Rev.
Cytol. 199:201-293.
Chabicovsky M, Niederstätter H, Thaler R, Hödl E, Parson W,
Rossmanith W, Dallinger R (2003) Localization and quanti-
fication of Cd- and Cu-specific metallothionein isoform
mRNA in cells and organs of the terrestrial gastropod Helix
aspersa. Toxicol. Appl. Pharmacol. 190:25-36.
Chabicovsky M, Klepal W, Dallinger R (2004) Mechanisms of
cadmium toxicity in terrestrial pulmonates: Programmed
cell death and metallothionein overload. Environ. Toxicol.
Chem. 23:648-655.
Chandran A, Sivakumar AA, Mohandass S, Aruchami M (2005)
Effect of cadmium and zinc on antioxidant enzyme activity
in the gastropod, Achatina fulica. Comp. Biochem. Physiol.
C 140:422-426.
Danscher G (1984) Autometallography. A new technique for
light and electron microscopic visualization of metals in bio-
logical tissues (gold, silver, metal sulphides and metal sele-
nides). Histochemistry 81:331-335.
Dimitriadis VK, Konstantinidou V (2002) Origin of excretory
cells in the digestive gland of the land snail Helix lucorum.
Malacologia 44:145-151.
Habeebu SSM, Liu J, Klaassen CD (1998) Cadmium-induced
apoptosis in mouse liver. Toxicol. Appl. Pharmacol. 153:48-
58.
Emaitzak eta eztabaida
184

Zelula-moten ordezkapena bareen liseri-guruinean: laborategi-ikerketa
185
Hamers T, Kalis EJJ, Van den Berg JHJ, Maas LM, Schooten
FJV, Murk AJ (2004) Applicability of the black slug Arion
ater for monitoring exposure to polycyclic aromatic hydro-
carbons and their subsequent bioactivation into DNA bin-
ding metabolites. Mut. Res. 552:219-233.
Ireland MP (1981) Uptake and distribution of cadmium in the
terrestrial slug Arion ater (L). Comp. Biochem. Physiol. A
73:855-858.
Ireland MP, Marigómez I (1992) The influence of dietary cal-
cium on the tissue distribution of Cu, Zn, Mn and P and his-
tological changes in the digestive cells in the snail Achatina
fulica Bowdich. J. Moll. Stud. 58:157-168.
Laskowski R, Hopkin SP (1996) Effect of Zn, Cu, Pb, and Cd on
fitness in snails (Helix aspersa). Ecotoxicol. Environ. Saf.
34:59-69.
Lowe DM, Moore MN, Clarke KR (1981) Effects of oil in the
digestive cells in mussels: quantitative alterations in cellu-
lar and lysosomal structure. Aquat. Toxicol. 1: 213-226.
Lowe DM, Clarke KR (1989) Contaminant-induced changes in
the structure of the digestive epithelium of Mytilus edulis.
Aquat. Toxicol. 15:345-358.
Lowry OH, Rosenbrough NJ, Farr AL, Randall RJ (1951)
Protein measurement with the Folin phenol reagent. J.
Biochem. Chem. 193:265-275.
Marigómez I, Angulo E, Moya J (1986a) Copper treatment of
the digestive gland of the slug Arion ater L. 1. Bioassay
conduction and histochemical analysis. Bull. Environ.
Contam. Toxicol. 36:600-607.
Marigómez I, Angulo E, Moya J (1986b) Copper treatment of
the digestive gland of the slug Arion ater L. 2.
Morphometrics and histophysiology. Bull. Environ. Contam.
Toxicol. 36:608-615.
Marigómez I, Cajaraville MP, Angulo E (1990) Histopathology of
the digestive gland-gonad complex of the marine proso-
branch Littorina littorea exposed to cadmium. Dis. Aquat.
Org. 9: 229-238.
Marigómez I, Soto M, Kortabitarte M (1996) Tissue-level bio-
markers of biological effect of mercury on sentinel slugs,
Arion ater. Arch. Environ. Contam. Toxicol. 31:54-62.
Marigómez I, Kortabitarte M, Dussart, GBJ (1998a) Tissue-
level biomarkers in sentinel slugs as cost-effective tools to
assess metal pollution in soils. Arch. Environ. Contam.
Toxicol. 34:167-176.
Marigómez I, Cajaraville MP, Soto M, Lekube X (1998b) Cell-
type replacement, a succesful strategy of molluscs to adapt
to chronic exposure to pollutants. Cuad. Invest. Biol.
18:431-435.
Marigómez I, Soto M, Cajaraville MP, Angulo E, Giamberini L
(2002) Cellular and subcellular distribution of metals in
molluscs. Micr. Res. Tech. 56:358-392.
Marigómez I, Izagirre U, Lekube X (2005) Lysosomal
enlargement in digestive cells of mussels exposed to
cadmium, benzo[a]pyrene and their combination. Comp.
Biochem. Physiol. C 141:188-193.
Müller S (1984) Lindanuntersuchungen im Aquatischen und
Terrestrischen Bereich in Bilbao (Nordostspanien).
Lizentziatura-Tesia. Universität Karlsruhe. 93 orr.
Orbea A, Ortiz-Zarragoitia M, Cajaraville MP (2002) Interactive
effects of benzo(a)pyrene and cadmium and effects od
di(2-ethylexyl) phtalate on antioxydant and peroxisomal
enzymes and peroxisomal volume density in digestive
gland of mussel Mytilus galloprovincialis Lmk. Biomarkers
7:33-48.
Okay OS, Donkin P, Peters LD, Livingstone DR (2000) The role
of algae (Isochrysis galbana) enrichment on the
bioaccumulation of benzo[a]pyrene and its effects on the
blue mussel Mytilus edulis. Environ. Poll. 110:103-113.
Piechotta G, Lacorn M, Lang T, Kammann U, Simat T, Jenke
HS, Steinhart H (1999) Apoptosis in dab (Limanda liman-
da) as possible new biomarker for anthropogenic stress.
Ecotox. Environ. Saf. 42:50-56.
Popham JD, D'Auria M (1980) Arion ater (Mollusca: Pulmonata)
as an indicator of terrestrial environmental pollution. Water
Air Soil Poll. 14:115-124.
Porcel D, Bueno JD, Almendros A (1996) Alterations in the
digestive gland and shell of the snail Helix aspersa Muller
(Gastropoda, Pulmonata) after prolonged starvation.
Comp. Biochem. Physiol. A 115:11-17.
Rasmussen LPD, Hage E, Karlog O (1983) Light and electron
microscopic studies of the acute and chronic toxic effects
of N-nitroso compounds on the marine mussel, Mytilus
edulis (L) II. N-methyl-N-nitro-N-nitrosoguanidine Aquat.
Toxicol. 3:301-311.
Recio A, Marigómez I, Angulo E, Moya J (1988a) Zinc treatment
of the digestive gland of the slug Arion ater L. 1. Cellular
distribution of zinc and calcium. Bull. Environ. Contam.
Toxicol. 41:858-864.
Recio A, Marigómez I, Angulo E, Moya J (1988b) Zinc treatment
of the digestive gland of the slug Arion ater L. 2. Sublethal
effects at the histological level. Bull. Environ. Contam.
Toxicol. 41:865-871.

Runham NW, Hunter PJ (1970) Terrestrial Slugs. Hutchinson
University Library, London 184 orr.
Russell LK, DeHaven JI, Botts RP (1981) Toxic effects of cad-
mium on the garden snail (Helix aspersa). Bull. Environ.
Contam. Toxicol. 30:245-251.
Small GM, Burdett K, Connock MJ (1985) A sensitive spectro-
photometric assay for peroxisomal Acyl-CoA oxidase.
Biochem. J. 227:205-210.
Snyman RG, Reinecke AJ, Reinecke SA (2005) Quantitative
changes in the digestive gland cells of the snail Helix
aspersa after exposure to fungicide copper oxychloride.
Ecotox. Environ. Saf. 60:47-52.
Soto M, Marigómez I (1997b) Metal bioavailability assessment
in “mussel-watch” programmes by automated image analy-
sis of autometallographical black silver deposits (BSD) in
digestive cell lysosomes. Mar. Ecol. Prog. Ser. 156:141-
150.
Soto M, Quincoces I, Lekube X, Marigómez I (1998)
Autometallographical procedure for the localization of trace
metals in molluscan tissue by lighht microscopy. J.
Histotech. 21:123-127.
Soto M, Zaldibar B, Cancio I, Taylor MG, Turner M, Morgan AJ,
Marigómez I (2002) Subcellular distribution of cadmium
and its cellular ligands in mussel digestive gland cells as
revealed by combined autometallography and X-ray micro-
probe analysis. Histochem. J. 34:273-280.
Sumner AT (1965) The cytology and histochemistry of the
digestive gland cells of Helix. Q. J. Microsc. Sci. 106:173-
192.
Swaileh KM, Ezzughayyar A (2000) Effects of dietary Cd and
Cu on feecing and growth rates of the landsnail Helix
engaddensis. Ecotox. Environ. Saf. 47:253-260.
Thompson RJ, Ratcliffe NA, Bayne BL (1974) Effects of starva-
tion on structure and function of the digestive gland of the
mussel (Mytilus edulis L) J. Mar. Biol. Assoc. U.K. 54:699-
712.
Wang WX, Rainbow PS (2005) Influence of metal exposure
history on trace metal uptake and accumulation by marine
invertebrates. Sci. China C. Life Sci. 48:110-117.
Weibel ER (1979) Stereological Methods. Vol. 1. Academic
Press. London 415 orr.
Widdows J, Bakke T, Bayne BL, Donkin P, Livingstone DR,
Lowe DM, Moore MN, Evans SV, Moore SL (1984)
Responses of Mytilus edulis on exposure to the WAF of
North Sea oil. Mar. Biol. 67:15-31.
Zaldibar B, Cancio I, Marigómez I (2004) Circatidal variation in
epithelial cell proliferation in the mussel digestive gland
and stomach. Cell Tiss. Res. 318:395-402.
Zubiaga AM (1986) Histofisiología dcel aparato reproductor de
Arion subfuscus (Draparnaud, 1805) (Gastropoda,
Stylomatophora). Doktoradutza-Tesia. UPV/EHU. 316 orr.
Emaitzak eta eztabaida
186

Zelula-moten ordezkapena bareen liseri-guruinean eta bere eragina biomarkatzaileengainean: metalez kutsatutako eta kutsatu gabekotokien artean lekuzaldatze-esperimentuak
Laburpena: Zelula-moten ordezkapena (CTR; ingeleraz, cell type replacement) estresatutakomoluskuen liseri-guruineko epitelioan ematen den fenomeno orokorra da. Aldez aurretik laborategianegindako ikerlanek, CTRk biometatze-prozesu eta biomarkatzaileen neurketen gainean eragina duelaerakutsi dute. Lan honen helburua CTR prozesua kutsadura metaliko pean kronikoki egondako bareetanitzulgarria ote den eta biometaketa-parametro eta esposizio- zein efektu- biomarkatzaileen gainean CTRkzer nolako eragina duen aztertzea da. Horretarako, Arion ater bareak, abandonatutako zink meatze batetik(Karrantza) eta kutsatugabeko toki batetik (Bakio) jaso ziren eta leku batetik bestera 3, 10 eta 28 egunezlekuzaldatu ziren. Metal-zama, absortzio atomikozko espektrofotometria (AAS) erabiliz kalkulatu zen.Lisosomen egitura-aldaketa, zelula-moten dentsitate bolumetrikoa, autometalografiko hauspeakinak etametalotioneinen mailak esposizio- eta efektu-biomarkatzaile gisa neurtu ziren. Ikerlan honen emaitzek,metal-kutsadura kroniko pean egondako bareen liseri-guruinak, laborategian hainbat astez metalen peanjarritako bareen liseri-guruinen antzekoak direla erakutsi dute. Hala ere, kronikoki kutsatutako liseri-guruinak, laborategian epe laburrean kutsatutakoen bareenak ez bezala, kutsadura-iturria etenez gero ezdira erantzunkorrak, edo behintzat, erantzuteko gaitasun murritzagoa dute. Bestalde, kutsatugabeko tokitikmeatzera lekuzaldatutako bareek, 3 eguneko lekuzaldatze ostean jada laborategian metal pean jarritakobareek bezala erantzun zuten; beraz, lan honek, erabilitako prozedura analitikoak eta biomarkatzaileaklurzorutan noizbehinkako metal-esposizioak detektatzeko egokiak direla demostratu du.
Gako-hitzak: lekuzaldatzea, zelula-moten ordezkapena, metalak, kutsadura kronikoa, liseri-guruina,bareak.
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
187

Cell-type replacement in the digestive gland of slugs and its influence inbiomarkers after transplantation between clean and metal-polluted sites
Abstract: Cell-type raplacement (CTR) is a general phenomenon that takes place in the digestive glandepithelium of stressed molluscs. Previous laboratory studies have demontrated that CTR influencesbioaccumulation processes and biomarker measurements. The aim of this work is to determine wether CTRis a reversible process is slugs exposed to chronic metal pollution and to investigate the influence of CTRin metal accumulation parametres and in effect and exposure biomarkers. For this purpouse slugs, Arionater, were transplanted from an abandoned Zn mine (Karrantza) to a non-polluted site (Bakio) and the otherway round for 3, 10 and 28 days. Metal burdens were measured by atomic absorption spectrophotomtry(AAS). Lysosomal structural changes, volume density of cell-types, autometallographical deposits andmetallothionein levels were calculated as exposure and effect biomarkers. The results of this study, showedthat the digestive gland of slugs chronically exposed to metal pollution is similar to the digestive gland ofslugs exposed to metals for some weeks under laboratory conditions. However, chronically polluteddigestive glands, are not responsible, or at least, are less responsible than short-term exposed ones whenthe pollution source disappears. On the other hand, slugs tranplanted from the reference site to the minerespond simmilarly to slugs exposed to metals in the laboratory after 3 days of transplantation, whichindicates that the analitical procedure and biomarkers employed in this work are able to detect eventualmetal exposures in soils.
Key Words transplant, cell-type replacement, metals, chronic pollution, digestive gland, slugs.
Sustitución de tipos celulares en la glándula digestiva de limacos y suinfluencia sobre biomarcadores después de un transplante entre un lugarlimpio y uno con contaminación metálica
Resumen El recambio de tipos celulares (CTR; del inglés, cell-type replacement) es un fenómenogeneral que se da en la glándula digestiva de moluscos en situaciones de estrés. Experimentos previosrealizados en el laboratorio, demostraron que CTR influye sobre las medidas de los procesos debioacumulación y biomarcadores. El objetivo del presente estudio es investigar si CTR es un procesoreversible en limacos crónicamente expuestos a contaminación metálica y determinar su influencia sobrelos parámetros de acumulación y biomarcadores de exposición y efecto. Para ello, se recogieron limacosArion ater en una mina de zinc abandonada (Karrantza) y en un lugar sin contaminar (Bakio) y setransplantaron de un lugar a otro durante 3, 10 y 28 días. La carga de metal se calculó medianteespectrofotometría de absorción atómica (AAS). Como biomarcadores de exposición y efecto seestudiaron los cambios estructurales de lisosomas, la densidad volumétrica de tipos celulares, losdepósitos autometalográficos y los niveles de metalotioneinas. Los resultados demuestran que los limacoscrónicamente expuestos a metales presentan una glándula digestiva similar a la de los limacos expuestosen el laboratorio durante semanas a metales. Sin embargo, al contrario de lo que ocurre en la glánduladigestiva de limacos expuestos a metales por un corto período de tiempo, cuando termina la exposición laglándula digestiva de limacos crónicamente expuestos a metales no responden o presentan una capacidadde respuesta limitada. Por otro lado, en los limacos tranplantados del lugar sin contaminar a la mina 3 díasson suficientes para observar respuestas similares a las obtenidas tras la exposición a metales en ellaboratorio, indicando que el procedimiento analítico y los biomarcadores empleados en el presente trabajoson válidos para detectar exposiciones puntuales a metales en suelos.
Palabras clave: transplante, sustitución de tipos celulares, metales, contaminación cronica, glánduladigestiva, limacos.
Emaitzak eta eztabaida
188

Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
189
SARRERA
Lehorreko gastropodoak metalen metatzaile
eraginkorrak direnez eta kutsaduraren aurrean
modu sentikor eta neurgarrian erantzuten dutenez,
lurzoruen kutsaduraren begirale moduan erabiliak
izan dira oso (Coughtrey & Martin, 1977; Popham
& D'Auria, 1980; Marigómez et al., 1998a; Pihan &
de Vaufleury, 2000; Viard et al., 2004; Snyman et
al., 2005). Liseri-guruina, metaleen metatze-toki
garrantzitsuena da bare eta barraskiloetan (Ireland,
1979; 1984a; 1994; Beeby & Richmond, 1987;
Berger & Dallinger, 1993; Marigómez et al., 1998a;
2002), metalen prozesatze eta detoxifikazioan
gako-funtzioak betetzen dituelarik (Kammenga et
al., 2000). Metalak, iraizte-zelulen lipofuszina
pikorretan (Marigómez et al., 1986a; 1996; 1998a;
Recio et al., 1988a) eta maiztasun gutxiagoz kaltzio
zeluletan (Schoettli & Seiler, 1977; Marigómez et
al., 1986a) lokaliza daitezke, baina metatze-
konpartimentu nagusia liseri-zelulen sistema endo-
lisosomikoa dugu, non metalak metalotioneinei
lotuta bahitzen baitira (Schoettli & Seiler, 1977;
Ireland, 1981; 1982; 1984a; Dallinger & Wieser,
1984; Marigómez et al., 1986a; 2002; Recio et al.,
1988a; Janssen & Dallinger, 1991;1995).
Metal pean egoteak, iraizte-jardueraren
emendioa sortarazi dezake liseri-zeluletan
(Marigómez et al., 1986a; 1986b; 1996; Recio et
al., 1988a; 1988b), zeintzuetatik lisosoma eta
hondakin-gorputzen barruan dauden proteinei
lotutako metalak jariapen apokrino edo holokrino
prozesuen bidez gorotzetara aska baitaitezke
(Marigómez et al., 1986a; 1990a; Recio et al.,
1988a; Ireland & Marigómez, 1992). Iraizte-
jarduera emendatu horren ondorioz, liseri-
guruinaren epitelioaren zelula-osaketan aldaketa
latzak sor daitezke kutsatzailepeko esposizio
subletala pairatu ondoren (Rasmussen et al., 1983;
Widdows et al., 1984; Lowe & Clarke, 1989;
Cajaraville et al., 1990a; 1990b; Marigómez et al.,
1990b; 1996; 1998a; 1998b; Soto et al., 2002).
Efektu horri zelula-moten ordezkapena (ingeleraz,
Cell Type Replacement; CTR) deritzo, eta toxiko
zein bestelako estres-iturri pean mantendutako
molusku itsastarretan egindako lan aurrendarietan
zelula basofilikoen dentsitate bolumetriko edo
kopuru erlatiboaren igoera gisa neurtu da
(Rasmussen et al., 1983; Widdows et al., 1984;
Lowe & Clarke, 1989; Cajaraville et al., 1990a;
1990b; Marigómez et al., 1990b; 1996; 1998a;
1998b).
Liseri-guruineko epitelioaren zelula-moten
ordezkapena, habitat eta kokapen taxonomiko
desberdinetako molusku estresatuetan gertatzen
den fenomeno orokorra dela dirudi. (Marigómez et
al., 1998a). Kutsatzaile organokimiko edo
metalikoen kontzentrazio subletal pean
mantendutako muskuilu itsastarrek, batez ere
zelula basofilikoz osatutako liseri-tepitelioa agertu
dute (Rasmussen et al., 1983; Lowe & Clarke,
1989; Cajaraville et al., 1990a; Soto & Marigómez,
1997a; 1997b; Soto et al., 2002; 1. Kapitulua).
Modu berean, gastropodo itsastarrek zelula
basofiliko hipertrofiatuz osatutako liseri-tubuluen
epitelioa erakutsi dute, laborategian Cd zein
hidrokarburo aromatikoen pean mantenduz gero
(Cajaraville et al., 1990a; Marigómez et al., 1990b).

