
Abundancia de obreras forrajeras en colonias de...
41
Universidad de Buenos Aires Facultad de Ciencias Exactas y Naturales Abundancia de obreras forrajeras en colonias de hormigas granívoras del género Pogonomyrmex en el Monte central Tesis de Licenciatura Autora: Beatriz Elisa Nobúa Behrmann Marzo 2008 Director: Javier Lopez de Casenave
Transcript of Abundancia de obreras forrajeras en colonias de...

Universidad de Buenos Aires Facultad de Ciencias Exactas y Naturales
Abundancia de obreras forrajeras en colonias de
hormigas granívoras del género Pogonomyrmex
en el Monte central
Tesis de Licenciatura
Autora: Beatriz Elisa Nobúa Behrmann
Marzo 2008
Director: Javier Lopez de Casenave

Índice
esumen . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 2
Resu
Discu
. . 31
Conc
R
Introducción . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 4
Métodos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Área de estudio. . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 10
Estimación del número de forrajeras . . . . . . . . . . . . . . . . . . . . . . . . . . . . 11
Técnicas de captura–marcado–recaptura . . . . . . . . . . . . . . . . . . . . . . . . . 12
Evaluación de los supuestos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 14
Determinación del tamaño de colonia por excavación. . . . . . . . . . . . . . . . . 16
Relación entre la abundancia de forrajeras y los niveles de actividad . . . . . . 17
ltados . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
Evaluación de los supuestos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 18
Abundancia de obreras forrajeras . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 20
Excavaciones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 21
Niveles de actividad . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 25
sión . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 27
Aplicabilidad de las técnicas de captura–marcado–recaptura . . . . . . . . . . . 27
Características de las colonias de las especies sudamericanas del género
Pogonomyrmex . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
Abundancia de forrajeras y actividad externa de la colonia . . . . . . . . . . . . .
33
lusiones . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 35
Bibliografía . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 36
Agradecimientos . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 40
1

Resumen En el Monte Central, las hormigas son consideradas los organismos granívoros más
importantes en primavera-verano, encontrándose entre ellas las especies del género
Pogonomyrmex. El efecto de estas hormigas sobre las semillas depende, entre otros
factores, de los niveles de actividad y de la estrategia de forrajeo (individual o grupal)
de la colonia. A su vez, ambos factores pueden depender de la cantidad de obreras
presentes en la colonia o de la abundancia de forrajeras en la misma. Una de las técnicas
que permiten estimar la abundancia de obreras forrajeras sin causar daños en el nido es
la de de captura–marcado–recaptura, cuya aplicabilidad depende de que se cumplan los
supuestos de la misma. El objetivo general de este trabajo fue profundizar en el
conocimiento de las características de las colonias de las hormigas granívoras del
género Pogonomyrmex y su influencia sobre el forrajeo de semillas en la porción central
del desierto del Monte. En particular, los objetivos específicos fueron: (1) evaluar la
aplicabilidad de las técnicas de captura–marcado–recaptura para estimar la abundancia
de obreras forrajeras en colonias de Pogonomyrmex pronotalis, P. rastratus y P.
inermis, (2) estimar el número de obreras forrajeras presentes en sus colonias, y (3)
evaluar la relación entre la abundancia de obreras forrajeras en las colonias de las tres
especies y sus niveles de actividad. Mediante experiencias de laboratorio se pudo
comprobar que el tipo de marca utilizada (pintura al óleo) no afecta a la supervivencia
ni el comportamiento de las obreras (dos de los supuestos necesarios para aplicar
correctamente la técnica de captura–marcado–recaptura). Se estimó la abundancia de
forrajeras usando dos métodos distintos de captura–marcado–recaptura: el Método de
Petersen (que requiere solo dos ocasiones de muestreo) y el de Schnabel (con múltiples
ocasiones de muestreo). Ambos arrojaron valores de abundancia diferentes, siendo
siempre mayores las obtenidas mediante el Método de Schnabel. El análisis de los
resultados sugiere fuertemente que ocurren eventos de mortalidad durante el tiempo
transcurrido entre el primer y el último muestreo. El Método de Schnabel es más
sensible al incumplimiento del supuesto de que la población es cerrada (i.e., sin
reclutamientos ni muertes), dado que, por las características biológicas y
comportamentales de las forrajeras de estas especies, necesariamente implica un tiempo
de muestreo más largo. Si bien ambos métodos pueden servir para estimar la
abundancia relativa de forrajeras en las colonias de P. pronotalis, P. inermis y P.
2

rastratus, el Método de Petersen es más recomendable en términos de eficiencia de
tiempo y esfuerzo de muestreo. El Método de Schnabel, aunque sobreestima la
abundancia absoluta de forrajeras, brinda información acerca del cumplimiento o no de
los supuestos de la técnica. A partir de las estimaciones mediante el Método de Petersen
y de excavaciones se encontró que las colonias de P. pronotalis, P. inermis y P.
rastratus son relativamente pequeñas, con aproximadamente unas 300-1000 obreras, de
las cuales el 7-15% corresponde a forrajeras. El análisis de la actividad forrajera mostró
que ésta se correlaciona positivamente con la abundancia de forrajeras en las colonias
de P. rastratus. La misma tendencia fue observada en P. inermis y P. pronotalis,
aunque en este caso no se pudo encontrar una correlación significativa entre ambas
variables.
3

Introducción
Las semillas constituyen una parte importante de los ecosistemas desérticos ya que
representan el principal medio de dispersión de muchas plantas y les permiten persistir
durante largos períodos de sequía (Kemp 1989). Al mismo tiempo, son una fuente de
alimento diverso y abundante para muchos de los animales que habitan estos sistemas,
un recurso que está disponible en el suelo y que incluso puede ser colectado y
almacenado para su posterior consumo (Brown et al. 1979). Los principales
consumidores de semillas en los desiertos son las aves, las hormigas y los roedores
(Brown et al. 1979). En particular, las hormigas granívoras influyen de manera
significativa en la estructura de las comunidades y el funcionamiento de estos
ecosistemas debido a la densidad y longevidad de sus colonias, la cantidad de semillas
que cosechan y la magnitud del movimiento de suelo que realizan durante la
construcción y el mantenimiento de sus nidos (Hölldobler & Wilson 1990, MacMahon
et al. 2000).
En las colonias de hormigas, las obreras tienden a especializarse en tareas específicas
(e.g., forrajeo, cuidado de las larvas, mantenimiento de la estructura del nido). En las
especies polimórficas, la división de tareas está dada por el tamaño de las obreras,
quienes mantienen una misma ocupación a largo de toda su vida adulta (Hölldobler &
Wilson 1990). Por el contrario, en la mayoría de las especies monomórficas, las obreras
van cambiando de tarea a medida que envejecen: las más jóvenes se ocupan de las
labores que se realizan dentro del nido, como el cuidado de las larvas, y las más viejas
de las tareas en el exterior, como forrajear (Gordon 1989, Hölldobler & Wilson 1990).
El número de obreras que se dedican a forrajear es relativamente constante dentro de
una colonia (Tripp et al. 2002) y suele estar relacionado con el número total de obreras
en la misma (e.g., Kruk-de Bruin et al. 1977, Porter & Jorgensen 1980).
El forrajeo y el consumo de semillas por parte de las hormigas granívoras pueden tener
importantes efectos sobre la vegetación. Si bien la mayoría de los estudios realizados
estiman que la fracción del total de semillas producidas por año que son consumidas por
estas hormigas es menor al 10% (Pulliam & Brand 1975, Whitford 1978, Crist &
MacMahon 1992), pueden tener un impacto mucho mayor sobre las especies que
4

prefieren, de las cuales llegan a remover hasta el 100% de las semillas disponibles (Crist
& MacMahon 1992). El consumo selectivo de semillas puede producir cambios
cualitativos en la estructura de la comunidad de plantas al alterar la abundancia relativa
de las especies (Pulliam & Brand 1975, Whitford 1978, Crist & MacMahon 1992,
MacMahon et al. 2000). Este efecto puede ser más marcado en las cercanías de los
nidos en donde suelen generar cambios en la abundancia y composición del banco de
semillas (Hölldobler & Wilson 1990, MacMahon et al. 2000, Pirk 2007) y modificar las
condiciones del suelo como resultado del movimiento de tierra y de la acumulación de
desechos (MacMahon et al. 2000, Wagner et al. 2004).
El efecto de las hormigas sobre las semillas depende, entre otros factores, de los niveles
de actividad, de las características del área de forrajeo y de la estrategia de forrajeo
(individual o grupal) de la colonia (Crist & MacMahon 1992, MacMahon et al. 2000).
A su vez, tanto los niveles de actividad como la estrategia de forrajeo pueden depender
de la cantidad de obreras presentes en la colonia o de la abundancia de forrajeras en la
misma. Los patrones diarios y estacionales de actividad están relacionados
principalmente con la temperatura del suelo (Hölldobler y Wilson 1990, Crist &
MacMahon 1991). Sin embargo, los niveles de actividad también pueden verse
afectados por otros factores como la disponibilidad de alimento (Crist & MacMahon
1991, Schafer et al. 2006) o la abundancia de forrajeras en las colonias. En algunas
especies se encontró que la tasa de forrajeo (nivel de actividad forrajera) aumenta con el
tamaño de colonia (Mailleux et al. 2003), aunque es esperable que la actividad de
forrajeo del nido esté más estrechamente relacionada con el número de hormigas
forrajeando que con el tamaño total de la colonia. En cuanto a la estrategia de forrajeo,
el patrón general observado es que las forrajeras de las especies que tienen colonias
pequeñas buscan su alimento de manera solitaria, mientras que las de especies con
colonias grandes forrajean en grandes grupos, usando señales químicas para reclutar a
sus compañeras (Beckers et al. 1989). En las especies estudiadas del género
Pogonomyrmex en América del Norte, por ejemplo, la estrategia de forrajeo varía en
función del tamaño de la colonia, con un promedio de 100–1500 obreras por colonia en
las especies con forrajeo individual y de >5000–10000 obreras en aquellas que
presentan forrajeo grupal (Johnson 2000).
5