Emaitzak eta eztabaida
190
Lehorreko bareetan, liseri-guruinaren epitelioa
desberdintzatutako hiru zelula-motez osaturik dago,
esaterako: liseri-zelula (ugariena), iraizte-zelula eta
kaltzio-zelula (zelula basofilikoa; Sumner, 1965).
Gainerako molusku taldeetan bezala, metalez
kutsatutako eskualdeetan bizi diren bare en liseri-
guruinean ere, zelula basofilikoen (kaltzio-zelulen)
kopuru erlatibo altuak eta hipertrofia aurkitu dira
(Marigómez et al., 1998a), eta baita laborategian
metalen pean mantendutakoetan ere (Marigómez
et al., 1996; 4. Kapitulua). Halaber, metalezko
kutsadura kronikoaren baldintzetan, iraizte-zelulen
kopuru erlatiboa ere emenda daiteke (Marigómez et
al., 1998a).
CTR, kutsatzaile peko esposizioari erantzunez
zelula basofilikoen (kaltzio-zelulen) proliferazioaren
ondorioa dela proposatu da (Thompson et al., 1974;
Rasmussen et al., 1983; Widdows et al., 1984;
Lowe & Clarke, 1989; Cajaraville et al., 1990a;
1990b; Marigómez et al., 1990b; 1996). Widdows et
al.-ek (1984), aldaketa horrek liseriketa-prozesuan
zein bioenergetikan berebiziko eragina izan
dezakeen moldarazpen-erantzuna dela
kontsideratu zuten. Izan ere, liseri-zelulen
berriztapena, zelula ama moduan jokatuko luketen
zelula basofilikoen zatiketaren bidez suertatuko
litzatekeela orokorki onartuta zegoen (Yonge, 1926;
Mix & Sparks, 1971; Thompson et al., 1974) eta,
zentzu horretan, zelula basofilikoen proliferazioak,
liseri-zelulen galerak ekarritako zelulen berriztapen
areagotua bermatuko luke (Cajaraville et al.,
1990a). Haatik, oraintsuko ikerketek, muskuiluen
liseri-guruinean liseri-zelulek zein zelula
basofilikoek, ziklo zelularra norberak bere aldetik
jarraitzen duela frogatu dute (Marigómez et al.
1999; Zaldibar et al., 2004), eta beraz, liseri-zelulak
zatitzeko gai direnez, beraiek ordezkatzeko zelula
basofilikoek proliferatzea gertagaitza litzateke. Are
gehiago, metal eta konposatu organikoen
kutsadura peko baldintzetan epitelioaren masa-
murrizpen netoa gertatzen da, liseri-zelulen
galeraren ondorioz seguru asko, murrizpen horrek
zelula basofilikoen kopuru erlatiboaren itxurazko
gorakada ekarriz (Vega et al., 1989; Marigómez et
al., 1990b; 1995; 1996; 1998b; Zaldibar et al.,
2002). Aitzitik, kutsadura peko baldintzetan zein
ingurumen-kaltea eteterakoan liseri-zelulek
proliferatzen dute, BrdU immunohistokimika bidez
magurio eta bareen kasuan laborategiko
esperimentuetan frogatu den moduan (Zaldibar et
al., 2002, 4. eta 5. kapituluak). Beraz, zelula
basofilikoen kopuru erlatiboaren emendioa, batez
ere liseri-zelulen galeraren ondorioa dela onar
genezake. Modu berean, iraizte- eta kaltzio-zelulen
kopuru erlatibo altuak, abandonatutako Cu
meatzean hainbat belaunaldiz (kutsadura kronikoa)
bizi izan diren bareetan aurkitu ziren, liseri-zelulak
pikor zitoplasmikoen iraizpenean oso aktiboak eta
oso bakuolizatuak agertu zirelarik (Marigómez et
al., 1998a). Egoera horrek liseri-zelulen etengabeko
askatzeak azalduko luke. Ildo beretik, iraizte-
zelulak, bizi-zikloaren azken urratsetan dauden
liseri-zelulak izango liratekeela proposatu da, eta ez
beste zelula-mota ezberdina (Porcel et al., 1996;
Dimitriadis & Konstantinidou, 2002). Horrek,
baraualdian eta kutsadurapean liseri- eta iraizte-
zelulen proportzio erlatiboek jarraitzen dituzten
jokabideak kontrajarriak izatea azalduko luke

Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
191
(Porcel et al., 1996; Marigómez et al., 1998a;
Dimitriadis & Konstantinidou, 2002; 5. Kapitulua).
Are gehiago, zelula basofilikoen presentzia
erlatiboaren gorakada, euren hiperplasia eta ez
euren kopuruaren emendioaren ondorioa izango
litzateke batik bat (Marigómez et al., 1990b; 1996;
Chabicovsky et al., 2004; 4. Kapitulua).
Liseri-zelula, ingurumen-kutsaduraren ebalua-
ketarako itu-konpartimentua nagusietarikoa denez,
kutsatzaileen pean egoteak bultzatutako liseri-
zelulen galerak, biometaketa-parametro eta
biomarkatzaileak neurtu direneko zelai- eta epe
luzeko ikerketen emaitza polemikoak azalduko
lituzke. Epe luzeko esposizio (kroniko) ostean,
metalak ez dira liseri-epitelioan metatzen, epitelio
horrek metatze-toki nagusia diren lisosomak
dituzten liseri-zelula urriak baititu (Marigómez et al.,
1996; 1998a; Soto et al., 1998; 2002; Soto &
Marigómez, 1997b). Aldez aurretik burututako
ikerlanek, laborategian metal pean mantendutako
muskuilu eta magurioetan neurtutako
biomarkatzaileen gainean CTRk zer nolako eragina
duen finkatu dute (Soto & Marigómez, 1997b; Soto
et al., 2002; 1. eta 4. Kapituluak), bareen inguruan
dagoen informazioa, berriz, bakana da (Marigómez
et al., 1996; 1998a; 5. Kapitulua).
Metalen pean kronikoki egondako bareen liseri-
guruinean CTR prozesu itzulgarria den finkatzea,
alde batetik, eta CTRk metaleen biometaketa eta
efektu- zein esposizio-biomarkatzaileen gainean
nolako eragina duen ezagutzea, bestetik, ikerlan
honen helburuak dira. Asmo horrekin, Karrantzako
Zn-meatze abandonatu baten inguruko bareak
(Arion ater) hartu eta Bakioko toki kutsatu gabe
batera lekuzaldatu ziren 3, 10 eta 28 egunez, eta
vice versa, Bakiotik Karrantzara. Gorputzeko
metal-zamak absortzio atomikozko
espektrofotometria (AAS) bitartez neurtu ziren.
Bareen liseri-guruinaren epitelioa osatzen duten 3
zelula-moten dentsitate bolumetrikoa estereologia
erabiliz kalkulatu zen (VvDIG, VvEXC, VvCAL)
hematoxilina-eosinaz tindatutako parafinazko
ebakien gainean. Autometalografiaz (AMG)
errebelatutako zilarrezko hauspeakin beltzen
dentsitate bolumetrikoa (VvBSD) eta
espektrofotometria bitartez neurtutako
metalotioneinen (MT) maila esposizio-
biomarkatzaile gisa erabili ziren. Efektu-
biomarkatzaile gisa lisosomen handiagotzea
aukeratu zen, kriostato-ebakien gainean, liseri-
lisosomen bolumenezko, azalerazko eta
zenbakizko dentsitateak (VvLYS, SvLYS eta
NvLYS) eta azalera/bolumena ratioa (S/VLYS)
irudi-analisi bidez neurtuz gero estimatu zelarik.
MATERIAL ETA METODOAK
Erreaktibo kimikoak
Erabilitako erreaktibo kimiko guztiak maila
analitikokoak eta Sigma-Aldrich (St. Louis,
Missouri, Amerikako Estatu Batuak), merkatal-
etxekoak dira besterik agertu ezean.
Diseinu esperimentala eta laginen prozesatzea
50 Arion ater bare ar, Euskal Herriko 2 herri
desberdinetan jaso ziren 2002.ko ekainaren 1ean
(1. Ird.): (a) Karrantzako bailaran abandonatutako
Zn-meatze zahar baten inguruan; eta (b)

kutsatugabeko toki batean Bakion, meatzetik aski
urruti dagoen landa-herria dena. Bareak toki batetik
bestera 3, 10 eta 28 egunez lekuzaldatu ziren, eta
baldintza naturaletan mantendu ziren lurzoruari
atxikitako saredun kutxen bitartez. Horretaz gain,
bai Bakion zein Karrantzan lurzorua eta bareak
lagindu ziren, transplantea egin zeneko egun
berean (0. eguna). Bareak (10 talde
esperimentaleko), laginketa-egunetan sakrifikatu
ziren, eta segidan azalduko den moduan prozesatu
ziren. Liseri-guruinaren zati bat, Carnoy likidoan
fixatu, parafinan inkluditu eta 7 µm-ko lodierako
ebakietan moztu zen, azterketa histologikoak eta
analisi histokimikoak egiteko. Bigarren zati bat,
glutaraldehidoan fixatu eta EPON erretxinan
inkluditu zen, ebaki semifinen behaketa egiteko.
Hirugarren zati bat, nitrogeno likidotan izoztu zen. 8
µm-ko lodierako ebakiak, kriostatoan moztu, eta
ondoren ß-glukuronidasa entzimaren jardueraren
demostrazio histokimikoa egiteko prozesatu ziren.
Liseri-guruinaren beste zati bat nitrogeno likidotan
izoztu zen, metalotioneinen neurketa
espektrofotometrikoa burutzeko. Azkenik, geratzen
zen liseri-guruinaren zatia, metalen analisi kimikoak
egiteko liseriketa azidoaren bitartez prozesatu zen.
Prozedura bakoitzaren nondik norakoak,
zehaztasun gehiagoz jarraian azalduko dira.
Analisi kimikoak
Liseri-guruinak ur distilatuan garbitu, eta labean
120ºC-tara lehortu ziren 48 orduz, pisua egonkortu
zen arte (Ireland, 1979). Lehortutako ehunak azido
nitriko kontzentratuan liseritu ziren. Gerora, ur
distilatuan disolbatutako azido nitrikotan (0.1 M)
diluitu ondoren, laginak absortzio atomikozko
espektrofotometroan (PerkinElmer 2280, Wellesley,
Massachusetts, Amerikako Estatu Batuak) neurtu
ziren, aldi bereko atzealdeko zuzenketaz eta 0.3
mg/l-ko sentikortasunaz. Merck merkatal-etxeko
(Merck & Co., Inc, Whitehouse Station, New Jersey,
Amerikako Estatu Batuak) disoluzio estandarrak
(zertifikatutakoak) 0.1 M azido nitrikotan diluitu ziren
kalibrazioa burutzeko. 7 metal analizatu ziren:
kadmioa (Cd), kromoa (Cr), kuprea (Cu), burdina
Emaitzak eta eztabaida
192
1. Irudia: Erreferentzia tokia(Bakio) eta antzinako meatze-tokiaren (Karrantza)kokapenaren irudi eskematikoa.Geziek bareenlekuzaldatzearen nondiknorakoak islatzen dituztekutxen barruan agertutakoegun kopuruen zehar

(Fe), beruna (Pb), nikela (Ni) eta zinka (Zn).
Lurzoruaren laginak modu berean prozesatu ziren.
Metalotioneina-mailen determinazio
espektrofotometrikoa
MT-mailak sulfidrilo taldeen (-SH) determinazio
espektrofotometrikoaren bitartez estimatu ziren
(UNEP/RAMOGE, 1999). MTen kontzentrazioa,
laginaren -SH edukina espertrofotometro bitartez
kuantifikatu da, Ellman erreaktiboa (5,5 ditiobis 2
azido nitrobenzoikoa; DTNB) erabiliz. Laburki,
liseri-guruinaren zatiak 1 g ehun emateko taldekatu
ziren, eta motoreak gidatutako teflonezko
homogeneizagailuan ß-merkaptoetanola,
fenilmetanosulfonilfluorido (PMSF) eta
leupeptinadun indargetzaile homogeneizatzai-
learen 3 bolumenetan homogeneizatu ziren.
Homogeneizatuak 30000x g-tan 20 min 0-4ºC-tan
zentrifugatu ziren. Gainjalkinean zeuden pisu
molekular baxuko proteinak etanol absolutu
hotzean (-20ºC), eta ondoz ondoko 6000x g-ko
zentrifugazioaren bitartez prezipitatu ziren. MTtan
joria zen pisu molekular baxuko frakzioa 0.25 M
NaCl, 1 N HCl eta 4 nM EDTA zuen indargetzailean
berresekitu zen. Erreferentziazko kurba
estandarrak, 1 mg glutation/ml 0.25 M NaCl stock
disoluzioa erabiliz sortu ziren. MTen ebaluazio
espektrofotometrikoa 0.43 mM DTNB 0.2 M fosfato
indargetzailean erreaktibo gisa erabiliz egin zen,
absorbantzia 412 nm-tan neurtuz.
Autometalografia
Parafinazko ebakien talde bat
autometalografiaz tindatu zen. Autometalografia
metalak zilarrezko hauspeakin beltz gisa
ikustarazteko argazkilaritzan oinarritutako teknika
da (Danscher, 1984). Ebakiak (7 µm), xilenoz
desparafinatu ziren, beheranzko graduazioko
etanolezko bainu-serie batean hidratatu eta labean
(40ºC) utzi ziren guztiz lehortu arte. Laginak
ondoren, eta gela ilunean, argazki-emultsio geruza
fin eta uniforme batez estali ziren (Ilford Nuclear
Emulsion L4, Ilford, Mobberley, Ingalaterra).
Laginak ilunpean mantendu ziren lehortu bitartean
(30 minutu) eta Ultrafin Tetenal SF (1:5 diluzioa ur
distilatuan) (Tetenal AG & Co, Norderstedt,
Alemania) errebelatzaile komertziala erabiliz
errebelatu ziren 15 minutuz. Ondoren,
errebelatzailearen erreakzioa %1 azido azetikoa
zuen diluzio batean geldiarazi zen minutu batez eta,
azkenik, erreakzioa fixatzailean (Agfa Agefix, Agfa-
Gevaert N.V., Mortsel, Belgika, 1:9 uretan) fixatu
zen 10 minutuz. Laginak, Kaiser gelatina
glizerinatuan muntatu ziren.
BSDen kuantifikazioa argi-mikroskopioan
irudi-analisi bitartez
Liseri-zelulen lisosometan zeuden BSDen
azalera irudi-analisia erabiliz kuantifikatu zen (Soto
& Marigómez, 1997a; 1997b), emaitzak BSDen
dentsitate bolumetriko gisa adieraziz (VvBSD). Argi-
mikroskopiazko irudiak (x100 handipenean) B&W-
CCD TV kamera, HR-TV kolorezko pantaila, Leitz
mikroskopioa, PC ordenagailua, PIP 1024 (Matrox
Electronic Systems Ltd., Dorval, Kanada) sistema
eta Sevisan (Bilbo, Espainia) entrepresak
garatutako softwarea erabiliz eskuratu ziren.
Ondoren, eskuz eta softwarearen laguntzarekin
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
193

liseri-zelulen lisosometako BSD eta zitoplasma
bereizteko segmentazio-prozedura aplikatu zen.
Neurketak bare bakoitzean 5 eremutan burutu
ziren, eta talde esperimental bakoitzean 10 bare
erabili ziren; tratamendu bakoitzeko guztira 50
neurketa eginez. Kalkuluak, Lowe et al.-ek (1981)
proposatutako formulak erabiliz egin ziren. Hau da,
BSD partikulen tamaina ebakiaren lodiera baino
txikiagoa zela kontutan hartuz, Holmes efektua
zuzentzeko faktorea kontuan hartu zen.
Azterketa histologikoa eta analisi histologiko
kuantitatiboak
Parafinan inkluditutako ebakiak (7 µm)
Hematoxilina-eosinaz tindatu ziren eta EPON
erretxinan inkluditutako ebaki semifinak (3-4 µm)
toluidina urdinez tindatu ziren, talde desberdinetako
bareen liseri-guruineko epitelioaren eta bertoko
zelula-moten deskribapen morfologikoa argi-
mikroskopioaren bitartez egiteko.
Liseri-zelulen, iraizte-zelulen eta kaltzio-zelulen
dentsitate bolumetrikoa kalkulatzeko (VvDIG,
VvEXC, VvCAL) prozedura estereologikoa aplikatu
zen. Horretarako, lagin bakoitzean azarez
aukeratutako zelai batetan egin ziren kontaketak
guztira talde esperimental bakoitzeko 10 kontaketa
burutuz. Analisi estereologikoak burutzeko, Nikon
Optiphot mikroskopioari atxikitako irudi-hodia
erabili zen (amaierako handipena 670x). Zelulen
VvDIG, VvEXC eta VvCAL Delesse printzipioan
oinarrituz (Weibel, 1979) kalkulatu zen. Lortutako
datu horietan oinarrituta, zelula-mota bakoitzaren
proportzio erlatiboa portzentaietan kalkulatu zen.
Lisosomen egitura-aldaketak
Liseri-zelulen lisosomen ikustaraztea ß-
glukuronidasa (ß-GUS) entzimaren jardueraren
agerketa histokimikoa burutuz egin zen, Cajaraville
et al.-en (1991) prozedura jarraituz. Ebakiak, egin
berria zen ß-GUS inkubazio-medioan (28 mg
naphtol AS-BI-ß-D-glucuronide 1.2 ml 50 mM sodio
bikarbonatoan disolbatuta eta 100 ml-arte %2.5
NaCl eta %15 polibinilo alkohola zuen azetato
indargetzailean, pH 4.5) murgildu ziren 10 minutuz
37ºC-tan, etengabe irabiatuz. Inkubazioaren
ondoren, laginak %2.5 NaCl gatz-disoluzioan
garbitu ziren 2 minutuz 37ºC-tan, etengabe
irabiatuz. Laburki, ebakiak disoluzio ikustarazlean
sartu ziren (0.1 g Fast Garnet GBC % 2.5 NaCl
zuen 0.1 M fosfato indargetzaileko (pH 7.4) 100 ml-
tan disolbatuak) 10 minutuz eta ilunpean. Ondoren,
laginak %2.5 NaCl zuen Baker formol kaltzikoan
fixatu ziren 10 minutuz 4ºC-tan, eta uretan labur
garbitu ziren. Azkenik, laginak % 0.1 Fast Green
disoluzio batean kontrastatu ziren bi minutuz, ur
distilatuan hainbat aldiz garbitu, Kaiser gelatina
glizerinatuan montatu eta azazkalen esmalteaz
zigilatu ziren.
Lisosomen egitura, automatizatutako irudi-
analisi (Biological Measurement System software-
BMS- Sevisan; Cajaraville et al., 1991) bitartez
neurtu zen. Ebakiak, x100 handipeneko objektiboa
erabiliz aztertu ziren Leitz argi-mikroskopioan. 5
neurketa burutu ziren ebaki bakoitzean, hurrengo
parametro estereologikoak kalkulatuz: lisosomen
dentsitate bolumetrikoa (VvLYS = V(L)/V(C)),
dentsitate superfiziala (SvLYS = S(L)/V(C)),
azalera/bolumena ratioa (S/VLYS = S(L)/V(L)) eta
Emaitzak eta eztabaida
194
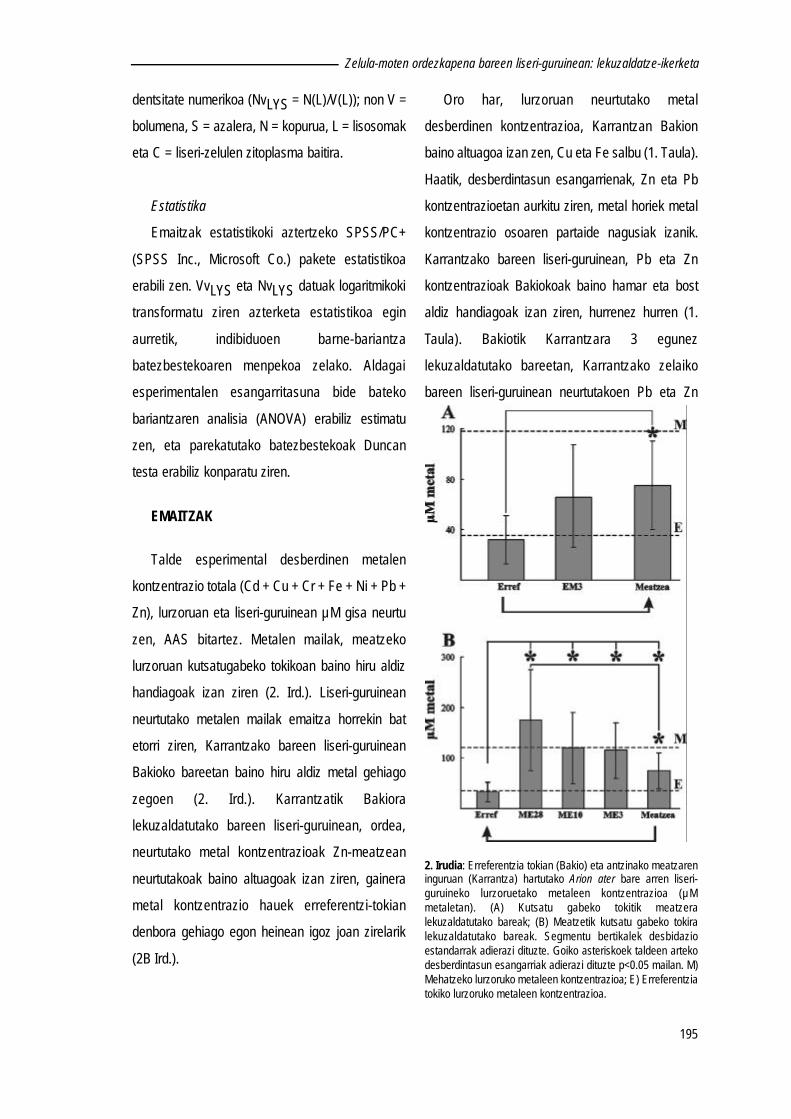
dentsitate numerikoa (NvLYS = N(L)/V(L)); non V =
bolumena, S = azalera, N = kopurua, L = lisosomak
eta C = liseri-zelulen zitoplasma baitira.
Estatistika
Emaitzak estatistikoki aztertzeko SPSS/PC+
(SPSS Inc., Microsoft Co.) pakete estatistikoa
erabili zen. VvLYS eta NvLYS datuak logaritmikoki
transformatu ziren azterketa estatistikoa egin
aurretik, indibiduoen barne-bariantza
batezbestekoaren menpekoa zelako. Aldagai
esperimentalen esangarritasuna bide bateko
bariantzaren analisia (ANOVA) erabiliz estimatu
zen, eta parekatutako batezbestekoak Duncan
testa erabiliz konparatu ziren.
EMAITZAK
Talde esperimental desberdinen metalen
kontzentrazio totala (Cd + Cu + Cr + Fe + Ni + Pb +
Zn), lurzoruan eta liseri-guruinean µM gisa neurtu
zen, AAS bitartez. Metalen mailak, meatzeko
lurzoruan kutsatugabeko tokikoan baino hiru aldiz
handiagoak izan ziren (2. Ird.). Liseri-guruinean
neurtutako metalen mailak emaitza horrekin bat
etorri ziren, Karrantzako bareen liseri-guruinean
Bakioko bareetan baino hiru aldiz metal gehiago
zegoen (2. Ird.). Karrantzatik Bakiora
lekuzaldatutako bareen liseri-guruinean, ordea,
neurtutako metal kontzentrazioak Zn-meatzean
neurtutakoak baino altuagoak izan ziren, gainera
metal kontzentrazio hauek erreferentzi-tokian
denbora gehiago egon heinean igoz joan zirelarik
(2B Ird.).
Oro har, lurzoruan neurtutako metal
desberdinen kontzentrazioa, Karrantzan Bakion
baino altuagoa izan zen, Cu eta Fe salbu (1. Taula).
Haatik, desberdintasun esangarrienak, Zn eta Pb
kontzentrazioetan aurkitu ziren, metal horiek metal
kontzentrazio osoaren partaide nagusiak izanik.
Karrantzako bareen liseri-guruinean, Pb eta Zn
kontzentrazioak Bakiokoak baino hamar eta bost
aldiz handiagoak izan ziren, hurrenez hurren (1.
Taula). Bakiotik Karrantzara 3 egunez
lekuzaldatutako bareetan, Karrantzako zelaiko
bareen liseri-guruinean neurtutakoen Pb eta Zn
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
195
2. Irudia: Erreferentzia tokian (Bakio) eta antzinako meatzareninguruan (Karrantza) hartutako Arion ater bare arren liseri-guruineko lurzoruetako metaleen kontzentrazioa (µMmetaletan). (A) Kutsatu gabeko tokitik meatzeralekuzaldatutako bareak; (B) Meatzetik kutsatu gabeko tokiralekuzaldatutako bareak. Segmentu bertikalek desbidazioestandarrak adierazi dituzte. Goiko asteriskoek taldeen artekodesberdintasun esangarriak adierazi dituzte p<0.05 mailan. M)Mehatzeko lurzoruko metaleen kontzentrazioa; E) Erreferentziatokiko lurzoruko metaleen kontzentrazioa.
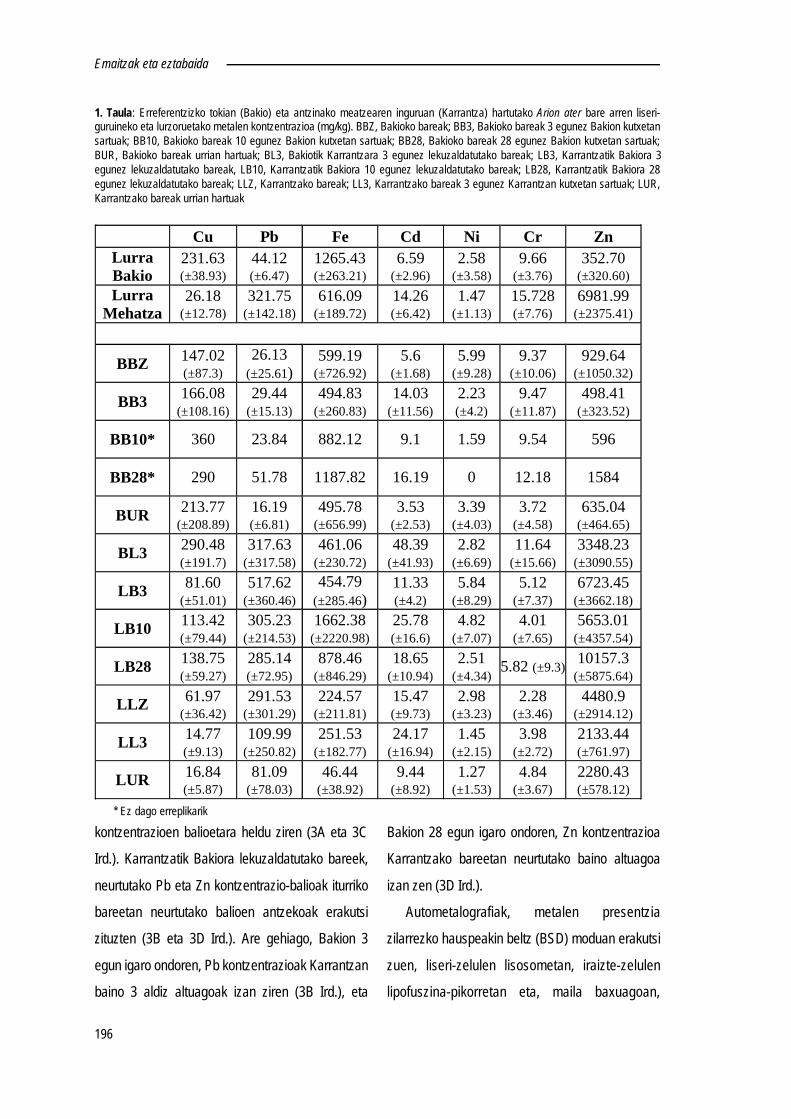
kontzentrazioen balioetara heldu ziren (3A eta 3C
Ird.). Karrantzatik Bakiora lekuzaldatutako bareek,
neurtutako Pb eta Zn kontzentrazio-balioak iturriko
bareetan neurtutako balioen antzekoak erakutsi
zituzten (3B eta 3D Ird.). Are gehiago, Bakion 3
egun igaro ondoren, Pb kontzentrazioak Karrantzan
baino 3 aldiz altuagoak izan ziren (3B Ird.), eta
Bakion 28 egun igaro ondoren, Zn kontzentrazioa
Karrantzako bareetan neurtutako baino altuagoa
izan zen (3D Ird.).
Autometalografiak, metalen presentzia
zilarrezko hauspeakin beltz (BSD) moduan erakutsi
zuen, liseri-zelulen lisosometan, iraizte-zelulen
lipofuszina-pikorretan eta, maila baxuagoan,
Emaitzak eta eztabaida
196
1. Taula: Erreferentzizko tokian (Bakio) eta antzinako meatzearen inguruan (Karrantza) hartutako Arion ater bare arren liseri-guruineko eta lurzoruetako metalen kontzentrazioa (mg/kg). BBZ, Bakioko bareak; BB3, Bakioko bareak 3 egunez Bakion kutxetansartuak; BB10, Bakioko bareak 10 egunez Bakion kutxetan sartuak; BB28, Bakioko bareak 28 egunez Bakion kutxetan sartuak;BUR, Bakioko bareak urrian hartuak; BL3, Bakiotik Karrantzara 3 egunez lekuzaldatutako bareak; LB3, Karrantzatik Bakiora 3egunez lekuzaldatutako bareak, LB10, Karrantzatik Bakiora 10 egunez lekuzaldatutako bareak; LB28, Karrantzatik Bakiora 28egunez lekuzaldatutako bareak; LLZ, Karrantzako bareak; LL3, Karrantzako bareak 3 egunez Karrantzan kutxetan sartuak; LUR,Karrantzako bareak urrian hartuak
Cu Pb Fe Cd Ni Cr Zn Lurra Bakio
231.63 (±38.93)
44.12 (±6.47)
1265.43 (±263.21)
6.59 (±2.96)
2.58 (±3.58)
9.66 (±3.76)
352.70 (±320.60)
Lurra Mehatza
26.18 (±12.78)
321.75 (±142.18)
616.09 (±189.72)
14.26 (±6.42)
1.47 (±1.13)
15.728 (±7.76)
6981.99 (±2375.41)
BBZ 147.02 (±87.3)
26.13 (±25.61)
599.19 (±726.92)
5.6 (±1.68)
5.99 (±9.28)
9.37 (±10.06)
929.64 (±1050.32)
BB3 166.08 (±108.16)
29.44 (±15.13)
494.83 (±260.83)
14.03 (±11.56)
2.23 (±4.2)
9.47 (±11.87)
498.41 (±323.52)
BB10* 360 23.84 882.12 9.1 1.59 9.54 596
BB28* 290 51.78 1187.82 16.19 0 12.18 1584
BUR 213.77 (±208.89)
16.19 (±6.81)
495.78 (±656.99)
3.53 (±2.53)
3.39 (±4.03)
3.72 (±4.58)
635.04 (±464.65)
BL3 290.48 (±191.7)
317.63 (±317.58)
461.06 (±230.72)
48.39 (±41.93)
2.82 (±6.69)
11.64 (±15.66)
3348.23 (±3090.55)
LB3 81.60 (±51.01)
517.62 (±360.46)
454.79 (±285.46)
11.33 (±4.2)
5.84 (±8.29)
5.12 (±7.37)
6723.45 (±3662.18)
LB10 113.42 (±79.44)
305.23 (±214.53)
1662.38 (±2220.98)
25.78 (±16.6)
4.82 (±7.07)
4.01 (±7.65)
5653.01 (±4357.54)
LB28 138.75 (±59.27)
285.14 (±72.95)
878.46 (±846.29)
18.65 (±10.94)
2.51 (±4.34) 5.82 (±9.3)
10157.3 (±5875.64)
LLZ 61.97 (±36.42)
291.53 (±301.29)
224.57 (±211.81)
15.47 (±9.73)
2.98 (±3.23)
2.28 (±3.46)
4480.9 (±2914.12)
LL3 14.77 (±9.13)
109.99 (±250.82)
251.53 (±182.77)
24.17 (±16.94)
1.45 (±2.15)
3.98 (±2.72)
2133.44 (±761.97)
LUR 16.84 (±5.87)
81.09 (±78.03)
46.44 (±38.92)
9.44 (±8.92)
1.27 (±1.53)
4.84 (±3.67)
2280.43 (±578.12)
* Ez dago erreplikarik

kaltzio-zelulen kaltzio-esferuletan (4. Ird.).
Behaketa mikroskopikoa burutu ondoren, BSD,
Karrantzako bareetan kutsatu gabeko tokian baino
ugariagoak zirela argi agertu zen (4A eta 4B Ird.).
Bakioko bareetan, BSD, liseri-epitelioko xafla
basalean (zentzu histologikoan) eta liseri-zelulen
erpinaldeko lisosoma (pikor berdeak) gutxi batzutan
behatu ziren (4A Ird.). Kutsatutako tokiko bareetan,
eta kutsatu gabeko tokira 3 egunez
lekuzaldatutakoetan BSD xafla basalean urriagoak
izan ziren arren, sistema endo-lisosomikoaren
bolumen gehiena betetzen agertu ziren (pikor berde
eta horiak), eta iraizte-zelulen lipofuszinetan ere
konspikuoak gertatu ziren (4B eta 4D Ird.).
BSDen hedapen-balioak, irudi-analisi bitartez
VvBSD gisa kuantifikatuak, Karrantzako bareetan
Bakioko bareetan baino 10 aldiz handiagoak izan
ziren. Bakiotik Karrantzara 3 egunez
lekuzaldatutako bareetan tarteko balioak
kuantifikatu ziren (5A Ird.). Kutsatutako tokitik
kutsatugabekora lekuzaldatutako bareen liseri-
guruinean, VvBSD balioa ez zen jaitsi. Aitzitik, 3 eta
10 egunez lekuzaldatu ondoren balio handiagoak
suertatu ziren (5B Ird.).
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
197
3. Irudia: Erreferentzia tokian (Bakio) eta antzinako meatzaren inguruan (Karrantza) hartutako Arion ater bare arren liseri-guruinekoehunetako eta lurzoruetako Pb (A eta B) eta Zn (C eta D) kontzentrazioak (µg/g ehun-pisu lehor).( A) eta (C) Kutsatu gabeko tokitikmeatzera lekuzaldatutako bareak; (B) eta (D) Meatzetik kutsatu gabeko tokira lekuzaldatutako bareak. Segmentu bertikalekdesbidazio estandarrak adierazi dituzte. Goiko asteriskoek taldeen arteko desberdintasun esangarriak adierazi dituzte p<0.05mailan. M) Mehatzeko lurzoruko Pb eta Zn kontzentrazioa; E) Erreferentzia tokiko lurzoruko Pb eta Zn kontzentrazioa.