La mitad de las especies de hormigas granívoras del mundo pertenecen al género
Pogonomyrmex. Este género, exclusivamente americano, incluye unas 60 especies con
dos grandes áreas de distribución independientes, una en la parte occidental de América
del Sur y otra en el sudoeste de los Estados Unidos y regiones colindantes de México
(Kusnezov 1951, Taber 1998). Las especies de América del Sur se caracterizan por
tener colonias mucho más pequeñas y poblaciones menos densas que las de América del
Norte (Hölldobler & Wilson 1990). Según Kusnezov (1951), todas ellas presentan
forrajeo individual, sin la formación de filas ni caminos radiales. En ese sentido, se las
considera menos evolucionadas que las especies congenéricas norteamericanas (de
colonias grandes y con forrajeo grupal), y aún menos en comparación con el género
norteamericano Veromessor y el género paleártico-etiópico Messor, los cuales son
considerados como los representantes más evolucionados de la forma de vida de
hormiga granívora (Kusnezov 1951). La mayoría de los estudios ecológicos y
comportamentales de las especies del género Pogonomyrmex, realizados en América del
Norte, se ocupan de las especies que poseen colonias grandes y conspicuas. Por el
contrario, poco se sabe de las que tienen colonias pequeñas, a pesar de que es
información necesaria para realizar estudios comparativos de la ecología y la historia
evolutiva de las especies de hormigas granívoras (Johnson 2000).
En el Monte Central, las hormigas son los organismos granívoros más importantes
durante la primavera y el verano, incluyendo unas 26 especies que remueven semillas
(Lopez de Casenave et al. 1998 datos no publicados, Marone et al. 2000). Tres de estas
especies, pertenecientes al género de granívoras especialistas Pogonomyrmex, han sido
estudiadas recientemente: P. pronotalis, P. inermis y P. rastratus (Figura 1). Por
ejemplo, se conoce su dieta y sus preferencias por distintas semillas (Pirk et al. 2004,
2007, Pirk & Lopez de Casenave 2006, Pirk 2007), la distribución espacial de sus
colonias (Pirk et al. 2004, Milesi et al., datos no publicados) y su sistema de
apareamiento (Pol et al. 2008). También se han estudiado sus patrones temporales y
espaciales de forrajeo, encontrándose variaciones estacionales similares a las descriptas
para otras especies congenéricas de América del Norte (Pol & Lopez de Casenave 2004,
Pol et al., datos no publicados), aunque con niveles de actividad más bajos (Crist &
MacMahon 1991). En cambio, aún se desconoce el tamaño de las colonias de estas
especies. Las únicas referencias disponibles son observaciones anecdóticas de la historia
6

natural de algunas especies del género (e.g., Kusnezov 1951) que sugieren que las
colonias tendrían bajas densidades de obreras (<500 por colonia), lo que a su vez
explicaría que sus niveles de actividad sean menores que los de las especies de colonias
grandes estudiadas en América del Norte. Es necesario estimar adecuadamente el
tamaño de colonia de estas especies para poder evaluar si existe una asociación entre ese
tamaño, los niveles de actividad y los patrones de forrajeo.
Figura 1. Obreras de las tres especies de hormigas granívoras del género Pogonomyrmex que habitan en
el Monte Central: P. pronotalis (izquierda), P. inermis (centro) y P. rastratus (derecha).
Se han utilizado diferentes técnicas para estimar el número de obreras en colonias de
distintas especies de hormiga. La más comúnmente usada consiste en excavar los nidos,
extraer a todos los individuos y contarlos (e.g., Gaglio et al 1998, Tschinkel 1998, 1999,
Diehl-Fleig & Diehl 2007, Johnson et al. 2007). La excavación permite registrar no solo
el número de obreras, sino también el de larvas y, algunas veces, hasta el número de
huevos presentes en una colonia. Sin embargo, es inapropiado para algunos estudios. En
primer lugar, necesariamente implica la destrucción de la colonia, impidiendo futuros
estudios y el seguimiento en el tiempo de la misma. En segundo lugar, para muchas
especies es difícil asegurar que todas las obreras y larvas han sido encontradas durante
la excavación (Stradling 1970, Billick 1999), por lo que resulta más un muestreo que el
pretendido censo. En tercer lugar, las excavaciones solo permiten medir el tamaño del
nido, cuando por lo general lo que se quiere conocer es el tamaño de la colonia (Billick
1999). Una colonia se define como la unidad dentro de la cual los individuos cooperan
para reproducirse, mientras que un nido es el lugar en donde se mantiene a las larvas
7

(Billick 1999). Las colonias de algunas especies presentan múltiples nidos y, sin
estudios comportamentales o genéticos, es imposible determinar mediante la excavación
si las obreras de nidos cercanos pertenecen o no a la misma colonia (Billick 1999). Por
último, por ser un método destructivo la excavación no permite distinguir entre grupos
de hormigas con distintas funciones (especialmente en las especies monomórficas): solo
es posible estimar el número total de obreras en la colonia, sin saber cuántas de ellas
pertenecen a diferentes grupos (e. g. forrajeras, patrulladoras, de mantenimiento, de
cuidado de larvas).
Existen técnicas menos invasivas para estimar el tamaño de las colonias de hormigas,
como las de captura–marcado–recaptura (e.g., Kruk-de Bruin et al. 1977, Porter &
Jorgensen 1980, 1981, Billick 1999, 2001, Tripp et al. 2000). Estas técnicas fueron
desarrolladas originalmente para pesquerías y su primer aplicación en ecología fue
llevada a cabo por Petersen en 1894 para calcular la tasa de mortalidad en peces (Le
Cren 1965). Sin embargo, el primero en publicarlas como técnicas para estimar el
tamaño poblacional fue Lincoln, quien en 1930 lo aplicó a una población de patos
(Bailey 1951). Las técnicas de captura-marcado-recaptura consisten básicamente en
capturar individuos de la población de interés, marcarlos y liberarlos. Luego de permitir
que los animales marcados se mezclen con el resto de la población, se toma otra muestra
y se cuenta el número de individuos marcados y no marcados que fueron capturados. El
tamaño de la población puede ser estimado haciendo el supuesto de que la proporción
de individuos marcados en la segunda muestra es la misma que en la población total. A
partir de la formulación original se han desarrollado muchos otros modelos, más
complejos, basados en el mismo principio (Seber 1982, 1986). Estas técnicas no son
destructivas y permiten el seguimiento de las colonias en el tiempo. Además, siguiendo
el movimiento de los individuos marcados es posible identificar cuáles nidos pertenecen
a la misma colonia. El principal problema de esta técnica, aunque no el único, es que
solo pueden ser capturadas y marcadas las obreras que salen del nido. Esto significa que
solamente puede estimarse el número de obreras que realizan tareas fuera del nido, sin
obtenerse información de los grupos de obreras que permanecen por debajo de la
superficie, especializadas en tareas como el almacenamiento de comida y el cuidado de
las larvas (Kruk-de Bruin et al. 1977, Billick 1999). Sin embargo, esta característica del
muestreo puede ser ventajosa cuando se busca discriminar entre grupos funcionales de
8

hormigas, como ser conocer el número de individuos de una colonia que buscan
alimento fuera del nido. La aplicabilidad de estas técnicas depende principalmente de
que se cumplan los supuestos de las mismas en el sistema en estudio.
El objetivo general de este trabajo es profundizar el conocimiento de las características
de las colonias de las hormigas granívoras del género Pogonomyrmex y su influencia
sobre el forrajeo de semillas en la porción central del desierto del Monte. En particular,
los objetivos específicos son: (1) evaluar la aplicabilidad de los métodos de captura–
marcado–recaptura para estimar la abundancia de obreras forrajeras en colonias de
Pogonomyrmex pronotalis, P. rastratus y P. inermis; (2) estimar el número de obreras
forrajeras presentes en colonias de P. pronotalis, P. rastratus y P. inermis; y (3) evaluar
la relación entre la abundancia de obreras forrajeras en las colonias de P. pronotalis, P.
rastratus y P. inermis y sus niveles de actividad de forrajeo.
9

Métodos
Área de estudio
El trabajo se desarrolló en la Reserva de la Biosfera de Ñacuñán (34º03’S, 67º54’O), en
la provincia de Mendoza. La reserva tiene una superficie de 12800 ha y está atravesada
de norte a sur por la ruta provincial 153. El clima es árido-semiárido, estacional, con
veranos cálidos y relativamente húmedos e inviernos fríos y secos. La temperatura
promedio anual es de 15.6 °C, con máxima y mínima mensual promedio de 23.8 °C y
7.6 °C, respectivamente (Lopez de Casenave 2001). La oscilación diaria de la
temperatura es muy grande: su valor promedio es de 16.2 °C. El régimen de lluvias
presenta una marcada estacionalidad y gran variación interanual. El promedio de
precipitaciones anuales en Ñacuñán es de 343.6 mm (periodo 1972-2005). A pesar de
presentar una estación relativamente húmeda (el 78% de las lluvias caen entre octubre y
marzo), el balance hídrico es deficitario a lo largo de todo el año (i.e., la
evapotranspiración siempre es mayor que las precipitaciones; Roig 1971).
El ambiente predominante de la reserva es el bosque abierto de Prosopis flexuosa
(algarrobal), que alterna con jarillales de Larrea cuneifolia en zonas bajas y con
comunidades asociadas a médanos. En el algarrobal, los árboles (Prosopis flexuosa y
Geoffroea decorticans) están dispersos en una matriz de arbustos altos (Larrea
divaricata, Condalia microphylla, Capparis atamisquea y Atriplex lampa), arbustos
bajos (Lycium spp., Verbena aspera y Acantholippia seriphioides) y gramíneas
perennes (Pappophorum spp., Trichloris crinita, Digitaria californica, Aristida spp.,
Setaria leucopila, Sporobolus cryptandrus, Stipa ichu). Las dicotiledóneas herbáceas
(e.g., Chenopodium papulosum, Phacelia artemisioides, Sphaeralcea miniata,
Parthenium hysterophorus, Glandularia mendocina, Descurainia spp.) presentan una
cobertura muy variable entre estaciones y años, aunque usualmente menor a la de las
gramíneas (Lopez de Casenave 2001).
La reserva se encuentra atravesada por numerosos ca os (picadas), que tienen un trazado
lineal y que, en su mayoría, confluyen radialmente en la estación biológica de la reserva
(Pirk et al. 2004). Las colonias estudiadas de las tres especies del género
10