Bi tokietan determinazio espektrofotometrikoz
kuantifikatutako metalotioneinen (MT)
kontzentrazioa ez zen esangarriki desberdina izan
(6A Ird.). Era berean, Bakiotik Karrantzara
lekuzaldatutako bareetan MT kontzentrazioa ez zen
esangarriki aldatu. Bestalde, kutsatu gabeko tokian
3 egunez mantendutako bareetan MT
kontzentrazioa esangarriki jaitsi zen (6B Ird.).
ß-glukuronidasaren histokimika burutu ondoren,
liseri-zelulen lisosomak arre-purpura koloreko
hauspeakin modura behatu ziren (4E eta F Ird.).
Karrantzako bareetan (4F Ird.), lisosomak
Emaitzak eta eztabaida
198
4. Irudia: Arion ater barearen liseri-epitelioan metalen lokalizazio autometalografikoa (A-D) eta ß-glukuronidasa entzima-jardueraren detekzio histokimikoa (E-F). (A) Kutsatu gabeko tokian hartutako barea; (B) Zn-meatzetik hartutako barea; (C) Kutsatugabeko tokitik meatzera 10 egunez lekuzaldatutako barea; (D) Zn-meatzetik kutsatu gabeko tokira 3 egunez lekuzaldatutako barea.Geziek liseri-zelula, iraizte-zelula eta kaltzio-zelulen barruan dauden metalak adierazten dituzte. Gezi-buruek liseri-azinoetakoxafla-basalean metatutako metalak adierazten dituzte. Eskala-marra: 50 µm. (E) Kutsatu gabeko bareen liseri-guruineantopatutako ß-glukuronidasa entzimaren jarduera; (F) Zn-meatzeko bareen ß-glukuronidasa entzimaren jarduera. Geziek ß-glukuronidasa jarduera adierazten dute. Eskala-marra: 25 µm.

nabarmenki handiagoak izan ziren Bakiokoetan
baino (4E Ird.). Estereologia bidez egindako analisi
kuantitatiboek emaitza hauek baieztatu zituzten:
VvLYS eta SvLYS altuagoak eta S/VLYSbaxuagoak neurtu ziren Karrantzako laginetan
Bakioko laginetan baino (7. Ird.). Bakiotik
Karrantzara 3 egunez lekuzaldatutako bareetan,
VvLYS eta SvLYS parametroak aldatu ez ziren
arren, lisosomen tamainaren handiagotzea
(S/VLYS baxua) kopuruaren txikipen
esangarriarekin batera (NvLYS baxua) gertatu zen.
Kontrako lekuzaldatzea egiterakoan, lisosomek ez
zuten inolako erantzunik eman 28 egunez (7B Ird.).
Karrantzako liseri-guruineko azinoek,
Bakiokoek baino epitelio meheagoa erakutsi zuten;
gainera, argia handiagoa eta azinoen kalibrea ere
txikiagoa izan ziren (8. Irudia). Are gehiago,
loditutako xafla basala eta odol hodiak eratzen
duten Leydig zelula puztuak aurkitu ziren (8. Ird.).
Liseri-zelulak, kaltzio-zelulak eta iraizte-zelulak
errez identifikatu ziren bi tokietako bareen liseri-
guruineko epitelioan; hala ere, zelula-mota
bakoitzaren maiztasun erlatiboa desberdina izan
zen bi tokietan (8. eta 9. Ird.). Eskuarki, kaltzio-
zelulen proportzio erlatiboa konstante mantendu
zen lagin desberdinetan, liseri-zelula eta iraizte-
zelulena esangarriki aldatu ziren bitartean. Iraizte-
zelulak, toki kutsatuan kutsatu gabeko tokian baino
bost aldiz ugariagoak izan ziren (%60 vs %12), eta
liseri-zelulak, ordea, askoz urriagoak (%30 vs %80;
9. Ird) Gainera, kutsatu gabeko tokitik toki
kutsatura 3 egunez lekuzaldatutako bareetan
zelula-mota desberdinen proportzioek tarteko
balioak erakutsi zituzten; hau da, batez ere iraizte-
zelulen emendioa gertatu zen (9A Ird.). Toki
kutsatutik kutsatugabekora lekuzaldatutako bareen
liseri-guruinean pareko erantzuna behatu zen, non
kaltzio-zelulen proportzioan aldaketarik behatu ez
zen bitartean iraizte-zelulen gutxiagotze graduala
aurkitu baitzen (9B Ird.). Aldaketa horiek parametro
adierazgarrietan islatzeko asmoz, VvDIG eta
VvEXC/VvDIG ratioa hautatu egin dugu (10. Ird.).
Bakion, VvDIG Karrantzan baino bi aldiz altuagoa
hain zuzen, eta 3 egunez Bakiotik Karrantzara
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
199
5. Irudia: Erreferentzia tokian (Bakio) eta antzinako meatzareninguruan (Karrantza) hartutako Arion ater bare arren liseri-guruinean neurtutako zilarrezko hauspeakin beltzen dentsitatebolumetrikoa. (A) Kutsatu gabeko tokitik meatzeralekuzaldatutako bareak; (B) Meatzetik kutsatu gabeko tokiralekuzaldatutako bareak. Segmentu bertikalek desbidazioestandarrak adierazi dituzte. Goiko asteriskoek taldeen artekodesberdintasun esangarriak adierazi dituzte p<0.05 mailan.

Emaitzak eta eztabaida
200
lekuzaldatu ondoren tarteko balioak agertu ziren
(10A Ird.). VvEXC/VvDIG ratioari dagokionez,
Karrantzako balioak Bakiokoak baino 8 aldiz
handiagoak dira (10B Ird.). Halaber, Karrantzatik
Bakiora 28 egunez lekuzaldatutako bareetan
VvDIG Bakioko jatorrizko balioetaraino igo ez
bazen ere, igoera gradualaren ebidentzia argiak
topatu ziren (10C Ird.). VvEXC/VvDIG ratioak ere
joera horren aztarnak islatzen ditu (10D Ird.).
EZTABAIDA
Metal-kutsadura kronikoaren aurreko
moldarazpena
Metal-kutsadura kronikoa eta bere efektua
lehorreko organismoen gainean, utzitako meatzeak
zelai-laborategi merke eta eraginkor gisa erabiliz
iker daiteke (Ireland & Wootton, 1975; Dallinger et
al., 1989; Marigómez et al., 1998a). Ikerlan
honetan, Zn-meatze batean (Karrantzan) hainbat
hamarkadaz bizitako bareen populazioek Pb eta Zn
kontzentrazio esangarriki altuagoak erakutsi
zituzten, erreferentziazko tokikoekin alderatzekotan
(Bakio). Ehunetako metalen kontzentrazioak
lurzoruan neurtutakoekin bat datoz. Behaketa
horiek, lehorreko gastropodoen liseri-guruinean
neurtutako metalen kontzentrazioak ingurumeneko
metalen mailekin erlazionaturik daudela baieztatu
dute (Ireland, 1981; 1982; Marigómez et al., 1998a;
Beeby & Richmond, 2002). Karrantzako bareek, Pb
eta Zn kutsadura kronikoaren pean egonik, hainbat
belaunalditan zehar bizirik irautea lortu dute. Hala
ere, Zn-meatzean bareek bizirik jarraitu arren,
nolabaiteko estres kronikoa pairatu dutela espero
daiteke, biomarkatzaile lisosomikoek agerian utzi
dutenez (ikus aurrerago).
Bizirauteko gaitasun hori ez zaigu horren
deigarria iruditu, euren ehunetan oso metal
kantitate altuak metatu dituzten metal kutsatzailen
pean egondako barraskilo eta bareetan garrantzi
gutxiko hilkortasuna aurkitu baita (Russell et al.,
1981; Marigómez et al., 1986c; 1998a; Greville &
Morgan, 1991; Berger & Dallinger, 1993; Dallinger,
1994). Gainera, metal batetik bestera
6. Irudia: Erreferentzia tokian (Bakio) eta antzinako meatzareninguruan (Karrantza) hartutako Arion ater bare arren liseri-guruinean neurtutako metalotioneinen (MT) kontzentrazioa. (A)Kutsatu gabeko tokitik meatzera lekuzaldatutako bareak; (B)Meatzetik kutsatu gabeko tokira lekuzaldatutako bareak.Segmentu bertikalek desbidazio estandarrak adierazi dituzte.Goiko asteriskoek taldeen arteko desberdintasun esangarriakadierazi dituzte p<0.05 mailan.

sentikortasuna eta tolerantzia alda daitezke
(Laskowski & Hopkin, 1996). Pb/Zn-z kutsatutako
eta kutsatu gabeko tokien arteko lekuzaldatze-
esperimentuetan oinarrituz, bareen metalekiko
tolerantziaren berri aurretikoz argitaratu da.
Tolerantzia hori, metalen metaketaren murrizpenaz
fenotipikoki adierazten da; Cd moduko beste metal
astunekiko tolerantzia, ordea, metatze-gaitasun
areagotuaz karakterizatzen den bitartean (Greville
& Morgan, 1991). Eskuarki, metal-kutsadura
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
201
7. Irudia: Erreferentzia tokian(Bakio) eta antzinako meatzareninguruan (Karrantza) hartutakoArion ater bare arren liseri-guruinean neurtutako ß-glukuronidasa jarduerarenparametro estereologikoak. (A)Kutsatu gabeko tokitik meatzeralekuzaldatutako bareen dentsitatebolumetrikoa; (B) Meatzetikkutsatu gabeko tokiralekuzaldatutako bareen dentsitatebolumetrikoa; (C) Kutsatu gabekotokitik meatzera lekuzaldatutakobareen dentsitate superfiziala; (D)Meatzetik kutsatu gabeko tokiralekuzaldatutako bareendentsitate superfiziala; (E)Meatzetik kutsatu gabeko tokiralekuzaldatutako bareenAzalera/Bolumena ratioa; (F)Meatzetik kutsatu gabeko tokiralekuzaldatutako bareenAzalera/Bolumena ratioa; (G)Kutsatu gabeko tokitik meatzeralekuzaldatutako bareen dentsitatenumerikoa; (H) Meatzetik kutsatugabeko tokira lekuzaldatutakobareen dentsitate numerikoa.Segmentu bertikalek desbidazioestandarra adierazi dituzte. Goikoasteriskoek taldeen artekodesberdintasun esangarriakadierazi dituzte p<0.05 mailan.

zelaian pairatzekotan, hautespen-presioak
animalien gainean eragin esanguratsua izan
dezake; kutsatzaileen harrera murrizteak eta iraizte
edo eta metatze bidezko detoxifikazio-tasak
areagotzeak, hautespenerako abantaila ekar
lezaketelarik (Wang & Rainbow, 2005). Bare eta
barraskiloek toki kutsatuetan bizirauteko duten
ahalmena, metalak detoxifikatu edo gehiegizko
esposizioa ekiditeko duten gaitasunean oinarritzen
dela dirudi. Oro har, metalekiko erresistentzia,
metal-ioiak (Cd, Cu, Zn) estuki bahitzen dituzten
metalotioneinen ekoizpenarekin zein liseri-
guruineko lisosometan eta pikor mineralizatuetan
metal toxikoen bahiketarekin erlazionatuta dago
(Hopkin et al., 1989; Dallinger, 1993; Marigómez et
al., 2002; Dallinger et al., 2004). Hirigunetako
barraskilo eta meatzetako bareen kasuan
proposatu denez, Pb- eta Zn-esposizio kronikoak
hautespen naturalerantz darama, mekanismo
genetiko edo eta anbientalen bitartez (Beeby &
Richmond, 1987; Greville & Morgan, 1991).
Fenotipo tolerante hautatuak, beste
bereizgarrien artean, liseri-guruineko epitelioko
zelula-moten banaketa eta presentzia erlatiboan
islatutako desberdintasunez karakterizatuta daude
(Marigómez et al., 1998a). Lehorreko
gastropodoetan, metalen metaketa eta
detoxifikaziorako toki nagusia liseri-guruina dugu,
eta bere epitelioa 3 zelula-motez osaturik dago:
liseri-zelula, iraizte-zelula eta kaltzio-zelula
(Sumner, 1965). Egoera normaletan, liseri-zelulak
ugarienak dira, baina Cd-z kutsatutako tokietan bizi
Emaitzak eta eztabaida
202
8. Irudia: Kutsatu gabeko tokiko (A eta C irudiak) eta Zn-meatzeko bareen (B eta D irudiak) irudi histologikoak, bai Hematoxilina-eosinaz (A eta B) zein toluidina urdinez (C eta D) tindatutako ebakietan. DC) liseri-zelula, EC) iraizte-zelula, eta CC) kaltzio-zelulaadierazten dute. Eskala-marra: 50 µm (A eta B irudiak); 20 µm (C eta D irudiak).

diren bareetan kaltzio-zelulak eta iraizte-zelulak
zelula-mota nagusiak bilaka daitezke (Marigómez
et al., 1998a). Epe luzeko/kronikoak diren
kutsatzaile-esposizioen ondorioz, zelula-moten
osaketaren halako aldaketak, moluskuen erantzun
orokortzat har daitezkeela dirudi (Rasmussen et al.,
1983; Widdows et al., 1984; Cajaraville et al.,
1990a; 1990b; Marigómez et al., 1990b; 1996;
1998a), azken finean kutsatzaileen aurreko
erantzuna zein erantzun horren zenbait parametro
adierazle ere eragin ditzaketelarik (Marigómez et
al., 1998b).
Laborategiko baldintzetan metal-kutsadura
pean jarritako lehorreko gastropodoen liseri-
guruinean, xafla basalak loditu, liseri-zelulak
lehendabizi bakuolizatu eta ondoren gutxiagotu,
kaltzio-zelulak hipertrofiatu, kaltzio- eta iraizte-
zelulak erlatiboki ugaritu, epitelioa mehetu, eta
odol-zelulak zein ehun konektiboko zelula
migratzaileak liseri-azinoen inguruan metatu egiten
dira (Marigómez et al., 1986a; 1986b; 1996; 1998a;
Recio et al., 1988a; 1988b; Ireland & Marigómez,
1992). Metal-kutsadurarekiko moldarazpenaren
“steady state” hori, metal toxikoen maila subletalen
pean kronikoki egondako zelaiko bareen
populazioetan ere gerta daiteke (Marigómez et al.,
1998a). Ikerlan honetan, Karrantzako bareetan
(utzitako zink-meatzea) aldez aurretik
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
203
9. Irudia: Erreferentzia tokian (Bakio) etaantzinako meatzaren inguruan(Karrantza) hartutako Arion ater barearren liseri-guruinean neurtutako osaketazelularra. (A) Kutsatu gabeko tokitikmeatzera lekuzaldatutako bareak; (B)Meatzetik kutsatu gabeko tokiralekuzaldatutako bareak.
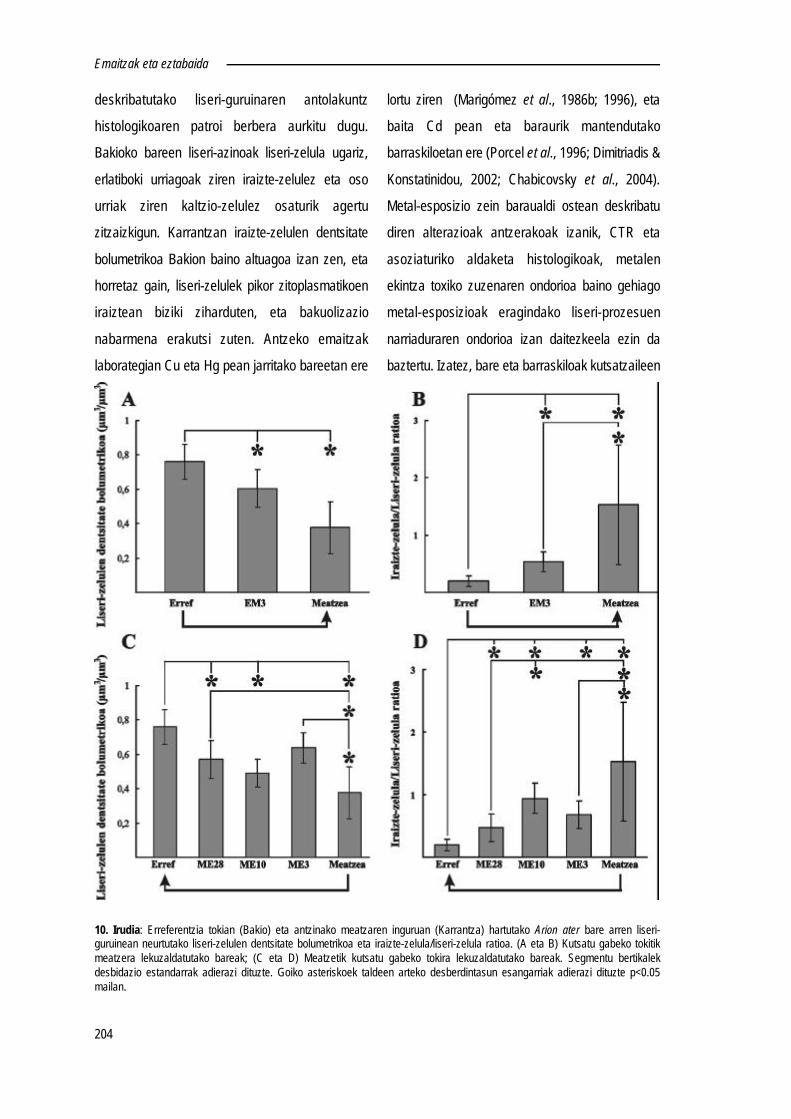
deskribatutako liseri-guruinaren antolakuntz
histologikoaren patroi berbera aurkitu dugu.
Bakioko bareen liseri-azinoak liseri-zelula ugariz,
erlatiboki urriagoak ziren iraizte-zelulez eta oso
urriak ziren kaltzio-zelulez osaturik agertu
zitzaizkigun. Karrantzan iraizte-zelulen dentsitate
bolumetrikoa Bakion baino altuagoa izan zen, eta
horretaz gain, liseri-zelulek pikor zitoplasmatikoen
iraiztean biziki ziharduten, eta bakuolizazio
nabarmena erakutsi zuten. Antzeko emaitzak
laborategian Cu eta Hg pean jarritako bareetan ere
lortu ziren (Marigómez et al., 1986b; 1996), eta
baita Cd pean eta baraurik mantendutako
barraskiloetan ere (Porcel et al., 1996; Dimitriadis &
Konstatinidou, 2002; Chabicovsky et al., 2004).
Metal-esposizio zein baraualdi ostean deskribatu
diren alterazioak antzerakoak izanik, CTR eta
asoziaturiko aldaketa histologikoak, metalen
ekintza toxiko zuzenaren ondorioa baino gehiago
metal-esposizioak eragindako liseri-prozesuen
narriaduraren ondorioa izan daitezkeela ezin da
baztertu. Izatez, bare eta barraskiloak kutsatzaileen
Emaitzak eta eztabaida
204
10. Irudia: Erreferentzia tokian (Bakio) eta antzinako meatzaren inguruan (Karrantza) hartutako Arion ater bare arren liseri-guruinean neurtutako liseri-zelulen dentsitate bolumetrikoa eta iraizte-zelula/liseri-zelula ratioa. (A eta B) Kutsatu gabeko tokitikmeatzera lekuzaldatutako bareak; (C eta D) Meatzetik kutsatu gabeko tokira lekuzaldatutako bareak. Segmentu bertikalekdesbidazio estandarrak adierazi dituzte. Goiko asteriskoek taldeen arteko desberdintasun esangarriak adierazi dituzte p<0.05mailan.