Pogonomyrmex de Ñacuñán estaban ubicadas a lo largo de estos caminos. Las picadas
son de tierra y tienen un ancho de aproximadamente 6 m incluyendo sus bordes. La
cobertura de plantas es mucho menor allí que en el algarrobal adyacente, encontrándose
la superficie del suelo más expuesta al sol, al viento y a la lluvia, y con un mayor grado
de compactación. Las especies de hormigas estudiadas habitan tanto en los caminos
como en el algarrobal, aunque son más abundantes sobre las picadas, y no presentan
diferencias importantes en su ecología trófica entre ambientes (Pirk et al. 2004).
Estimación del número de hormigas forrajeras
La estimación del número de obreras forrajeras se llevó a cabo durante febrero y marzo
de 2007, en el periodo del año de mayor actividad de Pogonomyrmex pronotalis, P.
inermis y P. rastratus (Pol & Lopez de Casenave 2004, Pirk 2007). Se seleccionaron
cinco nidos activos de cada una de las tres especies, ubicados a lo largo de los caminos
que atraviesan la reserva. El número de obreras forrajeras de cada colonia se estimó
mediante la técnica de captura–marcado–recaptura. Para ello se llevaron a cabo varios
eventos de captura y marcado en cada nido (las ocasiones de muestreo fueron siempre
en días diferentes, para evitar posibles disturbios en la colonia). Los muestreos se
hicieron durante los horarios de máxima actividad del día (Pol & Lopez de Casenave
2004) y evitando los días fríos, climáticamente desfavorables para la actividad forrajera.
En cada ocasión de muestreo se capturaron con un aspirador todos los individuos que
entraban o salían del nido durante al menos 30 minutos, colocándolos en un recipiente
plástico de boca ancha hasta terminar el evento de captura. La captura se daba por
terminada al cabo de una hora o cuando durante un lapso de 10 minutos no se
observaban hormigas entrando ni saliendo del nido. Para asegurarse de capturar solo a
las obreras involucradas en una excursión de forrajeo, se consideró que un individuo
regresaba de forrajear o abandonaba el nido para forrajear cuando estaba entrando o
saliendo, respectivamente (con o sin carga), de un círculo imaginario de 35 cm de
diámetro centrado en la entrada del nido (ver Pol & Lopez de Casenave 2004).
Una vez finalizado cada evento de captura, las hormigas fueron contadas, discriminando
entre las que habían sido marcadas previamente y las que no. El marcado de estas
últimas fue realizado en el campo. Para ello cada obrera fue asida con una pinza y
11

pintada cuidadosamente en el dorso del abdomen con un marcador de pintura al óleo
Uni Paint (Mitsubishi Pencil Co.). Esta misma pintura ya ha sido utilizada con este
objetivo en otras especies del género Pogonomyrmex (Gordon et al. 2005). Las
hormigas pintadas (Figura 2) fueron colocadas en otro recipiente y se esperaron unos
minutos hasta que secara la pintura. A continuación, todas las hormigas capturadas
fueron liberadas en las cercanías de la entrada del nido, para facilitar su retorno.
El proceso de captura y marcado se repitió para cada colonia hasta que el porcentaje de
obreras marcadas recapturadas era mayor o igual a 90% durante tres muestreos
consecutivos o hasta un máximo de 11 ocasiones.
Figura 2. Obreras de Pogonomyrmex pronotalis (izquierda), P. inermis (centro) y P. rastratus (derecha)
marcadas con pintura al óleo.
Técnicas de captura–marcado–recaptura
El número de obreras forrajeras en cada colonia se estimó usando el Método de Petersen
(considerando solo los dos primeros muestreos) y el Método de Schnabel (tomando en
cuenta todos los muestreos realizados).
El método de Petersen (también conocido como índice de Lincoln) es la más básica de
las técnicas de captura–marcado–recaptura e implica solo dos eventos de captura.
Consiste en tomar una muestra de n1 individuos de la población, marcarlos y liberarlos.
Luego se toma otra muestra de n2 individuos, en la cual se cuentan los m2 individuos
marcados. Con esos datos se utiliza el estimador de Petersen para el tamaño de la
población (N) según la fórmula N = n1 n2 / m2 . Como esta fórmula tiende a sobreestimar
el tamaño poblacional se utilizó la siguiente corrección según lo propuesto por Seber
12

(1982): N = [(n1 + 1) (n2 + 1) / (m2 + 1)] – 1. El método de Schnabel (1938) es una
extensión del de Petersen y consta de una serie de muestreos sucesivos. En cada muestra
de tamaño ni se registra el número de individuos marcados encontrados (mi), se marca a
aquellos que fueron capturados por primera vez y se los libera. Si Mi es el número de
individuos marcados presentes en la población antes de que la muestra i fuera tomada,
se puede estimar el tamaño de la población como N = Σ (ni Mi) / Σ mi . Los límites del
intervalo de confianza del 95% para cada estimador se calcularon según Seber (1982).
Para comparar a las tres especies estudiadas en cuanto a la abundancia de forrajeras en
sus colonias se realizó una prueba de Kruskal-Wallis (Siegel & Castellan 1988).
Es importante tener en cuenta que el tamaño de la población estimado con las técnicas
ara que estos métodos de captura–marcado–recaptura produzcan estimaciones
a mayor ventaja del muestreo múltiple para aplicar el método de Schnabel es que es
de captura–marcado–recaptura se refiere siempre a la población capturable, la cual
puede o no coincidir con la población entera (Krebs 1989). Para las especies de
hormigas que construyen su nido bajo rocas es fácil acceder a la mayoría de las cámaras
y, por lo tanto, es posible estimar el número total de obreras en la colonia (e.g., Brian et
al. 1965, Stradling 1970, Billick 1999). Sin embargo, en la mayoría de las especies solo
pueden ser capturadas las obreras que salen del nido. En estos casos, las técnicas de
captura–marcado–recaptura solo se aplican a la subpoblación de obreras forrajeras (la
población capturable de estas colonias).
P
confiables es necesario que se cumplan una serie de supuestos (Southwood 1978, Seber
1982): (1) la población es cerrada, por lo tanto N es constante; (2) todos los individuos
de la población tienen la misma probabilidad de ser capturados; (3) la marca no afecta el
comportamiento de los individuos; (4) la marca no afecta la expectativa de vida de los
individuos; (5) la marca no afecta la capturabilidad de los individuos (marcados y no
marcados tienen la misma probabilidad de ser capturados); (6) los individuos no pierden
sus marcas en el tiempo transcurrido entre dos muestreos.
L
posible detectar las violaciones a estos supuestos, en caso de haberlas. Un gráfico de
regresión de la proporción de animales recapturados en la muestra (mi / ni) en función
del número acumulado de animales previamente marcados (Mi) será lineal si los
13

supuestos se cumplen, pero se curvará cuando estos se violen (Figura 3).
Desafortunadamente, no existe una única interpretación para los gráficos curvilíneos: si
bien señalan que uno o más supuestos han sido violados, no indican cuál de ellos o
cómo ha sido violado (Krebs 1989).
0
0.5
1
0 30 60 90Marcados acumulados
Prop
orci
ón d
e re
capt
urad
os .
Figura ción del
valuación de los supuestos
captura–marcado–recaptura, se pusieron a prueba dos de
upervivencia.— Para evaluar si la marca afecta la supervivencia de las hormigas se
3. Ilustración esquemática de la proporción de animales recapturados (mi / ni) en fun
número acumulado de animales previamente marcados (Mi) a lo largo de un muestreo con múltiples
capturas. El tamaño de la población es igual al número de individuos marcados previamente cuando
la proporción de recapturas es igual a 1 (flecha). La relación entre ambos parámetros es lineal
siempre y cuando se cumplan los supuestos de las técnicas de captura–marcado–recaptura.
E
Antes de utilizar la técnica de
sus supuestos: (1) que la marca no afecta la supervivencia de los individuos y (2) que no
afecta el comportamiento de los individuos. Para ello se realizaron dos experiencias de
laboratorio durante enero de 2007.
S
capturaron obreras pertenecientes a tres colonias de cada una de las tres especies
estudiadas. De cada colonia se extrajeron 15 obreras, separándolas al azar en tres grupos
de cinco hormigas. Como el lugar en donde se aplica la marca puede influir en su efecto
sobre la supervivencia (según la cercanía de la pintura a los espiráculos y la complejidad
14

de manipuleo que requiera cada tipo de marca), se decidió marcar a las hormigas tanto
en el tórax como en el abdomen (siguiendo el procedimiento explicado más arriba), para
evaluar posibles diferencias. Por lo tanto, a las hormigas del primer grupo se las marcó
en el abdomen, a las del segundo grupo en el tórax y a las del tercero no se las marcó.
Luego se colocaron los tres grupos de cada colonia en un mismo formicario. Cada
formicario consistió de una bandeja plástica de 10 x 15 cm, con tierra en la base
(profundidad: 1 cm), extraída de los mismos sitios donde se encontraban las colonias, y
con una tapa perforada para permitir la ventilación. Se mantuvo a los formicarios en el
laboratorio durante siete días, en condiciones adecuadas de humedad y con semillas de
Setaria italica como alimento para las hormigas. Se calculó la proporción de hormigas
vivas por tratamiento (i.e., marca en el abdomen, marca en el tórax y sin marca) al cabo
de siete días.
Para analizar si existen diferencias en la supervivencia de las tres especies dependiendo
omportamiento.— Para poner a prueba el supuesto de que la marca no afecta el
del tipo de marca, la variable obtenida (proporción de hormigas vivas) fue analizada
mediante un ANOVA de dos factores (previa transformación aplicando el arcoseno de
la raíz cuadrada para que se cumplieran los supuestos de normalidad y homocedacia).
Se tuvieron en cuenta tres niveles para el factor especie: Pogonomyrmex pronotalis, P.
inermis y P. rastratus; y tres para el factor “tipo de marca”: en el abdomen, en el tórax y
sin marcar.
C
comportamiento de los individuos se usaron obreras de tres colonias de cada una de las
tres especies estudiadas. Se capturaron 10 obreras de cada colonia y se las llevó al
laboratorio, en donde fueron colocadas en formicarios, iguales a los descriptos en la
sección anterior, para observar su comportamiento. Las interacciones entre individuos
fueron categorizadas como: “agresión”, cuando un individuo tomaba a otro con sus
mandíbulas; “anteneo”, cuando dos individuos tocaban mutuamente sus antenas; “toque
otorgado”, cuando el individuo observado tocaba con sus antenas el cuerpo de otro
individuo; y “toque recibido”, cuando el individuo observado era tocado en el cuerpo
por otro individuo. Las hormigas se comunican usando feromonas, las cuales perciben
con sus antenas. Cuando una hormiga usa las antenas para tocar a otra, ya sea en las
antenas o en el cuerpo, puede recibir mensajes químicos y así determinar si la hormiga
15

con la cual se contacta es de la misma colonia (Gordon et al. 1993). Si la pintura
afectara las claves de reconocimiento entre individuos, se esperaría encontrar una mayor
frecuencia de contactos y anteneos, o incluso interacciones agresivas entre ellos.
Luego de un periodo de adaptación al formicario (3 minutos), se eligió al azar un
ebido a la complejidad del diseño y al bajo número de réplicas (tres colonias de cada
eterminación del tamaño de colonia por excavación
l de las colonias (i.e., el número
individuo y se hizo un seguimiento puntual, registrando todas las interacciones en las
que participaba durante un minuto. Este procedimiento se repitió tres veces
consecutivas, eligiendo cada vez un individuo del grupo al azar. A continuación se
retiraron cinco individuos, se los marcó en el abdomen y se los devolvió al formicario.
Se esperaron 3 minutos para permitir que las hormigas se adaptaran y se volvió a
registrar consecutivamente las interacciones de tres individuos marcados y de tres
individuos no marcados.
D
especie) no se realizó un análisis estadístico de los resultados. En su lugar, se
describieron y analizaron las tendencias observadas.
D
Para obtener una estimación aproximada del tamaño tota
total de obreras) se excavaron dos colonias de cada una de las tres especies estudiadas, a
las cuales previamente se les había estimado el número de obreras forrajeras. Las
excavaciones se realizaron 1–6 días después del último muestreo de captura–marcado–
recaptura, comenzando cuando los nidos estaban inactivos (Pol & Lopez de Casenave
2004). Se comenzó removiendo la capa superior de tierra, delimitando el área alrededor
de la entrada del nido en la cual se encontraban túneles o cámaras. Luego se removió
sistemáticamente la tierra por capas, capturando todos los individuos (larvas, adultos y
pupas) que aparecían. La tierra extraída fue tamizada para verificar la captura de todas
las hormigas de la colonia. La excavación continuó hasta una profundidad de varias
decenas de centímetros después de que ya no se encontraban más hormigas o signos de
ellas (túneles o cámaras). Se contaron todas las hormigas capturadas, discriminando
entre los distintos grados de desarrollo (larva, pupa o adulto) y su historia de captura
(marcada o no marcada). Para cada colonia se calculó la proporción de obreras
marcadas respecto del total de obreras encontradas.
16

Relación entre la abundancia de forrajeras y los niveles de actividad
ras forrajeras se En cada una de las colonias en las que se estimó el número de obre
registró el nivel de actividad forrajera durante 4–5 ocasiones de muestreo, antes de
comenzar los eventos de captura y marcado. Se contó el número de obreras forrajeras
que entraban y el número de forrajeras que salían del nido durante 3 minutos para P.
inermis y P. pronotalis y 6 minutos para P. rastratus (ver Pol & Lopez de Casenave
2004). Se consideró que un individuo regresaba de forrajear o abandonaba el nido para
forrajear cuando estaba entrando o saliendo, respectivamente (con o sin carga), de un
círculo imaginario de 35 cm de diámetro centrado en la entrada del nido. Al mismo
tiempo, se registró la temperatura de la superficie del suelo cerca de la entrada del nido
para asegurarse de que las mediciones se hicieran dentro de los rangos de temperatura
en los cuales la actividad de cada especie es máxima (ver Pol & Lopez de Casenave
2004). Los datos provenientes de las distintas ocasiones de muestreo se promediaron
para obtener un índice del nivel de actividad para cada colonia. Para evaluar la relación
entre el nivel de actividad y el número de obreras forrajeras de cada colonia se realizó
una correlación entre ambas variables con la Prueba de Spearman (Siegel & Castellan
1988).
17
Resultados
Evaluación de los supuestos
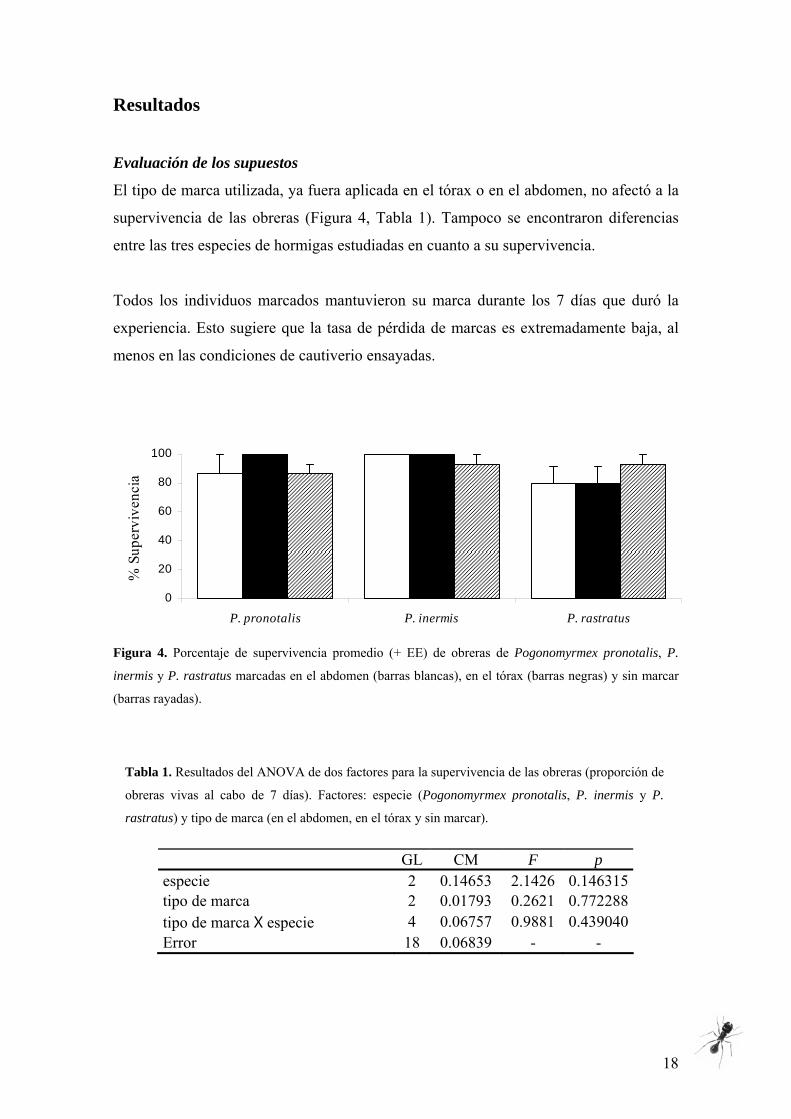
El tipo de marca utilizada, ya fuera aplicada en el tórax o en el abdomen, no afectó a la
supervivencia de las obreras (Figura 4, Tabla 1). Tampoco se encontraron diferencias
entre las tres especies de hormigas estudiadas en cuanto a su supervivencia.
Todos los individuos marcados mantuvieron su marca durante los 7 días que duró la
experiencia. Esto sugiere que la tasa de pérdida de marcas es extremadamente baja, al
menos en las condiciones de cautiverio ensayadas.
0
20
40
60
80
100
P. pronotalis P. inermis P. rastratus
% S
uper
vive
ncia
Figura 4. Porcentaje de supervivencia promedio (+ EE) de obreras de Pogonomyrmex pronotalis, P.
inermis y P. rastratus marcadas en el abdomen (barras blancas), en el tórax (barras negras) y sin marcar
(barras rayadas).
Tabla 1. Resultados del ANOVA de dos factores para la supervivencia de las obreras (proporción de
obreras vivas al cabo de 7 días). Factores: especie (Pogonomyrmex pronotalis, P. inermis y P.
rastratus) y tipo de marca (en el abdomen, en el tórax y sin marcar).
GL CM F p
especie 2 0.14653 2.1426 0.146315 tipo de marca 2 0.01793 0.2621 0.772288 tipo de marca X especie 4 0.06757 0.9881 0.439040 Error 18 0.06839 - -
18

Durante las experiencias comportamentales no se registraron conductas agresivas de
ningún tipo entre individuos de una misma colonia, ni antes ni después del marcado.
Las otras tres categorías de interacción (toque a otro individuo, anteneo y toque
recibido) fueron registradas tanto antes como después de marcar a las hormigas. No se
observaron tendencias de aumento ni de disminución en el número promedio de
interacciones antes y después del marcado, así como tampoco se observaron diferencias
importantes en el comportamiento entre hormigas marcadas y no marcadas cuando se
encontraban juntas (Figura 5).
Figura 5. Número promedio (± DE) de toques otorgados a otros individuos (arriba), de anteneos (centro)
y de toques recibidos (abajo) registrados en las experiencias comportamentales con P. pronotalis, P.
inermis y P. rastratus, antes del evento de marcado (cuadrados negros) y después del mismo, para
hormigas marcadas (círculos blancos) y no marcadas (círculos negros).
0
1
2
3
4
Toca
das
P. inermisP. pronotalis
0
1
2
3
4
Toqu
e
P. rastratus
sTo
ques
oto
rgad
osTo
ques
oto
rgad
os
0
2
4
6
Ant
eneo
sTo
ques
reci
bido
sTo
ques
reci
bido
s
Antes Después Antes Después Antes Después
19