pean mantentzeak, elikatze-jarduera eta
hazkuntzaren murrizpen esangarria eragin dezake
(Russell et al., 1981; Marigómez et al., 1986c;
Laskowski & Hopkin, 1996; 5. Kapitulua). Dena den,
Pb eta Zn, Cu eta Cd baino toxizitate txikiagokoak
dira Helix aspersa barraskiloentzat, eta Cu eta Hg
baino toxizitate txikiagokoak A. ater bareentzat,
behintzat elikatze-jarduera, hazkuntza eta
hilkortasunari dagokienez (Marigómez et al., 1986c;
Laskowski & Hopkin, 1996). Izan ere, bareen
kasuan, janariango 300 µg/kg goragoko Zn
kontzentrazioek baino ez dute eragina liseri-
guruinaren histologian eta glukogeno-gordekinetan
(Recio et al., 1988b). Kurioski, utzitako Cu-meatze
baten aldameneko bareek, Cu, Zn eta Cd
kontzentrazio tisular altuekin batera, gorputz
osoaren eta liseri-guruinaren pisu lehorraren %50-
eko murrizpena erakutsi zuten, leku kutsatu gabeko
erreferentzia-bareekin konparatu zirenean
(Marigómez et al., 1998a). Azkenik, Cu-
(Marigómez et al., 1998a) eta Zn-meatze (ikerlan
hau) abandonatuetako bareek, liseri-zelulen
presentzia erlatiboaren murrizpenaren ezaugarri
amankomuna duten arren, metal-esposizio
kronikoaren bi egoera horietan zelula-moten
osaketa ez da berbera. Cu-meatzeko bareetan,
kaltzio-zelulek liseri-zelulak ordezkatzen dituzte
(Marigómez et al., 1998a); eta Zn-meatzekoetan,
berriz, iraizte-zelulek burutzen dute ordezkapena
(ikerlan hau). Cu eta Zn, beranduago eztabaidatuko
den moduan, bareen liseri-guruineko konpartimentu
zelular desberdinetan lokalizatzen direnez, metal-
kutsadura kronikoaren iturri desberdinen pean
egondako bareen zelula-mota osaketaren
dibergentzia, hori bi detoxifikazio-mekanismo
desberdinen existentziaren ondorioa izan liteke.
Dena dela, liseri-zelulen galera erantzun orokorra
dela dirudi (Porcel et al., 1996; 1., 4. eta 5.
Kapituluak), eta beraz CTRren parametro adierazle
orokor gisa, gaur egun erabilitako VvBAS-en ordez,
VvDIG erabiltzea gomendatuko genuke (4.
Kapitulua) edo VvEXC/VvDIG ratioa (5. Kapitulua).
Kutsatu gabeko tokiko (Bakio) bareen iraizte-
zeluletako lipofuszinetan eta liseri-zeluletako
hondakin-gorputz gutxi batzuetan AMGren bidez
ikustarazitako metalen mailak ertain eta baxuak
bitartean suertatu ziren. Halaber, odol-hodiaren
inguruko ehun konektiboko geruzan BSD
konspikuoak behatu ziren. Egoera hori
erreferentziazko oinarri-balioak dira, eta aldez
aurretik egindako zelai-ikerketa eta laborategiko
esperimentuetan lortutako emaitzekin bat dator
(Marigómez et al., 1996; 1998a). Bestalde,
Karrantzako bareetan iraizte-zelulen lipofuszina-
pikorrek zein liseri-zelulen lisosoma eta hondakin-
gorputzek, eta baita kaltzio-zelulek ere, oso AMG-
markaketa intentsoa erakutsi zuten. AMGk
eskainitako datuen arabera, bareetan Pb
lehendabizi liseri-zelulen lisosometan metatzen da
(argitaratu gabe), beste metalen kasuan, Hg eta Cd
alegia, gertatu bezala (Marigómez et al., 1996;
1998a; 5. Kapitulua). Haatik, Zn liseri-, iraizte- eta
kaltzio-zeluletan lokalizatu da (Schoettli & Seiler
1977; Recio et al. 1988a; Almendros & Porcel,
1992; Triebskorn & Köhler, 1996), esperimentu
honetan egindako AMG bidezko behaketekin bat
datorrena. Lotugai-multzoak zelulaz-zelula
desberdinak dira, eta beraz metal ezberdinak
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
205

zelula-mota ezberdinetan bahitzen dira (Marigómez
et al., 2002). Are gehiago, oxigeno zein sufre
emailekiko elkarkidetasuna duen mugaldeko
metala den Zn-aren metatze-tokiak, espezieen
arabera ere desberdinak izan daitezke (Marigómez
et al., 2002). Esaterako, muskuilu eta magurioetan,
non zelula basofilikoen fosfato-pikorrak bare eta
barraskiloetan bezain garatuak ez baitaude, Zn-a
nagusiki liseri-zelulen lisosometan lokalizatu da
(Soto et al., 1998; 1. Kapitulua). Azkenik, liseri-
azinoen lumenean askatutako pikor zitoplasmatiko
eta lipofuszinetan BSD behatu izanak, liseri-
guruineko zelulak metatzaile baino gehiago
konpartimentu iraizleak direla sustatuko luke
(Marigómez et al., 1990a; 1996; Soto et al. 1996;
Soto & Marigómez, 1997a; 1997b). Gorotzekin
batera metalak kanporatzeak berebiziko garrantzia
du lehorreko gastropodoetan, zeintzuei lehorrean
bizitzeko moldarazpenak deshidratazio-arriskua
baitakarkie, iraizte-fluidoen kanporatzea ahalik eta
murritzen mantentzera behartuta egonik (Riddle,
1981).
VvBSD balioak Karrantzan 10 aldiz altuagoak
dira Bakion baino, AAS bitartez lortutako datuekin
bat datorrena, eta CTRk VvBSD gainean eraginik
ez duela iradoki dezakeena. Aitzitik, bareen kasuan
liseri-zeluletan modu selektiboan metatzen diren
Hg eta Cd moduko metalen pean luzaroan
egondakoan bareetan, BSDen presentzia espero
baino baxuagoa da CTR dela eta (Marigómez et al.,
1996). Ikerlan honetan, non Zn-a liseri-guruinean
metatutako metal nagusia baita (>10 aldiz Pb
kontzentrazio tisularraren gainetik), ez dirudi CTRk
VvBSD eragiten duenik, AMG bitartez Zn-a zelula-
mota guztietan lokalizatu baita.
Metalotioneinak (MT) zeluletan metal-ioi askeen
maila baxuak mantentzeaz arduratzen dira, eta
beraien ekoizpena metaleen presentziaren aurrean
induzigarria da (Dallinger, 1995; Langston et al.,
1998). Metalotioneinak, Cu, Zn, Hg eta Cd ioiei
modu espezifikoan lotzen zaizkie eta, beraz,
eraenketa- (Cu eta Zn metaletarako) edo
detoxifikazio-molekulatzat (Hg edo Cd-rako) jo dira,
moluskuetan hainbatetan erakutsi den moduan
(Ireland, 1981; Roesijadi, 1982; Dallinger et al.,
1989; Bebianno & Langston, 1989; Dallinger, 1993;
Berger & Dallinger, 1993). Ikerlan honetan,
meatzeko eta kutsatu gabeko tokiko bareen liseri-
guruinean espertrofotometrikoki kuantifikatutako
MT- kontzentrazioak ez dira esangarriki
desberdinak. Zn- eta Pb-esposizio kronikoa, MTen
indukzioari asoziatuta ez legokeela pentsa daiteke,
metal hauek fosfato moduko bestelako lotugai
zelularrei lotuta ere egon ahal diren arren. Honekin
lotuta, aipatu beharra dago Zn askotan kaltzio-
zelulen fosfato kaltzikoko pikorretan lokalizatu dela
(Taylor et al., 1990).
Karrantzako bareen liseri-zelulen lisosomak
Bakioko bareenak baino handiagoak izan ziren,
behaketa zuzenen bitartez zein estereologia
aplikatuz frogatu den bezala. Ingurumen-estresaren
neurketa orokorra den lisosomen handiagotzea
(Cajaraville et al., 1995), efektu aski ezaguna da
moluskuen liseri-guruinean (Viarengo et al., 1984;
Marigómez et al., 2005). Zehazkiago, lindanoz
kutsatutako tokietan, lisosomen handiagotzea
lindano kutsadura gradientearekin erlazionatu da
Emaitzak eta eztabaida
206

(Müller, 1994). NvLYS parametroan desberdintasun
esangarriak egon barik, tamaina handiagoko
lisosomek (S/V balio baxuak adierazi duenez),
Karrantzako VvLYS balioak Bakiokoak baino 5 aldiz
handiagoak izatea eragin dute. Efektu hori, BSD
balio altuekin estuki lotuta egon daiteke, eta
kronikoki mantendutako detoxifikazio-jarduera
areagotuaren isla izan daiteke. Kutsatzaileen peko
egoerak jarduera katalitikoen areagotzea eragiten
du moluskuen liseri-zeluletan, zeinek lisosomen
forma eta funtzioen aldaketak azalduko bailituzke
(Marigómez et al., 1986a; Moore & Viarengo, 1990).
Lisosomen handiagotzea, induzitutako autofagia
prozesuen nabaritasuna izan daitekeela iradoki da;
azkenean, induzitutako autofagia horrek, kinadak
behar adina irautekotan, zelulei kaltea eta heriotza
sortarazi diezaiekeelarik (Allen & Moore, 2005).
Testuinguru honetan, afera interesgarri bat
plazaratu da. Halako lisosomen handiagotzeari,
bizialdi osoan zehar eta baita, kronikoki kutsatutako
lekuetako bareetan, belaunaldiz-belaunaldi ere
eutsi dakioke. Beraz, kalte eta heriotza zelularraren
azalpena tentuz aztertu beharko genuke.
Azkenik, muskuiluetan gertatu ez bezala (1.
Kapitulua), ez dirudi CTRk lisosomen gainean
eragina duenik. Arrazoia nagusiki metodologikoa
dela dirudi. Bibalbioetan liseri-zelulak eta zelula
basofilikoak tartekatuta agertzen dira, eta irudi-
analisian neurketak egiten direnean neurtutako
eremuan zelula basofilikoen zatiak erortzea ez da
batere arraroa. Horrela, zelula basofilikoen kopurua
altuagoa den heinean, parametro lisosomikoak
azpiestimatuak izan daitezke sistematikoki.
Gastropodoetan, bestalde, kaltzio-zelulak urriagoak
dira eta taldekaturik agertzen dira; beraz, neurketa-
eremua soilik liseri-zeluletara mugatzea errazagoa
da, baita VvDIG balioa %50 baino baxuago denean
ere.
Metal-kutsadura berriaren aurreko erantzuna
Laborategiko ikerlanetan metalen pean egon
izanak eragindako aldaketa gehienak
probokatzeko, 3 eguneko esposizio-denbora
nahikoa izan zen zelaiko baldintzetan (Marigómez
et al., 1996); ikerlan honetan erabilitako analisi
kimikoak eta esposizio- zein efektu-
biomarkatzaileak, noizbehinkako metal-kutsadura
lurzoruan detektatzeko egokiak direla erakutsiz.
Bakiotik Karrantzara 3 egunez lekuzaldatu
ondoren, liseri-guruineko Pb eta Zn kontzentrazioak
Karrantzako meatzeko bareetan neurtutako
kontzentrazioen mailetara heldu ziren. Bakiotik
Karrantzara lekuzaldatu ondoren, BSDetan
behatutako aldaketak metal-kutsadura berriaren
aurrean ere erantzunkorrak izan ziren. VvBSDbalioak, Karrantzan Bakion baino 10 aldiz altuagoak
zirenak, toki kutsatura 3 egunez lekuzaldatutako
bareetan 3 aldiz altuagoak suertatu ziren. Metalen
metaketa moluskuen liseri-guruinean erantzun
azkarra dena jakina da, egun batez Cd pean
egondako muskuiluen VvBSD balio emendatuek
adierazi duten legez (1. Kapitulua). Aitzitik, Bakiotik
Karrantzara lekuzaldatutako bareetan, liseri-
guruinaren MT kontzentrazioa ez zuen aldaketarik
pairatu, Zn-k eta Pb-k MTen sintesirik eragiten ez
dutenaren seinaletzat har litekeena.
Toki kutsatura 3 egunez transplantatu ondoren,
liseri-zelulen proportzio erlatiboa esangarriki jaitsi
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
207

zen, Karrantzako VvDC balioetara hurbilduz.
Emaitza horiek, aldez aurretiko laborategiko
datuekin bat datoz, non 3 eta 21 egun bitartean
CTR eragiten baita, espezie, kutsatzaile, dosi eta
esposizio-modu eta ingurumen-baldintzen arabera
(1., 4. eta 5. Kapituluak). Liseri-zelulen
proportzioaren beherapena, epe labur eta ertainez
Hg zein Cd + keroseno nahasketaren pean
mantentzean ere behatu da bareen liseri-guruinean
(Marigómez et al., 1996; 5. Kapitulua).
Epe laburreko forma eta tamainaren aldaketa
lisosomikoak, ustekabekoak izan litezke nolabait;
zenbait egun eta zenbait astetako tartean
kutsatzaile pean egonez gero, behin-behineko
aldaketak gerta baitaitezke (Marigómez & Baybay-
Villacorta, 2003). Oro har, kutsatzaile-esposizioak
lisosomen handiagotzea eragiten du (Viarengo et
al., 1984; Marigómez et al., 2005), baina esposizio-
denbora jakinetan lisosomen txikiagotzea
handiagotzearen aurretik joan daiteke (Marigómez
& Baybay-Villacorta, 2003). Ikerlan honetan,
Bakiotik Karrantzara 3 egunez lekuzaldatutako
bareetan, VvLYS eta SvLYS ez ziren aldatu,
handiagotzea (S/VLYS baxua) lisosomen
kopuruaren jaitsiera esangarriarekin batera gertatu
baitzen (NvLYS baxua). Ondorioz, lisosomen
handiagotzea suertatu zen, baina seguruenik
detoxifikatzeko iraizte-jarduera areagotu
izanagaitik, lisosomak gutxiagotu ere egin ziren.
Zoritxarrez, hurrengo egunetako laginak istripuz
galdu ziren, eta beraz lisosomen kopurua
Karrantzako bareetan erregistratu ziren balioetara
beranduago helduko ote zitekeen ezin izan dugu
ikusi.
Kutsadura etetearen aurreko erantzuna
kronikoki kutsatutako bareetan
Toki kutsatutik kutsatu gabeko tokira
lekuzaldatutako bareetan ustekabeko emaitzak
eskuratu ziren.
Hasteko, kutsatu gabeko tokira lekuzaldatu
ondoren, liseri-guruineko metalen orotariko
kontzentrazioa aldaketarik gabe mantendu zen edo
are altuagoa bilakatu zen, meatzeko bareetan
erregistratukoekin alderatuz. Adibidez, Bakion 3
egunez egon eta gero, Pb kontzentrazioa
Karrantzan baino 3 aldiz altuagoa izan zen, eta
Bakion 28 egunez egon ostean Zn kontzentrazioa
jatorrizko lurzoru kutsatuan baino altuagoa ere
suertatu zen. Halaber, VvBSD ez zen txikiagotu.
Aitzitik, kutsatu gabeko tokira 3-10 egunez
transplantatu ondoren, VvBSD jatorrizko leku
kutsatuan baino are altuagoa bilakatu zen.
Bederen, MT kontzentrazioak esangarriki altuagoak
agertu ziren leku kutsatutik kutsatu gabeko tokira 10
egunez lekuzaldatu ondoren. Bitartean,
Karrantzatik Bakiora transplantatutako bareen
liseri-guruineko Cu kontzentrazioak gradualki gora
egin zuen 28 egunetan zehar, erreferentziazko
mailetara heldu arte. Demagun metal kutsatzaileek
lotugai zelularrei lotzeko Cu-arekin lehiatzen baitute
bare kutsatuetan baxua den liseri-guruineko Cu-
zama, kutsadura eten eta 10 egunetara maila
normaletara itzuli zela; nola ahal izan ziren
mantendu Zn eta Pb mailak jatorrizko bare
kutsatuetan bezain altu edo are altuagoak? Zn eta
Pb-aren kanpo-iturri bat ez bazegoen, metal horiek
liseri-guruinera beste ehunetatik heldu behar izan
Emaitzak eta eztabaida
208

zuten. Gastropodoetan, nefrozitoek, errogozitoek
eta hemozitoek Zn eta Pb metatzen dutena jakina
da (Soto et al., 1996). Hemozito eta errogozito
bidezko metalen mobilizazioa liseri-guruinerantz,
metal toxikoen kanporatze masiborako mekanismo
gisa proposatu da (Marigómez et al., 2002). Izatez,
liseri-guruinaren funtzio nagusia metalen metaketa
izan ezik metalen iraizpena dela iradoki da
(Marigómez et al., 2002). Hori dela eta, liseri-
guruineko Zn- eta Pb-zama altuek, metalen
mobilizazio areagotu izana islatuko lukete,
gorotzekin batera kanporatuak izateko organo
honetaranzko metalen mobilizazio intentsifikatua
adieraziko luketena. Horrekin bat, Karrantzara
lekuzaldatutako bareen liseri-dibertikuluetako
lumenera askatutako hondakin zelularretan
metalen presentzia nabarmena adierazi du AMGk.
Metalen mobilizazio areagotuak ere, kutsadura eten
osteko liseri-guruineko Cu kontzentrazioaren
emendioa azal lezake. Emaitza interesgarria, liseri-
guruineko Pb kontzentrazioaren piko maximoa (3.
eguna) Zn-arenaren (28. eguna) aurretik joan izana
dugu, Pb errezago detoxifikatuko balitz azaldu
litekeena, hain zuzen. Izatez, Pb kontzentrazioa
ehunetan Zn-arena baino askoz baxuagoa da, eta
gainera, bion konpartimentu zelularrak eta lotugaiak
desberdinak izan daitezke (Marigómez et al., 2002).
Metal-kontzentrazioen igoera horiek behin-
behinekoak diren ala kronikoki kutsatutako bareek,
metal-kutsatzailerik gabe mantenduta ere, metalak
liseri-guruinetik kanporatu ezin dituztenez behin
betikoak diren finkatzeko, kutsadura eten ondoko
epe esperimental luzeagoak ezinbestekoak dira.
Bigarrenez, CTR, aldez aurretik eztabaidatu
bezala, kronikoki kutsatutako bareetan mantentzen
den erantzun azkarra izan arren, VvDIG-ren igoera
gradualak patroi korapilotsuagoa jarrai dezakeela
dirudi. 10 egunez kutsatu gabeko tokira
transplantatutako bareetan VvDIG baxuagoa da 3
eta 28 egunez transplantatutakoetan baino eta, are
gehiago, ez da Zn-meatzeko bareen VvDIG-ren
esangarriki desberdina. Liseri-guruinaren zelula-
moten osaketa normala berreskuratzea liseri-
zelulen proliferazio gaitasunean oinarritzen bada,
prozesu hau erritmikoa izango litzateke, eta
ingurugiro aldagaien eta aldagai indibidualen
menpean egongo litzateke, muskuiluen liseri-
guruinean deskribatu den bezala (Zaldibar et al.,
2004; 3. Kapitulua). Lehorreko gastropodoetan,
giltzurruneko histofisiologian ziklo semilunarrak
deskribatu dira (Capellán et al., 1990), eta beraz
liseri-guruinean antzeko aldamoldeen presentzia
ezin da baztertu. Horrek, Zn-meatzetik kutsatu
gabeko tokira lekuzaldatutako bareen liseri-
guruinean behatutako VvDIG-ren patroi
korapilotsua azal lezake. Are gehiago, Karrantzatik
Bakiora 28 egunez lekuzaldatutako bareetan ere,
VvDIG balioek Bakion neurtutakoen desberdinak
izaten zerraiten. Laborategian metal pean 3-4 astez
mantendutako eta, detoxifikatzeko asmoz, 1-2
astez kutsatzailerik gabe mantendutako bare eta
magurioetan, detoxifikazio-denbora hori nahikoa
izan zen liseri-guruinak kontrolen antzeko zelula-
moten osaketa berreskuratzeko (4. eta 5.
Kapituluak). Kutsadura kronikoaren ostean, CTR
guztiz itzulgarria denentz ikusteko, kutsadura eten
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
209