Abundancia de obreras forrajeras
estimado mediante el índice de Schnabel resultó
Tabla 2. Abundancia de obreras forrajeras en colonias de Pogonomyrmex pronotalis, P. inermis
Colonia Petersen Schnabel
El número de obreras forrajeras
siempre mayor que el estimado con el de Petersen (Tabla 2). No se observaron
diferencias importantes en la precisión de la estimación realizada con el Método de
Petersen y el de Schnabel, ya que ambos presentaron intervalos de confianza de
amplitud similar.
y P. rastratus estimada mediante el Método de Petersen y el Método de Schnabel. Entre
paréntesis se muestran los límites superior e inferior del intervalo de confianza del 95% y entre
corchetes el número de muestreos sucesivos realizados para la estimación con el Método de
Schnabel.
P. pronotalis P3 52 (45-60) 109 (97-124) [10]
242 (237-247) rmis
15 (11-18) 35 (29-44) [10]
stratus 39 (33-45) 51 (42-65) [7]
133 (119-151) [10]
128 (117-140)
P6 68 (59-77) 103 (92-117) [10] P5 89 (82-96) 155 (140-173) [10] P2 90 (79-100) 203 (186-222) [10] P4 361 (337-388) [10] P. ine I10 I9 44 (20-68) 54 (46-64) [10] I5 56 (39-73) 88 (74-109) [11] I8 71 (60-81) 104 (91-122) [10] I7 96 (83-109) 125 (110-144) [10] P. ra R8 R7 42 (34-49) 63 (51-84) [9] R5 78 (54-102) R4 96 (81-112) 124 (108-146) [10] R6 156 (140-175) [8]
o se encontraron diferencias significativas entre las tres especies en cuanto al número
N
de obreras forrajeras de sus colonias, ya sea estimado mediante el Método de Petersen
(KW = 1.36; p > 0.10) o mediante el Método de Schnabel (KW = 4.22; p > 0.10). Sin
embargo, las colonias de P. pronotalis mostraron una tendencia a presentar un mayor
número de forrajeras, mientras que las de P. inermis tendieron a tener menos forrajeras.
20

Ambos métodos proveen una estimación similar de la abundancia relativa de forrajeras
entre colonias: al ordenar las colonias de una misma especie en función de su
abundancia creciente de obreras forrajeras se obtuvo el mismo ordenamiento con los dos
estimadores, a excepción de los pares de colonias P3-P6 y R5-R4 (que en ambos casos
superponen sus intervalos de confianza; Tabla 2).
La proporción de hormigas recapturadas aumentó a medida que se incrementaba el
xcavaciones
nidos se observó que las primeras cámaras se encuentran a muy escasa
lgunas de las cámaras superiores de los nidos de P. pronotalis contenían restos de
número acumulado de forrajeras marcadas (Figura 6). Sin embargo, en la mayor parte
de los casos no se observó la relación lineal esperada (Figura 3) entre ambas variables.
En general, la proporción de recapturas aumentó en forma desacelerada, generando
gráficos curvados, y raramente se obtuvieron muestras con todos los individuos
marcados, sino que la proporción se mantenía en valores cercanos a 0.8–0.9 a pesar de
que el número acumulado de marcadas seguía aumentando. Como consecuencia, la
abundancia de forrajeras estimada con el Método de Schnabel para una misma colonia
fue mayor a medida que aumentaba el número de muestreos sucesivos considerados (y
la duración del período de muestreo; Figura 7). Estos dos patrones sugieren que uno o
más de los supuestos de la técnica de captura–marcado–recaptura no se cumplen.
E
Excavando los
profundidad (2–3 cm), conectadas entre sí por túneles y formando una red que abarca un
radio de aproximadamente 40 cm alrededor de la entrada del nido. A medida que
aumenta la profundidad, va disminuyendo el número de cámaras y de túneles hasta que
solamente se pueden encontrar uno o dos túneles verticales que se ensanchan
periódicamente formando cámaras de tamaño variable. La profundidad de las
excavaciones varió entre 1–1.5 m, llegándose a remover hasta 2 m3 de suelo.
A
semillas y sus coberturas (glumas, palea y lemma), tratándose probablemente de
basureros internos. Estos basureros no fueron encontrados en los nidos de P. inermis
(que tienen basureros externos) ni en los de P. rastratus (cuyas obreras descartan las
estructuras que envuelven las semillas de varias de las especies que consumen antes de
llevarlas al nido; Pirk & Lopez de Casenave 2006, Pirk et al. 2007). Cuando se
21

P. pronotalis P. inermis P. rastratus
P2
0
0,5
1
0 50 100 150 200 250
P3
0
0,5
1
0 50 100 150
P4
0
0,5
1
0 100 200 300 400
P5
0
0,5
1
0 50 100 150 200
P6
0
0,5
1
0 30 60 90 120
I5
0 20 40 60 80
I7
0 50 100 150
I8
0 30 60 90 120
I9
0 20 40 60
I10
0 10 20 30 40 50
R4
0 50 100 150
R5
0 50 100 150
R6
0 40 80 120 160
R7
0 20 40 60 80
R8
0 20 40 60
Marcadas acumuladas
Prop
orci
ón d
e re
capt
urad
as
Figura 6. Proporción de obreras forrajeras de P. pronotalis, P. inermis y P. rastratus recapturadas (mi / ni) en
función del número acumulado de forrajeras previamente marcadas (Mi) a lo largo del muestreo para cada una
de las colonias estudiadas.
22

I5
40
50
60
70
80
90 R4
80
90
100
110
120
130
140P2
60
100
140
180
220
I7
80
90
100
110
120
130 R5
80
90
100
110
120
130
140P3
40
60
80
100
120
I8
60
70
80
90
100
110 R6
110
120
130
140
150
160
170P4
200
240
280
320
360
I9
10
20
30
40
50
60 R7
20
30
40
50
60
70
80P5
80
100
120
140
160
igura 7. Abundancia de obreras forrajeras de P. pronotalis, P. inermis y P. rastratus estimada mediante el F
Método de Schnabel a medida que aumenta el número de muestreos sucesivos para cada una de las colonias
estudiadas.
P6
50
70
90
110
130
0 2 4 6 8 10
I10
0
10
20
30
40
50
0 2 4 6 8 10
R8
20
30
40
50
60
70
80
0 2 4 6 8 10
Ocasiones de muestreo
P. pronotalis P. inermis P. rastratus A
bund
anci
a
23

encontraron granarios (cámaras en donde las hormigas acumulan las semillas), éstos
generalmente estaban ubicados a profundidades intermedias. Tanto las larvas como las
pupas fueron encontradas juntas a todas las profundidades, sin mostrar un patrón
evidente de distribución vertical. En algunos nidos, en especial en los de P. rastratus, se
hallaron larvas y pupas en las mismas cámaras que contenían las semillas.
En todos los nidos excavados, más del 60% de la colonia estaba compuesta por obreras,
de las cuales solo una pequeña fracción (5–13%) estaban marcadas (Tabla 3). La
mayoría de las obreras marcadas fueron encontradas en las capas más superficiales del
nido, mientras que en las zonas más profundas solo se encontraron obreras no marcadas.
A excepción de la colonia I7 de P. inermis, en el resto se encontraron larvas y pupas,
que constituyeron el 12–36% de los individuos de la colonia. El número de pupas fue
siempre ligeramente mayor que el de larvas. En tres de los nidos excavados se encontró
un adulto con el gáster ligeramente más desarrollado que el resto y las alas cortadas, que
podría ser la reina de la colonia.
Tabla 3. Resultado de la excavación de nidos de Pogonomyrmex pronotalis, P. inermis y P.
rastratus. Para cada colonia se muestran los números de obreras, adultos alados, larvas y pupas
encontrados. Se discrimina entre obreras marcadas y no marcadas, y se indica el porcentaje que las
primeras representan del total. También se muestra el número total de obreras del nido que fueron
marcadas a lo largo del muestreo de captura–marcado–recaptura (CMR) y el porcentaje de éstas que
fue recuperado durante la excavación. El asterisco indica que uno de los adultos alados encontrado
podría ser la reina activa de la colonia.
P. pronotalis P. inermis P. rastratus P2 P3 I5 I7 R4 R5 Obreras 691 538 378 219 916 1077 No marcadas 597 496 350 201 867 1017 Marcadas 94 42 28 18 49 60 Porcentaje marcadas (%) 13.6 7.8 7.4 8.2 5.3 5.6 Marcadas durante CMR 230 127 78 142 123 150 Marcadas encontradas (%) 41 38 36 13 40 40 Adultos alados 1 0 1 * 0 1 * 6 *
Larvas 159 87 22 0 100 156 Pupas 234 104 32 0 114 165
24

El número de obreras marcadas que fueron encontradas en los nidos durante las
excavaciones fue siempre menor que el total de obreras marcadas durante el muestreo
previo por captura–marcado–recaptura (Tabla 3). Esta fracción representó alrededor del
40% del total de individuos marcados en todas las colonias excepto en la colonia I7 de
P. inermis, donde la diferencia fue mayor (13%). Esto también indica que uno de los
puestos de la técnica de captura–marcado–recaptura no se cumple: la población de
.8 °C) que en P. pronotalis y P. inermis (37.7 °C y
9.8 °C, respectivamente), reflejando los distintos rangos de tolerancia térmica de estas
species.
L e
activ
P e
am e
en número de obreras forrajeras de sus colonias
(rs = 0.6, p > 0.10 en ambos casos).
su
forrajeras no es cerrada o hay mortalidad durante el período de estimación. El tiempo
transcurrido entre el primer evento de marcado y la excavación de los nidos fue de 11–
17 días (solo 1–6 días desde el último evento de marcado).
Niveles de actividad
Las temperaturas registradas durante las mediciones se encontraron dentro del rango en
el cual la actividad de cada una de las especies estudiadas es máxima (ver Pol & Lopez
de Casenave 2004). Los niveles de actividad de P. rastratus fueron medidos a una
menor temperatura promedio (28
3
e
as colonias con mayor número de obreras forrajeras tendieron a presentar niveles d
idad más elevados en las tres especies estudiadas, especialmente en P. rastratus y
. inermis (Figura 8). Sin embargo, solo se encontró una correlación significativa entr
bas variables en P. rastratus (rs = 0.9, p < 0.05). En P. pronotalis y P. inermis no s
contró correlación entre la actividad y el
25

0
10
20
30
40
50
30
40
Figura 8. Relación entre la abundancia de obreras forrajeras (y su intervalo de confianza
del 95%), estimada mediante el Método de Petersen, y el nivel de actividad promedio (±
EE) en colonias de Pogonomyrmex pronotalis, P. inermis y P. rastratus.
0
10
20
0
10
vida
P. pronotalis
P. inermis
dA
cti
20
30
40 P. rastratus
0 50 100 150 200 250
Abundancia de forrajeras
26