osteko denbora esperimental luzeagoak
ezinbestekoak dira.
Azkenik, toki kutsatutik kutsatu gabeko tokira 28
egunez lekuzaldatutako bareen liseri-zelulen
lisosomek ez zuten inolako erantzunik eman.
Haatik, Cd pean jarritako eta ondoren 14 egunez ur
garbitan arazten utzitako magurioen lisosomak
kontrol taldearen berdintsuak bilakatu ziren (4.
Kapitulua). Metal-kutsadura kroniko pean egondako
bareak, berriz, ez dirudi berreskuratzeko gai direnik,
edo behintzat 28 egun ez dirudi nahiko direnik, eta
lisosomek handiak izaten eta metal-iraizteak
areagoturik egoten darraite.
Ingurumen-biojarraipenerako garrantzizko
gogoetak
Aldez aurretik egindako ikerlanetan, animaliak
metal kontzentrazio desberdinen pean asteetan
zehar mantendu dira, baina ingurumen naturaletan
animaliak bizialdi osoan zehar metalen eragin
kroniko pean egon daitezke, euren erantzunak
nolakoak diren aztertzea jite oso interesgarria izanik
(Wang & Rainbow, 2005). Ikerlan honetan lortutako
emaitzen arabera, bareen liseri-guruinean behatu
diren ezaugarriak, metalen eragin pean aste
batzuetan zehar behatutako erantzunetatik oso
desberdinak ez dira izan. (Marigómez et al., 1986a;
1986b; 1996). Aitzitik, kronikoki kutsatutako bareen
liseri-guruinak ez du erantzun kutsadura eten
denean, edo behintzat erantzuteko gaitasun
murritzagoa adierazi du (5. Kapitulua). Hortaz,
antzekotasunak soilik azaleko ezaugarriei
legokieke, izan ere, erantzuteko gaitasuna eta
plastizitatea nabarmenki afektaturik geratu dira, eta
horrek organismoaren fisiologian eta baita
biomarkatzaileen eta bestelako kutsaduraren
esposizioaren eta efektuen neurketetan ere eragina
izan lezake. Aspaldidanik dakigunez, aldez
aurretiko metal-esposizioek hurrengo esposizioen
aurrean babes-mekanismoen areagotzea eragin
dezakete (Wang & Rainbow, 2005); baina,
berreskuratzeko gaitasun hori laborategiko
esposizio-esperimentuetan erregistratutakoa baino
mugatuagoa izatearen berri ez da oraingo lan
honetararte azaldu. Hortaz, kronikoki kutsatutako
populazioetan espero daitekeen suspertzea
laborategiko esperimentuetan oinarrituz
aurresateko, kontu handiz ibili beharra dago .
Bigarren ondorio deigarria kutsadura
kronikoaren inguruan biomarkatzaileen erabilerak
sor ditzaketen ondorioen oker eta mugen ingurukoa
da. Laborategiko esperimentuetan oinarrituz, Wu et
al.-ek (2005) kutsadura eteten denean
biomarkatzaile batzuk (entzimen indukzioa,
lisosomen integritatea) zeharo itzulgarriak direla eta
beste batzuk (kalte zelularra, patologiak), ordea,
itzulezinak edo iraunkorrak direla ondorioztatu
zuten. Autore horiek azpimarratu zutenez, nahiz eta
biomarkatzaile gehienak itzulgarriak izan,
itzulezinak direnak emaitza sasipositiboak eman
ditzakete, baldin eta ingurumen-baldintzak
nabarmenki ez hobetzekotan behintzat. Ikerlan
honetan, laborategian zenbait astetako
esposizioaren ondorioz itzulgarriak diren hainbat
erantzun biologiko (erantzun lisosomikoak,
adibidez), kutsadura kronikoaren ostean itzulgarriak
ez direla, edo behintzat modu azkarrean itzulgarriak
ez direla egiaztatu da. Kutsatzaileen aurreko
Emaitzak eta eztabaida
210

erantzun lisosomikoak ordu gutxiren buruan induzi
daitezkeena badakigunez, ikerlan honetan Bakiotik
Karrantzara lekuzaldatu ondoren erantzun
lisosomikoak eta metaleen metaketa
intralisosomikoa 3 egunetan gertatu izanak
egiaztatu duen bezala, 28 egunez Karrantzatik
Bakiora lekuzaldatzean inolako erantzunkortasunik
aurkitu ez izana harrigarria suertatu zaigu. Izan ere,
moldarazpen genetikoak metalen presentzian
bizirauteko gai ziren indibiduoak hautatu izan balitu,
susperketa ez zitekeen inoiz gertatuko. Hala ere,
erantzunkaiztasun hori, susperraldia luza dezakeen
plastizitatearen galeraren ondorioa ere izan
daiteke. Galdera horri erantzuteko asmoz,
susperraldi luzeagoko esperimentuak martxan
jarriko dira kronikoki kutsatutako bareak erabiliz.
Dena den, erantzuna zein den jakin ala ez,
biomarkatzaileak interpretatzean daukagun arazoa
ezin dugu gainetik kendu: biomarkatzaileak ez dira
euren maila basaletara itzultzen edo, behintzat, oso
astiro itzultzen dira. Susperraldia, biomarkatzaile,
kutsatzaile eta espeziearen sentikortasunaren
arabera aldakorra dela ere deskribatu da (Wu et al.
2005). Hala ere, gure ekarpenaren arabera,
susperraldia kutsadurak zenbat denboraz eragin
duen araberakoa izan daiteke. Beraz, kontu handiz
ibili beharra dago "Slug-Watch" moduko
biojarraipen-programetan biomarkatzaileak
kronikoki kutsatutako populazioetan aplikatzen
direnean: (a) biomarkatzaileen erantzuna
motelduta/aldatuta egon daiteke kutsadurarekiko
moldarazpenaren ondorioz (Wu et al., 2005); eta (b)
biomarkatzaileak kutsadura kronikoaren ondorioz
ez dira lehengoratzen, edo behintzat, ingurumen-
baldintzak hobatzekotan, kronikoki eta noizbehinka
edo behin-behinekoz kutsatutako populazioetan
susperraldia desberdina da (Wu et al., 2005; 5.
Kapitulua). Biomarkatzaileen gaineko
interpretazioak egiteko datu gehiago ezinbestekoak
dira. Bareetan, beste datu osagarrien artean, liseri-
guruinaren azterketa histopatologikoek bareak
kutsadura kroniko pean egon direnentz argitzeko
oso tresna baliagarria eskaini diezaigukete
(Marigómez et al., 1998a).
BIBLIOGRAFIA
Allen JI, Moore MN (2005) Environmental prognostics: Is the
current use of biomarkers appropiate for environmental risk
evaluation?. Mar. Environ. Res. 58:227-232.
Almendros A, Porcel D (1992) Phosphatase activity in the
hepatopancreas of Helix aspersa. Comp. Biochem.
Physiol. A 103:455-460.
Bebianno MJ, Langston WJ (1989) Concentration of metals
and metallothioneins in marine invertebrates using
differential pulse polarography. Portugaliae Electrochimica
Acta 7:511-524.
Beeby A, Richmond L (1987) Adaptation by an urban population
of the snail Helix aspersa to a diet contaminated with lead.
Environ. Pollut. 46:73-82.
Beeby A, Richmond L (2002) Evaluation of Helix aspersa as a
sentinel for mapping metal pollution. Ecol. Int. 1:261-270.
Berger B, Dallinger R (1989) Accumulation of cadmium and
copper by the terrestrial snail Arianta arbustorum L.:
kinetics and budgets. Oecologia 79:60-65.
Berger B, Dallinger R (1993) Terrestrial snails as quantitative
indicators of environmental metal pollution. Environ.
Monitor. Assess. 25:65-84.
Cajaraville MP, Díez G, Marigómez I, Angulo E (1990a)
Responses of basophilic cells of the digestive gland of
mussels to petroleum hydrocarbon exposure. Dis. Aquat.
Org. 9:221-228.
Cajaraville MP, Marigómez I, Angulo E (1990b) Short-term toxic
effects of 1-naphthol on the digestive gland-gonad complex
of the marine prosobranch Littorina littorea (L.): a light
microscopic study. Arch. Environ. Contam. Toxicol. 19:17-
24.
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
211

Cajaraville MP, Marigómez JA, Angulo E (1991) Automated
measurement of lysosomal structure alterations in oocytes
of mussels exposed to petroleum hydrocarbons. Arch.
Environ. Contam. Toxicol. 21:395-400.
Cajaraville MP, Marigómez I, Díez G, Angulo E (1992)
Comparative effects of the water accommodated fractions
(WAF) of three oils on mussels. 2.- Quantitative alterations
in the structure of the digestive tubules. Comp. Biochem.
Physiol C 102:113-123.
Cajaraville MP, Marigómez I, Angulo E (1993) Correlation
between cellular and organismic responses to oil-induced
environmental stress in mussels. Sci. Tot. Environ. Suppl.
1:1353-1371.
Cajaraville MP, Abascal I, Etxeberria M, Marigómez I (1995)
Lysosomes as cellular markers of environmental pollution:
time- and dose-dependent responses of the digestive lyso-
somal system of mussels after petroleum hydrocarbon
exposure. Environ. Toxicol. Water Qual. 10:1-8.
Capellán A, Angulo E, Mateo A (1990) Histochemistry of glyco-
gen and uric acid in the kidney sac of Cernuella virgata (da
Costa, 1778) (Mollusca, Pulmonata): monthly and daily
variations. Z. Mikrosk. Anat. Forsch. 104:147-154.
Chabicovsky M, Klepal W, Dallinger R (2004) Mechanisms of
cadmium toxicity in terrestrial pulmonates: Programmed
cell death and metallothionein overload. Environ. Toxicol.
Chem. 23:648-655.
Coughtrey PJ, Martin MH (1977) The uptake of lead, zinc,
cadmium, and copper by the pulmonate mollusc, Helix
aspersa Müller, and its relevance to the monitoring of
heavy metal contamination of the environment. Oecologia
27:65-74.
Dallinger R, Wieser W (1984) Patterns of accumulation,
distribution and liberation of zinc, copper, cadmium, and
lead in different organs of the land snail Helix pomatia L.
Comp. Biochem. Physiol. C 79:117-124.
Dallinger R, Janssen HH, Bauer-Hilty A, Berber B (1989)
Characterization of an inducible cadmium-binding protein
from hepatopancreas of metal exposed slugs (Arionidae,
Mollusca). Comp Biochem Physiol C 92:355-360.
Dallinger R (1993) Strategies of metal detoxification in
terrestrial invertebrates. 245-289 orr. Non: Dallinger R
Rainbow PS (ed) Ecotoxicology of metals in invertebrates,
Lewis Publ., Boca Raton, FL.
Dallinger R. (1994) Invertebrate organisms as biological
indicators of heavy metal pollution. Appl. Biochem.
Biotechnol. 48:27-31.
Dallinger R (1995). Mechanisms of metal incorporation into
cells. 135-154 orr. Non: Cajaraville MP (ed.) Cell biology in
environmental toxicology, University of the Basque Country
Press Serv, Bilbo, Basque Country.
Dallinger R, Chabicovsky M, Berger B (2004) Isoform-specific
quantification of metallothionein in the terrestrial gastropod
Helix pomatia. I. Molecular, biochemical, and
methodological background. Environ. Toxicol. Chem.
23:890-901.
Danscher G (1984) Autometallography. A new technique for
light and electron microscopic visualization of metals in
biological tissues (gold, silver, metal sulphides and metal
selenides). Histochemistry 81:331-335.
Dimitriadis VK, Konstantinidou V (2002) Origin of excretory
cells in the digestive gland of the land snail Helix lucorum.
Malacologia 44:145-151.
Greville RW, Morgan AJ (1991) A comparison of (Pb, Cd and Z)
accumulation in terrestrial slugs maintained in microcosms:
Evidence for metal tolerance. Environ. Poll. 74:115-127.
Hopkin SP, Hames CAC, Dray A (1989) X-ray microanalytical
mapping of the intracellular distribution of pollutant metals.
Eur. J. Microsc. Anal. 11:19-23.
Ireland MP, Wootton RJ (1975) Variation in the lead, zinc and
calcium content of Dendrobaena rubida (Oligochaeta) in a
base metal mining area. Environ. Pollut. 10:201-208.
Ireland MP (1979) Distribution of essential and toxic metals in
the terrestrial gastropod Arion ater. Environ. Pollut. 20:271-
278.
Ireland MP (1981) Uptake and distribution of cadmium in the
terrestrial slug Arion ater (L). Comp. Biochem. Physiol. A
73:855-858.
Ireland MP (1982) Sites of water, zinc and calcium uptake and
distribution of these metals after cadmium administration in
Arion ater (Gastropoda: Pulmonata). Comp. Biochem.
Physiol. A 73:217-221.
Ireland MP (1984a) Seasonal changes in zinc, manganese,
magnesium, copper and calcium contents in the digestive
gland of the slugs Arion ater. Comp. Biochem. Physiol. A
74:855-858.
Ireland MP (1984b) Effect of chronic and acute lead treatment
in the slug Arion ater on calcium and d-aminolaevulinic acid
dehydratase activity. Comp. Biochem. Physiol. C 79:287-
290.
Ireland MP, Marigómez I (1992) The influence of dietary
calcium on the tissue distribution of Cu, Zn, Mn and P and
Emaitzak eta eztabaida
212

histological changes in the digestive cells in the snail
Achatina fulica Bowdich. J. Moll. Stud. 58:157-168.
Ireland MP (1994) Interaction and effects of molybdenum
compounds on growth and mineral content of Achatina
fulica and Arion ater (Gastropoda: Pulmonata). Comp.
Biochem. Physiol. C 107:441-446.
Janssen HH, Dallinger R (1991) Diversification of cadmium-
binding proteins due to different levels of contamination in
Arion lusitanicus. Arch. Environ. Contam. Toxicol. 20:132-
137.
Kammenga JE, Dallinger R, Donker MH, Kohler HR, Simonsen
V, Triebskorn R, Weeks JM (2000) Biomarkers in terrestrial
invertebrates for ecotoxicological soil risk assessment.
Rev. Environ. Contam. Toxicol. 164:93-147.
Langston WJ, Bebianno MJ, Burt GB (1998) Metal handling
strategies in mollouscs. 219-293 orr. Non: Langston WJ &
Bebbiano MJ (ed). Metal metabolism in aquatic
environments. Chapman & Hall, London.
Laskowski R, Hopkin SP (1996) Effect of Zn, Cu, Pb, and Cd on
fitness in snails (Helix aspersa). Ecotoxicol. Environ. Saf.
34:59-69.
Lowe DM, Moore MN, Clarke KR (1981) Effects of oil in the
digestive cells in mussels: quantitative alterations in
cellular and lysosomal structure. Aquat. Toxicol. 1: 213-
226.
Lowe DM, Clarke KR (1989) Contaminant-induced changes in
the structure of the digestive epithelium of Mytilus edulis.
Aquat. Toxicol. 15:345-358.
Marigómez I, Angulo E, Moya J (1986a) Copper treatment of
the digestive gland of the slug Arion ater L. 1. Bioassay
conduction and histochemical analysis. Bull. Environ.
Contam. Toxicol. 36:600-607.
Marigómez I, Angulo E, Moya J (1986b) Copper treatment of
the digestive gland of the slug Arion ater L. 2.
Morphometrics and histophysiology. Bull. Environ. Contam.
Toxicol. 36:608-615.
Marigómez I, Angulo E, Saez V (1986c) Feeding and growth
responses to copper, zinc, mercury and lead in the
terrestrial gastropod Arion ater (Linné). J. Moll. Stud.
52:68-78.
Marigómez I, Cajaraville MP, Angulo E (1990a) Cellular
cadmium distribution in the common winkle, Littorina
littorea (L.) determined by X-ray microprobe analysis and
histochemistry. Histochemistry 94:191-199.
Marigómez I, Cajaraville MP, Angulo E (1990b) Histopathology
of the digestive gland-gonad complex of the marine
prosobranch Littorina littorea exposed to cadmium. Dis.
Aquat. Org. 9: 229-238.
Marigómez I, Soto M, Angulo E (1992) Seasonal variability in
the quantitative structure of the digestive tubules of
Littorina littorea. Aquat. Living. Resour. 2:299-305.
Marigómez I, Soto M, Etxeberria M, Angulo E (1993) Effects of
size, sex, reproduction, and trematode infestation on the
quantitative structure of digestive tubules in stressed
winkles. Zool. Jb. Anat. 123:319-336.
Marigómez I, Cajaraville MP, Quincoces I, Salisbury JR (1995)
Computer assisted 3-d reconstruction techniques may
provide new insides into the pattern of histological
organization of the bivalvian digestive gland. 12th Int.
Malacol. Congr. Vigo.
Marigómez I, Soto M, Cajaraville MP (1995) Morphofunctional
patterns of cell and tissue systems involved in metal
handling and metabolism. 89-134 orr. Non: Cajaraville MP
(ed.) Cell biology in environmental toxicology, University of
the Basque Country Press Serv, Bilbo, Basque Country.
Marigómez I, Soto M, Kortabitarte M (1996) Tissue-level
biomarkers of biological effect of mercury on sentinel slugs,
Arion ater. Arch. Environ. Contam. Toxicol. 31:54-62.
Marigómez I, Kortabitarte M, Dussart, GBJ (1998a) Tissue-
level biomarkers in sentinel slugs as cost-effective tools to
assess metal pollution in soils. Arch. Environ. Contam.
Toxicol. 34:167-176.
Marigómez I, Cajaraville MP, Soto M, Lekube X (1998b) Cell-
type replacement, a succesful strategy of molluscs to adapt
to chronic exposure to pollutants. Cuad. Invest. Biol.
18:431-435.
Marigómez I, Lekube X, Cancio I (1999) Immunochemical loca-
lisation of proliferating cells in mussel digestive gland tis-
sue. Histochem. J. 31:781-788.
Marigómez I, Soto M, Cajaraville MP, Angulo E, Giamberini L
(2002) Cellular and subcellular distribution of metals in
molluscs. Micr. Res. Tech. 56:358-392.
Marigómez I, Baybay-Villacorta L (2003) Pollutant-specific and
general lysosomal responses in digestive cells of mussels
exposed to model organic chemicals. Aquat. Toxicol.
64:235-257.
Marigómez I, Soto M, Cancio I, Orbea A, Garmendia L,
Cajaraville MP (2005) Cell and tissue biomarkers in mus-
sel, anh histopathology in hake and anchovy from Bay of
Biscay after the Prestige oil spill (Monitoring Campaign
2003). Mar. Poll. Bull. (prentsan).
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
213