Discusión
Aplicabilidad de las técnicas de captura–marcado–recaptura
Para poder aplicar correctamente las técnicas de captura–marcado–recaptura en la
estimación de tamaños poblacionales deben cumplirse sus supuestos. Dos de ellos son
que el procedimiento de marcado y la marca en sí no afectan el comportamiento ni la
expectativa de vida de los individuos marcados. Si bien en otros trabajos ya se ha
comprobado que la pintura para aeromodelismo no afecta la supervivencia (Kruk-de
Bruin et al. 1977) ni el comportamiento (Gordon 1989, Brown & Gordon 1997) de
varias especies de hormigas, los resultados podrían variar según el tipo de pintura
utilizada y las características de las hormigas (e.g., su tamaño). Los marcadores de
pintura al óleo empleados en este estudio ya fueron usados por Gordon et al. (2005)
para marcar obreras de Pogonomyrmex barbatus, aunque sin comprobar previamente la
inocuidad del marcado. En este trabajo se corroboró que el marcado cuidadoso con
pintura al óleo (ya sea aplicada en el tórax o en el abdomen) no afecta la supervivencia a
corto plazo de las obreras de Pogonomyrmex pronotalis, P. inermis y P. rastratus, al
menos en condiciones de laboratorio. Los resultados indican también que este tipo de
marca no influye sobre el comportamiento de las hormigas que la portan ni sobre el de
sus compañeras no marcadas que interactúan con ellas. En el campo, las hormigas
pintadas fueron encontradas forrajeando normalmente al poco tiempo de ser liberadas y
tampoco allí se observaron interacciones agresivas entre individuos marcados y no
mar a alta
dur as que
dur ron hormigas pintadas
hasta tres meses después de marcarlas. Otra ventaja de este tipo de marca es que, si las
hormigas se manipulan adecuadamente durante el procedimiento de marcado, no puede
ser transmitida de un individuo a otro. Otros marcadores, como el P32 usado por Golley
& Gentry (1964), aunque duraderos, se transmiten a otras hormigas dentro del nido,
llevando a estimaciones erróneas de abundancia.
Los dos métodos de captura–marcado–recaptura usados para estimar el número de
forrajeras en las colonias de P. pronotalis, P. inermis y P.rastratus arrojaron resultados
diferentes. Utilizando el Método de Schnabel siempre se obtuvieron abundancias
cados de una misma colonia. La marca utilizada, además, mostró un
abilidad. Ningún individuo perdió su marca en el laboratorio durante los 7 dí
ó el experimento de supervivencia, y en el campo se encontra
27

mayores que al aplicar el Método de Petersen para las mismas colonias. Esto indica que
no de los dos estimadores, o ambos, están subestimando o sobreestimando el número
os, el procedimiento de
7). El mismo patrón fue registrado por
olley & Gentry (1964) al realizar múltiples muestreos en colonias de Pogonomyrmex
u
de forrajeras. Si bien ambos métodos comparten sus supuest
obtención de datos es diferente y esto puede afectar la magnitud de importancia de
supuestos no válidos. Para aplicar el método de Petersen solo se precisan dos eventos de
muestreo, mientras que para el de Schnabel se deben realizar varios muestreos sucesivos
(entre 7 y 11, en este caso). Es necesario que transcurra cierto tiempo entre muestreos
sucesivos debido a la inevitable perturbación de la actividad de las forrajeras durante el
procedimiento de captura, así como para favorecer el supuesto de la técnica de que los
individuos marcados se mezclan en la población de interés hasta que su probabilidad de
captura sea la misma que la de los individuos no marcados. Dado que los períodos de
actividad de estas hormigas son relativamente cortos (ver Pol & Lopez de Casenave
2004) eso suele implicar que los muestreos consecutivos se hagan en días sucesivos.
Así, el método de Petersen se puede aplicar en solo dos días, mientras que el de
Schnabel abarca tantos días como muestreos sean necesarios. Esto implicaría una
diferencia entre los métodos en su sensibilidad al incumplimiento del supuesto de que la
población es cerrada. Cuando el tiempo que transcurre entre el primer y el último
muestreo es corto, es plausible suponer que no ocurra reclutamiento o muerte de
hormigas forrajeras, o que éstos sean tan bajos como para ser despreciables. Pero
cuando este período es más largo (algo inevitable al usar el Método de Schnabel) el
sesgo puede ser más importante. Bajo estas condiciones, este método arrojaría
estimaciones sesgadas del número de forrajeras (Sutherland 1996).
El incremento desacelerado en la proporción de hormigas recapturadas a medida que
aumenta el número de muestreos (y, por lo tanto, el número acumulado de obreras
marcadas), observado al aplicar el Método de Schnabel (Figura 6), permite detectar la
invalidez de alguno de los supuestos de la técnica de captura–marcado–recaptura.
Asociado a esto, al incorporar más eventos de muestreo a la estimación se obtienen
valores de abundancia cada vez mayores (Figura
G
badius en Carolina del Sur (EEUU) utilizando P32 como marca. Estos autores
propusieron que sus resultados se debieron a que la marca radioactiva es fácilmente
transferible entre individuos y a que las hormigas marcadas no se mezclan rápidamente
28

con el resto de la población. La primera explicación no puede ser aplicada a los datos de
este trabajo, dado que la pintura al óleo empleada no se transfiere entre las obreras. La
segunda explicación sería válida si la población de interés fuera la colonia entera, pero
no cuando se hace referencia a las obreras externas, las cuales rara vez se mezclan con
el resto de la colonia (Kruk-de Bruin et al. 1977, Porter & Jorgensen 1980, 1981, Tripp
et al. 2000). Las diferentes actividades fuera del nido (patrullaje, mantenimiento del
nido y forrajeo) son llevadas a cabo por distintos grupos de obreras que mantienen
relativamente constante su asignación de tareas (Gordon 1989, Tripp et al. 2000) y que,
como mostraron Kruk-de Bruin et al. (1977), se mezclan muy bien entre ellas al cabo de
un día. Aunque las obreras pueden cambiar de tarea, esto sucedería en ocasiones muy
particulares, como perturbaciones de la colonia o cambios en la oferta de alimento
(Gordon 1989). Por lo tanto, es improbable que en las colonias estudiadas las obreras
forrajeras marcadas no se hayan entremezclado con las no marcadas entre días de
muestreo y, por consiguiente, que ese haya sido el factor que afectó las estimaciones del
Método de Schnabel.
Durante las excavaciones, el número de obreras marcadas que se recuperaron en cada
nido fue muy similar y bastante menor que el total de obreras marcadas durante el
muestreo de captura–marcado–recaptura (alrededor de un 40% en 5 de los 6 nidos
excavados). Esto sugiere que, consistentemente, cerca del 60% de las hormigas
marcadas en cada nido durante los muestreos murió o perdió la marca. Como la tasa de
pérdida de marcas a corto plazo en condiciones de laboratorio fue nula (ver arriba), es
uy probable que corresponda a eventos de mortalidad de hormigas forrajeras. En m
general, la división de tareas dentro de las colonias de especies del género
Pogonomyrmex está asociada a la edad de las obreras: las más jóvenes se encargan de
las labores que se realizan enteramente dentro del nido, como el cuidado de las larvas,
mientras que las más viejas llevan a cabo las tareas en el exterior (Porter & Jorgensen
1981, Gordon 1996, 1999). Esta progresión con la edad desde obreras que trabajan en el
nido hasta forrajeras externas está acompañada por cambios en el peso corporal y el
desgaste de las mandíbulas (Porter & Jorgensen 1981, Tschinkel 1998). Estas
características llevan a considerar a las forrajeras como el grupo con mayor probabilidad
de mortalidad en la colonia (Porter & Jorgensen 1981). Se desconoce cuál es la duración
promedio de vida de las forrajeras de las tres especies de hormigas estudiadas, pero se
29

ha reportado que las obreras externas de Pogonomyrmex owyheei viven en promedio
unos 14 días (Porter & Jorgensen 1980, 1981). Si las obreras forrajeras de las tres
especies de Ñacuñán tuvieran niveles de mortalidad similares, seguramente muchas de
las hormigas marcadas en los primeros días de muestreo habrían muerto antes del
último evento de recaptura (aproximadamente 15 días más tarde). Aún cuando el
tamaño de la población de forrajeras se mantenga relativamente constante (y, por lo
tanto, represente un valor estimable por estas técnicas), reemplazándose las forrajeras
muertas en este lapso de tiempo por nuevas obreras reclutadas a esa tarea (como sucede
en Formica polyctena; Kruk-de Bruin et al. 1977), se obtendría una situación análoga a
la “pérdida de marca” (i.e., obreras marcadas se reemplazan por no marcadas). Eso
produciría un incremento desacelerado en la proporción de recapturas y estimaciones de
abundancia mayores al incorporar más eventos de muestreo, o al incrementar el período
de tiempo abarcado, cuando se utiliza el Método de Schnabel. En consecuencia, los
resultados obtenidos sugieren fuertemente que el Método de Schnabel sobreestima la
abundancia de forrajeras en las colonias de las especies estudiadas al ser más extendido
n el tiempo que el de Petersen. e
En resumen, el Método de Petersen parece más confiable pues requiere solo dos eventos
de muestreo y por lo tanto permite reducir los efectos negativos de la mortalidad de las
forrajeras (y, eventualmente, la posible pérdida de marcas) haciendo la recaptura poco
después del marcado. Por el contrario, la aplicación exclusiva de este método no
hubiese permitido detectar la invalidez de uno de los supuestos (i.e., que la población de
forrajeras es cerrada), así como tampoco permite estimar el sesgo que posee la
estimación del tamaño de la población de hormigas forrajeras. Merece destacarse que
ambos métodos proveen una evaluación similar de la abundancia relativa de obreras
forrajeras entre una serie de colonias de la misma especie. Cuando el objetivo del
estudio es conocer la abundancia relativa más que la abundancia absoluta de forrajeras
de una colonia (e.g., para correlacionarla con otra variable, ver abajo), cualquiera de los
dos métodos podría ser usado. No obstante, ante la obtención de resultados similares
seguiría siendo recomendable la utilización del de Petersen en términos de eficiencia de
tiempo y esfuerzo de muestreo.
Si bien las estimaciones mediante el Método de Petersen se ven poco afectadas por la
30