Emaitzak eta eztabaida
214
Mix MC, Sparks AK (1971) Repair of digestive tubule tissue of
the pacific oyster, Crassostrea gigas, damaged by ionizing
radiation. J. Invert. Pathol. 17:172-177.
Moore MN, Viarengo A (1990) Lysosomal membrane fragility
and catabolism of cytosolic proteins: evidence for a direct
relationship. Experientia 43:320-323.
Müller S (1984) Lindanuntersuchungen im Aquatischen und
Terrestrischen Bereich in Bilbao (nordostspanien).
Lizentziatura-Tesia. Universität Karlsruhe. 93 orr.
Pihan F, de Vaufleury A (2000) The snail as a target organism
for the evaluation of industrial waste dump contamination
and the efficiency of its remediation. Ecotoxicol. Environ.
Saf. 46:137-147.
Popham JD, D'Auria M (1980) Arion ater (Mollusca: Pulmonata)
as an indicator of terrestrial environmental pollution. Water
Air Soil Poll. 14:115-124.
Porcel D, Bueno JD, Almendros A (1996) Alterations in the
digestive gland and shell of the snail Helix aspersa Muller
(Gastropoda, Pulmonata) after prolonged starvation.
Comp. Biochem. Physiol. A 115:11-17.
Rasmussen LPD, Hage E, Karlog O (1983) Light and electron
microscopic studies of the acute and chronic toxic effects
of N-nitroso compounds on the marine mussel, Mytilus
edulis (L) II. N-methyl-N-nitro-N-nitrosoguanidine Aquat.
Toxicol. 3:301-311.
Recio A, Marigómez I, Angulo E, Moya J (1988a) Zinc treatment
of the digestive gland of the slug Arion ater L. 1. Cellular
distribution of zinc and calcium. Bull. Environ. Contam.
Toxicol. 41:858-864.
Recio A, Marigómez I, Angulo E, Moya J (1988b) Zinc treatment
of the digestive gland of the slug Arion ater L. 2. Sublethal
effects at the histological level. Bull. Environ. Contam.
Toxicol. 41:865-871.
Riddle WA (1981) Cold-hardiness in several species of land
snail. J. Ther. Biol.13:163-167.
Roesijadi G (1982) Uptake and incorporation of mercury into
mercury-binding proteins of gills of Mytilus edulis as a
function of time. Mar. Biol. 66:151-157.
Russell LK, DeHaven JI, Botts RP (1981) Toxic effects of
cadmium on the garden snail (Helix aspersa). Bull.
Environ. Contam. Toxicol. 30:245-251.
Schoettli G, Seiler HG (1977) Uptake and localization of
radioactive zinc in the visceral complex of the land
pulmonate Arion rufus. Experientia 26:1212-1213.
Snyman RG, Reinecke AJ, Reinecke SA (2005) Quantitative
changes in the digestive gland cells of the snail Helix
aspersa after exposure to fungicide copper oxychloride.
Ecotox. Environ. Saf. 60:47-52.
Soto M (1995) Simultaneous quantification of bioavailable
metals in mollusks by means of cellular and tissue
analysis. Implications for monitoring metal pollution in
water quality assessment. Doktoradutza-Tesia 330 orr.
Soto M, Agirregoikoa MG, Pérez MA, Marigómez I (1990) A
planimetric study of morphological variability in the
digestive diverticula of Littorina littorea (Linnaeus) and
Mytilus edulis Linnaeus. J. Moll. Stud. 56:339-344.
Soto M, Kortabitarte M, Marigómez I (1995) Bioavailable heavy
metals in estuarine waters as assessed by metal/shell-
weight indices in sentinel mussels Mytilus galloprovincialis.
Mar. Ecol. Prog. Ser. 125:127-136.
Soto M, Cajaraville MP, Angulo E, Marigómez I (1996)
Autometallographic localization of protein-bound copper
and zink in the common winkle, Litttorina littorea: a light
microscopical study. Histochem. J. 28:689-701.
Soto M, Marigomez I (1997a) BSD extent, an index for metal
pollution screening based on the metal content within
digestive cell lysosomes of mussels as determined by
autometallography. Ecotox. Environ. Saf. 37:141-151.
Soto M, Marigomez I (1997b) BSD extent, an index for metal
pollution screening based on the metal content within
digestive cell lysosomes of mussels as determined by
autometallography. Mar. Ecol. Prog. Ser. 156:141-150.
Soto M, Quincoces I, Lekube X, Marigomez I (1998)
Autometallographed metal content in digestive cells of
winkles: a cost-effective screening tool for monitoring Cu
and Zn pollution. Aquat. Toxicol. 40:123-140.
Soto M, Zaldibar B, Cancio I, Taylor MG, Turner M, Morgan AJ,
Marigómez I (2002) Subcellular distribution of cadmium
and its cellular ligands in mussel digestive gland cells as
revealed by combined autometallography and X-ray micro-
probe analysis. Histochem. J. 34:273-280.
Sumner AT (1965) The cytology and histochemistry of the
digestive gland cells of Helix. Q. J. Microsc. Sci. 106:173-
192.
Taylor MG, Graves GN, Simkiss K (1990) Biotransformation of
intracellular minerals by zinc ions in vivo and in vitro. Eur.
J. Biochem. 192:783-789.
Thompson RJ, Ratcliffe NA, Bayne BL (1974) Effects of
starvation on structure and function of the digestive gland
of the mussel (Mytilus edulis L) J. Mar. Biol. Assoc. U.K.
54:699-712.

Triebskorn R, Köhler HR (1996) The impact of heavy metals on
the grey garden slug, Deroceras reticulatum (Muller): Metal
storage, cellular effects and semi-quantitative evaluation of
metal toxicity. Environ. Poll. 93:323-343.
UNEP/RAMOGE (1999) Manual on the biomarkers recommen-
ded for the MED POL biomonitoring programme. Athens.
UNEP.
Vega MM, Marigómez I, Angulo E (1989) Quantitative
alterations in the structure of the digestive cell of Littorina
littorea on exposure to cadmium. Mar. Biol. 103:547-553.
Viard B, Pihan F, Promeyrat S, Pihan JC (2004) Integrated
assessment of heavy metal (Pb, Zn, Cd) highway pollution:
bioaccumulation in soil, Graminaceae and land snails.
Chemosphere 55:1349-1359.
Viarengo A, Pertica M, Mancinelli G, Orunesu M, Zanichi G,
Moore MN, Pipe RK (1984) Posible role of lysosomes in
the detoxication of copper in digestive gland cells of metal
exposed mussels. Mar. Environ. Res. 14:469-470.
Wang WX, Rainbow PS (2005) Influence of metal exposure
history on trace metal uptake and accumulation by marine
invertebrates. Sci. China C. Life Sci. 48: 110-117.
Weibel ER (1979) Stereological Methods. 1. Academic Press,
London 415 orr.
Widdows J, Bakke T, Bayne BL, Donkin P, Livingstone DR,
Lowe DM, Moore MN, Evans SV, Moore SL (1984)
Responses of Mytilus edulis on exposure to the WAF of
North Sea oil. Mar. Biol. 67:15-31.
Wu RSS, Siu WHL, Shin PKS (2005) Induction, adaptation and
recovery of biological responses: Implications for
environmental monitoring. Mar. Poll. Bull. 51:623-634.
Yonge CM (1926) The digestive diverticula in Lamellibranchia.
Trans. Roy. Soc. Edin. 54: 703-718.
Zaldibar B, Cancio I, Marigómez I (2002) Model slug to investi-
gatethe role of calcium cells in adaptation in chronic metal-
pollution. SETAC Europe 12th Meeting. Viena.
Zaldibar B, Cancio I, Marigómez I (2004) Circatidal variation in
epithelial cell proliferation in the mussel digestive gland
and stomach. Cell Tiss. Res. 318:395-402.
Zelula-moten ordezkapena bareen liseri-guruinean: lekuzaldatze-ikerketa
215

Emaitzak eta eztabaida
216

1.- Kutsatzaile organiko, ez-organiko zein bien nahasketaren pean jartzen direnean,
Mytilus galloprovincialis muskuiluan zelula-moten ordezkapena ematen da, zelula
basofilikoak liseri-zelulak ordezkatzen dituztelarik; dena den, ordezkapen zelularra
emateko beharrezkoa den kontzentrazio eraginkorra kutsatzaile bakoitzarentzat
desberdina da.
2.- Mytilus galloprovincialis muskuiluan kutsatzaileek eragindako zelula-moten
ordezkapena, estres-iturria eteten denean itzulgarria da; hala ere, zelula-moten
ordezkapenaren gainean, kutsatzaileak nahasketetan agertzearen eta muskuiluak aldez
aurretik estres-egoeran egotearen gisako faktoreek erantzun-patroian eragina eduki
dezakete.
3.- Mytilus galloprovincialis muskuiluaren liseri-guruinaren ehunean kadmioa
detektatzeko erabili den autometalografia bitartez errebelatutako zilarrezko hauspeakin
beltzak, liseri-zelulen lisosomen barnean espezifikoki lokalizatu dira. Lokalizazio espezifiko
hori dela eta, zelula-moten ordezkapenak zilarrezko hauspeakin beltzen dentsitate
bolumetrikoaren balioen gainean eragina dauka, metala metatzeko konpartimentuaren
txikitzearen ondorioz.
4.- Mytilus galloprovincialis muskuiluaren liseri-guruineko liseri-zeluletan irudi mitotikoak
behatu izanak eta BrdU immunohistokimikaren emaitzek, zelulen berriztapena liseri-
guruineko zelula helduen zatiketa autologoaren bitartez gauzatzen dela egiaztatu dute.
Urdaileko eta liseri-guruineko unitate albeolotubularretako epitelioek, zelulen berriztapen-
tasa ertaina eta altua bitartekoa dute. Epitelio-zelulen berriztapen hori sinkronizatuta dago,
24 ordutan 2 ziklo osoak bete dituen aldamolde zirkamareala jarraituz.
5.- Mytilus galloprovincialis muskuiluaren liseri-guruineko epitelioaren berriztapena,
muskuiluaren itsasaldi-erregimen eta urteko sasoiaren arabera alda daiteke. Horrela,
mareazpiko muskuiluetan egunero 2 ziklo ikusi diren bitartean, marearteko muskuiluetan
beti uretatik kanpo ematen den zelula proliferazioaren piko bakarra ikusi da, epitelioaren
berriztapen-dinamika desberdina izanik. Gainera liseri-guruineko zelulek, udazkenean eta
neguan baino proliferazio-jarduera bortitzagoa udan erakutsi dute; areagotutako
berriztapena, seguruenik, udan ematen den metabolismo-tasa altuarekin eralzionatuta
egonik. Muskuilu gazte eta helduek, proliferazio-maila eta berriztapen-dinamikaren
aldamolde beretsua agertu dute, urte-sasoi zein itsasaldi-erregimen desberdinetan.
219
Ondorioak eta tesia

6.- Kadmio pean jarritako Littorina littorea magurioan behatutako zelula-moten
ordezkapena, liseri-zelulen galerak eta aldibereko zelula basofilikoen hipertrofiak sortua
da; prozesu hori, muskuiluetan gertatzen den moduan, estres-iturria etetean itzulgarria
delarik. Liseri-zelulen proliferazioa, kadmio pean mantendutako magurioetan areagotua
agertu da, eta tendentzia hori, arazte-aldiaren ondoren mantendu da. Gainera, liseri-
zelulen galerak eta berreskurapenak, ikerlan honetan erabilitako maila zelular eta
tisularreko biomarkatzaileen gainean eraginik ez duela dirudi; seguruenik, magurioen
kasuan zelula basofilikoak taldekaturik agertzen direlako eta, beraz, mikroskopioa erabiliz
neurketak egiteko errez bazter daitezkeelako.
7.- Kutsatzaile organiko eta ez-organikoen nahasketa pean jarritako Arion ater bareen
liseri-guruinean zelula-moten ordezkapena gertatu da. Dena den, zelula-moten osaketa
berriak, ez du liseri-guruinaren metaketa-gaitasunean eta erabilitako maila zelular eta
tisularreko biomarkatzaileen gainean eraginik izan. Hiru zelula-motez osatutako bareen
liseri-guruinean, liseri-zelulen galera netoa suertatu beharrean, liseri-zelulak iraizte-zelula
faserantz pasatzea areagotu egin da.
8.- Metal-kutsadura kroniko pean hainbat belaunaldiz egondako Arion ater bareetan
liseri-guruineko epitelioa zelula-moten osaketan aldatutak gertatu dira. Hala ere, liseri-
zelulen galerak, erabilitako zelula- eta ehun-maileko biomarkatzaileen gainean ez du
eraginik izan, zelula-mota guztiak aztertutako metaleen (Zn, Pb) metatzaileak baitira.
Bestalde, kronikoki kutsatutako bareen populazioak kutsatu gabeko tokira
lekuzaldatutakoan, biomarkatzaileen erantzuna motelduta/aldatuta ageri da, seguruenik
kutsadurarekiko moldarazpenaren ondorioz. Gainera, biomarkatzaileak, kutsadura
kronikoaren ostean ez dira lehengoratzen; edo behintzat, ingurumen-baldintzak
hobatzekotan susperraldiaren iraupena, kronikoki eta noizbehinka kutsatutako
populazioetan desberdina da.
220
Ondorioak eta tesia

TESIA
Moluskuen liseri-guruinaren berriztapena, epitelioa osatzen duten zelulen zatiketa
autologoa dela medio burutzen da. Muskuiluetan, berriztapen horrek itsasaldiekin eta
fotoperiodoarekin erlazionatutako zikloak jarraitzen ditu. Dena den, itsasaldi-
erregimenaren eta urteko sasoiaren moduko ingurumen-faktoreek zikloen gainean eragina
dute. Ingurumenean dauden kutsatzaileek ere, liseri-epitelioa osatzen duten zelulen
proliferazio-jarduera areago dezakete, muskuilu, magurio zein bareetan, liseri-epitelioaren
osaketa-zelularrean aldaketak sorreraziz, liseri-zelulen kopuru erlatiboaren jaitsiera barne.
Aldaketa horiek, laborategiko egoeretan itzulgarriak diren arren, kronikoki metalen pean
egondako bareetan itzulgarritasun hori motelduta ageri da. Liseri-guruineko epitelioan
gerta daitezkeen zelula-moten osaketaren aldaketak, ingurumen-toxikologian erabilitako
biojarraipen-programetako esposizio- eta efektu-biomarkatzaileak aplikatzerakoan
kontuan hartu beharreko fenomenoa da, zenbait kasutan biomarkatzaile desberdinen
gainean eragina dutelako.
221
Ondorioak eta tesia

METODOLOGI-ERANSKINA

GERTAKUNTZA HISTOLOGIKOA
1.- Disekzionatu berri diren laginak Carnoy fixatzailean fixatu ordu batez.
Carnoy Fixatzailea (100 ml)Etanol absolutua 60 mlKloroformoa 30 mlAzido azetiko glaziala 10 ml
2.- Laginak deshidratatu eta parafinan inkluditu.
%80 etanola 45 min%100 etanola 2 x 45 minMetilo bentzoatoa Gau osoaBentzenoa 2 x 45 minParafina 4 ordu 60ºC-tan
HEMATOXILINA-EOSINA TINDAKETA
7 µm-ko lodierako ebakiak hurrengo protokoloa jarraituz tindatu.
Xilola 2 x 10 min%100 etanola 2 x 2 min%96 etanola 2 min%70 etanola 2 minH2O distilatua 5 minHarris Hematoxilina 4 minH2O distilatua 2 minH2O distilatua 10 seg
H2O distilatua 10 segAlkohol azidoa 10 segH2O distilatua 5 min
Litio karbonatoa 10 segH2O distilatua min 1%70 etanola 3 min%80 etanola min 1Eosina min 1%96 etanola 2 x 2 min%100 etanola 2 x 2 minXilola 2 x 2 minEbakiak DPX (Fluka 44581) muntai-medioan montatu.
225
Metodologi-eranskina

Harris HematoxilinaHematoxilina 5 g%100 etanola 37 mlPotasio aluminio oxidoa 75 gOxido merkurikoa 2.5 gH2O distilatua 500 ml
Osagaiak nahastu. Nahasketa, irakite punturaino berotu eta giro tenperaturaraino hoztendenean 5 ml azido azetiko glaziala gehitu.
EosinaDisoluzio amaEosina (horia) 10 gH2O distilatua 200 ml
Disoluzioa pixkanaka berotu, eta ondoren hoztu denean
%96 etanola 800 ml
Lan disoluzioaDisoluzio ama 100 ml%80 etanola 300 mlAzido azetiko glaziala 2 ml
Alkohol azidoaAzido klorhidrikoa 1N 5 ml%70 etanola 100 ml
Litio karbonatoaLitio karbonatozko disoluzio saturatua H2O distilatuan.
226
Metodologi-eranskina