violación del supuesto de población cerrada (cuando se lo aplica en un intervalo corto
de tiempo), existen otros métodos de captura–marcado–recaptura que podrían aplicarse
en esta situación para obtener estimaciones más exactas. Un ejemplo es el Método de
Jolly–Seber, el cual fue diseñado para ser aplicado en poblaciones abiertas. Este método
requiere de al menos tres eventos de muestreo y de marcas específicas para cada ocasión
de marcado (e.g., usar un color de pintura distinto cada vez). De esta manera, a partir de
la “historia de captura” de cada individuo, se podría estimar no solo el tamaño de la
población, sino también la tasa de pérdida y de ganancia de individuos a la población.
Aplicando este método (u otro similar) podría obtenerse información más detallada
acerca de la dinámica de la población forrajera en las colonias de las tres especies
estudiadas.
Características de las colonias de las especies sudamericanas del género
Pogonomyrmex
Considerando la relación entre obreras marcadas y no marcadas revelada por las
excavaciones de los nidos y las estimaciones de abundancia de forrajeras obtenidas por
l Método de Petersen, el 7–15% del total de obreras de una colonia serían forrajeras.
tivas entre las tres especies en cuanto a la abundancia
e forrajeras en sus colonias, se puede observar que el número promedio de forrajeras
e
Esto coincide con lo encontrado en otras especies del mismo género que habitan en
América del Norte: en P. owyheei las forrajeras llegan a ser el 13% de la colonia (Porter
& Jorgensen 1981), mientras que en P. badius y P. occidentalis son el 10% (Golley &
Gentry 1964, Crist & MacMahon 1991). En muchos casos la abundancia de forrajeras
se correlaciona con el tamaño de la colonia (Porter & Jorgensen 1980), y hasta es
posible para algunas especies obtener un estimador grosero del tamaño de la colonia a
partir de la abundancia de forrajeras usando técnicas de regresión (Kruk-de Bruin et al.
1977).
Como sucede con Formica sp. (Kruk-de Bruin et al. 1977), la abundancia de forrajeras
fue variable entre colonias en las especies del género Pogonomyrmex estudiadas. Esto
probablemente refleje la variabilidad en la edad de las colonias muestreadas, aunque
también podría estar asociado a diferencias en el éxito de las mismas. Si bien no se
encontraron diferencias significa
d
en las colonias de P. pronotalis (108 individuos) es levemente mayor que en las de
31

P.rastratus (77 individuos) y de P. inermis (56 individuos). Los estudios en otras
especies del mismo género en América del Norte (P. badius y P. owyheei) muestran
abundancias promedio mayores, cercanas a las 200 forrajeras por colonia, aunque en
este caso se trata de especies con colonias grandes, de más de 5000 obreras (Golley &
Gentry 1964, Porter & Jorgensen 1980). Las especies del género Pogonomyrmex de
Ñacuñán tienen colonias pequeñas (300–1000 obreras), a juzgar por el resultado de las
excavaciones. Estos valores se asemejan a los recopilados por Kusnezov (1949, 1951)
para la mayoría de las especies sudamericanas del género. No obstante, aunque las
excavaciones permiten conocer el número total de obreras de la colonia, los valores
deberían ser tomados con cautela debido al bajo número de nidos excavados y a que en
las especies que tienen colonias con múltiples nidos la excavación de uno de ellos no
ecesariamente produce valores referidos a la colonia (Billick 1999). La ausencia de
do y no de una colonia entera, lo que también explicaría que el
Por el contrario, muchas de las especies norteamericanas construyen nidos
randes y forrajean en forma grupal (Johnson 2000). Estas diferencias tan acentuadas
n
larvas y pupas en la colonia I7 (ver Tabla 3) es un indicio de que probablemente se haya
tratado de un ni
porcentaje de obreras marcadas encontradas en ese nido fuera mucho menor que en los
otros. Coincidentemente, de las tres especies estudiadas, solo en P. inermis se ha
observado ocasionalmente la utilización de diferentes nidos por parte de obreras de una
misma colonia (Lopez de Casenave & Milesi, datos no publicados).
El hecho de presentar colonias pequeñas y con pocas forrajeras (como es el caso de las
tres especies estudiadas en este trabajo) es, según Kusnezov (1951), una característica
típica de las especies del género Pogonomyrmex que habitan en Argentina. Como
consecuencia del bajo número de obreras, sus nidos están poco desarrollados y sus
forrajeras tienden a buscar el alimento en forma solitaria, sin formar caminos (Kusnezov
1951).
g
podrían ser consecuencia del desarrollo independiente de las especies de este género en
América del Norte y América del Sur (i.e., las dos áreas de distribución actuales de
mayor importancia), lo cual se corresponde con una de las hipótesis acerca del origen
evolutivo del género, que ubica su centro de dispersión en América del Sur (Kusnezov
1951, Taber 1998). Las características biológicas de las especies estudiadas en este
trabajo coinciden con lo reportado para otras especies sudamericanas (Kusnezov 1951)
y apoyan la hipótesis de dispersión planteada. Sin embargo, son necesarios más estudios
32

acerca de la biología de estas especies para poder analizar en profundidad la historia
evolutiva del género en el continente americano.
Abundancia de forrajeras y actividad externa de la colonia
Los patrones diarios y estacionales de actividad de las distintas especies están
relacionados principalmente con sus rangos de tolerancia a la temperatura y la humedad
(Hölldobler & Wilson 1990). Además, en los momentos en que sus obreras externas
están activas, las colonias de una misma especie pueden diferir en sus niveles de
actividad dependiendo de factores como la predación (Gentry 1974), la disponibilidad
de alimento (Crist & MacMahon 1991, Gordon 1991, Schafer et al. 2006), o el número
de obreras forrajeras presentes en las mismas. Para las colonias de P. pronotalis y P.
rastratus de Ñacuñán, se encontró que la temperatura superficial del suelo es el
principal factor (aunque no el único) que determina sus niveles diarios y estacionales de
actividad (Pol & Lopez de Casenave 2004); probablemente lo mismo ocurra con P.
inermis, que tiene ciclos diarios de actividad muy similares (observación personal).
Además, existen diferencias en las tolerancias térmicas entre especies: P. pronotalis
forrajea a temperaturas más elevadas que P. rastratus (Pol & Lopez de Casenave 2004),
patrón que coincide con lo observado en este trabajo. Pol & Lopez de Casenave (2004)
mbién han registrado una alta variabilidad en los niveles de actividad de las distintas ta
colonias de P. pronotalis y P. rastratus, aunque estos autores no analizaron la posible
influencia del tamaño de la colonia sobre este patrón. La actividad de estas especies está
confinada a los períodos con temperaturas del suelo entre 20 y 55 °C, pero, al mismo
tiempo, se encontró que la temperatura del suelo no es un buen predictor de los niveles
de actividad dentro de ese rango (Pol & Lopez de Casenave 2004). En este trabajo, las
temperaturas registradas durante las mediciones de actividad de P. pronotalis, P.
inermis y P. rastratus se encuentran dentro de dicho rango y, por lo tanto, puede
suponerse que no es la temperatura el factor que explicaría las variaciones observadas
entre las distintas colonias en cuanto a sus niveles de actividad.
Los resultados de este trabajo muestran que existe una tendencia de las colonias con
más forrajeras a presentar niveles de actividad más elevados, tanto al comparar colonias
de una misma especie como comparando entre especies. La relación intraespecífica, sin
embargo, resultó estadísticamente significativa solo en P. rastratus; la falta de
33

significancia para las otras dos especies probablemente se debe a la baja potencia del
análisis estadístico empleado (bajo tamaño muestreal y técnica de correlación no
aramétrica muy conservativa). Al comparar entre especies, se observó que las colonias
mente mayor que las de P.
p
de P. pronotalis presentan una abundancia de forrajeras ligera
rastratus, lo que concuerda con sus niveles de actividad mayores (reportados en Pol &
Lopez de Casenave 2004), y apoya la hipótesis de la influencia del tamaño de colonia
sobre la actividad de las colonias.
34

Conclusiones
1. Las obreras de Pogonomyrmex pronotalis, P. inermis y P. rastratus no presentan
cambios en su comportamiento ni en su supervivencia cuando son marcadas con
pintura al óleo. De esta manera se comprobó que se cumplen dos de los supuestos de
los métodos de captura–marcado–recaptura empleados en este trabajo.
2. El método de Petersen parece adecuado para estimar la abundancia relativa de
forrajeras en las colonias de P. pronotalis, P. inermis y P. rastratus. Al requerir de
solo dos días de muestreo, se reducen los efectos que la mortalidad de las obreras (y la
potencial pérdida de marcas) puede generar sobre la estimación.
3. El Método de Schnabel es más sensible que el Método de Petersen al incumplimiento
del supuesto de que la población es cerrada, dado que, por las características
biológicas y comportamentales de las forrajeras de P. pronotalis, P. inermis y P.
rastratus, necesariamente implica un tiempo de muestreo más largo. Esto genera que
los valores obtenidos tiendan a sobreeestimar la abundancia absoluta de forrajeras. Sin
embargo este método sirve para estimar la abundancia relativa de forrajeras entre
colonias de una misma especie, además de brindar información útil acerca del
cumplimiento o no de los supuestos de la técnica.
4. Las colonias de P. pronotalis, P. inermis y P. rastratus son relativamente pequeñas.
Tienen aproximadamente unas 300–1000 obreras, de las cuales el 7–15% corresponde
a forrajeras. Estas características son típicas de las especies del género Pogonomyrmex
que habitan en Argentina y las diferencian de la mayoría de sus congéneres estudiadas
en América del Norte.
5. El nivel promedio de actividad forrajera de las colonias de P. rastratus se
correlaciona positivamente con la abundancia de forrajeras presentes en las mismas.
La misma tendencia se observa en P. inermis y P. pronotalis, aunque no se pudo
encontrar una correlación significativa entre ambas variables.
35