MASSON-GOLDNER TRIKROMIKOA TINDAKETA
7 µm-ko lodierako ebakiak hurrengo protokoloa jarraituz tindatu.
Xilola 2 x 10 min%100 etanola 2 x 2 min%96 etanola 2 min%70 etanola 2 minH2O distilatua 5 min
Groat Hematoxilina 4 minDabilura 5 minPonceau fuzsina azidoa 5 min%1 azido azetiko H2O disoluzioa 10 seg
Laranja G molibdikoa 5 min%1 azido azetiko H2O disoluzioa 10 segBerde argia 5 min%1 azido azetiko H2O disoluzioa 10 seg%70 etanola 2 min%96 etanola 2 x 2 min%100 etanola 2 x 2 minXilola 2 x 2 minEbakiak DPX muntai-medioan montatu.
Groat Hematoxilina (100 ml)Hematoxilina 0.5 g%96 etanola 50 mlAzido sulfurikoa 0.8 mlAlunbre ferrikoa 1 gH2O distilatua 50 ml
Ponceau fuszina azidoa (300 ml)Fuszina basikoa 1 gPonceau 0.2 gAzido azetikoa 0.6 mlH2O distilatua 300 ml
Laranja G molibdikoa (100 ml)Laranja G 2 gAzido fosfomolibdikoa 4 gH2O distilatua 100 ml
Berde argia (100 ml)Berde argia 1 gAzido azetikoa 2 mlH2O distilatua 100 ml
227
Metodologi-eranskina

AUTOMETALOGRAFIA
A) PARAFINAZKO EBAKIAK
7 µm-ko lodierako ebakiak hurrengo protokoloa jarraituz tindatu.
Xilola 2 x 10 min%100 etanola 2 x 2 min%96 etanola 2 min%70 etanola 2 minH2O distilatua 5 minLehorketa Gau osoa 37ºC-tanEmultsioa 30 minErrebelatzailea 15 minGelsitze-soluzioa 1 minFixatzailea 10 minEbakiak Kaiser gelatina glizerinatua (Merck 1.09242.0100) muntai-medioan
montatu.
Errebelatzailea (100 ml)Errebelatzailea (Tetenal ultrafilm B/W developer) 20 mlH2O distilatua 80 ml
Gelditze-soluzioa (100 ml)Azido azetiko glaziala 1 mlH2O distilatua 99 ml
Fixatzailea (100 ml)Fixatzailea (Agefix B&W fixer) 10 mlH2O distilatua 90 ml
B) ERRETXINAZKO EBAKIAK
B-1) EBAKI SEMIFINAK
3-4 µm-tako lodierako ebakiak hurrengo protokoloa jarraituz tindatu ziren.
Emultsioa 30 minErrebelatzailea 30-45 minGelditze-soluzioa 1 minFixatzailea 10 minEbakiak Kaiser gelatina glizerinatua muntai-medioan montatu.
Errebelatzailea (100 ml)Errebelatzailea (KODAK D-19) 15 gH2O distilatua 100 ml
228
Metodologi-eranskina

Gelditze-soluzioa (100 ml)Azido azetiko glaziala 1 mlH2O distilatua 99 ml
Fixatzailea (100 ml)Fixatzailea (Agefix B&W fixer) 10 mlH2O distilatua 90 ml
B-2) EBAKI ULTRAFINAK
3-4 µm-ko lodierako ebakiak aurretik azaldu bezala tindatu. Ondoren, tindatutako laginakberriro ere EPON erretxinan inkluditu, erretxina polimerizatu, ebaki ultrafinak eskuratu eta azetatouraniloz tindatu.
IMMUNOHISTOKIMIKA
A) PCNA IMMUNOHISTOKIMIKA (Marigómez et al., 1999)
1.- Disekzionatutako laginak aurretik azaldu bezala prestatu.
2.- Ebakiak silanizatutako portaobjektuetan itsatsi eta 60ºC-tan lehortu. Ondoren ebakiakxiloletan eta etanolezko bainutan desparafinatu eta hidratatu.
Porten silanizazioaPortak ur detergentea duen ur distilatuan garbitu 30 min-ordu 1.Uretan garbitu.Portak 130ºC-tan lehortu.Portak %2 3-aminopropiltrietoxisilano (Sigma A-3648) azetonatan 5 segunduz murgildu. Azetonan bi bainu azkarretan garbitu. Ur bidistilatuan garbitu.37ºC-tan gau osoan lehortu.
Desparafinazioa eta hidratazioaXilola 2 x 10 min%100 etanola 2 x 2 min%96 etanola 2 min%70 etanola 2 minH2O distilatua 5 min
3.- Beharrezkoa bada laginetan antigenizitatearen berreskuratzea egin daiteke.Horretarako ebakiak %0.5 tripsina duen TBS-tan 5 minutuz murgildu ziren.
TBS, pH 7.6 (100 ml)Tris 0.605 gNaCl 0.877 gH2O distilatua 100 ml
229
Metodologi-eranskina

4.- Ebakiak PBS-an garbitu eta peroxidasa jarduera endogenoa %3 H2O2 duen
metanola 10 minutuz erabiliz blokeatu.
PBSNaCl 16 gKCl 0.4 gNaH2PO4 H2O 0.46 gNa2HPO4 12H2O 6.8 g
H2O distilatua 2 l
5.- Ebakiak PBS-tan garbitu (3 x 10 min) eta %0.1 BSA duen PBS-tan ordu batez inkubatu.
6.- Ondoren, ebakiak PCNA antigorputzarekin (PC10 klona) (Sigma P-8825) inkubatu(1:1500 diluzioa), %0.1 BSA duen PBS-tan, 4ºC-tan eta gau osoan.
7.- Ebakiak PBS-tan garbitu (3 x 10 min) eta antigorputza abidina-biotina entzima konplexuaren bitartez ikustarazi Vectastain Elite ABC Kit-a (Burlingame) erabiliz. Horrela, ebakiakbiotinilatutako arratoiaren aurkako zaldiaren antigorputz sekundarioan inkubatu (1:200 diluzioa) 30minutuz giro tenperaturan. Ondoren, ebakiak PBS-tan garbitu eta gero (3 x 10 min) ABC erreaktiboan inkubatu 25 minutuz eta giro-tenperaturan.
8.- PBS-tan garbiketak egin ondoren, ebakien peroxidasa jarduera diaminobenzidina(DAB) disoluzio kromogenoa erabiliz ikustarazi.
DAB disoluzio kromogenoaDAB 0.06 gPBS 75 mlH2O2 25 µl
9.- Ebakiak Hematoxilinaren bainu azkar baten bitartez kontrastatzen dira eta ur distila-tuan garbitu onodoren Kaiser gelatina glizerinatua muntai-medioan montatu.
B) BrdU IMMUNOHISTOKIMIKA
B-1) Peroxidasa bidezko detekzioa.
1.- Disekzionatutako laginak aurretik azaldu bezala prestatu.
2.- Ebakiak silanizatutako portaobjektuetan itsatsi eta 60ºC-tan lehortu. Ondoren ebakiakxiloletan eta etanolezko bainutan desparafinatu eta hidratatu.
3.- Beharrezkoa bada laginetan antigenizitatearen berreskuratzea egin daiteke.Horretarako ebakiak %0,5 tripsina duen TBS-tan 5 minutuz murgildu.
230
Metodologi-eranskina

TBS, pH 7.6 (100 ml)Tris 0.605 gNaCl 0.877 gH2O distilatua 100 ml
4.- Ebakiak PBS-an garbitu eta peroxidasa aktibitate endogenoa %3 H2O2 duen
metanola 10 minutuz erabiliz blokeatzen da.
5.- Ebakiak 11.36 N HCl-an ordu batez inkubatu 37ºC-tan DNA harizpien bereizketa lortzeko. Ondoren sodio tetraborato indargetzailean inkubatu (2 x 5 min).
Sodio tetraboratoa 0.1 M, pH 8.5 (100ml)B4O7Na2 3.814 g
H2O distilatua 100 ml
6.- Ebakiak PBS-an garbitu (3 x 10 min) eta %0.1 BSA duen PBS-an ordu batez inkubatu.
7.- Ondoren, ebakiak BrdU antigorputzarekin (Roche) (1:100 diluzioa) inkubatu %0.1 BSAduen PBS-an, ordu batez eta giro tenperaturan.
8.- Ebakiak PBS-an garbitu (3 x 10 min) eta antigorputza abidina-biotina entzima konplexuaren bitartez ikustarazi Vectastain Elite ABC Kit-a (Burlingame) erabiliz. Horrela, ebakiakbiotinilatutako arratoiaren aurkako zaldiaren antigorputz sekundarioan inkubatu (1:200 diluzioa) 30minutuz giro tenperaturan. Ondoren, ebakiak PBS-an garbitu ostean (3 x 10 min), ABC erreaktiboan 25 minutuz inkubatu giro-tenperaturan.
9.- PBS-an garbiketak egin ondoren, ebakien peroxidasa jarduera 3 amino 9 etilkarbazol(AEC) (Sigma A-6926) disoluzio kromogenoa erabiliz ikustarazi.
AEC kromogenoaren disoluzioaAEC disoluzioa 200 µlSodio azetatoa 0.05 M 4 mlH2O2 2 µl
AEC DisoluzioaAEC 20 mgDimetilformamida 2.5 ml
Sodio azetatoa 0.05 M, pH 5.2 (100 ml)Sodio azetatoa 0.6804 gH2O distilatua 100 ml
10.- Ebakiak, Hematoxilinaren bainu azkar baten bitartez kontrastatu eta ur distilatuangarbitu eta ondoren Kaiser gelatina glizerinatua muntai-medioan montatu.
231
Metodologi-eranskina

B-2) Fosfatasa alkalino bidezko detekzioa.
1.- Disekzionatutako laginak aurretik azaldu bezala prestatu.
2.- Ebakiak silanizatutako portaobjektuetan itsatsi eta 60ºC-tan lehortu. Ondoren, ebakiakxiloletan eta etanolezko bainutan desparafinatu eta hidratatu.
3.- Beharrezkoa bada laginetan antigenizitatearen berreskuraketa burutu daiteke.Horretarako ebakiak %0.5 tripsina duen TBS-an 5 minutuz murgildu.
TBS, pH 7.6 (100 ml)Tris 0.605 gNaCl 0.877 gH2O distilatua 100 ml
4.- Ebakiak PBS-an garbitu.
5.- Ebakiak 11.36 N HCl-an ordu batez inkubatu 37ºC-tan, DNA harizpien bereizketa lort-zeko. Ondoren, sodio tetraborato indargetzailean inkubatu (2 x 5 min).
Sodio tetraboratoa 0.1M, pH 8.5 (100ml)B4O7Na2 3.814 gH2O distilatua 100 ml
6.- Ebakiak PBS-an garbitu (3 x 10 min) eta %5 BSA duen PBS-an 20 minutuz inkubatu.
7.- Ondoren, ebakiak BrdU antigorputzarekin (Roche) inkubatu (1:100 diluzioa) %1 BSAduen PBS-an, ordu batez eta giro-tenperaturan.
8.- Ebakiak PBS-an garbitu (5 minutuz etangabe irabiatzen) eta antigorputza abidina-biotina-entzima konplexuaren bitartez ikustarazi, Sigma Mouse Extravidin Alkaline PhosphataseStaining Kit-a (Sigma EXTRA 2A) erabiliz. Horrela, ebakiak biotinilatutako arratoiaren aurkakoahuntzaren antigorputz sekundarioarekin inkubatu (1:20 diluzioa) 20 minutuz giro-tenperaturan.Ondoren, ebakiak PBS-an garbitu ostean (5 minutuz etengabe irabiatzen) Fosfatasa alkalinoanitsatsita daraman extrabidinarekin inkubatu (1:20 diluzioa) 20 minutuz eta giro-tenperaturan.
9.- Laginak TBS-an garbitu 5 minutuz eta fosfatasa alkalino jarduera Fast Red TR/Naphtoltabletak sustrato gisa erabiliz ikustarazi (Sigma F-4523).
10.- Ebakiak Mayer Hematoxilinaren bainu azkar baten bitartez kontrastatu eta ur distilatuan garbitu ostean Kaiser gelatina glizerinatua muntai-medioan montatu.
232
Metodologi-eranskina

Mayer Hematoxilina (1l)Hematoxilina 1 gAlunbre potasikoa 50 gSodio Iodatoa 0.2 gAzido Zitrikoa 1 gKloral hidratoa 30 gH2O distilatua 1 l
ß GLUKURONIDASA HISTOKIMIKA (Cajaraville et al., 1991)
1.- Disekzionatu berri diren laginak % 2.5 NaCl eta %10 sakarosa duen fosfato indarget-zailean (0.1 M) 15 minutuz murgildu.
Fosfato indargetzailea 0.1 M, pH 7.4 (100ml)NaH2PO4 H2O 0.256 gNa2HPO4 12H2O 2.892 g
H2O distilatua 100 ml
2.- Laginak nitrogeno likidotan izoztu.
3.- Kriostatoan eskuratutako 8 µm-ko lodierako ebakiak izozkailutik hozkailura eraman etabertatik giro tenperaturara igaro bertan hainbat minutu utziz.
4.- Ebakiak inkubazio-medioan inkubatu 37ºC-tan, eta 40 minutuz muskuilu, 20 minutuzmagurio eta 10 minutuz bareen kasuan, hain zuzen.
Inkubazio-medioaAzetato indargetzailea 285 mlNaftola (Sigma N-1875)) 0.081 gSodio bikarbonatoa 3.468 mlPolibinilo alkohola 30.60 g
Azetato indargetzailea 0.1 M, pH 4.5 (600 ml)NaCl 15 g Sodio azetatoa 7.344 gAzido azetikoa 3.78 mlH2O distilatua 600 ml
Sodio bikarbonatoa (100 ml)NaHCO3 0.42 gH2O distilatua 100 ml
5.- Ebakiak azetato indargetzailean 2 minutuz garbitu 37ºC-tan.
6.- Ebakiak %0.1 Fast Garner GBC (Sigma F-6504) eta %2.5 NaCl duen fosfato
233
Metodologi-eranskina

indargetzailean 10 minutuz sartu.
7.- Ebakiak, Baker formol kaltzikoan fixatu 10 minutuz. Formol kaltzikoak hotza egonbehar du.
Baker formol kaltzikoa (100 ml)Formaldehidoa (%40) 10 ml (pH = 7)CaCl2 3 gNaCl 7.5 gH2O distilatua 100 ml
8.- Ebakiak, ur distilatuko hainbat bainu azkarretan garbitu ondoren %1 Fast Green FCF(Sigma F-7252) duen ur distilatuan 2 minutuz kontrastatu giro-tenperaturan.
9.- Ebakiak, Kaiser gelatina glizerinatua muntai-medioan montatu.
X IZPIEN MIKROANALISIA ETA MAPAKETA
Erretxinan inkluditutako ebakiak, formvar/karbonoz estalitako aluminio eta titaniozkosaretxotan bildu. Ondoren, berriro ere, eta kontrastatu gabe, karbonoz estali ziren X izpien mikroanalisia egiteko.
X izpien mikronanalisia, energi dispertsiorako espektrometroa lotuta zeukan JEOL 1210STEM mikroskopioan burutu zen (kontaketa-denbora: 100 segundu). X izpien mapaketa Ag (Lα1-2.984 keV-, Lß1 -3.151 keV-), Cd (Lß1 -3.316 keV-) eta S (Kα -2.308 keV-) seinaleak batera lokalizatzeko burutu zen. Horretarako Link ISIS v1.04A softwarea erabili zen. Analisi hauek Cardiff-eko Galeseko Unibertsitatean egin ziren.
ABSORTZIO ATOMIKOZKO ESPEKTROFOTOMETRIA
1.- Disekzionatu berri ziren laginak, ur distilatuarekin garbitu eta 120ºC-tan 48 orduz lehortu.
2.- Dagoeneko guztiz lehorturik zeuden laginak pisatu.
3.- Beharrezkoa izatekotan, beirazko hodi baten laguntzaz laginak txikitu.
4.- Laginak azido nitriko kontzentratuan liseritu. Horretarako Erlenmeyer hodietan azidonitrikoa apurka isuri lagina guztiz disolbatu arte eta Erlenmeyer hodiak puxtarri batez estali liseriketa egokiagoa lortzeko.
5.- Laginen liseriketa amaituta dagoela egiaztatu ondoren, azido nitrikoa 80ºC-tan lurrundu.
6.- Lehor zeuden laginak azido nitriko diluituan (0.65 eta 0.1 M) berresekitu eta 4000 rpm-ra 4 minutuz zentrifugatu, gainjalkina eskuratu, eta neurtu bitartean hozkailuan gorde.
234
Metodologi-eranskina

AZIL KoA-OXIDASA (AOX) JARDUERAREN NEURKETA (Small et al., 1985).
1.- Bareen liseri-guruinak TVBE indargetzailean homogenizatu eta 500 g-tan zentrifugatu,15 minutuz eta 4ºC-tan.
TVBE indargetzailea pH 7.6 (100 ml)NaHCO3 8.4 mg0.1M EDTA 1 ml%0.1 etanol 0.1 ml%10 Triton X-100 0.1 mlH2O distilatua 100 ml
2.- Gainjalkinaren 100 µl (beharrezko diluzioan) hartu eta 1.9 ml inkubazio-mediora gehi-tu. Nahasketa 5 minutuz eta 25ºC-tan iluntasunean mantendu. Substratuaren (Palmitoil-KoA(Sigma P-9276) 3 mM) 10 µl gehitu, nahastu eta absorbantziaren aldaketa neurtu 502 nm-tan 3minutuz.
Inkubazio-medioa0.5M potasio fosfato indargetzailea 2 mlDCF disoluzioa* 2 mlErrefrau peroxidasa 1200 mU/ml (Sigma P-8250) 1 ml4 M NaN3 (Sigma S-2002) 1 ml%10 Triton X-100 (Sigma X-100) 2 mlH2O distilatua 100 ml arte
*1 ml egiteko: 1.3 mg diklorofluoreszeina (DCF) (Molecular Probes D-399) 100 µl dimetilformamida + 900 µl 0.01 N NaOH-an disolbatu.
Potasio fosfato indargetzailea 0.5 M pH 7.4KH2PO4 680 mg
H2O distilatua 10 mlK2HPO4 3H2O 1140 mg
H2O distilatua 10 ml
KH2PO4 disoluzioa K2HPO4 3H2O disluzioari gehitu pH 7.4 lortu arte.
Substratu disoluzioa (3 mM palmitoil-KoA oxidasa)Palmitoil-KoA (Sigma, P-9276) ur distilatuan disolbatu, beharrezko kontzentrazioa
lortu arte. 0.5 ml-tako alikuotetan izoztu eta gorde.
3.- AOX jardueraren neurketa honela burutu:mUnitate/ml = (Absorbantziaren aldaketa/minutu) x (erreakzio-bolumena/lagin-bolumena)
x (mg proteina/ml).
mUnitate/mg proteina = (mUnitate/ml) / ( mg proteina/ml)
235
Metodologi-eranskina

236
Metodologi-eranskina
Top Related