Bibliografía
Bailey, N.T.J. (1951) On estimating the size of mobile population from recapture data. Biometrika 38:293–306.
ubourg & J.M. Pasteels (1989) Colony size, communication and ant foraging strategy. Psyche 96:239–256.
ark-recapture to measure worker number in the rock
Beckers, R., S. Goss, J.L. Dene
Billick, I. (1999) The use of mnesting ant species, Formica neorufibarbis Emery. Insectes Sociaux 46:256–260.
B
rown, J.H., O.J. Reichman & D.W. Davidson (1979) Granivory in desert ecosystems.
B
C
Crist, T.O. & J.A. MacMahon (1992)
Diehl-Fleig, E. & E. Diehl (2007) Nest argrowing ant Mycetophylax simplex Emery, 1888 (Formicidae, Attini). Insectes
G
Gentry, J.B. (1974) Response to predation by colonies of the florida harvester ant, Ecology 55:1328–1338.
Gordon, D.M. (1989) Dyna
G ecology of seed-eating ants. American Naturalist 138:379–411.
Billick, I. (2001) Density dependence and colony growth in the ant species Formica neorufibarbis. Journal of Animal Ecology 70:895–905.
rian, M.V, J. Hibble & D.J. Stradling (1965) Ant pattern and density in a southern English heath. Journal of Animal Ecology 34:545–555.
BAnnual Review of Ecology and Systematics 10:201–227.
rown, M.J.F. & D.M. Gordon (1997) Individual specialization and encounters between harvester ant colonies. Behaviour 134:849–866.
rist, T.O. & J.A. MacMahon (1991) Foraging patterns of Pogonomyrmex occidentalis (Hymenoptera: Formicidae) in a shrub–steppe ecosystem: the roles of temperature, trunk trails, and seed resources. Environmental Entomology 20:265–275.
Harvester ant foraging and shrub-steppe seeds: interactions of seed resources and seed use. Ecology 73:1768–1779.
chitecture and colony size of the fungus-
Sociaux 54:242–247.
aglio, M.D., W.P. Mackay, E.A. Osorio & I. Iniguez (1998) Nest populations of Pogonomyrmex salinus harvester ants (Hymenoptera: Formicidae). Sociobiology 32:459–463.
Pogonomyrmex badius.
Golley, F.B. & J.B. Gentry (1964) Bioenergetics of the southern harvester ant, Pogonomyrmex badius. Ecology 45:217–225.
mics of task switching in harvester ants. Animal Behavior 38:194–204.
ordon, D.M. (1991) Behavioral flexibility and the foraging
36

Gordon, D.M. (1996) The organization of work in social insect colonies. Nature 380:121–124.
Hölldobler, B. & E.O. W mbridge.
36:89–122.
Pogonomyrmex
Kmpson (eds.)
Krebs, C.J. (1989)
al Ecology 46:457–470.
Gordon, D.M. (1999) Ants at work. How an insect society is organized. Norton & Company, Nueva York.
Gordon, D.M. (2002) The regulation of foraging activity in red harvester ant colonies. American Naturalist 159:509–518.
Gordon, D.M., R.E. Paul & K. Thorpe (1993) What is the function of encounter patterns in ant colonies? Animal Behavior 45:1083–1100.
Gordon, D.M., J. Chu, A. Lillie, M. Tissot & N. Pinter (2005) Variation in the transition from inside to outside work in the red harvester ant Pogonomyrmex barbatus. Insectes Sociaux 52:212–217.
ilson (1990) The ants. Harvard University Press, Ca
Johnson, R.A. (2000) Seed-harvester ants (Hymenoptera: Formicidae) of North America: an overview of ecology and biogeography. Sociobiology
Johnson, R.A., C.T. Holbrook, C. Strehl & J. Gadau (2007) Population and colony structure and morphometrics in the queen dimorphic harvester ant, pima. Insectes Sociaux 54:77–86.
emp, P.R. (1989) Seed banks and vegetation processes in deserts. Pp. 257–281 en: Ecology of soil seed banks. Allesio Leck, M., V.T. Parker & R.L. SiAcademic Press, San Diego.
Ecological Methodology. Harper Collins, Nueva York.
Kruk-de Bruin, M., L.C.M. Rost & F.G.A.M. Draisma (1977) Estimates of the number of foraging ants with the Lincoln-index method in relation to the colony size of Formica polyctena. Journal of Anim
Kusnezov, N. (1949) "Pogonomyrmex" del grupo "Ephebomyrmex" en la fauna de la Patagonia (Hymenoptera, Formicidae). Acta Zoológica Lilloana 8:291–307.
Kusnezov, N. (1951) El género "Pogonomyrmex" Mayr (Hym., Formicidae). Acta Zoológica Lilloana 11:227–333.
Le Cren, E.D. (1965) A note on the history of mark-recapture population estimates. Journal of Animal Ecology 34:453–454.
Lopez de Casenave, J. (2001) Estructura gremial y organización de un ensamble de aves del desierto del Monte. Tesis doctoral, Universidad de Buenos Aires, Buenos Aires.
37

Lopez de Casenave, J., V.R. Cueto & L. Marone (1998) Granivory in the Monte desert, Argentina: is it less intense than in other arid zones of the world? Global Ecology and Biogeography Letters 7:197–204.
MacMahon, J.A., J.F. Mull & T.O. Crist (2000) Harvester ants (Pogonomyrmex spp.): their community and ecosystem influences. Annual Review of Ecology and
Mailleux, A.C., J.L. Deneubourg & C. Detrain (2003) How does colony growth
Marone, L., J. Lopez de Casenave & V. Cueto (2000) Granivory in southern South
Pirk, G.I. & J. Lopez de Casenave (2006) Diet and seed removal rates by the harvester ants Pogonomyrmex rastratus and Pogonomyrmex pronotalis in the central Monte
ctoral, Universidad de Buenos Aires, Buenos Aires.
Pratus y P. inermis con caminos en el Monte central.
Ecología Austral
PPogonomyrmex spp.) diet. Environmental Entomology
9:247–256.
Roig, F.A. (1971) Flora y vegetación de la reserva forestal de Ñacuñán. Deserta 1:21–239.
Systematics 31:265–291.
influence communication in ants? Insectes Sociaux 50:24–31.
American deserts: conceptual issues and current evidence. BioScience 50:123–132.
desert, Argentina. Insectes Sociaux 53:119–125.
Pirk, G.I. (2007) Granivoría por hormigas del género Pogonomyrmex en el Monte Central: consumo de semillas e impacto sobre el banco de suelo. Tesis do
irk, G.I., J. Lopez de Casenave & R. Pol (2004) Asociación de las hormigas granívoras Pogonomyrmex pronotalis, P. rast
14:65–76.
irk, G.I., J. Lopez de Casenave & L. Marone (2007) Evaluation of three techniques for the study of harvester ant (36:1092–1099.
Pol, R. & J. Lopez de Casenave (2004) Activity patterns of harvester ants Pogonomyrmex pronotalis and Pogonomyrmex rastratus in the central Monte desert, Argentina. Journal of Insect Behavior 17:647–661.
Pol, R.G., J. Lopez de Casenave, H. Feldhaar, F.A. Milesi & J. Gadau (2008) Polyandry in two South American harvester ants. En prensa en Insectes Sociaux.
Porter, S.D. & C.D. Jorgensen (1980) Recapture studies of the harvester ant, Pogonomyrmex owyheei Cole, using a fluorescent marking technique. Ecological Entomology 5:263–269.
Porter, S.D. & C.D. Jorgensen (1981) Foragers of the harvester ant, Pogonomyrmex owyheei: a disposable caste? Behavioral Ecology and Sociobiology
Pulliam, H.R. & M.R. Brand (1975) The production and utilization of seeds in plains grassland of southeastern Arizona. Ecology 56:1158–1166.
38

Schafer, R.J., S. Holmes & D.M. Gordon (2006) Forager activation and food availability in harvester ants. Animal Behavior 71:815–822.
Schnabel, Z.E. (1938) The estimation of the total fish population of a lake. American
S estimation of animal abundance and related parameters. Segunda edición. Charles Griffin & Co., Londres.
S ce. Biometrics 42:267–292.
S al sciences. Segunda edición. McGraw-Hill, Singapur.
S Hall, Londres.
Tripp, J.M., D.R. Suiter, G.W. Bennett & J.H. Klotz (2000) Lincoln index derived
Tschinkel, W.R. (1998) Sociometry and sociogenesis of colonies of the harvester ant,
Tschinkel, W.R. (1999) Sociometry and sociogenesis of colonies of the harvester ant,
.
Whitford, W.G. (1978) Foraging in seed-harvester ants Pogonomyrmex spp. Ecology
Mathematical Monthly 45:348–50.
eber, G.A.F. (1982) The
eber, G.A.F. (1986) A review of estimating animal abundan
iegel, S. & N.J. Castellan Jr. (1988) Nonparametric statistics for the behavior
outhwood, T.R.E. (1978) Ecological methods with particular reference to the study of insect populations. Segunda edición. Chapman &
Stradling, D.J. (1970) The estimation of worker ant populations by the mark-release-recapture method: an improved marking technique. Journal of Animal Ecology 39:575–591.
Taber, S.W. (1998) The world of the harvester ants. Texas A&M University Press, College Station.
forager population estimates in the black carpenter ant, Camponotus pennsylvanicus (Hymenoptera: Formicidae). Sociobiology 35:109–118.
Pogonomyrmex badius: worker characteristics in relation to colony size and season. Insectes Sociaux 45:385–410.
Pogonomyrmex badius: distribution of workers, brood and seeds within the nest in relation to colony size and season. Ecological Entomology 24:222–240
Wagner, D., J.B. Jones & D.M. Gordon (2004) Development of harvester ant colonies alters soil chemistry. Soil Biology and Biochemistry 36:797–804.
59:185–189.
39

Agradecimientos
co or brindarme todo su apoyo y dedicación durante
F zar importantes aportes críticos. A
Rodrigo Pol, sin cuyo aporte no hubiera sido posible esta tesis. A todos los integrantes
es pañaron, me apoyaron y me
g ier, Fernando, María
es
ú
ca inigualable: Natalia, Hernán,
“e
Agradezco particularmente a mi director, Javier Lopez de Casenave, por tenerme la
nfianza y la paciencia necesarias, p
estos años y en todas las etapas de este trabajo. A Gabriela Pirk, Víctor Cueto y
ernando Milesi por revisar este manuscrito y reali
los que colaboraron “hombro a hombro” en el trabajo de campo: Bárbara Paván y
del Grupo de Investigación en Ecología de Comunidades de Desierto (ECODES), y
pecialmente a aquellos ecodianos que me acom
estimularon, tanto en el laboratorio como en el campo, y que forman este maravilloso
rupo humano con el cual comparto gran parte de mi tiempo: Jav
del Mar, Gabriela, Carolina, Taty, Víctor, Rodrigo y Federico. Quiero agradecer
pecialmente a Fernando Milesi por su interés, paciencia y dedicación; por estar
siempre dispuesto a brindar su ayuda y aportes, indispensables para el desarrollo de este
trabajo. Agradezco también mi familia (a mis padres y mi hermano) por apoyarme en
mis decisiones y estar siempre a mi lado, brindándome su cariño incondicional. Por
ltimo (pero no menos importante), quisiera agradecer a mis amigos y compañeros de la
rrera, por hacer de estos años una experiencia hermosa e
Lucas, María, Eugenia, Leticia, José, Osqui, Elisa, Mer, Mechi, mis queridos
copelelas” y todos los demás... No habría sido posible sin ellos.
40


















